Embed Size (px)
DESCRIPTION
http://www.cdb.riken.jp/math2013/
Citation preview

確率的に表わされる過程柴田達夫
理化学研究所 発生・再生科学総合研究センター
数理生物学サマーレクチャーコース第2回2013/7/30

数理生物学サマーレクチャーコース第2回 2013/7/30
Fig. 2. Recorded random walk trajectories by Jean Baptiste Perrin [72]. Left part: three designs obtained by tracinga small grain of putty (mastic, used for varnish) at intervals of 30 s. One of the patterns contains 50 single points. Rightpart: the starting point of each motion event is shifted to the origin. The "gure illustrates the pdf of the travelled distancer to be in the interval (r, r#dr), according to (2!!!)"# exp(!r!/[2!!])2!rdr, in two dimensions, with the length variance!!. These "gures constitute part of the measurement of Perrin, Dabrowski and Chaudesaigues leading to the determina-tion of the Avogadro number. The result given by Perrin is 70.5!10!!. The remarkable wuvre of Perrin discusses allpossibilities of obtaining the Avogadro number known at that time. Concerning the trajectories displayed in the left partof this "gure, Perrin makes an interesting statement: `Si, en e!et, on faisait des pointeH s de seconde en seconde, chacun deces segments rectilignes se trouverait remplaceH par un contour polygonal de 30 co( teH s relativement aussi compliqueH que ledessin ici reproduit, et ainsi de suitea. [If, veritably, one took the position from second to second, each of these rectilinearsegments would be replaced by a polygonal contour of 30 edges, each itself being as complicated as the reproduceddesign, and so forth.] This already anticipates LeH vy's cognisance of the self-similar nature, see footnote 9, as well as of thenon-di!erentiability recognised by N. Wiener.
$ In the historical context, note that the theory of the continuum formulation of #uid dynamics had already been fullydeveloped at that time. Thus, some of its milestones date back to the 18th and "rst half of the 19th century, such asBernoulli's equation (1738); Euler's equation (1755); Navier's (1827) use as a phenomenological model and Stokes' (1845)derivation of the Navier}Stokes equation. Maxwells' dynamical theory of gases dates back to 1867 and Boltzmann'stransport equation was published in 1872 for the description of collision processes. The latter is the footing for theatomistic random walk approach to Brownian motion.
A. Fick set up the di!usion equation in 1855 [68].$ Subsequently, the detailed experiments by Gouyproved the kinetic theory explanation given by C. Weiner in 1863. After attempts of "ndinga stochastic footing like the collision model by von NaK geli and John William Strutt, LordRayleigh's results, it was Albert Einstein who, in 1905, uni"ed the two approaches in his treatises onthe Brownian motion, a name coined by Einstein although he reportedly did not have access toBrown's original work. Note that a similar description of di!usion was presented by the French
8 R. Metzler, J. Klafter / Physics Reports 339 (2000) 1}77
Nobel Prizes Alfred Nobel Educational Video Player Nobel Organizations Search
About the Nobel Prizes
Facts and Lists
Nobel Prize in PhysicsAll Nobel Prizes in Physics
Facts on the Nobel Prize inPhysics
Prize Awarder for the NobelPrize in Physics
Nomination and Selection ofPhysics Laureates
Nobel Medal for Physics
Articles in Physics
Video Nobel Lectures
Nobel Prize in Chemistry
Nobel Prize in Medicine
Nobel Prize in Literature
Nobel Peace Prize
Prize in Economic Sciences
Nobel Laureates Have Their Say
Nobel Prize Award Ceremonies
Nomination and Selection ofNobel Laureates
1901 2010
Sort and list Nobel Prizes and Nobel Laureates Prize category:
1926
Jean Baptiste Perrin
Physics
The Nobel Prize in Physics 1926Jean Baptiste Perrin
The Nobel Prize in Physics 1926
Jean Baptiste Perrin
The Nobel Prize in Physics 1926 was awarded to Jean Baptiste Perrin "for hiswork on the discontinuous structure of matter, and especially for his discovery ofsedimentation equilibrium".
Photos: Copyright © The Nobel Foundation
TO CITE THIS PAGE:MLA style: "The Nobel Prize in Physics 1926". Nobelprize.org. 10 Jun 2011http://nobelprize.org/nobel_prizes/physics/laureates/1926/
RELATED DOCUMENTS:
ARTICLE
PHYSICS
The Nobel Prize inPhysics
Read more about theNobel Prize in Physics 1901-2000.
RECOMMENDED:
FACTS AND LISTS
NOBEL PRIZESWho Are the 2010Nobel Laureates?
See a list of the elevenNobel Laureates of 2010.
HAVE YOUR SAY!
What invention has mostaffected your life?
FACTS AND LISTS
NOBEL PRIZESNobel Prize Facts
Find out more aboutthe oldest, youngest, most
awarded Nobel Laureates.
SIGN UP
FOLLOW US
Youtube
Nobelprize.org Monthly
RSS
About Nobelprize.org Privacy Policy Terms of Use Technical Support Copyright © Nobel Media AB 2011
Home A-Z Index FAQ Press Contact Us
Home A-Z Index FAQ Press Contact Us
Home / Nobel Prizes / Nobel Prize in Physics / The Nobel Prize in Physics 1926
! Metzler, R. & Klafter, J. The random walk's guide to anomalous diffusion: a fractional dynamics approach. Phys Rep 339, 1–77 (2000).
米沢富美子 「ブラウン運動 」共立出版

数理生物学サマーレクチャーコース第2回 2013/7/30
Fig. 2. Recorded random walk trajectories by Jean Baptiste Perrin [72]. Left part: three designs obtained by tracinga small grain of putty (mastic, used for varnish) at intervals of 30 s. One of the patterns contains 50 single points. Rightpart: the starting point of each motion event is shifted to the origin. The "gure illustrates the pdf of the travelled distancer to be in the interval (r, r#dr), according to (2!!!)"# exp(!r!/[2!!])2!rdr, in two dimensions, with the length variance!!. These "gures constitute part of the measurement of Perrin, Dabrowski and Chaudesaigues leading to the determina-tion of the Avogadro number. The result given by Perrin is 70.5!10!!. The remarkable wuvre of Perrin discusses allpossibilities of obtaining the Avogadro number known at that time. Concerning the trajectories displayed in the left partof this "gure, Perrin makes an interesting statement: `Si, en e!et, on faisait des pointeH s de seconde en seconde, chacun deces segments rectilignes se trouverait remplaceH par un contour polygonal de 30 co( teH s relativement aussi compliqueH que ledessin ici reproduit, et ainsi de suitea. [If, veritably, one took the position from second to second, each of these rectilinearsegments would be replaced by a polygonal contour of 30 edges, each itself being as complicated as the reproduceddesign, and so forth.] This already anticipates LeH vy's cognisance of the self-similar nature, see footnote 9, as well as of thenon-di!erentiability recognised by N. Wiener.
$ In the historical context, note that the theory of the continuum formulation of #uid dynamics had already been fullydeveloped at that time. Thus, some of its milestones date back to the 18th and "rst half of the 19th century, such asBernoulli's equation (1738); Euler's equation (1755); Navier's (1827) use as a phenomenological model and Stokes' (1845)derivation of the Navier}Stokes equation. Maxwells' dynamical theory of gases dates back to 1867 and Boltzmann'stransport equation was published in 1872 for the description of collision processes. The latter is the footing for theatomistic random walk approach to Brownian motion.
A. Fick set up the di!usion equation in 1855 [68].$ Subsequently, the detailed experiments by Gouyproved the kinetic theory explanation given by C. Weiner in 1863. After attempts of "ndinga stochastic footing like the collision model by von NaK geli and John William Strutt, LordRayleigh's results, it was Albert Einstein who, in 1905, uni"ed the two approaches in his treatises onthe Brownian motion, a name coined by Einstein although he reportedly did not have access toBrown's original work. Note that a similar description of di!usion was presented by the French
8 R. Metzler, J. Klafter / Physics Reports 339 (2000) 1}77
x(t1), y(t1)( ), x(t2 ), y(t2 )( ),, x(tn ), y(tn )( )位置の観測値
確率論的 ↔ 決定論的
x, y( )確率変数 (stochastic variable)
確率過程 (stochastic process)
=確率変数の系列 (時系列)
! Metzler, R. & Klafter, J. The random walk's guide to anomalous diffusion: a fractional dynamics approach. Phys Rep 339, 1–77 (2000).

