
Universidade de São Paulo
Faculdade de Saúde Pública
Detecção de genes codificadores de resistência a
antimicrobianos de importância clínica em amostras
de carne de frango
Marina Manrique Coan
Dissertação de Mestrado apresentada ao Programa
de Pós-Graduação em Ciências.
Área de concentração: Serviços de Saúde Pública.
Orientação: Professor Associado Glavur Rogerio
Matté.
São Paulo
2014

Detecção de genes codificadores de resistência a
antimicrobianos de importância clínica em amostras
de carne de frango
Marina Manrique Coan
Dissertação de Mestrado apresentada ao Programa
de Pós-Graduação em Ciências.
Área de concentração: Serviços de Saúde Pública.
Orientação: Professor Associado Glavur Rogerio
Matté.
São Paulo
2014

RESUMO
COAN, M. M. Detecção de genes codificadores de resistência a antimicrobianos de
importância clínica em amostras de carne de frango [Dissertação de Mestrado]. São
Paulo: Faculdade de Saúde Pública da USP; 2014.
Introdução. A introdução dos antimicrobianos na prática clínica no século XX foi
um grande avanço para a medicina. Entretanto, seu uso indiscriminado, tanto na
medicina (humana e animal) quanto na agricultura e pecuária, possibilitou a seleção e
disseminação de microrganismos resistentes. A transferência genética horizontal é
um dos principais mecanismos responsáveis pela disseminação de genes que
codificam resistência aos antimicrobianos, pois possibilita a transmissão da
resistência de uma célula bacteriana (comensal ou patogênica) para outra. Genes
codificadores de resistência a antimicrobianos de importância clínica são encontrados
em microrganismos de origem ambiental, clínica e alimentar. Objetivo. O objetivo
do presente estudo foi detectar genes que codificam resistência aos antibióticos β-
lactâmicos, Tetraciclinas e Quinolonas, em amostras de carne de frango, visando
estudar a ocorrência da resistência antimicrobiana nesse produto considerado um
alimento comum na dieta do homem. Materiais e Métodos. Foram utilizadas 30
amostras de carne de frango. Após a inoculação da carne de frango em caldo Luria
0,5%, o DNA total das bactérias cultivadas nesse meio foi extraído por choque
térmico e utilizado na pesquisa de genes de resistência pela técnica de PCR seguida
de eletroforese em gel de agarose. Foi também realizada a pesquisa de Coliformes
Termotolerantes por meio da técnica dos tubos múltiplos para estimar a qualidade
higiênico sanitária das amostras. Resultados: Oito (26,7%) amostras apresentaram
um ou mais genes codificadores de resistência à antimicrobianos β-Lactâmicos, 28
(93,3%) amostras apresentaram um ou mais genes codificadores de resistência às
Tetraciclinas e 28 (93,3%) amostras apresentaram um ou mais genes codificadores
de resistência às Quinolonas. Conclusões: A realização deste estudo contribuiu com
informações a respeito da circulação de genes de resistência na carne de frango e
alerta as autoridades de Saúde Pública do Brasil quanto à disseminação da resistência
bacteriana e a necessidade de mais estudos a respeito desse problema, já que esses
são raros ou inexistentes no País.

ABSTRACT
COAN, M. M. Detecção de genes codificadores de resistência a antimicrobianos de
importância clínica em amostras de carne de frango [Dissertação de Mestrado]. São
Paulo: Faculdade de Saúde Pública da USP; 2014/ Detection of genes encoding
resistance to clinically important antibiotics in samples of chicken. São Paulo (BR):
Faculdade de Saúde Pública da USP; 2014.
Introduction. The introduction of antibiotics in clinical practice in the twentieth
century was a breakthrough for medicine. However, their indiscriminate use in both
medicine (human and animal) as in agriculture and livestock, enabled the selection
and spread of resistant microorganisms. Horizontal gene transfer is a major
mechanism responsible for the spread of genes encoding antimicrobial resistance,
since it allows the transmission of resistance from one bacteria to another. Genes
encoding resistance to antimicrobials of clinical importance are found in
microorganisms present in food, environment and clinical sources. Objective. The
aim of this study was to detect genes encoding resistance to β -lactam antibiotics,
tetracyclines and quinolones in meat samples from chicken samples, to study the
occurrence of antimicrobial resistance in this product considered a common food in
human diet. Materials and Methods. Thirty samples of chicken were used. After
inoculation of chicken meat in Luria broth 0.5 %, the total DNA of the bacteria
cultured was extracted by heat shock and used to search resistance genes by PCR
followed by agarose gel electrophoresis. The search for thermotolerant coliforms was
also conducted using the technique of multiple tubes in order to estimate the sanitary
quality of the samples. Results: Eight (26,7 %) of the samples had one or more genes
encoding for resistance to β -lactam antimicrobials, 28 (93,3%) had one or more
genes encoding resistance to tetracyclines and 27 (93,3 %) samples had one or more
genes encoding resistance to quinolones. Conclusions: This study contributes to the
scientific community and sanitary agencies, bringing information regarding the
occurrence and distribution of resistance genes in chicken, alerting the Public Health
authorities in Brazil about the spread of bacterial resistance and the need for more
studies in this field, as these are rare or nonexistent in the country.

DEDICATÓRIA
À minha família Mãe, Pai, Stela e Murilo, que se mostram presentes em todos os
momentos da minha vida.

AGRADECIMENTOS
A Deus, pelo Dom da vida e proteção...
...Aos meus pais, Teresa e Affonso, pelo amor e carinho com que me criaram.
Agradeço o incentivo de vocês em todas as minhas decisões. Sem vocês eu não seria
nada...
... A minha melhor amiga – irmã, Stela, pelo simples fato de existir. Você foi o
melhor acontecimento da minha vida...
... Ao meu namorado Murilo, pelo amor, paciência e companheirismo em todos os
momentos. MUITO OBRIGADA pelo apoio que me dá desde quando nos
conhecemos...
... Aos meus Avós João e Antonieta, que rezam e torcem pela minha vitória...
... A minha tia querida, Fernanda, pelas palavras alegres e conselhos sinceros...
... Ao meu avô Affonso, que com certeza gostaria de compartilhar esse momento
comigo. Saudades...
... A minha Avó Clarinda, por suas orações...
... As minhas grandes amigas Carol e Rebeca, pela nossa eterna amizade...
... Ao meu quarteto, Gabi Justi, Gabi Martins, Camilinha e Mayara, pelos momentos
de felicidade que me proporcionam...
... Ao meu querido amigo Marcelo, você é o “irmão” que Deus colocou em minha
vida...
... Aos amigos de Santana, pelos momentos inesquecíveis que vivemos ao longo
desses anos de amizade...
... A Professora Doutora Maria Helena, pelos ensinamentos que me transmitiu ao
longo do curso...
... Ao Professor Doutor Glavur, pela sua orientação e paciência...
... Ao Professor Alexandre, pelo seu ensinamento na época da faculdade e pelo
carinho com que vem me acompanhado...
... A Milena, pelos ensinamentos e por sempre me acolher nos momentos em que
estava aflita...

... A Livia, pelos conselhos sinceros e pela disposição para me ajudar quando
precisei...
... As companheiras de Laboratório, Ronalda, Bruna, Rafa e Vanessa pelos
momentos de diversão que me proporcionaram ao longo do tempo que estive com
vocês...
Muito Obrigada!

EPÍGRAFE
“Cada sonho que você deixa para trás, é um pedaço do seu futuro que deixa de
existir.”
Steve Jobs

ÍNDICE
1. INTRODUÇÃO 4
1.1 Resistência bacteriana aos Antimicrobianos 5
1.1.1 Resistência aos β-lactâmicos 8
1.1.2 Resistência às Tetraciclinas 21
1.1.3 Resistência às Quinolonas 24
1.2 Disseminação da Resistência Bacteriana 26
1.3 Avicultura no Brasil 33
1.4 Disseminação da Resistência Bacteriana em carne de frango no
Brasil
35
2. OBJETIVOS 37
2.1 Objetivo Geral 37
2.2 Objetivos Específicos 37
3. MATERIAL E MÉTODOS 38
3.1 Amostras 38
3.2 Contagem de Coliformes Termotolerantes 38
3.2.1 Preparo das Amostras 38
3.2.2 Técnica dos Tubos Múltiplos 39
3.3 Detecção de Genes de Resistência 39
3.3.1 Extração do DNA Total 39
3.3.2 Reação em cadeia pela Polimerase (PCR) 40
3.4 Visualização dos produtos amplificados 41
4. RESULTADOS 47
4.1 Coliformes Termotolerantes 47
4.2 Genes codificadores de resistência aos β-Lactâmicos 47
4.2.1 Genes codificadores de ESBL 47
4.2.2 Genes codificadores de AmpC β-Lactamases 49
4.3 Genes codificadores de resistência às Tetraciclinas 50
4.3.1 Genes tet codificadores de bombas de efluxo 50

4.3.2 Genes tet codificadores de proteção ribossomal 51
4.4 Genes codificadores de resistência às Quinolonas 52
5. DISCUSSÃO 54
5.1 Genes codificadores de resistência aos β-Lactâmicos 55
5.1.1 Genes codificadores de ESBL 55
5.1.2 Genes codificadores de AmpC β-Lactamases 59
5.2 Genes que codificam resistência às Tetraciclinas 59
5.2.1 Genes codificadores de bombas de efluxo 59
5.2.2 Genes codificadores de proteínas de proteção ribossomal 60
5.3 Genes que codificam resistência às Quinolonas 63
5.4 Contagem de Coliformes Termotolerantes 64
5.5 Globalização e a disseminação dos genes de resistência: um alerta
para a Saúde Pública
65
6. CONCLUSÕES 68
7. CONSIDERAÇÕES FINAIS 69
8. REFERÊNCIAS BIBLIOGRÁFICAS 71
ANEXOS 96
ANEXO 1 - Número e porcentagem dos diferentes tipos de amostras de carne de
frango (supermercado, açougue e feira) que continham genes codificadores de
resistência, segundo a classe de antimicrobianos.
97

LISTA DE QUADROS
Quadro 1. Principais antimicrobianos de utilidade clínica e seus respectivos
mecanismos de ação.
5
Quadro 2. Classificação e exemplificação dos antimicrobianos β-lactâmicos. 8
Quadro 3. Classificação das β-Lactamases segundo os esquemas molecular, e
funcional.
11
Quadro 4. Cronologia das AmpC β-Lactamases mediadas por plasmídeos. 17
Quadro 5. Classificação das Tetraciclinas 22
Quadro 6. Classificação das Quinolonas 24
Quadro 7. Microrganismos de importância clínica e resistência fenotípica aos
antimicrobianos utilizados no tratamento de infecções mediadas por esses
patógenos.
28
Quadro 8. Sequência de iniciadores (5´-3´) utilizados na pesquisa dos genes de
resistência
43
Quadro 9. Resultados obtidos para as diferentes amostras de carne de frango
quanto à identificação de genes codificadores de resistência aos β-Lactâmicos e
às Quinolonas.
53

2
LISTA DE FIGURAS
Figura 1. Classificação dos genes codificadores de resistência às
tetraciclinas segundo os seus mecanismos (ROBERTS, 2003).
23
Figura 2. Esquema geral da metodologia aplicada para pesquisa de
genes de resistência em amostras de frango.
46
Figura 3. Representação gráfica do número e tipos de amostras nas
quais há presença dos fragmentos correspondentes aos genes
codificadores de ESBL dos grupos blaCTX-M-1, blaCTX-M-2, blaCTX-M-8, e
para os genes blaTEM e blaSHV.
48
Figura 4. Representação gráfica do número e tipos de amostras nas
quais foram detectados os fragmentos correspondentes aos genes
codificadores de AmpC β-Lactamase, blaMOX e blaCMY.
49
Figura 5. Representação gráfica do número e tipos de amostras nas
quais os fragmentos correspondentes aos genes codificadores de
Bombas de Efluxo tetA, tetB, tetC, tetD e tetE foram obtidos.
51
Figura 6. Representação gráfica do número e tipos de amostras nas
quais foram observados os fragmentos correspondentes aos genes
codificadores de Proteínas de Proteção Ribosssomal, tetM, tetO, tetQ,
tetS e tetW.
52
Figura 7. Representação gráfica do número e tipos de amostras nas
quais foram amplificados os fragmentos correspondentes aos genes
codificadores de Proteínas de Proteção Ribosssomal, qnrA, qnrB e
qnrS.
53

3
LISTA DE TABELAS
Tabela 1. Número de variantes das principais famílias de β-Lactamases
identificadas no período de 2000 a 2014.
12

4
1. Introdução
Há 3000 anos, os chineses foram os responsáveis pelos primeiros relatos do
uso de antimicrobianos, por meio da utilização de bolores para o tratamento de
tumores e feridas infeccionadas. Já em 1928, Alexandre Fleming descobriu o
primeiro antibiótico de utilidade clínica, ao observar que uma cultura de
Staphylococcus aureus havia sido contaminada por um fungo e que, ao redor da
contaminação, não existia crescimento bacteriano. Estudando o fungo, denominado
Penicillium, descobriu que este era produtor de uma substância com efeito
antibacteriano (Penicilina) (TAVARES, 2001).
Inicialmente os antimicrobianos eram obtidos a partir de alguns tipos de
microrganismos e, após o conhecimento da sua estrutura química, passaram a ser
sintetizados em laboratórios. Esses compostos foram então classificados de acordo
com sua estrutura química, seu espectro de ação (amplo espectro ou baixo espectro),
sua ação sobre o microrganismo (bacteriostático ou bactericida) e seu mecanismo de
ação (TAVARES, 2001).
Quando os microrganismos são sensíveis aos antimicrobianos, podem
ocorrer dois efeitos na célula: morte da bactéria (bactericida) ou a interrupção do seu
crescimento e reprodução (bacteriostático). São seis os principais mecanismos de
ação dos antimicrobianos nas bactérias: interferência na biossíntese do
peptideoglicano, na tradução, na replicação do DNA, no metabolismo celular, na
transcrição e na membrana celular. Esses mecanismos variam de acordo com o tipo

5
de antibiótico utilizado e sua estrutura química. O Quadro 1 ilustra os principais
antimicrobianos de uso clínico e seus locais de ação (MORAR e WHRIGHT, 2010).
Quadro 1. Principais antimicrobianos de utilidade clínica e seus respectivos
mecanismos de ação.
CLASSES EXEMPLOS LOCAL DE AÇÃO
Β-lactâmicos Penicilinas
Cefalosporinas
Carpapenens
Monobactâmico
Biossíntese do
Peptiodeoglicano
Aminoglicosídeos Amicacina
Canamicina
Estreptomicina
Gentamicina
Tradução
Glicopeptídeos Vancomicina Biossíntese de peptideoglicano
Tetraciclinas Minociclinas
Tigeciclinas
Tradução
Macrolídeos Eritromicina
Azitromicina
Tradução
Lincosamidas Clindamicina Tradução
Estreptogaminas Synercid Tradução
Oxazolidinonas Linezolid Tradução
Cloranfenicol Cloranfenicol Tradução
Quinolonas Ciprofloxacina Replicação do DNA
Pirimidinas Trimetoprim Metabolismo celular
Sulfonamidas Sulfametoxazol Metabolismo celular
Rifamicinas Rifampicina Transcrição
Lipopeptídeos Daptomicina Membrana celular
Peptídeos Catiônicos Colistina Membrana celular
Fonte: DROPA (2006); MORAR e WHRIGHT (2010).
1.1 Resistência Bacteriana aos Antimicrobianos
A resistência aos antimicrobianos é um processo evolutivo natural, resultante
da exposição a esses compostos. Poucos anos depois da introdução da penicilina na
prática clínica, em 1940, foi observada uma grande proporção de Staphylococcus
aureus resistentes ao composto. Poucos anos depois, em 1944, um fato parecido
ocorreu com a estreptomicina, durante o tratamento de pacientes, quando foi relatado

6
que cepas de Mycobacterium turbeculosis mostraram resistência ao composto. À
medida que outros antimicrobianos foram descobertos e introduzidos, bactérias de
importância clínica apresentaram resistência, fenômeno que persiste nos dias atuais
(FRENCH, 2005; DAVIES e DAVIES, 2010).
Do ponto de vista clínico, define-se resistência bacteriana quando um
paciente, infectado por um patógeno específico, submete-se a um tratamento com
antibiótico, em dose e administração adequadas, porém os critérios de cura não são
atingidos. O início da utilização dos antimicrobianos no século XX foi um grande
avanço para a medicina humana, entretanto, o uso indiscriminado tanto na medicina
(humana e animal) quanto na agricultura e pecuária, contribuiu para a seleção e
disseminação de microrganismos resistentes no ambiente (HENRIQUES et al, 2006;
CANTÓN e MOROSINI, 2011).
Nos últimos anos, a disseminação da resistência bacteriana em
microrganismos de importância clínica tornou-se um sério problema mundial. A
resistência a antimicrobianos reduz significativamente a possibilidade de eficácia nos
tratamentos de infecções e resulta em um número maior de complicações e casos
fatais, além de aumentarem os gastos públicos em sistemas de saúde (ANDERSSON
e HUGHES, 2011). HOLMBERG et al. (1987) foram os primeiros a analisar o
impacto econômico da resistência antimicrobiana na clínica, e desde então estudos
têm revisado o assunto (MCGOWAN, 2001; COSGROVE e CARMELE, 2003;
SINGER et al., 2003). Um estudo desenvolvido por FRENCH et al. (2005)
demonstra que a taxa de mortalidade e o tempo de internação de um paciente

7
infectado por cepas resistentes são duas vezes maiores do que de pacientes infectados
por cepas sensíveis.
Os microrganismos possuem diversos mecanismos genéticos que os tornam
resistentes aos antimicrobianos. Esses mecanismos podem ser intrínsecos, ou seja,
associados a uma mutação cromossômica ou expressão cromossômica latente de um
gene de resistência, ou extrínseca, o qual envolve a aquisição de um novo material
genético por meio de bacteriófagos (transdução), incorporação de DNA
(transformação) ou aquisição de um plasmídeo (conjugação). Por meio desses
processos, os microrganismos tornam-se capazes de desenvolver três principais
mecanismos de resistência associados à informação codificada no seu material
genético: produção de uma enzima capaz de destruir ou inativar o antibiótico,
alteração no sítio alvo do antimicrobiano impedindo sua ação e impedimento do
acesso do antibiótico ao sítio alvo da bactéria (SHLAES et al. 1997; DZIDIC e
BEDEKOVIC, 2003).
Quando os genes associados à resistência bacteriana estão localizados em
elementos genéticos móveis, especialmente em plasmídeos, existe a possibilidade de
ocorrer a mobilização dessa resistência de uma célula bacteriana para a outra. A
transmissão da resistência entre bactérias se dá principalmente pela transferência
horizontal de genes (conjugação) e tem grande importância científica devido à sua
capacidade de disseminação da resistência bacteriana (PLOY et al., 2000; WHITE et
al., 2001).

8
1.1.1 Resistência aos β-lactâmicos
Os β-lactâmicos são os antimicrobianos de maior utilização na prática clínica,
devido à sua ação bactericida e baixa toxicidade. Esta classe de antibióticos é
composta por quatro grupos (Quadro 2) que são caracterizados por possuírem um
anel β-lactâmico (SAMAHA-KFOURY e ARAJ, 2003; DROPA, 2006).
Quadro 2. Classificação e exemplificação dos antimicrobianos β-lactâmicos.
GRUPOS EXEMPLOS
Penicilinas Amoxicilina, Ampicilina, Oxacilina,
Penicilina, Piperacilina, Ticarcilina.
Cefalosporinas
1ªgeração: Cefazolina, Cefalotina,
Cefaloridina, Cefalexina
2ª geração: Cefaclor, Cefuroxima,
Moxalactam, Cefotetan, Cefoxitina.
3ª geração: Ceftizoxima, Cefoperazona,
Cefotaxima, Cefpodoxima, Ceftazidima,
Ceftriaxona
4ª geração: Cefepime, Cefpirona
5ª geração: Ceftaroline, Ceftobripole
Monobactâmicos Aztreonam
Carbapenêmicos Imipenem, Meropenem, Ertapenem,
Doripenem.
Fonte: SAMAHA-KFOURY e ARAJ, 2003; DROPA, 2006; FERNANDES et al.,
2013.
Os β-lactâmicos agem na célula bacteriana por meio de dois mecanismos que
inibem a síntese da parede celular. Primeiramente, são englobados pela célula
bacteriana e inibem a transpeptidase, enzima responsável pela síntese da parede
celular e, em segundo, ligam-se às proteínas ligadoras de penicilina (PBPs)
permitindo a ação das hidrolases na parede bacteriana, o que promove a lise celular
(SAMAHA-KAFOURY e ARAJ, 2003).

