
AsPac J. Mol. Biol. Biotechnol., Vol. 17 (1), 2009 19 Bacteriocin from Lactobacillus plantarum PMU33 strainAsia Pacific Journal of Molecular Biology and Biotechnology, 2009Vol. 17 (1) : 19-25
*Author for Correspondence.Mailing address: Department of Biochemistry, Faculty of Science, Mahidol University, Bangkok 10400, Thailand. Tel: +66-2-2015600 ; Fax: +66-2-3547174; Email: [email protected]
Two-peptide bacteriocin from Lactobacillus plantarum PMU 33 strain isolated from som-fak, a Thai low salt fermented fish product
W. Noonpakdee 1*, P. Jumriangrit 1, K. Wittayakom1, J. Zendo, J. Nakayama2, K. Sonomoto2 , S. Panyim1
1Department of Biochemistry, Faculty of Science, Mahidol University, Bangkok 10400, Thailand 2Laboratory of Microbial Science and Technology, Division of Bioscience and Biotechnology,
Graduate School of Bioresource and Bioenvironmental Sciences, Kyushu University, 6-10-1 Hakozaki, Higashi-ku, Fukuoka 812-8581, Japan
Received 5 October 2007 / Accepted 20 May 2008
Abstract. A total of 12,520 lactic acid bacteria (LAB) isolated from fermented fish products “som-fak” were screened for bac-teriocin. One Lactobacillus plantarum PMU33 strain produced bacteriocin that inhibited a large number of Gram-positive bacteria including food borne pathogens, Listeria monocytogenes, Bacillus cereus and Staphylococcus aureus. Biochemical studies revealed that the bacteriocin was heat stable even at autoclaving temperature (121°C for 15 min) and was active over a wide pH range (2-10). The bacteriocin purified and characterized from the culture supernatant consists of two peptides with the molecular masses of 3222 and 3099 by mass spectrometry analysis. The molecular mass of this two-peptide bacteriocin were nearly identical to that of two-peptide plantaricin W (Plw) which consists of two peptides Plwα and Plwβ. The genes encoding these two peptides ampli-fied by PCR with Plw gene specific primer showed identical sequences to Plwα and Plwβ. The bacteriocins and their producing strains isolated from som-fak may find application as bio-preservatives in fermented food products.
Keywords. Lactic acid bacteria, Two-peptide bacteriocin, som-fak, fermented fish product.
INTRODUCTION Lactic acid bacteria (LAB) are known to produce many dif-ferent antibacterial substances including bacteriocins which can inhibit the growth of several pathogenic bacteria in the genera Bacillus, Enterococcus , Listeria, and Staphylococcus. Ow-ing to the widespread increased interest in bio-preservation, several studies have been directed at identifying bacteriocin producing lactic acid bacteria that may be used to control the growth of these organisms in fermented products (Huss in fermented products (Huss (Huss et al., 1995, Nes et al., 2004)
The bacteriocins from lactic acid bacteria are ribosomally synthesized antimicrobial peptides, mostly small, heat stable, hydrophobic and cationic peptides (Jack et al., 1995). They can be divided into two distinct families: the nonlantibiot-ics and the lantibiotics. The latter contains the unusual or modified amino acids lanthionine and 3- methyl lanthionine3- methyl lanthionine methyl lanthionine dehydrobutyrine and dehydroalanine. The unusual amino acids in lantibiotics are formed as a result of post transla-tional modifications of precursor peptides. The lantibiotics in general have wider spectrum of activity than the non-lantibiotics. Nisin produced by Lactococcus lactis subsp. lactis, is the best–studied and the only lantibiotic approved in more than 60 countries including USA to be used in food industry
(Federal Register,1998). Both types of bacteriocins are most frequently found as single active peptides, however there have been increasing reports on the two-peptide bacteriocin systems wherein both components are required for full ac-tivity (Garneau et al., 2002). The nonlantibiotic two-peptide bacteriocin; enterocin(Aymerich et al., 2000) lactocin 705 (PalaciosPalacios et al., 1999) and Plantaricin S (Ruis-Barba et al., 1994) together with the new family of lantibiotic two-peptide bacteriocin lacticin 3147 (Ryan et al., 1999), staphylococcin C55 (Navaratna et al., 1999) and plantaricin W (Holo et al., 2001) have been reported to have potential application in food preservation.
