Embed Size (px)
Citation preview

Diversity, Host Utilization and Ecological Niche of Fruit Flies (Diptera: Tephritidae) in Uganda
ISABIRYE BRIAN ERIPHAZ, [email protected]

2
Acknowledgements

3
Background… Agriculture…Global and Uganda
Employs 45% of the working global population and over 80% in most parts of Africa and Asia!
Uganda•Export:
90%
•Employment:80%
•People living in rural areas : 85%
•Source of raw materials…….

5
Trade: Dynamic and highly vibrant!
Center on Globalization, Governance & Competitiveness, Duke University, 2011

6
Key Players in the fruit trade
• Global import of fruits hit US$4.3 million tons in the year 2010. In which 87% or 3.8 million tons were imported by developed countries.
• The US and the EU market represents 70% of global import of tropical fruits.
• The EU is the largest tropical fruit importer with the major consumer of France and the main transshipment port of the Netherlands.
• The US and Japan, Canada and Hong Kong are also large importers (USDA, 2007).
7
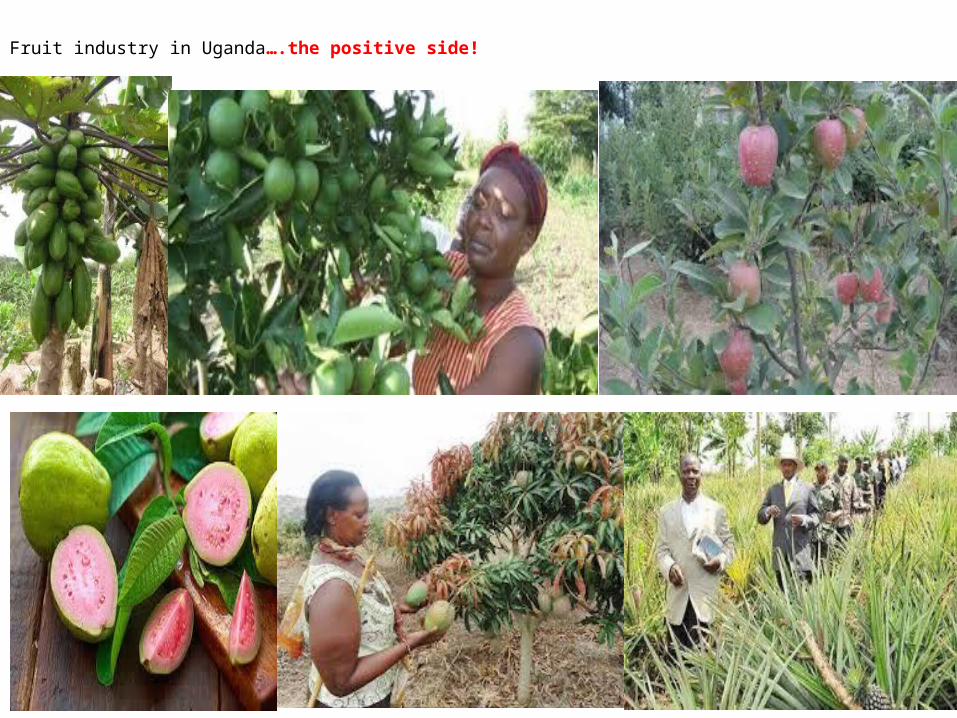
Fruit industry in Uganda….the positive side!

8
Bad news for most fruit producers!
Y2003 Y2004 Y2005 Y2006 Y2007 Y2008 Y2009 Y20100
1000
2000
3000
4000
5000
6000
7000
8000
9000
0
1000000
2000000
3000000
4000000
5000000
6000000
Volume (Tonnes) Value (US$)
Period (Years)
Volu
me
(Ton
nes)
(Source: UNEPB)
Valu
e (U
S$)

9
Common Challenge: Tephritid FF
Life Cycle
10
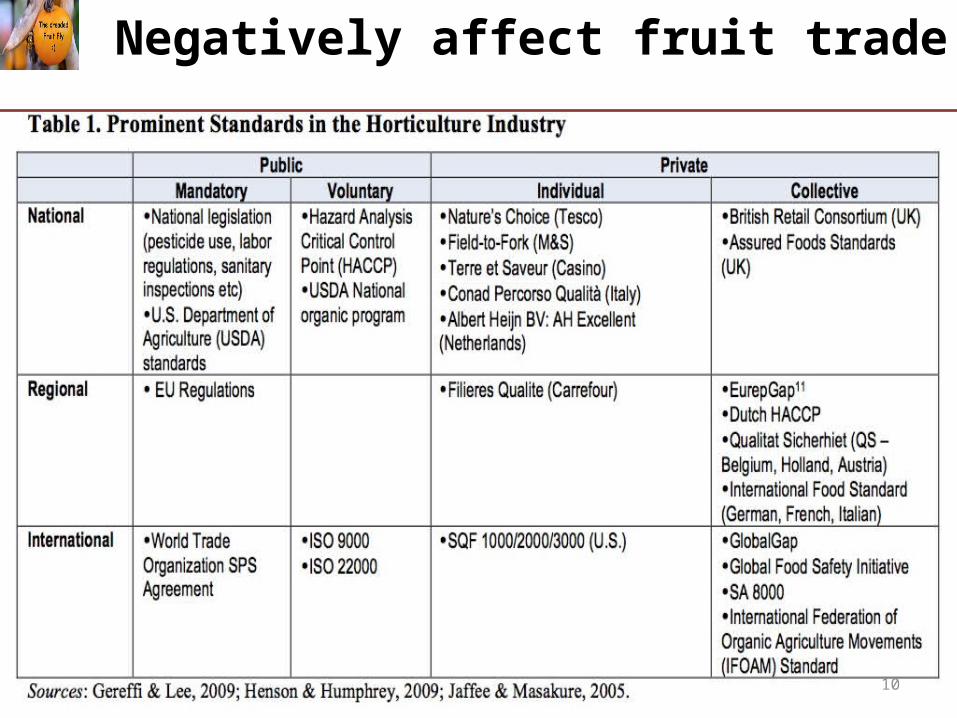
Negatively affect fruit trade

11
Problem/ Motivation…• Fruit flies cause about 40% fruit loss in Africa, and about 73% in Uganda
• Design of IPM strategies for fruit flies requires knowledge of their biology.
• Was limited to Nakasinga, 2002; Nemeye, 2005; Okullokwany, 2006.
• It is not clear how:
• Diversity has been shaped by hosts, distribution and envital variability.
• Highly cryptic and inter-intra-specific morphological variation (Clarke
et al., 2005; Drew et al., 2008) among Bactrocera spp. turns out.
• Will change in climate alter the suitability and distribution of species?

