Embed Size (px)
Citation preview

Nucleic Acids Research, Vol. 19, No. 17 4611 -4618
Analysis of in vitro binding of U1-A protein mutants to U1snRNA
Wilbert Boelens*, Daniel Scherly" +, Eric J.R.Jansen, Karin Kolen, lain W.Mattaj' andWalther J.van VenrooijUniversity of Nijmegen, Department of Biochemistry, PO Box 9101, 6500 HB Nijmegen, TheNetherlands and 'European Molecular Biology Laboratory, Meyerhofstrasse 1, 6900 Heidelberg, FRG
Received July 3, 1991; Accepted August 2, 1991
ABSTRACT
Despite the great sequence similarity between UlA andU2B", both proteins do have a difference in RNAbinding specificity and in the way they bind to theircognate RNAs. The U1A protein is able to bind in vitroUl RNA independently of other factors. The U2B"protein binds specifically to U2 RNA in the presenceof the U2A' protein only. We have compared the effecton RNA binding of multiple double point mutations atanalogous positions in the U1A and U2B1 protein. Theresults obtained show that amino acids at almost allof the analogous positions tested in UIA and U2B"have a comparable qualitative effect on RNA bindingalthough the quantitative effect of mutations on U2B"is more severe than on UlA. Using UlA mutants withinternal duplications a distinct area of the RNP motifof the UlA protein was identified which appears not tobe directly involved in Ul RNA binding. In addition,roles of the highly conserved RNPI and RNP2sequences of the N-terminal RNP motif of the UlAprotein, are investigated by replacing them with theanalogous U1-70K sequences.
INTRODUCTION
The UlA protein is a constituent of the Ul small nuclearribonucleoprotein particle (snRNP). This particle belongs to thefamily of spliceosomal snRNPs and consist of a Ul snRNAmolecule, a set of at least seven polypeptides common tospliceosomal snRNPs and three specific proteins referred to asUIA, U1-70K and U1C (for review see 1). The UIA proteinis very similar to the U2 snRNP specific B" protein (2) and bothproteins contain two copies of the RNP motif. This motif is a70-90 amino acid long sequence which contains two highlyconserved short regions, referred to as RNP1 (or RNP consensussequence) and RNP2 (3,4). The N-terminal RNP copy of UlAand of U2B" plus a few flanking amino acids have been shownto be required for specific RNA binding. In U1A and U2B" theC-terminal RNP motif is not required for specific RNA binding(5-8).
The N-terminal RNP domain of U1A is different from the RNPdomain of the U2B" protein at only 20 out of 101 positions.Although both RNP domains thus are very similar, the proteinsUlA and U2B" bind their cognate RNA differently. UlA proteincan bind specifically to Ul RNA in the absence of other UlsnRNP components while U2B" binds specifically to U2 snRNAonly in the presence of U2A' protein (7-12).The UlA protein binds to the second hairpin of Ul RNA and
the U2B" protein to the fourth hairpin of U2 RNA. The highlysimilar loop sequences of Ul hairpin H and U2 hairpin IV areessential for the binding of the proteins (5,7,8,13-15). TheU1-70K protein contains one copy of the RNP motif which isessential for the specific binding to the first hairpin of Ul RNA(16-18).
Recently the crystal structure of the RNP domain of UlA hasbeen solved (19). The secondary structure of this domain consistsof a four-stranded antiparallel (3-sheet and two a-helices arrangedin the order (31-aA-(32-(33-aB-04 in the primary structure. Theconserved RNP1 and RNP2 sequences are located side by sidein the middle of the four-stranded :-sheet (19).To localize amino acids of the RNP domain involved in the
interaction with RNA, the RNA binding ability of several mutantproteins were analysed. We have compared the influence on RNAbinding of double point mutations in UlA and U2B" protein andfound that in both proteins amino acids at analogous positionsare important for RNA binding. Furthermore we show that theconserved RNP1 sequence of the UlA protein is more importantthan the less conserved RNP2 sequence for U1 RNA bindingand that a distinct area of the RNP domain of UlA appears notto be directly involved in Ul RNA binding.
MATERIALS AND METHODSPlasmidsThe human UlA (2) and U2B" cDNAs (20) were subcloned intothe EcoRI site of pGEM-3Zf(+) (Promega) vector from whichthe BamHI restriction site was removed by BamHI digestion,filling in the recessed 3' termini and re-ligation. Single strandedDNA was produced with the helper phage M13KO7 and point
* To whom correspondence should be addressed
+ Present address: Department of Microbiology, CMU, 9 Avenue de Champel, CH/211 Geneve 4, Switzerland
.) 1991 Oxford University Press
4612 Nucleic Acids Research, Vol. 19, No. 17
mutations were produced using the oligonucleotide-directedmutagenesis system from Amersham (U.K.). Since each mutationintroduced a unique BamHI site, the mutation was checked bya BamHI-EcoRI digestion of the recombinants and sequencingof the mutated area.
