Embed Size (px)
Citation preview

Basin-scale paleobiogeography and paleoecology: an example fromKarpatian (Latest Burdigalian) benthic and planktonic foraminifera
and calcareous nannofossils from the Central ParatethysSilvia Spezzaferria,*, Stjepan Corica, Johann Hoheneggera, Fred Röglb
a Institute of Paleontology, University of Vienna, Althanstrasse 14, 1090 Vienna, Austriab Natural History Museum, Burgring 7, 1014 Vienna, Austria
Received 25 October 2001; accepted 21 February 2002
Abstract
We present the paleobiogeography and ecology of benthic and planktonic foraminifera and calcareous nannoplankton during theKarpatian (Latest Burdigalian) in the Austrian part of the Central Paratethys. Results are based on quantitative data of assemblages fromthe Molasse and Styrian Basins. Our data suggest that, although adjacent, the two basins underwent different paleoceanographic evolutionduring the Karpatian. Dysoxic bottom waters characterized the Molasse Basin, together with reducing environments, which produced theformation of pyritized levels. In the Styrian Basin, intense volcanic activity enriched surface waters in nutrient and produced strong primaryproductivity. Reduced planktonic foraminiferal abundance and dominance of agglutinated benthic forms in the Styrian Basin can beattributed to paleoceanographic conditions. The planktonic-depleted assemblages bias the paleodepth estimation by 100P(P + B) in theStyrian Basin and new tools for assessing paleodepth are needed. This study shows that paleobiogeography of foraminifera and calcareousnannoplankton is strictly related to local ecological conditions in enclosed basins and demonstrates that quantitative ecological studies arethe best way to monitor the biogeographic evolution of enclosed basins. © 2002 Éditions scientifiques et médicales Elsevier SAS. All rightsreserved.
Keywords:Karpatian; Paleobiogeography; Paleoecology; Foraminifera; Calcareous nannofossils; Paratethys
1. Introduction
Continents in motion, the dispersal of the Pangea conti-nent and the northward drift of India and Australia ended theperiod of the Mesozoic Tethys Ocean. The new Paratethyanbioprovince and the Mediterranean Tethys (Proto-Mediterranean) were born. From the Oligocene through theMiocene, the Paratethys underwent a complex evolutionthat produced deep environmental changes. The alternationof seaways and land bridges in turn opened and closedefficient marine connections with the Indian Ocean on itseastern part and the Mediterranean Sea on its western part.Small basins also in turn formed and disappeared throughthe Miocene. This area is, therefore, the ideal location toanalyze basin-scale paleobiogeography and ecology.
We present here a synthesis of results based on plank-tonic and benthic foraminifera and calcareous nannofossilsduring the Karpatian age which spans 16.4–17.2 Ma andcorresponds to the Late Burdigalian (Mandic et al., thisissue, Fig. 1). The investigated sections are located in theAustrian basins within the Central Paratethys: in the Mo-lasse Basin at Laa an der Thaya (Hole BL 503) andGöllersdorf in Lower Austria, Wagna and Retznei in theStyrian Basin. This study will reveal substantial ecologicaland paleobiogeographic difference between these adjacentbasins due to different geological and paleoceanographicconditions.
2. Geological settings
The Molasse Basin developed from Switzerland toLower Austria in a long west–east belt by the subduction of
* Corresponding author.E-mail address:[email protected] (S. Spezzaferri).
Geobios (2002) Mémoire spécial n° 24
www.elsevier.com/locate/geobios
© 2002 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.PII: S 0 0 1 6 - 6 9 9 5 ( 0 2 ) 0 0 0 6 3 - 3

the European platform under the load of the alpine trust belt(Steininger et al., 1989). In eastern Austria, area north of theDanube River, the Molasse Basin turns northeast andcontinues in the Carpathian foredeep. The basement of theMolasse Basin consists of the crystalline Bohemian Massifand a sequence of Late Paleozoic and Mesozoic sediments.The Molasse sedimentation in this eastern area started in theLate Oligocene (Egerian). The main overthrust followed inthe Late Early Miocene (Eggenburgian to Ottnangian).Marine cycles in Latest Early Miocene (Karpatian) andEarly–Middle Miocene flooded the Molasse Basin north ofthe Danube (Kapounek et al., 1965).
The Styrian Basin belongs to the Central Paratethysbioprovince and in particular, is part of the IntracarpathianPannonian Basin system. During the Karpatian and Bad-enian (Langhian, Middle Miocene), it was strongly influ-enced by hydrodynamic changes in the Proto-MediterraneanSea (Fig. 1) and by the tectonic activities in the Alpine–Car-pathian belt. In the Early Miocene, between the EasternAlps and the Pannonian realm, a syn-rift subsidence formeddeep-sea basins. The “Steirischer Schlier” was deposited inthese basins and consists of calcareous mudstones andsiltstones. The tectonic activity was accompanied by inten-sive volcanism (Sachsenhofer, 1996). The transition fromthe Early to the Middle Miocene is termed “Styrian TectonicPhase” (Stille, 1924) and is recorded in all circum-Mediterranean regions. Its geological expression is a sedi-mentary gap and different distribution between Karpatianand Badenian sediments.
3. Materials and methods
Two hundred grams of sediment for each sample weresoaked in gasoline (Laa Th. and Göllersdorf Sections) orhydrogen peroxide (Wagna and Retznei Sections) for sev-eral hours. Gasoline was used to desegregate pyritized
sediments without damaging the specimens and to retain theoriginal faunal composition. Samples were then soaked inwarm water and washed under running water through > 250,250–125 and > 63 µm mesh sieves. Washed residues werethen solitted using the splitter described in Rupp (1986) toobtain approximately 500–1000 specimens per fraction foreach split. Specimens of benthic and planktonic foramin-ifera were picked from one split per fraction for eachsample, identified with a binocular microscope and counted.
Smear slides for calcareous nannofossils analysis wereprepared following the standard preparation technique ofPerch-Nielsen (1985) and studied by light microscope at1000× magnification. Approximately 350 specimens foreach sample were counted. Raw data of microfossils werethen transformed into percentages over the total abundanceand percentage abundance curves were plotted (abundancecurves of single species are available upon request). Specieswith phylogenetic affinities and similar environmental sig-nificance were also grouped to better interpret distributionpatterns. Climatic curves were constructed using the methodof Cita et al. (1977), and followed by Spezzaferri (1995).Paleodepths were estimated following Murray (1976).
4. Results
4.1. Laa an der Thaya (Laa Th.) – Hole BL 503
This section was drilled northeast of Vienna, 2 km east ofthe town Laa, just outside the Wienerberger brickyard andconsists of sediments from the Laa Formation. A lithologicalsummary of Hole BL 503, together with range charts and adetailed description of the foraminiferal fauna and nannof-lora assemblages and their ecology are reported in Rögl(1969) and Spezzaferri and Coric (2001) (Fig. 2).
