Embed Size (px)
Citation preview

1
Testing for Xylem Vulnerability to Embolism
and Embolism Repair in Citrus species
An undergraduate Thesis
Presented to the Faculty of the Southern California Ecosystems Research Program
Department of Biological Science
California State University, Fullerton
Emily Nguyen Wieber
Approved by
Dr. H. Jochen Schenk (Thesis Advisor) Date
Dr. Darren Sandquist (SCERP Director) Date
Dr. Bill Hoese (SCERP co-Director) Date

2
General Introduction
In vascular plants, xylem is responsible for transporting water and inorganic and organic
solutes, termed xylem sap. Xylem vessels are composed of tracheids, vessel elements, and
parenchyma. Constant leaves’ evaporation generate a negative pressure gradient that draw the
water from the soil to the root to the leaves. The soil has higher water potential than the roots
because the roots contain higher concentration of solutes. Based on diffusion principle, water
flows from high to low water potential, thus water flow from the soil into the roots. Hydrogen
bonds links water molecules together; water molecules adhere to the hydrophilic of cell walls.
The cohesion and adhesion of water molecules are assisted with capillary action pull water
upwards against gravity (Joly and Dixon, 1894). Since leaves constantly evaporate water; there
is always tension, negative pressure on the surface of the leaves. As the water is pulled, many
menisci are formed in small diameter xylem such as capillary action. This tension imposes a
strong pressure on the xylem, demanding for more water. If the soil is dry, there is not enough
water to supply water demand; air is drawn into the xylem through the small spores between the
conduits, call pit membranes. Hacke and Sperry (2001) referred to this process as air seeding.
Xylem embolisms can impede water flow through the xylem, and if not removed, can lead to leaf
and branch mortality. Embolisms formation can be detected by measuring hydraulic
conductivity of stems before and after hydrating stems to fill all conduits with water. A lower
hydraulic conductivity before hydration indicates embolism formation. A higher resistance to
embolism formation indicates a high tolerance to water deficiency (Hacke & Sperry 2001;
Melcher et al. 2003; Poggi et al. 2007).
As water resources for irrigation of crops and fruit trees are becoming scarce in many
regions, there is a high need to study the drought resistance mechanisms of fruit species,
including Citrus varieties. Growing Citrus in Southern California, a Mediterranean climate,

3
requires efficient irrigation to maintain satisfactory tree growth and fruit development. It has
been hypothesized that the vulnerability to embolism formation may be an important factor for
determining the timing and amounts of irrigation, especially when irrigation water is scarce
(Poggi et al. 2007). The main objective of my research was to determine the vulnerability of
Citrus species to embolism formation and test for the capability to repair embolisms, with the
intent to contribute to more effective irrigation scheduling for Citrus in California. I measured
Citrus vulnerability to embolisms and tested different hydration methods to obtain maximum
hydraulic conductance as a baseline for all measurements. The best hydration method was then
used to hydrate Citrus stems throughout the subsequent study.
Another set of experiments was designed to test for the ability of Citrus to form and
repair xylem embolisms. It is commonly observed that embolisms form during daytime while
the hydraulic systems experience maximum transpiration and the strongest negative pressure.
Embolism repair in the past was thought to take place only in the absence of transpiration and
while xylem tension is low. However, recent studies have indicated that embolisms can be
repaired during transpiration and under negative pressure (Clearwater and Goldstein 2005). The
physiological conditions under which embolisms form and/ or are repaired were documented by
measurements of leaf stomata conductance, branch pressure potentials, and stem hydraulic
conductance. Three 24-hour experiments were conducted to measure the degree of embolism
formation and repair in fall 2011, and spring and summer of 2012.

4
Part I: Measuring Xylem Vulnerability in Three Citrus Species
Abstract
This study documented xylem vulnerability of Citrus x sinensis (Valencia orange), Citrus
x paradisi (star ruby red grapefruit), and Citrus x reticulata (tangerine) under drought conditions.
During water shortages, a better understanding of drought tolerance mechanisms in citrus would
enable to improve water conservation. Physiological mechanisms of drought resistance were
investigated by measuring the formation of xylem embolisms, also known as air bubbles that
prevent water flow through vascular tissue. Four methods were compared to determine the best
technique for hydrating the stems in order to achieve maximum hydraulic conductivity through
the xylem: 1) high pressure flushing for one hour at room temperature, 2) sixteen-hour vacuum
treatment of submerged stems at room temperature, 3) low pressure flushing at 12oC for 16
hours, and 4) one-hour vacuum treatment of submerged stems followed by one-hour high
pressure flushing at room temperature. Based on previous results and studies, we hypothesized
that submergence under a vacuum would be the best method of stem hydration because it would
effectively remove trapped air molecules from pit membranes. Based on differences in fruit
sizes, we hypothesized that grapefruit would be the most vulnerable to embolism formation.
High-pressure flushing was a widely used standard method that produced the lowest hydraulic
conductivity, whereas, long-term vacuum and long-term low pressure flushing both caused the
highest hydraulic conductivities. The three Citrus species were not significantly different in their
vulnerability to embolism formation. In the future, a long-term study would allow for a better
understanding of plant hydraulic conductivity in drought-prone regions.
Introduction

5
Citrus plants are widely grown in tropical and Mediterranean climates. The genus
originates from South East Asia (Monrovia, 2010). Citrus farming was once the main fruit crop
in California (Cooley et al. 2009). Due to California’s Mediterranean climate, there is a high
demand for irrigation dedicated to citrus production, which could result in adverse economic and
possibly ecological effects. Orange County was once responsible for producing large amounts of
high quality Citrus. With the advancements being made in agricultural farming and urban sprawl,
many of these farming communities have moved to rural areas. Yet, a large-scale citrus
production still persists in the adjacent counties.
In order to lessen the economic and environmental impact and to increase water
conservation it is necessary to examine drought tolerance of citrus trees. More knowledge
regarding drought tolerance mechanisms of Citrus could lead to more efficient Citrus selection of
varieties for dry-summer climates and could possibly increase water conservation via improved
irrigation scheduling. Therefore, it is important to investigate the physiological mechanism of
citrus adaptation to a drought prone environment (Poggi et al. 2007).
One crucial physiological aspect of drought resistance in plants is their vulnerability to
form xylem embolisms (Hacke & Sperry 2001; Melcher et al. 2003; Poggi et al. 2007). A tree
under normal conditions utilizes negative xylem pressure to draw water from the ground into the
leaves, replenishing water lost to transpiration and transportation of nutrients. Under drought
conditions, a limitation of soil water availability can cause the formation of xylem embolisms, as
air is drawn into the xylem through small pores called pit membranes, a process known as air
seeding (Hacke & Sperry 2001). Embolisms impede the flow of water through the xylem and, if
not removed, can cause leaves to die off or lead to the death of the plant. The process of forming
embolisms, often referred to incorrectly as cavitation, occurs when negative pressure in the

