Embed Size (px)
Citation preview

Do Salivary Antibodies Reliably ReflectBoth Mucosal and Systemic Immunity?
PER BRANDTZAEG
Laboratory for Immunohistochemistry and Immunopathology (LIIPAT),Department and Institute of Pathology, University of Oslo,Rikshospitalet-Radiumhospitalet Medical Center, N-0027 Oslo 1, Norway
ABSTRACT: Two major antibody classes operate in saliva: secretory IgA(SIgA) and IgG. The former is synthesized as dimeric IgA by plasmacells (PCs) in salivary glands and is exported by the polymeric Ig recep-tor (pIgR). Most IgG in saliva is derived from serum (mainly via gingivalcrevices), although some is locally produced. Gut-associated lymphoidtissue (GALT) and nasopharynx-associated lymphoid tissue (NALT) donot contribute equally to mucosal PCs throughout the body. Thus, en-teric immunostimulation is an inadequate mode of stimulating salivaryIgA antibodies, which are poorly associated with the intestinal SIgA re-sponse, for instance after enteric cholera vaccination. Nevertheless, theIgA response in submandibular/sublingual glands is better related toB cell induction in GALT than the parotid response. Such disparity issuggested by the elevated levels of IgA in submandibular secretions ofAIDS patients, paralleling their highly upregulated intestinal IgA sys-tem. Moreover, in patients with active celiac disease, IgA antibodies todisease-precipitating gliadin are reliably represented in whole saliva butnot in parotid secretion. Parotid SIgA may be more consistently linkedto immune induction in palatine tonsils and adenoids (human NALT),as supported by the homing molecule profile of NALT-derived B cellblasts. Also several other variables influence the levels of antibodies inoral secretions. These include difficulties with reproducibility and stan-dardization of immunoassays, the impact of flow rate, acute or chronicstress, protein loss during sample handling, and uncontrolled admixtureof serum-derived IgG and monomeric IgA. Despite such problems, salivaremains an interesting biological fluid with great scientific and clinicalpotentials.
KEYWORDS: IgA; IgG; mucosa-associated lymphoid tissue; MALT; gut-associated lymphoid tissue; GALT; nasopharynx-associated lymphoidtissue; NALT; salivary glands; crevicular fluid; polymeric Ig receptor;pIgR; secretory component; SC; vaccination; celiac disease
Address for correspondence: Prof. Per Brandtzaeg, LIIPAT, Rikshospitalet, N-0027 Oslo 1, Norway.Voice: 47-23072743; fax: 47-23071511.
Ann. N.Y. Acad. Sci. 1098: 288–311 (2007). C© 2007 New York Academy of Sciences.doi: 10.1196/annals.1384.012
288

BRANDTZAEG 289
INTRODUCTION
A molecular basis for the presence of antibodies in external body fluidsbecame available when it was shown in 1960 that salivary secretions con-tain immunoglobulin (Ig) molecules.1 Conclusive information to this end wasnot obtained, however, until specific identification of different Ig classes waspossible, and several laboratories reported that IgA predominates in many ex-ternal secretions.2 The discovery of unique properties of secretory IgA (SIgA)in 1965 further intensified investigation of local immunity.3 SIgA was shownto be polymeric (mainly dimers) and associated with an 80-kDa epithelial gly-coprotein initially called “transport piece” and later named “secretory com-ponent” (SC). Also importantly, the same year it was found that the Ig classdistribution of intestinal plasma cells (PCs) is strikingly different from thatobserved in lymph nodes and bone marrow4; in normal mucosal tissues, IgA+plasmablasts and PCs are approximately 20 times as numerous as IgG+ PCs.In 1973 our laboratory provided the first direct evidence that human mu-cosal IgA+ PCs produce mainly dimers and some larger polymers (collec-tively called pIgA) rather than monomers,5 and in 1974 this characteristic wasfound to be associated with the expression of the joining (J) chain by the samecells.6
SECRETORY IMMUNITY
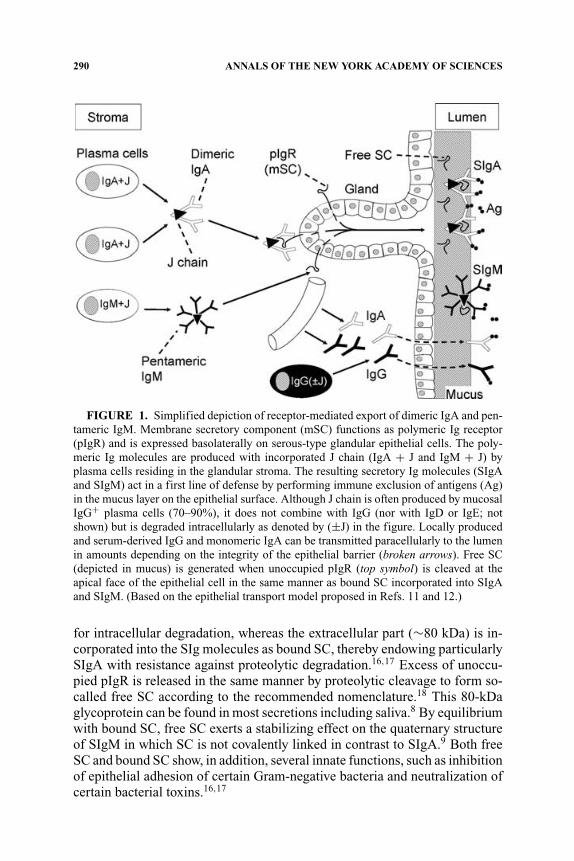
In the late 1960s our laboratory demonstrated that not only pIgA, but alsopentameric IgM is selectively exported to the secretions, apparently becauseof a common epithelial transport mechanism.7,8 Secretory IgM (SIgM) wasa few years later shown to be associated with the SC and to follow the sameintracellular route through secretory epithelia as SIgA.9,10 At the same time ashared receptor-mediated mechanism involving endocytosis and transcytosis,was proposed for the generation of SIgA and SIgM.5,6,11,12 Our transport modelwas based on the suggested crucial cooperation between J chain–expressingmucosal IgA+ and IgM+ PCs and SC-expressing serous-type of secretoryepithelial cells (FIG. 1).
Transmembrane SC is a carbohydrate-rich glycoprotein of ∼100 kDa consti-tutively expressed on the basolateral surface membrane of the epithelial cells,where it exhibits strong noncovalent affinity for J chain–containing pIgA andpentameric IgM.13 It belongs to the Ig supergene family and is now usuallyreferred to as the polymeric Ig receptor (pIgR). Its human gene has been clonedand characterized,14 and several DNA elements in its promoter and first intronresponsible for a remarkably high constitutive as well as cytokine-enhancedexpression have been identified.15
At the apical epithelial surface, SIgA and SIgM are exocytosed after cleav-age of the receptor (FIG. 1); only the C-terminal smaller pIgR segment remains
290 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
FIGURE 1. Simplified depiction of receptor-mediated export of dimeric IgA and pen-tameric IgM. Membrane secretory component (mSC) functions as polymeric Ig receptor(pIgR) and is expressed basolaterally on serous-type glandular epithelial cells. The poly-meric Ig molecules are produced with incorporated J chain (IgA + J and IgM + J) byplasma cells residing in the glandular stroma. The resulting secretory Ig molecules (SIgAand SIgM) act in a first line of defense by performing immune exclusion of antigens (Ag)in the mucus layer on the epithelial surface. Although J chain is often produced by mucosalIgG+ plasma cells (70–90%), it does not combine with IgG (nor with IgD or IgE; notshown) but is degraded intracellularly as denoted by (±J) in the figure. Locally producedand serum-derived IgG and monomeric IgA can be transmitted paracellularly to the lumenin amounts depending on the integrity of the epithelial barrier (broken arrows). Free SC(depicted in mucus) is generated when unoccupied pIgR (top symbol) is cleaved at theapical face of the epithelial cell in the same manner as bound SC incorporated into SIgAand SIgM. (Based on the epithelial transport model proposed in Refs. 11 and 12.)
for intracellular degradation, whereas the extracellular part (∼80 kDa) is in-corporated into the SIg molecules as bound SC, thereby endowing particularlySIgA with resistance against proteolytic degradation.16,17 Excess of unoccu-pied pIgR is released in the same manner by proteolytic cleavage to form so-called free SC according to the recommended nomenclature.18 This 80-kDaglycoprotein can be found in most secretions including saliva.8 By equilibriumwith bound SC, free SC exerts a stabilizing effect on the quaternary structureof SIgM in which SC is not covalently linked in contrast to SIgA.9 Both freeSC and bound SC show, in addition, several innate functions, such as inhibitionof epithelial adhesion of certain Gram-negative bacteria and neutralization ofcertain bacterial toxins.16,17
BRANDTZAEG 291
The binding sites of pIgA and pentameric IgM initially contacting the firstextracellular domain of pIgR have largely been defined.19 In addition, it hasbeen shown that the 15-kDa J chain is crucial for the stabilization of the initialnoncovalent complexing between the Ig polymers and pIgR or free SC.13,19
Our original suggestion that the J chain and membrane SC are involved ina “lock and key” mechanism in the selective epithelial export of pIgA andpentameric IgM is now firmly established.20–22
IMMUNOLOGICAL ACTIVITY OF SALIVARY GLANDS
Origin of Salivary Immunoglobulins
Salivary secretions are not particularly rich in IgA,8 which in fact repre-sents a minor fraction of total protein compared with the dominating enzymeamylase.23,24 Nevertheless, the parotid IgA-to-IgG ratio is about 500 times in-creased compared with that in serum (TABLE 1), reflecting the selective exportof pIgA (FIG. 1). The same transport mechanism explains that also the IgM-to-IgG ratio is substantially increased in normal parotid fluid compared withthat in serum; but because of the diffusion advantage of the relatively smallIgG molecules, active export of IgM to whole saliva is virtually masked.8,24
The monomeric fraction of salivary IgA is generally small, that is, about 10%in parotid fluid and 13–17% in whole saliva, depending on the clinical state ofthe gingiva.24 It has been estimated that up to 77% of monomeric IgA in salivais serum derived.25
These observations and the significant association of IgG in whole salivawith the product of the serum IgG concentration and the extent of gingival–periodontal inflammation (r = 0.85) show that monomeric IgA and IgG mainlyenter the oral cavity via crevicular fluid.8,24 Also of note, a substantial pro-portion (50–60%) of parotid IgA exists in >25S complexes in the secretion;
TABLE 1. Variations in mean results of salivary IgA determinations performed by thesame laboratory (LIIPAT, 1970–1991)
Conc. (�g/mL)IgA secretion
Samples (no. of adult subjects) IgA IgG rate (�g/min)
Stim. parotid secretion (n = 9)a 40 0.36 27Stim. parotid secretion (n = 27)a 36 NDc 34Stim. parotid secretion (n = 19)b 27 ND 14“Unstim.” parotid secretion (n = 5)a 120 ND 10“Unstim.” whole saliva:
Healthy individuals (n = 8)a 194 14.4 NDPeriodontitis patients (n = 13)a 371 69.7 ND
aSingle radial immunodiffusion; bELISA; cND = not determined.Adapted from Ref. 24.

