Embed Size (px)
Citation preview

Vol. 5, 1235-1241, November 1994 Cell Growth & Differentiation 1235
Arsenate Perturbation of Human KeratinocyteDifferentiation’
Donna J. Kachinskas, Marjorie A. Phillips, Qin Qin,James D. Stokes, and Robert H. Rice2
Department of Environmental Toxicology, University of California, Davis,
California 9561 6-8S88
Abstract
Treatment of cultured malignant human keratinocyteswith sodium arsenate greatly suppressed expression ofinvolucrin, a specific marker of keratinocytedifferentiation. This action was primarily attributable toinhibition of involucrin transcription according tomessage run-on and stability measurements. Involucrinwas suppressed in nontumorigenic keratinocytes as well,although the efficacy of suppression was less dramatic incells derived from clinically normal epidermis. Severaltransition metal oxyanions (vanadate, molybdate, andtungstate) also substantially suppressed involucrinexpression, but okadaic acid was ineffective.Immunoblotting detected marked increases in tyrosinephosphorylation of several proteins as a consequence ofarsenate treatment of the cultures, while mobility shiftanalysis revealed a dramatic loss of DNA binding by thetranscription factor AP2. These findings support aproposed role for altered levels of protein tyrosinephosphorylation in keratinocyte differentiation. Theyalso suggest that arsenate perturbs the differentiationprogram in target cells by altering this phosphorylationlevel and transcription factor activity.
Introduction
Keratinocytes carry out an intricate differentiation programwhich is now amenable to study in culture (1 ). A charac-teristic feature of terminal differentiation in this cell type issynthesis of the cross-linked envelope (2), the first compo-nent of which to be identified was involucrin (3). A highlyrepetitive protein containing many glutamine residues (4),involucrin is a prominent substrate for the membrane-bound keratinocyte transglutaminase (5), an isozyme spe-cific for this cell type (6), which is important for envelopesynthesis (7). Ordinarily present at very low levels in basaland lower spinous cells, both enzyme and substrate firstappear coordinately in upper spinous cells of epidermis (8).However, they are clearly under distinct regulation, since inculture transglutammnase appears much earlier than involu-cnn as the cells reach confluence, and the microenviron-ment can be altered so as to yield expression of eithermarker in the absence of the other (9, 1 0). Resolving the
Received 4/21/94; revised 8/1 1/94; accepted 8/30/94.
1 This research was supported by Grants AR271 30 (National Institute of
Arthritis, Musculoskeletal and Skin Diseases), P4204699 (NIEHS SuperfundBasic Research Program), ES07059 (NIEHS Graduate Training Grant), andES05707 (NIEHS Center for Health Effects).2 To whom requests for reprints should be addressed.
multiple layers of regulation responsible for expression ofsuch markers will assist in understanding the coordinationof keratinocyte programming and its perturbation bycarcinogens such as arsenic.
Previous work has shown that involucrin levels are sup-pressed in low calcium medium and by glucocorticoid