Traditional fermented fish products in South East Asia are unique with organoleptic properties and widely prepared by spontaneous microbial fermentation. They offer a new They offer a newThey offer a new source of bacterial strains and/or with special properties for potential biotechnological use such as starter culture. “Som-fak” is a Thai fermented fish product composed of minced fish fillet, salt (2-5 %), boiled rice and minced garlic. The mixture is tightly packed in banana leaves or

20 AsPac J. Mol. Biol. Biotechnol., Vol. 17 (1), 2009 Bacteriocin from Lactobacillus plantarum PMU33 strain
plastic bags and left fermented at ambient temperature for 2-4 days and the microflora becomes dominated by lactic acid bacteria (Paludau-Muller et al., 2002 ). The products can beThe products can be consumed raw or cooked with vegetables as a main dish or a snack. Little is known about microbiology and fermentation pattern of the product even though the products are now in small scale production. However, it was reported that the commonly found LAB in Thai fermented fish products werein Thai fermented fish products werehai fermented fish products werefermented fish products were Lb. plantarum , Lb. pentosus, Lc. lactis, Pediococcus pentosaceus, Weisella confusa, �arnobacterium piscicolaconfusa, �arnobacterium piscicola (Ostergaard et al., 1998, Paludan-Muller et al., 1999). Some of these strains had theSome of these strains had the antilisterial activity but their identities of bacteriocins have not yet been elucidated..
Fermented fish and ready-to-eat fish products have been linked to food safety problems (Rocourt et al., 2000). This might be due to insufficient process of preservation and the product was sometimes consumed without further cooking (Brillet et al., 2004). Attempt to inhibit food pathogenic bac-teria such as Listeria by various methods sometimes had a negative effect on the products or the substances used were not allowed in many countries. Many studies have shown the potential application of bacteriocin producing bacteria in a range of ready to eat food products including in fermented fish (Nilsson et al., 1999). Studying of bacteriocin producing strains isolated from Thai traditional fermented fish products may be useful to use them as starter cultures or co-protective culture for improving the microbial safety of these fermented fish and other products as well.
The purpose of this current study was to isolate and characterize bacteriocin producing LAB from somfak, a traditionally fermented low salt fish product, which is usually consumed without cooking in various parts of Thai-land. The production is typically at household levels and is sometimes associated with problems such as product short shelf life and poor hygiene which may link to food safety problem. The bacteriocin was also purified and identified by a three-step purification method.
MATERIALS AND METHODS
Bacterial strains and media. Bacterial strains used in this study are listed in Table 1. All LAB isolates and indicator strains were grown in MRS broth (Merck, USA) at 30 or 37°C. Listeria, Salmonella, Bacillus were grown in tryptic soy broth (Difco, USA) at 30°C. Enterococcus and Staphylococ-cus were grown in nutrient broth (Difco, USA). Escherichia coli was grown in Luria-Bertani (Difco, USA) medium at 37 °C. All cultures were maintained as frozen stocks held at -80 °C in appropriate broth containing 20 % glycerol (w/v). Throughout the experiments, strains were sub-cultured every 2 weeks on agar media and kept at 4 °C. Before use in experiments, cultures were propagated twice in broth overnight. Soft agar media were prepared by adding
0.6 % (w/v) agar to liquid media. Catalase at a final concentration of 50 U ml–1(Boehringer Mannheim, Germany) was added to soft agar overlays to eliminate the potential inhibitory effect of H2O2 produced by colonies.
Isolation of bacteriocin-producing bacteria and bacteriocin characterization. Bacteriocin-producing bacteria were isolated by the direct plating method (Coventry et al., 1997). A 10% food sample in diluent (0.85% NaCl) was homogenized and 10-fold serially diluted. Pour plates of serial dilutions (1 ml aliquots) in media MRS were incubated under anaerobic conditions (anaerobic generation kit, Merck, USA) for 48 h at 30 °C. Multiple plates of serial dilution (3-5 plates providing a total of approximately 1000 colonies) were overlaid with a set of 4 indicators and incubated at 30 °C for another 18-24 h. The four indicator strains used were Lactobacil-lus curvatus ATCC 25601, Lactobacillus plantarum TISTR 850, Pediococcus pentosaceus TISTR 374 and Propionibacterium freudenreichii TISTR 446. Colonies suspected to be produc-ing zones of growth inhibition in the indicator lawn were randomly selected and removed using a sterile Pasteur pi-pette. The agar plug (from Pasteur pipette) was inoculated into broth media and incubated anaerobically for 48 h at 30°C. Culture supernate was obtained by centrifugation at 10,000 g for 10 min at 4 °C and then was adjusted to pH 6.5 with 5 M NaOH and filtered through a 0.45 µm filter. Inhibition was tested by spotting 10 µl of the supernatant onto soft agar lawn (0.6%) seeded with 0.1 ml of an overnight grown indicator strain and incubated overnight. Cultures producing an inhibitor in broth were then purified by streaking from the broth and restreaking for single colony isolates two to three times. Strains that stably produced inhibitory substance in culture broth after five times sub-culturing were selected for further study.
