Embed Size (px)
Citation preview

Ann. soc. entomol. Fr. (n.s.), 2011, 47 (1) : xxx-xxx
1
ARTICLE
Antennal sensilla in fi ve Psychodini moth fl ies (Diptera: Psychodidae: Psychodinae) pollinators of Arum spp. (Araceae)
Abstract. The pollination of the genus Arum (Araceae) is mainly achieved by deception, the fl oral odour mimicking the pollinator ovipositing site. In order to discover the sensory organs involved in this attraction, we have studied the antennae of fi ve species of psychodine moth-fl ies (former Psychoda sensu lato = Psychodini), pollinators of Arum spp. The antennae of the fi ve Psychodini reveal seven types of sensilla: multiporous tribranched sensilla basiconica (sensilla ascoidea), multiporous sensilla basiconica, multiporous sensilla coeloconica, multiporous sensilla auricillica, uniporous sensilla basiconica, aporous sensilla chaetica, aporous Böhm’s sensilla. Each species possesses three, fi ve or six of these sensillum types. All the multiporous sensilla are probably olfactory receptors while the uniporous sensilla basiconica must possess a contact chemoreceptive function. The multiporous tribranched sensilla basiconica (s. ascoidea), present in all the species, are the best candidates for the reception of the odours given off by the ovipositing sites and the infl orescences of Arum. The multiporous sensilla basiconica and the multiporous sensilla coeloconica may be involved respectively as CO2 receptors or thermoreceptors. Psychoda phalaenoides, which is the main pollinator of A. maculatum, is the species which possesses the largest number of antennal sensilla. The sexual dimorphism, studied only in Psycha grisescens, as concerns the number of sensilla and the absence of a sensillum type which differ according to sex, is diffi cult to interpret.
Résumé. Sensilles antennaires de cinq espèces de Psychodini (Diptera : Psychodidae : Psychodinae) pollinisateurs du genre Arum (Araceae). La pollinisation du genre Arum (Araceae) est réalisée essentiellement par tromperie, l’odeur fl orale imitant les sites d’oviposition du pollinisateur. Afi n de rechercher les organes sensoriels impliqués lors de cette attraction, noue avons étudié les antennes de quelques espèces du genre Psychoda sensu lato, pollinisateurs des Arum. Les antennes des cinq Psychodini montrent l’existence de sept types sensillaires : des sensilles basiconiques multipores à trois branches (sensilles ascoïdes), des sensilles basiconiques multipores, des sensilles coeloconiques multipores, des sensilles auricilliformes multipores, des sensilles basiconiques à pore terminal, des sensilles chétiformes sans pore et des sensilles de Böhm sans pore. Chaque espèce possède trois, cinq ou six de ces types sensillaires. Toutes les sensilles multipores sont probablement des récepteurs olfactifs tandis que les sensilles basiconiques unipores possèdent une fonction chimio-réceptrice de contact. Les sensilles ascoïdes présentes en grands nombres chez toutes les espèces sont les meilleures candidates pour la réception des odeurs en provenance des sites d’oviposition et des infl orescences d’Arum. Les sensilles multipores basiconiques et coeloconiques peuvent être impliquées respectivement dans la réception du CO2 ou comme thermorécepteurs. Psychoda phalaenoides, principal pollinisateur de A. maculatum, est l’espèce la mieux dotée en sensilles antennaires. Le dimorphisme sexuel, étudié chez Psycha grisescens, portant sur le nombre de sensilles et l’absence d’un type sensoriel différent selon le sexe, reste diffi cile à interpréter.Keywords: Psychoda, Psycha, Logima, Chodopsycha, deceptive pollination.
Michel J. Faucheux (1) & Marc Gibernau (2)
(1) Université de Nantes, Laboratoire d’Endocrinologie des Insectes Sociaux, , 2 rue de la Houssinière, B.P. 92208, F-44322 Nantes Cedex 03, France, et Muséum d’Histoire naturelle de Nantes, 12 rue Voltaire, F-44000 Nantes, France
(2) CNRS – Ecofog UMR 8172, BP 709, F-97387 Kourou, France
E-mail: [email protected], [email protected]é le 31 mai 2010
The pollination of the genus Arum (Araceae) is mainly achieved by deception, the fl oral odour
mimicking the pollinator ovipositing site (Gibernau et al. 2004). Th e pollinators, mainly fl ies, and beetles in a few cases are temporarily trap during a complex pollination process (Gibernau et al. 2004) insects. At anthesis, the appendix of the infl orescence produces
heat and emits a specifi c odour, which attracts the insects. Th e lured insects are trapped at the base of the infl orescence within the fl oral chamber, also called kettle-trap, when stigmas are receptive (Lack & Diaz 1991, Vogel & Martens 2000). If they carry pollen grains, they achieve pollination. Th e insects will be released about 24h later after pollen emission, ensuring pollen dissemination.
In Western Europe, the two common Arum spe-cies, Arum maculatum L., and Arum italicum Mill., are mainly pollinated by non-biting moth fl ies from the
2
M. J. Faucheux & M. Gibernau
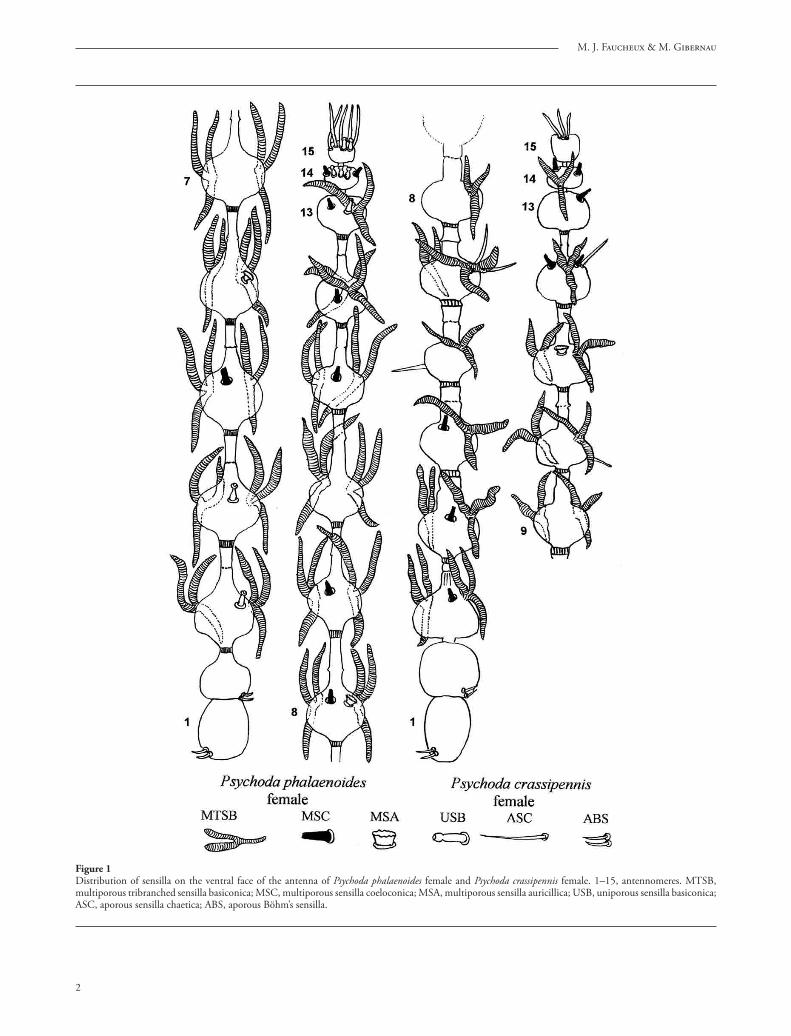
Figure 1Distribution of sensilla on the ventral face of the antenna of Psychoda phalaenoides female and Psychoda crassipennis female. 1–15, antennomeres. MTSB, multiporous tribranched sensilla basiconica; MSC, multiporous sensilla coeloconica; MSA, multiporous sensilla auricillica; USB, uniporous sensilla basiconica; ASC, aporous sensilla chaetica; ABS, aporous Böhm’s sensilla.

