Embed Size (px)
Citation preview

A HIGH-RESOLUTION HYDRODYNAMIC INVESTIGATION OF BROWN TROUT(SALMO TRUTTA) AND RAINBOW TROUT (ONCORHYNCHUS MYKISS) REDDS
M. A. MARCHILDON,a W. K. ANNABLE,a* J. G. IMHOFb and M. POWERc
a Department of Civil and Environmental Engineering, University of Waterloo, Waterloo, Ontario, N2L 3G1 Canadab Trout Unlimited Canada c/o Rm. 270, Axelrod Building, University of Guelph, Guelph, Ontario, N1G 2W1 Canada
c Department of Biology, University of Waterloo, Waterloo, Ontario, N2L 3G1 Canada
ABSTRACT
High-resolution velocity measurements were taken over a series of redds on a gravel-bed stream using a Pulse Coherent AcousticDoppler Profiler (PCADP) to quantify the hydrodynamics of brown trout (Salmo trutta) and rainbow trout (Oncorhynchus mykiss)redds. On redds studied, over 4500 velocity measurements per redd were acquired per day to quantify the flow velocity, flow depth andrelated fluid mechanics metrics of Reynolds numbers, Froude numbers and turbulent kinetic energy per unit area. Results showed thatvelocity and Froude numbers varied widely at the redd scale, but consistently showed higher velocities and Froude numbers over thetailspill regions relative to the surrounding study limits. Results of Reynolds numbers calculations showed no apparent correlations tospawning location preference and redd structure. Turbulent kinetic energy per unit area consistently demonstrated a strong correlationwith redd locations. The metric maintained low values (i.e. unidirectional flow with little turbulence) where all redds and attemptedredds were observed. The study also demonstrates that a number of hydraulic metrics and several spatial scales will likely be necessaryto understand any inherent relationship between river hydraulics and redd placement. Copyright # 2010 John Wiley & Sons, Ltd.
key words: fish habitat; hydraulics; fluvial geomorphology; redd selection
Received 22 August 2009; Revised 9 December 2009; Accepted 18 December 2009
INTRODUCTION
As both biotic and abiotic research has progressed in the
study of water courses, it has becomewidely recognized that
a collaborative effort amongst scientific, engineering and
social disciplines must be incorporated into lotic system
research to provide effective management and rehabilitation
strategies (Maddock, 1999; Schwartz and Herricks, 2007).
Imhof et al. (1996) stressed the need to evaluate lotic
systems at multiple nested spatial and temporal scales to
characterize channel form and function. They identified that
stream health potential is determined by the characteristics
of the watershed and channel geomorphological function at
large scales (from the basin and sub-basin scales) down to
the river-reach scale. Conversely, the investigation of
potential stream health is realized through river function
as it pertains to small-scale or micro-scale level processes
such as channel shear, sediment transport and biodiversity.
With increasing scale, and thus watershed complexity,
micro-scale processes are commonly simplified or hom-
ogenized to address scalability in observations. Examples of
such simplifications include the velocity-area method
(Buchanan and Somers, 1969) used to quantify discharge,
Manning’s equation used to estimate average cross-sectional
channel velocity, Wolman’s (1954) pebble count used to
represent substrate surface roughness distribution, indicator
species used to represent aquatic diversity and health
(Statzner et al., 1988; Champoux et al., 2003) and habitat
suitability indices used to evaluate instream flow require-
ments at various discharge events (Raleigh et al., 1984,
1986).
Another common problem related to the physical metrics
used in the study of rivers is the disparity between the data
collected to characterize fine-scale physical habitat features
and the data collected at flood flow conditions that define
channel morphology and physical habitat forms. Biological
data are typically collected during low flow seasons when
rivers are wadable and aquatic life is observable (Smith,
1973; Shirvell and Dungey, 1983; Witzel and MacCrimmon,
1983a; Kondolf et al., 1993). In these conditions, the
hydraulic, sedimentological and geomorphological pro-
cesses dictating river form, function and hence physical
habitat features are nominal. Conversely, measurements
related to channel form and function are primarily measured
during flood stages (i.e. bankfull discharge and larger) when
the hydraulic and sedimentological processes are actively
defining channel geomorphology (Leopold et al., 1964). The
disparity in temporal sampling strategies between biological
and physical scientists inevitably results in distinctly
different descriptions of the same physical system where
a continuum of spatial and temporal states should exist.
RIVER RESEARCH AND APPLICATIONS
River Res. Applic. 27: 345–359 (2011)
Published online 31 January 2010 in Wiley Online Library(wileyonlinelibrary.com) DOI: 10.1002/rra.1362
*Correspondence to: W. K. Annable, Department of Civil and Environ-mental Engineering, University of Waterloo, Waterloo, Ontario, N2L 3G1Canada. E-mail: [email protected]
Copyright # 2010 John Wiley & Sons, Ltd.

The tendency of many river spawning fishes to construct
redds in the stream bed substrate results in a biophysical
junction between small-scale processes and large-scale river
form and function. Construction and release of fertilized ova
into porous substrates is one strategy to protect vulnerable
embryos from predation and physical disruption or damage.
The site-specific selection process of a spawning fish is
believed to be based upon the small-scale physical and
chemical conditions (Burner, 1951) moderated somewhat by
the size of the female (Kondolf and Wolman, 1993). Thus
the majority of redd placement studies have concentrated on
fine-scale habitat measurement (e.g. Ottaway et al., 1981;
Shirvell and Dungey, 1983; Grost et al., 1990; Essington
et al., 1998). However, one cannot neglect that the micro-
habitats that fish select for spawning are influenced by larger
morphological features (such as a run or a riffle) formed by
hydraulic and sediment transport processes that occur during
larger and less frequent discharge events. Thus, little work
has been done to examine differences in redd placement
characteristics as a function of macro-habitat units (but see
Zimmer and Power, 2006). Recent advances in high-
resolution three-dimensional flow measurement technology,
however, provides an opportunity to study the hydrodynamic
characteristics of redds and surrounding fluvial features in
unprecedented detail. To that end, the spawning habitat of
brown trout (Salmo trutta) and rainbow trout (Oncorhynchus
mykiss) redds were measured and evaluated to determine if
identifiable hydrodynamic properties existed that would aid
in the definition of spawning habitat and description of its
complexity.
BACKGROUND
Spawning begins with a female fish scanning the stream bed
for a suitable spawning location, a process referred to as the
pre-spawning stage (Burner, 1951). The female may carry
out exploratory cutting (Jones and Ball, 1954; Grost et al.,
1991), whereby the female, through vigorous movements of
her tail, induces a negative pressure on the stream bed
surface (Kondolf et al., 1993), subsequently releasing
sediment into the current for transport downstream. If the
female considers the site suitable, she continues cutting into
the stream bed, leaving behind a semi-spherical depression
referred to as the pit (Figure 1). The pit provides a controlled
current region (Figure 1b) where the ova are laid and
subsequently fertilized (Kondolf et al., 1993). After
oviposition, the female moves immediately upstream of
the pit to displace new sediment. The coarser sediment is
carried downstream, covering the pit to form what is termed
the tailspill, while the finer suspended sediment is carried
downstream beyond the limits of the redd (Kondolf et al.,
1993). The sequential process of pit excavation and tailspill
development is repeated to produce a redd composed of
multiple successive pits clustered in an elongated, or
teardrop, shape that is readily identified in the field (Ottaway
et al., 1981; Shirvell and Dungey, 1983; Grost et al., 1990).
Displacement of sediment into the water column during
stream bed cutting reduces the sediment fine content in the
tailspill since finer particles are carried further downstream
(Chapman, 1988; Grost et al., 1991; Kondolf and Wolman,
1993; Kondolf et al., 1993; Kondolf, 2000; Schmetterling,
2000; Zimmermann and Lapointe, 2005). The distance and
rate of particle deposition downstream is related to the flow
velocity, channel bed shear, turbulent mixing and particle
fall velocity. Brown trout and rainbow trout tend to spawn
along swifter flowing riffles and runs (Hayes, 1987; Lisle,
1989; Trush, 1989) where the geomorphology and hydraulics
of the river assists in the evacuation and transport of fines.
