
UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO
DEPARTAMENTO DE BIOLOGIA
ÁREA DE BOTÂNICA
LABORATÓRIO DE FISIOLOGIA VEGETAL
RELATÓRIO FINAL DE ATIVIDADES
RECIFE, 2015

LETÍCIA SIQUEIRA WALTER
RELATÓRIO FINAL DE ATIVIDADES
RECIFE, 2015
Relatório final de atividades,
referente ao Programa Institucional
de Bolsa de Iniciação Científica da
UFRPE – PIBIC/UFRPE, no período de
agosto de 2014 a junho de 2015.

1. IDENTIFICAÇÃO
ALUNO: Letícia Siqueira Walter
CURSO: Bacharelado em Engenharia Florestal
PROGRAMA: Programa Institucional de Bolsas de Iniciação Científica
PIBIC/UFRPE
ORIENTADORA: Profa. Dra. Rejane Jurema Mansur Custódio Nogueira
DEPARTAMENTO/ÁREA: Biologia / Botânica / Laboratório de Fisiologia
Vegetal
2. TÍTULO DO PROJETO:
“Manejo de cultivo e interferência na fisiologia e produção de princípios ativos
em nim indiano.”
2.1.Título da Pesquisa
“Avaliação do crescimento, trocas gasosas, relações hídricas e teores de
pigmentos fotossintetizantes em mudas de nim indiano (Azadiractha indica A.
Juss) submetidas a diferentes níveis de salinidade”

3. RESUMO
De origem asiática o nim indiano (Azadirachta indica A. Juss) é pertencente da
família Meliaceae, e vem sendo utilizado como inseticida natural, na medicina
animal e humana, em reflorestamentos, como também na extração do óleo
presente nas sementes. Esta pesquisa tem como objetivo avaliar o
crescimento, as trocas gasosas, as relações hídricas e os teores de pigmentos
fotossintetizantes em mudas de nim indiano (Azadiractha indica A. Juss)
submetidas a diferentes níveis de salinidade. Foram utilizadas 6 plantas em
cada tratamento (0 mM; 50 mM; 100 mM e 150 mM de NaCl) totalizando 24
plantas, avaliadas aos 20, 40 e 60 dias após a diferenciação. Foram avaliados
o crescimento (diâmetro do caule, altura da planta, número de folha, matéria
seca e alocação de biomassa), trocas gasosas (fotossíntese, transpiração e
condutância estomática), relações hídricas (potencial hídrico foliar e teor
relativo de água) e pigmentos fotossintéticos (clorofila a, b, total e
carotenoides). Para a variável crescimento, apenas na última coleta os
tratamentos salinos mais severos apresentaram reduções em média de 13,05%
para o diâmetro do caule e 41,8% número de folhas. Para a produção de
mátéria seca só foi vista diferença signficativa na produção de matéria seca
total, com uma redução dos tratamentos salinos severos em relação ao
Controle. Para a alocação de biomassa houve uma alocação de 22,85% dos
tratamentos salinos em relação ao controle, e na alocação de biomassa do
caule foi observado um aumento na alocação dos tratamentos mais severos de
27% em relação ao tratamento Controle. Em relação as trocas gasosas foi
observado de maneira geral que as mudas foram afetadas com maior evidência
aos 60 dias de estresse. Já as relações hídricas foram afetadas com o passar
do tempo pelo acúmulo de sais, com os tratamentos mais severos (100 mM e
150 mM de NaCl) evidenciando as reduções a partir dos 40 dias de estresse.
Avaliando os pigmentos fotossintéticos foi observado uma redução nas
clorofilas a, porém as maiores reduções foram vistas nos pigmentos acessórios
(clorofila b e carotenoides) apenas na última coleta, após uma grande produção
de pigmentos aos 40 dias de estresse. Diante das respostas dadas pelas
mudas de nim indiano, pode-se concluir que esta espécie pode ser considerada
uma planta tolerante à salinidade, visto que mesmo com as altas

concentrações de sais no solo e pequenas mudanças fisiológicas, as mudas
após 60 dias de estresse foram evidenciadas apenas nos tratamentos mais
severos.

4. OBJETIVOS
4.1 Geral
• Avaliar o crescimento, as trocas gasosas, as relações hídricas e pigmentos
fotossintéticos em mudas de nim indiano (Azadirachta indica A. Juss)
submetidas a diferentes tratamentos salinos.
4.2 Objetivos específicos:
• Avaliar as variáveis de crescimento (altura da planta, diâmetro do caule,
número de folhas, produção de matéria seca e a alocação de biomassa para os
diversos órgãos) em mudas de nim indiano submetidas a diferentes níveis de
salinidade;
• Averiguar as trocas gasosas (fotossíntese, transpiração e condutância
estomática) em mudas de nim indiano submetidas a diferentes níveis de
salinidade;
• Verificar a influência da salinidade nas relações hídricas (potencial hídrico
foliar e o teor relativo de água) em mudas de nim indiano;
• Determinar os pigmentos fotossintéticos (clorofila a, clorofila b, clorofila total e
carotenoides) em mudas de nim indiano submetidas a diferentes tratamentos
salinos.

5. INTRODUÇÃO
5.1. CARACTERÍSTICAS E UTILIZAÇÃO DO NIM INDIANO
Com nome científico Azadirachta indica A. Juss, o nim indiano também é
conhecido como neem ou amargosa e pertence à família Meliaceae, assim
como o mogno, cedro, andiroba e cinamomo. Tem origem asiática, sendo
natural das regiões áridas da Índia, porém também é cultivado nos Estados
Unidos, Austrália e alguns países da África e América Central, no Brasil foi
introduzido no ano de 1986 pelo Instituto Agronômico do Paraná – IPAR, como
objetivo de pesquisar a ação inseticida do nim sendo atualmente encontrada
por toda a região Nordeste do Brasil. (MOREIRA et al, 2012; BITTENCOURT,
2006; NEVES, 2003; MARTINS, 2008). Segundo Neves e Carpanezzi (2008), o
Nordeste brasileiro é altamente rentável para produção de madeira para lenha,
por apresentar alto poder calorífico (ARAÚJO et al., 2000) e produção de frutos
em larga escala.
O neem, que na língua erudita Indiana (sânscrito) significa “curador”, a
mais de 2000 anos é utilizado na Índia como inseticida, nematicida, fungicida e
bactericida, além da fabricação de estátuas religiosas por sua madeira
apresentar boa trabalhabilidade. Além destas utilidades a espécie também vem
sendo utilizada na medicina animal e humana, na indústria de cosméticos (para
fabricação de shampoos, óleos para cabelo, tônico capilar e óleo para unhas),
na indústria madeireira (produzindo de carretas, cercas, ferramentas, móveis),
implementos agrícolas, em reflorestamentos de áreas degradadas pela
agricultura e sistemas agroflorestais, paisagismo, quebra-vento. Além da
utilização da torta de nim, subproduto da extração o óleo presente nas
sementes, que atua como fonte de nitrogênio, cálcio, sódio, potássio e
magnésio, e quando misturada aos fertilizantes reduz a perda de amônio
volatilizada pelas bactérias nitrificantes do solo (NEVES, 2003;
BITTENCOURT, 2006; NEVES, 2008; ALVES, 2008)
O principal componente responsável pela ação inseticida presente no
nim é a azadiractina, agindo contra mais de 300 espécies de insetos. Este
composto afeta o desenvolvimento dos insetos de diferentes formas, está
presente em todas as partes da planta, porém encontrado em maior

concentração sementes. Além da azadiractina, esta espécie apresenta outros
compostos como terpenóides (ARAÚJO et al, 2000; MARTINEZ, 2008).
A árvore de nim é resistente, de crescimento rápido e de grande porte,
com sua altura variando de 15 a 20 m, sua copa é frondosa, formada por uma
grande quantidade de folhas compostas imparipinadas, alternas organizadas
em aglomerados nos terminais de ramos e sem presença de estípulas. Suas
flores são pequenas, brancas, bixessuadas, dispostas em inflorescências
densas. Os frutos são apresentados na forma de bagas ovaladas, de cor
amarelada quando maduros, com tegumento branco que contém um óleo
dentro de uma ou raramente duas sementes (NEVES, 2006; MOSSINI &
KEMMELMEIER, 2005). A produção de frutos da espécie começa a partir do
terceiro ou quinto ano do plantio, com os meses de frutificação entre julho e
setembro e possível segunda florada entre novembro e janeiro. Seu sistema
radicular é composto de uma raiz pivotante, que pode chegar a 15 m de
profundidade, responsável pela absorção de água e nutrientes em grandes
profundidades (NEVES, 2013; MOSSINI & KEMMELMEIER, 2005). Neves
(2003), afirma que o nim pode ser produzido tanto por sementes, mudas,
cultura de tecidos ou produção por estaquia, porém este último não é viável
pois a raiz cresce de forma irregular e superficial, deixando a planta vulnerável
a queda por ventos muito fortes.
5.2. CARACTERIZAÇÃO DO SOLO E PLANTAS TOLERANTES À
SALINIDADE
No Brasil, as áreas com maior decorrência da salinidade estão
localizadas no Nordeste, onde as plantações necessitam de irrigação, sendo a
água de irrigação de má qualidade o caso mais clássico de acúmulo de sais na
superfície do solo. Porém a origem do solo, presença de matéria orgânica,
adubação e manejo do solo também influenciam na variação da quantidade de
sais acumulados (FERREIRA et al., 2010). Em regiões áridas e semiáridas
além do solo não apresentar boa drenagem e camadas impermeáveis, a baixa
pluviosidade e elevada evapotranspiração são fatores contribuintes da
concentração de sais solúveis na solução do solo (RIBEIRO, 2010). Holanda et
al. (2007) afirmam que das regiões áridas e semiáridas do mundo 7%,
correspondendo a 952,2 milhões de hectares, sofrem influência do acúmulo de