数理生物学サマーレクチャーコース第2回 2013/7/30
1次元ランダムウォーク
1. 各分子は速度vでτ秒毎に距離δ=vτだけ右か左に移動する
2. 各ステップで右に行く確率は1/2であり、左に行く確率は1/2 分子は水の分子と相互作用をすると、最後のステップでどちらへ動いたのか忘れてしまう
3. 各分子は他の全ての粒子と無関係に動く
-3δ -2δ -δ 0 δ 2δ 3δ

数理生物学サマーレクチャーコース第2回 2013/7/30
0
20
40
60
80
100
-10 -5 0 5 10
time
step
position
x(t)
x(0)

数理生物学サマーレクチャーコース第2回 2013/7/30
1次元ランダムウォーク
-3δ -2δ -δ 0 δ 2δ 3δ離散時刻t=nτ x(t) = �
n�
i=1
�i位置:
x(t) = �n�
i=1
�i = 0位置の期待値:
�i = �1, or 1ただし,確率1/2 1/2
• 粒子は平均としては動かない

数理生物学サマーレクチャーコース第2回 2013/7/30
1次元ランダムウォーク
離散時刻t=nτ x(t) = �n�
i=1
�i位置:
x(t) = �n�
i=1
�i = 0位置の期待値:
�i = �1, or 1ただし,確率1/2 1/2
01
平均2乗変位:(時刻tにおける位置の分散)
x(t)2 =��
n⇤
i=1
�i
⇥2= �2
� n⇤
i=1
�2i + 2n⇤
i=1
n⇤
j=i+1
�i� j
⇥2
= �2n⇤
i=1
�2i + 2n⇤
i=1
n⇤
j=i+1
�i� j2
= n�2 =�2
⇥t
-3δ -2δ -δ 0 δ 2δ 3δ

数理生物学サマーレクチャーコース第2回 2013/7/30
1次元ランダムウォーク
離散時刻t=nτ x(t) = �n�
i=1
�i位置:
x(t) = �n�
i=1
�i = 0位置の期待値:
�i = �1, or 1ただし,確率1/2 1/2
平均2乗変位: x(t)2 =�2
⇥t
x(t)2 = 2Dt D =�2
2⇥拡散係数
-3δ -2δ -δ 0 δ 2δ 3δ

数理生物学サマーレクチャーコース第2回 2013/7/30
0
20
40
60
80
100
-10 -5 0 5 10
time
step
position
|x(t)� x(0)|2 = 2Dt
x(t)
x(0)
�|x(t)� x(0)|2 =
⇤2Dt
平均2乗変位
確率過程の中の法則性
Mean Square DisplacementMSD
• 粒子の分布の広がりは時間の平方根に比例して大きくなる。

数理生物学サマーレクチャーコース第2回 2013/7/30
米沢富美子 「ブラウン運動 」共立出版

数理生物学サマーレクチャーコース第2回 2013/7/30
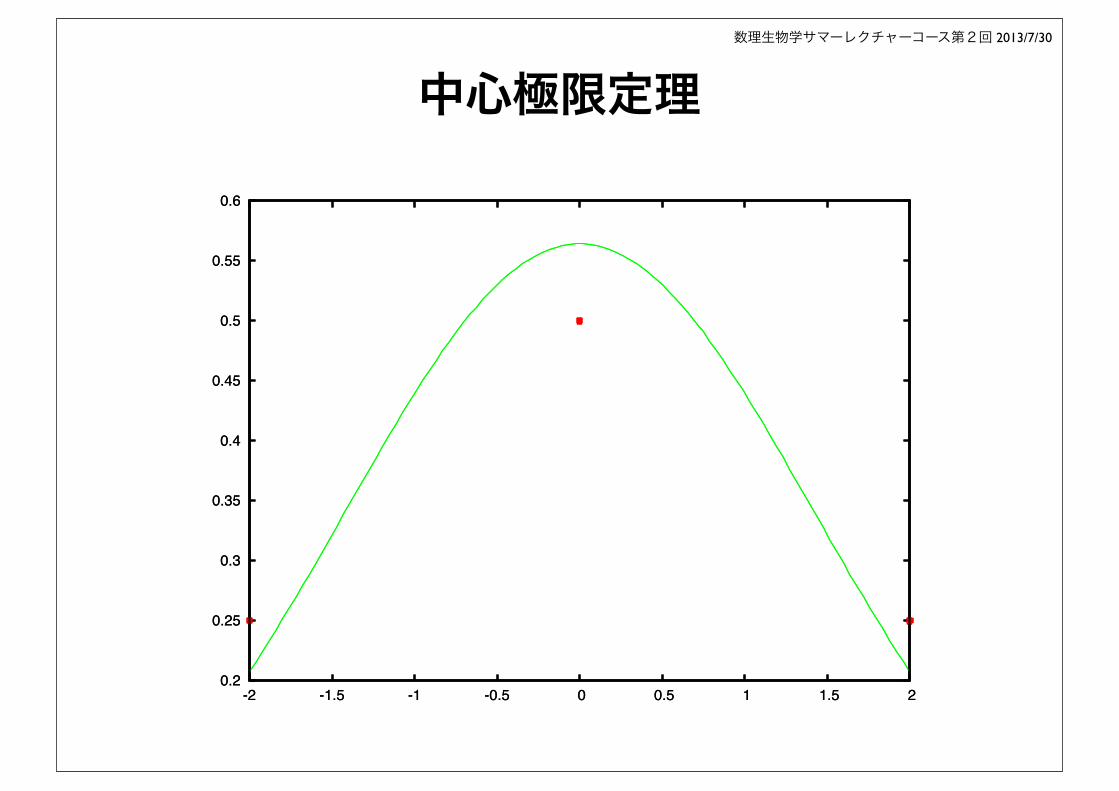
中心極限定理
0.45
0.5
0.55
0.6
0.65
0.7
0.75
0.8
-1 -0.5 0 0.5 1
g(x,1)
0.45
0.5
0.55
0.6
0.65
0.7
0.75
0.8
-1 -0.5 0 0.5 1
g(x,1)
数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0.2
0.25
0.3
0.35
0.4
0.45
0.5
0.55
0.6
-2 -1.5 -1 -0.5 0 0.5 1 1.5 2 0.2
0.25
0.3
0.35
0.4
0.45
0.5
0.55
0.6
-2 -1.5 -1 -0.5 0 0.5 1 1.5 2

数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
-4 -3 -2 -1 0 1 2 3 4 0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
-4 -3 -2 -1 0 1 2 3 4

数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0
0.05
0.1
0.15
0.2
0.25
0.3
-8 -6 -4 -2 0 2 4 6 8 0
0.05
0.1
0.15
0.2
0.25
0.3
-8 -6 -4 -2 0 2 4 6 8

数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0.2
-15 -10 -5 0 5 10 15 0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0.2
-15 -10 -5 0 5 10 15

数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
-30 -20 -10 0 10 20 30 0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
-30 -20 -10 0 10 20 30

