
doi:10.1182/blood-2005-09-3621Prepublished online October 27, 2005;
Abby L Olsen, David L Stachura and Mitchell J Weiss cellsDesigner blood: creating hematopoietic lineages from embryonic stem
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 1
Designer Blood: Creating Hematopoietic Lineages from Embryonic Stem Cells
Abby L. Olsen1,2, David L. Stachura 1 and Mitchell J. Weiss3,4
Running head: Producing blood lineages from ES cells
1. Cell and Molecular Biology Graduate Program, The University of Pennsylvania School of Medicine, Philadelphia, PA 19104, USA
2. Combined Degree Graduate Program, The University of Pennsylvania School of Medicine, Philadelphia, PA 19104, USA
3. The Children’s Hospital of Philadelphia, Division of Hematology and the University of Pennsylvania, Philadelphia, PA 19104, USA
4. Corresponding Author: Mitchell J. Weiss, Division of Hematology, 3615 Civic Center Blvd., Abramson Research Center, Philadelphia, PA 19103, [email protected], Fax: (215) 590-4834, Phone: (215) 590-0565
Financial Support: This work was supported by NIH grants R01 DK064037 (M.J.W.), T32-HL007971-04 (D.S.).
Word counts: Abstract: 163 Text: 5574
SCIENTIFIC HEADING: Review Article
Blood First Edition Paper, prepublished online October 27, 2005; DOI 10.1182/blood-2005-09-3621
Copyright © 2005 American Society of Hematology
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 2
Abstract
Embryonic stem (ES) cells exhibit the remarkable capacity to become virtually any
differentiated tissue upon appropriate manipulation in culture, a property that has been
beneficial for studies of hematopoiesis. Until recently, the majority of this work utilized
murine ES cells for basic research to elucidate fundamental properties of blood cell
development and establish methods to derive specific mature lineages. Now, the advent
of human ES cells sets the stage for more applied pursuits to generate transplantable cells
for treating blood disorders. Current efforts are directed toward adapting in vitro
hematopoietic differentiation methods developed for murine ES cells to human lines,
identifying the key interspecies differences in biological properties of ES cells and
overcoming current obstacles to generating ES cell-derived hematopoietic stem cells that
are competent to repopulate adult hosts. The ultimate medical goal is to create patient-
specific and generic ES cell lines that can be expanded in vitro, genetically altered and
differentiated into cell types that can be used to treat hematopoietic diseases.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 3
Introduction
Embryonic stem (ES) cells, first isolated in 1981, are primary cell lines originally derived
from the inner cell mass (ICM) of pre- or peri-implantation mouse blastocysts1,2. ES cells
are distinguished by their capacity to self-renew in vitro and differentiate into tissues
derived from all three embryonic germ cell layers. Following microinjection into host
blastocysts, donor ES cells contribute to all tissues of chimeric mice, including the germ
line3. This property, combined with the ability to easily manipulate the ES cell genome
via homologous recombination, provides the tools to generate new animal strains in
which genes of interest are knocked out or altered. In addition, ES cells can be induced
to form various differentiated cell types in vitro. Here we review the production of blood
lineages from ES cells.
The broad developmental capacity of ES cells and the relative ease by which resident
genes can be manipulated have been valuable for understanding the basic biology of
tissue development and offer tremendous potential for cell replacement therapies in
degenerative disorders, cancer and genetic diseases. The promise of clinical applications
for ES cells was stimulated recently by the development of human lines from blastocysts,
patients and normal adults4-6. Studies of murine hematopoiesis provide a paradigm for
utilizing ES cells as a model system to examine normal developmental processes and as a
source for tissue production. This work provides a launching point for efforts to
differentiate human ES cells into transplantable hematopoietic lineages that can be
employed therapeutically.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 4
Culture of ES cells
The first requirement for successful propagation of ES cells is to maintain them in an
undifferentiated, pluripotent state by adherence to specific culture conditions.
Subsequently, differentiation into desired lineages can be initiated in a controlled and
synchronous fashion. ES cells are held in an undifferentiated state by numerous cell
intrinsic and environmental signaling pathways7,8.
In practice, murine ES cells are maintained in vitro by cocultivation on a stromal layer,
typically murine embryonic feeder (MEF) cells, and addition of leukemia inhibitory
factor (LIF), a cytokine that activates signal transduction through a cognate receptor
signaling complex9. Recent studies demonstrate that a combination of LIF and bone
morphogenic protein 4 (BMP4) can bypass serum and feeder requirements to allow self-
renewal of murine ES cells in defined medium10.
Human ES cells can be maintained on stromal cells in the presence of basic fibroblast
growth factor (b-FGF)4. In contrast to its effects on murine ES cells, LIF does not
promote self-renewal of human ES cells11,12. Human ES cells are reported to have a
particular propensity for spontaneous differentiation in culture, perhaps due to species-
specific requirements for self-renewal that are incompletely understood13-15. In this
regard, further studies to optimize the propagation of human ES cells and eliminate their
requirements for animal feeder cells are important for future clinical applications. For
example, human ES cells can be maintained on human stroma16,17, and numerous
signaling pathways and transcription factors that participate in ES cell self-renewal have
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 5
been exploited for developing feeder free culture systems18-21. This includes the use of
basic fibroblast growth factor (b-FGF)18,22 and Wnt signaling agonists23,24. The nuclear
proteins Sox2, Oct3/4 and Nanog are critical for both human and murine ES cell
maintenance, and manipulation of their expression appears to bypass some cytokine and
stromal requirements for self-renewal8,25-27. It appears that these three transcription
factors are central components of a concerted autoregulatory network that promotes ES
cell self-renewal and pluripotency by activating and repressing numerous target genes,
predominantly those encoding other nuclear proteins28.
The expansion of ES cells and maintenance of their undifferentiated state in culture are
monitored by visual inspection, testing for developmental potential under various
differentiation conditions and gene expression analysis. In reference to the latter point,
an enormous amount of data on gene expression profiling of human and mouse ES cell
lines have been generated22,29-37. Distilling this information to produce a defined
molecular signature for ES cells and to identify all of the genes important for maintaining
their self-renewal and pluripotency has not yet been achieved, although as mentioned
above, Sox2, Nanog and Oct3/4 appear to be central to this process.
Creating blood lineages from ES cells
Major technological advances in deriving mature tissues from ES cells were established
through studies of the hematopoietic system using murine cells (for practical reviews, see
references 38-40). The first step in differentiating ES cells in vitro is to remove them from
the feeder cells and cytokines that maintain their pluripotency. Subsequently, if serum is
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 6
present, a substantial fraction of ES cells differentiate into mesoderm, the germ cell layer
from which blood is derived. Under serum free conditions, murine ES cells can be
induced to form mesoderm and hematopoietic progenitors by added cytokines, most
importantly, BMP4 and vascular endothelial growth factor (VEGF)41,42. As mentioned
above, in other culture contexts, BMP4 participates in murine ES cell self-renewal10,43.
BMP-4 also stimulates mesoderm formation by human ES cells, although these
experiments were performed with serum present44. Mesoderm formation, which
represents an early step in transforming ES cells into blood, is regulated by numerous
positive and negative regulatory factors45,46. An ultimate goal is to develop defined
culture conditions that optimize mesoderm formation from ES cells; these conditions may
differ somewhat between human and mouse systems.
Two different experimental systems are used for the majority of experiments to generate
blood precursors from ES cells: formation of embryoid bodies (EBs)47,48 and induction of
hematopoietic differentiation on stromal cells49. Upon removal from stroma and
withdrawal of LIF, suspension cultures of ES cells form EBs, aggregates of differentiated
cells including mesodermal derivatives capable of hematopoietic differentiation. In 1985,
Doetschman et al noted that EBs in liquid culture form cystic structures that contain
blood islands analogous to those found in the embryonic yolk sac48. Subsequent studies
established that erythroid and myeloid lineages develop within EBs50,51. The emergence
of hematopoietic progenitors in this culture system can be tracked by the appearance of
lineage specific markers through gene expression analysis and
immunohistochemistry47,52-54. In addition, disruption of EBs by trypsin or collagenase
produces single cell suspensions from which hematopoietic progenitors can be analyzed
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 7
and enumerated by flow cytometry or methylcellulose culture assays. In this fashion, the
effects of specific manipulations such as drugs, cytokines or gene targeting can be
analyzed. Cells within developing EBs synthesize various cytokines and consequently,
the initial stages of hematopoiesis occur when serum is the only source of added growth
factors39,47. However, addition of specific cytokine combinations optimizes the
development of specific mature lineages (see “In vitro production of specific
hematopoietic lineages from ES cells”).
Differentiation of ES cells on stromal lines provides a second means for inducing
hematopoiesis. The most commonly used system is OP9, a line established from calvaria
of newborn op/op mice, which lack functional macrophage colony-stimulating factor (M-
CSF)49,55. OP9 cells enhance hematopoietic development by providing a supportive
microenvironment for differentiation. The absence of M-CSF prevents the overgrowth of
monocyte-macrophage cells, which tend to outgrow other lineages in culture systems
using wild-type stroma. ES cells plated onto OP9 stroma form mesodermal colonies after
approximately 5 days, and cells within these colonies differentiate into multipotential
hematopoietic progenitors after an additional 3 days. Similar to what is observed in EBs,
ES cell-derived erythroid, myeloid and B lineage cells are generated on OP9 stroma
without exogenous growth factors, although cytokine supplementation influences lineage
output (see “In vitro production of specific hematopoietic lineages from ES cells”). The
EB and OP9 systems exhibit similar kinetics and efficiencies for many aspects of
hematopoietic development, although each approach may offer unique advantages56. For
example, empiric studies indicate that one specific ES cell differentiation system (EB or
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 8
OP9) may optimize the production of different hematopoietic lineages, although this has
not been examined rigorously.
Hematopoietic progenitor and stem cells develop optimally in vivo through cellular
interactions and paracrine effects provided by stromal niches57,58. Recapitulating these
microenvironments in vitro could enhance cell production. Toward this goal, three
dimensional culture systems incorporating extracellular matrix (ECM) proteins have been
used to provide a scaffold for EB development59-61. It is likely that refining this approach
to more closely mimic the in vivo niches that support cell specification and
organogenesis will further optimize tissue production from ES cells. For example,
mKirre is a type Ia membrane protein that is expressed by OP9 cells and contributes to
their ability to support hematopoiesis62. Incorporation of recombinant mKirre into
artificial matrix systems could, therefore, enhance hematopoietic culture of ES cells
under more defined conditions.