The excess presence of fines, di< 1mm (where di is the
particle grain diameter of interest) within the tailspill is
commonly considered a principal cause of fry mortality
(Hobbs, 1937; Wickett, 1954; Cooper, 1965; Vaux, 1968;
Phillips et al., 1975; Lotspeich and Everest, 1981). Fines
within the tailspill restrict the interstitial flow of water
(Figure 1b) and hyporheic exchange, reducing dissolved
oxygen and restricting the removal of metabolic wastes
(Stuart, 1953a,b; Wickett, 1954; Cooper, 1965; Vaux, 1968;
Sowden and Power, 1985; Thibodeaux and Boyle, 1987;
Elliott and Brooks, 1997a,b; Packman et al., 1997). Excess
fines can be removed during higher flow events with
increasing discharge and flow depth. Thus Lisle (1989),
Trush (1989), Bunn and Arthington (2002) andWheaton et al.
(2004) surmise that salmonids should, in the long run, benefit
from major storm events since heavy sediment discharge will
provide renewed clean gravel optimal for spawning.
The physical metrics characterizing redd site placement
typically consider local velocity and depth measurements,
sediment analysis and/or the detection of groundwater
seepage. Table I summarizes the ranges in flow depth and
Figure 1. Schematic of a typical trout redd in (a) plan view and (b) longitudinal profile view
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
346 M. A. MARCHILDON ET AL.

Table
I.A
summaryofliterature
reported
ofvelocities
anddepthsrecorded
proxim
alto
browntroutandrainbow
troutredds
Species
Velocity
(cms�
1)
Depth
(cm)Depth
ofpit
(cm)
Tailspill
length
(cm)
NMeasurementlocations
Methodofvelocity
determination
Currentmeter
used
Source
Brown
40(37–43)a
ND
28(27–30)a
ND
90
Deepestpartofpit
0.6
Depth,3cm
abovesurface
Marsh-M
cBirney
model
201
Beard
andCarline
(1991)(1987study)
Brown
33(31–36)a
ND
27(25–30)a
ND
113
Deepestpartofpit
0.6
Depth,3cm
abovesurface
Marsh-M
cBirney
model
201
Beard
andCarline
(1991)(1988study)
Rainbow
20–100
5–50
ND
50–350
61
Threelocationsin
frontofredd
0.6
Depth
Ottcurrentmeter
Crisp
andCarling(1989)
Brown
26.8
(3.4)
30.4
(3.5)
ND
ND
ND
ND
0.6
Depth,3cm
abovesurface
Marsh-M
cBirney
model
201
Essingtonet
al.(1998)b
Brown
34(15)
17(5)
20(5)
147(42)
80
Infrontofredd,in
reddpitandcrestof
tailspill
0.6
Depth
Marsh-M
cBirney
model
201
Grostet
al.(1990)
Brown
27.4
(13.6)
50.0
(15.5)
ND
ND
125
ND
ND
ND
Heggberget
(1991)
Brownand
rainbow
36.4
(15.7)
15.0
(5.0)
ND
ND
25
‘Several
points
around
theredd’
0.6
Depth
Pygmyorprice
currentmeter
Kondolfet
al.(1993)
Brown
37.1
(0.9)
7.2
(0.2)
ND
ND
264
Crest
oftailspill
4.5cm
Abovesurface,
theaverageheightof
amature
trout
Ottcurrentmeter
Ottaw
ayet
al.(1981)c
Brown
30.8
(13.7–45.7)d
16.2
(3–39.6)d
19.5
(7.6–39.6)d
ND
121
Infrontofreddpitand
crestoftailspill
0.6
Depth
Price
currentmeter
ReiserandWesche,
1977
Brown
39.4
(11.2)
31.7
(15.3)
ND
ND
118
Crest
oftailspill
2cm
Above
completedredd
Ottcurrentmeter,
Leopold-Stevens
currentmeter
ShirvellandDungey
(1983)
Brown
44.5
(54.1)
42.6
(54.4)
ND
ND
115
Infrontofredd
12cm
Abovesurface
No.22Gurley
currentmeter
Smith(1973)
Rainbow
69.8
(44.2)
34.2
(42.3)
ND
ND
51
Infrontofredd
12cm
Abovesurface
No.22Gurley
currentmeter
Smith(1973)
Brown
43(16.1)
20(5)
ND
ND
12
Centreoftheredd
5cm
Abovesurface
Marsh-M
cBirney
model
201
Sorensenet
al.(1995)
Brown
46.5
(14.9)
25.5
(8)
ND
ND
110
‘Atoneto
threepositions
aboveredd’
10cm
Abovesurface
Ottcurrentmeter
WitzelandMacCrimmon
(1983a)
Brown
30.3
(9.2)
35.3
(12.9)
37.9
(0.4)
189.2
(6.7)159
‘Velocity
isreferenced
withrespectto
thestream
bed
immediately
upstream
oftheredd’
Manning’s
uniform
flow
equation
None;
seemethodof
velocity
determination
Zim
mer
andPower
(2006)e;M.Power,
pers.comm.
a95%
Confidence
interval
within
parentheses.
bWeightedmean(BevingtonandRobinson,1992)usedto
combinethedataoffourreaches
from
Essingtonet
al.(1998).
cWeightedmean(BevingtonandRobinson,1992)usedto
combinethedataofeightreaches
from
Ottaw
ayet
al.(1981).Depth¼(depth
over
tailspill)þ(4cm
averagetailspillweight).
dValues
inbracketsare90%
confidence
interval
forvelocities
andminim
um–maxim
um
values
fordepths.
eWeightedmeanusedonBrowntroutreddslocatedonriffles
ofboth
channel
types
usingmethodsfrom
BevingtonandRobinson(1992),59of268reddsincluded
from
Zim
mer
andPower
(2005).
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
HIGH-RESOLUTION INVESTIGATION OF REDDS 347

velocities from several studies where the hydraulic
characteristics of brown trout and rainbow trout redds have
been investigated. As evidenced in the table, there is a broad
range in observed velocities and pit depths. Table I also lists
the velocity measurement methods that varied within and
among studies. Most studies determined mean velocities
using the four-tenths/six-tenths method (Henderson, 1966)
such that the velocity observation depth (z – relative to the
stream bed) is at z¼ 0.4h (or 60% below the water surface)
where h is the observed flow depth. Others measured nose-
level velocities at z¼ 10 cm (Smith, 1973; Witzel and
MacCrimmon, 1983a) or at z¼ 5 cm (Ottaway et al., 1981),
which is considered to coincide with the depth at which the
velocity most influences the fish’s ability to hold station
(Ottaway et al., 1981). Zimmer and Power (2006) calculated
the velocity at a series of redds using Manning’s equation
(Henderson, 1966) which is representative of the average
cross-sectional flow velocity. Grost et al. (1990) stated that
no matter what method was used to measure velocities,
spawning salmon and trout consistently avoided low
velocity regions and have a minimum spawning velocity
(Umin) threshold of 20 cm s�1.
METHODOLOGY
A SonTekTM Pulse Coherent Acoustic Doppler Profiler
(PCADP) was employed to measure three-dimensional
velocities on a highly discretized planometric spatial scale.
The PCADP is a downward looking velocity profiler capable
of obtaining three-dimensional velocity measurements at
1.6 cm vertical increments, ranging from 5 cm below the
PCADP to the channel bed. The operation of the PCADP
implicitly assumes flow homogeneity between the three
beams at every depth, where the distance between each beam
is 40% of the depth of measurement. The relative error
associated with the PCADP, therefore, is dependent on
measurement depth, water velocity and temperature, and the
length of the averaging interval. In this study a 30 s
averaging interval was used over a flow depth no greater than
1m (typical maximum wading flow depth) such that 104
beam pairs per sample with a maximum error of
�0.03 cm s�1 were obtained.
Application of the PCADP requires a fixed position to
accurately obtain velocity profiles. The PCADP is com-
monly employed in laboratory flumes or at fixed stations in
larger rivers, estuaries or other bodies of water to sample
velocity changes over extended periods of time. For the
intended study application, two-dimensional spatial data
gathered at short sampling periods were of particular interest
such that discharge could be assumed to be relatively
constant. The PCADP was integrated into a table-like
apparatus (referred to as the cage – Figure 2) to obtain
velocity profiles with a planometric spatial discretization in
the x- and y-plane directions. The discretization interval was
set at Dx¼Dy¼ 20 cm, with individual velocity profiles
acquired over a 30 s sampling period. A 5 s transfer interval
was used to move the PCADP to the next velocity profiling
position. At the maximum density of velocity profiling, 220
profiles were acquired over a 129min sampling duration. If a
constant sampling flow depth of 50 cm is assumed, for
example, a total of 6160 discrete velocity measurements are
obtained. During measurement, stream discharge was
assumed to remain constant.