The human U2A' cDNA (21) was subcloned into the EcoRIsite of pGEM-3Zf(+). The U1 and U2 RNA derivatives usedin this work have been described earlier (7).
In vitro transcriptionTo produce T7 messenger RNA for U lA, U2B", U2A' proteinsand mutants thereof, 2itg of linearized (HindIII or BamHI)template was incubated at 37°C for 60 min in a total volumeof 50,1d contaning 40 mM Tris-HCl pH 7.9 /6 mM MgCl2/10mM spermidine-HCl/ 10 mM NaCl/2 mM dithiothreitol(DTT)/0. 1 mg/ml BSA/601M GpppG/1 mM ATP, UTP, CTP,GTP/60 U RNase inhibitor (RNasin, Promega)/30 U T7 RNApolymerase (Promega). After incubation the non-incorporatednucleotides were removed with a Sephadex G50 spin column,the RNA was phenol extracted and precipitated with 0.1 volume3 M Na-acetate pH 5.2 and 2.5 volume ethanol. The RNA pelletwas resuspended in 10u1l sterile water. Approximately 2tg mRNAwas synthesized with this method.
Biotinylated U snRNAs were prepared as described aboveexcept that 100gM biotin-11-UTP (Enzo) was added to thetranscription reaction mix.
In vitro translation35S-labelled UIA and U2B" wild type and mutant proteins weremade in wheat germ extract (WGE, Amersham U.K.) bytranslation of their corresponding in vitro transcribed T7 mRNAs.200 ng T7 mRNA was incubated in a total volume of 30ulcontaining 15/l WGE/120 mM potassiumacetate/67gM each of19 amino acids (minus methionine) together with 30OgCL-[35Slmethionine (1200 Ci/mmol, Amersham). Whenunlabelled U2A' was made, 30OgC [35S]methionine was replacedby 67gtM methionine.
In vitro protein-RNA binding assay
8gtl KHN buffer (100 mM KCl/20 mM Hepes-KOH pH7.9/0.05% NP-40) was mixed with 20 ng/gl biotinylated RNA(in a solution containing 2gg/pJ Ecoli tRNA) and with lgl in vitromade radiolabelled UIA (mutant) protein or with both lgl
radiolabelled U2B" and Ill unlabelled U2A'. After incubationfor 60 min at 25°C the mixture was diluted with 5001l KHNbuffer and transferred to a vial containing 15AlS packedstreptavidin-agarose beads (BRL) pre-washed with KHN buffer.The solution was mixed for 90 min by rotating the tubes endover end at room temperature. The beads were pelleted bycentrifugation, the supernatant aspirated and the pellet washedthree times by rotating for 1 min with 1 ml KHN buffer. Thefinal bead-pellet was resuspended in 2Ogl SDS-sample buffer andheated at 100°C for 4 min. After brief centrifugation, thesupernatant was loaded onto an SDS-polyacrylamide gel. Afterelectrophoresis the gel was incubated with dimethylsulfoxide(DMSO, 2 x 30 min), 16% (w/v) 2,5-diphenyloxozal (PPO) inDMSO (1 x 60 min), water (3 x 30 min), dried and exposed toKodak X-omat film at -80°C.
Competition assay to measure relative binding affinitiesA series of incubations of labelled UlA mutant with biotinylatedU1 RNA was performed in the presence of increasing amountsof purified recombinant U lA protein. U 1A protein was purifiedby DEAE-cellulose and a phosphocellulose (Whatman P11)chromatography from a bacterial lysate containing overexpressedU lA polypeptide (22). Truncated radiolabelled U lA protein(A203 which contains the N-terminal 203 amino acids of UlAwt) was added as internal control. A203 has an equal bindingaffinity for U1 RNA as wild type U lA protein (6,19). Theprecipitated proteins were analysed by polyacrylamide gelelectrophoresis and the U IA mutant and A203 were quantifiedby densitometry of the autoradiogram. The relative RNA bindingaffinity is the amount of purified U 1A protein required for 50%reduction of the precipitated U IA mutant divided by the amountrequired for 50% reduction of precipitation of A203.
Table 1. Binding of double point mutants of UIA to Ul RNA and of U2B" to U2 RNA.
A protein B" protein
mutated positions binding relative mutated positions bindingamino acids affinity amino acids
AV 2/3 + 1.0 DT 2/3 +HT 10/11 + 0.3 HT 7/8 -IY 12/13 - IY 9/10 -NE 18/19 + 0.2 ND 15/16 -KS 28/29 + 1.0 RS 25/26 -GQ 38/39 + 1.0 GH 35/36 +KM 50/51 + 0.4 KM 47/48 -RG 52/53 - RG 49/50 -KE 60/61 + 1.0 KE 57/58 +RS 70/71 + 1.0 RQ 67/68 +KP 80/81 + 0.5 KP 77/78 -DI 92/93 + 1.0 DI 89/90 +VE 102/103 + 1.0 AD 99/100 +
The positions of the amino acids replaced by GlySer are shown. Specific RNA binding was measured withthe biotinylated RNA binding assay (see Materials and Methods). No detectable RNA binding is indicatedby -, and RNA binding ability is indicated by +. For the U IA mutants also relative values of RNA bindingaffinity, measured with the UIA competition assay, are given. The amount of purified U IA protein requiredfor 50% reduction of the precipitated UIA mutant was determined and divided by the amount requiredfor 50% reduction of wild type UIA precipitation (see Materials and Methods).

