Embed Size (px)
Citation preview

Enzyme and Microbial Technology 39 (2006) 1030–1035
Biodegradation of linear alkylbenzene sulfonate by a two-memberfacultative anaerobic bacterial consortium
Khaled M. Khleifat ∗Mutah University, Department of Biology, Karak, Mutah 61710, Jordan
Received 1 November 2005; received in revised form 29 January 2006; accepted 7 February 2006
Abstract
A bacterial consortium capable of degrading the linear alkylbenzene sulfonate (LAS) was isolated from the wastewater treatment plant. Thebacterial consortium consisted of two members, Pantoea agglomerans and Serratia odorifera 2. Cells were grown evenly together in a minimalmedium (M9) and nutrient broth (NB). The bacterial consortium was able to grow in the minimal medium containing LAS as the only carbonsource. The percentage degradation of 200 ppm LAS by this bacterial consortium was better when cells were grown in NB (∼70%) than in the M9medium (36%). Also, the degradation ability by the bacterial consortium was very much higher than by its individual cells. This work shows thatthe two bacteria complement each other in the degrading ability of LAS, indicating catabolic cooperation between the two consortium members.Ac©
K
1
fhamtaooSefb[afs
0d
n incubation temperature of 32 ◦C, an agitation rate of 250 rev min−1, and the addition of different carbon and nitrogen sources all independentlyaused complete mineralization of 200 mg L−1 LAS within 48–72 h.
2006 Elsevier Inc. All rights reserved.
eywords: LAS; Surfactants; Biodegradation
. Introduction
Linear alkylbenzene sulfonates (LAS) are widely used in sur-actant formulation. The biodegradability of these compoundsas been intensively studied, particularly in marine and wastew-ter treatment plants [1–4]. Most of the studies deal with pri-ary biodegradation. In this initial step of the biodegrada-
ion, short-chain sulfophenyl carboxylate (SPC) residue appearss transient intermediates after the first terminal oxidationf the alkyl chain, which is then shortened by means of �-xidation, causing the loss of surfactive characteristics [2].PC’s are then completely biodegraded in the second min-ralization step, achieved by other organisms through desul-onation and a ring-opening process [5–7]. Much work haseen done to confirm the complete biodegradability of LAS3,8], however, little is known about the degradation processesnd the microbial community involved [4]. The hypothesizedacile degradation of LAS is more complex than previouslypeculated. LAS is not a single compound, but, typically, a
mixture of 20 isomer compounds, all subterminally substi-tuted, a linear alkyl chain (C10–C13) carrying a 4-sulfophenylmoiety.
Most LAS is discarded as sewage and efficiently removed (upto 99%) during sewage treatment [9]; in some areas raw sewagethat contains LAS is discharged directly into the environment[10]. Thus the consequence of the insufficient biodegradationof LAS is the development of huge masses of foam in streamsand rivers in the vicinity of dams and other obstructions [8,11].The aim of this work was to study the ability of two bacteria(Pantoea agglomerans and Serratia odorifera 2) to facilitate thebiodegradability of LAS by individual isolates and their bac-terial consortium under different conditions. The two kinds ofbacteria had already been isolated from the wastewater treat-ment plant located on our University campus [12]. The condi-tions include pH, temperature, aeration and carbon and nitrogensources. This is therefore the first report describing these twospecies as being able to degrade LAS. P. agglomerans has beenpreviously described by many researchers [13,14]. It is ubiqui-tous in nature, animals and humans. Pantoea showed its bestresults in being able to use kerosene, toluene and vaseline as
∗ Tel.: +962 3 2372380; fax: +962 3 2375540.E-mail address: alkh [email protected].
the carbon sources for growth. It is gram-negative, noncapsu-lated, nonsporeforming and motile rode. S. odorifera 2 has been
141-0229/$ – see front matter © 2006 Elsevier Inc. All rights reserved.oi:10.1016/j.enzmictec.2006.02.007

K.M. Khleifat / Enzyme and Microbial Technology 39 (2006) 1030–1035 1031
described by Grimont et al. [15]. It is a member of the normalflora of the gram-negative, small rod-shaped bacteria of the orderEubacteriales. These motile peritrichous bacteria are commoninhabitants in water, soil, manure, bedding and feed. This groupcan be distinguished from other genera that belong to the familyEnterobacteriaceae by its production of three special enzymes:DNase, lipase and gelatinase.
