
http://jbr.sagepub.com/Journal of Biological Rhythms
http://jbr.sagepub.com/content/28/2/95The online version of this article can be found at:
DOI: 10.1177/0748730413481921
2013 28: 95J Biol RhythmsLawrence P. Morin
Nocturnal Light and Nocturnal Rodents: Similar Regulation of Disparate Functions?
Published by:
http://www.sagepublications.com
On behalf of:
Society for Research on Biological Rhythms
can be found at:Journal of Biological RhythmsAdditional services and information for
http://jbr.sagepub.com/cgi/alertsEmail Alerts:
http://jbr.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Apr 19, 2013Version of Record >>
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

95
JOURNAL OF BIOLOGICAL RHYTHMS, Vol. 28 No. 2, April 2013 95-106DOI: 10.1177/0748730413481921© 2013 The Author(s)
1 To whom all correspondence should be addressed: Lawrence P. Morin, Department of Psychiatry, Stony Brook Medical Center, Stony Brook University, Stony Brook, NY 11794-8101; email: [email protected].
CONJECTURE
Nocturnal Light and Nocturnal Rodents: Similar Regulation of Disparate Functions?
Lawrence P. Morin*,1
*Department of Psychiatry, Stony Brook Medical Center, Stony Brook University, Stony Brook, NY
Abstract Investigators typically study one function of the circadian visual system at a time, be it photoreception, transmission of photic information to the suprachiasmatic nucleus (SCN), light control of rhythm phase, locomotor activ-ity, or gene expression. There are good reasons for such a focused approach, but sometimes it is advantageous to look at the broader picture, asking how all the parts and functions complete the whole. Here, several seemingly disparate functions of the circadian visual system are examined. They share common characteristics with respect to regulation by light and, to the extent known, share a common input neuroanatomy. The argument presented is that the 3 hypothalamically mediated effects of light for which there are the most data, circadian clock phase shifts, suppression of nocturnal locomotion (“negative masking”), and suppression of nocturnal pineal function, are regulated by a common photic input pathway terminating in the SCN. For each, light triggers a relatively fixed interval response that is irradiance-dependent, the effective stimulus can be very brief light exposure, and the response continues to com-pletion in the absence of additional light. The presence of a triggered, fixed-length response interval is of particular importance to the understanding of the circuitry and mechanisms regulating circadian rhythm phase shifts because it implies that the SCN clock response to light is not instantaneous. It also may explain why certain stimuli (neuropeptide Y or novel wheel running) adminis-tered many minutes after light exposure are able to block light-induced phase shifts. The understanding of negative masking is complicated by the fact that it can be represented as a positive change, that is, light-induced sleep, not just as a reduction in locomotion. Acute nocturnal light exposure also induces adrenal hormone secretion and a rapid drop in body temperature, physiological responses that appear to be regulated similarly to the other light effects. The likelihood of a common regulatory basis for the several responses suggests that additional light-induced responses will be forthcoming and raises questions about the relationships between light, SCN cellular anatomy, the molecular clockworks of SCN neurons, and SCN throughput mechanisms for regulating disparate downstream activities.
Keywords circadian, sleep, masking, melatonin, locomotion, pineal, suprachiasmatic
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

96 JOURNAL OF BIOLOGICAL RHYTHMS / April 2013
Circadian rhythm investigators “know” what masking is despite the absence of a rigorous defini-tion. In his review of the topic, Mrosovsky (1999) defined negative masking in nocturnal animals as “a decrease in activity” and positive masking as “an increase in activity.” Other varieties of masking have also been defined (Mrosovsky, 1999). Most investiga-tors are familiar with negative masking as an index of light’s suppressive effects on wheel-running behavior. Masking has also been used much more generally in reference to the extent to which virtually any intrinsic or extrinsic stimulus impairs expression of an action of the circadian clock. For example, Redlin (2001) considers light-induced suppression of wheel-running and melatonin to be negative mask-ing responses (also see Mrosovsky, 1999, for discus-sion and references). Rensing prefers to consider the stimulus effects as “direct” effects rather than as “masking” effects (Rensing, 1989). Redlin has com-bined terms, referring to masking as resulting from direct effects of light (Redlin, 2001).