Antennal sensilla of Psychodini
3
genus Psychoda (Lack & Diaz 1991, Diaz & Kite 2002, Albre et al. 2003, Gibernau et al. 2004a). Th ese two species appear to have diff erent specifi city degrees for their pollinators. Up to now, A. maculatum has been shown to be almost exclusively pollinated by females of Psychoda phalaenoides (L. 1758), even if some other insects have been found in the infl orescences (Prime 1960). For example, up to 56 insect species of which 94% were psychodid midges were trapped in infl o-rescences of A. maculatum in Germany (Beck 1983, Rohacek et al. 1990). In the German populations, Psy-choda phalaenoides is the main pollinator (93.5%) but another Psychodid, Psycha grisescens (Tonnoir 1922), has also been found and may represent up to half of the Psychodidae in some populations (Beck 1983). On the contrary, the insects diversity found in the infl o-rescences of A. italicum fl uctuates greatly between sites (Gibernau et al. 2004a). Th e trapped insects can vary greatly according to the population from Spain up to England (Arcangeli 1883, Withers 1988, Méndez & Obeso 1992, Diaz & Kite 2002, Albre et al. 2003), but psychodid moth fl ies (e.g. mainly females) are general-ly among the most abundant insects representing up to 87% of the trapped insects. In two Arum populations from South of France, several Psychodidae species were found but in general one species dominate in a given population; P. crassipennis Tonnoir 1940 was the most abundant (41%) in one population and Apsycha pusilla (Tonnoir 1922) (30%) in the other (Albre et al. 2003). P. crassipennis has also been considered as a major pol-linator of A. italicum in England (Withers 1988).
In order to discover the sensory organs involved in the attraction of the insects by the infl orescences of Arum, we have chosen to study the antennae of the females of fi ve species of Diptera Psychodidae, members of the tribe of Psychodini. Th ese species were collected in infl orescences of A. maculatum (Psychoda phalaenoides and Psycha grisescens,) or A. italicum (Psychoda crassipennis, Logima sigma and Chodopsycha sp.) and are thus potential Arum pollinators.
Th e sensilla of Psychodids have been rarely studied. Th e distribution and the length of “ascoid sensilla” on the antennal segments have considerable taxonomic importance in mothfl ies (Psychodidae Psychodinae) and sandfl ies (Psychodidae Phlebotominae), which explains why they are fairly well known. Th eir structure and function, however, have only been dealt in the Phlebotominae Lutzomyia longipalpis (Lutz & Neiva 1912) for its economic signifi cance (Dougherty & Hamilton 1997, Spiegel et al. 2005a). As for the other sensilla which may be found on the antennae, they have only been mentioned among the Phlebotominae, and always for the same reasons. Apparently, apart from the ascoids, no other sensillum has been described on the antennae of Psychodinae.
Material and methodsTh e fi ve species studied belong to the former genus Psychoda Latreille 1796 but now to four closely related genera (Ježek 1983, 1990): Psycha grisescens (Tonnoir 1922) (3 males, 9 females), Psychoda phalaenoides (L. 1758) (9 females), Psychoda crassipennis Tonnoir 1940 (4 females), Logima sigma (Kincaid 1899) (3 females), and Chodopsycha sp. (3 females). For the study with scanning electron microscopy (SEM), the antennae were dehydrated in absolute ethanol, mounted on specimen holders and coated with a thin layer of gold and palladium in a JFC 1100 sputter coater. Preparations were examined in a Jeol JSM 6400 SEM at diff erent magnifi cations. Th e terminologies of Schneider (1964), Zacharuk (1985) and Faucheux (1999) are used in naming the types of sensilla. Th e mean number of each sensillum type was calculated from the counts on each antennomere with SEM, and expressed as mean +/- SD.
ResultsTh e antennae are moniliform and their length varies
from 750 μm to 850 μm. Each antenna comprises the scape (antennomere 1), the pedicel (antennomere 2) and the fl agellum composed of 12 to 14 fl agellomeres in the species studied; the fi rst fl agellomere is therefore the third antennomere.
Th e diagnosis of the antennae corresponds to the descriptions of Ježek (1990), Ježek & Harten (2005).
Psychoda phalaenoides possesses 15-segmented antennae (15 antennomeres), and are haired as well; the scape is a little longer than its maximum width, and narrowed at the base; the pedicel is almost globular, and the fl agellomeres pitcher-shaped (fi g. 1). Th e 13th, 14th and 15th segments are separated by a short narrowed part; the last two antennal segments are small, of the same size, but smaller than the foregoing segments (fi g. 5). Th e antennae of Psychoda crassipenis resembles to that of Psychoda phalaenoides (fi g. 1).
Psycha grisescens have 16-segmented antennae (16 antennomeres), with a cylindrical scape, a pedicel that is almost globular, pitcher-shaped fl agellomères; antennomeres 14 to 16 are rather minute in contrast to the foregoing antennomeres; the 13th and the 14th are fused, the 15th and 16th separated, the 16th been pyriform. Th e segments 13 and 14 are with (male) or without (female) a subapical spine (fi g. 2).
Logima sigma has 14-segmented antennae, the fourteenth segment being vestigial without scales or microtrichia, the 12th with a neck, and a swelling between the 13th and 14th (fi g. 3).
Th e antennae of Chodopsycha sp. are 16-segmented, antennomeres 13 to 16 form a contrast to the foregoing antennomeres, the 13th to 15th are fused, the 15th and 16th separated, and the 16th globular (fi g. 3).
