Embed Size (px)
Citation preview

ART ICLES
NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005 1167
A schizophrenia-associated mutation of DISC1 perturbs cerebral cortex developmentAtsushi Kamiya1,2, Ken-ichiro Kubo3, Toshifumi Tomoda4,5, Manabu Takaki1, Richard Youn1, Yuji Ozeki1,2, Naoya Sawamura1, Una Park6, Chikako Kudo3,7, Masako Okawa2, Christopher A. Ross1,6,8,9, Mary E. Hatten4, Kazunori Nakajima3,7 and Akira Sawa1,6,9,10
Disrupted-In-Schizophrenia-1 (DISC1), originally identified at the breakpoint of a chromosomal translocation that is linked to a rare familial schizophrenia, has been genetically implicated in schizophrenia in other populations. Schizophrenia involves subtle cytoarchitectural abnormalities that arise during neurodevelopment, but the underlying molecular mechanisms are unclear. Here, we demonstrate that DISC1 is a component of the microtubule-associated dynein motor complex and is essential for maintaining the complex at the centrosome, hence contributing to normal microtubular dynamics. Carboxy-terminal-truncated mutant DISC1 (mutDISC1), which results from a chromosomal translocation, functions in a dominant-negative manner by redistributing wild-type DISC1 through self-association and by dissociating the DISC1–dynein complex from the centrosome. Consequently, either depletion of endogenous DISC1 or expression of mutDISC1 impairs neurite outgrowth in vitro and proper development of the cerebral cortex in vivo. These results indicate that DISC1 is involved in cerebral cortex development, and suggest that loss of DISC1 function may underlie neurodevelopmental dysfunction in schizophrenia.
DISC1 was originally identified as the sole disrupted gene that has an open reading frame at the breakpoint of a balanced chromosomal trans-location, which was inherited in a large Scottish pedigree1. Most of the family members with the chromosomal abnormality have major mental illnesses, including schizophrenia. These illnesses have been diagnosed according to the criteria of DSM-IV, and it has been shown that they have no physical, neurological or dysmorphic conditions2. The patients display a reduction in the amplitude of the P300 event-related potential, which is frequently observed in general schizophrenia. Lack of available autopsied brains from this Scottish family has hampered further under-standing of the role of DISC1 in the pathophysiology of their mental conditions. The balanced translocation could lead to production of a carboxy-terminal truncated DISC1 mutant protein (mutDISC1) or hap-loinsufficiency due to instability of the DISC1 protein from the disrupted chromosome. Recent linkage and association studies also indicate that DISC1 has a role in schizophrenia in general populations3–6. Therefore, understanding of the molecular and cellular roles of DISC1 may provide insight into the pathology of schizophrenia.
The neurodevelopmental aetiology of schizophrenia has indi-cated that enlargement of ventricles occurs, with the presence of mild
cytoarchitectural abnormalities and no signs of gliosis7–11. Such defects of schizophrenia brains indicate that basic developmental mechanisms are altered in schizophrenia. However, its underlying molecular mecha-nisms remain elusive. Neuronal migration is a crucial step in cortico-genesis during neuronal development12–15. Human and mouse genetics have revealed that genes — such as Reelin, doublecortin (DCX), LIS1 and NUDEL — are implicated in this process13,14,16–19. The interaction of Reelin signalling and LIS1 is indicated by the fact that an adaptor protein of the Reelin receptor, DAB1, and LIS1 bind in a Reelin-induced phos-phorylation-dependent manner during brain development20. Studies on several nuclear-distribution mutants of the fungus Aspergillus nidulans have revealed the involvement of NUDEL and LIS1 in dynein signal-ling19,21,22. NUDEL is required for dynein function and facilitates the interaction between LIS1 and dynein17. The dynein/dynactin motor has a key role, not only in retrograde transport but also in the transport of microtubules from the centrosome to the axon/neurites23,24. The centro-some has a key role in neuronal migration, in which the dynein complex and Par6α are involved25. Nucleokinesis is proposed, at least in part, to be an important underlying mechanism for neuronal migration in which the dynein complex may have a crucial role17,26,27. Taken together, the
1Department of Psychiatry-Neurobiology, Johns Hopkins University School of Medicine, 600 N. Wolfe Street, CMSC 8-117, Baltimore, MD 21287, USA. 2Department of Psychiatry, Shiga University of Medical Science, Seta tsukinowa-cho, Otsu, Shiga 520-21, Japan. 3Department of Anatomy, Keio University School of Medicine, 35 Shinanomachi, Shinjuku-ku, Tokyo 160-8582, Japan. 4Laboratory of Developmental Neurobiology, The Rockefeller University, 1230 York Avenue, New York, NY 10021, USA. 5Division of Neurosciences, Beckman Research Institute of the City of Hope, LA 91010, USA. 6Department of Neuroscience, Johns Hopkins University School of Medicine, Baltimore, MD 21205, USA. 7Department of Molecular Neurobiology, Institute of DNA Medicine, Jikei Univ. School of Medicine, Tokyo 105-8471, Japan. 8Department of Neurology, Johns Hopkins University School of Medicine, Baltimore, MD 21287, USA. 9Graduate Program in Cellular Molecular Medicine, Johns Hopkins University School of Medicine, Baltimore, MD 21205, USA.10Correspondence should be addressed to A.S. (e-mail: [email protected])
Received 28 September 2005; accepted 3 November 2005; published online: 20 November 2005; DOI: 10.1038/ncb1328
print ncb1328.indd 1167print ncb1328.indd 1167 1/12/05 3:03:57 pm1/12/05 3:03:57 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

1168 NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005
A RT I C L E S
dynein complex, including LIS1 and NUDEL, is thought to be involved in neurodevelopment.
We previously reported the interaction of DISC1 with NUDEL28, which has been reproduced by other groups29–31. Biochemical analyses indicate that NUDEL is able to act as a bridge between DISC1 and LIS1 to allow formation of a trimolecular complex31. Another study based on yeast two-hybrid screening reported protein binding of DISC1 with FEZ1 and kend-rin, a centrosome protein32,33. Expression of DISC1 in PC12 cells indicates that DISC1 is involved in neurite outgrowth28,33. In these previous studies, it was unclear whether or not DISC1 binds only to NUDEL, or whether it forms an overall complex with the dynein/dynactin proteins.
In the present study, we find that DISC1 is a component of the dynein protein complex and has an essential role in stabilizing the complex at the centrosome, thereby contributing to normal microtubular dynam-ics. Consequently, DISC1 is required for proper neurodevelopment of the cerebral cortex. The familial mutation, mutDISC1, functions as a dominant-negative mutation and perturbs these processes, indicating that abnormalities in cerebral cortical development underlie dysfunc-tions that occur in schizophrenia.
RESULTSDISC1 participates in dynein motor functionTo test whether DISC1 is a component of the dynein protein complex, we examined endogenous protein interactions of DISC1 in neuronal PC12 cells. DISC1 was co-immunoprecipitated with γ-tubulin, dynac-tin (p150glued), dynein intermediate chain (Dyn IC), LIS1 and NUDEL (Fig. 1a). The binding of mutDISC1 to γ-tubulin, p150glued, LIS1 and NUDEL was much weaker than that for full-length or wild-type DISC1 (wtDISC1) in COS-7 cells (Fig. 1b). The levels of endogenous DISC1 were much lower than those of exogenously expressed DISC1 in COS-7 cells (Fig. 1b). Specific localization of DISC1 at the centrosome, together with LIS1 and γ-tubulin, was shown by cell staining in both COS-7 cells and primary cortical neurons (Fig. 1c).
Dynein is the minus-end retrograde motor protein that mediates not only retrograde axonal transport in neurons but also transport of microtubule segments from the centrosome to the cell periphery or extensions23,24. As a result, accumulation of the dynein complex at the centrosome leads to peripheral enrichment of microtubular bundles. LIS1 reportedly modulates dynein activity, and overexpression of LIS1 in COS-7 cells leads to enrichment of microtubules in the cell periph-ery19,26,34. We observed that overexpression of wtDISC1 in PC12 cells led to accumulation of Dyn IC and p150glued at unique perinuclear regions that corresponded to the centrosome (Fig. 1d; also see Supplementary Information, Fig. S1a). Exogenous wtDISC1 in COS-7 cells led to an altered microtubular network, with accumulation of polymerized micro-tubules in the cell periphery (Fig. 1e). Overexpression of mutDISC1 led to a completely disorganized microtubular network.
