Embed Size (px)
Citation preview

DISEASES OF AQUATIC ORGANISMSDis Aquat Org
Preprint, 2009doi: 10.3354/dao02134
Published online November 30, 2009
© Inter-Research 2009 · www.int-res.com*Email: [email protected]
Amphibian chytrid fungus and ranaviruses in theNorthwest Territories, Canada
Danna M. Schock1,*, Gregory R. Ruthig2, 8, James P. Collins2, Susan J. Kutz1,Suzanne Carrière3, Robert J. Gau3, Alasdair M. Veitch4, Nicholas C. Larter5,
Douglas P. Tate6, Glen Guthrie7, Daniel G. Allaire5, Richard A. Popko4
1Department of Ecosystem and Public Health, Faculty of Veterinary Medicine, University of Calgary, 3330 Hospital Drive NW,Calgary, Alberta T2N 4N1, Canada
2School of Life Science, Arizona State University, Tempe, Arizona 85287-4501, USA3Department of Environment & Natural Resources, Government of the Northwest Territories, PO Box 1320, Yellowknife,
Northwest Territories X1A 2L9, Canada4Department of Environment & Natural Resources, Government of the Northwest Territories, PO Box 130, Norman Wells,
Northwest Territories X0E 0V0, Canada5Department of Environment & Natural Resources, Government of the Northwest Territories, PO Box 240, Fort Simpson,
Northwest Territories X0E 0N0, Canada6Nahanni National Park Reserve, Parks Canada, 10002 – 102 St, Fort Simpson, Northwest Territories X0E 0N0, Canada
7Sahtu Renewable Resources Board, PO Box 134, Tulita, Northwest Territories X0E 0N0, Canada
8Present address: Department of Biology, Grinnell College, 1116 8th Ave, Grinnell, Iowa 50112-1690, USA
ABSTRACT: Pathogens can cause serious declines in host species, and knowing where pathogensassociated with host declines occur facilitates understanding host-pathogen ecology. Suspected dri-vers of global amphibian declines include infectious diseases, with 2 pathogens in particular, Batra-chochytrium dendrobatidis (Bd) and ranaviruses, causing concern. We explored the host range andgeographic distribution of Bd and ranaviruses in the Taiga Plains ecoregion of the Northwest Territo-ries, Canada, in 2007 and 2008. Both pathogens were detected, greatly extending their known geo-graphic distributions. Ranaviruses were widespread geographically, but found only in wood frogs. Incontrast, Bd was found at a single site, but was detected in all 3 species of amphibians in the surveyarea (wood frogs, boreal chorus frogs, western toads). The presence of Bd in the Northwest Territo-ries is not congruent with predicted distributions based on niche models, even though findings fromother studies at northern latitudes are consistent with those same models. Unexpectedly, we alsofound evidence that swabs routinely used to collect samples for Bd screening detected fewer infec-tions than toe clips. Our use and handling of the swabs was consistent with other studies, and thecause of the apparent lack of integrity of swabs is unknown. The ranaviruses detected in our studywere confirmed to be Frog Virus 3 by sequence analysis of a diagnostic 500 bp region of the majorcapsid protein gene. It is unknown whether Bd or ranaviruses are recent arrivals to the Canadiannorth. However, the genetic analyses required to answer that question can inform larger debatesabout the origin of Bd in North America as well as the potential effects of climate change and indus-trial development on the distributions of these important amphibian pathogens.
KEY WORDS: Ranavirus · Batrachochytrium dendrobatidis · Amphibian declines · Rana sylvatica ·Pseudacris maculata · Bufo boreas · Nahanni National Park Reserve · Taiga Plains
Resale or republication not permitted without written consent of the publisher
Contribution to DAO Special 4 ‘Chytridiomycosis: an emerging disease’

Dis Aquat Org: Preprint, 2009
INTRODUCTION
Pathogens are capable of causing serious populationdeclines in hosts (Smith et al. 2009). Two pathogens, achytrid fungus (Batrachochytrium dendrobatidis; Bd)and ranaviruses, are among the suspected drivers ofglobal amphibian declines (Collins & Storfer 2003,Daszak et al. 2003, Stuart et al. 2004, McCallum 2007),and both were listed as notifiable pathogens by theWorld Organisation for Animal Health (2008). Bd islinked to catastrophic declines in several parts of theworld, but its origin is unclear (Rachowicz et al. 2005,Skerratt et al. 2007). In many regions, patterns of dis-ease and declines suggest that Bd is a novel pathogenthat sweeps through naïve host species, reducing thenumber of amphibian species, and then later becomesestablished in areas with suitable environmental con-ditions and sufficient host numbers (Retallick et al.2004, Lips et al. 2006, 2008, Skerratt et al. 2007). Popu-lation genetic data for Bd also supports this hypothesis(Morehouse et al. 2003, Morgan et al. 2007) althoughthis is an important area of ongoing research. Alterna-tively, Bd may be a historically widely distributedorganism, and recent declines associated with Bd maybe linked to large-scale environmental changes thatincrease the impact of Bd on host populations (Ouelletet al. 2005, Pounds et al. 2006). Regardless of the ulti-mate source of Bd, this pathogen is now on every con-tinent where amphibians reside (Berger et al. 1998,Weldon et al. 2004, Bosch et al. 2007, Longcore et al.2007, Kusrini et al. 2008, Lips et al. 2008).
Ranaviruses (family Iridoviridae) infect fish, reptiles,and amphibians, and are of considerable concern inaquaculture (see Chinchar 2002 and Williams et al.2005 for recent reviews of ranavirus biology). Lethalamphibian ranaviruses infect a wide range of species,and die-offs have been documented world-wide,including die-offs associated with ranaculture opera-tions (Zhang et al. 2001, Miller et al. 2007). Amphibianranaviruses isolated from widely distributed, abundantspecies such as barred tiger salamanders Ambystomamavortium and wood frogs Rana sylvatica = Lithobatessylvaticus can cause lethal infections in other amphib-ian species (Jancovich et al. 2001, Schock et al. 2008).Thus, ranaviruses may be maintained in populations ofabundant species that can serve as sources of infectionfor rare species that share the same habitat. In addi-tion, die-offs caused by ranaviruses can severelyreduce numbers of amphibians at breeding sites andtherefore may affect amphibian populations in highlyfragmented habitats where recolonization is unlikely(Collins et al. 2003).
Knowledge of where pathogens associated with hostdeclines occur, and under what circumstances, isessential to understanding their ecology. However,
host–pathogen systems are often complex and difficultto tease apart. The relatively low levels of biodiversityat higher latitudes create opportunities to use northernamphibian populations as a model to study the ecologyof pathogens implicated in declines. We conductedsurveys for Bd and ranaviruses in the Northwest Terri-tories (NT), Canada, to determine the geographic dis-tributions, prevalences, and host ranges of thesepathogens. We found both pathogens in the NT anddiscuss the importance of our results to the ecology ofthese pathogens.