12
Objectives and HypothesisesMain ObjectiveTo describe the diversity, host utilization and ecological niche of major tephritid fruit flies in Uganda.
Specific Objective1. Determine the species diversity of fruit flies across selected agro ecological zones 2. Assess fruit fly host utilization in the different agro ecological zones. 3. Characterise the morphometric variability of the most important fruit fly species
among hosts and mango growing zones4. Determine the current and potential future spatial distribution of the major Tephritid
fruit fly species in Uganda.
Hypotheses1. There is no significant difference in the diversity of fruit flies in the different ecological
zones in Uganda.2. There is no significant difference in fruit fly host utilisation patterns in the different
ecological zones and among host types in Uganda.3. There is no significant morphometric heterogeneity among B. invadens populations
infesting different hosts in the different ecological zones in Uganda.4. Fruit fly species’ current and future distribution and ecological niches is random across
the different ecological zones in Uganda
13
Study IV: Morphometric (intra species) Diversity
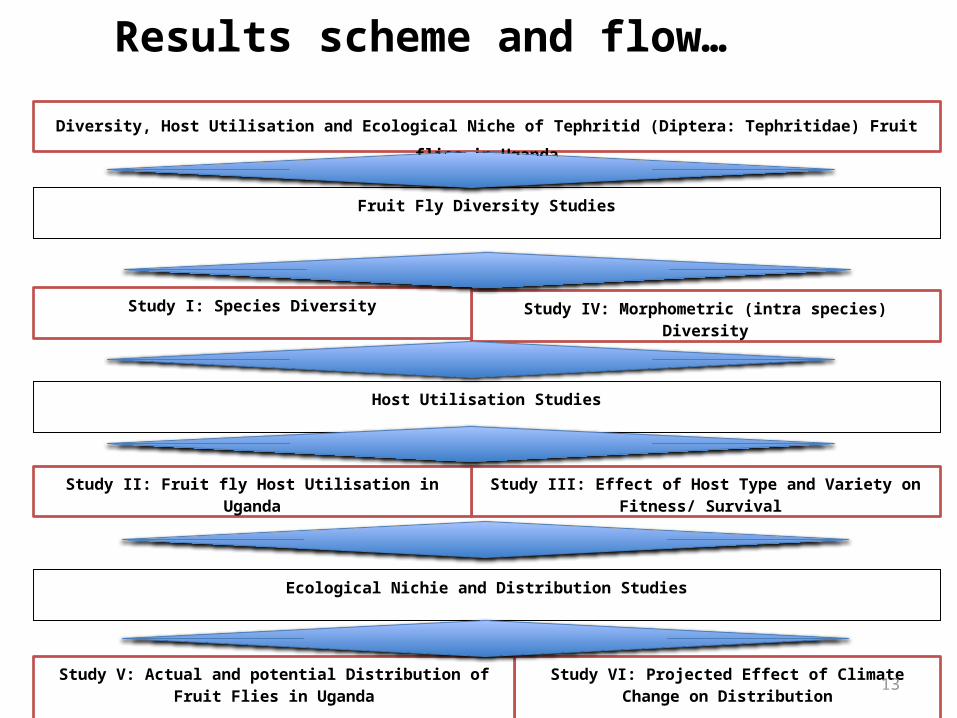
Diversity, Host Utilisation and Ecological Niche of Tephritid (Diptera: Tephritidae) Fruit flies in Uganda
Ecological Nichie and Distribution Studies
Fruit Fly Diversity Studies
Study I: Species Diversity
Study III: Effect of Host Type and Variety on Fitness/ Survival Study II: Fruit fly Host Utilisation in Uganda
Study VI: Projected Effect of Climate Change on Distribution
Study V: Actual and potential Distribution of Fruit Flies in Uganda
Host Utilisation Studies
Results scheme and flow…

14
STUDY ONE: INTER SPECIES DIVERSITY
“When you have seen one ant, one bird, one tree, you have not seen them all” E. O. Wilson

15
Introduction• FF are key pests of several fruit crops (Ekesi et al., 2006; Mayamba et al.,
2015)
• Yield losses can exceed 80% (Ekesi et al., 2006; Mayamba et al., 2014)
• Correct identification is key in sustainable management (Jang et al., 2003)
• Regional efforts (Mwatawala et al., 2006; 2009; Rwomushana et al., 2008; Geurts et al., 2012), but in Uganda (Nakasinga, 2002, Okullokwany, 2006)
• This study set out to: 1. Identify the fruit fly species present in the country, and2. Assess the fruit fly community structure across three mango
production AEZs
STUDY ONE: INTER SPECIES DIVERSITY

16
Mat. and MethodsWestern Medium High Altitude Farmlands (WMHF), Lake Victoria Crescent (LVC) and Northern Moist Farmlands (NMF) (Wortman and Eledu, 1993)
STUDY ONE: INTER SPECIES DIVERSITY
%[
%[%[
%[
%[%[
%[%[
%[
%[
%[
%[
%[
%[
%[
%[%[
%[
LIRA
APAC
GULU
OYAM
KASESE
AMURU
RAKAI
SOROTI
WAKISO
MUKONO
PADER
MITYANAKABAROLE
MPIGI
MASAKA
IGANGA
AGAGO
MAYUGE
IBANDA
BUSIA
RUKUNGIRI
DOKOLO
RUBIRIZI
KIBINGO
TORORO
AMOLOTAR
LAMWO
KAMULI
KIRYANDONGO
NWOYA
NTUNGAMO
BUNDIBUGYO
KALUNGU
KIYUNGA
KAYUNGA
KABALE
AMURIA
MBARARA
KIBOGA
BUDAKA
MASINDI
200 0 200 400 Kilometers
N
LakesDistrict Boundary
Lake Victoria Crescent & Mbale FarmlandsNorthern Moist FarmlandsWestern Medium-High Farmlands
Major Sites Sampled%[ Minor Sites Sampled%[
KEY
Agro Ecological Zones (AEZ)
• Trapping with baited traps• Methyl eugenol • Trimedlure • Torula yeast• Cuelure • Terpenyl Acetate
Collecting mango fruits and other fruits and incubate them to assess fruit fly infestation.

17
Results 1Fruit fly community composition
STUDY ONE: INTER SPECIES DIVERSITY
5 10 15 20
02
46
81
0
sites
sp
ecie
s r
ich
ne
ss
LVC
LVC
NMF
NMF
WMHF
WMHF
LVCNMFWMHF
2 4 6 8 10
1
e+
01
1
e+
02
1
e+
03
1
e+
04
1
e+
05
species rank
ab
un
da
nce
LVCNMFWMHF
LVCNMFWMHF
0 0.25 0.5 1 2 4 8 Inf
0.0
0.5
1.0
1.5
2.0
alpha
H-a
lpha
LVC
LVC
NMF
NMF
WMHF
WMHF
LVCNMFWMHF
Bactro
cera i
nvaden
s
B. curcu
bitae
C. anonae
C. cosyr
a
C. capita
ta
C. fasci
ventri
s
C. rosa
Trithuriu
m coffea
e
Dacus b
ivitatt
us
D.ciliat
us
0
20,000
40,000
60,000
80,000
100,000
120,000
140,000
119245
152 339 75 66 5 10 312 956 478
122112
3 401 3 62 0 0 32 2 1
122982
4 272 2 69 12 0 8 485 244
LVC NMF WMHF

18
Results 2Fruit fly community structure
STUDY ONE: INTER SPECIES DIVERSITY
100
22
43 2243 85
22
6485
0.55
0.6
0.65
0.7
0.75
0.8
0.85
0.9
0.95
Sim
ilarit
y
T.co
ffeae
_
C.co
syra
_
B.cu
rcub
itae_
C.ro
sa_
D.biv
itattu
s
D.cil
iatus
_
C.fas
civen
tris_
C.ca
pitata
_
B.inv
ande
ns_
C.an
onae
LVCNMFWMHF
-22-6 5
B.invandens
C.Anonae
C.cosyra
C.capitata
C. fasciventris
C. rosa
T.coffeae
B.curcubitae
D.bivitattusD.ciliatus
-1
1-1 1
P < 0.000

19
Results 3Displacement by B. invadens
STUDY ONE: INTER SPECIES DIVERSITY
0 1 2 3 4 5 6 7 8 9
Rank
-0.8
0
0.8
1.6
2.4
3.2
4
4.8
5.6
log
Abu
ndan
ce
0 1 2 3 4 5 6 7 8 9 10
Rank
0
0.6
1.2
1.8
2.4
3
3.6
4.2
4.8
5.4
6
log
Abu
ndan
ce
0 1 2 3 4 5 6 7 8
Rank
-0.8
0
0.8
1.6
2.4
3.2
4
4.8
5.6
log
Abu
ndan
ce
Conclusion1. At least 10 species in the country but B. invadens is the most widely distributed.
2. Significant diff in richness and abundance of fruit flies but not in evenness and diversity of fruit flies across zones.
3. Differences across zones may be in turn attributed to the inherent environmental and host plant composition.

20
Study IV: Morphometric (intra species) Diversity
Diversity, Host Utilisation and Ecological Niche of Tephritid (Diptera: Tephritidae) Fruit flies in Uganda
Ecological Nichie and Distribution Studies
Fruit Fly Diversity Studies
Study I: Species Diversity
Study III: Effect of Host Type and Variety on Fitness/ Survival Study II: Fruit fly Host Utilisation in Uganda
Study VI: Projected Effect of Climate Change on Distribution
Study V: Actual and potential Distribution of Fruit Flies in Uganda
Host Utilisation Studies
Results scheme and flow…

21
STUDY TWO: HOST USE

22
Introduction• Fruit industry provides livelihoods World-wide (Lux et al., 2003; Ekesi and Billah,
2006).
• FF cause variable losses (Lux et al., 2003; Vayssie`res et al., 2005).
• Limited host status knowledge in Uganda, save for regional studies (De Meyer et al., 2002; Copeland et al., 2002; Rwomushana et al., 2008).
• Makes design of mgt options hard (Mwatawala et al., 2009a).
• This study: – To profile the host range of the main fruit fly pests in the three main mango agro
ecological zones; and
– Determine the susceptibility of selected fruits and mango varieties grown to the various fruit fly pests in the country
STUDY TWO: HOST USE

23
Materials and Methods
Three major mango AEZs: WMHF, LVC and NMF (Wortman and Eledu, 1993)
STUDY TWO: HOST USE
1. Intensive collection of commercial and non-commercial fruit hosts.
2. Selected important fruits and mango cultivars in each zone at random sites.
3. Fruits included sweet orange, tropical almonds, avocado, guava and mango.
4. The mango cultivars: Apple Mango, Biire, Boribo, Dodo, Glen, Kagogwa, Kate, Keitt, Kent, Tommy Akinson, Palvin and Zillatte.
5. The 12 cultivars were classified according to their maturity seasonality into early, mid and late maturing cultivars (Ambele et al., 2012).
6. Fruits were transported to the rearing unit at the NARL (Copeland et al. 2002).