数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
0.1
-60 -40 -20 0 20 40 60 0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
0.1
-60 -40 -20 0 20 40 60
数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
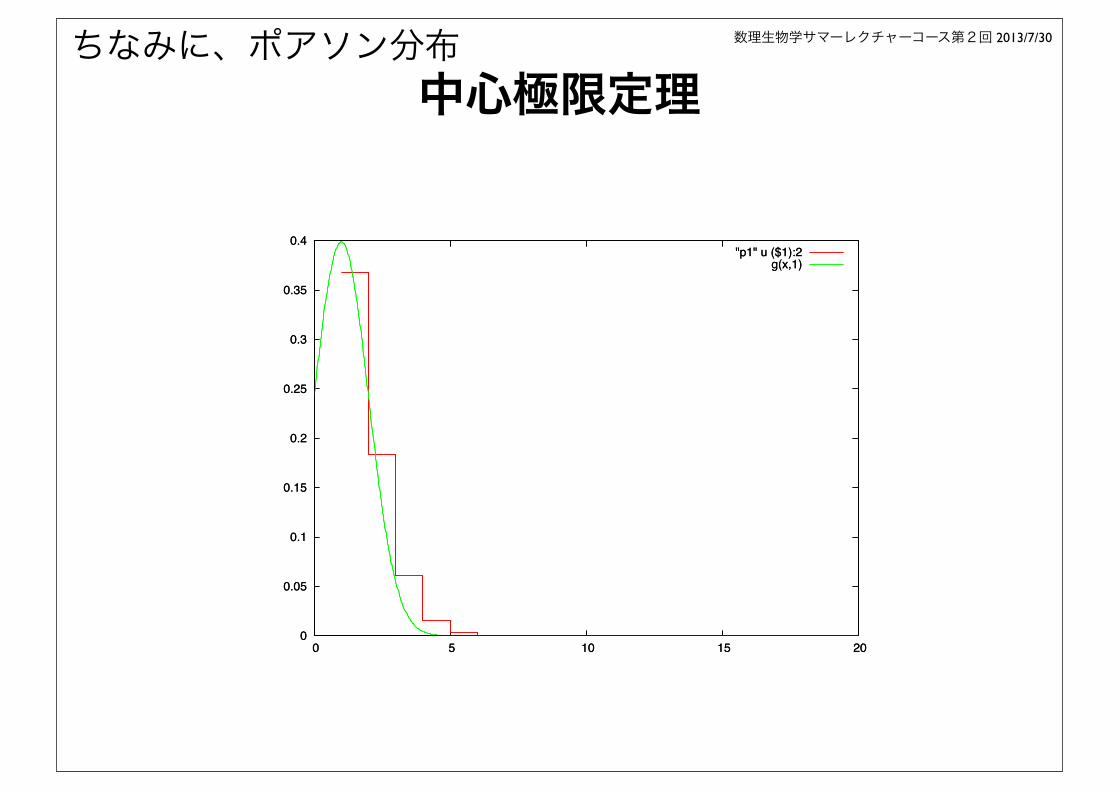
0 5 10 15 20
"p1" u ($1):2g(x,1)
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0 5 10 15 20
"p1" u ($1):2g(x,1)
ちなみに、ポアソン分布

数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0 5 10 15 20
"p5" u ($1):2g(x,5)
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0 5 10 15 20
"p5" u ($1):2g(x,5)
ちなみに、ポアソン分布

数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0 5 10 15 20
"p5" u ($1):2g(x,5)
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0 5 10 15 20
"p5" u ($1):2g(x,5)
ちなみに、ポアソン分布

数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
0
0.005
0.01
0.015
0.02
0.025
0.03
0.035
0.04
60 80 100 120 140
"p100" u ($1):2g(x,100)
0
0.005
0.01
0.015
0.02
0.025
0.03
0.035
0.04
60 80 100 120 140
"p100" u ($1):2g(x,100)
ちなみに、ポアソン分布

数理生物学サマーレクチャーコース第2回 2013/7/30
中心極限定理
Xn = x1 + x2 ++ xn
確率変数 xi
ただし、平均 m, 分散 s2 の同じ確率分布に従うとする
n が十分に大きいとき、Xnは平均 µ=nm, 分散σ2=ns2 のガウス分布に近づく
P(Xn ) =12πσ 2
e− (Xn−µ )
2
2σ 2

数理生物学サマーレクチャーコース第2回 2013/7/30
1次元ランダムウォーク
P(x,t x = 0,t = 0) = 14πDt
e− x2
4Dt
|x(t)� x(0)|2 = 2Dt平均2乗変位
D =�2
2⇥を一定に保って、δとτを小さくする
-3δ -2δ -δ 0 δ 2δ 3δ
t秒後の位置xはガウス分布に従う

数理生物学サマーレクチャーコース第2回 2013/7/30
1次元ランダムウォークを確率分布で考える
P(i,n) 粒子が離散時刻t=nτに位置x=δiにある確率
P(i,n +1) = P(i,n)+ 12P(i −1,n)+ 1
2P(i +1,n)− 2 1
2P(i,n)
P(x,t +τ ) = P(x,t)+ 12P(x −δ ,t)+ 1
2P(x +δ ,t)− 2 1
2P(x,t)
∂P(x,t)∂t
= δ 2
2τ∂2P(x,t)∂x2
時刻t+τ=(n+1)τの確率を時刻t=nτの確率で表わす
τ,δの十分小さい連続
-3δ -2δ -δ 0 δ 2δ 3δ

数理生物学サマーレクチャーコース第2回 2013/7/30
• 分子が細胞内にすっかり行き渡るのにかかる時間
tmix=L2/D L : 細胞サイズ(1~10μm)
tmix~10~0.1 sec• L : 胚 サイズ(100~1000μm)
tmix~103~105 sec• 粒子が2倍の距離を動こうとすれば4倍、10倍の距離を動こうとすれば100倍の時間がかかる。
細胞中のタンパク分子の拡散定数
水中 バクテリア 真核細胞 ミトコンドリア
細胞外マトリックス
拡散定数D
μm2/sec100程度 10程度 30程度 20~30程度 10~30程度
Xenopus early embryo, (inomata, et al. 2013)

数理生物学サマーレクチャーコース第2回 2013/7/30
拡散 VS 輸送
! Howard, J., Grill, S. W. & Bois, J. S. Turing's next steps: the mechanochemical basis of morphogenesis. Nature Reviews Molecular Cell Biology 12, 392–398 (2011).
• 輸送 (移流、advection)
- 典型的なモーターのスピード: v=1μm/sec
• 拡散- D=5 μm2/s; D=10 μm2/s; D=50 μm2/s
• 輸送が拡散を追い越すのは!
- D/v~ 5μm; D/v~ 10μm; D/v~50μm
• Péclet number : Pe=vL/D ; L=distance
- Pe << 1: 拡散
- Pe >> 1: 輸送
数理生物学サマーレクチャーコース第2回 2013/7/30
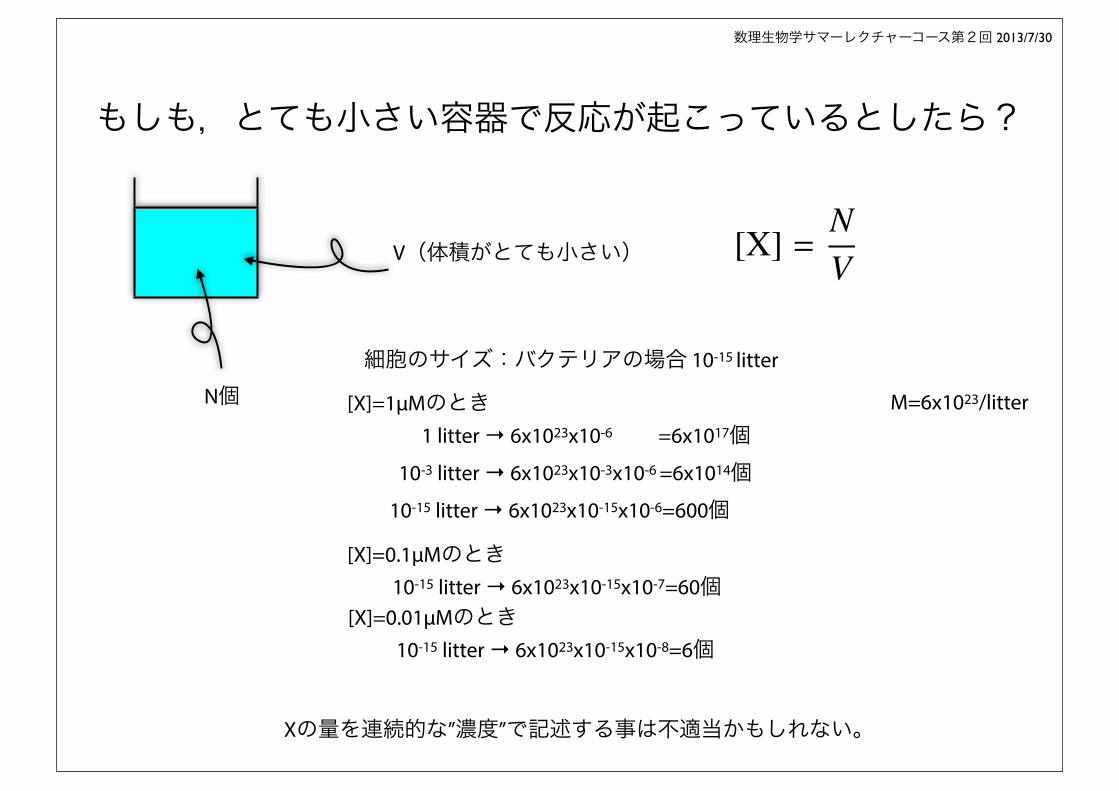
もしも,とても小さい容器で反応が起こっているとしたら?
Xの量を連続的な”濃度”で記述する事は不適当かもしれない。
N個
V(体積がとても小さい) [X] =NV
細胞のサイズ:バクテリアの場合 10-15 litter
[X]=1μMのとき 1 litter → 6x1023x10-6 =6x1017個 10-3 litter → 6x1023x10-3x10-6 =6x1014個10-15 litter → 6x1023x10-15x10-6=600個
[X]=0.1μMのとき10-15 litter → 6x1023x10-15x10-7=60個
10-15 litter → 6x1023x10-15x10-8=6個[X]=0.01μMのとき
M=6x1023/litter