Microfossil assemblages are generally abundant, wellpreserved and small-sized (125 µm) throughout. They con-sist of calcareous benthic and planktonic foraminifera, andrare siliceous forms (radiolarians and diatoms). The sedi-ments are attributed to the Middle Karpatian based on thepresence of pyritized levels containing Virgulinella pertusaand rare siliceous forms (“Virgulinella Horizons” of Va-sicek, 1951), and the typical assemblage with Uvigerinagraciliformis, Pappina primiformisand P. breviformis inthe absence of younger species such as Globigerinoidesbisphericus. Nannofossil assemblages are generally poorlypreserved with reworking of Paleogene and Cretaceousforms resulting from erosion of surrounding areas.
Nannofossil assemblages are characterized by relativelyabundant Calcidiscus leptoporus, Calcidiscus tropicus,Coccolithus pelagicus, Coccolithus miopelagicus, Cyclicar-golithus floridanus, Helicosphaera ampliaperta, Heli-cosphaera carteri, Pontosphaeraspp., Reticulofenestra ha-qii, Reticulofenestra minuta and Reticulofenestrapseudoumbilicus.
Fig. 1. Paleobiogeography of the Paratethys during the Karpatian (modifiedafter Rögl, 1999). The black arrows indicate the position of Laa Th.,Göllersdorf, Wagna and Retznei Sections. This map has been compiledusing also data from Senès (1960) and Senès and Marinescu (1974).
242 S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256

Based on the presence of H. ampliaperta and Spheno-lithus heteromorphus and the absence of Sphenolithusbelemnos, these sediments can be attributed to the H. am-pliaperta–S. heteromorphus Interval Zone (MNN4a) of For-naciari et al. (1996), and to the H. ampliaperta Zone (NN4)of Martini (1971).
4.2. Göllersdorf Section
This section outcrops within the Wienerberger brickyard,800 m northeast of Göllersdorf. These sediments also be-long to the Laa Formation (Fig. 2) and consist of dark-graymarly silt with thin intercalations of fine sand and rarelymiddle sand (Roetzel et al., 1999).
Foraminiferal assemblages are generally well preserved.Large-sized (> 250 µm) benthic foraminiferal assemblagesare scarce and consist of discontinuously distributed Prae-globobulimina pyrula-P. pupoides gr., Lenticulina spp., to-gether with abundant smaller-sized (125 µm) Buliminaelongata, Nonion commune, Cibicides lopjanicus, Pulleniabulloides, Melonis pompilioides, Oridorsalis umbonatus,Valvulineria complanata, Caucasina spp., Siphonodosariaspp. Planktonic foraminiferal assemblages vary from large-to small-size (> 250 to ≈ 63 µm) and consist of dominantGlobigerinella obesa, Globigerina ottnangiensis, andG. praebulloides. The accompanying assemblages includeGlobigerina bollii lentiana, G. concinna, Zeaglobigerinawoodi, Tenuitellinata spp., and Globigerinoides spp. In
Fig. 2. Simplified lithology of Wagna and Retznei Sections (Styrian Basin), Laa an der Thaya and Göllersdorf (Molasse Basin).
S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256 243

particular, G. concinna peaks in samples Göll. 2 and 4, andis associated to Spiroloculina compressiuscula, V. pertusa,and high abundance of the P. pyrula-pupoides group incorrespondence of the “Virgulinella Horizons” . Based onthe presence of these horizons, the sediments from Göllers-dorf can be correlated with those from Laa Th. andattributed to the Middle Karpatian. Nannofossil data are notcurrently available for this section. Distributions of plank-tonic and benthic foraminifera are reported in Appendix A.
4.3. Wagna Section
This section of the old brickyard Wagna, outcrops ap-proximately 20 km south of Graz, close to the town ofLeibnitz. Sediments consist of 2–5-m-thick levels of dark-gray silty marls with centimeter-thick intercalations ofsiltstones (Schlier). A 5-m-thick thin-layered level of graymarl is present at the bottom of the section. A crystallinepebble layer is present within a sandy-silt level at about14 m from the top and represents a channel deposit. Abovethis layer, sediments become more silty and sandy. Thesandy silt with silty pebbles (reworked “Schlier” ) andmolluscs at about 4 m from the top is interpreted to berelated to the Badenian transgression (Fig. 2).
Foraminiferal assemblages are moderately to well pre-served and consist of dominant large-sized (> 250 µm)agglutinated benthic forms such as Gaudryinopsis berego-viensis, Textularia laevigata and Cribrostomoides spp. to-gether with rarer Budashevaella wilsonii, B. laevigata, Al-veolophragmium obliquicameratum, Haplophragmoidescarinatus, H. laminatus and Reticulophragmium karpati-cum, R. venezuelanum, Hyperammina granulosa, Bathysi-phon filiformis, and Reophax scorpiurus. These species aregenerally associated with large-sized P. pyrula-pupoidesgr., P. bulloides, V. complanata, Heterolepa dutemplei,Chilostomella ovoidea, Allomorphina trigona, and Lenticu-lina spp. Other smaller-sized (125 µm) calcareous compo-nents consist of N. commune, B. elongata, U. graciliformis,Amphimorphina haueriana, Caucasina subulata, and boliv-inids. The planktonic fauna is rare and dominated byG. ottnangiensis, with rarer Globigerina tarchanensis, Tur-borotalita quinqueloba, and very rare G. obesa and sili-ceous components. The sediments are attributed to theMiddle Karpatian based on the presence of U. graciliformis,P. primiformis, the typical small-sized planktonic foramin-iferal assemblage in the absence of younger species such asG. bisphericus.
Sample Wag–26 contains abundant Ammonia spp. It islocated above the Early Badenian transgression horizon.Although it does not contain any marker species, it isattributed to the Early Badenian and, therefore, is notrelevant to this study.
Nannofossil assemblages are very abundant and wellpreserved and dominated by C. pelagicus, R. minuta andS. heteromorphus. These forms are generally associatedwith rare and irregularly present H. ampliaperta, Reticu-
lofenestra gelida, R. haqii, R. pseudoumbilicus, Spheno-lithus moriformis, Thoracosphaera spp. and Triquetrorhab-dulus milowii. The Early Badenian sandy layers from thetop of the section (sample Wag–26) contain more abundantH. ampliaperta than the gray marly sediments below.
The sediments from the Wagna Section are attributed tothe H. ampliaperta Zone (NN4) of Martini (1971), based onthe presence of S. heteromorphus and H. ampliaperta.
Distribution of benthic foraminifera is included in Spez-zaferri et al. (in press). Distributions of planktonic foramin-ifera and calcareous nannofossils are reported in AppendixB.
4.4. Retznei Section
This section of the Lafarge Perlmooser cement quarryoutcrops approximately 20 km south of Graz, close to thetown of Leibnitz and about 2 km from the Wagna Section.Sediments consist of dark-gray silty marls with sparsepebbles from the crystalline basement. A distinct pebblelayer is observed at the discordance with the overlyingBadenian limestone (Fig. 2).