Examining early hematopoietic ontogeny through studies of ES cells
During embryogenesis, the hematopoietic system is established spatiotemporally through
waves of distinct progenitors arising in different tissues63-66. The first recognizable blood
cells in the embryo are yolk sac-derived primitive erythrocytes that differ from adult-type
definitive erythrocytes in many respects including gene expression, morphology and
cytokine requirements. Hematopoietic stem cells (HSCs) capable of reconstituting adult
hosts are found initially in the murine yolk sac and in an intraembryonic region termed
the aorta-gonad-mesonephros (AGM)67,68. Around midgestation, the major site of HSC
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 9
activity and production of mature blood cells shifts to the fetal liver where definitive
erythrocytes and other adult-type lineages are produced69. Numerous studies in avian and
murine species suggested that the earliest embryonic hematopoietic progenitors and
endothelial cells derive from a common progenitor termed the hemangioblast70-72. As
discussed below, identification and analysis of this previously elusive progenitor was
greatly facilitated through the use of ES cells.
Remarkably, hematopoiesis in cultured ES cells largely recapitulates embryonic events.
For example, distinctly timed waves of primitive and definitive erythropoiesis develop in
vitro from ES cells in a fashion that parallels the onset of these developmental programs
in early embryos47. Moreover, Keller and colleagues discovered a transient progenitor in
early EBs, termed blast-colony forming cell (BL-CFC), which gives rise to primitive
erythrocytes, definitive lineages and endothelial cells73,74. Nishikawa et al extended these
studies, using cell surface markers to identify a developmental hierarchy in ES cell
differentiation in which hematopoietic cells arise from a precursor expressing the
endothelial markers Flk-1 and VE cadherin. They also identified single progenitor cells
capable of hematopoietic and endothelial differentiation75. Together, these studies helped
to establish lineage relationships between the first murine hematopoietic precursors and
verify the existence of the hemangioblast (Figure 1). These and subsequent studies are
beginning to define the characteristics of hemangioblasts, including cytokine and
transcription factor requirements. For example, BL-CFCs express the cytokine receptor
Flk-1 and are dependent on its ligand, vascular endothelial growth factor (VEGF). Gene
targeting studies have identified numerous transcription factors and signaling molecules
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 10
important for the generation or maintenance of murine ES-cell derived hemangioblasts 76-
81.
Recently, Huber et al identified hemangioblasts from the primitive streak of the early
mouse embryo82. Importantly, these progenitors have similar properties to ES cell
derived BL-CFCs but are an extremely rare and transient population in vivo. Most
embryos examined contained less than 10 hemangioblasts that could be identified by
current culture conditions. This study further illustrates the parallels between
developmental hematopoiesis in early embryos and ES cell differentiation cultures,
validating the biological relevance of the latter. Using ES cells, hemangioblasts can now
be generated and purified on a scale much greater than what could be achieved from
embryos. This, in turn, allows for further studies into the basic biology and therapeutic
potential of this progenitor.
The holy grail: hematopoietic stem cells from ES cells
By definition, transplantation of a single HSC into a suitable adult host can repopulate the
entire hematopoietic system for an extended period of time. While cultured ES cells give
rise to multipotent hematopoietic progenitors with extensive proliferative capacity, these
repopulate animals only weakly and inconsistently in most reported studies83,84. Burt et al
recently reported more robust in vivo reconstitution from purified EB-derived CD45+
cKit+ progenitors85. It is now important to further characterize these progenitors with
HSC activity, refine the conditions for their derivation and reproduce these encouraging
findings in other laboratories.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 11
Most ES cell-derived blood progenitors studied may exhibit relatively limited HSC
activity because they approximate an immature embryonic lineage that cannot effectively
survive and proliferate in adult hematopoietic niches86,87. Accordingly, “pre-HSCs” are
found in the yolk sac and para aortic splanchnopleura region of early embryos88-92.
Presumably, during fetal maturation, pre-HSCs are somehow made competent to
repopulate adult hosts, either though interactions with specific embryonic/fetal
microenvironments, execution of latent cell-autonomous genetic programs, or both
(Figure 1).
Recent work suggests that pre-HSCs from ES cells and embryos can be coaxed to
become adult repopulating cells through genetic manipulation93,94. The homeobox (Hox)
family of transcription factors participates in normal hematopoiesis and
leukemogenesis95-100. Work by Humphries’ group showed that ectopic expression of
HoxB4 can enhance the self-renewal capacity of hematopoietic progenitors and HSCs
from mice and ES cells101-105. Daley and colleagues extended these findings by showing
that enforced expression of HoxB4 in immature murine yolk sac or ES cell-derived
hematopoietic progenitors rendered them competent for long-term multilineage
engraftment of irradiated adult hosts106. In these experiments, a doxycycline-inducible
HoxB4 transgene was introduced into ES cells and expressed during a narrow window of
hematopoietic differentiation. Hence, ectopic HoxB4 can facilitate the maturation of pre-
HSCs but is not required subsequently. This result leads to several interesting questions
and areas for further study. First, for unknown reasons, lymphoid repopulation appeared
to be less robust than myeloid repopulation in the HoxB4-transduced cells, a finding
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 12
warranting further examination106,107. Second, 2 million cells were used to reconstitute
mice in these studies. It is now important to fractionate this mixture, identify the
repopulating cells and demonstrate that a single cell is capable of generating all
hematopoietic lineages. Third, basic mechanistic studies are essential to understand how
HoxB4 might educate pre-HSCs. For example, structure-function studies identified a
HoxB4 mutant with more potent regenerating activity108. In addition, it is important to
define the HoxB4 transcriptional program associated with pre-HSC maturation109. It is
possible that this line of research will identify functionally important cell signaling
interactions that can be modulated by drugs.
Additional strategies to generate HSCs from ES cells will be facilitated by ongoing and
future studies to better understand how pre-HSCs are conditioned in the embryo. For
example, stromal interactions almost certainly contribute to HSC development and
maturation as hematopoietic populations shift during embryogenesis110. In support of
this, multilineage progenitors and HSCs develop autonomously in ex vivo organ cultures
of the paraaortic splanchnopleura and AGM regions89,91,92,111. More recent work by
several groups suggests that HSC activity is induced and supported by stromal lines from
various embryonic sources112-114. It will be interesting to determine whether such stromal
lines can support the maturation of HSCs from ES cell differentiation cultures. In this
case, the next step will be to identify the specific molecules that influence HSC
development by either direct stromal interactions or paracrine effects115. Ultimately, the
goal is to reconstruct these inductive events using recombinant molecules.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 13
In vitro production of specific hematopoietic lineages from ES cells
Early studies showed that mixtures of different hematopoietic cell types develop in ES
cell differentiation cultures47-50,52,55,116. Subsequently, methods were refined to generate
relatively pure populations of specific lineages. Important variables that affect lineage
output include duration and mode of differentiation and supplemental cytokines or
chemicals. Conditions for generating erythroid cells, megakaryocytes, mast cells,
granulocytes, eosinophils, osteoclasts and lymphoid cells have been established (Table
1). These methods are useful for studying the development and physiology of specific
mature blood lineages generated from both wild type and gene targeted ES cells.
Analysis of hematopoiesis from genetically altered ES cells is particularly useful when
the mutation produces a complex phenotype with early embryonic lethality that interferes
with examination of definitive hematopoiesis. This application has been utilized
extensively; a few illustrative examples are discussed below. The methods summarized
in Table 1 were established using murine ES cells, but these are now being adapted for
use with human ES cells.
Erythroid cells.
When 7-8 day old EBs are disaggregated and plated into methylcellulose cultures with
erythropoietin (Epo) and Kit ligand (KL), the majority of resultant colonies are
erythroid47. We used this strategy to show that ES cells with a disrupted Gata1 gene,
which encodes an essential hematopoietic transcription factor, produce normal appearing
proerythroblasts that subsequently undergo maturation arrest and apoptosis117,118. In
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 14
these experiments, use of ES cells allowed us to easily generate a synchronous cohort of
erythroid precursors to pinpoint the developmental stage at which GATA-1 function
becomes essential. We used the same system to generate an immortalized GATA-1–
proerythroblast line that undergoes GATA-1-dependent terminal erythroid maturation119.
This line, termed G1E, for GATA-1– Erythroid, has been a useful tool for various
applications including erythroid gene discovery and examining mechanistic aspects of
GATA-1 function in erythroid development120-128. Zheng et al generated ES cells in
which the endogenous Gata1 gene was ablated and a tetracycline regulated Gata1 cDNA
expression cassette was inserted, permitting conditional rescue of the gene defect129.
Using this system the authors identified unique functions for GATA-1 at various stages
of erythroid development. Together, these studies illustrate how gene functions can be
studied through genetic manipulation and in vitro differentiation of ES cells.
Methods to derive definitive erythrocytes from ES cells were advanced by Beug’s group,
who developed culture conditions in which EB-derived immature erythroid progenitors
can be expanded in a relatively pure state roughly 107-fold over the course of several
months130. This approach relies on prior findings that dexamethasone promotes erythroid
progenitor self-renewal131-133. When dexamethasone is removed and the cytokine
mixture is adjusted, terminal maturation ensues. Carotta et al used this approach to
demonstrate an unrecognized function for the cytokine receptor Flk-1 in erythroid
development130. This role was not appreciated from in vivo studies, as Flk1–/– embryos
die early with complex, cell non-autonomous defects in hematopoiesis134.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 15
Megakaryocytes.
Culturing ES cells on OP9 stroma with thrombopoietin (Tpo) generates populations
enriched for megakaryocytes with characteristic functional features including proplatelet
production and fibrinogen binding after exposure to platelet agonists135-139. Eto et al used
this system to show that CalDAG-GEF1, a guanine nucleotide exchange factor that is
deficient in megakaryocytes lacking the transcription factor NFE2, promotes integrin-
mediated megakaryocyte signaling138. Fujimoto et al reported similar methods to derive
functional megakaryocytes and platelets from ES cells135. Our laboratory adapted these
culture methods to show that Gata1 gene-disrupted ES cells produce a unique self-
renewing bipotential progenitor with the capacity to undergo GATA-1-dependent
megakaryocyte and erythroid maturation140. A defined role at this stage of hematopoietic
development is relevant to recent findings that mutations in the GATA1 gene contribute to
Down’s Syndrome associated acute megakaryoblastic leukemia (AMKL), which exhibits
both erythroid and megakaryocytic features, suggesting that malignant transformation
may occur within a megakaryocyte-erythroid progenitor (MEP)141.
Granulocytes.
Lieber et al generated > 75% pure neutrophil cultures from ES cells142,143. This approach
involves inducing the maturation of EB-derived hematopoietic progenitors on OP9 cells
with added cytokines. The resultant neutrophils exhibit signature markers, chemotactic
response and superoxide production. The authors used this system to demonstrate
migration defects in neutrophils lacking mitogen-activated protein (MAP)/extracellular
signal-related kinase (MEKK1) through targeted mutagenesis.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 16
Mast cells.
Culturing EB-derived hematopoietic progenitors in suspension with the cytokines IL-3
and KL generates pure populations of mast cells that can be expanded significantly in
vitro47,50,144,145. Upon transplantation into mast cell-deficient mice (KitW/KitW-v), ES cell-
derived mast cells survive, proliferate, mature and mediate IgE-dependent immune
responses. This provides a powerful strategy to analyze the effects of genetic and
chemical alterations on mast cell development and function in vitro and in vivo. For
example, IgE-induced crosslinking of its high affinity receptor triggers mast cell
activation with release of inflammatory mediators, contributing to the pathophysiology of
allergic response. Several groups are dissecting this pathway by creating ES cells with
mutations in candidate genes suspected to participate in IgE receptor signaling and using
these lines to generate mast cells for functional studies144,146.