9
Β-Lactamases
A habilidade das bactérias produzirem enzimas que destroem os β-lactâmicos
mostra-se presente desde a descoberta da penicilina. A primeira β-Lactamase foi
identificada em uma cepa de Escherichia coli em 1940, e é a principal causa da
resistência bacteriana a essa classe de antimicrobianos (ABRAHAM e CHAIN,
1940). Essa enzima tem a capacidade de clivar o anel β-lactâmico, impedindo a
ligação do antibiótico nas PBPs e permitindo que a síntese da parede celular
bacteriana ocorra normalmente (TAVARES, 2001; SHAH et al., 2004; TURNER,
2005).
Diversos artigos descrevem a classificação das β-Lactamases em dois grupos
(funcional e molecular) para caracterizar as mais de 1320 variantes de enzimas
existentes (LAHEY, 2014). O esquema molecular, segundo Ambler, foi proposto em
1980 e divide as β-Lactamases em quatro principais classes (A a D) baseando-se na
homologia das proteínas, sendo as classes A, C e D serino β-Lactamases e a classe B
metalo β-Lactamases. Já o esquema funcional, de acordo com BUSH e JACOBY
(2010), classifica essas enzimas de acordo com o perfil do substrato e seu inibidor
(Quadro 3).
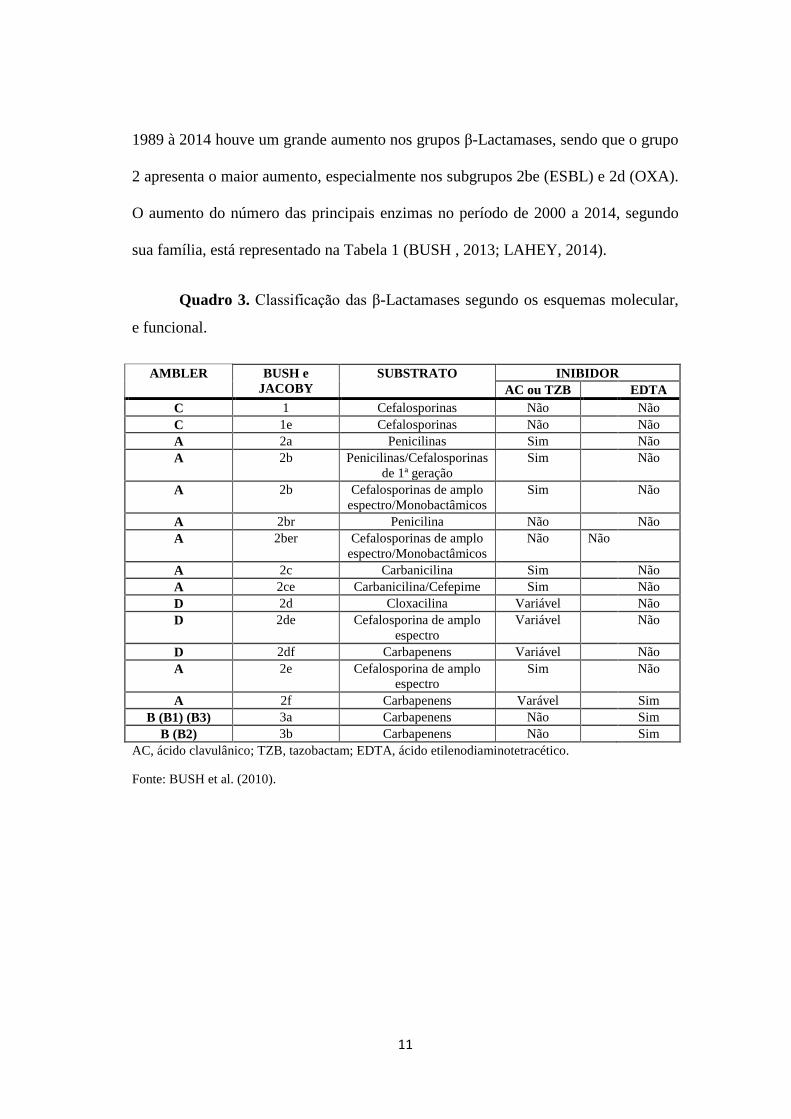
Como pode ser observado no Quadro 3, essas enzimas são classificadas de A-
D. A classe C (Molecular) é associada a dois subgrupos de acordo com o parâmetro
funcional, 1 e 1e, os quais hidrolisam Cefalosporinas e possuem pouca ação em
inibidores de Β-Lactamases. Entretanto o subgrupo 1e, também chamado de ESAC
Β-Lactamases (AmpC de espectro extendido), apresenta atividade de hidrólise em

10
Cefalosporinas com cadeias de aminotiazolexima. As classes Moleculares A e D ou
grupo 2, incluem as Serino Β-Lactamases que demonstram ampla atividade de
hidrólise em todos os antibióticos Β-lactâmicos. Duas principais enzimas dos
subgrupos 2be (ESBL) e 2f (Serino Carbapenemases) destacam-se nos estudos por
serem responsáveis por um alto número de doenças infecciosas no mundo, devido a
hidrólise de todas as classes dos Β-lactâmicos (BUSH, 2013).
A maioria das enzimas pertencentes ao grupo A são inibidas por Ácido
Clavulânico, apesar de apresentarem resposta a combinações de inibidores como
Amoxicilina-Clavulanato, pode ser variável em cada caso. A classe D é comum por
demonstrar resistência em microrganismos como Acinetobacter baumannii e
Pseudomonas aeruginosa. O grupo funcional 3 ou Classe B (Metalo Β-Lactamases)
são enzimas que hidrolisam todos os Β-lactâmicos com exceção do Monobactâmicos
e não são inibidos por inibidores de Β-Lactamases (BUSH, 2013).
Os genes codificadores de β-Lactamases (genes bla) estão localizados no
cromossomo bacteriano ou em elementos genéticos móveis (plasmídeos e
transposons). Um grande número de genes bla tem sido observado em integrons
(WELDHAGEN, 2004), e estes estão inseridos dentro de elementos genéticos
móveis que são importantes determinantes para a disseminação dos genes bla e de
outros tipos de resistência (BABIC et al, 2006).
A identificação de novas enzimas tem aumentado, devido ao baixo custo,
facilidade de pesquisa e o avanço da Biologia Molecular. Estima-se que o número de
Β-Lactamases de origem clínica seja de pelo menos 1320 variantes. No período de
11
1989 à 2014 houve um grande aumento nos grupos β-Lactamases, sendo que o grupo
2 apresenta o maior aumento, especialmente nos subgrupos 2be (ESBL) e 2d (OXA).
O aumento do número das principais enzimas no período de 2000 a 2014, segundo
sua família, está representado na Tabela 1 (BUSH , 2013; LAHEY, 2014).
Quadro 3. Classificação das β-Lactamases segundo os esquemas molecular,
e funcional.
AMBLER BUSH e
JACOBY
SUBSTRATO INIBIDOR
AC ou TZB
EDTA
C 1 Cefalosporinas Não Não
C 1e Cefalosporinas Não Não
A 2a Penicilinas Sim Não
A 2b Penicilinas/Cefalosporinas
de 1ª geração
Sim Não
A 2b Cefalosporinas de amplo
espectro/Monobactâmicos
Sim Não
A 2br Penicilina Não Não
A 2ber Cefalosporinas de amplo
espectro/Monobactâmicos
Não Não
A 2c Carbanicilina Sim Não
A 2ce Carbanicilina/Cefepime Sim Não
D 2d Cloxacilina Variável Não
D 2de Cefalosporina de amplo
espectro
Variável Não
D 2df Carbapenens Variável Não
A 2e Cefalosporina de amplo
espectro
Sim Não
A 2f Carbapenens Varável Sim
B (B1) (B3) 3a Carbapenens Não Sim
B (B2) 3b Carbapenens Não Sim
AC, ácido clavulânico; TZB, tazobactam; EDTA, ácido etilenodiaminotetracético.
Fonte: BUSH et al. (2010).

12
Tabela 1. Número de variantes das principais famílias de β-Lactamases
identificadas no período de 2000 a 2014. Fonte: BUSH 2013 e LAHEY, 2014.
ENZIMA CLASSE
MOLECULAR
2000 2012 2014
TEM A 86 197 217
SHV A 26 168 184
CTX-M A 9 134 150
GES A 1 22 24
KPC A 0 12 17
VIM B 2 37 40
IMP B 3 42 47
CMY C 6 95 111
OXA D 28 255 390
Β-Lactamases de Espectro Estendido (ESBL)
Com a descoberta dos β-lactâmicos de amplo espectro, esses compostos
passaram a ser utilizados amplamente na clínica para o tratamento de diversas
infecções, sobretudo as hospitalares. Entretanto, devido ao uso extenso, a resistência
a esses antimicrobianos difundiu-se rapidamente. KLIEBE et al. (1985) identificaram
a primeira ESBL, denominada SHV-2, na Alemanha, em um isolado de Klebsiella
ozaenae, que apresentou resistência a todos os compostos testados (cefalosporinas de

13
amplo espectro, penicilinas e monobactâmico), com exceção do cefotetan. Devido a
essa enzima, capaz de hidrolisar antimicrobianos de espectro estendido, outras que
foram surgindo com as mesmas características, receberam o nome de β-Lactamases
de espectro estendido (ESBL) (SHAH et al., 2004; TURNER, 2005).
As ESBL têm capacidade de hidrolisar penicilinas, cefalosporinas e aztreonam,
pertencem às classes A ou D de Ambler e aos grupos funcionais 2b, 2ber ou 2de de
BUSH e JACOBY, e são inibidas pelo ácido clavulânico e/ou tazobactam (BUSH et
al., 2010). A maioria dos genes codificadores dessas enzimas possuem localização
plasmidial (STṺRENBURG e MACK, 2003; DROPA, 2013).
A presença dessas enzimas em membros da família Enterobacteriaceae,
sobretudo em Klebsiella pneumoniae e Escherichia coli, tem grande importância na
área clínica. As ESBL também são encontradas em organismos gram negativos não
fermentadores, como Pseudomonas aeruginosa e Acinetobacter baumannii
(FALAGAS e KARAGEORGOPOULOS, 2009). As mais comumente encontradas
são as enzimas dos grupos TEM, SHV e CTX-M, sendo que o último é o mais
prevalente (PEREZ et al, 2007; FALAGAS e KARAGEORGOPOULOS, 2009).
A primeira TEM observada, TEM-1, foi identificada em um isolado de
Escherichia coli, em 1985 (MEDEIROS et al., 1985). Essa enzima, localizada no
transposon Tn3, possui atividade hidrolítica em penicilinas e cefalosporinas de
primeira geração, não possuindo portanto o fenótipo ESBL. A primeira enzima do
grupo TEM a apresentar espectro estendido foi a TEM-3, identificada em 1987
(http://www.lahey.org/studies/; STṺRENBURG e MACK, 2003; PATERSON e
BONOMO, 2005; DROPA, 2006).

14
Enzimas SHV são comumente associadas a isolados de Klebsiella
pneumoniae, tornando-se responsáveis por 20% da resistência desse microrganismo a
ampicilina. A maioria de suas variantes origina-se da substituição de serina por uma
glicina em sua cadeia de aminoácidos e, atualmente, existe um grande número de
variantes dessa enzima (BRADFORD, 2001; http://www.lahey.org/studies/ ;
SAMAHA- KAFOURY e ARAJ, 2003; PATERSON e BONOMO, 2005).
As enzimas TEM e SHV disseminaram-se no período de 1980 a 1990,
sobretudo em cepas de Klebsiella spp isoladas de pacientes internados em UTI
(Unidade de Terapia Intensiva). Variantes dessas enzimas como SHV-2, SHV-5,
SHV-12, TEM-3, TEM-10, TEM-26, TEM-52 e TEM-116 têm sido descritas em
diversos países, inclusive no Brasil (LIVERMORE, 2012).
A partir do ano 2000, houve uma grande mudança no que diz a respeito à
distribuição das ESBL, devido à disseminação das enzimas CTX-M (DROPA, 2013).
O primeiro isolado dessa enzima foi observado, simultaneamente, na Europa e na
América do Sul no início de 1989. A origem dos genes codificadores da CTX-M foi
identificada no cromossomo de um gênero bacteriano denominado Kluyvera, que
constitui a microbiota normal dos seres humanos, com baixa ou nenhuma atividade
patogênica e pode ser considerado um patógeno oportunista. As CTX-M são
divididas em 5 grupos (CTX-M-1, CTX-M-2, CTX-M-8, CTX-M-9 e CTX-M-25)
aos quais pertencem 154 variantes (http://www.lahey.org/Studies/) disseminadas pelo
mundo, e seu substrato preferencial é a cefotaxima (CANTÓN e COQUE, 2009;
ROSSOLINI et al. 2008).

15
Em microrganismos de importância clínica, essas enzimas têm sido
identificadas em plasmídeos nos quais também podem estar inseridos genes que
codificam resistência a outras classes de antimicrobianos (aminoglicosídeos,
cloranfenicol, sulfonamidas, trimetoprim e tetraciclinas) (BONNET, 2004). Na
Escherichia coli de origem humana e animal, por exemplo, o gene blaCTX-M-1 é
frequentemente reportado em plasmídeos do grupo IncN que também estão
associados ao gene blaVIM-1 (CARATTOLI, 2013).
Outros tipos clinicamente importantes de ESBL são: OXA, PER, VEB, CME,
TLA, SFO, GES e BES (http://www.lahey.org/studies/; PEREZ et al, 2007;
FALAGAS et al, 2009).
AmpC β-Lactamases
Em 1940, foi detectada, em Escherichia coli, a primeira enzima capaz de
hidrolisar penicilinas, denominada AmpC (ABRAHAM e CHAIN, 1940). Essa
enzima tem atividade hidrolítica em penicilinas, cefalosporinas, monobactâmicos e
inibidores de β-Lactamases. Classificam-se de acordo com Ambler, na classe C e no
esquema de Bush e Jacoby no grupo 1 e geralmente estão presentes no cromossomo
de microrganismos como Citrobacter freundii, Enterobacter aerogenes,
Enterobacter cloacae, Morganella morganii, Pseudomonas aeroginosa e Serratia
marcescens (BABIC et al., 2006; JACOBY, 2009; BUSH e JACOBY, 2010;
OLIVEIRA, 2011).
Em diversas espécies da família Enterobacteriaceae existe baixa expressão
cromossômica dessa enzima, que pode ser induzida pela exposição do

16
microrganismo ao β-lactâmico. Esse fenômeno de indução envolve cinco principais
genes (ampC, ampR, ampD, ampG e ampE) relacionados à reciclagem do
peptideoglicano da parede bacteriana. O gene ampR atua como ativador do
fenômeno da indução e como repressor em condições normais da célula, os genes
ampC e ampR codificam a síntese de proteínas de membrana e o gene ampD dá
origem a uma proteína solúvel, liberada no citoplasma da bactéria (JACOBY, 2009;
SERAL et al., 2012).
Na presença de um agente indutor (β-lactâmico) o gene ampR facilita a
entrada de produtos da degradação do peptideoglinano no citoplasma da célula
bacteriana. Esses produtos auxiliam na conversão do ampR repressor para ativador
do gene ampC. Além disso, produtos da degradação do peptideoglicano são
metabolizados por ampD, promovendo a reciclagem da parede celular, uma vez que
os metabólitos são reutilizados para a formação do peptideoglicano. Na ausência dos
β-lactâmicos, não há a liberação de produtos da degradação do peptideoglicano o que
impede a ativação do gene ampR em sua função de ativador do gene ampC. A função
do gene ampE ainda não está definida (HANSON e SANDERS, 1999; JACOBY,
2009; SERAL et al. 2012).
Diferente das outras enterobactérias, a Escherichia coli não possui o gene
ampR, fazendo com que o gene codificador da enzima AmpC não seja expresso por
meio da indução e sim por mecanismos de atenuação (JAURIN et al., 1981;
JACOBY, 2009).

17
Quadro 4. Cronologia dos relatos de AmpC β-Lactamases mediadas por plasmídeos.
AmpC β-
Lactamase
País de Origem Ano de Publicação Espécie do
Primeiro
Isolamento
CMY-1 Corea do Sul 1989 K. pneumoniae
CMY-2 Grécia 1996 K. pneumoniae
MIR-1 Estados Unidos 1990 K.pneumoniae
MOX-1 Japão 1993 K. pneumoniae
LAT-1 Grécia 1993 K. pneumoniae
FOX-1 Argentina 1991 K. pneumoniae
DHA-1 Arábia Saudita 1997 S. enteritidis
ACT-1 Estados Unidos 1997 K. pneumoniae
ACC-1 Alemanha 1999 K. pneumoniae
CFE-1 Japão 2004 E.coli
Fonte: adaptação de JACOBY (2009).
Até 1989, as AmpCs eram consideradas enzimas de origem cromossômica,
entretanto BAUERNFEIND et al. (1998) observaram um isolado de Klebsiella
pneumoniae capaz de transferir sua resistência a cefoxitina, cefotetan e outras
penicilinas, oximino-cefalosporinas e monobactâmico, para uma cepa de Escherichia
coli por meio de um plasmídeo. Desde então, genes com localizações plasmidiais
mediadores da classe C de β-Lactamases identificados, sobretudo, em
microrganismos que não possuem AmpC cromossômico, foram detectados em
diversas regiões do mundo (Quadro 4) (PHILIPPON et al., 2002).
Metalo β-Lactamases (MBL)
As classes A, C e D das β-Lactamases possuem um sítio ativo de serina
enquanto a classe B ou Metalo β-Lactamases requer um ou dois íons de zinco para
sua atividade. Essas enzimas caracterizam-se por hidrolisar todos os β-lactâmicos,

18
com exceção dos monobactâmicos, e chamam atenção por sua atividade constante e
eficiente em carbapenêmicos. Esse fato torna-se um alerta devido à atividade de
amplo espectro deste antimicrobiano em microrganismos de importância clínica.
Diferente das outras enzimas, as MBL não são inibidas por Ácido Clavulânico ou
Tazobactam, porém nos testes microbiológicos o EDTA (Ácido Etilenodiamino
Tetra-acético) mostra-se um inibidor dessas enzimas (PITOUT et al, 2005;
BEBRONE, 2007).
A primeira enzima deste grupo de β-Lactamases foi descoberta em 1966 por
SABBATH e ABRAHAM (1966), no cromossomo de uma cepa de Bacillus cereus,
em que foi demonstrado que sua atividade de carbapenemase poderia ser inibida pelo
EDTA. Durante duas décadas, essa cepa foi o único exemplo de MBL encontrado.
Entretanto, em 1980, foi reportada a presença de metalo β-Lactamases em um grande
número de microrganismos (Bacteroides fragilis, Pseudomonas aeruginosa,
Aeromonas hydrophila, Serratia marcescens e Elizabethkingia meningoseptica)
(BEBRONE, 2007; BERTONCHELI et al., 2008).
As MBL podem ser produzidas intrinsecamente, geralmente por
microrganismos de origem ambiental, por exemplo, Aeromonas hydrophilia, e estar
localizadas no cromossomo dessas bactérias. Hipóteses sugerem que essa produção
intrínseca pode ser resultado da exposição dessas bactérias a compostos β-
lactâmicos, assim mantendo os genes codificadores dessa enzima, ou ainda que as
MBL possam executar outra função não identificada na célula bacteriana.
Independente disso, esses genes codificadores são ativados por indução e a maioria

19
dos microrganismos que os carreiam, são ou podem tornar-se altamente resistentes
aos β-lactâmicos. (BERTONCHELI, 2008; WALSH et al. 2008).
Após a caracterização do gene cfiA, encontrado em uma cepa de Bacteriodes
fragilis, foi observado que a MBL poderia ser mediada por elementos genéticos
móveis (YAMAZOE et al., 1999). Diversos tipos de MBL já foram observados em
elementos transferíveis de microrganismos como Pseudomonas aeroginosa,
Acinetobacter spp e em diversas espécies da família Enterobacteriaceae
(BERTONCHELI, 2008). Atualmente existem 4 principais grupos de enzimas
pertencentes à classe das Metalo β-Lactamases: IMP, VIM, SPM e NDM.
As enzimas IMP (imipenemase) e VIM (Verona imipenemase) foram
observadas pela primeira vez no Japão e na Itália, respectivamente. Já a São Paulo
Metalo β-Lactamase (SPM) foi isolada de uma cepa de Pseudomonas aeruginosa,
em São Paulo (Brasil). A maioria dos genes codificadores das enzimas do tipo IMP e
VIM estão localizados em integrons de classe 1, embora IMP também possa ser
encontrada em integrons de classe 3. O gene codificador da enzima SPM possui
localização plasmidial (WALSH et al., 2005).
A New Deli Metalo β-Lactamase (NDM-1) é a MBL mais recente, descoberta
por YONG et al. (2009), em cepas de Klebsiella pneumoniae e Escherichia coli,
isoladas de amostras clínicas de um paciente em Nova Deli-Índia. A partir deste
primeiro relato, muitos casos de NDM-1 têm sido descritos em diversos locais
(Reino Unido, Índia, Paquistão, Bangladesh), sobretudo em cepas de E. coli e K.
pneumoniae. (NORDMANN et al., 2011). Atualmente, a NDM-1 tem gerado
discussões políticas e científicas e tornou-se uma das principais enzimas

20
disseminadas em microrganismos gram negativos em países no Norte da Europa e
Região Pacífica da Ásia, além já ter sido detectada no Brasil (CARVALHO-ASSEF
et al., 2013; BUSH, 2013).
Klebsiella pneumoniae Carbapenemase (KPC)
Os antimicrobianos carbapenêmicos (meropenem, imipenem, ertapenem) são
compostos de primeira escolha a serem utilizados em infecções graves, mediadas por
membros da família Enterobacteriaceae, produtores de ESBL. O surgimento de
enterobactérias resistentes aos carbapenens é preocupante, uma vez que as opções de
tratamento tornam-se muito restritas (NORDMANN, et al., 2009).
A KPC foi observada pela primeira vez em 1996, em um isolado de
Klebsiella pneumoniae, na Carolina do Norte (YIGIT et al., 2001). A KPC,
pertencente à classe A de Ambler, tem atividade hidrolítica em carbapenêmicos e o
gene blaKPC é responsável por codificá-la. Esse gene tem alta capacidade de
disseminação, devido à sua localização no transposon Tn4401, que caracteriza-se por
sua inserção em diversos plasmídeos de bactérias gram negativas (ARNOLD et
al.,2011).
Duas características principais separam a KPC das outras carbapenemases: a
localização dessa enzima em plasmídeos transferíveis e o espectro de hidrólise que
inclui antimicrobianos como a Aztreonam. Essa enzima é comumente encontrada em
Klebsiella pneumoniae e já foi identificada em isolados de Enterobacter spp e
Salmonella spp. Microrganismos produtores de KPC estão associados a infecções
sistêmicas, mediadas por enterobactérias, observadas com frequência em pacientes

21
imunocomprometidos. A partir do primeiro relato dessa enzima, enterobactérias
produtoras de KPC foram detectadas nos Estados Unidos, América do Sul (Brasil e
Argentina), China e com menor frequência na Europa. A enzima também foi
observada em isolados de Pseudomonas aeruginosa, na Colômbia e em Porto Rico,
tornando-se um alerta a disseminação desse tipo de resistência em uma bactéria não
pertencente à família das Enterobacteriaceae (QUEENAN e BUSH, 2007;
NORDMANN, 2009). Atualmente, essa enzima tem sido frequentemente observada
em infeções hospitalares nos Estados Unidos, Sul da Europa, Américas e China
mesmo com diversas medidas de controle sendo aplicadas nos diversos hospitais
(BUSH,2013)
Plasmídeos carreadores dos genes blaKPC podem estar associados a genes que
codificam resistência aos aminoglicosídeos e às enzimas ESBL, como blaCTX-M-15
(CAI et al., 2008; NORDMANN et al., 2009). Estudos identificaram, em plasmídeos
de cepas de Klebsiella pneumoniae, o gene blaKPC-3, onde também estavam
localizados os determinantes qnrA e qnrB, responsáveis por codificar resistência às
Quinolonas (CHMELNITSKY et al., 2008; ENDIMIANI et al., 2008).
1.1.2 Resistência às Tetraciclinas
A Tetraciclina foi descoberta em 1940 e devido aos poucos efeitos colaterais
causados pelo composto, exceto em crianças e grávidas, o seu uso terapêutico
iniciou-se em 1950. Possui amplo espectro de atividade em microrganismos gram
positivos e gram negativos, clamídias intracelulares, micoplasmas, riquétsias e em
alguns parasitas, além de ser utilizado amplamente na criação de animais, como
promotores de crescimento, e na agricultura (ROBERTS, 2003).

