
2017, Vol. 13 Núm. 1
Revista Latinoamericana de
Recursos Naturales
UNA REVISTA MULTIDISCIPLINAR
Instituto Tecnológico de Sonora

Revista Latinoamericana de Recursos Naturales©
Una revista interdisciplinar para el conocimiento científico de los recursos naturales en Latinoamérica.
Consejo Editorial
Editores: Fernando Lares Villa ([email protected]), Instituto Tecnológico de Sonora, México.
Sergio de los Santos Villalobos ([email protected]), Instituto Tecnológico de Sonora, México.
Editor técnico:
Roberto Munguía Valencia ([email protected]), Instituto Tecnológico de Sonora, México.
REVISTA LATINOMAERICANA DE RECURSOS NATURALES, Año 13, No. 25, enero-junio
2017, es una publicación semestral editada y publicada por el Instituto Tecnológico de Sonora
(ITSON), a través de la Dirección de Recursos Naturales, con domicilio en 5 de Febrero No. 818
sur Apdo. 335 C.P. 85000, Ciudad Obregón, Sonora, México. Tel:(644)4100923, www.itson.mx,
[email protected]. Editor responsable: Dr. Fernando Lares Villa. Reserva de
Derechos al Uso Exclusivo No. 04-2016-041414023300-203, otorgado por el Instituto Nacional
del Derecho de Autor. Responsable de la última actualización de éste número, Ing. Roberto
Munguía Valencia, con domicilio en 5 de Febrero 818 Sur, Col. Centro, Ciudad Obregón, Sonora,
CP. 85000, fecha de última modificación, 30 de junio de 2017.
Las opiniones expresadas por los autores no necesariamente reflejan la postura del editor de la
publicación.
Queda estrictamente prohibida la reproducción total o parcial de los contenidos e imágenes de la
publicación sin previa autorización del Instituto Tecnológico de Sonora.
© Todos los derechos reservados.

Prefacio
El Instituto Tecnológico de Sonora (ITSON), desde su creación, ha trabajado en el desarrollo de alternativas y sistemas de estudios para el aprovechamiento sustentable de los recursos naturales, creando la Ingeniería en Ciencias Ambientales con el objetivo de formar recursos humanos con la capacidad de solucionar problemas ambientales y gestionar el manejo y aprovechamiento de los recursos naturales, aplicando conocimientos de legislación ambiental, tecnología de descontaminación, diagnóstico y evaluación de impactos ambientales para contribuir al desarrollo sostenible. Los recursos naturales son bienes materiales y servicios que proporciona la naturaleza sin algún impacto antropogénico, los cuales son valiosos para el bienestar y desarrollo de las sociedades humanas, proveyendo materias primas, minerales, alimentos, servicios ecosistémicos, entre otros. De esta manera, el entendimiento y aprovechamiento sostenible de estos recursos representa un gran reto debido a la diversidad de enfoques, temas, y ramas de la ciencia que convergen, además de los aspectos teóricos y/o prácticos abordados. Por esta razón, el estudio de los recursos naturales se basa en la interdisciplinaridad, desde el conocimiento de las relaciones bióticas y abióticas en los ecosistemas, hasta el análisis e integración de las ciencias ómicas de cada uno de estos componentes y su impacto sobre la biósfera. Lo anterior con el objetivo de establecer sistemas de estudios integrales con mayor probabilidad de éxito cuando sean adoptados por el sector económico, productivo, privado y/o político. En este número, se presentan contribuciones científicas que inciden fuertemente en los diversos enfoques y objetivos del ITSON y de nuestra Revista Latinoamericana de Recursos Naturales©. La cual surge como una publicación multidisciplinaria enfocada a describir y aportar conocimiento para la conservación y aprovechamiento sostenible de los recursos naturales. Además, se busca que esta revista sea un vínculo con diversos países para la difusión de los resultados obtenidos en estas áreas, permitiendo así generar colaboraciones fructíferas, independientemente del recurso económico destinado por cada uno de nuestros países a la ciencia y tecnología. De esta manera, en el presente número se presentan cuatro trabajos enfocados a la conservación de los recursos naturales con enfoques teóricos y prácticos, v.gr. a) Evaluación de un sistema de recirculación para producción larvaria de Litopenaeus vannamei; b) Estimación de rendimiento de variedades nativas de maíz; c) Identificación de regiones genómicas asociadas a respuesta a la vacunación contra el PRRS en cerdas; y d) Producción de polihidroxibutirato a partir de la fermentación de suero de leche por Bacillus megaterium TRQ8.
Los Editores:
Fernando Lares-Villa Sergio de los Santos Villalobos
Roberto Munguía Valencia

Leyva-Marquez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 1-7, 2017
1
Evaluación de un sistema de recirculación para producción larvaria
de Litopenaeus vannamei, utilizando la macroalga Gracilaria
vermiculophylla como organismo biorremediador de efluentes
C.I. Leyva-Márquez1 *
, F. Lares-Villa1,2 *
, L.R. Martínez-Córdova3, J.C. Ibarra-Gámez
2, R.
Casillas-Hernández2 y A. Miranda-Baeza
3
1 Programa de Doctorado en Biotecnología, 2 Departamento de Ciencias Agronómicas y Veterinarias, Instituto Tecnológico de Sonora,
Cd. Obregón, Sonora, México 3 Departamento de Investigaciones Científicas y Tecnológicas de la Universidad de Sonora, Hermosillo, Sonora, México
4 Universidad Estatal de Sonora, Navojoa, Sonora, México.
Evaluation of a recirculation system for larval production of Litopenaeus vannamei using the macroalgae
Gracilaria vermiculophylla as an effluent bioremediation organism
Abstract
Looking for sustainable alternatives to larval shrimp production and considering the documented and
successfully use of algae as biofilters for the reduction of nutrients in aquaculture effluents, the present study
was aimed on evaluating the productive and physiological response of Litopenaeus vannamei postlarvae
grown in a recirculating and effluents-bioremediation systems using Gracilaria vermiculophylla. The
production system consisted of two-250 l tanks stocked with 36,000 nauplii, and connected to a sedimentation
tank, which was followed by three 120 l units stocked with the macroalgae in the case of treatments and three
without microalgae for the control. The culture was done for 18 days during which organisms were fed, taking
samples to monitor the biometrics and physical-chemical parameters following standard procedures for larval
production. Recirculation started at day 9th, and water samples were taken daily for the analysis of
ammonium, nitrate and nitrite, as well as for the total heterotrophs and Vibrio spp. counting. The results
showed a removal of nitrogen compounds in the system with macroalgae, averaging 59%, whereas the control
only removed 12%. The number of total heterotrophic bacteria and Vibrio spp., showed a very similar
behavior. The survival of the larvae was 61% in the tanks with G. vermiculophylla and 50% in the control;
however, the other productive parameters were not significantly different. The results suggest that the
incorporation of G. vermiculophylla can represent an efficient alternative to maintain the permissible levels of
ammonium for shrimp larvae cultivation, reducing the water exchange.
Key words: postlarvae, nitrogenous compounds, removal, shrimp culture.
Resumen
Ante la necesidad de buscar alternativas sustentables de producción larvaria de camarón que, y considerando
el uso exitoso y documentado de algas como biofiltros para la reducción de nutrientes en los efluentes
acuícolas, el presente estudio se enfocó en evaluar la respuesta productiva y fisiológica de postlarvas de
Litopenaeus vannamei cultivadas en sistemas de recirculación y biorremediación de efluentes, utilizando
Gracilaria vermiculophylla. Para este propósito se diseñó un sistema de producción de larvas utilizando dos
tanques de 250 l como, mismos que fueron sembrados con 36,000 nauplios, y cada uno se conectó a un tanque
de sedimentación seguido de tres tinas de 120 l, inoculados con la macroalga para el tratamiento y tres sin ella
*Autores de correspondencia
Email: [email protected]; [email protected]; [email protected]

Leyva-Marquez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 1-7, 2017
2
para el control. El cultivo se llevó a cabo durante 18 días, tiempo durante el cual los organismos se
alimentaron y se tomaron muestras para monitorear los parámetros biométricos y físicos-químicos, de acuerdo
a los procedimientos estándar. La recirculación inició el día 9 y diariamente se tomaron muestras para el
análisis de amonio, nitratos y nitritos, así como para la cuenta de heterótrofos totales y Vibrio spp. Los
resultados mostraron una remoción de compuestos nitrogenados en el sistema integrado con G.
vermiculophylla en un promedio de 59%, mientras que el control únicamente removió en promedio el 12%.
En el caso de las cuentas de heterótrofos totales y Vibrio, éstas se comportaron de manera similar. Aunque la
sobrevivencia de las larvas fue del 61% en el tanque con agua tratada con G. vermiculophylla y del 50% en la
no tratada, las diferencias en los otros parámetros de crecimiento no fueron significativas. Debido a la eficacia
en la remoción de N inorgánico en forma de amonio del agua del cultivo larvario de L. vannamei en sistemas
de cultivo cerrados, la incorporación de G. vermiculophylla puede representar una eficiente alternativa para
mantener los niveles de amonio permisibles para el cultivo, disminuyendo el uso de agua para recambio.
Palabras claves: postlarvas, compuestos nitrogenados, remoción, cultivo de camarón.
Introduccción
La camaronicultura es una actividad económica de
rápida expansión en todo el mundo como
consecuencia de la demanda mundial de alimentos,
que ha aumentado considerablemente como
resultado del crecimiento de la población humana
(Martínez-Córdova, 2009; FAO, 2016). Este
acelerado incremento ha traído consigo algunos
problemas relacionados con el impacto negativo que
la actividad provoca en los ecosistemas receptores
de las descargas ricas en nutrientes y actualmente
hay una gran preocupación global respecto a los
impactos ambientales adversos de tales prácticas
(Bardach, 1997; Martínez Córdova et al., 2011;
Naylor et al., 2000), así como con la propagación de
enfermedades y patógenos del camarón,
especialmente aquellos de origen viral. Esto ha
ocasionado que en los últimos años, los problemas
del sector se agraven debido a la aparición de
nuevos patógenos, especialmente virus y bacterias
más agresivos, mortales y resistentes a los
tratamientos convencionales. Además se han
invertido muchos recursos económicos para tratar
de combatir estos problemas sin resultados
positivos; por lo contrario, se han agravado debido a
que se han utilizado toneladas de químicos de
síntesis (antibióticos, desinfectantes, insecticidas,
viricidas, etc.) para tratar de eliminarlos,
aumentando con ello la contaminación de los
estuarios y la sobresaturación de los ecosistemas
con dichos productos. Esta crisis que vive la
industria camaronícola refleja la necesidad de
realizar cambios en los sistemas de producción, por
lo cual se han buscado herramientas de control,
surgiendo como una alternativa el uso sistemas de
biorremediación con los cuales se reducen o
eliminan efluentes, cargas contaminantes (Jiménez
y Balcazar, 2003; Martínez Córdova et al., 2011), y
se evita la dispersión de patógenos hacia el medio
ambiente. Con este enfoque, se ha impulsado la
reducción progresiva del recambio de agua hasta
llegar al cultivo sin recambio (Tacon, 2002).
Además, recientemente la biorremediación de agua,
sedimentos contaminados y aguas de descarga están
involucrando organismos como bacterias,
microalgas, macroalgas e invertebrados filtradores
(Granada et al., 2015). Por estas razones es
necesario buscar alternativas, e implementar
medidas de bioseguridad que sean ecológicamente
sustentables y económicamente rentables, para
impedir continuar con esta espiral de destrucción y
permitir reactivar la tan importante actividad
acuícola
La capacidad de las macroalgas para responder a la
disponibilidad de nutrientes antropogénicos
(nitrógeno y fosforo), hace que sean un eficiente
instrumento para la biorremediación (Marinho-
Soriano et al., 2011). El uso de algas como
biofiltros para la reducción de nutrientes en los
efluentes acuícolas, ha sido descrito como un
método efectivo y rentable (Seenivasan et al.,
2010), y se ha documentado que también es efectivo
para mejorar la calidad del agua en sistemas de
recirculación (Chaitanawisuti et al., 2011). Con el
presente trabajo se pretende generar la información
científica necesaria para ampliar el conocimiento
sobre la efectividad de sistemas de recirculación que

Leyva-Marquez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 1-7, 2017
3
utilicen macroalgas para la biorremediación,
mejorando la calidad del agua así como la respuesta
de postlarvas de L. vannamei cultivada en estos
sistemas cerrados, considerando la poca
información disponible sobre el uso de dichos
sistemas para el cultivo de postlarvas.
Materiales y métodos
Las corridas preliminares para evaluar la capacidad
de biorremediación de la macroalga se realizaronen
las instalaciones del laboratorio de Acuicultura del
Instituto Tecnológico de Sonora, Campus Centro,
Ciudad Obregón, Sonora y el bioensayo final fue
llevado a cabo en el laboratorio de producción de
postlarvas BG Almacenes y Servicios, S.A. de C.V.
ubicado en la Playa de Camahuiroa, Sonora.
Se colectaron ejemplares de Gracilaria
vermiculophylla en el mes de junio, en el área de la
escollera de Camahuiroa (26º29’21”N,
109º15’52”O), aprovechando una bajamar y
condiciones adecuadas de viento en la zona. Se
seleccionaron las algas más brillantes y sanas; se
colocaron alrededor de dos kilogramos en bolsas
plásticas en seco, dentro de una hielera. En el
laboratorio, se lavaron cuidadosamente con
abundante agua de mar filtrada, y con ayuda de un
cepillo muy fino se eliminaron epicomensales y
arena adheridos; fueron pesadas en una báscula
analítica y se colocaron 364.67 ± 6.35g en tres
cilindros de acrílico transparente (tratamiento) con
120 l de agua marina. La iluminación y temperatura
fueron bajo condiciones ambientales de
invernadero.
Los nauplios fueron proporcionados por un
laboratorio productor de postlarvas en estadio de
Nauplio 5; se aclimataron y contaron por método
volumétrico. Se colocaron 36,000 nauplios en las
tinas de tratamiento y control con 250 l de agua
marina y se suministraron 60,000 células por
mililitro de la microalga Thalassiosira sp., como
alimentación inicial. A partir del estadio Zoea I se
tomaron muestras diarias de organismos para
observar condición fisiológica en microscopio de
campo de campo claro Axiolab (Zeiss) y conteo de
células por mililitro de microalga en la Cámara de
Neubauer. Basado en estas observaciones se
ajustaron diariamente las raciones de alimento
teniendo como referencia el protocolo de
alimentación de un laboratorio de producción de
postlarvas. Se realizaron muestreos diariamente
para análisis bacteriológicos de microorganismos
heterótrofos totales y Vibrio de los tanques de
cultivo y de la microalga. Después de 18 días,
cuando las larvas estuvieron en el estadio de
postlarva 12 (PL12), fueron colectadas, medidas y
pesadas para estimar crecimiento y sobrevivencia.
El cultivo se mantuvo bajo condiciones ambientales,
monitoreando periódicamente las variables
principales de la calidad del agua: dos veces por día
temperatura y el oxígeno, y una vez por día la
salinidad, el pH, amonio, nitritos y nitratos,
utilizando técnicas estándar.
Del día 1 al 8 se incorporó agua y microalgas a los
tanques de cultivo para recuperación y aumento de
niveles hasta llegar a los 360 l, y al día 9 se inició la
recirculación, drenando inicialmente agua de los
tanques de cultivo hacia un tanque sedimentador y
posteriormente a los cilindros y réplicas de 120 l
que contenían las algas. El agua se retornó a los
cultivos con ayuda de cabezas de poder marca
Evans, conectadas a la tubería del sistema. Del día
10 al 18 se tomaron muestras para la medición de
niveles de nitrógeno amoniacal, nitritos y nitratos
tanto a la entrada como a la salida del tratamiento y
del control para calcular la remoción de estos
compuestos (Figura 1).
Resultados y discusión
Las algas colectadas (Gracilaria vermiculophylla)
fueron encontradas a una profundidad de 60-100
cm. La temperatura a la que se encontraron fue de
28ºC y durante los dos días de aclimatación se
mantuvieron a 30.2 ± 0.9°C. Diversos trabajos sobre
algas marinas han reportado las actividades
antibacterianas in vitro, como una alternativa a los
antibióticos comúnmente utilizados, y propiedades
nutritivas al co-cultivarlas con organismos marinos,
así como mejoradores de la calidad del agua al
utilizarlas como biorremediadoras. En estos y otros
trabajos se ha demostrado que varias especies del
género Gacilaria, poseen dichas actividades (Abreu
et al., 2009; Cruz-Suarez, et al., 2008; Han et al.,
2013; Lavanya y Veerappan, 2011; Vijayabaskar y
Shiyamala, 2011). Especies del género Gracilaria,
tienen distribución mundial y su presencia en el
ambiente marino obedece a las condiciones
climáticas a lo largo del año (Han et al., 2013;
Lavanya y Veerappan, 2011; Ríos et al., 2009),
predominando su incremento a partir de primavera y
finalizando en invierno; según lo reportado por