Eosinophils.
Hamaguchi-Tsuru et al generated eosinophils (~50% enriched) by differentiating ES cells
on OP9 stroma with IL-5 and either IL-3 or granulocyte-macrophage colony stimulating
factor (GM-CSF)147. The ability to generate eosinophil-enriched populations and study
their development and function at defined stages has implications for human disorders
such as allergies and asthma. For example, this approach may be a useful basis for
screens to identify new anti-inflammatory agents that inhibit eosinophil development or
function.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 17
T and B Lymphocytes.
Nakano et al cultured ES cells on OP9 stroma with IL-7 to generate early B cells. Most
of these were IgM– and completed DJ gene rearrangement, but a small proportion
differentiated into IgM+ cells that expressed the complete µ chain mRNA49. B cell
maturation from ES cells was developed further by stimulating differentiation in the
presence of IL-7 and Flt3 ligand148,149. This provides an efficient method to produce
relatively large amounts of mature Ig-secreting B cells, which, in principle, could be
adapted to engineer and produce specific murine or human antibodies.
It has been particularly challenging to differentiate ES cells into T lymphocytes, probably
because their development requires complex thymic stromal interactions that are not
easily recapitulated in vitro. Zuniga-Pflucker’s group differentiated ES cells on OP9
stroma and then reseeded Flk1+CD45- cells into fetal thymic organ cultures (FTOCs).
Although the fraction of T cells was low, normal CD4 and CD8 thymic subsets were
generated150. Recognizing that Notch signaling is uniquely important for the production
of T cells, the same group engineered OP9 cells to express the delta-like 1 (DL-1) Notch
ligand151. Differentiation of ES cells on this modified stroma produced more extensive
and complete T cell maturation including γδ and αβ T cell receptor (TCR)-bearing cells,
as well as mature CD8+ cells that express the TCR and produce interferon (IFN)-γ in
response to stimulation. However, further maturation in FTOCs is required to render
these ES cell-derived late stage T cell progenitors competent to functionally reconstitute
immunodeficient NOD/SCID mice. This system represents an important technical
advance in the manipulation of ES cells and should be useful for examining the cell-
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 18
autonomous and environmental factors that promote T lymphocyte differentiation and
maturation. The finding that ES cell-derived T lymphocyte progenitors can become self-
tolerant and functional after transplantation into mice has potential clinical utility if these
experimental methods can be adapted to humans.
Macrophages.
Macrophages can be produced from ES cells using either EB or OP9 culture systems. ES
cell-derived macrophages express Mac1 and lysozyme RNA, and stain with F4/80, a
specific macrophage antibody49,50. In addition, they exhibit numerous properties of
arterial lesion macrophages, and therefore, offer research opportunities in the field of
atherosclerosis152,153. In vitro differentiation of ES cells has been used to study the
effects of specific gene knockouts on macrophage function. For example, Guillemot et al
used this approach to demonstrate that GDID4, a guanine diphosphate dissociation
inhibitor that regulates the cycling of Rho family GTP binding proteins, is required for
normal superoxide production by macrophages154.
Dendritic Cells.
DCs are potent antigen presenting cells that can either promote immune response or self-
tolerance. ES cells offer a potential source of normal and genetically manipulated DCs
that could be used for a variety of clinical applications155-157. ES cell-derived DCs
expand in the presence of IL-3 and GM-CSF and are induced to mature by tumor necrosis
factor (TNF) α and IL-4 plus lipopolysaccharide or anti-CD40. Similar to their bone
marrow-derived counterparts, ES cell-derived DCs express characteristic markers,
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 19
process foreign antigen and stimulate primary T-cell responses in the mixed leukocyte
reaction. In one interesting application, Senju et al randomly introduced a reporter gene
flanked by loxP recombination sequences into ES cells and identified a clone in which
reporter expression was particularly high in DC progeny generated by in vitro
differentiation155. Then, they introduced ovalbumin cDNA into this active locus by CRE-
mediated recombination to generate a DC line that efficiently presented antigen to prime
ovalbumin-specific cytotoxic T-lymphocytes in vivo. This method has potential clinical
applications if it can be widely applied to other antigens.
Natural Killer (NK) Lymphocytes.
NK cells, an important arm of the innate immune system, destroy cells that fail to present
the correct MHC class I receptor, including some virally infected and malignant clones.
Lian et al. induced ES cells to differentiate into functional NK cells that killed certain
tumor lines and MHC class I-deficient lymphoblasts158. These findings provide a
mechanism to further examine NK cell biology and possibly to manufacture these cells
for antiviral and anticancer therapies.
Osteoclasts.
Osteoclasts, which are derived from HSCs, are involved in bone resorption and
remodeling. Therefore, understanding the biology of this lineage has important
implications for various bone disorders. Methods to derive osteoclasts from ES cells in
vitro have been developed and refined, mainly by Hayashi’s group159-163. Cytokines that
promote osteoclast differentiation and maturation from ES cells include M-CSF and
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 20
Receptor Activator of Nuclear factor Kappa B Ligand (RANKL). Osteoclast production
is also augmented by ascorbic acid, dexamethasone and 1α, 25-dihydroxyvitamin D3.
Analysis of gene targeted ES cells utilizing these culture methods identified several
signaling pathways that participate in osteoclastogenesis. For example, ES cells lacking
transcription factor NFATc1 are deficient in RANKL-induced osteoclastogenesis,
suggesting that NFATc1 is downstream of RANKL signaling164. In a different study,
Tsuneto et al showed that enforced expression of the myeloid transcription factor PU.1
stimulates osteoclast production in ES cells lacking SCL (Tal-1), a nuclear protein
essential for early stages of hematopoiesis165. This indicates that PU.1 is not only
important for osteoclast development, but it may also bypass some early hematopoietic
requirements for SCL.
Recent progress with human ES cells
In 1998, Thomson et al derived human ES cells from embryos4. More recently, human
and murine ES cells were developed by somatic cell nuclear transfer (SCNT), a method
in which the nucleus of an adult somatic cell (commonly from the skin for humans) is
injected into a normal enucleated oocyte to generate a blastocyst that can be used to
derive ES cells5,166. This technology opens up new opportunities to generate patient-
specific ES cell lines.
Blood from hES cells.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 21
Methods previously established for examining hematopoiesis in murine ES cells are
applicable to human studies167,168. Human ES cells differentiate to hematopoietic
lineages via EBs or culture on stromal cell lines169. It appears that human ES cell
differentiation cultures accurately mirror early human embryonic hematopoiesis, similar
to what is observed in murine systems. For example, distinct sequential waves of
primitive and definitive erythropoiesis occur in human EBs. Moreover, cells within
human EBs give rise to colonies that contain mesoderm, hematopoietic and endothelial
cells arranged into structures reminiscent of yolk sac blood islands. The nature of the
progenitor which gives rise to these colonies remains to be determined170. It is possible
that hemangioblasts originate from within these colonies. Along these lines, Wang et al
showed that CD45–, PECAM-1+, Flk-1+, VE-cadherin+ (CD45negPFV) cells within human
EBs give rise to hematopoietic and endothelial lineages171. This hints at the existence of
human hemangioblasts. However, in clonality studies, the viability and capacity for
hemato-endothelial differentiation of CD45negPFV cells were low. Hence, it will be
important to further fractionate these cells and optimize conditions for their culture and
differentiation.
While extensive studies of murine ES cells provide a broad basis for analyzing
hematopoiesis in human lines, it is important to point out that there are significant
interspecies differences that could influence culture protocols13,172. For example,
differences in self-renewal pathways are discussed earlier in the section on ES cell
culture. There are also differences in colony morphology, cloning efficiency, surface
markers and patterns of gene expression. In contrast to murine ES cells, serum free
methods to generate mesoderm and hematopoietic lineages from human ES cells are not
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 22
yet reported. It is likely that protocols to specifically optimize hematopoiesis from
human ES cells will be further developed and refined over the next few years.
Generating human HSCs.
As in murine systems, generating a human ES cell-derived HSC capable of long-term
multi-lineage engraftment is a vexing problem. It has been possible to produce human
ES cell-derived progenitors with some features of adult-type HSCs including CD34
expression, multi-lineage hematopoietic potential in clonal assays, the ability to efflux
dyes such as Hoechst 3334 and Rhodamine 123, and high aldehyde dehydrogenase
(ALDH) activity167. However, these cells exhibit limited repopulating activity after
transplantation into immunodeficient mice. Wang et al demonstrated that a population of
human ES cell-derived progenitors incapable of repopulating the marrow after
intravenous injection exhibits limited regional repopulation after intrafemoral
injections173. At minimum, this suggests defects in migratory capacity, homing or niche
interactions, similar to what is observed in pre-HSCs of the early mouse embryo174.
Consistent with this interpretation, multipotential progenitors derived from human ES
cells exhibit a distinct gene expression signature that resembles primitive embryonic
progenitors relative to adult HSCs found in bone marrow or cord blood173,175.
In contrast to murine ES cells, ectopic expression of HoxB4 does not promote the
expansion of bone fide HSCs from human ES cells173. This important finding could be
due to technical issues or could reflect intrinsic differences in the biologies of human and
mouse HSC development. In case of the latter, it is possible that manipulated expression
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 23
of different proteins in human ES cell cultures will stimulate HSC production. Candidate
molecules include Hox paralogues and numerous other nuclear regulators176-178. In
considering this experimental approach for human HSC production, it will be important
to utilize strategies that avoid stable genetic alterations by transgenes or viral vectors,
which pose a risk for malignant transformation. One innovative approach fused the cell
penetrating peptide, TAT, to HoxB4 protein179. This fusion protein was taken up into
murine bone marrow-derived HSCs and stimulated their expansion ex vivo. This
method, which allows cells to be manipulated by transcription factors without altering the
genome, could be applied to ES cells.
The future
The ability to create hematopoietic cells from human ES cells opens up profound
therapeutic possibilities (Figure 1). Currently, bone marrow transplantation (BMT) can
cure many hematopoietic diseases, but HSC availability is often limiting and many
patients lack matched tissue donors. Human ES cells offer a potentially renewable source
of tissue-compatible HSCs for BMT. In addition, compared to somatic HSCs used
currently for BMT, human ES cells are potentially more amenable to genetic
manipulation for rescuing specific defects or for altering antigenicity to augment
transplantation. Eventually, it should be possible to use SCNT to generate individualized
ES cell lines from patients with various inherited or acquired disorders, treat the defect by
genetic manipulation using homologous recombination, lentiviral or siRNA approaches,
and then generate transplantable HSCs by in vitro differentiation. However, numerous
unsolved questions related to the biology of ES cells derived from adult somatic tissues
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 24
must be further investigated. These include the nature of epigenetic influences such as
X-chromosome inactivation and genomic imprinting, telomere status and genomic
stability5.