Foraminiferal assemblages are generally well preservedand are dominated by large-sized agglutinated forms. Cal-careous organisms are small-sized (125 µm) and planktonicforms relatively scarce. Siliceous organisms are very rare.
From the bottom of the studied section up to 1.7 m, theagglutinated benthic foraminiferal assemblages containdominant large-sized and pyritized Spirorutilus carinatusassociated with P. pyrula-pupoides gr., V. complanata,H. dutemplei, P. bulloides, M. pompilioides, and Lenticu-lina spp. Rarer are Lagena spp., Siphonodosaria spp., anduvigerinids. Smaller calcareous benthic foraminifera in-clude moderately abundant A. trigona, N. commune,V. complanata, P. primiformis, C. subulata, and bolivinids.A remarkable shift in the agglutinated benthic foraminiferalassemblages occurs from 1.7 m up to the top of the section.This interval is characterized by dominant G. beregoviensis,R. karpaticum, and Cribrostomoides spp. Planktonic fora-miniferal assemblages are rare and dominated by G. ott-nangiensis with common T. quinqueloba, G. praebulloides,G. bollii lentiana, and rarer G. tarchanensis, G. obesa, andZ. woodi. Since no marker species are present, age attribu-tion based on foraminifera is difficult. The presence ofU. graciliformis and P. primiformis indicates a Karpatianage. The agglutinated benthic assemblages in the uppermostpart (from 0.3 to 1.7 m) resemble those from the WagnaSection and may be attributed to the Middle Karpatian. Nomore precise age can be attributed to the interval from 1.7 mdown to the bottom of the section.
Nannofossil assemblages from pelagic sediments inter-calated within the limestone are well preserved and domi-nated by R. minuta and Dictyococcites productus. Rarer areC. pelagicus, H. carteri, R. haqii, S. heteromorhus and Um-bilicosphaera jafari. The presence of S. heteromorhus andthe absence of the H. ampliaperta allow attribution of these
244 S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256

sediments to the S. heteromorphus Zone (NN5) of Martini(1971).
The sediments from 2 m to the bottom of the sectioncontain dominant C. pelagicus, H. carteri and R. minuta.They are associated with C. floridanus, H. ampliaperta,Pontosphaera multipora, R. haqii, R. pseudoumbilicus,Sphenolithus heteromorhus and Sph. moriformis. Nannofos-sils from this sediment are attributed to the H. ampliapertaZone (NN4) of Martini (1971), based on the continuouspresence of H. ampliaperta and Sph. heteromorphus.
Distributions of benthic, planktonic foraminifera andcalcareous nannofossils are reported in Appendix C.
4.5. Ecology
Ecological preference of benthic and planktonic foramin-ifera is retained according to the literature (e.g., Nishi, 1990;Loubere, 1991; Kaiho, 1994; Hermelin and Shimmield,1995; Gupta, 1997; Bernhard et al., 1998; Basso andSpezzaferri, 2000; Schönfeld, 2001; Spezzaferri and Coric,2001). Relevant to this study are abundances of agglutinatedbenthic and planktonic foraminifera, paleoclimatic indica-tors (warm- and cold-water species) (Table 1, Fig. 3),organic carbon-preferring, low-oxygen tolerant species andhigh primary productivity indicators (Fig. 4).
Climatic curves are based on warm- and cold-waterindicators. The climatic curve from Laa Th. (Fig. 3A) showsa positive peak between 13 and 15.5 m and one at 20.9 m.The remaining part of the curve is negative, and displaysvalues between – 80% and – 100% between 11 and 5.5 m.The climatic curve obtained for Göllersdorf is positivebetween 11 and 12 m with values around 2%, negative up to7 m and becomes strongly positive with a maximum ofabout 80% at 4 m (Fig. 3A). The climatic curve obtained forWagna is negative throughout and shows decreasing valuesfrom bottom to top (Fig. 3B). At Retznei, the climatic curve(Fig. 3B) shows positive values at the bottom and at 1.5 m.More negative values occur at 2 and 2.2 m from the top.Agglutinated benthic foraminifera are very scarce in theMolasse Basin (around 5% of the total benthic fauna) andabundant in the Styrian Basin, where they exceed 40% with
maximum of around 80% at Retznei (Fig. 4A). Planktonicforaminifera are abundant in the Molasse Basin (generallyover 50% of the total fauna) with peaks > 80% at Göllers-dorf. In the Styrian Basin, these forms are rarely more than20% (Fig. 4B). At Laa Th., organic carbon-preferringspecies are relatively abundant from the bottom up to17.6 m with a peak around 25% in correspondence of thepyritized level at 20.9 m, then they decrease to 5% from17.6 m up to the top. At Göllersdorf, their abundance variesbetween 10% and 30% at 11 and 7 m, respectively, to 0–2%in the upper part of the section. At Wagna, organic carbon-preferring species fluctuate in abundance from 5% to 20%and decrease from bottom to top. At Retznei, they areconstantly less than 5% (Fig. 4C). Low-oxygen tolerantspecies generally exceed 50% at Laa Th. and peaks atGöllersdof with two maxima between 70% and 80% at 1and 9 m. Abundances of these organisms fluctuate from20% to 40% at Wagna and from 20% to 30% at Retznei. Asingle peak around 50% occurs at 2.75 m (Fig. 4D). High
Table 1Summary of the ecological preferences of the main groups mentioned in the text
Warm-water planktonicindicators
Cool-water planktonicindicators
Organic carbon-preferringbenthic species
Low-oxygenbenthic species
High primary productivitybenthic species
G. obesa G. praebulloides B. elongata C. ovoidea M. pompilioidesG. siphonifera G. bulloides Bulimina spp. C. oolina Praeglobulimina spp.G. diplostoma G. bollii lentiana C. subulata A. trigona Uvigerina spp.P. acrostoma G. ottnangiensis C. schschkynskayae P. pupoidesP. mayeri G. dubia Caucasina spp. P. pyrulaP. inaequiconica T. quinqueloba S. compressiusculaZ. woodi G. glutinata V. complanataG. trilobus G. juvenilis V. pertusaG. subsacculifer Tenuitellinata spp. F. acutaC. globulosa Uvigerina spp.
Bolivina spp.Pappina spp.
Fig. 3. Climatic curves for Laa an der Thaya, Göllersdorf, Wagna andRetznei Sections. The curves are obtained by algebraic sum of abundance(%) of warm-water indicators (considered as positive) and abundance (%)cool-water indicators (considered as negative) following Cita et al. (1977)and Spezzaferri (1995).
