Embed Size (px)
Citation preview

wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 9
Avai lab le a t www.sc iencedi rec t .com
journa l homepage : www.e lsev ie r . com/ loca te /wat res
Anaerobic acidogenic digestion of olive mill wastewatersin biofilm reactors packed with ceramic filters or granularactivated carbon
Lorenzo Bertin a,*, Silvia Lampis b, Daniela Todaro c, Alberto Scoma a, Giovanni Vallini b,Leonardo Marchetti a,c, Mauro Majone d, Fabio Fava a
aDepartment of Applied Chemistry and Material Science (DICASM), Faculty of Engineering, University of Bologna, via Terracini 28,
40131 Bologna, ItalybDepartment of Biotechnology, University of Verona, Strada Le Grazie 15 e Ca’ Vignal, I-37134 Verona, Italyc INCA e Interuniversitary Consortium “Chemistry for the Environment”, via delle Industrie 21/8, I-30175 Marghera (VE), ItalydDepartment of Chemistry, Sapienza University of Rome, Piazzale Aldo Moro 5, I-00185 Rome, Italy
a r t i c l e i n f o
Article history:
Received 15 April 2010
Received in revised form
4 June 2010
Accepted 11 June 2010
Available online 18 June 2010
Keywords:
Olive mill wastewaters
Acidogenesis
Packed bed biofilm reactors
Vukopor S10 ceramic cubes
Polyhydroxyalkanoates
Microbial speciation
* Corresponding author. Tel.: þ39 051 209031E-mail addresses: [email protected]
unibo.it (A. Scoma), giovanni.vallini@univr(M. Majone), [email protected] (F. Fava).0043-1354/$ e see front matter ª 2010 Elsevdoi:10.1016/j.watres.2010.06.025
a b s t r a c t
Four identically configured anaerobic packed bed biofilm reactors were developed and
employed in the continuous acidogenic digestion of olive mill wastewaters to produce
volatile fatty acids (VFAs), which can be exploited in the biotechnological production of
polyhydroxyalkanoates. Ceramic porous cubes or granular activated carbon were used as
biofilm supports. Aside packing material, the role of temperature and organic loading rate
(OLR) on VFA production yield and mixture composition were also studied. The process
was monitored through a chemical, microbiological and molecular biology integrated
procedure. The highest wastewater acidification yield was achieved with the ceramic-
based technology at 25 �C, with an inlet COD and an OLR of about 17 g/L and 13 g/L/day,
respectively. Under these conditions, about the 66% of the influent COD (not including its
VFA content) was converted into VFAs, whose final amount represented more than 82% of
the influent COD. In particular, acetic, propionic and butyric acids were the main VFAs by
composing the 55.7, 21.5 and 14.4%, respectively, of the whole VFA mixture. Importantly,
the relative concentrations of acetate and propionate were affected by the OLR parameter.
The nature of the packing material remarkable influenced the process performances, by
greatly affecting the biofilm bacterial community structure. In particular, ceramic cubes
favoured the immobilization of Firmicutes of the genera Bacillus, Paenibacillus and Clos-
tridium, which were probably involved in the VFA producing process.
ª 2010 Elsevier Ltd. All rights reserved.
1. Introduction similar to those of some petroleum-derived plastics [e.g.
Polyhydroxyalkanoates (PHAs) are promising microbial
biopolymers,mainly because: a) they showphysical properties
7; fax: þ39 051 2090322.t (L. Bertin), [email protected] (G. Vallini), leonardo.
ier Ltd. All rights reserved
the copolymer poly(3-hydroxybutyrate/3-hydroxyvalerate)
[P3-(HB/HV)] can replace polypropylene in a wide range of
applications (Lee, 1996)]; b) they can be produced by means of
univr.it (S. Lampis), [email protected] (D. Todaro), alberto.scoma2@[email protected] (M. Marchetti), [email protected]
.

wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 94538
renewable resources (Serafim et al., 2008); c) they are
completely biodegradable (Braunegg et al., 1998) and biocom-
patible (Mochizuki, 2002). To date, industrial PHA productions
have taken advantage only of pure bacterial cultures andwell-
defined synthetic media (Reddy et al., 2003; Khanna and
Srivastava, 2005; Philip et al., 2007). PHA production
processes carried out under such well-defined conditions can
provide excellent results, but are not economically competi-
tivewith those of petroleum-derived plastics (Noda et al., 2005;
Philip et al., 2007). Thus, the exploitation of alternative low-
cost feeding stocks (e.g. organic wastes) and mixed bacterial
consortia (i.e. highly biodiverse microbial populations selec-
tively enriched for the ability of producing PHA) is of great
relevance to get an economically feasible PHA production
process. In this respect, many papers have dealt with PHAs
production by activated sludgemicroorganisms fromdifferent
waste (Reis et al., 2003; Chua et al., 2003; Rhu et al., 2003;
Dionisi et al., 2005a; Albuquerque et al., 2007; Salmiati et al.,
2007; Bengtsson et al., 2008a,b). In particular, Dionisi et al.
(2005a) proposed the exploitation of olive mill wastewaters
(OMWs) in a three-stage integrated anaerobic-aerobic PHA
producing process: in the first anaerobic stage, the organic
waste is fermented under acidogenic conditions to obtain an
effluent rich in volatile fatty acids (VFAs), which are suitable
substrates for the PHA biological synthesis occurring in the
following second and third aerobic steps. Although this
process has been already assessed for the performances of the
aerobic phases (Dionisi et al., 2004, 2005b, 2006), a little has
been done so far to optimize the acidogenic step. In this
respect, the employment of reactors capable of supporting
high flow rate feedings, such as Packed Bed Biofilm Reactors
(PBBRs) (Bertin et al., 2004), can allow to minimize the VFA-
consuming methanogenic activity, which is normally medi-
ated by bacteria with very low specific growth rates. Thus, we
recently developed a biofilm reactor packed with Ceramic
Cubes (CCs) with the aim of fermenting OMWs to generate
a VFA-rich effluent employed in the production of PHAs
(Beccari et al., 2009). However, no efforts to optimize the
process in terms of concentration and relative amounts of
produced VFAs were made in the previous study.
Given the key role played by the relative content of
carboxylic acids containing even or odd number of carbon
atoms on the properties of resulting PHAs (Bengtsson et al.,
2008b), this research was undertaken to evaluate the influ-
ence of some key process parameters, such as temperature
and Organic Loading Rate (OLR), on concentration and relative
amounts of produced VFAs by the recently developed OMW
acidogenic process (Beccari et al., 2009). Moreover, the role of
the packing material was also evaluated by developing
parallel reactors packed with Granular Activated Carbon
(GAC), a biomass carrier which was already employed in the
biomethanization of OMWs (Bertin et al., 2004), and by
determining the microbial speciation of the biofilms gener-
ated onto the surface of CC and GAC samples collected at the
end of the study.
To the very best of our knowledge, this is the first work in
which an immobilized cell-based acidogenic anaerobic diges-
tion of OMWswas studied in terms of the influence of itsmain
process parameters and assessed through an integrated
chemical,microbiologicalandmolecularbiologymethodology.
2. Materials and methods
2.1. Olive mill wastewaters
TwoOMWs, namedOMW1andOMW2,were kindly purchased
by the Sant’Agata d’Oneglia (Imperia, Italy) and Grassanese
(Matera, Italy) three phase olive mills, respectively, and
employed in the research. Their COD was about 25 and 35 g/L,
respectively, partially due to VFAs (about 10 and 12 gCOD/L,
respectively) and phenols (about 2 g/L in both OMWs). Total
and volatile suspended solids were 20 and 12 g/L, respectively,
in OMW1; 32 and 13 g/L, respectively, in OMW2. Their pH
values were 4.3 and 4.6, respectively.