292 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
this fraction is even higher for SIgA dissolved from the sedimented mucus clotobtained by centrifugation,26 most likely reflecting the mucophilic propertiesof bound SC.17
Variability of Salivary IgA Levels
Because of the slow development of the salivary IgA system,24 age is animportant variable in studies of Igs in oral fluids. Also, various stressors re-portedly27 influence the IgA levels in different manners (TABLE 2). In addition,quantification of salivary IgA is afflicted with many methodological prob-lems. It is difficult to standardize the quantitations due to problems with sam-ple collection, processing, and storage, as reflected in different normal valuesobtained for total IgA and IgM concentrations, even in studies performed bythe same laboratory (TABLE 1). Thus, the results vary strikingly between stud-ies based on single radial immunodiffusion or enzyme-linked immunosorbentassay (ELISA).
It is particularly important to be aware of the striking impact of the secre-tory flow rate on the salivary IgA level (TABLE 1), which may partly explaindifferences among studies. “Unstimulated” parotid secretion thus contains atleast three times more IgA than the stimulated counterpart,23,28 and a similarproportional difference has been reported for whole saliva.29 Some investi-gators have tried to avoid this problem by reporting salivary IgA related tototal protein or albumin, but this will also be misleading because the secretoryresponse of individual parotid proteins is quite different, with large individualvariations,23,24,30 and the salivary level of albumin will depend on leakagefrom serum in a manner similar to IgG and monomeric IgA (FIG. 1).
Some studies recommend reporting the output of salivary proteins in secre-tion rates (�g/min),8 whereas others have suggested that the actual concen-tration (�g/mL) may be a better alternative for IgA.31 Both parameters showconsiderable variations over time in the same individual, and between the leftand the right parotid gland.8,24 Therefore, it is probably best to record boththe absolute concentration and the secretion rate, and to sample from the same
TABLE 2. Effect of different stressors on salivary IgA levels
Definition of stressors Salivary IgA
Chronic academic stress (e.g., during exam period) ReducedAcute academic stress (e.g., just before or after exam) IncreasedAcute “naturalistic stress” (e.g., work shift) IncreasedLaboratory stressors:
“Acute coping” of challenges (sympathetic activation followed Reduced (?)by parasympathetic rebound) Increased
“Passive coping,” feeling of disgust Reduced
Adapted from Ref. 27.

BRANDTZAEG 293
side at the same time of the day on every occasion in longitudinal studies.Relatively large study populations are clearly required to obtain reliable re-sults. Stimulated secretion may be preferable as a test sample31,32; it is alsomore easily collected and less adversely affected by storage than unstimulatedfluid.23
Although the secretion rate of parotid IgA in an individual appears to bemore stable over time than the actual IgA concentration,8,24 SIgA is more sub-jected to short-term variation than other salivary proteins.33,34 This may reflectdifferences in the glandular structures involved in the secretion of various pro-tein components35 and also the fact that SIgA is mainly a product of adaptiveimmunity. Thus, studies in inbred mice have suggested that fluctuations inglandular IgA+ PCs may contribute to variations in salivary Ig levels.36 Di-urnal and seasonal variations should also be considered, as should relation tomeals, cigarette smoking, and pregnancy,31,37,38 in addition to various stressorsas mentioned above (TABLE 2).
Standardized Ig quantitations are even more difficult in whole saliva thanin parotid fluid. First, the contributions to whole saliva from the minor, sub-mandibular, sublingual and parotid glands vary greatly according to the rate offlow,39 and contamination with nasal secretions (and tears) may be difficult toavoid, particularly in uneasy children.40 Notably, the mean IgA concentrationin secretion from labial glands has been reported to be three times higher thanthat in parotid fluid.41 On this basis it can be estimated that the minor glandscontribute 30–35% of the total salivary IgA.41 Second, the flow rate of wholesaliva cannot be so accurately measured as that of the parotid secretion. Third,whole saliva samples usually require centrifugation before quantification, andthe sediment represents a variable Ig loss, probably because SIgA binds notonly to mucus,17,26 but also to oral bacteria.42 Centrifugation may be avoidedby collecting the fluid by careful suction from the floor of the mouth, and evenmore controlled sampling might be obtained by the use of absorbing discs.43
Despite various disadvantages, whole saliva is commonly used as a “rep-resentative” external secretion because it is easily obtained. To increase fluidvolume, chewing on paraffin wax or Parafilm is often recommended. Althoughthis is convenient, it entails certain pitfalls, however; the wax adsorbs organicmaterial44 and the chewing may increase leakage of serum proteins into theoral cavity, for instance from inflamed gingivae. Therefore, oral health shouldbe considered when whole saliva is used for immunological studies.
Local Ig Production in Salivary Glands
IgA+ PCs are normally found scattered among the acini of major salivaryglands and often in clusters adjacent to ducts.24 Interestingly, the submandibu-lar glands contain on average approximately two times more IgA+ PCs pertissue unit than the parotid,45,46 in accordance with a larger output of SIgA.31
It is tempting to speculate that antigens gain easier access to submandibular