Typically, each fl agellomere comprises a basal bulb (node) and a neck (internode). Th e neck is often
4
M. J. Faucheux & M. Gibernau
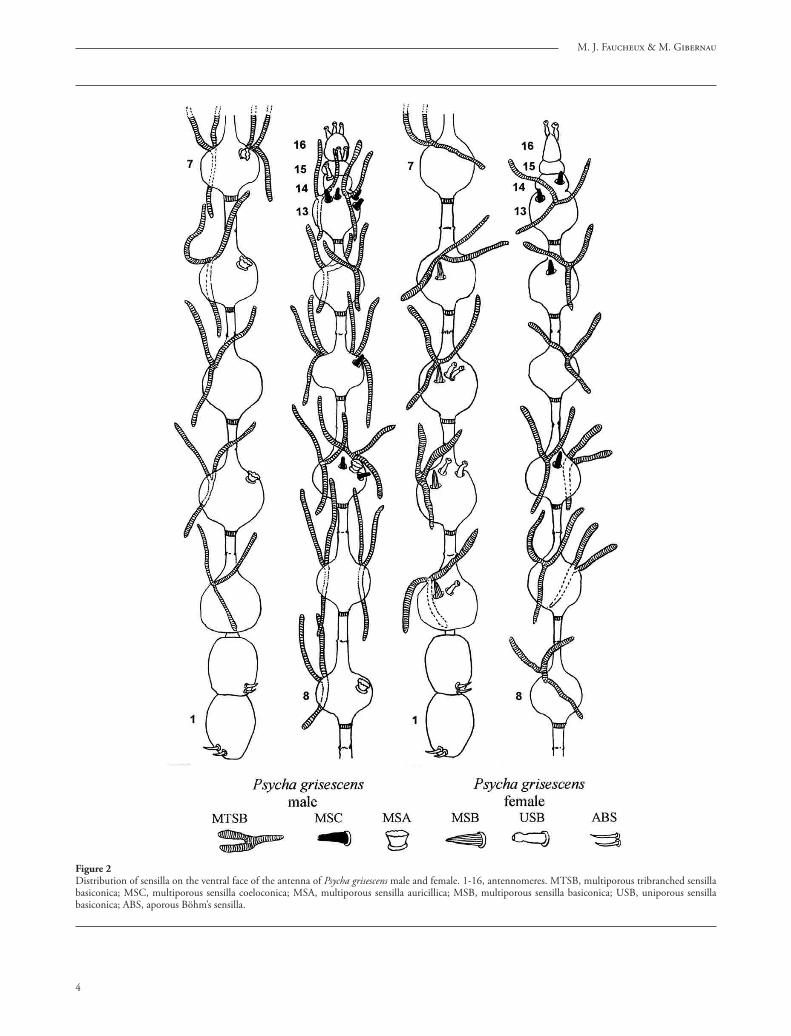
Figure 2Distribution of sensilla on the ventral face of the antenna of Psycha grisescens male and female. 1-16, antennomeres. MTSB, multiporous tribranched sensilla basiconica; MSC, multiporous sensilla coeloconica; MSA, multiporous sensilla auricillica; MSB, multiporous sensilla basiconica; USB, uniporous sensilla basiconica; ABS, aporous Böhm’s sensilla.

Antennal sensilla of Psychodini
5
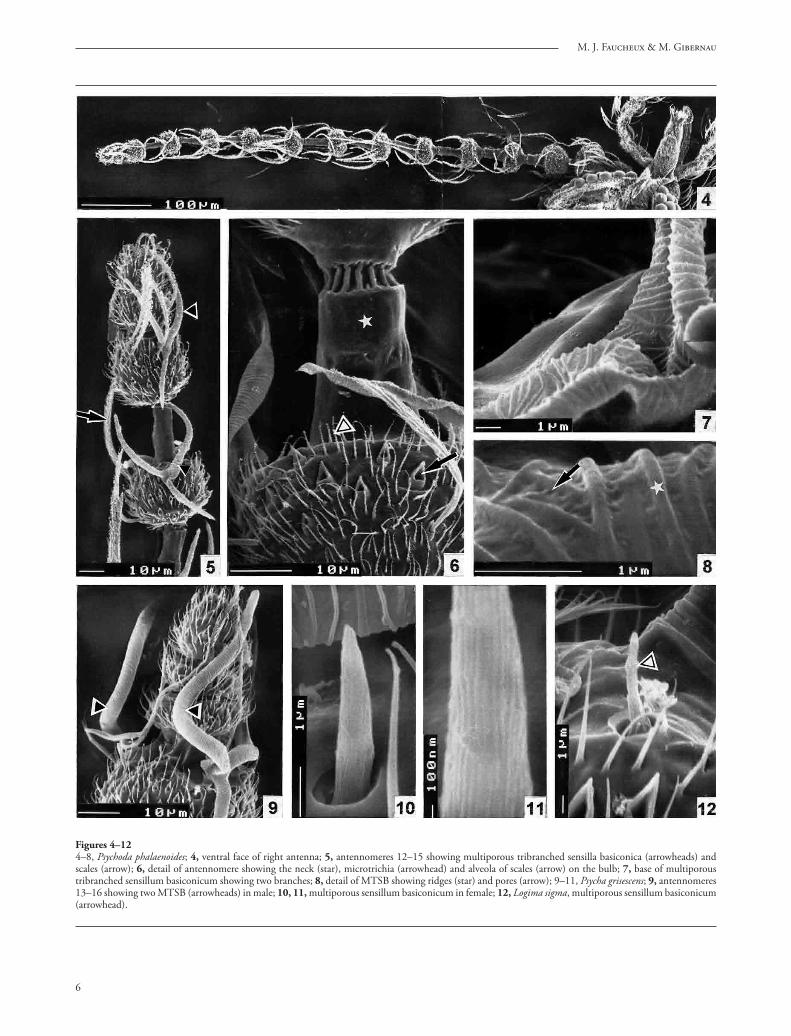
Figure 3Distribution of sensilla on the ventral face of the antenna of Logima sigma female and Chodopsycha sp. female. 1–14 or 1–16, antennomeres. MTSB, multiporous tribranched sensilla basiconica; MSC, multiporous sensilla coeloconica; MSA, multiporous sensilla auricillica; MSB, multiporous sensilla basiconica; USB, uniporous sensilla basiconica; ABS, aporous Böhm’s sensilla.