6
xylem drops below a certain threshold. In many studies of plant species, a lower vulnerability
towards forming embolisms indicates a high

7
tolerance to water deficiency (Poggi et al. 2007). Yet, there are not many studies of xylem
vulnerability for fruit species and only one study of one citrus variety (Poggi et al. 2007).
Therefore, the purpose of this study was to document the xylem vulnerability of three Citrus
species: Citrus x. sinensis (Valencia orange), Citrus x paradisi (Star Ruby Red grapefruit), and
Citrus x. reticulata (Tangerin).
Xylem vulnerability can be measured by determining how much positive air pressure is
required to push air laterally into the xylem of detached stems (Sperry & Saliendra 1994). If
embolisms are formed in the xylem conduits, the hydraulic conductivity of the stems decreases.
A xylem vulnerability curve documents the percent loss of hydraulic conductivity (PLC) as a
function of pressure. Measuring xylem vulnerability curves of three citrus species allows gaining
insights into drought tolerance mechanisms in Citrus and may aid in developing improved
irrigation schedules and water conservation in California.
Three Citrus species were studied Citrus x sinensis (Valencia Orange), Citrus x paradisi
(Star Ruby Red Grapefruit), and Citrus x reticulata (Tangerin). Stems of these three species were
collected from the Fullerton Arboretum in California. The purpose of this study was to document
xylem vulnerability in three citrus species C. x sinensis, C. x paradisi, and C. x reticulata. To do
this, maximum hydraulic conductance of fully hydrated stems had to be measured as a baseline
for the measurements. A recent study has found that the standard method of hydrating stems by
flushing them with water under positive pressure may not always result in full hydration (Espino
& Schenk, 2010). Therefore, the first step of this research project was to determine a standard
method to hydrate Citrus stems. This method was then used for all measurements of xylem
vulnerability.

8
Four stem hydration methods were tested, including short term high pressure flushing
(T1), long term vacuum (T2), overnight low temperature, low pressure flushing (T3), and a
combination of high pressure flushing and short term vacuum (T4). Based on findings by Espino
& Schenk (2010), it was hypothesized that the best method to hydrate citrus stems would be to
submerge them in an under-water vacuum for 16 hours. The long term vacuum was predicted to
out-compete the other methods due to its ability to generate a long-lasting pressure differential
that allows for efficient diffusion of air molecules from embolized vessels through pit
membranes into water-filled cells.
Based on published research regarding irrigation demands (FAO 2010) and on differences in
fruit sizes, it was hypothesized that C. x paradisi would be the most vulnerable to embolism
formation. This was based on the assumption that a higher demand for water corresponded to a
higher vulnerability to embolism formation.
Materials and Methods
Collecting stems samples
Stems of three Citrus species: Citrus x sinensis (Valencia orange), Citrus x paradisis
(Star Ruby Red Grape Fruit), and Citrus x reticulata (Tangerin) were collected from the
Fullerton Arboretum, CA. Fresh stems with diameters between 2.3 mm and 5.0 mm and length
of 25 to 40 cm for stem hydration experiments and longer than 80 cm for xylem vulnerability
measurements were collected. Stems were cut and placed in a bucket of water which was
transported to the plants and H2O lab at Cal State Fullerton; green stems with minimal or no
branching were selected. In the lab, stems were submerged in a container containing de-ionized
(DI) water. For stem hydration experiments, stems were successively cut down to 15cm lengths

9
on both ends of the stems, cutting 1 cm at a time. The bark at the proximal end of the stems was
removed.
Hydraulic conductivity measurements
Hydraulic conductivity was measured as described by Espino and Schenk (2010). A
XYL’EM embolism meter (Bronkhorst, Montigny les Cormeilles, France) was used to measure
the flow rate of water through the stems. Stems were connected from their proximal ends to a
manifold, which was connected via tubing to the XYL’EM. Water used for measurements and
for hydrating stems was degassed using a membrane contactor degassing unit (Liqui-Cel mini-
module 1.7 x 5.5, Membrana, Charlotte, NC, USA) and passed through a 0.2 μm filter (model
Polycap AS 75, Whatman Inc., Piscataway, NJ). Initial flow rate of water moving into stems was
measured in the absence of a pressure differential, (Fo at 0 kPa). Stems were then measured
under a pressure differential of 6 kPa which was generated by raising the water supply relative to
the stems. Once the flow rate was stable, typically a duration of one to five minutes, the flow rate
was recorded as Fp. Hydraulic conductivity, k was determined as flow rate at 6 kPa, Fp minus
flow rate at 0 kPa pressure divided by the pressure differential, delta p: k = (Fp - Fo)/ delta p.
Specific hydraulic conductivity (ks) of a stem with certain length L and area A was
calculated as flow rate at 6 kPa minus flow rate at 0 kPa times length and divided by the
differential pressure and by area: ks= (Fp – Fo) L/ delta p A.
All hydrating conductivities reported in this study include a temperature correction to
20◦C to allow for changes in water viscosity with temperature.
Hydration treatments
Citrus stems of all three species were hydrated using four treatments to determine the
optimal hydration technique; treatment 1: Ten stems were flushed with DI, degassed water for 1

10
hour at 150kPa using the high-pressure chamber in the XYL’EM apparatus. This is the standard
hydration technique used in almost all xylem vulnerability studies (Espino & Schenk, 2010).
Treatment 2: Ten stems were submerged under DI water for 16 hours in a vacuum chamber
under 3 kPa absolute pressure, generated using a vacuum diaphragm pump (model DAA-
V715A-EB, Gast, Benton Harbor, MI, USA). This treatment was recommended by Espino and
Schenk (2010) as to avoid the bubble formation that can be caused by high pressure flushing.
Treatment 3: Ten stems were connected to one manifold and were flushed under low pressure
with DI, degassed water that was cooled to 12oC before entering the stems. Cooling increases gas
solubility in water (Mercury et al. 2003). Flushing was done for 16 hours at a rate of 10g hr- for
10 connected stems (i.e.,approximately 1 g hr- per stem). Treatment 4: Ten stems were
submerged under DI water in a vacuum chamber for 1 hour under 3 kPa absolute pressure. This
treatment would first remove air from open vessels at both ends of the stems, which could help to
avoid bubble formation under the subsequent flushing under high pressure.
Xylem Vulnerability Curves
Xylem vulnerability curves are measurements of the percent loss of hydraulic
conductance (PLC, relative to the maximum conductance, kmax) as a function of xylem
pressure.
The air injection method (Sperry and Saliendra, 1994; Meltcher et al., 2003) was used to
measure xylem vulnerability in Citrus. For Valencia Orange stems with 100 cm length, vessel
length of the stems were determined by gradually cutting 1 cm until the bubbles being released
were observed. The vessel length of an orange stem was 60 cm, as was the vessel length for
grapefruit and tangerine. Stems were notched 1 cm apart on two sides of the stem in order to
allow for air entry into the xylem. Notches were made by cutting at an angle directly into the