sais no solo. Este acúmulo de sais pode não só afetar a planta como também
alterar a estrutura dos solos, diminuindo a porosidade, permeabilidade e
fertilidade, além do risco de erosão e contaminação dos lençóis freáticos (TAIZ
& ZEIGER, 2006; RIBEIRO, 2010).
Solos afetados por sais são divididos em três grupos: 1) solos salinos -
ocorrem quando a alta concentração de sal provoca estresse osmótico nas
plantas; 2) solos sódicos - ocorre quando a relação de sódio trocável é alta; e
3) solos salino-sódico - ocorre os dois tipos anteriores simultaneamente
(FERREIRA et al., 2010). Por isso o estudo de espécies arbóreas tolerantes à
salinidade, é um importante passo para a recuperação de áreas degradadas já
que servem como cobertura vegetal para o solo e o sistema radicular profundo
melhora a permeabilidade e porosidade do solo, aumentando a evaporação
(HOLANDA et al., 2007).
Levando em consideração às respostas dadas pelas plantas em relação
às concentrações de sais presentes nos solos, as plantas são classificadas em
glicófitas e halófitas. As plantas glicófitas são classificadas como menos
tolerantes a sais, já as halófitas são espécies que se ajustam osmoticamente
para tolerar os efeitos salinos, sendo geralmente espécies nativas de solos
salinos com capacidade de completar seu ciclo nestes ambientes. Este último
grupo de plantas, aumentam a concentração de sais no seu interior para que
as raízes possam continuar absorvendo água do substrato (TAIZ & ZEIGER,
2006; RIBEIRO, 2010). Segundo Willadino e Camara (2010) as plantas
glicófitas reduzem o crescimento quando o teor de sal é maior que 10 mM
enquanto as halófitas suportam uma variação entre 50 e 500 mM de sal.
Souza (2013) afirma ser necessário a utilização de práticas de correção,
para que solos afetados por sais possam ser utilizados para produção agrícola
ou florestal, porém tais métodos geram altos custos e não resolvem de fato o
problema. Sendo uma alternativa a utilização de espécies tolerantes à
salinidade tanto para a recuperação destes solos como também para a
produção.

5.3. EFEITOS DO ESTRESSE SALINO EM PLANTAS
Segundo Taiz e Zeiger (2006) estresse é definido como "um fator
externo, que exerce uma influência desvantajosa sobre a planta", sendo
medido de acordo com a sobrevivência, produtividade agrícola e crescimento
geral da espécie, que é afetado pela toxidade gerada pela alta concentração de
íons na planta, além de apresentar dificuldades na absorção de água
(RIBEIRO, 2010). A capacidade de se adaptar ao estresse é responsabilidade
dos mecanismos de defesa da planta, que podem ser de escape ou de
tolerância. Os mecanismos de escape reduzem as atividades metabólicas,
fazendo com que as plantas cheguem a um estado de dormência, já os de
tolerância aumentam as atividades metabólicas quando a planta está
submetida aos níveis moderados de estresse, quando este estresse passa a
ser severo as atividades metabólicas baixam, fazendo com que a planta
suporte o estresse (WILLADINO & CAMARA, 2010).
Os efeitos da salinidade são resultados de dois componentes: osmótico
e iônico. O osmótico é resultado da elevada concentração de solutos na
solução do solo, fazendo com que a planta sofra com déficit hídrico através da
redução osmótica, já o iônico é resultado das altas concentrações de Na+ e Cl-,
que alteram as relações de K+/Na+ e outros nutrientes, provocando toxidez nos
tecidos vegetais. Esses efeitos osmóticos fazem com que as plantas
apresentem reações semelhantes às apresentadas quando submetidas a
déficit hídrico (SILVA, 2013; WILLADINO & CAMARA, 2010). Conseguindo se
ajustar osmoticamente, as plantas que crescem em solos salinos, como forma
de evitar a perda do turgor, geram um potencial hídrico mais baixo fazendo
com que a planta continue absorvendo água do solo. Quando o teor de sal está
em excesso a planta pode chegar a perder água para o solo, tal efeito é
resultado da plasmólise caracterizado como o movimento da água do meio
menos concentrado para o mais concentrado. Além da capacidade de se
ajustar osmoticamente, espécies submetidas a altos níveis de sais podem
também reduzir a área foliar ou perder folhas por abscisão (RIBEIRO, 2010;
TAIZ & ZEIGER, 2006; WILLADINO & CAMARA, 2010).
Uma redução significativa no crescimento da planta é o primeiro efeito
do estresse salino, onde a planta irá apresentar uma nova taxa de crescimento,

desta vez menor do que a taxa apresentada antes da imposição do estresse.
Além desta redução outras respostas fisiológicas são afetadas quando a planta
é submetida à salinidade como por exemplo o potencial hídrico, nutrição
mineral, fechamento estomático, eficiência fotossintética e alocação e utilização
do carbono (TÁVORA et al, 2001; WILLADINO & CAMARA, 2010). Na tabela
abaixo (Tabela 01), são apresentados sintomas do acúmulo de sais sobre
plantas tolerantes e sensíveis em relação ao crescimento.
Tabela 01. Respostas à salinidade em diferentes épocas. (Adaptação de Willadino & Camara,
2010).
Tempo Efeitos sobre crescimento de
plantas tolerantes
Efeito sobre plantas sensíveis
ao sal
Dias Redução da taxa de emissão de
novas folhas.
Injúria nas folhas mais velhas
(toxidez).
Semanas Redução na altura final e/ou do
número de folhas.
Morte das folhas mais velhas.
Meses Alteração no tempo de floração;
redução na produção de
sementes.
Morte das olhas mais jovens;
planta morre antes da
maturação das sementes.
Diante do exposto, é de grande importância conhecer os mecanismos
fisiológicos das plantas quando submetidos aos estresses salinos, para que
assim possam ser utilizadas da melhor forma as espécies para recuperação de
áreas degradadas por sais, como também para que possam ser utilizadas
como fonte de renda secundária dos produtores, com espécies de múltiplo uso.

6. MATERIAL E MÉTODOS
6.1. CONDIÇÕES EXPERIMENTAIS
O experimento foi conduzido na casa de vegetação do Laboratório de
Fisiologia Vegetal pertencente ao Departamento de Biologia da Universidade
Federal Rural de Pernambuco - UFRPE. Foram utilizadas sementes coletadas
de três árvores matrizes em um plantio no município de Itabaiana-PB,
pertencente à Cruanji Neem do Brasil, com sede localizada no município de
Timbaúba-PE.
A coleta dos frutos foi feita diretamente dos galhos de três matrizes,
utilizando um podão e uma lona (Figura 01). Após os frutos estarem
uniformemente amadurecidos, os mesmos foram despolpados com uma
peneira e água corrente para que as sementes fossem colocadas para secar
durante dois dias. Depois de secas, as sementes foram postas para germinar
em bandejas de polietileno contendo areia lavada para que elas estivessem
livres de qualquer tipo de nutrientes, sendo regada diariamente com água.
Figura 01. Coleta dos frutos de nim indiano.
Após a germinação, as mudas foram selecionadas quanto à sanidade,
altura, número de folhas e uniformidade e transferidas para vasos de polietileno
com capacidade para 2,5 Kg, preenchidos com areia lavada. Durante o período
de dois meses em que passaram aclimatando, as mudas foram regadas com
solução nutritiva ½ força de Hoagland e Arnon (1950) a cada dois dias,

intercalando com água. Além disso, foram mantidas com pequenos plásticos
cobrindo apenas o solo, para que fosse minimizada a perda de água por
evapotranspiração.
6.2. DELINEAMENTO EXPERIMENTAL
As plântulas foram transplantadas com 13,48 cm em média de
comprimento de raiz (medidos da ponta da raiz até a base da mesma) e 8,20
cm de comprimento da parte aérea (medidos da base da raiz até o último ponto
de inserção da folha).
Após o período de dois meses de aclimatação foram impostos os
tratamentos. Com delineamento inteiramente casualizado, os diferentes níveis
de salinidade foram: 0 mM (1,41 dS.m-1), 50 mM (6,33 dS.m-1), 100 mM (10,63
dS.m-1) e 150 mM (14,35 dS.m-1) de NaCl, com 6 repetições para cada
tratamento totalizando 24 plantas. Os tratamentos salinos foram preparados em
solução nutritiva ½ força e a manutenção dos tratamentos foram através da
aferição da condutividade elétrica da solução drenada dos vasos, feita
semanalmente com um condutivímetro portátil (Figura 02). Aos 20, 40 e 60 dias
foram realizadas coletas, a fim de se avaliar a influência do estresse salino nas
variáveis estudadas, sendo na última coleta o desmonte do experimento
Figura 02. Medição da condutividade elétrica da solução drenada dos vasos
Os parâmetros analisados no presente experimento foram crescimento
(diâmetro do caule, altura da planta, número de folha, matéria seca e alocação

de biomassa, razão raiz/parte aérea), trocas gasosas (fotossíntese,
transpiração e condutância estomática), relações hídricas (potencial hídrico
foliar e teor relativo de água) e pigmentos fotossintéticos (clorofila a, b, total e
carotenoides).
6.3. CRESCIMENTO (ALTURA, DIÂMETRO DO CAULE, NÚMERO DE
FOLHAS, ALOCAÇÃO DE BIOMASSA, RELAÇÃO RAIZ/PARTE AÉREA)
As avaliações de crescimento foram feitas semanalmente a partir do dia
do transplantio. Para o diâmetro do caule, foi utilizado um paquímetro digital, e
a medição foi feita a um centímetro acima do solo (marcado previamente), para
a altura da planta foi utilizada uma trena onde as plantas foram medidas a um
centímetro do solo até a inserção da última folha, e a contagem do número de
folhas foi realizada por simples contagem (Figura 03). Para a normalização, os
dados de número de folhas foram transformados para X+0,5 (ZAR, 1999).
Figura 03. Medição do diâmetro do caule (A), altura da planta (B) e contagem do número de
folhas (C).
Ao final do período experimental, as plantas foram separadas em seus
respectivos órgãos: folhas, caule e raiz, que foram pesados e em seguida
colocados em sacos de papel e levados para a estufa de aeração forçada a 65°
C até atingir peso constante para obtenção do peso da matéria seca dos
referidos órgãos. De posse dos dados da matéria seca da folha (MSF), matéria
seca do caule (MSC) e matéria seca da raiz (MSR), serão calculados a
alocação de biomassa para as folhas (ABF), caule (ABC) e para as raízes
(ABR) e a razão raiz/parte aérea (R/Pa), segundo Benincasa (2003).
A B C A