The PCADP-cage apparatus developed and used (Figure 2)
is similar to that used by Lacey and Roy (2007) where
acoustic Doppler velocity meters were employed. The
configuration employed here allowed a greater density of
velocity measurements to be acquired over a significantly
shorter time span. The cage was designed to allow the
discrete position of each velocity profile to be obtained and
geo-referenced to UTM coordinates. The four corners of the
Figure 2. Cage configuration used for study measurements in (a) plan view and (b) field operation
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
348 M. A. MARCHILDON ET AL.

cage were outfitted with total station prism mounts so that
the corners could be surveyed and referenced to established
geodetic coordinates on land, thus creating a three-
dimensional Cartesian coordinate system of velocity
measurements. Outriggers were integrated into the cage
design to minimize wake turbulence induced by the
upstream legs (Figure 2) and minimize disturbances within
the sampling area. As illustrated in Figure 2b, the user
maintains a constant 2.5m distance from the PCADP at all
times and retreats backward beyond the limits of the cage
and outriggers as measurements progressed across the cage.
A number of metrics were derived from the data. Dimen-
sionless fluid ratios have been used elsewhere to characterize
lotic flow conditions important for micro-habitat selection,
population density and the evolved adaptations of aquatic
organisms (e.g. Leopold and Maddock, 1953; Statzner et al.,
1988; Davis and Barmuta, 1989; Jowett, 1993; Newbury and
Gaboury, 1993; Vogel, 1994; Allan, 1995; Giller and
Malmqvist, 1998; Wadeson and Rowntree, 1998; Rempel
et al., 2000; Lamouroux and Capra, 2002). The Reynolds
number (Re) is the ratio of inertial to viscous forces and is
defined as
Re ¼ ul
v(1a)
where u is the flow velocity at a given location, l is the
characteristic length and n is the kinematic viscosity. For
high width/depth ratio channels, u is often assumed to be the
depth-averaged velocity, U, and l is equal to the flow depth,
h, such that the Reynolds number is expressed as
Re ¼ Uh
v(1b)
for assumed flow under fully turbulent flow conditions (i.e.
Re> 2500, Knighton, 1998).
The Froude number (Fr) defines the ratio of a fluids
inertial forces to gravitational forces and has frequently been
used to describe the tranquility of flow when characterizing
physical habitat (Statzner et al., 1988). The Froude number
is defined as
Fr ¼ Uffiffiffiffiffigh
p (2)
where g is the gravitational constant. Flow is characterized
as sub-critical (or tranquil) when Fr< 1 and super-critical
when Fr> 1. The majority of natural rivers across the world
are typified by Froude numbers <1 at the reach scale. The
most common exceptions are rivers located in steep
mountainous regions, where channel bed slopes typically
exceed 5%. However, at the sub-reach scale, multiple meso-
scale fluid transitions between sub- and super-critical flow
are readily observable (even at low-flow conditions) as water
passes over individual rocks along the stream bed creating a
series of micro-scale hydraulic jumps (Newbury and
Gaboury, 1993).
Methods used to define physical habitat are mostly two-
dimensional (planform) metrics that implicitly assume
depth-wise homogeneity. To illustrate depth-wise trends, a
modification of turbulent kinetic energy per unit mass
equation was developed as an alternative to the Reynolds
number to describe the turbulence state of the observed flow
conditions. The mean turbulent kinetic energy per unit mass
(e) was used to assess the consistency of time-averaged
velocity measurements (Moody and Smith, 2004; Stone
et al., 2006; Smith and Brannon, 2007) and was defined as
e ¼ 1
2ðs2
x þ s2y þ s2
z Þ (3a)
where s2x ; s
2y ; s
2z are the velocity variances in the x-, y-, z-
directions, respectively. In place of a point velocity
observation as described by Equation (3a), a series of
vertical velocities were obtained in this study at a given
planometric location to estimate a vertical velocity profile
that is a depth integrated measure of turbulent kinetic energy
per unit area (ea). Summing the principal velocity
components of each vertical velocity profiles yields
ea¼ rh
2ðn� 1ÞXn
i¼0
½ðux;i�UxÞ2 þ ðuy;i � UyÞ2 þ ðuz;i � UzÞ2�
(3b)
for a discrete velocity profile where r is the density of water
and n is the number of velocity observations in each vertical
profile. Low values of ea ! 0:0 depict parallel flow lines
where flow tends towards unidirectional behaviour in the
downstream direction. As values of ea increase, flow
becomes more turbulent.
The existence of interstitial flow can be estimated by the
difference in average velocities between the pit and tailspill
region and the resulting kinetic energy heads (u2/2g). The
vertical transition from the pit to the top of the tailspill can be
simplified as a small bed rise (DZ) observed in a channel whichcan be evaluated using the Bernoulli equation such that
h1 þ u212g
¼ h2 þ u222g
þ DZ (4)
where 1 represents the hydraulic metrics of the pit and 2
represents the hydraulic metrics above the tailspill.
STUDY SITE
Whiteman’s Creek, located near Paris, Ontario, Canada
(Figure 3a) was selected to investigate the hydrodynamic
characteristics of sites selected by spawning rainbow and
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
HIGH-RESOLUTION INVESTIGATION OF REDDS 349

brown trout. The study reach is characterized as a low bed
slope (Figure 3b), flood plain dominated, gravel-bed river.
Summary of reach characteristics (Table II) are from Hartley
(1999).
Research by Hartley (1999) suggested that the selected
reach supported diverse aquatic niches. Thus both spring-
spawning rainbow trout and fall-spawning brown trout use
the reach for spawning. Redd identification was undertaken
within a week of spawning, when strong visual contrasts
could be identified in bed material prior to periphytic
regrowth (Witzel and MacCrimmon, 1983a,b; Grost et al.,
1990; Beard and Carline, 1991; Schmetterling, 2000). A
subset of riffles identified with redds were selected for
detailed hydrodynamic analysis. Rainbow trout redds were
identified in only two riffles (R1 and R3) within the study
reach (Figure 3). A similar survey of the study reach in the
autumn noted the presence of brown trout redds in five
separate riffles. Only two of the five riffles, R1 and R2, were
selected for use in the study (Figure 3). Choice of study
riffles was random and within the constraint of equipment
accessibility. All selected riffles demonstrated high spawn-
ing activity over the course of the respective spring and
autumn spawning periods based on the presence of multiple
redds and evidence of redd superimposition and/or
abandoned/incomplete redd construction.
Habitat variables for both brown trout and rainbow trout
study redds are presented in Table III. For brown trout,
velocity profile suites were measured between 6th and 15th
November 2006 in a window between two larger flow events
that exceeded wadable conditions. During the study period,
a relatively constant discharge of 5.5m3 s�1 was measured at
the gauge station at the upper limit of the study reach
(Figure 3).
For rainbow trout redds in riffles R1 and R3, velocity
profiles were obtained between 9th and 11th May 2007, when
a relatively constant discharge of 2.33m3 s�1 was observed.