11
xylem, only into the section that was subjected to air pressure; then 65 cm to 100 cm long Citrus
stems were hydrated as described in treatment 2. After hydration, they were inserted into
cavitation chambers (PMS Instruments, Albany, OR) with both ends protruding. The cavitation
chambers were connected to a pressure chamber (model 1000, PMS instruments), which was
used to raise the air pressure inside the cavitation chambers successively from 0 to a maximum
of 10 MPa. After each successive increase in pressure, the pressure was held for 5 minutes then
reduced to 0.1 MPa for about 1 to 2 minutes (Sperry and Saliendra, 1994), which was then
reduced to an ambient pressure for measurements of hydraulic conductance using the XYL’EM
apparatus. Percent loss of conductance (PLC) was calculated as:
where a PLC of zero % indicates completely filled xylem and a PLC of 100% indicate a
completely embolized xylem. For all xylem vulnerability curves, kmax was defined as the ks
measured after application of 0.25 MPa pressure to correct for cavitation fatigue of the xylem
(Jacobsen et al. 2007).
Data Analysis
Hydration treatments were compared among the three Citrus species by ANOVA using
the statistical software SYSTAT (version 13, SYSTAT Software Inc., San Jose, CA). Xylem
vulnerability curves were analyzed using nonlinear regression of PLC vs. air pressure. Logistic
dose response curves were fitted to the data using the software TableCurve 2D (version 5.01,
SYSTAT Software Inc., San Jose, CA), and the pressures at which PLC reached 50% (Ψ50) was
determined from these regressions.

12
Results
The long-term vacuum treatment (T2), long-term low-pressure and cold flushing
treatment (T3) were equally effective in removing embolisms from Citrus vessels (Fig. 1).
However, the vacuum treatment was superior to high-pressure flushing, whether combined with
vacuum or not. The maximum specific hydraulic conductivity of stems after the long-term
vacuum treatment was on average about twice as high as achieved after one hour of high-
pressure flushing (Fig. 1), suggesting that the vacuum was much more effective for removing air
bubbles from stems. Maximum vessel lengths determined by the air injection method was 60 cm
for the Valencia orange, 60 cm for the Star Ruby Red Grapefruit, and 60 cm for the tangerine.
Based on these findings on the length of stems, 100 to 120 cm were used for xylem vulnerability
measurements were not significantly different in their vulnerability to xylem embolism
formation (Fig. 2), as differences between calculated Ψ50 values (= pressure at 50% PLC) were
not statistically significant. The Valencia orange, Ruby Red grapefruit and tangerine were not
significantly different in their vulnerability to xylem embolism formation (Fig. 2), as differences
between calculated Ψ50 values (= pressure at 50% PLC) were not statistically significant. This
was largely caused by the very high variability of conductivities in response to air injection into
the stems, as indicated by the large scatter of points on all graphs of Fig.2.

13

14

15
Discussion
As hypothesized the best method for hydrating Citrus stems was the sixteen hour
submerged vacuum treatment (T2). This method proved to be the most effective technique to
remove air from the xylem. This method stands out the most among other treatments in
providing a long-lasting partial pressure gradient between the gas phase in the vacuum chamber
and the gas phase in embolized vessels, which drives the movement of air molecules from
vessels through pit membranes into water-filled vessels and ultimately through the water’s
surface in the vacuum pump. As Espino and Schenk (2010) found previously, submergence
under vacuum effectively removed air from vessels, while high-pressure flushing was found to
be the least effective in removing air from vessels. High-pressure flushing, the widely used
standard method produced the lowest hydraulic conductivity, whereas, long-term vacuum and
long-term low pressure flushing both caused the highest hydraulic conductivities. As air removal
from wood occurs by way of diffusion, a relatively slow process, the longer the time, the more
efficient the removal of air will be. The second vacuum treatment, being the only one to be
significantly different from treatment 1 and, was also more feasible to implement as a standard
method than treatment 3, was adopted as the method of choice for hydrating Citrus stems.
The second hypothesis, that grapefruit would be the most vulnerable to embolism
formation than orange and tangerine, was not supported. The three Citrus species were not
significantly different in their vulnerability to embolism formation, although there appeared to be
a trend that showed that Valencia oranges were the most resistant species and the tangerines
were the most vulnerable. The inability to detect statistical differences between the
vulnerabilities of the three species was entirely due to large variances within species, resulting in
large standard errors (Fig. 2). The widely distributed data points suggests that the air injection

16
method, as applied, did not result in highly reliable and reproductive measurements of xylem
vulnerability. The xylem vulnerability curves of all three species did not go up to 100% PLC
over the range of pressures measured. Poggi et al. (2007) documented a pattern of xylem
cavitation for Clementine that was similar to that of the three Citrus species studied here. These
authors suggested that xylem conduits in Citrus may be divided into more vulnerable and less
vulnerable “compartments”, but could not find evidence for their species to support this
hypothesis.
For future works, it would be constructive to study the xylem’s anatomical structure of
the three Citrus species. The anatomical xylem analysis of the three Citrus species would
examine any difference in xylem straits that have been associated with vulnerability to embolism
formation in previous studies, including vessel diameters and the area of pits connecting vessels
(Hacke et al. 2009). Such measurements could be used to test the xylem compartments
hypothesis of Poggi et al. (2007), for example if vessels turned out to be present in two distinct
diameter classes. Also, the three Citrus species turned out to be anatomically similar, which
would explain why the three Citrus species were not statistically different in their vulnerability to
embolism formation.
As a follow up study on the findings of the research reported here, I would like to
conduct a study of the effects of Citrus root grafting on hydraulic traits of Citrus. Root grafting is
currently utilized within the agricultural field to increase the yield and control the longitudinal
growth of Citrus trees. Citrus farmers use the practice of grafting one species of Citrus trees to a
developed root system of a secondary citrus species to shorten the growth rate which it will take
a species to develop from a seedling to an adult tree capable of producing fruit. Another useful
and relevant study for the future could be on the effects of grafting on the formation of