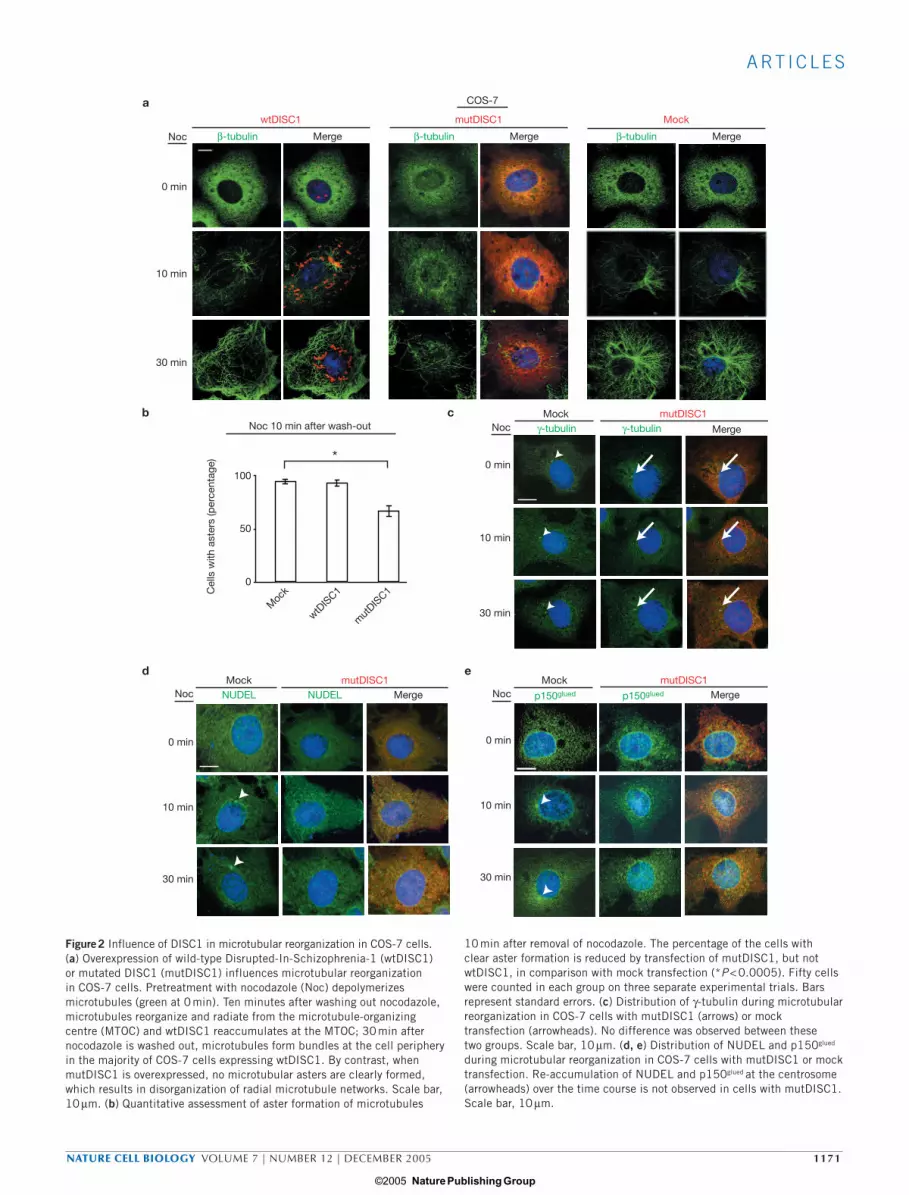
To obtain direct evidence that DISC1 plays a key role in centrosome-associated microtubule organization and dynamics, we analysed the time course of microtubular organization from the centrosome in COS-7 cells that were recovering from pretreatment with nocodazole, a micro-tubular depolymerizing agent (Fig. 2a). Ten minutes after nocodazole washout, no difference was evident between mock and wtDISC1-trans-fected cells, but a significant reduction in the percentage of cells with the microtubular aster from the centrosome was observed in cells expressing mutDISC1 (Fig. 2b). In cells with mock transfection, time-dependent
growth of microtubular asters in cells was observed. Cells with wtDISC1 displayed more peripheral growth of microtubular bundles 30 min after nocodazole washout, following transient formation of tiny asters, which is consistent with a role for DISC1 in centrosome-associated dynein motor functions. In cells with mutDISC1, disorganized radial microtu-bule networks are found over this time course. Under these conditions with mutDISC1, the centrosomes themselves are intact, as indicated by the immunostaining of γ-tubulin (Fig. 2c), but disturbance in normal re-accumulation of NUDEL, LIS1, Dyn IC and p150glued was observed (Fig. 2d, e; also see Supplementary Information, Fig. S1b).
Dominant-negative function of mutDISC1 in the centrosome-associated dynein protein complexTo examine the effect of mutDISC1, wt and mutDISC1 were exogenously expressed individually or together in COS-7 cells. wtDISC1 occurs selectively in the perinuclear regions, in contrast to the more diffuse cytoplasmic distribution of mutDISC1, as previously demonstrated by ourselves and others28,29. The levels of exogenous wt and mutDISC1 are almost equivalent, and are much greater than those of endogenous DISC1 in COS-7 cells, as shown by western blotting (Figs 1b, 3a). Following co-expression of mutDISC1, there was a robust change in subcellular distribution of wtDISC1 in that wtDISC1 became distributed more diffusely in the cytoplasm (Fig. 3a). By contrast, the distribution of mutDISC1 remained cytoplasmic, with small punctates occuring even after co-expression with wtDISC1. This result indicates that mutDISC1 disturbs the normal subcellular localization of wtDISC1; that is, it inter-feres with the normal and crucial function of wtDISC1. This observation was confirmed by the glycerol gradient method (Fig. 3b) and subcellular fractionation (data not shown). The subcellular change of wtDISC1 in the presence of mutDISC1 was specific, as co-overexpression of glycer-aldehyde-3-phosphate dehydrogenase (GAPDH) with wtDISC1 did not disturb the distribution of wtDISC1 in the glycerol gradient (Fig. 3b).
DISC1 contains coiled-coil domains in the middle portion of the mol-ecule, most of which are retained in mutDISC1. Therefore, we hypoth-esized that self-association of DISC1 may underlie the cellular effect of mutDISC1. To test this hypothesis, wtDISC1 and mutDISC1 proteins with Myc or HA-tag were expressed in HEK293 cells, and immunopre-cipitates with an anti-Myc antibody were analysed. The results indicate that, indeed, DISC1 self-associates, and that mutDISC1 can bind to wtDISC1 (Fig. 3c). This observed self-association was the result of its direct protein interaction (Fig. 3d). Amino acids 403–504, which harbour a well-conserved coiled-coil domain, were essential for DISC1 self-asso-ciation (Fig. 3e). Exogenous mutDISC1 in differentiated PC12 cells was co-immunoprecipitated with endogenous DISC1 (see Supplementary Information, Fig. S1c). This self-association is required for mutDISC1 to function as a dominant-negative mutation, as no subcellular redistribu-tion of wtDISC1 occurred when mutDISC1 deficient in the self-associa-tion domain (mutDISC1∆(403–504)) was co-transfected (Fig. 3f).
To test how the interference of wtDISC1 by mutDISC1 affects the dynein complex, we examined the localization of endogenous DISC1, LIS1, p150glued and γ-tubulin in COS-7 cells that were exogenously expressing mutDISC1. Endogenous wtDISC1 was selectively stained with an anti-DISC1 C-terminal antibody, the epitope of which is miss-ing in mutDISC1. MutDISC1 caused the redistribution of endogenous wtDISC1, LIS1 and p150glued from the centrosome, but the localization of γ-tubulin, the key structural component of the centrosome, was not
print ncb1328.indd 1168print ncb1328.indd 1168 1/12/05 3:04:02 pm1/12/05 3:04:02 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005 1169
A RT I C L E S
γ-tubulin
LIS1NUDEL
GAPDH
γ-tubulinp150glued p150glued
Dyn IC
LIS1GAPDH
HO
NUDEL
PC12a
c
e
bCOS-7
DISC1
d
DISC1
LIS1
Merge
DISC1
Merge
γ-tubulin
COS-7 Cortical neuron
DISC1
LIS1
Merge
DISC1
Merge
γ-tubulin
IP: anti-
IB: a
nti-
IB: a
nti-
IP: anti-HA
Tf:
HA
Dyn
IC
p15
0glu
ed
98
62
wtDISC1
98
62
COS-7
COS-7
IB: anti-DISC1
PC12
GAPDH Input wtDISC1 mutDISC1 GAPDH Input
Mock
Mr(K)
Mr(K)
Mock wtDISC1 mutDISC1
β-tubulin β-tubulin Merge β-tubulin Merge
1
2
3 3
1
2
wtDISC1Dyn IC Merge MergewtDISC1p150glued
1 2 3 1 2 3200
150
100
50
0
200
150
100
50
0
200
150
100
50
0
300
225
150
75
0
300
225
150
75
0
300
225
150
75
0
Figure 1 DISC1 regulates the dynein protein complex at the centrosome and modulates microtubular dynamics. (a) Endogenous Disrupted-In-Schizophrenia-1 (DISC1) co-immunoprecipitates with γ-tubulin, dynactin (p150glued), dynein intermediate chain (dynein IC), NUDEL and LIS1 in differentiated PC12 cells, but not with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or haem oxygenase (HO). Only GAPDH was co-precipitated with anti-GAPDH antibody. IB and IP indicate antibodies used for western blotting and immunoprecipitation, respectively. (b) Left panel: HA-tagged mutDISC1, compared with wtDISC1, displays much weaker binding with γ-tubulin, p150glued, NUDEL and LIS1 in COS-7 cells, as shown by immunoprecipitation with anti-HA antibody. Immunoprecipitates from cells expressing HA-GAPDH include GAPDH, but no DISC1 interactors. The exogenous proteins used for co-IP are depicted by western blotting with anti-HA antibody (lower panel). Tf indicates overexpressed proteins in COS-7 cells. Right panel: The cell extracts with wtDISC1 or mock transfection are probed with an anti-DISC1 antibody. The level of exogenous DISC1 (arrow) is more than tenfold that of endogenous DISC1. As reported, a couple of small variants of DISC1 are
also observed. K, relative molecular mass in thousands. (c) Endogenous DISC1 is colocalized with LIS1 and γ-tubulin at the centrosome in COS-7 cells (arrowheads) and cortical neurons (arrowheads). Scale bar, 10 µm. (d) In differentiated PC12 cells, overexpressed wtDISC1 leads to accumulation of dynein IC and p150glued at the centrosome, as analysed by the line scan. Vertical dotted lines indicate the borders between the cytoplasm and the nucleus. The x-axis denotes the position within the cells that were scanned along the white lines. The bold line represents the region that corresponds to the nucleus. The y-axis indicates the immunofluorescent signal intensity along the lines scanned. Dyn IC (left panel) and p150glued (right panel) accumulate in the perinuclear region of cells expressing wtDISC1 (line 1, but not lines 2 and 3). Scale bar, 10 µm. (e) Overexpressed wtDISC1 in COS-7 cells leads to an altered microtubular network with polymerized microtubule accumulation in the cell periphery. Overexpressed mutDISC1 leads to a disorganized microtubular network. Arrowheads: microtubule-organizing centre (MTOC). Green, microtubules; red, exogenous DISC1. Scale bar, 10 µm. For the whole blots of a and b, see Supplementary Information, Fig. S4.
print ncb1328.indd 1169print ncb1328.indd 1169 1/12/05 3:04:05 pm1/12/05 3:04:05 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

1170 NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005
A RT I C L E S
affected by expression of mutDISC1 (Fig. 4a). Under these experimental conditions, approximately 70–80% of LIS1 and p150glued were dissociated from the centrosome as a result of detachment of endogenous wtDISC1 from the centrosome (Fig. 4b). By contrast, mutDISC1∆(403–504) was much less effective in redistributing wtDISC1, LIS1 and p150glued, clearly indicating that the self-association of DISC1 is required for the domi-nant-negative effect of mutDISC1, which disrupts the role of endogenous wtDISC1 to anchor the dynein protein complex at the centrosome (see Supplementary Information, Fig. S1d). This notion is consistent with the above observation that overexpression of mutDISC1 results in absence of microtubule organization at the centrosome in COS-7 cells (Figs 1e, 2a), as mutDISC1 disturbs endogenous DISC1 at the centrosome.