2. Materials and methods
2.1. Materials
LAS (linear alkylbenzene sulfonate) was supplied by Jordan Sulpho-chemicals Co. Ltd., Jordan. It was an aqueous sodium salt solution with aminimum purity of 96.5% and an average molecular weight of 320 g mol−1
that was used in the preparation of working standards. Benzethoinium chloride(hyamine) and patent blue (disulfide blue) were supplied by Acros Organics,(Fisher Scientific, UK). Most of the chemicals used were either from Sigma,USA or from Fluka Chemika, Switzerland. Other chemicals were of analyticalgrade and were obtained from commercial suppliers.
2.2. Bacterial isolates
Two previously studied wastewater isolates, P. agglomerans (formerly Enter-obacter agglomerans [13]) and S. odorifera 2 were used in this study [12]. Theirmorphological characteristics were re-verified and their biochemical identitywas verified using the REMEL kit (RapIDTM ONE and RapIDTM NF plus sys-tems) procedure.
2
cwhwl
Swp
csatndld
2
tinpisoobNa
0.24 g MgSO4, 0.05 g CaCl2·6H2O, and 0.05 g FeCl3·6H2O. After autoclaving,the following were added by predissolving each in 2 mL distilled water and ster-ilizing by filtration through a 0.45 �m Millipore filter: 0.001 g thiamine–HCl.LAS was used in the concentration of 200 ppm as the carbon and energysource.
The initial cell mass (starting inocula) for each experiment in both the nutri-ent broth (NB) and minimal media (MM) experiments were grown from singlecolonies in two consecutive shake flask cultures (overnight, followed by 3 hrecovery) in nutrient broth. Cells from the 3-h cultures were harvested by cen-trifugation and then washed with and resuspended in NB at a concentration of 0.5A600 units. One milliliter of these bacterial cells was used to inoculate 100 mLof either media in 250 mL Erlenmeyer flasks for shake flask experiments. In thecase of the mixed culture (P–S), 1-mL inocula of the two bacterial cells havealways been taken to constitute a 50:50 co-culture (taking into considerationthe total number of bacterial cell per OD600). Samples were then taken at timeintervals and assayed for the cell mass OD600. In all experiments, acid-washedglassware was used.
2.5. Growth curves
Two different media, nutrient broth (NB) and M9 minimal medium (MM)were used for growth curves. Each medium was supplemented with 200 mg L−1
LAS; and in the minimal medium LAS was the only carbon source uti-lized. Growth curves experiments were performed on the NB and MMmedia under the same conditions. The same initial cell mass (starting inoc-ula) for the three cases was taken into consideration, as mentioned above.Samples were then taken at time intervals and assayed for their cell massOD600.
2.6. Effect of different growth conditions on LAS biodegradability
2r
6cwaub(c
cbT[
2
tstei
3
ttpubb
.3. Analytical methods
LAS (linear alkyl benzene sulfonate) was analyzed in culture media using thelassic methylene blue method [17], modified by Li and Rosen [18]. The LASas extracted in an organic solvent (chloroform) and titrated against standardizedyamine (benzethonium) in the presence of patent (anionic dye). The endpointas detected by the pink to blue oily droplets colour development of the organic
ayer.The disappearance of LAS was qualified by reverse-phase HPLC on a
pherisorb ODS2 column (250 by 4.6 mm; particle size, 5 �m); the motile phaseas 0.11 M NaClO4 with an acetonitrile gradient (0–100%; vol./vol.), and com-ounds were detected spectrophotometrically at 255 nm [11].
In order to compare the degradation ability achieved between two differentonditions, the percentage degradation (%) of the LAS compared to the corre-ponding control was conducted. In other conditions such as Figs. 3 and 7, theverage of degradation rates of LAS was measured by dividing the net amount ofransformed LAS for 24 h, since within this time period there was many havingo further degradation, such as in Fig. 3. The reason for calculating the averageegradation by this method to avoid any erroneous caused by different lengths ofag phases, and the difficulty to ascertain the time required achieving completeegradation or when the degradation had stopped.
