Embed Size (px)
Citation preview

Bq
TD
a
ARRAA
KSEMWFI
1
chitrhtitpiag
eim
0h
Acta Tropica 128 (2013) 606– 612
Contents lists available at ScienceDirect
Acta Tropica
jo ur n al hom epa ge: www.elsev ier .com/ locate /ac ta t ropica
ehavioral and electrophysiological responses of Culexuinquefasciatus to certain fatty acid esters
hangaraj Seenivasagan ∗, Lopamudra Guha, S. Thanvir Iqbalefence Research & Development Establishment, Jhansi Road, Gwalior, 474002 M.P., India
r t i c l e i n f o
rticle history:eceived 10 April 2013eceived in revised form 4 September 2013ccepted 7 September 2013vailable online 17 September 2013
eywords:emiochemicalslectrophysiologyosquito behavior
a b s t r a c t
Oviposition response of gravid Culex quinquefasciatus females to a series of synthetic fatty acid esterswas evaluated at 10 ppm under laboratory conditions. Octyl tridecanoate and propyl octadecanoateelicited 85% and 73% increased ovipositional responses respectively, compared to control, among the16 esters tested. Other 14 esters showed highly significant repellency (67–96%) to gravid females.Standard 3-methyl indole received 69% increased egg deposition compared to control. In the Y-tubeolfactometer, gravid C. quinquefasciatus females exhibited 78, 64% and 58% orientation respectively tooctyl tridecanoate, propyl octadecanoate and 3-methyl indole. Gravid females exhibited 19–41% reducedorientation toward treatment odors of other esters significantly different from respective control. Elec-troantennogram studies revealed 4–18-fold increased antennal response, in which 3-methyl indole, octyl
est Nile virusilariasisntegrated vector management
tridecanoate and propyl octadecanoate elicited 8-, 18- and 15-fold EAG response respectively, comparedto control. Relative EAG response of octyl tridecanoate compared to standard 3-methyl indole was sig-nificantly different. Reduced EAG responses were elicited by FAE-06, -08, -13, -14 and -15, while therelative EAG responses of other esters were at par with the standard stimulus. These, esters could beutilized potentially as oviposition attractants and repellents against C. quinquefasciatus females to reducethe breeding in polluted water along with existing integrated vector control methods.
. Introduction
The Southern house mosquito, Culex quinquefasciatus Say (Culi-idae: Diptera) is the principal vector of filariasis in India. It is aighly anthropophilic and prolific breeder, and commonly found
n both polluted waters such as stagnant drains, cesspools, sep-ic tanks, burrow pits and in clear waters such as ground pools,ock pools, streams, paddy fields, old tyres or any container thatolds stagnated water. This species dwells in the tropical parts ofhe world causing millions of cases of lymphatic filariasis and var-ous viral encephalitis (WHO, 2000). Most of the breeding sites ofhe vector lie in the adjoining areas of human habitation havingoor sewage and sanitation facilities. Oviposition is a critical stage
n the mosquito life cycle, especially for Culex mosquitoes, whichre thought to lay a single clutch of eggs in a single habitat peronotrophic cycle (Clements, 1999).
The abundance of the vector and its medical importance nec-ssarily focuses a critical feature of their life history. Selection anddentification of a suitable oviposition sites by gravid females ulti-
ately influences the survivorship of their progeny, distribution
∗ Corresponding author. Tel.: +91 751 239 0154; fax: +91 751 234 1148.E-mail address: [email protected] (T. Seenivasagan).
001-706X/$ – see front matter © 2013 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.actatropica.2013.09.009
© 2013 Elsevier B.V. All rights reserved.
of larvae and population dynamics (Edgerly et al., 1998; Spenceret al., 2002). Oviposition behavior in mosquitoes is influenced byvisual, tactile and olfactory factors, with the latter being of pri-mary importance. These cues include color and optical density ofthe water, texture and moisture, temperature and reflectance ofthe oviposition substrate (Bentley and Day, 1989). The chemicalfactors involved in oviposition site selection by mosquitoes havegained much interest in recent years (Beehler et al., 1994; Georgeet al., 1986; Knols et al., 2004; Millar et al., 1992; Seenivasagan et al.,2009; Seenivasagan and Vijayaraghavan, 2010; Takken and Knols,1999).
Pheromone and semiochemical-mediated oviposition activityin mosquitoes is a well-known aspect in mosquito behavioralecology. Intensive work has been done on the Culex spp. insearch of oviposition pheromone/kairomone eliciting differentialoviposition response. An oviposition attractant, erythro-6-acetoxy-5-hexadecanolide, identified from the egg rafts of C. quinquefasciatuswas the first pheromone identified in any species of medical impor-tance (Laurence and Pickett, 1982, 1985). Bermuda grass infusionsemitting 3-methylindole were found to attract and stimulate gravidC. quinquefasciatus for oviposition (Du and Millar, 1999; Millar
et al., 1992). Organic infusions used as baits in traps for laboratoryand field conditions elicited effective ovipositional responses fromC. quinquefasciatus and Culex nigripalpus in two-choice bioassays(Allan et al., 2005).
T. Seenivasagan et al. / Acta Tropica 128 (2013) 606– 612 607
Table 1List of fatty acid esters synthesized and evaluated for behavioral and electrophysiological responses against Culex quinquefasciatus.