MATERIALS AND METHODS
Study areas and amphibian species. We focused onthe western portion of the NT (Fig. 1), within the taigaplains ecoregion (Ecosystem Classification Group2007). Three amphibian species occur in our studyarea: wood frogs, western toads Bufo boreas =Anaxyrus boreas, and boreal chorus frogs Pseudacrismaculata. All 3 species have large geographic rangesthat include a variety of habitats in Canada and theUSA (Fournier 1997, Russell & Bauer 2000, Stebbins2003). Western toads are federally listed as a species ofSpecial Concern in Canada and listed as Endangeredin several US states (NatureServe 2009). Boreal chorusfrogs are considered stable throughout their range(NatureServe 2009). Although wood frogs appear to bestable throughout most of their range, they have beenextirpated from Idaho, and populations are decliningin Wyoming (NatureServe 2009).
Bd has been detected in all 3 amphibian specieselsewhere in their respective ranges. In wood frogs, Bdhas been reported from Alaska (Reeves & Green 2006,Reeves 2008), British Columbia (Ouellet et al. 2005),Colorado (Rittman et al. 2003, Green & Muths 2005,Young et al. 2007), Wyoming (Young et al. 2007),Michigan (Zellmer et al. 2008), Quebec (Ouellet et al.2005), and Maine (Longcore et al. 2007). In westerntoads, Bd has been reported in British Columbia(Raverty & Reynolds 2001, Adams et al. 2007) and inColorado and Wyoming, where populations havedeclined greatly in the past 3 decades (Green et al.2002, Green & Muths 2005, Muths et al. 2003, Young etal. 2007). Pearl et al. (2007) did not detect Bd in west-ern toads during their survey in Oregon and Washing-ton, but only 13 toads were tested and the pathogenwas found in other species in the same areas duringtheir study. Until recently, boreal chorus frogs andwestern chorus frogs Pseudacris triseriata were consid-ered subspecies of a larger Pseudacris species complex(Moriarty & Lannoo 2005, Lemmon et al. 2007).Because various reports of Bd have used different spe-cies names, reports of Bd in P. triseriata from western
2
Schock et al.: Amphibian pathogens in the Northwest Territories
North America as well as P. maculata are of interest.Bd has been detected in Pseudacris sp. in Colorado(Rittman et al. 2003, Green & Muths 2005, Young et al.2007), Arizona (Retallick & Miera 2007), and Wyoming(Young et al. 2007).
Ranaviruses have been isolated from wood frog pop-ulations, generally from die-offs, in Saskatchewan(Schock et al. 2008), Ontario (Greer et al. 2005, Duffuset al. 2008), North Carolina (Harp & Petranka 2006),and in North Dakota, Maine, and Massachusetts(Green et al. (2002). Duffus et al. (2008) reported that aranavirus was detected in Pseudacris tadpoles inOntario. However, no PCR product was sequenced intheir study so the identity of the putative ranavirus isunknown. Green & Muths (2005) did not detectranavirus infections in Pseudacris in their study in Col-orado. Similarly, numerous Pseudacris tadpoles andadults from multiple sites in Saskatchewan (D. M.
Schock unpubl. data) and New York (J. L. Brunnerpers. comm.) have been screened, but no Pseudacrisindividuals have tested positive thus far. We are notaware of any reports of ranaviruses in western toads.
Field surveys and collection of samples. Surveyswere conducted from 25 June to 14 July in 2007 and16 June to 5 July in 2008. We looked and listened foramphibians while walking through apparently suit-able habitats (e.g. wetlands, meadows), along cut-linesnear apparently suitable habitat, and while walkingand dip-netting along the perimeters of apparentlysuitable breeding sites. In both years, most surveyedsites were accessible by road or short hikes from roads(<2 km) for logistical reasons. Some, however, wereaccessible only by helicopter and/or boat. The 2008survey in Nahanni National Park Reserve was distinctin terms of accessibility, as it involved rafting down theSouth Nahanni River from Kraus Hot Springs to the
3
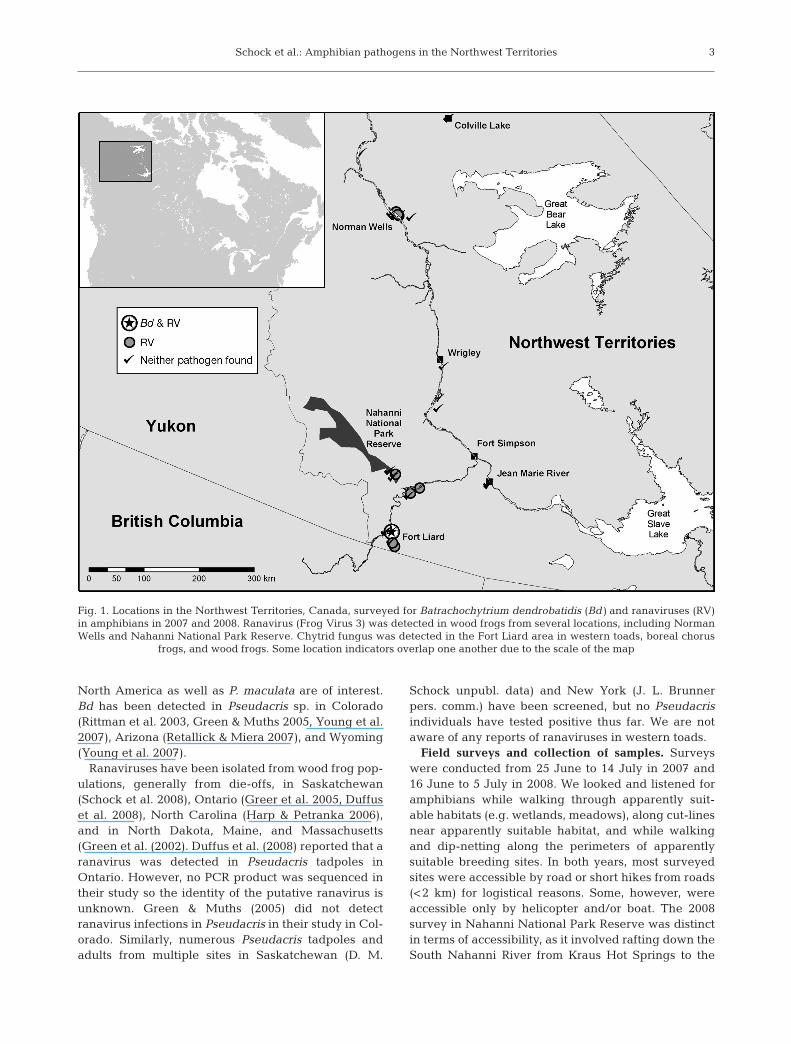
Fig. 1. Locations in the Northwest Territories, Canada, surveyed for Batrachochytrium dendrobatidis (Bd) and ranaviruses (RV)in amphibians in 2007 and 2008. Ranavirus (Frog Virus 3) was detected in wood frogs from several locations, including NormanWells and Nahanni National Park Reserve. Chytrid fungus was detected in the Fort Liard area in western toads, boreal chorus
frogs, and wood frogs. Some location indicators overlap one another due to the scale of the map

Dis Aquat Org: Preprint, 2009
Reserve boundary, banking at several spots along theriver where it appeared that amphibian breeding habi-tat might be accessible (e.g. openings in tree canopiessuggestive of ponds in clearings, sandy areas withshallow ponds cut off from the river or small streams).