Nucleic Acids Research, Vol. 19, No. 17 4613
RESULTSThe effect on RNA binding of double point mutants of UlAcompared with analogous U2B" mutant proteinsTo obtain the mutants, BamHI restriction sites were created at13 analogous positions in the UlA and U2B" cDNAs. A BamHIrestriction site positioned in frame results in a replacement oftwo amino acids by GlySer at a defined position in the aminoacid sequence (table 1). The 35S-labelled mutant proteins weresubsequently tested in an in vitro RNA binding assay (seeMaterials and Methods). The UlA mutants were incubated withbiotinylated Ul RNA and the U2B" mutants with biotinylatedU2 RNA in the presence of in vitro made U2A'. After incubationthe RNA bound proteins were precipitated with streptavidin-agarose beads and analysed by SDS polyacrylamide gelelectrophoresis (see fig. lA and lB for some typical examples).Table 1 shows which mutants specifically bound to their cognateRNA and which did not. Mutations in the conserved RNP1 andRNP2 sequences of both UIA and U2B" (positions 52/53 and12/13 respectively in UIA and positions 49/50 and 9/10respectively in U2B") abolished RNA binding. All the otherdouble point mutants of U1A were able to bind to UIl RNA. Incontrast, a number of analogous mutations in U2B" had a strongeffect on U2 RNA binding. Double point mutations in U2B" atposition 7/8, 15/16, 25/26, 47/48 and 77/78 abolished bindingto U2 RNA.
Precipitation of labelled mutants with biotinylated RNA doesnot give very precise information on the RNA binding affinity.To investigate the RNA binding affinity of these mutants incomparison with UlAwt, competition experiments wereperformed. Labelled UIA mutants were incubated withbiotinylated Ul RNA in the presence of increasing amounts ofpurified unlabelled recombinant UlAwt protein produced in E.coli. Subsequently bound protein was precipitated by streptavidin-agarose beads. As an internal control a truncated labelled UIAprotein (A203, which contains the N-terminal 203 amino acidsof UlAwt) was used. A203 and wild type UlA protein have anequal binding affinity for Ul RNA (6,19). The precipitated UlAmutants and A203 were then separated on a polyacrylamide gel,autoradiographed (see fig IC shows typical examples) andquantified by scanning. A 50% reduction of radio-labelled UlAwtprecipitation was achieved by the addition of 9ng of purified UlAprotein. This value was arbitrarily set as a relative value of 1.0.The relative values for binding affinity of the UlA-mutants areshown in Table 1. When UIA was mutated either at positions10/11, 18/19, 50/51 or 80/81 a lower relative binding affinitywas found. Analogous mutations in U2B" abolished detectableRNA binding completely (Table 1). Thus amino acid changesat these positions reduce RNA binding affinity of both proteins,albeit to different extents.
Mutations at positions 2/3, 38/39, 60/61, 70/71, 92/93 or102/103 did not reduce the binding affinity of the UIA proteinfor Ul RNA and the analogous mutations in U2B" showed noeffect on U2 RNA binding (Table 1). Thus the amino acids atthese positions in both UlA and U2B" protein are probably notdirectly involved in RNA binding.Only one pair of mutants, UIA28/29 and U2B"25/26, gave
very different results. The U2B"25/26 mutation abolished RNAbinding while the analogous mutation in UlA (U1A28/29) hadno effect (Table 1). The most likely explanation for this is theinability of U2B"25/26 to interact with U2A' protein (9). Thisinteraction is required for specific binding of U2 RNA by U2B"(7,8).
Effects of small deletions and sequence duplications in theRNP domain of UlA on RNA bindingWe next analysed the effects of small internal deletions andsequence duplications in the UIA RNP domain. Deletions mayintroduce a severe disturbance of the tertiary structure of the RNPdomain. However, comparison of different members of the RNPmotif family shows that deletions of particular regions of the motifis not incompatable with the retention of RNA binding (4). Onthe other hand, because all primary sequence information is stillpresent, internal sequence duplications may have relatively minoreffects on the tertiary structure of the RNP domain.
Deletion and duplication mutants were obtained using thedouble point mutant clones (see table 1 for amino acid positions).Mutant proteins were produced in vitro and tested using the RNAbinding assay described above.