Sl. no. Code no. Carboxylic acid RCOOH Alkyl halide R′Br Fatty acid ester RCOOR′
1 FAE-1 CH3CH2COOH CH3(CH2)16CH2Br Octadecyl propanoate2 FAE-2 CH3(CH2)2COOH CH3(CH2)15CH2Br Heptadecyl butanoate3 FAE-3 CH3(CH2)3COOH CH3(CH2)14CH2Br Hexadecyl pentanoate4 FAE-4 CH3(CH2)4COOH CH3(CH2)13CH2Br Pentadecyl hexanoate5 FAE-5 CH3(CH2)5COOH CH3(CH2)12CH2Br Tetradecyl heptanoate6 FAE-6 CH3(CH2)6COOH CH3(CH2)11CH2Br Tridecyl octanoate7 FAE-7 CH3(CH2)7COOH CH3(CH2)10CH2B Dodecyl nonanoate8 FAE-8 CH3(CH2)8COOH CH3(CH2)9CH2Br Undecyl decanoate9 FAE-9 CH3(CH2)9COOH CH3(CH2)8CH2Br Decyl undecanoate
10 FAE-10 CH3(CH2)10COOH CH3(CH2)7CH2Br Nonyl dodecanoate11 FAE-11 CH3(CH2)11COOH CH3(CH2)6CH2Br Octyl tridecanoate12 FAE-12 CH3(CH2)12COOH CH3(CH2)5CH2Br Heptyl tetradecanoate13 FAE-13 CH3(CH2)13COOH CH3(CH2)4CH2Br Hexyl pentadecanoate14 FAE-14 CH3(CH2)14COOH CH3(CH2)3CH2Br Pentyl hexadecanoate
baesiAaaaemte
doensoq(4aatHaui
alsoogeTfttSttr
15 FAE-15 CH3(CH2)15COOH
16 FAE-16 CH3(CH2)16COOH
George et al. (1986) showed that the following esters, viz.,utyl benzoate, methyl-3-amino benzoate, butyl-2-ethyl hex-noate, methyl-3,5-dinitro benzoate, butyl-3,5-dinitro benzoate,thyl crotonate and methoxy phenyl acetate at 15 ppm elicitedignificant oviposition attractancy to C. quinquefasciatus. Earliernvestigations focused on the role of short chain fatty acid esters onedes aegypti (Perry and Fay, 1967), lower aliphatic acids such ascetic, propionic, isobutyric, butyric, iso valeric and caproic acidsgainst C. quinquefasciatus (Hwang et al., 1980), and fatty acidsnd their derivatives against Culex and Aedes mosquitoes (Hwangt al., 1982) demonstrated oviposition repellency to the targetosquitoes. Ganesan et al. (2006) identified and reported that cer-
ain fatty acids (C12, C14 and C16) and methyl esters from conspecificggs were oviposition attractants for A. aegypti females.
Culex quinquefasciatus occur in a wide range of habitats and haveeveloped higher level of resistance to conventional control meth-ds; therefore it is essential to screen newer compounds which canffectively be used in integrated vector management. Researchersowadays are more focused on eco-friendly approaches usingemiochemicals of natural and synthetic route to control vectorsf medical importance. Electroantennogram studies on gravid C.uinquefasciatus females to the odor cues of natural breeding waterMordue et al., 1992) and synthetic odors such as 3-methyl indole,-methyl phenol, 4-ethyl phenol and indole (Blackwell et al., 1993)s well as compounds identified form Bermuda grass infusions (Dund Millar, 1999) demonstrated dose dependent antennal recep-or sensitivity and significant behavioral responses by the females.owever, electroantennogram studies on a series of long chain fattycid esters have not been reported in C. quinquefasciatus femalesntil date. The current work reported hereunder is a step forward
n this direction.These long chain fatty acid esters influenced the oviposition of A.
egypti and Aedes albopictus Skuse (Sharma et al., 2008) and Anophe-es stephensi Liston (Sharma et al., 2009). Esters are generally sweetmelling and odoriferous chemicals with a characteristic fruitydor. A C21-chain length adds stability to this group of compoundsn the water surface. It was hypothesized that, ester functionalroup placed at various positions in succession (C3–C18) wouldlicit a differential behavioral responses from C. quinquefasciatus.his study focuses on the behavioral responses of gravid C. quinque-asciatus females with the following objectives. First, to investigatehe oviposition response of gravid C. quinquefasciatus females tohese fatty acid esters treated water in a dual choice bioassay.
econdly, to determine the behavior of gravid females to orientoward or away from these ester plumes in a Y-tube olfactometero confirm the nature of these compounds as either attractants orepellents. Finally through electroantennogram experiments, weCH3(CH2)2CH2Br Butyl heptadecanoateCH3CH2CH2Br Propyl octadecanoate
investigated the peripheral antennal response of the test mosquitospecies to the fatty acid esters.
2. Materials and methods
2.1. Insects
The test mosquitoes C. quinquefasciatus used for bioassayson oviposition, orientation and electroantennogram studies wereobtained from the laboratory colony maintained in our insectary.The experiments were performed at 27 ± 2 ◦C, 75 ± 5% RH, L10:D14regime. Five-day-old mated C. quinquefasciatus were kept in sepa-rate standard-sized wooden cages (750 mm × 600 mm × 600 mm)with a sleeve opening on one side. Sucrose (10%) was provided toadults ad libitum. Female mosquitoes were fed on chicken bloodon the fifth day after emergence. All the fatty acid esters (FAE-01to FAE-16) used in the different experiments were synthesized asdescribed by Sharma et al. (2008) (Table 1).