Redlin’s suggestion that light-induced activity suppression and inhibition of melatonin production are both masking phenomena illustrates some of the semantic confusion surrounding the concept. This arises because the two variables are fundamentally different. Melatonin level is a physiological measure that varies on a continuum from low to high, accord-ing to the prevailing conditions, and it is not known to covary with other measures. In contrast, measures of “negative masking” have been used as indices of behavioral change, without considering the fact that by definition, all behaviors must always sum to 100%. In other words, when light reduces activity to 0% of ongoing behavior (meaning that “negative masking” reaches 100%), at least 1 alternate behavior must have correspondingly increased. If we could identify and measure that elevated behavior, would it be an index of positive masking or negative masking? Clarification of this semantic difficulty is necessary in order to uncover organizing principles governing the effects of light on physiology and behavior.
A second problem regarding the historical use of “negative masking” is the implication that the behav-ioral change to which it refers is contingent upon the presence of light. A second assumption is explicit: masking effects of light occur “by a route that does not involve a pacemaker” (Mrosovsky, 1999). Despite wide acceptance of both characteristics (Aschoff, 1960, 1981; Minors and Waterhouse,1989; Mrosovsky, 1999; Redfern et al., 1994; Redlin, 2001), neither
expectation has been proven. More important for the purposes of the present discussion is the recent dem-onstration that sustained suppression of locomotion does not require the continued presence of light (Morin and Studholme, 2009; Vidal and Morin, 2007). Moreover, research questioning the relationship between light and negative masking has revealed several unexpected results. One is the occurrence of light-induced sleep, hereafter referred to as “photo-somnolence.” The fact that nocturnal light induces sleep in mice and hamsters (Morin and Studholme, 2009, 2011; Studholme et al., 2013) is a clear example of the above point that if one behavior becomes less likely (e.g., active wakefulness), at least one other behavior must become more likely (e.g., sleep).
The revelation of photosomnolence requires reconsideration of what negative masking actually represents in order to understand how light modifies physiology and behavior. Here, it is proposed that light induces circadian rhythm phase shifts and suppresses pineal function in ways that are remark-ably similar to the effects of light on negative mask-ing. Indeed, it is plausible that all 3 responses, mediated through the circadian visual system, share a single, light-activated, time-limited mechanism necessary for their expression which, once initiated, continues to completion without additional light being necessary.
The present discussion focuses on the above high-lighted issues, emphasizing the need for better understanding of the nature of light-induced changes in physiology and behavior while addressing some of the ramifications that the issues may present to circadian rhythm research and the understanding of global SCN function. For the remainder of this pre-sentation, masking refers to negative masking unless otherwise specified and, to avoid semantic confu-sion, will be generally replaced by the term locomotor suppression.
LOCOmOTOR SUppRESSiON aNd phaSE ShifTiNg aRE SimULTaNEOUSLy iNdUCEd
by LighT
Mrosovsky used several novel ways of assessing light-induced locomotor suppression, but the basic procedure has been to provide nocturnal rodents with a running wheel under a standard light-dark (LD) photoperiod and expose them to a 30- or 60-min light pulse during the early activity phase (Mrosovsky,
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

Morin / EFFECTS OF NOCTURNAL LIGHT 97
1999; Mrosovsky et al., 1999 2000, 2001; Redlin and Mrosovsky, 1999b). In a typical study with saturating light, running declines to zero within a few minutes, where it remains during continued expo-sure to the stimulus (Redlin and Mrosovsky, 1999b).