24
Results 1Fruit fly Host Range
• 38 fruit species, from 30 genera in 18 plant families were sampled.
• Among these, 633 (35.0%) samples were positive for fruit fly infestation.
• B. invadens was the dominant species: recorded in 29 out of the 38 plant species, while out of the 633 positive samples, 483 (76.3%) were due to B. invadens.
• Host infestation incidence for the rest of the fruit fly species was low
STUDY TWO: HOST USE
Anno
nace
ae
Sola
nace
ae
Ruta
ceae
Anac
ardi
acea
e
Myr
tace
ae
Cucu
rbita
ceae
Mor
acea
e
Sapo
tace
ae
Rosa
ceae
Laur
acea
e
Rubi
acea
e
Cari
cace
ae
Com
bret
acae
a
Ster
culia
ceae
Vita
ceae
Euph
orbi
acea
e
Mus
acea
e
Verb
enac
eae
0
2
4
6
8
10
12
14
16
6 99
9 85 4
3 26 5 4 4 3 2 1 1 1
Plant Richness Fruit Fly Richness
Plant Families
Spec
ies
Rich
ness

25
STUDY TWO: HOST USE
…………………………………………………… ……………………………… ……………… ………………. ………………………………………………………………… ……………………………… ……………… ……………….
……………

26
Results 2:Species Associations
STUDY TWO: HOST USE
Annacardium_occidentale
Mangifera_indica_
Sclerocarya_birrea
Annona_cherimolaAnnona_muricata
Annona_reticulataAnnona_senegalensis
Annona_squamosa
Cananga_odorata
Carica_papaya
Terminalia_catappa
Momordica_charantia
Cucumis_melo_Cucurbita_spp.
Drypetes__natalensis
Persea_americana_
Antiaris_toxicaria_
Artocarpus_sp._
Ficus_sp._
Musa_sp.
Acca_sellowiana
Eugenia_uniflora
Psidium_guanjavaCydonia_oblonga
Prunus_Spp._
Coffeae_arabica
Citrus_limon
Citrus_reticulata
Citrus_sinensis
Citrus_Spp._
Chrysophyllum_albidum
Manilkara_zapota_
Capsicum_annum
Lycopersicon_esculentum
Solanum_Spp._
Theobroma_cacao_
Vitex_sp._
Vitis_vinifera
Bactrocera_cucurbitae
Bactrocera_invadensCeratitis_anonae_Ceratitis_capitata
Ceratitis_cosyra
Ceratitis_fasciventris_
Ceratitis_punctata_Ceratitis_rosa
Dacus_bivittatus_Dacus_cilliatus
Trirhithrum_coffeae_
-300 -240 -180 -120 -60 60 120 180 240 300 360
Axis 2 (38.2%)
-300
-250
-200
-150
-100
-50
50
100
150
200
250
300
350
Axi
s 3
(20.
8%
)
First time infestation of B. invadens on T. catappa, A. toxicaria, E. uniflora, A. selllowiana, Musa spp. and C. Arabica, T. cacao and C. oblonga in Uganda.

27
Results 3
STUDY TWO: HOST USE
Com
bret
acae
a
Anac
ardi
acea
e
Myr
tace
ae
Anno
nace
ae
Mor
acea
e
Cucu
rbita
ceae
Ster
culia
ceae
Sola
nace
ae
Laur
acea
e
Rosa
ceae
Caric
acea
e
Vita
ceae
Sapo
tace
ae
Rubi
acea
e
Mus
acea
e
Euph
orbi
acea
e
Ruta
ceae
Verb
enac
eae
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
100.0
Plant Families
Mea
n Po
sitivi
ty (%
)
Term
inal
ia c
atap
pa
Psid
ium
gua
njav
a
Man
gife
ra in
dica
Pers
ea a
mer
ican
a
Citr
us li
mon
Citr
us s
inen
sis
Citr
us r
eticu
lata
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
0
10
20
30
40
50
60
70
80
90
100
B. invadens (%) Overall Positivity (%)
Fruit Species
Pos
itiv
ity
(%)
B. i
nva
den
s (%
)

28
Results 4: Mango Fruit Host Utilization
STUDY TWO: HOST USE
Keitt
Kate
Biire
Glen
Zille
tte
Borib
o
Kago
gwa
Appl
e
Dodo
Palv
in
Kent
Tom
my
0
10
20
30
40
50
60
70
80
05101520253035404550
Mean/ Kg Positive (%)
Infe
stati
on (L
arva
e/Kg
)
Positi
vity
(%)
ALL ZONES
Keit
Tom
my
Kago
gwa
Biire
Dodo
Kent
Borib
o
Palv
in
Appl
e
Kate
Glen
Zille
tte
0
10
20
30
40
50
60
0
10
20
30
40
50
60
Mean Positive (%)
Infe
stati
on (L
arva
e/Kg
)
Positi
vity
(%)
WMHF
Kate
Kago
gwa
Biire
Tom
my
Appl
e
Keitt
Borib
o
Palv
in
Zilla
tte
Glen
Dodo
Kent
0102030405060708090
100
05101520253035404550
Mean/ Kg Positive (%)
Infe
stati
on (L
arve
a/Kg
)
Positi
vity
(%)
LVC
Zille
tte
Biire
Kago
gwa
Appl
e
Dodo
Kent
Glen
Tom
my
Kate
Borib
o
Palv
in
Keitt
0.010.020.030.040.050.060.070.080.0
0.020.040.060.080.0100.0120.0140.0160.0180.0200.0
Mean/Kg Positive (%)
Positi
vity
(%)
Infe
stati
on (L
arva
e/ K
g)
NMF
29
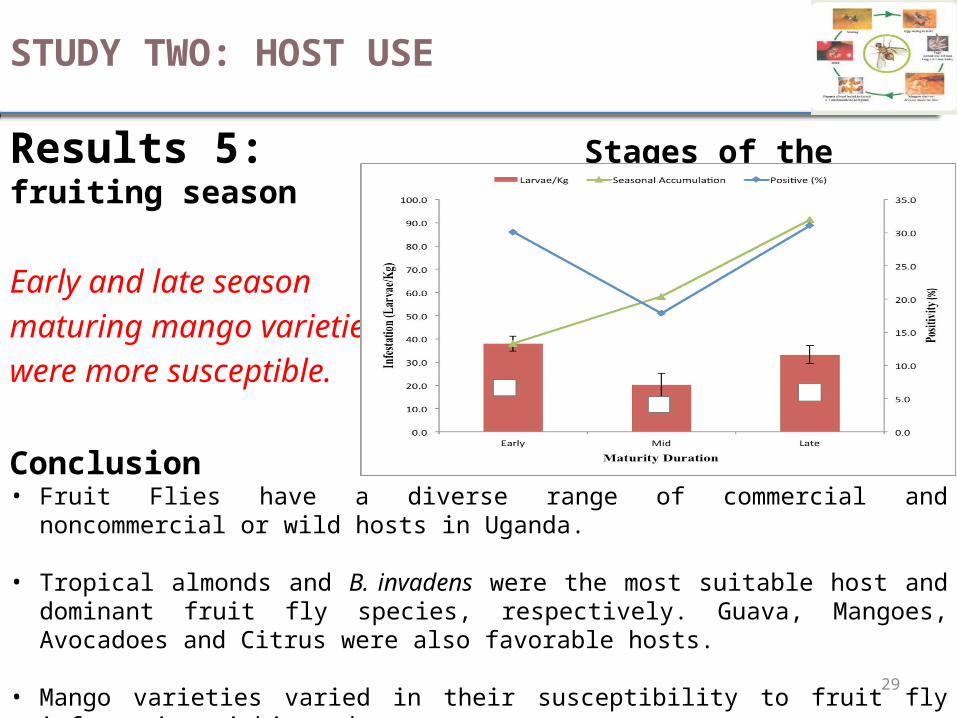
Results 5: Stages of the fruiting season
Early and late season maturing mango varieties were more susceptible.
STUDY TWO: HOST USE
Conclusion• Fruit Flies have a diverse range of commercial and noncommercial or wild hosts in Uganda.
• Tropical almonds and B. invadens were the most suitable host and dominant fruit fly species, respectively. Guava, Mangoes, Avocadoes and Citrus were also favorable hosts.
• Mango varieties varied in their susceptibility to fruit fly infestation within and across zones.
• New fruit fly-host associations were probably due to the adaptive evolution or new records