6
M. J. Faucheux & M. Gibernau
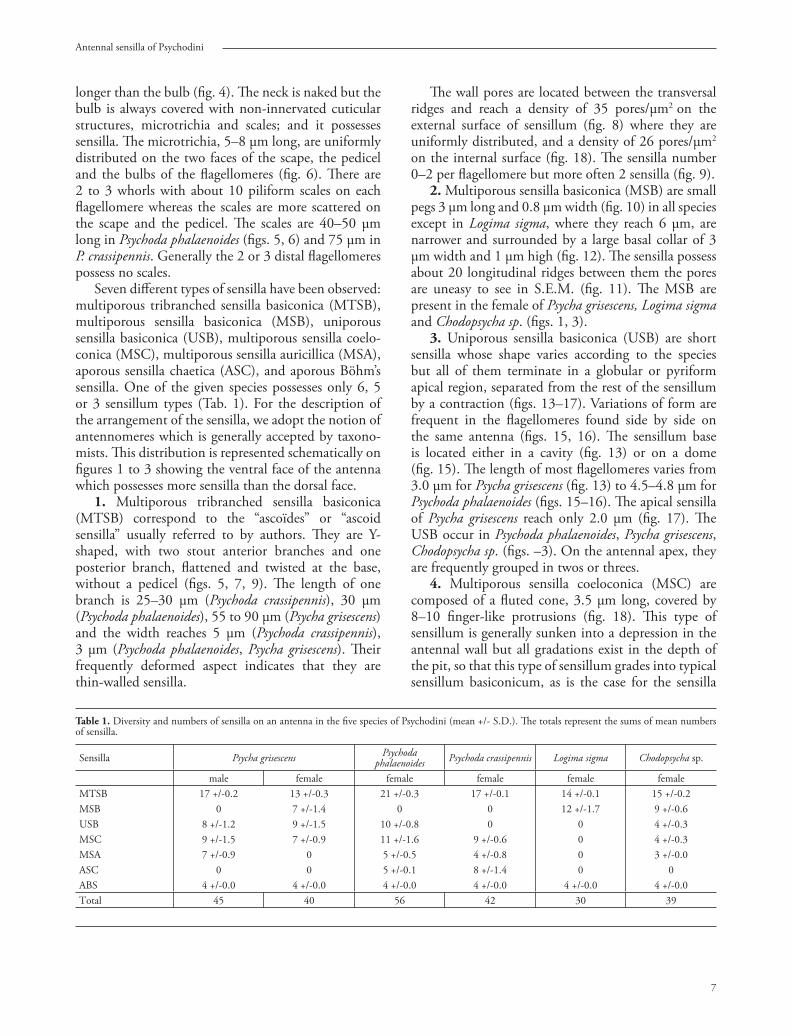
Figures 4–124–8, Psychoda phalaenoides; 4, ventral face of right antenna; 5, antennomeres 12–15 showing multiporous tribranched sensilla basiconica (arrowheads) and scales (arrow); 6, detail of antennomere showing the neck (star), microtrichia (arrowhead) and alveola of scales (arrow) on the bulb; 7, base of multiporous tribranched sensillum basiconicum showing two branches; 8, detail of MTSB showing ridges (star) and pores (arrow); 9–11, Psycha grisescens; 9, antennomeres 13–16 showing two MTSB (arrowheads) in male; 10, 11, multiporous sensillum basiconicum in female; 12, Logima sigma, multiporous sensillum basiconicum (arrowhead).
Antennal sensilla of Psychodini
7
longer than the bulb (fi g. 4). Th e neck is naked but the bulb is always covered with non-innervated cuticular structures, microtrichia and scales; and it possesses sensilla. Th e microtrichia, 5–8 μm long, are uniformly distributed on the two faces of the scape, the pedicel and the bulbs of the fl agellomeres (fi g. 6). Th ere are 2 to 3 whorls with about 10 piliform scales on each fl agellomere whereas the scales are more scattered on the scape and the pedicel. Th e scales are 40–50 μm long in Psychoda phalaenoides (fi gs. 5, 6) and 75 μm in P. crassipennis. Generally the 2 or 3 distal fl agellomeres possess no scales.
Seven diff erent types of sensilla have been observed: multiporous tribranched sensilla basiconica (MTSB), multiporous sensilla basiconica (MSB), uniporous sensilla basiconica (USB), multiporous sensilla coelo-conica (MSC), multiporous sensilla auricillica (MSA), aporous sensilla chaetica (ASC), and aporous Böhm’s sensilla. One of the given species possesses only 6, 5 or 3 sensillum types (Tab. 1). For the description of the arrangement of the sensilla, we adopt the notion of antennomeres which is generally accepted by taxono-mists. Th is distribution is represented schematically on fi gures 1 to 3 showing the ventral face of the antenna which possesses more sensilla than the dorsal face.
1. Multiporous tribranched sensilla basiconica (MTSB) correspond to the “ascoïdes” or “ascoid sensilla” usually referred to by authors. Th ey are Y-shaped, with two stout anterior branches and one posterior branch, fl attened and twisted at the base, without a pedicel (fi gs. 5, 7, 9). Th e length of one branch is 25–30 μm (Psychoda crassipennis), 30 μm (Psychoda phalaenoides), 55 to 90 μm (Psycha grisescens) and the width reaches 5 μm (Psychoda crassipennis), 3 μm (Psychoda phalaenoides, Psycha grisescens). Th eir frequently deformed aspect indicates that they are thin-walled sensilla.
Th e wall pores are located between the transversal ridges and reach a density of 35 pores/μm2 on the external surface of sensillum (fi g. 8) where they are uniformly distributed, and a density of 26 pores/μm2 on the internal surface (fi g. 18). Th e sensilla number 0–2 per fl agellomere but more often 2 sensilla (fi g. 9).
2. Multiporous sensilla basiconica (MSB) are small pegs 3 μm long and 0.8 μm width (fi g. 10) in all species except in Logima sigma, where they reach 6 μm, are narrower and surrounded by a large basal collar of 3 μm width and 1 μm high (fi g. 12). Th e sensilla possess about 20 longitudinal ridges between them the pores are uneasy to see in S.E.M. (fi g. 11). Th e MSB are present in the female of Psycha grisescens, Logima sigma and Chodopsycha sp. (fi gs. 1, 3).