S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256 245
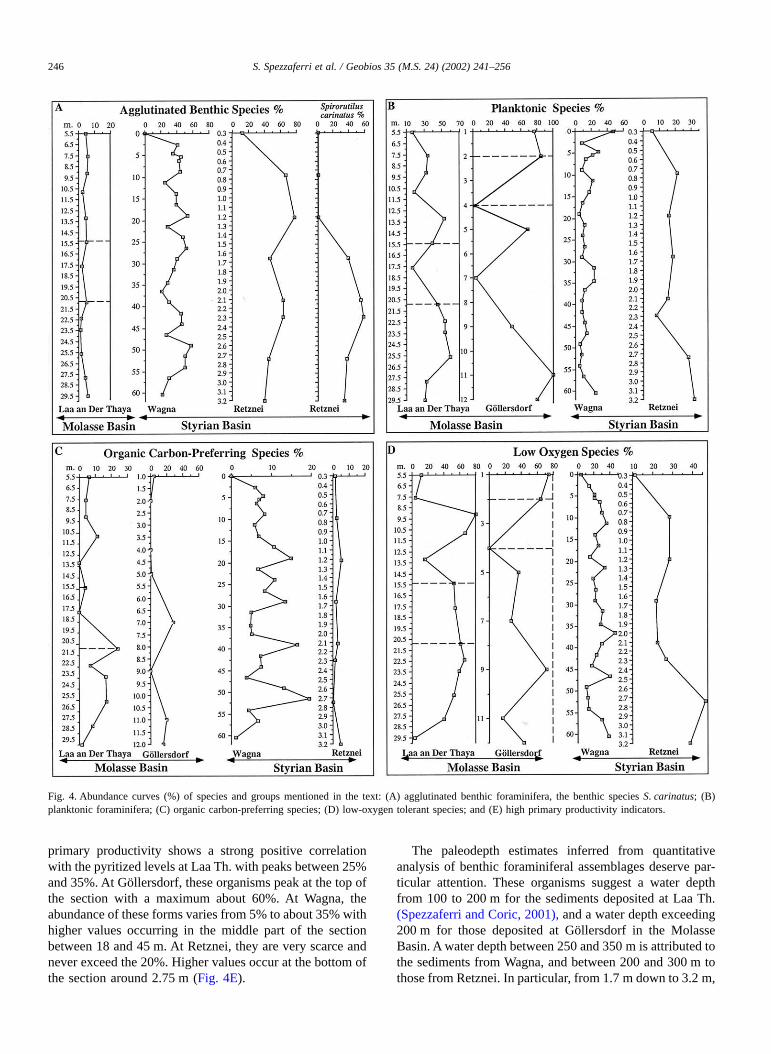
primary productivity shows a strong positive correlationwith the pyritized levels at Laa Th. with peaks between 25%and 35%. At Göllersdorf, these organisms peak at the top ofthe section with a maximum about 60%. At Wagna, theabundance of these forms varies from 5% to about 35% withhigher values occurring in the middle part of the sectionbetween 18 and 45 m. At Retznei, they are very scarce andnever exceed the 20%. Higher values occur at the bottom ofthe section around 2.75 m (Fig. 4E).
The paleodepth estimates inferred from quantitativeanalysis of benthic foraminiferal assemblages deserve par-ticular attention. These organisms suggest a water depthfrom 100 to 200 m for the sediments deposited at Laa Th.(Spezzaferri and Coric, 2001), and a water depth exceeding200 m for those deposited at Göllersdorf in the MolasseBasin. A water depth between 250 and 350 m is attributed tothe sediments from Wagna, and between 200 and 300 m tothose from Retznei. In particular, from 1.7 m down to 3.2 m,
Fig. 4. Abundance curves (%) of species and groups mentioned in the text: (A) agglutinated benthic foraminifera, the benthic species S. carinatus; (B)planktonic foraminifera; (C) organic carbon-preferring species; (D) low-oxygen tolerant species; and (E) high primary productivity indicators.
246 S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256

the abundance of S. carinatus may indicate slope environ-ment at Retznei.
5. Discussion
The Austrian Molasse and the Styrian Basins are twosmall adjacent basins connected to a larger enclosed seashowing a restricted but efficient southwestern connectionwith the Proto-Mediterranean Sea (Fig. 1). Quantitativestudies of microfossils are carried out on Karpatian sediments from these basins to establish proxies’ relationshipwith selected paleoenvironmental factors. In modern envi-ronments, for each species, different factors may be limitingdistribution both temporally and spatially. To be consideredas environmental proxies, species and assemblages mustshow a strong correlation with environmental parameters. Inparticular, distribution patterns of foraminifera are con-trolled by those environmental factors that may havereached their critical threshold (Murray, 2001).
5.1. The water column
Climatic curves suggest a cool-temperate paleoclimate inthe Molasse Basin during the Early–Middle Karpatian (Fig.3A). Warmer episodes are recorded in correspondence ofthe two pyritized levels at Laa Th. and just above them atGöllersdorf. Cool conditions prevailed upward as recordedby planktonic organisms from Laa Th. (Spezzaferri andCoric, 2001). The climatic curve of Retznei shows twowarming episodes that may be correlated with those ob-served at Laa Th. and Göllersdorf (Fig. 3B). However, theclimatic curve obtained for the Wagna Section (Fig. 3B)indicates that cool conditions characterized the Styrian
Basin, which presently is located only a few hundredkilometers from the Molasse Basin. Such a difference inpaleoclimatic condition cannot be reasonably explainedwith different geographical location only, and an alternativeexplanation is needed.
Interpretation of data suggests that a large portion of theplanktonic assemblages is missing and the climatic recordsat Wagna and partially at Retznei are corrupted. Thisassumption is also supported by the paleodepth estimatesobtained by the Plankton/Benthos Ratio (Murray, 1976).This ratio is based on the percentages of planktonic andbenthic foraminifera. Planktonic forms generally correlatewith depth and distance from the coast and thePlankton/Benthos Ratio should show a general decreasefrom ocean to shelf due to the decreased amounts ofplankton (Murray, 1976). The Plankton/Benthos Ratio esti-mates are consistent with those inferred from benthicforaminiferal assemblages from Laa Th and Göllersdorf, butcontrast with those inferred for Wagna and Retznei Sec-tions. The middle to outer shelf and the outer shelf envi-ronments indicated by the 100P/(P + B) for Laa Th. andGöllersdorf agree with the depth estimates of 100–200 and> 200 m, respectively, inferred from the benthic foramin-ifera (Fig. 5A). The curve obtained for Wagna (Fig. 5B)suggests that these sediments were deposited in a waterdepth corresponding to the inner shelf (down to 50 m asmaximum). On the contrary, benthic foraminiferal assem-blages, which include relatively deep-water forms likeH. carinatus, Budashevaella spp., Karrerulina spp.,G. beregoviensis and Bathysiphon spp. (Fig. 6) indicate apaleodepth exceeding 200 m. A similar situation applies toRetznei (Fig. 5B), where benthic assemblages suggest thatthe shift in the arenaceous components from S. carinatus toG. beregoviensis at Retznei indicates the passage from slopeto deeper-water environment. All these lines of evidenceconfirm depletion in planktonic foraminiferal assemblages,which bias the 100P/(P + B) values.