For a semi quantitative assay of the bacteriocins, two-fold serial dilutions of the supernatant was tested using Lb. plantarum TISTR 850 as the indicator strain. The activ-ity was expressed in arbitrary units (AU ml-). One AU was defined as the reciprocal of the highest serial twofold dilution which did not show inhibition of the indicator strain per mil-liliter (Schillinger et al., 1993). The spectrum of bacteriocin and effect of temperature and protease treatment on anti-microbial activity were determined as previously described (Coventry et al., 1997, Noonpakdee et al., 2003). Traditional fermented fish product “som-fak” were obtained from the markets in northern part of Thailand and in Bangkok. Identification of the bacteriocin producing isolate. Only one isolate (designated PMU33 ) of the 12,520 screened was stably produced bacteriocin in broth media. Strain PMU 33 was therefore further characterized and identified on the basis of Gram stain, catalase reaction and other identification tests (Schillinger and Lucke, 1987). Carbohydrate fermenta-tion was determined using API 50 CHL system (BioMerieux, France) The identification of one chosen PMU33 strain was

AsPac J. Mol. Biol. Biotechnol., Vol. 17 (1), 2009 21 Bacteriocin from Lactobacillus plantarum PMU33 strain
confirmed by 16S rDNA technique using the primers which correspond to nucleotides position 8 to 1510. The sequence obtained was compared with sequences in the database of the DNA Data Bank of NCBI (http://www.ncbi.nlm.nih.gov) using the BLAST program.
Purification, N terminal amino acid sequencing and mass spectrometry analysis of bacteriocin from PMU33 strain. The bacteriocin was purified from 1 litre cultures of Lb. plantarum PMU33 strain grown to early stationary phase about 18-20 h. The bacterial cells were removed by centrifu-gation at 10,000 g for 10 min at 4 °C. Later, 20 g of Amberlite XAD-16 equilibrated with water was added to the supernatant with constant shaking at 4 °C for 3 h. The suspension was then transferred to a column and washed with 100 ml of water and 100 ml of 40 % ethanol in water. The bacteriocin was eluted in 100 ml 70 % (v/v) 2-propanol. The 2-propa-nol in the eluate was then evaporated out. The bacteriocin remaining in the eluate was then pH adjusted to 5.7 by 1 M NaOH and transferred to a SP-sepharose column (10 ml). The matrix was washed with 30 ml of 20mM sodium phosphate (pH5.7) buffer and the bacteriocin was eluted with 10ml of 0.5 mM NaCl. The eluate was adjusted to 0.1 % TFA before subjected to reverse-phase chromatography on a Resource-RPC column (Amersham Biosciences, USA) in HPLC system (USI, Fukuoka, Japan). The peptides were eluted from the column in a water/acetonitrile gradient (20-80%) containing 0.1 % TFA at a flow rate 1 ml/min. Fractions showing bacteriocin activities were collected and purified separately by repeating the reverse phase chromatog-raphy step. The purified peptide from reverse phase HPLC was concentrated for mass spectral analysis and amino acid sequencing. The amino acid sequence was determined by Edman degradation using automatic protein sequencer PPSQ21 with LC 10 system (Shimadzu Corporation, Kyoto, Japan).
The MALDI –TOF MS was recorded in linear mode using a Voyager HP Biosystems (Framingham, MA, USA) equipped with a nitrogen laser. The analyzer operated in positive ion mode with an accelerating voltage of 20 kV. The spectrum was obtained by summing 10 scans. A solution of 10 mg/ml alpha cyano-4-hydroxycinnamic acid in 50 % acetonitrile, 0.1 % trifluoroacetic acid was used as matrix. A 1 µl aliquot of the matrix was then mixed with 1 µl sample and 1 µl of this solution was applied to the target, air dried and analyzed. For internal calibration insulin B with a molecular mass of 5730.6 Da and ACTH fragment with a molecular mass of 2465.2 Da were used. DNA extraction, Polymerase chain reaction (PCR) and DNA sequencing. Extraction of genomic DNA and the polymerase chain reaction analysis followed procedures described by Noonpakdee et al. (2003). The primer set for plw structural gene, 5’-GCGCTTGCCAATGAACA- AAT-3’ and 5’-TATCTTCTCCCCAAACTCAC-3’ generating
a 591-bp amplicon , are located at -39 to -20 and 138 to 121 downstream of the plw gene (Holo et al., 2001). The reaction was performed in a 25 µl mixture in a DNA thermo cycler (Perkin Elmer GenAmp 2400). For plw gene, the amplifica-tion was subjected to 25 cycles consisting of denaturation at 94ºC for 1 min followed by annealing at 58ºC for 1 min and extension at 72ºC for 1 min. The PCR products were analyzed by 1.2 % agarose gel electrophoresis and the size of the amplicons was determined by comparison with a 100bp DNA Ladder.