17
embolisms within the trees’ roots and stems by comparing grafted and un-grafted trees. This
would allow me to test the hypothesis that root grafting leads to the development of more
embolism due to a hydraulic mismatch between roots and stems. Steppe et al. (2006) reported
evidence for such a hydraulic mismatch in orange trees by observing peculiar stomatal
oscillations that may have been due to an insufficient hydraulic conductance of the root system.
In the future, I would like to expand my study within the area of water usage for citrus
species and work with Citrus farmers to develop and test root irrigation systems either above or
below ground and generate improved irrigation schedules to allow for effective water
conservation within drought prone areas, which would ultimately lower costs in yield production
for the farmer. I believe that this would facilitate a thriving agricultural economy here in
California that could persist in the face of climate change. Moreover, lower cost production of
locally grown produce would be more attractive to local area consumers within a targeted
community and would allow for a competitive job growth of small agricultural farmers with the
potential of creating job opportunities in a given area.
In conclusion, this study has resulted in a new standard method to hydrate stems for
measurements of xylem vulnerability curves, which will now be tested on other plant species.
The research on xylem vulnerability in Citrus has remained inconclusive, but will continue to
develop better experimental protocols and increase the reliability of the technique.

18
Literature Cited
Cooley, H., Christian-Smith, J., Gleick, P. 2009. Sustaining California Agriculture in an
Uncertain Future. Pacific Institute, July Report, 1-81.
Espino, S. and Schenk, H. J. 2010. Mind the bubbles: Achieving stable measurements of
maximum hydraulic conductivity through woody plant samples. Journal of Experimental
Botany. Vol, 62, 1119-1132.
FAO 2010. Crop water information; Citrus. Available from
http://www.fao.org/nr/water/cropinfo_citrus.html.
Hacke, Uwe., Sperry, John. 2001. Functional and ecological xylem anatomy. Urban and Fischer
Verlag. Vol. 4/2, 97-115.
Hacke, U. G., A. L. Jacobsen, and R. B. Pratt. 2009. Xylem function of arid-land shrubs from
California, USA: an ecological and evolutionary analysis. Plant, Cell & Environment.
Vol. 32, 1324-1333.
Mercury, L., M. Azaroual, H. Zeyen, and Y. Tardy. 2003. Thermodynamic properties of
solutions in metastable systems under negative or positive pressures. Geochimica et
Cosmochimica Acta. Vol. 67, 1769-1785.
Monrovia [Internet]. [cited 2010 Oct 10]. Campbell Valencia Orange. Available from
http://www.monrovia.com/plant-catalog/plants/789/campbell-valencia -orange.php
Poggi, I., Polidori, J., Gandoin, J., Paolacci, V., Battini, M., Albertini, M., Ameglio, T., and
Cochard, H. 2007. Stomatal regulation and xylem cavitation in Clementine (Citrus

19
clementia Hort) under drought conditions. Journal of Horticultural Science &
Biotechnology, Vol 82, 845-858.
Sperry, J. S., Donnely, J. R., and Tyree, M. T. 1988. A Method for measuring hydraulic
conductivity and embolism in xylem. Plant, Cell and Environment. Vol 11, 35-40.
Sperry, J. S. and N. Z. Saliendra. 1994. Intra- and inter-plant variation in xylem cavitation in
Betula occidentalis. Plant, Cell and Environment 17:1233-1241.
Steppe, K., S. Dzikit, R. Lemeur, and J. R. Milford. 2006. Stomatal oscillations in orange trees
under natural climatic conditions. Annals of Botany, Vol. 92, 831-83.

20
Part II: Xylem Embolism Repair in Valencia Orange, Citrus x sinensis
Abstract
Plants normally transport water under negative pressure, which make their hydraulic
systems, the xylem, vulnerable to embolism formation if there is a water shortage in the soil. Air
can be drawn into xylem conduits through pit membranes, and the resulting embolisms impede
water flow, which can lead to branch and leaf mortality. Recent studies have shown that in some
species, embolisms form and are repaired diurnally during transpiration and even under negative
xylem pressure, but the mechanisms remain unknown. Little is known about the vulnerability of
fruit trees to embolism formation or their capability of embolism repair. If fruit trees grown in
semi-arid climates were capable of diurnal embolism repair then this could have implications for
efficient irrigation scheduling, as watering could potentially be timed to occur when it is most
beneficial for hydraulic recovery. The objective of this study was to test for diurnal embolism
formation and repair in Valencia Orange (Citrus x sinensis) grown in Southern California. Plant
water relations parameters, including leaf temperature, stomatal conductance, leaf water
potentials, stem hydraulic conductance, and percent loss of conductance (PLC) were monitored
over 24 hours periods at 4 hour intervals on nine young dwarf orange trees from fall to spring
2011/12.
Orange stems were highly embolized, with 40 to 60% loss of conductance, in fall and
winter, but there was no indication during these periods of embolism repair, even though the
trees were irrigated regularly. Nighttime leaf water potentials during these periods ranged
between -0.5 and -0.7 MPa; and no relationship was found during this time between water
potentials and the degree of xylem embolism. Findings from a warm day in spring 2012 were
substantially different. Xylem embolisms formed during the day to the same levels as previously

21
observed (40 to 60% PLC) and were repaired during the night down to less than 25% PLC at leaf
water potentials of about -0.3 to -0.6 MPa, corresponding to stem water potentials of about -0.1
to -0.4 MPa. In spring, there was a significant relationship between percent loss of conductance
and water potential, with a 50% PLC estimated at -1.1 MPa. The findings suggest that orange
trees may require high nocturnal temperatures and high water potentials for embolism repair.
The finding that about half of the vessels appear to be almost permanently embolized in dwarf
Valencia Orange trees is hypothesized to be a dwarfing root stock effect.