6.4. TROCAS GASOSAS (FOTOSSÍNTESE, TRANSPIRAÇÃO E
CONDUTÂNCIA ESTOMÁTICA)
Antes da diferenciação foi feito um curso diário das trocas gasosas nas
mudas de nim, para que pudesse encontrar o horário de maior abertura
estomática com maior valor de fotossíntese. O curso diário foi feito no decorrer
de um dia, utilizando o Analisador de Gás à Infravermelho (IRGA, ADC, modelo
LCipro) iniciando as análises às 8 h da manhã até ás 16 h com intervalo de 2 h
entre as análises. Após encontrar o horário mais adequado para a realização
das análises de trocas gasosas, entre 10 h e 12 h, foram feitas análise das
trocas gasosas (Figura 04) nos dias das coletas, utilizando a terceira folha
completamente expandida (folha +3).
Figura 04. Análise das trocas gasosas com o IRGA.
6.5. RELAÇÕES HÍDRICAS (POTENCIAL HÍDRICO FOLIAR E TEOR
RELATIVO DE ÁGUA)
Para avaliação das relações hídricas, foi coletada uma folha localizada
no terço médio superior sendo cortada rente ao pecíolo e logo em seguida
envolta com papel filme e colocada em recipiente refrigerado (Figura 05), para
que pudessem ser feitas as medidas utilizando a câmara de pressão de
Scholander modelo 3035 (Soil Moisture Equipment Corp, Santa Bárbara, CA,
USA), seguindo a metodologia de Scholander et al. (1965).

Figura 05. Coleta (A) e análise da folha para potencial hídrico foliar (B) e TRA (C).
Na mesma folha utilizada nas medições do potencial hídrico foliar foi
feita a determinação do teor relativo de água (TRA), retirando-se dois folíolos
da base da folha, antes da medição do potencial, onde foi retirada a nervura
central e foram pesados para obtenção do peso da matéria fresca (PMF) e em
seguida postos em placa de Petri contendo 10 mL de água destilada, onde
permaneceram por 24 h, num ambiente refrigerado. Após esse período, os
folíolos foram retirados, cuidadosamente secos para retirar o excesso de água,
e pesados em seguida para que fosse obtido o peso da matéria túrgida (PMT).
Os folíolos foram então colocados em sacos de papel devidamente
identificados e levados para a estufa de aeração forçada a 65° C até atingir
peso constante, para que fosse obtido o peso da matéria seca (PMS). A partir
dos dados do peso da matéria fresca (PMF), matéria túrgida (PMT) e matéria
seca (PMS), pôde-se calcular o TRA das folhas utilizando a seguinte fórmula
descrita na metodologia de Cairo (1995).
6.6. PIGMENTOS FOTOSSINTÉTICOS (CLOROFILA a, CLOROFILA b,
CLOROFILA TOTAL E CAROTENOIDES)
TRA = (PMF – PMS) x 100
(PMT – PMS)
A B C

Para análise dos teores dos pigmentos fotossintéticos foi pesado 0,1 g
das folhas, retirando-se as nervuras, em seguida foram picotadas em pequenos
pedaços e colocados em tubos de ensaio rosqueáveis, contendo 10 mL de
álcool P.A. e envoltos em papel alumínio, para evitar o contato com a luz,
seguindo a metodologia de Lichtenthaler e Buschmann (2001) com
modificações. As leituras do material foram feitas em espectrofotômetro modelo
SP-220, nos comprimentos de onda 664 nm para clorofila a, 649 nm para
clorofila b e 470 nm para carotenoides. Após serem feitas as leituras, os
valores foram colocados nas seguintes fórmulas:
6.7. ANÁLISE ESTATÍSTICA
De posse dos dados, os mesmos foram submetidos à análise de
variância (ANOVA) e as médias comparadas entre si pelo teste de Tukey ao
nível 5% de probabilidade. O software utilizado foi o Assistat, versão 7.7 beta.
Chla = 13,36 x A664 - 5,19 x A649
Chlb = 27,43 x A649 - 8,12 x A664
Chltotal = Chla + Chlb
Carotenóides = (1000 x A470 – 2,13 x Chla – 97,64 x Chlb)/ 209

7. RESULTADOS E DISCUSSÃO
7.1. CRESCIMENTO (ALTURA DA PLANTA, NÚMERO DE FOLHAS,
DIÂMETRO DO CAULE, ALOCAÇÃO DE BIOMASSA E RAZÃO RAIZ/PARTE
AÉREA)
De maneira geral, o crescimento do nim indiano foi afetado pelo acúmulo
de sais no solo apenas na última semana de avaliação, 60 dias após da
imposição do estresse. Porém a variável altura não apresentou diferenças
significativas em nenhum dos tratamentos como pode ser observado na figura
06 A.
Já em relação à variável diâmetro do caule (Figura 06 B), observou-se
que apenas na última semana do experimento os tratamentos 100 mM e 150
mM de NaCl diferiram estatisticamente do tratamento controle, apresentando
reduções de 11,2% e 14,9% respectivamente. Para a taxa de produção de
novas folhas, foi observado que o tratamento 150 mM de NaCl apresentou o
maior decréscimo (41,8%) em relação ao tratamento controle na última semana
de avaliação, porém também foi observado que mesmo sem diferença
estatística do tratamento controle o tratamento mais severo apresentou a partir
da semana em que o experimento foi diferenciado, indicada pela seta, sempre
os menores valores para o número de folhas. Os tratamentos 50 mM e 100 mM
de NaCl também diminuíram a produção de folhas novas, porém não diferiram
do tratamento controle (Figura 06 C).
A

Figura 06. Avaliação da altura (A), diâmetro do caule (B) e número de folhas (C) da planta em
mudas de nim indiano submetidas a diferentes níveis de salinidade nos tratamentos Controle
(CR), 50 mM, 100 mM e 150 mM de NaCl referente às semanas de duração do experimento. A
seta representa a diferenciação do experimento. Letras iguais não diferem entre os tratamentos
pelo teste de Tukey ao nível 5% de probabilidade.
Freire et al. (2010) encontraram resultados que diferem dos encontrados
na presente pesquisa. Quando avaliaram mudas de nim e cinamomo observou
que mudas de nim após 45 dias de submissão ao estresse salino mais severo,
com condutividade elétrica igual a 10,45 dS.m-1, apresentaram 20% de redução
em relação ao tratamento controle (0,49 dS.m-1), e o cinamomo reduziu a altura
em apenas 12,5% em relação ao controle. Assim como Diniz et al. (2013)
também encontraram reduções nas alturas de mudas de nim indiano cultivadas
B
C