It is also possible that replacement therapies using mature hematopoietic lineages derived
from human ES cells could be of clinical utility. For example, in vitro manipulated
human DCs are being explored as a mechanism to tolerize the immune system against
autoimmune disease or to stimulate anti-tumor immune responses. However, the number
of DCs that can be isolated from patients using current technologies is limiting180. Using
human ES cells as a source for generating DCs could circumvent this problem.
Additionally, ES cells appear to be a particularly robust source for expanding erythroid
progenitors compared to fetal liver and adult tissues130. By merging several established
technologies, it may be possible to produce large quantities of mature red blood cells
from human ES cells181. In principle, this represents a limitless source for erythrocyte
transfusions and for laboratory reagents used in blood bank testing. This may be
especially useful for rare red blood cell types with limited donor availability. In addition,
it may be possible through genetic manipulation to design improved universal donor
erythrocytes with a limited antigenic repertoire. Similarly, functional granulocytes and
megakaryocytes or their committed progenitors could be used to treat neutropenia and
thrombocytopenia, respectively. One potentially interesting application for the latter
would be to manipulate hemostasis by engineering hES cells to produce modified
platelets that deliver ectopically expressed pro or anticoagulant factors directly to the site
of blood clots182,183.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 25
Several additional problems must be further addressed to facilitate the transit of ES cells
from bench to bedside. Like all organs, hematopoietic tissues develop in the context of a
complex and poorly understood three-dimensional microenvironment comprised of
paracrine factors, stromal contacts and physical forces that vary during development.
Defining these microenvironments and recapitulating them in vitro may provide a key to
more efficiently recapitulating hematopoiesis, particularly HSC generation. Along the
same lines, continued refinement of serum free and stroma free culture systems will
minimize pathogen contamination and provide tighter control of cellular differentiation.
It is also critical to develop new approaches for testing ES cell-based therapies. For
example, transplantation into NOD/SCIDS immunodeficient mice may not be the most
sensitive or effective method to evaluate ES-cell derived human hematopoietic cells. To
gain more information on the safety and utility of ES cell-based therapies, it is important
to use model systems that more closely approximate humans. Along these lines, several
groups are examining hematopoietic development from non-human primate ES cells184-
188. In general, these lines exhibit hematopoiesis similar to what is observed from human
ES cells. While these studies are relatively preliminary, continued investigation of non-
human primate ES cells will provide improved preclinical models for ES cell-derived
hematopoietic transplantation therapies.
In addition to therapeutic applications, new technologies for making blood from human
ES cells will shed light on the biology of hematopoietic ontogeny, which is difficult to
study in the human embryo for ethical and practical reasons. Due to inherent
developmental differences, not all conclusions drawn from murine ES cells will be
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 26
applicable to the human embryo. Comparative studies to examine the onset of
hematopoiesis in human and murine ES cell differentiation cultures should elucidate
fundamental similarities and differences between the two species.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 27
Acknowledgements
We thank Gerd Blobel, Gordon Keller, Marcela Maus and Toru Nakano for comments
and suggestions on the manuscript.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 28
References
1. Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature. 1981;292:154-156
2. Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci U S A. 1981;78:7634-7638
3. Bradley A, Evans M, Kaufman MH, Robertson E. Formation of germ-line chimaeras from embryo-derived teratocarcinoma cell lines. Nature. 1984;309:255-256
4. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145-1147
5. Hwang WS, Roh SI, Lee BC, Kang SK, Kwon DK, Kim S, Kim SJ, Park SW, Kwon HS, Lee CK, Lee JB, Kim JM, Ahn C, Paek SH, Chang SS, Koo JJ, Yoon HS, Hwang JH, Hwang YY, Park YS, Oh SK, Kim HS, Park JH, Moon SY, Schatten G. Patient-specific embryonic stem cells derived from human SCNT blastocysts. Science. 2005;308:1777-1783
6. Li T, Zhou CQ, Mai QY, Zhuang GL. Establishment of human embryonic stem cell line from gamete donors. Chin Med J (Engl). 2005;118:116-122
7. Rao M. Conserved and divergent paths that regulate self-renewal in mouse and human embryonic stem cells. Dev Biol. 2004;275:269-286
8. Chambers I, Smith A. Self-renewal of teratocarcinoma and embryonic stem cells. Oncogene. 2004;23:7150-7160
9. Williams RL, Hilton DJ, Pease S, Willson TA, Stewart CL, Gearing DP, Wagner EF, Metcalf D, Nicola NA, Gough NM. Myeloid leukaemia inhibitory factor maintains the developmental potential of embryonic stem cells. Nature. 1988;336:684-687
10. Ying QL, Nichols J, Chambers I, Smith A. BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell. 2003;115:281-292
11. Daheron L, Opitz SL, Zaehres H, Lensch WM, Andrews PW, Itskovitz-Eldor J, Daley GQ. LIF/STAT3 signaling fails to maintain self-renewal of human embryonic stem cells. Stem Cells. 2004;22:770-778
12. Humphrey RK, Beattie GM, Lopez AD, Bucay N, King CC, Firpo MT, Rose-John S, Hayek A. Maintenance of pluripotency in human embryonic stem cells is STAT3 independent. Stem Cells. 2004;22:522-530
13. Pera MF, Trounson AO. Human embryonic stem cells: prospects for development. Development. 2004;131:5515-5525
14. Perkel J. Fifteen tips and tricks for human embryonic stem cells. The Scientist. 2005;19:18-19
15. Lewis R. Stem Cells: An emerging portrait: Struggling to sharpen the lines that define embryonic stem cells. The Scientist. 2005;19:15-17
16. Lee JB, Song JM, Lee JE, Park JH, Kim SJ, Kang SM, Kwon JN, Kim MK, Roh SI, Yoon HS. Available human feeder cells for the maintenance of human embryonic stem cells. Reproduction. 2004;128:727-735
17. Choo AB, Padmanabhan J, Chin AC, Oh SK. Expansion of pluripotent human embryonic stem cells on human feeders. Biotechnol Bioeng. 2004;88:321-331
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 29
18. Wang L, Li L, Menendez P, Cerdan C, Bhatia M. Human embryonic stem cells maintained in the absence of mouse embryonic fibroblasts or conditioned media are capable of hematopoietic development. Blood. 2005;105:4598-4603
19. Beattie GM, Lopez AD, Bucay N, Hinton A, Firpo MT, King CC, Hayek A. Activin A maintains pluripotency of human embryonic stem cells in the absence of feeder layers. Stem Cells. 2005;23:489-495
20. Rosler ES, Fisk GJ, Ares X, Irving J, Miura T, Rao MS, Carpenter MK. Long-term culture of human embryonic stem cells in feeder-free conditions. Dev Dyn. 2004;229:259-274
21. Klimanskaya I, Chung Y, Meisner L, Johnson J, West MD, Lanza R. Human embryonic stem cells derived without feeder cells. Lancet. 2005;365:1636-1641
22. Wei CL, Miura T, Robson P, Lim SK, Xu XQ, Lee MY, Gupta S, Stanton L, Luo Y, Schmitt J, Thies S, Wang W, Khrebtukova I, Zhou D, Liu ET, Ruan YJ, Rao M, Lim B. Transcriptome profiling of human and murine ESCs identifies divergent paths required to maintain the stem cell state. Stem Cells. 2005;23:166-185
23. Sato N, Meijer L, Skaltsounis L, Greengard P, Brivanlou AH. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat Med. 2004;10:55-63
24. Dravid G, Ye Z, Hammond H, Chen G, Pyle A, Donovan P, Yu X, Cheng L. Defining the role of Wnt/{beta}-catenin signaling in the survival, proliferation and self-renewal of human embryonic stem cells. Stem Cells. 2005
25. Niwa H, Miyazaki J, Smith AG. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nat Genet. 2000;24:372-376
26. Chambers I, Colby D, Robertson M, Nichols J, Lee S, Tweedie S, Smith A. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell. 2003;113:643-655
27. Carpenter MK, Rosler ES, Fisk GJ, Brandenberger R, Ares X, Miura T, Lucero M, Rao MS. Properties of four human embryonic stem cell lines maintained in a feeder-free culture system. Dev Dyn. 2004;229:243-258
28. Boyer LA, Lee TI, Cole MF, Johnstone SE, Levine SS, Zucker JP, Guenther MG, Kumar RM, Murray HL, Jenner RG, Gifford DK, Melton DA, Jaenisch R, Young RA. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell. 2005;122:947-956
29. Skottman H, Stromberg AM, Matilainen E, Inzunza J, Hovatta O, Lahesmaa R. Unique gene expression signature by human embryonic stem cells cultured under serum free conditions correlates with their enhanced and prolonged growth in an undifferentiated stage. Stem Cells. 2005
30. Skottman H, Mikkola M, Lundin K, Olsson C, Stromberg AM, Tuuri T, Otonkoski T, Hovatta O, Lahesmaa R. Gene expression signatures of seven individual human embryonic stem cell lines. Stem Cells. 2005
31. Miura T, Luo Y, Khrebtukova I, Brandenberger R, Zhou D, Thies RS, Vasicek T, Young H, Lebkowski J, Carpenter MK, Rao MS. Monitoring early differentiation events in human embryonic stem cells by massively parallel signature sequencing and expressed sequence tag scan. Stem Cells Dev. 2004;13:694-715