Leyva-Marquez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 1-7, 2017
4
Saad-Navarro y Riosmena-Rodríguez (2005), para
las algas rojas, división a la cual pertenecen el
Orden de las Glacilariales y que las encuentran para
el área de Baja California Sur, región vecina a
dónde se hicieron los aislamientos para este
experimento. El uso de Gracilaria vermiculophyla
como agente biofiltrador o biorremediador en
cultivos multitróficos integrados con diferentes
especies acuáticas, y en donde la remoción de
nutrientes en cultivos intensivos, es la preocupación
contante para lograr una acuacultura sustentable, ha
sido bien documentada (Abreu et al., 2011;
Sánchez-Romero, 2013; Skriptsova y
Miroshnikova, 2011). No existen reportes de este
tipo de metodologías aplicadas a la producción de
larvas de Litopenaeus vannamei, por lo que el
presente estudio adquiere relevancia.
Las variables ambientales de los cultivos larvarios,
fueron similares en el tratamiento y en el control,
aun cuando no se mantuvieron en condiciones
controladas. Durante el desarrollo del cultivo
larvario las variables físico químicas se
mantuvieron dentro de los valores recomendados
para el cultivo de L. vannamei (Tabla 1).
Se encontró diferencia significativa entre
tratamiento y control respecto a la concentración de
compuestos nitrogenados. En ambos, los valores de
TAN se mantuvieron dentro de los límites
aceptables para el camarón según Martínez-Córdova
(2009). Durante el día 9 se presentó el valor más
alto, de 1.35 mg l-1
en el tratamiento y 1.87 mg l-1
en el control, y se inició la recirculación en el
sistema en el estadio de PL3. En los próximos tres
días, la concentración disminuyó abruptamente
tanto en el tratamiento como en el control, y
mientras en el tratamiento las concentraciones se
mantuvieron bajas hasta el final del experimento, en
el control se elevaron de nuevo a partir del día 12
(Figura 2a). Aunque los valores de nitritos NO2 y
nitratos NO3 presentaron su valor máximo en el día
8 se mantuvieron dentro de los límites aceptables
para L. vannamei. Los valores máximos de nitratos
fueron de 8.9 mg l-1
en el tratamiento y 7.1 mg l-1
en
el control (Figura 2b) y los valores máximos de
nitritos fueron de 0.30 mg l-1
en el tratamiento y
0.23 en el control (Figura 2c). La remoción de TAN
en el sistema integrado con G. vermiculophylla fue
del 59%, mientras que en el control únicamente se
removió en promedio el 12%.
La respuesta productiva de las larvas se resume en
la tabla 2. Las postlarvas del tratamiento alcanzaron
una ganancia de peso de 0.002 g en promedio, con
una talla de 7.6 mm, ligeramente inferior a los
0.003g y 8.12 mm observados en el control. Esta
diferencia no fue estadísticamente significativa
(p>0.05). La sobrevivencia alcanzada por las larvas
en el tratamiento y control fue del 61% y 50%
respectivamente. De los 364.7 ± 6.4 g colocados
inicialmente de macroalgas en el tratamiento, se
cosecharon 427.3 ± 41.2 g, es decir que se obtuvo
un incremento medio en la biomasa de 62.6 g
(17%).
Figura 1. Representación esquemática del sistema de biorremediación con macroalgas. Las flechas indican los flujos de
agua, (L) Tanque de cultivo de larvas L. vannamei, (S) Tanque de sedimentación, (T) Tratamiento, (C) Control, (1) Punto
de muestreo a tanque de cultivo, (2) Punto de muestreo de entrada de agua a tratamiento y control, (3) Punto de muestreo
de salida de agua de tratamiento y control.

Leyva-Marquez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 1-7, 2017
5
Tabla 1. Variables fisicoquímicas ambientales durante el cultivo de postlarvas de L. vannamei.
Tratamiento Control
Media ± DS Media ± DS
Temperatura (°C) 32.6 ± 1.4 32.5 ± 1.6
Oxígeno (mg l-1) 4.9 ± 1.5 4.9 ± 0.3
Salinidad (ppm) 37.4 ± 1.6 36.7 ± 1.4
pH 8.0 ± 0.2 8.0 ± 0.2
Figura 2. a) Concentración de NH₄+ mg l-1, b) Concentración de NO₂ mg l-1, c) Concentración de NO₃ mg l-1.
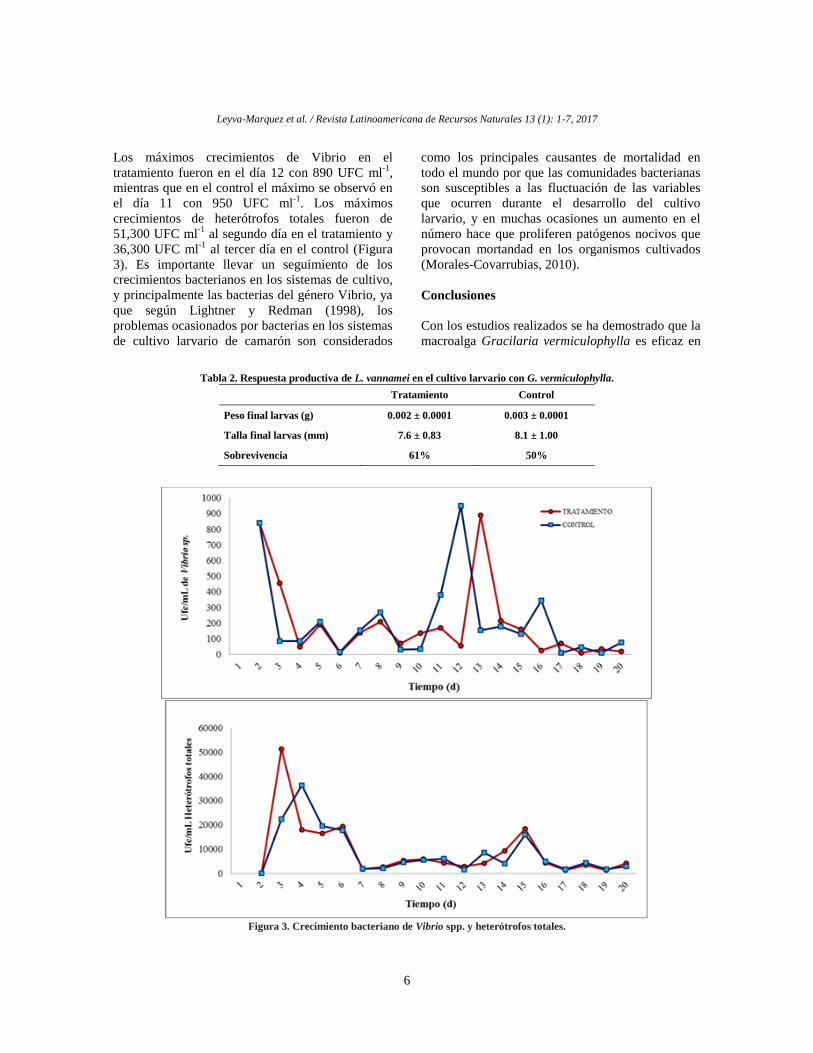
Leyva-Marquez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 1-7, 2017
6
Los máximos crecimientos de Vibrio en el
tratamiento fueron en el día 12 con 890 UFC ml-1
,
mientras que en el control el máximo se observó en
el día 11 con 950 UFC ml-1
. Los máximos
crecimientos de heterótrofos totales fueron de
51,300 UFC ml-1
al segundo día en el tratamiento y
36,300 UFC ml-1
al tercer día en el control (Figura
3). Es importante llevar un seguimiento de los
crecimientos bacterianos en los sistemas de cultivo,
y principalmente las bacterias del género Vibrio, ya
que según Lightner y Redman (1998), los
problemas ocasionados por bacterias en los sistemas
de cultivo larvario de camarón son considerados
como los principales causantes de mortalidad en
todo el mundo por que las comunidades bacterianas
son susceptibles a las fluctuación de las variables
que ocurren durante el desarrollo del cultivo
larvario, y en muchas ocasiones un aumento en el
número hace que proliferen patógenos nocivos que
provocan mortandad en los organismos cultivados
(Morales-Covarrubias, 2010).
Conclusiones
Con los estudios realizados se ha demostrado que la
macroalga Gracilaria vermiculophylla es eficaz en
Tabla 2. Respuesta productiva de L. vannamei en el cultivo larvario con G. vermiculophylla.
Tratamiento Control
Peso final larvas (g) 0.002 ± 0.0001 0.003 ± 0.0001
Talla final larvas (mm) 7.6 ± 0.83 8.1 ± 1.00
Sobrevivencia 61% 50%
Figura 3. Crecimiento bacteriano de Vibrio spp. y heterótrofos totales.

Leyva-Marquez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 1-7, 2017
7
la remoción de N inorgánico en forma de amonio
durante el cultivo larvario de Litopenaeus vannamei
y que su integración a los sistemas de cultivo
cerrados de cría de postlarvas (en recirculación),
puede representar una eficiente alternativa para
mantener los niveles de amonio dentro de los
límites permisibles para el cultivo, disminuyendo el
uso de agua de recambio.
Bibliografía
Abreu, M.H., Rui, P., Yarish, Ch., Buschmann A.H., Sousa-
Pinto, I. 2011. IMTA with Gracilaria vermiculophylla:
Productivity and nutrient removal performance of the
seaweed in a land-based pilot scale system. Aquaculture 312: 77-87.
Abreu, M.H., Varela, D.A., Henríquez, L., Villarroel, A., Yarish,
Ch., Sousa-Pinto, I., Buschmann A.H. 2009. Traditional vs. Integrated Multi-Trophic Aquaculture of Gracilaria chilensis
C.J. Bird, J. McLachlan & E.C. Oliveira: Productivity and
physiological performance. Aquaculture 293: 211–220. Bardach, J.E. 1997. Acuaculture, prollution and biodiversity. P.
89-99. En: J. E. Bardach (ed.), Sustainable Acuaculture.
John Willy & Sons. USA. Bhakuni, D. y Silva, M. 1974. Biodynamic substances from
marine flora. Botanica Marina 27:40-51.
Bourgaud, F., Gravot, A., Milesi, S. Gontier, E. 2001. Production
of plant secondary metabolites: a historic perspective. Plant
Science 161, 839.851.
Chaitanawisuti, N., Santhaweesuk, W., Kritsanapuntu, S. 2011. Performance of seaweeds Gracilaria salicornia and Caulerpa
lentillifera as biofilters in a hatchery scale recirculating
aquaculture system for juvenile spotted babylons (Babylonia areolata). Aquaculture Int. 19: 1139-1150.
Cruz-Suárez, L.E., Tapia-Salazar, M., Nieto-López, M.G.,
Ricque-Marie, D. 2008. A Review of the Effects of Macroalgae in Shrimp Feeds and in Co-Culture. En:
Avances en Nutrición Acuícola IX, IX Simposio
Internacional de Nutrición Acuícola. Cruz-Suárez, L.E., Ricque-Marie, D., Tapia-Salazar, M., Nieto-López, M.G.,
Villareal-Cavazos D.A., Lazo, J.P., Viana M.A. eds. UANL,
Muevo León, México. Noviembre 24-27. Pp. 304-333. FAO. 2016. Resumen El estado mundial de la pesca y la
acuicultura 2016. Roma. 23 p.
Granada, L., Sousa, N., Lopes, S., Lemos, M. 2015. Is integrated multitrophic aquaculture the solution to the sectors’ major
challenges?- a review. Reviews in Aquaculture 6: 1-18.
Han, T., Jiang, Z., Fang, J., Zhang, J., Mao, Y., Zou, J., Huang, Y., Wang, D. 2013. Carbon dioxide fixation by the seaweed
Gracilaria lemaneiformis in integrated multi-trophic
aquaculture with the scallop Chlamys farreri in Sanggou Bay, China. Aquacult Int 21:1035–1043.
Jiménez, M.G., Balcazar, J.L. 2003. Uso de filtros biológicos en
larvicultura de Litopenaeus vannamei. Principios generales. Revist Aquatic No. 18: 11-14.
Lavanya, R., Veerappan, N. 2011. Antibacterial Potential of Six
Seaweeds Collected from Gulf of Mannar of Southeast Coast of India. Advances in Biological Research 5 (1): 38-44.
Lightner, D.V. y Redman, R.M. 1998. Shrimp diseases and current diagnostic methods. Aquaculture 164:201-220.
Marinho-Soriano, E. Azevedo C.A.A., Trigueiro, T.G., Pereira,
D.C., Carneiro, M.A.A., Camara, M.R. 2011. Bioremediation of aquaculture wastewater using macroalgae
and Artemia. International Biodeterioration 65: 253-257.
Martínez-Córdova, L. 2009. Camaronicultura Sustentable: Manejo y Evaluación. Ed. Trillas. Primera Edición. México,
D.F. 176 p.
Martínez-Córdova, L.R., López-Elías, J.A., Leyva-Miranda, J.G., Armenta-Ayon L., and Martínez-Porchas, M. 2011.
Bioremediation and reuse of shrimp Aquaculture effluents to
farm White leg shrimp, Litopenaeus vannamei: A first approach. Aquaculture Research, 42, 1415-1423.
Morales-Covarrubias, M.S. 2010. Enfermedades del camarón:
detección mediante análisis en fresco e histopatología. 2a.
Ed. Trillas-CIAD. México, D.F. 180 p.
Naylor, R.L., Goldburg, R.J., Primavera, J.H., Kautsky, N.,
Beveridge, M.C.M., Clay, J., Folke, C., Lubchenco, J., Mooney, H., Troell, M. 2000. Effect of aquaculture on world
fish supplies. Nature 405, 1017-1024.
Paez-Osuna F. 2001. Camaronicultura y medio ambiente. Programa universitario de alimentos. Universidad de Texas.
452 p.
Ríos, N., Medina, G., Jiménez, J., Yánez, C., García, M.Y., Di Bernardo, M.L., Gualtieri M. 2009. Actividad antibacteriana
y antifúngica de extractos de algas marinas venezolanas. Rev
peru biol 16(1): 097- 100. Saad-Navarro, G., Riosmena-Rodríguez, R. 2005. Variación
espacial y temporal de la riqueza florística de macroalgas en
la zona rocosa de Bahía de Muertos, B.C.S. México. Ciencia y mar IX (26): 19-32.
Sánchez-Romero, A. 2013. Evaluación del efecto de la
iluminación en la remoción de nitrógeno por Gracilaria vermiculophylla en un cultivo integrado con camarón
Litopenaeus vannamei en sistema de recirculación. Tesis
Doctoral. Universidad de Sonora, México. Seenivasan, R., Indu, H., Archana, G., Geetha S. 2010. The
Antibacterial Activity of Some Marine Algae from South East Coast of India. American-Eurasian J. Agric. & Environ.
Sci., 9 (5): 480-489.
Skriptsova A.V., Miroshnikova, N.V. 2011. Laboratoru experiment to determine the potential of two macroalgae
from the Russian Far-East as biofilters for integrated multi-
trophic aquaculture (IMTA) Bioresource Technology 102 : 3149-3154.
Tacon, A.G.H. 2002. Thematic review of feeds and feed
management practices in shrimp aquaculture. Report prepared under the World Bank, NACA, WWF and FAO
consortium program on shrimp farming and the
environment. Work in progress for public discussion. Publisher by the consortium. 69 p.
Vijayabaskar, P., Shiyamala, V. 2011. Antibacterial Activities of
Brown Marine Algae (Sargassum wightii and Turbinaria ornata) from the Gulf of Mannar Biosphere Reserve.
Advances in Biological Research 5 (2): 99-102

Maria-Ramírez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 8-14, 2017
8
Estimación de rendimiento de variedades nativas de maíz en el
estado de Tlaxcala
A. Maria-Ramírez2 *
, V.H. Volke-Haller2 y M.L. Guevara-Romero
3
1 El Colegio de Tlaxcala, A.C. Privada Melchor Ocampo 28, Centro, 90600 Tlaxcala, Tlaxcala.
2 Colegio de Postgraduados, Montecillo. Carretera México-Texcoco km 36.5, Montecillo, Texcoco 56230, Estado de México. 3 Facultad de Arquitectura, Benemérita Universidad Autónoma de Puebla. Boulevard Valsequillo s/n. Ciudad Universitaria, Puebla,
Puebla.
Yield estimation of native corn varieties in the State of Tlaxcala
Abstract
Crop yield estimation using different models has been generated with different degrees of complexity, in
terms of the information considered and their degree of precision, the methodologies and the time. In the
present study two mathematical models for the estimation of landraces performance from the State of
Tlaxcala were generated based on characteristics, such as: i) weight of ear, percentage of cob and percentage
of grain moisture; and ii) weight of ear, percentage of cob, and considered the moisture of dry grain in the
shadow. Both models have a good fit (R2 = 0.999), and were valid for estimating performance in landraces of
the State of Tlaxcala. It is worth noting that there is little information about the development and use of
mathematical models such as those generated in this research, and the usefulness they may have.
Key words: physical characteristics of the ear, mathematical models, rainfed maize, races of native maize.
Resumen
Para las estimaciones de rendimiento de cultivos se han generado diversos modelos, con diferentes grados de
complejidad en cuanto a la información que consideran y su grado de precisión, a las metodologías y al
momento en que se aplican. En la presente investigación se generaron dos modelos matemáticos de
estimación de rendimiento de maíces nativos del estado de Tlaxcala, con base en características de la mazorca
como: uno, peso de la mazorca, porcentaje de olote y porcentaje de humedad del grano, para cuando se
dispone de un medidor de humedad; y, otro, peso de la mazorca, y porcentaje de olote, y considera la
humedad del grano seco a la sombra, para cuando no se dispone de un medidor de humedad. Ambos modelos
presentan un buen ajuste (R2 = 0.999), y son válidos para estimar el rendimiento en maíces nativos del estado
de Tlaxcala. Cabe resaltar que existe escasa información sobre el desarrollo y uso de modelos matemáticos
como los generados en esta investigación, y la utilidad que ellos pueden tener.
Palabras claves: características físicas de la mazorca, modelos matemáticos, maíz de temporal, razas de
maíces nativos.
*Autores de correspondencia
Email: [email protected]; Fax (246) 4647726