数理生物学サマーレクチャーコース第2回 2013/7/30
濃度、分子数
体積体積体積体積
1 µm3
(E. coli)100 µm3
(核) 1,000 µm3
()10,000 µm3
(Fibroblast)
10 µM 6,000 600,000 6,000,000 60,000,0001 µM 600 60,000 600,000 6,000,000
100 nM 60 6000 60,000 600,00010 nM 6 600 6000 60,0001 nM 0.6 60 600 6,0001 pM 0.06 0.6 6

数理生物学サマーレクチャーコース第2回 2013/7/30
細胞中の化学反応は確率であらわされる• 化学反応は確率過程で記述される
• 反応速度定数 k:単位時間、分子あたりの反応確率
• 細胞中の反応確率 (単位時間あたり)
• その結果、濃度は時間的に確率的変動を示すV
E
A B
A+E k�⇥ B+E
200
250
300
350
400
0 100 200 300 400 500 600
濃度はゆらぐ 平均値: X
ゆらぎ=分散: σ2
k
VNANE

数理生物学サマーレクチャーコース第2回 2013/7/30
濃度の変化は体積に依存する
• 十分大きな体積中の化学反応と、極めて小さい体積中の化学反応は性質が大きく異なる
0 20 40 60 80
100
0 10 20 30 40 50
0 200 400 600 800
1000
0 10 20 30 40 50
0 2000 4000 6000 8000
10000
0 10 20 30 40 50
0 2 4 6 8 10
0 10 20 30 40 50time (s)
V=10
V=100
V=1000
V =∞
確率的
決定論的
ある化学反応を体積を変えながら調べた(ブリュッセレーター)