2.2. Oviposition experiments
Dual choice oviposition bioassays were conducted according toGuha et al. (2012) with slight modifications. Ten female mosquitoeswere offered with oviposition substrate 72 h post blood meal whenthey became fully gravid. Experiments were conducted in separatecages; food grade polycarbonate plastic bowls with 10 cm diame-ter (7 cm height) filled with 500 mL of dechlorinated tap water wasused as the oviposition substrate. One milli liter of the 5000 ppmstock solution of test compounds, 3-methyl indole in hexane (tomake 10 ppm concentration) was added to the enamel bowls andthoroughly mixed. Another bowl containing 1 mL of hexane in500 mL of dechlorinated tap water served as control. Stock solu-tions of all the test chemicals were dissolved in HPLC-grade hexane(Merck) and stored in a deep freezer (−20 ◦C) after every exper-iment. The bowls were rinsed with tap water and washed withhexane before setting the experiment. The experiment was com-pletely randomized, and in each bioassay females were allowed tochoose between the treated and control substrates. The position ofthe substrate in dual choice experiments was changed in a clock-wise rotation every day. Each compound was evaluated in separatecages, and the experiment was repeated for 8 days with a differentbatch of mosquitoes. Every day, freshly treated bowls were offeredto newer batch of blood-fed gravid mosquitoes. The number of egg
rafts laid in the control and treatment sets was counted manu-ally to assess the oviposition preference of the mosquito species.The oviposition activity was expressed as oviposition activity index(OAI) using the formula: OAI = (Nt − Ns)/(Nt + Ns), where Nt is the
6 ta Tro
nr
2
admtmla2btomoto(oato
v3pmtriw
2
qotwmeIctapwsmopppshtscftai
08 T. Seenivasagan et al. / Ac
umber of egg rafts laid in test solution and Ns is the number of eggafts laid in control (Kramer and Mulla, 1979).
.3. Y-tube olfactometer experiments
Flight orientation studies on gravid C. quinquefasciatus femalest 72 h post-blood meal using a Y-tube olfactometer were con-ucted as described by Seenivasagan et al. (2010) with slightodifications, in that the mosquitoes were 8–10 days old at the
ime of exposure to experimental odors. The olfactometer wasade of Perspex glass with internal diameter of 10 and 120 cm
ength from the release chamber to treatment/control chamberst the two arms. The experiment was done between 18:00 h and0:00 h using a group of 20 gravid mosquitoes for each replicateecause maximum nocturnal oviposition activity was observed inhe first 2 h after the sunset and majority of the egg depositionccurred during the first 4 h (Reddy et al., 2007). In a dimly lit room,osquitoes were aspirated and released in the releasing chamber
f the olfactometer. The mosquitoes were given 5 min to acclima-ize in the release chamber. A constant flow of fresh air flushed thelfactometer for 5 min between replicates. The treatment solutions250 �l of 10 ppm = 2.5 �g odor stimuli) were applied onto a piecef filter paper (10 cm × 3 cm), the solvent was allowed to evaporatend subsequently placed in an odor cartridge in one of the arms ofhe olfactometer. Five replicates were conducted, and the positionsf test stimuli were alternated in the arms of the olfactometer.
Another odor cartridge holding a filter paper treated with sol-ent hexane served as control stimulus. A separate positive control-methyl indole was used at equivalent dose (2.5 �g) for com-arison. The bioassay was conducted until all the mosquitoes hadoved from the release chamber and entered the upwind end of
he olfactometer. Different set of mosquitoes were used for eacheplicate of experiment and the number of mosquitoes collectedn control, treatment arms as well as non-responding mosquitoes
ere recorded for statistical analysis.
.4. Electroantennogram experiments
Electroantennogram (EAG) studies were performed on gravid C.uinquefasciatus females based on the previous detailed methodol-gy described by Seenivasagan et al. (2012) with a modificationhat females 72 h PBM were used for EAG recording. The headas clipped off at the neck from a cold immobilized adult femaleosquito. The base of the excised head was mounted on indiffer-
nt electrode using Electrode gel (Spectra 360; Parker Laboratoriesnc., Fairfield, NJ, USA). The tip of one antenna was cut off and thenonnected to the recording electrode. A stimulus controller (Syn-ech, Hilversum, The Netherlands) delivered the charcoal filterednd humidified air @600 mL/min continuously over the antennalreparation at 2 cm distance through a 15 cm long borosil glass tubeith 0.5 cm ID. All the test stimuli (i.e. fatty acid esters) were dis-
olved in HPLC grade hexane (Merck Ltd, Mumbai, MS, India) toake 100 ppm (10−4 g) concentration. Precisely, 25 �l of 100 ppm
f test chemical (2.5 �g) was loaded onto hexane washed filteraper (5 cm × 1 cm) and after evaporation of the solvent, the filteraper was placed inside the Pasteur pipette and ca. 5 mL of air wasuffed onto antennal preparation for a duration of 0.5 s. A positivetandard [2.5 �g methyl indole (25 �l of 0.01% 3-methyl indole inexane)] was puffed at the start and end of every recording sessiono ensure the responsiveness of antenna. Complete mixing of thetimulus odor with continuous air flow was ensured by pushing testhemical odor into the air stream through a side port located 10 cm
rom the end of tube. Recording of EAG was initiated with puffinghe odor of air, hexane and the test compounds in a random ordernd ended with reverse order of the first two stimuli with 1 minnterval between subsequent stimulations. Each recording sessionpica 128 (2013) 606– 612
on a mosquito antenna constituted one replicate and seven suchnew antennal preparations were made to generate 7-replicates foreach test chemicals. For analysis, the EAG amplitude of air was sub-tracted from the EAG values (−mV) of control, positive standard3-methyl indole and test stimuli. Mean EAG amplitude relative tostandard 3-methyl indole and the solvent control was consideredfor statistical significance.