Maria-Ramírez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 8-14, 2017
9
Introduccción
Las estimaciones de rendimiento de los cultivos
resultan de la mayor importancia para la economía
de un país, a través de las diferentes actividades en
que se encuentran directa o indirectamente
involucradas (Díaz, 1990; Tinoco y García, 2009;
Aguilar-Ávila y Santoyo-Cortés, 2013). Para estas
estimaciones se han generado diversos modelos, con
diferentes grados de complejidad en cuanto a la
información que consideran y el grado de precisión
que dan las estimaciones, de metodologías y del
momento en que se aplican (Aguilar-Ávila y
Santoyo-Cortés, 2013).
Los modelos más amplios pueden tener sus
limitaciones en ámbitos donde predominan parcelas
de superficies pequeñas, de variabilidad climática y
por tanto de fechas de siembra variables y de
prácticas de manejo del cultivo diferentes, o por
carencia de la información necesaria (Aguilar-Ávila
y Santoyo-Cortés, 2013).
Alguno de los modelos más sencillos es uno basado
en la estimación de rendimientos a nivel predial,
que, dependiendo de los objetivos que se busquen,
puede incluir además de la estimación del
rendimiento por superficie, variables como la
variedad del cultivo, de manejo del cultivo, de suelo
y clima, y de daños por fenómenos climáticos.
En el caso del maíz, se han generado modelos
sencillos para realizar estimaciones de rendimiento
en regiones de productores pequeños con base en
características de la mazorca, en que estas varían en
su tamaño y forma y otras de sus características,
pudiendo considerar también la variedad y variables
de suelo y clima, y de manejo agronómico del
cultivo (Díaz, 1990; Ángeles-Gaspar et. al., 2010;
Aguilar-Ávila y Santoyo-Cortés, 2013). A este
respecto, Díaz (1990) desarrolló y utilizó un modelo
matemático para estimar el rendimiento de maíz en
regiones del estado de Puebla, basado en
información de la mazorca como largo y ancho, el
porcentaje de olote de la mazorca y de humedad del
grano, y la densidad de población del cultivo,
considerando también ajustes por daños de heladas
y sequía, con fines de observar cambios en el
rendimiento derivados de la transferencia y uso de
tecnologías de producción nuevas por los
productores. Pérez (2001) presenta una metodología
y modelo para la estimación de rendimiento de maíz
en parcelas de los productores, que considera el
muestreo e información de la variedad, peso de 22
mazorcas y peso medio de cinco de ellas, y peso de
grano, factor de desgranado y porcentaje de
humedad de las cinco mazorcas, para finalmente
calcular el rendimiento de la parcela por hectárea
considerando el número de plantas por hectárea y el
número medio de mazorcas por planta. Aguilar-
Ávila y Ávalos-Gutiérrez (2013) presentan una
metodología y modelo para la estimación de
rendimientos de maíz a nivel de parcela de
productores, que considera el muestreo e
información del cultivo como prácticas de
producción, presencia de fenómenos climáticos y de
plagas y enfermedades, y variables de la mazorca
como daños por fenómenos climáticos y por plagas
y enfermedades, factor de desgranado, humedad del
grano, número de granos por mazorca y peso
específico de un grano, además del número de
plantas por hectárea y el número medio de mazorcas
por planta.
Una vez obtenida la información de las variables de
la mazorca consideradas, se podrá obtener un
modelo matemático que permita estimar el peso de
grano de una mazorca, y con base en la medición
del número de plantas por hectárea y el número
medio de mazorcas por planta, se obtiene el
rendimiento por hectárea.
La precisión del modelo podrá estar afectada por
factores de tipo biótico (plagas y enfermedades),
abióticos (temperatura, sequías, heladas, suelos y de
manejo del cultivo), que afecten el rendimiento, y
esto determina que en la información a captar se
puedan considerar también estos factores y su
variación, a fin de obtener un modelo más preciso.
En el proceso de obtención del modelo matemático
estarán involucrados aspectos de muestreo y
metodológicos que será necesario considerar para
llegar a un modelo adecuado, que pueda ser
representativo para la región; por ejemplo, en el
aspecto metodológico, para la estimación del
modelo normalmente se ha considerado el
procedimiento “selección a pasos” (Díaz, 1990) sin
embargo, este procedimiento no siempre permite
obtener los modelos más adecuados (Volke, 1981).
El objetivo de la presente publicación fue
desarrollar un modelo o modelos matemáticos de
estimación de rendimiento de maíces nativos del
estado de Tlaxcala, a partir de características físicas
de la mazorca, bajo condiciones de temporal, con
fines de estimación de rendimientos.

Maria-Ramírez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 8-14, 2017
10
Materiales y métodos
Para desarrollar la presente investigación se usó la
información generada en el Proyecto FZ016,
Conocimiento de la Diversidad y Distribución
Actual del Maíz Nativo y sus Parientes Silvestres en
México, Segunda Etapa 2008-2009, que el Instituto
Nacional de Investigaciones Forestales, Agrícolas y
Pecuarias (INIFAP) realizó para la Comisión
Nacional para el Conocimiento y Uso de la
Biodiversidad (CONABIO), y que comprendió la
investigación: Diversidad y Distribución Actual de
los Maíces Nativos en Tlaxcala, como parte del
Proyecto antes mencionado (María y Hernández,
2010).
Esta información se refiere a 256 muestras de
mazorca de maíces nativos en parcelas de
productores, obtenidas en el estado de Tlaxcala en
los ciclos de producción de 2008-2009 y 2009-
2010.
Región de estudio
El estado de Tlaxcala se encuentra ubicado en en
centro del pais, con altitudes entre 2200 y 3000 m, y
comprende 60 municipios, de los cuales se
cubrieron las principales áreas productoras de maíz
del estado (María y Hernández, 2010).
Los valores medios de precipitación total anual en
la región oscilan entre 262 (Rancho Zoquiapan,
Benito Juárez) y 793 mm (ciudad de Tlaxcala), que
se distribuyen entre 67.3 y 84.2 % en los meses de
junio a octubre, respectivamente (María et al.,
20051). Las temperaturas medias anuales van de
13.0 a 14.5 °C en la parte noroeste del estado, de
14.5 a 15.2 °C en la parte sur y de 13.5 a 15.2 °C en
la parte sureste (INIFAP, 2012).
Los suelos presentes en el estado de Tlaxcala, según
el sistema FAO corresponden a Andosoles (5.20
%), Arenosoles (1.75 %), Cambisoles (9.99 %),
Durisoles (11.87 %), Fluvisoles (2.51 %), Gleysoles
(0.06 %), Leptosoles (11.50 %), Luvisoles (5.68 %),
Phaeozem (33.97 %), Regosoles (13.30 %),
Solonchak (0.06 %) , Vertisoles (0.80 %) y
Umbrisoles (2.00 %), con una superficie de
hacentamientos humanos de 1.35 % (INEGI, 1986;
1 María R., A.; Medina G., G. y Ruiz C., J. A. 2005. SICATLAX
Sistema de Información para Caracterizaciones Agroclimáticas.
En: Foro Sistemas Producto e Innovaciones Tecnológicas, Experiencias y Perspectivas Regionales Tlaxcala-Puebla-
Hidalgo. p. 119-128, y con datos de la Comisión Nacional del
Agua (CONAGUA) hasta 2010, usando SICATLAX,.
SECODUVI, 2013). Estos suelos difieren en sus
características de textura, profundidad y pendiente,
y conjuntamente con las condiciones climáticas y
disponibilidad de riego, tienen un uso de praderas y
de cultivos, principalmente maíz y hortalizas en las
áreas con riego, con algunas superficies menores
con bosques.
En 2015, en el estado de Tlaxcala se sembraron
122,326 ha de maíz, donde 106,302 ha (86.9 %)
fueron de temporal, con un rendimientopromedio
de 2.5 t ha-1
, y 16,024 ha (13.1 %) fueron con riego
con un rendimiento medio de 2.8 t ha-1
(SIAP,
2017).
Para maíz de temporal se presentan áreas de muy
buena productividad (MBP), de buena
productividad (BP) y de mediana productividad
(MP). Las áreas de MBP presentan una
precipitación media anual de 675 a 800 mm en el
período de junio a octubre y suelos de más de 1.0 m
de profundidad, con rendimientos de hasta 6.0 t ha-
1; las áreas de BP presentan una precipitación media
anual en el período de junio a octubre de 625 a 675
mm y suelos de más de 0.7 m de profundidad, con
rendimientos de hasta 4.0 t ha-1
; las áreas de MP
presentan una precipitación media anual en el
período de junio octubre de 475 a 625 mm y una
profundidad del suelo mayor a 0.5 m, con
rendimientos de hasta 3.0 t ha-1
(María et al., 2002,
2003).
Hernández (2014), menciona que 90 % del maíz
sembrado es con semilla nativa o criolla, y que ese
alto porcentaje de siembra de maíces nativos
muestra el gran interés de las comunidades
campesinas por preservar este grano y el esfuerzo
que realizan por resistir la introducción de
variedades mejoradas.
Razas de maíz
María y Hernández (2010), reportan que las razas de
maíz identificadas en 256 colectas en 34 municipios
del estado de Tlaxcala, fueron: Cacahuacintle (2.7
%), Chalqueño (13.7 %), Chalqueño por Bolita (0.4
%), Chalqueño por Cacahuacintle (0.4 %),
Chalqueño por Cónico (4.3 %), Cónico (44.5),
Cónico por Elotes Cónicos (0.8 %), Cónico por
Cacahuacintle (0.4 %), Cónico por Bolita (3.1 %),
Cónico por Chalqueño (9.4 %), Cónico por Pepitilla
(0.4 %) y Elotes Cónicos (19.9 %)2. Así, 78.5% de
2La clasificación directa fue de Hernández Casillas, J.M., experto
de maíz del Instituto Nacional de Investigaciones Forestales,
Agrícolas y Pecuarias.

Maria-Ramírez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 8-14, 2017
11
las muestras colectadas, independientemente del
color, están relacionadas con las razas de maíces
Cónicos.
El color predominante del grano de las razas de
maíz es el blanco, con 91.3 %, y 5.3 % presentan
grano de color amarillo y 3.4% de color rojo, azul y
negro (Medina, 2016).
Muestreo de mazorcas
Las parcelas de los productores en las que se
colectaron las muestras, se distribuyeron en la
superficie de siembras de maíz considerada, con
base en los caminos de acceso, y las muestras se
obtuvieron tanto de manera directa en el momento
de la cosecha como en las viviendas de los
productores cuando la cosecha ya se había
realizado, pero no se habían desgranado las
mazorcas.
En el ciclo de producción primavera-verano 2008-
2009 se colectaron 204, y en el ciclo primavera-
verano de 2009-2010 se colectaron 52 muestras; en
algunos casos, se obtuvo más de una muestra por
parcela, si a simple vista se consideraba que las
mazorcas podían corresponder a diferente raza.
En cada muestra se obtuvieron al azar 50 mazorcas
que fueren representativas, y que no estuvieron
dañadas; esto, ya sea, al momento de la cosecha en
los montones en que se van hacinando o del llenado
de bolsas para su transporte, o en los sitios de
almacenamiento y/o secado en los hogares. Las
mazorcas se secaron al aire y a la sombra, hasta que
su estado de humedad permitiese su desgranado
manual. De las 50 mazorcas se tomaron al azar 15,
se ordenaron según su longitud, y se tomaron las 10
centrales para medir las variables a considerar en
cada una de ellas.
Las variables de la mazorca para las cuales se tomó
información fueron: longitud, desde la base hasta la
punta de la mazorca (cm), diámetro, en la parte
central (cm), diámetro del olote, en la parte central
(cm), número de hileras, número de granos por
hilera, peso del grano (g) y contenido de humedad
(%).
El diámetro de la mazorca y del olote se midió con
un vernier metálico marca Truper, el peso de la
mazorca se registró con una balanza mecánica
Ohaus 750 SW, de 2 610 g, y el contenido de
humedad se determinó con un medidor portátil de
humedad marca Dickey-John Grain Moisture Tester
Estados Unidos.
Con el peso de la mazorca y el peso del grano se
determinó el porcentaje de olote, cuyo cálculo es
igual a:
• Porcentaje de olote = [(peso de mazorca – peso
de grano) / (peso de mazorca)] * 100 (%).
Con el porcentaje de humedad del grano se calculó
el peso del grano a humedad comercial (14.0 %),
como:
• Peso del grano a humedad comercial de 14.0 %
= (100 – porcentaje de humedad del grano a la
cosecha) / 86 (g).
Por otra parte, se realizó una clasificación de las
razas de los maíces nativos presentes en la región,
con la finalidad de observar si la variación de la
forma y tamaño de la mazorca fuere una variable a
considerar en la estimación del rendimiento a nivel
de la región, de acuerdo a la distribución de ellas en
ésta.
Análisis de la información
La información se analizó para el peso de grano a
humedad comercial (14 %) por mazorca en función
de las variables de la mazorca y la humedad del
grano, mediante regresión, con fines de obtener uno
o más modelos matemáticos. Para esto se siguió el
procedimiento propuesto por Volke (2008).
Con el modelo de regresión obtenido, se
determinaron los valores predichos de peso de grano
a humedad comercial de una mazorca, considerando
las variables que incluyó el modelo en sus valores
de interés, medios o de mayor frecuencia. Con el
modelo obtenido y, en su caso considerando la
distribución en la región de los valores de las
variables que incluyese, además de una densidad de
población dada o su distribución en la región, se
podrá obtener el rendimiento medio en la región, lo
cual no fue objetivo de esta investigación.
Resultados
En el proceso de obtención de un modelo
matemático para el peso de grano a humedad
comercial (14 %) de una mazorca, se probaron todas
las variables de la mazorca medidas. De estas
variables, en todos los modelos probados quedaron
incluidas las variables de peso de la mazorca
húmeda, porcentaje de humedad de la mazorca y
porcentaje de olote de la mazorca. Otras variables
asociadas a la forma de la mazorca que pudiesen
intervenir en el peso de grano de la mazorca, puesto
que en la región, ya que se presentan distintas razas
de maíz, como diámetro y longitud de la mazorca,

Maria-Ramírez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 8-14, 2017
12
diámetro del olote, relación largo/diámetro de la
mazorca y las mismas diferentes razas de maíz, no
mejoraron el ajuste de un modelo que incluyese a
las variables de peso de la mazorca húmeda,
porcentaje de humedad de la mazorca y porcentaje
de olote de la mazorca.
En estos términos, el modelo matemático obtenido
para el peso de grano a humedad comercial (14%)
de una mazorca fue el siguiente:
PG = 45.933 + 0.9065 PM – 1.487 HG – 8.233 PO0.50
(Pr. F = 0.0001, CME = 0.515, CV = 0.566 %, R2 = 0.999)
Dónde: PG = peso de grano de una mazorca, a 14.0
% de humedad (g); PM = peso medio de 10
mazorcas, a humedad de cosecha (%); PO =
porcentaje de olote (%); HG = humedad de la
mazorca a la cosecha (%).
Cabe aclarar que en el año 2009 se presentaron
heladas en la etapa de llenado del grano en la zona
de El Carmen Tequexquitla, en 3.9% de las
localidades, que afectaron y disminuyeron la
producción de grano de la mazorca. Las colectas de
las localidades afectadas se incluyeron en la
estimación del modelo matemático como una
variable auxiliar; sin embargo, para lo cual no se
observó un efecto significativo en el ajuste del
modelo, lo que tiene su explicación en que las
heladas causaron una disminución del tamaño
(diámetro) y peso de la mazorca, y un incremento
del porcentaje de olote de la mazorca, con un
incremento medio de 5.1%, y fue esta la variable
que quedó incluida en el modelo. Este incremento
del porcentaje de olote de la mazorca indica que la
helada afectó un mayor grado al grano que al olote.
Sin embargo, podrá ocurrir que el efecto de la
helada pueda variar según el momento en que
ocurra al estado de desarrollo de la mazorca.
Para estimar el rendimiento en un año con daño de
heladas a partir del modelo obtenido en esta
investigación, se debe considerar el peso medio de
las mazorcas muestreadas a su humedad de cosecha,
la humedad media de las mazorcas, y el porcentaje
de olote sin y con daño de heladas, y con los valores
de peso de grano de una mazorca medidos se
procede a obtener la media ponderada, según el
porcentaje de superficie estimada con daño de
helada.
Con el modelo matemático obtenido se calcula el
peso medio de grano por mazorca de un productor,
introduciendo en él un correspondiente peso medio
de mazorca a la cosecha, un porcentaje medio de
olote y una humedad media del grano.
Para el caso en que no se disponga de un medidor
de humedad, se procedió a determinar un modelo
que excluyese esta variable, a condición de que las
mazorcas se hayan secado a la sombra por cierto
tiempo hasta que resultase fácil su desgranado a
mano, de modo que tendiesen a tener un contenido
de humedad relativamente constante. Con la
presente información, la distribución de mazorcas
por contenido de humedad del grano fue la que se
indica en el tabla 1.
Tabla 1. Distribución de mazorcas por contenido de
humedad del grano.
Contenido de humedad
(%)
Porcentaje de mazorcas
(%)
10.5 – 11.0 0.8
11.1 – 12.0 20.3
12.1 – 13.0 52.7 13.1 – 14.0 19.9
14.1 – 17.5 6.3
Teniendo en cuenta que las mazorcas deban llegar a
un contenido de humedad lo más uniforme posible,
mediante su secado, se consideraron las mazorcas
con humedad entre 12.0 y 13.0, que además
comprendieron 52.7 % del total, y con ellas se
calculó un modelo.
En este modelo se consideró una humedad del grano
en 12.5 %, y a partir de este valor se estimó el
modelo a 14.0 % de humedad del grano, que es el
siguiente:
PG = 28.566 + 0.9052 PM – 8.542 PO0.50
(Pr.F = 0.0001, CME = 0.558, CV = 0.591 %, R2 = 0.999)
Donde: PG = peso de grano de una mazorca, a 14.0
% de humedad (g), PM = peso medio de 10
mazorcas, secadas a la sombra, en promedio a 12.5
% de humedad (g); PO = porcentaje de olote (%).
El modelo obtenido presenta un buen ajuste, sin
embargo, su cuadrado medio del error es
ligeramente mayor que el del modelo que incluye el
porcentaje de humedad de la mazorca.
Otro modelo de estimación de rendimiento,
utilizado por investigadores como Díaz (1990), es el
que considera las variables largo y diámetro de la
mazorca, al cual se le estaría agregando el peso de
la mazorca, e igual que el modelo anterior,
consideraría la humedad de la mazorca secada a la
sombra hasta que fuese fácil su desgranado a mano.