30
STUDY THREE: PP HYPOTHESIS
Does mother know best?

31
Introduction
• Preference of oviposition vs offspring performance (P-P) is of interest (Bonebrake et al., 2010; Heard, 2012).
• P–P hypothesis: females evolve oviposition behaviors that maximize offspring growth and survival (Thompson, 1988).
• Positive P-P (Rossi and Strong, 1991; Hanks et al., 1993), and negative (Karban and Courtney, 1987; Horner and Abrahamson, 1992) correlations have been recorded.
• Due to the polyphagous nature of B. invadens it was important to assess its relative P-P in the various hosts and mango varieties.
• Hypothesis: Because of its polyphagous nature, B. invadens can obscure the P-P.
STUDY THREE: PP HYPOTHESIS

32
Materials and MethodsSTUDY THREE: PP HYPOTHESIS
• Lake Victoria Crescent (Wortman and Eledu, 1993)
• Five host plants: sweet orange, tropical almonds, avocado, guava and mango.
• Fruits naturally infested by B. invadens were incubated to determined fruit host preference for oviposition (Aluja et al., 2009).
• Pupae handled as by Copeland et al. (2002), adults as by White & Elson-Harris (1992).
• The adults were sexed and separately weighed.
• Developmental time of development stages was measured as time (days) for each pupae to develop into teneral adult stage.

33
Results 1Fruit host preference for oviposition differed significantly
STUDY THREE: PP HYPOTHESIS
Tommy
Zillatt
eKeitt Ken
t
Kagogw
aApple
Palvin
DodoKate Biire Glen
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
Mango Varieties
Infe
stat
ion
(Lar
vae/
Kg)
Trop
ical a
lmon
ds
Guav
a
Man
go
Citr
us
Avoc
ado
0
2
4
6
8
10
12
14
16
18
20
Fruit Species
Num
ber
of la
rvae
/ Fru
it

34
Results 2Pupal development too varied significantly 1
STUDY THREE: PP HYPOTHESIS
12 14 16 18 20 22 24 26 28 30 32 34 360
50
100
150
200
250
TA Citrus Guava Avocado Mango
Cumulative Days
Ad
ult
Em
erge
nce
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 170.00
2.00
4.00
6.00
8.00
10.00
12.00
Apple Biire Dodo Glen Kagogwa Kate KeittKent Palvin Tommy Zillatte
Cumulative Days*
Adul
t Em
erge
nce

35
Results 2Pupal development too varied significantly 2
STUDY THREE: PP HYPOTHESIS
36
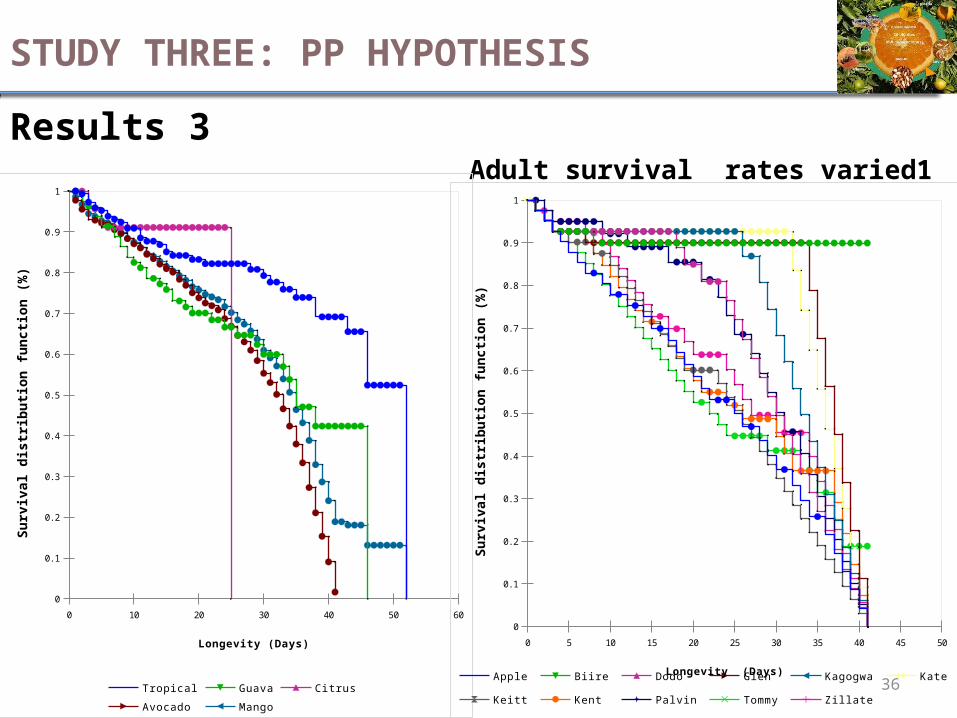
Results 3Adult survival rates varied1
STUDY THREE: PP HYPOTHESIS
0 10 20 30 40 50 600
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Tropical Guava Citrus Avocado Mango
Longevity (Days)
Surv
ival
dist
ributi
on fu
nctio
n (%
)
0 5 10 15 20 25 30 35 40 45 500
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Apple Biire Dodo Glen Kagogwa Kate Keitt Kent
Palvin Tommy Zillate
Longevity (Days)
Surv
ival
dist
ributi
on fu
nctio
n (%
)

37
Results 3Adult survival rates varied2
STUDY THREE: PP HYPOTHESIS

38
Results 4Adult weight and sex ratio were significantly higher for TA and least in citrus
STUDY THREE: PP HYPOTHESIS

39
Results 5P-P was consistent among species but less for cultivars
STUDY THREE: PP HYPOTHESIS
B. invadens choice of fruit species for female oviposition is guided by preference performance hypothesis. However, among varieties of the same species (for instance mangoes), females tend to maximize their fitness and not necessarily that of offsprings as proposed by the optimal foraging hypothesis!

40
Study IV: Morphometric (intra species) Diversity
Diversity, Host Utilisation and Ecological Niche of Tephritid (Diptera: Tephritidae) Fruit flies in Uganda
Ecological Nichie and Distribution Studies
Fruit Fly Diversity Studies
Study I: Species Diversity
Study III: Effect of Host Type and Variety on Fitness/ Survival Study II: Fruit fly Host Utilisation in Uganda
Study VI: Projected Effect of Climate Change on Distribution
Study V: Actual and potential Distribution of Fruit Flies in Uganda
Host Utilisation Studies
Results scheme and flow…

41
STUDY FOUR: INTRASPECIES DIVERSITY
“The millions of species now inhabiting this planet have, evolved from a common ancestor, and the multiplication in the number of species has been generated as single species have split into two.” Darwin

42
B. Papayae Oriental fruit fly
IntroductionIdentity of B. dorsalis complex (>70) is difficult, even with molecular tools (Clarke et al., 2005; Drew et al., 2008).
STUDY FOUR: INTRASPECIES DIVERSITY
B. Philippinensis
B. Carambolae
B. Invadens
B. dorsalis/B. papayae/B. philippinensis
B. carambolae
B. opiliaeB. cacuminata
B. musaeB. occipitalis

43
Introduction 2• Variations may lead to biotypes, host or pheromone races with
variable responses to management (Menken et al., 1996).
• Has site and host specific phynotypic variations taken course among B. invadens populations in Uganda?
• This study: – Assess the morphometric variations among three geographic
and hosts B. invadens populations in Uganda.
– Examine fluctuating asymmetry (FA, small random departures from perfect symmetry among individuals).
STUDY FOUR: INTRASPECIES DIVERSITY