Phase shift studies are conducted similarly. Depending on whether an Aschoff Type I or II procedure is used, animals are either in constant dark (DD) or LD and are exposed to a 5- to 60-min light pulse. In rhythm studies, a great deal of emphasis has been placed on effects of light on the circadian clock and the phase response curves (PRCs) that describe those effects. Masking receives little or no mention. Nevertheless, it should be abundantly clear that procedures for dem-onstrating masking and phase shifting are essentially identical and that every phase shift study is, by default, a masking study. The major difference between the two is the elapsed time before a result can be considered reliable. From a practical perspec-tive, useful results about negative masking can be obtained within a few minutes of stimulus onset, whereas reliable phase shift detection requires many hours or days after the stimulus to collect the data.
More bluntly, 2 different responses are elicited via the identical stimulus (Fig. 1). One is considered to be a demonstration of the most important function of the circadian visual system, namely, the ability to maintain a stable temporal relationship with the environmental photoperiod through the phase shift-ing action of light on the circadian clock. Masking, in contrast, is usually considered (if it is considered at all) to be a hindrance to circadian rhythm assessment, a mere by-product resulting from a procedure by which light is used to induce a phase shift. Because identical methods are used to test phase shifting and masking, an obligatory conclusion is that in the absence of other qualifying information, experiments designed to evaluate how light alters intrinsic func-tion of the circadian clock (i.e., the SCN) cannot distinguish between effects on clock phase and effects on masking. Mrosovsky (1999) lamented, with good reason, that so “little study has been devoted to masking compared to that lavished on entrainment.”
��
A B
0 6 12 18 24
**
figure 1. a millisecond light stimulus effective for eliciting phase shifts is also effec-tive for eliciting a masking response. here, several days of a running record obtained from a constant dark-housed mouse (Mus musculus) are shown. The stimulus consisted of 10 flashes of 2 msec each, distributed equally across a 5-min interval beginning at CT13. in (a), the circadian periods before and after the stimulus are indicated by the slope of each black line fitted through the daily activity onsets. The effect of the light stimulus yielded a subsequent phase shift indicated by the greek letters DF in the enlargement (b). The exact time of onset for the flash sequence is indicated by the black arrow in (b). The asterisks in (a) and (b) indicate the interval of locomotor sup-pression induced by the flashes. modified after fig. 1 in morin and Studholme (2009).
ThE phOTiC iNpUT paThwayS fOR maSkiNg aNd phaSE ShifTiNg
may bE idENTiCaL
Circadian rhythm phase shifts and masking respond to photic information arriving in the brain via very similar, if not identical, input pathways involving classical rod/cone and ganglion cell photo-reception. As is now well known, about 1% to 2% of mammalian retinal ganglion cells are photoreceptive (pRGCs) (Berson et al., 2002; Hattar et al., 2002) and use melanopsin as a photopigment (Panda et al., 2005; Qiu et al., 2005). Both classical (rod/cone) pho-toreceptors and pRGCs mediate masking and circa-dian rhythm phase and account for all non-image-forming visual responses (Hattar et al., 2003; Mrosovsky and Hattar, 2003; Mrosovsky et al., 2001; Thompson et al., 2008). Moreover, photic infor-mation received by classical photoreceptors appar-ently must pass through pRGCs in order to elicit those non-image-forming visual responses (Goz et al., 2008; Guler et al., 2008; Hatori et al., 2008).
Retinal ganglion cells project to about 30 retinore-cipient brain regions (Morin and Blanchard, 1999) and the pRGCs project to many of these regions (Ecker et al., 2010; Gooley and Saper, 2003; Hattar et al., 2006; Morin et al., 2003). In particular, pRGCs densely innervate the SCN, intergeniculate leaflet (IGL), and olivary pretectum and less densely innervate the dorsal lateral geniculate and superior colliculus.