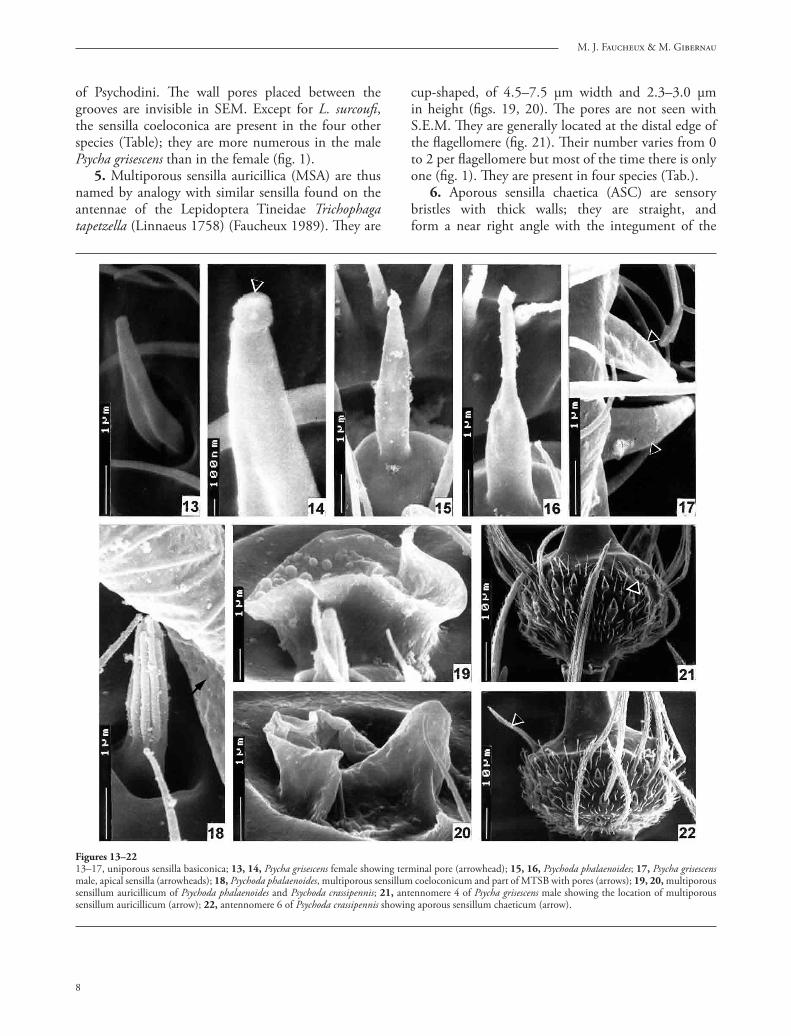
3. Uniporous sensilla basiconica (USB) are short sensilla whose shape varies according to the species but all of them terminate in a globular or pyriform apical region, separated from the rest of the sensillum by a contraction (fi gs. 13–17). Variations of form are frequent in the fl agellomeres found side by side on the same antenna (fi gs. 15, 16). Th e sensillum base is located either in a cavity (fi g. 13) or on a dome (fi g. 15). Th e length of most fl agellomeres varies from 3.0 μm for Psycha grisescens (fi g. 13) to 4.5–4.8 μm for Psychoda phalaenoides (fi gs. 15–16). Th e apical sensilla of Psycha grisescens reach only 2.0 μm (fi g. 17). Th e USB occur in Psychoda phalaenoides, Psycha grisescens, Chodopsycha sp. (fi gs. –3). On the antennal apex, they are frequently grouped in twos or threes.
4. Multiporous sensilla coeloconica (MSC) are composed of a fl uted cone, 3.5 μm long, covered by 8–10 fi nger-like protrusions (fi g. 18). Th is type of sensillum is generally sunken into a depression in the antennal wall but all gradations exist in the depth of the pit, so that this type of sensillum grades into typical sensillum basiconicum, as is the case for the sensilla
Table 1. Diversity and numbers of sensilla on an antenna in the fi ve species of Psychodini (mean +/- S.D.). Th e totals represent the sums of mean numbers of sensilla.
Sensilla Psycha grisescens Psychoda phalaenoides Psychoda crassipennis Logima sigma Chodopsycha sp.
male female female female female femaleMTSB 17 +/-0.2 13 +/-0.3 21 +/-0.3 17 +/-0.1 14 +/-0.1 15 +/-0.2MSB 0 7 +/-1.4 0 0 12 +/-1.7 9 +/-0.6USB 8 +/-1.2 9 +/-1.5 10 +/-0.8 0 0 4 +/-0.3MSC 9 +/-1.5 7 +/-0.9 11 +/-1.6 9 +/-0.6 0 4 +/-0.3MSA 7 +/-0.9 0 5 +/-0.5 4 +/-0.8 0 3 +/-0.0ASC 0 0 5 +/-0.1 8 +/-1.4 0 0ABS 4 +/-0.0 4 +/-0.0 4 +/-0.0 4 +/-0.0 4 +/-0.0 4 +/-0.0Total 45 40 56 42 30 39
8
M. J. Faucheux & M. Gibernau
of Psychodini. Th e wall pores placed between the grooves are invisible in SEM. Except for L. surcoufi , the sensilla coeloconica are present in the four other species (Table); they are more numerous in the male Psycha grisescens than in the female (fi g. 1).
5. Multiporous sensilla auricillica (MSA) are thus named by analogy with similar sensilla found on the antennae of the Lepidoptera Tineidae Trichophaga tapetzella (Linnaeus 1758) (Faucheux 1989). Th ey are
cup-shaped, of 4.5–7.5 μm width and 2.3–3.0 μm in height (fi gs. 19, 20). Th e pores are not seen with S.E.M. Th ey are generally located at the distal edge of the fl agellomere (fi g. 21). Th eir number varies from 0 to 2 per fl agellomere but most of the time there is only one (fi g. 1). Th ey are present in four species (Tab.).
6. Aporous sensilla chaetica (ASC) are sensory bristles with thick walls; they are straight, and form a near right angle with the integument of the
Figures 13–2213–17, uniporous sensilla basiconica; 13, 14, Psycha grisescens female showing terminal pore (arrowhead); 15, 16, Psychoda phalaenoides; 17, Psycha grisescens male, apical sensilla (arrowheads); 18, Psychoda phalaenoides, multiporous sensillum coeloconicum and part of MTSB with pores (arrows); 19, 20, multiporous sensillum auricillicum of Psychoda phalaenoides and Psychoda crassipennis; 21, antennomere 4 of Psycha grisescens male showing the location of multiporous sensillum auricillicum (arrow); 22, antennomere 6 of Psychoda crassipennis showing aporous sensillum chaeticum (arrow).

Antennal sensilla of Psychodini
9
fl agellomere (fi g. 22). Th ey possess a fl exible circular membrane at the base. Th ey have no pore at the tip. Sensilla chaetica are found only in Psychoda crassipennis on antennomeres 6, 7, 10, 12 and the apex of 15, and in Psychoda phalaenoides at the apex of the15th antennomere (fi gs. 2, 23).
7. Aporous Böhm’s sensilla, 6–9 μm long are grouped by two and located at the base of the scape and of the pedicel (fi gs. 1, 2, 3, 24–26).
DiscussionTh e number of sensillum types in Psychodini is fairly
similar to that of the Phlebotominae. For example, fi ve types (trichodeum, basiconicum, auricillicum, coeloconicum, chaeticum = ascoid) are described in Phlebotomus argentipes Annandale & Brunetti 1908 (Ilango 2000).