22
Introduction
The xylem is the transport system for water and minerals in plants. Trees under normal
conditions use negative pressure, which is generated by leaf transpiration, to draw water through
the xylem from the soil into the leaves to replenish water lost through transpiration. Under
drought conditions, soil water availability becomes limited, and air can be drawn into xylem
conduits through small pores connecting the conduits, which are called pit membranes. This
process is called air seeding (Hacke and Sperry 2001). The creation of air bubbles in the xylem
conduits is referred as xylem embolism formation (Hacke and Sperry 2001). Embolisms impede
the flow of water through the xylem, which can lead to leaf and branch mortality if the
embolisms are not removed. It is commonly observed that embolisms form during daytime
while the hydraulic system experiences maximum transpiration and the strongest negative
pressure (Bucci et al. 2003). Until quite recently, embolism repair was thought to take place
only in the absence of transpiration and while xylem tension is low. However, several recent
studies have indicated that embolisms can be repaired during transpiration and under negative
pressure (Clearwater and Goldstein 2005).
According to Yang and Tyree (1992), Steudle (2001), and Hacke and Sperry (2003),
embolisms can be repaired only if pressure in the xylem conduit is high enough to dissolve the
air bubbles into the xylem sap. The threshold pressure of the xylem sap required to dissolve
embolisms depends largely on the radius of the bubbles, following the Young-Laplace equation:
where pt is the threshold xylem sap pressure above which the gas bubble will collapse, P is the
gas pressure inside the bubble, T is the surface tension of water (0.00728 N m-1
, 0oC), and r is the
radius of curvature of the gas-water interface, i.e., the radius of the air bubble, which in the case

23
of a completely gas-filled xylem conduit would be the radius of the conduit. If the xylem is at,
or only slightly under, atmospheric pressure then air bubbles will slowly and spontaneously
dissolve. During transpiration, xylem pressure is almost always more negative than the “-2T/r”
threshold. Embolism repair under this condition is referred as refilling under tension (Hacke and
Sperry 2003), and the mechanisms for this process are largely unknown (Zwieniecki & Holbrook
2009)
Refilling of embolized vessels under negative pressure was documented for Laurus
nobilis in Hacke and Sperry’s study (2003). The calculated minimum average -2T/r limit was a
xylem pressure of ΨPX= - 0.1 MPa. L. nobilis plants were stressed to about -2.7 MPa and were
re-watered to a target xylem pressure (Ψw) of about -0.5 MPa. The ΨPX observed during
hydraulic recovery and embolism repair was approximately -0.66 MPa, which was below the
minimum -2T/r limit of ΨPX = -0.1 MPa. The result indicated that L. nobilis embolized vessels
were refilled under negative pressure. In a study of two Brazilian savanna tree species,
Schefflera macrocarpa and Caryocar brasiliense, Bucci et al. (2003) also found that embolized
vessels were refilling under tension. During afternoon to early evening, their hydraulic specific
conductivity dramatically increased as water potential increased as well. This pattern coincided
with a partial disappearance of starch in sheath cells surrounding the vascular bundles of petioles
in the afternoon. The authors hypothesized that hydrolysis of starch creates osmosic pressure that
draws water from the adjacent cells to refill embolized conduits (Bucci et al. 2003).
Because air bubbles cannot spontaneously dissolve in xylem conduits that are under
tension, plant physiologists have proposed a number of embolism repair scenarios that could
explain embolism repair under tension. These include: 1) cell membrane osmosis, 2) pit
membranes osmosis, 3) tissue pressure, 4) membrane asymmetry (Clearwater and Goldstein

24
2005), and 5) the liquid-gas membrane contactor theory (Schenk, unpublished manuscript).
Grace et al. (1993) proposed the cell membrane osmosis hypothesis, according to which solutes
are transported from the phloem into ray cells and from there through living parenchyma cells
into embolized conduits. This process could lower the osmotic pressure in the conduit, causing
water to flow from living cells into the conduit and refill it. Hacke and Sperry (2003) proposed a
variant of this hypothesis, termed the pit membrane osmosis hypothesis, to explain why high
concentrations of osmotic agents, such as salts or sugars, have not been observed in refilling
xylem. According to this hypothesis, living parenchyma cells may release solutes of high
molecular weight into conduits, which are unable to penetrate pit membranes, thereby remaining
confined to the refilling conduits, and not detectable in extracted sap.
Canny (1997) and Bucci et al. (2003) proposed a fundamentally different mechanisms,
termed the tissue pressure hypothesis. Conduit refilling according to this hypothesis is based on
the expansion of living xylem tissue. This exerts a compensating tissue pressure on cells xylem
that causes water flow into the refilling conduits. This tissue pressure and the refilling process
would be caused by increasing osmotic pressure inside living parenchyma cells and could be
caused by hydrolysis of starch. Pickard (2003) proposed a variant of the tissue pressure
hypothesis, which has been termed the membrane asymmetry hypothesis (Clearwater and
Goldstein 2005). According to this model, xylem tissue external to the xylem parenchyma
provides a source of solutes and water. During conduit refilling, solutes are loaded by membrane
transporters into xylem parenchyma, causing water uptake and increased turgor in these cells,
and this turgor pressure in turn would cause water to flow into the refilling conduit.
While all the of the proposed mechanisms discussed so far address the movement of
water into refilling conduit, the membrane contractor theory proposed by Schenk et al.

25
(unpublished manuscript) focuses on the mechanism of gas removal from conduits. According
to this theory, during nighttime transpiration, air bubbles from embolized xylem conduits diffuse
through pit membranes into the transpiration stream of functional conduits. This is possible
because xylem sap at night a cooler temperatures can dissolve large amounts of air.
Regardless of the mechanisms involved, the ability to repair embolisms may be very
important for survival, growth, and reproduction of plants during drought. Crop species,
including fruit trees, are likely to benefit from the ability to repair embolisms, as they could be
able to recover from drought after irrigation. Little information is available on embolism repair
in crops. To date, embolism repair under tension has been found in grapevine (Brodersen et al.
2010) and rice (Stiller et al. 2005).
Little is known about the drought tolerance of Citrus species. According to Spiegel-Roy
and Goldschmidt (1996) although the wild type of Citrus is of unknown origin, most citrus
commercially grown nowadays are hybrids and originated from South East Asia, indigenous to
China and India. According to Scora (1975), Citrus x. sinensis (sweet orange), the study species,
is a hybrid of C. reticulata (mandarins) and C. grandis (pomelo). Citrus became widely used in
the region as food, for its medicinal properties (used as herbal medicine), as ornamentals, for
fragrance, and as an antidote against poison. Through trade and missionaries citrus spread
through other parts of Asia and was introduced throughout the world to Europe, Africa, and not
until the early 16th
century, to America. Citrus was first introduced in Florida, then Arizona, then
California in 1769. Climatology of citrus encompasses a wide range north 40 degree (Corsica,
Japan) to south 40 degree (New Zealand). Citrus can be found near the equator in hot and humid
climates, in warm subtropical regions, and in cooler-maritime climates. The reproductive cycle
of Citrus includes blooming in spring, bearing fruit in summer, reaching maturity of fruit in fall.