deprivation (1 0). Involucrin message levels are stimulatedby acute treatment of cultures with the tumor promoter1 2-O-tetradecanoylphorbol-1 3-acetate (1 1 ) but suppressedby chronic treatment with this agent (9). The phorbol estereffects, interpreted plausibly as a consequence of proteinkinase C stimulation versus depletion (acute versus chronictreatment), indicate the importance of serine/threoninephosphorylation events in expression ofthis marker. Recentfindings also show the importance of tyrosine phosphory-lation in keratinocyte differentiation, particularly in the re-sponse to calcium (12). Actually, phorbol ester effects caninclude modulation of protein tyrosine phosphatase activi-ties (1 3), and the activity of the 6 isozyme of protein kinaseC (prominent in keratinocytes) can be modulated bytyrosine phosphorylation (14).
A proper balance between phosphorylation and dephos-phorylation reactions clearly is critical to cell function.Protein tyrosine kinases have received much attention withrespect to replication in view of their key actions in signaltransduction of growth factor responses. Protein phospho-tyrosine phosphatases are similarly critical and appear tocomprise a diverse family as well (1 5). The importance ofboth classes of enzyme for cell differentiation is empha-sized by recent reports of transcription factors regulatedthrough tyrosine phosphorylation (16). Present findingsare highly compatible with these reports and suggest thatarsenate perturbs involucrin expression, at least in partby acting as a protein tyrosine phosphatase inhibitor.A preliminary account of these results has appeared(17).
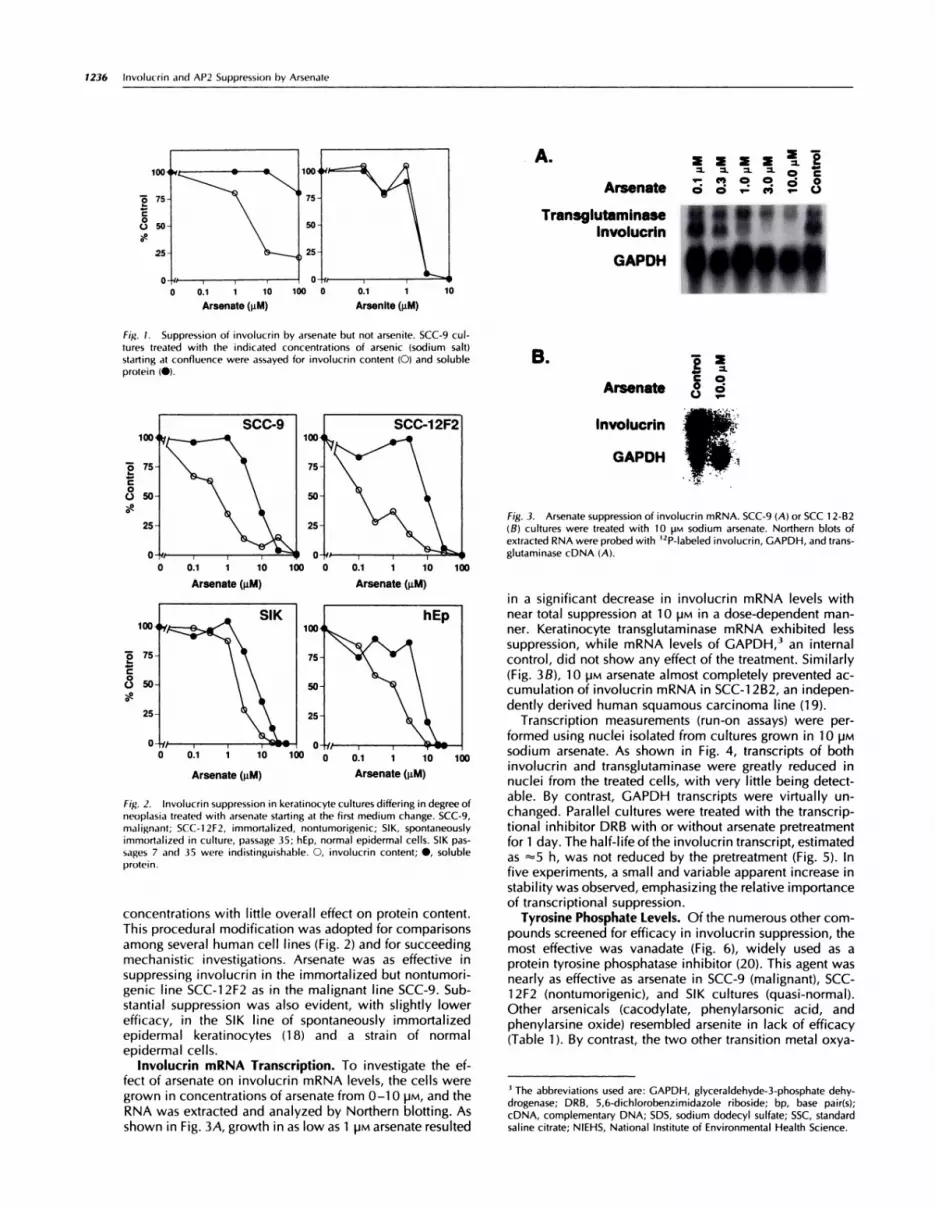
ResultsInvolucrin Suppression. Ordinarily, involucrin expressionin SCC-9 cultures is undetectable or very low at confluenceand reaches maximal values 10 days later (9). When thecultures were treated with arsenate at confluence, the sub-sequent expression of involucrin was suppressed. The effecton this differentiation marker protein was evident at arsen-ate concentrations as low as 1 pM, while toxic effects,visible as a reduction in total cell protein, were not ob-served at 30 �M and were slight at 100 �M (Fig. 1). In theseinitial experiments, maximal involucrin suppression of 80%was observed at 10 �M. This action at confluence is indic-ative of a direct effect on differentiation rather than anindirect one through growth perturbation. By contrast, so-dium arsenite was clearly toxic to the cells at 3 �M butshowed no specific effect on involucrin.
When SCC-9 cultures were treated with arsenate startingat the firstmedium change instead of at confluence, involu-cnn suppression was more dramatic, generally >90% at
100
.� 75
C0
050
at
25
100
75
50
25
10 - _�_j
Fig. 1. Suppression of involucrin by arsenate but not arsenite. SCC-9 cul-
tures treated with the indicated concentrations of arsenic (sodium salt)starting at confluence were assayed for involucrin content (0) and solubleprotein (#{149}).
0
C00at
0 0.1 1 10 100 0 0.1 1 10 100
Arsenate (gM) Arsenate (�tM)
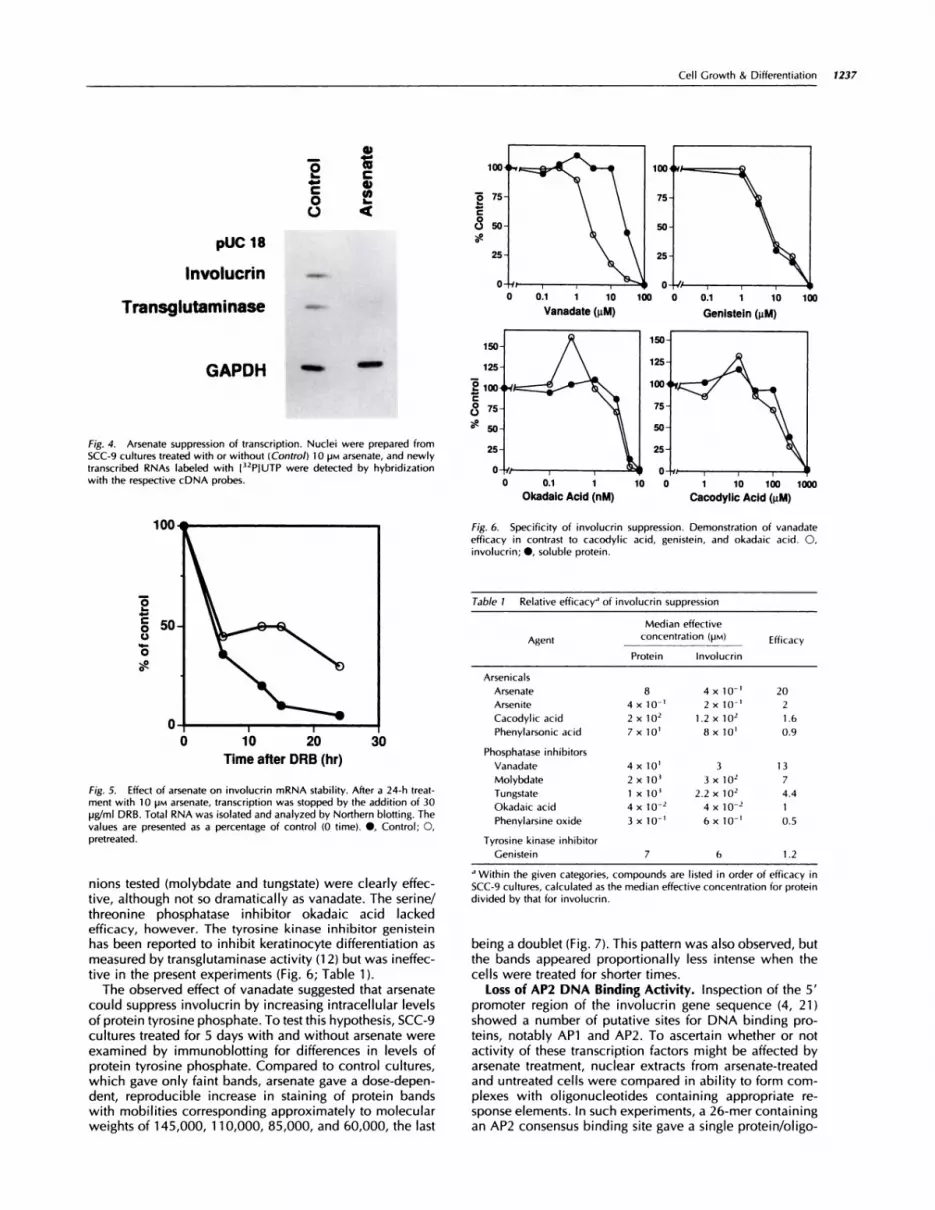
Fig. 3. Arsenate suppression of involucrin mRNA. SCC-9 (A) or 5CC 1 2-B2
(B) cultures were treated with 10 ira sodium arsenate. Northern blots of
extracted RNA were probed with ‘2P-labeled involucrin, GAPDH, and trans-glutaminase cDNA (A).