Maria-Ramírez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 8-14, 2017
13
No obstante, estando presente la variable peso de la
mazorca, de fácil determinación, las variables
longitud y diámetro de la mazorca no presentaron
significancia, reflejo ello de la alta correlación entre
el peso de la mazorca y estas variables, de:
• peso – longitud: r = 0.859 (p = 0.0001),
• peso – diámetro: r = 0.843 (p = 0.0001),
motivo por lo cual no se obtuvo un modelo
matemático con estas variables.
Una vez obtenido el modelo matemático que estima
el peso medio de grano a humedad comercial (14
%) por mazorca; a nivel de productor, se puede
estimar el rendimiento de grano por superficie
(hectárea). Para esto se requiere conocer el número
de mazorcas por hectárea, igual al número de
mazorcas en una superficie muestreada, por un
factor para expresar a número de mazorcas por
hectárea, donde:
• Superficie muestreada = [(largo de los surcos
muestreados (m)) * (ancho promedio de los surcos
muestreados (m))] (m2)
• Factor para expresar por hectárea = 10000 /
(superficie cosechada) (m2)
• Número de mazorcas por hectárea = (número
de mazorcas cosechadas en una superficie) * (factor
para expresar por hectárea).
Díaz (1990), propone un procedimiento para el
muestreo del número de plantas por superficie,
válido también para determinar el número de
mazorcas por superficie, que consiste en tomar
mediciones del número de mazorcas y ancho de
surco en cinco sitios de 10 m lineales distribuidos
en la parcela según el formato de muestreo,
obteniendo el número promedio de plantas y de
mazorcas y calculando el área promedio (largo por
ancho) muestreada.
El número de mazorcas por hectárea puede variar
entre productores, lo mismo que la superficie de las
parcelas; sin embargo, si la muestra es
representativa, la estimación del rendimiento a nivel
de región corresponderá al producto del número
medio de mazorcas por hectárea por las hectáreas
totales.
Discusión
Dado que el objetivo de la investigación fue generar
uno o más modelos matemáticos para estimar el
rendimiento de grano de maíz para los maíces
nativos del estado de Tlaxcala, que considerasen
factores bióticos para tratar de tener mejor
precisión, y además fueran de uso relativamente
sencillo, se generaron dos modelos. El primero
requiere del peso medio de 10 mazorcas, el
porcentaje de olote y la humedad del grano; el
segundo considera también estas variables, excepto
la humedad del grano por si no se dispone del
instrumento para obtenerla, aunque con un ajuste
ligeramente menor que el primero.
Estos modelos incluyeron: uno, tres variables de la
mazorca, como el peso de la mazorca a humedad de
cosecha, la humedad del grano y el porcentaje de
olote, y otro, el peso de la mazorca seca a la sombra
y el porcentaje de olote, con valores ambos de R2 =
0.999.
En estos términos, los modelos obtenidos presentan
un buen ajuste y son de fácil aplicación,
considerando que la humedad del grano en su caso y
el porcentaje de olote son variables que es necesario
determinar, aunque en uno de los modelos se
consideró la humedad del grano seco a la sombra.
Díaz (1990) generó modelos matemáticos del peso
de grano de la mazorca, ya corregido por porcentaje
de olote y porcentaje de humedad del grano, en
función de las variables longitud y diámetro de la
mazorca: uno, con las variables simples y
cuadráticas y las interacciones entre ellas, con un
total de ocho variables y un R2 de 0.840; otro,
partiendo de las mismas ocho variables y aplicando
el procedimiento de “selección a pasos”, que
incluyó la variable lineal de diámetro y las
interacciones: negativa entre las variables lineal de
longitud y diámetro al cuadrado y positiva entre la
variable lineal de diámetro y cuadrática de longitud,
con un R2 de 0.829.
El modelo con ocho variables puede estar
mostrando signos contrarios a los esperados, para
una o más variables, por ejemplo, negativo para la
longitud de la mazorca, debido a las correlaciones
entre las variables, y el modelo obtenido con el
procedimiento “selección a pasos” no incluyó la
variable lineal de longitud, y en cambio incluyó las
interacciones: negativa cuadrática de diámetro por
lineal de longitud y positiva lineal de diámetro por
cuadrática de longitud, lo que resulta difícil de
interpretar en términos del fenómeno.
Este resultado concuerda con el hecho de que el
procedimiento de “selección a pasos” no siempre da
los mejores modelos, ya sea por exclusión de
variables con sentido según el fenómeno o con
signos contrarios a los esperados, derivado ello de
una alta correlación entre las variables que el

Maria-Ramírez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 8-14, 2017
14
procedimiento precisamente trata de superar.
Por último, cabe resaltar que en la literatura existe
escasa información sobre el desarrollo y uso de
modelos matemáticos como los generados en el
presente caso, y la utilidad que ellos pueden tener.
Conclusiones
Se generaron dos modelos matemáticos para la
estimación de rendimiento de grano de maíces
nativos del estado de Tlaxcala, con base en
variables de la mazorca como peso de la mazorca y
porcentaje de olote, y porcentaje de humedad del
grano, relativamente sencillos y con buen ajuste
estadístico (R2 = 0.999, en ambos), cuya validez es
solo para este tipo de maíces en el estado de
Tlaxcala.
Bibliografía
Aguilar-Ávila, J. y Avalos-Gutiérrez, C. 2013. Estimación de
cosecha de maíz. En: J. Aguilar-Ávila y V.H. Santoyo-
Cortés (Coords.), Estimación de rendimientos en el sector agropecuario. Universidad Autónoma Chapingo-Porrúa,
México, D.F., pp.11-22.
Aguilar-Ávila, J. y Santoyo-Cortés, V.H. 2013. Introducción. En: J. Aguilar-Ávila y V.H. Santoyo-Cortés (Coords.),
Estimación de rendimientos en el sector agropecuario.
Universidad Autónoma Chapingo-Porrúa, México, D.F., pp. 7-10.
Ángeles-Gaspar, E., Ortíz-Torres, E., López, P.A. y López-Romero, G. 2010. Caracterización y rendimiento de
poblaciones de maíz nativas de Molcaxac, Puebla. Fitotecnia
Mexicana, 33(4):287-296. Díaz C., M. 1990. Manual para estimar rendimiento de maíz y
determinar el uso de la tierra en programas de desarrollo
agrícola regional. Colegio de Postgraduados, Chapingo, Estado de México, México.
Hernández R. C. 2014. La Tierra del Maíz. Instituto Tlaxcalteca
de la Cultura - Gobierno del Estado de Tlaxcala. Tlaxcala, México.
INEGI (Instituto Nacional de Estadística y Geografía). 1986.
Síntesis geográfica de Tlaxcala. Secretaría de Programación y Presupuesto. México, D.F., pp. 97.
INIFAP (Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias). 2012. Pronóstico Agroclimático
Tlaxcala. SAGARPA-INIFAP-Secretaría de Fomento
Agropecuario-Fundación Produce, Tlaxcala, Tlaxcala, México.
María R. A., García N., H., Ortiz T.,C., Medina G.,G., y
Mendoza H.,J. 2002. Sistema de Información de zonas potenciales para la producción de cultivos en el Estado de
Tlaxcala. INIFAP, Tlaxcala, Tlaxcala, México.
María R., A., Rojas, M., I., Ávila P., M.A. y Gámez V., J.A. 2003. Producción de maíz de temporal en el Estado de
Tlaxcala. INIFAP, Tlaxcala, Tlaxcala, México.
María R., A. y Hernández C., J.M. 2010. Diversidad y
distribución actual de los maíces nativos en Tlaxcala.
Informe final de actividades 2008-2010, INE-CONABIO-
INIFAP, Tlaxcala, Tlaxcala, México. Medina D., M.L. 2016. La defensa del maíz nativo en Tlaxcala,
2006-2012. Tesis de Doctorado, El Colegio de Tlaxcala
A.C., Tlaxcala, Tlaxcala, México. Pérez-Camarillo, J.P. 2001. Metodología para la evaluación de
cosecha de maíz en parcelas comerciales. INIFAP-
Fundación Hidalgo produce, Pachuca, Hidalgo, México. SECODUVI (Secretaría de Obras Públicas, Desarrollo Urbano y
Vivienda). 2013. Programa de Ordenamiento Territorial y
Desarrollo Urbano para el estado de Tlaxcala. Periódico oficial no. extraordinario, junio 4 de 2013.
SIAP (Servicio de Información Agroalimentaria y Pesquera).
2017. Programas de producción agrícola. Avance de siembras y cosechas. Resumen por estado. Acciones y
Programas. Resumen por estado. Consultado el 15 de enero
de 2017. http://infosiap.siap.gob.mx:8080/agricola_siap_gobmx/Resu
menDelegacion.do;jsessionid=23C82F12CB1A924A7C39C
56A10574B4F Tinoco A., C.A. y García A., J.L. 2009. Validación de una
metodología generada para la predicción de cosechas de arroz para el estado de Veracruz. En: reunión de avances en
la investigación agrícola, pecuaria, forestal y acuícola en el
trópico mexicano. INIFAP-UV-CP-UACH-ITUG-ITBOCA. Veracruz, Veracruz, México. pp. 79-85.
Volke 1981. Estimación de funciones de producción mediante
regresión en experimentos con fertilizantes y densidad de plantas, con fines de determinación de óptimos económicos.
Colegio de Postgraduados, Chapingo, México.
Volke H., V. 2008. Estimación de funciones de respuesta para información de tipo no experimental, mediante regresión.
Colegio de Postgraduados. Montecillo, Estado de México.

Luna-Nevárez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 15-23, 2017
15
Identificación de regiones genómicas asociadas a respuesta a la
vacunación contra el PRRS en cerdas reproductoras del sur de
Sonora, México
G. Luna-Nevárez1 *
, C.M. Aguilar-Trejo1, R.I. Luna-Ramírez
1, R. Zamorano-Algandar
1, X.
Zheng2, M.E. Enns
2, S. Speidel
2, M.G. Thomas
2 y P. Luna-Nevárez
1
1 Departamento de Ciencias Agronómicas y Veterinarias; Instituto Tecnológico de Sonora; C.P. 85000, Ciudad Obregón, Sonora.
2 Department of Animal Sciences; Colorado State University; P.C. 80523-1171, Fort Collins, Colorado.
Identification of Genomic Regions Associated to PRRS Vaccination Response in Replacement Gilts from South of
Sonora, Mexico
Abstract
The porcine respiratory and reproductive syndrome (PRRS) is a viral disease that seriously affects porcine
production in Mexico. The most important method to prevent such disease is the vaccination of replacement
sows, whose highly-variable response involves a complex genetic component. Therefore, objective of this
research project is to discover genetic basis associated to a physiological-clinical indicator of vaccination
response (IVR) against PRRS virus, such indicator is composed by the variables rectal temperature (TR) and
daily weight gain (GPD). This study included 100 replacement sows with breed composition ¾Landrace and
¼Yorkshire, about 6-month of age, and vaccinated against PRRS using a modified live virus (day 0). Data
from rectal temperature and live weight were collected the days -7, 0, 7, 14, 21, 28 and 35 after vaccination.
Blood samples were obtained from each sow at day 40 after vaccination and poured on blood collection cards.
The cards were processed for genomic analyses using a low density device which included 10,000
polymorphisms. A Bayesian model for genomic analysis called Bayes C was implemented in the software
GenSel to study the genetic variation of the VRI explained by molecular markers previously associated to
VRI. The genomic analysis detected 13 genic windows within chromosomes 3, 4, 5, 9, 12, 15, 16 and 17,
which explain about 32% of the total genomic variation associated to IVR. In conclusion, there exist genetic
components associated to IVRI, and this assumption is supported by the genomic analyses that suggests the
IVR is gene-influenced. Therefore, this study could be proposed as an important strategy for genetic selection
to identify sows with favorable response after vaccination against the PRRS virus.
Key words: replacement sows, genome, PRRS, genetic variation.
Resumen
El síndrome respiratorio reproductivo porcino (PRRS) es una enfermedad de origen viral que afecta
seriamente la producción porcina en México. El principal método para prevenir dicha enfermedad es la
vacunación de las hembras de reemplazo, cuya respuesta altamente variable tiene un complejo componente
genético. Por lo anterior, el objetivo de la presente investigación fue descubrir bases genéticas asociadas a un
indicador fisiológico-clínico de respuesta a la vacunación (IRV) contra el virus del PRRS; dicho indicador
está compuesto por los caracteres de ganancia de peso diaria (GPD) y temperatura rectal (TR). El estudio
incluyó 100 marranas de reemplazo de raza ¾Landrace y ¼Yorkshire, con edad de 6 meses y vacunadas con
el virus vivo modificado del PRRS (día 0). Se recolectaron datos de temperatura rectal y peso vivo de todas
*Autores de correspondencia
Email: [email protected]

Luna-Nevárez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 15-23, 2017
16
las hembras los días -7, 0, 7, 14, 21, 28 y 35 posteriores a la vacunación. Muestras de sangre fueron obtenidas
de cada hembra el día 40 posterior a la vacunación y depositadas en tarjetas de recolección “Blood Cards”.
Las tarjetas fueron procesadas para análisis genómico utilizando un dispositivo de baja densidad de 10,000
polimorfismos. Un modelo bayesiano de análisis genómico Bayes C fue implementado en el software Gensel
para estudiar la variación genética del IRV que es explicada por los marcadores moleculares que resultaron
previamente asociados a dicho indicador. El análisis genómico detectó 13 ventanas génicas dentro de los
cromosomas 3, 4, 5, 9, 19, 12, 15, 16 y 17, que explican cerca del 32% del total de la variación genómica
asociada al IRV. Se concluye que existen componentes genético asociado al IRV, ya que el análisis el
genómico arrojó resultados que sugieren en forma confiable que el IRV está regulado por genes. Por lo tanto,
el presente estudio puede ser propuesto como una potencial estrategia de selección genética de cerdas con
respuesta favorable a la vacunación contra el virus del PRRS.
Palabras claves: cerdas de reemplazo, genoma, virus del PRRS, variación genética.
Introduccción
El síndrome reproductivo y respiratorio porcino
(PRRS) es la más significativa enfermedad que
afecta comercialmente a la producción porcina en
Norteamérica, Europa y Asia (Rowland et al.,
2012). El cerdo (Sus scrofa), tanto doméstico como
silvestre, es la única especie conocida que resulta
naturalmente susceptible al PRRS (AHA, 2004). El
PRRS causa problemas reproductivos en animales
en crianza, y problemas respiratorios y reducción
del desarrollo en animales en crecimiento
(Boddicker et al., 2012). El fallo reproductivo se
caracteriza por infertilidad, momificación fetal,
abortos, agalactia y el nacimiento de lechones
muertos, o tan débiles que mueren poco después de
nacer, debido a trastornos respiratorios e infecciones
secundarias como Salmonella choleraesuis,
Haemophilus parasuis, Streptococcus suis,
Mycoplasma hyopneumoniae y virus de la influenza
porcina (Hill, 1996). En cerdos en crecimiento el
virus del PRSS causa una neumonía intersticial
(Collins et al., 1992). El virus del PRRS (PRRSV),
pertenece al orden Nidovirales, familia
Arteriviridae, género Arterivirus (Zimmerman et
al., 2006). El período de incubación oscila entre 4 y
8 días experimentalmente, pero en focos naturales
puede durar entre 3 y 37 días (AHA, 2004). Las
medidas utilizadas actualmente para controlar el
PRRS incluyen despoblación, repoblación y
eliminación del hato, bioseguridad, prueba y
remoción, y vacunación (Corzo et al., 2010). La
estrategia de vacunación es la menos costosa para
los porcicultores y es accesible para todos los
tamaños de productores (ej. pequeños, medianos y
grandes), comparada con otras estrategias de control
del PRRS. Hay dos tipos de vacunas disponibles
comercialmente, una es una vacuna con virus vivo-
modificado (VVM) y la otra es una vacuna con
virus muerto (VM). (Charerntantanakul et al.,
2010).
Por otro lado, estudios asociativos de genoma
completo relacionados con el PRRS en cerdos se
han elaborado con anterioridad. Bodicker et al.
(2012) realizaron un estudio asociativo de genoma
completo (GWAS) utilizando los caracteres de
carga viral y ganancia de peso en cerdos expuestos
al PRRSV. Los genotipos obtenidos usando un
dispositivo de perfil genómico (chip) de ~60,000
SNPs identificaron regiones genómicas asociadas
con carga viral en los cromosomas 4 y X, y con
ganancia de peso en los cromosomas 1, 4, 7, y 17.
Por otra parte, Serão et al. (2014) realizaron estudio
asociativo en cerdos expuestos a la vacuna del
PRRS utilizando los caracteres de ganancia de peso
y presencia de anticuerpos; ellos reportaron 61,565
SNPs útiles e identificaron regiones genómicas
asociadas a ganancia de peso en el cromosoma 4, y
para presencia de anticuerpos en los cromosomas 2,
6, 7, 13, 10 y 18. Hasta el momento, no se ha
identificado un estudio considerando sólo los
caracteres de temperatura rectal y ganancia de peso
asociados con respuesta favorable a la vacuna del
PRRS, en el proceso de aclimatación.
Adicionalmente Serão et al. (2016) reportaron la
validación de genes asociados a la resistencia a la
enfermedad del PRRS, durante el proceso de
aclimatación de cerdas de reemplazo expuestas a la
vacuna del PRRSV.