26
A sufficient long and warm summer is required for citrus fruit to grow and reach maturity.
Growing citrus in Mediterranean climate with long and dry summers requires irrigation to
maintain satisfactory tree growth and fruit development.
Although Citrus did not originate from a Mediterranean climate, the plants possess a
number of apparent drought adaptations. Leaves are leathery, have thick, waxy cuticles that
prevent water loss, and have low transpiration rates (Fererest et al. 1979). The fruits are coated
with thick wax and an oily peel (Spiegel-Roy and Goldschmidt, 1996). Fereres et at. (1979)
indicated that citrus crops can withstand considerable water stress. For instance Valencia orange
tree can tolerate very low negative water pressure of – 6.6 MPa and rapidly recover within 2
hours after the irrigation started. In contrast, Poggi et al. (2007) found Citrus x clementine to be
quite vulnerable to embolism formation. My preliminary result investigating the vulnerability of
three citrus species to embolism indicated that Valencia orange, star ruby red grapefruit, and
tangerine are equally vulnerable to embolism. No study to date has investigated embolism repair
in any citrus species. The objective of this study will be to test for embolism formation and
repair in Valencia orange, one of the most widely grown varieties of sweet orange.
Stem hydraulic conductivity, leaf stomatal conductance, and branch pressure potential
will be measured over 24 hour periods. The diurnal course of stem hydraulic conductivity will
indicate whether or not embolism formation and repair occurs. The physiological conditions
under which embolisms form and/or a repaired will be documented by measurements of leaf
stomatal conductance and branch pressure potentials.
Methods
Valencia orange trees were sampled on the South side of Langsdorf Hall on the campus
of Cal State Fullerton in Fullerton, California. Tests for embolism repair were conducted during

27
three twenty-four-hour periods on November 21-22, February 25 - 26, and April 21-22,
following a protocol developed by Hessom et al. (unpublished manuscript).
On each date, nine individual Valencia orange trees were sampled in 4 hour increments
over a 24 hour period. At each time interval, each tree was measured for branch pressure
potential, stomatal conductance, and stem hydraulic conductance. To measure stomatal
conductance, a leaf porometer (Decagon Devices Inc. Leaf Porometer: Version 4.0, Pullman,
WA, USA) was used on two leaves of the same branch sampled for leaf pressure potentials and
stem hydraulic conductance. To measure leaf pressure potentials, a branch tip with leaves from
each tree was cut, and leaf pressure potentials were measured using a pressure chamber (model
1000, PMS Instruments, Eugene, OR, USA). The branches were then cut at about 50 cm long
from the tip and quickly transported to the lab.
Stem hydraulic conductance was measured on 15 cm-long stem segments using a
XYL’EM Embolism Meter (Instrutec, France. In the lab, the 50 cm branches were submerged in
a container with deionized (DI) water. Stems were cut down to 15 cm length from both ends of
the stems by cutting 1 cm at a time. The bark at the proximal end of the stems was removed.
After stem hydraulic conductivity measurement, the stem segments were submerged in water-
filled test tubes and were rehydrated under vacuum for 20 hours. After rehydration, maximum
hydraulic conductance was measured for all stem samples using the XYL’EM Embolism Meter.
Hydraulic conductivity was measured as described by Espino and Schenk (2010). A
XYL’EM embolism meter was used to measure the flow rate of water through the stems
(Bronkhorst, Montigny les Cormeilles, France). Stems were connected from their proximal ends
to a manifold, which was connected via tubing to the XYL’EM . Water used for measurements
and for hydrating stems was degassed using a membrane contactor degassing unit (Liqui-Cel

28
mini-module 1.7 x 5.5, Membrana, Charlotte, NC, USA) and passed through a 0.2 µm (model
Polycap AS 75, Whatman Inc., Piscataway, NJ). The initial flow rate of water moving into
stems was measured in the absence of a pressure differential, (Fo at 0 kPa) to account for
osmotically-driven flow. Stems were then measured under a pressure differential of 6 kPa which
was generated by raising the water supply relative to the stems. Once the flow rate was stable,
typically after one to five minutes, the flow rate was recorded as Fp. Hydraulic conductivity, k
was determined as flow rate at 6 kPa, Fp, minus flow rate at 0 kPa pressure, divided by the
pressure differential, p: k = (Fp- Fo)/ p.
Specific hydraulic conductivity (ks) of a stem with a length (L) and conducting area (A)
was determined as flow rate at 6 kPa minus flow rate at 0 kPa times length divided by the
pressure differential and conducting area: ks= (Fp – Fo) L/ p A.
All hydraulic conductivities reported in this paper include a temperature correction to
20oC to allow for changes of water viscosity with temperature. In this study, percent of loss of
conductivity was referred as percent of embolism.
Results
As hypothesized, Valencia orange was quite vulnverabe to embolism formation but was
able to repair embolisms at high water potential and low transpiration. Leaf water potential
showed a steady increase from fall 2011 to spring 2012. As the seaons changed, the temperature
increased causing an increase in stomatal conductance. Percent of embolisms were consistently
high in the Fall 2011 and early spring 2012; however, nighttime embolisms significantly dropped
in April. As leaf water potential increased, the percent of native embolism decreased.

29
In November 2011, leaf water potential during daytime was significantly lower than
during nighttime. During the day, it reached its lowest leaf water potential of -2.3 MPa while
during night, its highest leaf water potential was at -0.5 MPa (Fig. 1 A). Stomatal conductance
between daytime and nighttime also showed a great difference. The diurnal stomatal
conductance was above 150 mmol m-2
s-1
while the nocturnal stomatal conductance was very
close to 0 mmol m-2
s-1
(Fig. 1 B). Despite these great variations in leaf water potential and
stomatal conductance, percent of embolism showed minimal fluctuation over the course of the 24
hour measurement period and ranged between 40 – 60 % (Fig. 1 C).
Fig.1: Plant water relations of Citrus x sinensis over a 24 hr period on 21-22 November 2011. The
period from sunset to sunrise is marked by the shaded area. A. Leaf water potentials. B. Stomatal
conductance. C. Percent loss of conductance due to embolisms. All error bars are ± SE.

30
In February 2012, the leaf water potential increased; and there was minimal fluctuation of the
stem water potential over the 24 hr experimental period. The stem water potential reached its
lowest at -1.4 MPa which occurred at noon on February 26th
(Fig. 2 A). The nocturnal stomatal
conductance observed in February 2012 was higher than that of November 2011. The percent of
embolisms measured in February 2012 was similar to that of November 2011, ranging from 40
% to 60 % (Fig. 1 C and 2 C). On 25th
February 2012, the peak of percent loss of conductivity
occurred in the afternoon while the stem water potential was high. The percent loss of
conductance dropped from 55% to around 40% and remained at 40% from 9 a. m. to noon the
next day (Fig. 2 C).
Fig. 2: Plant water relations of Citrus x sinensis over a 24 hr period on 25- 26 February 2011.
The period from sunset to sunrise is marked by the shaded area. A. Leaf water potentials. B.
Stomatal conductance. C. Percent loss of conductance due to embolisms. All error bars are ± SE.