0
C00at
0 0.1 1 10 100 o o.i i io
Arsenate (pM) Arsenate (gM)
100
Fig. 2. Involucrin suppression in keratinocyte cultures differing in degree ofneoplasia treated with arsenate starting at the first medium change. SCC-9,
malignant; 5CC-i 2F2, immortalized, nontumorigenic; 51K, spontaneouslyimmortalized in culture, passage 35; hEp, normal epidermal cells. 51K pas-
sages 7 and 35 were indistinguishable. 0, involucrin content; #{149},solubleprotein.
1236 Involucrin and AP2 Suppression by Arsenate
0.1
Ars1 10 100
enate (pM)
0 0.1
Arseni
1
te (gM)
concentrations with little overall effect on protein content.This procedural modification was adopted for comparisons
among several human cell lines (Fig. 2) and for succeedingmechanistic investigations. Arsenate was as effective insuppressing involucrin in the immortalized but nontumori-genic line SCC-i 2F2 as in the malignant line SCC-9. Sub-stantial suppression was also evident, with slightly lowerefficacy, in the 51K line of spontaneously immortalizedepidermal keratinocytes (18) and a strain of normalepidermal cells.
Involucrin mRNA Transcription. To investigate the ef-fect of arsenate on involucrin mRNA levels, the cells weregrown in concentrations of arsenate from 0-1 0 pM, and theRNA was extracted and analyzed by Northern blotting. Asshown in Fig. 3A, growth in as low as 1 �M arsenate resulted
A.�. �. �. �- 0
,- C., 0 e � 0Arsenate o o �.: �,; �.
Transglutaminase
lnvolucrln
GAPDH
B. o��CO
Arsenate � 2
lnvolucrin
GAPDH
in a significant decrease in involucrin mRNA levels withnear total suppression at 1 0 �M in a dose-dependent man-ner. Keratinocyte transglutaminase mRNA exhibited lesssuppression, while mRNA levels of GAPDH,3 an internalcontrol, did not show any effect of the treatment. Similarly(Fig. 3B), 10 �M arsenate almost completely prevented ac-cumulation of involucrin mRNA in SCC-i 2B2, an indepen-dently derived human squamous carcinoma line (19).
Transcription measurements (run-on assays) were per-formed using nuclei isolated from cultures grown in 1 0 �M
sodium arsenate. As shown in Fig. 4, transcripts of bothinvolucrin and transglutaminase were greatly reduced innuclei from the treated cells, with very little being detect-able. By contrast, GAPDH transcripts were virtually un-changed. Parallel cultures were treated with the transcrip-tional inhibitor DRB with or without arsenate pretreatmentfor 1 day. The half-life ofthe involucrin transcript, estimatedas “5 h, was not reduced by the pretreatment (Fig. 5). Infive experiments, a small and variable apparent increase instability was observed, emphasizing the relative importanceof transcriptional suppression.
Tyrosine Phosphate Levels. Of the numerous other com-
pounds screened for efficacy in involucrin suppression, themost effective was vanadate (Fig. 6), widely used as aprotein tyrosine phosphatase inhibitor (20). This agent wasnearly as effective as arsenate in SCC-9 (malignant), 5CC-1 2F2 (nontumorigenic), and 51K cultures (quasi-normal).Other arsenicals (cacodylate, phenylarsonic acid, andphenylarsine oxide) resembled arsenite in lack of efficacy(Table 1 ). By contrast, the two other transition metal oxya-
3 The abbreviations used are: GAPDH, glyceraldehyde-3-phosphate dehy-
drogenase; DRB, 5,6-dichlorobenzimidazole riboside; bp, base pair(s);
cDNA, complementary DNA; SDS, sodium dodecyl sulfate; SSC, standard
saline citrate; NIEHS, National Institute of Environmental Health Science.
0 0.1 1 10
Vanadate (gM)1000 � 100
Genlstein (gM)
10 0 1 10 100 1000
Cacodylic Acid (pM)
25
fly-
0 0.1 1
Okadaic Acid (nM)
GAPDH �
Fig. 4. Arsenate suppression of transcription. Nuclei were prepared fromSCC-9 cultures treated with or without (Control) 1 0 ira arsenate, and newlytranscribed RNAs labeled with I32PIUTP were detected by hybridizationwith the respective cDNA probes.
0I-
0
0
0
Fig. 6. Specificity of involucrin suppression. Demonstration of vanadateefficacy in contrast to cacodylic acid, genistein, and okadaic acid. 0,
involucrin; #{149},soluble protein.
10 20
Time after DRB (hr)
Fig. S. Effect of arsenate on involucrin mRNA stability. After a 24-h treat-ment with 1 0 pra arsenate, transcription was stopped by the addition of 30pg/mI DRB. Total RNA was isolated and analyzed by Northern blotting. The
values are presented as a percentage of control (0 time). #{149},Control; 0,pretreated.
Cell Growth & Differentiation 1237
pUC 18
Involucrin
Transglutaminase
_5 ��- C
-C0
C) <
nions tested (molybdate and tungstate) were clearly effec-tive, although not so dramatically as vanadate. The serine/threonine phosphatase inhibitor okadaic acid lackedefficacy, however. The tyrosine kinase inhibitor genisteinhas been reported to inhibit keratinocyte differentiation asmeasured by transglutaminase activity (1 2) but was ineffec-tive in the present experiments (Fig. 6; Table 1).
The observed effect of vanadate suggested that arsenatecould suppress involucrin by increasing intracellularlevelsof protein tyrosine phosphate. To test this hypothesis, SCC-9cultures treated for 5 days with and without arsenate wereexamined by immunoblotting for differences in levels ofprotein tyrosine phosphate. Compared to control cultures,which gave only faint bands, arsenate gave a dose-depen-dent, reproducible increase in staining of protein bandswith mobilities corresponding approximately to molecularweights of 1 45,000, 1 1 0,000, 85,000, and 60,000, the last
0
C
at
150
125
EiooC
(� 75
at50
Table 1 Relative efficacy” of involucrin suppression
Median effective
Agent concentration (pra) Efficacy
Protein lnvolucrin
Arsenicals
Arsenate 8 4 x 1 0 � 20
Arsenite 4 x 10-’ 2 x 10-i 2
Cacodylic acid 2 x 102 1.2 x 102 1.6
Phenylarsonic acid 7 x 10� 8 x 10’ 0.9
Phosphatase inhibitors
Vanadate 4 x 10’ 3 13
Molybdate 2 x 10’ 3 x 102 7
Tungstate 1 x 10� 2.2 x 102 4.4
Okadaic acid 4 x 10_2 4 x 102 1
Phenylarsine oxide 3 x 10_i 6 x 10�’ 0.5
Tyrosine kinase inhibitor
Genistein 7 6 1.2
.1Within the given categories, compounds are listed in order of efficacy in
SCC-9 cultures, calculated as the median effective concentration for protein
divided by that for involucrin.
being a doublet (Fig. 7). This pattern was also observed, butthe bands appeared proportionally less intense when thecells were treated for shorter times.