2.2. Packed bed biofilm reactors
Four identically configured up-flow PBBRs were developed
following the approach reported elsewhere (Beccari et al.,
2009) and employed as described in Section 2.3. Each PBBR
consisted of a 2.5 L-hermetically closed glass column (diam-
eter: 80 mm; height: 450 mm) wrapped with a silicon tubing
serpentine continuously recycling thermostated water and
equipped with a recycle line. The recycling ratio, expressed as
the ratio between the recycled broth flow and the whole flow
entering the column, was about 0.97. The liquid and gas
effluents were collected in a tank, connected to a “Mariotte”
bottle through which the produced biogas volume was
determined. A pH probe (81-04 model, ATI Orion, Boston, MA)
was placed at the top of the bioreactor. Two of the reactors
were packed with Ceramic Cubes (CCs, Vukopor S10 product,
Lanik, Boskovice, CZ) whose dimensions, porosity and density
were 25 � 25 � 18 mm, 10 ppi and 2.38 g/mL, respectively (CC-
PBBRs), while the other two with Granular Activated Carbon
(GAC, CP4-60 product, Chemviron Carbon, Feluy, Belgium),
consisting of cylinders of about 3 mm diameter and 10 mm
length, whose density was 1.32 g/mL (GAC-PBBRs). As a result
of support addition, the reaction volumes of CC-PBBRs became
2.25 and 2.28 L (CC1 and CC2, respectively), while the ones of
GAC-PBBRs became 1.50 and 1.80 L (GAC1 and GAC2,
respectively).
2.3. Olive mill wastewater digestion experiments
Where not differently described, the reactor influent flows
consisted of OMWs diluted with an equal volume of tap water
and amended with urea (0.45 g/L) and with a 10 N NaOH
solution (added to correct their pH to 5.5).
One PBBR per group, i.e. CC1 and GAC1, where thermo-
stated at 55 �C, whereas the other two, i.e. CC2 and GAC2, at
35 �C. They were all filled anaerobically with amended OMW1,
which was previously inoculated at 10% v/v with the same
deoxygenated, high-density suspension of microbial biomass
employed to develop the process described in Beccari et al.
(2009), which did not harbor any detectable taxa belonging
to archaeal domain. The main detected bacteria occurring in
the inoculum were a strain belonging to the Flexibacter-Cyto-
phaga-Bacteroides group and a Syntrophus sp.. The reactor
medium was replaced with deoxygenated fresh amended
OMW1 for three successive two-weeks batch cycles. The
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 9 4539
reactors were then forced to operate under continuous mode.
Five experiments of about one-month long were carried out
with the aim of studying the effect of temperature and loading
conditions on process performances. Steady state conditions
were considered to be attained when VFAs, COD and phenols
concentrations along with produced biogas were appreciably
constant (with standard deviations generally not exceeding
15%) for at least a week (i.e. at least 4 times each process HRT).
Steady state was typically achieved within two weeks after
a new loading condition was set.
The main process working parameters are reported in
Table 1. The first experiment, through which information on
the influence of temperature on process performances was
attained, was carried out by feeding both thermophilic
(experiment No. 1a) and mesophilic (experiment No. 1b)
reactors with the amended OMW1. The four successive
experiments, through which the former information was
integrated and the influence of OLR was studied, were per-
formed with CC2 and GAC2 reactors, which were fed with the
amended OMW1 (experiments No. 2 and 3) or OMW2 (exper-
iments No. 4 and 5). Experiments No. 3, 4 and 5 were carried
out by thermostating the reactors at 25 �C. PBBRs were fed at
an OLR of about 9 (experiments No. 1 and 3) and 13 (experi-
ments No. 2 and 4) g/L/day. During the last experiment, OMW2
was not diluted with tap water, so that the influent COD was
doubled respect to the one of experiment No. 4 (Table 1).
Moreover, during the last experiment the pH of GAC2 inlet
flow was not modified by means of NaOH amendment.
Liquid samples (collected on daily bases) were filtered on
0.22 mm cellulose-nitrate filters and then analyzed for COD,
VFAand total phenols (parameters related to influent flows are
reported with the subscript suffix “IN”, e.g. CODIN, while the
ones related to the effluents are reported with the subscript
suffix“OUT”, e.g.CODOUT).VFAamountswereexpressedasgof
equivalentCOD (gCOD) bymeansof stoichiometric calculations.
The net COD conversion into VFAs (COD/VFA) percentage,
representing the ratio between the net VFA production and
the influent COD excluding its VFA fraction, was employed to
define the process efficiency and calculated as follows:
COD/VFA ¼ VFAOUT � VFAIN
CODIN � VFAIN� 100 (1)
The whole process yield was calculated as the percentage of
the ratio between the effluent VFA total amount and the
influent COD:
Table 1 e List of performed experiments and relatedmainworkreactor couples (CC1 and GAC1, experiment No. 1a; CC2 and G
Exp. OMW T (�C) CODIN (g/L) VFAI
1a OMW1 55 13.52 � 0.43 6.1
1b OMW1 35 13.52 � 0.43 6.1
2 OMW1 35 11.71 � 0.26 6.4
3 OMW1 25 11.55 � 0.32 5.8
4 OMW2 25 16.71 � 0.52 7.9
5 OMW2 25 36.64 � 0.74 12.
a CC2.
b GAC2.
Process yield ¼ VFAOUT
CODIN� 100 (2)
The concentration of the total immobilized biomass
occurring at the top, the middle and the bottom of CC2 and
GAC2 was quantified at the end of the study by collecting
samples of about 1 g of support from the bottom, the middle
and the top of the fixed-beds (50, 200 and 380 mm from the
column bottom, respectively) and by subjecting them to
protein analysis. The biofilms immobilized on the surface of
the same samples together with the last experiment influent
and effluent flows were microbiologically characterized by
means of DNA extraction as described below.
2.4. Total DNA extraction, PCR amplification anddenaturing gradient gel electrophoresis
Total DNA extraction fromCCandGAC sampleswas carried as
reported elsewhere (Beccari et al., 2009). The 16S rRNA-genes
wereamplifiedbyPCRusingTaqDNApolymerasewithprimers
targeting conserved domains. Bacterial 16S rRNA genes were
selectively amplified using F8/R11 primers (Beccari et al., 2009)
with the following thermocycling program: initial denatur-
ation at 94 �C for 2 min; 30 cycles of denaturation at 94 �C for
45 s, annealing at 50 �C for 30 s, and extension at 72 �C for
2.5 min; final extension at 72 �C for 5min. Afterwards a nested
PCR was performed as described in Beccari’s work (2009).
Conditions were as above, except for number of cycles, 35, the
annealing temperature, 57 �C, and extension time, 35 s.
For Archaea, primers A109-f (Grosskopf et al., 1998) and
1510-r (Lane, 1991) were used for nearly complete 16S rRNA
gene amplification. Afterwards a nested PCR was performed
on the hypervariable V2-V3 region using primers A109(T)-f
and 515-GC-r (Roest et al., 2005), with a GC-clamp. The first
archaeal PCR reaction was performed with the following
thermocycle program: initial denaturation at 94 �C for 5 min;
30 cycles of denaturation at 94 �C for 45 s, annealing at 52 �Cfor 30 s, extension at 72 �C for 1 min; and final extension at
72 �C for 5 min. The nested PCR was as above but with 35
cycles. All primers were purchased from Sigma-Genosys
(Milan, Italy). The PCR products were quantified using Low
DNA MassTM Ladder (Celbio, Italy) in a 2.0% agarose gel.
DGGE analyses were performed on amplicons obtained
both for bacterial V3 and archaeal V2-V3 regions following the
procedures reported in Beccari’s et al.’s work (2009). Repre-
sentative DGGE bands were excised and incubated for 4 h in
ing parameters. Experiment No.1 was carried out with bothAC2, experiment No. 1b).
N (gCOD/L) PhenolIN (g/L) OLR (g/L/day)
83 � 0.29 1.034 � 0.08 9.66
83 � 0.29 1.034 � 0.08 9.62
65 � 0.25 1.009 � 0.04 12.5
12 � 0.14 1.399 � 0.06 8.17
24 � 0.29 0.978 � 0.03 13.3
52 � 0.46 2.393 � 0.04 22.3a 44.4b
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 94540
50 mL of sterile water. A total of 12 and 21 bands from ampli-
cons obtained respectively for Archaea and Bacteria regions
were sequenced.
2.5. Cloning, sequencing, and phylogenetic analysis
DGGE bands containing DNA to be sequenced were re-ampli-
fied. PCR amplification was carried out as described before,
except for the use of non-GC-clamped primers. PCR products
were transformed in Escherichia coli Xl1blue using the pGEM-T
vector system according to the manufacturer’s instructions
(Promega, Italy), sequenced on both strands, and finally
searched for homology using the BLASTN database (Altschul
et al., 1997).
The sequences were initially aligned using the multiple
alignment program CLUSTAL_X 1.83 (Thompson et al., 1997).