Morphological and functional considerationsTh e multiporous tri-branched sensilla basiconica
(MTSB) correspond to the “fi lets sensoriels” of Philosepedon humerale (Meigen 1818) which are also tri-branched (Séguy 1925), “sensory fi laments” (Ježek 1990). Like the latter, they are to be found by two arranged symetrically on the fl agellomere; they are also known as “Tonnoir organs” (Séguy 1951) in reference to Tonnoir (1919) which was the fi rst to mention them. Th ese sensory fi laments, also known as “épines géniculées courtes” are reduced to a single branch but retain the same arrangement on the fl agellomeres de Phlebotomus papatasi Scopoli 1786 (Séguy 1925, 1951). Th e MTSB ressemble the multiporous multibranched sensilla basiconica or “sensilla ascoidea” of Micropteryx
calthella (Linnaeus 1761), Lepidoptera Micropterigidae in which the number of branches varies from one to 12, the fi rst fl agellomeres possessing ascoids with only one or two branches (Faucheux 1997).
In none of the studied species have we observed the small pedicel sketched by Ježek (1990) for the male antenna of Psychoda phalaenoides (Fig. 119, p. 62) no more than the long pedicel shown on the male antenna of Psycha grisescens (Fig. 43, p. 45) at the base of the MTSB. We have not observed variations in the length of the ascoids in the same species, or any sexual dimorphism concerning the number of branches.
On the basis of morphological similarities with sensilla of known function in other insects such as in the subfamily of Phlebotominae (Dougherty & Hamilton 1997, Dougherty et al. 1999), the MTSB (ascoids) of Psychodini are probably olfactory receptors. In Lutzomyia longipalpis, adult males release the following compound, (1 S, 3 S, 7 R)-3-methyl-α-himachalene from their tergal glands to lure females to the same site for mating and feeding. Th e authors have not obtained electrophysiological recordings from ascoid sensillum receptor cells in antennae of male and female sandfl ies. Receptor cells of both sexes responded only to the main component of the male tergal gland extract (Spiegel et al. 2005a). Both males and females fl ew upwind in a wind tunnel towards a fi lter paper disk treated with either 4-6 male equivalents of the tergal gland extract, pure (1 S, 3 S, 7 R)-3-methyl-α-himachalene or the synthetic mixture of eight isomers. Th is indicates that the compound derived from Lutzomyia longipalpis males has a dual function in causing male aggregation as well as serving as a sex pheromone for females.
Figures 23–26Psychoda phalaenoides. 23, apical aporous sensilla chaetica (arrowheads); 24, aporous Böhm’s sensilla at the base of scape (arrowhead); 25, detail; 26, Böhm’s sensilla of pedicel (arrowheads).

10
M. J. Faucheux & M. Gibernau
Furthermore, the females of Lutzomyia longipalpis produce an oviposition pheromone (the dodecanoic acid) in the accessory glands that is secreted onto the eggs during oviposition. Th is compound attracts and/or stimulates gravid females for egg-laying. When single sensillum recordings were taken from the “ascoid” on the antennae of female sandfl ies, the response to egg-extract and dodecanoic acid was similar. Th ere was a dose dependent response to dodecanoic acid and preferential sensitivity compared to other fatty acids (Dougherty & Hamilton 1997; Dougherty et al. 1999). If the case is the same in the Psychodini, the absence of sexual dimorphism concerning the MTSB in Psycha grisescens would not be surprising.
In all the species of psychodids, the MTSB are the most frequently represented sensillum type. Th us, because of its frequence and thanks to the surface of the sensilla, it is the best adapted to the reception of olfactive stimuli. Of the 5 females studied, Psychoda phalaenoides is the species which off ers the largest total number of sensilla, and of MTSB. Th is can explain why A. maculatum is pollinated essentially by Psychoda phalaenoides and secondly by Psycha grisescens.
Th e sexual dimorphism, studied only in Psycha grisescens, from a numerical point of view, is based entirely on the slightly large number of MTSB in the male. If these sensilla are the receptors of a sexual pheromone (a hypothesis in the Psychodini), the diff erence is justifi ed. As it is well known for females of Lepidoptera, the pheromone receptors are present but are much less sensitive than those of the males (Van der Pers & Den Otter 1978). Indeed, the chemical composition of the pheromones of the Psychodini is not known but their existence is suspected because the males of the “bolas spiders” feed on the males of Psychoda which leads one to suppose that the females emit a pheromone (Yeargan & Quate 1996). Th e females of the “bolas spiders” are known to practise chemical ensnaring by mimicking the pheromones of moths in order to capture them; one may suppose than males which are much smaller do the same with the moth fl ies (Yeargan & Quate 1996).
Th e MSB are olfactory receptors which may be involved in locating host plants for egg-laying (review in Faucheux 1999). However, in the Diptera Tephritidae Bactrocera tryoni (Frogatt 1897), multiporous thin-walled sensilla basiconica are innervated by both a sensory cell with a CO2-detecting function, and two sensory cells which respond to general environmental odors (Stange & Stowe 1999). During the anthesis, Arum infl orescences are known to increase their temperature by a particular respiratory pathway and emit large quantities of CO2 (Lance 1974). Hence
psychodids may also be attracted to Arum infl orescences as source of CO2.
Th e USB of Psychodini are identical to the uniporous sensilla basiconica described on the antennae of primitive lepidoptera such as the Micropterigidae (Faucheux 1997). Th ey can be compared morphologically with the “fl at setae” located near the apiculus which prolonges the distal fl agellomere of the Psychodidae Trichomyiinae, Trichomyiam carlestobrai Wagner 2001. In Logima adumbrata (Satchell 1953) and Falsologima savaiiensis (Edwards 1928), Satchell (1953) notes a “sensory cone” on each of the three apical fl agellomeres. Similar cones (named “spines”) are also cited by Cordeiro & Bravo (2008) in Psychoda amazonensis Cordeiro & Bravo 2008. Goblet-shaped organs, named “sensilla coeloconica” by Campbell & Pelham-Clinton (1960) and “sensilla ampullaceal” by Wirth & Navai (1978), are present in Culicoides sp. (Diptera, Ceratopogonidae). All these structures unobserved in S.E.M by the previous authors could correspond to the USB present in Psychodini. Th ese sensilla with a terminal pore probably have a contact-chemoreceptive function (Zacharuk 1985). Although short in length, they have no diffi culty entering into contact with the anthers and the other pieces of the spadix when the insects are imprisoned within the spathe of Arum sp. Th eir presence revealed in Psycha grisescens, Phalaenoides phalaenoides and Chodopsycha sp. shows that their function is shared by several species of Psychodini. Th eir abundance at the antennal apex of Psychoda phalaenoides and at that of the male of Psycha grisescens justifi es their gustative function. Th eir absence in Psychoda crassipennis may be related to the presence in that species of sensilla chaetica with a purely tactile function. We have been unable to reveal the presence of USB in Logima sigma. Th e similar number of sensilla in both sexes of Psycha grisescens shows that the latter have identical possibilities of reception of gustative stimuli, which may intervene at diff erent moments in their behaviour.