DISC1 regulates neurite outgrowth in neuronal-cell culturesIf DISC1 has a key role in the dynein functions at the centrosome as indicated above, suppression of endogenous DISC1 in neuronal cells may lead to morphological changes. To test this question, we used RNA interference (RNAi) to target the suppression of DISC1 (ref. 35). We observed pronounced inhibition of neurite outgrowth in differentiating PC12 cells with the strong suppressor of DISC1, RNAi#1 (Fig. 5a, b, d; also see Supplementary Information, Fig. S2a, b). The disturbance of neurite outgrowth is unrelated to cell death and no signs of apoptotic nuclei were observed in cells with RNAi (Fig. 5c). The effect of RNAi is correlated with the levels of DISC1 suppression, as RNAi#1 showed a more marked inhibition of neurite outgrowth than did RNAi#2 (Fig. 5a, d). As co-trans-fection of the expression construct of human wtDISC1, which has four nucleotide mismatches with endogenous rat DISC1 sequences, normal-izes the inhibition of neurite outgrowth by RNAi#1, the effect of DISC1 RNAi could be attributed to the direct suppression of DISC1 (Fig. 5d). Suppression of DISC1 or overexpression of mutDISC1 both impair neurite outgrowth, which is consistent with the notion that mutDISC1 functions in a dominant-negative manner (Fig. 5d). We confirmed the occurrence of reduced accumulation of the protein interactors of DISC1, such as NUDEL, LIS1 and p150glued in PC12 cells expressing mutDISC1 (Fig. 5e), which is consistent with the above observation in COS-7 cells (Fig. 4). Similar effects of DISC1 RNAi were observed in primary cortical neurons (see Supplementary Information, Fig. S2c).
DISC1 plays a key role in the proper development of the cerebral cortex in vivoWe tested the influence of DISC1 in vivo by modulating DISC1 expres-sion in the developing cortex by an in utero gene-transfer technique using electroporation17,36–38. At embryonic day 14.5 (E14.5), embryos were subjected to electroporation with RNAi or expression constructs. At P2, brain slices with control RNAi showed that labelled cells exited the ventricular zone, migrated through the cortical wall and formed super-ficial layers of the cortex (Fig. 6a). When the tissue was electroporated with RNAi#2, which generated a mild phenotype in neurite outgrowth assays, subtle defects in migration were observed. When the tissue was electroporated with RNAi#1, neuronal migration was virtually inhibited, as few labelled neurons reached the upper layers of the cortex. Many process-bearing cells were still evident in the ventricular zone. Therefore, loss of DISC1 by RNAi results in a severe defect in neuronal migration in the developing neocortex.
To evaluate the role of DISC1 loss on dendritic arborization of cortical neurons, we examined sections from the same animals at P14, a time
point of extensive dendritic arborization and of the formation of local circuits in the neocortex (Fig. 6b). Whereas slices electroporated with control RNAi contain labelled pyramidal neurons in layers II/III with vertical arbors, neurons in layers II/III of brains that had been electro-porated with RNA#2 had shorter arbors that are not oriented in the radial plane. Within layers II/III of brains that had been electroporated with RNAi#1, fewer labelled neurons were seen, labelled neurons were malpositioned relative to those in control animals and their patterns of dendritic outgrowth were impaired with the formation of shorter, randomly oriented processes. The labelled cells that we observed in the upper layers of the P14 cortex were misaligned pyramidal neurons (see Supplementary Information, Fig. S3). Therefore, loss of DISC1 inhibits the rate of radial neuronal migration in the developing neocortex, and impairs the orientation, polarity and degree of dendritic arborization of those neurons that manage to migrate into position.
To address more mechanistic issues, we examined the effects of DISC1 on the nucleus–centrosome coupling that has been proposed as, at least in part, an important mechanism to account for neuronal-positioning defects in association with the dynein protein complex17,26,27. We exam-ined the migrating neurons in the cortical plate at E18.5, and observed a significantly increased distance between the nucleus and the centrosome when DISC1 expression was knocked down with RNAi (Fig. 6c).
mutDISC1 leads to abnormal development of the cerebral cortex in vivoWe next compared the effects of loss of DISC1 with those by expression of mutDISC1, which would exert a dominant-negative effect (Figs 2–5). MutDISC1 was introduced at E14.5, and the cerebral cortex was analysed between P2 and P14. The levels of mutDISC1 seemed to be low com-pared with endogenous DISC1 in the cortex, judging from immunos-taining with an anti-DISC1 antibody (data not shown). Electroporation of mutDISC1 resulted in delayed neuronal migration of some cells with mutDISC1 at P2 (Fig. 7a). Moreover, at P14, there was a small (20–25%) but significant reduction in the percentage of pyramidal neurons that were vertically oriented (Fig. 7b). The extent of the defects was similar to that seen in brain slices with RNAi#2 (milder RNAi). These minor cytoarchitectural changes in neurons were reminiscent of pathology observed in brains from patients with schizophrenia7–10. The resemblance of morphological defects resulting from decreased levels of endogenous DISC1 by RNAi and those induced by mutDISC1 further supports the concept that mutDISC1 functions, at least in part, as a dominant-nega-tive inhibitor of microtubular dynamics and of proper neurodevelop-ment of the cerebral cortex.
DISCUSSIONHere, we demonstrate that DISC1 is a key component of the microtu-bule-based dynein motor complex that comprises a number of proteins, including dynein, dynactin, LIS1 and NUDEL. In the central nervous system, dynein-motor-associated microtubular dynamics regulates neuronal migration and axonal formation during neuronal develop-ment12,14,15,19,25–27,34. We now demonstrate that DISC1 upregulates the function of this motor complex by accumulating and stabilizing it at the centrosome, thereby contributing to microtubular organization and dynamics. Lowering the function of endogenous DISC1 leads to disso-ciation of the complex from the centrosome, which results in decreased neurite outgrowth in vitro and disturbed neuronal migration in vivo.
print ncb1328.indd 1170print ncb1328.indd 1170 1/12/05 3:04:06 pm1/12/05 3:04:06 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group
NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005 1171
A RT I C L E S
Moc
k
wtDIS
C1
mut
DISC1
100
50
0
*
Cel
ls w
ith a
ster
s (p
erce
ntag
e)
p150glued p150glued
0 min
10 min
30 min
0 min
COS-7
0 min
10 min
30 min
a
b c
d e
Noc
Noc
Noc Noc
0 min
10 min
30 min
Noc 10 min after wash-out
wtDISC1 mutDISC1
mutDISC1
mutDISC1 mutDISC1
Mock
10 min
30 min
β-tubulin Merge
Mock
Mock Mock
β-tubulin
γ-tubulin γ-tubulin
Merge
Merge
Merge Merge
β-tubulin Merge
NUDEL NUDEL
Figure 2 Influence of DISC1 in microtubular reorganization in COS-7 cells. (a) Overexpression of wild-type Disrupted-In-Schizophrenia-1 (wtDISC1) or mutated DISC1 (mutDISC1) influences microtubular reorganization in COS-7 cells. Pretreatment with nocodazole (Noc) depolymerizes microtubules (green at 0 min). Ten minutes after washing out nocodazole, microtubules reorganize and radiate from the microtubule-organizing centre (MTOC) and wtDISC1 reaccumulates at the MTOC; 30 min after nocodazole is washed out, microtubules form bundles at the cell periphery in the majority of COS-7 cells expressing wtDISC1. By contrast, when mutDISC1 is overexpressed, no microtubular asters are clearly formed, which results in disorganization of radial microtubule networks. Scale bar, 10 µm. (b) Quantitative assessment of aster formation of microtubules
10 min after removal of nocodazole. The percentage of the cells with clear aster formation is reduced by transfection of mutDISC1, but not wtDISC1, in comparison with mock transfection (*P < 0.0005). Fifty cells were counted in each group on three separate experimental trials. Bars represent standard errors. (c) Distribution of γ-tubulin during microtubular reorganization in COS-7 cells with mutDISC1 (arrows) or mock transfection (arrowheads). No difference was observed between these two groups. Scale bar, 10 µm. (d, e) Distribution of NUDEL and p150glued
during microtubular reorganization in COS-7 cells with mutDISC1 or mock transfection. Re-accumulation of NUDEL and p150glued at the centrosome (arrowheads) over the time course is not observed in cells with mutDISC1. Scale bar, 10 µm.