A phylogenetic tree was constructed using the neighbour-
joining method with the MEGA version 4.0 software package
(Kumar et al., 2008). Bootstrap analysis was performed from
1000 bootstrap replications.
2.6. Analytical methods
VFA concentration was monitored through a HP GC-5890
(Agilent, Milano, Italy) equipped with a FID detector and
a Supelcowax-10 column (SigmaeAldrich, Milano, Italy) under
the following conditions: initial temperature 60 �C; isothermal
for 1 min; temperature rate 25 �C/min; final temperature
150 �C; isothermal for 6 min; temperature rate 4 �C/min; final
temperature 180 �C; temperature rate 25 �C/min; final
temperature 240 �C; injector and FID temperature 280 �C;carrier gas flow rate (nitrogen) 17.6 mL/min. Before the anal-
yses, the samples were diluted with an equal amount of
a 60 mM oxalic acid solution. VFAs concentrations are
expressed as g of COD equivalents/L (gCOD/L). COD and total
phenol concentrations were determined spectrophotometri-
cally according to the following methods: Hach Mn(III) (Miller
et al., 2001) and Folin-Ciocalteu (Folin and Ciocalteu, 1927),
respectively. Total phenols were determined by employing
4-hydroxybenzoate as the calibration standard. Biogas
amount and composition were daily determined as reported
elsewhere (Bertin et al., 2004). The concentration of the total
biomass occurring at the bottom, themiddle and the top of the
reactor packed beds was quantified according to the Lowry
method applied in previous studies (Bertin et al., 2004).
Table 2 e COD, VFA and phenol concentrations in PBBR effluenperformed experiments.
Exp. CC-PBBRs
CODOUT
(g/L)VFAOUT
(gCOD/L)CH4/CODDEP
a
(L/g)PhenolOUT
(g/L)
1a 10.93 � 0.37 6.138 � 0.11 0.197 � 0.02 0.774 � 0.10
1b 9.350 � 0.26 6.207 � 0.52 0.334 � 0.05 0.797 � 0.07
2 10.33 � 0.30 6.232 � 0.30 0.380 � 0.06 1.007 � 0.03
3 11.20 � 0.16 6.121 � 0.32 0.010 � 0.00 1.120 � 0.06
4 15.47 � 0.43 13.73 � 0.49 0.015 � 0.00 0.830 � 0.05
5 28.52 � 0.89 13.67 � 0.42 0.024 � 0.01 2.256 � 0.06
a Depleted COD.
3. Results
Significant amounts of VFAswere found to be generated in the
bioreactors only when the anaerobic treatment was per-
formed in CC2. The main results of the five successive
experiments performed with the CC- and GAC-PBBRs under
continuousmode of operation are summarized in Table 2. The
highest VFAs production was observed when CC2 was ther-
mostated at 25 �C and fed with an OLR of about 13 g/L/day
(experiment No. 4): under such conditions, the total VFAs
concentration in the effluent was 13.73 gCOD/L and it was
mainly due to acetic, propionic and butyric acids (representing
the 55.7, 21.5 and 14.4% of the whole detected VFAs, respec-
tively, Fig. 1a). The net COD conversion into VFAs was about
the 66%. VFAs total amount represented about the 89% of the
overall effluent COD (Table 3), while the process yield was
about 82% (Table 3). The pH of such a VFA-rich effluent was
5.13 � 0.04.
3.1. Effect of packing material on PBBRs performances
The effect of packing material on process performances was
studied throughout the first four experiments performed with
the parallel CC- and GAC-PBBR under the same temperature
and loading conditions (CC1 and GAC1, employed in the sole
experiment No. 1; CC2 and GAC2, Table 1). Higher VFA
concentrations were always attained in the CC-PBBRs than in
the GAC-PBBRs, where a large part of the influent VFA load
was conversely depleted (Table 2). Compatible pH values for
biological acidogenic processes were alwaysmeasured for CC-
PBBRs (5.23 averagely), while in GAC-PBBRs such a parameter
was in the range 6.43e6.97. High methane productions
occurred in the latter reactors, where in particular CH4
production yields close to the maximum theoretical ones (i.e.
0.35 L of CH4 produced per g of COD removed)were obtained in
the first two experiments (Table 2). At the same time, GAC-
PBBRs provided very high COD removals which ranged
between 58 and 86% of the initial COD. Conversely, COD
removals were very low (between 3 and 31%) in CC-PBBRs,
where a negligible methanogenic activity occurred during the
experiments carried out at 25 �C.Different mixtures of VFAs accumulated in the two packed
bed reactor types: acetic, propionic and butyric acids were the
main VFAs generally occurring in CC-PBBR effluents, whereas
ts along with CH4 production yields related to all the
GAC-PBBRs
CODOUT
(g/L)VFAOUT
(gCOD/L)CH4/CODDEP
a
(L/g)PhenolOUT
(g/L)
2.827 � 0.16 0.837 � 0.18 0.391 � 0.04 0.071 � 0.04
2.133 � 0.06 0.501 � 0.04 0.339 � 0.06 0.076 � 0.02
1.677 � 0.02 0.189 � 0.03 0.324 � 0.02 0.287 � 0.03
1.787 � 0.14 0.553 � 0.06 0.261 � 0.01 0.389 � 0.03
5.396 � 0.35 1.497 � 0.10 0.116 � 0.01 0.207 � 0.06
15.27 � 1.05 4.808 � 0.63 0.135 � 0.02 0.954 � 0.16
Acetic
Propionic
i-Butyric
Butyric
i-Valeric
ValericOthers
1a1b2
34
5
0
20
40
60
80
100
A
Acetic
Propionic
i-Butyric
Butyric
i-Valeric
ValericOthers
1a1b2
34
5
0
20
40
60
80
100
B
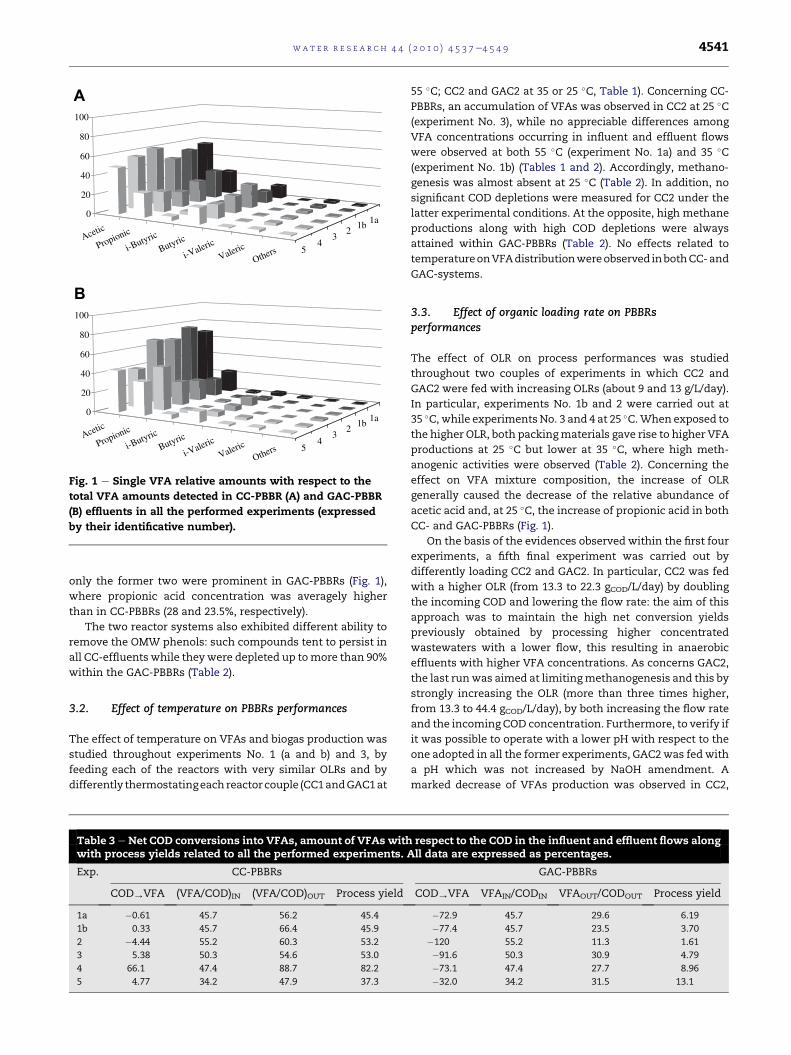
Fig. 1 e Single VFA relative amounts with respect to the
total VFA amounts detected in CC-PBBR (A) and GAC-PBBR
(B) effluents in all the performed experiments (expressed
by their identificative number).