294 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
glands, thereby inducing a more active local immune system. The daily outputof IgA/kg wet weight of glandular tissue is similar for salivary and lactatingmammary glands, so the superiority of the latter as an SIgA source dependson the organ size and ductal storage system.46
Because the minor salivary glands are numerous and have close proximityto the oral mucosal surface, they are probably quite important in the defenseof the oropharynx. This is supported by the observation of numerous IgA+PCs adjacent to their ducts47,48 and an abundant output of SIgA from theseglands.41 In fact, the density of IgA+ PCs in the labial glands has been reportedto be three times that in the parotid.49
The subclass IgA2 is more stable than IgA1 because of its resistance tocertain bacterial proteases.50 Therefore, it is interesting that a relatively largeproportion (35–38%) of the IgA+ PCs in salivary glands produce IgA2.48,51
In this respect the salivary glands are intermediate between the upper airwaysand the distal gut, a disparity that clearly reflects regional immunoregulatorydifferences.52 In agreement with the similar affinity of IgA1 and IgA2 for freeSC,53 both subclasses appear to be equally well exported by pIgR into the se-cretion.26 This transport not only takes place primarily through the intercalatedducts but also through serous-type acini.35,47
Although SIgM is a result of pIgR-mediated export (FIG. 1), it is not secon-darily stabilized with SC by covalent bonding,9 and its resistance to proteolyticdegradation is inferior compared to SIgA. Also, when comparing the propor-tions of parotid PC classes and the IgA:IgM ratio in the secretion, the glandularexport of pIgA is favored over that of pentameric IgM by a factor of approxi-mately five (or 12-fold on a molar basis).54 This is not explained by differenthandling of the two polymers by pIgR but is due to diffusion restriction for therelatively large IgM through stromal matrix and basement membranes.54
HOMING OF MUCOSAL B CELLS TO SALIVARY GLANDS
Multiple Sites of B Cell Activation
The initial stimulation of mucosal pIgA-expressing B cells takes placemainly in mucosa-associated lymphoid tissue (MALT), particularly in Peyer’spatches of the distal ileum as well as in other parts of gut-associated lym-phoid tissue (GALT), such as the numerous isolated lymphoid follicles and theappendix.52 From these organized structures, activated B cells reach periph-eral blood and migrate to secretory effector sites, where their extravasationdepends on complementary adhesion molecules and chemokine–chemokinereceptor pairs.52
It is not well delineated which part of MALT is most important for inductionof immune responses subsequently expressed as salivary IgA antibody produc-tion, but there is convincing evidence both in animals and humans that acti-vated B cells can migrate from GALT to salivary glands.55–58 Nevertheless, in

BRANDTZAEG 295
subjects immunized orally with a cholera toxin (CT) B subunit–whole-cell Vib-rio cholerae vaccine, the specific IgA antibody detection sensitivity in salivawas not better than in serum and only about 50% of that in intestinal lavage(FIG. 2). After infection with V. cholerae or enterotoxigenic Escherichia coli,the detection sensitivity for antibodies against the respective toxins increased,but not as much as that seen in serum.59 These results suggest that intestinalimmune induction is not so well reflected in the salivary IgA system. Alsonotably, this study probably overestimated the enteric–oral B cell homing axisbecause CT and enteropathogens breach the gut mucosa to become undulydisseminated to the systemic immune system, as reflected by the high levelsof specific serum IgG antibodies (FIG. 2).
Recent studies point to considerable compartmentalization of B cell hom-ing.52 Thus, nasopharynx-associated lymphoid tissue (NALT), such as theadenoids and palatine tonsils in humans, may be relatively more importantthan GALT as inductive sites for B cells destined to the salivary glands.60,61
Although it remains uncertain to what extent these lymphoepithelial struc-tures of Waldeyer’s ring are functionally comparable to NALT of rodents,18
they are indeed strategically located to orchestrate regional immune functionsagainst both airborne and alimentary antigens. Moreover, they have obviousadvantages as immune-inductive organs because of antigen-retaining cryptsand absence of antigen-degrading digestive enzymes.
FIGURE 2. Antibody responses against CT or Escherichia coli heat-labile toxin inadult Bangladeshi volunteers after peroral Vibrio cholerae vaccination (Vac.), and afterinfection (Inf.) with V. cholerae or enterotoxigenic E. coli. Vaccination was performedtwice by oral administration of a CT B– subunit–whole-cell vaccine approved for humanuse. The antibody detection sensitivity in stimulated whole saliva, breast milk, and serumfor antibodies to the relevant bacterial toxin was normalized in relation to the detection ofspecific IgA antibody in intestinal lavage (set to 100% for each individual). (Based on datafrom Ref. 59.)

296 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
Evidence for Homing of B Cellsfrom NALT to Salivary Glands
Regionalization of the mucosal immune system is supported by B cell hom-ing studies in rats as reviewed elsewhere.60 Also, preferential appearance ofspecific IgA antibodies in saliva has been noted in rabbits after tonsillar anti-gen exposure.62,63 In keeping with the latter observation, direct immunizationof human palatine tonsils, and particularly nasal vaccination, gave rise to localB cell responses in palatine tonsils and adenoids as well as circulating specificB cells that apparently did not enter the intestinal mucosa.64 Moreover, in ba-bies dying of the sudden infant death syndrome (SIDS), the tonsillar germinalcenters were shown to be overstimulated as revealed by an increased numberof IgG+ and IgA+ PCs,65 probably reflecting airway infection; such activatedB cells were apparently distributed in excessive numbers to regional secretoryeffector sites including the parotid glands,66 thereby giving rise to increasedlevels of salivary IgA and IgM in SIDS.67
Altogether, there is accumulating evidence to support the notion that humanNALT supplies secretory effector sites of the upper aerodigestive tract withactivated pIgA precursor cells.24,68 The reason for the suggested homing di-chotomy between this region and the small intestine appears to be differencesin the employed homing molecules.69 The integrin �4�7 is important for B cellextravasation into the intestinal lamina propria by interaction with the mucosaladdressin cell adhesion molecule (MAdCAM)-1 expressed on the mucosalmicrovascular endothelium, but this integrin does not appear to be importantfor homing to the airways and salivary glands.52 Also the involved chemokinereceptor–chemokine interactions (CCR9–CCL25 vs. CCR10–CCL28) show astriking dichotomy between the two body regions.52,69
FUNCTION OF SALIVARY ANTIBODIES
Various Defense Mechanisms of SIgA
The remarkable stability of SIgA makes it well suited to function in protease-containing secretions, such as whole saliva.70 Nevertheless, several oral bacte-ria produce enzymes that can selectively cleave SIgA1 in its extended hinge re-gion, especially certain strains of Streptococcus sanguis and S. mitior, but alsoBacteroides and Capnocytophaga species that are involved in periodontal dis-ease.50 On average, at least 60% of salivary SIgA consists of the IgA1 isotype,26
and parotid antibodies to S. mutans occur predominantly in this subclass,71
whereas activity to lipoteichoic acid from S. pyogenes and to lipopolysac-carides from Bacteroides gingivalis, B. fragilis, and E. coli is carried mainlyby the SIgA2 isotype.71
Although Fab� fragments released by the IgA1 proteases may retain antigen-binding capacity,72 this immune reaction may be adverse rather than protective.

BRANDTZAEG 297
Such fragments may shield microorganisms from the defense function of SIgAantibodies and may even enhance epithelial colonization,73 whereas intactSIgA can specifically inhibit cellular attachment and penetration of influenzavirus in contrast to monomeric IgA- or IgG-neutralizing antibodies.74
The chief defense function of SIgA appears simply to be binding of solubleor particulate antigens (FIG. 1). In vivo coating of oral bacteria with SIgAcan be directly demonstrated by immunofluorescence staining.42 This is noproof of antibody reactivity because many strains of group A or B streptococcipossess Fc� receptors,75 but SIgA coating of bacteria via Fc interactions maynevertheless be of similar functional importance.
Many identified mechanisms may contribute to simple mucosal immune ex-clusion of soluble or particulate antigens.16,24 In addition to efficient antigenbinding, complexing and neutralization, SIgA antibodies show better aggluti-nating properties than monomeric IgA16 and may render bacteria mucophilicin saliva.76 The SIgA function appears to be considerably enhanced by a highlevel of cross-reactivity observed for salivary antibodies.77
Salivary Immunoglobulins in Relation to Disease
Numerous studies have attempted to relate salivary IgA to a variety of oralas well as systemic diseases. Reviews of such reports have been published, butfar from all available data can be considered conclusive.24,38
Dental Caries
How SIgA antibodies might protect against dental disease has for long beena matter of dispute.78 The balance of evidence indicates that there is an in-verse relationship between caries suspectibility and the output of salivary IgAin children and young adults.79 An inverse relationship between salivary IgAantibodies to S. mutans and its early oral colonization, or the colonized indi-vidual’s caries experience, has also been reported; the mechanistic interest inthis respect is focused on bacterial adhesins, glucosyltransferases, and glucan-binding proteins.80
Parotid secretion of young adults regularly contains inhibiting IgA antibodiesto glucosyltransferase, and this enzyme may play a major role in dental plaqueformation. Oral vaccination with killed S. mutans of host origin induced sali-vary IgA antibodies that inhibited glucosyltransferase and reduced the numbersof viable S. mutans organisms in whole saliva and dental plaque.81Also interest-ingly, an experimental study in young Americans suggested that a low parotidIgA antibody level to S. mutans serotype c was associated with enhanced col-onization on molar tooth surfaces, whereas rapid clearance of serotype d wasassociated with relatively higher levels of corresponding parotid IgA antibod-ies.82 Characterization of the salivary IgA response to this bacterium continues