2.5. Data analysis
Egg raft counts in oviposition experiments were log(x + 0.5)transformed for each replicate and subjected to t-test. Whereas,the number of gravid females making choice between treated andcontrol odor in Y-tube olfactometer was subjected to t-test withoutany transformation. However, per cent oviposition and orientationresponse was calculated for each replicate and used in graphicalrepresentation. Calculated values of orientation and ovipositionactivity indices as well as EAG amplitude (−mVolt) data were sub-jected to one-way analysis of variance (SigmaStat V2.03, SPSS Inc,Chicago, IL). Treatment means separated by least significant differ-ence (LSD) at 95% confidence interval was considered statisticallysignificant.
3. Results
3.1. Oviposition response
Gravid C. quinquefasciatus females deposited 83%, 72% and69% increased egg rafts respectively on octyl tridecanoate (OAI+0.67), propyl octadecanoate (OAI +0.43) and 3-methyl indole(+0.41) treated bowls than control (t14 = 3.79, P < 0.01). The ovipo-sition deterrent effect by the 14-fatty acid ester treated bowlswas highly prominent at 10 ppm concentration effecting nega-tive ovipositional responses with OAI values ranging from −0.48to −0.97 from gravid Culex females. Tridecyl octanoate, dodecylnonanoate and decyl undecanoate showed highest ovipositionrepellent (OAI −0.97) effect where, only 2% of total egg raftswere laid. Similarly, pentadecyl hexanoante, undecyl decanoateand hexyl pentadecanoate received <10% egg rafts exerting greaterthan 90% repellency. Egg deposition was reduced to 80% in bowlstreated with FAE-01, -02, -03, -14 and -15 while 75% reduction inegg laying was observed in heptyl tetradecanoate treated bowls(Fig. 1). One way ANOVA apparently revealed significant differences(F16, 102 = 17.01, P < 0.001) among the treated chemicals that wereshowing attractant and repellent effect to the gravid females.
3.2. Orientation response
The odor plumes of octyl tridecanoate, propyl octadecanoateand 3-methyl indole attracted respectively 78%, 64% and 62%gravid females to the treatment chamber (t8 = 9.33, P < 0.001).All other esters at 2.5 �g odor plume showed mild repellencyto the orienting gravid females. Reduced orientation by gravidC. quinquefasciatus females to the other 14-compounds appar-ently demonstrated the repellent action of these fatty acid esters.About 81% and 75% of orienting gravid females were repelledrespectively by tridecyl octanoate and undecyl decanoate odors,while most other esters demonstrated 65–70% repellency to gravidfemales. Flight orientation response was not significant (P > 0.05)to octadecyl propanoate and hexyl pentadecanoate odors (Fig. 2).One way ANOVA on orientation indices apparently revealed sig-
nificant differences (F16, 68 = 19.98, P < 0.001) among the treatedchemicals that were showing attractant and repellent effect tothe gravid females. Attraction of gravid females to propyl octade-canoate odor was at par with 3-methyl indole; however orientation
T. Seenivasagan et al. / Acta Tropica 128 (2013) 606– 612 609
F sters.
d ased o
ra
3
oi(ratoFF-ctFi
4
rocO
ig. 1. Oviposition response of gravid Culex quinquefasciatus females to fatty acid eifferent alphabet superscripts are significantly different (n = 8, ANOVA, P < 0.001) b
esponse to octyl tridecanoate was significantly different from bothttractants.
.3. Electroantennogram response
Antenna of gravid C. quinquefasciatus females showed high levelf sensitivity to the odor of all the sixteen fatty acid esters testedn this experiment with significantly increased antennal responsesF16, 102 = 1.86, P < 0.03) compared to hexane control. The EAGesponse varied from 4-folds and 18-folds for pentyl hexadecanoatend octyl tridecanoate respectively in comparison to solvent con-rol. About 15-fold increased EAG response was observed for propylctadecanoate and octadecyl propanoate, while FAE-02, -05 andAE-10 elicited 10–11-fold increased EAG responses followed byAE-03, -07, -09 and -12 with 9-fold response. Whereas, FAE-06,08, -13 and FAE-15 showed 6–7-fold increased EAG responsesompared to control (Fig. 3). Based on relative EAG, the responseo octyl tridecanoate (FAE-11) was slightly higher, and response toAE-01, FAE-16 was at par with that of positive standard 3-methylndole, while other esters showed lower relative EAG responses.