32. Brandenberger R, Khrebtukova I, Thies RS, Miura T, Jingli C, Puri R, Vasicek T, Lebkowski J, Rao M. MPSS profiling of human embryonic stem cells. BMC Dev Biol. 2004;4:10
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 30
33. Abeyta MJ, Clark AT, Rodriguez RT, Bodnar MS, Pera RA, Firpo MT. Unique gene expression signatures of independently-derived human embryonic stem cell lines. Hum Mol Genet. 2004;13:601-608
34. Sato N, Sanjuan IM, Heke M, Uchida M, Naef F, Brivanlou AH. Molecular signature of human embryonic stem cells and its comparison with the mouse. Dev Biol. 2003;260:404-413
35. Suarez-Farinas M, Noggle S, Heke M, Hemmati-Brivanlou A, Magnasco MO. Comparing independent microarray studies: the case of human embryonic stem cells. BMC Genomics. 2005;6:99
36. Bhattacharya B, Miura T, Brandenberger R, Mejido J, Luo Y, Yang AX, Joshi BH, Ginis I, Thies RS, Amit M, Lyons I, Condie BG, Itskovitz-Eldor J, Rao MS, Puri RK. Gene expression in human embryonic stem cell lines: unique molecular signature. Blood. 2004;103:2956-2964
37. Richards M, Tan SP, Tan JH, Chan WK, Bongso A. The transcriptome profile of human embryonic stem cells as defined by SAGE. Stem Cells. 2004;22:51-64
38. Kitajima K, Tanaka M, Zheng J, Sakai-Ogawa E, Nakano T. In vitro differentiation of mouse embryonic stem cells to hematopoietic cells on an OP9 stromal cell monolayer. Methods Enzymol. 2003;365:72-83
39. Kennedy M, Keller GM. Hematopoietic commitment of ES cells in culture. Methods Enzymol. 2003;365:39-59
40. Fraser ST, Yamashita J, Jakt LM, Okada M, Ogawa M, Nishikawa S. In vitro differentiation of mouse embryonic stem cells: hematopoietic and vascular cell types. Methods Enzymol. 2003;365:59-72
41. Nakayama N, Lee J, Chiu L. Vascular endothelial growth factorsynergistically enhances bone morphogenetic protein-4-dependent lymphohematopoietic cell generation from embryonic stem cells in vitro. Blood. 2000;95:2275-2283
42. Park C, Afrikanova I, Chung YS, Zhang WJ, Arentson E, Fong Gh G, Rosendahl A, Choi K. A hierarchical order of factors in the generation of FLK1- and SCL-expressing hematopoietic and endothelial progenitors from embryonic stem cells. Development. 2004;131:2749-2762
43. Qi X, Li TG, Hao J, Hu J, Wang J, Simmons H, Miura S, Mishina Y, Zhao GQ. BMP4 supports self-renewal of embryonic stem cells by inhibiting mitogen-activated protein kinase pathways. Proc Natl Acad Sci U S A. 2004;101:6027-6032
44. Chadwick K, Wang L, Li L, Menendez P, Murdoch B, Rouleau A, Bhatia M. Cytokines and BMP-4 promote hematopoietic differentiation of human embryonic stem cells. Blood. 2003;102:906-915
45. De Robertis EM, Kuroda H. Dorsal-ventral patterning and neural induction in Xenopus embryos. Annu Rev Cell Dev Biol. 2004;20:285-308
46. Tiedemann H, Asashima M, Grunz H, Knochel W. Pluripotent cells (stem cells) and their determination and differentiation in early vertebrate embryogenesis. Dev Growth Differ. 2001;43:469-502
47. Keller G, Kennedy M, Papayannopoulou T, Wiles MV. Hematopoietic commitment during embryonic stem cell differentiation in culture. Mol Cell Biol. 1993;13:473-486
48. Doetschman TC, Eistetter H, Katz M, Schmidt W, Kemler R. The in vitro development of blastocyst-derived embryonic stem cell lines: formation of visceral yolk sac, blood islands and myocardium. J Embryol Exp Morphol. 1985;87:27-45
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 31
49. Nakano T, Kodama H, Honjo T. Generation of lymphohematopoietic cells from embryonic stem cells in culture. Science. 1994;265:1098-1101
50. Wiles MV, Keller G. Multiple hematopoietic lineages develop from embryonic stem (ES) cells in culture. Development. 1991;111:259-267
51. Burkert U, vonRuden RT, Wagner EF. Early fetal hematopoietic development from in vitro differentiated embryonic stem cells. New Biol. 1991;3:698-708
52. Schmitt RM, Bruyns E, Snodgrass HR. Hematopoietic development of embryonic stem cells in vitro: cytokine and receptor gene expression. Genes Dev. 1991;5:728-740
53. Ling V, Neben S. In vitro differentiation of embryonic stem cells: immunophenotypic analysis of cultured embryoid bodies. J. Cell. Physiol. 1997;171
54. Mitjavila-Garcia MT, Cailleret M, Godin I, Nogueira MM, Cohen-Solal K, Schiavon V, Lecluse Y, Le Pesteur F, Lagrue AH, Vainchenker W. Expression of CD41 on hematopoietic progenitors derived from embryonic hematopoietic cells. Development. 2002;129:2003-2013
55. Nakano T. In vitro development of hematopoietic system from mouse embryonic stem cells: a new approach for embryonic hematopoiesis. Int J Hematol. 1996;65:1-8 56. Zhang WJ, Park C, Arentson E, Choi K. Modulation of hematopoietic and endothelial cell differentiation from mouse embryonic stem cells by different culture conditions. Blood. 2005;105:111-114
57. Wang J, Zhao HP, Lin G, Xie CQ, Nie DS, Wang QR, Lu GX. In vitro hematopoietic differentiation of human embryonic stem cells induced by co-culture with human bone marrow stromal cells and low dose cytokines. Cell Biol Int. 2005
58. Bilko NM, Votyakova IA, Vasylovska SV, Bilko DI. Characterization of the interactions between stromal and haematopoietic progenitor cells in expansion cell culture models. Cell Biol Int. 2005;29:83-86
59. Levenberg S, Huang NF, Lavik E, Rogers AB, Itskovitz-Eldor J, Langer R. Differentiation of human embryonic stem cells on three-dimensional polymer scaffolds. Proc Natl Acad Sci U S A. 2003;100:12741-12746
60. Gerecht-Nir S, Cohen S, Ziskind A, Itskovitz-Eldor J. Three-dimensional porous alginate scaffolds provide a conducive environment for generation of well-vascularized embryoid bodies from human embryonic stem cells. Biotechnol Bioeng. 2004;88:313-320
61. Liu H, Roy K. Biomimetic three-dimensional cultures significantly increase hematopoietic differentiation efficacy of embryonic stem cells. Tissue Eng. 2005;11:319-330
62. Ueno H, Sakita-Ishikawa M, Morikawa Y, Nakano T, Kitamura T, Saito M. A stromal cell-derived membrane protein that supports hematopoietic stem cells. Nat Immunol. 2003;4:457-463
63. Palis J, Robertson S, Kennedy M, Wall C, Keller G. Development of erythroid and myeloid progenitors in the yolk sac and embryo proper of the mouse. Development. 1999;126:5073-5084
64. Brotherton TW, Chui DHK, Gauldie J, Patterson M. Hemoglobin ontogeny during normal mouse fetal development. Proc. Natl. Acad. Sci. 1979;76:2853-2857
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 32
65. Russel E. Hereditary anemias of the mouse: A review for geneticists. Adv. Genet. 1979;20:357-459
66. Wood WG. Erythropoiesis and haemoglobin production during development. In: Jones CT, ed. Biochemical development of the fetus and neonate. New York: Elsevier Biomedical Press; 1982:127-162
67. Muller AM, Medvinsky A, Strouboulis J, Grosveld F, Dzierzak E. Development of hematopoietic stem cell activity in the mouse embryo. Immunity. 1994;1:291-301
68. Toles JF, Chui DHK, Belbeck LW, Starr E, Barker JE. Hematopoietic stem cells in murine embryonic yolk sac and peripheral blood. Proc. Natl. Acad. Sci. USA. 1989;86:7456-7459
69. Moore MAS, Metcalf D. Ontogeny of the haemopoietic system: yolk sac origin of in vivo and in vitro colony forming cells in the developing embryo. British J. Hematol. 1970;18:279-296
70. Haar JL, Ackerman GA. A phase and electron microscopic study of vasculogenesis and erythropoiesis in the yolk sac of the mouse. Anat. Rec. 1971;170:199-224
71. Sabin FR. Studies on the origin of the blood vessels and of red blood corpuscles as seen in the living blastoderm of chick during the second day of incubation. Contrib. Embryol. 1920;9:215-262
72. Murray PDF. The development of in vitro of the blood of the early chick embryo. Proc. Roy. Soc. London. 1932;11:497-521
73. Kennedy M, Firpo M, Choi K, Wall C, Robertson S, Kabrun N, Keller G. A common precursor for primitive erythropoiesis and definitive haematopoiesis. Nature. 1997;386:488-493
74. Choi K, Kennedy M, Kazarov A, Papadimitriou JC, Keller G. A common precursor for hematopoietic and endothelial cells. Development. 1998;125:725-732
75. Nishikawa SI, Nishikawa S, Hirashima M, Matsuyoshi N, Kodama H. Progressive lineage analysis by cell sorting and culture identifies FLK1+VE-cadherin+ cells at a diverging point of endothelial and hemopoietic lineages. Development. 1998;125:1747-1757
76. Robertson SM, Kennedy M, Shannon JM, Keller G. A transitional stage in the commitment of mesoderm to hematopoiesis requiring the transcription factor SCL/tal-1. Development. 2000;127:2447-2459
77. D'Souza SL, Elefanty AG, Keller G. SCL/Tal-1 is essential for hematopoietic commitment of the hemangioblast but not for its development. Blood. 2005;105:3862-3870
78. Ng ES, Azzola L, Sourris K, Robb L, Stanley EG, Elefanty AG. The primitive streak gene Mixl1 is required for efficient haematopoiesis and BMP4-induced ventral mesoderm patterning in differentiating ES cells. Development. 2005;132:873-884
79. Hidaka M, Stanford WL, Bernstein A. Conditional requirement for the Flk-1 receptor in the in vitro generation of early hematopoietic cells. Proc Natl Acad Sci U S A. 1999;96:7370-7375
80. Chan RJ, Johnson SA, Li Y, Yoder MC, Feng GS. A definitive role of Shp-2 tyrosine phosphatase in mediating embryonic stem cell differentiation and hematopoiesis. Blood. 2003;102:2074-2080
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 33