Luna-Nevárez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 15-23, 2017
17
Debido a la relevancia que tiene la vacuna del
PRRS como medio para reducir la pérdidas
económicas relacionadas con los signos clínicos de
la enfermedad (Charerntantanakul et al., 2010), el
objetivo del presente estudio es conocer las bases
genéticas que propician una reacción favorable a la
vacuna del PRRS utilizando un indicador
compuesto por dos caracteres de comportamiento:
temperatura corporal y ganancia de peso, en
marranas expuestas a la vacuna, aplicando una
herramienta estadístico-Bayesiana para conducir un
estudio asociativo de genoma-completo (GWAS).
Los resultados del estudio son parte de una primera
etapa para descubrir las bases genéticas de respuesta
de resistencia al virus del PRRS (PRRSV). La
identificación de genes asociados con el indicador
compuesto de respuesta a la vacuna IRV será útil
para implementar posteriormente estudios a nivel
molecular.
Materiales y métodos
Población
Se eligieron inicialmente 100 individuos de una
granja porcina comercial ubicada en el Valle del
Yaqui, al sur del estado de Sonora (LN: 27°17', LO:
109°56', altitud 50 msnm). Los individuos fueron
cerdas primerizas en desarrollo con propósito final
de producción, con 6 meses de edad y que nacieron
el mismo día: 02/11/2012. Su composición racial
fue ¼York + ¾Landrace, con un peso inicial de
108.23 ± 10.9 kg Todas las hembras fueron
seleccionadas en forma aleatoria, y no presentaban
infección al virus del PRRS al inicio del estudio,
momento desde el cual fueron alojadas en corrales
dentro de un área de cuarentena. Después de un
período de adaptación de 7 días, se aplicó la vacuna
comercial por vía intra-muscular con la dosis
mínima inmunizante propuesta por el fabricante
contra el virus del PRRS (virus activo modificado
cepa ATCC-VR-2332 del PRRS DICT50 propagado
en cultivos celulares, Ingelvac PRRS MLV del
laboratorio Boehringer Ingelheim) dando inicio al
experimento, por lo que el día de la vacunación se
consideró como el día 0. Durante el desarrollo del
proyecto fue necesario prescindir de 25 animales,
debido a que fueron sujetos a otras pruebas
experimentales, siendo 75 cerdas el universo final
para estudio.
Fenotipos
Siete días antes de la vacunación contra la
enfermedad del PRRS, se extrajo una muestra de
sangre de cada hembra, las cuales fueron
transportadas al Laboratorio de Diagnóstico Integral
de Patología Animal (DIPA), México. Con las
muestras de ARN identificadas se realizó el análisis
de PCR en tiempo real para PRRS utilizando un kit
comercial (Tetracore Nextgen Real-Time QT-PCR
Target Specific reagents for the detection &
differentiation of North American & European
PRRSV Viral ARN), que reconoce un segmento del
ORF 7, reportándose como número de copias de
ARN del virus PRRS por mililitro de muestra (con
el equipo: Cepheid Smart Cycler Version 2.0d.).
Esta prueba se realizó para garantizar que todas las
hembras a estudiar presentaran un diagnóstico
negativo a la presencia del virus del PRRS.
Las hembras fueron pesadas semanalmente
utilizando una báscula ganadera de corral para
pequeñas y medianas especies, para calcular la
ganancia de peso diaria (GDP; kg). Antes de pasar a
la báscula se les tomó la temperatura rectal (TR;
oC) utilizando un termómetro GLA M750 digital
(GLA Electrónica agrícolas). Las mediciones de
ambas variables se recolectaron los días: -7, 0, 7,
14, 21, 28 y 35, con respecto al día de la aplicación
de la vacuna contra el PRRS. Los estadísticos
descriptivos de las mediciones de se muestran en la
tabla 1.
Todas las hembras fueron categorizadas de acuerdo
a las mediciones promedio durante el estudio de
GDP y TR, como parte de la preparación de la
información para los análisis estadístico-
moleculares.
Para GDP (donde el promedio es igual a 0.467 kg):
1=Debajo del promedio,
2=Por arriba del promedio.
Para TR:
1=Debajo de 39oC, y
2=Arriba de 39oC.
Basándose en las categorías antes descritas se
generó el indicador compuesto, el cual incorporó los
dos caracteres fisiológicos y al cual se le denominó
indicador de respuesta a la vacuna (IRV). Se
eligieron estos dos fenotipos debido a que el
carácter de ganancia de peso ha sido previamente
reportado como asociado a la respuesta a la

Luna-Nevárez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 15-23, 2017
18
vacunación contra el virus del PRRS (Serão, 2014),
así como en estudios asociativos con enfoque
productivo (Fontanesi, 2014; Onteru, 2013; Do,
2015), mientras que el carácter de temperatura
rectal ha sido medido, evaluado y analizado como
un signo clínico en estudios con inoculación del
PRRSV (Ladinig, 2014; Bonckaert, 2016; Han,
2011).
El IRV permitió clasificar a las hembras de la
siguiente forma:
A=Respuesta Altamente Favorable (si GDP=2,
TR=1),
M=Respuesta Medianamente Favorable (si GDP=2,
TR=2),
P=Respuesta Pobremente Favorable (si GDP=1,
TR=2).
Después de la aplicación de criterios en las 75
cerdas de reemplazo para obtener el IRV, la
distribución de individuos en cada sub-universo fue:
28 se identificaron como “Pobremente Favorables”,
18 como “Medianamente Favorables” y,
29 como “Altamente Favorables”.
Genotipos
El día 40 posterior a la vacunación, de cada una de
las 75 hembras se extrajo de la parte trasera-inferior
Tabla 1. Detalle de estadísticos descriptivos para los caracteres (fenotipos).
Día n Min Media Max SD
Ps -7 (kg) 75 85.00 108.30 131.40 11.00
Ps 0 (kg) 75 82.00 109.00 136.00 11.97
Ps 7 (kg) 75 89.00 113.40 137.60 11.54
Ps 14 (kg) 75 93.00 116.60 140.60 11.21
Ps 21 (kg) 75 96.70 121.10 147.20 11.41
Ps 28 (kg) 75 102.20 125.90 155.40 12.33
Ps 35 (kg) 75 100.60 125.30 151.20 12.39
Ps Promedio (kg) 75 93.73 117.09 141.71 10.78
GDP 0 (kg) 75 -3.171 0.875 2.771 0.875
GDP 7 (kg) 75 -2.800 0.733 4.067 1.123
GDP 14 (kg) 75 -1.770 0.461 2.543 0.806
GDP 21 (kg) 75 -1.200 0.634 1.714 0.506
GDP 28 (kg) 75 -1.425 0.602 1.725 0.462
GDP 35 (kg) 75 -1.700 -0.097 1.500 0.621
GDP Promedio (kg) 75 -0.246 0.467 1.247 0.293
TR -7 (°C) 75 38.85 39.63 41.00 0.47
TR 0 (°C) 75 35.29 39.09 42.38 0.67
TR 7 (°C) 75 38.40 39.04 40.04 0.34
TR 14 (°C) 75 31.76 38.62 40.75 1.26
TR 21 (°C) 75 38.16 38.91 40.53 0.33
TR 28 (°C) 75 36.74 39.08 39.95 0.38
TR 35 (°C) 75 38.44 38.89 39.86 0.29
TR Promedio (°C) 75 37.71 38.90 39.62 0.32
Cálculos estadísticos descriptivos (mínima, media, máxima y desviación estándar) para los caracteres de temperatura rectal (TR en °C) y ganancia de peso diaria (GDP en kg) para los días de recolección de datos (-7, 7, 14, 21, 28, 35), así como el promedio del periodo de
recolección. GDP fue calculada con el carácter de peso (Ps en kg).

Luna-Nevárez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 15-23, 2017
19
de la oreja 3 ml de sangre, mediante punción con
jeringa comercial estéril, la cual fue depositada en
tarjetas recolectoras con filtro separador de
componentes sanguíneos “blood cards”. Las tarjetas
fueron transportadas al Laboratorio de
Biotecnología de Reproducción del ITSON y
almacenadas en cajas especiales a temperatura
ambiente (32 oC) durante una semana.
Posteriormente, las tarjetas fueron enviadas al
Laboratorio Neogen/GeneSeek Inc. (Nebraska
USA), donde se procesaron para la extracción y
cuantificación del ADN. Para identificar los
genotipos se utilizó un dispositivo de perfil
genómico de baja densidad (GeneSeek Genomic
Profiler for Porcine Low Density BeadChip, versión
GSGT 1.9.4) con aproximadamente 10,000
polimorfismos de nucleótido simple (SNPs), de
acuerdo al protocolo definido por el proveedor. Este
chip, o circuito integrado en una pequeña tableta, es
comúnmente utilizado por la USDA (Departamento
de Agricultura de los Estados Unidos) por que
contiene SNPs que permiten identificar marcadores
de crianza que están asociados con un desarrollo
estructural robusto y sano del animal. Algunos de
los principales marcadores incluidos son: Gen del
síndrome de estrés porcino, gen de rendimiento
Napole, gen de resistencia a E. coli (F4 ab/ac), y el
gen de resistencia o tolerancia al virus del PRRS
(WUR10000125).
En la tabla 2 se presentan los estadísticos
descriptivos sobre la ubicación de los SNPs
identificados en el análisis genómico.
Control de Calidad
Para el análisis de calidad y depuración de los
genotipos se utilizó el software PLINK v1.07
(Purcell et al., 2007).
Los criterios de control de calidad implementados
con PLINK y que fueron aplicados a los datos de
cada SNP de la población fueron:
Los animales que presentaron una frecuencia (call
rate) >0.9 y porcentaje de error Mendeliano <0.05,
sólo los SNPs con frecuencia >0.9, frecuencia
menor de alelo (MAF) > 0.03, valores de P > 10-6
para la prueba de equilibrio de Hardy-Weinberg y
porcentaje de error Mendeliano < 0.1, fueron
incluidos en el estudio (Yang et al., 2013).
Un total de 8,826 SNPs útiles resultaron después de
aplicar el procedimiento de calidad, para ser
incluidos en el análisis asociativo, en los 75
individuos.
Estudio Asociativo de Genoma Completo (GWAS)
La asociación de los genotipos de cada SNP con el
fenotipo IRV fue analizada considerando todos los
SNP en forma simultánea usando el método
bayesiano para selección genómica llamado Bayes
C (Habier et al., 2011), el cual se implementó con el
software GenSel versión 3.04 (Fernando y Garrick
et al., 2009). Se obtuvo la variación genética
explicada por cada SNP en cada ventana genómica
de un mega base (1 Mb). La ecuación del modelo
estadístico usado fue:
y = Xb + Σ zi αi δi + e; la sumatoria es, de i hasta n
número de SNP.
donde:
y = vector de fenotipos (respuesta a la vacuna del
PRRS, IRV);
X = matriz de incidencias relacionadas con los
efectos fijos de los fenotipos;
b = vector de efectos fijos;
z = vector de covariables del genotipo del SNP (-
10/0/10), basado en el número de alelos B usando la
nomenclatura del proveedor del Chip (GeneSeek);
α = efecto aleatorio de la sustitución alélica para
cada SNP ;
δ = indicador de inclusión/exclusión del SNP
(inclusión=1, exclusión=0);
e = vector de efectos aleatorios del error residual.
Cuando el valor aleatorio de la variable
independiente δ indicó ausencia (exclusión) del
SNP se consideró probabilidad π, y cuando indicó
presencia (inclusión) se consideró probabilidad 1-π.
El efecto residual e se asumió que está normalmente
distribuido, es decir N(0, σ2e). El método Bayes C
Tabla 2. Estadísticos Descriptivos de Distancias entre SNPs
Concepto No. SNPs Min Mean Max SD
Distancia (bp) 8,826 45 342,842 2,749,313 269,757
Estadísticos descriptivos (tamaño de la muestra, valor mínimo, valor máximo, media y desviación estándar) sobre las distancias entre
los marcadores identificados, obtenidos de las muestras de sangre enviadas para análisis genómico del ADN. La unidad de medición de
la distancia es bp (pares de bases).

Luna-Nevárez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 15-23, 2017
20
asume que el efecto aleatorio de la sustitución
alélica de los SNPs (α) es diferente de cero (con δ=1
y probabilidad 1-π), y la varianza es la misma para
el análisis conjunto de todos los SNPs. Las
distribuciones previas para las varianzas (σ2α y
σ2e) fueron asumidas bajo una distribución de
probabilidad χ² (ji-cuadrada) inversa con 4 o 10
grados de libertad. La probabilidad previa de que δi
= 0 se consideró como igual 0.98, es decir π = 0.98,
buscando identificar alta variación. Un total de
40,000 iteraciones de Cadena de Markov fueron
consideradas para el análisis, con 1,000 iteraciones
como máximo para cada eslabón para obtener la
media posterior del efecto de cada SNP. La varianza
genética fue registrada cada 100 iteraciones
(frecuencia de salida a los resultados) para cada
fragmento de 1 Mb de genoma y expresado como
un porcentaje del total de la varianza explicada en el
genoma completa en esa iteración (Zeng et al.,
2014).
Ubicación de Genes Candidatos
La versión de genoma porcino 10.2 (Pig Sscrofa
10.2) fue utilizada para caracterizar a los genes en la
página web “e!Ensembl”. La página fue consultada
con el navegador de páginas web Firefox versión
40.0.3
(http://useast.ensembl.org/Sus_scrofa/Info/Index).
Resultados
Control de Calidad
Se presentan dos cuadros de resúmenes (Tablas 3 y
4) sobre los resultados del proceso de control de
calidad, y corresponden a los animales en los que
existieron genotipos, y cuáles genotipos se
identificaron.
Estudio Genómico
Los resultados del GWAS para IRV usando Bayes
C se muestran en la figura 1. Se identificaron 13
ventanas genómicas con distancia de 1 Mb con alta
representación que explican un 32% de variación
genómica para el IRV. Los 13 locus de caracteres
cuantitativos (QTL) fueron seleccionados por el
criterio de mayor variación genómica por ventana,
para cada cromosoma, y se eligieron los SNPs más
representativos para los cuales la suma conjunta de
su variación genómica supera el 1%. Un resumen
sobre los QTLs se muestra en la tabla 5.
El genoma del cerdo mide aproximadamente 143
432 695 Mb, por lo que incluye 16,251 ventanas de
1 Mb no sobrepuestas basándonos en 8,826
marcadores genotipificados (Onteru et al., 2013).
La ventana 1618 del cromosoma 12 (SSC12, o Sus
Scrofa Chromosome 12 por sus siglas en inglés;
acrónimo para mencionar un cromosoma del
genoma del cerdo), con una distancia de 2 Mb, e
integrada por 8 polimorfismos, fue la que presentó
mayor variación genómica. El QTL correspondiente
a la ventana 1618 inicia en la posición 2 097 504
terminando en la posición 2 979 580 del genoma
porcino. La ventana 499 del cromosoma 3 (SSC3)
con una distancia de 30 Mb, conformada por 3
SNPs fue la que presentó la menor variación
genómica del grupo presentado.
Los tres SNPs con mayor variación genómica en el
análisis fueron: ASGA0088990 con 7.70% de
variación, en la ventana 1618, en SSC12, en la
posición 2,979,580 Mb del genoma;
Tabla 3. Estadísticos Descriptivos de las Muestras Genotipificadas
Concepto n Min Mean Max SD
Call rate 75 0.939 0.983 0.988 0.006
Het rate 75 0.343 0.4 0.436 0.016
Estadísticos descriptivos sobre el porcentaje de existencia del grupo de marcadores del Chip, en la muestra. El concepto “Call rate” es el indicador de existencia, y “Het rate” es indicador de inexistencia.
Tabla 4. Estadísticos Descriptivos para los Marcadores Moleculares
Concepto n Min Mean Max SD
Call rate 8,826 0 0.983 1 0.101
MAF 8,520 0.006 0.306 0.5 0.133
Presentación de cálculos de estadísticos descriptivos para porcentaje de existencia del total de los 10,000 SNPs en el ADN de los
individuos. El concepto “Call rate” es el indicador de existencia, y “MAF” es la frecuencia del alelo menor.