22
Essa classe de antimicrobiano pode ser dividida em 1ª geração (1948 à 1963), 2ª
geração (1965 à 1972) e 3 ª geração (1993) (Quadro 5), e seu mecanismo de ação
ocorre por meio da inibição da síntese proteica, impedindo a associação do
aminoacetil tRNA com o ribossomo da bactéria (ROBERTS, 2003).
Quadro 5. Classificação das Tetraciclinas.
Geração Nome Ano de descoberta
1ª geração Oxitetraciclina
Clortetraciclina
6-dimetilclortetraciclina
1948
1948
1957
2ª geração Doxicilina
Minociclina
1967
1972
3ª geração Tigeciclina 1993
Fonte: CHOPRA e ROBERTS (2001), THAKER et al. (2010).
Existem 43 genes tet/otr que codificam resistência às Tetraciclinas. Desses,
17 (39%) são encontrados em microrganismos de origem ambiental. A maioria
desses genes estão associados a elementos genéticos móveis que permitem sua
disseminação. Vinte e sete genes codificam resistência a bombas de efluxo, 12
codificam, às proteínas de proteção ribossomal, 3 codificam enzimas de inativação
enzimática e um único possui mecanismo desconhecido (Figura 1) (ROBERTS,
2011).
O primeiro microrganismo a apresentar resistência a esse antimicrobiano,
Shigella dysenteriae, foi isolado em 1953. A resistência antimicrobiana às
tetraciclinas, na maioria das vezes, deve-se à aquisição de novos genes que,

23
geralmente, estão associados a elementos móveis (plasmídeos, transposons e/ou
integrons) (ROBERTS, 2003; ROBERTS, 2005).
Setenta e seis espécies de bactérias gram negativas e 47 gram positivas de
origem humana e animal já foram identificadas carreando genes tet codificadores de
bombas de efluxo, sendo o tetB o mais comum em microrganismos gram negativos.
Quarenta e nove espécies gram negativas e 35 gram positivas foram observadas
carreando um ou mais genes codificadores de proteção ribossomal, sendo o tetM o
gene encontrado com maior frequência (ROBERTS, 2011).
Figura 1. Classificação dos genes codificadores de resistência às tetraciclinas
segundo os seus mecanismos (ROBERTS, 2003).

24
1.1.3 Resistência às Quinolonas
A primeira Quinolona a ser introduzida na prática clínica foi o Ácido
Nalidíxico em 1962. Desde então, modificações em sua estrutura resultaram no
surgimento de quatro gerações de quinolonas, baseadas na atividade antimicrobiana
desse composto (Quadro 6) (OLIPHANT e GREEN, 2002).
Esses antimicrobianos agem por meio da inibição da síntese de DNA
bacteriano, ligando-se em complexos que se formam entre a DNA girase ou a
Topoisomerase IV, gerando uma alteração conformacional nessas enzimas, o que
impede a replicação do DNA e provoca a morte rápida do microrganismo
(HAWKEY, 2003).
Quadro 6: Classificação das Quinolonas.
Geração Antimicrobiano Atividade Antimicrobiana
1ª geração Ácido Nalidíxico
Cinoxacina
Enterobactérias
2ª geração Classe 1:
Lomefloxacina,
Norfloxacina, Enoxacina
Classe 2:
Ofloxacina
Ciprofloxacina
Enterobactérias
Enterobactérias, patógenos
atípicos, Pseudomonas
aeruginosa (ciprofloxacina)
3ª geração Levofloxacina
Sparfloxacina
Gatifloxacina
Moxifloxacina
Enterobactérias, patógenos
atípicos, Streptococcus
4ª geração Trovafloxacina Enterobactérias, Pseudomonas
aeroginosa, patógenos
atípicos, Staphylococcus
aureus meticilina resistente,
Streptococcus, anaeróbios
Fonte: OLIPHANT e GREEN, 2002.

25
A resistência as quinolonas tornou-se um problema desde a introdução do
Ácido Nalidíxico e têm sido estudada em isolados de humanos e animais, nas últimas
três décadas. Três mecanismos de resistência a esses antimicrobianos são atualmente
conhecidos: mutações que alteram o sítio alvo do composto, bombas de efluxo e
proteínas que protegem o sítio alvo da célula bacteriana contra o efeito das
quinolonas (JACOBY, 2005).
Os mecanismos de resistência às quinolonas eram considerados apenas
cromossômicos, entretanto, em 1998, foi detectado um gene plasmidial, denominado
qnrA, mediador de resistência à quinolonas presente em um isolado de Klebsiella
pneumoniae. Outros dois determinantes, QnrB e QnrS, também foram identificados
logo após o QnrA, em isolados de enterobactérias (POIREL, 2008).
As proteínas qnr, são formadas por peptídeos e codificadas por genes qnr.
Essas agem protegendo a região alvo das Quinolonas, impedindo a ligação do
antimicrobiano na DNA-girase, assim bloqueando a ação do mesmo. Os três
principais determinantes qnr (qnrA, qnrB, qnrS) já foram observados em todo o
mundo, sobretudo em isolados de Klebisiella pneumoniae, Escherichia coli,
Enterobacter spp e Salmonella spp, e os genes qnrA e qnrB estão geralmente
associados a plasmídeos que carreiam genes codificadores de ESBL (blaCTX, blaTEM,
blaSHV e blaVEB-1) ou AmpC (blaFOX-5 e blaDHA-1) (NORDMANN e POIREL, 2005;
POIREL, 2008).

26
1.2 Disseminação da Resistência Bacteriana
A resistência bacteriana aos antibióticos é um persistente problema na Saúde
Pública (LANDERS et al.2012). A exposição a antimicrobianos é considerada a
principal causa da emergência e disseminação da resistência bacteriana. O uso
extensivo desses compostos na medicina (humana e animal), na criação de animais e
na agricultura, torna-se um fator contribuinte para a seleção de microrganismos
resistentes. Estudos no Brasil e no mundo demonstram a disseminação de genes de
resistência no ambiente, na clínica e nos alimentos, mediados por bactérias de
importância clínica (SCHWARTZ et al., 2003; RIBEIRO et al., 2008; MAIA et al.,
2009; CANTÓN e MOROSINI, 2011).
A ampla utilização de antimicrobianos na Medicina Humana ou Animal e
Agricultura (diretamente, por meio da aplicação de antibióticos em cultivos e
indiretamente pela utilização de fertilizantes naturais como por exemplo, as fezes de
animais) e a própria seleção natural de microrganismos resistente têm contribuído
para a seleção e disseminação destes no meio ambiente, por meio de forças físicas
(vento e o movimento de bacias hidrográficas) e forças biológicas (animais e
atividades humanas) (ALLEN et al., 2010).
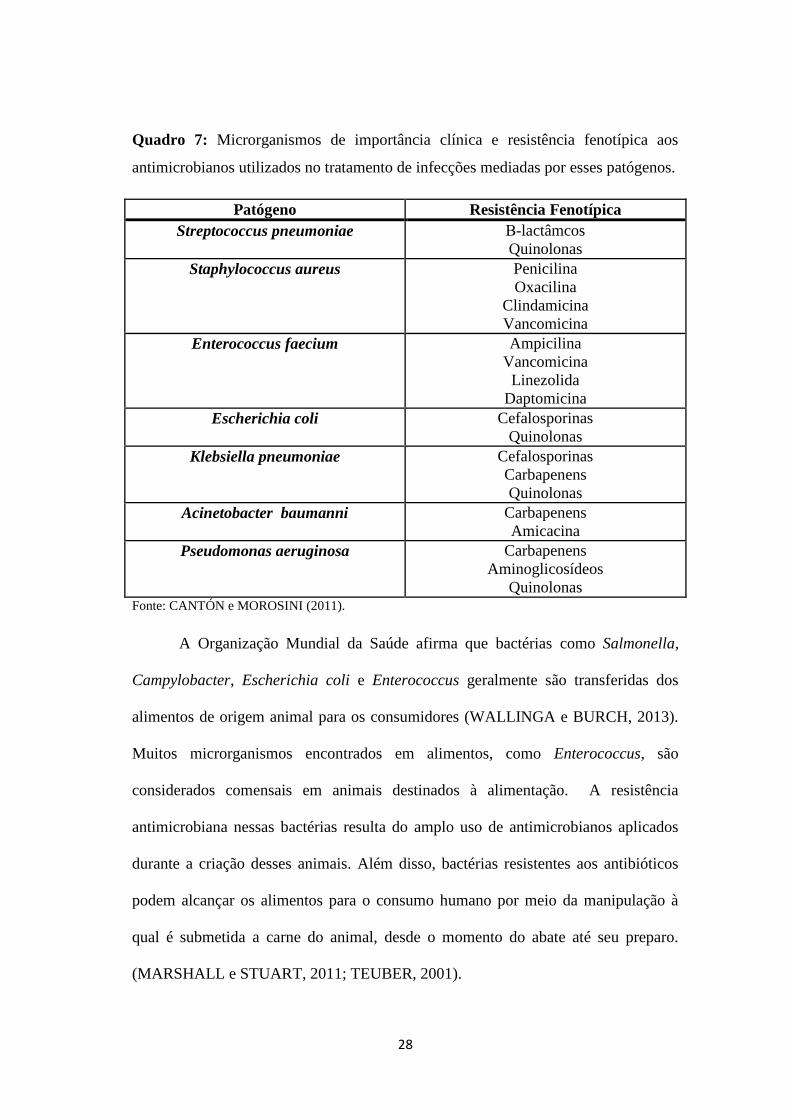
O maior problema da disseminação da resistência bacteriana é a aquisição de
genes codificadores de resistência por microrganismos de importância clínica. A
última década mostrou a mudança da sensibilidade de diversas bactérias aos
antimicrobianos comumente utilizados em infecções mediadas por esses patógenos
(Quadro 7). A resistência em agentes patogênicos gram negativos, tais como
Escherichia coli, Salmonella enterica e Klebsiella pneumoniae apresenta uma

27
ligação com o antimicrobiano de escolha (β-lactâmico) em infecções mediadas por
esses patógenos (RICE, 2009; DAVIES e DAVIES, 2010).
Atualmente, os antimicrobianos são utilizados na criação de animais com
busca de três principais objetivos: tratamento de infecções, profilaxia e promoção de
crescimento. As informações sobre a utilização desses compostos são muito escassas
e as estimativas sobre quantidades estão disponíveis em poucos países (BOGAARD
e STOBBERINGH, 1999).
A prática da introdução dos antimicrobianos em rações de animais, com o
objetivo de promover o crescimento, foi descrita pela primeira vez em 1940, quando
uma linhagem de frangos alimentados com resíduos da fermentação da tetraciclina
cresceram mais rapidamente que o grupo controle. Passados 50 anos, o aumento do
uso de doses subterapêuticas de antimicrobianos em rações de animais tem sido
descrito constantemente (MORENO-BONDI, 2009; ZHAO et al., 2010; MODI et al.,
2011; WEI et al., 2011). Esses promotores agem no trato gastrointestinal dos
animais, sobretudo, em organismos gram positivos, reduzindo a utilização bacteriana
de resíduos essenciais, permitindo a síntese adequada de proteínas, vitaminas e
outros fatores de crescimento, reduzindo a produção de toxinas bacterianas, entre
outros compostos (BARTON, 2000).
28
Quadro 7: Microrganismos de importância clínica e resistência fenotípica aos
antimicrobianos utilizados no tratamento de infecções mediadas por esses patógenos.
Patógeno Resistência Fenotípica
Streptococcus pneumoniae Β-lactâmcos
Quinolonas
Staphylococcus aureus Penicilina
Oxacilina
Clindamicina
Vancomicina
Enterococcus faecium Ampicilina
Vancomicina
Linezolida
Daptomicina
Escherichia coli Cefalosporinas
Quinolonas
Klebsiella pneumoniae Cefalosporinas
Carbapenens
Quinolonas
Acinetobacter baumanni Carbapenens
Amicacina
Pseudomonas aeruginosa Carbapenens
Aminoglicosídeos
Quinolonas Fonte: CANTÓN e MOROSINI (2011).
A Organização Mundial da Saúde afirma que bactérias como Salmonella,
Campylobacter, Escherichia coli e Enterococcus geralmente são transferidas dos
alimentos de origem animal para os consumidores (WALLINGA e BURCH, 2013).
Muitos microrganismos encontrados em alimentos, como Enterococcus, são
considerados comensais em animais destinados à alimentação. A resistência
antimicrobiana nessas bactérias resulta do amplo uso de antimicrobianos aplicados
durante a criação desses animais. Além disso, bactérias resistentes aos antibióticos
podem alcançar os alimentos para o consumo humano por meio da manipulação à
qual é submetida a carne do animal, desde o momento do abate até seu preparo.
(MARSHALL e STUART, 2011; TEUBER, 2001).

29
Os consumidores estão expostos, portanto, a microrganismos resistentes por
meio do consumo de produtos de origem animal. É cientificamente comprovado que
alimentos de origem animal contêm alta quantidade de bactérias carreadoras de genes
que codificam resistência a diversos antibióticos. As técnicas de biologia molecular
têm contribuído cada vez mais para demonstrar que o mesmo gene de resistência
encontrado em uma bactéria isolada de um alimento pode estar presente em uma
cepa isolada de humano, comprovando a disseminação da resistência bacteriana
(MARSHALL e STUART, 2011).
A ideia de que microrganismos comuns no trato gastrointestinal de animais
poderiam adquirir resistência ao antimicrobiano utilizado como promotor de
crescimento e essa ser transferida para o homem por meio da alimentação, deu-se
pela primeira vez em 1975 em uma demonstração utilizando oxatetraciclina em uma
criação de frangos. A utilização desse antimicrobiano como promotor de crescimento
demonstrou um aumento de Escherichia coli resistente à Tetraciclina nos frangos,
bem como a mesma resistência desse microrganismo, só que dessa vez isolados do
trato gastrointestinal de pessoas que moravam na fazenda dessa criação (ROLAIN,
2013).
Geralmente, infecções mediadas por alimentos restringem-se ao trato
gastrointestinal do homem. Entretanto, estudos recentes mostram o aumento de
infecções no trato urinário mediadas por Escherichia coli, ligadas ao consumo de
produtos de origem animal (sobretudo as aves) colonizados por esse microrganismo
(NORDSTROM et al.,2013). Em 2007, JOHNSON et al. (2007) realizaram um

30
estudo comparativo em Minnesota e Wisconsin entre isolados de Escherichia coli da
população humana e aviária. Com o resultado observaram que os isolados de
Escherichia coli da população humana eram mais semelhantes àqueles isolados das
aves do que da própria microbiota do trato gastrointestinal. Esse estudou sugeriu que
a ingestão ou manipulação de aves seria a principal fonte de aquisição de Escherichia
coli resistente a antimicrobianos pela população daquela região.
Genes codificadores de resistência isolados de microrganismos de origem
alimentar já foram isolados de seres humanos, comprovando indiretamente a
disseminação por meio da manipulação ou consumo do alimento. Em 2001,
SORESEN et al. (2001) confirmaram o risco do consumo de produtos cárneos
colonizados com bactérias resistentes. Esse estudo foi realizado com 18 voluntários,
os quais ingeriram uma mistura contendo cepas de Enterecococcus faecium
resistentes aos glicopeptídeos, isoladas de carne de frango ou porco. No 6º e 14º dia
após a ingestão, as cepas ingeridas por meio da mistura foram isoladas das fezes
destes voluntários, comprovando que esses microrganismos são capazes de
sobreviver e se multiplicar nas condições gástricas de um ser humano.
Uma vez que microrganismos resistentes são capazes de sobreviver no
intestino do homem, é importante ressaltar a possibilidade da aquisição de novos
genes de resistência por outras bactérias, por meio de mecanismos já citados
anteriormente. Porém, se ocorrer morte bacteriana, tanto na carne que será
consumida, quanto no intestino do homem, o material genético desse microrganismo

31
pode ser liberado e incorporado por outras bactérias, por meio de um mecanismo
denominado transformação natural.
WANG et al. (2005) demonstraram esta teoria, por meio da realização de um
estudo observando o mecanismo da transformação. Cepas de Lactococcus lactis,
isoladas de queijo e leite cru, continham um plasmídeo carreador do gene tetS/M,
codificador de resistência à tetraciclina. Para demonstrar o risco do consumo de
alimentos colonizados por bactérias carreadoras de genes de resistência, foi realizada
a transformação, in vitro, entre o plasmídeo carreador do gene que codifica
resistência a tetraciclina e a bactéria Steptococcus mutans, comum no trato oral de
seres humanos. A transformação foi confirmada por meio da aquisição do plasmídeo
pelo Streptococcus mutans, assim demonstrando a possibilidade da ocorrência da
transferência natural, que contribui com a disseminação da resistência bacteriana.
Dessa forma, estudos tem demonstrado que a microbiota humana natural
possui bactérias carreadoras de genes de resistência e que esses podem ser
transferidos para microrganismos patogênicos que por alguma ocasião estejam no
trato gastrointestinal do homem. Consequentemente, devido ao amplo uso dos
antimicrobianos na medicina humana e animal e na agricultura, estudos evidenciaram
que microrganismos carreadores de genes de resistência podem estar presentes em
humanos, alimentos de origem animal, vegetais, entre outros.
Em 1995, a Dinamarca estabeleceu o DANMAP (Danish Integrated
Antimicrobial Resistance Monitoring and Research Programme), um sistema de

32
monitoramento de resistência bacteriana em animais de fazenda, para analisar o
impacto da retirada de promotores de crescimento das rações. Com isso, o uso de
alguns antimicrobianos foi banido da criação animal. Devido a esse programa, a
Dinamarca apresentou grande redução do uso de antimicrobianos e da resistência
bacteriana a esses compostos (COGLIANI et al., 2011).
Diversos países demonstram a preocupação em relação a disseminação da resistência
bacteriana em produto de origem animal. Em 2000, a Suíça estabeleceu um órgão,
SVARM (Swedish Veterinary Antimicrobial Resistance), para monitorar a
resistência bacteriana em animais de fazenda. Com esse programa, desenvolveram
diretrizes para rações, medicação, gestão e higiene para manter os animais saudáveis
e prevenir infecções, assim diminuindo a aplicação de antimicrobianos na criação
desses (COGLIANI et al, 2011).
Em 2002, na Holanda, foi estabelecido o MARAN (Monitoring of
Antimicrobial Resistance and Antibiotic Usage in Animals in the Netherlands), um
sistema de monitoramento da resistência bacteriana em microrganismos de origem
alimentar. Com a retirada dos promotores de crescimento da alimentação animal,
observou-se um aumento do uso de antimicrobianos com fins terapêuticos, devido a
não utilização de medidas de controle contra infecções, falta de monitoramento do
governo e não comprometimento dos criadores. Isso demonstra que a retirada de
promotores de crescimento de criações animais holandeses precisa de um
acompanhamento adequado e, principalmente, adequação do sistema de criação para
controle de infecções (COGLIANI et al.,2011).