We captured up to 61 individuals of each life stage ofeach species at each site to collect basic information(e.g. length, weight, stage of development; D. M.Schock unpubl. data), and to non-lethally collect tissueand/or swab samples for pathogen screening. Not allindividuals that were encountered were captured, thusnumbers presented here do not reflect total numbersencountered. Tissue samples were collected by cuttinga single hind toe from frogs, or a small (≤5 mm) piece oftail tip from tadpoles, using a new blade and newgloves for each animal. Tissue samples were stored in-dividually in 1 ml of 70% chemical grade ethanol untilprocessing at the lab. Most sampled individuals werealso swabbed for Bd following Hyatt et al. (2007). Asterile swab (MW100 tube dry swabs, Medical Wire &Equipment) was gently and repeatedly run across theanimal’s ventral surface, legs, and toe webbing, or inthe case of tadpoles, the mouthparts were gentlyswabbed with a swirling motion. These are areas of thebody most likely to be infected with Bd zoosporangia(Pessier et al. 1999, Berger et al. 2005). The swabs werestored in sealed freezer bags, grouped according tosite, in the dark, at 4 to 22°C for 4 mo (2007) or 1 mo(2008) until they could be shipped to the lab, wherethey were frozen at –20°C for 8 mo (2007) or <1 mo(2008) when they were screened.
Concerted effort was made to prevent accidentalspread of pathogens within and among sites. First,amphibians were individually held in new plasticZiploc® bags or individual plastic containers from thetime of capture until release a short time later. Bagswere full of air plus sufficient moisture/water for thelife stage, and all containers and bags were kept in theshade. Second, new latex gloves, blades, and swabswere used for each animal when taking samples.Finally, equipment that came into contact with animals(e.g. buckets and nets) was bleached, rinsed, and usu-ally dried between sites. Multiple sets of nets andbuckets were carried at all times. The use of powderedbleach (16% dry bleach, Zep Manufacturing) wasinstrumental in the disinfection process, particularly insituations where weight or bulk of gear was of concern.The bleach powder was mixed with a bucket of pondwater and the equipment was disinfected. Equipmentwas rinsed using either potable water brought in con-tainers, or in some instances, with water from the nextsite visited — that is, bleached equipment was taken tothe next site and then doused with buckets of waterfrom that site. In this manner, animals were only everexposed to water from their own sites. In all instances
of use in the field, bleach water was disposed of on theroad or on flat rocky areas devoid of vegetation whereit could evaporate and degrade. No animals came intocontact with bleach water and no bleach water drainedinto ponds.
PCR-based pathogen detection and identification.DNA was extracted from tissue samples (toe, tail) usinga salt extraction method (Sambrook & Russell 2001),whereas DNA was extracted from swabs using Prep-Man Ultra® (Applied Biosystems) in accordance withthe manufacturer’s instructions. Tissue and swab DNAextractions were kept separate and used separately inscreening assays; thus, for most animals, there were 2separate DNA sources.
To screen samples for Bd, we used quantitative PCR(qPCR) following the protocol of Boyle et al. (2004).Samples were run in duplicate on a 384-well opticalPCR plates on an ABI Prism 7900 Sequence Detectionsystem (Applied Biosystems). Animals were scored aspositive if the reactions in both wells amplified the tar-get DNA sequence prior to 40 amplification cycles.Samples where there was no amplification in eitherwell were scored as negative for Bd. Samples were re-run in duplicate if there was amplification in only 1well, or evidence of amplification after 40 cycles. Inmost instances, the results of the re-run were conclu-sive and animals could reliably be scored as positive ornegative. In the small number of instances where therewas any doubt, animals were scored as negative.
While confirming all Bd positives and several nega-tives by re-running a subset of samples, it was noticedthat some animals tested negative for Bd when DNAfrom swab extractions was used whereas they testedpositive when DNA from tissue extractions was used.As a consequence, all animals from the Fort Liardregion (both years), Norman Wells (both years), and 20additional animals per year from elsewhere werescreened multiple times, using DNA from tissues aswell as from swabs. Disagreement between sourcesof DNA occurred for 3 animals from the Fort Liardregion — in these cases, swabs consistently testednegative while toe tissues consistently tested positive.These 3 animals were scored as positive for Bd.
Animals were assayed for ranaviruses using the DNAextracted from tissues and the PCR methods describedby Schock et al. (2008). Briefly, genetic markers wereused that target a conserved ~500 base pair (bp) regionof the major capsid protein (MCP) gene, the MCP4/5markers (Mao et al. 1996). PCR products were treatedwith SYBR Green nucleic acid stain (Invitrogen) ac-cording to the manufacturer’s directions and then visu-alized by electrophoresis on 1.5% agarose gels usingan ultraviolet transilluminator (BioRad). A PCR productof ~500 bp indicated that an individual was positive forranavirus, whereas no PCR band indicated no infection.
4

Schock et al.: Amphibian pathogens in the Northwest Territories
All PCR products that produced a 500 bp band wereprepared for sequencing with ExoSAP-IT® (USB) ac-cording to manufacturer directions and sequenced onan ABI 3730 automated capillary column sequencer(Applied Bio-Systems). Sequences were aligned by eyeand analyzed using MEGA version 4.0 (Tamura et al.2007) and then compared to the MCP sequences ofother ranaviruses using the BLAST tool from the Na-tional Center for Biotechnology Information (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
Stata 10 (StataCorp) was used for statistical analyses.Confidence intervals (CIs) were calculated using thebinomial-exact platform.
RESULTS
In total, 396 wood frogs (223 tadpoles, 173 frogs), 61boreal chorus frogs (51 tadpoles, 10 frogs), and 99western toads (95 tadpoles, 4 frogs) were screened forBd and ranaviruses (Table 1). Western toads wereencountered only in the Fort Liard area. Only woodfrogs were encountered at any of the sites north ofWrigley.
Bd was detected at 1 site near Fort Liard (Table 1;Fig. 1), and was detected in all 3 amphibian species.This site was visited in 2007 and 2008, and Bd wasdetected in both years. We did not notice any grosssigns of Bd infection such as skin sloughing in frogs(Pessier et al. 1999, Berger et al. 2005) or abnormalmouth parts in tadpoles (Knapp & Morgan 2006 andreferences therein, but see Padgett-Flohr & Goble2007).
Ranaviruses were detected in wood frogs at 3 sitesnear Fort Liard, 1 site near Blackstone, 1 site inNahanni National Park Preserve, and 4 sites in theNorman Wells area (Table 1; Fig. 1). Ranavirus-posi-tive frogs were detected at the same site where Bd wasdetected, although no individuals tested positive forboth pathogens. Ranaviruses were not detected inboreal chorus frogs or western toads. In some in-stances, tadpoles that tested positive for ranavirus byPCR had exhibited stereotypic gross signs of ranavirusinfections such as red hemorrhages on the legs and tailand abdominal swelling (Greer et al. 2005). However,most individuals that tested positive for ranavirus didnot outwardly appear infected.