298 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
to be of interest80 because it is of high relevance in strategies aiming at a futureactive caries vaccine.83
Periodontal Disease
Several studies have shown a positive relationship between the concentrationof salivary IgA (TABLE 1) and periodontal disease.8,79 Accumulation of dentalplaque may stimulate IgA production by increasing the amounts of swallowedbacteria that activate B cells in NALT and GALT. Notably, elevated levels ofparotid IgA antibodies to Actinobacillus actinomycetemcomitans were seen insubjects whose subgingival plaque harbored this microorganism.84 Secretoryimmunity may hence be involved in host resistance to periodontal disease; butSIgA antibodies probably have little or no effect on the growth of an establisheddental plaque.
Mucous Membrane Diseases
Low capacity for IgA antibody production as reflected in serum apparentlypredisposes to recurrence and complications of herpes simplex infection, par-ticularly development of intraoral lesions.24 Parotid IgA antibody productionto Candida albicans may similarly be low in some patients with mucocu-taneous candidiasis.24 Conversely, chronic alcoholics and/or heavy smokershave increased IgA levels in whole saliva, and the same is true for patientswith oropharyngeal carcinoma.85 As molecular characterization was not per-formed, the results might be explained by leakage of monomeric serum IgAinto the oral cavity. This possibility was suggested by a study including mea-surements of whole saliva IgG as well as parotid and submandibular IgA.86
Chronic Sialadenitis and Sjogren’s Syndrome
IgG and albumin are relatively much more elevated than IgA in parotidfluid during the early phase of parotitis.87 This suggests admixture with serumproteins from an inflammatory exudate. In addition, some IgG appears to beof local origin, which harmonizes with the presence of an increased numberof IgG+ PCs in chronic sialadenitis.24 Alterations of the parotid Ig pattern inpatients with Sjogren’s syndrome are less pronounced and mimic those foundlate in the recovery phase of recurrent parotitis, but the fluid flow rate is morereduced.87
Most PCs in the benign lymphoepithelial or myoepithelial parotid lesionsof Sjogren patients occur in areas of remaining secretory epithelial elements,including proliferating ducts, and are often dominated by the IgG class.24 In

BRANDTZAEG 299
contrast to the myoepithelial islands, the secretory epithelium shows increasedexpression of pIgR/SC and uptake of IgA, indicating enhanced pIgR-mediatedpIgA export despite extensive pathology.88 Some of the IgA+ and IgM+ PCsmay be involved in local production of rheumatoid factors,89 which have beendetected in whole saliva of patients with Sjogren’s syndrome with or withoutassociated rheumatoid arthritis.90
Also, labial glands of Sjogren patients often show increased production ofIgG and to a lesser extent IgM, apparently as a function of disease severity.91
Immunohistochemistry of labial biopsy specimens may thus aid the diagnosticevaluation. However, it has been shown that labial glands from patients withrheumatoid arthritis or systemic lupus erythematosus may similarly contain in-creased numbers of IgG+ and IgM+ PCs, even in the absence of overt Sjogren’ssyndrome.92
Non-Hodgkin’s Malignant B Cell Lymphoma
Patients with Sjogren’s syndrome have a 40- to 50-fold increased risk ofdeveloping malignant lymphoma, but early recognition of such monoclonalB cell proliferations are often difficult by conventional histopathological ex-amination. Proliferative areas with immunoblasts and plasmacytoid cells havebeen reported in up to 90% of myoepithelial lesions.93 However, molecularstudies of Ig heavy chain gene rearrangements showed a lower frequency of Bcell monoclonality in parotid (68%), and particularly in labial (∼15%), sali-vary glands.94,95 It would have been of diagnostic interest if the monoclonalIgM product could be identified in secretions from the affected glands. InWaldenstrom’s macroglobulinemia (which may be associated with Sjogren’ssyndrome), IgM is increased in parotid fluid87 and its monoclonal nature can beidentified.96 Synthesis of monoclonal IgM has been demonstrated in salivaryglands of such patients.89
Celiac Disease
Patients with untreated celiac disease, or gluten-sensitive enteropathy, haveelevated levels of IgA antibodies against wheat gluten, or more specificallyagainst the ethanol-soluble gliadin fraction of gluten, both in intestinal se-cretions and serum; these antibodies disappear during a successful treatmentwith a gluten-free diet.97 As a reflection of gluten-induced GALT activation,untreated patients also have a raised number of circulating IgA-producinggliadin-specific plasmablasts, which can be detected in an enzyme-linked im-munospot (ELISPOT) assay.98 The number of such activated B cells is reducedto a normal level after successful diet treatment, but increases again in mostceliac patients after 1–2 weeks of challenge with ordinary gluten-containingfood.98

300 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
These results show that the gluten-specific IgA response is disseminated byB cell trafficking throughout the body. It could therefore be anticipated thatIgA antibodies to gliadin would occur in saliva, and in the 1990s this possibilitywas explored in search of a noninvasive approach to screen for celiac disease(FIG. 3). In two studies employing samples of unstimulated whole saliva, thediscrimination of confirmed celiac patients was quite satisfactory,99 or evenbetter than by measurement of IgA antibodies in serum.100 However, compa-rable studies employing stimulated parotid secretion showed only little,101 orvirtually no102 elevation of IgA antibody levels to gliadin in contrast to theresults obtained with serum (FIG. 3).
Such observations suggest that GALT-induced B cells home to the parotidglands with only poor efficiency. Comparable homing to the submandibular,sublingual, and minor salivary glands appears to be more consistent, as deemedfrom the levels of IgA antibodies to gliadin in whole saliva (FIG. 3). Thisdisparity points to compartmentalization of the local B cell system even amongvarious components of the salivary gland system, which would not be surprisingin view of the fact that the mucosal homing mechanism also differs betweenthe small and large bowel.52 In view of the above information, it should bedetermined whether whole saliva could provide a convenient and noninvasivescreening medium for IgA autoantibody activity to tissue transglutaminase,which at present represents the most reliable nonbiopsic indicator of celiacdisease when performed with blood serum samples.97
FIGURE 3. IgA antibody levels to gliadin in serum, whole saliva, and parotid salivaof patients with untreated celiac disease, normalized in relation to the comparable antibodylevels present in healthy control subjects. Only whole saliva could be used in screening forceliac disease with a result matching that obtained with serum. (Based on data from Ref.99 [study 1], Ref. 100 [study 2], Ref. 101 [study 3], and Ref. 102 [study 4].)

BRANDTZAEG 301
Allergy and Airway Infections
Most subjects with selective IgA deficiency in developed countries are clin-ically healthy, but some have increased tendency to allergy and infections,especially when compensatory production of SIgM is lacking.103 It has beenreported that recurrent respiratory infections and isolation of pathogens arerarely seen in IgA-deficient children when salivary IgM is increased.104 Thisobservation was subsequently supported by a study in adults with selectiveIgA deficiency; raised salivary levels of total IgM and of IgM antibodies topoliovirus and E. coli tended to be associated with relatively good resistance toinfections of the respiratory tract.105 Proneness to such infections in selectiveIgA deficiency has been shown, moreover, to be associated with replacementof nasal IgA+ PCs mainly with cells producing IgG or IgD, whereas exclusivereplacement with IgG+ and IgM+ PCs apparently confers satisfactory mucosalresistance.106
In addition, several studies have been performed on subjects with no overtimmunodeficiency to see if the level of salivary IgA shows any negative re-lationship to contraction of allergy or infectious disease, mainly in the upperairways. Usually no clear-cut associations have appeared with infections,107
although it was reported that infants born to atopic parents show significantlyincreased prevalence of salivary IgA deficiency, presumably mainly beingtransient.108 Such a temporary IgA defect was not convincingly related to laterdevelopment of allergy, in agreement with a subsequent study.109 However,it was not excluded that variable compensation with SIgM (which was notstudied) explained the apparent lack of association.
It has also been reported that total salivary IgA tends to be reduced ininfection-prone children with no overt immunodeficiency.110 This is in keepingwith some other studies, but the possibility exists that the result could beexplained by degradation of SIgA1 by microbial proteases. Salivary IgA may,moreover, be decreased in children with recurrent tonsillitis111 or adenoidhyperplasia,112 and also in asthmatic children when wheezing is precipitatedmainly by recurrent respiratory tract infections.111 A subsequent study reporteda significant relationship of transient salivary IgA deficiency with bronchialhyperreactivity but not with asthma.109 It has also been suggested that repeatedantibiotic courses may lead to persistently low salivary IgA levels.113
Human Immunodeficiency Virus (HIV) Infection and AIDS
Saliva of HIV-infected subjects may variably carry this virus,114 probablydepending on the collection method and oral health, which both influence thenumber of HIV+ lymphocytes in the samples.115 Saliva from infected subjectscontains IgA antibodies to HIV, which to some extent may neutralize the virus,and there are HIV-inhibiting factors in saliva unrelated to antibodies.116