81. Lacaud G, Gore L, Kennedy M, Kouskoff V, Kingsley P, Hogan C, Carlsson L, Speck N, Palis J, Keller G. Runx1 is essential for hematopoietic commitment at the hemangioblast stage of development in vitro. Blood. 2002;100:458-466
82. Huber TL, Kouskoff V, Fehling HJ, Palis J, Keller G. Haemangioblast commitment is initiated in the primitive streak of the mouse embryo. Nature. 2004;432:625-630
83. Hole N, Graham GJ, Menzel U, Ansell JD. A limited temporal window for the derivation of multilineage repopulating hematopoietic progenitors during embryonal stem cell differentiation in vitro. Blood. 1996;88:1266-1276
84. Muller AM, Dzierzak EA. ES cells have only a limited lymphopoietic potential after adoptive transfer into mouse recipients. Development. 1993;118:1343-1351
85. Burt RK, Verda L, Kim DA, Oyama Y, Luo K, Link C. Embryonic stem cells as an alternate marrow donor source: engraftment without graft-versus-host disease. J Exp Med. 2004;199:895-904
86. Keller G. Embryonic stem cell differentiation: emergence of a new era in biology and medicine. Genes Dev. 2005;19:1129-1155
87. Lengerke C, Daley GQ. Patterning definitive hematopoietic stem cells from embryonic stem cells. Exp Hematol. 2005;33:971-979
88. Yoder MC, King B, Hiatt K, Williams DA. Murine embryonic yolk sac cells promote in vitro proliferation of bone marrow high proliferative potential colony-forming cells. Blood. 1995;86:1322-1330
89. Cumano A, Dieterlen-Lievre F, Godin I. Lymphoid potential, probed before circulation in mouse, is restricted to caudal intraembryonic splanchnopleura. Cell. 1996;86:907-916
90. Medvinsky AL, Samoylina NL, Muller AM, Dzierzak EA. An early pre-liver intraembryonic source of CFU-S in the developing mouse. Nature. 1993;364:64-66
91. Medvinsky A, Dzierzak E. Definitive hematopoiesis is autonomously initiated by the AGM region. Cell. 1996;86:897-906
92. Cumano A, Ferraz JC, Klaine M, Di Santo JP, Godin I. Intraembryonic, but not yolk sac hematopoietic precursors, isolated before circulation, provide long-term multilineage reconstitution. Immunity. 2001;15:477-485
93. Perlingeiro RC, Kyba M, Daley GQ. Clonal analysis of differentiating embryonic stem cells reveals a hematopoietic progenitor with primitive erythroid and adult lymphoid-myeloid potential. Development. 2001;128:4597-4604
94. Schuringa JJ, Wu K, Morrone G, Moore MA. Enforced activation of STAT5A facilitates the generation of embryonic stem-derived hematopoietic stem cells that contribute to hematopoiesis in vivo. Stem Cells. 2004;22:1191-1204
95. Shimamoto T, Tang Y, Naot Y, Nardi M, Brulet P, Bieberich CJ, Takeshita K. Hematopoietic progenitor cell abnormalities in Hoxc-8 null mutant mice. J Exp Zool. 1999;283:186-193
96. Lawrence HJ, Sauvageau G, Humphries RK, Largman C. The role of HOX homeobox genes in normal and leukemic hematopoiesis. Stem Cells. 1996;14:281-291
97. Abramovich C, Humphries RK. Hox regulation of normal and leukemic hematopoietic stem cells. Curr Opin Hematol. 2005;12:210-216
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 34
98. van Oostveen J, Bijl J, Raaphorst F, Walboomers J, Meijer C. The role of homeobox genes in normal hematopoiesis and hematological malignancies. Leukemia. 1999;13:1675-1690
99. Bjornsson JM, Larsson N, Brun AC, Magnusson M, Andersson E, Lundstrom P, Larsson J, Repetowska E, Ehinger M, Humphries RK, Karlsson S. Reduced proliferative capacity of hematopoietic stem cells deficient in Hoxb3 and Hoxb4. Mol Cell Biol. 2003;23:3872-3883
100. Ernst P, Mabon M, Davidson AJ, Zon LI, Korsmeyer SJ. An Mll-dependent Hox program drives hematopoietic progenitor expansion. Curr Biol. 2004;14:2063-2069
101. Antonchuk J, Sauvageau G, Humphries RK. HOXB4 overexpression mediates very rapid stem cell regeneration and competitive hematopoietic repopulation. Exp Hematol. 2001;29:1125-1134
102. Antonchuk J, Sauvageau G, Humphries RK. HOXB4-induced expansion of adult hematopoietic stem cells ex vivo. Cell. 2002;109:39-45
103. Sauvageau G, Thorsteinsdottir U, Eaves CJ, Lawrence HJ, Largman C, Lansdorp PM, Humphries RK. Overexpression of HOXB4 in hematopoietic cells causes the selective expansion of more primitive populations in vitro and in vivo. Genes Dev. 1995;9:1753-1765
104. Buske C, Feuring-Buske M, Abramovich C, Spiekermann K, Eaves CJ,Coulombel L, Sauvageau G, Hogge DE, Humphries RK. Deregulated expression of HOXB4 enhances the primitive growth activity of human hematopoietic cells. Blood. 2002;100:862-868
105. Helgason CD, Sauvageau G, Lawrence HJ, Largman C, Humphroes RK. Overexpression of HOXB4 enhances the hematopoietic potential of embryonic stem cells differentiated in vitro. Blood. 1996;87:2740-2749
106. Kyba M, Perlingeiro RC, Daley GQ. HoxB4 confers definitive lymphoid-myeloid engraftment potential on embryonic stem cell and yolk sac hematopoietic progenitors. Cell. 2002;109:29-37
107. Pilat S, Carotta S, Schiedlmeier B, Kamino K, Mairhofer A, Will E, Modlich U, Steinlein P, Ostertag W, Baum C, Beug H, Klump H. HOXB4 enforces equivalent fates of ES-cell-derived and adult hematopoietic cells. Proc Natl Acad Sci U S A. 2005;102:12101-12106
108. Beslu N, Krosl J, Laurin M, Mayotte N, Humphries KR, Sauvageau G. Molecular interactions involved in HOXB4-induced activation of HSC self-renewal. Blood. 2004;104:2307-2314
109. Ferrell CM, Dorsam ST, Ohta H, Humphries RK, Derynck MK, Haqq C, Largman C, Lawrence HJ. Activation of stem-cell specific genes by HOXA9 and HOXA10 homeodomain proteins in CD34+ human cord blood cells. Stem Cells. 2005;23:644-655
110. Takeuchi M, Sekiguchi T, Hara T, Kinoshita T, Miyajima A. Cultivation of aorta-gonad-mesonephros-derived hematopoietic stem cells in the fetal liver microenvironment amplifies long-term repopulating activity and enhances engraftment to the bone marrow. Blood. 2002;99:1190-1196
111. Mukouyama Y, Hara T, Xu M, Tamura K, Donovan PJ, Kim H, Kogo H, Tsuji K, Nakahata T, Miyajima A. In vitro expansion of murine multipotential hematopoietic progenitors from the embryonic aorta-gonad-mesonephros region. Immunity. 1998;8:105-114
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 35
112. Oostendorp RA, Harvey KN, Kusadasi N, de Bruijn MF, Saris C, Ploemacher RE, Medvinsky AL, Dzierzak EA. Stromal cell lines from mouse aorta-gonads-mesonephros subregions are potent supporters of hematopoietic stem cell activity. Blood. 2002;99:1183-1189
113. Kusadasi N, Oostendorp RA, Koevoet WJ, Dzierzak EA, Ploemacher RE. Stromal cells from murine embryonic aorta-gonad-mesonephros region, liver and gut mesentery expand human umbilical cord blood-derived CAFC(week6) in extended long-term cultures. Leukemia. 2002;16:1782-1790
114. Matsuoka S, Tsuji K, Hisakawa H, Xu M, Ebihara Y, Ishii T, Sugiyama D, Manabe A, Tanaka R, Ikeda Y, Asano S, Nakahata T. Generation of definitive hematopoietic stem cells from murine early yolk sac and paraaortic splanchnopleures by aorta-gonad-mesonephros region-derived stromal cells. Blood. 2001;98:6-12
115. Oostendorp RA, Robin C, Steinhoff C, Marz S, Brauer R, Nuber UA, Dzierzak EA, Peschel C. Long-term maintenance of hematopoietic stem cells does not require contact with embryo-derived stromal cells in cocultures. Stem Cells. 2005;23:842-851
116. Nakano T, Kodama H, Honjo T. In vitro development of primitive and definitive erythrocytes from different precursors. Science. 1996;272:722-724
117. Weiss MJ, Keller G, Orkin SH. Novel insights into erythroid development revealed through in vitro differentiation of GATA-1- embryonic stem cells. Genes and Dev. 1994;8:1184-1197
118. Weiss MJ, Orkin SH. Transcription factor GATA-1 permits survival and maturation of erythroid precursors by preventing apoptosis. Proc. Natl. Acad. Sci. 1995;92:9623-9627
119. Weiss MJ, Yu C, Orkin SH. Erythroid-cell-specific properties of transcription factor GATA-1 revealed by phenotypic rescue of a gene-targeted cell line. Mol Cell Biol. 1997;17:1642-1651
120. Gregory T, Yu C, Ma A, Orkin SH, Blobel GA, Weiss MJ. GATA-1 and erythropoietin cooperate to promote erythroid cell survival by regulating bcl-xL expression. Blood. 1999;94:87-96
121. Kihm AJ, Kong Y, Hong W, Russell JE, Rouda S, Adachi K, Simon MC, Blobel GA, Weiss MJ. An abundant erythroid protein that stabilizes free alpha hemoglobin. Nature. 2002;417:758-763
122. Shirihai OS, Gregory T, Yu C, Orkin SH, Weiss MJ. ABC-me, a novel mitochondrial transporter induced by GATA-1 during erythroid differentiation. EMBO J. 2000;19:2492-2502
123. Rylski M, Welch JJ, Chen Y-Y, Letting DL, Diehl JA, Chodosh LA, Blobel GA, Weiss MJ. GATA-1-Mediated Proliferation Arrest During Erythroid Maturation. Mol Cell Biol. 2003;23:5031-5042
124. Vakoc CR, Letting DL, Gheldof N, Sawado T, Bender MA, Groudine M, Weiss MJ, Dekker J, Blobel GA. Proximity among distant regulatory elements at the beta-globin locus requires GATA-1 and FOG-1. Mol Cell. 2005;17:453-462
125. Johnson KD, Grass JA, Boyer ME, Kiekhaefer CM, Blobel GA, Weiss MJ, Bresnick EH. Cooperative activities of hematopoietic regulators recruit RNA polymerase II to a tissue-specific chromatin domain. Proc Natl Acad Sci U S A. 2002;99:11760-11765
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 36
126. Kapur R, Cooper R, Xiao X, Weiss MJ, Donovan P, Williams DA. The presence of novel amino acids in the cytoplasmic domain of stem cell factor results in hematopoietic defects in Steel(17H) mice. Blood. 1999;94:1915-1925.