com e sem biofertilizante bovino e regadas com água não salina (0,5 dS.m-1) e
com água salina (6,0 dS.m-1), sendo este último o tratamento mais severo, tais
resultados diferem dos encontrados na presente pesquisa. Mesquita et al.
(2012) também encontraram reduções no crescimento em alturas de mudas de
nim indiano, porém nas mudas que receberam biofertilizante o efeito do sal foi
menor do que nas mudas sem o biofertilizante.
Holanda et al. (2007) concluíram em sua pesquisa, que o nim é uma
espécie tolerante à salinidade quando compararam espécies nativas da
caatinga (angico, aroeira, cumaru e gliricídia) com o nim indiano. Observaram
que esta última espécie apresentou melhores resultados, em relação às demais
espécies, para altura em solos salinos e solos salinos com gesso. O mesmo
comportamento foi apresentado pelos autores para a variável diâmetro do
caule nas mudas de nim indiano.
Nunes et al. (2012) afirmam que o nim reduziu o crescimento do
diâmetro do caule em 41,6% quando comparou o tratamento mais severo (4,0
dS.m-1) com o controle (0,5 dS.m-1). Ainda segundo os mesmos autores, esta
espécie é afetada pela ação de sais nos estágios iniciais se irrigado com água
salina superior a 0,5 dS.m-1, tal afirmação difere dos resultados da presente
pesquisa, visto que o acúmulo de sais só começou a afetar o nim a partir de 60
dias após a imposição do estresse salino e apenas nos tratamentos mais
severos. Já Costa et al. (2013) não encontraram diferenças significativas nos
seus resultados de diâmetro do caule e comprimento do ramo quando avaliou o
crescimento inicial do nim em solo salino e não salino.
Avaliando o crescimento inicial de mudas de nim indiano submetidas à
diferentes níveis de salinidade sem a utilização de biofertilizante, Nunes et al.
(2012) observaram uma redução de 49% na emissão de novas folhas quando
comparou o tratamento mais severo com o de menor salinidade. Quando Silva
(2015) avaliou mudas de nim indiano sob ação de dois sais (NaCl e CaCl2) em
diferentes concentrações, o número de folhas só começou a ser afetado pelo
CaCl2 aos 49 dias após a imposição do estresse, apresentando redução de
26% entre o tratamento mais severo e o moderado, porém aos 56 dias essa
redução deixou de ser significativa. Os tratamentos com NaCl não foram
afetados até o final do experimento. Tais resultados corroboram com a
presente pesquisa e podem levar a conclusão de que o nim é uma espécie
tolerante à salinidade em estágios iniciais, quando avaliamos o crescimento
inicial.
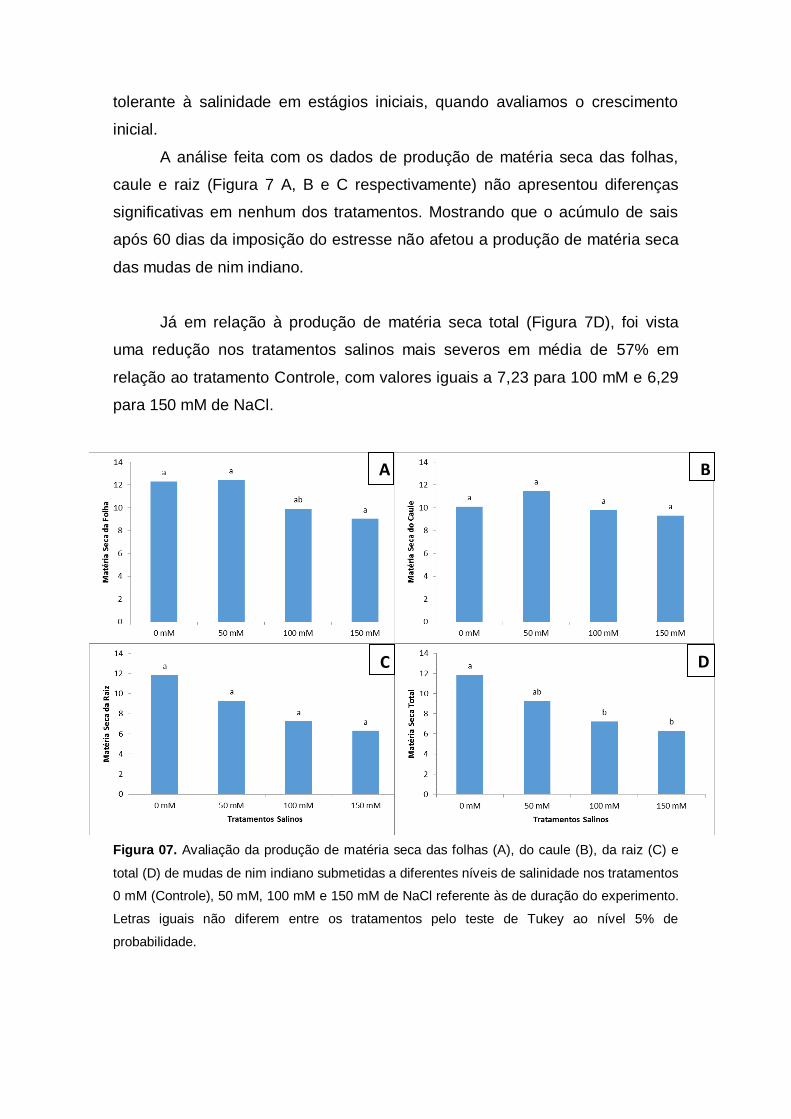
A análise feita com os dados de produção de matéria seca das folhas,
caule e raiz (Figura 7 A, B e C respectivamente) não apresentou diferenças
significativas em nenhum dos tratamentos. Mostrando que o acúmulo de sais
após 60 dias da imposição do estresse não afetou a produção de matéria seca
das mudas de nim indiano.
Já em relação à produção de matéria seca total (Figura 7D), foi vista
uma redução nos tratamentos salinos mais severos em média de 57% em
relação ao tratamento Controle, com valores iguais a 7,23 para 100 mM e 6,29
para 150 mM de NaCl.
Figura 07. Avaliação da produção de matéria seca das folhas (A), do caule (B), da raiz (C) e
total (D) de mudas de nim indiano submetidas a diferentes níveis de salinidade nos tratamentos
0 mM (Controle), 50 mM, 100 mM e 150 mM de NaCl referente às de duração do experimento.
Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
A B
C D
Quando Martins (2010) avaliou mudas de nim indiano submetidas a
déficit hídrico concluiu que as mudas com capacidade de pote de 80%
apresentaram melhor resultado para produção de matéria seca das folhas e do
caule. Já na produção de matéria seca da raiz os tratamentos controle e 80 %
da capacidade de pote apresentaram os melhores resultados. Na produção de
matéria seca total, os tratamentos sem rega e reirrigados apresentaram um
aumento na produção de matéria seca.
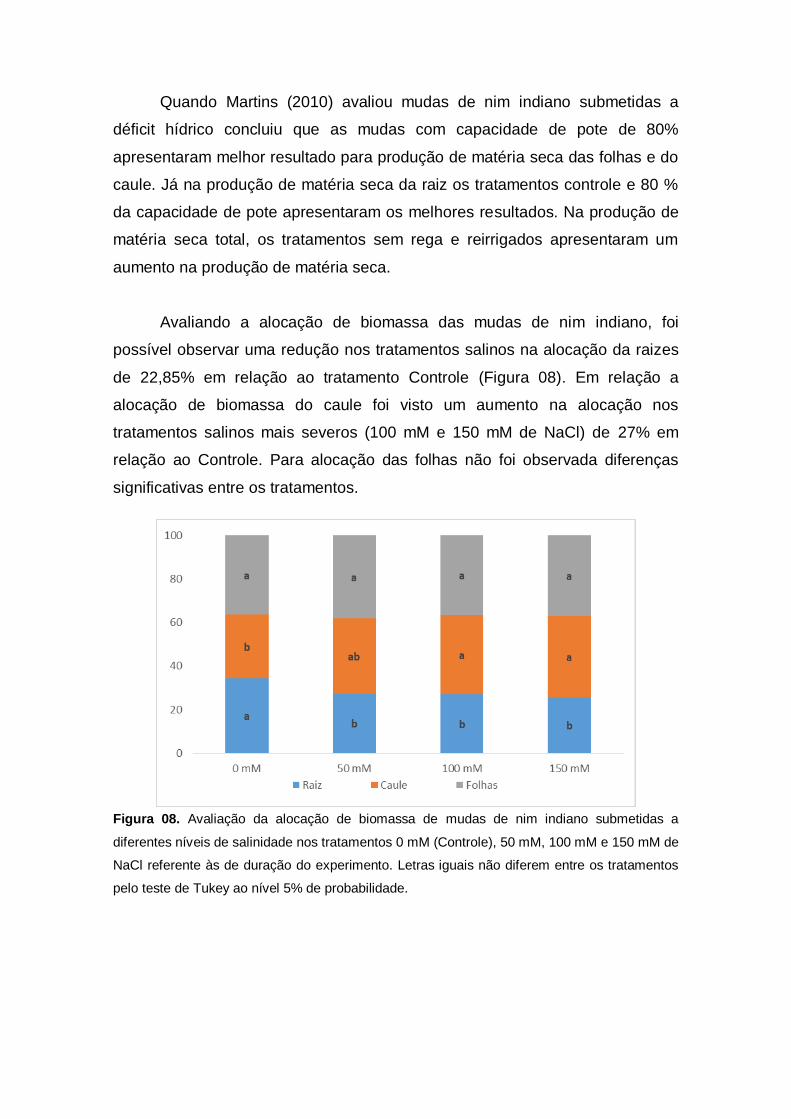
Avaliando a alocação de biomassa das mudas de nim indiano, foi
possível observar uma redução nos tratamentos salinos na alocação da raizes
de 22,85% em relação ao tratamento Controle (Figura 08). Em relação a
alocação de biomassa do caule foi visto um aumento na alocação nos
tratamentos salinos mais severos (100 mM e 150 mM de NaCl) de 27% em
relação ao Controle. Para alocação das folhas não foi observada diferenças
significativas entre os tratamentos.
Figura 08. Avaliação da alocação de biomassa de mudas de nim indiano submetidas a
diferentes níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de
NaCl referente às de duração do experimento. Letras iguais não diferem entre os tratamentos
pelo teste de Tukey ao nível 5% de probabilidade.

A razão raiz/parte aérea foi observada uma redução nas médias de
31,3% dos tratamentos salinos em relação ao tratamento Controle, como pode
ser observado na tabela 01.
Tabela 01. Razão raiz/parte aérea (R/PA) de mudas de nim indiano (Azadirachta indica),
submetidas a diferentes níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM
e 150 mM de NaCl referente às de duração do experimento. Letras iguais não diferem entre os
tratamentos pelo teste de Tukey ao nível 5% de probabilidade.
Tratamentos Raiz/Parte aérea
0 mM 0,53359 a
50 mM 0,38036 b
100 mM 0,37285 b
150 mM 0,34279 b
Mudas de nim indiano submetidas a diferentes regimes hídricos, quando
avaliadas por Walter et al. (2014) não foram vistas diferenças significativas na
razão raiz/parte aérea.
7.2 TROCAS GASOSAS (FOTOSSÍNTESE LÍQUIDA, TRANSPIRAÇÃO E
CONDUTÂNCIA ESTOMÁTICA)
Observa-se que, de modo geral, as trocas gasosas foram afetadas pelo
estresse salino, sendo este comportamento evidenciado aos 60 dias de
imposição do estresse.
Após a análise de dados das três coletas (20, 40 e 60 dias), pode ser
observado uma redução gradativa da fotossíntese líquida, com a última coleta
apresentando valores médios iguais a 7,33, 6,23, 6,17, 4,45 μmolCO2.m-2.s-1
respectivamente para os tratamentos 0 mM, 50 mM, 100 mM e 150 mM de
NaCl. Como pode ser observado na figura 09, os tratamentos mais severos
começaram a diminuir sua taxa fotossintética aos 40 dias de estresse. Tais
resultados mostram que o acúmulo de sais, 40 dias após a imposição do
estresse já começava a afetar a fotossíntese das mudas.