33
Nos Estados Unidos, o CDC (Centers for Disease Control and Prevention)
fornece informações sobre o uso abusivo de antibióticos e as consequências da
resistência bacteriana a esses compostos. Além disso, esse órgão lançou
recentemente, pela primeira vez, um relatório para conscientização da população,
para que exista uma ação imediata em relação a esse assunto, que afeta a Saúde
Pública do país (http://www.cdc.gov/drugresistance/threat-report-2013/pdf/ar-
threats-2013-508.pdf).
Em 2009 no Brasil, foi fundado o Projeto EUREQA (Epidemiologia do Uso e
da Resistência Bacteriana a Quimioterápicos e Antibióticos na população) que estuda
as correlações de tempo e espaço do uso de antimicrobianos pela população
brasileira e as infecções bacterianas de alta prevalência na comunidade
(www.dpi.inpe.br/eureqa/index.php). Entretanto, o projeto não abrange o uso de
antimicrobianos como promotores de crescimento em rações de animais.
1.3 Avicultura no Brasil
A carne de frango, comum na dieta do homem pode ser considerada
reservatório de genes de resistência. Estudos realizados no mundo todo apontam a
disseminação de genes que codificam resistência a diversos antimicrobianos de
importância clínica (β-lactâmicos, Quinolonas, Tetraciclinas, entre outros) em carne
de frango (GAROFALO et al., 2006; OLESEN et al.,2008; HUANG et al., 2009;
DHANJI et al., 2010 KIM et al., 2011).
O Brasil destaca-se no mercado internacional de carnes com o frango sendo
um dos seus principais produtos de exportação do país. O desenvolvimento da

34
avicultura brasileira teve início em 1960 com grupos agroindustriais e desde então se
tornou competitiva mundialmente. Esse fato deve-se aos baixos custos da produção,
qualidade e higiene na cadeia produtiva e desenvolvimento de estudos e novas
tecnologias para o setor (WINCK e MACHADO, 2011).
No ano de 2010, as exportações de carne de frango no Brasil chegaram a
quase 4 milhões de toneladas, o que permitiu ao país se manter como o líder mundial,
posição conquistada desde 2004.
A avicultura também tem grande impacto nacional. Um estudo realizado pelo
MAPA mostra a preferência crescente dos consumidores brasileiros por carne de
frango, cujo crescimento projetado é de 3% ao ano no período de 2007/2008 à
2017/2018. De acordo com dados também do Ministério da Agricultura, o Brasil
daqui a 10 anos terá capacidade de suprir o consumo interno e as demandas de países
importadores desse tipo de carne (MAPA, 2010; BASSO et al., 2012).
Em 2012, a produção de carne de frango no Brasil chegou a mais de 12
milhões de toneladas, apresentando uma pequena redução (3,17%) em relação a
2011, sendo São Paulo, Rio Grande do Sul, Santa Catarina e Paraná os estados com
maior índice de abate de carne de frango. Mesmo assim, o Brasil manteve a posição
de maior exportador mundial e terceiro maior produtor de carne de frango, ficando
atrás apenas dos Estados Unidos e China. Do volume total da produção do país, 69%
foi referente ao consumo nacional e 31% às exportações. Assim, o consumo de carne
de frango per capita no ano de 2012 atingiu 45 quilos por habitante (UBABEF,
2013).

35
1.4 Disseminação da Resistência Bacteriana em carne de frango no Brasil
Assim como em outros países, no Brasil, a aplicação dos antimicrobianos na
avicultura tem três principais objetivos: terapêutico, profilático e promotor de
crescimento. Antimicrobianos de importância clínica são amplamente utilizados na
indústria animal.
O setor avícola está em constante crescimento e há mais de 25 anos os
antimicrobianos estão sendo empregados rotineiramente na criação de aves. Essa
prática tem gerado sérias consequências no que diz a respeito ao surgimento e
disseminação de microrganismos resistentes, colocando em risco a saúde humana. A
pesquisa de novos compostos para serem utilizados como promotores de crescimento
não é considerada fácil, uma vez que a maioria das substâncias com efeitos
antimicrobianos descobertas são consideradas tóxicas ou instáveis para os animais
(SANTOS et al., 2008).
A exigência dos órgãos de defesa dos consumidores, como o IDEC (Insituto
de Defesa do Consumidor) e de outros países importadores da carne de frango do
Brasil, como a União Europeia, quanto ao uso de antimicrobianos na ração animal,
tem sido importante na restrição do uso desses compostos como aditivos na ração
animal (SANTOS et al., 2008).
No Brasil, o MAPA (Ministério da Agricultura, Pecuária e Abastecimento) é o órgão
responsável pela regulamentação e fiscalização dos produtos utilizados na
alimentação animal, por meio da Divisão de Fiscalização de Aditivos, da

36
Coordenação de produtos para Alimentação Animal (CPAA). De acordo com esse
órgão, os antimicrobianos proibidos na alimentação animal são: Avoparcina (Ofício
Circular DFPA nº 047/1998), Cloranfenicol e Nitrofuranos (Instrução Normativa nº
09, 27/06/2003), anfenicóis, tetraciclinas, β-lactâmicos (benzilpenicilinas e
cefalosporinas), quinolonas e sulfonamidas sistêmicas (Instrução Normativa nº 26,
09/07/2009).
Estudos fenotípicos têm sido realizados abordando a resistência bacteriana em
isolados bacterianos de carne de frango (RIBEIRO et al., 2008; KUCHENBECKER
et al., 2009; MAIA et al., 2009; ABREU et al., 2010; BARROS et al., 2012). Em
relação aos estudos genotípicos, WARREN et al. (2008) observaram, no Reino
Unido, o gene blaCTX-M-2 em amostras de carne de frango importadas do Brasil.
Entretanto, dados referentes à detecção de genes codificadores de resistência aos
antimicrobianos em carne de frango ainda são escassos no país.

37
2. Objetivos
2.1 Objetivo Geral
Detectar genes codificadores de resistência a diferentes classes de
antimicrobianos em amostras de carne de frango.
2.2 Objetivos Específicos
Avaliar a contagem de coliformes termotolerantes, estimando a qualidade
higiênico sanitária das amostras.
Verificar a presença de genes codificadores de resistência a antibióticos β-
lactâmicos, Tetraciclinas e Quinolonas em amostras de carne de frango,
congeladas e frescas, provenientes de mercados (congeladas), feiras livres
(resfriadas) e açougues (resfriadas).

38
3. Material e Métodos
3.1 Amostras
Para o presente estudo foram utilizadas 30 amostras de peito de frango cru
resfriados e congelados, sendo 10 de frango de supermercado (congelados), 10 de
feira livre (resfriados) e 10 de açougues (resfriados). As amostras foram
acondicionadas em recipientes térmicos e transportadas ao laboratório para que
fossem processadas.
Todas as amostras foram provenientes do município de São Paulo. Os locais
de coleta foram escolhidos baseados nas opções do consumidor quando esse tem a
intenção de adquirir a carne de frango para o consumo. Sendo assim, foi possível
analisar se há ou não presença de genes de resistência em amostras provenientes de
diferentes locais de comércio.
3.2 Contagem de Coliformes Termotolerantes
A contagem dos coliformes termotolerantes (capazes de se multiplicar a 45
ºC) permitiu estimar o padrão microbiológico sanitário das amostras de peito de
frango e os resultados foram comparados com a Resolução RDC nº 12, de 2 de
Janeiro de 2001.
3.2.1 Preparo das Amostras
Alíquotas de 25g do alimento foram pesadas em sacos plásticos estéreis e
homogeneizadas com 225 ml de salina 0,85% previamente esterilizada. Diluições
decimais a partir da diluição 10-1
até 10-8
foram preparadas em frascos contendo 90
ml de salina 0,85%.

39
3.2.2 Técnica dos Tubos Múltiplos
Para todas as amostras de peito de frango analisadas foi determinado o
número mais provável de coliformes termotolerantes de acordo com a técnica dos
tubos múltiplos. Alíquotas de 1ml de cada diluição em salina foram transferidas para
séries de três tubos contendo 9 ml de caldo Lauril Sulfato de Sódio (LST) (Difco ®)
(com tubos de Durham invertidos). Os tubos foram incubados a 37ºC, sendo a
produção de gás observada por até 48 horas. Os tubos que apresentaram crescimento
e produção de gás tiveram uma alçada do crescimento transferida para tubos
contendo 9 ml de caldo EC (Difco®) (com tubos De Durham invertidos).
Os tubos contendo caldo EC foram incubados por até 48 horas a 45 ºC.
Aqueles que apresentaram crescimento e produção de gás, indicando presença de
coliformes termotolerantes, foram considerados para o cálculo do número mais
provável (NMP) de coliformes termotolerantes por grama de alimento (Figura 2).
3.3 Detecção de Genes de Resistência
3.3.1 Extração do DNA Total
Cada uma das 30 amostras de peito de frango (1,5 Kg) foi homogeneizada
com 350 ml de caldo Luria contendo 0,5% de NaCl e o lavado dessas amostras foi
incubado a 37ºC por 24 horas. Após o período de incubação, 1 ml da cultura
bacteriana foi transferido homogeneizados e transferidos para microtubos para

40
realização da extração do DNA total (plasmidial e cromossômico) por meio de
choque térmico, segundo a técnica descrita por CHAPMAN et al. (2001).
O esquema geral de procedimentos da pesquisa de genes de resistência a
antibióticos nas amostras está apresentado na Figura 2.
A cultura foi centrifugada por 5 minutos em velocidade de 14 000 rpm em
temperatura ambiente. Após a centrifugação, o sobrenadante foi descartado e o pellet
correspondente às células bacterianas foi lavado com 1ml de água MilliQ®
estéril e
centrifugado novamente nas mesmas condições já descritas. O sobrenadante foi
descartado e o precipitado resuspenso em 200 µl de água MilliQ® e então colocado
em banho-maria a 95ºC por 10 minutos. Em seguida foi mantido durante 30 minutos
a -20ºC. Após esse período, o tubo permaneceu em temperatura ambiente até o
descongelamento de seu conteúdo e foi centrifugado a 14 000 rpm por 5 minutos. O
sobrenadante contendo o material genético foi transferido para um novo microtubo e
conservado a -20 ºC até o momento do uso.
3.3.2 Reação em cadeia pela polimerase (PCR)
Reações de PCR foram realizadas para a detecção de genes que conferem
resistência aos antimicrobianos β-lactâmicos, Tetraciclinas e Quinolonas, utilizando
iniciadores específicos. Esses antimicrobianos foram escolhidos pelo fato de serem
proibidos na criação de aves.
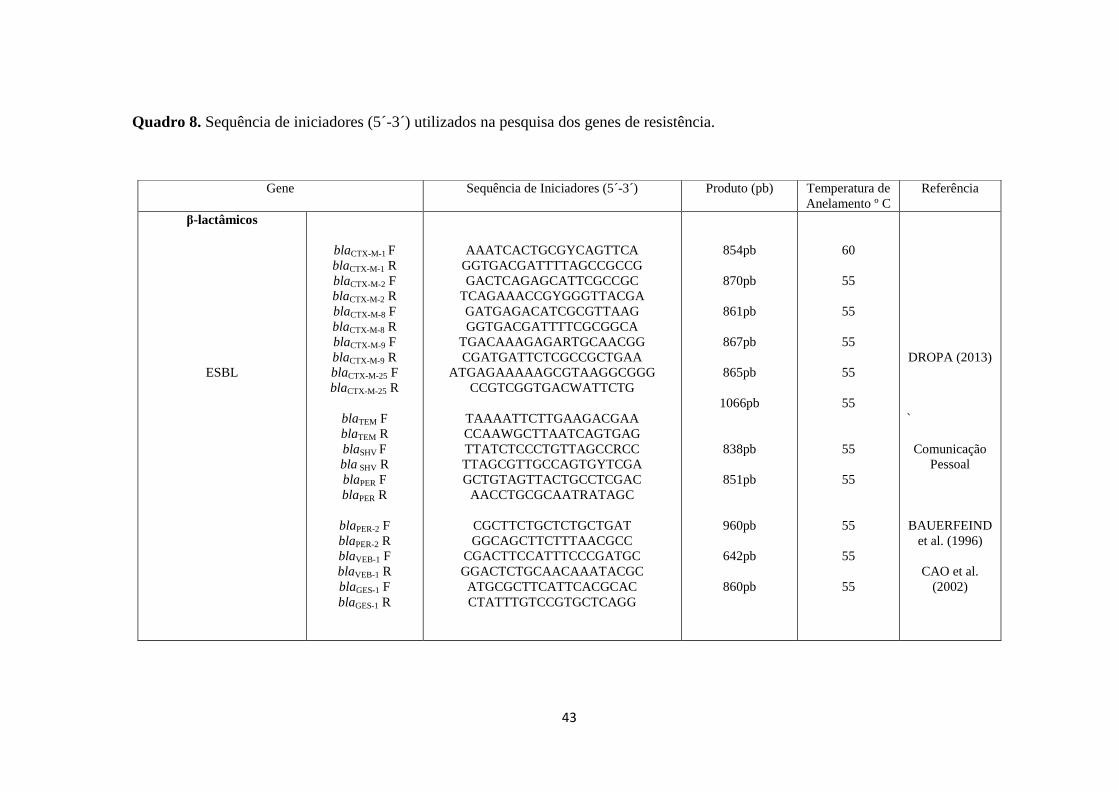
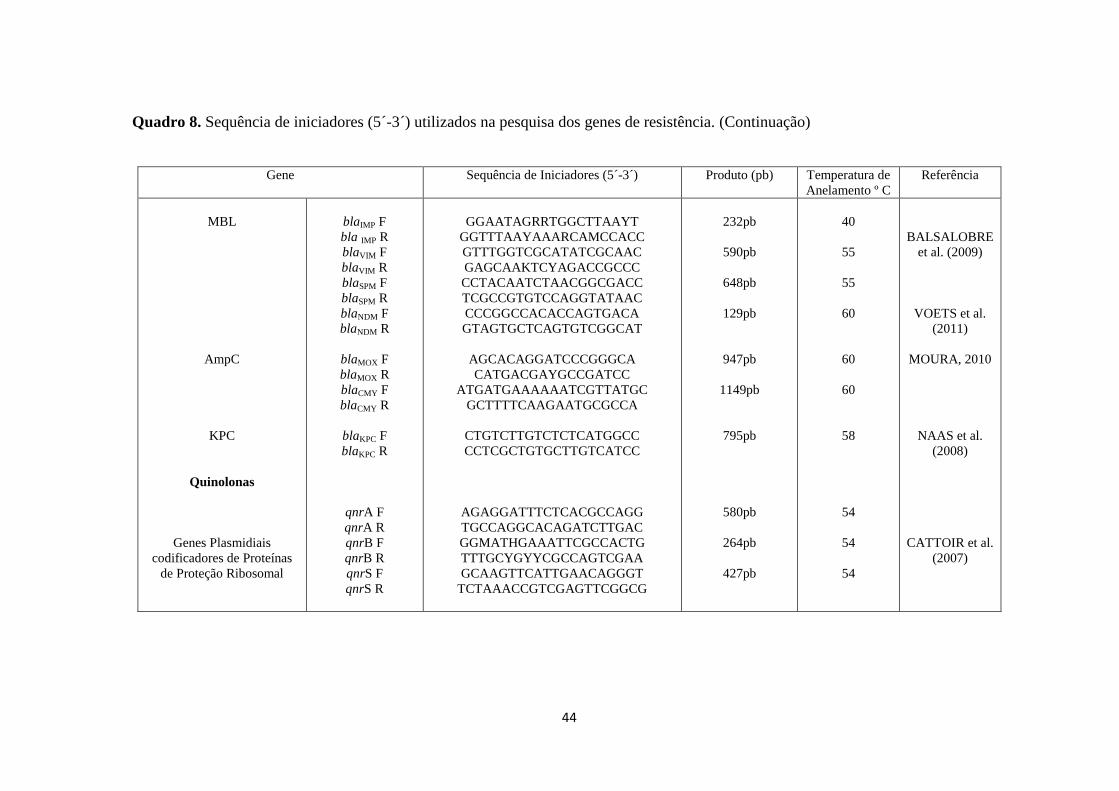
O Quadro 8 apresenta os genes pesquisados, os respectivos iniciadores e a
temperatura de anelamento de cada par de iniciadores. Os controles positivos de cada

41
gene pesquisado encontram-se na coleção de cultura do Laboratório de Prática de
Saúde Pública da FSP-USP, com exceção dos genes.
Cada microtubo de reação continha, para um volume final de 25µl, os
seguintes reagentes: 5µl de DNA; tampão 5X (Promega); 1,5mM de MgCl2
(Promega), 200 uM de DNTp (Fermentas); 0,2 uM de cada iniciador e 1,25U de
GoTaq®
Polymerase (Promega) e água MilliQ
® para completar o volume de 25µl.
As reações ocorreram em termociclador (Mastercycler Gradient, Eppendorf)
com um ciclo de desnaturação inicial a 95ºC por 5 min., seguido de 30 ciclos
composto de denaturação a 95 ºC por 1 min., anelamento por 1 min. (Quadro 8),
extensão a 72ºC a 1 min. (dependendo do tamanho do fragmento), e extensão final a
72º C por 10 min.
3.4 Visualização dos produtos amplificados
A visualização da presença dos fragmentos amplificados para os diferentes
genes estudados foi realizada por meio de eletroforese em gel de agarose corado com
brometo de etídeo (1µg/ml), sob intensidade de corrente de 6V/cm. O tamanho do
fragmento foi determinado por comparação com o marcador de peso molecular 100
bp (MassRuler TM
DNA Ladder, Fermentas) utilizado em cada gel.
O gel foi visualizado sob luz ultravioleta 302nm e as imagens foram
registradas pelo sistema Epi Chemi II Darkroom, UVP Bioimaging Systems e
Software Labworks.
Alguns fragmentos de tamanho correspondentes aos genes pesquisados foram
selecionados e purificados com o kit comercial (illustra TM
GFTX TM
DNA and Gel