302 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
Controversial results have been published concerning the effect of HIV in-fection on the salivary IgA level.116 Such inconsistency may be ascribed both tothe disease stage and the sample source. Because HIV infection is often associ-ated with elevation of serum IgA (mainly IgA1), it is important to avoid wholesaliva for conclusive IgA studies, especially when the patients have candidiasisor other oral health problems. Atkinson et al.117 first reported significantly ele-vated IgA in submandibular but not in parotid secretion from such patients, andMandel et al.118 confirmed the lack of parotid elevation. These observationsmight reflect that the IgA response in the submandibular/sublingual glands ismore closely related to GALT in terms of B cell homing than that in the parotidas discussed earlier, although the influence of a reduced flow rate on the sali-vary IgA level could not be excluded.117 Untreated AIDS patients do indeedhave a highly upregulated intestinal IgA system,119 which would be expectedto become disseminated to salivary glands with the possible exception of theparotid.
Nevertheless, Jackson120 subsequently reported reduced whole saliva IgAin AIDS, which was ascribed to a selective decrease of IgA2, while our lab-oratory26 showed that such patients had reduced output of both parotid IgA1and IgA2—a result that might reflect the degree of secondary immunodefi-ciency.116 This observation has subsequently been supported and extended towhole saliva.121 Also salivary IgA antibodies to viral p24 and gp160 are re-duced in symptomatic patients.122 It is possible that in some studies, a leakageof monomeric IgA1 into whole saliva could have masked a decrease of locallyproduced salivary IgA1.
The level of IgG antibodies to p24 in whole saliva correlates with that inserum,122 again suggesting paracellular leakage from blood,116 either via gin-gival crevices (see earlier) or directly through the oral surface epithelium whentouched by a sampling device, for example, a cotton swab. Tight junctions be-tween epithelial cells are dynamic structures that rapidly open up under the in-fluence of bacterial toxins and inflammatory mediators.123 Thus, IgG immunecomplexes increase leakage of bystander protein through rabbit sublingual mu-cosa.124 Moreover, the slightest irritation of human nasal or intestinal mucosaleads to bulk flow of serum proteins to the epithelial surface.125 Also inter-estingly, salivary IgG has been reported to carry specific anti-gp160 activity25-fold higher than that of serum IgG,126 suggesting some local production—perhaps in tonsils, inflamed gingivae, or submandibular and labial salivaryglands (which contain a higher density of IgG+ PCs than the parotid).46,49
Adequately performed measurements of salivary IgG antibodies to HIVdo show high specificity and sensitivity,127 and commercial kits are availablefor this purpose. Sampling of whole saliva has many advantages in scien-tific field studies of HIV infection and may be preferable to blood for anti-body screening also in clinical settings—and particularly for home testing—although issues concerning false-positive and false-negative results need to beresolved.128

BRANDTZAEG 303
CONCLUSIONS
Oral microorganisms and aerodigestive antigens are continuously influencedby the two major antibody classes in saliva: SIgA and IgG. The former is syn-thesized as pIgA by PCs in salivary glands and is exported by an epithelialreceptor-mediated mechanism. Conversely, most IgG occurring in saliva repre-sent systemic immunity because it is derived from serum by passive diffusion—preferentially through gingival crevices—although a minor fraction may orig-inate from glandular, gingival, or tonsillar PCs. Along with the paracellularleakage of monomeric IgA and IgG antibodies, other serum-derived or locallyproduced factors and mediators will also appear in whole saliva, such as IL-6in patients with ulcerative colitis.129
The secretory antibody system is subject to complex immunoregulation thatinfluences distinctly the activity of the various cell types involved in SIgAgeneration. However, a number of biological phenomena induced by mucosalantigen exposure are poorly defined in experimental animals and still moreobscure in humans. There is evidence to suggest that GALT and NALT do notcontribute equally to the induction of secretory immunity in various regions ofthe body. Because of such compartmentalization, enteric immunization can-not be regarded as an adequate mode of stimulating salivary IgA antibodies,whereas NALT stimulation might be more effective. In fact, the various salivaryglands may rely on different inductive sites for humoral immunity.
In addition to the apparent compartmentalization of the oral secretory im-mune system, there are several other important variables influencing the levelsof total IgA and specific antibodies in salivary fluids. These include the im-pact of flow rate, protein loss during sample handling, difficulties with repro-ducibility and standardization of immunoassays, and uncontrolled admixtureof serum-derived monomeric IgA and IgG to the samples. Despite these prob-lems, saliva remains an interesting biological fluid with great scientific andclinical potentials.
ACKNOWLEDGMENTS
Studies in the author’s laboratory have been supported by the ResearchCouncil of Norway, the University of Oslo, Anders Jahre’s Foundation for thePromotion of Science, and Rikshospitalet-Radiumhospitalet Medical Center.Ms. Hege Eliassen and Mr. Erik Kulø Hagen are thanked for excellent assis-tance with the manuscript.
REFERENCES
1. ELLISON, S.A., P.A. MASHIMO & I.D. MANDEL. 1960. Immunochemical studiesof human saliva. I. The demonstration of serum proteins in whole and parotidsaliva. J. Dent. Res. 39: 892–898.

304 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
2. HANSON, L.A. & P. BRANDTZAEG. 1993. The discovery of secretory IgA and themucosal immune system. Immunol. Today 14: 416–417.
3. TOMASI, T.B., E.M. TAN, A. SOLOMON, et al. 1965. Characteristics of an immunesystem common to certain external secretions. J. Exp. Med. 121: 101–124.
4. CRABBE, P.A., A.O. CARBONARA & J.F. HEREMANS. 1965. The normal hu-man intestinal mucosa as a major source of plasma cells containing �A-immunoglobulin. Lab. Invest. 14: 235–248.
5. BRANDTZAEG, P. 1973. Two types of IgA immunocytes in man. Nat. New Biol.243: 142–143.
6. BRANDTZAEG, P. 1974. Presence of J chain in human immunocytes containingvarious immunoglobulin classes. Nature 252: 418–420.
7. BRANDTZAEG, P., I. FJELLANGER & S.T. GJERULDSEN. 1968. Immunoglobulin M:local synthesis and selective secretion in patients with immunoglobulin A de-ficiency. Science 160: 789–791.
8. BRANDTZAEG, P., I. FJELLANGER & S.T. GJERULDSEN. 1970. Human secretoryimmunoglobulins. I. Salivary secretions from individuals with normal or lowlevels of serum immunoglobulins. Scand. J. Haematol. Suppl. 12: 1–83.
9. BRANDTZAEG, P. 1975. Human secretory immunoglobulin M. An immunochem-ical and immunohistochemical study. Immunology 29: 559–570.
10. BROWN, W.R., Y. ISOBE & P.K. NAKANE. 1976. Studies on translocation of im-munoglobulins across intestinal epithelium. II. Immuno electron−microscopiclocalization of immunoglobulins and secretory component in human intestinalmucosa. Gastroenterology 71: 985–995.
11. BRANDTZAEG, P. 1974. Mucosal and glandular distribution of immunoglobulincomponent. Immunohistochemistry with a cold ethanol-fixation technique. Im-munology 26: 1101–1114.
12. BRANDTZAEG, P. 1974. Mucosal and glandular distribution of immunoglobulincomponents. Differential localization of free and bound SC in secretory epithe-lial cells. J. Immunol. 112: 1553–1559.
13. BRANDTZAEG, P. & H. PRYDZ. 1984. Direct evidence for an integrated function ofJ chain and secretory component in epithelial transport of immunoglobulins.Nature 311: 71–73.
14. KRAJCI, P., K.H. GRZESCHIK, A.H.M. GEURTS VAN KESSEL, et al. 1991. The humantransmembrane secretory component (poly-Ig receptor): molecular cloning,restriction fragment length polymorphism and chromosomal sublocalization.Hum. Genet. 87: 642–648.
15. JOHANSEN, F.-E. & P. BRANDTZAEG. 2004. Transcriptional regulation of themucosal IgA system. Trends Immunol. 25: 150–157.
16. BRANDTZAEG, P. 2003. Role of secretory antibodies in the defence against infec-tions. Int. J. Med. Microbiol. 293: 3–15.
17. PHALIPON, A. & B. CORTHESY. 2003. Novel functions of the polymeric Ig receptor:well beyond transport of immunoglobulins. Trends Immunol. 24: 55–58.
18. BRANDTZAEG, P. & R. PABST. 2004. Let’s go mucosal: communication on slipperyground. Trends Immunol. 25: 570–577.
19. NORDERHAUG, I.N., F.-E. JOHANSEN, H. SCHJERVEN, et al. 1999. Regulation ofthe formation and external transport of secretory immunoglobulins. Crit. Rev.Immunol. 19: 481–508.
20. VAERMAN, J.-P., A.E. LANGENDRIES, D.A. GIFFROY, et al. 1998. Antibody againstthe human J chain inhibits polymeric Ig receptor-mediated biliary and epithelialtransport of human polymeric IgA. Eur. J. Immunol. 28: 171–182.