Figura 09. Avaliação da fotossíntese líquida (A) em mudas de nim indiano submetidas a
diferentes níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de
NaCl referente à coleta com 20, 40 e 60 dias após a imposição do estresse. Onde letras
minúsculas correspondem aos tratamentos e letras maiúsculas correspondem às coletas.
Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
Quando Silva (2015) avaliou mudas de nim indiano submetidas a
diferentes concentrações de NaCl e CaCl2, concluiu que a taxa fotossintética
sofreu reduções gradativas durante o período experimental. Ainda segundo a
mesma autora, na coleta realizada após 60 dias da imposição do estresse o
tratamento de 100 mM sofreu reduções de 48% e 41% para os sais NaCl e
CaCl2 respectivamente.
Mudas de Azadirachta indica quando avaliada por Lima et. al (2014) em
solo com e sem encharcamento e com baixa (2,0 dS.m-1) e alta (12,0 dS.m-1)
salinidade, apresentaram reduções menores, de 3 % e 10,7%, nos valores de
fotossíntese líquida nos tratamentos com alta salinidade e sem encharcamento
e alta salinidade com encharcamento respectivamente. Os autores destacaram
ainda uma leve recuperação após 15 dias sem encharcamento das mudas de
nim. Silva et al. (2012) avaliando as trocas gasosas em mudas de aroeira e
pau-mocó, observaram redução na fotossíntese com o aumento do teor salino
do solo, apresentando reduções de 24,3% para aroeira e 43,3% para o pau-
mocó comparando o maior nível salino (8,4 dS.m-1) e o menor (1,2 dS.m-1).
Os resultados encontrados pelos autores acima colaboram com os resultados
da presente pesquisa, mostrando sempre a redução gradativa da fotossíntese
ao longo do período experimental.
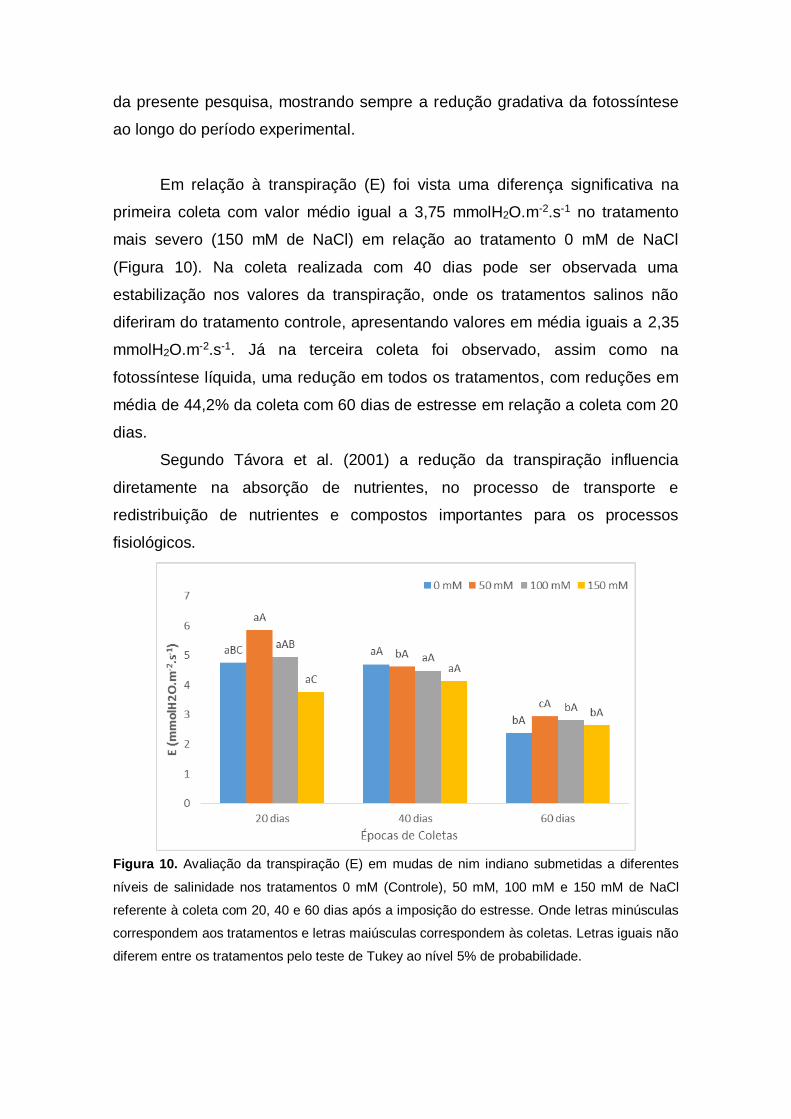
Em relação à transpiração (E) foi vista uma diferença significativa na
primeira coleta com valor médio igual a 3,75 mmolH2O.m-2.s-1 no tratamento
mais severo (150 mM de NaCl) em relação ao tratamento 0 mM de NaCl
(Figura 10). Na coleta realizada com 40 dias pode ser observada uma
estabilização nos valores da transpiração, onde os tratamentos salinos não
diferiram do tratamento controle, apresentando valores em média iguais a 2,35
mmolH2O.m-2.s-1. Já na terceira coleta foi observado, assim como na
fotossíntese líquida, uma redução em todos os tratamentos, com reduções em
média de 44,2% da coleta com 60 dias de estresse em relação a coleta com 20
dias.
Segundo Távora et al. (2001) a redução da transpiração influencia
diretamente na absorção de nutrientes, no processo de transporte e
redistribuição de nutrientes e compostos importantes para os processos
fisiológicos.
Figura 10. Avaliação da transpiração (E) em mudas de nim indiano submetidas a diferentes
níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de NaCl
referente à coleta com 20, 40 e 60 dias após a imposição do estresse. Onde letras minúsculas
correspondem aos tratamentos e letras maiúsculas correspondem às coletas. Letras iguais não
diferem entre os tratamentos pelo teste de Tukey ao nível 5% de probabilidade.

Silva et al. (2012) observaram uma redução na taxa de transpiração em
mudas de pau-mocó e aroeira quando os níveis de sal aumentavam. As mudas
apresentaram reduções média de 31% no tratamento mais salino (8,4 dS.m-1),
em relação ao tratamento com condutividade de 1,2 dS.m-1, porém foi a
aroeira quem apresentou a menor redução quando foi comparado o tratamento
mais severo com o de menor salinidade. Já Silva (2015), avaliando nim indiano
sob ação de NaCl e CaCl2 em diferentes concentrações, encontrou reduções
nas coletas com 45 e 60 dias após a imposição do estresse nos tratamentos
severos (100 mM), com o CaCl2 sendo o sal mais prejudicial para a espécie.
Tais resultados colaboram com os encontrados na presente pesquisa, onde
foram observadas reduções ao longo do período experimental.
Para a variável condutância estomática (gs), foram vistas diferenças
significativas apenas para a coleta realizada 60 dias após a diferenciação, com
reduções acentuadas, em média de 60%, em relação à primeira coleta, e
valores médios iguais a 0,09, 0,06, 0,06, 0,06 mmolH2O.m-2.s-1, para 0 mM, 50
mM, 100 mM e 150 mM de NaCl na última coleta (Figura 11).
Távora et al. (2012) afirmam que as plantas fecham seus estômatos
visando a redução da perda de água por transpiração, reduzindo assim a taxa
fotossintética e consequentemente afetando todo o funcionamento da planta.
Figura 11. Avaliação da condutância estomática (gs) em mudas de nim indiano submetidas a
diferentes níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de
NaCl referente à coleta com 20, 40 e 60 dias dias após a imposição do estresse. Onde letras
minúsculas correspondem aos tratamentos e letras maiúsculas correspondem às coletas.

Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
Quando Lima et al. (2014) avaliaram espécies arbóreas (pau-mocó, ipê
roxo, nim e aroeira) submetidas ao encharcamento do solo com dois níveis de
salinidade (2,0 dS.m-1 e 12,0 dS.m-1), os autores observaram uma redução na
variável condutância estomática, após o 1º encharcamento, em todas as
espécies no tratamento encharcamento e alta salinidade (CE/AS). No 2º
encharcamento, realizado 58 dias após da semeadura, o nim e a aroeira
apresentaram as menores reduções, em relação às outras espécies. Silva et. al
(2012) trabalhando com pau-mocó e aroeira, concluiu que as duas espécies
apresentaram redução na condutância estomática com o aumento da
salinidade, com a aroeira apresentando a menor redução. Silva (2012)
encontrou nos seus resultados, avaliando mudas de moringa submetidas a
diferentes níveis de NaCl (0 mM, 75 mM e 150 mM), que as trocas gasosas
foram reduzidas com o aumento da salinidade. Tais resultados corroboram com
os encontrados na presente pesquisa, onde os tratamentos mais severos (100
mM e 150 mM de NaCl) reduziram suas trocas gasosas, porém não houve o
fechamento estomático.
Avaliando mudas de nim indiano submetidas a diferentes concentrações
de NaCl e CaCl2, Silva (2015) concluiu que a partir da segunda coleta (30 dias
de estresse) o CaCl2 já afetava a condutância estomática das mudas, sendo
três vezes menor que os valores apresentados pelo NaCl. Já aos 60 dias a
autora só encontrou diferenças significativas nos tratamentos com NaCl.
7.3. RELAÇÕES HÍDRICAS: POTENCIAL HÍDRICO FOLIAR (W) E TEOR
RELATIVO DE ÁGUA (TRA)
As relações hídricas foram afetadas, de maneira geral, com o passar do
tempo pelo acúmulo de sais na solução do solo. Os tratamentos mais severos
(100 mM e 150 mM de NaCl) sofreram reduções mais evidentes a partir dos 40
dias de estresse.