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

98 JOURNAL OF BIOLOGICAL RHYTHMS / April 2013
It has been known since 1972 that destruction of the SCN eliminates circadian rhythmicity in rodents (Moore and Eichler, 1972; Stephan and Zucker, 1972). Lesions elsewhere in the brain generally have no effect on rhythm expression. Two exceptions are the IGL and pretectal-tectal region, both of which medi-ate rhythm response to light (Harrington and Rusak, 1986; Marchant and Morin, 1999; Morin and Pace, 2002; Pickard et al., 1987). This information is back-ground for a series of studies by Mrosovsky and col-leagues designed to determine which brain region is necessary for masking. Masking has been tested after lesions of the retinorecipient pretectum, tectum, dor-sal lateral geniculate, ventral lateral geniculate, IGL, and visual cortex (Edelstein and Mrosovsky, 2001; Mrosovsky, 1999; Redlin et al., 2003; Redlin et al., 1999). The results are unequivocal: masking persists in the absence of these brain regions (although 1 or more may modulate “positive masking” responses to low irradiance).
Considering the foregoing results, the SCN is the most likely location for the control of masking. This, together with the observation that the photic input pathway for circadian rhythm phase control seems to be the same as that for masking, argues by analogy that the SCN is essential to both functions. Unfortunately, 2 tests of SCN contribution to mask-ing have yielded opposite conclusions (Li et al., 2005; Redlin and Mrosovsky, 1999a). The studies were similarly conducted except that Li et al. (2005) used a powerful method (Pickard and Turek, 1982) that determined complete destruction of the SCN by absence of any SCN retinal terminal field. The idea that the SCN regulates masking is also supported by work showing that effects of light are absent from published running records generated by animals with complete transection of the retinohypothalamic tract (RHT), a procedure that blocks entrainment (Johnson et al., 1988a). Similarly, transplantation of an embryonic SCN to an arrhythmic adult restores locomotor rhythmicity without entrainment or apparent masking (Lehman et al., 1987). Thus, com-plete SCN lesions not only cause arrhythmicity (and loss of entrainment by light) (Rusak, 1977 but also block masking (Li et al., 2005). The existing data sup-port the view that there is no difference between the photoentrainment pathway and the pathway for light-induced locomotor suppression. Although unlikely, it remains possible that the functional path-way for light-induced locomotor suppression termi-nates not in the SCN but in another retinorecipient
hypothalamic region (Johnson et al., 1988b; Muscat et al., 2003).
ThE phOTiC iNpUT paThway fOR piNEaL REgULaTiON iS SimiLaR TO ThaT
fOR maSkiNg aNd phaSE ShifTiNg
A third non-image-forming visual response initi-ated via rod/cone or pRGC photoreception is light suppression of pineal activity (Hattar et al., 2003; Panda et al., 2003). SCN lesions that render animals arrhythmic and abolish locomotor suppression also abolish photic regulation of the pineal (Antle et al., 2007; Klein and Moore, 1979; Li et al., 2005; Rusak and Morin, 1976; Stetson and Watson-Whitmyre, 1976). Such results support the view that photic regu-lation of pineal function occurs through the same input pathway influencing locomotor suppression and phase shifting.
VERy bRiEf LighT ExpOSURE haS SimiLaR EffECTS ON SUppRESSiON Of piNEaL aCTiViTy, iNdUCTiON Of maSkiNg,
aNd phaSE ShifTiNg
Redlin’s (2001) detailed review describes numer-ous points of similarity between light suppression of pineal activity and masking. Of particular impor-tance for the present discussion is the fact that mela-tonin suppression in rodents is initiated by light and persists well beyond the actual presence of light. The “duration” characteristic is important because it also exists for locomotor suppression, but it has received only limited investigative attention or discussion (Antle et al., 2007; Lall and Biello, 2003a; Redlin, 2001). Informal observations (Fig. 1B in Redlin, 2001) show that a light pulse lasting only 15 min is suffi-cient to induce locomotor suppression that endures for more than an hour. A 1-min light pulse also sup-presses melatonin for about the same amount of time (Fig. 1A in Redlin, 2001). The responses of the 2 vari-ables to identical stimuli have not been studied in sufficient depth to know the extent of their similarity.