.4. Growth
In all experiments conducted, the LAS-containing uninoculated media wereaken as the control. The nutrient broth (NB) and agar media were used for thesolation of bacteria and all degradation tests performed. The nutrient agar andutrient broth were supplied by Difco. The ingredients of NB (g L−1) are 5 geptone, 5 g sodium chloride, 1.5 g beef extract and 1.5 g yeast extract. In thenitial experiments, the LAS compound was used as the sole carbon and energyources by including it in the M9 minimal medium (MM), which has no otherrganic compounds. This medium was used to further verify the degradabilityf LAS by the bacterial consortium. The minimal broth medium, as describedy Miller [16], consists of the following: 3.0 g Na2HPO4, 1.5 g KH2PO4, 1.0 gH4Cl, and 0.5 g NaCl were dissolved in 500 mL distilled water and the pH
djusted to 7.4 with 6N NaOH. Then the following compounds were added:
.6.1. Effect of substrate concentration, pH, temperature and agitationate
The effect of the different substrate concentrations (200, 300, 400, 500,00 and 700 ppm) on the LAS removal by the bacterial consortium (P–S) wasonducted. The growth medium (NB plus corresponding LAS concentration)as incubated at 37 ◦C incubation temperature, under a 150 rpm shaking rate
nd a pH of 7.5. Different pHs (5.5, 6.5, 7.5 and 8.5) of the growth media weresed to assess the effect of variations in pH on the percentage degradation of LASy bacterial combination (P–S). The effect of different incubation temperatures25, 32, 37 and 42 ◦C) upon the percentage degradation of LAS by bacterialonsortium (P–S) was investigated.
To test the effect of agitation rates on the LAS degradation by the bacterialonsortium (P–S), the NB medium was used, as mentioned previously, to growacterial cells at 37 ◦C under three different agitation rates: 75, 150 and 250 rpm.hese agitation rates are referred to as low, normal and high aeration, respectively
25].
.6.2. Effect of different carbon and nitrogen sourcesDifferent carbon and nitrogen sources were supplied to the NB to study
heir effect on the biodegradation of LAS as a function of time. The carbonources included glucose, sucrose, maltose, mannitol and succinic acid, whereashe nitrogen sources being tested included ammonium chloride, trypton, yeastxtract, ammonium nitrate and casein. All of the nutrient sources were suppliedndependently to NB at a concentration of 0.2%.
. Results and discussion