The amplified PCR products of plw gene were puri-fied by Qiagen quick PCR purification kit (Qiagen, West Sussex, UK). The PCR products were sequenced with ABI Prism Dye Terminator Cycle Sequencing Ready Reaction kit (Perkin-Elmer, USA) and an ABI PRISM 377 DNA sequencer (Perkin Elmer, USA) as described by the instruc-tions of the manufacturer.
RESULTS
Isolation of bacteriocin producing strains. A total of 12,520 colonies from 10 fermented fish som-fak samples were examined for potential inhibitory substance against the four indicator strains. A total of 60 colonies was recorded as positive producing inhibition clear zones on agar media (detection rate 0.48 %). Of these colonies only one desig-nated PMU33 strain stably secreted inhibitory substance into culture broth. This bacteriocin-producing strain PMU33 was a Gram-positive, catalase-negative bacillus, non motile and produced no gas from glucose. The analysis of the carbohy-drate fermentation pattern by the API CHL kit, PMU33 was potentially identified as Lb. plantarum. The identification of PMU 33 strain was further confirmed by 16S rDNA sequence analysis with the GenBank and ribosomal databases (99.9 %) (data not shown).
Characterization of bacteriocin from Lb. plantarum PMU33 strain. The inhibitory spectrums of the bacte-riocin produced by Lb. plantarum PMU33 is presented in Table 1. The bacteriocin exhibited inhibitory activity against a broad range of closely related bacteria in the genera Lactobacillus, Pediococcus, Leuconostoc and some pathogenic strains of Bacillus, Staphylococcus, Enterococcus and Listeria. The bacteriocin was not effective against Gram-negative bacte-ria. Antimicrobial activity was stable after heat treatment at 100 °C for 30 min and remained active over a wide pH range between 2 and 10 (Table 2).
Purification of bacteriocin from PMU33 strain and mass spectrum analysis. Bacteriocin was purified from cul-ture supernatant by a three-step protocol as described in Methods. The fractions, corresponding to the 14 (α peptide) and 20 min (β peptide) in the elution profile from reverse
22 AsPac J. Mol. Biol. Biotechnol., Vol. 17 (1), 2009 Bacteriocin from Lactobacillus plantarum PMU33 strain
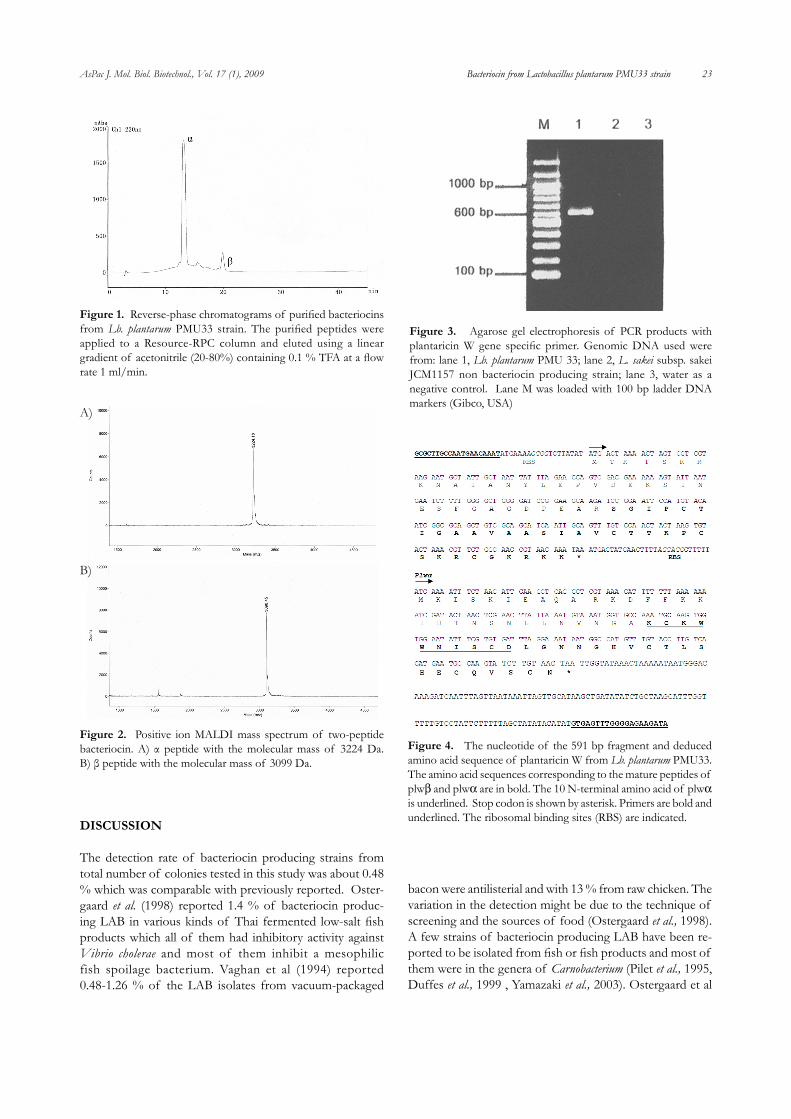
phase HPLC recorded at 220 nm were both active toward indicator strain Lb. plantarum TISTR850 (Figure 1). Each fraction has low activity but when mixed them together high activity was recovered which was similar to that of producing strain (data not shown).