RESULTS
A total 2683 velocity profiles were acquired from the brown
trout and rainbow trout study redds. Data are compared to
previous literature studies in Figure 4. Mean velocities
Figure 3. Whiteman’s Creek study reach (a) site location map and (b) longitudinal profile of riffle study locations (R1, R2 and R3)
Table II. A summary of study reach parameters from Hartley(1999)
Parameter Metric
Bed slope (S0) 0.27%Sinuosity (V) 1.6Effective catchment area (Ad) 383 km2
Bankfull discharge (Qbf) 42m3 s�1
Average bankfull width (Wbf) 23.2mAverage bankfull depth (hbf) 1.1mParticle size of riffles d16¼ 11.8mm, d50¼ 42.8mm,
d84¼ 90.9mmParticle size of reach d16¼ 8.1mm, d50¼ 33.4mm,
d84¼ 72.8mm
Table III. Summary of habitat variables measured at brown trout and rainbow trout study redds
Variables Brown trout redds Rainbow trout redds
Data collection period 6–15 November 2006 9–11 May 2007Study discharge (Q) 5.5m3 s�1 2.33m3 s�1
Temperature (oC) 68C� T� 98C 158C� T� 198CBed slope (S0) BR1¼ 0.0061mm�1 R1¼ 0.0061mm�1
BR2¼ 0.0007mm�1 R3¼ 0.0075mm�1
Bed friction slope (Sf) BR1¼ 0.0028mm�1 R1¼ 0.0028mm�1
BR2¼ 0.0027mm�1 R3¼ 0.0004mm�1
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
350 M. A. MARCHILDON ET AL.
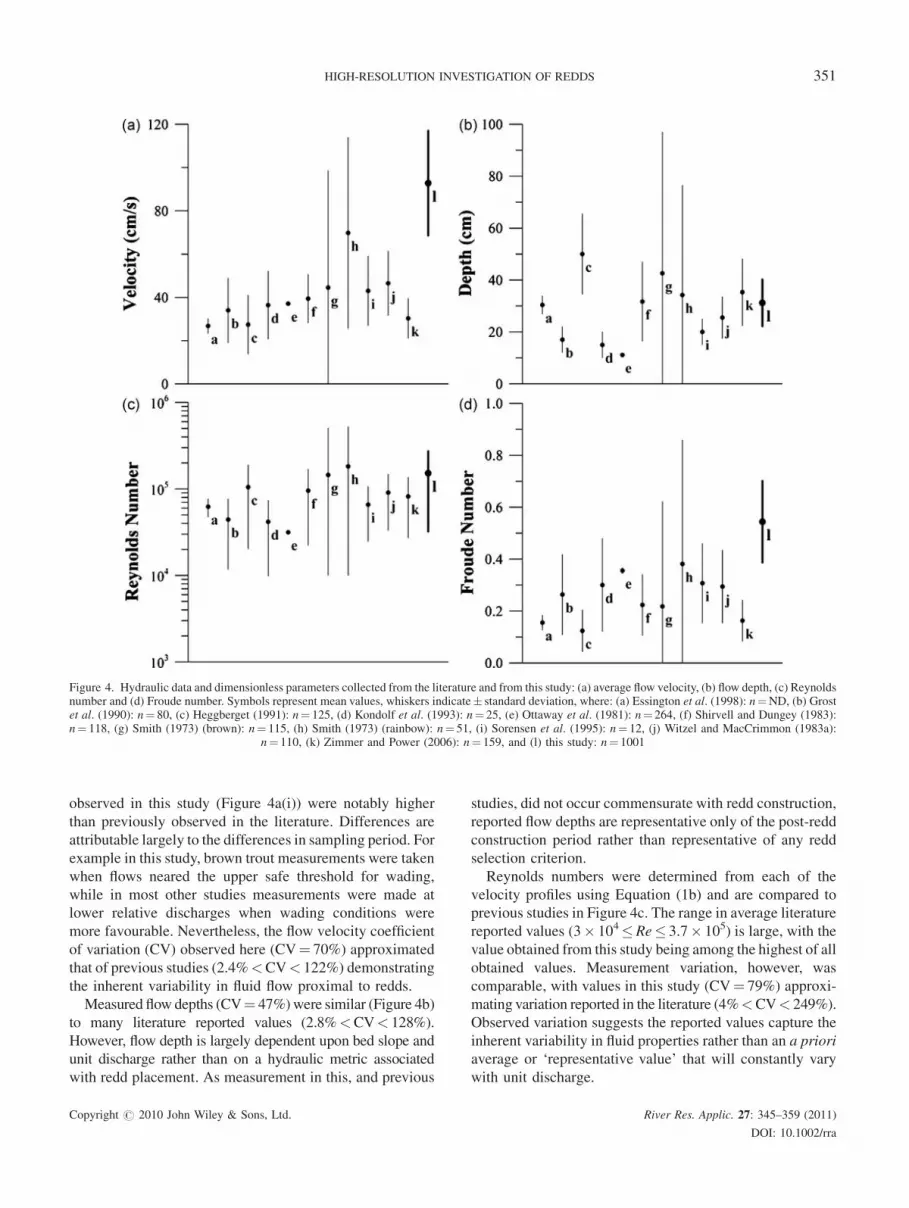
observed in this study (Figure 4a(i)) were notably higher
than previously observed in the literature. Differences are
attributable largely to the differences in sampling period. For
example in this study, brown trout measurements were taken
when flows neared the upper safe threshold for wading,
while in most other studies measurements were made at
lower relative discharges when wading conditions were
more favourable. Nevertheless, the flow velocity coefficient
of variation (CV) observed here (CV¼ 70%) approximated
that of previous studies (2.4%<CV< 122%) demonstrating
the inherent variability in fluid flow proximal to redds.
Measured flow depths (CV¼ 47%)were similar (Figure 4b)
to many literature reported values (2.8%<CV< 128%).
However, flow depth is largely dependent upon bed slope and
unit discharge rather than on a hydraulic metric associated
with redd placement. As measurement in this, and previous
studies, did not occur commensurate with redd construction,
reported flow depths are representative only of the post-redd
construction period rather than representative of any redd
selection criterion.
Reynolds numbers were determined from each of the
velocity profiles using Equation (1b) and are compared to
previous studies in Figure 4c. The range in average literature
reported values (3� 104�Re� 3.7� 105) is large, with the
value obtained from this study being among the highest of all
obtained values. Measurement variation, however, was
comparable, with values in this study (CV¼ 79%) approxi-
mating variation reported in the literature (4%<CV< 249%).
Observed variation suggests the reported values capture the
inherent variability in fluid properties rather than an a priori
average or ‘representative value’ that will constantly vary
with unit discharge.
Figure 4. Hydraulic data and dimensionless parameters collected from the literature and from this study: (a) average flow velocity, (b) flow depth, (c) Reynoldsnumber and (d) Froude number. Symbols represent mean values, whiskers indicate� standard deviation, where: (a) Essington et al. (1998): n¼ND, (b) Grostet al. (1990): n¼ 80, (c) Heggberget (1991): n¼ 125, (d) Kondolf et al. (1993): n¼ 25, (e) Ottaway et al. (1981): n¼ 264, (f) Shirvell and Dungey (1983):n¼ 118, (g) Smith (1973) (brown): n¼ 115, (h) Smith (1973) (rainbow): n¼ 51, (i) Sorensen et al. (1995): n¼ 12, (j) Witzel and MacCrimmon (1983a):
n¼ 110, (k) Zimmer and Power (2006): n¼ 159, and (l) this study: n¼ 1001
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
HIGH-RESOLUTION INVESTIGATION OF REDDS 351
Froude numbers calculated using Equation (2) were also
compared to previous studies (Figure 4d) and were, on
average, higher than in previous literature studies. Measure-
ment variation (CV¼ 29%), however, compared favourably
to variation reported in the literature (3%<CV< 185%).
The Froude number is linearly proportional to the velocity
(Figure 4a) and as a result of the higher velocities measured,
particularly in brown trout redds, higher Froude numbers
were obtained.