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 9 4541
only the former two were prominent in GAC-PBBRs (Fig. 1),
where propionic acid concentration was averagely higher
than in CC-PBBRs (28 and 23.5%, respectively).
The two reactor systems also exhibited different ability to
remove the OMW phenols: such compounds tent to persist in
all CC-effluents while theywere depleted up tomore than 90%
within the GAC-PBBRs (Table 2).
3.2. Effect of temperature on PBBRs performances
The effect of temperature on VFAs and biogas production was
studied throughout experiments No. 1 (a and b) and 3, by
feeding each of the reactors with very similar OLRs and by
differently thermostatingeach reactor couple (CC1andGAC1at
Table 3 e Net COD conversions into VFAs, amount of VFAs withwith process yields related to all the performed experiments. A
Exp. CC-PBBRs
COD/VFA (VFA/COD)IN (VFA/COD)OUT Process yield
1a �0.61 45.7 56.2 45.4
1b 0.33 45.7 66.4 45.9
2 �4.44 55.2 60.3 53.2
3 5.38 50.3 54.6 53.0
4 66.1 47.4 88.7 82.2
5 4.77 34.2 47.9 37.3
55 �C; CC2 and GAC2 at 35 or 25 �C, Table 1). Concerning CC-
PBBRs, an accumulation of VFAs was observed in CC2 at 25 �C(experiment No. 3), while no appreciable differences among
VFA concentrations occurring in influent and effluent flows
were observed at both 55 �C (experiment No. 1a) and 35 �C(experiment No. 1b) (Tables 1 and 2). Accordingly, methano-
genesis was almost absent at 25 �C (Table 2). In addition, no
significant COD depletions were measured for CC2 under the
latter experimental conditions. At the opposite, high methane
productions along with high COD depletions were always
attained within GAC-PBBRs (Table 2). No effects related to
temperatureonVFAdistributionwereobserved inbothCC-and
GAC-systems.
3.3. Effect of organic loading rate on PBBRsperformances
The effect of OLR on process performances was studied
throughout two couples of experiments in which CC2 and
GAC2 were fed with increasing OLRs (about 9 and 13 g/L/day).
In particular, experiments No. 1b and 2 were carried out at
35 �C, while experimentsNo. 3 and 4 at 25 �C.When exposed to
the higher OLR, both packingmaterials gave rise to higher VFA
productions at 25 �C but lower at 35 �C, where high meth-
anogenic activities were observed (Table 2). Concerning the
effect on VFA mixture composition, the increase of OLR
generally caused the decrease of the relative abundance of
acetic acid and, at 25 �C, the increase of propionic acid in both
CC- and GAC-PBBRs (Fig. 1).
On the basis of the evidences observed within the first four
experiments, a fifth final experiment was carried out by
differently loading CC2 and GAC2. In particular, CC2 was fed
with a higher OLR (from 13.3 to 22.3 gCOD/L/day) by doubling
the incoming COD and lowering the flow rate: the aim of this
approach was to maintain the high net conversion yields
previously obtained by processing higher concentrated
wastewaters with a lower flow, this resulting in anaerobic
effluents with higher VFA concentrations. As concerns GAC2,
the last runwas aimed at limitingmethanogenesis and this by
strongly increasing the OLR (more than three times higher,
from 13.3 to 44.4 gCOD/L/day), by both increasing the flow rate
and the incoming COD concentration. Furthermore, to verify if
it was possible to operate with a lower pH with respect to the
one adopted in all the former experiments, GAC2was fed with
a pH which was not increased by NaOH amendment. A
marked decrease of VFAs production was observed in CC2,
respect to the COD in the influent and effluent flows alongll data are expressed as percentages.
GAC-PBBRs
COD/VFA VFAIN/CODIN VFAOUT/CODOUT Process yield
�72.9 45.7 29.6 6.19
�77.4 45.7 23.5 3.70
�120 55.2 11.3 1.61
�91.6 50.3 30.9 4.79
�73.1 47.4 27.7 8.96
�32.0 34.2 31.5 13.1

Fig. 2 e Top: DGGE fingerprints of the eubacterial communities. Samples collected from top, middle and bottom sections of
the reactors are marked with T, M and B, respectively. OMWIN refers to the inlet flow, CC2OUT and GAC2OUT to the effluents
discharged under steady state conditions. Roman numerals (I and II) correspond to different replicates. Letters indicate
bands that have been excised, cloned and sequenced. Bottom: Similarity dendrograms indicate relationships among the
different DGGE profiles.
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 94542
which probably suffered from the high-applied OLR. However,
in agreement with the above described trend, acetic and
propionic acid relative amounts further decreased and
increased, respectively. Concerning GAC2, methane produc-
tion yield was still comparable to the one observed in exper-
imentNo. 4 although the influent flowpHwas not increased to
5.5. However, the highest tested loading condition led to the
lowest COD consumption ever detected in such a reactor and
the VFAs concentration was correspondently the highest
obtained within GAC-PBBRs. Nevertheless, VFAs were yet
significantly consumed as a result of the methanogenesis
(Table 2).
3.4. Microbiological features of the biofilms
The total amount of immobilized biomass in CC2 and GAC2,
along with its microbial composition, was analyzed at the end
of the study, when the two reactors were sacrificed and
samples of both packing materials were collected at three
different heights of the reactor packed beds. The total
immobilized biomass (quantified as mg of dried biomass/g of
dried support) found in the reactor packed beds collected from
the bottom, the middle and the top of the reactors was 12.6,
15.2 and 18.1, respectively, for CC2, and 1.8, 3.5 and 2.6,
respectively, for the GAC2. Considering the dry weight of the
support initially introduced in the GAC- and CC-PBBR (739.8
and 1228.1 g, respectively), it was estimated that the total
immobilized biomass available in the CC- and GAC-PBBR was
11.31 and 3.215 g, respectively.
The composition of both Bacteria and Archae cenoses
enriched at the end of the study in CC2 and GAC2 reactors and
in their inlet and outlet flows was monitored through PCR-
DGGE analysis. All the evidences gathered for Bacteria in
relation to either packing matrices or influent and effluent
wastewaters indicate the presence of heterogeneous
communities, quite rich in bacterial biodiversity (Fig. 2,
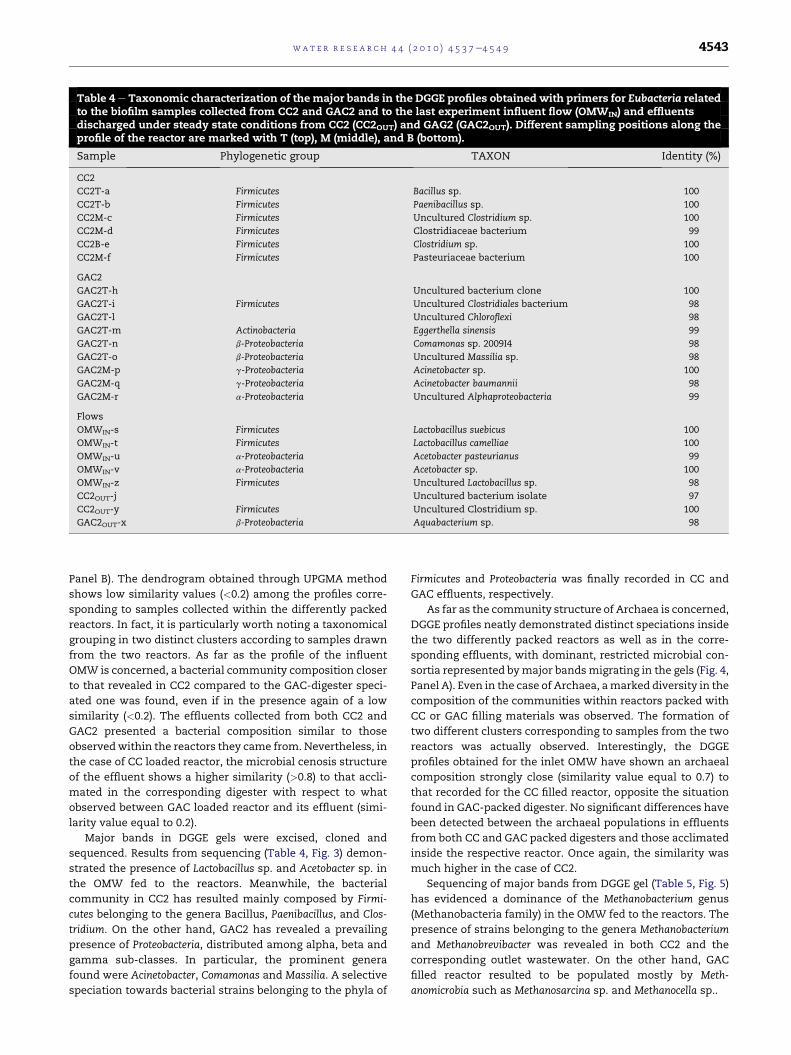
Table 4 e Taxonomic characterization of themajor bands in the DGGE profiles obtained with primers for Eubacteria relatedto the biofilm samples collected from CC2 and GAC2 and to the last experiment influent flow (OMWIN) and effluentsdischarged under steady state conditions from CC2 (CC2OUT) and GAG2 (GAC2OUT). Different sampling positions along theprofile of the reactor are marked with T (top), M (middle), and B (bottom).