A
B
A2/3 A18/19 A52/53
I Ul U2 I Ul U2 I J1. U2
' 4in' _
B"2/3 B"15/16 B"49/50
I U2 Ul I U2 Ul I U2 Ul
sw
CAwt*
A203- AO
A203h
59 41 18 6 4 2 0.6 0 ng
5411 6. 4 2 0. 0i
59 41 18 6 4 2 0. 6 0 ng
- M _1
-...... _0o _0 #A_ w 10
Fig. 1. Typical examples of binding behavior of U1A and U2B" mutants measuredby precipitation of labelled protein with biotinylated RNA transcripts. (A) Shownare the precipitation of a U1A mutant which binds as wild type (UIA2/3), onewhich has a reduced RNA binding affinity (U1A18/19) and one which does notbind Ul RNA (U1A52/53). The in vitro made 35S-labelled mutants were incubatedwith biotinylated Ul and U2 RNA transcripts. After precipitation of RNA-boundproteins with streptavidin-agarose beads, the proteins were analysed by SDS-polyacrylamide gel electrophoresis. As an internal control AIOl was used (notshown, see ref. 5). Lane I shows the input material used for the binding assay.(B) Precipitation of a U2B" mutant which is able to bind U2 RNA (U2B"2/3)and two which can not bind U2 RNA (U2B"(15/16) and U2B"49/5s). The in vitromade 35S-labelled mutants were incubated with biotinylated U2 and Ul RNAtranscripts and analysed as described above. As an internal control U2B"88 wasused (not shown, see ref. 7). Lane I contains the input material used for the bindingassay. (C) Example of the relative RNA binding affinity assay. Labelled UlAwtand mutant A80/81 was precipitated with biotinylated Ul RNA in the presenceof varying amounts of purified recombinant UlAwt protein and analysed asdescribed above. The amount of recombinant UlAwt (in ng) used is indicatedabove each lane. As an internal control a truncated labelled UlA protein (A203,which contains the N-terninal 203 amino acids of UlAwt and has the same bindingaffinity as UlAwt) was used. The amount needed for a 50% reduction of theprecipitated radio-labelled mutant was measured as described in Materials andMethods and divided by the amount required for 50% reduction of precipitationof wild type UIA. The relative Ul RNA binding affinity of the UIA doublepoint mutants are given in table 1. Lane I contains 10% of the input mixtureused for the binding assay.
I

4614 Nucleic Acids Research, Vol. 19, No. 17
Figure 2 shows that all deletions inside the RNP domainabolished RNA binding. Thus, when sequences of about 10 aminoacids are deleted from the N-terminal 91 amino acids the proteincan not form a functional RNA binding domain.
In contrast, deletion from amino acids 93 up to 102 did notabolish the U 1 RNA binding (fig. 2A lane 22- 30). This result
was rather unexpected because a truncated protein including theN-terminal 91 amino acids of U IA itself does not detectably bindU 1 RNA (5,6). Results from other deletion mutants suggest thatthe specific amino acid sequence from 93 up to 102 is not requiredfor RNA binding but that the presence of amino acids in thisregion may influence the conformation of the RNP domain.
A
km P4Iam:
I
a
a... ..I'_"6
a-U}r.w2
V'
*0*40
'E3
W4
a I
B
I
Fig. 2. Binding of U lA deletion mutants to Ul RNA. (A) The in vitro made 35S-labelled deletion mutants of UIA protein were incubated with biotinylated Uland U2 RNA transcripts and analysed as described in fig IA. As an internal control AIOI was used. The first and last amino acid position of the deleted sequenceare given between the brackets. Lanes I contain 10% of the quantity of the mixture used for the binding assay. The position of the deletion mutant is marked withan arrowhead. (B) Schematic representation of UlAwt and deletion mutants. The RNP domain of UIA is indicated by the dashed area and the RNP1 by a blackbox. Deletions in the mutants are shown as open areas.
A.dk-i.I'MM
t.
zt" ll.
.'t.

Nucleic Acids Research, Vol. 19, No. 17 4615
The relative values for binding affinity of the mutantsAA(93 - 102), AA(93 - 118) and AA(93 -203), measured withthe competition assay described above, were all 0.4. This meansthat the deletions from amino acid 93 up to 118 and from 93up to 203 has the same effect on the RNA binding affinity asthe deletion from 93 up to 102.
A Ai(3-10) Ai(19-28) Ai(29-38)
The internal duplications had, in general, a less dramatic effect.Mutants Ai(3-10), Ai(19-28), Ai(61-70), Ai(51-80),Ai(61 -80), Ai(71 -80), Ai(81 -92) and Ai(93- 102) specificallybound Ul RNA (fig. 3A). In contrast mutant Ai(39-50) andAi(51 -60) were not able to bind Ul RNA and mutant Ai(29-38)bound very weakly. These results suggest that only in the latter
Ai(39-50) Ai(51-60) Ai(61-70)
I UI U2 I U1 U2 I Ul U2
_ _ _ _ .. _
40_
A101-
1 2 3 4 5 6 7 8 9 10 11 12
I Ul U2 I U1 U2 I Ul U2
4...
04'
S.