The primary biodegradation of LAS by the individual cul-ures and bacterial consortium was established by measuringhe disappearance of LAS in the cultures. Fig. 1 shows theercentage of LAS left in the broth medium over time bysing two different culture media (minimal medium and nutrientroth). The co-culture was capable of growing in both media,ut the percentage degradation of LAS obtained by the bacte-
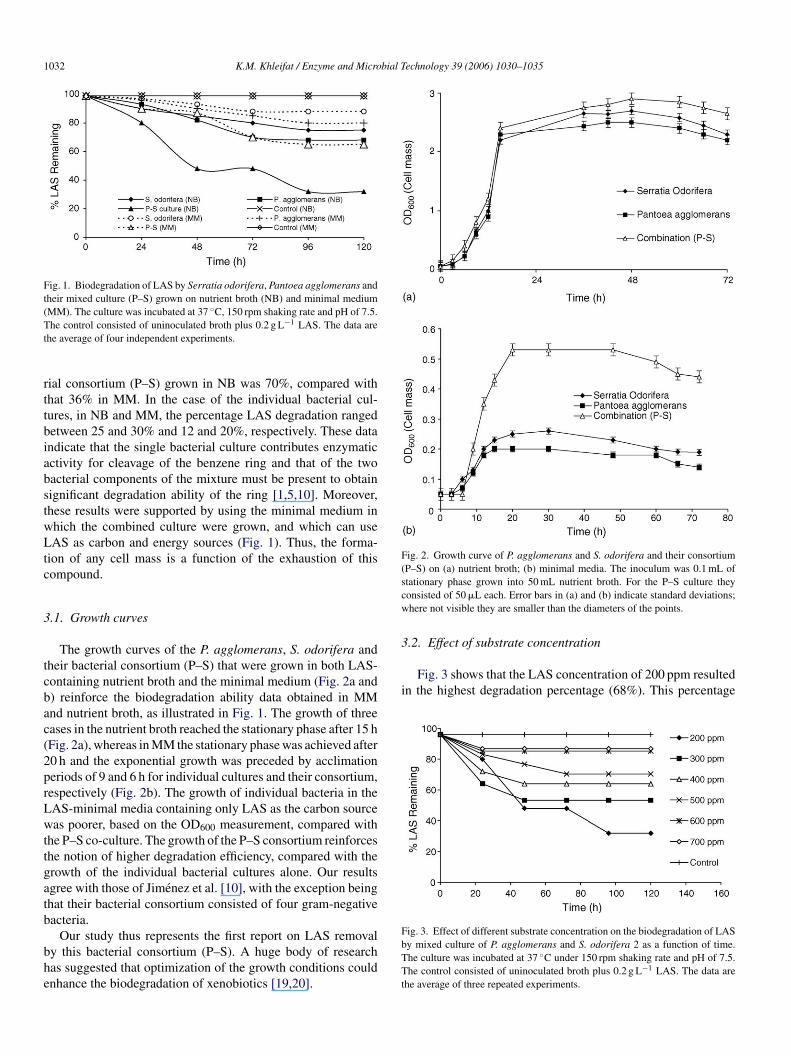
1032 K.M. Khleifat / Enzyme and Microbial Technology 39 (2006) 1030–1035
Fig. 1. Biodegradation of LAS by Serratia odorifera, Pantoea agglomerans andtheir mixed culture (P–S) grown on nutrient broth (NB) and minimal medium(MM). The culture was incubated at 37 ◦C, 150 rpm shaking rate and pH of 7.5.The control consisted of uninoculated broth plus 0.2 g L−1 LAS. The data arethe average of four independent experiments.
rial consortium (P–S) grown in NB was 70%, compared withthat 36% in MM. In the case of the individual bacterial cul-tures, in NB and MM, the percentage LAS degradation rangedbetween 25 and 30% and 12 and 20%, respectively. These dataindicate that the single bacterial culture contributes enzymaticactivity for cleavage of the benzene ring and that of the twobacterial components of the mixture must be present to obtainsignificant degradation ability of the ring [1,5,10]. Moreover,these results were supported by using the minimal medium inwhich the combined culture were grown, and which can useLAS as carbon and energy sources (Fig. 1). Thus, the forma-tion of any cell mass is a function of the exhaustion of thiscompound.
3.1. Growth curves
The growth curves of the P. agglomerans, S. odorifera andtheir bacterial consortium (P–S) that were grown in both LAS-containing nutrient broth and the minimal medium (Fig. 2a andb) reinforce the biodegradation ability data obtained in MMand nutrient broth, as illustrated in Fig. 1. The growth of threecases in the nutrient broth reached the stationary phase after 15 h(Fig. 2a), whereas in MM the stationary phase was achieved after20 h and the exponential growth was preceded by acclimationperiods of 9 and 6 h for individual cultures and their consortium,rLwttgatb
bhe
Fig. 2. Growth curve of P. agglomerans and S. odorifera and their consortium(P–S) on (a) nutrient broth; (b) minimal media. The inoculum was 0.1 mL ofstationary phase grown into 50 mL nutrient broth. For the P–S culture theyconsisted of 50 �L each. Error bars in (a) and (b) indicate standard deviations;where not visible they are smaller than the diameters of the points.