Sample Phylogenetic group TAXON Identity (%)
CC2
CC2T-a Firmicutes Bacillus sp. 100
CC2T-b Firmicutes Paenibacillus sp. 100
CC2M-c Firmicutes Uncultured Clostridium sp. 100
CC2M-d Firmicutes Clostridiaceae bacterium 99
CC2B-e Firmicutes Clostridium sp. 100
CC2M-f Firmicutes Pasteuriaceae bacterium 100
GAC2
GAC2T-h Uncultured bacterium clone 100
GAC2T-i Firmicutes Uncultured Clostridiales bacterium 98
GAC2T-l Uncultured Chloroflexi 98
GAC2T-m Actinobacteria Eggerthella sinensis 99
GAC2T-n b-Proteobacteria Comamonas sp. 2009I4 98
GAC2T-o b-Proteobacteria Uncultured Massilia sp. 98
GAC2M-p g-Proteobacteria Acinetobacter sp. 100
GAC2M-q g-Proteobacteria Acinetobacter baumannii 98
GAC2M-r a-Proteobacteria Uncultured Alphaproteobacteria 99
Flows
OMWIN-s Firmicutes Lactobacillus suebicus 100
OMWIN-t Firmicutes Lactobacillus camelliae 100
OMWIN-u a-Proteobacteria Acetobacter pasteurianus 99
OMWIN-v a-Proteobacteria Acetobacter sp. 100
OMWIN-z Firmicutes Uncultured Lactobacillus sp. 98
CC2OUT-j Uncultured bacterium isolate 97
CC2OUT-y Firmicutes Uncultured Clostridium sp. 100
GAC2OUT-x b-Proteobacteria Aquabacterium sp. 98
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 9 4543
Panel B). The dendrogram obtained through UPGMA method
shows low similarity values (<0.2) among the profiles corre-
sponding to samples collected within the differently packed
reactors. In fact, it is particularly worth noting a taxonomical
grouping in two distinct clusters according to samples drawn
from the two reactors. As far as the profile of the influent
OMW is concerned, a bacterial community composition closer
to that revealed in CC2 compared to the GAC-digester speci-
ated one was found, even if in the presence again of a low
similarity (<0.2). The effluents collected from both CC2 and
GAC2 presented a bacterial composition similar to those
observedwithin the reactors they came from. Nevertheless, in
the case of CC loaded reactor, the microbial cenosis structure
of the effluent shows a higher similarity (>0.8) to that accli-
mated in the corresponding digester with respect to what
observed between GAC loaded reactor and its effluent (simi-
larity value equal to 0.2).
Major bands in DGGE gels were excised, cloned and
sequenced. Results from sequencing (Table 4, Fig. 3) demon-
strated the presence of Lactobacillus sp. and Acetobacter sp. in
the OMW fed to the reactors. Meanwhile, the bacterial
community in CC2 has resulted mainly composed by Firmi-
cutes belonging to the genera Bacillus, Paenibacillus, and Clos-
tridium. On the other hand, GAC2 has revealed a prevailing
presence of Proteobacteria, distributed among alpha, beta and
gamma sub-classes. In particular, the prominent genera
found were Acinetobacter, Comamonas and Massilia. A selective
speciation towards bacterial strains belonging to the phyla of
Firmicutes and Proteobacteria was finally recorded in CC and
GAC effluents, respectively.
As far as the community structure of Archaea is concerned,
DGGE profiles neatly demonstrated distinct speciations inside
the two differently packed reactors as well as in the corre-
sponding effluents, with dominant, restricted microbial con-
sortia represented bymajor bandsmigrating in the gels (Fig. 4,
Panel A). Even in the case of Archaea, amarked diversity in the
composition of the communities within reactors packed with
CC or GAC filling materials was observed. The formation of
two different clusters corresponding to samples from the two
reactors was actually observed. Interestingly, the DGGE
profiles obtained for the inlet OMW have shown an archaeal
composition strongly close (similarity value equal to 0.7) to
that recorded for the CC filled reactor, opposite the situation
found in GAC-packed digester. No significant differences have
been detected between the archaeal populations in effluents
from both CC and GAC packed digesters and those acclimated
inside the respective reactor. Once again, the similarity was
much higher in the case of CC2.
Sequencing of major bands from DGGE gel (Table 5, Fig. 5)
has evidenced a dominance of the Methanobacterium genus
(Methanobacteria family) in the OMW fed to the reactors. The
presence of strains belonging to the genera Methanobacterium
and Methanobrevibacter was revealed in both CC2 and the
corresponding outlet wastewater. On the other hand, GAC
filled reactor resulted to be populated mostly by Meth-
anomicrobia such as Methanosarcina sp. and Methanocella sp..
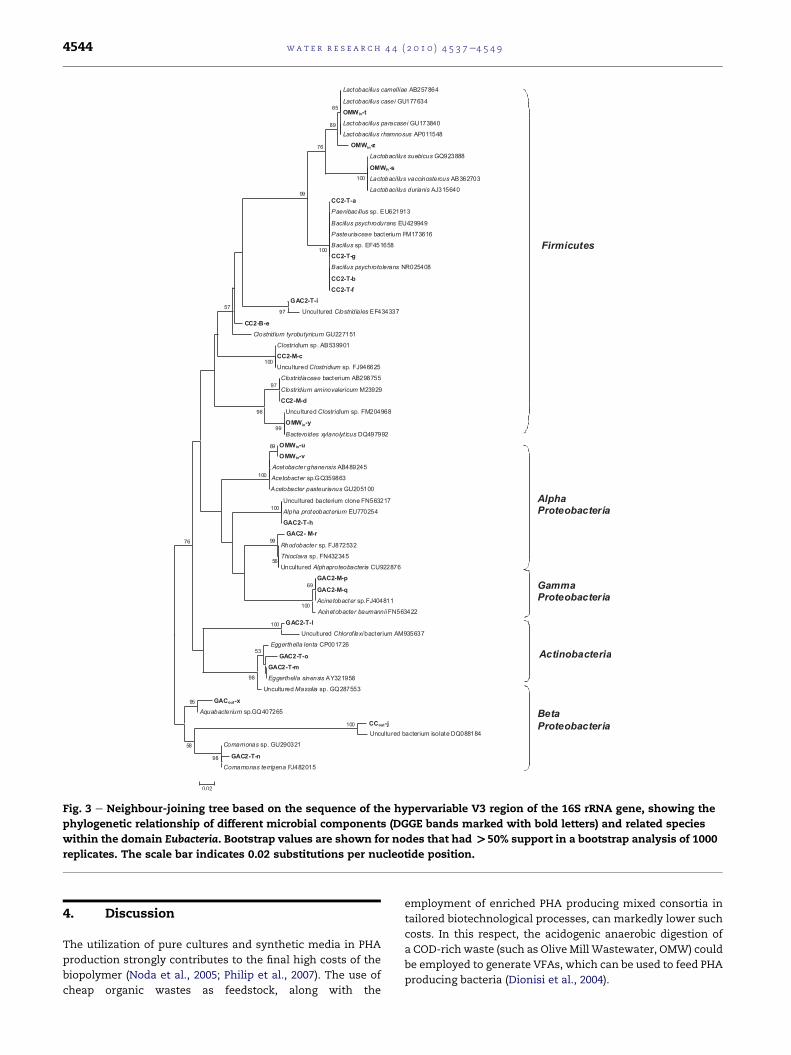
Fig. 3 e Neighbour-joining tree based on the sequence of the hypervariable V3 region of the 16S rRNA gene, showing the
phylogenetic relationship of different microbial components (DGGE bands marked with bold letters) and related species
within the domain Eubacteria. Bootstrap values are shown for nodes that had >50% support in a bootstrap analysis of 1000
replicates. The scale bar indicates 0.02 substitutions per nucleotide position.