42
Band Purification Kit, GE Healthcare, UK), seguindo as orientações do fabricante, e
encaminhados para seqüenciamento, para confirmação e do gene em questão.
As sequencias foram alinhadas manualmente utilizando o programa Bioedit
Sequence Alignment Editor (HALL, 1999) e comparadas às sequencias dos genes
codificadores dos diferentes grupos em estudo, disponíveis em BUSH e JACOBY
(http://www.lahey.org/Studies) e no banco de dados GenBank
(www.ncbi.nih.gov/genbank).
43
Quadro 8. Sequência de iniciadores (5´-3´) utilizados na pesquisa dos genes de resistência.
Gene Sequência de Iniciadores (5´-3´) Produto (pb) Temperatura de
Anelamento º C
Referência
β-lactâmicos
ESBL
blaCTX-M-1 F
blaCTX-M-1 R
blaCTX-M-2 F
blaCTX-M-2 R
blaCTX-M-8 F
blaCTX-M-8 R
blaCTX-M-9 F
blaCTX-M-9 R
blaCTX-M-25 F
blaCTX-M-25 R
blaTEM F
blaTEM R
blaSHV F
bla SHV R
blaPER F
blaPER R
blaPER-2 F
blaPER-2 R
blaVEB-1 F
blaVEB-1 R
blaGES-1 F
blaGES-1 R
AAATCACTGCGYCAGTTCA
GGTGACGATTTTAGCCGCCG
GACTCAGAGCATTCGCCGC
TCAGAAACCGYGGGTTACGA
GATGAGACATCGCGTTAAG
GGTGACGATTTTCGCGGCA
TGACAAAGAGARTGCAACGG
CGATGATTCTCGCCGCTGAA
ATGAGAAAAAGCGTAAGGCGGG
CCGTCGGTGACWATTCTG
TAAAATTCTTGAAGACGAA
CCAAWGCTTAATCAGTGAG
TTATCTCCCTGTTAGCCRCC
TTAGCGTTGCCAGTGYTCGA
GCTGTAGTTACTGCCTCGAC
AACCTGCGCAATRATAGC
CGCTTCTGCTCTGCTGAT
GGCAGCTTCTTTAACGCC
CGACTTCCATTTCCCGATGC
GGACTCTGCAACAAATACGC
ATGCGCTTCATTCACGCAC
CTATTTGTCCGTGCTCAGG
854pb
870pb
861pb
867pb
865pb
1066pb
838pb
851pb
960pb
642pb
860pb
60
55
55
55
55
55
55
55
55
55
55
DROPA (2013)
`
Comunicação
Pessoal
BAUERFEIND
et al. (1996)
CAO et al.
(2002)
44
Quadro 8. Sequência de iniciadores (5´-3´) utilizados na pesquisa dos genes de resistência. (Continuação)
Gene Sequência de Iniciadores (5´-3´) Produto (pb) Temperatura de
Anelamento º C
Referência
MBL
AmpC
KPC
Quinolonas
Genes Plasmidiais
codificadores de Proteínas
de Proteção Ribosomal
blaIMP F
bla IMP R
blaVIM F
blaVIM R
blaSPM F
blaSPM R
blaNDM F
blaNDM R
blaMOX F
blaMOX R
blaCMY F
blaCMY R
blaKPC F
blaKPC R
qnrA F
qnrA R
qnrB F
qnrB R
qnrS F
qnrS R
GGAATAGRRTGGCTTAAYT
GGTTTAAYAAARCAMCCACC
GTTTGGTCGCATATCGCAAC
GAGCAAKTCYAGACCGCCC
CCTACAATCTAACGGCGACC
TCGCCGTGTCCAGGTATAAC
CCCGGCCACACCAGTGACA
GTAGTGCTCAGTGTCGGCAT
AGCACAGGATCCCGGGCA
CATGACGAYGCCGATCC
ATGATGAAAAAATCGTTATGC
GCTTTTCAAGAATGCGCCA
CTGTCTTGTCTCTCATGGCC
CCTCGCTGTGCTTGTCATCC
AGAGGATTTCTCACGCCAGG
TGCCAGGCACAGATCTTGAC
GGMATHGAAATTCGCCACTG
TTTGCYGYYCGCCAGTCGAA
GCAAGTTCATTGAACAGGGT
TCTAAACCGTCGAGTTCGGCG
232pb
590pb
648pb
129pb
947pb
1149pb
795pb
580pb
264pb
427pb
40
55
55
60
60
60
58
54
54
54
BALSALOBRE
et al. (2009)
VOETS et al.
(2011)
MOURA, 2010
NAAS et al.
(2008)
CATTOIR et al.
(2007)
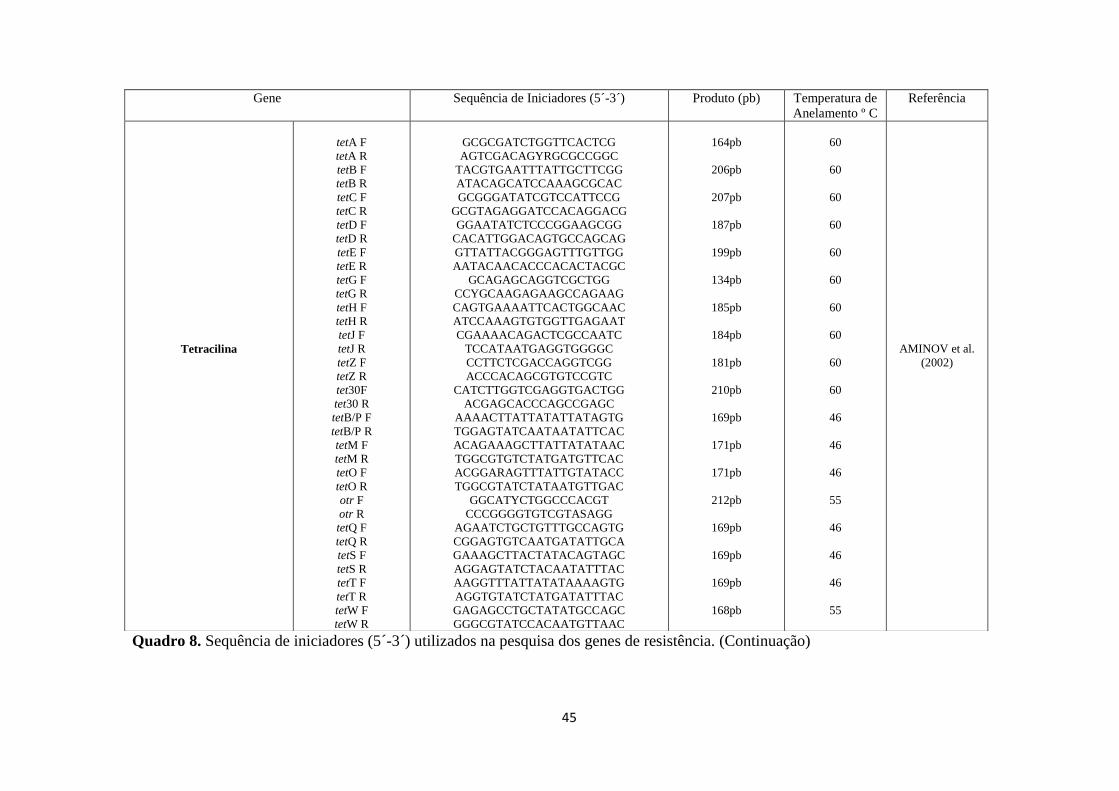
45
Quadro 8. Sequência de iniciadores (5´-3´) utilizados na pesquisa dos genes de resistência. (Continuação)
Gene Sequência de Iniciadores (5´-3´) Produto (pb) Temperatura de
Anelamento º C
Referência
Tetracilina
tetA F
tetA R
tetB F
tetB R
tetC F
tetC R
tetD F
tetD R
tetE F
tetE R
tetG F
tetG R
tetH F
tetH R
tetJ F
tetJ R
tetZ F
tetZ R
tet30F
tet30 R
tetB/P F
tetB/P R
tetM F
tetM R
tetO F
tetO R
otr F
otr R
tetQ F
tetQ R
tetS F
tetS R
tetT F
tetT R
tetW F
tetW R
GCGCGATCTGGTTCACTCG
AGTCGACAGYRGCGCCGGC
TACGTGAATTTATTGCTTCGG
ATACAGCATCCAAAGCGCAC
GCGGGATATCGTCCATTCCG
GCGTAGAGGATCCACAGGACG
GGAATATCTCCCGGAAGCGG
CACATTGGACAGTGCCAGCAG
GTTATTACGGGAGTTTGTTGG
AATACAACACCCACACTACGC
GCAGAGCAGGTCGCTGG
CCYGCAAGAGAAGCCAGAAG
CAGTGAAAATTCACTGGCAAC
ATCCAAAGTGTGGTTGAGAAT
CGAAAACAGACTCGCCAATC
TCCATAATGAGGTGGGGC
CCTTCTCGACCAGGTCGG
ACCCACAGCGTGTCCGTC
CATCTTGGTCGAGGTGACTGG
ACGAGCACCCAGCCGAGC
AAAACTTATTATATTATAGTG
TGGAGTATCAATAATATTCAC
ACAGAAAGCTTATTATATAAC
TGGCGTGTCTATGATGTTCAC
ACGGARAGTTTATTGTATACC
TGGCGTATCTATAATGTTGAC
GGCATYCTGGCCCACGT
CCCGGGGTGTCGTASAGG
AGAATCTGCTGTTTGCCAGTG
CGGAGTGTCAATGATATTGCA
GAAAGCTTACTATACAGTAGC
AGGAGTATCTACAATATTTAC
AAGGTTTATTATATAAAAGTG
AGGTGTATCTATGATATTTAC
GAGAGCCTGCTATATGCCAGC
GGGCGTATCCACAATGTTAAC
164pb
206pb
207pb
187pb
199pb
134pb
185pb
184pb
181pb
210pb
169pb
171pb
171pb
212pb
169pb
169pb
169pb
168pb
60
60
60
60
60
60
60
60
60
60
46
46
46
55
46
46
46
55
AMINOV et al.
(2002)

46
Figura 2. Esquema geral da metodologia aplicada para pesquisa de genes de resistência em amostras de frango.

47
4. Resultados
4.1 Coliformes Termotolerantes
Todas as amostras analisadas no presente estudo foram consideradas próprias
para o consumo, de acordo com a Resolução RDC nº 12.
4.2 Genes codificadores de resistência aos β-Lactâmicos
Das 30 amostras de carne de frango estudadas, 17 (56,7%) apresentaram um
ou mais genes codificadores de resistência aos antimicrobianos β-Lactâmicos,
(Anexo 1). Os genes codificadores de KPC e MBL não foram detectados nas
amostras estudadas.
4.2.1 Genes codificadores de ESBL
A presença do fragmento de 854pb correspondente ao grupo blaCTX-M-1 foi
detectado em 4 amostras de frango analisadas. Já o grupo blaCTX-M-2 foi observado
em 10 e o grupo blaCTX-M-8 estava presente em 6 amostras. Quatro amostras de carne
de frango, apresentaram o fragmento correspondente ao grupo blaTEM e 5
apresentaram o fragmento correspondente ao grupo blaSHV. Os fragmentos
correspondentes aos grupos blaCTX-M-9, blaCTX-M-25, blaPER, blaVEB e blaGES não foram
detectados nas amostras de frango estudadas.
Para a confirmação e identificação dos genes específicos dos grupos
pesquisados, os produtos amplificados foram enviados ao sequenciamento. Foi
confirmada a presença dos genes blaCTX-M-1 (pertencente ao grupo CTX-M-1) em 4
amostras, sendo 2 de supermercados e 2 d açougues, blaCTX-M-2 (pertencente ao grupo
CTX-M-2) em 3 amostras de supermercados e 1 de açougue, blaCTX-M-8 (pertencente
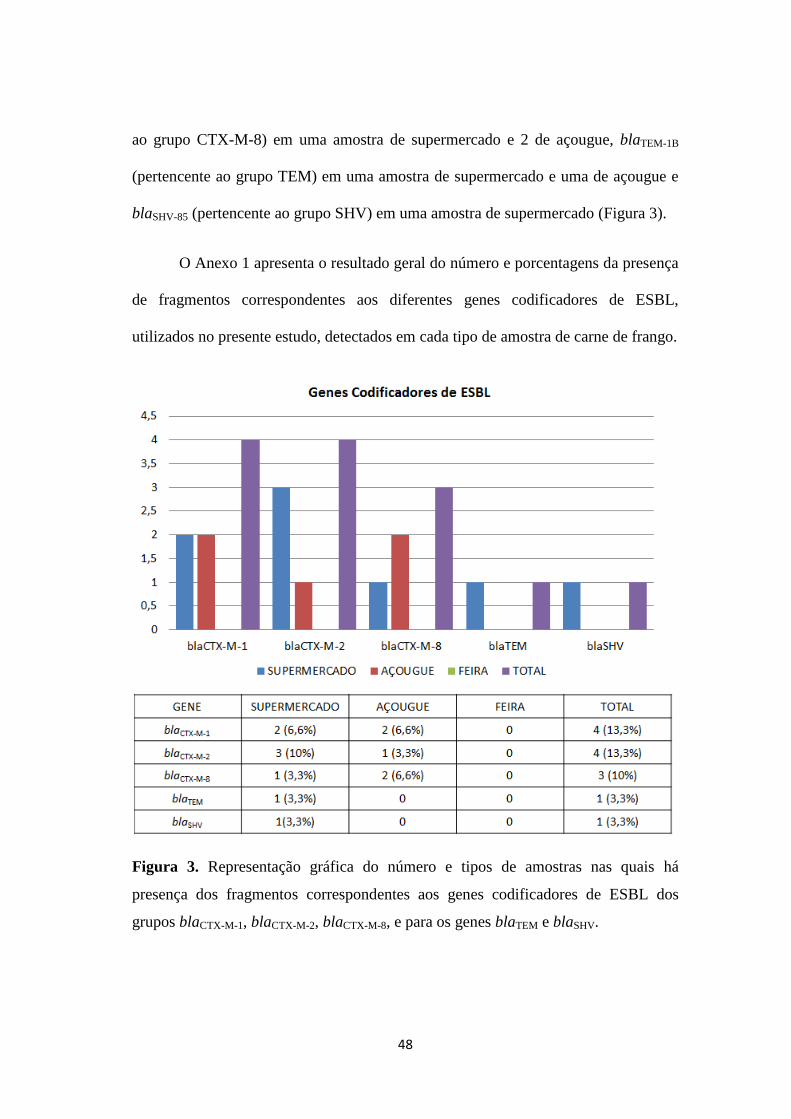
48
ao grupo CTX-M-8) em uma amostra de supermercado e 2 de açougue, blaTEM-1B
(pertencente ao grupo TEM) em uma amostra de supermercado e uma de açougue e
blaSHV-85 (pertencente ao grupo SHV) em uma amostra de supermercado (Figura 3).
O Anexo 1 apresenta o resultado geral do número e porcentagens da presença
de fragmentos correspondentes aos diferentes genes codificadores de ESBL,
utilizados no presente estudo, detectados em cada tipo de amostra de carne de frango.
Figura 3. Representação gráfica do número e tipos de amostras nas quais há
presença dos fragmentos correspondentes aos genes codificadores de ESBL dos
grupos blaCTX-M-1, blaCTX-M-2, blaCTX-M-8, e para os genes blaTEM e blaSHV.

49
4.2.2 Genes codificadores de AmpC β-Lactamases
O fragmento correspondente ao gene blaMOX foi amplificado em 7 das
amostras estudadas, enquanto que o fragmento de 1149 pb correspondente ao gene
blaCMY foi amplificado em 4 amostras de carne de frango.
Para a confirmação e identificação dos genes específicos dos grupos
pesquisados, os produtos amplificados foram enviados ao sequenciamento onde foi
confirmada apenas a identificação do gene blaMOX5-6 (pertencente ao grupo MOX)
em uma amostra de frango de feira (Anexo 1 e Figura 4). Genes pertencentes ao
grupo CMY não foram identificados no sequenciamento.
Figura 4. Representação gráfica do número e tipos de amostras nas quais foram
detectados os fragmentos correspondentes aos genes codificadores de AmpC β-
Lactamases, blaMOX e blaCMY.

50
4.3 Genes codificadores de resistência às Tetraciclinas
Em 28 amostras (93,3%) estudadas foi possível observar a presença de
fragmentos correspondentes a um ou mais genes codificadores de resistência às
Tetraciclinas pesquisados (Anexo 1). Os fragmentos identificados não foram
enviados ao sequenciamento, pois os iniciadores utilizados são específicos para cada
gene pesquisado.
4.3.1 Genes tet codificadores de Bomba de Efluxo
Os fragmentos de 164pb, 206pb, 207pb, 187pb e 199pb correspondentes aos
genes tetA, tetB, tetC, tetD e tetE, foram observados em 19, 10, 19, 17 e 9 amostras,
respectivamente (Figura 5) (Anexo 1).
Os fragmentos correspondentes aos genes tetG, tetH, tetJ, tetZ e tet30, não
foram amplificados nas amostras estudadas com os iniciadores utilizados no estudo.

51
Figura 5. Representação gráfica do número e tipos de amostras nas quais os
fragmentos correspondentes aos genes codificadores de Bombas de Efluxo tetA,
tetB, tetC, tetD e tetE foram obtidos.
4.3.2 Genes tet codificadores de Proteção Ribossomal
Treze amostras de carne de frango apresentaram o fragmento de 171pb
correspondente ao gene tetM. O fragmento correspondente ao gene tetO foi
amplificado em 4 amostras de carne de frango. Já o fragmento correspondente ao
gene tetQ foi identificado em 3 amostras. Os fragmentos correspondentes aos genes
tetS (169pb) e tetW (168pb) foram observados em 7 e 1 amostras, respectivamente
(Figura 6) (Anexo 1).

52
Não foram detectados os fragmentos correspondentes aos genes tetB(P), tetZ
e tetT nas amostras analisadas com os iniciadores propostos no estudo.
Figura 6. Representação gráfica do número e tipos de amostras nas quais foram
observados os fragmentos correspondentes aos genes codificadores de Proteínas de
Proteção Ribosssomal, tetM, tetO, tetQ, tetS e tetW.
4.4 Genes Codificadores de Resistência às Quinolonas
Duas amostras apresentaram o fragmento de tamanho esperado para o gene
qnrA. O fragmento correspondente ao gene qnrB foi identificado em 18 amostras de
carne de frango e o fragmento esperado para o gene qnrS em 27 amostras (Figura 7)
(Anexo 1).

53
Figura 7. Representação gráfica do número e tipos de amostras nas quais foram
amplificados os fragmentos correspondentes aos genes codificadores de Proteínas de
Proteção Ribosssomal, qnrA, qnrB e qnrS.
Quadro 9. Resultados obtidos para as diferentes amostras de carne de frango quanto
à identificação de genes codificadores de resistência aos β-Lactâmicos e às
Quinolonas.
Genes
Amostra blaCTX-
M-1
blaCTX-
M-2
blaCTX-
M-8 qnrA qnrB qnrS
5C + - - - - +
7C - + - - - +
8C - + - - - +
9C + + + - - +
6A + - - - + +
10A + - + - + +

54
5. Discussão
Na última década o aumento e a disseminação da resistência bacteriana em
microrganismos de origem clínica tornou-se um grave problema na Saúde Pública da
maioria dos países, devido à pressão seletiva do uso de antibióticos. Entretanto,
existem outras formas de pressão seletiva, como por exemplo, o amplo uso de
antimicrobianos na criação de animais. Com o avanço de estudos metagenômicos em
seres humanos, animais e no ambiente foi possível a descoberta de novos
reservatórios de genes codificadores de resistência a antimicrobianos (ROLAIN,
2013).
A resistência a antimicrobianos vem aumentando nos últimos anos sendo
observado desde microrganismos resistentes a apenas uma classe de antibióticos até
aqueles que apresentam resistência a diversas classes desses compostos
(multirresistência). Os mecanismos de ação envolvidos na resistência bacteriana são
estudados na clínica com frequência, porém é importante a pesquisa focar em outros
reservatórios como, por exemplo, em microrganismos de origem animal.
Antibióticos adicionados em rações de animais, utilizados como promotores
de crescimento resultam na propagação da resistência bacteriana por meio da
aquisição de genes codificadores de resistência, por microrganismos comensais do
animal, e esses podem ser transferidos para microrganismos do trato gastrointestinal
do homem por meio da alimentação (SANTOS et al., 2008).
Todos os genes escolhidos para a detecção em carne de frango nesse estudo
são codificadores de resistência a antimicrobianos proibidos na criação desses

55
animais, segundo a legislação brasileira (Instrução Normativa nº 26, 09/07/2009).
Mesmo assim, observaram-se dezessete (56,7%) amostras apresentando um ou mais
genes codificadores de resistência a antimicrobianos β-Lactâmicos, 28 (93,3%)
amostras apresentando um ou mais genes codificadores de resistência às
Tetraciclinas e 28 (93,3%) amostras apresentando um ou mais genes codificadores de
resistência às Quinolonas.
5.1 Genes codificadores de resistência aos β-Lactâmicos
5.1.1 Genes codificadores de ESBL
No presente estudo foram pesquisados 10 grupos de ESBL (CTX-M-1, CTX-
M-2, CTX-M-8, CTX-M-9. CTX-M-25, TEM, SHV, PER VEB e GES) nas 30
amostras de carne de frango. Em relação às ESBL, os genes codificadores dessa
enzima mais observados foram do grupo CTX-M (Figura 3). Essa família de
enzimas, além de disseminada na clínica, compreende as ESBL mais prevalentes no
mundo (ANDREA et al., 2013).
Quatro amostras (13,3%) apresentaram o fragmento correspondente ao gene
blaCTX-M-1 (Figura 3). No Brasil, o grupo CTX-M-1 até o momento não havia sido
reportado em amostras de carne de frango, porém está disseminado em amostras
clínicas e já foi descrito em amostras ambientais (PEIRANO et al., 2009; DROPA,
2013; OLIVEIRA et al., 2014). Em países Europeus como Bélgica, Dinamarca,
França, Itália, Holanda, Portugal e Espanha, esse gene ou esse grupo é isolado com
frequência de carne de suínos e aves (WITTUM et al., 2010; OVERDEVEST et al.,
2011; ACCOGLI et al., 2013).

56
Dentre os grupos de CTX-M pesquisados, o gene codificador de ESBL
blaCTX-M-2, foi detectado em 4 amostras de carne de frango (Figura 3). O grupo
CTX-M-2 foi identificado pela primeira vez em 1992 em cepas de Salmonella
Typhimurium, na Argentina (BAUERNFEIND et al., 1992) e são detectados com
frequência na América do Sul. A variante CTX-M-2 encontra-se disseminada em
cepas de origem clínica no Brasil (DROPA, 2013) e estudos recentes demonstram a
detecção desses genes em microrganismos como Salmonella enterica, Salmonella
Typhimurium e Escherichia coli isolados de frango (FERNANDES et al., 2009;
SILVIA e LINCOPAN, 2012; FERREIRA et al., 2014).
WARREN et al. (2008) também detectaram o gene blaCTX-M-2 no Reino
Unido, em cepas de Escherichia coli isoladas de amostras de carne de frango
importadas do Brasil, alertando que a importação desse tipo de alimento pode
representar uma das formas de disseminação desses genes. A identificação desse
gene nesse tipo de amostra mostra-se um alerta para o agronegócio do país, tendo em
vista que somos importantes exportadores de carne de frango no mundo (SILVA e
LINCOPAN, 2012).
O gene bla CTX-M-8 foi identificado em 3 amostras (10%) (sendo uma de
supermercado e 2 de açougues). Desde sua primeira descrição na literatura, ocorrida
no Brasil (BONNET et al., 2000), o gene blaCTX-M-8 (pertencente ao grupo CTX-M-8)
tem sido observado em várias espécies da família Enterobacteriaceae e em diversos
continentes. Segundo recentes estudos desenvolvidos na Espanha e Estados Unidos,
a disseminação dessa enzima e a posterior propagação na comunidade pode ser

57
consequência da transferência horizontal de genes, além da movimentação de
viajantes ou alimentos contaminados de um país para o outro (ELLER et al., 2013).
PEIRANO et al. (2006) realizaram um estudo no Brasil com isolados de
Salmonella enterica de diferentes amostras (origem clínica, fezes de animais,
animais para o consumo humano, entre outros), no qual também observaram o gene
blaCTX-M-8.
No Brasil os genes TEM e SHV já foram observados em amostras clínicas e
ambientais (BONNET et al., 2000; BONNET et al., 2001; MINARINI et al., 2009;
BALSALOBRE et al., 2010; DROPA et al., 2009; DROPA, 2013). Os grupos TEM e
SHV são menos observados no Brasil que os grupos CTX-M. DROPA, 2013,
descreveu pela primeira vez no Brasil, o gene blaTEM-15 isolado de amostra clínica.
Esse gene já tinha sido descrito na Europa e na Ásia, entretanto foi o primeiro relato
na América Latina. Em relação ao grupo SHV, o gene blaSHV-5 é considerado o gene
mais frequente na América Latina (DROPA, 2013).
No presente estudo, o gene blaTEM-1B foi detectado em 2 amostras, sendo uma
de supermercado e uma de açougue, e o gene blaSHV-85 foi identificado em uma
amostra de supermercado (Figura3). Ambos os genes não possuem perfil ESBL, ou
seja, hidrolisam apenas antimicrobianos como a Penicilina e as Cefalosporinas de
primeira geração.
A maioria das enzimas SHV, como a SHV-85 são derivadas da SHV-1, a qual
não possui perfil ESBL, e são disseminadas em cepas de Klebsiella pneumoniae. A
SHV-85 foi observada pela primeira vez em 2008, no Brasil, por GARCIA et al.,

58
2008, isolada de uma cepa clínica de Klebsiella pneumoniae. Também já foi
identificada em cepas de Klebsiella pneumoniae isoladas de amostras clínicas, na
Arábia Saudita (TAEFIK et al., 2011). Essa é a primeira descrição na literatura do
gene blaSHV-85 isolado de amostras de carne de frango.
No Brasil, não há relatos na literatura da identificação de genes blaTEM com
perfil ESBL (DROPA, 2013). Esse dado corrobora o presente estudo, já que o gene
blaTEM-1B isolado de amostra de carne de frango não possui perfil ESBL.
Os genes blaTEM são 99% similares uns aos outros e são carreados por 3
diferentes tipos de transposons descritos na literatura, Tn1, Tn2 e Tn3 (SMET et al.,
2009). O gene blaTEM-1B, localiza-se no transposon Tn2 e foi observado pela primeira
vez em 1997 por PEIXE et al., em uma cepa de Escherichia coli de origem clínica,
na Europa. Esse gene é derivado da enzima TEM-6 e não possui perfil ESBL.
Na literatura não há relatos desse gene em amostras de carne de frango,
entretanto, tem sido observado em amostras clínicas em alguns países como,
Espanha, Nigéria e China (TORO et al., 2011; LEEKITCHAROENPHON et al.,
2013; TORO et al., 2013; HUANG et al., 2014). Assim, este é o primeiro relato do
gene blaTEM-1b no Brasil.
O fato dos grupos TEM, SHV e CTX-M terem sido observados nas amostras
deste estudo, reforça a ideia da disseminação da resistência bacteriana também por
amostras de origem alimentar no Brasil. Em outros países os grupos CTX-M-8, TEM
e SHV são comumente observados em isolados de carne de frango (DHANJI et al.,
2010; STUART et al., 2012; REICH et al., 2013).