Analisando os resultados obtidos para o potencial hídrico foliar às 4 h da
manhã (Figura 1), os tratamentos salinos mais severos não diferiram entre si
nas duas primeiras coletas. Já aos 60 dias pode ser constatado uma redução
no potencial hídrico em todos os tratamentos, com o maior valor sendo
apresentado pelo tratamento 100 mM e NaCl com -1,81 MPa.
Figura 12. Avaliação do potencial hídrico foliar (w) em mudas de nim indiano submetidas a
diferentes níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de
NaCl às 4h referente às coletas com 20, 40 e 60 dias após a imposição do estresse. Onde
letras minúsculas correspondem aos tratamentos e letras maiúsculas correspondem às coletas.
Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
Com os dados obtidos às 12 h, foi observado que os tratamentos salinos
apresentaram valores maiores na última coleta, com valores em média de -1,89
MPa. Os tratamentos salinos aos 60 dias apresentaram valores próximos aos
observados aos 40 dias, com o tratamento 150 mM de NaCl não apresentando
diferenças significativas nas 3 coletas, com valores médios de – 2,32 MPa e o
tratamento 100 mM de NaCl não diferindo nas duas últimas coletas (Figura 13).
4 h

Figura 13. Avaliação do potencial hídrico foliar (w) em mudas de nim indiano submetidas a
diferentes níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de
NaCl às 12h referente às coletas com 20, 40 e 60 dias após a imposição do estresse. Onde
letras minúsculas correspondem aos tratamentos e letras maiúsculas correspondem às coletas.
Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
Mudas de goiabeira submetidas à salinidade, avaliadas por Távora et al
(2001), apresentaram uma redução no potencial com o incremento de sal e
com o aumento da duração do experimento. Silva (2013) avaliando mudas de
moringa sob estresse salino, afirma que as mudas no tratamento controle e
estresse moderado não apresentaram diferenças com 2h de imposição do
estresse, porém com 48h o estresse severo apresentou menor potencial hídrico
foliar em relação aos demais tratamentos. Silva (2015) afirma em sua pesquisa,
de acordo com os resultados encontrados, que as coletas realizadas às 4 horas
da manhã apresentam os efeitos puros dos tratamentos impostos,
diferentemente do que ocorre às 12 h, visto que as plantas apresentam maior
evapotranspiração e o ambiente tem temperaturas mais altas alterando assim o
status hídricos das mudas.
Para o teor relativo de água (TRA), às 4 h da manhã, pode ser visto na
figura 14, que na segunda coleta a porcentagem do TRA aumentou em relação
à coleta com 20 dias, onde os valores médios dos tratamentos salinos não
diferiram estatisticamente do tratamento controle, voltando a reduzir seus

valores na última coleta. Aos 60 dias de estresse os valores médios do TRA
foram iguais a 86,5%, 83,3%, 82%, 79,2% para 0 mM, 50 mM, 100 mM e 150
mM de NaCl respectivamente, estes valores não diferiram estatisticamente dos
tratamentos na coleta realizada aos 20 dias.
Figura 14. Avaliação do teor relativo de água (TRA) em mudas de nim indiano submetidas a
diferentes níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de
NaCl no horário às 4 h referente às coletas com 20, 40 e 60 dias após a imposição do estresse.
Onde letras minúsculas correspondem aos tratamentos e letras maiúsculas correspondem às
coletas. Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
Os valores médios, ás 12h, na coleta com 40 dias aumentaram 3,08%
em relação à primeira coleta (Figura 15). Os tratamentos 100 e 150 mM de
NaCl apresentaram diferenças estatísticas do tratamento controle em média de
9,3% na última coleta, com 60 dias de estresse, porém os mesmos tratamentos
apresentavam valores menores, sem diferir estatisticamente entre si nas três
coletas, do que os apresentados no tratamento Controle. O tratamento 50 mM
de NaCl apresentou um aumento gradativo nas coletas, igualando seus valores
médios na coleta com 40 e 60 dias com o tratamento controle, mostrando
assim que as plantas conseguiram manter o conteúdo hídrico dentro das
células, mesmo com o prolongamento do estresse.
A redução dos valores nos tratamentos salinos mais severos para as
relações hídricas levam a conclusão de que as plantas estão reduzindo a

capacidade de absorção de água devido ao acumulo de sal no solo (TÁVORA
et al, 2001).
Figura 15. Avaliação do teor relativo de água (TRA) em mudas de nim indiano submetidas a
diferentes níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de
NaCl no horário às 12 h referente às coletas com 20, 40 e 60 dias após a imposição do
estresse. Onde letras minúsculas correspondem aos tratamentos e letras maiúsculas
correspondem às coletas. Letras iguais não diferem entre os tratamentos pelo teste de Tukey
ao nível 5% de probabilidade.
Para a variável TRA Távora et al. (2001) afirmam que os valores
apresentaram reduções com o aumento do teor de sal, corroborando com os
resultados apresentados na presente pesquisa para a variável às 12h, assim
como com a maior exposição ao estresse. Seguindo o mesmo autor, as mudas
de goiabeira apresentaram redução de 13% no tratamento mais severo (150
mM de NaCl) aos 50 dias de estresse, em relação a primeira coleta, realizada
30 dias após a imposição do estresse.
Avaliando mudas de Moringa oleífera, Silva (2013) observou que as
mudas submetidas ao estresse severo (150 mM de NaCl) reduziram o teor
relativo de água nas 3 avaliações (2, 24 e 48 h), enquanto o tratamento com 75
mM de NaCl, não diferiu do tratamento controle com 2 h e 48 h de estresse.
Silva (2015) avaliando mudas de nim indiano submetidas a diferentes
concentrações de NaCl e CaCl2, afirma que nos 30 primeiros dias de estresse
as plantas dos tratamentos salinos (50 e 100 mM de NaCl e CaCl2), de maneira
geral, não sofreram com o acúmulo de sal. Na coleta com 60 dias de estresse,
foram vistas reduções, ainda que pequenas, onde às 4 h as plantas submetidas
ao CaCl2 apresentaram médias do TRA 3% (50 mM) e 6% (100 mM) menores
que os tratamentos salinos com NaCl. O mesmo autor afirma ainda que o nim
não é uma espécie sensível à estes tipos de sais, mesmo que em altas
concentrações, confirmando os resultados encontrados na presente pesquisa.
Onde mesmo com reduções das relações hídricas as plantas não
apresentaram perda de turgescência nem sintomas de injúrias causadas pelo
acúmulo de sais.
7.4. PIGMENTOS FOTOSSINTÉTICOS (CLOROFILA a, CLOROFILA b,
CLOROFILA TOTAL E CAROTENOIDES)
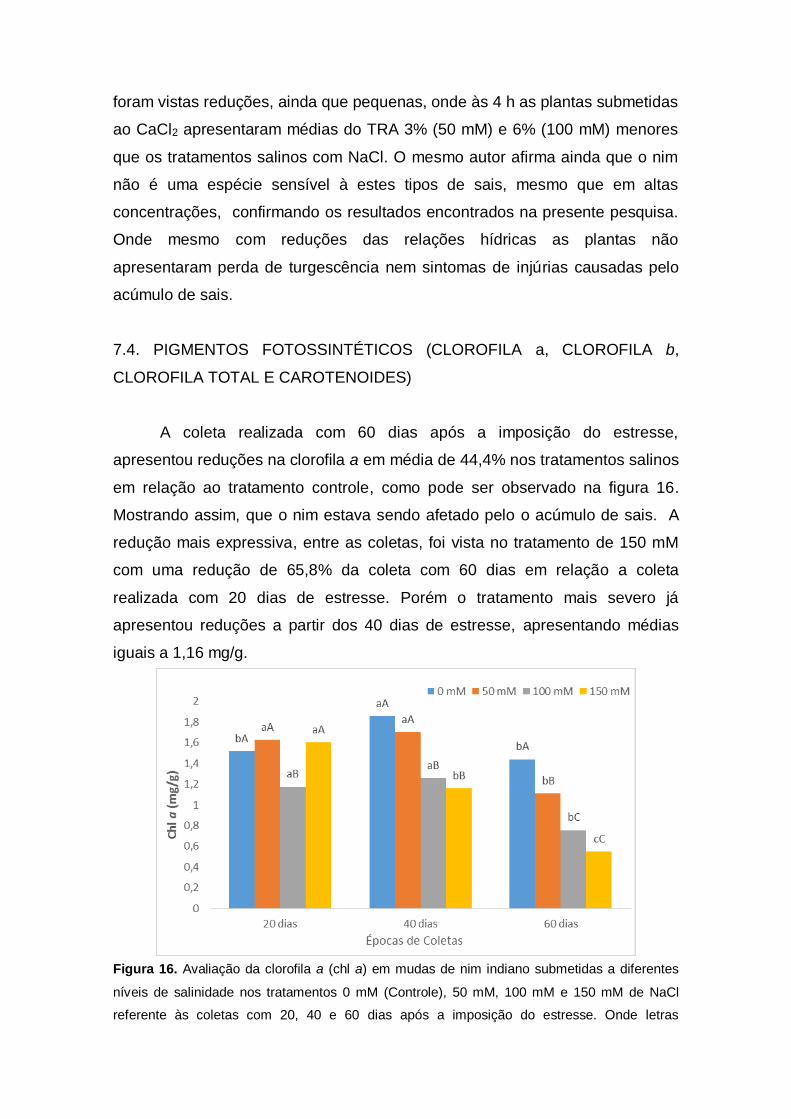
A coleta realizada com 60 dias após a imposição do estresse,
apresentou reduções na clorofila a em média de 44,4% nos tratamentos salinos
em relação ao tratamento controle, como pode ser observado na figura 16.
Mostrando assim, que o nim estava sendo afetado pelo o acúmulo de sais. A
redução mais expressiva, entre as coletas, foi vista no tratamento de 150 mM
com uma redução de 65,8% da coleta com 60 dias em relação a coleta
realizada com 20 dias de estresse. Porém o tratamento mais severo já
apresentou reduções a partir dos 40 dias de estresse, apresentando médias
iguais a 1,16 mg/g.
Figura 16. Avaliação da clorofila a (chl a) em mudas de nim indiano submetidas a diferentes
níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de NaCl
referente às coletas com 20, 40 e 60 dias após a imposição do estresse. Onde letras

minúsculas correspondem aos tratamentos e letras maiúsculas correspondem às coletas.
Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
Mudas de moringa submetidas a três níveis de salinidade, avaliadas por
Silva (2013), não apresentaram diferenças significativas entre os tratamentos
salinos. Avaliando mudas de jatobá submetidas a diferentes níveis de
salinidade (0 mM, 50 mM e 100 mM de NaCl), Nascimento (2013) afirma que
não foram apresentas diferenças significativas entre os tratamentos nos dois
períodos analisados.
Avaliando mudas de nim indiano sob ação de NaCl e CaCl2 em
diferentes concentrações Silva (2015) afirma que os teores de clorofila a
sofreram reduções de 12% (100 mM de NaCl) e 15% (100 mM de CaCl2) aos
15 dias de estresse. O tratamento 100 mM de NaCl reduziu 52% em relação ao
tratamento mais ameno sendo o mais afetado coleta realizada 60 dias após a
imposição do estresse. Tais resultados colaboram com os encontrados na
presente pesquisa, visto que o tratamento mais severo de NaCl foi o mais
afetado ao final do período experimental.
Para os resultados da clorofila b, foi observado que na primeira coleta,
20 dias após a imposição do estresse, houve uma redução de 28,5% no
tratamento de 100 mM em relação ao tratamento Controle (Figura 17). Aos 40
dias de estresse foi vista uma redução nos tratamentos mais severos de
52,11% em média. Já na última coleta, os tratamentos não diferiram entre si,
porém quando analisamos os tratamentos de maneira isolada pode ser visto
que os tratamentos salinos apresentaram reduções no teor de clorofila b desde
a segunda coleta.