print ncb1328.indd 1171print ncb1328.indd 1171 1/12/05 3:04:10 pm1/12/05 3:04:10 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

1172 NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005
A RT I C L E S
b
a
10% 25%
IB:anti-HA
anti-p150glued
Glycerol gradient
wtDISC1
wtDISC1
wtDISC1
wtDISC1−HA
mutDISC1−HA
wtDISC1−HA, mutDISC1−MycwtDISC1−HA, GAPDH−Myc
Mock
mutDISC1
COS-7
Co-IP
wtDISC1
mutDISC1
c e
98
62
Self-interactingregion
wtDISC1mutDISC1
(1−536)N(1−504)N(1−402)N
Breakpoint
Co-IP
d
f
DISC1
In vitro binding
mutDISC1∆(403−504)(402−854)C
wtDISC1
β-tubulin
wt
mut
wt
mut
wt Mergemut∆(403−504)
wtDISC1 mutDISC1only
wtDISC1+ mutDISC1
Merge
wtDISC1−Myc
HA-proteins
Predicted coiled-coil region
wtDISC1
98
98
6249
38
98
62G
APDH + w
tm
ut + m
utm
ut + w
tw
t + w
t
IP
Input
IP
Input
p150glued
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Mr(K)
Mr(K)
Mr(K)
mutDISC1
wtDISC1−HAwtDISC1−MycmutDISC1−HA
mutDISC1−MycGAPDH−Myc
GST
−DIS
C1
+ His
−DIS
C1
GST
+ H
is−D
ISC
1
wt +
wt
wt +
mut
wt +
(1−5
36)N
wt +
(1−5
04)N
wt +
(1−4
02)N
wt +
(402
−854
)Cw
t + m
ut∆(
403−
504)
wt +
GAP
DH
1 402
504
536 598 854
Figure 3 mutDISC1 functions in a dominant-negative manner via its self-association domain. (a) Left panel: wild-type Disrupted-In-Schizophrenia-1 (wtDISC1) is distributed predominantly in the perinuclear regions of COS-7 cells, especially in the centrosome. By contrast, mutated DISC1 (mutDISC1) is distributed widely in the cytoplasm. When wtDISC1 is co-expressed with mutDISC1, wtDISC1 interestingly redistributes from a discrete perinuclear to a diffuse organization in the cytoplasm. Scale bar, 10 µm. Right panel: The expression level of mutDISC1 is equivalent or slightly lower than that of wtDISC1 as shown by western blotting, indicating that the dominant influence of mutDISC1 compared with wtDISC1 is independent of the dose effects. β-Tubulin was used as an internal control. K, relative molecular mass in thousands. (b) A change in the subcellular distributions of wtDISC1 by co-expression of mutDISC1 in a glycerol density gradient. The distributions of wtDISC1 are markedly changed by the co-expression of mutDISC1, but not by glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The blot of each fraction with anti-p150glued antibody is also shown as a control for the fractionations. IB, immunoblot. (c) Co-immunoprecipitation (Co-IP) of exogenous wtDISC1–HA/wtDISC1–Myc, wtDISC1–HA/mutDISC1–Myc or mutDISC1–HA/mutDISC1–Myc in HEK293 cells. Both wt and/or mutDISC1
self-associate (top panel). GAPDH–Myc does not co-immunoprecipitate with wtDISC1–HA. Anti-Myc antibody and anti-HA antibody are used for precipitation and western blotting, respectively. The inputs of each protein are depicted (bottom panels). (d) DISC1 self-associates via direct protein–protein binding. Purified glutathione S-transferase (GST)- and His-tagged DISC1 recombinant protein fragments (amino acids 347–600) co-precipitate with glutathione-beads. (e) Domain mapping for DISC1 self-association. A series of transfected DISC1 mutants with HA-tag depicted in the lower scheme were tested for co-immunoprecipitation with an anti-HA antibody. Immunoprecipitates were analysed on SDS–PAGE, followed by western blotting with an anti-Myc antibody. The putative self-interacting region comprises amino acids 403–504 of DISC1. This binding domain is verified by failure of the self-association of DISC1 that is deficient in amino acids 403–504. The inputs of HA-tagged DISC1 mutant or GAPDH, as well as Myc-tagged wtDISC1, are depicted in the middle and bottom panels. (f) No redistribution of wtDISC1 from the discrete perinuclear regions to diffuse organization in the cytoplasm occurs when wtDISC1 is co-expressed with mutDISC1 that lacks the self-association domain (mutDISC1∆(403–504)). Scale bar, 10 µm. For the whole blots of b and d, see Supplementary Information, Fig. S4.
print ncb1328.indd 1172print ncb1328.indd 1172 1/12/05 3:04:11 pm1/12/05 3:04:11 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005 1173
A RT I C L E S
Protein interactions within the motor complex have not yet been well characterized. Tsai and colleagues17 recently reported that NUDEL com-petes with PAFAH α2 subunits for LIS1 binding in an in vitro binding assay, and NUDEL may thereby recruit LIS1 from the PAFAH complex and alter the conformation of LIS1 in favour of its binding to dynein17,39. It will be important in the future to elucidate how DISC1 and NUDEL relate to each other in terms of the regulation of the dynein complex, especially utilizing both structural and functional assays.
All the patients in the Scottish family have one intact and one mutant chromosome2. The lack of available autopsied brains from this family has hampered further understanding of the role of DISC1 in the patho-physiology of mental illnesses. The balanced translocation could lead to production of mutDISC1 or haploinsufficiency due to message or pro-tein instability of DISC1 from the disrupted chromosome. Our present study indicates that mutDISC1 fails to bind directly to the protein com-plex that contains LIS1, dynein and NUDEL, due to loss of the DISC1
C-terminal domain. We also demonstrated that mutDISC1 detaches wtDISC1 from the centrosome via direct association of mutDISC1 with endogenous wtDISC1. Furthermore, the detachment of endogenous wtDISC1 by mutDISC1 leads to the redistribution of the dynein protein complex from the centrosome, as shown by the disappearance of LIS1 and p150glued, but not γ-tubulin, from the centrosome in the presence of mutDISC1. Expression of mutDISC1 in cells did not alter wtDISC1 expression level per se (data not shown). As a result, mutDISC1 inter-feres with a key role of DISC1 in anchoring the dynein protein complex at the centrosome. Taken together, we postulate that the chromosomal abnormality in the Scottish family results in overall loss of DISC1 func-tion for the dynein protein complex at the centrosome, impaired neur-ite outgrowth and neuronal migration. Because the functional effect of mutDISC1 as a dominant-negative is mild in neurons, it is reasonable to suggest that partial impairment of DISC1 function may occur in the brains of Scottish patients.
DISC1
LIS1
p150glued
Merge
Merge
DISC1
LIS1
Merge
Merge
p150gluedMerge
Merge
Merge
Mergeγ-tubulin γ-tubulin
a
b
Ret
entio
n of
LIS
1/d
ynei
nm
achi
nery
at
cent
roso
me
(per
cent
age)
0
50
100
mutDISC1 mutDISC1∆(403−504)
LIS1 p150glued γ-tubulin
Figure 4 MutDISC1 affects a key function of wtDISC1 in anchoring the dynein motor proteins at the centrosome. (a) Overexpression of mutated Disrupted-In-Schizophrenia-1 (mutDISC1; green) redistributes endogenous DISC1, LIS1 and p150glued from the centrosome, but the localization of γ-tubulin, the key structural component of the centrosome, is not affected by expression of mutDISC1. The effect of the redistribution is much weaker in the presence of mutDISC1∆(403–504). Endogenous DISC1, LIS1, p150glued and γ-tubulin are stained in red, and those at the centrosome are specifically indicated with arrowheads. The cells with expression of mutDISC1 lose
endogenous DISC1, LIS1 and p150 glued at the centrosome (white arrows). Localization of γ-tubulin is unchanged in the presence of mutDISC1 (yellow arrow). The cells with expression of mutDISC1∆(403–504), as well as cells without exogenous DISC1, maintain endogenous DISC1, LIS1, p150 glued and γ-tubulin at the centrosome (arrowheads). Scale bar, 10 µm. (b) Under these experimental conditions, approximately 80% of LIS1 and p150glued are dissociated from the centrosome as a result of detachment of DISC1 from the centrosome. Three independent experiments were performed, with 100 cells being counted per group.
print ncb1328.indd 1173print ncb1328.indd 1173 1/12/05 3:04:12 pm1/12/05 3:04:12 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

1174 NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005
A RT I C L E S
There is increasing evidence that supports the possibility that DISC1 may be a candidate gene for schizophrenia in general populations3–6. In addition, we have found another mutation of DISC1 that could lead to a C-terminal protein truncation in individuals from a family with schizo-phrenia 40. The missing portion of the second C-terminal DISC1 trunca-tion (amino acids 809–854) includes the domain that is responsible for binding to NUDEL (M.T., A.K., and A.S., unpublished observations). Although this second DISC1 mutation was found in just one small fam-ily, it may help to confirm a key role for DISC1 mutations in schizophre-nia, just as the second mutation in α-synuclein (A30P), although also
found in a small family, helped to confirm that this gene is involved in the pathogenesis of Parkinson’s disease41. We do not exclude additional mechanisms for the actions of mutDISC1 that lead to cytoarchitectural abnormalities. Hayashi and colleagues42 have reported that DISC1 inhibits an oligopeptidase activity in NUDEL that can be disturbed by the Scottish mutation. Taken together, it is possible that various genetic variations of DISC1 may lead to loss of DISC1 function and increase susceptibility to schizophrenia.
Neuroanatomical studies of individuals with schizophrenia indicate a range of cytoarchitectural deficits7–10. These include mild alterations of
HA
d
Cel
ls w
ith n
eurit
es (p
erce
ntag
e)
a b
*
β-tubulin
β-tubulin
†
**
0
10
20
30
40
50
††
Nucleus Nucleus + GFP
RNAi#1
RNAi Con
c
e
RNAi#1
DISC1
pDsRed−CETN2
Merge
98
62
Mr(K)
98DISC1
PC12
LIS1
NUDEL NUDEL Merge
p150glued p150glued
Mock mutDISC1
PC12
Merge
LIS1 Merge
IB:anti-
exo
DIS
C1
end
o D
ISC
1
PC12
Cortical neurons
HeLa
PC12
RNAi Con
Mock RNAicon
RNAi#1 RNAi#2 wtDISC1
mutDISC1
MockRNAi1+wt
RNAi#1 RNAi#2RNAiCon
Figure 5 Requirement of DISC1 for neurite outgrowth in neuronal cells. (a) Influence of small hairpin RNA (shRNA; RNA interference (RNAi)#1, #2) on exogenous (exo) rodent Disrupted-In-Schizophrenia-1 (DISC1) protein levels in HeLa cells was evaluated by western blotting (upper panels). RNAi#1 and #2 suppress 90% and 40% of DISC1 expression, respectively. Endogenous (endo) DISC1 (relative molecular mass of 100,000) in the differentiated PC12 cells is also markedly suppressed with RNAi#1, slightly with RNAi#2 but not with RNAi Con (control) (lower panels). IB indicates antibodies used for western blotting. K, relative molecular mass in thousands. (b) Suppression of endogenous DISC1 in primary cortical neurons with RNAi to DISC1. Accumulation of DISC1 (green) at the centrosome (arrowheads), which is co-stained with a marker of the centrosome (pDsRed–CETN2, red), disappears after transfection of RNAi#1 (left), but not of RNAi Con (right). Arrows indicate the centrosomes without accumulation of DISC1 following treatment with by RNAi#1.