Fig. 4 (suite).
Fig. 5. Plankton/Benthos Ratio, 100P/(P + B) obtained for the studiedsections following Murray (1976). P = planktonic foraminifera;B = benthic foraminifera.
S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256 247
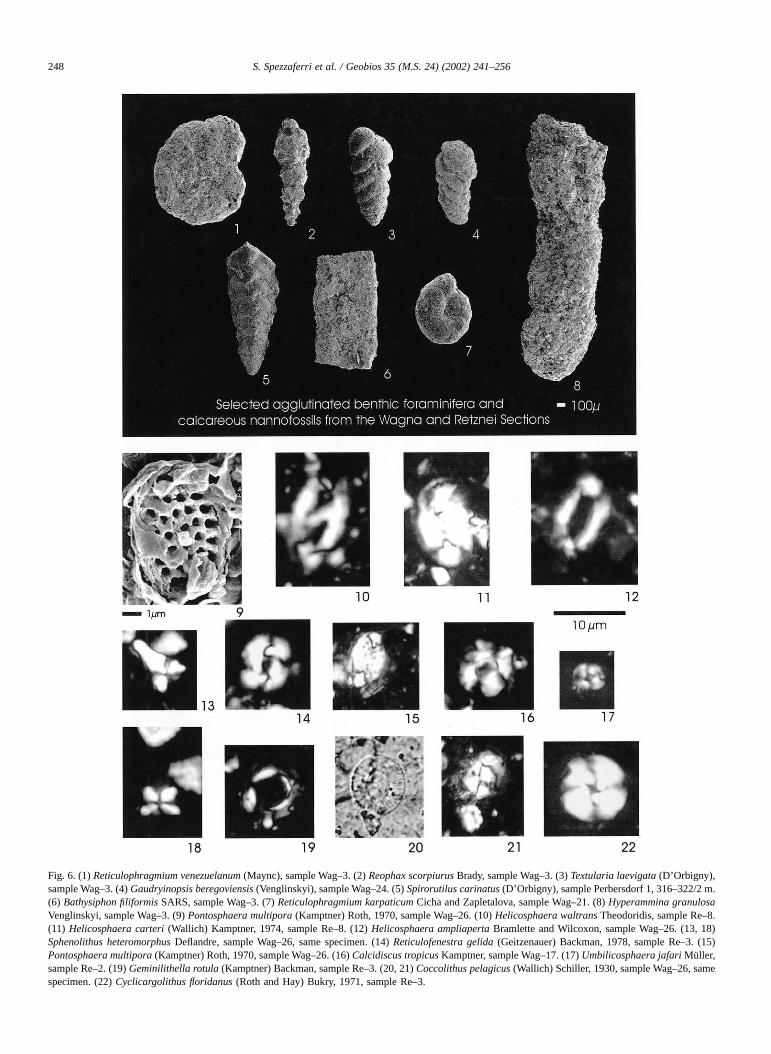
Fig. 6. (1) Reticulophragmium venezuelanum (Maync), sample Wag–3. (2) Reophax scorpiurus Brady, sample Wag–3. (3) Textularia laevigata (D’Orbigny),sample Wag–3. (4) Gaudryinopsis beregoviensis (Venglinskyi), sample Wag–24. (5) Spirorutilus carinatus (D’Orbigny), sample Perbersdorf 1, 316–322/2 m.(6) Bathysiphon filiformis SARS, sample Wag–3. (7) Reticulophragmium karpaticum Cicha and Zapletalova, sample Wag–21. (8) Hyperammina granulosaVenglinskyi, sample Wag–3. (9) Pontosphaera multipora (Kamptner) Roth, 1970, sample Wag–26. (10) Helicosphaera waltrans Theodoridis, sample Re–8.(11) Helicosphaera carteri (Wallich) Kamptner, 1974, sample Re–8. (12) Helicosphaera ampliaperta Bramlette and Wilcoxon, sample Wag–26. (13, 18)Sphenolithus heteromorphus Deflandre, sample Wag–26, same specimen. (14) Reticulofenestra gelida (Geitzenauer) Backman, 1978, sample Re–3. (15)Pontosphaera multipora (Kamptner) Roth, 1970, sample Wag–26. (16) Calcidiscus tropicus Kamptner, sample Wag–17. (17) Umbilicosphaera jafari Müller,sample Re–2. (19) Geminilithella rotula (Kamptner) Backman, sample Re–3. (20, 21) Coccolithus pelagicus (Wallich) Schiller, 1930, sample Wag–26, samespecimen. (22) Cyclicargolithus floridanus (Roth and Hay) Bukry, 1971, sample Re–3.
248 S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256

What produced the depletion in planktonic foraminiferalassemblages in the Styrian Basin? Drastic and extremedecrease in salinity may produce low diversified and dwarfplanktonic assemblages resembling cool-water assem-blages. This situation, e.g., is recorded in the MediterraneanLago-Mare facies deposited during the Messinian (LateMiocene), when connections with the Atlantic Ocean wereclosed and water exchange was confined to temporaryincursion of Atlantic water into a fully Lago-Mare (brack-ish) environment (e.g., Spezzaferri et al., 1998). The Aus-trian part of the Central Paratethys during the Karpatian wasefficiently connected to the Proto-Mediterranean (Rögl,1999, Fig. 1) and water exchange was sufficient enough toprevent a reduction in salinity, which could affect planktonicforaminiferal assemblages. In addition, such a drastic de-creased salinity in the Central Paratethys would not explainthe abundance of planktonic foraminifera in the sedimentsform Laa and Göllersdorf.
If we consider the high abundance of agglutinatedbenthic foraminifera with respect to the calcareous forms,we might assume that cool-, aggressive-water undersaturated in calcium carbonate characterized this basin duringthe Karpatian and prevented the optimal life of calcareousforms or dissolved the shells sinking at the sea floor.Modern environments having bottom waters corrosive withrespect to the CaCO3 include some shelf seas (Alexander-sson, 1978), parts of epicontinental seas and those parts ofthe oceans deeper than the carbonate compensation depth(Murray, 1991). However, the very good preservation andabundance of nannofossil assemblages argue against thedissolution hypothesis (Fig. 6). Dissolution of calcareousnannofossils can be sometimes retarded by the fact thatmost coccoliths drop to the sea floor within the fecal pelletsof planktonic grazers. However, if coccoliths may avoiddissolution within the water column, once the fecal pelletsreach the sea floor, they disintegrate rapidly due to bacterialaction and the dissolution process is activated (Haq, 1978).Therefore, we can also exclude dissolution as a factorresponsible for depleted planktonic foraminiferal assem-blages.