44
Materials and Methods• WMHF, LVC and NMF (Wortman and Eledu, 1993).
• Terminalia catappa, Psidium guanjava and M. indica.
• Both wings were slide mounted using Canada Balsam. Once dry, photos were taken using a sony camera
• Euclidean distance matrix analysis and variance structure coordinate system geometric techniques.
• Size and shape were analyzed throughout 15
landmarks for 360 specimens after symmetrization.
STUDY FOUR: INTRASPECIES DIVERSITY
45
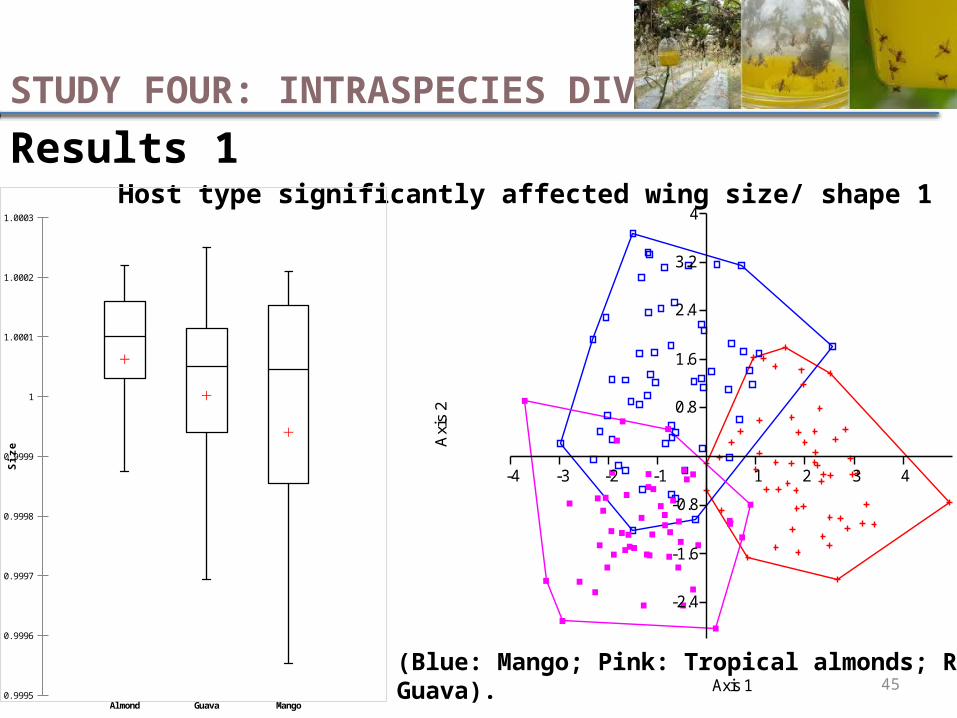
Results 1Host type significantly affected wing size/ shape 1
STUDY FOUR: INTRASPECIES DIVERSITY
0.9995
0.9996
0.9997
0.9998
0.9999
1
1.0001
1.0002
1.0003
Almond Guava Mango
Size
-4 -3 -2 -1 1 2 3 4
Axis 1
-2.4
-1.6
-0.8
0.8
1.6
2.4
3.2
4
Axi
s 2
(Blue: Mango; Pink: Tropical almonds; Red: Guava).

46
Results 1Host type significantly affected intraspecific shape variation 2
2
STUDY FOUR: INTRASPECIES DIVERSITY

47
Results 2Geographical Variations in Wing Size/ Shape 1
STUDY FOUR: INTRASPECIES DIVERSITY
0.9994
0.9995
0.9996
0.9997
0.9998
0.9999
1
1.0001
1.0002
1.0003
WMHF NMF LVC
Size
-4 -3.2 -2.4 -1.6 -0.8 0.8 1.6 2.4 3.2
Axis 1
-5
-4
-3
-2
-1
1
2
3
Axi
s 2
(Blue: WMHF; Pink: NMF; Red: LVC).

48
Results 2Intraspecific Geographical Variations in Wing Shape 2
STUDY FOUR: INTRASPECIES DIVERSITY
49
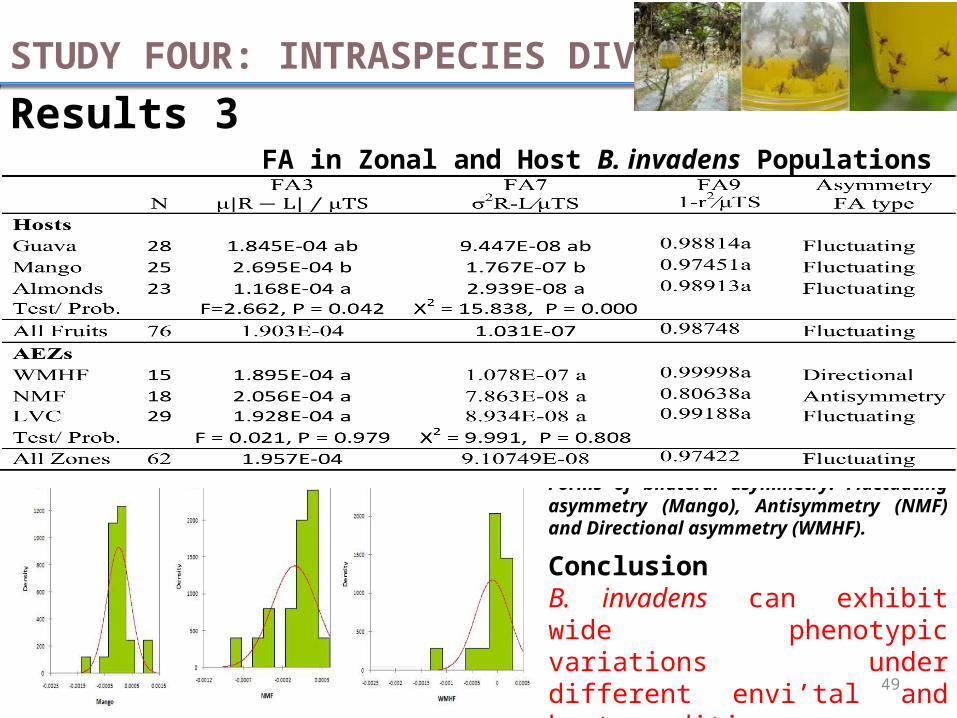
Results 3FA in Zonal and Host B. invadens Populations
STUDY FOUR: INTRASPECIES DIVERSITY
Forms of bilateral asymmetry: Fluctuating asymmetry (Mango), Antisymmetry (NMF) and Directional asymmetry (WMHF).
ConclusionB. invadens can exhibit wide phenotypic variations under different envi’tal and host conditions

50
Study IV: Morphometric (intra species) Diversity
Diversity, Host Utilisation and Ecological Niche of Tephritid (Diptera: Tephritidae) Fruit flies in Uganda
Ecological Niche and Distribution Studies
Fruit Fly Diversity Studies
Study I: Species Diversity
Study III: Effect of Host Type and Variety on Fitness/ Survival Study II: Fruit fly Host Utilisation in Uganda
Study VI: Projected Effect of Climate Change on Distribution
Study V: Actual and potential Distribution of Fruit Flies in Uganda
Host Utilisation Studies
Results scheme and flow…

51
STUDY FIVE: POTENTIAL DISTRIBUTION
“There is no part of natural history more interesting or instructive, than the study of the geographical distribution of animals.” Alfred Russell Wallace (1823-1913)

52
Introduction• Spatial suitability good in the design explicit management strategies for the pest.
• ENM provides an option for potential species distribution.
• ENM allows obtaining range of conditions for species survival/ reproduction (Pearson, 2007; Rubio and Acosta, 2010).
• Prediction model is a function of species response to the environmental variables, hence fundamental niche (Austin, 2007).
• This study: – Determine the geographical regions that are ecologically suitable for fruit fly establishment.
– Explore the climatic profiles underpinning the selected species distribution, to understand their niche requirements
STUDY FIVE: POTENTIAL DISTRIBUTION