Loss of AP2 DNA Binding Activity. Inspection of the 5’promoter region of the involucrin gene sequence (4, 21)showed a number of putative sites for DNA binding pro-teins, notably APi and AP2. To ascertain whether or notactivity of these transcription factors might be affected byarsenate treatment, nuclear extracts from arsenate-treatedand untreated cells were compared in ability to form com-plexes with oligonucleotides containing appropriate re-sponse elements. In such experiments, a 26-mer containingan AP2 consensus binding site gave a single protein/oligo-

A.LLM Arsenate
kDa 0 3 10
200 -
91-
46 -
30 -
Com�tor APi AP2 AP3 SP1 NFIICTF
r��.L�:1i ..I .jr-�:-1 r�i
* � ��- AP2 -�
77V �NNIIU#{149}� I
Control Arisnats
AnU-AP2 - 4 - +
SupsrshItt-�-
B.
Fig. 8. Loss of AP2 DNA binding activity in arsenate-treated SCC-9 cells as
detected by mobility shift assays. A, left, a 64-bp oligonucleotide formedthree electrophoretically distinguishable protein complexes when incubatedwith SCC-9 control nuclear extract, the slowest migrating of which was not
seen using extracts from arsenate-treated cultures. A, right, formation of themost slowly migrating complex was inhibited by an oligonucleotide con-
taming an AP2 response element, but oligonucleotides containing four otherresponse elements present in the 64-mer were ineffective at the indicatedratios of competitor to labeled oligonucleotide (1 0-1 OOX). The two leftmost
lanes correspond to the labeled oligonuleotide alone (‘) and with controlextract but no competitor (-). B, left, a 26-bp oligonucleotide containing anAP2 response element formed a complex with extracts from control but notarsenate-treated cultures. Anti-AP2 antibody reduced the mobility of thecomplex formed by control extract (Supershift). B, right, a change in DNAbinding activity was evident in arsenate-treated versus control extracts usinga 22-bp oligonucleotide containing an APi response element.
1238 Involucrin and AP2 Suppression by Arsenate
Fig. 7. Protein phosphotyrosine immunoblotting. Cultures were treatedwith 0, 3, or 1 0 ira arsenate as indicated for 5 days prior to harvest. Letters a-d(right margin) point to protein bands with elevated phosphotyrosine. Rainbow
molecular weight protein standards (Amersham, Arlington Heights, IL) with
mobilities corresponding to the indicated molecular weights (left margin, top tobottom) were myosin, phosphorylase b, ovalbumin, and carbonic anhydrase.
nucleotide complex readily detectable by native gel elec-trophoresis. This complex contained AP2 as judged by theshift in mobility of the complex in the presence of anti-AP2antibody and was undetectable in extracts of arsenate-treated cells (Fig. 88). In parallel trials, a reduction inamount (and a slight increase in mobility) was noted for acomplex formed by a 22-bp oligonucleotide containing anAPi response element (Fig. 88).
Confirming these findings, extracts were compared inbinding to a 64-bp oligonucleotide with consensus sites forAPi , AP2, AP3, SP1 , and NF1/CTF. Using extracts fromcontrol cultures, three complexes were separable by elec-trophoresis, of which the slower migrating one was missingin lanes using extracts from arsenate-treated cultures (Fig.8A). As illustrated, competition with increasing amounts ofshorter oligonucleotides, each containing one of the fiveconsensus response elements, showed that only the slowestmigrating complex was removed by the competition andthat the only successful competitor was an oligonucleotidecontaining a consensus sequence for AP2.
Discussion
The transition metal oxyanions vanadate, molybdate, andtungstate resemble phosphate in certain chemical proper-ties (e.g., tetrahedral geometry) and share a number ofbiochemical actions (22-24), such as phosphatase inhibi-tion. Including arsenate, they have shown inhibition, atrelatively high concentration, of model enzymes such asacid phosphatase (24). They are thought to affect a varietyof phosphatases, however, and hence are believed to havehigher affinity for intracellular phosphatases not yet identi-fied. Although it may have other actions as well, vanadatehas shown specificity for inhibition of protein tyrosine phos-phatases (20, 25). Findings with this agent, closest to arsen-ate in efficacy for involucrin suppression, are consistentwith arsenate acting at least in part by producing an imbal-ance in protein tyrosine phosphate, an effect that has nowbeen observed directly. Specificity is evident in that phe-nylarsine oxide reportedly inhibits tyrosine phosphatase ac-tion in T cells (26) but was ineffective in the present exper-iments, as was the protein tyrosine kinase inhibitor,genistein. Although an imbalance in protein serine/threo-nine phosphorylation could result in downstream alter-
ations in protein tyrosine phosphorylation, okadaic acidalso was ineffective in altering involucrin levels. The inter-connections between phosphatase and kinase reactionpathways create difficulties in finding the specific tar-get(s) of arsenate in intact cells. Nevertheless, furtheridentification of agents that may act in a similar way,such as other metal or metalloid oxyanions, could beinstrumental at least in ruling out certain classes of phos-phatase whose inhibition by such agents can bedetermined in vitro.
Identification of specific signal transduction pathwayswhich arsenate perturbs may lead to a clearer understand-ing of those regulating expression of differentiation features.A relationship between two proteins exhibiting greatly ele-vated tyrosine phosphorylation in the present work(Mr “�‘60,000 and “85,000) and two of similar mobilitywhose level of tyrosine phosphorylation was noted to bestimulated by calcium and tetradecanoyl phorbol acetate inmouse keratinocytes (12) is unclear, as is the participationof any of the four seen presently in an appropriate signalingpathway. However, especially if the large increases intyrosine phosphorylation noted are due to altered posttrans-lational modification (as opposed to new synthesis), further

Cell Growth & Differentiation 1239
4 Manuscript in preparation.
study of these proteins may help characterize such path-ways. Ultimately, changes in involucrin level are antici-pated to resultfrom altered regulation of transcription fac-tors, a type of perturbation of which vanadate is capable(27). Present results show that arsenate suppresses theactivity of the AP2 transcription factor. This it could doplausibly as a protein phosphotyrosine phosphatase in-hibitor by interfering with reactions that, directly or in-directly, alter the phosphorylation state of AP2 or ofanother protein with which AP2 may interact. Alterna-tively, substantial reduction in AP2 transcription, a phe-nomenon that would be of great interest, cannot be ruledout.