31
In April 2012, leaf water potential was highest compared to previous month, especially
the nighttime leaf water potential reached -0.3 MPa (Fig. 3 A). The temperature also increased
(Fig. 3 B). The stomatal conductance in April resembled that of February. The diurnal percent
of embolism still remained high; however nocturnal percent of embolism dropped to 25%, which
was denoted with the asterisks to indicate a significant drop in percent of embolisms.
Fig. 3: Plant water relations of Citrus x sinensis over a 24 hr period on 21 - 22 April 2011. The
period from sunset to sunrise is marked by the shaded area. A. Leaf water potentials. B.
Stomatal conductance. C. Percent loss of conductance due to embolisms. All error bars are ± SE.
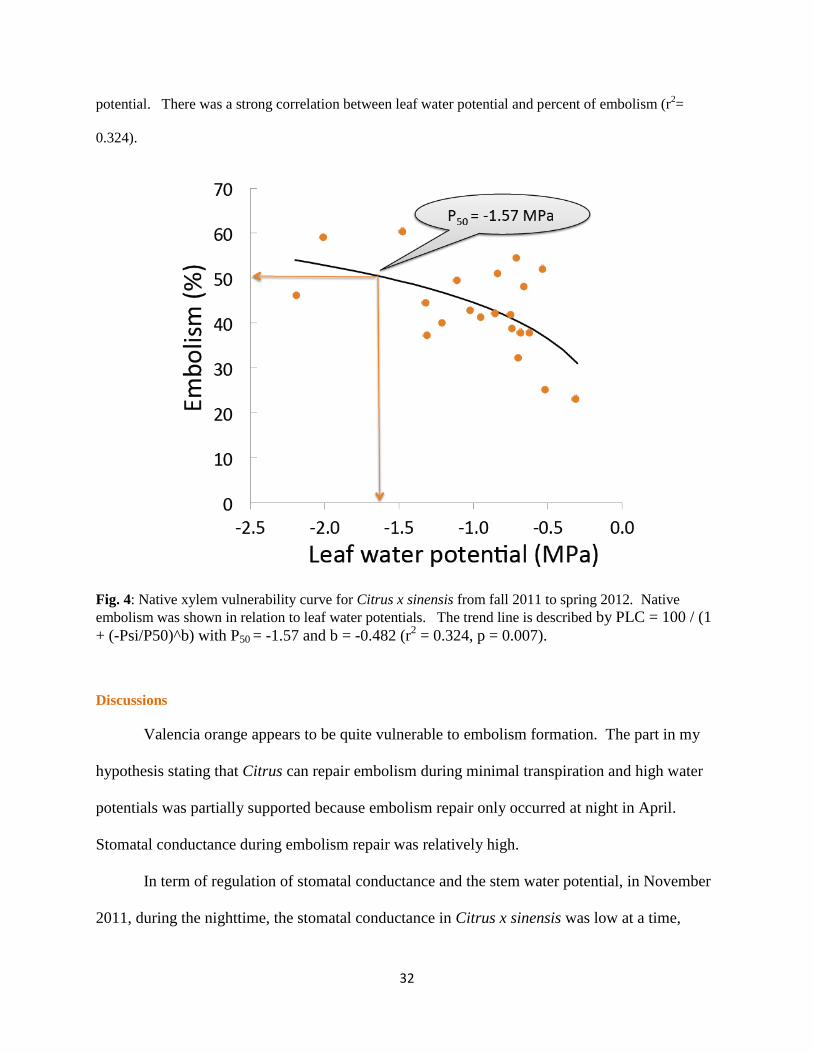
Most importantly, the native xylem vulnerability curve showed Citrus x sinensis trees are quite
vulnerable to embolism formation. Fifty percent of embolism was at -1.57 MPa, relative high water
32
potential. There was a strong correlation between leaf water potential and percent of embolism (r2=
0.324).
Fig. 4: Native xylem vulnerability curve for Citrus x sinensis from fall 2011 to spring 2012. Native
embolism was shown in relation to leaf water potentials. The trend line is described by PLC = 100 / (1
+ (-Psi/P50)^b) with P50 = -1.57 and b = -0.482 (r2 = 0.324, p = 0.007).
Discussions
Valencia orange appears to be quite vulnerable to embolism formation. The part in my
hypothesis stating that Citrus can repair embolism during minimal transpiration and high water
potentials was partially supported because embolism repair only occurred at night in April.
Stomatal conductance during embolism repair was relatively high.
In term of regulation of stomatal conductance and the stem water potential, in November
2011, during the nighttime, the stomatal conductance in Citrus x sinensis was low at a time,

33
when the stem water potential was high. Schenk et al (2011, unpublished manuscripts) also
found the same pattern of lower stomatal conductance and higher stem water potential at night
for Encelia farinosa. Surprisingly, nocturnal stomatal conductance in February and April was
higher than in November. The differences in stomatal conductance patterns between November
2011, February and April 2012 suggest that the plants are capable of regulating their stomata,
perhaps in response to seasonal temperature changes and vapor pressure deficit.
In November 2011, the stem water potential of Citrus x sinensis showed the same pattern
as the stem water potential observed in Malosma laurina in February 2009 (Hessom, 2010).
Stem water potential was low while stomata opened and high when stomata closed. However,
the stem water potential of Citrus x sinensis in February and April 2012 remained steady
throughout the 24 hour experimental period. In November 2011, the leaf water potential was
higher at night than during the day. However the percent of embolisms remained high between
40%, despite the fact that the trees were irrigated twice a week and rained on the previous day of
the measurements.
In February 2012, percent of embolism was same as that of November even though leaf
water potential in February was higher than that in November. Approximately half of the vessels
in Citrus x sinensis appeared to be permanently non-functional, regardless of irrigation. 50%
percent loss of conductivity was approximately at -0.7 MPa to -0.8 MPa, which was quite
vulnerable compared to many other woody species (Maherali et al. 2004). In contrast, Poggi et
al. (2007) found Citrus x clementina to be much less vulnerable to embolism formation, with less
than 50% PLC at -4 MPa stem water potential. The consistently high percent of embolisms over
the two 24-hour experiments suggested that Citrus x sinensis trees were vulnerable to embolism
but did not repair during fall 2011 and early spring 2012.