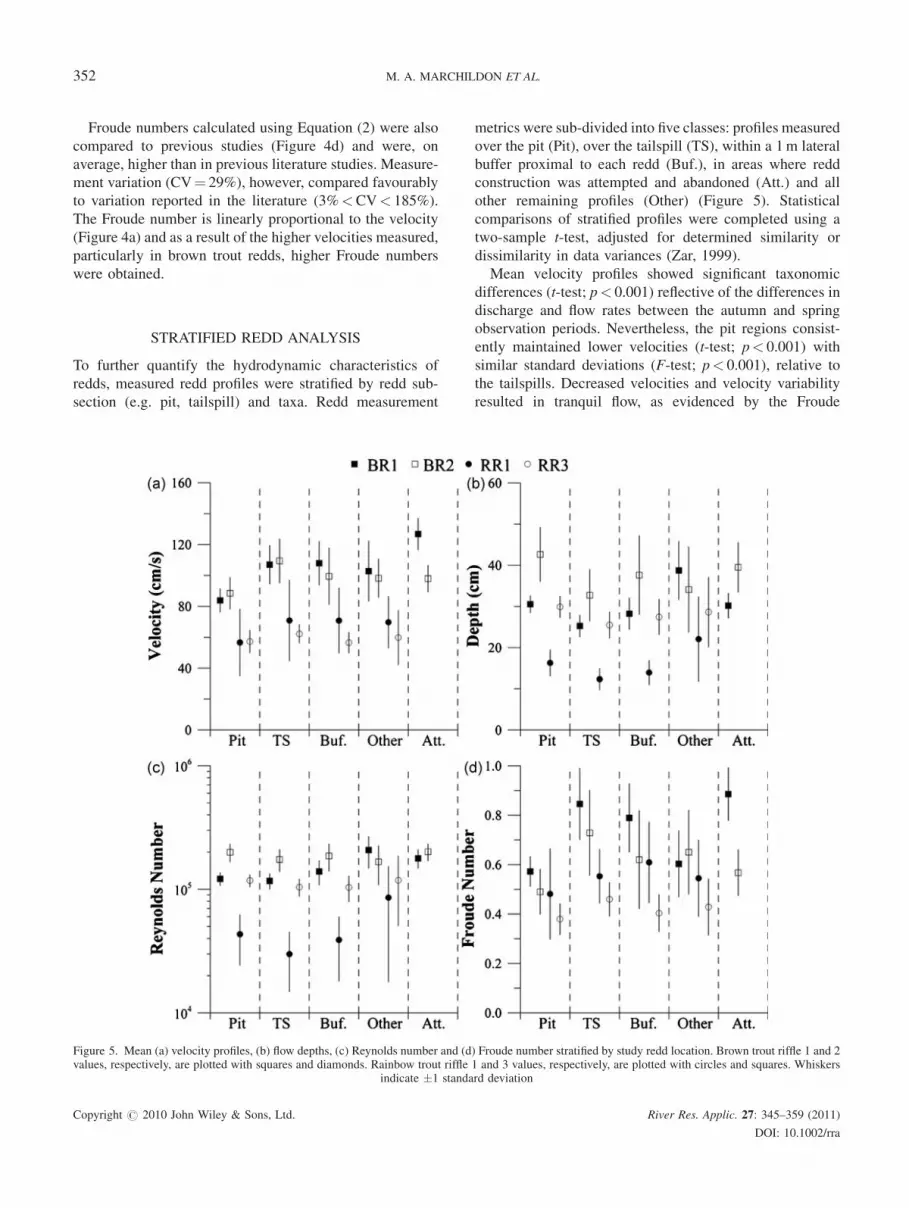
STRATIFIED REDD ANALYSIS
To further quantify the hydrodynamic characteristics of
redds, measured redd profiles were stratified by redd sub-
section (e.g. pit, tailspill) and taxa. Redd measurement
metrics were sub-divided into five classes: profiles measured
over the pit (Pit), over the tailspill (TS), within a 1m lateral
buffer proximal to each redd (Buf.), in areas where redd
construction was attempted and abandoned (Att.) and all
other remaining profiles (Other) (Figure 5). Statistical
comparisons of stratified profiles were completed using a
two-sample t-test, adjusted for determined similarity or
dissimilarity in data variances (Zar, 1999).
Mean velocity profiles showed significant taxonomic
differences (t-test; p< 0.001) reflective of the differences in
discharge and flow rates between the autumn and spring
observation periods. Nevertheless, the pit regions consist-
ently maintained lower velocities (t-test; p< 0.001) with
similar standard deviations (F-test; p< 0.001), relative to
the tailspills. Decreased velocities and velocity variability
resulted in tranquil flow, as evidenced by the Froude
Figure 5. Mean (a) velocity profiles, (b) flow depths, (c) Reynolds number and (d) Froude number stratified by study redd location. Brown trout riffle 1 and 2values, respectively, are plotted with squares and diamonds. Rainbow trout riffle 1 and 3 values, respectively, are plotted with circles and squares. Whiskers
indicate �1 standard deviation
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
352 M. A. MARCHILDON ET AL.

numbers in Figure 5d, describe a controlled region in the pit
for ova placement and fertilization experiencing minimal
disturbance relative to the remainder of the redd and
surrounding redd region.
The Reynolds numbers when stratified by redd location
and taxa indicated similar means (all pairwise t-test;
p> 0.05) and all pairwise variances (F-test; p> 0.05) with
the exception of the rainbow trout redd on riffle 1 (R1).
Lower Reynolds numbers were related to the lower
velocities (Figure 5a) and flow depths (Figure 5b) at the
time of measurement. These results continue to demonstrate
the inherent variability of flow over the redd that can occur
over a relatively short period of sampling. Given the high
resolution of measurement, the large range in observed
values effectively demonstrates temporal variability in
hydraulic metrics and challenges the appropriateness of
using a single or representative value to represent redd
hydraulic or habitat condition.
In addition to the higher Froude number in the tailspill
regions relative to other areas of the redd (t-test; p for all red
class comparisons <0.01, Figure 5d), it is important to
consider occurrences of flow conditions nearing super-
critical flow conditions (Fr! 1.0) over the tailspill and
some of the surrounding buffer regions of the studied
rainbow trout redd from riffle 1. Close inspection of the data
identifies a number of outlier locations where super-critical
flow occurred. These are micro-scale hydraulic jumps
created by the combined hydraulic conditions of shallow
flow depths (Figure 5b) and higher velocities lf flow lines
around individual. The hydraulic jumps occur over
individual clasts, transitioning from sub-critical over the
rock and returning to sub-critical flow through a hydraulic
jump on the micro-hydraulic scale. Micro-scale hydraulic
jumps entrain dissolved oxygen in the water column, which
may enter the tailspill substrate and provide additional
benefits to the ova during low-flow conditions.
Figure 6 presents the plots of turbulent kinetic energy per
unit area by stratifying the data into redd sub-regions
(similar to Figure 5). For both brown and rainbow
trout, there was a considerable decrease in secondary flow
above and surrounding the tailspill (TS) and buffer (Buf.)
regions (all comparisons p< 0.01). These are the regions
where the flow is mostly parallel and unidirectional in the
downstream direction. Flow principally focussed in one
direction over the tailspills may benefit the region by
maintaining smaller secondary flow patterns that enhance
the deposition of fine-grained material to the detriment of the
developing ova. Furthermore, the high variation of the
profiles categorized as ‘other’ in Figure 6 illustrates two
important characteristics of the measured flow regimes.
First, it shows that assuming depth-wise homogeneity when
employing the use of two-dimensional planform hydraulic
models is a major simplification. Second, there is a high
degree of spatial depth-wise heterogeneity in the planform
perspective.
Early workers noted fertilized eggs in tailspill interstitial
spaces (Hobbs, 1937; Burner, 1951). Dye experiments
further noted that flow entered the redd at the upstream end
of the tailspill, exiting downstream of the redd’s crest
(Figure 1b), and affected regions occupied by fertilized eggs.
Wu (2000) noted that bedforns reportedly experiencing
substantial interstitial flow had a form analogous to salmonid
redds, as illustrated in Figure 1b. Interstitial flow within the
bedform is attributed to flux variations at the sediment–water
interface, consistent with Bernoulli’s principle.
Although dye-tracing experiments were not conducted
through the tailspills in this study, the hydraulic data
collected provides a means of interpreting the existence of
interstitial flow. Velocity measurements were extracted from
the profiles at 1.6 cm above the stream bed that represent the
velocities closest to the sediment–water interface. The
velocities were stratified by redd sub-regions, as previously
described, and plotted in Figure 7a. The results show a
considerable increase in flow velocities immediately over
the tailspill regions relative to the redd pit regions (t-test;
p< 0.01). The difference in average velocities between the
pit and tailspill regions, and resulting kinetic energy heads
(u2/2g), corroborates the existence of interstitial flow. Using
the maximum depth of each pit, the associated shallowest
depth over each tailspill and the velocities observed at a
Figure 6. Mean turbulent kinetic energy per unit area of stream bedstratified by riffle location. Brown trout riffle 1 and 2 values, respectively,are plotted with squares and diamonds. Rainbow trout riffle 1 and 3 values,respectively, are plotted with circles and squares. Whiskers indicate �1
standard deviation
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
HIGH-RESOLUTION INVESTIGATION OF REDDS 353

depth of 1.6 cm above the stream bed in each pit and tailspill,
an energy balance between the pits and tailspills can be
calculated using Equation (4). The analysis results in an
average difference in total energy between the pits and
tailspills of 6.8% and an entire range between 1.75 and
11.2%. Results are well within the error measurement range
of the calculation (given the scale and relative roughness of
the substrate) and, therefore, conservation of energy can be
assumed. Given that the bed rise (DZ) in all cases is
permeable, interstitial flow must be occurring within the
tailspill regions.