数理生物学サマーレクチャーコース第2回 2013/7/30
Single channel recording
216
directly the amplitude of current pulses
(about 3 :aA), and hence the conductance
of a singh: open ion channel, which is com-
monly about 30 pS (i.e. resistance about
33 000 rsegohms; one Sicmen = one
reciproca ohm). This is information that
cannot be obtained by the 'relaxation'
methods though it can be estimated h'om
noise analysis.
( 4 ) O b s e r r a t i o n s o f noise , o r f l~tf tualions,
in the r e s t ~ n s e to agonis~v.
When many ion channels are simulta-
neoasly open, the individual step-like tran.
sitions shown in Fig. 3 may (depending on
the recmding method) not be distinguish-
abl¢~ But they are there, and they give rise
to the nc,isiness of the record in the pres-
ence of a;gonists shown in Fig. 4A and B.
it has already been mentioned above
that the term equilibrium refers only to
arerage be,~¢iour. Imagine that there are
one million ion channels in the experimen-
tal preparation, and that we apply enough
agonist to open, at equilibrium, one chan-
nel in a thousand, Therefore at equilibrium
the~e art', on a r e r a g e . 1000 ion channels
open. But all of the channels are opening
and shutting at random and there is not
exact ly o n e thousand open at any pa~licular
moment. Suppose that an instantaneous
picture c~uld be taken of all the channels at
a partic~lar instant, and the state (open or
shut) of each one noted. This would he
e x ~ l y analogou[ to t~ssing a penny a mil-
lion times and noting, each time whether it
came do~vn "heads" (channel open) or tails
(channel shut). (In this case it is a very
b l a n d penny, that comes down heads o.~iy
once in 1000 throws on average.) Clearly
every time the experiment was repeated
(an instantaneous picture taken, or a mil-
lion tosses performed), we would not
expect to see exac t l y 1000 open channels
(or to get exactly 1000 'heads'). The prob-
lem can be treated by standard coin tossing
theory*, and the expected distribution of
the number of open channels is shown in
Fig. 4C. it has a standard deviation of 31.6
and a roughly Gaussian form. Sowe expect
that 95% of the time, the number of open
channels will be within two standard devia-
tions (i.e. 63.2) of the mean and we, should
therefore say, not that there are 1000
channels open at equilibrium, but that
there are 1000 ± 63.2. These
momeut- to-moment fluctuations (of
about +6 percent in this example) in the
number of open ion channels are quite big
"The binomia!_distribution with N = IO~channels.p = U.OOI ( p m b a b i l i l y Of being open ). gives the mean
numherofopenchannelsasNp = 1000. withstandard det-ialion ¢ ~ = 31.6.
enough to see, and they are what gives rise
to the fluctuations, o, noise, shown in Fig.
4A, B'.
Holy, then. can usehtl informatior
ext;actcd from the rather unpromis
looking signals shown in Fig. 4A, B? T .e
are two things that we can measure, the
amplitude of the noise, and its frequency.
Let us deal with these in turn.
N o i s e amplitud,~,. A conventional way to
measure the amplitude of a fluctuating cur-
rent (e.g. the alternating mains supply) is to
specify the rms ( : ~ t mean square) current,
which i~ ~:othi~: other than the standard
deviatioo of thc cerrent, calculated in the
ordinary way from a series of values of the
current measured at different instants. This
amplitude (standard deviation) is, for
example, clearly bigger for the record in
Fig. 4B than for that in Fig. 4A. The stan-
dasd dc~ia~a~, ~.~[ t i ~ current is a direct
measure of the standard deviation of the
number of open channels which was dis-
cussed above. If wc divide the square of the
observed standard deviation by the mean
current, we obtain an estimate of tbe cur-
rent through a single ion channel (around
3pA) and hence an estimate of the channel
conductance (as long as few channels are
open and a mechanism like equation 4 is
valid*).
N o i s e f r e q u e n c y . It is obvious by eye that
the noise in Fig. 4A contains higher fre-
quencies than that in Fig. 4B. it is therefore
natural to ask whet~ter anything interesting
can be learned from the frequency, and, if
~ , how it can be treasured. The answer to
the first question is yes; we can learn about
the rate at which channels open and shut.
The answer to the ~ccond question follows.
Fig. 5A shows, sehematically, the opening
and shutting of five: individual ion channels
(mean channel lifetime = 5 ms, say), and
Fig. 5B shows tht ~, sum of these records.
"l~e fluctuating record in Fig. 5B is jus,~ the
~ r l of noise illustrated in Fig. 4A,B
(~lhough the individual steps are too small
to be distinguished in the latter). Now a~
tiime zero it is seen that there are four ion
channels open (channels ~umber 1, 2, 4
and 5). If we look a bit later on, say ! .0 ms
later (as marked in Fig. 5B), there are still
~ u r channels open; not surprisingly, they
~ e the same four as were open originally,
I~¢cause, in a short time like 1.0 ms it is
¢!uite likely that none of these channels will
+ Under these conditions Np(I - p) ~- Np so the var- ianc~ o f the total conductance is approximalely y'Np (wl~m y is the conductance o f a single channel) whereas the mean conductance is 1/Np. Their ratio i~ simply 3/.
T I P S - A u g u s t 1981
have shut yet (and that no others will have
opened), it is therefore to be expected that
the current at any given moment will be
very similar to the current at a short time
later if, by short, we mean shor t relative to
the c h a n n e l l i f e t i m e . In other words the cur-
rent at any given moment will be highly
correlated with the current i .0 ms later, If
we calculate an ordinary correlation coef-
ficient between t h e ~ two quantities we
expect that it will be very good, i.e. near
unity, as shown in Fig. 5C. Now consider
what happens if, rather than waiting 1.0
ms, we wait 15 ms (see Fig. 5B). This is
quite long compared with the mean open
¢hem~ • Sins ~ .moan ~ l i f l~me
I I
~[ - L . _ Y ~ r ~ ' L . L Y - - - - ' L Y - ,
I', ~l --L÷+ ~ _ _ _ J -
I t Or t~ ~ ,
~.0 t -$ml l-ISmt
C. D
io 5 f t i m d d ~ m l t e sm~ll~t~.O~l frequency ll'lzJ tiog ~ b t
Fig. 5. Explanation o] the analysis o f the frequency
characteristics o f flucmations in the current elicited (i.e.
in the number orlon channels opened) by agonist~'.
A. Simulatedhehariourof~veindividuMionchannels
(opening is pirated downwards). They are opening and
shutting at random with a mean open li~time o f 5 ms.
B. Sum o f the fire r~'cords shown in A. The total
number open (and hence the tmal currenO shows fluc-
tuations o f the sort that gis'e rise to observations like
those in Fig. 4A,B. An arbiwarily chosen line marks
zero time, and the timex !. 5 and 15 ms later are also
marked with rertical lines.
C. The correlation analysis o fnoise. Two obsert'aCions
separated by I ms are likely to he highly correlated (ns
shown by the dot). The correlation wil' be less when dw
separation is 5 ms, and there is hardly any correlation
with a separation o f 15 ms. The correlation (according
to the mechanism in equation 4) dies out along the
exponential curre shown, with a lime constant o f • =
l / (a + 13'), The time constant is 3 ms in the ¢:'ample
shown; it is less than Ihe mean open channel lifetime (5
ms). because, as is clear from A, the channeh" are open
for a substantial pan o f the lime• (From e~iuation 5 it
follows that the equilil~ium fraction o f shut channels is
0.6. o f open channels is 0.4. ¢i = 1#5 ms) = 200 s -~
and 13' = 133.3 s-~" thus r = i 1333.3 = 3 ms),
D. Presemation o f the same analysis o f noise as a
power spectrum. The ordinate is normally in units o f
AmpllHz, but is giren in arbitrary uni~ here. The
amoum o f noiw is hah'ed (from I00 to 50) at 53.1 H: .
so v = 11(2n ! 53.D = 3 ms. e.mctly ns found in C.
ひとつのチャンネルが開いている時間の分布は指数関数にしたがっている
p(t) ⇥ e��t
1.! Sakmann, B. & Neher, E. Single-channel Recording. (Springer, 2009).
数理生物学サマーレクチャーコース第2回 2013/7/30
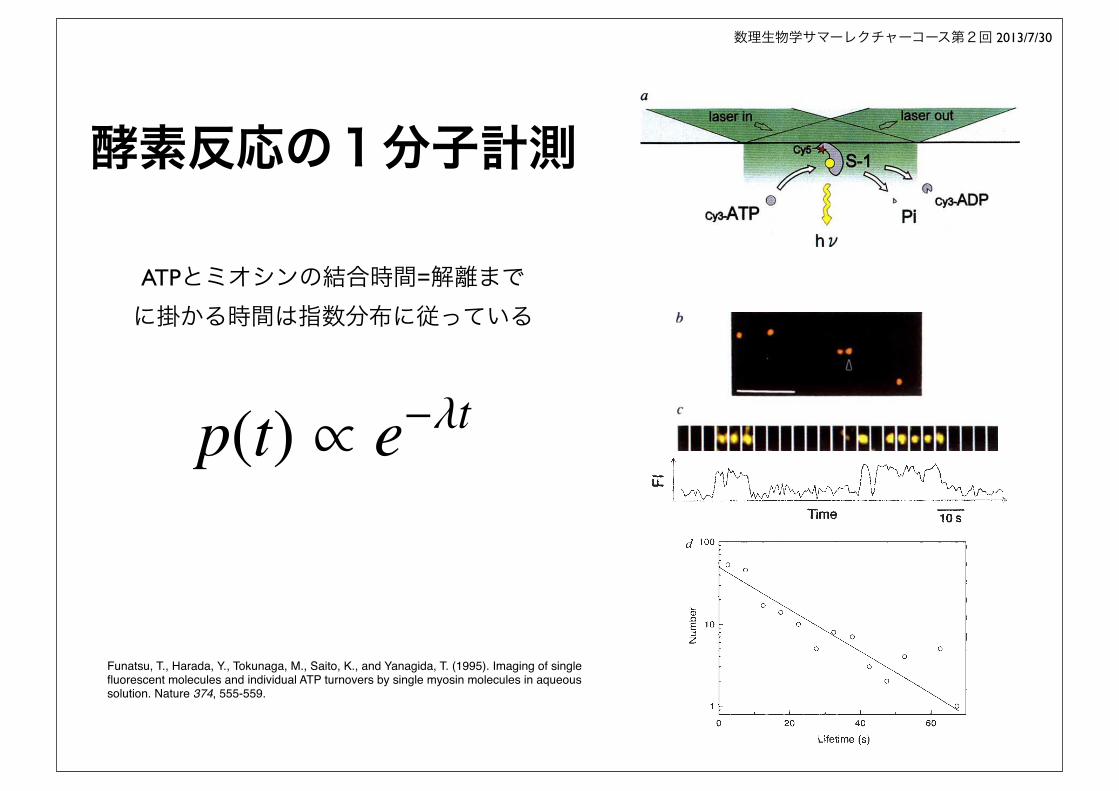
酵素反応の1分子計測
Funatsu, T., Harada, Y., Tokunaga, M., Saito, K., and Yanagida, T. (1995). Imaging of single fluorescent molecules and individual ATP turnovers by single myosin molecules in aqueous solution. Nature 374, 555-559.
ATPとミオシンの結合時間=解離までに掛かる時間は指数分布に従っている
p(t) ⇥ e��t
数理生物学サマーレクチャーコース第2回 2013/7/30
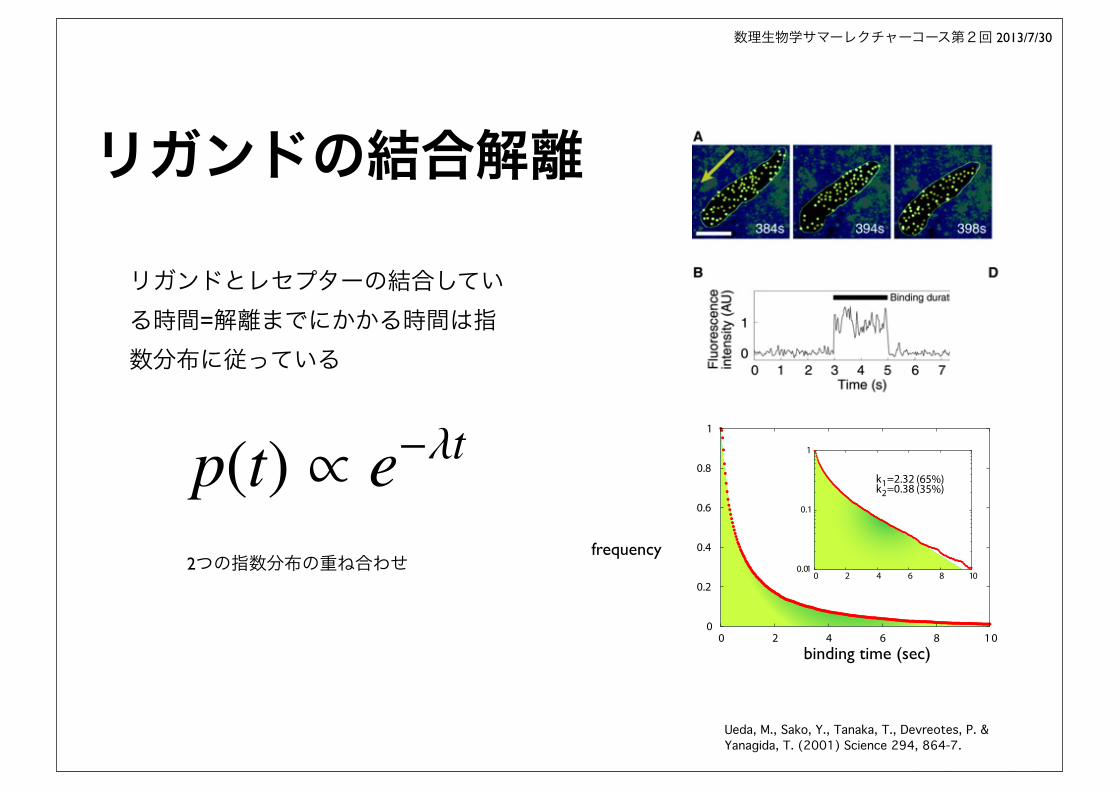
リガンドの結合解離
Ueda, M., Sako, Y., Tanaka, T., Devreotes, P. & Yanagida, T. (2001) Science 294, 864-7.
2つの指数分布の重ね合わせ
binding time (sec)
binding time (sec)
frequency
リガンドとレセプターの結合している時間=解離までにかかる時間は指数分布に従っている
p(t) ⇥ e��t
数理生物学サマーレクチャーコース第2回 2013/7/30
ON
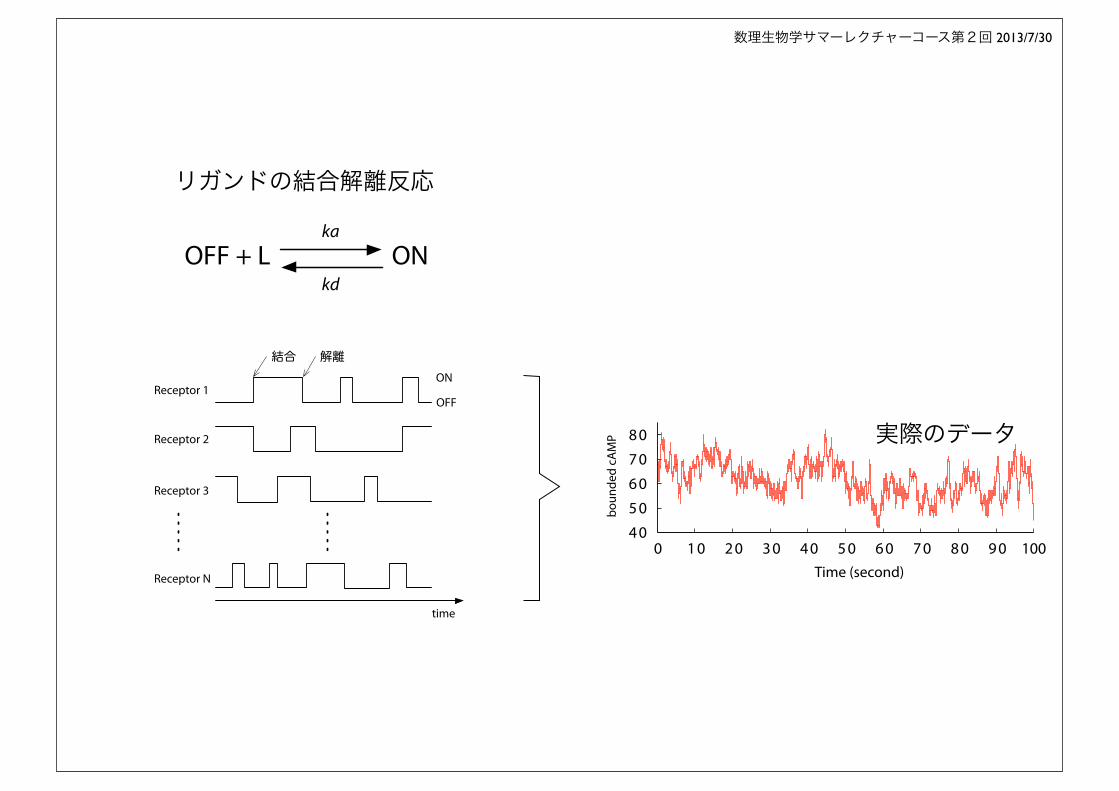
OFFReceptor 1
結合 解離
time
4 0
5 0
6 0
7 0
8 0
0 1 0 2 0 3 0 4 0 5 0 6 0 7 0 8 0 9 0 100Time (second)
boun
ded
cAM
P 実際のデータ
ON
OFF
time
Receptor 1
Receptor 2
Receptor 3
Receptor N
結合 解離
OFF + L ONka
kd
リガンドの結合解離反応