Analysis of the involucrin promoter is at a rudimentarystage and iscomplicated by possible regulatory activityof a1 -kilobase intron located between the transcriptional andtranslational start sites (21). Putative APi response ele-ment(s), recognizable by inspection of the genomic 5’-flanking DNA sequence, have been proposed to participatein transcriptional regulation by phorbol esters (1 1). Al-though APi may well be a target for arsenate action, ourresults point most clearly to the transcription factor AP2 asan important target, for which at least three putative re-sponse elements are evident in the 5’-flanking sequence byinspection of the first 200 bp of the promoter region. Ex-periments in progress show that mutation of some of thesesites, preventing AP2 binding, substantially reduces reportertranscription in transient transfections,4 although whetherthis can account entirely for the degree of involucrin sup-pression is not yet clear. AP2 has demonstrated impor-tance for the expression of the K14 gene and may beimportant for other keratins as well (28). Thus, exploringpossible effects on several differentiation markers, in-cluding the keratins, may help to elucidate sensitive tran-scriptional response elements and to characterize howextensive is the transcriptional domain of arsenateperturbation.
Arsenic is a known human carcinogen for skin, lung, andlikely other organs (29-31) and is of great concern at ex-posure levels encountered by a substantial fraction of thepopulation (32, 33). Setting scientifically defensible expo-sure limits has been difficult in the absence of a suitableanimal model, thus encouraging mechanistic studies onbiochemical reactions of arsenicals with isolated macro-molecules and cultured cells. Inorganic arsenic has a lowpotency as a mutagen and tumor initiator (34), suggesting acomplicated interactionwith targetcells.Itscarcinogenicityand efficacy as a comutagen could result from action as aclastogen which, in the case of arsenite (the more acutelytoxic +3 oxidation state), is attributable to inhibition ofDNA ligase (35). Arsenite forms stable complexes with freesulfhydryls, particularly those in a vicinal configuration inorganic compounds such as lipoic acid (36) and proteins.This behavior explains its selective inactivation of glucocor-ticoid as opposed to other steroid receptors (37) and per-haps its induction of heat shock proteins and the MDR1
multidrug resistance gene (38, 39). This could be the more
carcinogenic oxidation state, since it is more potent inproducing morphological transformation of Syrian hamster
embryo or mouse 3T3 cells (40, 41) and gene amplificationin cultured mouse 3T6 cells (42). The latter phenomenon is
consistent with suggestions that arsenic acts as a late stagepromoter, since normal human cells are known to be resis-tant to gene amplification, unlike those with seriouslydamaged genomes (43).
Arsenate (the +5 oxidation state)isthe more stable formunder ordinary ambient cond itions (oxid izi ng atmosphereand neutral pH) and in chronic dosing of experimentalanimals is the predominant form in the circulation (44).Partial reduction to arsenite can occur in the liver and,judging by culture experiments (41), in some peripheraltissues. Thus, exposure to both states usually occurs simul-taneously in practice. It has long been recognized thatarsenate could have pathological effects on cells through itsincorporation in place of phosphate into macromoleculesto yield markedly less stable arsenate esters and diesters(45).This would resultin futileuse of cellenergy stores (36)and in DNA strand breaks. However, present results em-phasize that arsenate, by a previously unsuspected mech-anism, can perturb keratinocyte gene regulation in waysthat arsenite does not. Moreover, its effect on involucrin isobservable even in normal epidermal cells. Thus, in addi-tion to providing insights into keratinocyte differentiation,these findings raise the possibility that arsenic carcinoge-nicity may result from more than a single type of perturba-tion in which the arsenate/arsenite combination is moreeffective than either form alone. Further investigation willbe required to determine the validity of this speculation.
Materials and Methods
Cell Culture. Human 5CC lines (1 9, 46), all epidermalexcept for SCC-9 (lingual), were cultivated with a feederlayer of lethally irradiated 3T3 cells in a mixture ofDulbecco-Vogt Eagle’s and Ham’s F-12 media (3:1) supple-mented with fetal bovine serum (5%), hydrocortisone (0.4pg/mI), insulin (5 pg/mI), transferrin (5 pg/mI), T3 (20 pM),
and adenine (0.1 8 mM) (47). The 51K line of spontaneouslyimmortalized keratinocytes, derived from clinically normalepidermis (1 8), was propagated in the same way. In mostexperiments, cells were inoculated at a density of iO�/1 0-cm culture and reached confluence in 7-1 0 days. Cul-tures were treated with reagent grade sodium arsenate orother compound at each medium change (twice weekly),starting at confluence (initial experiments) or 3 days afterinoculation.
Involucrin Measurement. Cultures were treated until 10days past confluence, then rinsed in phosphate-bufferedsaline, scraped from the dishes into microfuge tubes, andstored frozen until assay. While thawing in an ice bath, thesamples were sonicated. Aliquots of the supernatants, clar-ified in a microfuge, were measured in triplicate using acompetitive enzyme-linked immunoassay (9, 48). Therange of values for a given sample ordinarily was within 5%of the mean. Total soluble protein was determined withbicinchoninic acid (Pierce Chemical Co., Rockford, IL) orCoomassie G-250 (Bio-Rad, Richmond, CA). In normal epi-dermal and 51K cultures, involucrin comprised 1-2% of thesoluble protein, while in SCC-9 and SCC12F2, involucrinwas 0.05-0.08% of the soluble protein.