Luna-Nevárez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 15-23, 2017
21
ALGA0092677 con 3.27% de variación, en la
ventana 2295, en SSC17, en la posición 3,264,239
Mb; y ASGA0073565 con 3.16% de variación, en
la ventana 2264, en SSC16 ubicado en la posición
59,713,601 Mb.
Discusión
Estudio Genómico
En el estudio GWAS realizado por Serão et al.
(2014) en donde se analizaron los caracteres
ganancia de peso diaria (ADG) y respuesta de
anticuerpos (indicador S/P-Acc.) en individuos
Figura 1. Gráfico “Manhattan Plot” que presenta en ventanas con distancia de 1 Mb la ubicación de los polimorfismos de
nucleótido simple (SNPs) para cada cromosoma, por porcentaje de variación genómica. La variación fue calculada en base a su
grado de asociación con el indicador de respuesta a la vacuna (IRV) mediante el modelo estadístico-matemático Bayes C del
software GenSel.
Tabla 5. Resultados del Análisis Genómico
Ventana SNP Inicial SNP Final #SNPs %Var Pos. Inicial Pos. Final Cromosoma Mb
1618 ASGA0052479 ASGA0088990 8 7.82 2,097,504 2,979,580 12 2
2295 ALGA0092682 ASGA0100169 6 3.30 3,153,062 3,751,012 17 3
2264 ALGA0090848 H3GA0046792 4 3.18 59,182,785 59,930,060 16 59
614 ALGA0022165 M1GA0005374 5 2.87 3,078,372 3,961,749 4 3
799 ALGA0031834 ALGA0031841 3 2.37 47,148,878 47,782,626 5 47
42 DRGA0000604 DRGA0000615 4 2.34 42,106,831 42,826,453 1 42
1308 ASGA0102944 ALGA0051011 2 2.15 8,381,215 8,780,883 9 8
656 MARC0036045 ASGA0019569 3 2.06 45,255,747 45,908,536 4 45
655 SIRI0000030 ASGA0019551 4 1.56 44,003,320 44,916,881 4 44
2111 ALGA0085492 ASGA0069687 3 1.24 62,062,684 62,506,865 15 62
1300 ASGA0091563 M1GA0026290 9 1.08 70,220 974,310 9 0
1527 ASGA0094517 ASGA0090294 5 1.04 77,141,165 77,708,236 10 77
499 ASGA0014064 H3GA0009179 3 1.03 30,120,718 30,781,487 3 30
Total SNPs 59
Resumen sobre los resultados del análisis genómico. Se presentan los SNPs con su correspondiente variación genómica, agrupados en
ventanas dentro de un mismo cromosoma, indicando el SNP Inicial y Final del grupo. Adicionalmente se muestra la ubicación de inicio
de los SNPs en el genoma del cerdo, así como como la distancia ocupada conjuntamente (Mb).

Luna-Nevárez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 15-23, 2017
22
vacunados contra el PRRSV, para el carácter de
ganancia de peso diaria se identificó una región en
las posiciones 77-81 Mb del SSC4, explicando un
1.8 % del total de la varianza genética, y para
respuesta de anticuerpos en el proceso de
aclimatación se identificaron las regiones: 27-30
Mb del SSC7, y 82-83 Mb del SSC13, explicando
1.3% y 1.2% de total de la varianza genética,
respectivamente. En el estudio GWAS para
identificar bases genéticas de resistencia a la
enfermedad del PRRS (Boddicker et al., 2012),
donde se analizaron los caracteres de ganancia de
peso (WG) y carga viral (VL), para ganancia de
peso se identificaron regiones genómicas en las
posiciones 139-140 Mb del SSC4 y 45 Mb del
SSCX explicando el 15.7% y 1.8% de varianza
genética, y para carga viral en las posiciones: 123-
124 Mb del SSC1, 139-140 Mb del SSC4, 24 Mb
del SSC7, y 32-33 Mb del SSC8, explicando del
total de varianza genética 2.1%, 11.2%, 2.6% y
1.0%, respectivamente.
Realizando un comparativo entre las regiones
genómicas del presente estudio y las mencionadas
anteriormente, para el carácter de ganancia de peso
Serão et al. (2014) encontraron en el SSC4 una
variación genómica de 1.8%, Boddicker et al.
(2012) obtuvieron 15.7% y el análisis realizado en
el presente estudio sobre IRV (GPD y TR) arrojó
7.08%. Las posiciones de las regiones de los
cromosomas comparados no son coincidentes. El
resto de los caracteres no son comparables (S/P-
Acc. y VL).
El resultado total de variación genómica
identificada para el IRV de 32%, en regiones no
consecutivas, se puede considerar bueno, más no
excelente, ya que los resultados más altos de los
otros autores fueron de 17.5% (Boddicker et al.
2012) y 60.1% (Serão et al. 2014).
El marcador molecular WUR10000125, el cual fue
identificado por Bodicker et al. (2012) como gen
con alto potencial para resistencia a la enfermedad
del PRRSV, en el presente estudio no se encontró
asociado genéticamente de manera significativa con
el IRV, aun cuando este SNP se presentó en el
100% de los individuos.
Conclusiones
La identificación de 59 marcadores moleculares
incluidos en las 13 ventanas genómicas en los
animales expuestos a la vacuna en contra del
PRRSV, explica cerca de 32% de la variación
genómica del indicador compuesto IRV. Estos
resultados genómicos sugieren que durante el
proceso de aclimatación a la vacunación del PRRSV
se favorece la ganancia de peso diaria (GDP) y el
control de temperatura corporal (TR); por lo que el
estudio puede ser utilizado como estrategia
complementaria de selección genética de hembras
porcinas con respuesta favorable a la vacunación en
contra del virus del PRRS.
En el estudio se ejemplificó el potencial de un
análisis extensivo del genoma para identificar las
bases que propician superioridad genética en
caracteres de impacto económico como: respuesta
favorable a la vacuna de enfermedades (nuestro
caso), resistencia a enfermedades, adaptación a
condiciones climáticas adversas, o alto desempeño
reproductivo o productivo. Se sugiere dar
continuidad a investigaciones a nivel molecular para
las regiones genómicas encontradas.
Agradecimientos
Al laboratorio DIPA-ITSON por su apoyo en la
realización de las pruebas de laboratorio y a la
Unión Ganadera Regional de Porcicultores de
Sonora por el financiamiento de los análisis de
genoma completo.
Bibliografía
Animal Health Australia (AHA). (2004). Disease strategy:
Porcine reproductive and respiratory syndrome (Version
3.0). Australian Veterinary Emergency Plan (AUSVETPLAN), Edition 3, Primary Industries Ministerial
Council, Canberra, ACT. 2004.
Boddicker N., Waide E. H., Rowland R. R. R., Lunney J. K., Garrick D. J., Reecy J. M. and Dekkers J. C. M. (2012).
“Evidence for a major QTL associated with host response to
Porcine Reproductive and Respiratory Syndrome Virus challenge”. J. Anim. Sci. 90:1733-1746. 2012.
Bonckaert C., Van der Meulen K., Rodríguez-Ballarà I.,
Pedrazuela R., Fenech M. and Nauwynck H.J. (2016). “Modified-live PRRSV subtype 1 vaccine UNISTRAIN®
PRRS provides a partial clinical and virological protection
upon challenge with East European subtype 3 PRRSV strain Lena”. Porcine Health Management 2:12. 2016. Doi
10.1186/s40813-016-0029-y. 2016.
Charerntantanakul, W. (2012). “Porcine reproductive and respiratory syndrome virus vaccines: Immunogenicity,
efficacy and safety aspects”. World Journal of Virology,
1(1), 23–30. http://doi.org/10.5501/wjv.v1.i1.23. 2012. Collins J. E., Benfield D. A., Christianson W. T., Harris L.,
Hennings J. C., Shaw D. P., Goyal S. M., McCullough S., Morrison R. B., Joo H. S., Gorcyca D., and Chladek D.
(1992). “Isolation of swine infertility and respiratory

Luna-Nevárez et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 15-23, 2017
23
syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic
pigs”. J. Vet. Diagn. Invest. 4:117–126. 1992.
Corzo C.A., Mondaca E., Wayne S., Torremorell M., Dee S., Davies P., Morrison R.B. (2010). “Control and elimination
of porcine reproductive and respiratory syndrome virus”.
Virus Res. 2010;154:185–192. 2010. Do D.N., Janss L.L., Jensen J., Kadarmideen H.N. “SNP
annotation-based whole genomic prediction and selection: an
application to feed efficiency and its component traits in pigs”. J Anim Sci. 2015;93:2056–63.
Fernando R. and Garrick D. (2009). “User manual for a portfolio
of genomic selection related analysis”. Second Edition. Iowa
State University. 2009.
Fontanesi L., Schiavo G., Galimberti G., Calo D.G., Russo V. “A
genomewide association study for average daily gain in Italian Large White pigs”. Journal of animal science.
2014;92(4):1385–94. doi: 10.2527/jas.2013-7059
Habier D., Fernando R. L., Kizilkaya K., Garrick D. J. (2011). “Extension of the Bayesian Alphabet for Genomic
Selection”. BMC Bioinformatics 12:186. 2011.
Han K., Seo H. W., Shin J. H., Oh Y., Kang I., Park C., & Chae C. (2011). “Effect of the Modified Live Porcine
Reproductive and Respiratory Syndrome Virus (PRRSV)
Vaccine on European and North American PRRSV Shedding in Semen from Infected Boars” . Clinical and
Vaccine Immunology : CVI, 18(10), 1600–1607.
http://doi.org/10.1128/CVI.05213-11. 2011. Hill H. (1996). “PRRS: Practical strategies for prevention and
management of a positive herd”. Proceedings of the 1996
North Carolina Pork Producers Conference, pp. 1-7, January 9-10, Fayetteville, North Carolina. 1996.
Ladinig A., Wilkinson J., Ashley C., Detmer S. E., Lunney J. K.,
Plastow G., & Harding J. C. S. (2014). “Variation in Fetal Outcome, Viral Load and ORF5 Sequence Mutations in a
Large Scale Study of Phenotypic Responses to Late Gestation Exposure to Type 2 Porcine Reproductive and
Respiratory Syndrome Virus”. PLoS ONE, 9(4), e96104.
http://doi.org/10.1371/journal.pone.0096104. 2014. Onteru S.K., Gobach D.M., Young J.M., Garrick D.J., Dekkers
J.C.M. (2013). “Whole Genome Association Studies of
Residual Feed Intake and Related Traits in the Pig”. PLoS ONE 8(6):e61756. Doi:10.1371/journal.pone.0061756.
2013.
Purcell S., Neale B., Todd-Brown K., Thomas L., Ferreira M.A. (2007). “PLINK: a tool set for whole-genome association
and population-based linkage analyses”. Am J Hum Genet
81: 559–575. doi: 10.1086/519795. 2007. Rowland RRR, Lunney J, Dekkers J. (2012). “Control of porcine
reproductive and respiratory syndrome (PRRS) through
genetic improvements in disease resistance and tolerance”. Frontiers in Genetics. 2012;3:260.
doi:10.3389/fgene.2012.00260. 2012.
Serão N.V.L., Kemp R.A., Mote B.E., Harding J.C.S., Willson P., Bishop S.C., Plastow G.S. and Dekkers J.C.M. (2014).
“Whole-genome scan and validation of regions previously
associated with PRRS antibody response and growth rate
using gilts under health challenge in commercial settings”.
Paper presented at 10th World Congress of Genetics Applied
to Livestock Production, Vancouver, BC, Canada (2014, August 17–22).
Serão NVL, Kemp R.A., Mote B.E., Willson P., Harding J.C.S.,
Bishop S.C., Plastow G., Dekkers J.C.M. (2016) “Genetic and genomic basis of antibody response to porcine
reproductive and respiratory syndrome (PRRS) in gilts and
sows”. Genet Sel Evol.;48:51. 2016. Yang, B., Zhang, W., Zhang, Z., Fan, Y., Xie, X., Ai, H., … Ren,
J. (2013). “Genome-Wide Association Analyses for Fatty
Acid Composition in Porcine Muscle and Abdominal Fat Tissues”. PLoS ONE, 8(6), e65554.
http://doi.org/10.1371/journal.pone.0065554. 2013.
Zeng X., Cockrum R. R., Berge R. R., Neary J. M., Garry F. B., Holt T., Blackburn H. D., Speidel S. E., Garrick D. J., Enns
R. M., Thomas M. G. (2014). “Genetic Correlation and
Genome Wide Association Study of Pulmonary Arterial Pressure and Post Weaning Growth Traits in Angus Heifers
from a High Altitude Breeding Program”. Paper presented at
10th World Congress of Genetics Applied to Livestock Production, Vancouver, BC, Canada (2014, August 17–22).
Zimmerman J., Benfield D., Murtaugh M., Osorio F., Stevenson G. y Torremorell M. (2006). “Porcine reproductive and
respiratory syndrome virus”. En Straw, B.E., D’Allaire, S.,
Zimmerman, J., Taylor, DJ (eds). Diseases of Swine, 9.ª edición, Blackwell Publishing Company, Ames, Iowa
(USA). 2006.

Ruiz et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 24-31, 2017
24
Production of polyhydroxybutyrate from milk whey fermentation by
Bacillus megaterium TRQ8
J. B. Ruiz Aguirre1, L. Z. Oliver Gómez
1, S. de los Santos Villalobos
3, M. L. Sánchez
1, 2*
1 Tecnológico de Monterrey, Escuela de Ingeniería y Ciencia. Vía Atlixcayotl 2301, Reserva Territorial Atlixcayotl, 72453 Puebla, Pue.
2 Universidad Nacional de Quilmes, Dpto de Ciencia y Tecnología, Laboratorio de Ingeniería Genética y Biología Celular y Molecular -
Area Virosis de Insectos. 3 CONACYT- Instituto Tecnológico de Sonora. 5 de Febrero 818 Sur, Col. Centro, Ciudad Obregón, Sonora, México, CP. 85000
Producción de polihidroxibutirato a partir de la fermentación de suero de leche por Bacillus megaterium TRQ8.
Abstract
Plastics derived from oil represent a major environmental problem, due to their long period of degradation,
high emissions of CO2 and consumption of non-renewable resources. Thus, it is necessary to generate
sustainable alternatives to produce natural polymers, such as polyhydroxybutyrate (PHB), a biopolymer
produced from the fermentation of agro-industrial wastes by some microorganisms.
The aim of this work was to analyze the optimum culture medium composition and culture conditions to
improve the yield of PHB production in Bacillus megaterium TRQ8. Milk whey was used as the carbon
source and the fermentation was performed inside a bioreactor under controlled conditions. In order to
achieve this purpose an initial conditioning pretreatment to remove the nutrients from the whey was
implemented, afterwards, the fermentation and extraction of PHB were carried out; furthermore, different
culture media containing ethanol and sodium acetate at different concentrations were tested to enhance the
synthesis of PHB. As a result of the tests performed it was concluded that the medium with ethanol 1% v/v
was the one that increased significantly the PHB production.
Key words: Bacillus megaterium, milk whey, fermentation, polyhydroxybutyrate (PHB), biofilms.
Resumen
Los plásticos que son producidos a partir del petróleo representan un problema ambiental importante, debido
a su largo periodo de degradación, altas emisiones de CO2 y por el consumo de recursos no renovables. Por lo
tanto, es necesario generar alternativas sostenibles para producir polímeros naturales, como el
polihidroxibutirato (PHB), que es un biopolímero producido a partir de la fermentación de residuos
agroindustriales llevada a cabo por algunos microorganismos.
El objetivo de este trabajo fue analizar la composición óptima del medio de cultivo y las condiciones de
cultivo para mejorar el rendimiento de la síntesis de PHB en Bacillus megaterium TRQ8. La fermentación se
realizó dentro de un biorreactor con condiciones controladas y se utilizó suero de leche como fuente de
carbono. Inicialmente se hizo un pretratamiento con la finalidad de eliminar los nutrientes del suero y
posteriormente se llevó a cabo la fermentación y extracción del PHB; además, diferentes medios de cultivo
que contenían etanol y acetato de sodio a distintas concentraciones fueron analizados con el objetivo de
incrementar la producción de polihidroxibutirato. Con base en los resultados obtenidos se concluyó que el
medio con etanol al 1% v / v fue el que aumentó significativamente la producción de PHB.
Palabras claves: Bacillus megaterium, Suero de leche, fermentación, Polihidroxibutirato (PHB), biopelículas.
*Autores de correspondencia
Email: [email protected]