Th e multiporous sensilla coeloconica (MSC) have so far not been identifi ed in the Psychodinae. Th ey probably correspond to the very numerous “papillae” in the Phlebotominae, for example Brumptomyia sp. (Shimabukoro et al. 2007) and Sergentomyia sp. (Kakarsulemankhel 2004). Th ey are identifi ed as sensilla coeloconica in P. argentipes (Ilango 2000). In the latter species, they all belong to the type “with a fence of microtrichia” whereas in the studied Psychodini, they are of the “naked” type. Th e sensilla coeloconica of the Psychodini are similar to those of the Lepidoptera in which the two types are present (Faucheux 1999). Th ese sensilla are olfactory receptors

Antennal sensilla of Psychodini
11
in Lepidoptera (Pophof 1997). In Apis mellifi ca, some sensilla coeloconica respond only to carbon dioxide while others react to temperature and humidity (Lacher 1964). In bloodsucking insects such as Culicoides sp. that must fi nd a warm-blooded host, the number of antennal sensilla coeloconica is considerable (Jamnback 1965). Moreover, at anthesis, the appendix of the infl orescence of Arum sp. produces heat and emits a specifi c odour which attracts insects (Gibernau et al. 2004). Th e MSC of the studied Psychodini (Psychoda phalaenoides, Psycha grisescens, Psychoda crassipennis and Chodopsycha sp.) could be the sensilla involved in this attraction.
Th e multiporous sensilla auricillica described in the Psychodini recall by their form the cup-shaped sensilla auricillica observed on the antennae of the tineid moth Trichophaga tapetzella (Faucheux 1989). As far as we know, they have not been discovered in any other Diptera or other Insecta. As for the other sensilla, a study in transmission electron microscopy would have to be undertaken to comprehend their structure and confi rm the presence of supposed pores. Th e MSA are present in four species; it would be worthwile to look for them in other Psychodidae.
Th e aporous sensilla chaetica observables only in Psychoda crassipennis and Psychoda phalaenoides (Table) are tactile mechanoreceptors (Zacharuk 1985). Th e tactile function of the antennae of other Psychodini is therefore negligible or compensated by other sensory organs.
Aporous Böhm’s sensilla of scape and pedicel are proprioceptive organs which monitor the position of the antennae (Schneider 1964; Faucheux 1999).
Phylogenetic considerationsTh is fi rst comparative study shows that none of the
fi ve species have the same antenna in terms of sensilla diversity and abundances. Even gender diff erences were found, but as only one species was observed, further studies are needed to validate this sexual antennal dimorphism. Psychoda phalaenoides appears to have the more ornamented antenna whereas Logima sigma presents the antenna with the less diversity and abundance of sensilla. More psychodid species must be studied before assigning to such diff erences some taxonomical or biological value.
Th e MTSB (ascoids) of Psychodidae have often been compared with the multiporous multibranched sensilla basiconica (ascoids) present in Lepidoptera Micropterigidae (Le Cerf 1926, Bourgogne 1951). However they diff er by the pore location: on the two faces in Psychodidae, only on the external face in Mi-cropterigidae (Faucheux 1999). Th e naked sensilla coe-
loconica of the dipterans resemble those of basal Lepi-doptera. Moreover, the uniporous sensilla basiconica of psychodids are very similar to the same sensilla only described on the antennae of basal Lepidoptera such as the Agathiphagidae (Faucheux 1990), Micropterigi-dae (Faucheux 1997; 2004a), and Heterobathmiidae (Faucheux 2004b).
ConclusionIn Phlebotominae, besides the ascoid sensilla present
in the antennae, other sensilla with a chemoreceptive function are present in mouthparts such as the labella and the maxillary palps (Spiegel et al. 2005 b). A study of the sensory equipment on the mouthparts of Psychodini is necessary. Mapping the sensilla distribution associated with electroantennography studies may help in the search for kairomones involved in plant attraction in these insects.Acknowledgements. Our acknowledgments to Mr. Pierre Watelet, Director of the Natural History Museum, Nantes, for allowing us the use of the scanning electron microscope of the University of Nantes; to Mr. Nicolas Stephant (Centre of S.E.M, Faculty of Sciences) and Mrs. Catherine Aké for their help with the photography; to Mr. Vittorio Ballardini for help with the translation; and to Quentin Laurent who prepared the specimens for identifi cation and separated the heads for S.E.M. analyses.
ReferencesAlbre J., Quilichini A., Gibernau M. 2003. Pollination ecology of Arum
italicum (Araceae) Botanical Journal of the Linnean Society 141: 205-215.Arcangeli G. 1883. Osservazioni sull’impollinizione in alcune Araceae.
Nuovo Giornale Botanico Italiano 15: 72-97.Beck I. 1983. Blütenökologische Untersuchungen am Aronstab (Arum
maculatum L.). unpublished thesis, University of Tübingen, 122 p.Bourgogne J. 1951. Ordre des Lépidoptères, p. 174-448 in Grassé P.P.
(éd.). Traité de Zoologie, tome X (1). Masson et Cie, Paris.Campbell J. A., Pelham-Clinton E. C. 1960. A taxonomic review of the
British species of Culicoides Latreille (Diptera, Ceratopogonidae). Proceedings of the Royal Society of Edinburgh, Section B 67: 181-302.
Cordeiro D., Bravo F. 2008. A new species of viviparous moth fl y (Diptera, Psychodidae, Psychodinae) from the Brazilian Amazon. Biota Neotropica 8 (2): 115-119.
Diaz A., Kite G. C. 2002. A comparison of the pollination ecology of Arum maculatum and A. italicum in England. Watsonia 24: 171-181.
Dougherty M., Hamilton G. 1997. Dodecanoic acid is the oviposition pheromone of Lutzomyia longipalpis. Journal of Chemical Ecology 23 (12): 2657-2671.
Dougherty M. J., Guerin P., Ward R. D., Hamilton J. G. C. 1999. Behavioural and electrophysiological responses of the phlebotominae sandfl y Lutzomyia longipalpis (Diptera: Psychodidae) when exposed to canid host odour kairomones. Physiological Entomology 24: 251-262.
Faucheux M. J. 1989. Antennal sensilla in male and female carpet moth, Trichophaga tapetzella L. (Lepidoptera: Tineidae): a scanning electron microscopic study. Annales de la société entomologique de France (n.s.) 25: 83-93.
Faucheux M. J. 1990. Antennal sensilla in adult Agathiphaga vitiensis Dumbl. and A. queenslandensis Dumbl. (Lepidoptera: Agathiphagidae). International Journal of Insect Morphology & Embryology 19: 257-268.

12
M. J. Faucheux & M. Gibernau
Faucheux M. J. 1997. Sensory organs on the antennae of Micropterix calthella L. (Lepidoptera: Micropterigidae). Acta Zoologica (Stockholm) 78: 1-8.