Although internal circular flow within redd pits have been
suggested in the literature (e.g. Hobbs, 1937; Burner, 1951 –
Figure 1b) data collected here do not substantiate such
conclusions. For flow profiles collected in this study the
average of the collected z-direction velocity standard
deviations (sz) is presented in Figure 7b. If a circular flow
pattern were present, a notable increase in sz for profiles
measured in the pit regions relative to the surrounding
regions should be observed. Results of the analysis here
show no distinct difference in the standard deviations of the
flow directions in the vertical (z) direction (F-test; p> 0.05),
thereby demonstrating that dominant circulation patterns in
the pit region do not exist. Although the analysis does not
exclude the possibility of weak circulation patterns
occurring within the pit, associated flows would be below
the detectable limits of the PCADP. Other circulation
patterns may also arise in the pit regions that cannot be
identified by characterizing the standard deviations of the
vertical velocity component. Mean turbulent kinetic energy
per unit area (Figure 6) in the pit regions of the redd show
higher values than the tail regions. These more complex
patterns may represent planometric circulation patterns
within the pit regions.
PLANOMETRIC DISTRIBUTION OF HYDRAULIC
PARAMETERS
The hydraulic metrics used in the analysis above were also
used to plot depth-average velocity profiles, with the
exception of the velocity observed 10 cm above the stream
bed (U10). Figures 8 and 9 illustrate the two-dimensional
plan form spatial distribution patterns of hydraulic metrics
studied for a sub-set of measured redds. The hydraulic
metrics shown are of two brown trout redds (Figure 8)
located on riffle 2 (BR2-1 and BR2-2) and two rainbow trout
redds (Figure 9) located on riffle 1 (RR1) and riffle 3 (RR2).
Examination of the metric spatial trends for redds and/or
taxa showed no clear trends in any of the metrics, with the
exception of the mean turbulent kinetic energy per unit area
which consistently identified the redd tailspills with very
unidirectional parallel flow conditions (p< 0.001). Metrics
were consistently higher for the brown trout redds illustrated
relative to rainbow trout redds (t-test; p for taxonomic
comparisons <0.05). Similar results were obtained with the
analysis of the other brown and rainbow trout redds (t-test; p
for taxonomic comparisons <0.01). The differences in
metric magnitude (Figures 8 and 9) is largely attributable to
the differences in flow rates between the two study periods,
with flows of 5.5 and 2.33m3 s�1, respectively, prevailing
during the completion of the brown trout and rainbow trout
redd studies. The temporal pattern of study results further
reinforces the importance of identifying a range in metrics,
Figure 7. (a) Average velocity at 1.6 cm above stream bed (u1.6) and (b) averages of profile vertical velocity standard deviation (sz) stratified by redd location.Brown trout riffle 1 and 2 values, respectively, are plotted with squares and diamonds. Rainbow trout riffle 1 and 3 values, respectively, are plotted with circles
and squares. Whiskers indicate �1 standard deviation
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
354 M. A. MARCHILDON ET AL.

Figure 8. Two-dimensional hydraulic parameter distributions of two brown trout redds on riffle 2 (BR2-1) and (BR2-2) of Reynolds number (Re), Froudenumber (Fr), average flow velocity (U, flow velocity 10 cm above stream bed (U10), and mean turbulent kinetic energy per unit area (ea)). Mean flow direction is
indicated by arrows and the ellipses indicates the extent of the redd. This figure is available in colour online at wileyonlinelibrary.com/journal/rra
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
HIGH-RESOLUTION INVESTIGATION OF REDDS 355
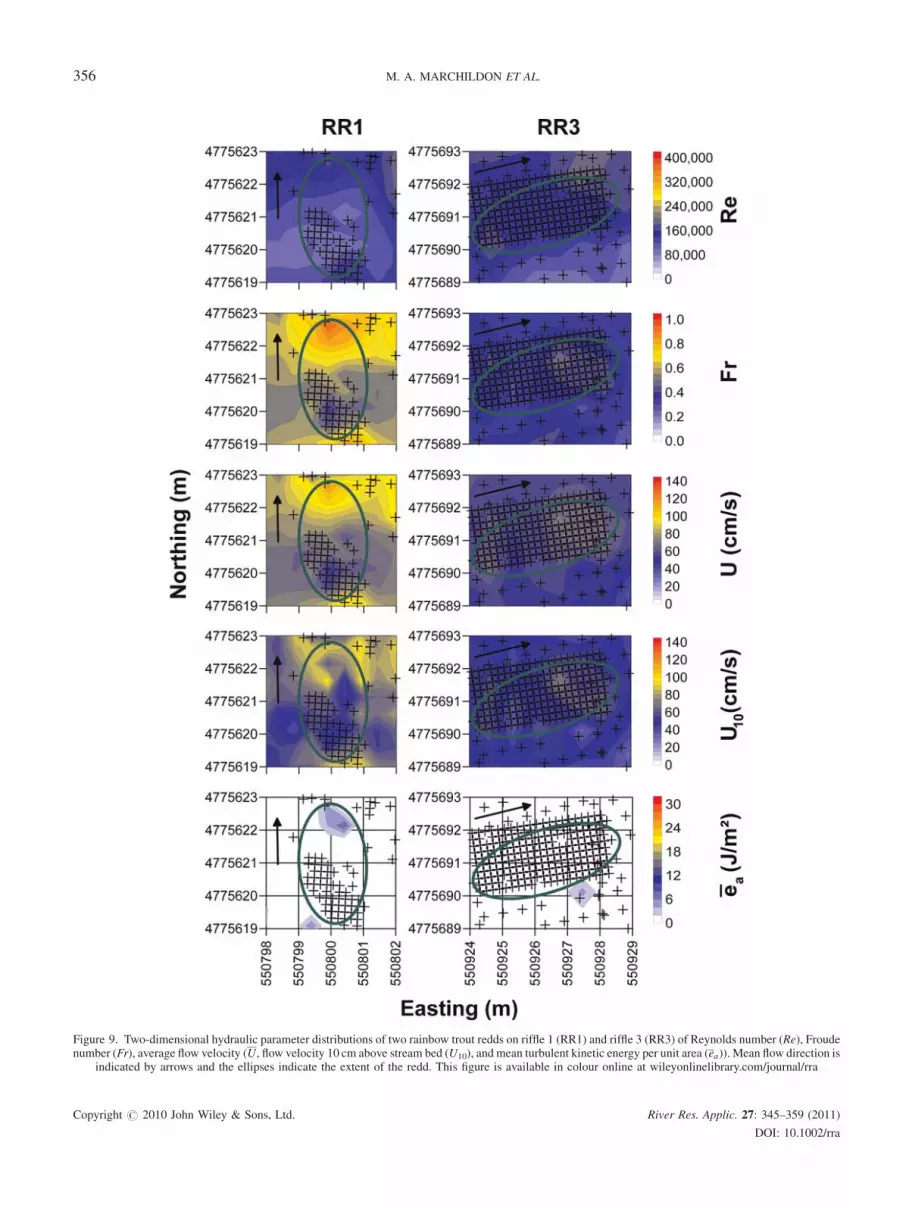
Figure 9. Two-dimensional hydraulic parameter distributions of two rainbow trout redds on riffle 1 (RR1) and riffle 3 (RR3) of Reynolds number (Re), Froudenumber (Fr), average flow velocity (U, flow velocity 10 cm above stream bed (U10), and mean turbulent kinetic energy per unit area (ea)). Mean flow direction is
indicated by arrows and the ellipses indicate the extent of the redd. This figure is available in colour online at wileyonlinelibrary.com/journal/rra
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
356 M. A. MARCHILDON ET AL.

rather than specific representative values to characterize the
redd suitability characteristics of a specific river reach.
Similar to the previous discussions, Froude numbers were
consistently observed to increase over the tailspills relative
to other parts of the redd (t-test; p for all red class
comparisons<0.01), a fact attributable to the shallower flow
depths and higher velocities as expressed in Equation (2).