The only difference between the Molasse and StyrianBasin during the Karpatian is the presence of intensevolcanic activity due to the “Styrian Tectonic Phase” .Geological evidence of a large volcanic complex, activeduring the Karpatian time, is found adjacent to the Wagnaand Retznei Sections (Sachsenhofer, 1996). Active volca-noes are responsible for some of the world’s most fertilesoils. Tephra commonly contains potassium and phospho-rus, two nutrients essential to plant growth (Fyfe et al.,1982). In addition, volcanic minerals can supply the ironconcentrations needed to trigger intense primary productiv-ity in the sea (Martin and Fitzwater, 1988; Martin et al.,1994). It is well known that high nutrient concentration atthe sea-surface produces high primary productivity, but verysmall and poorly diversified planktonic foraminiferal as-
semblages. Moreover, when the nutrient concentration in-creases, foraminiferal abundance decreases (e.g., Hemlebenet al., 1989). Nannofossil assemblages seem to react differ-ently to anomalous injection of volcanic-derived nutrients(Fig. 6). In the Massignano Section (Italy), the fertilityindices sharply peak in correspondence of biotite-rich vol-canic horizons (Monechi et al., 2000; Gardin pers. comm.).We speculate that the reduced planktonic foraminiferalassemblages in the Styrian Basin reflect intense volcanic-derived high nutrient concentration in surface waters, asalso supported by the presence of siliceous organisms whichgenerally proliferate in upwelling nutrient-rich areas (e.g.,Spezzaferri, 1995).
5.2. The sea floor
The assumptions of high fertility surface waters in theStyrian Basin are also supported by the generally highabundance of high primary productivity benthic foramin-iferal indicators at Wagna. Their abundance also positivelycorrelates with that of organic carbon-preferring species(Fig. 4C). At Laa Th., high primary productivity benthicforaminiferal species indicate increased primary productiv-ity in correspondence of the pyritized levels where also thesiliceous components are present (Fig. 4E).
Bottom-water oxygenation is one of the most difficultpaleoenvironmental parameters to trace. Benthic foramin-ifera are potential proxies for oxygen content for the lowerlimit only, but once oxygen levels arise, there is no longer arelationship between oxygen levels and abundance (Murray,2001). Abundances of low-oxygen indicators suggest thatbottom waters were generally more strongly suboxic todysoxic in the Molasse Basin than in the Styrian Basin.Occurrence of pyritized levels at Laa Th. and Göllersdorfindicates that chemically reducing environment and produc-tion of ferrous sulfide may be related to dysoxic conditions(Sliter, 1975). Suboxic conditions characterized also benthicassemblages from the Styrian Basins. Sliter (1975) sug-gested that calcareous and agglutinated species may displaydifferent adaptive responses to low-oxygen conditions.Metabolic rates and reproductive processes appear to beinhibited in some calcareous species, whereas the oppositetrend applies to agglutinated species. He also stated that thiscould indicate a reduced ability of foraminifera to secretecalcite tests in low-oxygen environments. We propose herea similar situation to explain high abundances of aggluti-nated benthic foraminifera in the Styrian Basin. We alsointerpret S. carinatus (Fig. 4A) as species adapted to reduc-ing microenvironments based on its positive correlationwith low-oxygen indicators, and pyritized tests as observedat Retznei (Fig. 4D).
However, the model proposed for the Styrian basin doesnot apply to the Molasse Basin where calcareous speciesseem to have survived the severe oxygen deficiency byreducing size.
S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256 249

6. Summary and conclusions
The Early to Middle Karpatian was characterized bytemperate to cool paleoclimate interrupted by short warm-ing episodes as indicated by the climatic curve from theMolasse Basin. The Molasse and Styrian Basins, althoughadjacent, underwent different paleoceanographic evolutionduring the Karpatian. The Molasse Basin was characterizedby dysoxic bottom-water conditions and reducing microen-vironments which allowed formation of ferrous sulfide-richlevels. Calcareous benthic assemblages consist of low-oxygen adapted species, the remaining benthic assemblagesrespond to oxygen deficient environment in reducing size.High primary production and high surface water fertility