The MCP sequences of all ranaviruses detected inour study were 99%+ identical to one another (576 or577 bp / 577 bp). BLAST searches indicated that theMCP sequences from our study were >99% identical(576 or 577 bp / 577 bp) to the MCP sequence of FrogVirus 3 (FV3; GenBank accession number AY548484.1),the type isolate of amphibian ranaviruses (Tan et al.2004). Previous studies showed that the MCP sequence
of ranaviruses detected in wood frogs and northernleopard frogs in Saskatchewan (Schock et al. 2008) andOntario (Greer et al. 2005) were >98% identical to theFV3 MCP sequence. Representative MCP sequencesfrom our study were submitted to GenBank underaccession numbers GQ144407 and GQ144408.
In 2 areas, Norman Wells and Fort Liard, samplessizes were sufficiently large to allow for statistical com-parisons of prevalences. Because of the potential biasin Bd detection in tadpoles, only frog-stage individualswere included in comparisons of Bd prevalence. CIsoverlapped when Bd prevalence in each area wascompared between years (data not shown) so datafrom both years were pooled for each area. The preva-lence of Bd in the Fort Liard area was 14% (7/51, 95%CI = 6 to 26%) while the prevalence of Bd in the Nor-man Wells area was 0% (0/59, 97.5% 1-tailed CI = 0 to6%). The difference in Bd prevalence between Nor-man Wells and Fort Liard was significant (Fisher’sexact test, χ2 1 df = 7.5738, p = 0.006).
Ranaviruses were only detected in wood frogs, andtherefore only wood frogs were included in statisticalcomparisons. Both tadpoles and frog-stage wood frogswere included in calculations since there were noconcerns about swab-related sampling biases be-tween life stages; ranaviruses cause internal infec-tions and therefore tissues, not swabs, were screenedfor ranavirus. CIs of ranavirus prevalences overlappedbetween years within each area (not shown) so datafor each area were pooled across years. In both areas,ranavirus prevalence in wood frogs was 7% (NormanWells = 9/131, 95% CI = 3 to 13%; Fort Liard =10/135, 95% CI = 4 to 13%). There was no significantdifference in ranavirus prevalence between NormanWells and Fort Liard (Fisher’s exact test, χ2 1 df =0.0251, p = 0.874).
DISCUSSION
We detected Bd and ranaviruses, 2 pathogens associ-ated with amphibian declines, in the NT, therebyextending the known range of both pathogens.Although ranaviruses were found widely, they weredetected only in wood frogs. In contrast, Bd was foundonly in the Fort Liard area but was detected in all 3amphibian species.
Detection of Bd in our study area is not congruentwith niche models developed by Ron (2005). Ron(2005) predicted the geographic distribution of Bdusing several abiotic variables related to elevation,precipitation, and temperature based on locationswhere Bd was reported in the literature. The modelsdid not predict that Bd would occur in the NT, with theclosest predicted areas >450 km south or west of
5

Dis Aquat Org: Preprint, 20096
Lat
itu
de
/ lo
ng
itu
de
Gen
eral
are
aS
ite
nam
eS
pec
ies
Ch
ytri
d s
cree
nR
anav
iru
s sc
reen
Tad
pol
esF
rog
sT
adp
oles
Fro
gs
2007
60°1
7’16
”N
, 123
°29’
46”
WF
t. L
iard
K29
roa
dW
ood
fro
g–
0/5
–0/
560
°19’
16”
N, 1
23°1
8’16
”W
Ft.
Lia
rdM
usk
eg R
iver
dem
onst
rati
on f
ores
tW
ood
fro
g–
0/4
–0/
460
°18’
17”
N, 1
23°1
9’22
”W
Ft.
Lia
rdM
usk
eg R
iver
gra
vel
pit
pon
ds
Woo
d f
rog
0/21
2/7
5/21
0/7
Wes
tern
toa
d0/
610/
10/
610/
160
°05’
37”
N, 1
23°0
5’33
”W
Ft.
Lia
rdR
oad
sid
e P
ond
2W
ood
fro
g0/
25–
2/25
–60
°09’
28”
N, 1
23°1
4’31
”W
Ft.
Lia
rdR
oad
sid
e P
ond
3W
ood
fro
g0/
70/
100/
70/
1060
°08’
18”
N, 1
23°1
1’38
”W
Ft.
Lia
rdR
oad
sid
e P
ond
4W
ood
fro
g0/
120/
23/
120/
261
°07’
33”
N, 1
22°5
1’02
”W
Bla
ckst
one
Lin
db
erg
Lan
din
gW
ood
fro
g–
0/3
–1/
360
°56’
55”
N, 1
23°0
5’39
”W
Bla
ckst
one
Nah
ann
i B
utt
e W
inte
r R
oad
Pon
d 1
Woo
d f
rog
0/12
–1/
12–
Ch
oru
s fr
og0/
130/
40/
130/
460
°57’
14”
N, 1
23°0
6’57
”W
Bla
ckst
one
Nah
ann
i B
utt
e W
inte
r R
oad
Pon
d 2
Woo
d f
rog
0/5
0/7
0/5
0/7
61°0
9’44
”N
, 123
°48’
49”
WN
ahan
ni
NP
RN
ear
wh
ere
Jack
fish
Riv
er e
nte
rsW
ood
fro
g–
0/6
–0/
6S
outh
Nah
ann
i R
iver
61°1
3’24
”N
, 123
°46’
15”
WN
ahan
ni
NP
RY
ohin
Lak
eW
ood
fro
g–
0/5
–1/
565
°15’
36”
N, 1
26°4
1’52
”W
Nor
man
Wel
lsA
irst
rip
are
aW
ood
fro
g0/
5–
0/5
–65
°17’
26”
N, 1
26°5
2’34
”W
Nor
man
Wel
lsB
osw
orth
Cre
ek –
up
per
bri
dg
eW
ood
fro
g–
0/13
–0/
1365
°18’
12”
N, 1
26°1
1’1”
WN
orm
an W
ells
Bos
wor
th C
reek
– o
rig
inW
ood
fro
g–
0/7
–0/
765
°15’
18”
N, 1
26°5
8’26
”W
Nor
man
Wel
lsW
sid
e of
Mac
ken
zie
Riv
er –
Can
ol t
rail
hea
dW
ood
fro
g–
0/10
–0/
1065
°13’
33”
N, 1
26°5
1’08
”W
Nor
man
Wel
lsW
sid
e of
Mac
ken
zie
Riv
er –
Sto
p 1
Woo
d f
rog
–0/
3–
0/3
65°1
3’37
”N
, 126
°46’
24”
WN
orm
an W
ells
W s
ide
of M
ack
enzi
e R
iver
– S
top
2W
ood
fro
g–
0/8
–0/
865
°15’
8”N
, 126
°39’
53”
WN
orm
an W
ells
DO
T P
ond
Woo
d f
rog
0/1
0/6
1/1
0/6
65°1
7’47
”N
, 126
°49’
53”
WN
orm
an W
ells
Hon
ey B
uck
et R
oad
slo
ug
hW
ood
fro
g–
0/4
–1/
465
°17’
35”
N, 1
26°3
6’22
”W
Nor
man
Wel
lsJa
ckfi
sh L
ake
– ea
st e
nd
Woo
d f
rog
–0/
1–
0/1
65°1
6’30
”N
, 126
°47’
21”
WN
orm
an W
ells
Loo
mis
Gre
enh
ouse
pon
dW
ood
fro
g0/
37–
2/37
–65
°15’
47”
N, 1
26°4
3’35
”W
Nor
man
Wel
lsV
OR
tow
er r
oad
pon
ds
Woo
d f
rog
0/1
–0/
1–
66°0
0’23
”N
, 128
°56’
15”
WN
orm
an W
ells
/M
idd
le o
f n
owh
ere
swam
pW
ood
fro
g–
0/2
–0/
2F
t. G
ood
Hop
e 20
0861
°29’
57”
N, 1
20°3
7’55
”W
Jean
Mar
ie R
iver
Roa
dsi
de
Pon
d 1
3W
ood
fro
g–
0/4
–0/
4C
hor
us
frog
0/30
a–
0/30
–61
°28’
15”
N, 1
20 °
36’5
3”W
Jean
Mar
ie R
iver
Roa
dsi
de
Pon
d 1
4W
ood
fro
g0/
80/
230/
80/
23C
hor
us
frog
–0/
2–
0/2
62°2
7’29
”N
, 123
°00’
31”
WW
rig
ley
Roa
dsi
de
Pon
d 1
5W
ood
fro
g–
0/2
–0/
263
°08’
40”
N, 1
23°1
5’38
”W
Wri
gle
yR
oad
sid
e P
ond
16
Woo
d f
rog
–0/
4–
0/4
Ch
oru
s fr
og–
0/3
–0/
362
°37’
51”
N, 1
23°0
4’19
”W
Wri
gle
yR
oad
sid
e P
ond
17
Ch
oru
s fr
og0/
4–
0/4
–60
°18’
17”
N, 1
23°1
9’22
”W
Ft.