Exploration of light-induced suppression of pineal function in nocturnal rodents has also demonstrated its persistence in response to mere seconds of light exposure, which is sufficient to elicit a rapid decline in pineal activity (Fig. 2; as indicated by the amount of melatonin or N-acetyltransferase [NAT]) (Illnerova
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from
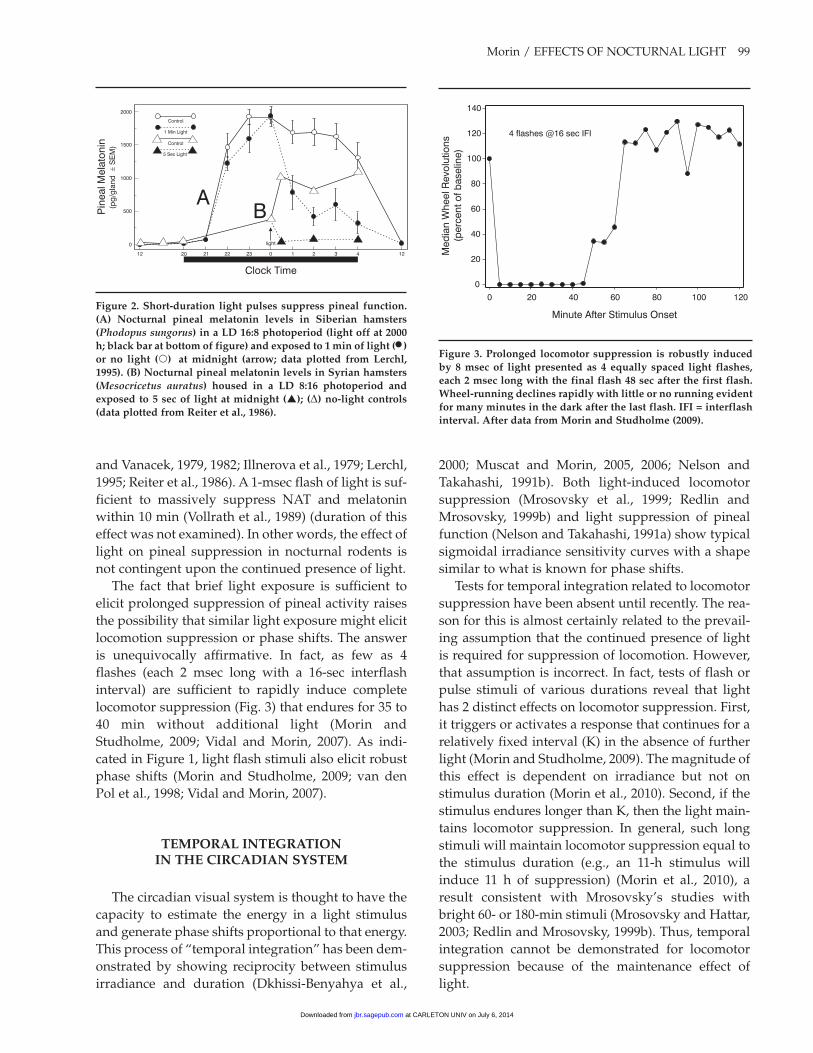
Morin / EFFECTS OF NOCTURNAL LIGHT 99
and Vanacek, 1979, 1982; Illnerova et al., 1979; Lerchl, 1995; Reiter et al., 1986). A 1-msec flash of light is suf-ficient to massively suppress NAT and melatonin within 10 min (Vollrath et al., 1989) (duration of this effect was not examined). In other words, the effect of light on pineal suppression in nocturnal rodents is not contingent upon the continued presence of light.