Lower velocities were observed at 10 cm above the stream
bed (U10) in most cases relative to the depth-average
velocities (U) which will further trend towards 0 cm s�1 at
the sediment–water interface. The velocities 10 cm above
the stream bed also spatially show a lower velocity in the pit
region relative to the tailspill region (t-tets; p< 0.01). No
clear discernable trends were observed in using Reynolds
number to characterize the redds and surrounding regions
other than to note that the spatial range in values was large
(3� 104�Re� 3.7� 105).
CONCLUSIONS
Results of this study have demonstrated high spatial varia-
bility in the physical metrics typically used to characterize
lotic flow regimes. The only parameter that consistently
spatially correlated with redd location was turbulent kinetic
energy per unit area of stream bed. Low turbulent kinetic
energy per unit area is related to flow regimes with little
turbulence where flow is essentially unidirectional.
At the redd scale, the lowest turbulent kinetic energy per
unit area existed atop the tailspills, where the flow passing
over the elongated teardrop form showed an increase in the
linearity of flow. From a fluid dynamics perspective, the
teardrop form is known to produce the lowest amount of drag
per unit volume, as long as the fluid flow is moving along the
teardrop’s principal axis (Vogel, 1994). This would be
complementary to the development of interstitial flow
through the tailspill in order to aerate the ova.
Results of Froude number analysis demonstrated high
spatial variability and no particular spatial consistency with
redd site location. Froude numbers were the highest over the
tailspill areas where higher velocities and shallower flow
depths consistently exist. However, beyond the limits of the
pit and tailspill, no spatial trends in the measured hydraulic
parameters were identified relating to redd location
placement. Of particular note was the fact there were no
spatial correlations were identified between red locations
and the Reynolds number.
The spatial variability observed in the fluid properties of
flow velocity and flow depth and the resulting metrics of the
Reynolds number, the Froude number and turbulent kinetic
energy per unit area were not surprising considering the high
resolution of the measurements and the decrease in
measurement scale towards the micro-scale relative to
previous studies. In particular, the differences in average
reported values and the ranges of variability in the metrics
measured and calculated between the study periods for the
spring and fall redd surveys demonstrates the inherent
natural variability of aquatic systems that strongly suggests
they cannot be accurately described by single ‘representa-
tive’ hydraulic metric values.
NOMENCLATURE
Ad effective catchment area (L2)
di grain size diameter (where i¼ 5,16, 25, . . . 95thpercentile) (L)
e mean turbulent kinetic energy per unit mass (L2T �2)
ea mean turbulent kinetic energy per unit area (MT �2)
Fr Froude number
g gravitational acceleration (LT �2)
h flow depth (L)
hbf bankfull depth (L)
l characteristic length (L)
n number of observations in vertical velocity profile
Q bankfull discharge (L3T �1)
Qbf bankfull discharge (L3T �1)
Re Reynolds number
Sf friction slope
S0 bed slope
T temperature (8C)u velocity (LT�1)
u1.6 skin velocity 1.6 cm above stream bed (LT�1)
u10 flow velocity 10 cm above stream bed (LT�1)
U depth-averaged velocity (LT �1)
Umin average minimum spawning velocity threshold
(LT�1)
Wbf bankfull width (L)
z height above stream bed (L)
Dx spatial discretization of PCADP in the x-direction
(L)
Dy spatial discretization of PCADP in the x-direction
(L)
DZ bed rise (L)
n kinematic viscosity of water, where n¼m/r (L2T �1)
r mass density of water �1000 kgm�3 (ML�3)
sx,y,z standard deviation of velocity in the x-, y-, z-
directions (LT �1)
V sinuosity
ACKNOWLEDGEMENTS
Funding for this project was provided by the National
Science and Engineering Research Council of Canada
(NSERC) Discovery Grant programme, Trout Unlimited
Canada – Cold Water Conservation Fund, Canada Founda-
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
HIGH-RESOLUTION INVESTIGATION OF REDDS 357

tion for Innovation – Leaders Opportunity Fund and the
Grand River Conservation Authority – Apps Mill Nature
Centre.
REFERENCES
Allan JD. 1995. Stream Ecology: Structure and Function of Running
Waters. Chapman & Hall: London.
Beard TD Jr, Carline RF. 1991. Influence of spawning and other stream
habitat features on spatial variability of wild brown trout. Transactions of
the American Fisheries Society 120: 711–722.
Bevington PR, Robinson DK. 1992. Data Reduction and Error Analysis for
the Physical Sciences, 2nd edn. McGraw-Hill: Boston, New York.
Buchanan TJ, Somers WP. 1969. Discharge measurements at gaging
stations, US Geological Survey Techniques of Water-Resources Inves-
tigation, Book 3, Chapter A8, 73.
Bunn SE, Arthington AH. 2002. Basic principles and ecological con-
sequences of altered flow regimes for aquatic biodiversity. Environmental
Management 30(4): 492–507.
Burner CJ. 1951. Characteristics of spawning nests of Columbia River
salmon. U.S. Fish and Wildlife Service: Fishery Bulletin 61: 97–110.
Champoux O, Biron PM, Roy AG. 2003. The long-term effectiveness of fish
habitat restoration practices: Lawrence Creek, Wisconsin. Annals of the
Association of American Geographers 93(1): 42–54.
Chapman DW. 1988. Critical review of variables used to define effects of
fines in redds of large salmonids. Transactions of the American Fisheries
Society 117(1): 1–21.
Cooper AC. 1965. The Effect of Transported Stream Sediments on the
Survival of Sockeye and Pink Salmon Eggs and Alevin. International
Pacific Salmon Committee Bulletin 1: New Westminster, B.C.
Crisp DT, Carling PA. 1989. Observation on siting, dimensions and
structure of salmonid redds. Journal of Fish Biology 34: 119–134.
Davis JA, Barmuta LA. 1989. An ecologically useful classification of mean
and near-bed flows in streams and rivers. Freshwater Biology 21: 271–
282.
Elliott AH, Brooks NH. 1997a. Transfer of nonsorbing solutes to a
streambed with bed forms: theory. Water Resources Research 33:
123–136.
Elliott AH, Brooks NH. 1997b. Transfer of nonsorbing solutes to a
streambed with bed forms: laboratory experiments. Water Resources
Research 33: 137–151.
Essington TE, Sorensen PW, Paron DG. 1998. High rate of redd super-
imposition by brook trout (Salvelinus fontinalis) and brown trout (Salmo
trutta) in a Minnesota stream cannot be explained by habitat availability
alone. Canadian Journal of Fisheries and Aquatic Sciences 55: 2310–
2316.
Giller PS, Malmqvist B. 1998. The Biology of Streams and Rivers, 3rd edn.
Oxford University Press: Oxford.
Grost RT, Hubert WA, Wesche TA. 1990. Redd site selection by brown trout
in Douglas creek, Wyoming. Journal of Freshwater Ecology 5(3): 365–
371.
Grost RT, Hubert WA, Wesche TA. 1991. Description of brown trout in a
mountain stream. Transactions of the American Fisheries Society 120:
582–588.
Hartley AM. 1999. The pool-riffle sequence as the principal design com-
ponent of low-gradient, meandering, gravel-bed channels, M.Sc. Thesis,
School of Engineering, University of Guelph, Guelph, Ontario, Canada,
158.
Hayes JW. 1987. Competition for spawning space between brown (Salmo
trutta) and rainbow trout (S. gairdneri) in a lake inlet tributary, New
Zealand. Canadian Journal of Fisheries and Aquatic Sciences 44: 40–47.
Heggberget TG. 1991. Some environmental requirements of Atlantic
salmon. American Fisheries Society Symposium 10: 132–135.
Henderson FM. 1966. Open Channel Flow. MacMillan Publishing Com-
pany: New York; 522.
Hobbs DF. 1937. Natural reproduction of quinnat salmon, brown and
rainbow trout in certain New Zealand waters. New Zealand Marine
Department Fisheries Bulletin 6: 104.
Imhof JG, Fitzgibbon J, Annable WK. 1996. A hierarchical evaluation
system for characterizing watershed ecosystems for fish habitat. Cana-
dian Journal of Fisheries and Aquatic Sciences 53: 312–326.
Jones JW, Ball JN. 1954. The spawning behaviour of brown trout and
salmon. British Journal of Animal Behaviour 2: 103–114.
Jowett IG. 1993. A method for objectively identifying pool, run, and riffle
habitats from physical measures. New Zealand Journal of Marine and
Freshwater Research 27: 241–248.