59
A adição de antimicrobianos em rações para a criação de aves tem
contribuído para a aquisição de genes codificadores de ESBL por patógenos de
origem animal. Recentes estudos no Brasil demonstram que a identificação de genes
que codificam resistência às ESBL, sobretudo CTX-M, em microrganismos isolados
da carne de frango, representam um problema emergente na população, pois podem
ser transferidos para o homem por meio da alimentação (FERNANDES et al., 2009;
SILVA e LINCOPAN, 2012).
5.1.2 Genes codificadores de AmpC β-Lactamases
Nesse estudo foi observada apenas a presença de um gene blaMOX em uma
amostra de frango de feira. Esse gene que ainda não foi relatado nesse tipo de
amostra na literatura, apenas em amostras ambientais (MOURA, 2010), apresenta
similaridade com os genes blaMOX-3 e blaMOX-4. Faz-se necessário mais estudos para
analisar a possibilidade de ser uma nova variante do grupo MOX.
Diversos estudos têm descrito a identificação de AmpC β-Lactamases em
Escherichia coli na cadeia de produção de animais. Segundo EWERS et al. (2012) e
LIEBANA et al. (2013), dentre os genes codificadores de AmpCs, o mais
frequentemente identificado é o blaCMY-2, entretanto genes pertencentes ao grupo
CMY não foram identificados no sequenciamento.
5.2 Genes que codificam resistência às Tetraciclinas
5.2.1 Genes codificadores de Bombas de Efluxo
Os fragmentos correspondentes a um ou mais genes codificadores de bombas
de efluxo (tetA, tetB, tetC, tetD e tetE) propostos para detecção foram obervados em

60
26 (86,7%) amostras de carne de frango (Anexo 1). Considerando que os iniciadores
para os diferentes genes tet utilizados no presente estudo são específicos e somente
detectam as respectivas variantes dos genes tet, é possível afirmar que as amostras
são positivas sem a necessidade do sequenciamento.
Dentre todos os genes codificadores de bombas de efluxo, estudos relatam
que o gene tetD é o mais raro de ser encontrado, mesmo em Escherichia coli isolados
de carne animal (microrganismo comum nesse tipo de amostra) (KOO e WOO,
2001). Dados obtidos no presente estudo não demonstram esse fato, já que os genes
com menor frequência observados nas amostras foram o tetE (9 amostras - 30%) e o
tetB (10 amostras - 33,3%), enquanto o tetD foi detectado c um número maior de
amostras (17 - 56,7%) amostras.
No Brasil, não há relatos na literatura da detecção dos genes tetB, tetC e tetD
em amostras de origem alimentar e tetA e tetE em amostras de carne de frango,
sendo este estudo o primeiro a apontar esses genes nesse tipo de amostra. Em outros
países, entretanto, diversos estudos identificaram esses genes em diversos
microrganismos isolados de frangos, sobretudo em Salmonella spp, Campylobacter
spp e Enterococcus spp (BRYAN et al., 2004; MILES et al., 2006; SMITH et
al.,2007; LYNNE et al., 2009.
5.2.2 Genes Codificadores de Proteínas de Proteção Ribossomal
Os genes tetM, tetO, tetQ, tetS e tetW foram obervados em uma ou mais
amostras estudadas (Anexo 1). O gene tetM foi o gene codificador de proteína de
proteção ribossomal mais frequente nas amostras, 13 (43,3%) (Figura 6). No Brasil

61
CAMPOS (2012) observou em seu estudo o gene tetM em 93% dos isolados de
Enterococcus de carcaça de frango. Em Porto Alegre, GAMA (2008) também
observou o gene tetM em 90% dos isolados de Enterococcus das amostras de frango
analisadas.
O gene tetO foi detectado em 4 (13,3%) amostras de carne de frango (Figura
6). Dados na literatura brasileira não apontam a detecção desse gene nesse tipo de
amostra no Brasil, sendo esse o primeiro relato. POUBEL et al. detectaram o gene
tetO em microrganismos isolados do leite de bovinos, no Brasil. Estudos realizados
em outros países demonstram a detecção dos genes tetM e tetO em carne de frango e
outros tipos de alimentos (CHOPRA e ROBERTS, 2001; NG et al., 2001;
GAROFALO et al., 2006).
Estudos demonstram que os genes tetM e tetO são comumente encontrados
em Lactobacillus. Esses microrganismos são utilizados, principalmente, na etapa de
fermentação de determinados tipos de alimentos, e na produção de probióticos
adicionados em rações para a criação de animais, além de estarem presentes no trato
gastrointestinal de homens e animais (KLEIN, 2011). A carne crua e os alimentos
fermentados são veículos potenciais para a transmissão de microrganismos
resistentes aos antibióticos na cadeia alimentar e, portanto, ao consumidor
(GAROFALO et al., 2006).
Organismos do gênero Enterococcus também são microrganismos
fermentadores presentes no trato gastrointestinal de mamíferos e participam da
fermentação de determinados tipos de alimentos, mas também podem estar presentes
no solo, água, plantas e vegetais. Em relação a resistência às tetraciclinas, os

62
organismos desse gênero normalmente apresentam os genes codificadores tetM, tetO
e tetS (HUYS et al., 2004).
Dados observados no presente estudo corroboram a literatura. Foi possível
observar o microrganismo que carreava o gene tetM, em uma amostra de
supermercado (congelado), enriquecendo a mesma com o antimicrobiano que
apresentava resistência (no caso, a Tetraciclina), e assim identificar a bactéria
Enterococcus, Por meio deste método, foi possível também comprovar que é viável a
identificação dos microrganismos carreadores dos genes identificados no estudo.
A expressão dos genes tetM e tetO parece ser regulada pela exposição à
tetraciclina. Diversos estudos demonstram que quando microrganismos carreadores
desses genes são expostos a esse antimicrobiano, a expressão de ambos aumenta. O
gene tetM é muitas vezes associado ao elemento conjugativo das famílias de
transposon Tn916 e Tn1545. Esse grupo de elementos formam regiões intermediárias
não replicativas que são essenciais para transposição intracelular e transferência
conjugativa intracelular favorecendo sua movimentação do cromossomo para o
plasmídeo, facilitando assim sua disseminação entre gerações de mesmos ou
diferentes microrganismos. O gene tetO não está associado a elementos conjugativos
e sim a plasmídeos conjugativos que também facilitam sua disseminação entre
microrganismos (CHOPRA e ROBERTS, 2001).
Os genes tetQ, tetS e tetW (Figura 6) foram menos prevalentes nesse estudo,
sendo identificados em três (10%), 7 (23,3%) e uma amostra (3,3%),
respectivamente, e ainda não foram identificados nesse tipo de amostra no Brasil,
sendo esse o primeiro relato no país, além de ser o primeiro relato dos genes tetQ e

63
tetW em amostras de carne de frango. Na literatura há relatos dos genes tetQ em
bactérias de origem intestinal de humanos (Bacteriodes spp e Provotella
intermedius) e de bovinos (Provotella ruminicula) (van den BOGAARD e
STOBBERINGH, 2000) e tetW em queijos (COMUNIAN et al., 2010), suínos e
bovinos (TEUBER, 2000). O gene tetS, como citado anteriormente, já foi observado
em cepas de Enterococcus isolados de amostra de alimentos e também já foi
obervado em carne de frango e em microrganismos isolados do queijo (WANG et al.,
2005).
5.3 Genes que codificam resistência às Quinolonas
No presente estudo, das 30 amostras analisadas, 28 (93,3%) apresentaram os
fragmentos correspondentes a um ou mais genes codificadores de resistência às
quinolonas (qnrA – 2 (6,6%) amostras, qnrB - 11 (36,6%) amostras - qnrS - 27
(90%) amostras) (Figura 7). No Brasil, esses genes ainda não foram identificados
nesse tipo de amostra, sendo considerado esse o primeiro relato dos mesmos no País.
Entretanto, já foram obervados em isolados de amostras clínicas (MINIRANI et al.,
2008; SOUZA; 2014).
HUANG et al. (2009) realizaram um estudo com Escherichia coli isoladas de
carne de frango, no período de 2001 à 2007, e identificaram os genes qnrA, qnrB e
qnrS em 4, 21 e 27 cepas das 532 analisadas. No presente estudo, o gene qnrS
também foi identificado com maior frequência (90%) dentre as amostras estudadas.
Estudos realizados em outros países identificaram esses genes em carne de
frango e outros tipos de amostras (YUE et al., 2008; VELDMAN et al., 2011).

64
Observa-se também relatos em estudos nos quais Salmonella spp, isoladas de frango,
eram resistentes às Quinolonas (THRELFALL, 2002; ANJUM et al., 2011;
VELDMAN et al., 2011; KIM et al., 2013).
Relatos na literatura apontam que os genes qnrA e qnrB estão geralmente
associados a plasmídeos que carreiam genes codificadores de ESBL (blaCTX, blaTEM,
blaSHV e blaVEB-1). No presente estudo, todas as amostras que foram identificados os
genes codificadores de ESBL observou-se genes codificadores de resistência às
Quinolonas, como mostra o Quadro 9.
Informações quanto à identificação de genes que codificam resistência às
Quinolonas em amostras de alimentos não são relatadas com frequência na literatura.
O presente estudo pesquisou apenas os 3 principais plasmidiais, os quais foram
observados com alta frequência.
Os dados obtidos no presente estudo abrem caminho para a confirmação da
disseminação de genes de resistência que, podem não ter sido observados nesses
tipos de amostras devido ao reduzido número de estudos moleculares realizados em
amostras de frango para identificação dos mesmos.
5.4 Contagem de Coliformes Termotolerantes
Todas as amostras analisadas no presente estudo foram consideradas próprias
para o consumo, de acordo com a Resolução RDC nº 12. Esse parâmetro permitiu
comprovar que mesmo uma amostra estando dentro dos padrões higiênicos sanitários
exigidos pela Lei, no que diz respeito de Coliformes Termotolerantes, pode haver
genes codificadores de resistência bacteriana na carga microbiana comum da carne

65
de frango. Os genes de resistência obervados nas mesmas não irão causar
enfermidade no momento da ingestão do alimento, porém, podem colaborar com a
disseminação da resistência bacteriana a longo prazo.
5.5 Globalização e a disseminação dos genes de resistência: um alerta
para a Saúde Pública.
Atualmente, um dos aspectos mais preocupantes da microbiologia clínica é a
resistência bacteriana aos antimicrobianos. A disseminação da resistência está cada
vez mais se caracterizando como um importante problema para a Saúde Pública. A
resistência bacteriana pode sofrer variações influenciadas pelo tipo de antibiótico
utilizado, os diferentes grupos de microrganismos envolvidos e mesmo pela
localização geográfica da área de estudo de modo que nenhum país do mundo está
livre desse problema (SHITO et al., 2002).
Nos países com o sistema de saúde precário, o impacto da resistência
bacteriana na economia é subestimado. Calcula-se que nesses locais pelo menos dois
terços da mortalidade infantil estão relacionados a infecções, como sepse neonatal,
tuberculose, meningite, diarréia e febre tifóide, que muitas vezes são mediadas por
bactérias resistentes aos antibióticos (SHITO et al., 2002).
Nos últimos 50 anos, o mundo tem sido modificado por grande movimento da
sociedade do campo para a cidade e estima-se que em 2030 esse número chegará aos
60%. Nosso planeta é percorrido por milhões de pessoas que viajam. Em 2000, o
número de pessoas deslocadas em todo o mundo chegou em 150 milhões. Além das
consequências políticas e demográficas, esse deslocamento global contribui com a

66
disseminação de doenças infecciosas e microrganismos resistentes aos
antimicrobianos (WALDVOGEL, 2004).
Como observado no presente estudo, o problema da resistência bacteriana não
está só na clínica ou no ambiente, mas também nos alimentos. O mercado alimentício
também tem seguido o processo de globalização e suas consequências na saúde da
população e política de um país. O cultivo em massa de produtos para a alimentação
e a mistura desses alimentos, unidos a importação e exportação, também tornam-se
um grave problema para Saúde Pública no que diz a respeito à disseminação da
resistência bacteriana (WALVOGEL, 2004).
O Brasil pode ser tomado como exemplo, pois atualmente é um dos maiores
exportador de carne de frango no mundo. Apesar de todo o controle do MAPA e
outros órgãos de fiscalização, estudos tem demonstrado o isolamento de
microrganismos resistentes aos antibióticos, no frango importado do nosso país
(WARREN et al., 2008; HAWKEY e JONES, 2009).
Assim, o comércio e as viagens internacionais são fatores que contribuem
amplamente para a disseminação da resistência bacteriana. Um microrganismo que é
primeiramente observado no Japão, por exemplo, seja de origem alimentar ou não,
pode chegar ao Brasil em menos de 30 horas, devido a facilidade da movimentação
de pessoas e mercadorias de um país para o outro.
O custo da resistência bacteriana gerado para a Saúde Pública é muito alto.
Em 2007, na Europa, houve cerca de 400.000 mil infecções mediadas por
microrganismos resistentes a antibióticos. Nos Estados Unidos, 20 bilhões de dólares

67
são gastos anualmente devido a disseminação da resistência bacteriana no país
(BUSH et al., 2011). No Brasil, os dados a respeito desse grave problema ainda são
precários. Em 2013, a ANVISA publicou uma nota técnica para Medidas de
Prevenção e Controle de Infecções por Enterobactérias multiresistentes, porém ainda
o país continua com dados insuficientes a respeito da resistência bacteriana. A página
virtual do Projeto EUREQA não possui notícias disponíveis e a última publicação
disponível é do ano de 2009.
Conhecer a diversidade de genes de resistência a antibióticos que circulam no
ambiente, sejam eles quais forem, pode gerar um alerta para a necessidade de ampliar
os cuidados, sejam eles no tratamento precoce de um paciente ou nas ações que
podem ser tomadas quanto à manipulação de alimentos, boas práticas na cadeia de
produção e orientação dos consumidores quanto a higienização das mãos e os
cuidados com a contaminação cruzada. Vivemos em uma economia global em que a
movimentação de alimentos e pessoas contaminados com bactérias resistentes pode
ter rápido e grande impacto em qualquer área do planeta. A globalização contribui
amplamente com o problema da disseminação da resistência bacteriana na Saúde
Pública Mundial.

68
6. Conclusões
Genes codificadores de resistência aos antimicrobianos β-Lactâmicos,
Tetraciclina e Quinolona estavam disseminados nas amostras carne de frango,
independente da forma de conservação (fresca ou congelada) ou da origem
das mesmas (feira, supermercado ou açougue).
Genes codificadores de resistência podem ser detectados em carne de frango,
mesmo que esta atenda aos Padrões Higiênicos Sanitários exigidos pela
ANVISA.
Genes, como blaCTX-M-1, blaSHV-85, blaTEM-1b, blaMOX-3/4, tetA, tetB, tetC, tetD,
tetE, tetQ, tetS, tetW, qnrA, qnrB e qnrS, que não haviam sido detectados
nesse tipo de amostra no Brasil e /ou no mundo foram observados nesse
estudo.

69
7. Considerações Finais
O objetivo do estudo não foi apontar se o alimento está próprio ou impróprio
para o consumo, mas sim informar sobre a problemática que vem ocorrendo em
diferentes tipos de amostra (inclusive em alimentos), para que assim possam ser
geradas algumas recomendações com o intuito de minimizar a disseminação da
resistência bacteriana.
Diante dos dados levantados para a realização do estudo, bem como os
resultados obtidos no mesmo, foi possível levantar a problemática da resistência
bacteriana em carne de frango. Assim, foram abordados alguns pontos básicos e
necessários para tentar minimizar esse problema, sobretudo no Brasil, o qual
demonstrou escassez de dados e estudos sobre o assunto.
Se levarmos em conta as amostras analisadas de acordo com os parâmetros
para Coliformes Termotolerantes, podemos dizer que durante a cadeia de produção, a
carne de frango foi corretamente manipulada e controlada, não gerando risco ao
consumidor em relação a algum tipo de contaminação. Entretanto, como observado,
existem genes de resistência circulando nesse alimento, que podem estar presentes
em bactérias comensais da carne de frango, e algumas recomendações podem ser
levadas em conta para evitar a disseminação da resistência bacteriana.
Alertar os colaboradores quanto à higienização das mãos e do uniforme e a
otimização da cadeia de produção, são pontos que poderiam reduzir a carga
microbiana e evitar a passagem de microrganismos, que muitas vezes são carreadores

70
de genes de resistência, do homem para o alimento. O controle da aplicação de
antimicrobianos como promotores de crescimento e agentes profiláticos também é
necessário, para evitar a seleção e disseminação da resistência bacteriana em
microrganismos comensais do frango.
A orientação de pessoas envolvidas na comercialização e no preparo do
alimento é de extrema importância para minimizar a contaminação dessas carcaças.
Além da orientação dos manipuladores em açougues, mercados e feiras, o
consumidor deve ter informações a respeito do modo de preparo do alimento, a fim
de que esse processo seja realizado com higiene e sem riscos de transmissão de um
microrganismo resistente, do alimento para o consumidor.
Por fim, a busca de medidas preventivas para a problemática da resistência
bacteriana depende também de maior incentivo do governo para o desenvolvimento
de estudos em amostras de origem alimentar. De acordo com o levantamento
bibliográfico no Brasil, estudos a respeito da disseminação da resistência bacteriana
são raros ou inexistentes, o que dificulta a tomada de medidas preventivas.