Mass spectrometry analysis for these two fractions showed the molecular mass of 3224 Da and 3099 Da for peptide α and β respectively (Figure 2A,B). The molecular mass of these two peptides are very similar to those predicted of two-peptide plw.
N-terminal sequence of the α peptide. Since the concen-tration of α peptide from the HPLC fraction was higher (Figure 1), we decided to further investigate the amino acid sequence of this peptide. The first 10 N-terminal sequence of this peptide was almost the same as proposed primary structure for plwα with the exception at Cys residues as shown in Table 3. Confirmation that bacteriocin produced by Lb. plantarum PMU33 is plantaricin W. To prove that the bacteriocin
produced by Lb. plantarum PMU 33 was plw, PCR analysis using the published sequences of the plw structural gene (Holo, et al., 2001) was performed. Two primers comple-mentary to sequences occurring proximal to the 3’ and 5’ ends of the plw gene were used to amplify plw gene from the genomic DNA of Lb. plantarum PMU33. The result is shown in Figure 3. A 591 bp fragment was amplified from the genomic DNA of Lb. plantarum PMU 33 (lane 1). No DNA fragment was amplified using genomic DNA ex-tracted from L. sakei subsp. sakei JCM 1157 which did not produce plw ( lane 2), or when no DNA template was present (lane 3). The amplified fragment of 591 bp of the purified DNA of Lb. plantarum PMU33 was then subjected to DNA sequence analysis. Figure 4 shows the nucleotide sequence and the deduced amino acid sequence of the 591 bp ampli-fied DNA product of Lb. plantarum PMU33. The sequences contained 591 bp encoding 2 genes, which were identical to those of plwβ and plwα (Holo, et al., 2001). Two genes were located close together with a ribosome-binding site at 9 and 7 bp upstream of plwβ and plwα and the ORFs encoding two peptide of 67 and 59 amino acid respectively. It was reported that both plwβ and plwα contain leader sequences, once processed gave rise to mature peptides of 32 and 29 amino acids. The sequence homology of plw gene from Lb. plantarum PMU 33 to that of Lb. plantarum LMG2379 was 100 % (data not shown)
Table 1. Antibacterial spectrum of bacteriocin produced by Lactobacillus plantarum PMU 33 strain
Indicatorstraina Sensitivityb
BacilluscoagulanJCM2257 ++c
BacilluscirlulansJCM2504� ++ ++++ BacilluscereusIFRPD2037� + + + +++ BacilluscereusATCC117�7�8� + + + +++ BacillussubtilisJCM14�65 ++ ++++ LactobacilllusplantarumTISTR8�50 + + + + + ++++ LactobacillussakeisupspsakeiJCM1157� + + + + + ++++ LactobacilluscurvatusATCC25601 + + + + + ++++ LeuconostoccremorisATCC19254� + + + +++ LeuconostocmesenteroidesATCC108�30 + + + +++ ListeriamonocytogenesIFRPD2068� + + + +++ ListeriamonocytogenesDMST7�995 + + + +++ ListeriamonocytogenesDMST28�7�1 + + + +++ ListeriainocuaIFRPD207�1 + + + +++ MicrococcusluteusTISTR8�8�4� + ++ PediococcusacidilacticiATCC8�08�1 + + + + + ++++ PediococcuspentosaceusTISTR37�4� + + + + + ++++ PropionibacteriumfreudenreichiiTISTR4�4�6 + + + + + ++++ StaphylococcusaureusTISTR118� + + + +++ StaphylococcusaureusIFRPD2034� + + + +++ EnterococcusfaecalisTISTR927� + + + + + ++++ EnterococcusfaeciumTISTR928� + + + + + ++++ Salmonellasp.TISTR101 � �� SalmonelladerbyA�2 � �� EscherichiacoliATCC4�38�95 � ��
a ATCC, American Type Culture Collection; TISTR, Thailand Institute of Scientific and Technological Research; DMST, Department of Medical Science Thailand; IFRPD, Institute of Food Research and Product Development; JCM, Japan Collections of Microorganism b All indicators were tested for inhibition of growth as determined by a zone of growth inhibition in the indicator lawn culture. c inhibition zone (mm); + + + , 10-15 mm; + + , 5-9 mm; +, 1-4 mm; - , no inhibition
Conditions Bacteriocinactivity(AUml�1)*
Control(noheat) 64�00 100°Cfor30min 64�00 121°Cfor15min 3200 pH 2 6400 3 6400 4 6400 5 6400 6 6400 7 6400 8 3200 9 1600 10 800
The results of the assay are from three experiments, each with tripicate samples. * One AU was defined as the reciprocal of the highest serial twofold dilution which did not show inhibition of the indicator strain per milliliter.