13 14 15 16 17 18
Ai(51-80) Ai(61-80) Ai(71-80s) Ai(81-92) Ai(93-102)
I U1 U2 I Ul U2 I Ul U2 I U1 U2 I Ul U2
o a
7- -~~~~~a
A101_ - 5e
19 20 21 22 23 24 25 26 27 28 29 30 31 32 33
BU1-A mutants
I 101
Ziih ... .__- ..'.iI_ .__
::t eE :\ 1x: lS1 \1 ......................... .__ ___.___ __ ._ ...................[ .__ .v .::1 .n1.1: .. ... _ __ . .. - -- - - - - -' '-- - - '- :---n[:I:S X:' : X: I L:x\lIXsXSr ' -- - ' . '- -: - -.--' .......... _ = _ .5:::E z x:51: 1 . _ . _ . _ _._. . __: ....... .... _ . :_ _: _JC _._.: . n b ............. _:l .15 = ..... .... _ _ . _ ____ _ _ . _ _ :_ __. _: . _ . _ . _ ............... _ . _ _._ zS;
C . 1 .na11 .E_E .E .155:v1U = = _ _ ::_ : ::=_ =_._ n:L::xE::x:E\:t\SE\:EX :t\:t\.}2::x:E\x1 .. ... . . _ . .. xno :E::Eu:E;:E1:Eox:_::xM ::Ex:x::6;:E1 _ _ - ---- -- ----_srns s Ct::'.1:t'11:1:1 . .: -.- ___._''- ........... - ._=- . _ .: .. .... : _:: '' '282 binding to Ul RNA
/+
Fig. 3. Binding of UlA internal duplication mutants to Ul RNA. (A) The binding of UlA mutants with an internal duplication to Ul and U2 RNA transcriptswas analysed as described in the legend to Figure IA. As an internal control AIOI was used. The first and last amino acid position of the sequence which has beenduplicated in the polypeptide are given between the brackets. The internal duplication mutants, all of similar size, are indicated by an arrowhead. Lane I contains10% of the quantity of the mixture used for the binding assay. (B) Schematic representation of UlAwt and internal duplication mutants. UlAwt is represented as
explained in fig. 2B. The duplicated amino acids are shown as an extra box above each UIA representation.
s. 1..
*..m*...
A wt
Ai3- 10)
Ali19-28)
Al 29-38)
Ai 39-500
AI; 51-60
Ail(O1-70)
AiS1-8O)
AiNSI-80S
Ail7l-801
Ai(81-9 2)
AiI93-l102
4m lw40Pk-W,
4616 Nucleic Acids Research, Vol. 19, No. 17
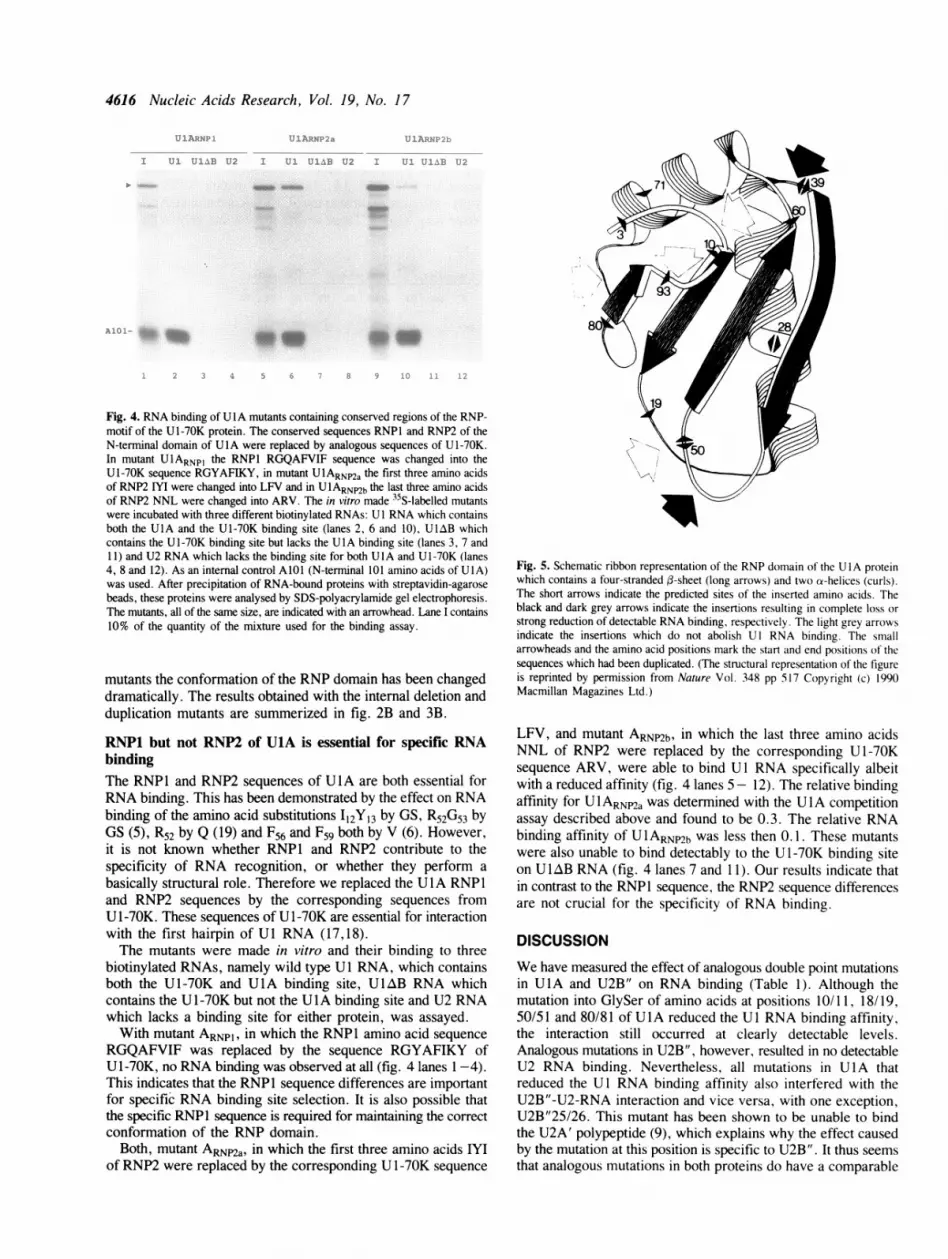
Fig. 4. RNA binding of U IA mutants containing conserved regions of the RNP-motif of the U 1-70K protein. The conserved sequences RNPl and RNP2 of theN-terminal domain of U1A were replaced by analogous sequences of U1-70K.In mutant UlARNpI the RNP1 RGQAFVIF sequence was changed into theU1-70K sequence RGYAFIKY, in mutant UlARNna the first three amino acidsof RNP2 IYI were changed into LFV and in UlARNP2b the last three amino acidsof RNP2 NNL were changed into ARV. The in vitro made 35S-labelled mutantswere incubated with three different biotinylated RNAs: U I RNA which containsboth the UlA and the Ul-70K binding site (lanes 2, 6 and 10), ULAB whichcontains the U1-70K binding site but lacks the U1A binding site (lanes 3, 7 and11) and U2 RNA which lacks the binding site for both U1A and U 1-70K (lanes4, 8 and 12). As an internal control AIOI (N-terminal 101 amino acids of UIA)was used. After precipitation of RNA-bound proteins with streptavidin-agarosebeads, these proteins were analysed by SDS-polyacrylamide gel electrophoresis.The mutants, all of the same size, are indicated with an arrowhead. Lane I contains10% of the quantity of the mixture used for the binding assay.