数理生物学サマーレクチャーコース第2回 2013/7/30
1次反応を考えよう:分解反応
d[X]dt= ��[X]
[X]t = [X]0e��t
X��⇥
0.0 0.5 1.0 1.5 2.0
0.2
0.4
0.6
0.8
1.0
[X]t
t

数理生物学サマーレクチャーコース第2回 2013/7/30
1つの分子の反応を考えるたとえば,,
t=0
いつ反応するか?
X��⇥ いつ消滅するか?例1
Xk�⇥ Y いつYに変化するか?例2
数理生物学サマーレクチャーコース第2回 2013/7/30
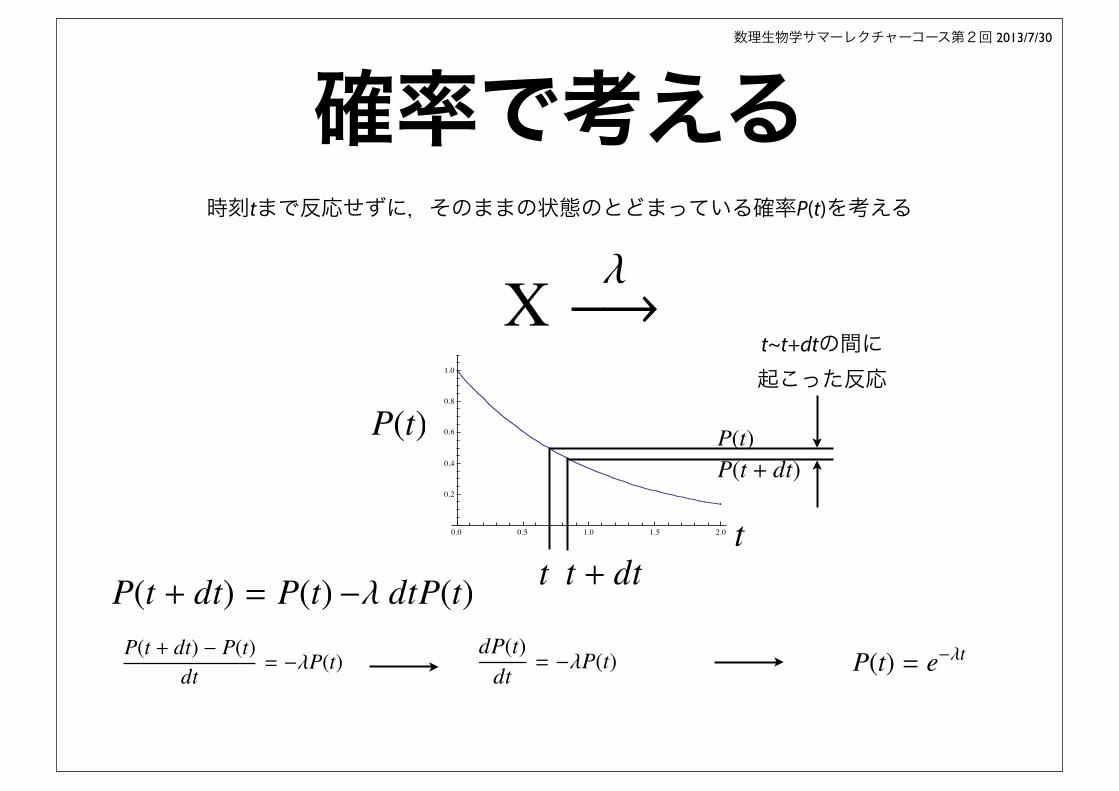
確率で考える時刻tまで反応せずに,そのままの状態のとどまっている確率P(t)を考える
0.0 0.5 1.0 1.5 2.0
0.2
0.4
0.6
0.8
1.0
t
P(t)
t t + dt
t~t+dtの間に起こった反応
P(t + dt) = P(t)
X��⇥
�� dtP(t)P(t + dt) � P(t)
dt= ��P(t)
dP(t)dt= ��P(t) P(t) = e��t
P(t)P(t + dt)