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 94544
4. Discussion
The utilization of pure cultures and synthetic media in PHA
production strongly contributes to the final high costs of the
biopolymer (Noda et al., 2005; Philip et al., 2007). The use of
cheap organic wastes as feedstock, along with the
employment of enriched PHA producing mixed consortia in
tailored biotechnological processes, can markedly lower such
costs. In this respect, the acidogenic anaerobic digestion of
a COD-richwaste (such as Olive Mill Wastewater, OMW) could
be employed to generate VFAs, which can be used to feed PHA
producing bacteria (Dionisi et al., 2004).

Fig. 4 e Top: DGGE fingerprints of the archaebacterial communities. Samples collected from top, middle and bottom sections
of the reactors are markedwith T, M and B, respectively. OMWIN refers to the inlet flow, CC2OUT and GAC2OUT to the effluents
discharged under steady state conditions. Roman numerals (I and II) correspond to different replicates. Letters indicate
bands that have been excised, cloned and sequenced. Bottom: Similarity dendrograms indicate relationships among the
different DGGE profiles.
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 9 4545
Freely suspended cell bioreactors were already employed
for producing VFAs from several organic waste and effluents,
including OMWs (Ntaikou et al., 2009). The use of Packed Bed
Biofilm Reactor (PBBR) filled with Ceramic Cubes (CCs) was
also recently proposed by us for the same purpose (Beccari
et al., 2009); in the present study, the same process was
flunked by a GAC-packed technology with identical configu-
ration and both were assessed by evaluating the influence of
temperature and Organic Loading Rate (OLR) on concentration
and relative amounts of produced VFAs.
Packing material played a crucial role in the biofilm bio-
logical activities, as also evidenced by other authors (Ince
et al., 1995; Tay and Show, 1999; Picanco et al., 2001; Yang
et al., 2004). In particular, the biofilm generated on CCs
always gave rise to higher VFA concentrations than those
obtained in the parallel GAC reactors where, on the contrary,
a remarkable methanogenic activity was observed. Moreover,
GAC-PBBRs gave rise to much higher COD depletions, with
potential additional adverse effects on PAH production due to
the influent COD subtraction. Thus, the most favourable
conditions for a subsequent PHA production were achieved
with the CC-based process, where methanogenesis was
negligible or absent at all at 25 �C and no significant COD
removals occurred (Table 2). In particular, the latter evidence
allowed process yields, which link VFA amounts to the
influent COD (equation (2)), closed to the ratios between VFA
and COD concentrations in the effluents (Table 3). Packing
material played also a paramount role in the removal of
polyphenols, which were significantly depleted only in GAC-
PBBRs (Table 2). Since they are known to inhibit mostly
methanogenic populations (Dionisi et al., 2005a), their
persistence in CC-systems may have contributed to their

Table 5 e Taxonomic characterization of themajor bands in the DGGE profiles obtained with primers for Archaea related tothe biofilm samples collected from CC2 and GAC2 and to the last experiment influent flow (OMWIN) and effluentsdischarged under steady state conditions from CC2 (CC2OUT) and GAG2 (GAC2OUT). Different sampling positions along theprofile of the reactor are marked with T (top), M (middle), and B (bottom).
Phylogenetic group Taxon Identity (%)
CC2
CC2B-a Methanobacteria Methanobrevibacter acididurans 99
CC2B-b Methanobacteria Methanobrevibacter sp. 99
GAC2
GAC2T-c Methanobacteria Methanobacterium bryantii 99
GAC2T-d Methanomicrobia Uncultured Methanosarcinales archaeon 99
GAC2T-e Methanomicrobia Methanocella paludicola 97
Flows
OMWIN-f Actinobacteria Atopobium rimae 96
OMWIN-g Methanobacteria Methanobacterium sp. 99
OMWIN-h Methanobacteria Methanobacterium bryantii 99
GAC2OUT-i Methanobacteria Methanobacterium beijingenses 99
GAC2OUT-l Actinobacteria Uncultured Coriobacteriaceae bacterium 97
wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 94546
higher efficiency in OMW acidification. On the contrary,
temperature and OLR did not affect the removal of such
compounds.
In accordance with the lower methanogenic activities,
acidogenic fermentation in CC-PBBRs raised proportionally by
decreasing temperature (experiments No. 1a, 1b and 3) by
achieving the best performance at 25 �C (5.38% of net COD
conversion into VFA, Table 3). Aside temperature, another
parameter typically affecting methanogenesis is OLR (Ince
Fig. 5 e Neighbour-joining tree based on the sequence of the hyp
phylogenetic relationship of different microbial components (D
within the domain Archaebacteria. Bootstrap values are shown f
1000 replicates. The scale bar indicates 0.05 substitutions per nu
here used for Archaea efficaciously functioned even in amplifica
et al., 1995; Grover et al., 2001; Kennedy et al., 2006). Meth-
anogenesis generally decreased when higher OLRs were
applied (Table 2). However, temperature played a key role in
determining to which extent this occurred, being at 25 �Cobtained the best results in term of methanogenesis reduc-
tion: once again, enhancements achieved with CCs support
were much greater than with GAC: the net COD conversion
into VFAs was one order of magnitude higher than what
previously obtained (up to about 66%), leading to a whole
ervariable V2-V3 region of the 16S rRNA gene, showing the
GGE bands marked with bold letters) and related species
or nodes that had >50% support in a bootstrap analysis of
cleotide position. It is worth noting that the specific primers
tion of 16S rRNA gene of Actinobacteria.

wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 9 4547
process yield of about 82% (experiments No. 4, Table 3). Such
performances, which are the best among those obtained in
this study, were significantly better than those achieved
during the previous study performed with the same CC-based
technology and at the same temperature (25 �C) (Beccari et al.,2009), where the net COD conversion into VFAs and process
yield were about 13 and 29%, respectively. On the other hand,
when the OLR was further increased (experiment No. 5),
acidogenic fermentation was inhibited and the net COD
conversion into VFAs dropped back (Table 3). The high
concentration of toxic polyphenols present in the wastewater
could have contributed to this evidence (Kennedy et al., 2006)
together with the inability of CCs, differently from GAC, to
adsorb/desorb aromatic compounds, thus “buffering” such
toxic effects (Bertin et al., 2004). However, the results related
to the fifth experiment were closed to the ones achieved in the
previous study, which was carried out under similar loading
conditions (Beccari et al., 2009). Thus, even the previously
developed process was probably negatively influenced by
a high OLR.
The employment of the CC-PBBR technology allowed also
to obtain a higher OMW acidification with respect to that of
a conventional CSTR fed with different OLRs (Ntaikou et al.,
2009). In particular, the latter system higher process yield
was about 30%, due to an influent COD of 19.5 g/L fed at about
14 g/L/day and to a total effluent VFA amount of about
6 gCOD/L. Furthermore, the CC-based system performances
were higher than those of the study of Dionisi et al. (2005a),
where OMW was pre-treated by means of bentonite amend-
ment (even followed by centrifugation) and fermented under
batch conditions: in that case, comparable total VFA amounts
were achieved but with a high initial COD, this resulting
in lower process yields (higher process yield: 43.6%, corre-
sponding to an initial COD of 28.5 g/L and to a total final VFA
amount of 12.4 g/L).
The relative composition of the VFA mixture was mainly
affected by the parameter OLR: in particular, CC-biofilms
induced a decrease in the amount of acetate both at 35
(experiments No. 1b and 2) and 25 �C (experiments No. 3, 4 and
5) (Fig. 1a); within the experiments performed with CC2 at
25 �C, it was also reported a relative increase in the propionate
content while butyrate amount was maintained constant.