127. Kong Y, Zhou S, Kihm AJ, Katein AM, Yu X, Gell DA, Mackay JP, AdachiK, Foster-Brown L, Louden CS, Gow AJ, Weiss MJ. Loss of alpha-hemoglobin-stabilizing protein impairs erythropoiesis and exacerbates beta-thalassemia. J Clin Invest. 2004;114:1457-1466
128. Munugalavadla V, Dore LC, Tan BL, Hong L, Vishnu M, Weiss MJ, Kapur R. Repression of c-kit and its downstream substrates by GATA-1 inhibits cell proliferation during erythroid maturation. Mol Cell Biol. 2005;25:6747-6759
129. Zheng J, Kitajima K, Sakai E, Kimura T, Minegishi N, Yamamoto M, Nakano T. Differential effects of GATA-1 on proliferation and differentiation of erythroid lineage cells. Blood. 2005;In press
130. Carotta S, Pilat S, Mairhofer A, Schmidt U, Dolznig H, Steinlein P, Beug H. Directed differentiation and mass cultivation of pure erythroid progenitors from mouse embryonic stem cells. Blood. 2004;104:1873-1880
131. von Lindern M, Zauner W, Mellitzer G, Steinlein P, Fritsch G, Huber K, Lowenberg B, Beug H. The glucocorticoid receptor cooperates with the erythropoietin receptor and c-Kit to enhance and sustain proliferation of erythroid progenitors in vitro. Blood. 1999;94:550-559
132. Wessely O, Bauer A, Quang CT, Deiner EM, von Lindern M, Mellitzer G, Steinlein P, Ghysdael J, Beug H. A novel way to induce erythroid progenitor self renewal: cooperation of c-Kit with the erythropoietin receptor. Biol Chem. 1999;380:187-202
133. Wessely O, Deiner EM, Beug H, von Lindern M. The glucocorticoid receptor is a key regulator of the decision between self-renewal and differentiation in erythroid progenitors. Embo J. 1997;16:267-280
134. Shalaby F, Rossant J, Yamaguchi TP, Gertsenstein M, Wu XF, Breitman ML, Schuh AC. Failure of blood-island formation and vasculogenesis in Flk-1-deficient mice. Nature. 1995;376:62-66
135. Fujimoto TT, Kohata S, Suzuki H, Miyazaki H, Fujimura K. Production of functional platelets by differentiated embryonic stem (ES) cells in vitro. Blood. 2003;102:4044-4051
136. Tanaka M, Zheng J, Kitajima K, Kita K, Yoshikawa H, Nakano T. Differentiation status dependent function of FOG-1. Genes Cells. 2004;9:1213-1226
137. Eto K, Leavitt AL, Nakano T, Shattil SJ. Development and analysis of megakaryocytes from murine embryonic stem cells. Methods Enzymol. 2003;365:142-158
138. Eto K, Murphy R, Kerrigan SW, Bertoni A, Stuhlmann H, Nakano T, Leavitt AD, Shattil SJ. Megakaryocytes derived from embryonic stem cells implicate CalDAG-GEFI in integrin signaling. Proc Natl Acad Sci U S A. 2002;99:12819-12824
139. Era T, Takagi T, Takahashi T, Bories JC, Nakano T. Characterization of hematopoietic lineage-specific gene expression by ES cell in vitro differentiation induction system. Blood. 2000;95:870-878
140. Stachura DL, Chou ST, J. WM. An early block to erythro-megakaryocytic development conferred by loss of transcription factor GATA-1. Blood. 2005;In press
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 37
141. Hitzler JK, Zipursky A. Origins of leukaemia in children with Down syndrome. Nat Rev Cancer. 2005;5:11-20
142. Lieber JG, Webb S, Suratt BT, Young SK, Johnson GL, Keller GM, Worthen GS. The in vitro production and characterization of neutrophils from embryonic stem cells. Blood. 2004;103:852-859
143. Lieber JG, Keller GM, Worthen GS. The in vitro differentiation of mouse embryonic stem cells into neutrophils. Methods Enzymol. 2003;365:129-142
144. Tsai M, Wedemeyer J, Ganiatsas S, Tam SY, Zon LI, Galli SJ. In vivo immunological function of mast cells derived from embryonic stem cells: an approach for the rapid analysis of even embryonic lethal mutations in adult mice in vivo. Proc Natl Acad Sci U S A. 2000;97:9186-9190
145. Tsai M, Tam SY, Wedemeyer J, Galli SJ. Mast cells derived from embryonic stem cells: a model system for studying the effects of genetic manipulations on mast cell development, phenotype, and function in vitro and in vivo. Int J Hematol. 2002;75:345-349
146. Garrington TP, Ishizuka T, Papst PJ, Chayama K, Webb S, Yujiri T, Sun W, Sather S, Russell DM, Gibson SB, Keller G, Gelfand EW, Johnson GL. MEKK2 gene disruption causes loss of cytokine production in response to IgE and c-Kit ligand stimulation of ES cell-derived mast cells. Embo J. 2000;19:5387-5395
147. Hamaguchi-Tsuru E, Nobumoto A, Hirose N, Kataoka S, Fujikawa-Adachi K, Furuya M, Tominaga A. Development and functional analysis of eosinophils from murine embryonic stem cells. Br J Haematol. 2004;124:819-827
148. Cho SK, Webber TD, Carlyle JR, Nakano T, Lewis SM, Zuniga-Pflucker JC. Functional characterization of B lymphocytes generated in vitro from embryonic stem cells. Proc Natl Acad Sci U S A. 1999;96:9797-9802
149. Cho SK, Zuniga-Pflucker JC. Development of lymphoid lineages from embryonic stem cells in vitro. Methods Enzymol. 2003;365:158-169
150. de Pooter RF, Cho SK, Carlyle JR, Zuniga-Pflucker JC. In vitro generation of T lymphocytes from embryonic stem cell-derived prehematopoietic progenitors. Blood. 2003;102:1649-1653
151. Schmitt TM, de Pooter RF, Gronski MA, Cho SK, Ohashi PS, Zuniga-Pflucker JC. Induction of T cell development and establishment of T cell competence from embryonic stem cells differentiated in vitro. Nat Immunol. 2004;5:410-417
152. Lindmark H, Rosengren B, Hurt-Camejo E, Bruder CE. Gene expression profiling shows that macrophages derived from mouse embryonic stem cells is an improved in vitro model for studies of vascular disease. Exp Cell Res. 2004;300:335-344
153. Moore KJ, Fabunmi RP, Andersson LP, Freeman MW. In vitro-differentiated embryonic stem cell macrophages: a model system for studying atherosclerosis-associated macrophage functions. Arterioscler Thromb Vasc Biol. 1998;18:1647-1654
154. Guillemot JC, Kruskal BA, Adra CN, Zhu S, Ko JL, Burch P, Nocka K, Seetoo K, Simons E, Lim B. Targeted disruption of guanosine diphosphate-dissociation inhibitor for Rho-related proteins, GDID4: normal hematopoietic differentiation but subtle defect in superoxide production by macrophages derived from in vitro embryonal stem cell differentiation. Blood. 1996;88:2722-2731
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 38
155. Senju S, Hirata S, Matsuyoshi H, Masuda M, Uemura Y, Araki K, Yamamura K, Nishimura Y. Generation and genetic modification of dendritic cells derived from mouse embryonic stem cells. Blood. 2003;101:3501-3508
156. Fairburn LJ, Cowling GJ, Reipert BM, Dexter TM. Suppression of apoptosis allows differentiation and development of a multipotent hemopoietic cell line in the absence of added growth factors. Cell. 1993;74:823-832
157. Fairchild PJ, Brook FA, Gardner RL, Graca L, Strong V, Tone Y, Tone M, Nolan KF, Waldmann H. Directed differentiation of dendritic cells from mouse embryonic stem cells. Curr Biol. 2000;10:1515-1518
158. Lian RH, Maeda M, Lohwasser S, Delcommenne M, Nakano T, Vance RE, Raulet DH, Takei F. Orderly and nonstochastic acquisition of CD94/NKG2 receptors by developing NK cells derived from embryonic stem cells in vitro. J Immunol. 2002;168:4980-4987
159. Yamane T, Kunisada T, Yamazaki H, Era T, Nakano T, Hayashi SI. Development of osteoclasts from embryonic stem cells through a pathway that is c-fms but not c-kit dependent. Blood. 1997;90:3516-3523
160. Yamane T, Kunisada T, Hayashi S. Embryonic stem cells as a model for studying osteoclast lineage development. Methods Mol Biol. 2002;185:97-106
161. Tsuneto M, Yamazaki H, Yoshino M, Yamada T, Hayashi S. Ascorbic acid promotes osteoclastogenesis from embryonic stem cells. Biochem Biophys Res Commun. 2005;335:1239-1246
162. Okuyama H, Tsuneto M, Yamane T, Yamazaki H, Hayashi S. Discrete types of osteoclast precursors can be generated from embryonic stem cells. Stem Cells. 2003;21:670-680
163. Hemmi H, Okuyama H, Yamane T, Nishikawa S, Nakano T, Yamazaki H, Kunisada T, Hayashi S. Temporal and spatial localization of osteoclasts in colonies from embryonic stem cells. Biochem Biophys Res Commun. 2001;280:526-534
164. Takayanagi H, Kim S, Koga T, Nishina H, Isshiki M, Yoshida H, Saiura A, Isobe M, Yokochi T, Inoue J, Wagner EF, Mak TW, Kodama T, Taniguchi T. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev Cell. 2002;3:889-901
165. Tsuneto M, Tominaga A, Yamazaki H, Yoshino M, Orkin SH, Hayashi S. Enforced expression of PU.1 rescues osteoclastogenesis from embryonic stem cells lacking Tal-1. Stem Cells. 2005;23:134-143
166. Hochedlinger K, Jaenisch R. Monoclonal mice generated by nuclear transfer from mature B and T donor cells. Nature. 2002;415:1035-1038
167. Vodyanik MA, Bork JA, Thomson JA, Slukvin I. Human embryonic stem cell-derived CD34+ cells: efficient production in the coculture with OP9 stromal cells and analysis of lymphohematopoietic potential. Blood. 2005;105:617-626
168. Kaufman DS, Hanson ET, Lewis RL, Auerbach R, Thomson JA. Hematopoietic colony-forming cells derived from human embryonic stem cells. Proc Natl Acad Sci U S A. 2001;98:10716-10721
169. Tian X, Morris JK, Linehan JL, Kaufman DS. Cytokine requirements differ for stroma and embryoid body-mediated hematopoiesis from human embryonic stem cells. Exp Hematol. 2004;32:1000-1009
170. Zambidis ET, Peault B, Park TS, Bunz F, Civin CI. Hematopoietic differentiation of human embryonic stem cells progresses through sequential
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 39
hematoendothelial, primitive, and definitive stages resembling human yolk sac development. Blood. 2005;106:860-870
171. Wang L, Li L, Shojaei F, Levac K, Cerdan C, Menendez P, Martin T, Rouleau A, Bhatia M. Endothelial and hematopoietic cell fate of human embryonic stem cells originates from primitive endothelium with hemangioblastic properties. Immunity. 2004;21:31-41
172. Odorico JS, Kaufman DS, Thomson JA. Multilineage differentiation from human embryonic stem cell lines. Stem Cells. 2001;19:193-204
173. Wang L, Menendez P, Shojaei F, Li L, Mazurier F, Dick JE, Cerdan C, Levac K, Bhatia M. Generation of hematopoietic repopulating cells from human embryonic stem cells independent of ectopic HOXB4 expression. J Exp Med. 2005;201:1603-1614
174. Yoder MC, Hiatt K. Engraftment of embryonic hematopoietic cells in conditioned newborn recipients. Blood. 1997;89:2176-2183
175. Lu SJ, Li F, Vida L, Honig GR. CD34+CD38- hematopoietic precursors derived from human embryonic stem cells exhibit an embryonic gene expression pattern. Blood. 2004;103:4134-4141
176. Lessard J, Faubert A, Sauvageau G. Genetic programs regulating HSC specification, maintenance and expansion. Oncogene. 2004;23:7199-7209
177. Kyba M, Perlingeiro RC, Hoover RR, Lu CW, Pierce J, Daley GQ. Enhanced hematopoietic differentiation of embryonic stem cells conditionally expressing Stat5. Proc Natl Acad Sci U S A. 2003;100 Suppl 1:11904-11910
178. Zhu J, Zhang Y, Joe GJ, Pompetti R, Emerson SG. NF-Ya activates multiple hematopoietic stem cell (HSC) regulatory genes and promotes HSC self-renewal. Proc Natl Acad Sci U S A. 2005
179. Krosl J, Austin P, Beslu N, Kroon E, Humphries RK, Sauvageau G. In vitro expansion of hematopoietic stem cells by recombinant TAT-HOXB4 protein. Nat Med. 2003;9:1428-1432
180. Murphy A, Westwood JA, Teng MW, Moeller M, Darcy PK, Kershaw MH. Gene modification strategies to induce tumor immunity. Immunity. 2005;22:403-414
181. Giarratana MC, Kobari L, Lapillonne H, Chalmers D, Kiger L, Cynober T, Marden MC, Wajcman H, Douay L. Ex vivo generation of fully mature human red blood cells from hematopoietic stem cells. Nat Biotechnol. 2005;23:69-74
182. Yarovoi HV, Kufrin D, Eslin DE, Thornton MA, Haberichter SL, Shi Q, Zhu H, Camire R, Fakharzadeh SS, Kowalska MA, Wilcox DA, Sachais BS, Montgomery RR, Poncz M. Factor VIII ectopically expressed in platelets: efficacy in hemophilia A treatment. Blood. 2003;102:4006-4013
183. Kufrin D, Eslin DE, Bdeir K, Murciano JC, Kuo A, Kowalska MA, Degen JL, Sachais BS, Cines DB, Poncz M. Antithrombotic thrombocytes: ectopic expression of urokinase-type plasminogen activator in platelets. Blood. 2003;102:926-933
184. Wolf DP, Kuo HC, Pau KY, Lester L. Progress with nonhuman primate embryonic stem cells. Biol Reprod. 2004;71:1766-1771
185. Umeda K, Heike T, Yoshimoto M, Shiota M, Suemori H, Luo HY, Chui DH, Torii R, Shibuya M, Nakatsuji N, Nakahata T. Development of primitive and definitive hematopoiesis from nonhuman primate embryonic stem cells in vitro. Development. 2004;131:1869-1879
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 40
186. Hematti P, Obrtlikova P, Kaufman DS. Nonhuman primate embryonic stem cells as a preclinical model for hematopoietic and vascular repair. Exp Hematol. 2005;33:980-986
187. Sasaki K, Inoue M, Shibata H, Ueda Y, Muramatsu SI, Okada T, Hasegawa M, Ozawa K, Hanazono Y. Efficient and stable Sendai virus-mediated gene transfer into primate embryonic stem cells with pluripotency preserved. Gene Ther. 2005;12:203-210
188. Asano T, Ageyama N, Takeuchi K, Momoeda M, Kitano Y, Sasaki K, Ueda Y, Suzuki Y, Kondo Y, Torii R, Hasegawa M, Ookawara S, Harii K, Terao K, Ozawa K, Hanazono Y. Engraftment and tumor formation after allogeneic in utero transplantation of primate embryonic stem cells. Transplantation. 2003;76:1061-1067
189. Fairchild PJ, Nolan KF, Waldmann H. Probing dendritic cell function by guiding the differentiation of embryonic stem cells. Methods Enzymol. 2003;365:169-186
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 41
Table 1: Generation of Specific Hematopoietic Lineages from Murine ES cells.Hematopoietic
LineageMethod
ofCulture
Cytokines References Other
Megakaryocytes and platelets
OP9, EBs Major: TPO
Other: IL-6, IL-11
39,135-139 Megarkaryocytes show agonist dependent fibrinogen binding.Platelets exhibit agonist dependent fibrinogen binding, aggregation and spreading.