mutants the conformation of the RNP domain has been changeddramatically. The results obtained with the internal deletion andduplication mutants are summerized in fig. 2B and 3B.
RNP1 but not RNP2 of UlA is essential for specific RNAbindingThe RNP1 and RNP2 sequences of U lA are both essential forRNA binding. This has been demonstrated by the effect on RNAbinding of the amino acid substitutions 112Y13 by GS, R52G53 byGS (5), R52 by Q (19) and F56 and F59 both by V (6). However,it is not known whether RNP1 and RNP2 contribute to thespecificity of RNA recognition, or whether they perform a
basically structural role. Therefore we replaced the UlA RNP1and RNP2 sequences by the corresponding sequences fromU 1-70K. These sequences of U1-70K are essential for interactionwith the first hairpin of Ul RNA (17,18).The mutants were made in vitro and their binding to three
biotinylated RNAs, namely wild type Ul RNA, which containsboth the U1-70K and ULA binding site, U1AB RNA whichcontains the U1-70K but not the UIA binding site and U2 RNAwhich lacks a binding site for either protein, was assayed.With mutant ARNPI, in which the RNP1 amino acid sequence
RGQAFVIF was replaced by the sequence RGYAFIKY ofU 1-70K, no RNA binding was observed at all (fig. 4 lanes 1-4).This indicates that the RNP1 sequence differences are importantfor specific RNA binding site selection. It is also possible thatthe specific RNP1 sequence is required for maintaining the correctconformation of the RNP domain.
Both, mutant ARNp2a in which the first three amino acids IYIof RNP2 were replaced by the corresponding U 1-70K sequence
Fig. 5. Schematic ribbon representation of the RNP domain of the U 1 A proteinwhich contains a four-stranded /-sheet (long arrows) and two a-helices (curls).The short arrows indicate the predicted sites of the inserted amino acids. Theblack and dark grey arrows indicate the insertions resulting in complete loss orstrong reduction of detectable RNA binding, respectively. The light grey arrowsindicate the insertions which do not abolish Ul RNA binding. The smallarrowheads and the amino acid positions mark the start and end positions of thesequences which had been duplicated. (The structural representation of the figureis reprinted by permission from Nature Vol. 348 pp 517 Copyright (c) 1990Macmillan Magazines Ltd.)