Knighton D. 1998. Fluvial Forms and Processes: A New Perspective.
Oxford University Press, Inc.: New York.
Kondolf GM. 2000. Assessing salmonid spawning gravel quality. Trans-
actions of the American Fisheries Society 129: 262–281.
Kondolf GM, Wolman MG. 1993. The size of salmonid spawning gravels.
Water Resources Research 29(7): 2275–2285.
Kondolf GM, Sale MJ, Wolman MG. 1993. Modification of fluvial gravel
size by spawning salmonids. Water Resources Research 29(7): 2265–
2274.
Lacey RW, Roy AG. 2007. A comparative study of the turbulent flow field
with and without a pebble cluster in a gravel bed river. Water Resources
Research 43(W05502): 1–7.
Lamouroux N, Capra H. 2002. Simple predictions of instream habitat model
outputs for target fish populations. Freshwater biology 47(8): 1543–1556.
Leopold LB, Maddock T Jr. 1953. The hydraulic geometry of stream
channels and some physiographic implications. U.S.G.S Professional
Paper 252, 57.
Leopold LB, Wolman MG, Miller JP. 1964. Fluvial Processes in Geo-
morphology. W. H. Freeman: San Francisco; 522.
Lisle TE. 1989. Sediment transport and resulting deposition in spawning
gravels, north Coastal California. Water Resources Research 25(6):
1303–1319.
Lotspeich FB, Everest FH. 1981. A new approach for reporting and
interpreting textural composition of spawning gravel. U.S.D.A. Forestry
Service Research note PNW-369, 11.
Maddock I. 1999. The importance of physical habitat assessment for
evaluating river health. Freshwater biology 41: 373–391.
Moody JA, Smith JD. 2004. Field measurements of Reynolds stress near a
riverbank. Hydraulic Measurements and Experimental Methods Con-
ference 2002, 1–12.
Newbury RW, Gaboury M. 1993. Stream Analysis and Fish Habitat Design:
A Field Manual. Newbury Hydraulics: Gibsons.
Ottaway EM, Carling PA, Clark A, Reader NA. 1981. Observations on the
structure of brown trout, Salmo trutta Linnaeus, redds. Journal of Fish
Biology 19: 593–607.
Packman AI, Brooks NH, Morgan JJ. 1997. Experimental techniques for
laboratory investigation of clay colloid transport and filtration in a stream
with a sand seal. Water, Air and Soil Pollution 99: 113–122.
Phillips RW, Lantz RL, Claire EW,Moring JR. 1975. Some effects of gravel
mixtures on emergence of coho salmon and steelhead trout fry. Trans-
actions of the American Fisheries Society 3: 461–466.
Raleigh RF, Hickman T, Solomon RC. 1984. Habitat suitability infor-
mation: rainbow trout. U.S. Department of the Interior. Fish and Wildlife
Service FWS/OBS-82/10.60, 74.
Raleigh RF, Zuckerman LD, Nelson PC. 1986. Habitat suitability index
models and instream flow suitability curves: brown trout. U.S. Depart-
ment of the Interior. Fish and Wildlife Service: Biological report
82(10.24).
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
358 M. A. MARCHILDON ET AL.

Reiser DW, Wesche TA. 1977. Determination of physical and hydraulic
preferences of brown and brook trout in the selection of spawning
locations. Water Resources Series No. 64. Water Resources Research
Institute, University of Wyoming, Laramie, Wyoming, 100.
Rempel LL, Richardson JS, Healey MC. 2000. Macroinvertebrate com-
munity structure along gradients of hydraulic and sedimentary conditions
in a gravel-bed river. Freshwater biology 45: 57–73.
Schmetterling DA. 2000. Redd characteristics of fluvial westslope cutthroat
trout in four tributaries to the Black river, Montana. North American
Journal of Fisheries Management 20: 776–783.
Schwartz JS, Herricks EE. 2007. Evaluation of pool-riffle naturalization
structures on habitat complexity and the fish community in an urban
Illinois stream. River Research and Applications 23: 451–466.
Shirvell CS, Dungey RG. 1983. Microhabitats chosen by brown trout for
feeding and spawning in rivers. Transactions of the American Fisheries
Society 112: 355–367.
Smith AK. 1973. Development and application of spawning velocity and
depth criteria for Oregon salmonids. Transactions of the American
Fisheries Society 2: 312–316.
Smith DL, Brannon EL. 2007. Influence of cover on mean column hydraulic
characteristics in small pool riffle morphology streams. River Research
and Applications 23: 125–139.
Sorensen PW, Cardwell JR, Essington T, Weigel DE. 1995. Reproductive
interactions between sympatric brook and brown trout in a small Min-
nesota stream. Canadian Journal of Fisheries and Aquatic Sciences 52:
1958–1965.
Sowden TK, Power G. 1985. Prediction of rainbow trout embryo survival in
relation to groundwater seepage and particle size of spawning substrates.
Transactions of the American Fisheries Society 114: 804–812.
Statzner B, Gore JA, Resh VH. 1988. Hydraulic stream ecology: observed
patterns and potential applications. Journal of the North American
Benthological Society 7(4): 307–360.
Stone MC, Hotchkiss RH, Morrison RR. 2006. Turbulence observations in
cobble-bed rivers. World Environmental and Water Resource Congress
2006 Examining the Confluence of Environmental and Water Concerns.
Stuart TA. 1953a. Spawning migration, reproduction, and young stages of
loch trout (Salmo trutta L.). Scottish Home Department, Freshwater and
Salmon Fisheries Research 5, Edinburgh, 39.
Stuart TA. 1953b. Water currents through permeable gravels and their
significance to spawning salmons, etc. Nature 172: 407–408.
Thibodeaux LJ, Boyle JD. 1987. Bedform-generated convective transport in
bottom sediment. Nature 325: 341–343.
Trush WJ. 1989. Salmonid geomorphology. Proceedings of the Wild Trout
IV Symposium, Yellowstone National Park, Wyoming, September 18–19,
163–169.
Vaux WG. 1968. Intergravel flow and interchange of water in a
streambed U.S. Fish and Wildlife Service: Fishery Bulletin 66: 479–489.
Vogel S. 1994. Life in Moving Fluids: The Physical Biology of Flow. W.
Grant Press: Boston; 467.
Wadeson RA, Rowntree KM. 1998. Application of the hydraulic biotope
concept to the classification of instream habitats. Aquatic Ecosystems
Health and Management 1: 143–157.
Wheaton JM, Pasternack GB, Merz JE. 2004. Spawning habitat rehabilita-
tion – I. Conceptual approach and methods. International Journal of
River Basin Management 2(1): 3–20.
Wickett WP. 1954. The oxygen supply to salmon eggs in spawning beds.
Journal of the Fisheries Research Board of Canada 11(6): 933–953.
Witzel LD, MacCrimmon HR. 1983a. Redd-site selection by brook trout
and brown trout in southwestern Ontario streams. Transactions of the
American Fisheries Society 112: 760–771.
Witzel LD, MacCrimmon HR. 1983b. Embryo survival and alevin emergence
of brook charr, Salvelinus fontinalis, and brown trout, Salmo trutta, relative
to redd gravel composition. Canadian Journal of Zoology 61: 1783–1792.
Wolman MG. 1954. A method of sampling coarse river-bed material.
Transactions of the American Geophysical Union 35(6): 951–956.
Wu F.-C. 2000. Modeling embryo survival affected by sediment deposition
into salmonid spawning gravels: application to flushing flow prescrip-
tions. Water Resources Research 36(6): 1595–1603.
Zar JH. 1999. Biostatistical Analysis, 2nd edn. Prentice Halll, Upper Saddle
River: New Jersey; 663.
Zimmer MP, Power M. 2006. Brown trout habitat selection preferences and
redd characteristics in the Credit river, Ontario. Journal of Fish Biology
68: 1–14.
Zimmermann AE, Lapointe M. 2005. Intergranular flow velocity through
salmonid redds: sensitivity to fines infiltration from low intensity sedi-
ment transport events. River Research and Applications 21: 865–881.
Copyright # 2010 John Wiley & Sons, Ltd. River Res. Applic. 27: 345–359 (2011)
DOI: 10.1002/rra
HIGH-RESOLUTION INVESTIGATION OF REDDS 359





