Thus, the OMW acidogenic digestion carried out in a CC-PBBR
operating at 25 �C (i.e. the best performing conditions) seems
to allow to control the acetateepropionate relative amounts
by regulating the applied OLR; however, the process optimi-
zation should take into consideration that the higher total VFA
production would not correspond to the higher propionate
production. On the contrary, lower hydraulic retention times
often led to a reduction of propionic acid, whose highest
production corresponded to the highest OMW acidification
(Ntaikou et al., 2009; Bengtsson et al., 2008b); however, the
opposite effect was also reported (Dinopoulou et al., 1988).
Themicrobiological investigations, regarding themicrobial
structure of both the biofilms within the two differently
packed bioreactors and the wastewaters, clearly indicated
a different speciation of either Bacteria and Archaea
depending on the two different support materials. This seems
to be in accordance with results concerning the performances
of the two bioreactors. In fact, the bacterial community that
has been selected within CC2 is exclusively composed by Fir-
micutes such as Clostridium, Bacillus, and strains belonging to
the Pasteuriaceae family. Actually, several bacterial species
belonging to these genera are known to show acidogenic
activity (Akao et al., 2007). In particular, some of the major
bands in the DGGE profiles (Fig. 2, bands c, d, e) revealed the
presence of strains closely related to C. tyrobutiricum as well as
Clostridium aminovalericum (Fig. 3). C. tyrobutiricum has been
shown to produce butyric and acetic acid as its major
fermentation products from glucose and xylose (Liu et al.,
2006). On the other hand, C. aminovalericum is capable to
anaerobically degrade 5-aminovalerate to valerate, acetate,
propionate, and ammonia (Barker et al., 1987).
Conversely, the bacterial community selected within the
GAC2 was mainly composed by Proteobacteria. Among the
different major bands in the DGGE profile, Acinetobacter is well
represented (Fig. 2, bands p, q). Nevertheless bacteria strains
belonging to this genus are common members of microbial
consortia involved in the biodegradation of biogenic and
xenobiotic compounds, and its high potential for the treat-
ment of phenol-containing wastewaters has been recently
elucidated (Liu et al., 2009).
As far as the Archaea speciation is concerned, the occur-
rence of species belonging mainly to the genus Methano-
brevibacter in the bioreactor packed with ceramic support is
consistent with the environmental conditions settled in such
digester. Striking VFA concentrations were in fact observed in
CC-PBBRs where acidic pHs (w5.0) might have favoured
methanogens to become well established, among those more
acidophilic (Savant et al., 2002; Rea et al., 2007). Moreover, as
Methanobrevibacter grows on a H2/CO2 gas mixture or, in
addition, can utilize formate, strict acidogenic conditions
such as those in CC-PBBRs are coherent with the activity of
possible consortia of incoming hydrogen-producing ace-
togens and this hydrogenotrophic methanogen (Fang, 2000).
Furthermore, the archaeal composition of the CC2 biofilm
was strongly closed to the one of the influent wastewater,
this suggesting that the immobilization of a large spectrum of
methanogenic consortia was not favoured in such a reactor.
The opposite situation found in GAC2, whereas a high
methanogenic activity was observed, seems to support the
latter hypothesis: the presence of mostly Methanomicrobia is
in agreement either with the pH conditions fluctuating nearly
around neutrality (w6.8), better conducive to the acclimation
of this kind of methanogens (Kendall and Boone, 2006), or
with the particular tendency of GAC to be colonized espe-
cially by Methanosarcinales (Schmidt and Ahring, 1999).
Interestingly, the sole member of Methanobacteria found in
GAC2, namely Methanobacterium bryantii, is a species which
shows an optimum pH range for growth between 6.9 and 7.2
(Boone, 1987). This picture fits well with recent findings on
bacterial speciation in GAC-PBBRs treating OMWs (Rizzi et al.,
2006).
5. Conclusions
An effective OMW acidification process was developed
through the employment of a PBBR technology which
employed porous ceramic cubes for supporting the biofilm

wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 94548
generation. The highest COD conversion into VFAs was
observed when the reactor was thermostated at 25 �C and
loaded with an OLR of about 13 g/L/day, conditions under
which a total VFA concentration of about 14 g/L was ach-
ieved, this corresponding to about the 88 and the 82% of the
influent and effluent COD, respectively. The relative
amounts of the main VFAs can be controlled by regulating
the applied OLR, and this is of particular interest in the
perspective of feeding the acidogenic effluent to a biotech-
nological PHA producing process. The packing material
appeared as a crucial parameter able to influence the
process performances much more than temperature and
OLR, and this by greatly affecting the biofilm bacterial
community structure.
Acknowledgements
The authors thank the Olive mills Sant’Agata d’Oneglia
(Imperia, Italy) and Grassanese (Matera, Italy) for having
provided the OMWs. This research was financially supported
by the Italian Ministry of University and Research (PRIN 2005)
and partially by the Fondazione Del Monte di Bologna e Rav-
enna (Bologna, Italy).
r e f e r e n c e s
Albuquerque,M.G.E., Eiroa,M., Torres, C.,Nunes, B.R., Reis,M.A.M.,2007. Strategies for the development of a side stream processfor polyhydroxyalkanoate (PHA) production from sugar canemolasses. J. Biotechnol. 130, 411e421.
Akao, S., Tsuno, H., Horie, T., Mori, S., 2007. Effects of pH andtemperature on products and bacterial community in L-lactatebatch fermentation of garbage under unsterile condition.Water Res. 41 (12), 2636e2642.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z.,Miller, W., Lipman, D.J., 1997. Gapped BLAST and PSI-BLAST:a new generation of protein database search programs.Nucleic Acids Res. 25, 3389e3402.
Barker, H.A., D’Ari, L., Kahn, J., 1987. Enzymatic reactions in thedegradation of 5-aminovalerate by Clostridium aminovalericum.J. Biol. Chem. 262 (19), 8994e9003.
Beccari, M., Bertin, L., Dionisi, D., Fava, F., Lampis, S., Majone, M.,Valentino, F.,Vallini, G.,Villano,M., 2009. Exploitingoliveoilmilleffluents as a renewable resource for production ofbiodegradablepolymers throughacombinedanaerobiceaerobicprocess. J. Chem. Technol. Biotechnol. 84, 901e908.
Bengtsson, S., Werker, A., Christensson, M., Welander, T., 2008a.Production of polyhydroxyalkanoates by activated sludgetreatingpapermillwastewater.Bioresour.Technol. 99,509e516.
Bengtsson, S., Hallquist, J., Werker, A., Welander, T., 2008b.Acidogenic fermentation of industrial wastewaters: effect ofchemostat retention time and pH on volatile fatty acidsproduction. Biochem. Eng. J. 40, 492e499.
Bertin, L., Berselli, S., Fava, F., Petrangeli-Papini, M., Marchetti, L.,2004. Anaerobic digestion of olive mill wastewaters in biofilmreactors packed with granular activated carbon and“Manville” silica beds. Water Res. 38, 3167e3178.
Boone, D.R., 1987. Replacement of the type strain ofMethanobacterium formicicum and reinstatement ofMethanobacterium bryantii sp. nov. nom. rev. (ex Balch and
Wolfe, 1981) with M.o.H. (DSM 863) as the type strain. Int. J.Syst. Bacteriol. 37, 172e173.
Braunegg, G., Lefebvre, G., Genser, K.F., 1998.Polyhydroxyalkanoates, biopolyesters from renewableresources: physiological and engineering aspects. J.Biotechnol. 65, 127e161.
Chua, A.S.M., Takabatake, H., Satoh, H., Mino, T., 2003. Productionof polyhydroxyalkanoates (PHA) by activated sludge treatingmunicipal wastewater: effect of pH, sludge retention time(SRT), and acetate concentration in influent. Water Res. 37(15), 3602e3611.
Dionisi, D., Carucci, G., Petrangeli-Papini, M., Riccardi, C.,Majone, M., Carrasco, F., 2005a. Olive oil mill effluents asa feedstock for production of biodegradable polymers. WaterRes. 39, 2076e2084.
Dionisi, D., Beccari, M., Di Gregorio, S., Majone, M., Petrangeli-Papini, M., Vallini, G., 2005b. Storage of biodegradablepolymers by an enriched microbial community ina sequencing batch reactor operated at high organic load rate.J. Chem. Technol. Biotechnol. 8, 1306e1318.