. Discussion
This study reports the oviposition behavior, orientation
esponse and antennal sensitivity of C. quinquefasciatus to a seriesf C21fatty acid esters. We found that esters from FAE-02 to FAE-07aused 62–97% reduction in oviposition by C. quinquefasciatus.nly two fatty acid esters octyl tridecanoate (FAE-11) and propylPer cent values are mean of 8 replicates (n = 8, t-test, P < 0.001). Ester legends withn oviposition activity index.
octadecanoate (FAE-16) elicited increased oviposition responses.Anopheles stephensi deposited increased eggs in FAE-07, -08,-09, -10, -14 and FAE-16 treated bowls compared to control(Sharma et al., 2009). In contrast, A. albopictus females depositedsignificantly reduced number of eggs in bowls treated with theseesters, while A. aegypti showed reduced egg laying for all testedchemicals except in FAE-16 treated bowls (Sharma et al., 2008).Culex quinquefasciatus females laid fewer eggs onto FAE-12, FAE-13and FAE-15 treated bowls. Similar trend was observed in theresponse of A. stephensi, A. aegypti and A. albopictus to these esters.FAE-01 reduced the oviposition of C. quinquefasciatus; similarto A. stephensi and A. albopictus females. However, A. aegyptifemales partially preferred FAE-01 treated bowls for ovipositionat 10 ppm. FAE-09 and FAE-10 elicited negative oviposition fromC. quinquefasciatus, while, stimulated A. stephensi females for eggdeposition. However, A. aegypti and A. albopictus females showedlesser preference toward FAE-09 and FAE-10 in our previous stud-ies. Increased oviposition in most control substrates might havebeen mediated by the odor of pre-existing egg rafts that were laidby early visiting females which avoided treated bowls. Previouslydeposited egg rafts display the suitability of a site and further influ-ence the females for egg deposition. The blend of site derived andpheromonal oviposition cues stimulate optimum oviposition activ-ity by gravid C. quinquefasciatus females (Olagbemiro et al., 2004).
Yet only a few egg rafts laid at treated bowl in a day demonstratethe physiological condition that drives the female to deposit hereggs after perceiving the control bowl as an over crowded site. Inmost circumstances, it would be lethal to females to retain her eggs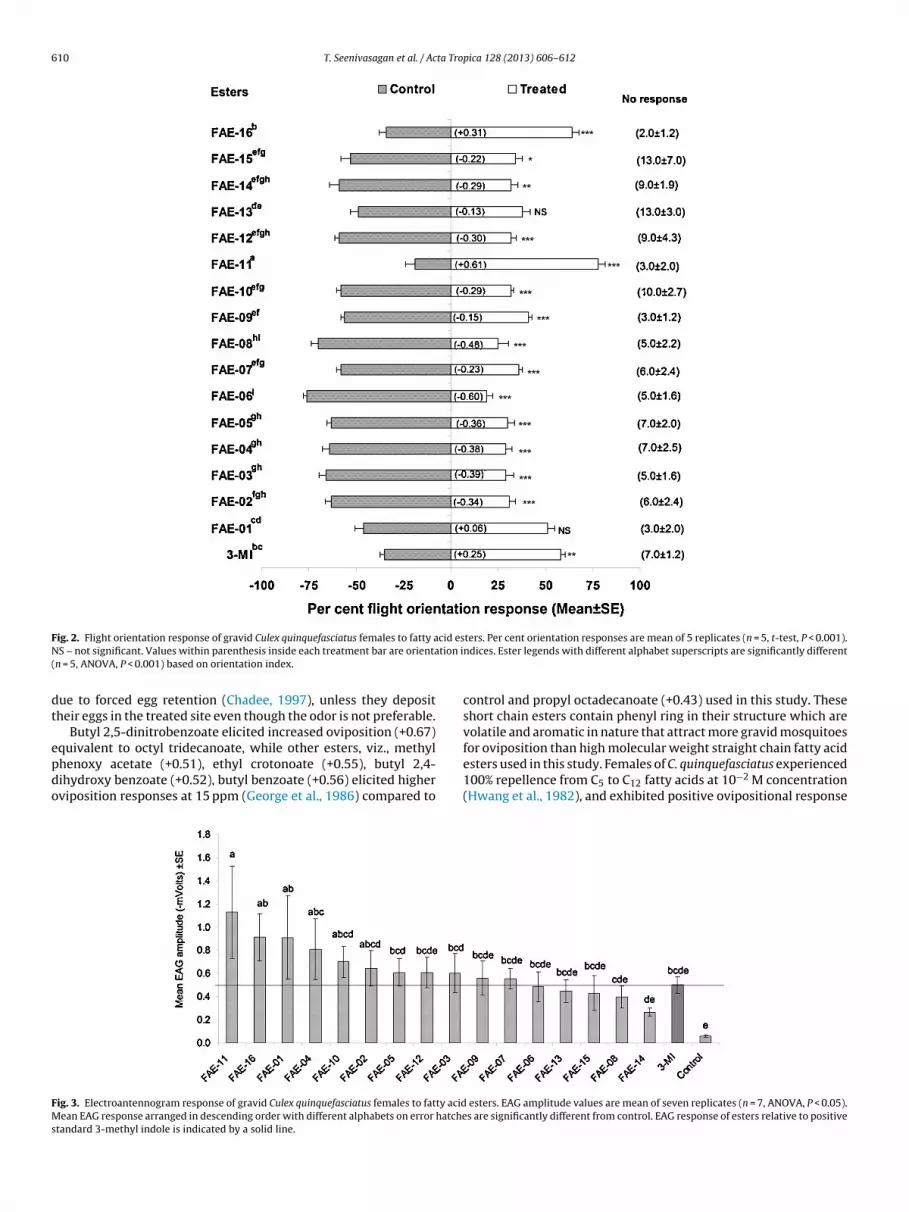
610 T. Seenivasagan et al. / Acta Tropica 128 (2013) 606– 612
F cid esN tion i(
dt
epdo
FMs
ig. 2. Flight orientation response of gravid Culex quinquefasciatus females to fatty aS – not significant. Values within parenthesis inside each treatment bar are orienta
n = 5, ANOVA, P < 0.001) based on orientation index.
ue to forced egg retention (Chadee, 1997), unless they depositheir eggs in the treated site even though the odor is not preferable.