LFV, and mutant ARNP2b, in which the last three amino acidsNNL of RNP2 were replaced by the corresponding U 1-70Ksequence ARV, were able to bind U 1 RNA specifically albeitwith a reduced affinity (fig. 4 lanes 5- 12). The relative bindingaffinity for UlARNP2a was determined with the UlA competitionassay described above and found to be 0.3. The relative RNAbinding affinity of U 1 ARNP2b was less then 0.1. These mutantswere also unable to bind detectably to the U 1-70K binding siteon UIAB RNA (fig. 4 lanes 7 and 11). Our results indicate thatin contrast to the RNP1 sequence, the RNP2 sequence differencesare not crucial for the specificity of RNA binding.
DISCUSSIONWe have measured the effect of analogous double point mutationsin UlA and U2B" on RNA binding (Table 1). Although themutation into GlySer of amino acids at positions 10/11, 18/19,50/51 and 80/81 of UIA reduced the Ul RNA binding affinity,the interaction still occurred at clearly detectable levels.Analogous mutations in U2B", however, resulted in no detectableU2 RNA binding. Nevertheless, all mutations in UlA thatreduced the Ul RNA binding affinity also interfered with theU2B"-U2-RNA interaction and vice versa, with one exception,U2B"25/26. This mutant has been shown to be unable to bindthe U2A' polypeptide (9), which explains why the effect causedby the mutation at this position is specific to U2B". It thus seemsthat analogous mutations in both proteins do have a comparable

Nucleic Acids Research, Vol. 19, No. 17 4617
effect on RNA binding, although the effect on U2B" is moresevere than the effect on UIA. Clearly, the way both proteinsbind their cognate RNAs is highly similar as might be expectedfrom the great homology between UlA and U2B" on one handand between their binding sites on the other hand (2,7).
In several double point mutants Lys and Arg residues havebeen changed (e.g. K28, K50, R52, K60, R70 and K80, see Table1). Some of these positively charged residues may interact withthe phosphate backbone. Nagai et al. (19) reported the effect ofreplacing Lys and Arg residues in the RNP domain by Gln. Thereplacement of Arg on position 52 abolished RNA binding, thereplacement of K50 and K80 reduced RNA binding affinity andthe replacement of K28, K60 and R70 had no significant effecton Ul RNA binding. These results are in good agreement withour results shown in Table 1. It seems that the reduced RNAbinding affinity of double point mutants A50/51, A52/53 andA80/81 is at least partially caused by the replacement of apositively charged amino acid which may be directly involvedin U1 RNA binding.
Several mutants with insertions in the RNP domain were testedin the RNA binding assay. The location of the inserted duplicatedamino acids is of course not precisely known but can be predicted,at least in the cases where RNA binding is retained, by assumingthat the tertiary structure of the domain will remain largely intactin spite of the inserted amino acids and that the inserted aminoacids will loop out at regions not involved in the formation ofthe $-sheet or the oa-helices. Based upon this assumption thepredicted locations of the inserted amino acids in the mutantsare N-terminal of (31 strand in Ai(3-10), between the (31 strandand the a-helix A in Ai(19-28), between the a-helix A and the(32 strand in Ai(29 -38), between the a-helix B and the (34 strandin Ai(7 1-80) and C-terminal from the (4 strand in Ai(93 -102)(see fig. 5). The inserted amino acids in the mutants Ai(39 -50),Ai(51-60), Ai(51-80), Ai(61-70), Ai(61-80) and Ai(81-92)have more than one predicted location. However, in mutantsAi(39 -50) and Ai(51 -60) the inserted amino acids are probablylocated in the loop between (2 and (33 strand because of theflexibility of this loop (19). The extra loops containing the insertedamino acids may sterically hinder binding of Ul RNA. Theinsertion sites in the mutants which still are capable of bindingUl RNA, are therefore probably not directly involved in Ul RNAbinding. These predicted sites are located N-terminal of (31,between (31 and a-helix A, between a-helix B and (4 and C-terminal of (34 (see fig. 5). Almost all these sites are located inareas of the molecule distant from the area of the molecule whichis thought to be important for the RNA binding (19). An exceptionis the loop between the (31 strand and the a-helix A (mutantAi(19 -28)) which is very close to the loop between the (32 and(3 strands. Previously, we have shown that the loop betweenthe (2 and the (33 strand is essential for Ul RNA binding (7).
Mutant Ai(29 -38) binds only very weakly to U1 RNA. Theinserted amino acids in this mutant are probably positionedbetween the A a-helix and the (32 strand, which might suggestthat this region is involved in Ul RNA binding.Only the internal duplication mutants Ai(39 -50) and
Ai(51-60) were not able to bind Ul RNA. In these mutants theinserted amino acids may be located in the loop between the (2and the (3 strand which indeed is essential for Ul RNA binding(7,8,19). Insertions at this position probably disrupt the structurerequired for RNA binding.When the RNP1 sequence in UIA was replaced by the related
U1-70K RNP1 sequence the binding to Ul RNA was abolished.