Northern Blotting. After 1 0-1 5 days of treatment, cul-tures were rinsed with phosphate-buffered saline and dis-solved in 1-2 ml of6 Mguanidinethiocyanate-0.1 MTrisCl(pH 7.5)-1% f3-mercaptoethanol and stored at -80#{176}Cuntilextraction. RNA was isolated by CsCI centrifugation (49)with modifications for use of the Beckman TL1 00 tabletop

1240 Involucrin and AP2 Suppression by Arsenate
9. Rice, R. H., Rong, x., and Chakravarty, R. Suppression of keratinocytedifferentiation in SCC-9 human squamous carcinoma cells by benzo-
ultracentrifuge. For Northern blotting (50), total cellularRNA (25 pg) was electrophoresed through a 1 .1 % agarosegel containing 0.66 M formaldehyde and transferred to anylon membrane (Biotrans). Quantitation of the fluores-
cence in the 285 ribosomal band by densitometry of anegative was used to normalize the amounts of RNA com-pared. Uniform loading of mRNA was confirmed by com-parison of GAPDH message using a cDNA probe. Themembrane was prehybridized in 1 M NaCI, 100 m�iNaH2PO4 (pH 6.5), 1 0 mM EDTA, 1 % SDS, 200 pg/mI yeasttRNA, and 200 pg/mI salmon sperm DNA at 65#{176}Cfor 2-4h. A mixture of involucrin, transglutaminase, and GAPDHcDNA probes (each 1 X 106 cpm/ml), labeled with
L32PIdCTP by random priming using Klenow polymerase,were added and hybridized for 24 h. Prior to autoradiog-raphy, the membrane was washed twice each with 1 M
NaCI, 50 mM NaH2PO4, 1 mivi EDTA, and 1 % SDS and with0.5 M NaCI, 50 mM NaH2PO4, 1 m�i EDTA, and 0.5% SDSand once with 0.5X SSC [(1 x SSC = 1 50 mt�i NaCl-1 5 mr�iNa citrate, pH 7.0) (20 mm at 65#{176}C)].Each biochemicalmeasurement was repeated at least three times, of which arepresentative result is illustrated.
mRNA Stability and Run-On Assays. To determine in-volucrin mRNA stability, cells were held for 7-1 2 days atconfluence and then treated with 30 pg/mI DRB (51), andthe RNA was isolated for Northern blotting. Arsenic-pre-treated cells were exposed to 1 0 �M disodium arsenate for24 h before administration of the DRB.
For transcriptional run-on assays, cultures were grown in10 �M sodium arsenate from the first medium change untila week past confluence (“1 0 days). Nuclei were preparedfrom arsenate-treated and untreated cells in hypotonicbuffer (52) and stored at -80#{176}Cin aliquots (“�‘2 x 1 0” nucleiprepared from 2-4 10-cm dishes). After run-on transcrip-tion, RNA was purified (53) and hybridized to 5 pg oflinearized plasmid that was denatured and slot blotted ontonitrocellulose (Schleicher and Schuell). The plasmids usedwere pSV2INVECE [a subclone of pAI-3H6B (4) which con-tains the entire coding region of involucrin], TG1 3 (54),GAPDH (55), and pUC 1 8 as a control for nonspecifichybridization. Filters were prehybridized for 1 h at 42#{176}Cin2 ml of 50% formamide, 0.6 M NaCI, 50 mt�i sodium phos-phate (pH 6.5), 10 mM EDTA, 1% SDS, and 100 pg/mIdenatured salmon sperm DNA; then filters were hybridizedfor 48 h with 3 x 1 0� cpm of radiolabeled RNA. Filters werewashed twice for 1 5 mm in 2X SSC-0.1 % SDS at 65#{176}C,rinsed in 2X SSC, and then treated with 1 0 mg/mI RNase A(Sigma Chemical Co., St. Louis, MO) in 2X SSC at 37#{176}Cfor45 mm, followed by a final wash in 2X SSC at 37#{176}C.
Anti-Phosphotyrosine Immunoblotting. Cultu res weretreated at the first medium change with the indicated chem-icals and concentrations. Five days later, by which timethey were approaching confluence, each was lysed in 1 mlof 1 0 mM Tris-HCI (pH 7.5), 0.27 M sucrose, 7.5 mt�i f3-mer-captoethanol, 2 mM EDTA, 2 mivi EGTA, 5 pg/mI leupeptin,0.1 mM phenylmethylsulfonyl fluoride, 5 m�i sodium PP1,25 m�i sodium fluoride, and 0.2% Triton X-100, scrapedinto microfuge tubes, and frozen. Samples of 1 50 pg ofprotein were electrophoresed in 1 2.5% polyacrylamideSDS gels and transferred to nitrocellulose. The membraneswere blocked and incubated with a commercial anti-phos-photyrosine monoclonal antibody; the color developed asrecommended for the immunoblotting kit used (UpstateBiotechnology, Lake Placid, NY). In control experiments,where cultures were treated with epidermal growth factor
(not normally added since 5CC lines do not require it forgrowth) or with fresh medium after serum starvation,i mmunoblotti ng revealed strongly reactive bands.
Mobility Shift Analysis. Nuclear extracts were preparedby standard methods (56) from SCC-9 cultures treated with0 or 10 �M sodium arsenate for 2 weeks and harvested 10days after confluence. Aliquots of nuclear extract (5 pgof protein) were incubated at room temperature for 1 hwith 20-35 ng of labeled oligonucleotide in an incubationbuffer [1 2% glycerol, 1 2 mt�’i 4-2-hydroxyethyl)-1 -pipera-zineethanesulfonic acid (pH 7.9), 4 mrsi Tris-HCI (pH 7.5),1 50 mt�i KCI, 0.01 % Nonidet P-40, 1 mtvi EDTA, and 1 mrvidithiothreitol) supplemented with 2 pg of poly(dl-dC). La-beled with [‘y-32P]dATP to approximately 1 .5 X 1 0� cprn/ngusing T4 polynucleotide kinase, the oligonucleotides werecommercial, double-stranded preparations (all fromStratagene) containing either an APi (5’-CTAGTGAT-GAGTCAGCCGGATC-3’) or AP2 response element (5’-GATCGAACTGACCGCCCGCGGCCCGT-3’) or responseelements for APi , AP2, AP3, SP1 , and NF1/CTF (5’-GATC-GGGGCGGGGATCAGCCAATGATCTGAGTCAGGATCA-CCGCCCGCGGGATCACTTTCCACA-3’). Unlabeled dou-ble-stranded competitor oligonucleotides for APi and AP2were the same as above, while those for AP3 (5’-CTAGTGGGACTTTCCACAGATC-3 ‘), SP1 (5 ‘-GATC-GATCGGGGCGGGGCGATC-3’), and NFI/CTF (5’-AlTTT-GGCTTGAAGCCAATATG-3’) were also from Stratagene.In each case, parallel incubations were performed using acommercial HeLa cell nuclear extract (Stratagene) as asource of authentic AP2. In some samples, the bindingreactions were supplemented with 200 ng of AP2-specificantibody (Santa Cruz Biotechnology). After incubation, oh-gonucleotide-protein complexes were examined by nativeelectrophoresis in 4% polyacrylamide gels and subsequentautoradiography.