数理生物学サマーレクチャーコース第2回 2013/7/30
0 = t0 < t1 < t2 << tn−1 < tn = T
x(t1), x(t2 ),, x(tn )
観測時刻
ti = i × Δt
観測値
x =01⎧⎨⎩
(off)
(on)
216
directly the amplitude of current pulses
(about 3 :aA), and hence the conductance
of a singh: open ion channel, which is com-
monly about 30 pS (i.e. resistance about
33 000 rsegohms; one Sicmen = one
reciproca ohm). This is information that
cannot be obtained by the 'relaxation'
methods though it can be estimated h'om
noise analysis.
( 4 ) O b s e r r a t i o n s o f noise , o r f l~tf tualions,
in the r e s t ~ n s e to agonis~v.
When many ion channels are simulta-
neoasly open, the individual step-like tran.
sitions shown in Fig. 3 may (depending on
the recmding method) not be distinguish-
abl¢~ But they are there, and they give rise
to the nc,isiness of the record in the pres-
ence of a;gonists shown in Fig. 4A and B.
it has already been mentioned above
that the term equilibrium refers only to
arerage be,~¢iour. Imagine that there are
one million ion channels in the experimen-
tal preparation, and that we apply enough
agonist to open, at equilibrium, one chan-
nel in a thousand, Therefore at equilibrium
the~e art', on a r e r a g e . 1000 ion channels
open. But all of the channels are opening
and shutting at random and there is not
exact ly o n e thousand open at any pa~licular
moment. Suppose that an instantaneous
picture c~uld be taken of all the channels at
a partic~lar instant, and the state (open or
shut) of each one noted. This would he
e x ~ l y analogou[ to t~ssing a penny a mil-
lion times and noting, each time whether it
came do~vn "heads" (channel open) or tails
(channel shut). (In this case it is a very
b l a n d penny, that comes down heads o.~iy
once in 1000 throws on average.) Clearly
every time the experiment was repeated
(an instantaneous picture taken, or a mil-
lion tosses performed), we would not
expect to see exac t l y 1000 open channels
(or to get exactly 1000 'heads'). The prob-
lem can be treated by standard coin tossing
theory*, and the expected distribution of
the number of open channels is shown in
Fig. 4C. it has a standard deviation of 31.6
and a roughly Gaussian form. Sowe expect
that 95% of the time, the number of open
channels will be within two standard devia-
tions (i.e. 63.2) of the mean and we, should
therefore say, not that there are 1000
channels open at equilibrium, but that
there are 1000 ± 63.2. These
momeut- to-moment fluctuations (of
about +6 percent in this example) in the
number of open ion channels are quite big
"The binomia!_distribution with N = IO~channels.p = U.OOI ( p m b a b i l i l y Of being open ). gives the mean
numherofopenchannelsasNp = 1000. withstandard det-ialion ¢ ~ = 31.6.
enough to see, and they are what gives rise
to the fluctuations, o, noise, shown in Fig.
4A, B'.
Holy, then. can usehtl informatior
ext;actcd from the rather unpromis
looking signals shown in Fig. 4A, B? T .e
are two things that we can measure, the
amplitude of the noise, and its frequency.
Let us deal with these in turn.
N o i s e amplitud,~,. A conventional way to
measure the amplitude of a fluctuating cur-
rent (e.g. the alternating mains supply) is to
specify the rms ( : ~ t mean square) current,
which i~ ~:othi~: other than the standard
deviatioo of thc cerrent, calculated in the
ordinary way from a series of values of the
current measured at different instants. This
amplitude (standard deviation) is, for
example, clearly bigger for the record in
Fig. 4B than for that in Fig. 4A. The stan-
dasd dc~ia~a~, ~.~[ t i ~ current is a direct
measure of the standard deviation of the
number of open channels which was dis-
cussed above. If wc divide the square of the
observed standard deviation by the mean
current, we obtain an estimate of tbe cur-
rent through a single ion channel (around
3pA) and hence an estimate of the channel
conductance (as long as few channels are
open and a mechanism like equation 4 is
valid*).
N o i s e f r e q u e n c y . It is obvious by eye that
the noise in Fig. 4A contains higher fre-
quencies than that in Fig. 4B. it is therefore
natural to ask whet~ter anything interesting
can be learned from the frequency, and, if
~ , how it can be treasured. The answer to
the first question is yes; we can learn about
the rate at which channels open and shut.
The answer to the ~ccond question follows.
Fig. 5A shows, sehematically, the opening
and shutting of five: individual ion channels
(mean channel lifetime = 5 ms, say), and
Fig. 5B shows tht ~, sum of these records.
"l~e fluctuating record in Fig. 5B is jus,~ the
~ r l of noise illustrated in Fig. 4A,B
(~lhough the individual steps are too small
to be distinguished in the latter). Now a~
tiime zero it is seen that there are four ion
channels open (channels ~umber 1, 2, 4
and 5). If we look a bit later on, say ! .0 ms
later (as marked in Fig. 5B), there are still
~ u r channels open; not surprisingly, they
~ e the same four as were open originally,
I~¢cause, in a short time like 1.0 ms it is
¢!uite likely that none of these channels will
+ Under these conditions Np(I - p) ~- Np so the var- ianc~ o f the total conductance is approximalely y'Np (wl~m y is the conductance o f a single channel) whereas the mean conductance is 1/Np. Their ratio i~ simply 3/.
T I P S - A u g u s t 1981
have shut yet (and that no others will have
opened), it is therefore to be expected that
the current at any given moment will be
very similar to the current at a short time
later if, by short, we mean shor t relative to
the c h a n n e l l i f e t i m e . In other words the cur-
rent at any given moment will be highly
correlated with the current i .0 ms later, If
we calculate an ordinary correlation coef-
ficient between t h e ~ two quantities we
expect that it will be very good, i.e. near
unity, as shown in Fig. 5C. Now consider
what happens if, rather than waiting 1.0
ms, we wait 15 ms (see Fig. 5B). This is
quite long compared with the mean open
¢hem~ • Sins ~ .moan ~ l i f l~me
I I
~[ - L . _ Y ~ r ~ ' L . L Y - - - - ' L Y - ,
I', ~l --L÷+ ~ _ _ _ J -
I t Or t~ ~ ,
~.0 t -$ml l-ISmt
C. D
io 5 f t i m d d ~ m l t e sm~ll~t~.O~l frequency ll'lzJ tiog ~ b t
Fig. 5. Explanation o] the analysis o f the frequency
characteristics o f flucmations in the current elicited (i.e.
in the number orlon channels opened) by agonist~'.
A. Simulatedhehariourof~veindividuMionchannels
(opening is pirated downwards). They are opening and
shutting at random with a mean open li~time o f 5 ms.
B. Sum o f the fire r~'cords shown in A. The total
number open (and hence the tmal currenO shows fluc-
tuations o f the sort that gis'e rise to observations like
those in Fig. 4A,B. An arbiwarily chosen line marks
zero time, and the timex !. 5 and 15 ms later are also
marked with rertical lines.
C. The correlation analysis o fnoise. Two obsert'aCions
separated by I ms are likely to he highly correlated (ns
shown by the dot). The correlation wil' be less when dw
separation is 5 ms, and there is hardly any correlation
with a separation o f 15 ms. The correlation (according
to the mechanism in equation 4) dies out along the
exponential curre shown, with a lime constant o f • =
l / (a + 13'), The time constant is 3 ms in the ¢:'ample
shown; it is less than Ihe mean open channel lifetime (5
ms). because, as is clear from A, the channeh" are open
for a substantial pan o f the lime• (From e~iuation 5 it
follows that the equilil~ium fraction o f shut channels is
0.6. o f open channels is 0.4. ¢i = 1#5 ms) = 200 s -~
and 13' = 133.3 s-~" thus r = i 1333.3 = 3 ms),
D. Presemation o f the same analysis o f noise as a
power spectrum. The ordinate is normally in units o f
AmpllHz, but is giren in arbitrary uni~ here. The
amoum o f noiw is hah'ed (from I00 to 50) at 53.1 H: .
so v = 11(2n ! 53.D = 3 ms. e.mctly ns found in C.
確率変数
確率過程