The fact that brief light exposure is sufficient to elicit prolonged suppression of pineal activity raises the possibility that similar light exposure might elicit locomotion suppression or phase shifts. The answer is unequivocally affirmative. In fact, as few as 4 flashes (each 2 msec long with a 16-sec interflash interval) are sufficient to rapidly induce complete locomotor suppression (Fig. 3) that endures for 35 to 40 min without additional light (Morin and Studholme, 2009; Vidal and Morin, 2007). As indi-cated in Figure 1, light flash stimuli also elicit robust phase shifts (Morin and Studholme, 2009; van den Pol et al., 1998; Vidal and Morin, 2007).
TEmpORaL iNTEgRaTiON iN ThE CiRCadiaN SySTEm
The circadian visual system is thought to have the capacity to estimate the energy in a light stimulus and generate phase shifts proportional to that energy. This process of “temporal integration” has been dem-onstrated by showing reciprocity between stimulus irradiance and duration (Dkhissi-Benyahya et al.,
2000; Muscat and Morin, 2005, 2006; Nelson and Takahashi, 1991b). Both light-induced locomotor suppression (Mrosovsky et al., 1999; Redlin and Mrosovsky, 1999b) and light suppression of pineal function (Nelson and Takahashi, 1991a) show typical sigmoidal irradiance sensitivity curves with a shape similar to what is known for phase shifts.
Tests for temporal integration related to locomotor suppression have been absent until recently. The rea-son for this is almost certainly related to the prevail-ing assumption that the continued presence of light is required for suppression of locomotion. However, that assumption is incorrect. In fact, tests of flash or pulse stimuli of various durations reveal that light has 2 distinct effects on locomotor suppression. First, it triggers or activates a response that continues for a relatively fixed interval (K) in the absence of further light (Morin and Studholme, 2009). The magnitude of this effect is dependent on irradiance but not on stimulus duration (Morin et al., 2010). Second, if the stimulus endures longer than K, then the light main-tains locomotor suppression. In general, such long stimuli will maintain locomotor suppression equal to the stimulus duration (e.g., an 11-h stimulus will induce 11 h of suppression) (Morin et al., 2010), a result consistent with Mrosovsky’s studies with bright 60- or 180-min stimuli (Mrosovsky and Hattar, 2003; Redlin and Mrosovsky, 1999b). Thus, temporal integration cannot be demonstrated for locomotor suppression because of the maintenance effect of light.
0
500
1000
1500
2000
Pin
eal M
ela
ton
in(p
g/g
lan
dS
EM
)�
12 20 21 22 23 0 1 2 3 4 12
Control
1 Min Light
Control
5 Sec Light
light
AB
Clock Time
figure 2. Short-duration light pulses suppress pineal function. (a) Nocturnal pineal melatonin levels in Siberian hamsters (Phodopus sungorus) in a Ld 16:8 photoperiod (light off at 2000 h; black bar at bottom of figure) and exposed to 1 min of light (•) or no light () at midnight (arrow; data plotted from Lerchl, 1995). (b) Nocturnal pineal melatonin levels in Syrian hamsters (Mesocricetus auratus) housed in a Ld 8:16 photoperiod and exposed to 5 sec of light at midnight (); (D) no-light controls (data plotted from Reiter et al., 1986).
Minute After Stimulus Onset
0 20 40 60 80 100 120
0
20
40
60
80
100
120
140
Med
ian
Wh
eel R
evo
lutio
ns
(perc
en
t o
f b
aselin
e)
4 flashes @16 sec IFI
figure 3. prolonged locomotor suppression is robustly induced by 8 msec of light presented as 4 equally spaced light flashes, each 2 msec long with the final flash 48 sec after the first flash. wheel-running declines rapidly with little or no running evident for many minutes in the dark after the last flash. ifi = interflash interval. after data from morin and Studholme (2009).