Figura 17. Avaliação da clorofila b (chl b) em mudas de nim indiano submetidas a diferentes
níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de NaCl
referente às coletas com 20, 40 e 60 dias após a imposição do estresse. Onde letras
minúsculas correspondem aos tratamentos e letras maiúsculas correspondem às coletas.
Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
Silva (2013) observou que mudas de moringa quando submetidas à
salinidade não apresentaram diferenças significativas após 2h da imposição do
estresse, entretanto com o maior tempo de exposição ao estresse a mudas
apresentaram reduções no tratamento mais severo (150 mM de NaCl) em
relação ao tratamento controle. Lima et al. (2012) avaliando mudas de pinhão-
manso submetidas a quatro níveis de salinidade (0,45; 2,5; 5,0 e 7,5 dS.m-1) e
4 teores de Si (0; 221,76; 443,52 e 665,28 mg.L-1), verificaram que apenas a
clorofila b no tratamento 0 dS.m-1 relacionado com 655,28 mg.L-1 apresentou
o maior valor (102,5 μmol m-2) em relação aos demais tratamentos. A clorofila
a, total e os carotenoides não apresentaram diferenças estatísticas, indicando
segundo os autores, que a interação entre o sal e o Silício reduziu os efeitos do
sal sobre a espécie.
Na presente pesquisa o nim sofreu reduções nos teores de clorofila b
com mais intensidade na última coleta, assim como na pesquisa realizada por
Silva (2015), onde mudas de Azadirachta indica submetidas a 50 mM e 100
mM de NaCl e CaCl2 começaram a apresentar reduções nos teores de clorofila
b apenas na terceira coleta (45 dias de estresse), porém as maiores reduções
ficaram evidentes apenas na última coleta, onde o tratamento 100 mM de NaCl
apresentou redução de 58% em relação ao tratamento moderado (50 mM), e
tratamento 50 mM de CaCl2 foi 25% menor que o tratamento de 50 mM de
NaCl. De maneira geral pode ser observado um decréscimo gradual da
clorofila total da primeira para a última coleta, como pode ser visto no figura 18.
Com os valores obtidos para a clorofila total pode ser observada uma redução
entre a primeira (20 dias) e a última coleta (60 dias), com as maiores reduções
sendo dos tratamentos 50 mM, 1,97 mg/g e 1,28 mg/g e 100 mM com 2,48

mg/g e 1,12 mg/g, sendo o primeiro valor para 20 dias e o segundo 60 dias de
estresse respectivamente.
Figura 18. Avaliação da clorofila total (chl total) em mudas de nim indiano submetidas a
diferentes níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de
NaCl referente às coletas com 20, 40 e 60 dias após a imposição do estresse. Onde letras
minúsculas correspondem aos tratamentos e letras maiúsculas correspondem às coletas.
Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
Nascimento (2013) avaliando mudas de jatobá, observou que os teores
de clorofila total nos tratamentos 50 mM e 100 mM de NaCl, nas duas coletas,
apresentaram um acréscimo nos seus valores, com 25,25% e 27,6%
respectivamente. As mudas de nim indiano avaliadas por Silva (2015)
apresentaram reduções nos teores de clorofila total a partir da segunda coleta,
com redução mais acentuada na ultima coleta (60 dias de estresse) com
reduções de 53% do tratamento 100 mM de NaCl em relação ao 50 mM do
mesmo sal, e com as plantas sob ação do CaCl2 apresentando menores teores
em relação a mesma concentração do NaCl.
Para os carotenoides foi observado um comportamento diferente ao
observado até a segunda coleta, onde pode ser visto que os teores de
carotenoides, foram aumentados em 149% em média em relação à primeira
coleta (Figura 19). Podemos entender com esses resultados que as plantas
aumentaram a produção de carotenoides, por ser um pigmento acessório

fotoprotetor, para que as clorofilas não fossem afetadas de forma direta,
fazendo com que as plantas não interrompessem a fotossíntese (STREIT et al,
2005).
Os teores de carotenoides na coleta após 60 dias de estresse reduziram
bruscamente em relação à segunda coleta, diferindo estatisticamente da
segunda coleta (40 dias de estresse) porém sem apresentar diferenças da
coleta com 20 dias de estresse. Os carotenoides analisados na terceira coleta
apresentaram reduções de 62% para 0 mM, 56,2% para 50 mM, 54,8% para
100 mM e 50,3% para 150 mM de NaCl em relação a segunda coleta. Tais
resultados mostram que o nim após 40 dias começou a ser afetado pelo
acúmulo de sais, não conseguindo mais produzir pigmentos acessórios
suficientes para que as clorofilas a não fossem danificadas com os possíveis
danos do acúmulo de NaCl.
Figura 19. Avaliação dos carotenoides em mudas de nim indiano submetidas a diferentes
níveis de salinidade nos tratamentos 0 mM (Controle), 50 mM, 100 mM e 150 mM de NaCl
referente às coletas com 20, 40 e 60 dias após a imposição do estresse. Onde letras
minúsculas correspondem aos tratamentos e letras maiúsculas correspondem às coletas.
Letras iguais não diferem entre os tratamentos pelo teste de Tukey ao nível 5% de
probabilidade.
Semelhante aos resultados encontrados para clorofila b, as mudas de
jatóba quando submetidas a salinidade aumentaram o teor de carotenoides,
26,5% para 50 mM e 28,7% para 10 mM) nos tratamentos salinos em relação
ao tratamento controle. Tais resultado apresentados por Nascimento (2013)

corroboram com a presente pesquisa, onde os teores de carotenoides foram
acrescidos na segunda coleta.
Diferente do encontrado na presente pesquisa Silva (2015) afirma que
as mudas de nim indiano nas coletas com 30 e 45 dias de estresse reduziram
os teores de carotenoides em relação a primeira coleta (20 dias), porém 60
dias após a imposição do estresse os tratamentos salinos (50 mM e 100 mM de
NaCl e CaCl2) aumentaram a produção de carotenoides. O autor ressalta ainda
a possibilidade das plantas produzirem pigmentos acessórios para proteger o
sistema fotossintético.
8. CONSIDERAÇÕES FINAIS
Diante do exposto na presente pesquisa é possível concluir que mesmo
com as mudanças na fisiologia das plantas o nim se mostrou tolerante à
salinidade sem alterações na produção de matéria seca, podendo ser possível
cultivar mudas de nim indiano em solos salinos com concentrações de até 150
mM de NaCl.

9. CRONOGRAMA
10. BIBLIOGRAFIA
ARAÚJO, L. V. C.; RODRIGUEZ, L. C. E.; PAES, J. B. Características físico-químicas e energéticas da madeiras de nim indiano. Scientia Florestalis. Piracicaba, n. 57, p. 153-159, jun. 2000.
ALVES, A. R. Níveis Críticos de Fósforo e Potássio para Crescimento Inicial de Nim (Azadirachta indica A. Juss). 2008. 48 f. Dissertação
(Mestrado) – Programa de pós-graduação de Ciências Florestais, Universidade Federal Rural de Pernambuco, Recife, 2008.
BENINCASA, M. M. P. Análise de crescimento de Plantas (Noções
básicas). Jaboticabal: FUNEP, 1988. 42p.

BITTENCOURT, A. M. O cultivo do nim indiano (azadirachta indica a. juss):
uma visão econômica. 2006. 125 f. Dissertação (Mestrado) - Curso de
Engenharia Florestal, Universidade Federal do Paraná, Curitiba, 2006.
CAIRO, P.A.R. Curso básico de relações hídricas de plantas. Vitória da
Conquista, UESB, 1995. 32p.
COSTA, J. D. S; SILVA, J. A. B; TARGINO, H. C O; SANTOS, I. E. A;
BARROS, E. S. C; SEABRA, T. X; NERY, F. M. Efeito da Salinidade do Solo
Sobro Crescimento Inicial de Plantas nim [Azadirachta indica
(MELIACEAE)]. Anais do 64º Congresso Nacional de Botânica, Belo Horizonte,
2013.
DINIZ, B. L. M. T. et al. Crescimento inicial e consume hídrico de nim submetido ao estresse salino e biofertilizante bovino. Revista Brasileira de Ciências Agrárias. V. 8, n. 3, p. 470-475. 2013.
FERREIRA, P. A; SILVA, J. B. L; RUIZ, H. A. Aspectos Físicos e Químicos de Solos em Região Áridas e Semiárias. In Manejo de Salinidade na
Agricultura: Estudos Básicos e Aplicados. GHEYI et. al. INCTSal, 2010.
FREIRE, A. L. O. et al. Crescimento e nutrição mineral do nim (Azadirachta indica A. Juss.) e Cinamono (Melia azedarach Linn.) submetidos à salinidade. Ciência Florestal. V. 20, n. 2, p. 207-215. 2010.
HOAGLAND, D.R.; ARNON, D.I. The water-culture method for growing plants without soil. California, California Agricultural Experiment Station, 32p. (CAES. Circular, 347). 1950.
HOLANDA, A. C; SANTOS, R. V; SOUTO, J. S; ALVES, A.R. Desenvolvimento
inicial de espécies arbóreas em ambientes degradados por sais. Revista de
Biologia e Ciências da Terra. São Cristovão, vol. 07, n. 01, p. 39-50,
setembro, 2007.
KARASAWA, S; EGUCHI, E.S; MIRANDA, J. H; IGARASHI, I. G. S. Teor
Relativo de Água (TRA) nas Folhas da Muda de Cafeeiro Submetido à
Irrigação com Água Salina. II Simpósio de Pesquisa do Café no Brasil, Vitória
–ES, p.222-226, 2001.
LICHTENTHALER H. K. & BUSCHMANN, C. Chlorophylls and Carotenoids:
Measurement and Characterization by UV-VIS Spectroscopy. In: WROLSTAD,
R. E.; ACREE, T. E.; AN, H.; DECKER, E. A.; PENNER, M. H.; REID, D. S.;
SCHWARTZ, S. J.; SHOEMAKER, C. F.; SPORNS, P. Current Protocols in
Food Analytical Chemistry. John Wiley & Sons, Inc: New Jersey. Unit F4.3, p.
F4.3.1-F4.3.8, 2001.
LIMA, A. D; SILVA, J. A; SOUSA, C. H. C; BEZERRA, A. M. E; LACERDA, C.
F. Trocas Gasosas em Espécies Florestais Submetidas a Estresse Salino
e Períodos de Encharcamento do Solo. II Inovagri Internacional Meeting.
Fortaleza, 2014.