Scale bar, 5 µm. (c) No signs of apoptotic cell death, such as nuclear condensation and fragmentation, in PC12 cells treated with RNAi to DISC1. Scale bar, 20 µm. GFP, green fluorescent protein. (d) Percentages of differentiated PC12 cells harbouring neurites 3 d after transfection of mock vector, RNAi control vector, RNAi#1, RNAi#2, wtDISC1, mutDISC1 or RNAi#1 with human wtDISC1. RNAi#1 effectively suppresses neurite outgrowth (†P < 0.001, ††P < 0.001). RNAi#2 has a lesser effect compared with RNA#1. Furthermore, co-expression with human wtDISC1 rescues neurite outgrowth from the effects of RNA#1 (*P < 0.005). mutDISC1 shows a similar effect to RNAi#2 at suppressing neurite outgrowth (**P < 0.05). (e) Disturbance of LIS1, NUDEL and p150glued at the centrosome by transfection with mutDISC1. Cells with mutDISC1 (stained in green) lack the centrosomal accumulation of these proteins (arrows), but cells without mutDISC1 or mock transfection show substantial centrosomal accumulation of these proteins (arrowheads). Scale bar, 10 µm.
print ncb1328.indd 1174print ncb1328.indd 1174 1/12/05 3:04:15 pm1/12/05 3:04:15 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005 1175
A RT I C L E S
MZ
CP
MZ
CP
IZ + SVZVZ
IZ + SVZVZ
a
c
b
RNAi Con RNAi#2 RNAi#1
P2 P14
01020304050607080
01020304050607080
†
††
GFP+PI
GFP
Post-migratorycells
Migratingcells
I
II/III
IV
V
VI
RNAi Con RNAi#2
pDsRed−CETN2/GFP/TO−PRO-3
0
5
10
15
20
25
30
35
Nuc
leus
-cen
tros
ome
dis
tanc
e (µ
m)
GFPRNAi C
on
RNAi#1
1/10
0RNAi#
1 1/
30RNAi#
2
Misoriented Oriented
100908070605040302010
0
RNAiCon RNAi#2 RNAi#1 RNAi
Con RNAi#2 RNAi#1
Per
cent
age
Per
cent
age Per
cent
age
CP VZIZ +SVZ
RNAiCon
RNAi#2 RNAi#1
*
***
**
*
**
Figure 6 Essential role of DISC1 in proper neuronal migration of the developing cerebral cortex in vivo. (a) Disturbed radial migration with RNA interference (RNAi) to Disrupted-In-Schizophrenia-1 (DISC1). A much reduced percentage of neurons reaches the cortical plate (CP), and the majority remains in the intermediate zone (IZ) and subventricular zone (SVZ) at P2 in the presence of RNAi#1 (†P < 0.0001, ††P < 0.0001). Therefore, the post-migratory neurons (neurons in the quarter of CP) are decreased significantly after treatment with both RNAi#2 and #1, according to their potency in suppressing DISC1 (*P < 0.001, **P < 0.0001). Green, cells with green fluoresent protein (GFP) and RNAi constructs; red, propidium iodide (PI). Scale bar, 100 µm. MZ, marginal zone; VZ, ventricular zone. (b) Disturbed dendritic arborization of cortical neurons at P14 with RNAi to DISC1. Neurons with RNAi Con (control) have vertical arbors that are orthogonal to the pial surface (arrows in the left bottom panel). Neurons with RNAi#2 have shorter arbors that were not oriented in the radial plane (arrowhead in the middle bottom panel). In sections with RNAi#1, some neurons were malpositioned with misoriented processes (arrowhead in the
right bottom panel). The ‘oriented’ vertical arbors were defined according to their angle towards the pial surface within ±15º (*P < 0.001, **P < 0.001). Scale bar, 200 µm, upper panel; 50 µm, lower panel. (c) Influence of DISC1 on the nucleus–centrosome coupling in neurons in the developing cerebral cortex. Neurons in the cortical plate (CP) were examined in the brain slices. The centrosomes and the nuclei were labelled with pDsRed–CETN2 in red (arrowheads) and with TO–PRO-3 iodide in blue, respectively. The distances between the nuclei and the centrosomes are shown by white lines. Neurons with RNAi Con show radial-oriented stereotypic morphology with a thick leading process that is typical of the migrating neurons in the CP (left panels). By contrast, the major leading process of neurons with RNAi#2 to DISC1 shows deformed morphology (right panels). The nucleus–centrosome distances are increased in neurons with RNAi to DISC1, in comparison with those with RNAi Con. Quantitative assessment was conducted with no RNAi construct (n = 62), RNAi Con (n = 84), 1/100 diluted RNAi#1 (n = 87), 1/30 diluted RNAi#1 (n = 70) or RNAi#2 (n = 83) (*P < 0.0001, **P < 0.0001) (right panel). Bars indicate standard errors. Scale bar, 20 µm.
print ncb1328.indd 1175print ncb1328.indd 1175 1/12/05 3:04:17 pm1/12/05 3:04:17 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group
1176 NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005
A RT I C L E S
dendrites and/or cell positioning. The abnormalities are far more subtle than mental retardation or the lissencephalies13,15. The anatomical infor-mation comes from analyses of autopsied brains from adult patients with schizophrenia; therefore, some anatomical changes during neurodevel-opment may be partially compensated. Furthermore, there are many confounding factors, including the effects of medications, drug abuse, malnutrition and stress. This may be one of the reasons why the neuropa-thology data, although suggestive of cytoarchitectural abnormalities dur-ing neurodevelopment, are not perfectly consistent with each other. To overcome the limitations of the neuropathology, cell and in vivo model systems based on known genetic mutations have significant advantages. DISC1 is particularly advantageous for model building, because DISC1 is known to have disease-associated mutations1,40.
The range of defects that we observed in neurite outgrowth and neu-ronal migration is consistent with multiple important roles for DISC1 in cortical histogenesis. Our data indicate that, although suppression of DISC1 leads to delayed neuronal migration, such a change may be compensated and retained as relatively minor cytoarchitectural abnor-malities at later time points. Our findings imply dose-dependent effects of DISC1 impairment on dynein-associated microtubular dynamics and cortical neuropathology, as severe reductions in DISC1 that are induced by RNAi resulted in a more pronounced failure of migration than did the addition of mutDISC1. In patients with the DISC1 genetic alterations that have been described so far, the levels of mutDISC1 are unknown, but may well be relatively low, which is consistent with either haploin-sufficiency or mild dominant-negative effects, either of which would
be fully consistent with our present data. Therefore, schizophrenia is likely to encompass a spectrum of developmental malformations, with specific psychiatric defects probably being related to subtle alterations in the basic steps of brain histogenesis — cell production, migration, neurite outgrowth and formation of the synaptic circuitry, which may be, at least in part, associated with microtubular dynamics.
METHODS
Reagents and antibodies. All reagents were from Sigma (St Louis, MO) or Invitrogen (Carlsbad, CA), except where indicated. Protein concentration was determined using the BCA Protein Assay Reagent (Pierce Biotechnology, Rockbond, IL). Preparation of antibodies against DISC1 has been described previously28. An affinity-purified rabbit antiserum against green fluorescent protein (GFP; Molecular Probes, Eugene, OR) and a mouse monoclonal anti-body against GFP (Molecular Probes) were used to visualize the morphology of GFP-transfected PC12 cells in the neurite outgrowth assay and staining of cortical slices. The following antibodies were also used: mouse monoclonal anti-p150glued antibody (Transduction Laboratories, San Jose, CA); mouse monoclonal anti-CaM kinase II (Upstate, Charlottesville VA); mouse monoclonal antibodies against β-tubulin, γ-tubulin and dynein IC (Sigma); rabbit polyclonal anti-His tag antibody (MBL, Nagoya, Japan); rabbit polyclonal anti-GABA (Sigma); mouse monoclonal antibodies against HA-tag, myc-tag and anti-TuJ1 (BAbCO, Berkeley, CA). The rabbit polyclonal anti-LIS1 and rat monoclonal anti-NUDEL antibody were gifts from Dr Li-Huei Tsai (Harvard Medical School) and Dr Hiroyuki Arai (University of Tokyo), respectively. pDsRed–CETN2 was a gift from Dr Joseph G. Gleeson (University of California). The vector system for small hairpin RNA (shRNA) was used to suppress endogenous DISC1 protein expression35. In this study, we produced seven shRNA plasmids and selected two representative ones with the following sequences for the present study: RNAi#1 with strong
P2
MZ
CP
IZ + SVZVZ
P14a b
0
10
20
30
40
50
60
70
80
0
20
40
60
80
100
* *****
Post-migratorycells
Migratingcells
GFP mutDISC1 RNAi#2
Misoriented Oriented
10
30
50
70
90
Merge GFP mutDISC1 Merge GFP mutDISC1 P
erce
ntag
e
Per
cent
age
GFP mutDISC1 RNAi#2
Figure 7 mutDISC1 impairs proper neurodevelopment in the cerebral cortex in vivo. (a) Electroporation of mutated Disrupted-In-Schizophrenia-1 (mutDISC1) resulted in delayed neuronal migration in that a portion of the process-bearing cells expressing HA-tagged mutDISC1 (co-transfected with a green fluorescent protein (GFP) expression construct) does not complete migration at P2 (arrowheads). The percentage of neurons that have completed migration (neurons in the quarter of cortical plate (CP), defined as post-migratory cells) at P2 is decreased in the presence of mutDISC1, to a similar extent to that seen in the brain slices with RNAi#2 (milder RNA interference (RNAi)) (*P < 0.001, **P < 0.001). In addition to GFP,
sections are immunostained with an anti-HA antibody (in red). Scale bar, 200 µm. IZ, intermediate zone; MZ, marginal zone; SVZ, subventricular zone; VZ, ventricular zone. (b) At P14, brain slices expressing mutDISC1 display disturbed dendritic arborization similar to those with RNAi treatment. Neurons expressing mutDISC1 frequently display disarray of vertical arbors (arrowheads), to a similar extent to that seen in brain slices with RNAi#2 (milder RNAi) (*P < 0.0001, **P < 0.001). The ‘oriented’ vertical arbors were defined according to their angle towards the pial surface within ±15º. In addition to GFP, sections were immunostained with an anti-HA antibody (in red). Scale bar, 25 µm.
print ncb1328.indd 1176print ncb1328.indd 1176 1/12/05 3:04:26 pm1/12/05 3:04:26 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005 1177
A RT I C L E S
suppression, 5′-GGCAAACACTGTGAAGTGC-3′; RNAi#2 with milder suppres-sion, 5′-CGGCTGAGCCAAGAGTTGG-3′.