53
Materials and Methods 1• Ten species: B.invandens, C.anonae, C.cosyra, C.capitata, C.fasciventris, C.rosa,
T.coffeae, B.curcubitae, D.bivitattus and D.ciliatus.
• Nineteen (19) environmental variables at 30 arc-seconds (~1 km²) partial resolution were derived from the WorldClim project (Hijmans et al. 2005).
• Models by Maxent and Bioclim (Graham and Hijmans, 2006; Phillips et al., 2006).
• Model evaluation: 75% of the original presence data (training sample), while 25% was test data (Pearson, 2007; Acosta, 2008; Echarri et al., 2009).
• Accuracy of the model was evaluated by calculating the AUC in a receiver operating characteristic plot
STUDY FIVE: POTENTIAL DISTRIBUTION
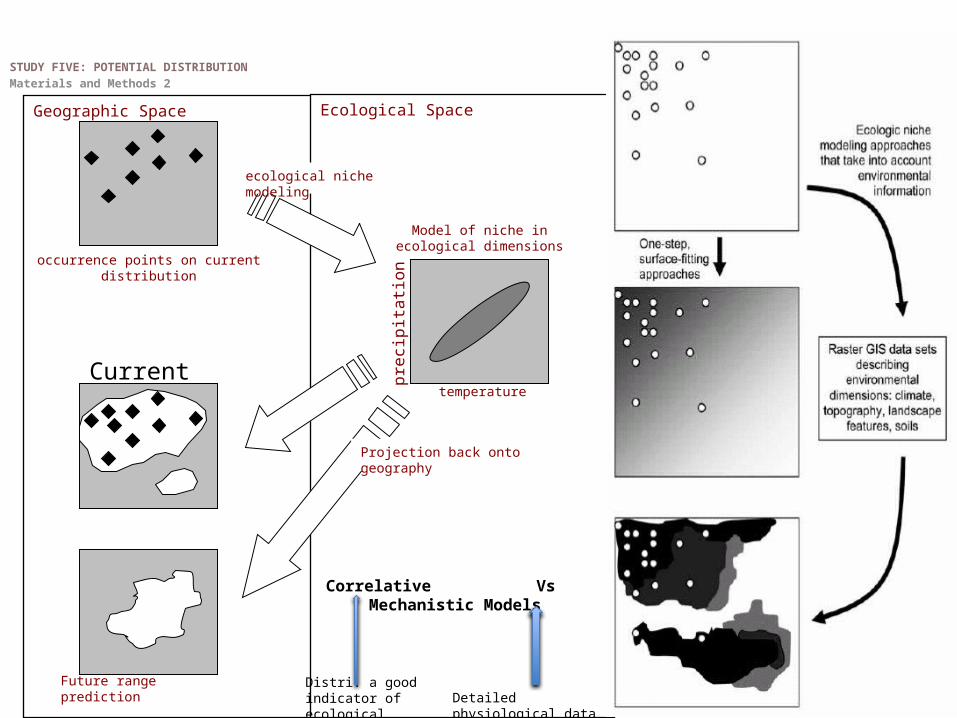
Current range prediction
Geographic Space Ecological Space
occurrence points on current distribution
ecological niche modeling
Projection back onto geography
Future range prediction
temperature
Model of niche in ecological dimensions
pre
cip
itatio
n
Current
Correlative Vs Mechanistic Models
Distri. a good indicator of ecological needs Detailed physiological data
STUDY FIVE: POTENTIAL DISTRIBUTIONMaterials and Methods 2

Materials and Methods 3Defining Niches
STUDY FIVE: POTENTIAL DISTRIBUTION
( )jir e ( ; )j j j
i ix R
( ; )j x T
Grinnell, scenopoetic
Elton, bionomic
MovementsG
BAM Diagram
B
M
A
1( ) ( ; ) ( ; )
jj j j j ji
i i iji
dxr e x x
x dt R T
Fundamental nicheIntrinsic Growth Rate(Scenopoetic)
Resource-consumer dynamics, competitors,predator-prey (bionomic).
Migrationcolonization, history
• Physiological tolerances, migration limitations and evolutionary forces that limit adaptation
• A starting point for abiotic factors is often climate. Climate variables often also correlate with other variables
Environmental Gradient
(Hawkins et al., 2003) 55

56
Results 1Records of Fruit Flies and Potential Distribution 1
STUDY FIVE: POTENTIAL DISTRIBUTION
Species are diverse and widely distributed!

57
Results 1Bioclimatic profiles of fruit fly species
STUDY FIVE: POTENTIAL DISTRIBUTION

58
Results 2: Potential DistributionSTUDY FIVE: POTENTIAL DISTRIBUTION
I (D. punctatifrons), II (T. coffeae), III (C. fasciventris), IV (B. cucurbitae), V (D. cilliatus), VI (C. cosyra), VII (C. capitata), VIII (D. bivittatus), IX (C. anonae), X (B.invadens) & XI (ALL).

59
Results 2: Limiting Factors
STUDY FIVE: POTENTIAL DISTRIBUTION

STUDY FIVE: POTENTIAL DISTRIBUTION: Limiting Factors for selected species

61
Results 2: Limiting Factors
STUDY FIVE: POTENTIAL DISTRIBUTION

62
Results 3STUDY FIVE: POTENTIAL DISTRIBUTION
Conclusion
• Fruit flies pose a significant threat to the country; countrywide potential distribution of native and exotic species was demonstrated.
• Precipitation and temperature significantly determined distribution.
• Central & mid north zones were most suitable habitats, while the western, north eastern & areas around Albert Nile were marginal.

STUDY SIX: CLIMATE INDUCED RANGE SHIFTS
Projections of Climate-Induced Future Range Shifts among Fruit Fly Species in Uganda.
Charles Masembe, Brian E. Isabirye, I. Rwomushana, A. M. Akol, Caroline K. Nankinga
Journal of Plant Protection Science
“Led by a new paradigm, scientists adopt new instruments and look in new places...” Thomas S. Kuhn (1922-1996)

64
Introduction• Knowledge on geographical suitability of the pest is needed in the design of
spatially and temporally explicit management strategies for pests.
• Global climate continues to change (IPCC, 2007).
• For agriculture, climate change will be significant, as such changes are associated with shifts in pest and disease ranges, posing new risks (Cooper et al., 2013).
• Considerable effort has gone into predicting the effect of future climate scenarios (Walther et al., 2002; Chambers et al., 2005, Shi et al., 2006; McKenney et al., 2007).
• This study: How fruit fly local-level distribution patterns may be expected to change under
future climate change and the comparative potential range shifts among species.
STUDY SIX: CLIMATE INDUCED RANGE SHIFTS

65
Materials and Methods
STUDY SIX: CLIMATE INDUCED RANGE SHIFTS
• Species assessed were: Bactocera invandens, Ceratitis anonae, Ceratitis cosyra, Ceratitis capitata, Ceratitis fasciventris, Dacus punctatifrons, Trirhithrum coffeae, Bactrocera curcubitae, Dacus bivitattus, and Dacus ciliatus.
• Annual mean temperature and mean temperature of wettest quarter were chosen, while moisture gradients were represented by mean annual precipitation and precipitation of coldest quarter.
• Climatic controls on current fruit fly distributions were summarized using the climate envelope (CE) approach (Nix, 1986).
• From the extent of the current CE for each fruit fly species, Future climate variables were generated by two GCMs: HADCM and CCCMA under emission scenarios A2 and B2.
• In the full-dispersal scenario, changes in CE area were calculated by expressing the future CE area as a percentage of the current CE area.
• For the no-dispersal scenario, future maps were overlaid on current maps and only the area of overlap was taken as the future distribution.
• Predicted current and future local species richness and turn over of each of the regions were estimated
66
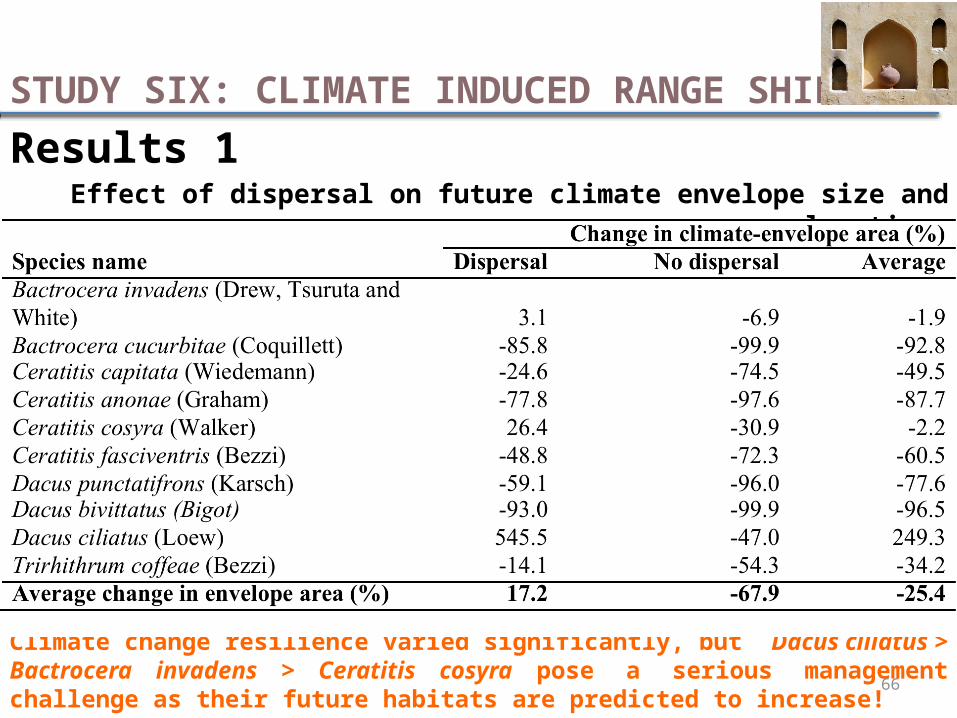
Results 1Effect of dispersal on future climate envelope size and location
STUDY SIX: CLIMATE INDUCED RANGE SHIFTS
Climate change resilience varied significantly, but Dacus ciliatus > Bactrocera invadens > Ceratitis cosyra pose a serious management challenge as their future habitats are predicted to increase!