Ruiz et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 24-31, 2017
25
Introduction
Synthetic polymers include a wide range of
materials synthetized by the polymerization of
monomers derived from oil or gas, such as:
polypropylene, polyethylene, polyvinyl chloride,
polystyrene and polyethylene terephthalate
(Andrady and Neal, 2009). Likewise, plastics are
usually made from these polymers using different
chemical additives. These materials are cost-
effective, durable, lightweight and corrosion-
resistant materials with high thermal and electrical
insulation properties that can be extruded, molded
and casted (Thompson et al, 2009b). They are used
in the manufacture of everyday items that generate
social benefits and comfort; many aspects of daily
life involve the use of plastics in many different
areas like transportation, telecommunications,
clothing, footwear and especially for packaging
(Thompson et al, 2009b).
The great variety of today´s polymers and the
versatility of their properties provide an ideal raw
material for the manufacture of many different
plastic products that facilitate the transportation of
food, beverages and other goods (Thompson et al,
2009b). Packaging is the most used application for
these polymers and accounts for about 40.1% of the
overall consumption (Lambert, 2013), however,
most of them are non-biodegradable, and their
increasing accumulation in the environment is
generating several environmental problems (Tokiwa
et al, 2009). Each year, an estimated 500 billion
plastic bags are consumed worldwide (Myint et al,
2012), in addition, the global demand for these
polymers has increased, which negatively impacts
the economy around these products, because the
crude oil is limited (North and Halden, 2013) and it
has been estimated that 260 million tons of plastic
are used per year, accounting for approximately 8%
of world oil production (Thompson et al, 2009).
Among the most significant problems are that
plastic pollution and accumulation in the
environment (natural habitats, shores, oceans, etc.)
affects wildlife due to entanglement and ingestion
of plastic residues by animals, causing serious
injuries, restriction of movement, breathing,
lacerations, ulcers, reproductive problems and death
(Webb et al, 2013). Also, many of the chemicals
that are used in the fabrication of plastics are known
to be toxic (Thompson et al, 2009). It has been
proven that the chemicals used in the manufacture
of plastics are present in human population through
ingestion, inhalation or dermal contact (Talsness et
al, 2009), this toxic substances, like the bisphenol
A, polybrominated diphenyl ether (PBDE) and
tetrabromobisphenol A (TBBPA), affect the
reproductive system, disrupt thyroid hormone
homeostasis, generate an anti-androgen action and
cardiovascular diseases (Stahlhut et al, 2009).
However the most concerning problem is the
current unsustainable production and high
consumption of natural resources like oil to satisfy
the immense demand of plastic items (North and
Halden, 2013).
Thus, the generation or production of other kind of
sustainable and biodegradable materials using
renewable sources to mitigate the high fossil energy
consumption (Gautam, 2009) and CO2 emissions is
required (Naranjo, 2010). Natural polymers, such as
polyhydroxybutyrate (PHB), represent a feasible
and sustainable alternative for the fabrication of
different biodegradable plastics (Getachew and
Woldesenbet, 2016). One of the most important
characteristics of this kind of polymers is their
production using renewable carbon sources like
milk whey, which is generated as an agro-industrial
waste (González et al, 2013).
PHB is a polyester belonging to the
polyhydroxyalkanoates family (Figure 1) that can
be produced by several bacterial strains growing
under stress culture conditions like nutrient
depletion and an excess carbon source (Campuzano
and Vasquez, 2013). Under these conditions,
bacterial cells store PHB intracellularly as a reserve
of carbon and energy in the form of granules
(Babruwad et al, 2015).
Figure 1. Chemical structure of Polyhydroxyalkanoates
PHB is considered as one of the main alternatives to
replace traditional plastics because it has favorable
physicochemical characteristics such as being a
completely biodegradable, strong and highly
hydrophobic material (Wang et al, 2007). The
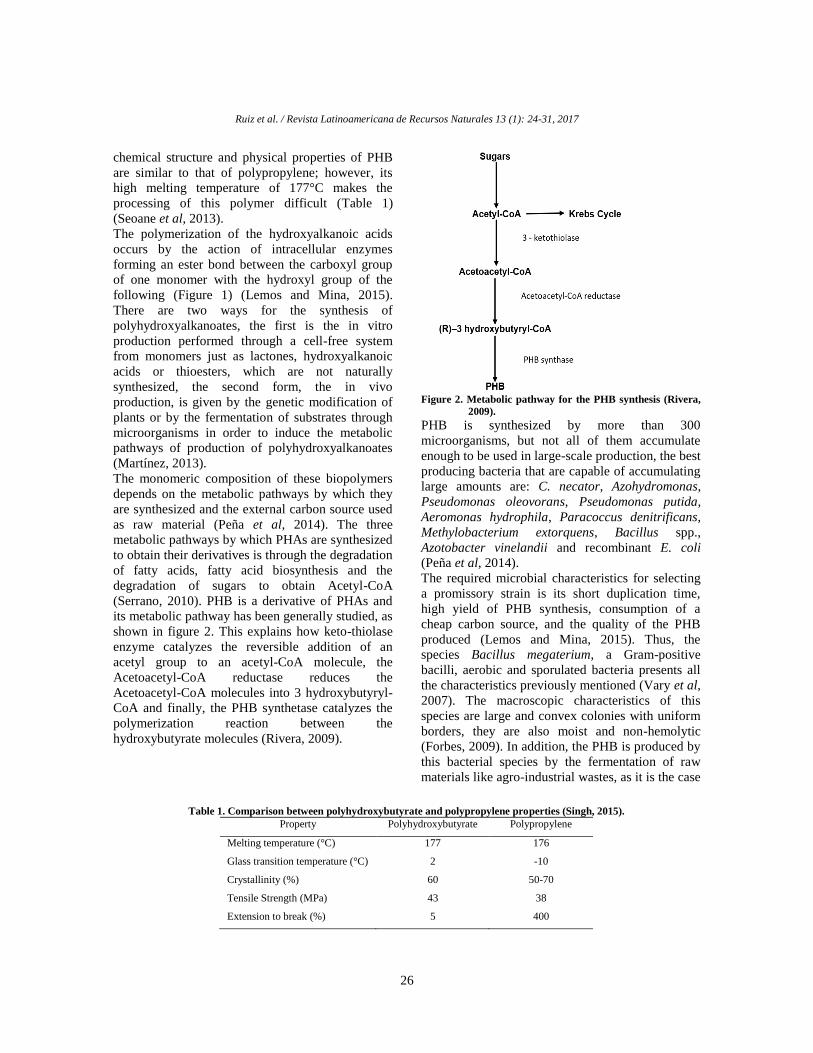
Ruiz et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 24-31, 2017
26
chemical structure and physical properties of PHB
are similar to that of polypropylene; however, its
high melting temperature of 177°C makes the
processing of this polymer difficult (Table 1)
(Seoane et al, 2013).
The polymerization of the hydroxyalkanoic acids
occurs by the action of intracellular enzymes
forming an ester bond between the carboxyl group
of one monomer with the hydroxyl group of the
following (Figure 1) (Lemos and Mina, 2015).
There are two ways for the synthesis of
polyhydroxyalkanoates, the first is the in vitro
production performed through a cell-free system
from monomers just as lactones, hydroxyalkanoic
acids or thioesters, which are not naturally
synthesized, the second form, the in vivo
production, is given by the genetic modification of
plants or by the fermentation of substrates through
microorganisms in order to induce the metabolic
pathways of production of polyhydroxyalkanoates
(Martínez, 2013).
The monomeric composition of these biopolymers
depends on the metabolic pathways by which they
are synthesized and the external carbon source used
as raw material (Peña et al, 2014). The three
metabolic pathways by which PHAs are synthesized
to obtain their derivatives is through the degradation
of fatty acids, fatty acid biosynthesis and the
degradation of sugars to obtain Acetyl-CoA
(Serrano, 2010). PHB is a derivative of PHAs and
its metabolic pathway has been generally studied, as
shown in figure 2. This explains how keto-thiolase
enzyme catalyzes the reversible addition of an
acetyl group to an acetyl-CoA molecule, the
Acetoacetyl-CoA reductase reduces the
Acetoacetyl-CoA molecules into 3 hydroxybutyryl-
CoA and finally, the PHB synthetase catalyzes the
polymerization reaction between the
hydroxybutyrate molecules (Rivera, 2009).
Figure 2. Metabolic pathway for the PHB synthesis (Rivera,
2009). PHB is synthesized by more than 300
microorganisms, but not all of them accumulate
enough to be used in large-scale production, the best
producing bacteria that are capable of accumulating
large amounts are: C. necator, Azohydromonas,
Pseudomonas oleovorans, Pseudomonas putida,
Aeromonas hydrophila, Paracoccus denitrificans,
Methylobacterium extorquens, Bacillus spp.,
Azotobacter vinelandii and recombinant E. coli
(Peña et al, 2014).
The required microbial characteristics for selecting
a promissory strain is its short duplication time,
high yield of PHB synthesis, consumption of a
cheap carbon source, and the quality of the PHB
produced (Lemos and Mina, 2015). Thus, the
species Bacillus megaterium, a Gram-positive
bacilli, aerobic and sporulated bacteria presents all
the characteristics previously mentioned (Vary et al,
2007). The macroscopic characteristics of this
species are large and convex colonies with uniform
borders, they are also moist and non-hemolytic
(Forbes, 2009). In addition, the PHB is produced by
this bacterial species by the fermentation of raw
materials like agro-industrial wastes, as it is the case
Table 1. Comparison between polyhydroxybutyrate and polypropylene properties (Singh, 2015).
Property Polyhydroxybutyrate Polypropylene
Melting temperature (°C) 177 176
Glass transition temperature (°C) 2 -10
Crystallinity (%) 60 50-70
Tensile Strength (MPa) 43 38
Extension to break (%) 5 400

Ruiz et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 24-31, 2017
27
of the milk whey, which is the liquid obtained
during cheese production by coagulating and
separating casein proteins from milk (Naranjo,
2010). Whey contains a large amount of nutrients
such as proteins, lipids and minerals, since it retains
55% of the total milk nutrients it also has a high
lactose content close to 5% or 94 g l-1
(Bovo, 2014).
It is estimated that 1 or 2 kg of cheese can be
produced from every 10 liters of cow's milk, and an
approximate from 8 to 9 kg of milk whey is
produced in this industrial process. Thus, this
product is the most viable alternative to be used as
substrate for the production of PHB (Naranjo,
2010). The milk whey composition is characterized
by the origin of the milk, generally 70% of crude
proteins (beta-lactoglobulin, alpha-lactoglobulin,
and immunoglobulin), protease-peptones, and
native enzymes remains in the serum (Valencia,
2009).
Therefore, the synthesis of PHB by the bacterial
fermentation of milk whey is a promissory
alternative to replace the plastics derived from
petroleum, such as propylene and polypropylene
(Naranjo, 2010). In addition, the use of milk whey
as a carbon source in this process mitigates the
environmental problems caused when it is discarded
to the environment without any treatment (Valencia,
2009). PHB can also be used in 3D printing, tissue
repair and polymer-based depots for controlled drug
release or implants (Luef et al, 2015).
The objective of this work is to present the PHB as
a new alternative polymer for the production of
biofilms and to determine the optimum culture
conditions to enhance its synthesis in B. megaterium
TRQ8.
Materials and methods
Bacterial strain
The strain Bacillus megaterium TRQ8 used in this
work was isolated from wheat rhizosphere in the
Yaqui Valley, Sonora, México. This strain belongs
to Colección de Microorganismos Edáficos y
Endófitos Nativos del Instituto Tecnológico de
Sonora (www.itson.mx/COLMENA) (de los
Santos-Villalobos et al., 2017).
Pre-treatment of the Milk Whey
To be able to use the milk whey as a substrate for
the production of PHB by B. megaterium TRQ8
fermentation, it was necessary to sterilize and to
remove the proteins present in the serum. The milk
whey was sterilized at 115 °C for 15 minutes,
causing the precipitation of a fraction of proteins
(Bell et al, 1983). The supernatant obtained from
the sterilization was filtered using a cellulose-
acetate fiber membrane, then the pH of the
supernatant was adjusted to 7.0 using a 12N NaOH
solution. Finally the solution was centrifuged at
4,400 rpm for 15 minutes and the supernatant
obtained in this step was used to prepare the culture
medium. (Campuzano et al., 2013).
Growth Conditions for Fermentation
For the fermentation of B. megaterium TRQ8 were
used 1.0 x 103 Unit Forming Colonies ml-1, which
was carried out in a bioreactor with a capacity of 5 l
during 48 hours at 32 °C and 200 rpm. The total
volume of whey and culture medium were 2 l, with
10% v/v of the culture medium composed by
KH2PO4, 1.5 g l-1
; Na2 PO4, 9 g l-1
; MgSO4 7H4O,
0.2 g l-1
; and 1 ml l-1
of a solution of trace elements
composed of: FeSO4 7H2O, 10 g l-1
; ZnSO4 7H2O,
2.25 g l-1
; CuSO4 5H2O, 1 g l-1
; MnSO4 4H2O, 0.5 g
l-1
; CaCl2 2H2O, 2 g l-1
; H3BO4, 0.23 g l-1
;
(NH4)Mo7O24, 0.2 g l-1
; and 35% HCl, 10 ml.
(Heinrich et al, 2012; Campuzano et al., 2013).
PHB Extraction, Quantification and Production of
the Biofilms
After 48 hours of fermentation, the culture was
centrifuged at 4,400 rpm during 30 minutes, the
supernatant was discarded and the dry weight of the
cells in g/ml was measured from the obtained pellet.
Then, it was digested with a 4% sodium
hypochlorite solution at a temperature of 37 °C for
20 minutes, the solution was centrifuged again at
4,400 rpm for 30 minutes. The pellet was then
washed with acetone, water and ethanol 70%. Once
the PHB was obtained, its weight was measured and
the percentage of accumulation, in g/ml, was
calculated using the following formula (Mikkili et
al, 2014):
% 𝑜𝑓 𝑃𝐻𝐵 𝑝𝑟𝑜𝑑𝑢𝑐𝑡𝑖𝑜𝑛 =𝐷𝑟𝑦 𝑤𝑒𝑖𝑔ℎ𝑡 𝑜𝑓 𝑃𝐻𝐵
𝐷𝑟𝑦 𝑤𝑒𝑖𝑔ℎ𝑡 𝑜𝑓 𝑐𝑒𝑙𝑙𝑠∗ 100
Finally, 100 ml of chloroform were added at 60 °C
during 15 minutes for each 3 g of PHB obtained, the
mixture was placed on a cellulose paper inside a
Petri dish and the solvent was evaporated at room
temperature in a vacuum for 24 hours (Cyras et al,
2007).

Ruiz et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 24-31, 2017
28
Optimization of PHB Production
The experiment was carried out with the same
inoculation, culture medium, and conditions
previously mentioned, however, 3 modifications
were evaluated with the aim of improving the yield
of the PHB, such as the addition of: i) 1% v/v of
ethanol, ii) 3% v/v of ethanol, iii) 1% v/v of sodium
acetate, and iv) 3% v/v of sodium acetate, 27 hours
after incubation, when the bacteria was in the
stationary phase. In addition, the amount of oxygen
available in a flask was increased to quantify its
effect on the PHB synthesis (Table 2) All the
samples were done by duplicate. After 48 hours of
fermentation, the amount of PHB was quantified,
according to the protocol mentioned before.
Statistics analysis
Data were analyzed by ANOVA analysis of
variance, using Fisher LSD test (P= 0.5), using the
Statgraphics Plus 5.1 software.
Results and discussion
All the treatments, evaluated by duplicate, allowed
the synthesis of PHB from the milk whey
fermentation by B. megaterium TRQ8, since all of
them had the required conditions aforementioned.
The application of external stress has been reported
as an efficient strategy to improve the production of
PHB (Obruca et al, 2011). Thus the culture medium
containing 1% of ethanol increased the production
of polyhydroxybutyrate to 46% (control treatment:
28%) (Figure 3).
In this case, the strain TRQ8 oxidized the ethanol
present in the medium and transformed it to Acetyl-
CoA, which is the main substrate of the PHB
pathway (Obruca et al, 2011). As it is shown in
figure 4, also during these oxidative reactions,
reduced coenzymes NAD(P)H are produced,
stimulating the flux of acetyl-CoA towards the PHB
biosynthetic pathway (Obruca et al, 2010c).
Table 2. Composition of the culture medium inside the flasks.
Milk Whey
(ml)
Buffer
(ml)
Solution of trace
elements (µl)
Ethanol
(ml)
Acetate
(g)
Control 189 21 210 0 0
Ethanol 1% 189 21 210 2.19 0
Ethanol 3% 189 21 210 6.57 0
Acetate 1% 189 21 210 0 3.076
Acetate 3% 189 21 210 0 9.23
Oxygen 54 6 60 0 0
Figure 3. Percentage of PHB accumulation in Bacillus megaterium TRQ8, growing under different compositions of culture
media.