Dionisi, D., Majone, M., Papa, V., Beccari, M., 2004. Biodegradablepolymers from organic acids by using activated sludgeenriched by aerobic periodic feeding. Biotechnol. Bioeng. 85(6), 569e579.
Dionisi, D., Majone, M., Vallini, G., Di Gregorio, S., Beccari, M.,2006. Effect of the applied organic load rate on biodegradablepolymer production by mixed microbial cultures ina sequencing batch reactor. Biotechnol. Bioeng. 93, 76e88.
Dinopoulou, G., Rudd, T., Lester, N.J., 1988. Anaerobicacidogenesis of a complexwastewater: I. The influence ofoperational parameters on reactor performance. Biotechnol.Bioeng 31, 958e968.
Fang, H.H.P., 2000. Microbial distribution in UASB granules and itsresulting effects. Water Sci. Technol. 42 (12), 201e208.
Folin, O., Ciocalteu, V., 1927. On tyrosine and tryptofandeterminations in protein. J. Biol. Chem. 73, 627e650.
Grosskopf, R., Janssen, P.A., Liesack, W., 1998. Diversity andstructure of the methanogenic community in anoxic ricepaddy soil microcosms as examined by cultivation and direct16S rRNA gene sequence retrieval. Appl. Environ. Microbiol.64, 960e969.
Grover, R., Marwaha, S.S., Kennedy, J.F., 2001. Methanogenesis ofblack liquor in a two-stage biphasic reactor system using animmobilized cell system. J. Chem. Technol. Biotechnol. 76,251e256.
Ince, O., Anderson, G.K., Kasapgil, B., 1995. Control of organicloading rate using the specific methanogenic activity testduring start-up of an anaerobic digestion system. Water Res.29 (1), 349e355.
Kendall, M.L., Boone, D.R., 2006. The order Methanosarcinales. In:Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H.,Stackebrandt, E. (Eds.), The ProkaryotesArchaea. Bacteria:Firmicutes, Actinomycetes, third ed., vol. 3. Springer, NewYork, pp. 244e256.
Kumar, S., Nei, M., Dudley, J., Tamura, K., 2008. MEGA: a biologist-centric software for evolutionary analysis of DNA and proteinsequences. Brief. Bioinformatics 9, 299e306.
Kennedy, J.F., Panesar, P.F., Grover, R., Marwaha, S.S., 2006.Continuous methanogenesis of black liquor of pulp and papermills in an anaerobic baffled reactor using an immobilized cellsystem. J. Chem. Technol. Biotechnol. 81, 1277e1281.
Khanna, S., Srivastava, A.K., 2005. Recent advances in microbialpolyhydroxyalkanoates. Process Biochem. 40, 607e619.
Lane, D.J., 1991. 16S/23S rRNA sequencing. In: Stackebrandt, E.,Goodfellow, M. (Eds.), Nucleic Acid Techniques in BacterialSystematics. Wiley, Chichester, United Kingdom, pp. 115e175.
Lee, S.Y., 1996. Bacterial polyhydroxyalkanoates. Biotechnol.Bioeng. 49, 1e14.

wat e r r e s e a r c h 4 4 ( 2 0 1 0 ) 4 5 3 7e4 5 4 9 4549
Liu, Y.J., Nikolausz, M., Wang, X.C., 2009. Biodegradation anddetoxication of phenol by using free and immobilized cells ofAcinetobacter sp. XA05 and Sphingomonas sp. FG03. Environ. Sci.Health A Tox. Hazard. Subst. Environ. Eng. 44 (2), 130e136.
Liu, X., Zhu, Y., Yang, S.T., 2006. Butyric acid and hydrogenproduction by Clostridium tyrobutyricum ATCC 25755 andmutants. Enzyme Microb. Technol. 38 (3e4), 521e528.
Miller, D.G., Brayton, S.V., Boyles, W.T., 2001. Chemical oxygendemand analysis of wastewater using trivalent manganeseoxidant with chloride removal by sodium bismuthate pre-treatment. Water Environ. Res. 73 (1), 63e71.
Mochizuki, M., 2002. Properties and application of aliphaticpolyester products. In: Steinbuchel, A., Doi, Y. (Eds.),Biopolymers, vol. 4. Wiley-vch, Weinheim.
Noda, I., Green, P.R., Satkowski, M.M., Schectman, L.A., 2005.Preparation and properties of a novel class ofpolyhydroxyalkanoate copolymers. Biomacromolecules 6,580e586.
Ntaikou, I., Kourmentza, C., Koutrouli, E.C., 2009. Exploitation ofolive oil mill wastewater for combined biohydrogen andbiopolymers production. Biores. Technol. 100 (15), 3724e3730.
Philip, S., Keshavarz, T., Roy, I., 2007. Polyhydroxyalkanoates:biodegradable polymers with a range of applications. J. Chem.Technol. Biotechnol. 82, 233e247.
Picanco, A.P., Vallero, M.V.G., Gianotti, E.P., Zaiat, M., Blundi, C.E., 2001. Influence of porosity and composition of supportson the methanogenic biofilm characteristics developed ina fixed bed anaerobic reactor. Water Sci. Technol. 44,197e204.
Rea, S., Bowman, J.P., Popovski, S., Pimm, C., Wright, A.D.G., 2007.Methanobrevibacter millerae sp. nov. and Methanobrevibacterolleyae sp. nov., methanogens from the ovine and bovinerumen that can utilize formate for growth. Int. J. Syst. Evol.Microbiol. 57, 450e456.
Reddy, C.S.K., Ghai, Rashmi, R., Kalia, V.C., 2003.Polyhydroxyalkanoates: an overview. Bioresour. Technol. 87,137e146.
Reis, M.A.M., Serafim, L.S., Lemos, P.C., Ramos, A.M., Aguiar, F.R.,Van Loosdrecht, M.C.M., 2003. Production of
polyhydroxyalkanoates by mixed microbial cultures.Bioprocess. Biosyst. Eng. 25 (6), 377e385.
Rhu, D.H., Lee, W.H., Kim, J.Y., Choi, E., 2003.Polyhydroxyalkanoate (PHA) production from waste. WaterSci. Technol. 48 (8), 221e228.
Rizzi, A., Zucchi, M., Borin, S., Marzorati, M., Sorlini, C.,Daffonchio, D., 2006. Response of methanogen populations toorganic load increase during anaerobic digestion of olive millwastewater. J. Chem. Technol. Biotechnol. 81, 1556e1562.
Roest, K., Heilig, H.G.H.J., Smidt, H., de Vos, W.M., Stams, A.J.M.,Akkermans, A.D.L., 2005. Community analysis of a full-scaleanaerobic bioreactor treating paper mill wastewater. Syst.Appl. Microbiol. 28, 175e185.
Salmiati, Z., Ujang, M.R., Salim, M.F., Din, M.D., Ahmad, M.A.,2007. Intracellular biopolymer productions using mixedmicrobial cultures from fermented POME. Water Sci. Technol.56 (8), 179e185.
Savant, D.V., Shouche, Y.S., Prakash, S., Ranade, D.R., 2002.Methanobrevibacter acididurans sp. nov., a novel methanogenfrom a sour anaerobic digester. Int. J. Syst. Evol. Microbiol. 52,1081e1087.
Schmidt, J.E., Ahring, B.K., 1999. Immobilization patterns anddynamics of acetate-utilizing methanogens immobilized insterile granular sludge in upflow anaerobic sludge blanketreactors. Appl. Environ. Microbiol. 65 (3), 1050e1054.
Serafim, L.S., Lemos, P.C., Albuquerque, M.G.E., Reis, M.A.M.,2008. Strategies for PHA production by mixed cultures andrenewable waste materials. Appl. Microbiol. Biotechnol. 81,615e628.
Tay, J.,Show,K.,1999. Influenceofsupportmediaonbiomassgrowthand retention in anaerobic filters. Water Res. 33, 1471e1481.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F.,Higgins, D.G., 1997. The CLUSTAL_X windows interface:flexible strategies for multiple sequence alignment aided byquality analysis tools. Nucleic Acids Res. 25, 4876e4882.
Yang, Y., Tada, C., Shohidullah Miah, M., Tsukahara, K.,Yagishita, T., Sawayama, S., 2004. Influence of bed materialson methanogenic characteristics and immobilized microbesin anaerobic digester. Mater. Sci. Eng. 24, 413e419.





