Lia
rdM
usk
eg R
iver
gra
vel
pit
pon
ds
Woo
d f
rog
0/10
3/8
0/10
0/8
Ch
oru
s fr
og0/
41/
10/
40/
1W
este
rn t
oad
0/34
1/3
0/34
0/3
60°1
6’36
”N
, 123
°29’
14”
WF
t. L
iard
K29
Roa
dW
ood
fro
g0/
50/
90/
50/
9
Tab
le 1
. 200
7 an
d 2
008
surv
eys
for
chyt
rid
fun
gu
s an
d r
anav
iru
ses
in a
mp
hib
ian
s in
the
Nor
thw
est T
erri
tori
es, C
anad
a. P
CR
-bas
ed d
iag
nos
tics
wer
e u
sed
to s
cree
n s
amp
les
for
the
pat
hog
ens.
Val
ues
ind
icat
e th
e n
um
ber
of
ind
ivid
ual
s th
at t
este
d p
osit
ive
out
of t
he
nu
mb
er s
cree
ned
. NP
R: N
atio
nal
Par
k R
eser
ve
Schock et al.: Amphibian pathogens in the Northwest Territories
where we detected Bd near Fort Liard. Furtherresearch is needed to identify the underlying reason(s)for the discrepancy between predictions made by Ron(2005) and our finding of Bd in the NT. One explana-tion may be related to information that was availableat the time Ron (2005) developed his models — nichemodels extrapolate from available data, and Bd is nowknown from many more locations than when Ron(2005) developed his projections. However, other stud-ies that have searched for Bd at northern latitudeshave been consistent with predictions made by Ron(2005): Bd was detected in Kenai National WildlifeRefuge, Alaska (Reeves & Green 2006, Reeves 2008),but not in Tetlin or Innoko National Wildlife Refuges(Reeves 2008), Alaska, or Denali National Park, Alaska(Chestnut et al. 2008). Similarly, Adams et al. (2007)and Pearl et al. (2007) detected Bd at several locationsin western North America, as predicted by Ron (2005).
An alternate explanation for Bd in the NT may belinked to host species assemblages at a location andeach species’ uses of microhabitats that protect Bdfrom inhospitable conditions. For example, the extentto which Bd can withstand freezing in nature isunknown (Daszak et al. 2003, but see Seimon et al.2007). Bd may be able to persist in areas with other-wise unsuitable habitats as a result of the overwinter-ing strategy of host species that avoid freezing. Woodfrogs and boreal chorus frogs overwinter on land,buried under leaf litter and snow cover. Both of thesespecies freeze to –3°C, a feat made possible by physi-ological cryoprotectants that control the freezing pro-cess and mitigate the physiological stresses associatedwith freezing (Storey & Storey 1996). In areas whereonly wood frogs exist, persistence of Bd may dependon suitable environmental conditions. This explana-tion is consistent with the results of surveys in Alaska(Reeves & Green 2006, Chestnut et al. 2008, Reeves2008): Bd was only detected in areas where the envi-ronmental conditions are conducive to Bd survival, aspredicted by Ron (2005). In contrast, western toads donot freeze during winter, but overwinter below thefrost line by burrowing and using existing cavitiessuch as those associated with decayed root channelsand abandoned beaver lodges (Muths & Nanjappa2005, Brown & Symes 2007). It is plausible that Bd ispersisting near Fort Liard by overwintering on west-ern toads.
Ranaviruses were detected in wood frogs in severallocations in our study, including Norman Wells(65° 17’ N), which is the most northerly record of am-phibian ranaviruses in North America. In all instances,the ranaviruses we detected were FV3-like based ontheir MCP gene sequences. However, Schock et al.(2008) demonstrated with restriction enzyme analysesthat amphibian ranaviruses whose MCP sequences are
7
Lat
itu
de
/ lo
ng
itu
de
Gen
eral
are
aS
ite
nam
eS
pec
ies
Ch
ytri
d s
cree
nR
anav
iru
s sc
reen
Tad
pol
esF
rog
sT
adp
oles
Fro
gs
2008
60°0
5’37
”N
, 123
°05’
33”
WF
t. L
iard
Roa
dsi
de
Pon
d 2
Woo
d f
rog
0/5
–0/
5–
60°0
9’28
”N
, 123
°14’
31”
WF
t. L
iard
Roa
dsi
de
Pon
d 3
Woo
d f
rog
–0/
1–
0/1
61°1
4’47
”N
, 123
°59’
43”
WN
ahan
ni
NP
RR
iver
Sto
p 1
Woo
d f
rog
–0/
3–
0/3
61°1
5’27
”N
, 123
°58’
14”
WN
ahan
ni
NP
RR
iver
Sto
p 2
Woo
d f
rog
–0/
2–
0/2
61°1
3’08
”N
, 123
°46’
26”
WN
ahan
ni
NP
RY
ohin
Lak
e ca
bin
Woo
d f
rog
–0/
5–
0/5
61°1
4’0”
N, 1
23°4
5’30
”W
Nah
ann
i N
PR
Yoh
in T
rail
Sto
p 1
Woo
d f
rog
–0/
2–
0/2
60°5
6’55
”N
, 123
°05’
39”
WB
lack
ston
eN
ahan
ni
Bu
tte
Win
ter
Roa
d P
ond
1W
ood
fro
g0/
16–
0/16
–60
°57’
14”
N, 1
23°0
6’57
”W
Bla
ckst
one
Nah
ann
i B
utt
e W
inte
r R
oad
Pon
d 2
Woo
d f
rog
0/16
–0/
16–
65°1
5’8”
N, 1
26°3
9’53
”W
Nor
man
Wel
lsD
OT
pon
dW
ood
fro
g0/
5–
0/5
–65
°17’
47”
N, 1
26°4
9’53
”W
Nor
man
Wel
lsH
oney
Bu
cket
Roa
d s
lou
gh
Woo
d f
rog
0/11
0/4
1/11
1/4
65°1
5’47
”N
, 126
°43’
35”
WN
orm
an W
ells
VO
R t
ower
roa
d p
ond
sW
ood
fro
g0/
120/
13/
120/
167
°02’
32”
N,1
26°0
5’49
”W
Col
vill
e L
ake
Col
vill
e S
top
2W
ood
fro
g–
0/1
–0/
167
°02’
04”
N, 1
26°0
5’58
”W
Col
vill
e L
ake
Col
vill
e L
ily
Pon
dW
ood
fro
g0/
90/
10/
90/
1
a Du
e to
log
isti
cal
pro
ble
ms,
th
ese
bor
eal
chor
us
frog
tad
pol
es c
ould
not
be
swab
bed
. T
he
chor
us
frog
tad
pol
es w
ere
tail
cli
pp
ed a
nd
th
e ti
ssu
es a
ssay
ed f
or b
oth
pat
hog
ens,
bu
t b
ecau
se t
adp
ole
tail
cli
ps
are
mu
ch l
ess
lik
ely
to b
e p
osit
ive
for
chyt
rid
th
an s
wab
s, t
hes
e re
sult
s m
ay r
efle
ct f
alse
neg
ativ
es
Tab
le 1
(co
nti
nu
ed)

Dis Aquat Org: Preprint, 2009
>99% identical to FV3 can differ elsewhere in theirgenome. Detailed characterizations required to ade-quately compare northern amphibian ranaviruses tothose found elsewhere await further study.