34
However significant nighttime embolism repair occurred in April, which was differed
from previous dates in having higher nocturnal leaf water potentials (≥-0.5 MPa) and nocturnal
leaf temperature (≥20oC). Stomatal conductance during embolism repair was high. The findings
suggest that Citrus trees as a tropical species require high nocturnal temperature and nocturnal
water potentials to successfully repair embolism. The findings suggest that irrigation for Citrus
farms during warm nights may be more effective than during cold nights.
The knowledge about embolism formation and repair in Citrus trees could improve
irrigation scheduling. My potential graduate research project will focus on minimize water cost
while maximize Citrus fruit production. Another 24 hour measurements of plant hydraulic
conductance conducted during the hottest month in summer would be extremely beneficial
further confirm the results that Citrus repair embolisms during warmers seasons. Before
conducting these measurements, irrigation will be asked to turned off for 2 weeks to see if the
Citrus trees can still effectively repair embolisms when water supply is limited.
Acknowledgements
This research was supported by the Southern California Ecosystems Research Program, funded
by the National Science Foundation DBI – 1041203 (W. Hoese and D. Sandquist, PIs). The authors
thank Susana Espino, Miguel Macias, Kenny Duong, Matthew Laine, Sarah Taylor, Diane Tran, An Ly,
Donald Quick and Robert Wieber for helping with the measurements.

35
Literature Cited
Bucci, S. J., Scholz, F. G., Meinzer, F. C., and Sternberg, L. DA S. L. 2003 Dynamic changes in
hydraulic conductivity in petioles of two savanna tree species: factors and mechanism
contributing to the refilling of embolized vessels. Plant, Cell, and Environment 26, 1633-
1645.
Brodersen, C. R., McElrone, A. J., Choat, B., Matthews, M. A., & Shackel, K. A. (2010). The
dynamics of embolism repair in xylem: in vivo visualizations using High Resolution
Computed Tomography. Plant Physiology, 154(3), 1088-1095. doi:
10.1104/pp.110.162396
Canny, M. J. 1997. Vessel contents during transpiration: Embolisms and refilling. American J
ournal of Botany, 84, 1223-1230.
Clearwater, M. J., & Goldstein, G. 2005. Embolism repair and long distance water transport. In
N. M. Holbrook & M. A. Zwieniecki (Eds.), Vascular transport in plants (pp. 375-399).
Amsterdam: Elsevier Academic Press.
Dixon, H; Joly (1894). "On the ascent of sap". Ann. Bot. 8: 468–470.
Fereres, E.,Cruz-Romero,G.,HoffmanOFFMAN,G. J. and Rawlins, R. L.(1979). Recovery of
orange trees following severe water stress. Journal of Applied Ecology, 16, 833–842.
Grace, J. 1993. Refilling of embolized xylem. In Water Transport in Plants Under Climate
Stress. Cambridge University Press 51-62.
Hacke, U.G. and Sperry, John. 2001. Functional and ecological xylem anatomy. Urban and
Fischer Verlag. Vol. 4/2, 97-115.
Hacke, U. G. and Sperry, J. S. 2003. Limits to xylem refilling under negative pressure in Laurus
nobilis and Acer negundo. Plant Cell Environment 26: 303-311.

36
Hessom, E. 2010 Xylem Embolism Repair in Two Chaparral Shrubs, Malosma laurina and
Ceanothus crassifolius. Undergraduate Thesis, California State University of Fullerton,
Fullerton.
Maherali, H., W. T. Pockman, and R. B. Jackson. 2004. Adaptive variation in the vulnerability of
woody plants to xylem cavitation. Ecology 85:2184-2199.
Nicolosi, E., Deng, Z. N., Gentile, A., La Malfa, S., Continella, G., & Tribulato, E. (2000).
Citrus phylogeny and genetic origin of important species as investigated by molecular
markers. TAG Theoretical and Applied Genetics, 100(8), 1155-1166. doi:
10.1007/s001220051419.
Pickard, W. F. 2003. The riddle of root pressure. I. Putting Maxwell’s demon to rest. Functional
Plant Biology 30: 288-297.
Poggi, I., Polidori, J., Gandoin, J., Paolacci, V., Battini, M., Albertini, M., Ameglio, T., and
Cochard, H. 2007. Stomatal regulation and xylem cavitation in Clementine (Citrus clementia
Hort) under drought conditions. Journal of Horticultural Science & Biotechnology, Vol
82, 845-858.
Salleo, S., Lo Gullo, M. A., de Paoli, D., & Zippo, M. (1996). Xylem recovery from cavitation-
induced embolism in young plants of Laurus nobilis: a possible mechanism. New
Phytologist, 132, 47-56.
Schenk, H. J., Espino S., Martinez-Cabrera, H., Kana, T. M., and Jones, C. S. 2011 Nighttime
transpiration and embolism repair in plants: Two mysteries explain each other.
(Unpublished manuscript).
Scora, R. W. (1975). On the history and origin of Citrus. Bulletin of the Torrey Botanical Club,
102(6), 369-375.

37
Spiegel-Roy P. & Goldschmidt E.E. (1996) Biology of Citrus. Cambridge University Press,
Cambridge.
Steudle, E. 2001. The cohension-tension mechanism and the acquisition of water by plant roots.
Annu Rev Plant Physiol Plant Mol Biol 52: 847-875.
Stiller, V., Sperry, J. S., & Lafitte, R. (2005). Embolized conduits of rice (Oryza sativa, Poaceae)
refill despite negative xylem pressure. American Journal of Botany, 92(12), 1970-1974.
Webber, H. J. (1967). History and development of the citrus industry. In W. Reuther, H. J.
Webber & L. D. Batchelor (Eds.), The citrus industry (Vol. Vol. 1, pp. 1-39). Berkeley, CA:
University of California Press.
Wheeler, D. T., Strook, D. A. 2008. The transpiration of water at negative pressures in a
synthetic tree. Nature. 455, 208-212.
Yang, S. and Tyree, M. T. 1992. A theoretical-model of hydraulic conductivity recovery from e
embolism with comparison to experimental-data on Acer saccharum. Plant Cell
Environment 15: 633-643.
Zufferey, V., Cochard, H., Ameglio, T., Spring, J.-L., and Viret. O. 2011 Diurnal cycles of
embolism formation and repair in petioles of grapevine (Vitis vinifera cv. Chasselas).
Journal of Experimental Botany 62, 3885-3894.
Zwieniecki, M. A. and N. M. Holbrook. 2009. Confronting Maxwell's demon: biophysics of
xylem embolism repair. Trends in Plant Science 14:530-534.