71
8. Referências Bibliográficas
1.Abraham EP, Chain E. An enzyme from bacteria able to destroy Penicillin. Nature.
1940; 837.
2.Abreu DLC, Franco RM, Nascimento ER, Pereira VLA, Alves FMX, Almeida JF.
Perfil de sensibilidade antimicrobiana e perfil do gene ISS pela reação em cadeia
pela polimerase na tipificação de Escherichia coli patogênica em codornas de corte
sob inspeção sanitária. Pesq. Vet. Bras. 2010; 30 (5): 406-410.
3.Accogli M, Fortini D, Giufré M, Graziane C, Dolejska M, Carattoli A, Cerquetti
M. Incl1 plasmids associated with the spread of CMY-2, CTX-M-1, an SHV-12 in
Escherichia coli of animal and human origin. Clinical Microbiology and Infection.
2013; 19 (5): 238-240.
4.Allen HK, Looft T, Bayles DO, Humphey S, Levine UY, Alt D, Stanton TB.
Antibiotic in feed induce prophages in swine fecal microbiomes. mBio. 2011; 2: 1-9.
5.Andersson DI, Hughes D. Persistence of antibiotic resistance in bacterial
populations. FEMS Microbiology Reviews. 2011; 35: 901-911.
6.Andrea MM, Arena F, Pallecchi L, Rossolini GM. CTX-M type β-Lactamases: A
successful story of antibiotic resistance. International Journal of Medical
Microbiology. 2013; 303: 305-317.
7.Anjum MF, Choudehary S, Morrison V, Snow LC, Mafura M, Slickers P, Ehricht
R, Woodward MJ. Identifying antimicrobial resistance genes of human clinical

72
relevance within Salmonella isolated from food animals in Great Britain. J.
Antimicrob. Chemother. 2011; 66: 550-559.
8.Antibiotic Resistance Threats in the United States. Centers for Disease Control and
Prevention [homepage na internet] 2013 [acesso em 12 abr 2014]. Disponível em:
http://www.cdc.gov/drugresistance/threat-report-2013/pdf/ar-threats-2013-508.pdf.
9.Arnold RS, Thom KA, Sharma S, Philips M, Jonhson JK, Morgan DJ. Emergence
of Klebsiella pneumoniae Carbapenemase (KPC) producing bacteria. South Med J.
2011: 104 (1): 40-45.
10.Babic M, Hujer AM, Bonomo RA. What´s new in antibiotic resistance? Focus on
Β-Lactamases. Drug Resistance Updates. 2006; 9: 142-156.
11.Balassiano IT, Bastos MCF, Madureira DJ, Silva IG, Freitas-Almeida ACF,
Oliveira SS. The involvement of tetA and tetE tetracycline resistance genes in
plasmid and chromosomal resistance of Aeromonas in Brazilian strains. Mem. Inst.
Oswaldo Cruz. 2007; 102(7): 861-866.
12.Balsalobre, LC. Pesquisa de genes codificadores de enterotoxinas e B-lactamases
em Aeromonas jandaei e Aeromas hydrophila provenientes de ambientes aquáticos.
[dissertação de mestrado] São Paulo: Faculdade de Saúde pública da USP; 2010.
13.Barros MR, Silveira WD, Araujo JM, Costa EP, Oliveira AAF, Santos ASFF,
Silva VAS, Mota RA. Resistência antimicrobiana e perfil plasmidial de Escherichia
coli isoladas de frango de corte e poedeiras comerciais no Estado de Pernambuco.
Pesq. Vet. Bras. 2012; 35 (5): 405-410.

73
14.Bauernfeind A, Chong Y, Lee K. Plamided-Enconded AmpC β-Lactamase: How
far have we gone 10 years after discovery? Yonsei Medical Journal. 1998; 39 (6):
520-525.
15.Bebrone C. Metallo β-Lactamases (classification, activity, genetic organization,
structure, zinc coordination) and their superfamily. Biochemcal Pharmacology. 2007;
74 (12): 1686-1701.
16.Bertoncheli CM, Hӧrner R. Uma revisão sobre metalo β-Lactamases. Revista
Brasileira de Ciências Farmacêuticas. 2008; 44 (4): 577-599.
17.Bogaard AE, Stobberingh EE. Epidemiology of resistance to antibiotics Links
between animals and humans. Inter. J. Antimicrob. Agents. 2000; 14: 327-335.
18.Bonnet R, Dutour C, Sampaio JLM, Chanal C, Sirot D, Labia R, Champs C De,
Sirot J. Novel cefotaximase (CTX-M-16) with encreased catalytic efficiecy due to
substitution Asp 240 - Gly. Antimicrob Agents and Chemother. 2001; 45 (8): 2269-
2275.
19.Bonnet R, Sampaio JLM, Labia R, Champs De, Sirot D, Chanal C, Sirot J. A
novel CTX-M β-Lactamase (CTX-M-8) in Cefotaxime-Resistance
Enterobacteriaceae isolated in Brazil. Antimicrob Agents and Chemother. 2000; 44
(7): 1936-1942.
20.Bonnet R. Growing group of Extended Expectrum β-Lactamases: The CTX-M
enzymes. Antimicrob. Agents Chemother. 2004; 48 (1): 1-14.
21.Bradford PA. What´s New in β-Lactamases? Curr Infect Dis Rep. 2001; 3: 13-19.

74
22.Bryan A, Shapir S, Sadowsky J. Frequency and distribution of Tetracycline
Resistance Genes in Genetically Diverse, Nonselected and Nonclinical Escherichia
coli Strains Isolated from Diverse Human and And Animal Sources. Appl. Environ.
Microbiol. 2004; 70 (4): 2503-2507.
23.Bush K, Couvarlin P, Dantas G, Davies J, Eisenstein B, Huovinen P, Jacoby JA,
Kishony R, Kreiswirth BN, Kutter E, Lerner SA, Levy S, Lewis K, Lomovckaya O,
Miller JH, Mobashery S, Piddock LJV, Projan S, Thomas CM, Tomasz A, Tulkens,
PM, Walsh TR, Watson JD, Witkowski J, Witte W, Wright G, Yeh P, Zsurskaya.
Tackling antibiotic resistance. Nature Reviews. 2011; 9: 895-896.
24.Bush K, Jacoby GA, Amicosante G, Bonomo RA, Bradford P, Cornaglia G,
Garau J, Giamerellou H, Jarlier V, Martinez-Martinez L, Miriagou V, Palzkill T,
Pascual A, Rodriguez-Bano J, Rossoline GM, Sougakoff W, Vatopoulos A.
Comment on: Redefining extended spectrum β-Lactamases: balance science and
clinical need. J Antimicrob Chemother. 2009; 64: 212-216.
25.Bush K, Jacoby, GA. Amino Acid Sequences for TEM, SHV and OXA
Extended-Spectrum and Inhibitor Resistant β-Lactamases. Lahey Clinic [homepage
na internet] 2003. [acesso em 10 jan 2013]. Disponível em: www.lahey.org.
26.Bush K, Jacoby, GA. Updated Functional Classification of β-Lactamases.
Antimicrob Agents Chemother. 2010; 54 (3): 969-976.
27.Busk K. Proliferation and significance of clinically relevant β-Lactamases.
Antimicrobial Therapeutics Reviews. 2013; 84-90.

75
28.Cai, JC, Zhou HW, Zhang R, Chen, G. Emergenc of Serratia marcescens,
Klebsiella pneumoniae, and Escherichia coli isolate possessing the plasmid-mediated
carbapenem-resistant-hydrolyzing β-Lactamase KPC-2 intensive care units of a
chinese hospital. Antimicrob Agents Chemother. 2008: 52 (6): 2004-2018.
29.Campos AFB. Resistência antimicrobiana de cepas de Enterococcus isoladas de
carcaças de frango comercializadas no Distrito Federal. [dissertação de mestrado]
Brasília: Faculdade de Agronomia e Medicina Veterinária; 2012.
30.Cantón R, Morosini, M. Emergence and Spread of antibiotic resistance following
exposure to antibiotics. FEMS Microbiol Rev. 2011; 35: 977-991.
31.Cantón, R, Coque, TM. The CTX-M β-Lactamase pandemic. Currt Opin
Microbiol. 2006; 9: 466-475.
32.Cao V, Lambert T, Nhu DQ, Loan HK, Hoang NK, Arlet G, Courvalin P.
Distribution of extended-spectrum β-Lactamases in clinical isolates of
Enterobacteriaceae in Vietnam. Antimicrob Agents Chemoter. 2002; 46: 3739-3743.
33.Carvalho-Assef ED, Pereira PS, Albano RM, Berião GC, Chagas TPG, Timm LN,
Silva RCF, Falci DR, Asensi MD. Isolation of NDM-producing Providencia rettgeri
in Brazil. J. Antimicrob. Chemother. 2013; 68 (12): 2956-2957.
34.Cattoir V, Poirel L, Rotimi V, Soussy CJ, Nordmann P. Multiplex PCR for
detection of plasmid-mediated quinolone resistance qnr genes in ESBL-producing
enterobacterial isolates. J Antimicrob Chemother. 2007;60(2):394-7.

76
35.Chapmann, PA, Ellin, M, Ashton, R, Shafique, W. Comparison of culture, PCR,
and immunoassays for detecting Escherichia coli O157 following enrichment culture
and immunomagnetic separation performed on naturaly contaminated raw meat
products. Int J Food Microbiol. 2001; 68: 11-20.
36.Chmelnitsky, I, Navon-Venezia S, Strahilevitz J, Carmeli Y. Plasmid-Mediated
qnrB2 and Carbapenemase gene blaKPC-2 on the same plasmid in carbapenem-
resistent ciprofloxacin-suscetible Enterobacter cloacae isolates. Antimicrob Agents
Chemother. 2008; 52 (8): 2962-2965.
37.Chopra I, Roberts M. Tetracycline Antibiotics: mode of acton, applications,
molecular biology, and epidemiology of bacterial resistance. Microbiol Mol Biol
Rev. 2001; 65 (2): 232-260.
38.Cogliani C, Goossens H, Greko C. Restricting Antimicrobial Use in Food
Animals: Lessons from Europe. Microbe. 2011; 6 (6): 274-279.
39.Comunian R, Daga E, Dupré I, Paba A, Deviergiliis C, Piccione V, Perozzi G,
Zonenschain D, Rebbechi A, Morelli L, Lorentiis A, Giraffa G. Susceptibility to
tetracycline and erythromycin of Lactobacillus paracasei strains isolated from
traditional Italian fermented foods. Internarional Journal of Food Microbiology.
2010; 138: 151-156.
40.Cosgrove, SE, Canneli, Y. The Impact of Antimicrobial Resistance on Health and
Economic Outcomes. Clin Infect Dis; 36: 1433-1437.

77
41.Davies J, Davies, D. Origins and Evolution of Antibiotic Resistance. Microbial
Mol Rev. 2010; 74 (3): 417-433.
42.Dhanarani, TS, Shankar, C, Park, KJ, Dexilin, M, Kuman, RR, Thamaraiselvi, K.
Study on acquisition of bacterial antibiotic resistance determinants in poultry letter.
Int J Poult Sci. 2009; 88: 1381-1387.
43.Dhanji, H, Murphy, NM, Doumith, M, Purmus, S, Lee, SS, Hope, R, Woodford,
N, Livermore, DM. Cephalosporin resistance mechanisms in Escherichia coli
isolated from raw chicken imported into the UK. J Antimicrob Chemothe. 2010; 65:
2534-2537.
44.Dierikx C, Essen-Zandbergen A, Veldman K, Smith H, Mevius D. Increased
detection of extended spectrum β-Lactamase producing Salmonella enterica and
Escherichia coli isolates from poultry. Veterinary Microbiology. 2010; 145: 273-
278.
45.Dropa, M, Balsalobre, LC, Lincopan, N, Mamizuka, EM, Cassetari, VC, Franco,
F, Guido, SM, balabakes, AJ, Passadore, LF, Santos, SR, Matté, GR, Matté, MH.
Extended Spectrum Beta Lactamases Among Enterobacteriaceae isolated in a Public
Hospital in Brazil. Rev Inst Med Trop S. Paulo. 2009; 51 (4): 203-209.
46.Dropa, M. Caracterização genotípica de cepas da família Enterobacteriaceae
produtoras de β-Lactamases de espectro estendido, isoladas de pacientes de um
hospital da rede pública da cidade de São Paulo. [dissertação de mestrado] São
Paulo: Faculdade de Saúde Pública da USP; 2006.

78
47.Dropa, M. Disseminação da resistência a antimicrobianos em cepas clínicas e
ambientais de Enterobacteriaceae: identificação e mapeamento do ambiente genético
de genes codificadores de ESBL. [tese de doutorado] São Paulo. Faculdade de Saúde
Pública da USP; 2013.
48.Dzidic, S, Bedekovic, V. Horizontal Gene Transfer emerging multidrug resistance
in hospital bacteria. Acta. Pharmacol. Sin. 2003; 24 (6): 519-526.
49.Eller C, Leistner R, Guerra B, Fischer J, Wendt C, Rabsch W, Wener G, Pfeifer
Y. Emergence of Extended-spectrum β-Lactamase (ESBL) CTX-M-8 in Germany. J.
Antimicrob. Chemother. 2013; 1-3.
50.Endimiani, A, Carias, LL, Hujer AM, Bethel CR, Hujer KM, Perez F, Hutton RA,
Fox, WR, Hall GS, Jacobs MR, Paterson DL, Rice LB. Jekins, SG, Tenover, FC,
Bonomo RA. Presence of Plasmid-Mediated quinolone resistance in Klebsiella
pneumoniae isolates possessing blaKPC in the United States. Antimicrob Agents
Chemother. 2008; 52 (7): 2680-2682.
51. EUREQA (Epidemiologia do Uso e da Resistência bacteriana a Quimioterápicos
e Antibióticos na População) [homepage na internet] 2013 [acesso em 24 abr 2014].
Disponível em: www.dpi.inpe.br/eureqa/index.php.
52.Falagas ME, Karageorpopoulos, DE. Extended-Spectrum β-Lactamases-
producing organims. J Hosp Infect. 2009; 73: 345-354.
53.Fernandes SA, Paterson DL, Ghilardi-Rodrigues AC, Adams-Haduch JM,
Tavechio AT, Doi Y. CTX-M-2 Producing Salmonella Typhymurium isolated from

79
pediatric patients and poultry in Brazil. Microbial Drug Resistance. 2009: 15 (4):
317-321.
54.Ferreira JC, Filho RACP, Andrade LN, Junior AB, Darini ALC. Detection of
chromossomal blaCTX-M-2 in diverse Escherichia coli isolates from healthy broiler
chickens. Clin Microbiol Infect. 2014; 2-4.
55.French, GL. Clinical impact and relevance of antibiotic resistance. Adv Drug
Deliv Rev. 2005; 57: 1514-1527.
56.Gama BA. Análise da resistência antimicrobiana e de genes de virulência de
Enterococcus spp. [dissertação mestrado] Rio Grande do Sul: Universidade Federal
do Rio Grande do Sul; 2008.
57.Garcia DO, Doi Y, Szabo D, Adams Haduch JM, Vaz TMI, Leite D, Padoveze
MC, Freire MP. Silveira FP, Paterson DL. Multiclonal Outbreak of Klebsiella
Pneumoniae Producing Extended-Spectrum β-Lactamase CTX-M-2 and novel
variant CTX-M-59 in a neonatal intensive care unit in Brazil. Antimicrob. Agents
Chemother. 2008; 52(5): 1790-1793.
58.Garofalo, C, Vignaroli, C, Zandri, G, Aquilant, L, Bordoni, D, Osimani, A,
Clementi, F, Biavasco, F. Direct Detection of Antibiotic Resistance genes in
speciments of chicken and pork meat. Int J Food Microbiol. 2007; 113: 75-83.
59.Gillespie, SH. Antibiotic Resistance in the absence of selective pressure. Int J.
Antimicrob Agents. 2001; 17: 171-176.

80
60.Hanson, ND, Sanders, CC. Regulation of Inducible AmpC Β-Lactamase
Expression Among Enterobacteriaceae. Curr Phar Des. 1999; 5: 881-894.
61.Hawkey, PM. Mechanisms of quinolone action and microbial response. J
Antimicrob Chemother. 2003; 51: 29-35.
62.Henriques, IS, Fonseca, F, Alves, A, Saavedra, MJ, Correia, A. Ocurrence and
Diversity of integrons and β-Lactamase genes among ampicilin-resistant isolates
from estuarine waters. Res Microbiol. 2006; 57: 938-947.
63.Holmberg, SD, Solomon, SL, Blak, DA. Health and Economic Impacts of
Antimicrobial Resistance. Rev Infect Dis. 1987; 9(6): 1065-1078.
64.Huang Y, Xu Y, Wang Z, Lin X. Antimicrobial resistance genotype analysis of
Extended-Spectrum-β-Lactamase-Producing Proteus Mirabilis. Open Journal of
Clinical Diagnostics. 2014; 4: 52-57.
65.Huang, S, Dai, L, Xia, L, Du, X, Qi, Y, iu, H, Wu, CSJ. Increased Prevalence of
Plasmid-Mediated Quinolone Resistance Determinants in chicken Escherichia coli
isolates from 2001 to 2007. Foodborn Pathog Dis. 2009: 6(10): 1203-1209.
66.Huys G, D´Haene K, Collard J, Swuings J. Prevalence and Molecular
characterization of tetracycline resitance in Enterococcus isolates from food. Appl.
Environ. Microbiol. 2004; 70 (3): 1555-1562.
67.Jacoby, GA. AmpC β-Lactamases. Clin Microbiol Rev. 2009; 22 (1): 161-182.

81
68.Jacoby, GA. Mechanisms of Resistance to Quinolones. Clin Infec Dis. 2005; 41:
S120-S126.
69.Jaurin, B, Grundstrӧm, T, Edeund, T, Normark, S. The Escherichia coli β-
Lactamase attenuator mediates growth rate-depend regulation. Nature. 1981; 290:
221-225.
70.Jonhson JR, Sannes MR, Croy C, Johnston B, Clabots C, Kuskowski MA, Bender
J, Smith KE, Winokur PL, Belongia EA. Antimicrobial Drug Resistance Escherichia
coli from humans and poultry products, Minnesota and Wisconsin, 2002-2004.
Emerging Infectious Disease. 2007; 13 (6): 839-846.
71.Kim, K, Park, J, Kwak, H, Woo, G. Characterization of the quinolone resistance
mechanism in foodborne Salmonella isolates with high nalidix acid resistance. ntJ
Food Microbiol. 2011; 146: 52-56.
72.Klein G. Antibiotic resistance and molecular characterization of probiotic and
clinical Lactobacillus strains in relarion to safety aspects of probiotics. Foodborne
Pathogens and Disease. 2011; 8 (2): 269-281.
73.Kliebe, C, Nies, BA, Meyer, JF, Tolxdorff-Neutzling, RM, Wiedmann, B.
Evolution of plasmid-coded resistance to broad-spectrum cephalosporins.
Antimicrob Agents Chemother. 1985; 28 (2): 302-307.
74.Koo H, Woo G. Distribution and transferability of tetracycline resistance
determinants in Escherichia coli isolated from meat and meat products. 2011; 145:
407-413.

82
75.Kuchenbecker, BS, Ribeiro, AR, Cardoso, M. Perfil de resistência de isolados de
Staphylococcus aureus obtidos de produtos de origem animal analisados pelo serviço
de inspeção federal do Brasil. Acta Veterinariae. 2009; 37 (2): 143-149.
76.Leekicharoenphon P, Friis C, Zankari E, Svendsen CA, Price LB, Rahmani M,
Herrero-Fresno A, Fashae K, Vandenberg O, Aarestrup FM, Hedrikasen RS.
Genomics of an emerging clone of Salmonella serovar Typhimurium ST313 from
Nigeria and The Democratic Republic of Congo. J. Infect. Dev. Ctries. 2013; (10):
696-706.
77.Liebana E, Guns D, Garcia-Migura L, Woodward MJ, Clifton-Hadley FA, Davies
RH. Molecular typing of Salmonella sorotypes prevalent in animals in England. L.
Clin. Microbial. 2001; 39 (10): 3609-3616.
78.Livermore DM. Fourteen years in resistance. Int J Antimicrob Agents. 2012; 39
(4): 283-294.
79. Lynne AM, Kaldhone P, David D, White DG, Foley SL. Characterization of
antimicrobial resistance in Salmonella enterica serotype Heidelberg isolated from
food animals. Foodborne Pathogens and disease. 2009; 6 (2): 207-215.
80.Maia, AA, Cantisani, ML, Esposto, EM, Silva, WCP, RodrLigues, ECP,
Rodrigues, DP, Lázaro, NS. Resistência antimicrobiana de Pseudomonas aeruginosa
isolados de pescado e de cortes de miúdo de frango. Ciência e Tecnologia de
Alimentos. 2009; 29 (1): 114-119.

83
81.Marshall, BM, Levy, SB. Food Animals and Antimicrobials: Impacts on Human
Health. Clin Microbiology Reviews. 2011; 24 (4): 493-499.
82.McGowan, JEJ. Economic Impact Of Antimicrobial Resistance. Emerg Infect
Dis. 2001; 7 (2): 286-292.
83.Medeiros AA, Cohenford, M, Jacoby, GA. Five novel plasmid-determined β-
Lactamases. Antimicrob Agents Chemother. 1985; 27 (5): 715-719.
84.Miles TD, Mclaughlin W, Brown PD. Antimicrobial resistance of Escherichia
coli isolates from boiler chickens and humans. BMC Veterinary and Research. 2006;
2 (7): 1-9.
85.Minarini LAR, Poirel L, Trevisani NAC, Darini ALC, Nordmann P.
Predominance of CTX-M type extended-spectrum β-lactmase genes among
enterobacterial isolates from outpatients in Brazil. Diagnostic Microbiology and
Infectious Disease. 2009; 65: 202-206.
86.Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa n. 9,
de 27 de junho de 2003. Aditivos Proibidos na Agricultura. Diário Oficial da União.
30 de junho de 2003; Seção 1:4.
87.Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa n. 26,
de 9 de julho de 2009. Aprova o Regulamento Técnico para Fabricação, o controle
de qualidade, a comercialização e o emprego de produtos antimicrobianos de uso
veterinário. Diário Oficial da União. 10 de julho de 2009; Seção 1.