We did not encounter any wood frog die-offs, butwe suspect that ranavirus related die-offs occur inwood frogs in the NT as they do elsewhere (Green etal. 2002, Greer et al. 2005, Harp & Petranka 2006,Schock et al. 2008). It is possible that our surveysoccurred too early in the year, and therefore too earlyin epidemic curves, to detect large die-offs. Further-more, our study likely underestimates the prevalenceof ranaviruses in northern populations of wood frogsbecause we screened toe/tail tissues. Amphibianranaviruses attack internal organs, especially liver,kidney, and gastro-intestinal tissues (Bollinger et al.1999, Greer et al. 2005). However, testing theseorgans requires lethal sampling. By testing toe/tail tis-sues, our methods could only detect infections wherevirus was circulating in the blood in sufficiently hightitres. Future studies that facilitate repeated visits tosites over the entire course of the amphibian activeseason will be invaluable to understanding ranavirusdisease dynamics in general.
An unexpected finding we encountered was evi-dence that some of our swab samples may havedegraded prior to DNA extraction. We became awareof the situation when positive results were obtainedwhen screening frog toes but negative results wereobtained from swabs of those same individuals.Because Bd causes external infections of mouthparts intadpoles, tail clips from tadpoles are inherently lesslikely to collect zoospores than swabs, and thereforeare more likely to give false negative results. As a con-sequence, failure to detect Bd in tail clips is not infor-mative. The handling and storage of our samples wereconsistent with those described by Hyatt et al. (2007),wherein they report no decrease in zoospore recoveryin samples stored at room temperature for 18 mo.Recently, another study has shown that storage andhandling conditions appear to impact the integrity ofswabs (Van Sluys et al. 2008). Although we do notknow which aspect(s) of handling may have affectedthe swabs in our study, prolonged exposure to hightemperatures, the topic addressed by Van Sluys et al.(2008), did not occur. This suggests that other aspectsof storage or handling may also affect the integrity ofsamples collected on swabs in field settings.
It is unknown whether Bd or ranaviruses are recentarrivals to the Canadian north or if they have beencomponents of northern ecosystems for considerableperiods of evolutionary time. This important questioncan be explored through further genetic analyses. Theresults of such analyses can also inform the largerdebate about the origin of Bd in North America as well
as the extent to which the ranges of these pathogensmay be shifting as a result of climate change or rapidindustrial development. If Bd and/or ranaviruses havehistorically been components of northern ecosystems,disease dynamics will have been influenced by localselection pressures on hosts and pathogens alike.Amphibian ranaviruses and Bd infect multiple hostspecies and therefore transmission dynamics will likelybe affected by changes in northern host species assem-blages. Without limits on migration, amphibians arepredicted to expand into higher latitudes to a greaterextent than other vertebrates as our climate changes(Araújo et al. 2006, Lawler et al. 2009). As a result,threats posed by ranaviruses or Bd to the long-termpersistence of northern amphibian populations mayarise from altered disease dynamics due to the move-ment of additional host species into northern areas.Regardless of any changes that may occur in host orpathogen geographic ranges, climate change in thenorth will likely lead to changes in Bd disease dynam-ics because its lifecycle is tightly coupled with bothtemperature and humidity (Woodhams et al. 2003,Pounds et al. 2006). Similarly, temperature alone canprofoundly affect mortality rates due to ranavirusinfections (Rojas et al. 2005).
Although logistical issues involved with working inthe north can be considerable, the biological tractabil-ity of the ecosystem creates opportunities to under-stand complex questions surrounding the role of infec-tious diseases in global amphibian declines. Lessonslearned from watching the north may be instrumentalin managing the effect of infectious diseases onamphibian populations much farther south.
Acknowledgements. Field surveys were conducted in compli-ance with Wildlife Research Permits and Wildlife Care Com-mittee Protocols from the Government of the Northwest Terri-tories, Parks Canada Research Collection Permits, andUniversity of Calgary Animal Use Protocols. Permission toconduct surveys was granted by Acho Dene Koe Band,Dechita Society, Fort Liard Métis Local 67, Fort SimpsonMétis Local 52, Liidlii Kue First Nation, Jean Marie River FirstNation, Nahanni Butte Dene Band, Pehdzeh Ki First Nation,and the Behdzi Ahda First Nation in Colville Lake. Assistancefrom B. Elkin with procuring sampling materials is gratefullyacknowledged. Enthusiastic volunteer field help from Nor-man Wells residents D. Fowler, K. Kivi, M. Meulenbroek, andY. Meulenbroek was appreciated. It was a privilege to workwith Community Representatives G. Tsetso and C. Deneyoua,and Parks Canada staff J. Hardisty, M. Matou, and A.Okrainec, while surveying in Nahanni National Park Reserve.This research received financial and considerable logisticalsupport from the Government of the Northwest Territories –Department of Environment & Natural Resources, NSERCPromoScience (S.J.K.), NSF Integrated Research Challengesin Environmental Biology grant no. DEB 0213851 (J.P.C.),Parks Canada, Sahtu Renewable Resources Board, and theDetroit Zoological Society.