These shRNAs target common sequences of mouse and rat DISC1. Therefore, the potency of shRNA was initially tested by the extent of suppression in co-transfected mouse DISC1 in HeLa cells. Once suppression against exogenous DISC1 was confirmed, selected shRNAs were further tested against endogenous DISC1 in rat PC12 cells as well as in mouse brain slices. Small oligonucleotides synthesized to the corresponding DISC1 sequences for small interfering RNA (siRNA) were from Dharmacon RNA Technologies (Lafayette, CO).
An expression construct of human DISC1 was used to test functional comple-mentation in the neurite outgrowth assay in PC12 cells with RNAi#1 against rat DISC1. Human DISC1 sequences have four mismatches to rodent DISC1 as follows: 5′-GGAAAATACTATGAAGTAC-3′ (bold: mismatched nucleotides). We have already confirmed that both RNAi#1 and #2 have no effects on human DISC1.
Cell culture, staining and neurite outgrowth assay. PC12 cells were maintained in Dulbecco’s Modified Eagle Medium (DMEM) with 10% fetal bovine serum (FBS) and 5% horse serum (HS). Differentiation was initiated by the addition of 50 ng ml–1 of nerve growth factor (NGF) with culture medium being changed to DMEM with 1% FBS and 1% HS. NGF was supplemented daily after differentia-tion. COS-7 and HeLa cells were maintained in DMEM with 10% FBS. Rodent primary cortical neurons were prepared as described previously43.
Transfection of si/shRNAs was conducted with a Mouse Neuron NucleofectorTM Kit (Amaxa, Gaithersburg, MD). Transfection of expression constructs or RNAi constructs was carried out with lipofectamine 2000 (Invitrogen) for PC12 cells and COS-7 cells, and Polyfect Transfection Reagent (Qiagen, Valencia, CA) for HeLa and HEK293 cells.
Cell staining was carried out as described previously44. In brief, cells were fixed with 3.7% paraformaldehyde in phosphate-buffered saline (PBS) and permea-bilized with 0.1% Triton X-100. For some staining, ice-cold methanol at –20˚C was used as the fixative.
Neurite outgrowth in PC12 cells was assayed as described previously28. In brief, neurite outgrowth was evaluated by the percentage of cells with processes longer than three cell-body diameters. The length of the longest process of each neurite-harbouring cell was measured. In this study, we added a minor modification: to obtain clearer images of cell morphology, cells co-transfected with various expres-sion or RNAi constructs with the GFP construct were stained with an anti-GFP antibody. A confocal microscope (Zeiss LSM 510 Meta, Grottingem, Germany) was used for epifluorescent image collections. A Zeiss Axiovert 135 microscope mounted with a charge-coupled device (CCD) camera (Roper Scientific CoolSnap HQ cooled 12 bit; Roper Scientific, Trenton, NJ) was used to obtain PC12 cell images in the neurite outgrowth assay. Cell morphology was analysed in a blinded manner. Statistical analyses were conducted by using a one-way ANOVA followed by post-hoc testing.
Line-scan. Line-scan analysis was carried out to evaluate the cellular distribution of dynein IC and dynactin (p150glued) antibodies, as described previously45. Line-scan was carried out through the midline of the cells, from one edge, over the nucleus and microtubule-organizing centre (MTOC) or centrosome, to the side opposite to the nucleus of the cells. Following this, the average gray levels for dynein or p150glued stains were plotted along the line by using Metamorph analysis software (Universal Imaging Corporation, Buckinghamshire, UK). A black area with no cells was chosen as background level. Neighbouring non-transfected cells were chosen as negative controls. At least three transfected or non-transfected cells were scanned for dynein and dynactin staining. For each cell scanned, the ratio of the highest value on the MTOC side to the highest value on the opposite side was calculated to evaluate the degree of dynein/dynactin accumulation at the MTOC.
Microtubule reorganization assays. The procedure followed a published proto-col34. COS-7 cells that were transfected either with HA-tagged wt or mutDISC1 or empty vector (mock) were pretreated with nocodazole at a final concentration of 5 µg ml–1 at 37ºC for 3 h. Cells were incubated for specified times to allow micro-tubules to reorganize and then fixed for 10 min with methanol at –20ºC.
Cell extractions. Cells were homogenized or solubilized in ice-cold lysis buffer (0.32 M sucrose, 50 mM HEPES, pH 7.4, 5 mM MgCl2, 5 mM 1,4-dithiothreitol (DTT), 1 mM phenylmethane sulfonylfluoride (PMSF), 1 mM EDTA, 1% Triton X-100 and a protease inhibitor mixture (Roche, Basel, Switzerland)).
Immunoprecipitation. Cells were lysed in a RIPA buffer (50 mM HEPES, pH 7.4, 150 mM NaCl, 5 mM MgCl2, 5 mM DTT, 1 mM PMSF, 1 mM EDTA, 1% Triton X-100 and protease inhibitor mixture), and supernatant fractions obtained after centrifugation at 10,000 g for 15 min were incubated with primary antibodies and protein A agarose (Oncogene, San Diego, CA). The immunoprecipitates were analysed with SDS–PAGE, followed by western blotting. ProFoundTM Mammalian HA Tag IP/Co-IP Kit and ProFoundTM
Mammalian Co-Immunoprecipitation Kit (Pierce Biotechnology) were also used for immunoprecipitation.
In vitro binding assay. Glutathione S-transferase (GST) and His-tagged recom-binant DISC1 protein fragments (amino acids 347–600) were incubated in 50 mM Tris-HCl, pH 7.4, 150 mM NaCl and 0.1 mg ml–1 bovine serum albumin (BSA) for 1 h. His-tagged DISC1 bound to GST-tagged DISC1 was precipitated with glutath-ione Sepharose beads. The protein precipitates were analysed with SDS–PAGE, followed by western blotting with an antibody against His-tag.
Subcellular fractionation. The fractionation was performed as described previ-ously43. For the glycerol density-gradient centrifugation, cells with HA-tagged wtDISC1 and/or Myc-tagged mutDISC1 were lysed in a buffer (20 mM HEPES, pH 7.4, 1% Triton X-100, 1 mM EDTA, 150 mM NaCl, 1 mM DTT and protease inhibitor mixture), and centrifuged at 3,000 g for 10 min. The supernatants were loaded on top of 4 ml of a 10–25% continuous glycerol gradient and centrifuged at 20,000 g for 24 h. Each fraction was analysed by western blotting with anti-HA, Myc or p150glued antibodies.
In utero electroporation. All animal experiments were performed according to the guidelines of the Keio University School of Medicine. Pregnant ICR mice were deeply anaesthetized, and their intrauterine embryos were surgically manipulated, as described previously46. The RNAi plasmids that had been characterized fully in vitro, including expression studies and neurite outgrowth assays in PC12 cells, were used for in utero electroporation at E14.5. Plasmid solutions at a concentra-tion of 2.5 µg µl–1 in 1–2 µl were introduced directly into the ventricular zone by in utero electroporation, as reported previously36. To confirm the specificity of the effects, dilution series of each plasmid (RNAi plasmids, expression plasmid) in 1–2 µl were introduced, and their dose-correlated effects were confirmed, respec-tively. The GFP expression vector with CAG vector was co-transfected with RNAi constructs at a concentration of 2.5 µg µl–1.
Brain slice preparation and staining. The coronal slices of developing cerebral cortex were prepared as described previously36. Briefly, the brains were fixed with 4% paraformaldehyde and sectioned with a cryostat at 20 µm. Green fluorescent images were captured directly in some samples, and immunofluorescent staining was performed in others. The primary antibodies that were applied in the brain slices included anti-DISC1 (1:50 dilution), anti-TuJ1 (1:1000), anti-GABA (1:500), anti-CaM Kinase II (1:200) and anti-GFP (1:500). Nuclei were labelled with propidium iodide (Molecular Probes). Images of the slices were acquired with a confocal microscope (Zeiss LSM510 and Olympus Optical FV300).
Quantitative analysis of brain slices. To quantify the pattern of migration, the numbers of GFP-positive cells in the ventricular zone, the subventricular zone/intermediate zone and the cortical plate were counted from three independent sections. We defined labelled neurons in the quarter of cortical plate, the super-ficial cortical plate, to be post-migratory, whereas the labelled cells in other layers of cortical plate, as well as in the intermediate zone and the subventricular zone, were labelled as migrating neurons. The numbers of neurons in each layer or each category from more than five independent experiments were counted in a blind manner.