LIMA, M. S. R; BELTRÃO, N. E. M; ROCHA, M. S. R; FERRAZ, R. L. S;
CARVALHO JUNIOR, G. S; NEGREIROS, K. V. Concentração de Pigmentos
Fotossintéticos nas Folhas do Pinhão Mando Cultivado sob Salinidade e
Silício. V CONGRESSO BRASILEIRO DE MAMONA; II SIMPÓSIO
INTERNACIONAL DE OLEAGINOSAS ENERGÉTICAS & I FÓRUM
CAPIXABA DE PINHÃO MANSO, 2012, Guarapari. Desafios e Oportunidades:
Anais. Campina Grande: Embrapa Algodão, 2012. p. 278.
MARTINEZ, S. S. O nim – Azadirachta indica – um inseticida natural.
Londrina, fev. 2008 (Documentos IAPAR).
MARTINS, M. O. Aspectos fisiológicos do nim indiano sob déficit hídrico
em condições de casa de vegetação. 2008. 84f. Dissertação (Mestrado em
Botânica) Universidade Federal Rural de Pernambuco, Recife.
MESQUITA, F. O; CAVACANTI, L. F; DINIZ, B. L. M. T; SOUTO, A. G. L; LIMA
NETO, A. J; MEDEIROS, R. F. Produção de Mudas de Nim sob Irrigação
Suplementar com Àguas Salinas, Biofertilizante e Drenagem. I Inovagri
International Meeting e IV Winotec. Fortaleza, 2012.
MOSSINI, S. A. G; KEMMELMEIER, C. A árvore NIm (Azadiravtha indica A. Juss): Multiplos usos. Acta Farmacéutica Bonaerense - vol. 24 n° 1, p. 139-
148, 2005.
MOREIRA, F. T. A; ARAÚJO, L. V. C; SILVA, S. H; ASSIS, M. M; FERREIRA,
A. E. S. Características dendrométricas de um povoamento de nim indiano
(Azadirachta indica A. Juss) no semiárido paraibano. Revista Verde (Mossoró
– RN), v. 7, n. 3, p. 194-198, jul-set, 2012.
NASCIMENTO, H. H. C. MECANISMOS FISIOLÓGICOS E BIOQUÍMICOS EM
MUDAS DE JATOBÁ (Hymenaea courbaril L.), SOB CONDIÇÕES
ADVERSAS. 2013. 162f. Dissertação (Mestrado em Ciências Florestais).
Universidade Federal Rural de Pernambuco, Recife.
NEVES, B. P. das; OLIVEIRA, I. P. de; NOGUEIRA, R. J. M. C. Cultivo e
utilização do nim indiano. Santo Antônio de Goiás: Embrapa Arroz e Feijão,
2003. 12 p. (Embrapa Arroz e Feijão. Circular Técnica, 62).
NEVES, B. P. das; CARPANEZZI, A. A. O Cultivo de Nim para Produção de
Fruto nos Brasil. Colombo: Embrapa Florestas, 2008. 8p. (Embrapa Florestas.
Circular Técnica, 162).
NUNES, J. C; CAVALCANTI, L. F; LIMA NETO, A. J; REBEQUI, A. M; DINIZ, B. M. T; GHEYI, H. R. Comportamento de Mudas de Nim à Salinidade da Àgua em Solo não Salino com Biofertilizante. Revista Brasileira de Engenharia Agrícola e Ambiental, v.16, n.11, p.1152–1158, 2012.
RIBEIRO, M. R. SALINIDADE NO SOLO E NA ÁGUA IN GHEYI: MANEJO
DA SALINIDADE NA AGRICULTURA: ESTUDOS BÁSICOS E APLICADOS.
FORTALEZA, INCT SAL. 2010. p.472.

SCHOLANDER, P.F.; HAMMEL, H. T.; HEMINGSEN, E. A.; BRADSTREES, E.
D. Hydrostatic pressure and osmotic potencial in leaves of mangroves and
some other plants. Proceedings of Nacional Academy Science, v.51, p. 119-
125, 1965.
SILVA, N. V. Germinação e Estabelecimento de Mudas de Nim Indiano
(Azadirachta indica A. Juss.) Submetidas a Estresses Abióticos. 112f.
Dissertação (Mestrado em Ciências Florestais). Universidade Federal Rural de
Pernambuco, Recife.
SILVA, E. C. A. Respostas Fisiológicas, Bioquímicas e Enzimáticas em
mudas de Moringa oleífera Lam. Submetidas a Estresses Abióticos. 2013.
98f. Dissertação (Mestrado em Ciências Florestais). Universidade Federal
Rural de Pernambuco, Recife.
SILVA, J. A; NOGUEIRA FILHO, F. P; BESSA, M. C; PERREIRA, M. S;
BEZERRA, A. M. E; LACERDA, C. F. Respostas Fisiológicas de
Myracrodruon urundeuva e Luetzelburgia auriculata à Salinidade. I
Inovagri International Meeting e IV Winotec. Fortaleza, 2012.
STREIT, N. M; CANTERLE, L. P.; CANTO, M. W de.; HECKTHEUER, L. H. H.
As Clorofilas. Ciência Rural, Santa Maria, v.35, n.3, p.748-755, 2005.
TAIZ, L; ZEIGER, E. Fisiologia Vegetal. 3.ed. Porto Alegre: Artmed, 2006.
719p.
TÁVORA, F. J. A. F; FERREIRA, R. G; HERNANDEZ, F. F. F. Crescimento e
Relações Hídricas em Plantas de Goiabeira Submetidas a Estresse Salino com
NaCl. Revista Brasileira de Fruticultura, Jaboticabal - SP, v. 23, n. 2, p. 441-
446, 2001.
WALTER, L.S; SILVA, N. V; SILVA, M. F. L. T; SILVA, E. C. A; ARAÚJO, R. P.
S; NOGUEIRA, R. J. M. C. Crescimento, trocas gasosas e relações hídricas em
mudas de nim indiano submetidas a diferentes níveis de irrigação. In Jornada
de Pesquisa e Extensão, 14, 2014, Recife. Anais/Resumos da XIV JEPEX.
Recife: UFRPE, 2014.
WILLADINO, L; CAMARA, T. R. Tolerância das Plantas à Salinidade: Aspectos
Fisiológicos e Bioquímicos. Enciclopédia Biosfera, Centro Científico Conhecer
– Goiânia, vol. 06, n. 11, 2010.
WILLADINO, L. G; CAMARA, T. R. Aspectos Fisiológicos do Estresse
Salino em Plantas. In.: NOGUEIRA, et. al. Estresses Ambientais: danos e
benefícios causados em plantas. Recife: Universidade Federal Rural de
Pernambuco, 2005. 500p.
ZAR, J. H. Biostatistical Andyses. New Jersey: Prentice Hall, 1999. 663p.

11. ATIVIDADES RELEVANTES DESENVOLVIDAS PELO BOLSISTA
Resumos publicados de evento:
LIMA, O. M. L. R; BARRETO, T. H. L; SANTOS, C. A; PACHECO, C. M; SILVA, N. V; WALTER, L. S; NOGUEIR, R. J. M. C. AVALIAÇÃO DE CRESCIMENTO,
RELAÇÕES HÍDRICAS E TROCAS GASOSAS EM MUDAS DE UMA ESPÉCIE FLORESTAL DA CAATINGA SUBMETIDA AO ESTRESSE HÍDRICO. In.: XIV Jornada de Ensino, Pesquisa e Extensão, 2014, Recife. Anais da XIV Jornada de Ensino Pesquisa e Extensão da UFRPE, 2014.
SILVA, M. F. L. T; SANTOS, C. A; WALTER, L. S; PACHECO, C. M.; SILVA, N. V; NOGUEIRA, R. J. M. C. AVALIAÇÃO DAS RELAÇÕES HÍDRICAS DE SORGO SACARINO (Sorghum bicolor (L.) Moench) EM ESTRESSE HÍDRICO. In.: XIV Jornada de Ensino, Pesquisa e Extensão, 2014, Recife. Anais da XIV Jornada de Ensino Pesquisa e Extensão da UFRPE, 2014.
WALTER, L. S; SILVA, L. A. C; SILVA, M. F. L. T; SILVA, N. V; SILVA, E. C. A;
ARAÚJO, R. P. S; NOGUEIRA, R. J. M. C. CRESCIMENTO, TROCAS GASOSAS E RELAÇÕES HÍDRICAS EM MUDAS DE NIM INDIANO SUBMETIDAS A DIFERENTES NÍVEIS DE IRRIGAÇÃO. In.: XIV Jornada de Ensino, Pesquisa e Extensão, 2014, Recife. Anais da XIV Jornada de Ensino Pesquisa e Extensão da UFRPE, 2014.
Participação de evento:
• XIV JORNADA DE ENSINO, PESQUISA E EXTENSÃO da UFRPE, 2014.
Recommended

