8

Schock et al.: Amphibian pathogens in the Northwest Territories
LITERATURE CITED
Adams MJ, Galvan S, Reinitz D, Cole RA, Pyare S, Hahr M,Govindarajulu P (2007) Incidence of the fungus Batra-chochytrium dendrobatidis in amphibian populationsalong the northwest coast of North America. Herpetol Rev38:430–431
Araújo MB, Thuiller W, Pearson RG (2006) Climate warmingand the decline of amphibians and reptiles in Europe.J Biogeogr 33:1712–1728
Berger L, Speare R, Daszak P, Green DE and others (1998)Chytridiomycosis causes amphibian mortality associatedwith population declines in the rain forest of Australia andCentral America. Proc Natl Acad Sci USA 95:9031–9036
Berger L, Speare R, Skerratt LF (2005) Distribution of Batra-chochytrium dendrobatidis and pathology in the skin ofgreen tree frogs Litoria caerulae with severe chytridiomy-cosis. Dis Aquat Org 68:65–70
Bollinger TK, Mao J, Schock D, Brigham RM, Chinchar VG(1999) Pathology, isolation and preliminary molecularcharacterization of a novel iridovirus from tiger salaman-ders in Saskatchewan. J Wildl Dis 35:413–429
Bosch J, Carrascal LM, Duran L, Walker S, Fisher MC (2007)Climate change and outbreaks of amphibian chytridio-mycosis in a montane area of Central Spain; is there alink? Proc R Soc Lond B Biol Sci 274:253–260
Boyle DG, Boyle DB, Olsen V, Morgan JAT, Hyatt AD (2004)Rapid quantitative detection of chytridiomycosis (Batra-chochytrium dendrobatidis) in amphibian samples usingreal-time Taqman PCR assay. Dis Aquat Org 60:141–148
Brown C, Symes S (2007) Hibernation sites of western toadsin Alberta. Alberta Wildlifer 18:6–8
Chestnut T, Johnson JE, Wagner RS (2008) Results of amphibianchytrid (Batrachochytrium dendrobatidis) sampling in DenaliNational Park, Alaska, USA. Herpetol Rev 39: 202–204
Chinchar VG (2002) Ranaviruses (family Iridoviridae): emerg-ing cold-blooded killers. Arch Virol 147:447–470
Collins JP, Storfer A (2003) Global amphibian declines: sort-ing the hypotheses. Divers Distrib 9:89–98
Collins JP, Brunner JL, Miera V, Parris MJ, Schock DM, Stor-fer A (2003) Ecology and evolution of infectious diseases.In: Semlitsch R (ed) Amphibian conservation. SmithsonianPress, Washington, DC, p 139–151
Daszak P, Cunningham AA, Hyatt AD (2003) Infectious dis-ease and amphibian population declines. Divers Distrib 9:141–150
Duffus ALJ, Pauli BD, Wozney K, Brunetti CR, Berrill M(2008) Frog virus 3-like infections in aquatic amphibiancommunities. J Wildl Dis 44:109–120
Ecosystem Classification Group (2007) Ecological regions ofthe Northwest Territories — Taiga Plains. Department ofEnvironment and Natural Resources, Government of theNorthwest Territories, Yellowknife, NT
Fournier MA(1997)Amphibians in the Northwest Territories. In:Green DM (ed) Amphibians in decline — Canadian studiesof a global problem. Herpetological Conservation No. 1.Society for the Study of Amphibians and Reptiles, St.Louis, MO
Green DE, Muths E (2005) Health evaluation of amphibians inand near Rocky Mountain National Park, Colorado, USA.Alytes 22:109–129
Green DE, Converse KA, Schrader AK (2002) Epizootiology ofsixty-four amphibian morbidity and mortality events in theUSA, 1996–2001. Ann NY Acad Sci 969:323–339
Greer AL, Berrill M, Wilson PJ (2005) Five amphibian mortal-ity events associated with ranavirus infection in south cen-tral Ontario, Canada. Dis Aquat Org 67:9–14
Harp EM, Petranka JW (2006) Ranavirus in wood frogs (Ranasylvatica): potential sources of transmission within andbetween ponds. J Wildl Dis 42:307–318
Hyatt AD, Boyle DG, Olsen V, Boyle DB and others (2007) Diag-nostic assays and sampling protocols for the detection of Ba-trachochytrium dendrobatidis. Dis Aquat Org 73: 175–192
Jancovich JK, Davidson EW, Seiler A, Jacobs BL, Collins JP(2001) Transmission of the Ambystoma tigrinum virus toalternative hosts. Dis Aquat Org 46:159–163
Knapp RA, Morgan JAT (2006) Tadpole mouthpart depig-mentation as an accurate indicator of chytridiomycosis, anemerging disease of amphibians. Copeia 2006:188–197
Kusrini MD, Skerratt LF, Garland S, Berger L, Endarwin L(2008) Chytridiomycosis in frogs of Mount Gede Pan-grengo, Indonesia. Dis Aquat Org 82:187–194
Lawler JJ, Shafer SL, White D, Kareiva P, Maurer EP, BlusteinAR, Bartlein PJ (2009) Projected climate-induced faunalchange in the Western Hemisphere. Ecology 90:588–597
Lemmon EM, Lemmon AR, Collins JT, Lee-Yaw JA, Can-natella DC (2007) Phylogeny-based delimitation of speciesboundaries and contact zones in the trilling chorus frogs(Pseudacris). Mol Phylogenet Evol 44:1068–1082
Lips KR, Brem F, Brenes R, Reeve JD and others (2006)Emerging infectious disease and the loss of biodiversity ina Neotropical amphibian community. Proc Natl Acad SciUSA 103:3165–3170
Lips KR, Diffendorfer J, Mendelson JR III, Sears MW (2008)Riding the wave: reconciling the roles of disease andclimate change in amphibian declines. PLoS Biol 6:e72,doi:10.1371/journal.pbio.0060072
Longcore JR, Longcore JE, Pessier AP, Halteman WA (2007)Chytridiomycosis widespread in anurans of northeasternUnited States. J Wildl Manag 71:435–444
Mao J, Tham TN, Gentry GA, Aubertin A, Chinchar VG(1996) Cloning, sequence analysis, and expression of themajor capsid protein of the iridovirus frog virus 3. Virology216:431–436
McCallum M (2007) Amphibian decline or extinction? Cur-rent declines dwarf background extinction rate. J Her-petol 41:483–491
Miller DL, Rajeev S, Gray MJ, Baldwin CA (2007) Frog virus 3infection, cultured American bullfrogs. Emerg Infect Dis13:342
Morehouse EA, James TY, Ganley ARD, Vilgalys R, Berger L,Murphy PJ, Longcore JE (2003) Multilocus sequence typ-ing suggests the chytrid pathogen of amphibians is arecently emerged clone. Mol Ecol 12:395–403
Morgan JAT, Vredenburg VT, Rachowicz LJ, Knapp RA andothers (2007) Population genetics of the frog-killing fun-gus Batrachochytrium dendrobatidis. Proc Natl Acad SciUSA 104:13845–13850
Moriarty E, Lannoo MJ 2005. Pseudacris triseriata complex(including feriarum, kalmi, triseriata, and maculata). In:Lannoo M (ed) Amphibian declines — the conservationstatus of United States species. University of CaliforniaPress, Los Angeles, CA, p 485–488