67
Results 2Range Shift: Increase
STUDY SIX: CLIMATE INDUCED RANGE SHIFTS
Baseline: 1950-2000 Future: 2000-2050
D. ciliatus
B. invadens
C. cosyra

68
Results 2Range Shift: Decrease
STUDY SIX: CLIMATE INDUCED RANGE SHIFTS
Baseline: 1950-2000 Future: 2000-2050
D. bivittatus
B. cucurbitae
C. anonae
69
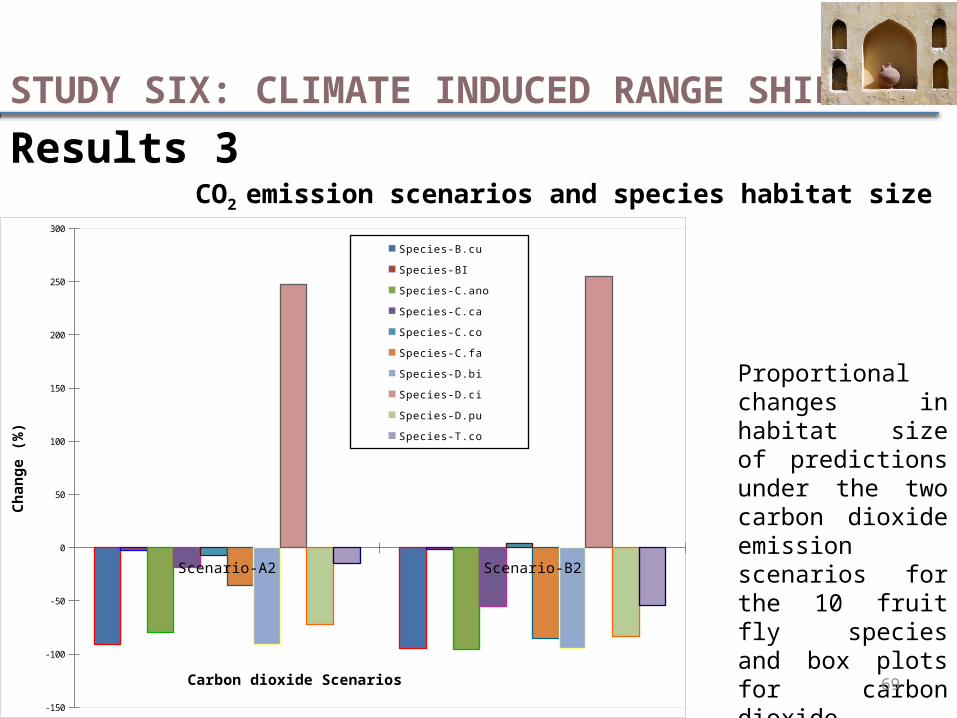
Results 3CO2 emission scenarios and species habitat size
STUDY SIX: CLIMATE INDUCED RANGE SHIFTS
Scenario-A2 Scenario-B2
-150
-100
-50
0
50
100
150
200
250
300
Species-B.cu Species-BI
Species-C.ano Species-C.ca
Species-C.co Species-C.fa
Species-D.bi Species-D.ci
Species-D.pu Species-T.co
Carbon dioxide Scenarios
Cha
nge
(%)
Proportional changes in habitat size of predictions under the two carbon dioxide emission scenarios for the 10 fruit fly species and box plots for carbon dioxide scenarios from a 1950-2000 baseline to the 2050 future period.
70
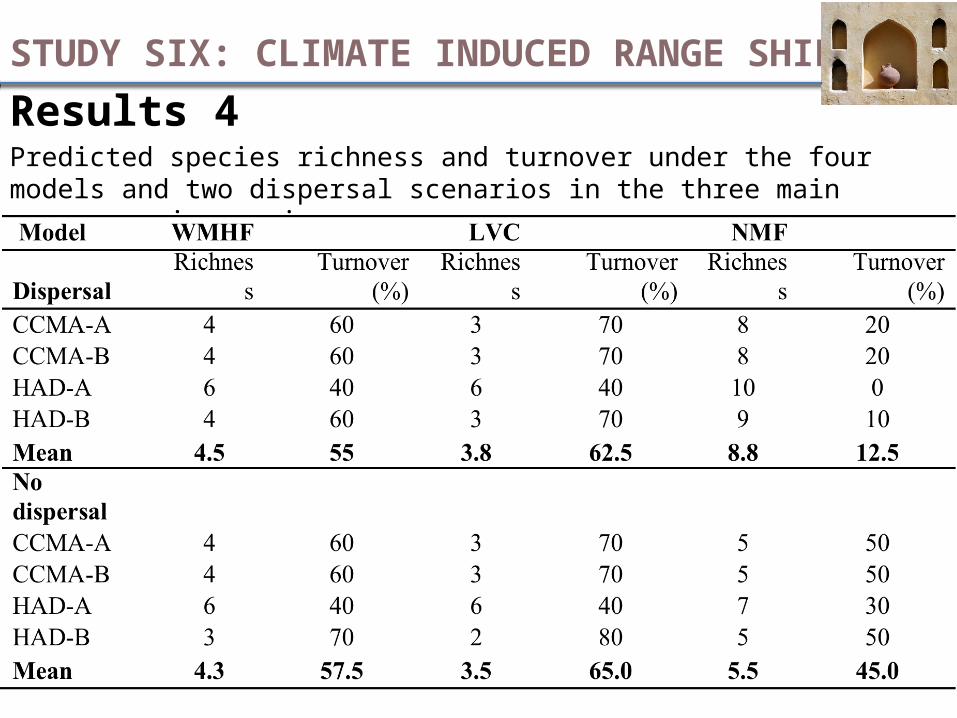
Results 4 Predicted species richness and turnover under the four models and two dispersal scenarios in the three main mango-growing regions.
STUDY SIX: CLIMATE INDUCED RANGE SHIFTS

71
Results 5STUDY SIX: CLIMATE INDUCED RANGE SHIFTS
Conclusion
• Most species are vulnerable and will likely be unable to keep pace with climate change, with habitat losses averaging 25.4% by 2050 future period.
• Fruit fly climate change resilience varied: Dacus ciliatus > Bactrocera invadens > Ceratitis cosyra > Trirhithrum coffeae > Ceratitis capitata > Ceratitis fasciventris > Dacus punctatifrons > Ceratitis anonae > Bactrocera cucurbitae > Dacus bivittatus.
• Dacus ciliatus > Bactrocera invadens > Ceratitis cosyra pose a serious management challenge as range will likely increase
• Future ranges are predicted to shift northwards, mainly to the Northern Moist Farmlands.