Ruiz et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 24-31, 2017
29
Reduced coenzymes suppress the TCA cycle,
furthermore, it has been proven that the
accumulation of reduced coenzymes during ethanol
oxidation stimulates the activity of the 3-
ketothiolase and acetoacetyl-CoA reductase
enzymes from the PHB pathway resulting in an
overproduction of the polymer (Figure 4) (Oeding
and Schlegen, 1973). Hence, less free CoA, which
inhibits PHB biosynthesis, is formed as a result of
TCA cycle inhibition and instead it is used to build
acetyl-CoA (Obruca et al, 2010b). For this reason
the concentration of NAD(P)H is considered to be
the major regulatory factor determining the flux of
acetyl-CoA to either TCA cycle or PHB Pathway
(Obruca et al, 2010c).
However, the production of PHB diminished with
3% of ethanol in the culture medium, which
suggests the toxic effect of high ethanol
concentration for the strain, affecting its growth,
viability, and metabolism, due to ethanol-induced
leakage of the plasma membrane (Ingram, 1990).
Ethanol causes an increase in membrane fluidity
and can alter its integrity because it interacts with
membrane proteins, causing conformational
changes and thereby influencing their function
(Tóth et al, 2014). For this reason, it is possible that
a concentration of 3% of ethanol in the medium
killed a large number of bacteria, and therefore
caused a decrease in the synthesis of PHB.
On the other hand, the culture media containing
sodium acetate did not show higher production. It is
known that a large number of bacteria use acetate
for the synthesis of lipids and other compounds
(Guifang et al, 2002), therefore, there is a high
probability that B. megaterium TRQ8 has used the
sodium acetate available in the culture medium to
produce lipids instead of PHB (Fei et al, 2016),
since the percentage of accumulation at both
concentrations were lower in comparison with the
control (Figure 3).
The production of PHB was lower in the flask with
a higher amount of oxygen because the carbon flux
was bigger towards the Krebs cycle than the
fermentation pathway (Nath et al, 2008). A limited
amount of dissolved oxygen concentration increases
the fluxes of acetyl CoA towards PHB. The
shortage of oxygen prevents the electron transport
chain to work due to the absence of an electron
acceptor, consequently, an alternative pathway
involving the production of an organic compound,
in this case, PHB is promoted to regenerate the
electron carrier NAD+ (Baron, 1996). Therefore, a
limited dissolved amount of oxygen increases the
flux of acetyl-CoA, the common precursor of both
polyhydroxybutyrate and TCA pathways, towards
PHB accumulation instead of TCA cycle (Nath et
al, 2008).
Conclusions
Bacillus megaterium TRQ8 is able to produce high
quantities of PHB under exogenous stress
conditions, such as a culture medium containing 1%
of ethanol. Thus, it is possible to increase the yield
of PHB synthesis by adding a volume of 1% of
ethanol to the culture medium for this strain. The
Figure 4. Metabolic pathway of ethanol oxidation and PHB fermentation.

Ruiz et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 24-31, 2017
30
other media did not have the expected results due to
various factors like alternative metabolic pathways
or inhibition of the development and death of the
microbial population. Also the amount of available
oxygen is another important factor to take into
account, since the PHB production is stimulated by
oxygen limitation.
However the high cost of production is the main
drawback of this kind of plastics and has prevented
the PHB from being used in different items, thus the
reduction of the manufacturing costs is the main
aspect to be improved.
The application of an external stress, like ethanol, is
a promising strategy to enhance the production of
PHB from cheese whey using B. megaterium.
Nevertheless many other strategies can be applied in
order to obtain better results. For instance, the
utilization of a recombinant bacterium with better
growth characteristics will require less time to
produce the polymer. Moreover, the deletion of
other metabolic pathways (fermentation) and the
overexpression of the 3-ketothiolase enzyme can
redirect the carbon flux towards the desired pathway
and hence increase the yield of synthesis thereby
reducing the costs of production to obtain similar
values to those of synthetic polymers.
Acknowledgements
We express our thanks to Guillermo A. Ocegueda
Pintado and Marysol Ramírez Santacruz for their
suggestions and support, which improved the
experimental design.
References
Andrady, A. and Neal, M. A., 2009. Applications and societal
benefits of plastics. Philosophical Transactions of the Royal
Society B: Biological Sciences, 364(1526): 1977-1984. Babruwad, P., Prabhu, S., Upadhyaya, K. and Hungund, B.,
2015. Production and characterization of thermostable
polyhydroxybutyrate from Bacillus cereus PW3A. Biochem Tech, 3: 990-995.
Baron, S. (1996). Medical Microbiology. Galveston, USA:
Elvesier. Bell, D., Hoare, M. and Dunnill, P., 1983. The Formation of
Protein Precipitates and Their Centrifugal Recovery.
Torrington Place, London: University College London, T. Bovo, F., Tuanny, L., Eliana, R. and Fernandes, C., 2014. Ability
of a Lactobacillus rhamnosus strain cultured in milk whey
based medium to bind aflatoxin B1. 19/06/17, de Department of Food Engineering, College of Animal
Science and Food Engineering, University of São Paulo Sitio
web: http://www.scielo.br/scielo.php?script=sci_arttextypid=S010
1-20612014000300019 Campuzano, A. and Vasquez, H., 2013. Evaluation of the PHB
Production from Milk Whey. 28/1/17, de Universidad
Nacional de Colombia Web Site: http://uest.ntua.gr/tinos2015/proceedings/pdfs/campuzano_hi
guita.pdf
Cyras, V., Soledad, M., Mauri, A. and Vázquez, A., 2007. Biodegradable Double-Layer Films Based on Biological
Resources: Polyhydroxybutyrate and Cellulose. Wiley
InterScience, 106: 749-756. de los Santos Villalobos, S.; Parra Cota, F.I.; Herrera Sepúlveda,
A.; Valenzuela Aragón, B.; Estrada Mora, J.C. 2017.
COLMENA: Colección de Microorganismos Edáficos y
Endófitos Nativos, para contribuir a la seguridad alimentaria
nacional. Revista Mexicana de Ciencias Agrícolas. En
prensa. Fei, T., Cazeneuve, S., Wen, Z., Wu, L. and Wang, T., 2016.
Effective Recovery of Poly-b-Hydroxybutyrate (PHB)
Biopolymer from Cupriavidus necator Using a Novel and Environmentally Friendly Solvent System. Food Science and
Human Nutrition, 4: 678-685.
Forbes, B., Sahm, D. and Weissfeld, A., 2009. Diagnóstico Microbiológico. Bueno Aires, Argentina: Médica
Panamericana.
Gautam, S, 2009. Biodegradable Plastics Impact on Environment. 04/02/17, de Ministry of Environment y
Forests Government of India Web Site:
http://cpcb.nic.in/upload/NewItems/NewItem_150_PlasticsWaste.pdf
Getachew, A. and Woldesenbet, F., 2016. Production of
biodegradable plastic by polyhydroxybutyrate (PHB) accumulating bacteria using low cost agricultural waste
material. BMC Research Notes, 9: 1-9.
González, Y., Meza, J., González, O. and Córdova, J., 2013. Síntesis y biodegradación de polihidroxialcanoatos: Plásticos
de origen microbiano. Revista Internacional de Contamaminación Ambiental, 29(1): 77-115.
Guifang, W., Tian, B., Zhongyao, S. and Qingyu, W., 2002.
Sodium Acetate Stimulates PHB Biosynthesis in Synechocystis sp. PCC 6803*. Tsinghua Science and
Technology, 7: 435 - 438
Heinrich, D., Madkour, M., Al-Ghamdi, M., Shabbaj, I. and Steinbüchel, A., 2012. Large scale extraction of poly (3-
hydroxybutyrate) from Ralstonia eutropha H16 using sodium
hypochlorite. 09/02/17, de NCBI Web Site: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3564783/
Ingram, L., 1990. Ethanol tolerance in bacteria. Critical Reviews
in Biotechnology, 9: 305-319. Lambert S., 2013. Environmental Risk of Polymers and their
Degradation Products. Submitted for the degree of Doctor of
Philosophy. University of York Environment Department. Lemos, A. and Mina, A., 2015. Polyhydroxyalkanoates (PHA’s)
produced by bacteria its potential application to industrial
level. Informador Técnico. 79(1): 93-101. Luef, K., Stelzer, F. and Wiesbrockb, F., 2015.
Poly(hydroxyalkanoates in Medical Applications. Chemistry
and Technology of Materials, 29 (2): 287-297. North, E. J. and Halden, R. U., 2013. Plastics and Environmental
Health. The Road Ahead. Reviews on Environmental Health,
28(1): 1-8. Martínez, G. and Loperena, L., 2013. Producción de plásticos
biodegradables en bacterias. Thesis for Biotechnology
Engineering. Facultad de ciencias Universidad de la República.

Ruiz et al. / Revista Latinoamericana de Recursos Naturales 13 (1): 24-31, 2017
31
Myint, B., Champakalakshmi, R., Kishore, M., Sing, C. and Sakharkar, K., 2012. Biodegradation of Low Density
Polythene (LDPE) by Pseudomonas Species. Association of
Microbiologists of India, 52(3): 411-419. Mikkili, I., Karlapudi, A., Venkateswarulu, T., Babu, J., Nath, S.
and Kodali, V., 2014. Isolation, Screening and Extraction of
Polyhydroxybutyrate (PHB) producing bacteria from Sewage sample. International Journal of PharmTech
Research, 6: 850-857
Naranjo, J., 2010. Producción de polihidroxibutirato a partir de residuos agroindustriales. Thesis for Magister in Chemical
Engineering (20 - 40). Manizales, Colombia.
Nath, A., Dixit, M., Bandiya, A., Chavda, S. and Desai, A., 2008.
Enhanced PHB production and scale up studies using cheese
whey in fed batch culture of Methylobacterium sp. ZP24.
Bioresource Technology, 99: 5749-5755. Obruca, S., Marova, I., Svoboda, Z. and Mikulikova, R., 2010.
Use of Controlled Exogenous Stress for Improvement of
Poly(3-hydroxybutyrate) Production in Cupriavidus necator. Folia Microbiology, 55 (1): 17-22.
Obruca, S., Marova, I., Stankova, M., Mravcova, L. and
Svoboda, Z., 2010. Effect of ethanol and hydrogen peroxide on poly(3-hydroxybutyrate) biosynthetic pathway in
Cupriavidus necator H16. World Journal of Microbiology
and Biotechnology, 26:1261-1267. Obruca, S., Marova, I., Melusova, S. and Mravcova, L., 2011.
Production of polyhydroxyalkanoates from cheese whey
employing Bacillus megaterium CCM 2037. Springer-Verlag, 61:947-953
Oeding, V. and Schlegel, H., 1973. β-Ketothiolase from
Hydrogenomonas eutropha H16 and its significance in the regulation of poly-β-hydroxybutyrate metabolism.
Biochemical Journal, 134(1): 239-248.
Peña, C., Castillo, T., García, A., Millán, M. and Segura, D., 2014. Biotechnological strategies to improve production of
microbial poly-(3-hydroxybutyrate): a review of recent research work. Microbial Biotechnology, 7(4): 278-293.
Rivera, L. and Nevárez, G., 2009. Economic carbon sources for
bacterial bioplastic production. Tecnociencia Chihuahua. Vol., III (2): 58-63.
Seoane, I., Manfredi, L. and Cyras, V., 2013. Properties and
processing relationship of polyhydroxybutyrate and cellulose biocomposites. Procedia Materials Science, 8: 808-813.
Serrano, J., 2010. Polyhydroxyalkanoates (PHAs) polymers
produced by microorganisms. A solution to environmental pollution. Teoría y praxis investigativa, 5 (2): 79-84.
Singh, R., 2015. International Journal of Pharmaceutical Research | Apr-Jun 2015 | Vol 7 | Issue 2 | 17 Review Article
Polyhydroxybutyrate (PHB): Biodegradable, Bioplastics
Produced by Microorganisms. International Journal of Pharmaceutical Research, 7: 17-23.
Stahlhut, R. W., Welshons, W. V., and Swan, S. H., 2009.
Bisphenol A Data in NHANES Suggest Longer than Expected Half-Life, Substantial Nonfood Exposure, or Both.
Environmental Health Perspectives, 117(5): 784-789.
Talsness, C. E., Andrade, A. J. M., Kuriyama, S. N., Taylor, J. A., and vom Saal, F. S., 2009. Components of plastic:
experimental studies in animals and relevance for human
health. Philosophical Transactions of the Royal Society B:
Biological Sciences, 364(1526): 2079-2096.
Thompson, R. C., Swan, S. H., Moore, C. J., and vom Saal, F. S,
2009. Our plastic age. Philosophical Transactions of the Royal Society B: Biological Sciences, 364 (1526): 1973-
1976.
Thompson, R. C., Moore, C. J., vom Saal, F. S., and Swan, S. H, 2009. Plastics, the environment and human health: current
consensus and future trends. Philosophical Transactions of
the Royal Society B: Biological Sciences, 364 (1526): 2153-2166.
Tokiwa, Y., Calabia, B. P., Ugwu, C. U. and Aiba, S., 2009.
Biodegradability of Plastics. International Journal of Molecular Sciences, 10(9): 3722-3742.
Tóth, M. E., Vígh, L. and Sántha, M, 2014. Alcohol stress,
membranes, and chaperones. Cell Stress y Chaperones, 19(3): 299-309.
Valencia, E. and Ramírez, M., 2009. La industria de la leche y la
contaminación del agua. Elementos. (73), (27-31). Puebla, México.
Vary, PS., Biedendieck, R., Fuerch, T., Meinhardt, F., Rohde,
M., Deckwer, W. and Jahn, D., 2007. Bacillus megaterium from simple soil bacterium to industrial protein production
host. Applied Microbiology and Biotechnology, 76: 957-967.
Wang, L., Zhu, W., Wang, X., Chen, X., Chen, G. and Xu, K.,
2007. Processability Modifications of Poly (3-hydroxybutyrate) by Plasticizing, Blending, and Stabilizing.
Journal of Applied Polymer Science, 107: 166-173.
Webb, H., Arnott, J., Crawford, R. and Ivanova, E., 2013. Plastic Degradation and Its Environmental Implications with
Special Reference to Poly(ethylene terephthalate). Polymers,
5: 1-18.

Revista Latinoamericana de Recursos Naturales© Una revista multidisciplinar para el conocimiento científico de los recursos naturales en Latinoamérica
Información de la revista
La Revista Latinoamericana de Recursos Naturales©
publica semestralmente contribuciones originales
en cualquier campo relacionado con el conocimiento científico, la tecnología, la gestión y el aprovechamiento sostenible de los recursos naturales de Latinoamérica. La Revista Latinoamericana de Recursos Naturales publica tanto estudios multidisciplinarios como artículos monodisciplinarios que son de interés para cualquier disciplina relacionada con los recursos naturales.
MÁS INFORMACIÓN EN: http://www.itson.mx/publicaciones/rlrn/Paginas/informacion2.aspx
Información para autores
La Revista Latinoamericana de Recursos Naturales©
publica trabajos originales en cualquier campo
relacionado con el agua, el suelo, las plantas y los recursos agropecuarios. En particular, se reciben
trabajos de los ámbitos de la Hidrología, Meteorología, Biología, Biotecnología, Ecología, Geología,
Microbiología, Edafología, Geomorfología, Agrobiología, Química y Recursos Agropecuarios, con énfasis
en la ciencia básica, la aplicada y la generación de tecnología. Igualmente, será bienvenido cualquier
estudio que verse sobre la gestión de cualquier recurso natural con énfasis en la conservación de los
ecosistemas y el desarrollo sostenible de la sociedad. Los estudios de revisión de algún tema deben
ser concertados primero con los editores, enviando un breve índice de los principales puntos a tratar.
Más información/sugerencias/comentarios: [email protected]
CONSULTE LA GUÍA DETALLADA EN: http://www.itson.mx/publicaciones/rlrn/Paginas/guia.aspx

Revista Latinoamericana de Recursos Naturales© Una revista multidisciplinar para el conocimiento científico de los recursos naturales en Latinoamérica
Contenido Volumen 13, Número 1
Consejo Editorial Prefacio
Evaluación de un sistema de recirculación para producción larvaria de Litopenaeus
vannamei, utilizando la macroalga Gracilaria vermiculophylla como organismo
biorremediador de efluentes / (Evaluation of a recirculation system for larval production of
Litopenaeus vannamei using the macroalgae Gracilaria vermiculophylla as an effluent
bioremediation organism) C.I. Leyva-Márquez F. Lares-Villa, L.R. Martínez-Córdova, J.C. Ibarra-
Gámez, R. Casillas-Hernández y A. Miranda-Baeza ............................................................................... 1
Estimación de rendimiento de variedades nativas de maíz en el estado de Tlaxcala /
(Yield estimation of native corn varieties in the State of Tlaxcala) A. Maria-Ramírez, V.H. Volke-
Haller y M.L. Guevara-Romero........................................................................................................... 8
Identificación de regiones genómicas asociadas a respuesta a la vacunación contra el
PRRS en cerdas reproductoras del sur de Sonora, México / (Identification of Genomic
Regions Associated to PRRS Vaccination Response in Replacement Gilts from South of Sonora,
Mexico) G. Luna-Nevárez. C.M. Aguilar-Trejo, R.I. Luna-Ramírez, R. Zamorano-Algandar, X. Zheng,
M.E. Enns, S. Speidel, M.G. Thomas y P. Luna-Nevárez .............................................................................. 15
Production of polyhydroxybutyrate from milk whey fermentation by Bacillus
megaterium TRQ8 / (Producción de polihidroxibutirato a partir de la fermentación de suero de
leche por Bacillus megaterium TRQ8) J. B. Ruiz Aguirre, L. Z. Oliver Gómez, S. de los Santos
Villalobos y M. L. Sánchez ................................................................................................................. 24
Copyright © 2017 Publicado por el Instituto Tecnológico de Sonora
Recommended

