due to the intense volcanic activity characterized the StyrianBasin. High nutrient concentration is also responsible ofplanktonic depleted foraminiferal assemblages. Suboxicbottom-water conditions allowed proliferation of aggluti-nated with respect to calcareous benthic forms. Paleodepthestimated by 100P/(P + B) can be biased by depletion inplanktonic forms due to ecological conditions in enclosedbasins, and therefore, new paleoceanographic proxies andtools for paleodepth estimates are needed.
In conclusion, this study shows that paleobiogeographyof foraminifera and calcareous nannoplankton from smalladjacent basins is strictly related to local ecological condi-tions and demonstrates that quantitative ecological studiesare the best way to monitor the biogeographic evolution ofenclosed basins.
250 S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256

Appendix A. Distribution of planktonic and benthic foraminifera in the Göllerdorf Section. R = rare; C = common; X = presence. A = benthic foraminifera;B = planktonic foraminifera
S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256 251
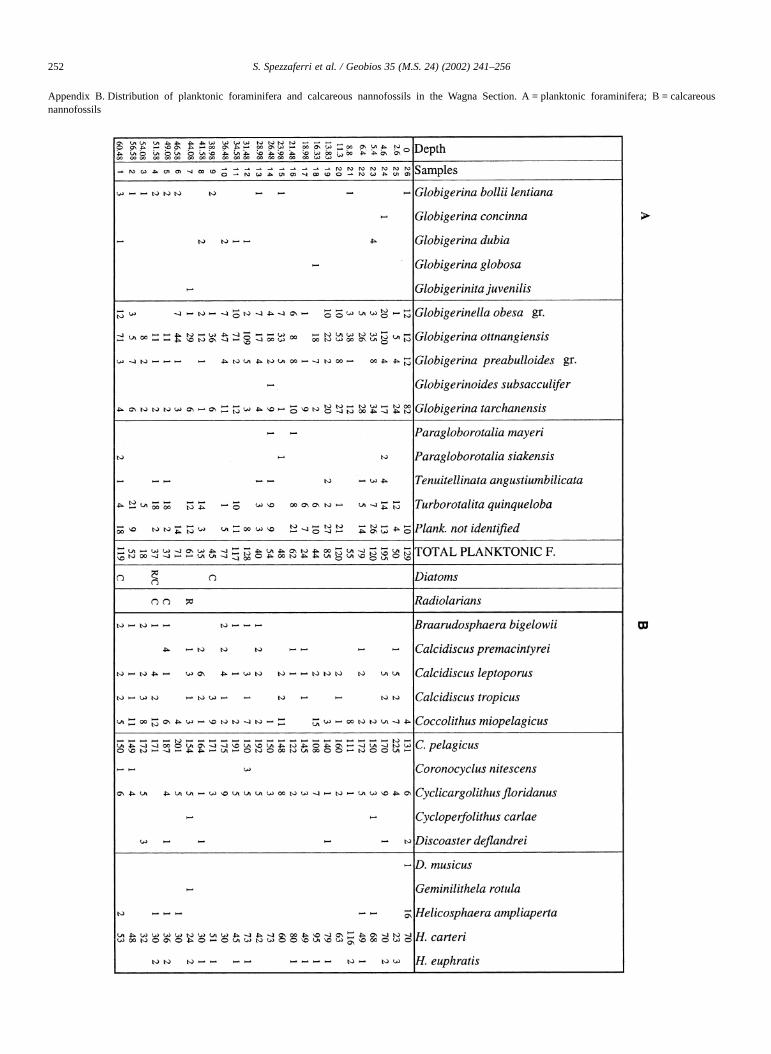
Appendix B. Distribution of planktonic foraminifera and calcareous nannofossils in the Wagna Section. A = planktonic foraminifera; B = calcareousnannofossils
252 S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256
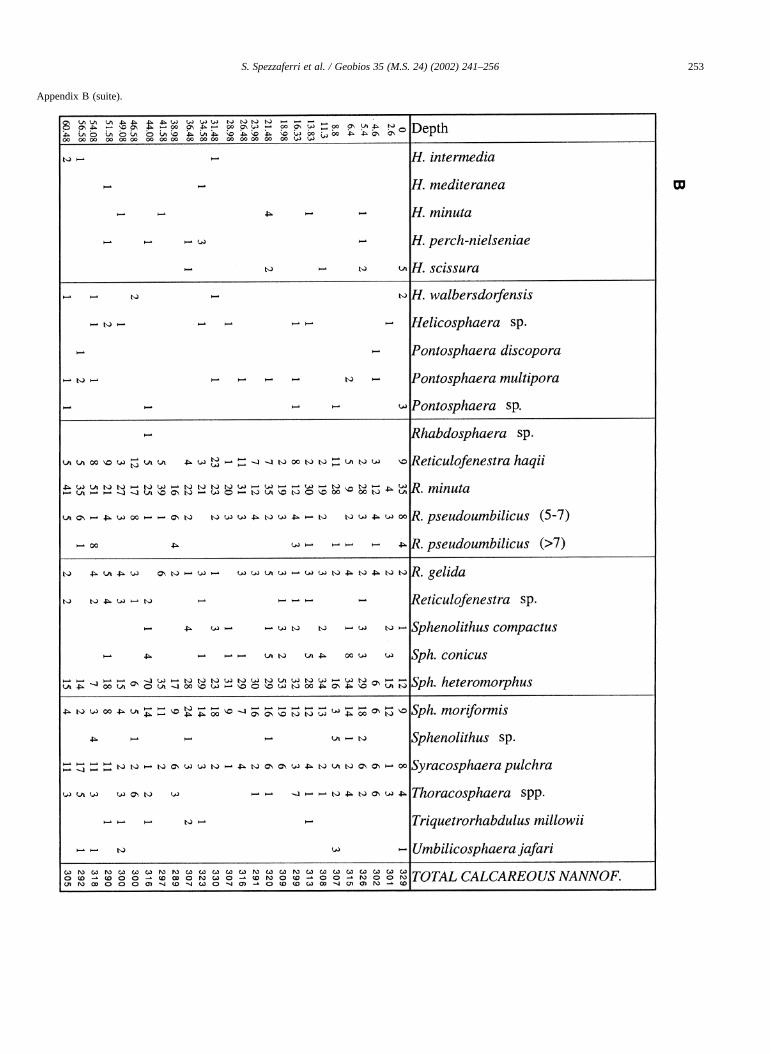
Appendix B (suite).
S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256 253

Appendix C. Distribution of benthic and planktonic foraminifera and calcareous nannofossils in the Retznei Section. A = benthic foraminifera; B = planktonicforaminifera; C = calcareous nannofossils
254 S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256

Acknowledgements
This study is part of Project FWF P13743–BIO focusedon the ecology of the marine Miocene in Austria and fundedby the Austrian Science Foundation. Project FWFP13743–BIO gave financial support to S.S and S.C. Warmthanks to Dr. Pervesler (Vienna) who helped the authors infield work and sampling and to Dr. Roetzel (GeologicalSurvey, Vienna) for providing samples from the GöllersdorfSection.
References
Alexandersson, E.T., 1978. Destructive diagenesis of carbonate sedimentsin the eastern Skagerrak, North Sea. Geology 6, 324–327.
Basso, D., Spezzaferri, S., 2000. The distribution of living (stained) benthicforaminifera in Iskenderun Bay: a statistical approach. BollettinoSocietà Paleontologica Italiana 39 (3), 359–370.
Bernhard, J.M., Sen Gupta, B.K., Borne, P.F., 1998. Benthic foraminiferalproxy to estimate dysoxic bottom-water oxygen concentrations: SantaBarbara Basin, US Pacific continental margin. Journal ForaminiferalResearch 27, 301–310.
Cita, M.B., Vergnaud-Grazzini, C., Chamley, H., Ciaranfi, N.,D’Onofrio, S., 1977. Paleoclimatic record of a long deep sea corefrom the eastern Mediterranean. Quaternary Research 8, 205–235.
Fornaciari, E., Di Stefano, A., Rio, D., Negri, A., 1996. Middle Miocenequantitative calcareous nannofossil biostratigraphy in the Mediterra-nean region. Micropaleontology 42 (1), 37–63.
Fyfe, W.S., Kronberg, B.I., Brown, J.R., 1982. Leonardos, Mount St.Helens Manna. In: Keller, S.A.C. (Ed.), Mount St. Helens: One YearLater. Eastern Washington University Press, pp. 236.
Gupta, A.N., 1997. Paleoceanographic and paleoclimatic history of theSomali Basin during the Pliocene–Pleistocene: multivariate analysesof benthic foraminifera from DSDP Site 241 (Leg 25). JournalForaminiferal Research 27 (3), 196–208.
Haq, B.U., 1978. Calcareous nannoplankton. In: Haq, B.U., Boersma, A.(Eds.), Introduction to Marine Micropaleontology. Elsevier, Amster-dam, pp. 79–107.
Hemleben, C., Spindler, M., Anderson, O.R., 1989. Modern PlanktonicForaminifera. Springer, NY (363 p.).
Hermelin, J.O.R., Shimmield, G.B., 1995. Impact on productivity eventson the benthic foraminiferal fauna in the Arabian Sea over the last150,000 years. Paleoceanography 10 (1), 105–116.
Kaiho, K., 1994. Benthic foraminiferal dissolved-oxygen index and dis-solved levels in the modern ocean. Geology 22 (7), 19–722.
Kapounek, J., Kröll, A., Papp, A., Turnovsky, K., 1965. Die verbreitungvon Oligozän, Unter – und Mittelmiozän in Niederösterreich. Erdoel-Erdgas – Zeitschrift 81, 109–116.