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

100 JOURNAL OF BIOLOGICAL RHYTHMS / April 2013
The maintenance effect may also confound inter-pretation of studies designed to test temporal inte-gration with respect to phase shift responses to light pulses (Morin et al., 2010; Nelson and Takahashi, 1991b; see Morin et al., 2010, for a complete discus-sion).
Temporal integration of photic stimuli is absent when either phase shift or locomotor suppression responses to millisecond stimuli are tested (Morin and Studholme, 2009; Vidal and Morin, 2007). For example, a series of 100 flashes (2 msec each) does not yield either phase shifts or locomotor suppres-sion equal to those in response to 10 flashes at 10 times the energy per flash (Morin et al., 2010; Vidal and Morin, 2007). In other words, 2 stimulus sequences having equal energy do not elicit equal responses for either phase shifting or suppression of locomotion. Similar studies concerning temporal integration and suppression of pineal activity are lacking.
TOwaRd a SyNThESiS Of SCN fUNCTiON: aCUTE phOTiC mOdULaTiON
Of mULTipLE SySTEmS
To this point, the similarities between photic regu-lation of circadian rhythm phase, locomotor suppres-sion, and pineal suppression have been discussed, and the fact that light-induced locomotor suppres-sion is actually an index of a positive change, namely sleep induction, has been acknowledged. These facts alter appreciation of how photic information must interact with SCN activity to regulate multiple bodily functions, not just circadian rhythm phase. One pre-diction is that the initial events in the light-activated sequence are likely to be same regardless of whether the response of interest is photosomnolence, phase shifting, or pineal hormone suppression. At some location, presumably in the SCN circuitry, there will be 1 or more points of pathway divergence that lead to expression of the 3 outputs. Each output response would theoretically be subject to idiosyncratic modi-fication at or beyond the point of pathway diver-gence.
It is parsimonious to conclude that light triggers 1 event sequence in the SCN to initiate all 3 responses. Therefore, a search for light-dependent changes in the SCN having time constants similar to the interval, K, is likely to be a worthwhile endeavor. One class of physiological processes known to be activated by light during the subjective night is induction of SCN
gene expression. The immediate early gene product FOS is induced in cells throughout the SCN by light pulses or a few millisecond flashes (Aronin et al., 1990; Kornhauser et al., 1990; Vidal and Morin, 2007). In the SCN, the region of FOS expression corresponds to that of retinal input (Hattar et al., 2006; Morin et al., 2006). It should be apparent that light will induce FOS or other gene products in the SCN irrespective of whether the behavioral or physiological response being measured is photosomnolence, phase shifting, or suppression of pineal activity. As for the photic input pathway, all retinorecipient SCN cells are likely to participate in regulation of all SCN functions, and the separation of functions occurs further down-stream.
Most studies of light and SCN function focus exclusively on issues relating to circadian rhythm entrainment without consideration of the possibility that photic information affecting the SCN affects multiple functions via a common mechanism. As a result, there has been no attempt to establish the point or points at which the several SCN-mediated outputs diverge. It should be possible to obtain such information using a combination of neuroanatomical and electrophysiological methods to determine intra-SCN circuitry. One investigation has already nar-rowed the scope of the issue by showing that nocturnal light can induce FOS and Per1 in SCN neurons even when phase shifts are blocked by a nonphotic stimulus (Edelstein et al., 2003; but see Maywood et al., 1999; Yokota et al., 2000). Thus, the location at which the nonphotic input is acting must be downstream of the signal transduction cascade promoting expression of those genes. It remains to be seen whether the same nonphotic stimulus would block light-induced suppression of pineal activity or some other response to light.