Orientation of the vertical arbors was evaluated as follows. The angular dis-placement of the dendrite from the direction to the pial surface was measured. The ‘oriented’ vertical arbors were defined according to their angles towards the pial surface within ±15º. The percentage of neurons possessing the oriented verti-cal arbors towards the pial surface was calculated. To analyse the vertical arbor orientation, a minimum of 50 neurons was scored from five independent sec-tions in a blinded manner. Representative data from more than five independent experiments are presented in the figures. Statistical analyses were conducted by using a one-way ANOVA followed by post-hoc testing.
print ncb1328.indd 1177print ncb1328.indd 1177 1/12/05 3:04:32 pm1/12/05 3:04:32 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

1178 NATURE CELL BIOLOGY VOLUME 7 | NUMBER 12 | DECEMBER 2005
A RT I C L E S
Nucleus–centrosome coupling. RNAi constructs, together with expression constructs of GFP and pDsRed–CETN2, a marker for the centrosome, were introduced into embryos at E14.5 by using in utero electroporation. The molar ratio of GFP, pDsRed–CETN2 and RNAi constructs introduced in this experi-ment was 1:1:3 (1, 1 and 3 µg µl–1), respectively. The developing cerebral cortices at E18.5 were sectioned in coronal planes with a cryostat at 20 µm. The cen-trosomes and the nuclei were labelled with pDsRed–CETN2 and TO–PRO-3 (Molecular Probes), respectively. Images of neurons in the cortical plate were randomly acquired with confocal microscopes (Zeiss LSM510 and Olympus Optical FV300). Post-migratory neurons that completed the migration in the one-fourth of the cortical plate (the superficial cortical plate, as described) were excluded from the assay. The distance between the nucleus and the centrosome was examined for more than 60 migrating neurons in cortical plate per group with the software Image J (http://rsb.info.nih.gov/ij/) in a blind manner. One-way ANOVA followed by post-hoc testing was used for statistical analyses.
Note: Supplementary Information is available on the Nature Cell Biology website.
ACKNOWLEDGEMENTSWe thank Y. Lema for preparation of the figures. We thank P. Talalay, D. Solecki, T. Sedlak, N. Katsanis, J. Cheah and S. Snyder for critical reading of the manuscript. We thank T. Oe for technical support of the neurite outgrowth assay, and M. Peak for technical assistance. This work was supported by a U.S. Public Health Service Grant (MH-069853) to A.S. It was also supported by foundation grants from NARSAD, Stanley, S-R and funds from the departments (A.S.); NARSAD and Stanley (C.A.R.); MEXT, JSPS, Kanehara, Casio, Novaltis and Sumitomo foundations (K.N.).
COMPETING INTERESTS STATEMENTThe authors declare that they have no competing financial interests.
Published online at http://www.nature.com/naturecellbiologyReprints and permissions information is available online at http://npg.nature.com/reprintsandpermissions
1. Millar, J. K. et al. Disruption of two novel genes by a translocation co-segregating with schizophrenia. Hum. Mol. Genet. 9, 1415–1423 (2000).
2. Blackwood, D. H. et al. Schizophrenia and affective disorders — cosegregation with a translocation at chromosome 1q42 that directly disrupts brain-expressed genes: clinical and P300 findings in a family. Am. J. Hum. Genet. 69, 428–433 (2001).
3. Ekelund, J. et al. Chromosome 1 loci in Finnish schizophrenia families. Hum. Mol. Genet. 10, 1611–1617 (2001).
4. Harrison, P. J. & Weinberger, D. R. Schizophrenia genes, gene expression, and neuropa-thology: on the matter of their convergence. Mol. Psychiatry 10, 40–68 (2005).
5. Hodgkinson, C. A. et al. Disrupted in schizophrenia 1 (DISC1): association with schizophrenia, schizoaffective disorder, and bipolar disorder. Am. J. Hum. Genet. 75, 862–872 (2004).
6. Hennah, W. et al. Haplotype transmission analysis provides evidence of association for DISC1 to schizophrenia and suggests sex-dependent effects. Hum. Mol. Genet. 12, 3151–3159 (2003).
7. Akbarian, S. et al. Altered distribution of nicotinamide-adenine dinucleotide phos-phate-diaphorase cells in frontal lobe of schizophrenics implies disturbances of cortical development. Arch. Gen. Psychiatry 50, 169–177 (1993).
8. Lewis, D. A. & Levitt, P. Schizophrenia as a disorder of neurodevelopment. Annu. Rev. Neurosci. 25, 409–432 (2002).
9. Weinberger, D. R. Implications of normal brain development for the pathogenesis of schizophrenia. Arch. Gen. Psychiatry 44, 660–669 (1987).
10. Harrison, P. J. The neuropathology of schizophrenia. A critical review of the data and their interpretation. Brain 122, 593–624 (1999).
11. Sawa, A. & Snyder, S. H. Schizophrenia: diverse approaches to a complex disease. Science 296, 692–695 (2002).
12. Bielas, S., Higginbotham, H., Koizumi, H., Tanaka, T. & Gleeson, J. G. Cortical neuronal migration mutants suggest separate but intersecting pathways. Annu. Rev. Cell Dev. Biol. 20, 593–618 (2004).
13. Ross, M. E. & Walsh, C. A. Human brain malformations and their lessons for neuronal migration. Annu. Rev. Neurosci. 24, 1041–1070 (2001).
14. Hatten, M. E. New directions in neuronal migration. Science 297, 1660–1663 (2002).
15. Gupta, A., Tsai, L. H. & Wynshaw-Boris, A. Life is a journey: a genetic look at neocortical development. Nature Rev. Genet. 3, 342–355 (2002).
16. Reiner, O. et al. Isolation of a Miller-Dieker lissencephaly gene containing G protein β-subunit-like repeats. Nature 364, 717–721 (1993).
17. Shu, T. et al. Ndel1 operates in a common pathway with LIS1 and cytoplasmic dynein to regulate cortical neuronal positioning. Neuron 44, 263–277 (2004).
18. Hirotsune, S. et al. Graded reduction of Pafah1b1 (Lis1) activity results in neuronal migration defects and early embryonic lethality. Nature Genet. 19, 333–339 (1998).
19. Wynshaw-Boris, A. & Gambello, M. J. LIS1 and dynein motor function in neuronal migration and development. Genes Dev. 15, 639–651 (2001).
20. Assadi, A. H. et al. Interaction of reelin signaling and Lis1 in brain development. Nature Genet. 35, 270–276 (2003).
21. Niethammer, M. et al. NUDEL is a novel Cdk5 substrate that associates with LIS1 and cytoplasmic dynein. Neuron 28, 697–711 (2000).
22. Sasaki, S. et al. A LIS1/NUDEL/cytoplasmic dynein heavy chain complex in the develop-ing and adult nervous system. Neuron 28, 681–696 (2000).
23. Ahmad, F. J., Echeverri, C. J., Vallee, R. B. & Baas, P. W. Cytoplasmic dynein and dyn-actin are required for the transport of microtubules into the axon. J. Cell Biol. 140, 391–401 (1998).
24. Waterman-Storer, C. M. et al. The interaction between cytoplasmic dynein and dynactin is required for fast axonal transport. Proc. Natl Acad. Sci. USA 94, 12180–12185 (1997).
25. Solecki, D. J., Model, L., Gaetz, J., Kapoor, T. M. & Hatten, M. E. Par6α signaling controls glial-guided neuronal migration. Nature Neurosci. 7, 1195–1203 (2004).
26. Tanaka, T. et al. Lis1 and doublecortin function with dynein to mediate coupling of the nucleus to the centrosome in neuronal migration. J. Cell Biol. 165, 709–721 (2004).
27. Tsai, L. H. & Gleeson, J. G. Nucleokinesis in neuronal migration. Neuron 46, 383–388 (2005).
28. Ozeki, Y. et al. Disrupted-in-Schizophrenia-1 (DISC-1): mutant truncation prevents binding to NudE-like (NUDEL) and inhibits neurite outgrowth. Proc. Natl Acad. Sci. USA 100, 289–294 (2003).
29. Morris, J. A., Kandpal, G., Ma, L. & Austin, C. P. DISC1 (Disrupted-In-Schizophrenia 1) is a centrosome-associated protein that interacts with MAP1A, MIPT3, ATF4/5 and NUDEL: regulation and loss of interaction with mutation. Hum. Mol. Genet. 12, 1591–1608 (2003).
30. Millar, J. K., Christie, S. & Porteous, D. J. Yeast two-hybrid screens implicate DISC1 in brain development and function. Biochem. Biophys. Res. Commun. 311, 1019–1025 (2003).
31. Brandon, N. J. et al. Disrupted in Schizophrenia 1 and Nudel form a neurodevelop-mentally regulated protein complex: implications for schizophrenia and other major neurological disorders. Mol. Cell Neurosci. 25, 42–55 (2004).
32. Miyoshi, K. et al. DISC1 localizes to the centrosome by binding to kendrin. Biochem. Biophys. Res. Commun. 317, 1195–1199 (2004).
33. Miyoshi, K. et al. Disrupted-In-Schizophrenia 1, a candidate gene for schizophrenia, participates in neurite outgrowth. Mol. Psychiatry 8, 685–694 (2003).
34. Smith, D. S. et al. Regulation of cytoplasmic dynein behaviour and microtubule organi-zation by mammalian Lis1. Nature Cell Biol. 2, 767–775 (2000).
35. Yu, J. Y., DeRuiter, S. L. & Turner, D. L. RNA interference by expression of short-inter-fering RNAs and hairpin RNAs in mammalian cells. Proc. Natl Acad. Sci. USA 99, 6047–6052 (2002).
36. Tabata, H. & Nakajima, K. Efficient in utero gene transfer system to the developing mouse brain using electroporation: visualization of neuronal migration in the developing cortex. Neuroscience 103, 865–872 (2001).
37. Bai, J. et al. RNAi reveals doublecortin is required for radial migration in rat neocortex. Nature Neurosci. 6, 1277–1283 (2003).
38. Xie, Z., Sanada, K., Samuels, B. A., Shih, H. & Tsai, L. H. Serine 732 phosphorylation of FAK by Cdk5 is important for microtubule organization, nuclear movement, and neuronal migration. Cell 114, 469–482 (2003).
39. Tarricone, C. et al. Coupling PAF signaling to dynein regulation: structure of LIS1 in complex with PAF-acetylhydrolase. Neuron 44, 809–821 (2004).
40. Sachs, N. A. et al. A frameshift mutation in Disrupted in Schizophrenia 1 in an American family with schizophrenia and schizoaffective disorder. Mol. Psychiatry 10, 758–764 (2005).