Butyl 2,5-dinitrobenzoate elicited increased oviposition (+0.67)
quivalent to octyl tridecanoate, while other esters, viz., methylhenoxy acetate (+0.51), ethyl crotonoate (+0.55), butyl 2,4-ihydroxy benzoate (+0.52), butyl benzoate (+0.56) elicited higherviposition responses at 15 ppm (George et al., 1986) compared toig. 3. Electroantennogram response of gravid Culex quinquefasciatus females to fatty acidean EAG response arranged in descending order with different alphabets on error hatche
tandard 3-methyl indole is indicated by a solid line.
ters. Per cent orientation responses are mean of 5 replicates (n = 5, t-test, P < 0.001).ndices. Ester legends with different alphabet superscripts are significantly different
control and propyl octadecanoate (+0.43) used in this study. Theseshort chain esters contain phenyl ring in their structure which arevolatile and aromatic in nature that attract more gravid mosquitoes
for oviposition than high molecular weight straight chain fatty acidesters used in this study. Females of C. quinquefasciatus experienced100% repellence from C5 to C12 fatty acids at 10−2 M concentration(Hwang et al., 1982), and exhibited positive ovipositional responseesters. EAG amplitude values are mean of seven replicates (n = 7, ANOVA, P < 0.05).s are significantly different from control. EAG response of esters relative to positive

ta Trop
treatcamecham1aaqbtltfwmvmtei
7iatSftmTtats
estadid1otawatetdt
mrr
gambiae (Diptera: Culicidae) by semiochemicals of microbial origin. Int. J. Trop.
T. Seenivasagan et al. / Ac
o trans-octadec-11-enoic acid at concentration below 1 × 10−3 M,elative to control Hwang et al. (1984). However, in our studysters synthesized from C5 to C12 fatty acids with correspondinglkyl halides also repelled Culex females from oviposition in thereated bowls. Further, Schultz et al. (1982) found 8–10 carbonompounds such as nonanoic acid, ethyl nonanoate were morective. A chain length of 6–18 carbon atoms seems to be opti-al for eliciting oviposition responses, where hexanoic acid ethyl
ster elicited increased egg deposition by Aedes mosquitoes thanontrol (Knight and Corbet, 1991). Whereas, Ganesan et al. (2006)ave reported positive ovipositional responses for certain C12–C18cids and negative responses for methyl esters of these acids in theethanol extract of A. aegypti eggs at concentrations of 1, 10 and
00 ppm treated water. However, our present study demonstrates contrasting property of two esters derived from tridecanoiccid (C13) and octadecanoic acid (C18) that were attractants for C.uinquefasciatus females. Many active compounds possess a car-oxylic acid, carbonyl functional group and the evidence suggestshat level of activity is determined by other factors such as chainength, molecular weight, methyl branching, concentration andhe species involved. Presumably, semiochemicals with differentunctional groups that attracts mosquitoes to treated substratesould represent a rational and potential oviposition attractant withechanisms deeply embedded in mosquito ecology, thereby pro-
iding useful alternatives to currently available methods to containosquito breeding. Accurate and reliable information regarding
he breeding habitats and distribution of C. quinquefasciatus is anssential requisite for improved management strategies for thesemportant disease vectors (Pickett and Woodcock, 1996).
In this study antenna of gravid C. quinquefasciatus females, after2 h PBM exhibited increased sensitivity to fatty acid ester odors
n few orders of magnitude compared to solvent control as wells 3-methyl indole, which signifies that these esters stimulatehe peripheral olfactory receptors of C. quinquefasciatus females.imilar observations were reported by Sharma (2008) for theseatty acid esters in which sugar fed females were more sensitivehan blood fed females of A. aegypti, A. albopictus and A. stephensi
osquitoes. Previous electrophysiological studies by Davis andakahashi (1980) have also established that the neurons sensitiveo host produced cues such as lactic acid became less sensitives the blood meal in A. aegypti is digested, while neurons sensi-ive to methyl butyrate, an oviposition site attractant became moreensitive.
The compounds which elicited significant oviposition also influ-nced flight orientation of gravid females in a Y-tube olfactometer,ignifying that these compounds were perceived through the olfac-ory mechanism. However, the suitability of the treated substratessessed through contact chemoreception could have effected eggeposition. The female integrates sensory information from chem-
cal cues, together with other physio chemical (e.g., pH, alkalinity,issolved oxygen, etc.) and mechanical cues (Bentley and Day,989), to resolve whether the water container represents a suitableviposition site. In this study, Culex females hovered shortly abovehe treated bowls, before alighting on the rim of bowl. Increasedntennal sensitivity, orientation and oviposition onto the treatedater by gravid females reflects the combined effects of activation,
ttraction, arrestment and stimulation of oviposition. Although,hese fatty acid esters does not have any direct association withither breeding ecology or oviposition site cues, the ester func-ional group placed in succession of a C21straight chain elicitedistinct behavioral responses in the target mosquitoes, signifyinghe structure activity function of this group of bio-active molecules.