84
88.Modi CM, Mody SK, Patel HB, Dudhatra GB, Kumar A, Sheikn TJ. Growth
promoting use of antimicrobial agents in animals. J Apllie Pharma Sci. 2011; 1 (8):
33-36
89.Morar, M, Whidht, GD. The Genomic Enzymology of Antibiotic Resistance.
Annu Rev Genet. 2010; 44: 25-51.
90.Moreno-Bondi MC. Antibiotics in food and environmental samples. Anal Bioanal
Chem. 2009: 395 (4): 875-876.
91.Moura, EMM. Análise do perfil de resistência a antibióticos e detecção dos genes
de virulência e resistência em Aeromonas provenientes de amostras ambientais.
[dissertação de mestrado] 2010. São Paulo: Faculdade de Saúde Pública da
Universidade de São Paulo.
92.Naas T, Cuzon G, Villegas M, Lartigue M, Quinn JP, Nordmann P. Genetc
structures at the origin of acquisition of the β-Lactamase blaKPC gene. Antimicrob
Agents Chemother. 2008; 52 (4): 1257-1263.
93.Ng LK, Martin I, Alfa M, Mulvey M. Multiplex PCR for the detection of
tetracycline resistant genes. Molecular and Cellular Probes. 2001; 15: 209-215.
94.Nordmann, P, Cuzon, G, Naas, T. The real threat of Klebsiella pneumoniae
Carbapenemase-producing bacteria. Lacet Infec Dis. 2009; 9:228-236.
95.Nordmann, P, Poirel, L, Walsh, TR, Livermore, DM. The Emerging NDM
Carbapenemase. Trends Microbiol. 2011; 19 (12): 58

85
96.Nordmann, P, Poirel, L. Emergence of Plasmid-mediated resistance to quinolones
in Enterobacteriaceae. J Antimicrob Chemother. 2005; 56: 463-469.
97.Nordstrom L, Liu CM, Price LB. Foodborne urinary tract infectious: a new
paradigm antimicrobial-resistant foodborn illness. Frontiers in Microbiology. 2013;
29 (4): 1-6.
98.Olesen, I, Hasman, H, Aaustrup, FM. Prevalence of β-Lactamases among
Ampicillin Resistant Escherichia coli and salmonella Isolated from food Animals in
Denmark. Microb Drug Resist. 2004; 10 (4): 334-340.
99.Oliphant, CM. Quinolones: A Comprensive Review. Am Fam Physician. 2002; 65
(3): 455-464.
100.Oliveira, DE. Caracterização, pesquisa dos genes de virulência e β-Lactamases
em Aeromonas hydrophila provenientes de lodo e esgoto tratados. [dissertação de
mestrado]. São Paulo: Faculdade de Saúde Pública da USP. 2011.
101.Olverdevest I, Willemsen I, Rijnsburger M, Eustace A, Xu L, Hawkey P, Heck
M, Savelkoul P, Vanderbroucke-Grauls C, Zwaluw K, Huijsdens X, Kluytmans J.
Extended-Spectrum β-Lactamases genes of Escherichia coli in chicken meat and
humans, the Netherlands. Emerging Infectious Disease. 2011; 17 (7): 1216-1222.
102.Paterson DL, Bonomo, RA. Extended Spectrum β-Lactamases: a Clinical
Updated. Clin Microbiol Rev. 2005; 18 (4): 657-686.

86
103.Peirano G, Agerso Y, Aeretrup M, Reis EMF, Rodrigues BP. Ocurrence of
integrons and antimicrobial resistance genes among Salmonella enteric from Brazil.
J. Antimicrob Chemother. 2006; 58: 305-309.
104.Peirano G, Pitout GDD. Molecular Epidemiology of Escherichia coli producing
CTX-M β-Lactamases. The worldwide emergence of clone ST131 025:H4.
International Journal of Antimicrobial Agents. 2010: 35 (4): 316-321.
105.Peixe LV, Sousa JCF, Perez-Dias JC, Baquero F. A bla TEM-1b derived TEM-6
β-Lactamase: a case of convergente evolution. Antimicrob. Agents Chemother. 1997;
41 (5): 1206.
106.Perez, F, Endimiani, A, Hujes, KM, Bonomo, RA. The continuing challenge of
ESBLs. Curr Opin Pharmacol. 2007; 7: 459-469.
107.Philippon, A, Arlet, G, Jacoby, GA. Plasmid-Determined AmpC-Type β-
Lactamases. Antimicrob Agents Chemother. 2002; 46 (1): 1-11.
108.Pitout, JDD, Gregson, DB, Poirel, L, McClure, JLP, Church, DL. Detection of
Pseudomonas aeruginosa Producing Metallo-β-Lactamases in a Large Centralized
Laboratory. J Clin Microbiol. 2005; 43 (7): 3129-3125.
109.Ploy, M, Lambert, T, Couty, J, Denis, F. Integrons: an antibiotic resistance gene
capture and expression system. Clin Chem Lab Med. 2000; 38 (6): 483-487.
110.Poirel L, Rodriguéz-Martinéz JM, Mammeri H, Liard A, Nordmann P. Origin of
Plasmid-Mediated Quinolone Resistance Determinant QnrA. Antimicrob. Agents
Chemother. 2005; 49 (8): 3523-3525.

87
111.Poirel, L, Cattoir, C, Nordmann, P. Is plasmid-mediaated quinolone resistance a
clinically significant problem? Clin Microbiol Infect. 2008; 14: 295-297.
112.Queenan AM, Bush K. Carbapenemases: the Versatile β-Lactamases. Clin
Microbiol Rev. 2007; 20 (3): 440-458.
113.Ramos LSM. Aditivos alternativos a antibióticos em rações para frangos de
corte. [tese de doutorado] Piauí. Universidade Federal do Piauí; 2009.
114.Reich F, Atanassova V, Klein G. Extended Spectrum β-Lactamase and AmpC
Producing Enterobacteria in Healthy broiler chickens, Germany. Emerging Infectious
Disease. 2013; 19 (8): 1254-1259.
115.Ribeiro AR, Kallermann, A, Santos LR, Nascimento VP. Resistência
antimicrobiana em Salmonella Enteritidis isoladas de amostras clínicas e ambientais
de frangos de corte e matrizes pesadas. Arq Bras Med Vet Zootec. 2008; 60 (5):
1259-1262.
116.Rice, LB. The clinical consequences of antimicrobial resistance. Curr Opin
Microbiol. 2009; 12: 476-481.
117.Roberts MC. Tetracycline Therapy: Update. Clin Infec Dis. 2003; 36: 462-467.
118.Roberts MC. Update on acquired tetracycline resistance genes. FEMS Microbiol
Letters. 2005: 245: 195-203.

88
119.Roberts, MC. Tet Mechanisms of resistance. Washington Faculty [homepage na
internet] 2001. [acesso em 9 jun 2012]. Disponível em:
http://faculty.washington.edu/marilynr/tetweb4.pdf.
120.Rolain J. Food and Human gut as reservoirs of transferable antibiotic resistance
enconding genes. Frontiers in Microbiology. 2013; 173 (4): 1-10.
121.Rossolini, GM, D´Andrea, MM, Mugnaioli, C. The Spread of CTX-M type
extended-spectrum β-Lactamases. Clin Microbiol. Infect. 2008; 14: 33-41.
122.Sabath, LD, Abraham, EP. Zinc as a cofactor Cephalosporinase from Bacillus
cereus 569. Biochem J. 1966; 98:11-13.
123.Samaha-Kfoury, JN, Araj GF. Recent Developments in β-Lactamases and
extended spectrum β-Lactamases. BMJ. 2003; 327 1209-1213.
124.Santos MM. Resistência Antimicroniana em cepas bacterianas isoladas de
celulite aviária. [dissertação de mestrado] Brasília: Faculdade de Agronomia e
Medicina Veterinária; 2012.
125.Schwartz, T, Kohnen, W, Jasen, B, Obst, U. Detection of Antibiotic-resistant
bacteria and their resistance genes in wastwater, surface water, and drinking water
biofilms. FEMS Microbiol Ecol. 2003; 43: 325-335.
126.Seral, C, Gude, MJ, Castillo, FJ. Emergencia de β-Lactamases AmpC
plasmídicas (p AmpC ó cefamicinasas): origen, importância, detección y alternativas
terapêuticas. Rev Esp Quimioter. 2012; 25 (2): 89-99.

89
127.Shah, AA, Hasan, I, Ahmed, A. Extended-Spectrum β-Lactamases (ESBL):
Characterization, Epidemiology and Detection. Crit Rev Microbiol. 2004; 30 (1): 25-
32.
128.Shito GC. Is antimicrobial resistance also subject to globalization. European
Society of Clinical Microbiology and Infectious Diseases. 2002; 8 (3): 1-8.
129.Shlaes, DM, Gerding, DN, John, JFJ, Croug, WA, Bornstein, DL, Duncan, RA,
Eckman, MR, Farrer, WE, Greene, WH, Lorian, V, Levy, S, McGowan, JE, Paul,
SM, Ruskin, J, Tenover, FC, Watanakunakorn, C. Society for Healthcare
Epidemiology of America Joint Committee on the Prevention of Antimicrobial w
130.Silva KC, Lincopan N. Epidemiologia das betalactamases de espectro estendido
no Brasil: impacto clínico e implicações para o agronegócio. J. Bras. Patol. Med.
Lab. 2012; 48 (2): 91-99.
131.Silva, MAP, Rosado, PL, raga MJ, Campos, AC. Determinantes da oferta de
Exportação de carne de frango do Brasil, de 1992 a 2007. RESP. 2011; 49 (1): 31-54.
132.Singer ME, Harding, I, Michael, JR, Jaffe, DH. Impact of antimicrobial
resistance on health outcomes in the out-pacient treatment of adult community
acquired pneumonia: a probably model. J Antimicrobal Chemother. 2003; 51: 1269-
1282.
133.Smet A, Martel A, Persoons D, Dewulf J, Heyndrickx M, Herman L,
Haesebrouck F, Butaye P. Broad-Spectrum β-lactamases among Entereobacteriaceae

90
of animal origin: molecular aspects, mobility and impact on public health. FEMS
Microbiology Reiviews. 2010; 34: 295-316.
134.Smith JL, Drum DJV, Daí Y, Kim JM, Sanchez S, Maurer JJ, Hofacre CL, Lee
MD. Impact of antimicrobial usage on antimicrobial resistance in commensal
Escherichia coli strains colonizing broiler chickens. Applied and and Environmental
Microbiology. 2007; 73 (5): 1404-1414.
135.Söresen, TL, Blom, M, Monnet, L, Frimodt-Möller, N, Poulsen, KL, Espersen,
F. Transient intestinal carriage after ingestion of antibiotic resistance Enterococcus
faecium from Chicken and Pork. N Engl Med 2001; 345 (16): 1161-1166.
136.Souza, RRF. Pesquisa de genes de resistência a quinolonas em bacilos gram
negativos de origem clínica e ambiental. [dissertação de mestrado] São Paulo:
Faculdade de Saúde Pública da USP; 2014.
137.Stuart BL, Marshall, B. Antibacterial resistance worldwid: causes, challenges
and responses. Nat Med. 2004; 10 (12): 122-129
138.Stϋrenburg, E, Mack, D. Extended Spectrum β-Lactamases: implications for the
clinical microbiology laboratory, therapy, and infection control. J Infect. 2003; 47:
273-295.
139.Tavares, W. Manual de Antibióticos e Quimioterápicos. São Paulo: Atheneu;
2001.

91
140.Tawfic AF, Alswilem AM, Ahibl AM, Al-Agamy MHM. Prevalence and genetic
characteristics of TEM, SHV and CTX-M in clinical Klebsiella pneumoniae isolates
from Saudi Arabia. Microbial Drug Resistance. 2011: 17 (3): 384-388.
141.Teuber M. Veterinary use and antibiotic resistance. Curr Opin Microbiol. 2001;
4: 493-499.
142.Thaker M, Spanogiannopoulos P, Wright GD. The tetracycline resistome. Cell
Mol Life Sci. 2010; 67: 419-431.
143.Toro M, Garcia P, Rodríguez I, Rojo Bezares B, Helmuth R, Ridicio MR,
Guerra B, Torres C. Characterization of plasmids implicated in the mobilization of
extended-spectrum and AmpC β-lactamases genes in clinical Salmonella enterica
isolates and temporal stability of the resistance genotype. International Journal of
Antimicrobial Agents. 2013.
144.Toro M, Saénz Y, Cercenado E, Rojo-Bezares B, García-Capello M, Undabeitia
E, Torres C. Genetic characterization of the mechanisms of resistance to
amoxicillin/clavulanate and third-generation cephalosporin in Salmonella enterica
from three Spanish hospitals. International Microbiology. 2011; 14: 173-181.
145.Turner, PJ. Extended-Spectrum β-Lactamases. Clin Infect Dis. 2005; 41: 273-
275.
146.União Brasileira de Avicultura – UBABEF Relatório Anual 2013. Disponível
em: <http://www.abef.com.br/ubabef/publicacoes_relatoriosanuais.php>. Acesso em:
09 ago. 2013.

92
147.Veldman K, Cavaco LM, Mevius D, Battisti A, Franco A, Botteldoorn N,
Bruneau M, Perrin-Guyomard A, Cerny T, Escobar CF, Guerra B, Schroeter A,
Gutierrez M, Hopkins K, Myllyniemi A, Sunde M, Wasyl D, Aarestrup FM.
Internarional collaborative study on the occurrence of plasmid-mediated quinolone
resistance in Salmonella enterica and Escherichia coli isolated from animals,
humans, food and the invironment in 13 European countries. J. Antimcrob.
Chemother. 2011; 66: 1278-1286.
148.Voets GM, Fluit AC, Scharringa J, Stuart JC, Hall MAL. A set of multiplex
PCRs for genotypic detection of extended-spectrum β-Lactamases, carbapenemases,
plasmid-mediated AmpC β-Lactamases and OXA β-Lactamases. Inter J Antimicrob
Agents. 2011; 37: 356-359.
149. Waldvogel FA. Infectious diseases in the 21st century: old challenges and new
opportunities. International Journals of Infectious Diseases. 2004; 5-12.
150.Wallinga D, Burch DGS. Does adding routine antibiotics to animal feed pose a
serious risk to human health. BMJ. 2013; 1-3.
151.Walsh F. Investigating antibiotic resistance in non-clinical environments.
Fronstiers in Microbiology. 2013; 19 (4): 1-5.
152.Walsh, RT. Clinically significant carbapenemases: na update. Curr Opin Infect
Dis. 2008; 21 (4): 367-371.
153.Walsh, TR, Toleman, MA, Poirel, L, Nordmann, P. Metallo β-Lactamases: the
Quiet before the Storm? Clin Microbiol Rev. 2005; 18 (2): 306-325.

93
154.Wang, HH, Manuzon, M, Lehman, M, Wan, K, Liv, H, Wittum, TE, Yousef, A,
Bakaletz, LO. Food commensal microbes as a potencially important avenue in
trnsmiting antibiotic resistance genes. FEMS Microbiol Lett 2006; 254: 226-231.
155.Warren RE, Ensor VM, Neill PO, Butler V, Taylor J, Nye K, Harvey M,
Livermore DM, Woodford N, Hawkey PM. Imported chicken meat as a potencial
source of quinolone-resistant Escherichia coli producing extended-spectrum β-
Lactamases in the UK. J. Antimicrob Chemother. 2004; 61: 504-508.
156.Wei R, Ge F, Huang S, Chen M, Wang R. Ocurrence of veterinary antibiotics in
animal wastewater and surface water around farms in Jiangsu, China. Chemosphere.
2011: 82 (10): 1408-1414.
157.Weldhagen, GF. Integrons and β-Lactamases - a novel perspective on resistome.
Int J Antimicrob Agents. 2004; 23: 556-562.
158.White, PA, Mclver, CJ, Rawlinson, WD. Integrons and gene cassets in the
Enterobacteriaceae. Antimicrob Agents Chemother. 2001; 45 (9): 2658-2661.
159.Winck CA, Machado JAD. Avicultura brasileira: perspectivas para o mercado
consumidor Chinês. Revista de Administração, Contabilidade e Economia. 2011; 10
(2): 241-268.
160.Wittum TE, Mollenkopf DF, Daniels JB, Parkinson AE, Mathews JL, Fry PR,
Abley MJ, Gebreyes WA. CTX-M-Type Extended-Spectrum β-Lactamases present
in Escherichia coli from the faces of Cattle in Ohio, United States. Foodborn
Pathogens and Disease. 2010; 7(12): 1575-1579.

94
161.Wu, H, Liu BG, Liu JH, Pan YS, Yuan L, Hu GZ. Phenotypic and molecular
characterization of CTX-M-14 extended-spectrum β-Lactamase and plasmid-
mediated ACT-like AmpC β-Lactamase produced by Klebsiella pneumoniae isolates
from chickens in Henan Province, China. Genetics and Molecular Researchs. 2012;
11 (3): 3357-3364.
162.Yamazoe, K, Kato, N, Kato, H, Tanaka, K, Katagiri, Y, Watanabe, K.
Distribution of cfiA gene among Bacterioides fragilis strains in Japan and
Relatedness of cfiA to imipenem Resistance. Antimicrob Agents Chemother. 1999;
43 (11): 2808-2810.
163.Yang B, Wang Q, Cui S, Wang W, Shi C, Xia X, Xi M, Wang X, Shi X, Wang
D, Zhang Z, Meng J. Characterization of extended-spectrum beta-lactamases-
producing Salmonella strains isolated from retail foods in Shaanxi and Henan
Province. China. Food Microbiology. 2014; 42: 14-18.
164.Yigit, H, Queenan, AN, Anderson, GJ, Domenech-Sanchez, Am Biddle, JW,
Steward, CD, Albert, S, Bush, K, Tenover, SC. Novel Carbapenem-Hydrolyzing β-
Lactamase, KPC-1,from a Carbapenem Resistant Strain of Klebsiella pneumoniae.
Antimicrob Agents Chemoter. 2001; 45(4): 1151-1161.
165.Yong, D, Toleman, NA, Giske, CG, Cho, HS, Sudman, K, Lee, K, Walsh, TR.
Characterization of a New Metallo β-Lactamase gene, blaNDM-1, and a novel
Erytromicyn Esterase gene carried on a Unique Genetic Structure in Klebsiella
pneumoniae Sequence Type 14 from India. Antimicrob Agents Chemother. 2009; 53.

95
166.Yue L, Jiang H, Liao X, Liu J, Li S, Chen X, Chen C, Lu D, Liu Y. Prevalence
of plasmid-mediated quinolone resistance qnr genes in poultry and swine clinical
isolates of Escherichia coli. Veterinary Microbiology. 2008; 132: 414-420.
167.Zhao L, Dong YH, Wang H. Residues of Veterinary antibiotics in masures from
feedlot livestock in eight provinces of China. Sci Total Environ. 2010; 408 (5): 1069-
1075.

96
ANEXOS
ANEXO 1 - Número e porcentagem dos diferentes tipos de amostras de carne de
frango (supermercado, açougue e feira) que continham os fragmentos de tamanhos
esperados para os diferentes genes codificadores de resistência, segundo a classe de
antimicrobianos.

97
ANEXO 1 - Número e porcentagem dos diferentes tipos de amostras de carne de frango (supermercado, açougue e feira) que continham os
fragmentos de tamanhos esperados para os diferentes genes codificadores de resistência, segundo a classe de antimicrobianos.
AMOSTRAS
SUPERMERCADO (congelado) AÇOUGUE (resfriado) FEIRA (resfriado) Total
1C 2C 3C 4C 5C 6C 7C 8C 9C 10C 1A 2A 3A 4A 5A 6A 7A 8A 9A 10A 1F 2F 3F 4F 5F 6F 7F 8F 9F 10F Nº (%)
Β-L
AC
TÂ
MIC
OS
E
SB
L
blaCTX-M-1 - - - - + - - - + - - - - - - + - - - + - - - - - - - - - - 4 (13,3)
blaCTX-M-2 - - - - - - + + + - - - - + - - - - - - - - - - - - - - - - 4 (13,3)
blaCTX-M-8 - - - - - - - - + - - - - + - - - - - + - - - - - - - - - - 3 (10,0)
blaTEM - - - - - - - - + - - - - - - - - - - + - - - - - - - - - - 2 (6,7)
blaSHV - - - - - - - - + - - - - - - - - - - - - - - - - - - - - - 1 (3,3)
Am
pC
blaMOX - - - - - - - - - - - - - - - - - - - - - - - - - - - + - - 1 (3,3)
blaCMY - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - 0
TE
TR
AC
ICL
INA
S
Bo
mb
as d
e E
fluxo tetA - + + + + + + + + + + + - + - - + + - + + - - - + - + + - - 19 (63,3)
tetB - - + - + - - - + - - - + - - - - - - + - - - - + - + + + + 10 (33,3)
tetC - + + + + + + + + - - - - + - - - + - + + + + - + - + + + + 19 (63,3)
tetD + + + + + + + + + + - - - - - - - - - - + + - - + - + + + + 17 (56,7)
tetE - - - - - - - - - - - - - + + - + - - + - - - - + - + + + + 9 (30,0)
Pro
teçã
o
Rib
oss
om
al
tetM - + - - - - - - + - - + + + + + + + + + - - + - - - + - - - 13 (43,3)
tetO - + - - - - - - - - - + - + + - - - - - - - - - - - - - - - 4 (13,4)
tetQ - - - - - - - - - - - + - - + - - - - + - - - - - - - - - - 3 (10,0)
tetS - - - - - + + + + + - - - - - - - - + - - - - - - + - - - 7 (23,3)
tetW - - - - - - - - - - - - - - + - - - - - - - - - - - - - - - 1 (3,3)
QU
INO
LO
N
AS
Pro
teín
as d
e
Pro
teçã
o qnrA - + - - - - - - - - - - - - - - + - - - - - - - - - - - - - 2 (6,7)
qnrB - - + - - - - - - - + + + + + + + + - + + + + - + - + + + + 18 (60,0)
qnrS - + + + + + + + + + + + - + + + + + + + + + + - + + + + + + 27 (90,0)

98
Recommended

