Acknowledgments
We thank Dr. P. Richard Vulliet and Kristin KuIp for generous assistance inprotein phosphotyrosine immunoblotting.
References1 . Watt, F. M. Terminal differentiation of epidermal keratinocytes. Curr.Opin. Cell Biol., 1: 1107-1115, 1989.
2. Reichert, U., Michel, S., and Schmidt, R. The cornified envelope: a keystructure of terminally differentiating keratinocytes. In: M. Blumenberg andM. Darmon (eds.), Molecular Biology of the Skin: The Keratinocyte,pp. 107-1 50. New York: Academic Press, 1993.
3. Rice, R. H., and Green, H. Presence in human epidermal cells ofa solubleprotein precursor ofthe cross-linked envelope: activation ofthe cross-linkingprocess by calcium ions. Cell, 18: 681-694, 1979.
4. Eckert, R. L., and Green, H. Structure and evolution ofthe human involu-cnn gene. Cell, 46: 583-589, 1986.
5. Simon, M., and Green, H. Enzymatic cross-linking of involucrin and otherproteins by keratinocyte particulates in vitro. Cell, 40: 329-336, 1985.
6. Parenteau, N. L., Pilato, A., and Rice, R. H. Induction of keratinocyte typeI transglutaminase in epithelial cells of the rat. Differentiation, 33: 1 30-141,1986.
7. Rice, R. H., Mehrpouyan, M., Qin, Q., and Phillips, M. A. Transglutami-nases in keratinocytes. In: I. M. Leigh, F. M. Watt, and B. Lane (eds.), TheKeratinocyte Handbook. Cambridge, England: Cambridge University Press,1994.
8. Thacher, S. M., and Rice, R. H. Keratinocyte-specific transglutaminase ofcultured human epidermal cells: relation to cross-linked envelope formationand terminal differentiation. Cell, 40: 685-695, 1985.

Cell Growth & Differentiation 1241
(a)pyrene, 1 2-Q-tetradecanoylphorbol-1 3-acetate and hydroxyurea.Carcinogenesis (Lond.), 9: 1885-1 890, 1988.
10. Rubin, A. L., Parenteau, N. L., and Rice, R. H. Coordination of kerati-nocyte programming in human 5CC-i 3 squamous carcinoma and normalepidermal cells. J. Cell. Physiol., 138: 208-214, 1989.
1 1 . Takahashi, H., and lizuka, H. Analysis of the 5-upstream promoterregion of human involucrin gene: activation by 1 2-O-tetradecanoylphorbol-1 3-acetate. J. Invest. Dermatol., 100: 1 0-1 5, 1993.
1 2. Filvaroff, E., Stern, D. F., and Dotto, G. P. Tyrosine phosphorylation is anearly and specific event involved in primary keratinocyte differentiation.Mol. Cell. Biol., 10: 1 164-1 173, 1990.
1 3. Seimiya, H., and Tsuruo, T. Differential expression of protein tyrosinephosphatase genes during phorbol ester-induced differentiation of humanleukemia U937 cells. Cell Growth & Differ., 4: 1033-1039, 1993.
14. Denning, M. F., Dlugosz, A. A., Howett, M. K., and Yuspa, S. H.Expression of an oncogenic rasHa gene in murine keratinocytes inducestyrosine phosphorylation and reduced activity of protein kinase C 8. J. Biol.Chem., 268:26079-26081, 1993.
1 5. Fischer, E. H., Charbonneau, H., and Tonks, N. K. Protein tyrosinephosphatases: a diverse family of intracellular and transmembrane enzymes.Science (Washington DC), 253: 401-406, 1991.
1 6. Hunter, T. Cytokine connections. Nature (Lond.), 366: 1 14-1 16, 1993.
17. Kachinskas, D. J., Phillips, M. A., Qin, Q., and Rice, R. H. Arsenatesuppression of involucrin expression in malignant human keratinocytes.FASEBJ., 7:A1230, 1993.
1 8. Rice, R. H., Steinmann, K. E., de Graffenried, L. A., Qin, Q., Taylor, N.,
and Schlegel, R. Elevation of cell cycle control proteins during spontaneousimmortalization of human keratinocytes. Mol. Biol. Cell, 4: 185-1 94, 1993.
1 9. Rheinwald, J. G., Germain, E., and Beckett, M. A. Expression of enve-lope and keratin proteins in normal and malignant human keratinocytes andmesothelial cells. ln:C. Harris and H. Autrup (eds.), Human Carcinogenesis,pp. 85-96. New York: Academic Press, 1983.
20. Gordon, J. A. Use of vanadate as protein-phosphotyrosine phosphataseinhibitor. Methods Enzymol. 201: 477-482, 1991.
21 . Carroll, J. M., and Taichman, L. B. Characterization of the humaninvolucrin promoter using a transient j3-galactosidase assay. J. Cell Sci., 103:
925-930, 1992.
22. FilIat, C., Rodriguez-Gil, J. E., and Guinovart, J. J. Molybdate and tung-state act like vanadate on glucose metabolism in isolated hepatocytes.Biochem.J., 282:659-663, 1992.
23. Hutchison, K. A., Stancato, L. F., Jove, R., and Pratt, W. B. The protein-protein complex between pp60v-src and hsp90 is stabilized by molybdate,vanadate, tungstate and an endogenous cytosolic metal. J. Biol. Chem., 267:
13952-1 3957, 1992.
24. Saha, A. K., Crans, D. C., Pope, M. T., Simone, C. M., and Glew, R. H.Inhibition of human seminal fluid and Leishmania donovani phosphatases bymolybdate heteropolyanions. J. Biol. Chem., 266: 351 1-351 7, 1991.
25. Swarup, G., Cohen, S., and Garbers, D. L. Inhibition of membranephosphotyrosyl-protein phosphatase activity by vanadate. Biochem.Biophys. Res. Commun., 107: 1 1 04-1 109, 1982.
26. Garcia-Morales, P., Minami, Y., Luong, E., Klausner, R. D., and Samel-son, L. E. Tyrosine phosphorylation in T cells is regulated by phosphataseactivity: studies with phenylarsine oxide. Proc. Nail Acad. Sci. USA, 87:9255-9259, 1990.
27. Igarishi, K-I., David, M., Lamer, A. C., and Finbboom, D. S. In vitro
activation of a transcription factor by y-interferon requires a membrane-associated tyrosine kinase and is mimicked by vanadate. Mol. Cell. Biol., 13:3984-3989, 1993.
28. Leask, A., Byrne, C., and Fuchs, E. Transcription factor AP2 and its rolein epidermal-specific gene expression. Proc. NatI. Acad. Sci. USA, 88:7948-7952, 1991.