3.2. Effect of substrate concentration
Fig. 3 shows that the LAS concentration of 200 ppm resultedin the highest degradation percentage (68%). This percentage
Fig. 3. Effect of different substrate concentration on the biodegradation of LASby mixed culture of P. agglomerans and S. odorifera 2 as a function of time.The culture was incubated at 37 ◦C under 150 rpm shaking rate and pH of 7.5.The control consisted of uninoculated broth plus 0.2 g L−1 LAS. The data arethe average of three repeated experiments.
espectively (Fig. 2b). The growth of individual bacteria in theAS-minimal media containing only LAS as the carbon sourceas poorer, based on the OD600 measurement, compared with
he P–S co-culture. The growth of the P–S consortium reinforceshe notion of higher degradation efficiency, compared with therowth of the individual bacterial cultures alone. Our resultsgree with those of Jimenez et al. [10], with the exception beinghat their bacterial consortium consisted of four gram-negativeacteria.
Our study thus represents the first report on LAS removaly this bacterial consortium (P–S). A huge body of researchas suggested that optimization of the growth conditions couldnhance the biodegradation of xenobiotics [19,20].
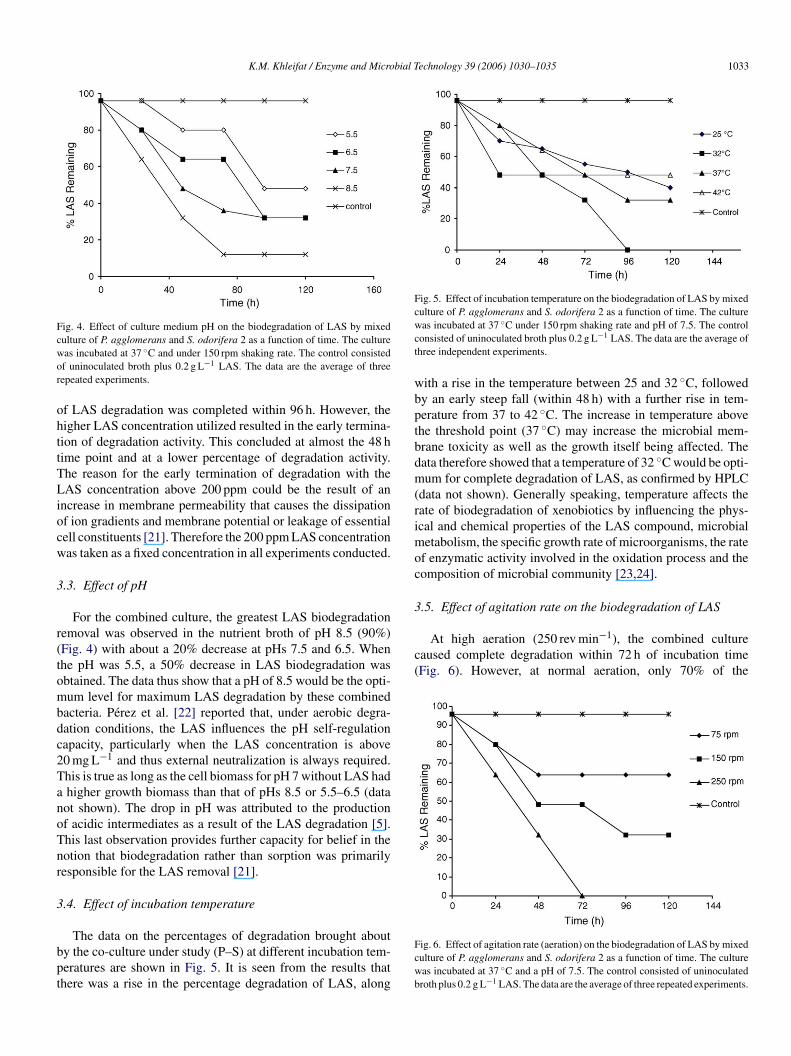
K.M. Khleifat / Enzyme and Microbial Technology 39 (2006) 1030–1035 1033
Fig. 4. Effect of culture medium pH on the biodegradation of LAS by mixedculture of P. agglomerans and S. odorifera 2 as a function of time. The culturewas incubated at 37 ◦C and under 150 rpm shaking rate. The control consistedof uninoculated broth plus 0.2 g L−1 LAS. The data are the average of threerepeated experiments.
of LAS degradation was completed within 96 h. However, thehigher LAS concentration utilized resulted in the early termina-tion of degradation activity. This concluded at almost the 48 htime point and at a lower percentage of degradation activity.The reason for the early termination of degradation with theLAS concentration above 200 ppm could be the result of anincrease in membrane permeability that causes the dissipationof ion gradients and membrane potential or leakage of essentialcell constituents [21]. Therefore the 200 ppm LAS concentrationwas taken as a fixed concentration in all experiments conducted.