BRANDTZAEG 305
21. JOHANSEN, F.-E., R. BRAATHEN & P. BRANDTZAEG. 2001. The J chain is essentialfor polymeric Ig receptor-mediated epithelial transport of IgA. J. Immunol. 167:5185–5192.
22. BRAATHEN, R., V.S. HOHMAN, P. BRANDTZAEG, et al. 2007. Secretory antibodyformation: conserved binding interactions between J chain and polymeric Igreceptor from humans and amphibians. J. Immunol. 178: 1589–1597.
23. BRANDTZAEG, P. 1971. Human secretory immunoglobulins. VII. Concentrationsof parotid IgA and other secretory proteins in relation to the rate of flow andduration of secretory stimulus. Arch. Oral Biol. 16: 1295–1310.
24. BRANDTZAEG, P. 1998. Synthesis and secretion of human salivary immunoglobu-lins. In Glandular Mechanisms of Salivary Secretion. J.R. Garrett, J. Ekstrom &L.C. Anderson, Eds.: Frontiers of Oral Biology 10: 167–199. Karger. London.
25. DELACROIX, D.L. & J.P. VAERMAN. 1983. Function of the human liver in IgAhomeostasis in plasma. Ann. N. Y. Acad. Sci. 409: 383–401.
26. MULLER, F., S.S. FRøLAND, M. HVATUM, et al. 1991. Both IgA subclasses arereduced in parotid saliva from patients with AIDS. Clin. Exp. Immunol. 83:203–209.
27. BOSCH, J.A. 2001. Stress and salivary defense systems. The effects of acutestressors on salivary composition and salivary function. Research thesis(Academisch Proefschrift), Vrije University, the Netherlands. ISBN 90-9015363-2 (Printpartners Ipskamp B.V.), pp. 213.
28. MANDEL, I.D. & H.S. KHURANA. 1969. The relation of human salivary �A globulinand albumin to flow rate. Arch. Oral Biol. 14: 1433–1435.
29. GRONBLAD, E.A. 1982. Concentration of immunoglobulins in human wholesaliva: effect of physiological stimulation. Acta Odontol. Scand. 40: 87–95.
30. RUDNEY, J.D. & Q.T. SMITH. 1985. Relationships between levels of lysozyme,lactoferrin, salivary peroxidase, and secretory immunoglobulin A in stimulatedparotid saliva. Infect. Immun. 49: 469–475.
31. STUCHELL, R.N., I.D. MANDEL. 1978. Studies of secretory IgA in caries-resistantand caries-susceptible adults. Adv. Exp. Med. Biol. 107: 341–348.
32. OON, C.H. & J. LEE. 1972. A controlled quantitative study of parotid salivarysecretory IgA-globulin in normal adults. J. Immunol. Methods 2: 45–48.
33. GAHNBERG, L. & B. KRASSE. 1981. Salivary immunoglobulin A antibodies react-ing with antigens from oral streptococci: longitudinal study in humans. Infect.Immun. 33: 697–703.
34. RUDNEY, J.D., K.C. KAJANDER & Q.T. SMITH. 1985. Correlations between hu-man salivary levels of lysozyme, lactoferrin, salivary peroxidase and secretoryimmunoglobulin A with different stimulatory states and over time. Arch. OralBiol. 30: 765–771.
35. KORSRUD, F.R. & P. BRANDTZAEG. 1982. Characterization of epithelial elementsin human major salivary glands by functional markers: localization of amylase,lactoferrin, lysozyme, secretory component and secretory immunoglobulins bypaired immunofluorescence staining. J. Histochem. Cytochem. 20: 657–666.
36. DESLAURIERS, N., M. OUDGHIRI, J. SEGUIN, et al. 1986. The oral immune sys-tem: dynamics of salivary immunoglobulin production in the inbred mouse.Immunol. Invest. 15: 339–349.
37. WIDERSTROM, L. & D. BRATTHALL. 1984. Increased IgA levels in saliva duringpregnancy. Scand. J. Dent. Res. 92: 33–37.
38. GLEESON, M., A.W. CRIPPS & R.L. CLANCY. 1995. Modifiers of the human mu-cosal immune system. Immunol. Cell Biol. 73: 397–404.

306 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
39. KERR, A.C. 1961. The physiological regulation of salivary secretions in man.In International Series of Monographs on Oral Biology, Vol. 1. R.C. Greulich,J.B. MacDonald & M.A. Rushton, Eds. Pergamon Press, Oxford.
40. TENOVUO, J., O.P. LEHTONEN, A.S. AALTONEN, et al. 1986. Antimicrobial factorsin whole saliva of human infants. Infect. Immun. 51: 49–53.
41. CRAWFORD, J.M., M.A. TAUBMAN & D.J. SMITH. 1975. Minor salivary glands asa major source of secretory immunoglobin A in the human oral cavity. Science190: 1206–1209.
42. BRANDTZAEG, P., I. FJELLANGER & S.T. GJERULDSEN. 1968. Adsorption of im-munoglobulin A onto oral bacteria in vivo. J. Bacteriol. 96: 242–249.
43. KRISTIANSEN, B.E. 1984. Collection of mucosal secretion by synthetic discs forquantitation of secretory IgA and bacteria. J. Immunol. Methods 73: 251–257.
44. BERG, E. & J.C. TJELL. 1969. Paraffin chewing as a saliva-stimulating agent.J. Dent. Res. 48: 325.
45. KORSRUD, F.R. & P. BRANDTZAEG. 1980. Quantitative immunohistochemistryof immuno globulin- and J-chain-producing cells in human parotid and sub-mandibular salivary glands. Immunology 39: 129–140.
46. BRANDTZAEG, P. 1983. The secretory immune system of lactating human mam-mary glands compared with other exocrine organs. Ann. N. Y. Acad. Sci. 409:353–381.
47. BRANDTZAEG, P. 1977. Immunohistochemical studies of various aspects of glan-dular immunoglobulin transport in man. Histochem. J. 9: 553–572.
48. MORO, I., S. UMEMURA, S.S. CARGO, et al. 1984. Immunohistochemical distribu-tion of immmunoglobulins, lactoferrin, and lysozyme in human minor salivaryglands. J. Oral Pathol. 13: 97–104.
49. MATTHEWS, J.B., A.J.C. POTTS & M.K. BASU. 1985. Immunoglobulin-containingcells in normal human labial salivary glands. Int. Arch. Allergy Appl. Immunol.77: 374–376.
50. KILIAN, M., J. REINHOLDT, H. LOMHOLT, et al. 1996. Biological significance ofIgA1 proteases in bacterial colonization and pathogenesis: critical evaluationof experimental evidence. APMIS 104: 321–338.
51. KETT, K., P. BRANDTZAEG, J. RADL, et al. 1986. Different subclass distribution ofIgA-producing cells in human lymphoid organs and various secretory tissues.J. Immunol. 136: 3631–3635.
52. BRANDTZAEG, P. & F.-E. JOHANSEN. 2005. Mucosal B cells: phenotypic character-istics, transcriptional regulation, and homing properties. Immunol. Rev. 206:32–63.
53. BRANDTZAEG, P. 1977. Human secretory component. VI. Immunoglobulin-binding properties. Immunochemistry 14: 179–188.
54. NATVIG, I.B., F.-E. JOHANSEN, T.W. NORDENG, et al. 1997. Mechanism for en-hanced external transfer of dimeric IgA over pentameric IgM. Studies of diffu-sion, binding to the human polymeric Ig receptor, and epithelial transcytosis.J. Immunol. 159: 4330–4340.
55. MESTECKY, J., J.R. MCGHEE, R.R. ARNOLD, et al. 1978. Selective induction ofan immune response in human external secretions by ingestion of bacterialantigen. J. Clin. Invest. 61: 731–737.
56. WEISZ-CARRINGTON, P., M.E. ROUX, M. MCWILLIAMS, et al. 1979. Organ andisotype distribution of plasma cells producing specific antibody after oral im-munization: evidence for a generalized secretory immune system. J. Immunol.123: 1705–1708.