数理生物学サマーレクチャーコース第2回 2013/7/30
P(x0 )
P(x1, x0 )
P(xn ,, x1, x0 )
経路の確率
遷移確率 P(x1 x0 ) =P(x1, x0 )P(x0 )
P(xn ,, x1 x0 ) =
P(xn ,, x1, x0 )P(x0 )
P(xn xn−1,, x1, x0 ) =
P(xn ,, x1, x0 )P(xn−1,, x1, x0 )

数理生物学サマーレクチャーコース第2回 2013/7/30
P(x2, x1 x0 ) = P(x2 x1)P(x1 x0 )
P(x2, x1, x0 ) = P(x2 x1)P(x1 x0 )P(x0 )
P(xn xn−1,, x1, x0 ) = P(xn xn−1)
マルコフ過程 (Markov process)
P(x2 x0 ) = P(x2 x1)P(x1 x0 )x1∑
(Chapman-Kolmogorov equation)

数理生物学サマーレクチャーコース第2回 2013/7/30
0.0 0.5 1.0 1.5 2.0
0.2
0.4
0.6
0.8
1.0
t
時刻tにXである確率p(t) = e−λt q(t) = 1− p(t) = 1− e−λt
時刻tにYである確率
X λ⎯ →⎯ Y
P(Y,t X,0) = 1− e−λtP(X,t X,0) = e−λt
P(Y,2t;X,t X,0) = (1− e−λt )e−λt = P(Y,2t X,t)P(X,t X,0)P(Y,2t;Y,t X,0) = 1− e−λt = P(Y,2t Y,t)P(Y,t X,0)
P(Y,2t X,0) = 1− e−λ (2t ) (Chapman-Kolmogorov equation)

数理生物学サマーレクチャーコース第2回 2013/7/30
• 位置Xはマルコフ過程ではない
• なぜなら、時刻tの位置Xは、t-1, t-2,,に依存する
• 位置Xと速度Vはマルコフ過程になる
���� ��� � �� ����
�����
�����
�����
����
����
��������
��� � ��� i
���� ��� � �� ����
�����
����
�����
��� ����
������ i
� ���� ���� ���� ���� ���������
���
�
��
���
�������
� �� ��
i
A
B濃度の増加
濃度の増加
-1 0 1-2 2
C
D
・ ・ ・ ・・ ・ ・ ・
Fig.3
E
通常の運動
q>pp0 適応
濃度の減少
適応濃度減少時の運動濃度増加時の運動
濃度の減少
方向転換の確率
v -vv

数理生物学サマーレクチャーコース第2回 2013/7/30
X λ0⎯ →⎯ X' λ1⎯ →⎯ Y
直接観測できない中間状態X’がある
X λ⎯ →⎯ Y
2 4 6 8 10
0.6
0.7
0.8
0.9
1.0
時刻tにX or X’である確率時刻tにYでない確率
2 4 6 8 10
0.05
0.10
0.15
0.20
0.25
Yになるまでの時間tの確率分布関数
p(t) = e−λ0t − e−λ1t
λ1 − λ0
1−0
t
∫ p(s)ds

数理生物学サマーレクチャーコース第2回 2013/7/30
P(Y,t X,0) =0
t
∫ p(s)dsP(X,t X,0) = 1−
0
t
∫ p(s)ds
P(Y,2t;X,t X,0) = 1−0
t
∫ p(s)ds( ) 0
t
∫ p(s)dsP(Y,2t;Y,t X,0) = 1−
0
t
∫ p(s)ds
P(Y,2t;X,t X,0)+ P(Y,2t;Y,t X,0) =0
t
∫ p(s)ds( )2≠ P(Y,2t X,0)
p(t) = e−λ0t − e−λ1t
λ1 − λ0Yになるまでの時間tの確率分布関数
XとX’は区別ができないとする。それらをまとめてXと書く

数理生物学サマーレクチャーコース第2回 2013/7/30
生成,分解反応/ birth-and-death process
k�⇥ X, X��⇥
転写,翻訳 分解例えば,遺伝子発現
N個ある確率をP(N,t)とおく。
P(N) = (k/�)N
N! e�k� ポアソン分布
N = k�
平均: ⇥2 = N2 � N2= k�分散:
定常分布� dPdt = 0
⇥
dP(N, t)dt
= k�P(N � 1, t) � P(N, t)
⇥+ ��(N + 1)P(N + 1, t) � NP(N, t)
⇥
生成反応 分解反応
N � �2

数理生物学サマーレクチャーコース第2回 2013/7/30
組織中の細胞の多様性はどのように維持されているか
1.! Klein, A. M. & Simons, B. D. Universal patterns of stem cell fate in cycling adult tissues. Development 138, 3103–3111 (2011).

数理生物学サマーレクチャーコース第2回 2013/7/30
Single-proliferative compartment model of IFE maintenance
With this background, we turn now to the consideration of the novel single-proliferative
compartment model introduced in the main text. When formulated as a non-equilibrium
process, the model is described by the set of non-equilibrium rate equations
A��!
8>>>><
>>>>:
A + A Prob. r
A + B Prob. 1� 2r
B + B Prob. r
B��! C,
(5)
where, as usual, A denote EPCs, B represent post-mitotic cells in the basal layer, and C
specify the suprabasal layer cells. To maintain the total basal layer cell population, we
must impose the condition that � = ⇢1�⇢�, i.e. the net rate at which post-mitotic cells are
generated in the basal layer, ⇢�, is compensated by the rate at which they are removed,
(1 � ⇢)�. By neglecting processes involving the shedding of cells from the surface of the
epidermis, the model is restricted to the consideration of the total clone size distribution
at appropriately short time scales. However, if we focus only on the clone size distribution
associated with those cells which occupy the basal layer, the model can be applied up to
arbitrary time scales. For this case, the transfer process must be replaced by one in which
B type cells are lost from the distribution at a rate �, i.e. B��! ;. In either case, the
time evolution associated with (5) can be cast in the form of a Master equation. Defining
PnA,nB(t) as the probability of finding nA type A cells and nB type B cells in a given clone
after some time t, the basal layer cell probability distribution evolves according to the Master
equation
@tPnA,nB = r� [(nA � 1)PnA�1,nB � nAPnA,nB ] + r�[(nA + 1)PnA+1,nB�2 � nAPnA,nB ]
+(1� 2r)�[nAPnA,nB�1 � nAPnA,nB ] + �[(nB + 1)PnA,nB+1 � nBPnA,nB ],
6
pn (t) ~τtexp −n τ
t⎛⎝⎜
⎞⎠⎟
十分に時間が経てば
τ = ρrλ
Γ = ρ1− ρ
λただし、
ρは、Aの割合
スケーリング則
パラメータフィッティングの結果
比較的簡単なマルコフモデルが実験結果をよく説明する
実験とモデルの比較
1.! Clayton, E. et al. A single type of progenitor cell maintains normal epidermis. Nature 446, 185–189 (2007).2.! Klein, A. M., Doupé, D. P., Jones, P. H. & Simons, B. D. Kinetics of cell division in epidermal maintenance. Phys Rev E 76, 021910 (2007).

まとめ/キーワード• 確率変数
• 確率過程
• ランダムウォーク、拡散
• 平均二乗変位 (mean
square displacement, MSD)
• 中心極限定理、ガウス分布、正規分布
• 指数分布
• 経路の確率
• 遷移確率
• マルコフ過程、非マルコフ過程
• 生成消滅過程, birth-and-
death process

数理生物学サマーレクチャーコース第2回 2013/7/30
• ハーケン「Synergetics, 共同現象の数理」東海大学出版
• W. フェラー「確率論とその応用」紀伊国屋書店
• ハワード バーグ 「生物学におけるランダムウォーク」 法政大学出版局
• 寺本英「ランダムな現象の数学」、吉岡書店
• Van Kampen, “Stochastic Processes in Physics and Chemistry”, North Holland





