3.3. Effect of pH
For the combined culture, the greatest LAS biodegradationremoval was observed in the nutrient broth of pH 8.5 (90%)(Fig. 4) with about a 20% decrease at pHs 7.5 and 6.5. Whenthe pH was 5.5, a 50% decrease in LAS biodegradation wasobtained. The data thus show that a pH of 8.5 would be the opti-mum level for maximum LAS degradation by these combinedbacteria. Perez et al. [22] reported that, under aerobic degra-dation conditions, the LAS influences the pH self-regulationcapacity, particularly when the LAS concentration is above20 mg L−1 and thus external neutralization is always required.This is true as long as the cell biomass for pH 7 without LAS hada higher growth biomass than that of pHs 8.5 or 5.5–6.5 (datanot shown). The drop in pH was attributed to the productionoTnr
3
bpt
Fig. 5. Effect of incubation temperature on the biodegradation of LAS by mixedculture of P. agglomerans and S. odorifera 2 as a function of time. The culturewas incubated at 37 ◦C under 150 rpm shaking rate and pH of 7.5. The controlconsisted of uninoculated broth plus 0.2 g L−1 LAS. The data are the average ofthree independent experiments.
with a rise in the temperature between 25 and 32 ◦C, followedby an early steep fall (within 48 h) with a further rise in tem-perature from 37 to 42 ◦C. The increase in temperature abovethe threshold point (37 ◦C) may increase the microbial mem-brane toxicity as well as the growth itself being affected. Thedata therefore showed that a temperature of 32 ◦C would be opti-mum for complete degradation of LAS, as confirmed by HPLC(data not shown). Generally speaking, temperature affects therate of biodegradation of xenobiotics by influencing the phys-ical and chemical properties of the LAS compound, microbialmetabolism, the specific growth rate of microorganisms, the rateof enzymatic activity involved in the oxidation process and thecomposition of microbial community [23,24].
3.5. Effect of agitation rate on the biodegradation of LAS
At high aeration (250 rev min−1), the combined culturecaused complete degradation within 72 h of incubation time(Fig. 6). However, at normal aeration, only 70% of the
Fcwb
f acidic intermediates as a result of the LAS degradation [5].his last observation provides further capacity for belief in theotion that biodegradation rather than sorption was primarilyesponsible for the LAS removal [21].
.4. Effect of incubation temperature
The data on the percentages of degradation brought abouty the co-culture under study (P–S) at different incubation tem-eratures are shown in Fig. 5. It is seen from the results thathere was a rise in the percentage degradation of LAS, along
ig. 6. Effect of agitation rate (aeration) on the biodegradation of LAS by mixedulture of P. agglomerans and S. odorifera 2 as a function of time. The cultureas incubated at 37 ◦C and a pH of 7.5. The control consisted of uninoculatedroth plus 0.2 g L−1 LAS. The data are the average of three repeated experiments.

1034 K.M. Khleifat / Enzyme and Microbial Technology 39 (2006) 1030–1035
Fig. 7. Effect of carbon source on the degradation of LAS by mixed culture of P.agglomerans and S. odorifera 2 as a function of time. The culture was incubatedat 37 ◦C, under 150 rpm shaking rate and pH of 7.5. The control consisted ofuninoculated broth plus 0.2 g L−1 LAS. The data are the average of three repeatedexperiments.
degradation was achieved within a 96 h interval, whereas whencells were grown under limited aeration (75 rev min−1) less than40% of LAS degradation was obtained.
It should be considered that degradation is basically an aero-bic process and hence the introduction of air to the solution willfavour it. Aerobic degradation of LAS is preferred due to thefact that the first stage of degradation, oxygenation at the end ofthe alkyl chain, requires oxygen [5,8,10]. Other studies indicatethat LAS can be anaerobically degradable only if preceded by aperiod of aerobic exposure [26].