Muths E, Nanjappa P (2005) Bufo boreas Baird and Girard,western toad. In: Lannoo M (ed) Amphibian declines —the conservation status of United States species. Univer-sity of California Press, Los Angeles, CA, p 392–396
Muths E, Corn PS, Pessier AP, Green DE (2003) Evidence fordisease-related amphibian decline in Colorado. Biol Con-serv 110:357–365
NatureServe (2009) NatureServe Explorer: an online ency-clopedia of life (web application). Version 7.1. Nature-Serve, Arlington, VA. Available at: www.natureserve.org/explorer
9

Dis Aquat Org: Preprint, 2009
Ouellet M, Mikaelian I, Pauli BD, Rodrigue J, Green DM(2005) Historical evidence of widespread chytrid infectionin North American amphibian populations. Conserv Biol19:1431–1440
Padgett-Flohr GE, Goble ME (2007) Evaluation of tadpolemouthpart depigmentation as a diagnostic test for infec-tion by Batrachochytrium dendrobatidis for four Californiaanurans. J Wildl Dis 43:690–699
Pearl CA, Bull EL, Green DE, Bowerman J, Adams MJ, HyattA, Wente WH (2007) Occurrence of the amphibianpathogen Batrachochytrium dendrobatidis in the PacificNorthwest. J Herpetol 41:145–149
Pessier AP, Nichols DK, Longcore JE, Fuller MS (1999) Cuta-neous chytridiomycosis in poison dart frogs (Dendrobatesspp.) and White’s tree frogs (Litoria caerulae). J Vet DiagnInvest 11:194–199
Pounds JA, Bustamante MR, Coloma LA, Consuegra JA andothers (2006) Widespread amphibian extinctions from epi-demic disease driven by global warming. Nature 439:161–167
Rachowicz LJ, Hero JM, Alford RA, Taylor JW and others(2005) The novel and endemic pathogen hypotheses: com-peting explanations for the origin of emerging infectiousdiseases of wildlife. Conserv Biol 19:1441–1448
Raverty S, Reynolds T (2001) Cutaneous chytridiomycosis indwarf aquatic frogs (Hymenochirus boettgeri) originatingfrom southeast Asia and in a western toad (Bufo boreas)from northeastern British Columbia. Can Vet J 42:385–386
Reeves MK (2008) Batrachochytrium dendrobatidis in woodfrogs (Rana sylvatica) from three national wildlife refugesin Alaska, USA. Herpetol Rev 39:68–70
Reeves MK, Green DE (2006) Rana sylvatica (wood frog)chytridiomycosis. Herpetol Rev 37:450
Retallick RWR, Miera V (2007) Strain differences in theamphibian chytrid Batrachochytrium dendrobatidis andnon-permanent, sub-lethal effects of infection. Dis AquatOrg 75:201–207
Retallick RWR, McCallum H, Speare R (2004) Endemic infec-tion of the amphibian chytrid fungus in a frog communitypost-decline. PLoS Biol 2:e351, doi:10.1371/journal.pbio.0020351
Rittman SE, Muths E, Green DE (2003) Pseudacris triseriata(western chorus frog) and Rana sylvatica (wood frog)chytridiomycosis. Herpetol Rev 34:53
Rojas S, Richards K, Jancovich JK, Davidson EW (2005) Influ-ence of temperature on ranavirus infection in larval sala-manders Amybstoma tigrinum. Dis Aquat Org 63:95–100
Ron SR (2005) Predicting the distribution of the amphibianpathogen Batrachochytrium dendrobatidis in the NewWorld. Biotropica 37:209–221
Russell AP, Bauer AM (2000) Amphibians and reptiles ofAlberta. A field guide and primer of boreal herpetology,2nd edn. University of Calgary Press, Calgary, AB
Sambrook J, Russell DW (2001) Molecular cloning: a labora-tory manual. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY
Schock DM, Bollinger TK, Chinchar VG, Jancovich JK,
Collins JP (2008) Experimental evidence that amphibianranaviruses are multi-host pathogens. Copeia 2008:133–143
Seimon TA, Seimon A, Daszak P, Halloy SRP and others(2007) Upward range extension of Andean anurans andchytridiomycosis to extreme elevations in response totropical deglaciation. Glob Change Biol 13:288–299
Skerratt LF, Berger L, Speare R, Cashins S and others (2007)Spread of chytridiomycosis has caused the rapid globaldecline and extinction of frogs. EcoHealth 4:125–134
Smith KF, Acevedo-Whitehouse K, Pedersen AB (2009) Therole of infectious diseases in biological conservation. AnimConserv 12:1–12
Stebbins RC (2003) A field guide to western reptiles andamphibians, Peterson field guide series, 3rd edn.Houghton Mifflin Company, New York
Storey KB, Storey JM (1996) Natural freezing survival in ani-mals. Annu Rev Ecol Syst 27:365–386
Stuart SN, Chanson JS, Cox NA, Young BE, Rodrigues ASL,Fischman DL, Waller RW (2004) Status and trends ofamphibian declines and extinctions worldwide. Science306:1783–1786
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molec-ular evolutionary genetics analysis (MEGA) software ver-sion 4.0. Mol Biol Evol 24:1596–1599
Tan WG, Barkman TJ, Chinchar VG, Essani K (2004) Compar-ative genomic analyses of frog virus 3, type species of thegenus Ranavirus (family Iridoviridae). Virology 323: 70–84
Van Sluys M, Kriger KM, Phillott AD, Campbell R, SkerrattLF, Hero JM (2008) Storage of samples at high tempera-tures reduces the amount of amphibian chytrid fungusBatrachochytrium dendrobatidis DNA detectable by PCRassay. Dis Aquat Org 81:93–97
Weldon C, Du Preez LH, Hyatt AD, Muller R, Speare R (2004)Origin of the amphibian chytrid fungus. Emerg Infect Dis10:2100–2105
Williams T, Barbosa-Solomieu V, Chinchar VG (2005) Adecade of advances in iridovirus research. Adv Virus Res65:173–248
Woodhams DC, Alford RA, Marantelli G (2003) Emerging dis-ease of amphibians cured by elevated body temperature.Dis Aquat Org 55:65–67
World Organisation for Animal Health (2008) OIE aquatic ani-mal health code, 11th edn. Available at: www.oie.int/eng/normes/en_acode.htm?e1d10
Young MK, Allison GT, Foster K (2007) Observations of borealtoads (Bufo boreas boreas) and Batrachochytrium dendro-batidis in south-central Wyoming and north-central Col-orado. Herpetol Rev 38:146–150
Zellmer AJ, Richards CL, Martens LM (2008) Low prevalenceof Batrachochytrium dendrobatidis across Rana sylvaticapopulations in southeastern Michigan, USA. Herpetol Rev39:196–198
Zhang QY, Xiao F, Li ZQ, Gui JF, Mao J, Chinchar VG (2001)Characterization of an iridovirus from the cultured pigfrog Rana grylio with lethal syndrome. Dis Aquat Org 48:27–36
10
Submitted: May 19, 2009; Accepted: August 23, 2009 Proofs received from author(s): November 23, 2009





