Table 2. Effect of heat and pH on the bacteriocin activity of strain PMU33
Bacteriocin Sequences
PMU33α peptide NH2- K X K W W N I A X D Plwα NH2- K C K W W N I A C D
Table 3. N- terminal amino acid sequences of bacteriocin produced by Lactobacillus plantarum PMU 33 strain
AsPac J. Mol. Biol. Biotechnol., Vol. 17 (1), 2009 23 Bacteriocin from Lactobacillus plantarum PMU33 strain
DISCUSSION
The detection rate of bacteriocin producing strains from total number of colonies tested in this study was about 0.48 % which was comparable with previously reported. Oster-gaard et al. (1998) reported 1.4 % of bacteriocin produc-ing LAB in various kinds of Thai fermented low-salt fish products which all of them had inhibitory activity against Vibrio cholerae and most of them inhibit a mesophilic fish spoilage bacterium. Vaghan et al (1994) reported 0.48-1.26 % of the LAB isolates from vacuum-packaged
Figure 1. Reverse-phase chromatograms of purified bacteriocins from Lb. plantarum PMU33 strain. The purified peptides were applied to a Resource-RPC column and eluted using a linear gradient of acetonitrile (20-80%) containing 0.1 % TFA at a flow rate 1 ml/min.
Figure 2. Positive ion MALDI mass spectrum of two-peptide bacteriocin. A) α peptide with the molecular mass of 3224 Da. B) β peptide with the molecular mass of 3099 Da.
A)
B)
Figure 3. Agarose gel electrophoresis of PCR products with plantaricin W gene specific primer. Genomic DNA used were from: lane 1, Lb. plantarum PMU 33; lane 2, L. sakei subsp. sakei JCM1157 non bacteriocin producing strain; lane 3, water as a negative control. Lane M was loaded with 100 bp ladder DNA markers (Gibco, USA)
Figure 4. The nucleotide of the 591 bp fragment and deduced amino acid sequence of plantaricin W from Lb. plantarum PMU33. The amino acid sequences corresponding to the mature peptides of plwβ and plwα are in bold. The 10 N-terminal amino acid of plwα is underlined. Stop codon is shown by asterisk. Primers are bold and underlined. The ribosomal binding sites (RBS) are indicated.
bacon were antilisterial and with 13 % from raw chicken. The variation in the detection might be due to the technique of screening and the sources of food (Ostergaard et al., 1998). A few strains of bacteriocin producing LAB have been re-ported to be isolated from fish or fish products and most of them were in the genera of �arnobacterium (Pilet et al., 1995, Duffes et al., 1999 , Yamazaki et al., 2003). Ostergaard et al

24 AsPac J. Mol. Biol. Biotechnol., Vol. 17 (1), 2009 Bacteriocin from Lactobacillus plantarum PMU33 strain
(1998) isolated many strains of Lb. plantarum , one strain of Lc. lactis and one strain of �arnobacterium spp. from a Thai fermented whole fish products with antilisterial activity but theirs identities were not known. Recently Campos et al., (2006) reported preliminary characterization of bacteriocins from Lactococcus lactis, Enterococcus faecium and Enterococcus mundtii. strains isolated from turbot.