72
GENERAL CONCLUSIONS, DISCUSSIONS AND RECOMMENDATIONS I
At least ten tephritid fruit fly species; but B. invadens is the most abundant, and was observed to be displacing the other fruit fly fauna
Difference in zonal faunal composition can be attributed to their (zones) inherent differences in envital conditions, hosts and farming systems.
NMFs recorded lesser alternative hosts for fruit flies, which was converse to the LVC and the WMHF, hence differences in composition
Dominance of B. invadens can be attributed to competition efficiency (Ekesi et al., 2009), and reproduction and resource distribution (Kiesecker et al., 2001).
The LVC offers opportunities for resource distribution, which avoids clumping, converse to the NMFs, which might also explain the difference in displacement pressures between the two zones.

GENERAL CONCLUSIONS, DISCUSSIONS AND RECOMMENDATIONS II
A wide range of hosts was recorded, albeit with significant variability in preference and infestation levels among types, varieties and zones.
Fruits have specific adaptations in their ecological requirements, which ultimately determine their (fruit) susceptibility in their respective environments.
Preference of the flies for the local selection and Kagogwa varieties may be attributed to an increase in performance on these varieties or due to experience or learning (Szentesi and Jermy 1990; Dukas and Bernays, 2000).

74
GENERAL CONCLUSIONS, DISCUSSIONS AND RECOMMENDATIONS III
B. invadens does undergo rapid phenotypic variability which can lead to biotypes, host races, etc…
May lead different populations to adapt and survive in difficult conditions such as the stress caused by control practices and subsequently cause resistance among populations.
The recorded fine-scale intraspecific population phenotypic variations may simply be evidence of phenotypic divergences rather than interspecific differences (Schutze et al., 2012).
Such differences may be irresolvable using techniques such as molecular analysis, hence the current lack of molecular markers to discriminate between these eco- and host types.

75
GENERAL CONCLUSIONS, DISCUSSIONS AND RECOMMENDATIONS IV
The most suitable niches encompass areas around Central and mid north zones, while the western, northeastern and areas around Albert Nile were characterized as marginal.
Current and future niches offer the optimum bioclimatic tolerance limits.
Current and future potential distributions of fruit flies will be determined not only by climate but also dispersal ability, biotic interactions, genetic adaptation, and abiotic factors.
The Ethiopian fruit fly (D. ciliatus), B. invadens and D. ciliatus should be of great concern as range is predicted to increase.

76
GENERAL CONCLUSIONS, DISCUSSIONS AND RECOMMENDATIONS III The ecology of fruit flies needs to be further explored to understand
how community composition evolves in the other landscapes and mgt options.
Habitat suitability maps for these species could be improved by inclusion of edaphic and host plant data.
Further studies on biology of fruit flies in the different edapho-climatic conditions of Uganda are recommended
Achievement of optimum management of fruit flies across farming landscapes is possible, with IPM and area-wide mgt (Ekesi and Billah, 2006; Dyck, Hendrichs and Robinson, 2005).

77
Publications I Refereed Journal Papers
1. Isabirye BE, Masembe C, Akol AM, Muyinza H, Rwomushana I, Nankinga CK (2015) Modeling the Potential Geographical Distribution and Ecological Niche of Selected Fruit Fly (Diptera: Tephritidae) Species in Uganda, Journal of Plant and Pest Science, 2 (1): 18-33
2. Alex Mayamba, Caroline Kukiriza Nankinga, Brian Isabirye, Anne Margaret Akol (2014). Seasonal Population Fluctuations of Bactrocera invadens (Diptera: Tephritidae) in Relation to Mango Phenology in the Lake Victoria Crescent, Uganda. Fruits, 2014, vol. 69, p. 473–480
3. B.E. Isabirye, A. M. Akol, H. Muyinza, C. Masembe and I. Rwomushana, C. K. Nankinga (2015). Fruit Fly (Diptera: Tephritidae) Host Status and Relative Infestation of Selected Mango Cultivars in three Agro Ecological Zones in Uganda. International Journal of Fruit Science. (In Press).
4. B.E. Isabirye, C. Masembe, C. K. Nankinga, A. M. Akol, 2013. Geometric Morphometrics of Geographic and Host-Associated Population Variations of Bactrocera invadens in Uganda. American Journal of Agriculture and Environment.
5. B.E. Isabirye, A. M. Akol, C. K. Nankinga, C. Masembe, I. Rwomushana (2015). Species Composition and Community Structure of Fruit Flies (Diptera: Tephritidae) Across Major Mango-Growing Regions in Uganda. International journal of Tropical Insect science, 1-12.
6. A. M. Akol, C. Masembe, B. E. Isabirye, C. N. Kukiriza, and I. Rwomushana (2014). Oviposition Preference and Offspring Performance in Bactrocera invadens (Diptera: Tephritidae). International Research Journal of Horticulture. IRJH 2014, 2(3): 36-44.
7. Brian E. Isabirye, Charles Masembe,, I. Rwomushana, Caroline K. Nankinga, A. M. Akol (Review). Projections of Climate-Induced Future Range Shifts among Fruit Fly Species in Uganda. Journal of Plant Protection Science

78
Publications II Extended Abstracts
1. Anne Akol, Brian Isabirye, Caroline Nankinga, Charles Masembe and Ivan Rwomushana, 2014. Species Composition and Community Structure of Fruit Flies across Major Mango-Growing Regions in Uganda. 9th International Symposium on Fruit Flies of Economic Importance (ISFFEI).
2. Anne Akol, Charles Masembe, Brian Isabirye, Caroline Nankinga and Ivan Rwomushana, 2014. Oviposition Preference and Offspring Performance in Phytophagous Fruit Flies: The African invader, Bactrocera invadens. 9th International Symposium on Fruit Flies of Economic Importance (ISFFEI).
3. Brian Isabirye, Charles Masembe, Caroline K. Nankinga, I. Rwomushana, Harriet Muyinza, Anne M. Akol, 2014. Projections of Climate-Induced Future Range Shifts among Fruit Fly Species in Uganda. 9th International Symposium on Fruit Flies of Economic Importance (ISFFEI).
4. Brian Isabirye, Charles Masembe, Caroline Nankinga, Harriet Muyinza and Anne Akol, 2014. Geometric Morphometrics of Geographic and Host-Associated Population Variations of Bactrocera invadens in Uganda. 9th International Symposium on Fruit Flies of Economic Importance (ISFFEI).
5. Caroline Nankinga, Brian Isabirye, Mayamba Alex, Harriet Muyinza, Winnifred Aool, Ivan Rwomushana, Philip Stevenson and Anne Akol, 2014. Status of Fruit Fly Infestation of Mango and Other Fruits in Uganda. 9th International Symposium on Fruit Flies of Economic Importance (ISFFEI).
6. B. E. Isabirye, C.K. Nankinga, H. Muyinza, C. Masembe and A.M.Akol, 2012. Effect of Three Host Species on Infestation Levels, Offspring Survivorship, Sex Ratio and Body Weight of Bactrocera invadens (Diptera: Tephritidae). 2nd International Symposium of TEAM.

All of us should be worried about fruit flies as serious
pests, because of their diversity; direct larval feeding on
the host fruit (economic loss); long list of host plants;
flexibility in developing
different life history strategies, including rapid adaptation
to
new habitats; efficient habitat utilization; an ability to
rapidly develop host and geographic races (particularly
true for Bactocera species); and short generation time
(rapid multiplication)
(Brian E. Isabirye, 2015)
Take Home ….

Supervisors:
Mwebale inhoo!!!
“Though the road's been rocky it sure feels good to me.” ― Bob Marley

81
Extras

82
ENM Publication up to 1917-2012
Análisis ecológico del Pleistoceno
Bases de datos, colecciones e inventarios
Biodiversidad
Biogeografía y filogeografía
Cambio climático
Coberturas
Conceptos de especie y subespecie
Conceptos de nicho ecológico
Conservación
Conservadurismo de nicho
Distribución geográfica
Especies invasoras
Evaluación y validación de modelos
Georreferenciación
Importancia de la escala
MNE enfermedades infecciosas
Modelos de distribución y comparación de algoritmos
Sistemas de información Geográfica
Software, guías y tutoriales
0 10 20 30 40 50 60
8
20
24
42
54
9
9
16
53
45
53
23
33
4
9
14
40
11
21






