Faucheux M. J. 1999. Biodiversité et unité des organes sensoriels des Insectes Lépidoptères. Société des Sciences Naturelles de l’Ouest de la France, Nantes (éd.), 296 p.
Faucheux M. J. 2004a. Antennal sensilla of Sabatinca sterops Turner (Lepidoptera: Micropterigidae). Transactions of the Royal Society of South Australia 128: 187-194.
Faucheux M. J. 2004b. Antennal sensilla in female Heterobathmia pseuderiocrania (Insecta, Lepidoptera, Heterobathmiina). Revista del Museo Argentino de Ciencias Naturales, n.s. 6: 165-173.
Gibernau M., Macquart D., Przetak G. 2004. Pollination in the genus Arum – a review. Aroideana 27: 148-166.
Ilango K. 2000. Morphological characteristics of the antennal fl agellum and its sensilla chaetica with character displacement in the sandfl y Phlebotomus argentipes Annandale and Brunetti sensu lato (Diptera: Psychodidae). Journal of Biosciences 25 (2): 163-172.
Jamnback H. 1965. Th e Culicoides of New York State (Diptera: Ceratopo-gonidae). New York State Museum Scientifi c Bulletin 399: 1-154.
Ježek J. 1990. Redescriptions of none common Palaearctic and holarctic species of Psychodini End. (Diptera: Psychodidae). Actaz Entomologica Musei Nationalis Pragae 43: 33-83.
Ježek J., Van Harten A. 2005. Further new taxa and little known species of non-biting moth fl ies (Diptera: Psychodidae) from Yemen. Acta Entomologica Musei Nationalis Pragae 45: 199-220.
Kakarsulemankhel J. K. 2004. New record of Sergentomyia murghabiensis Perfi liev (1939) from Pakistan (Diptera : Psychodidae : Phlebotominae). Pakistan Journal of Biological Sciences 7 (1): 58-62.
Lacher V. 1964. Elektrophysiologische Untersuchungen an einzelen Rezep-toren für Geruch, Kohlendioxyd Luftfeuchtigkeit und Temperatur auf der Antennen den Arbeitsbiene und der Drohne (Apis mellifi ca L.). Zeitschrift für Vergleichende Physiology 48: 587-623.
Lack A. J., Diaz A. 1991. Th e pollination of Arum maculatum L.- a historical review and new observations. Watsonia 18: 333-342.
Lance C. 1974. Respiratory control and oxidative phosphorylation in Arum maculatum mitochondria. Plant Science Letters 2: 165-171.
Le Cerf F. 1926. Contribution à l’étude des organes sensoriels des Lépidop-tères. Encyclopédie entomologique B III (Lepidoptera) 1: 133-158.
Méndez M., Obeso J. R. 1992. Infl uencia del osmoforo en la produccion de infrutescencias en Arum italicum Miller (Araceae). Anales del Jardín Botánico de Madrid 50: 229-237.
Pophof B. 1997. Olfactory responses recorded from sensilla coeloconica of the silkmoth Bombyx mori. Physiological Entomology 22: 239-248.
Prime C. T. 1960. Lords and Ladies. Collins, London, 241 p.Rohacek J., Beck-Hayg I., Dobat K. 1990. Sphaeroceridae associated
with fl owering Arum maculatum (Araceae) in the vicinity of Tübingen, S W Germany. Senckenbergiana Biology 71: 259-268.
Satchell G. 1953. New and little known Samoan Psychodidae and a new species from Rarotonga. Proceedings of the Royal Entomological Society of London (B) 22: 181-188.
Schneider D. 1964. Insect antennae. Annual Review of Entomology 9: 103-122.
Séguy E. 1925. Diptères (Nématocères piqueurs) : Ptychopteridae, Orphnephi-lidae, Simuliidae, Culicidae, Psychodidae, Phlebotominae). Faune de France n° 12, Lechevalier, Paris, 109 p.
Séguy E. 1951. Ordre des Diptères, p. 449-744 in Grassé P. P. (éd.), Traité de Zoologie 10 (1er fascicule), Masson et Cie, Paris.
Shimabukoro P. H. F., Marassá A. M., Galati E. A. B. 2007. Brumptomyia carvalheiro sp. nov. (Diptera: Psychodidae: Phlebotominae) from Atlantic forest domain, São Paula State, Brazil. Zootaxa 1637: 47-54.
Spiegel C. N., Jeanbourquin P., Guerin P. M., Hooper A. M., Claude S., Tabacchi R., Sano S., Mori K. 2005a. (1 S, 3 S, 7 R)-3-methyl-α-himachalene from the male sandfl y Lutzomyia longipalpis (Diptera: Psychodidae) induces neurophysiological responses and attracts both males and females. Journal of Insect Physiology 51 (12): 1366-1375.
Spiegel C. N., Oliveira S. M. P., Brazil R. P., Soares M. J. 2005b. Structure and distribution of sensilla on maxillary palps and labella of Lutzomyia longipalpis (Diptera: Psychodidae) sand fl ies. Microscopy Research and Technique 66 (6): 321-330.
Stange G., Stowe S. 1999. Carbon-dioxide sensing structures in terrestrial arthropods. Microscopy Research and Technique 47: 416-427.
Tonnoir A. 1919. Notes sur les Ptychopteridae. Annales de la Société royale belge d’Entomologie 59: 115.
Vogel S., Martens. J. 2000. A survey of the function of the lethal kettle traps of Arisaema (Araceae), with records of pollinating fungus gnats from Nepal. Botanical Journal of the Linnean Society 133: 61-100.
Van der Pers J. N. C., Den Otter C. J. 1978. Single cell responses from olfactory receptors of small ermine moths to sex attractants. Journal of Insect Physiology 24: 337-343.
Wagner R. 2001. Contribution to the knowledge of Spanish Psychodidae (Diptera) with description of two new species. Zoologica baetica 12: 83-90.
Wirth W. W., Navai S. 1978. Terminology of some antennal sensory organs of Culicoides biting midges (Diptera: Ceratopogonidae). Journal of Medical Entomology 15: 43-49.
Withers P. 1988. Revisionary notes on british species of Psychoda Latreille (Diptera, Psychodidae) including new synonyms and a species new to science. British Journal of Entomological Natural History 1: 69-76.
Yeargan K. V., Quate L. W. 1996. Juvenile bolas spiders attract psychodid fl ies. Oecologia 106: 266-271.
Zacharuk R. Y. 1985. Antennae and sensilla, p. 1-70 in Kerkut G. A., Gilbert L. I. (eds.) Comprehensive Insect Physiology, Biochemistry and Pharmacology, vol. 6, Pergamon Press, London.





