Loubere, P., 1991. Deep-Sea benthic foraminiferal assemblage response toa surface ocean productivity gradient: a test. Paleoceanography 6 (2),193–204.
Mandic, O., Harzhauser, M., Spezzaferri, S., Zuschin, M., this issue. Thepaleoenvironment of an early Middle Miocene Paratethys sequence inNE Austria with special emphasis on paleoecology of mollusks andforaminifera. In: Monegatti, P., Cecca, F., Raffi, S. (Eds.), Interna-tional Conference “Paleobiogeography and Paleoecology 2001” ,Piacenza and Castell’Arquato 2001. Geobios, M.S, 24.
Martin, J.H., Fitzwater, S.E., 1988. Iron deficiency limits phytoplanktongrowth in the north-east Pacific subarctic. Nature 331, 341–343.
Martin, J.H., Coale, K.H., Johnson, K.S., Fitzwater, S.E., Gordon, R.M.,Tanner, S.J., Hunter, C.N., Elrod, V.A., Nowicki, J.L., Coley, T.L.,Barber, R.T., Lindley, S., Watson, A.J., Van Scoy, K., Law, C.S.,Liddicoat, M.I., Ling, R., Stanton, T., Stockel, J., Collins, C.,
Appendix C (suite).
S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256 255

Anderson, A., Bidigare, R., Ondrusek, M., Latasa, M., Millero, F.J.,Lee, K., Yao, W., Zhang, J.Z., Fredrich, G., Sakamoto, C., Chavez, F.,Buck, K., Kolber, Z., Green, R., Falkowski, P.G., Chisholm, S.W.,Hoge, F., Swift, R., Yungle, J., Turner, S., Nightingale, P.L., Hat-ton, A., Liss, P., Tindale, N.W., 1994. Testing the iron hypothesis inecosystems of the equatorial Pacific. Nature 371, 123–129.
Martini, E., 1971. Standard Tertiary and Quaternary Calcareous Nanno-plankton Zonation. Proceedings of the II Planktonic Conference. Ed.Tecnoscienza, Roma (pp.739–785).
Monechi, S., Buccianti, A., Gardin, S., 2000. Biotic signals from nannof-lora across the iridium anomaly in the upper Eocene of the Massig-nano section: evidence from statistical analysis. Marine Micropaleon-tology 39 (1/2), 219–237.
Murray, J.W., 1976. A method of determining proximity of marginal seasto an ocean. Marine Geology 22, 103–119.
Murray, J.W., 2001. The niche of benthic foraminifera, critical thresholdsand proxies. Marine Micropaleontology 41, 1–7.
Murray, J.W., 1991. Ecology and paleoecology of benthic foraminifera.Longman Scientific and Technical, UK (397 p.).
Nishi, H., 1990. The depth distribution of living benthic foraminiferawithin marine sediments of Suruga and Sagami Bays, off the southerncoats of Japan. Benthos 90, 109–115.
Perch-Nielsen, K., 1985. The Tertiary calcareous nannofossils genusHelicosphaera – A basis for discussions. INA Newsletter 7, 89–92.
Rögl, F., 1969. Die miozäne Foraminiferenfauna von Laa an der Thaya inder Molassezone von Niederösterreich. Mitteilungender GeologischeGesellschaft in Wien 61, 63–123.
Rögl, F., 1999. Mediterranean and Paratethys. Facts and hypothesis of anOligocene to Miocene paleogeography (short overview). GeologicaCarpatica 50, 339–349.
Roetzel, R., Cicha, I., Stojaspal, F., Decker, K., Wimmer-Frey, I., Ottner, F.,Papp, H., 1999. C5 Göllersdorf – Ziegleri und Tonbergbau Wiener-berger. Arbeitstatung Geologisches Bundesandstalt, 335–341.
Rupp, C., 1986. Paläoökologie der Foraminiferen in der Sandschalerzone(Badenien, Miozän) des Wiener Beckens. Beiträge zur Paläontologievon Österreich 12, 1–98.
Sachsenhofer, R.F., 1996. The Neogene Styrian Basin: an overview.Mitteilungen Gesselschaft Geologie Bergbaustudenten Oesterreich41, 19–32.
Schönfeld, J., 2001. Benthic foraminifera and pore-water oxygen profiles:a re-assessment of species boundary conditions at the western Iberianmargin. Journal Foraminiferal Research 31 (2), 86–107.
Senès, J., 1960. Entwicklungsgeschichte der Paratethys. MitteilungenGeologisches Gesselschaft Wien 52, 181–187.
Senès, J., Marinescu, F., 1974. Cartes paléogéographiques de la Paratéthyscentrale. Mémoires BRGM 78, 785–792.
Sliter, W.V., 1975. Foraminiferal life and residue assemblages fromCretaceous slope deposits. Geological Society of America Bulletin86, 897–906.
Spezzaferri, S., 1995. Planktonic foraminiferal paleoclimatic implicationsacross the Oligocene–Miocene transition in the oceanic record (Atlan-tic, Indian and South Pacific). Palaeogeography, Palaeoclimatology,Palaeoecology 114, 43–74.
Spezzaferri, S., Coric, S., 2001. Ecology of Karpatian (Early Miocene)foraminifera and calcareous nannoplankton from Laa an der Thaya,Lower Austria: a statistical approach. Geologica Carpathica 52 (6),361–374.
Spezzaferri, S., Cita, M.B., McKenzie, J.A., 1998. The Miocene/Plioceneboundary in the Eastern Mediterranean: results from site 967 and 969.In: Robertson, A.H.F., Emeis, K.C., Richter, C., Camerlenghi, A.(Eds.), Proceedings ODP, Scientific Results, 160, College Station, TX(Ocean drilling Program). pp. 9–28.
Spezzaferri, S., Rögl, F., Coric, S., Hohenegger, J., in press. Paleoenviron-mental changes and agglutinated foraminifera across theKarpation/Badenian (Early/Middle Miocene) boundary in the StyrianBasin (Austria, Central Paratethys). Proceedings of the Sixth Inter-national Workshop on Agglutinated Foraminifera. Grzybowski Foun-dation Special Publication.
Steininger, F.F., Wessely, G., Rögl, F., Wagner, L., 1989. Tertiary sedimen-tary history and tectonic evolution of the Eastern Alpine Foredeep.Giornale di Geologia 48 (1/2), 285–297.
Stille, H., 1924. Grundfragen vergleichender Tektonik. Borntraeger, Berlin(443 p.).
Vasicek, M., 1951. The contemporary state of the microbiostratigraphicresearch of the Miocene sedimentary deposits in the Out-CarpathianNeogene Basin in Moravia. Sborn. Ust. Ust. Geol. (Paleont.) 18,45–195.
256 S. Spezzaferri et al. / Geobios 35 (M.S. 24) (2002) 241–256




