The bacteriocin produced by Lb. plantarum PMU33 strain was further characterized and purified. The bacteriocin was small, heat stable and acidic resistant. It exhibited inhibitory activity against closely related Gram positive bacteria and some pathogenic strains of Bacillus, Staphylococcus, Enterococcus and Listeria which were not commonly found in bacteriocins from LAB isolated from fish products (Pilet et al., 1995, Duffes et al., 1999, Yamazaki et al., 2003). The bacteriocin was further purified by three-step purification and from HPLC column chromatography pattern shows two active peptides which act synergistically. The molecular mass of these two peptides were 3224 Da and 3099 Da which were identical to the molecular mass of lantibiotic two-peptide bacteriocin plw (plwα and plwβ) previously reported (Holo et al., 2001). Even though the gene sequence of bactericin plw from our PMU33 strain was identical to that of Lb. plantarum LMG2379 strain isolated from fermenting Pinot Noir wine in Oregon, the antimicrobial spectrum of bacteriocins from both strains was a little difference. Plw from Lb.plantarum PMU33 was shown to inhibit pathogenic bacteria in the genus of Bacillus including B. cereus , B. coagulan, B. circulans, B. subtilis while that of Lb . plantarum LMG2379 could not. This might be due to genetic diversities of the tested strains isolated from different regions which might have different pattern of gene expres-sion to different environment (Hong et al., 2007).
Several authors have reported the production of bacteriocins by Lb. plantarum strains from plant or meat origin and they have been used as starter culture for the fermentation of vegetable and sausage products. Some of these bacteriocins have been purified to homogeneity and shown to be antilisterial pediocin-like or two peptide non-lantibiotic bacteriocins (Atrih et al., 2001). However, very few bacteriocins from Lb. plantarum have been reported to be isolated from fish or fish product and none of them have not yet been purified and identified. The lantibiotic two-peptide bacteriocin Plw produced Lb. plantarum PMU33 strain isolated from som-fak in this study might have some applications in food preservation and safety.
REFERENCES
Atrih, A., Rekhif, N., Moir, A.J.G., Lebrihi, A. and Lefebvre, G. 2001. Mode of action, purification and amino acid sequence of plantaricin C19, an anti-Listeria bacterio-cin produced by Lactobacillus plantarum C19. Intenational Journal of Food Microbiology 68: 93-104.
Aymerich, M.T., Garriga, M., Ylla, J.,Vallier, J., Monfort, J.M. and Hugas, M. 2000. Application of enterocins as biopreservatives against Listeria innocua in meat products. Journal of Food Protection 63: 721-726.
Brillet, A., Pilet, M.-F., Bouttefroy, A. and Leroi, F. 2004. Biodiversity of Listeria monocytogenes sensitivity to bacte-riocin-producing �arnobacterium strains and application in sterile cold-smoked salmon. Journal of Applied Microbiology 97: 1029-1037.
Campos, C.A., Rodriguez, O., Calo-Mata, P., Prado, M. and Velazquez, J.B. 2006. Preliminary characterization ofPreliminary characterization of bacteriocins from Lactococcus lactis, Enterococcus faecium and Enterococcus mundtii strains iolated from turbot (Psetta maxima). Food Research International 39: 356-364.
Coventry, M.J., Gordon, J.B., Wilcock, A., Harmark, K., Davidson, B.E., Hickey, M.W., Hillier, A.J. and Wan, J. 1997. Detection of bacteriocins of lactic acid bacteria isolated from foods and comparison with pediocin and nisin. Journal of Applied Microbiology 83: 248-258.
Duffes, F., Leroi, f., Boyaval, P. and Eousset, X. 1999. Inhibition of Listeria monocytogenes by �arnobacterium spp. strains in a simulated cold smoked fish system stored at 4 C. International Journal of Food Microbiology 47: 33-42.
Federal Register 1998. Nisin Preparation: affirmation of GRAS status as a direct human food ingredient. Federal Register 54: 11247-11251.
Garneau, S., Martin, N.I. and Vederas, J.C. 2002. Two-peptide bacteriocins produced by lactic acid bacteria. Biochimie 84: 577-592.
Holo, H., Jeknic, Z., Daeschel, M., Stevanovic, S. and Nes, I.F. 2001. Plantaricin W from Lactobacillus plantarum belongs to a new family of two-peptide lantibiotics. Microbiology 147, 643-651.
Hong, E., Doumith, M., Duperrier, S., Giovannacci, I., Mor-van, A., Glaser, P., Buchrieser, C., Jacquet, C. and Martin, P. 2007. Genetic diversity of Listeria monocytogenes recov-ered from infected persons and pork, seafood and dairy products on retail sale in France during 2000 and 2001. International Journal of Food Microbiology 114: 187-194.
Huss, H.H., Jeppesen, V.F., Johansen, C. and Gram, L. 1995. Bio-preservation of fish products. A review of recent approaches and results. Journal of Aquatic and Food Produc-tion Technology 42: 5-26.