3.6. Effect of carbon and nitrogen source
All of the carbon sources being tested caused an increase inthe LAS biodegradation rate (Fig. 7). Sucrose, maltose and glu-cose were the better carbon sources, although all carbon sourcesbeing tested produced better and more complete degradationactivity than the positive control (without C-sources). This com-plete degradation was achieved within 72 h.
All nitrogen sources being tested produced the greatest rateof LAS degradation, which was completed within 48 h (Fig. 7).The nature of the carbon and nitrogen sources may affect bothgrowth biomass and enzymes involved in LAS degradation. Theincrease in degradation of LAS by an additional supply of carbonand nitrogen sources may be due to the increasing cosubstratedLftHdoofAst
first report on a consortium that consists only of these two definedbacterial species (P–S) that mineralize LAS. Further work willfocus on the role of each member in the consortium.
4. Concluding remarks
In conclusion, evidence is presented in this work for thebiodegradation of linear alkylbenzene sulfonate (LAS) by co-culture consisted of P. agglomerans and S. odorifera 2. Sucha co-culture would seem to hold promise for the removal of ahigher concentration of LAS (200 ppm) than any other concen-tration detected by other bacteria, either singly or in combina-tion. Moreover, the optimization of growth conditions resultedin the complete biodegradation of LAS under most of the con-ditions tested separately.
Acknowledgement
This work was supported in part by Deanship of scientificresearch grant from Mutah University, Jordan.
References
[1] Sigoillot JC, Nguyen MH. Isolation and characterization of surfactantdegrading bacteria in a marine environment. FEMS Microbiol Ecol
[
[
[
[
iversity/complexity. Aside from the nutrient broth, in whichAS was completely removed (confirmed by HPLC) under dif-
erent conditions, nitrogen sources were the most efficient ofhe cosubstrates for the removal of LAS on a g L−1 basis. ThePLC peaks have validated our data pertaining to the completeegradation. We have yet to establish whether the participationf these organisms in the consortium depends upon the presencef the plasmid or not and further research will involve lookingor LAS-degrading enzymes (e.g., oxygen) and localizing them.lso, development of a treatment system will require additional
tudies on the survival of the organism and on the regulation ofhe catabolic reaction. In conclusion, this work constitutes the
1990;73:59–68.[2] Sigoillot JC, Nguyen MH. Complete oxidation of linear alkylbenzene
sulfonate by bacterial communities selected from coastal seawater. ApplEnviron Microbiol 1992;58:1308–12.
[3] Schberl P. Linear alkylbenzenesulphonate (LAS) monitoring in Germany.Tenside Surfact Det 1997;34:233–7.
[4] Schleheck D, Lechner M, Schnenberger R, Suter MJF, Cook AM.Desulfonation and degradation of sulfodiphenylethercarboxylates fromlinear alkyldiphenyletherdisulfonate surfactants. Appl Environ Microbiol2003;69:938–44.
[5] Swisher RD. Surfactant biodegradation. 2nd ed. New York, NY: MarcelDekker; 1987.
[6] Schberl P. Basic principles of LAS biodegradation. Tenside Surfact Det1989;26:86–94.
[7] White GF, Russell NJ. Biodegradation of anionic surfactants and relatedmolecules. In: Ratledge C, editor. Biochemistry of microbial degradation.Dordrecht: Kluwer; 1994. p. 143–77.
[8] Schleheck D, Tindall BJ, Rossell-Mora R, Cook AM. Parvibaculumlavamentivorans gen. nov., spec. nov., a new heterotrophic bacteriumwhich initiates catabolism of linear alkylbenzenesulfonate (LAS). Int JSyst Evol Microbiol 2004;1489–97.
[9] Rapaport RA, Eckhoff WS. Monitoring linear alkylbenzene sulfonate inthe environment: 1973–1986. Environ Toxicol Chem 1990;9:1245–57.
10] Jimenez L, Breen A, Thomas N, Federle TW, Sayler GS. Mineralizationof linear alkylbenzene sulfonate by a four-member aerobic bacterialconsortium. Appl Environ Microbiol 1991;57:1566–9.