BRANDTZAEG 307
57. JACKSON, D.E., E.T. LALLY, M.C. NAKAMURA, et al. 1981. Migration of IgA-bearing lymphocytes into salivary glands. Cell. Immunol. 63: 203–209.
58. CZERKINSKY, C., A.M. SVENNERHOLM, M. QUIDING, et al. 1991. Antibody-producing cells in peripheral blood and salivary glands after oral cholera vac-cination of humans. Infect. Immun. 59: 996–1001.
59. JERTBORN, M., A.M. SVENNERHOLM & J. HOLMGREN. 1986. Saliva, breast milk,and serum antibody responses as indirect measures of intestinal immunity af-ter oral cholera vaccination or natural disease. J. Clin. Microbiol. 24: 203–209.
60. BRANDTZAEG, P. 1996. The B-cell development in tonsillar lymphoid follicles.Acta Otolaryngol. Suppl. 523: 55–59.
61. BRANDTZAEG, P. 2003. Immunology of tonsils and adenoids: everything the ENTsurgeon needs to know. International Congress Series (ICS) 1254: 89–99, 2003(Elsevier)/Int. J. Pediatr. Otorhinolaryngol. 67(Suppl. 1): S69–76.
62. FUKUIZUMI, T., H. INOUE, Y. ANZAI, et al. 1995. Sheep red blood cell instillationat palatine tonsil effectively induces specific IgA class antibody in saliva inrabbits. Microbiol. Immunol. 39: 351–359.
63. FUKUIZUMI, T., H. INOUE, T. TSUJISAWA, et al. 1999. Tonsillar application offormalin-killed cells of Streptococcus sobrinus reduces experimental dentalcaries in rabbits. Infect. Immun. 67: 426–428.
64. QUIDING-JARBRINK, M., G. GRANSTROM, I. NORDSTROM, et al. 1995. Inductionof compartmentalized B-cell responses in human tonsils. Infect. Immun. 63:853–857.
65. STOLTENBERG, L., A. VEGE, O.D. SAUGSTAD, et al. 1995. Changes in the concen-tration and distribution of immunoglobulin−producing cells in SIDS palatinetonsils. Pediatr. Allergy Immunol. 6: 48–55.
66. THRANE, P., T.O. ROGNUM & P. BRANDTZAEG. 1990. Increased immune responsein upper respiratory and digestive tracts in SIDS. Lancet 335: 229–230.
67. GLEESON, M., R.L. CLANCY & A.W. CRIPPS. 1993. Mucosal immune response ina case of sudden infant death syndrome. Pediatr. Res. 33: 554–556.
68. BRANDTZAEG, P. 1999. Regionalized immune function of tonsils and adenoids.Immunol. Today 20: 383–384.
69. JOHANSEN, F.-E., E.S. BAEKKEVOLD, H.S. CARLSEN, et al. 2005. Regional induc-tion of adhesion molecules and chemokine receptors explains disparate homingof human B cells to systemic and mucosal effector sites: dispersion from tonsils.Blood 106: 593–600.
70. MA, J.K., B.Y. HIKMAT, K. WYCOFF, et al. 1998. Characterization of a recombinantplant monoclonal secretory antibody and preventive immunotherapy in humans.Nat. Med. 4: 601–606.
71. BROWN, T.A. & J. MESTECKY. 1985. Immunoglobulin A sublass distribution ofnaturally occurring salivary antibodies to microbial antigens. Infect. Immun.49: 459–462.
72. MANSA, B. & M. KILIAN. 1986. Retained antigen-binding activity of Fab frag-ments of human monoclonal immunoglobulin A1 (IgA1) cleaved by IgA1 pro-tease. Infect. Immun. 52: 171–174.
73. WEISER, J.N., D. BAE, C. FASCHING, et al. 2003. Antibody-enhanced pneumococcaladherence requires IgA1 protease. Proc. Natl. Acad. Sci. USA 100: 4215–4220.
74. TAYLOR, H.P. & N.J. DIMMOCK. 1985. Mechanism of neutralization of influenzaevirus by secretory IgA is different from that of monomeric IgA or IgG. J. Exp.Med. 161: 198–209.

308 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
75. CHRISTENSEN, P. & V.-A. OXELIUS. 1975. A reaction between some streptococciand IgA myeloma proteins. Acta Pathol. Microbiol. Scand. [C] 83: 184–188.
76. BIESBROCK, A.R., M.S. REDDY & M.J. LEVINE. 1991. Interaction of a salivarymucin−secretory immunoglobulin A complex with mucosal pathogens. Infect.Immun. 59: 3492–3497.
77. QUAN, C., A. BERNEMAN, R. PIRES, et al. 1997. Natural polyreactive secretoryimmunoglobulin A autoantibodies as a possible immune barrier in humans.Infect. Immun. 65: 3997–4004.
78. RUSSELL, M.W. & J. MESTECKY. 1986. Potential for immunological interventionagainst dental caries. J. Biol. Buccale 14: 159–175.
79. TAUBMAN, M.A. & D.J. SMITH. 1993. Significance of salivary antibody in dentaldisease. Ann. N. Y. Acad. Sci. 694: 202–215.
80. NOGUEIRA, R.D., A.C. ALVES, M.H. NAPIMOGA, et al. 2005. Characterization ofsalivary immunoglobulin A responses in children heavily exposed to the oralbacterium Streptococcus mutans: influence of specific antigen recognition ininfection. Infect. Immun. 73: 5675–5684.
81. GREGORY, R.L. & S.J. FILLER. 1987. Protective secretory immunoglobulin Aantibodies in humans following oral immunization with Streptococcus mutans.Infect. Immun. 55: 2409–2415.
82. GREGORY, R.L., S.M. MICHALEK, S.J. FILLER, et al. 1985. Prevention of Strep-tococcus mutans colonization by salivary IgA antibodies. J. Clin. Immunol. 5:55–62.
83. RUSSELL, M.W., N.K. CHILDERS, S.M. MICHALEK, et al. 2004. A caries vaccine?The state of the science of immunization against dental caries. Caries Res. 38:230–235.
84. SMITH, D.J., J.L. EBERSOLE, M.A. TAUBMAN, et al. 1985. Salivary IgA antibody toActinobacillus actinomyetemcomitans in young adult population. J. PeriodontalRes. 20: 8–11.
85. MANDEL, M.A., K. DVORAK & J.J. DE COSSE. 1973. Salivary immunoglobulinsin patients with oropharyngeal and bronchopulmonary carcinoma. Cancer 31:1408–1413.
86. BROWN, A.M., E.T. LALLY & A. FRANKEL. 1975. IgA and IgG content of the salivaand serum of oral cancer patents. Arch. Oral Biol. 20: 395–398.
87. MANDEL, I.D. & H. BAURMASH. 1978. Salivary immunoglobulins in diseasesaffecting salivary glands. Adv. Exp. Med. Biol. 107: 839–847.
88. THRANE, P.S., L.M. SOLLID, H.R. HAANES, et al. 1992. Clustering of IgA-producing immunocytes related to HLA-DR-positive ducts in normal and in-flamed salivary glands. Scand. J. Immunol. 35: 43–51.
89. ANDERSON, L.G., N.A. CUMMINGS, R. ASOFSKY, et al. 1972. Salivary gland im-munoglobulin and rheumatoid factor synthesis in Sjogren’s syndrome. Am. J.Med. 53: 456–463.
90. DUNNE, J.V., D.A. CARSON, H.L. SPIEGELBERG, et al. 1979. IgA rheumatoid fac-tor in the sera and saliva of patients with rheumatiod arthritis and Sjogren’ssyndrome. Ann. Rheum. Dis. 38: 161–165.
91. TALAL, N., R. ASOFSKY & P. LIGHTBODY. 1970. Immunoglobulin synthesis bysalivary gland lymphoid cells in Sjogren’s syndrome. J. Clin. Invest. 49: 49–54.
92. MATTHEWS, J.B., A.J.C. POTTS, J. HAMBURGER, et al. 1986. Immunoglo-bulin−producing cells in labial salivary glands of patients with rheumatoidarthritis and systemic lupus erythematosus. J. Oral Pathol. 15: 520–523.