41. Kruger, R. et al. Ala30Pro mutation in the gene encoding α-synuclein in Parkinson’s disease. Nature Genet. 18, 106–108 (1998).
42. Hayashi, M. A. et al. Inhibition of NUDEL (nuclear distribution element-like)-oli-gopeptidase activity by disrupted-in-schizophrenia 1. Proc. Natl Acad. Sci. USA 102, 3828–3833 (2005).
43. Sawa, A., Khan, A. A., Hester, L. D. & Snyder, S. H. Glyceraldehyde-3-phosphate dehy-drogenase: nuclear translocation participates in neuronal and nonneuronal cell death. Proc. Natl Acad. Sci. USA 94, 11669–11674 (1997).
44. Sawa, A. et al. Increased apoptosis of Huntington disease lymphoblasts associated with repeat length-dependent mitochondrial depolarization. Nature Med. 5, 1194–1198 (1999).
45. Tomoda, T., Kim, J. H., Zhan, C. & Hatten, M. E. Role of Unc51.1 and its binding partners in CNS axon outgrowth. Genes Dev. 18, 541–558 (2004).
46. Nakajima, K., Mikoshiba, K., Miyata, T., Kudo, C. & Ogawa, M. Disruption of hippoc-ampal development in vivo by CR-50 mAb against reelin. Proc. Natl Acad. Sci. USA 94, 8196–8201 (1997).
print ncb1328.indd 1178print ncb1328.indd 1178 1/12/05 3:04:37 pm1/12/05 3:04:37 pm
Nature Publishing Group© 2005
© 2005 Nature Publishing Group

NATURE CELL BIOLOGY ADVANCE ONLINE PUBLICATION 1
E R R AT U M
Owing to a technical error, the pages of this manuscript were origi-nally mis-numbered by a 100 pages. Th is has now been corrected on-line. Th e corrected online manuscript is numbered 100 pages higher than the mis-numbered version.
© 2005 Nature Publishing Group

S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 1
Total soluble Total pellet Total
p150
Dyn IC
wtDISC1-HA
β-tubulin
glued wtDISC1 + – + – + –
*
*
Supplementary Information Figure S1 (a) Distribution changes of Dyn IC and p150 by overexpression of wtDISC1 in COS-7 cells. Overexpression of wtDISC1 in COS7 cells leads to translocation of Dyn IC and p150 , but not β-tubulin, to the pellet fraction, which reportedly contains the centrosomal protein complex . (b) MutDISC1 affects the centrosomal localization of LIS1 and DynIC. After nocodazole washout, Lis1 and DynIC accumulate to the centrosome in COS7 cells with mock transfection, but not in COS7 cells expressing mutDISC1. Arrowheads: accumulated LIS1 or DynIC. Scale bar: 10µm.(c) Co-immunoprecipitation of endogenous DISC1 with HA-tagged mutDISC1, but not HA-GAPDH, in differentiated PC12 cells. Immunoprecipitation with an anti-HA antibody was followed by analysis of the immunoprecipitates on SDS-PAGE and Western blotting with an antibody against the C-terminal domain of DISC1 (the domain deficient in mutDISC1).(d) Quantitative representation of the effects of mutDISC1 or mutDISC1∆(403-504) on redistribution of endogenous DISC1, LIS1, and p150 from COS7 cells. Their effects are different from each other (*: p<0.001) in 3 independent experiments with 100 cells counted per group.
REFERENCE1. Smith, D. S. et al., Regulation of cytoplasmic dynein behaviour and microtubule organization by mammalian Lis1. Nature Cell Biol. 2, 767-775 (2000).
glued
glued
1
Mock DynIC
mutDISC1DynIC Merge
Mock LIS1
mutDISC1 LIS1 Merge
0 min
10 min
30 min
Noc
0 min
10 min
30 min
Noc
a
b
Tf:
DISC1
mutD
ISC1
GAPDH
mutD
ISC1
GAPDH
IP Input
Co-IPc d
0
20
40
60
80 DISC1LIS1
γ-tubulin
p150glued
*
* *
mutDISC1 mutDISC1 (403-504)
% C
ells
Re-
dis
trib
ute
d
fro
m c
entr
oso
me
glued
© 2005 Nature Publishing Group
© 2005 Nature Publishing Group

S U P P L E M E N TA RY I N F O R M AT I O N
2 WWW.NATURE.COM/NATURECELLBIOLOGY
DISC1
GFP
Merge
DISC1
GFP
Merge
a
c
RNAi RNAi RNAi#1 #2 Con
PC12
bRNAi RNAi RNAi#1 #2 Con
Supplementary Information Figure S2(a) Suppression of endogenous DISC1 by shRNA in differentiated PC12 cells.The representative images are depicted 3 days after transfection of RNAi#1 or RNAi#2. Red,DISC1; Green, co-transfected GFP. Cells with RNAi#1 (arrows) display dramatic decreasein endogenous DISC1, in contrast to cells with control RNAi (RNAi Con) (arrowheads). Scalebar: 20µm.(b) Representative morphology of differentiated PC12 cells expressing RNAi#1, RNAi#2, orRNAi control with GFP. Scale bar: 50µm.(c) Inhibition of neurite outgrowth in primary cortical cultures is observed with transfection ofRNAi#1 to DISC1 (*p<0.0001). The length of the longest extension in each neuron wasmeasured in three groups (RNAi Con, RNAi#1, and RNAi#2). A total of 330 neurons wereexamined in a blinded manner.
SUPPLEMENTARY METHODSNeurite outgrowth assay of primary cortical neurons. Neurite outgrowth of primary corticalneurons was assayed as follows. RNAi constructs together with a GFP expression constructwere introduced into embryos at E14.5 by in utero gene transfer technique. The dissociatedneuronal cultures were prepared from the embryos at E17.5. The longest neurite of each neuronwith RNAi stained in green was measured 1 day after the plating by using the software Image J(http://rsb.info.nih.gov/ij/). Over 100 neurons per group were evaluated in a blinded manner.Statistical analyses were conducted by using a one-way ANOVA followed by post hoc test.
RNAi RNAi RNAiCon #1 #2
*(µm)
cortical neuron
Leng
thof
neur
ites
0
30
60
90
120
PC12
© 2005 Nature Publishing Group
© 2005 Nature Publishing Group

S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 3
Supplementary Information Figure S3 Knockdown expression of endogenous DISC1 by introduction of shRNA to DISC1 in the developing cerebral cortex.(a) Endogenous DISC1 levels at postnatal day 2 (P2) are suppressed by transfer of shRNA to DISC1 (the DISC1 RNAi#1 plasmid) at embryonic day 14.5 (E14.5) using electroporation. Green, cells with GFP and RNAi constructs; red, endogenous DISC1. The regions with suppressed DISC1 with RNAi are indicated by white arrowheads. Expression construct for GFP is with the CAG promoter. Scale bar: 100 µm. (b) Suppression of endogenous DISC1 by shRNA to DISC1 at P2 in high magnification pictures. Green, cells with GFP and RNAi constructs; red, endogenous DISC1. In target cells of gene transfer indicated in green, DISC1 is suppressed in the presence of RNAi#1, but not of RNAi control (RNAi Con). In the merged pictures, yellow signals occur only in the target cells with RNAi Con (arrows), but not in cells with RNAi#1, indicating the selective suppression by RNAi to DISC1. CP, cortical plate; IZ, intermediate zone. Scale bar: 10 µm.(c) Confirmation that the target cells with RNAi Con or RNAi#1 are mainly neuronal progenitor cells in the ventricular zone (VZ). Target cells with GFP are co-stained with neuronal markers such as TuJ1. Scale bar: 10 µm.(d) The target cells with RNAi Con or RNAi#1 in mouse cortices at P14 were co-stained with a marker of the pyramidal neurons, CaMKII (right panels), but not co-stained with a marker for the interneurons, GABA (left panels). Yellow signals in the merged pictures indicate the co-staining. Scale bar: 10µm.
REFERENCE1. Niwa, H. et al., Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 108, 193-199 (1991).
1
a b
c d
RNAi Con
RNAi #1
RNAi Con
RNAi #1
GFP DISC1 Merge
GFP TuJ1 Merge
RNAi Con
RNAi #1
RNAi Con
RNAi #1
CP
IZ
GFP DISC1 Merge
RNAi Con
RNAi #1
GABA Merge
CaMKII
CaMKII
Merge
Merge
P2 P2
P2 P14GABA Merge
1
© 2005 Nature Publishing Group
© 2005 Nature Publishing Group

S U P P L E M E N TA RY I N F O R M AT I O N
4 WWW.NATURE.COM/NATURECELLBIOLOGY
γ-tubulin
Dyn IC
p150
NUDEL
GST-DISC1+His-D
ISC1
GST+His-DISC1
DISC1 GAPDH Input DISC1 GAPDH Input DISC1 GAPDH Input
wtDISC mutDISC1
GAPDH Input wtDISC
mutDISC1
GAPDH Input
wtDISC mutDISC1
GAPDH Input
NUDEL
IP: anti- IP: anti- IP: anti-
98
62
49
188
38
28
98
62
49
188
38
28
98
62
49
188
38
28
98
62
49
38
28
γ-tubulin98
62
49
188 glued98
62
49
188
38
28
(kDa)(kDa) (kDa)
(kDa) (kDa)(kDa)
98
6249
188
38
28
98
6249
188
38
28
10% 25%
wt-HA
wt-HA, mut-myc
Glycerol gradient
IB:with anti-HA
1 2 3 4 5 6 7 8 9 10 11 12 13 14
(kDa)
His-DISC1(347-600aa)
98
62
49
38
28
(kDa)
17
a b c
d e f
g
h
Supplementary Information Figure S4 The full scan images of a, b, and c correspond to Figure 1a. The full scan images of d, e, and f correspond to Figure 1b. The full scan image of g corresponds to Figure 3b. The full scan image of h corresponds to Figure 3d.
© 2005 Nature Publishing Group
© 2005 Nature Publishing Group





