29. Shannon, R. L., and Strayer, D. S. Arsenic-induced skin toxicity. Hum.Toxicol., 8: 99-104, 1989.
30. WHO. In: IARC Monographs on the Evaluation of the Carcinogenic Riskof Chemicals to Humans: Some Metal and Metallic Compounds, pp. 101-1 1 4. Lyon, France: International Agency for Research on Cancer, 1980.
31 . Wu, M. M., Kuo, T. L., Hwang, Y., and Chen, C. J. Dose-responserelation between arsenic concentration in well water and mortality fromcancers and vascular diseases. Am. J. Epidemiol., 130: 1 123-1 1 32, 1989.
32. Brown, K. G., Boyle, K. E., Chen, C. W., and Gibb, H. J. A dose-responseanalysis of skin cancer from inorganic arsenic in drinking water. Risk Anal.,9:519-528, 1989.
33. Smith, A. H., Hopenhayn-Rich, C., Bates, M. N., Goeden, H. M., Hertz-Picciotto, I., Duggan, H. M., Wood, R., Kosnett, M. J., and Smith, M. Cancer
risks from arsenic in drinking water. Environ. Health Perspect., 97: 259-267,
1992.
34. Jacobson-Kram, D., and Montalbano, D. The reproductive effects as-sessment group’s report on the mutagenicity of inorganic arsenic. Environ.Mutagen., 7: 787-804, 1985.
35. Li, J-H., and Rossman, T. G. Inhibition of DNA ligase activity by arsenite:a possible mechanism of its comutagenesis. Mol. Toxicol., 2: 1-9, 1989.
36. Aposhian, H. V. Biochemical toxicology of arsenic. Rev. Biochem.Toxicol., 10:265-299, 1989.
37. Lopez, S., Miyashita, Y., and Simons, S. S., Jr. Structurally based, selec-
tive interaction of arsenite with steroid receptors. J. Biol. Chem., 265:
16039-1 6042, 1990.
38. Deaton, M. A., Bowman, P. D., Jones, G. P., and Powanda, M. C. Stressprotein synthesis in human keratinocytes treated with sodium arsenite, phe-nyldichloroarsine and nitrogen mustard. Fundam. AppI. Toxicol., 14:
471-476, 1990.
39. Chin, K-V., Tanaka, S., Darlington, G., Pastan, I., and Gottesman, M. M.Heat shock and arsenite increase expression of the multidrug resistance(MDRI) gene in human renal carcinoma cells. J. Biol. Chem., 265: 221-226,
1990.
40. Lee, T. C., Oshimura M., and Barrett, J. C. Comparison of arsenic-induced cell transformation, cytotoxicity, mutation and cytogenetic effects inSyrian hamster embryo cells in culture. Carcinogenesis (Lond.), 6:
1421-1426, 1985.
41 . Bertolero, F., Pozzi, G., Sabbioni, E., and Saffioti, U. Cellular uptake andmetabolic reduction of pentavalent to trivalent arsenic as determinants ofcytotoxicity and morphological transformation. Carcinogenesis (Lond.), 8:
803-808, 1987.
42. Lee, T. C., Tanaka, N., Lamb, P. W., Gilmer, T. M., and Barrett, J. C.Induction of gene amplification by arsenic. Science (Washington DC), 241:
79-81, 1988.
43, Wright, J. A., Smith, H. S., Watt, F. M., Hancock, M. C., Hudson, D. L.,and Stark, G. R. DNA amplification is rare in normal cells. Proc. NatI. Acad.Sci. USA, 87: 1791-1795, 1990.
44. Hanlon, D. P., and Ferm, V. H. Concentration and chemical status of
arsenic in the blood of pregnant hamsters during critical embryogenesis.Environ. Res., 40: 372-379, 1986.
45. Westheimer, F. H. Why nature chose phosphates. Science (WashingtonDC), 235: 1 1 73-1 1 78, 1987.
46. Rheinwald, J. G., and Beckett, M. A. Tumorigenic keratinocyte linesrequiring anchorage and fibroblast support cultured from human squamous
carcinomas. Cancer Res., 41: 1657-1663, 1981.
47. Allen-Hoffman, B. L., and Rheinwald, J. G. Polycyclic aromatic hydro-
carbon mutagenesis of human epidermal cells in culture. Proc. NatI. Acad.Sci. USA, 81: 7802-7806, 1984.
48. Parenteau, N. L., Eckert, R. L., and Rice, R. H. Primate involucrins:antigenic relatedness and detection of multiple forms. Proc. NatI. Acad. Sci.
USA, 84:7571-7575, 1987.
49. Chirgwin, I. M., Przybyla, A. E., MacDonald, R. J., and Rutter, W. J.Isolation of biologically active ribonucleic acid from sources enriched inribonuclease. Biochemistry, 18: 5294-5299, 1979.
50. Fourney, R. M., Miyakoshi, J., Day, R. S., Ill, and Paterson, M. C.Northern blotting: efficient RNA staining and transfer. Focus (Idaho), 10:7-10, 1988.
51 . Zandomeni, R., Mittleman, B., Bunick, D., Ackerman, S., and Wein-mann, R. Mechanism of action of dichIoro-�-D-ribofuranosylbenzimidazole:effect on in vitro transcription. Proc. NatI. Acad. Sci. USA, 79: 3167-31 70,1982.
52. Greenburg, M. E., and Ziff, E. B. Stimulation of 3T3 cells inducestranscription of the c-los protooncogene. Nature (Lond.), 31 1: 433-438,
1984.
53. Celano, M. B., Berchtold, C., and Casero, R. A., Jr. A simplification of thenuclear run-off transcription assay. Biotechniques, 7: 942-944, 1989.
54. Phillips, M. A., Stewart, B. E., Qin, Q., Chakravarty, R., Floyd, E. E.,Jetten, A. M., and Rice, R. H. Primary structure of keratinocyte transglutami-nase. Proc. NatI. Acad. Sci USA, 87:9333-9337, 1990.
55. Arcari, P., Martinelli, R., and Salvatore, F. The complete sequence of afull length cDNA for human liver glyceraldehyde-3-phosphate dehydroge-nase: evidence for multiple mRNA species. Nucleic Acids Res., 12:9179-9189, 1984.
56. Chodosh, L. A. Mobility shift DNA-binding assay using gel electrophore-sis. In: F. M. Ausubel, R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman,J. A. Smith, and K. Struhl teds.), Current Protocols in Molecular Biology,Suppl. 13, pp. 12.2.1-12.2.10. New York: Current Protocols, 1993.





