In conclusion, C. quinquefasciatus being the most tolerant
osquito species which breeds profusely in any type of envi-onment and needs a special attention as they have developedesistance to most conventional methods of vector control. Gradual
ica 128 (2013) 606– 612 611
increase in the adaptability of vectors demands newer strategiesto combat them. Our present study proposes the potential appli-cation of fatty acid esters as oviposition attractants or repellentsfor surveillance and control of mosquito population using ovitraps.However, the ability of esters to mask the inherent odor of pol-luted water bodies would need further investigations. Therefore,treating a suitable oviposition sites with repellents after establish-ing eco-toxicological standards to divert the gravid females towardattractant treated bowls with insecticides based on push and pullstrategy, would prove highly efficient in controlling mosquitoesalong with integrated vector management strategies.
Acknowledgments
This work receives financial grant through Project-201 (S&T)of our Establishment. The authors are thankful to Director, DRDEand Head, Vector Management Division for their interest andencouragement. TSV expresses his sincere thanks to three anony-mous reviewers for their constructive comments to improve thismanuscript. We thank A.N. Rao and K. Ganesan for synthesis andproviding the test chemicals. We thank Matadin, Senior TechnicalAssistant and all the members of VMD associated with maintenanceof the mosquito culture.
References
Allan, S.A., Bernier, U.R., Kline, D.L., 2005. Evaluation of oviposition substrates andorganic infusions on collection of Culex in Florida. J. Am. Mosq. Control Assoc.21, 268–273.
Beehler, J.W., Millar, J.G., Mulla, M.S., 1994. Field evaluation of synthetic com-pounds mediating oviposition response in Culex mosquitoes (Diptera:Culicidae).J. Chem. Ecol. 20, 281–291.
Bentley, M.D., Day, F.J., 1989. Chemical ecology and behavioural aspects of mosquitooviposition. Annu. Rev. Entomol. 34, 401–421.
Blackwell, A., Mordue, A.J., Hansson, B.S., Wadhams, L.J., Pickett, J.A., 1993. Abehavioural and electrophysiological study of oviposition cues for Culex quin-quefasciatus. Physiol. Entomol. 18, 343–348.
Chadee, D.D., 1997. Effect of forced egg-retention on the oviposition patterns offemale Aedes aegypti (Diptera: Culicidae). Bull. Entomol. Res. 87, 649–651.
Clements, A.N., 1999. The Biology of Mosquitoes: Sensory Reception & Behavior.CABI, Wallingford.
Davis, E.E., Takahashi, F.T., 1980. Humoral alteration of chemoreceptor sensitivityin the mosquito. In: Proceedings of International Symposium on Olfaction andTaste, IRL, London, pp. 139–142.
Du, Y.J., Millar, J.G., 1999. Electroantennogram and oviposition bioassay responsesof Culex quinquefasciatus and Culex tarsalis (Diptera: Culicidae) to chemicals inodors from Bermuda grass infusions. J. Med. Entomol. 36, 158–166.
Edgerly, J.S., McFarland, M., Morgan, P., Livdahl, T., 1998. A seasonal shift in egg-laying behaviour in response to cues of future competition in a treeholemosquito. J. Anim. Ecol. 67, 805–818.
Ganesan, K., Mendki, M.J., Suryanarayana, M.V.S., Prakash, S., Malhotra, R.C., 2006.Studies of Aedes aegypti (Diptera: Culicidae) ovipositional responses to newlyidentified semiochemicals from conspecific eggs. Aust. J. Entomol. 45, 75–80.
George, N., Ramiah, K.D., Sujatha, C.H., Kalyanasundaram, M., Das, P.K., 1986. Ovipo-sition attractancy of some substituted esters and the pheromone extracted fromegg rafts against Culex quinquefasciatus. Curr. Sci. 55, 1205–1207.
Guha, L., Seenivasagan, T., Bandyopadhyay, P., Iqbal, S.T., Sathe, M., Sharma, P.,Parashar, B.D., Kaushik, M.P., 2012. Oviposition and flight orientation responseof Aedes aegypti to certain aromatic aryl hydrazono esters. Parasitol. Res. 111,975–982.
Hwang, Y.S., Kramer, W.L., Mulla, M.S., 1980. Ovipositional attractants and repel-lents of mosquitoes: isolation, identification of oviposition repellents for Culexmosquitoes. J. Chem. Ecol. 6, 71–80.
Hwang, Y.S., Schultz, G.W., Axelrod, H., Kramer, W.L., Mulla, M.S., 1982. Oviposi-tional repellency of fatty acids and their derivatives against Culex and Aedesmosquitoes. Environ. Entomol. 11, 223–226.
Hwang, Y.S., Schultz, G.W., Mulla, M.S., 1984. Structure activity relationships ofunsaturated fatty acids as mosquito oviposition repellents. J. Chem. Ecol. 10,145–151.
Knight, J.C., Corbet, S.A., 1991. Compounds affecting mosquito oviposition: structureactivity relationships and concentration effects. J. Am. Mosq. Control. Assoc. 7,37–41.
Knols, B.G.J., Sumba, L.A., Guda, T.O., Deng, A.I., Hassanali, A., Beier, J.C., 2004. Medi-ation of oviposition site selection in the African malaria mosquito Anopheles
Insect. Sci. 24, 260–265.Kramer, W.L., Mulla, M.S., 1979. Oviposition attractants and repellents of mosquito
oviposition resonses of Culex mosquitoes to organic infusions. Environ. Entomol.8, 1111–1117.