Erythroid OP9, EBs Major: EPO, KL Other: Insulin, IGF-1
39,47,49,116,130
Dexamethasone used for erythroid progenitor expansion..
Primitive progenitors (EryP) are KL-independent. Definitive progenitors (EryD) require EPO/KL.
Iron saturated transferrin added to improve hemoglobin synthesis during terminal maturation.
Mast cells EBs IL-3, KL 145,146 EB-derived mast cells proliferate extensively in culture and reconstitute mast cell-deficient KitW/KitW-v mice after adoptive transfer.
Macrophages OP9, EBs IL-1, IL-3, M-CSF, GM-CSF 49,152-154 Macrophage overgrowth tends to inhibit the production of other lineages in ES cell differentiation cultures
T lymphocytes OP9OP9-DL1
Flt-3 ligand, IL-7 150,151 Full T lymphocyte development requires maturation in thymic stroma (FTOC or RTOCs).
T lymphocyte production is augmented by cultivation on OP9 cells ectopically expressing the notch ligand, DL-1.
B lymphocytes OP9 Flt-3 ligand, IL-7 49,148,149 Stimulation with LPS augments production of IgM+
secreting B-lymphocytes.
Eosinophils OP9 IL-5, IL-3, GM-CSF, eotaxin 147 Express characteristic granules and cell surface markers, produce oxidative response to Leishmania.
Neutrophils EBs, OP9 Progenitor expansion:OSM, bFGF, IL-6, IL-11, LIF, KL
Neutrophil differentiation:G-CSF, GM-CSF, IL-6
142,143 Functional neutrophils produced. This includes characteristic markers and morphology, superoxide production and chemotactic responses.
Dendritic Cells (DC)
EBs, OP9 Progenitor expansion:GM-CSF, IL-3 Differentiation/maturation:IL-4, TNFα, LPS, anti-CD40
155,157,189 DC production facilitated by plating EBs on tissue culture dishes.Exhibit characteristic surface phenotype, process protein antigens and stimulate T lymphocytes in the mixed lymphocye reaction (MLR).
Natural Killer (NK) cells
OP9 Flt-3 ligand, IL-15, IL-6, IL-7, KL
149,158 Express characteristic NK proteins, kill selected tumor lines and MHC I-deficient lymphoblasts.
Osteoclasts OP9 ST2
M-CSF, RANKL 159-163 Recognized by TRAP expression and bone resorbtion activity. Production augmented by vitamins C and D and dexamethasone.
Abbreviations: TPO, thrombopoietin; EPO, erythropoietin; OP9, M-CSF–/– stromal line; EB, embryoid bodies; IL-1, interleukin 1; IL-3, interleukin 3; IL-5, interleukin 5; IL-6, interleukin 6; IL-7, interleukin 7; IL-11, interleukin 11; G-CSF, granulocyte colony stimulating factor; M-CSF, macrophage colony stimulating factor; GM-CSF granulocyte/macrophage colony stimulating factor; IGF-1, insulin-like growth factor 1; KL, c-kit ligand; OSM, oncostatin M; bFGF, basic fibroblast growth factor; LIF, leukemia inhibitory factor; FTOC, fetal thymic organ cultures; RTOC, reaggregate thymic organ cultures; OP9-DL-1, Notch ligand Delta-like-1 expressing OP9 cells; VEGF, vascular endothelial growth factor; LPS, lipopolysaccharide; TNFα, tumor necrosis factor α; ST2, bone marrow-derived stromal line; TRAP, tartrate-resistant acid phosphatase.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom

Olsen et al. Producing blood lineages from ES cells 42
Figure legend
Figure 1. In vitro hematopoiesis from embryonic stem (ES) cells. The solid arrows
indicate established methods to produce specific hematopoietic lineages. The dashed
arrows indicate potential future therapeutic applications to be derived from human ES
cells. HSC, hematopoitic stem cell.
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom
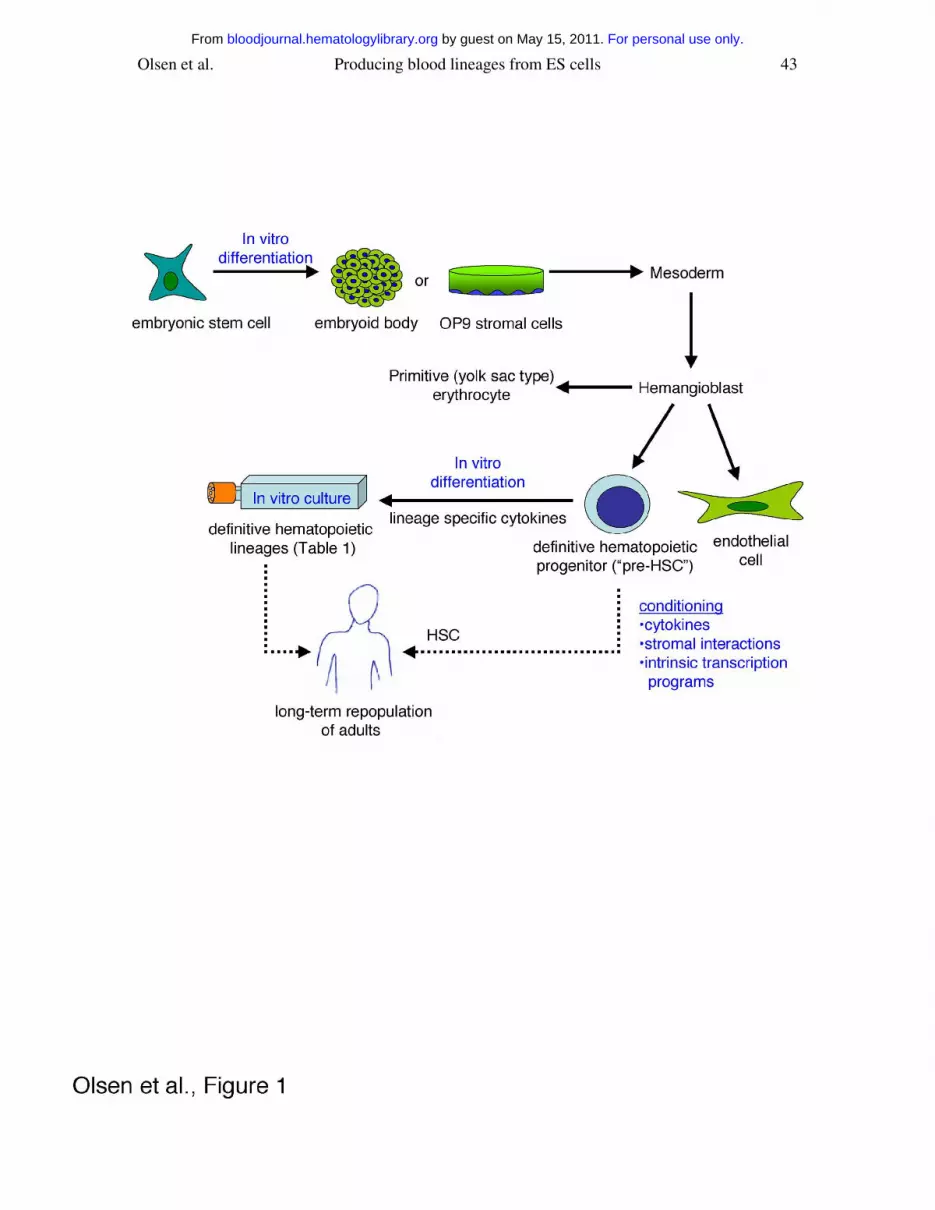
Olsen et al. Producing blood lineages from ES cells 43
For personal use only. by guest on May 15, 2011. bloodjournal.hematologylibrary.orgFrom
Recommended

