Jack, R.W., Tagg, J.R. and Ray, B. 1995. Bacteriocins of gram positive bacteria. Microbiology Reviews 59 : 171-200.
Navaratna, M.A.D.B., Sahl, H.G. and Tagg, J.R. 1998. Two-component anti-Staphyloccoccus aureus lantibiotic activity produced by Staphylococcus aureus C55. Applied and Envi-ronmental Microbiology. 64: 4803-4808.

AsPac J. Mol. Biol. Biotechnol., Vol. 17 (1), 2009 25 Bacteriocin from Lactobacillus plantarum PMU33 strain
Nes, I.F., Johnsborg, O., 2004. Exploration of antimicrobial potential in LAB by genomics. Current Opinion in Biotech-nology 15: 100-104.
Nilsson, L., Gram, L. and Huss, H. H.1999. Growth control of Listeria monocytogenes on cold-smoked salmon using a competitive lactic acid bacteria flora. Journal of Food Protection 62: 336-342.
Noonpakdee, W., Santivarangkna, C., Jumriangrit, P., So-nomoto, K. and Panyim, S. 2003. Isolation of nisin-Isolation of nisin-producing Lactococcus lactis WNC20 strain from nham, a traditional Thai fermented sausage. International Journal of Food Microbiology 81: 137-145.
Ostergaard, A., Embarek, P.K.B., Wedell-Neergaard, C., Huss, H.H and Gram, L. 1998. Characterization of anti-listerial lactic acid bacteria isolated from Thai fermented fish products. Food Microbiology 15 : 223-233.
Palacios, J., Vignolo, G., Farias, M.E., Ruiz , Holgado, A.P., de Oliver, G. and Sesma, F. 1999. Purification and aminoPurification and amino acid sequence of Lactocin 705, a bacteriocin produced by Lactobacillus casei CRL 705. Microbiology Research 154: 1-6
Paludan-Muller, C., Huss, H.H. and Gram, L. 1999. Char-Char-acterization of lactic acid bacteria isolated from a Thai low-salt fermented fish product and the role of garlic as substrate for fermentation. International Journal of Food Microbiology 45: 219-229.
Paludan-Muller, C., Valyasevi, R., Huss, H. H. And Gram, L. 2002. Genotypic and phenotypic characterizationGenotypic and phenotypic characterization of garlic-fermenting lactic acid bacteria isolated from som-fak, a Thai low-salt fermented fish product. Journal of Applied Microbiology 92 : 307-314.
Pilet, M.F., Dousset, X., Barre, R., Novel, G., Desmazaud, M. and Piard, J.C. 1995. Evidence for two bacteriocins produced by �arnobacterium piscicola and �arnobacterium divergens isolated from fish and active against Listeria monocytogenes. Journal of Food Protection 58: 256-262.
Rocourt, J., Jacquet, C. and Reilly, A. 2000. Epidemiology of human listeriosis and seafoods. International Journal of Food Microbiology 37: 215-219
Ruis-Barba, J.I., Catheart, C.P., Warner, P.J., Jimenez-Diaz, R., 1994. Use of Lactobacillus plantarum LPCO010, a bacteriocin producer, as a starter culture in Spanish-style green olive fermentations. Applied and Environmental Microbiology 60: 2059-2064.
Ryan, M.P., Jack, R.W., Josten, M., Sahl, H.G., Jung, G., Ross, R.P. and Hill, C. 1999. Extensive post translational modification, including serine to D-Alanine conversion, in the two-component lantibiotic Lacticin 3147. Journal of Biological Chemistry 274: 37544-37550.
Schillinger, U. and Lucke, F.K. 1987. Identification of lac-tobacilli from meat and meat products. Journal of Food Microbiology 4: 199-208.
Schillinger, U., Stiles, M.E. and Holzapfel, W.H. 1993. Bac-teriocin production by �arnobacterium piscicola LV61. International Journal of Food Microbiology 20: 131-147
Vaughan, E.E., Caplice, E., Looney, R., O’Rourke, N., Coveney, H., Daly, C. and Fitzgerald, G.F.1994. Isolation from food sources of lactic acid bacteria that produced antimicrobials. Journal of Applied Bacteriology 76: 118-123.
Yamazaki, K., Suzuki, M., Kawai, Y., Inoue, N. and Montville, T.J. 2003. Inhibition of Listeria monocytogenes in cold-smoked salmon by �arnobacterium piscicola CS526 isolated from frozen surimi. Journal of Food Protection 66: 1420-1425.
Recommended

