Four amino acids of UIA RNP1 differ from those of U1-70KRNP1. In the crystal structure of the N-terminal fragment of UlA(19) two of these are part of the interior core (Val and Phe atposition 57 and 59 which were replaced by Ile and Tyr) and twopoint into solution (Gln and Ile at position 54 and 58 which werereplaced by Tyr and Lys). The changes of the inward-pointingresidues are rather conserved in contrast to the replacements ofthe other two. This could mean that the two amino acids, Gln54and 11e8, which are on the protein surface, may be directlyinvolved in specific interaction with the second hairpin of UlRNA. However, it remains possible that the replacement mighthave a non-specific effect on RNA binding through altering theconformation of the protein.
Replacements of parts of the RNP2 sequence in U1A by relatedU1-70K RNP2 sequences, IYI by LFV and NNL by ARV,reduced the RNA binding affinity of U1A but did not changethe RNA binding specificity. This indicates that the RNP2sequence in U1A is less important for the RNA binding specificityand has probably mainly a structural role.
ACKNOWLEDGEMENTS
We are grateful to Ger Pruijn for critical reading of themanuscript. We thank Dr. K.Nagai for permission to use fig 2Aof ref. 19 and Rob Nelissen for cloning U1A cDNA in pET-3a.The investigations were supported in part by the NetherlandsFoundation for Chemical Research (SON) with financial aid fromthe Netherlands Organization for Scientific Research (NWO) andthe Dutch Rheumatism Foundation. W.B. and D.S. wererecipients of EMBO fellowships.
REFERENCES1. Luhrmann, R., Kastner, B. and Bach, M., (1990) Bioch. Bioph. Acta, 1087,
265-292.2. Sillekens, P.T.G, Habets, W.J., Beijer, R.P. and van Venrooij, W.J. (1987)
EMBO J., 6, 3841-3848.3. Adams, S.A., Nakagawa, T., Swanson, M.S., Woodruff, T.K. and Dreyfuss,
G. (1986) Mol. Cell. Biol., 6, 29321-2943.4. Dreyfuss, G., Swanson, M.S. and Pinol-Roma, S. (1988) Trends biochem.
Sci., 13, 86-91.5. Scherly, D., Boelens, W., van Venrooij, W.J., Dathan, N.A., Hamm, J.,
Mattaj, I.W., (1989) EMBO J., 8, 4163-4170.6. Lutz-Freyermuth C., Query C.C., Keene J.D., (1990) Proc. Natl. Acad.
Sci. USA, 87, 6393-6397.7. Scherly, D., Boelens, W., Dathan, N.A, van Venrooij, W.J, Mattaj, I.W
(1990) Nature, 345, 502-506.8. Bentley,R.C. and Keene,J.D. (1991) Mol. Cell. Biol., 11, 1829-1839.9. Scherly D, Dathan N.A, Boelens W, van Venrooij W.J, Mattaj I.W (1990)
EMBO J., 9, 3675-3681.10. Boelens,W., Scherly,D., Beijer,R.P., Jansen,E.J.R., Dathan,N.A.,
Mattaj,I.W., van Venrooij,W.J. (1991) Nucleic. Acids Res., 19, 455-46011. Fresco,L.D., Harper,D., Keene,J.D. (1991) Mol. Cell. Biol., 11,
1578-1589.12. Scherly,D., Kambach,C. Boelens,W., van Venrooij,W.J. and Mattaj,I.W.
J. Mol. Biol. in press.13. Lutz Freyermuth C., and Keene J.D., (1989) Mol. Cell. Biol., 9, 2975-2982.14. Patton, J.R., Habets, W., van Venrooij, W.J., Pederson, T., (1989) Mol.
Cell. Biol., 9, 3360-3368.15. Bach, M., Krol, A., Luhrmann, R. (1990) Nucleic Acids Res., 18,449-457.16. Query, C.C., Bentley, R.C. and Keene, J.D. (1989) Cell, 57, 89-10117. Surowy C.S., van Santen V.L., Scheib-Wixted S.M., Spritz R.A., (1989)
Mol. Cell. Biol. 9, 4179-4186.18. Query,C.C., Bentley,R.C. and Keene,J.D. (1989) Mol. Cell. Biol., 9,
4872-4881.19. Nagai, K., Oubridge, C., Jessen, T.H. Li, J. and Evans, P.R. (1990) Nature,
348, 515-520.

4618 Nucleic Acids Research, Vol. 19, No. 17
20. Habets, W.J., Sillekens, P.T.G., Hoet, M.H., Schalken, J.A., Roebroek,A.J.M., Leunissen, J.A.M., van de Ven, W.J.M, and van Venrooij, W.J.(1987) Proc. Natl. Acad. Sci. USA, 84, 2421-2425.
21. Sillekens,P.T.G., Beijer,R.P., Habets,W.J., and van Venrooij,W.J. (1989)Nucleic Acids Res., 17, 1893-1906.
22. Studier, F.W., Rosenberg, A.H. and Dunn, J.J. (1990) Methods inEnzymology, 185, 60-89.





