11] Kertesz MA, Kolbener P, Stockinger H, Beil S, Cook AM. Desulfonationof linear alkylbenzene sulfonate surfactants and related compounds bybacteria. Appl Environ Microbiol 1994;60(7):2296–3303.
12] Khleifat K, Al-Shamailah W, Abboud MM. Effect of chlorination treat-ment in gram-negative bacterial composition of recycled wastewater.Mutah Lil-Buhuth wad-Dirasat (Mutah University, Jordan) [submittedfor publication].
13] Gavini F, Mergaert J, Beji A, Mielcarek C, Izard D, Kersters K,et al. Transfer of Enterobacter agglomerans (Beijerinck 1888) Ewingand Fife 1972 to Pantoea gen. nov. as Pantoea agglomerans comb.nov. and description of Pantoea dispersa sp nov. Int J Syst Bacteriol1989;39:337–45.

K.M. Khleifat / Enzyme and Microbial Technology 39 (2006) 1030–1035 1035
[14] Wright SA, Zumoff IC, Schneider HL, Beer SV. Pantoea agglomeransstrain EH318 produces two antibiotics that inhibit Erwinia amylovorain vitro. Appl Environ Microbiol 2001;67(1):284–92.
[15] Grimont PAD, Grimont F, Richard C, Davis BR, Steigerwalt AG, Bren-ner DJ. Deoxyribonucleic acid relatedness between Serratia plymuthicaand other Serratia species, with a description of Serratia odorifera sp.nov. (Type strain ICPB 3995). Int J Syst Evol Microbiol 1978;28:453–63.
[16] Miller JH. Experiment in molecular genetics. Cold Spring Harbor, NewYork: Cold Spring Harbor Laboratory; 1972.
[17] Longwell J, Maniece WD. Determination of anionic detergent in sewageeffluents and river water. Analyst 1955;80:167–71.
[18] Li Z, Rosen M. Two-phase mixed indicator titration method for deter-mination of anionic surfactants. Anal Chem J 1981;53(9):1516–9.
[19] Armenante P, Fava F, Kafkewitz D. Effect of yeast extract on growthkinetics during aerobic biodegradation of chlorobenzoic acids. Biotech-nol Bioeng 1995;47:227–33.
[20] Fava F, Armenante P, Kafkewitz D. Aerobic degradation, and dechlori-nation of 2-chlorophenol 3-chlorophenol and 4-chlorophenol by a Pseu-domonas pickettii strain. Appl Microbiol Biotechnol 1995;43:171–7.
[21] Brandt KK, Hesselsøe MP, Roslev K, Henriksen K, Sørensen J. Toxiceffects of linear alkylbenzene sulfonate on metabolic activity, growthrate, and microcolony formation of Nitrosomonas and Nitrosospirastrains. Appl Environ Microbiol 2001;67(6):2489–98.
[22] Perez M, Romero JM, Quiroga JM, Sales D. Effect of LAS (linear alkyl-benzene sulphonates) on organic matter biodegradation. Tenside SurfactDet 1996;33(6):473–8.
[23] Gibbs CF, Pugh KB, Andrews AR. Quantitative studies on marinebiodegradation of oil. II. Effect of temperature. Proc R Soc Lond Ser B1975;188:83–94.
[24] Takamatsu Y, Nishimura O, Inamori Y, Sudo R, Matsumura M. Effectof temperature on biodegradability of surfactants in aquatic microcosmsystem. Water Sci Technol 1996;34(7–8):61–8.
[25] Kallio PT, Kim DJ, Tsai PS, Bailey JE. Intracellular expression of Vit-reoscilla hemoglobin alters Escherichia coli energy metabolism underoxygen-limited conditions. Eur J Biochem 1994;15(1–2):201–8.
[26] Larson RJ, Rothgeb TM, Shimp RJ, Ward TE, Ventullo RM. Kinet-ics and practical significance of biodegradation of linear alkylbenzenesulphonate in the environment. J Am Oil Chem Soc 1993;70(7):645–57.





