6 ta Tro
L
L
M
M
O
P
P
R
S
12 T. Seenivasagan et al. / Ac
aurence, B.R., Pickett, J.A., 1982. Erythro-6-acetoxy-5-hexadecanolide, the majorcompound of a mosquito attractant pheromone. J. Chem. Soc. Chem. Commun.,59–60.
aurence, B.R., Pickett, J.A., 1985. An oviposition attractant pheromone in Culex quin-quefasciatus Say (Diptera: Culicidae). Bull. Entomol. Res. 75, 283–290.
illar, J.G., Chaney, J.D., Mulla, M.S., 1992. Identification of oviposition attractantsfor Culex quinquefasciatus from fermented Bermuda grass infusions. J. Am. Mosq.Control. Assoc. 8, 11–17.
ordue, A.J., Blackwell, A., Hansson, B.S., Wadhams, L.J., Pickett, J.A., 1992.Behavioural and electrophysiological evaluation of oviposition attractants forCulex quinquefasciatus Say (Diptera: Culicidae). Experientia 48, 1109–1111.
lagbemiro, T.O., Birkett, M.A., Mordue Luntz, A.J., Pickett, J.A., 2004. Laboratory andfield responses of the mosquito, Culex quinquefasciatus, to plant-derived Culexspp. oviposition pheromone and the oviposition cue skatole. J. Chem. Ecol. 30,965–976.
erry, A.S., Fay, R.W., 1967. Correlation of chemical constitution and physical prop-erties of fatty acid esters with oviposition response of Aedes aegypti. Mosq. News27, 175–183.
ickett, J.A., Woodcock, C.M., 1996. The role of mosquito olfaction in oviposi-tion site selection and in the avoidance of unsuitable hosts. In: Block, G.R.,Cardew, G. (Eds.), Olfaction in Mosquito–Host Interactions. Wiley, Chichister,pp. 109–123.
eddy, M.R., Lepore, T.J., Pollack, R.J., Kiszewski, A.E., Spielman, A., Reiter, P., 2007.Early evening questing and oviposition activity by the Culex (Diptera: Culicidae)
vectors of West Nile virus in northeastern North America. J. Med. Entomol. 44,211–214.chultz, G.W., Hwang, Y.S., Kramer, W.L., Axelrod, H., Mulla, M.S., 1982. Field evalu-ation of ovipositional repellents against Culex (Diptera: Culicidae) mosquitoes.Environ. Entomol. 1, 968–971.
pica 128 (2013) 606– 612
Seenivasagan, T., Sharma, K.R., Ganesan, K., Prakash, S., 2010. Electrophysiological,flight orientation and oviposition responses of three species of mosquito vectorsto hexadecyl pentanoate: residual oviposition repellent activity. J. Med. Entomol.47, 329–337.
Seenivasagan, T., Sharma, K.R., Prakash, S., 2012. Electroantennogram, flight orien-tation and oviposition responses of Anopheles stephensi and Aedes aegypti to afatty acid ester-propyl octadecanoate. Acta Trop. 124, 54–61.
Seenivasagan, T., Sharma, K.R., Sekhar, K., Ganesan, K., Prakash, S., Vijayaraghavan,R., 2009. Electroantennogram, flight orientation, and oviposition responses ofAedes aegypti to the oviposition pheromone n-heneicosane. Parasitol. Res. 104,827–833.
Seenivasagan, T., Vijayaraghavan, R., 2010. Oviposition pheromones inhaematophagous insects. Vitam. Horm. 83, 597–630.
Sharma, K.R., 2008. Studies on the Effect of Parapheromones on the OvipositionActivity in Mosquito Vectors. Jiwaji University, Gwalior, pp. 173 (Ph.D. Thesis)Zoology.
Sharma, K.R., Seenivasagan, T., Rao, A.N., Ganesan, K., Agarwal, O.P., Malhotra, R.C.,Prakash, S., 2008. Oviposition responses of Aedes aegypti and Aedes albopictus tocertain fatty acid esters. Parasitol. Res. 103, 1065–1073.
Sharma, K.R., Seenivasagan, T., Rao, A.N., Ganesan, K., Agrawal, O.P., Prakash, S., 2009.Mediation of oviposition responses in the malaria mosquito Anopheles stephensiListon by certain fatty acid esters. Parasitol. Res. 104, 281–286.
Spencer, M., Blaustein, L., Cohen, J.E., 2002. Oviposition habitat selection bymosquitoes (Culiseta longiareolata) and consequences for population size. Ecol-
ogy 83, 669–679.Takken, W., Knols, B.G., 1999. Odor-mediated behavior of Afrotropical malariamosquitoes. Annu. Rev. Entomol. 44, 131–157.
WHO, 2000. Control of Lymphatic Filariasis: A Manual for Health Personnel, 10thed. World Health Organization, Geneva, Switzerland.







![Hemisynthesis and antiproliferative properties of mono-[O-(14-benzoylaconine-8-yl)]esters and bis-[O-(14-benzoylaconine-8-yl)]esters](https://img.dokumen.tips/doc/110x75/6355005121a0f893210b4ee9/hemisynthesis-and-antiproliferative-properties-of-mono-o-14-benzoylaconine-8-ylesters.jpg)





