BRANDTZAEG 309
93. SCHMID, U., D. HELBRON & K. LENNERT. 1982. Development of malignant lym-phoma in myoepithelial sialadenitis (Sjogren’s syndrome). Virchows Arch.(Pathol. Anat.) 395: 11–43.
94. JORDAN, R.C., E.W. ODELL & P.M. SPEIGHT. 1996. B-cell monoclonality in sali-vary lymphoepithelial lesions. Eur. J. Cancer B Oral Oncol. 32B: 38–44.
95. JORDAN, R.C., Y. MASAKI, S. TAKESHITA, et al. 1996. High prevalence of B-cellmonoclonality in labial gland biopsies of Japanese Sjogren’s syndrome patients.Int. J. Hematol. 64: 47–52.
96. BRANDTZAEG, P. 1971. Human secretory immunoglobulins. II. Salivary secretionsfrom individuals with selectively excessive or defective synthesis of serumimmunoglobulins. Clin. Exp. Immunol. 8: 69–85.
97. BRANDTZAEG, P. 2002. The mucosal B cell and its functions. In Food Allergy andIntolerance. Second edition. J. Brostoff & S.J. Challacombe, Eds.: 127–171.Saunders: Elsevier Science. London.
98. HANSSON, T., A. DANNAEUS, W. KRAAZ, et al. 1997. Production of antibodies togliadin by peripheral blood lymphocytes in children with celiac disease: theuse of an enzyme-linked immunospot technique for screening and follow-up.Pediatr. Res. 41: 554–559.
99. HAKEEM, V., R. FIFIELD, H.F. AL-BAYATY, et al. 1992. Salivary IgA antigliadinantibody as a marker for coeliac disease. Arch. Dis. Child. 67: 724–727.
100. AL-BAYATY, H.F., M.J. ALDRED, D.M. WALKER, et al. 1989. Salivary and serumantibodies to gliadin in the diagnosis of celiac disease. J. Oral Pathol. Med. 18:578–581.
101. O’MAHONY, S., E. ARRANZ, J.R. BARTON, et al. 1991. Dissociation between sys-temic and mucosal humoral immune responses in coeliac disease. Gut 32:29–35.
102. GIBNEY, M.J. & C. BRADY. 1991. Systemic and salivary antibodies to dietaryantigens (gliadin, casein and ovalbumin) in treated and untreated coeliac diseaseand in Crohn’s disease. Eur. J. Int. Med. 2: 115–119.
103. BRANDTZAEG, P. & D.E. NILSSEN. 1995. Mucosal aspects of primary B-cell de-ficiency and gastrointestinal infections. Curr. Opin. Gastroenterol. 11: 532–540.
104. OSTERGAARD, P.A. 1980. Predictable clinical disorders related to serum and salivaIglevels and the number of circulating T cells in asthmatic children. Clin. Al-lergy 10: 277–284.
105. MELLANDER, L., J. BJORKANDER, B. CARLSSON, et al. 1986. Secretory antibodiesin IgA-deficient and immunosuppressed individuals. J. Clin. Immunol. 6: 284–291.
106. BRANDTZAEG, P., G. KARLSSON, G. HANSSON, et al. 1987. The clinical conditionof IgA-deficient patients is related to the proportion of IgD- and IgM-producingcells in their nasal mucosa. Clin. Exp. Immunol. 67: 626–636.
107. NAKAMURA, C., T. AKIMOTO, S. SUZUKI, et al. 2006. Daily changes of salivarysecretory immunoglobulin A and appearance of upper respiratory symptomsduring physical training. J. Sports Med. Phys. Fitness. 46: 152–157.
108. VAN ASPEREN, P.P., M. GLEESON, A.S. KEMP, et al. 1985. The relationship betweenatopy and salivary IgA deficiency in infancy. Clin. Exp. Immunol. 62: 753–757.
109. GLEESON, M., R.L. CLANCY, M.J. HENSLEY, et al. 1996. Development of bronchialhyperreactivity following transient absence of salivary IgA. Am. J. Respir. Crit.Care Med. 153: 1785–1789.

310 ANNALS OF THE NEW YORK ACADEMY OF SCIENCES
110. LEHTONEN, O.-P.J., J. TENOVUO, A.S. AALTONEN, et al. 1987. Immunoglobulins andinnate factors of immunity in saliva of children prone to respiratory infections.Acta Pathol. Microbiol. Scand. [C] 95: 35–40.
111. OSTERGAARD, P.A. 1977. IgA levels and carrier rate of pathogenic bacteria in 27children previously tonsillectomized. Acta Pathol. Microbiol. Scand. [C] 85:178–186.
112. HESS, M., J. KUGLER, D. HAAKE, et al. 1991. Reduced concentration of secretoryIgA indicates changes of local immunity in children with adenoid hyperplasiaand secretory otitis media. ORL J. Otorhinolaryngol. Relat. Spec. 53: 339–341.
113. OSTERGAARD, P.A. 1983. Oral bacterial flora and secretory IgA in small childrenafter repeated courses of antibiotics. Scand. J. Infect. Dis. 15: 115–118.
114. GROOPMAN, J.E., S.Z. SALAHUDDIN, M.G. SARNGADHARAN, et al. 1984. HTLV-IIIin saliva of people with AIDS-related complex and healthy homosexual men atrisk for AIDS. Science 226: 447–449.
115. PEKOVIC, D., D. AJDUKOVIC, C. TSOUKAS, et al. 1987. Detection of human im-munosuppressive virus in salivary lymphocytes from dental patients with AIDS.Am. J. Med. 52: 188–189.
116. CHALLACOMBE, S.J. & J.R. NAGLIK. 2006. The effects of HIV infection on oralmucosal immunity. Adv. Dent. Res. 19: 29–35.
117. ATKINSON, J.C., C. YEH, F.G. OPPENHEIM, et al. 1990. Elevation of salivary antimi-crobial proteins following HIV-1 infection. J. Acquir. Immune Defic. Syndr. 3:41–48.
118. MANDEL, I.D., C.E. BARR & L. TURGEON. 1992. Longitudinal study of parotidsaliva in HIV-1 infection. J. Oral Pathol. Med. 21: 209–213.
119. NILSSEN, D.E., O. ØKTEDALEN & P. BRANDTZAEG. 2004. Intestinal B cell hyper-activity in AIDS is controlled by highly active antiretroviral therapy. Gut 53:487–493.
120. JACKSON, S. 1990. Secretory and serum IgA are inversely altered in AIDS patients.In Advances in Mucosal Immunology. T.T. MacDonald, S.J. Challacombe, P.W.Bland, C.R. Stokes, R.V. Heatly & A. Mcl. Mowat, Eds.: 665. Kluwer AcademicPublishers. Lancaster.
121. SWEET, S.P., D. RAHMAN & S.J. CHALLACOMBE. 1995. IgA subclasses in HIVdisease: dichotomy between raised levels in serum and decreased secretionrates in saliva. Immunology 86: 556–559.
122. MATSUDA, S., S. OKA, M. HONDA, et al. 1993. Characteristics of IgA antibodiesagainst HIV-1 in sera and saliva from HIV-seropositive individuals in differentclinical stages. Scand. J. Immunol. 38: 428–434.
123. FASANO, A. & T. SHEA-DONOHUE. 2005. Mechanisms of disease: the role ofintestinal barrier function in the pathogenesis of gastrointestinal autoimmunediseases. Nat. Clin. Pract. Gastroenterol. Hepatol. 2: 416–422.
124. BRANDTZAEG, P. & K. TOLO. 1977. Mucosal penetrability enhanced by serum-derived antibodies. Nature 266: 262–263.
125. PERSSON, C.G., J.S. ERJEFALT, L. GREIFF, et al. 1998. Contribution of plasma-derived molecules to mucosal immune defence, disease and repair in the air-ways. Scand. J. Immunol. 47: 302–313.
126. LU, X.S., J.F. DELFRAISSY, L. GRANGEOT-KEROS, et al. 1994. Rapid and constantdetection of HIV antibody response in saliva of HIV-infected patients; selectivedistribution of anti-HIV activity in the IgG isotype. Res. Virol. 145: 369–377.

BRANDTZAEG 311
127. HUNT, A.J., J. CONNELL, G. CHRISTOFINIS, et al. 1993. The testing of saliva samplesfor HIV-1 antibodies: reliability in a non-clinic setting. Genitourin. Med. 69:29–30.
128. WRIGHT, A.A. & I.T. KATZ. 2006. Home testing for HIV. N. Engl. J. Med. 354:437–440.
129. NIELSEN, A.A., J.N. NIELSEN, A. SCHMEDES, et al. 2005. Saliva interleukin-6 inpatients with inflammatory bowel disease. Scand. J. Gastroenterol. 40: 144–148.



















![Nye Annals[1]](https://img.dokumen.tips/doc/110x75/577d216e1a28ab4e1e9538aa/nye-annals1.jpg)