LighT-iNdUCEd LOCOmOTOR SUppRESSiON aNd phaSE ShifTS aRE LiNkEd
The recent evidence that a single stimulus induces both phase shifts and photosomnolence highlights an issue simmering in the circadian rhythm literature. It concerns whether wheel running, a standard behav-ior used to evaluate rhythmicity, is causally related to generation of light-induced phase shifts. A number of experiments now support the principle that non-photic stimuli (enforced locomotion being one of several types) greatly reduce or abolish the phase shifting effects of light (Edelstein et al., 2003;
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

Morin / EFFECTS OF NOCTURNAL LIGHT 101
Mistlberger and Antle, 1998; Mistlberger and Holmes, 1999; Ralph and Mrosovsky, 1992). In other words, the data show that locomotor activity must be sup-pressed (or photosomnolence induced) in order for light to maximally elicit phase shifts.
Importantly, the foregoing studies suggest a path-way by which locomotion suppresses light-induced phase shifts. Light and behavioral activity oppose each other at the SCN cellular level (Schaap and Meijer, 2001). Locomotion (or a correlate) (Mistlberger et al., 2003) is thought to modify SCN function via release of NPY from terminals of IGL neurons (see Yannielli and Harrington, 2004, for a review). Light-induced phase shifts are blocked by NPY infusion onto the SCN (Gamble et al., 2005; Lall and Biello, 2002, 2003a, 2003b; Yannielli and Harrington, 2000). Several additional results are consistent with the idea that light initiates a prolonged sequence of events, mediated by the SCN, that promotes both locomotor suppression and phase shifts. These include the fact that running in a novel wheel induces NPY release in the SCN (Glass et al., 2010) and that both NPY treat-ment and novel wheel activity block light-induced phase shifts even when administered an hour after the photic stimulus (Lall and Biello, 2002, 2003a, 2003b; Yannielli and Harrington, 2000, 2001a, 2001b). Lall and Biello (2003b) also noted that individual NPY treatments simultaneously block or greatly attenuate both light-induced phase shifts and the duration of locomotor suppression. This observation is consistent with the view that the 2 effects of light are mediated by the SCN. In the case of pineal mela-tonin synthesis, direct infusion of NPY onto the SCN mimics the inhibitory effect of light (Gamble et al., 2006). It remains to be seen whether photic suppres-sion of pineal melatonin is or is not blocked by loco-motion. The interval during which NPY is able to antagonize nocturnal light effects may be related to the “early phase resetting” interval determined from SCN cell physiology and characterized by increasing spike frequency in VIP neurons and reduction of the potassium current (see Fig. 8 in Kuhlman et al., 2003).
CONCLUSiONS fROm STUdiES Of LighT-iNdUCEd LOCOmOTOR SUppRESSiON
Exposure of mice or hamsters to light during the early night suppresses locomotion, whether mea-sured as wheel running or open field activity. A vari-ety of studies have yielded the following conclusions:
1. Classic and pRGC photoreceptors contribute to loco-motor suppression.
2. The SCN is necessary for light-induced locomotor suppression.
3. Very brief light exposure is sufficient to induce loco-motor suppression.
4. Single light pulses must be more than 3 sec long to suppress locomotion.
5. An effective light stimulus “triggers” an interval, K, of locomotor suppression.
6. Long light exposure “maintains” the interval of loco-motor suppression.
7. Higher irradiance light pulses yield greater locomo-tor suppression.
8. Locomotor suppression by flashes does not display temporal integration.
9. Conclusions 1 through 8 are also true for light-induced phase shift responses.
10. Light induces rhythm phase shifts simultaneously with locomotor suppression.
11. Inactivity is necessary to obtain normal light-induced phase shifts.
The sole indication that the effect of light on rhythm phase differs from its effect on locomotor suppression comes from phase shift data which have been interpreted as suggesting that the phase shift mechanism shows temporal integration of light pulse energy. The difference between the phase shift and locomotor suppression results may lie with the con-found that long light pulses have both activating and maintenance effects (see (Morin et al., 2010, for a discussion). Given the numerous similarities between photic regulation of phase shifts and locomotor sup-pression, it is parsimonious to infer that phase shift magnitude is modified by light according to the same rules governing the duration and magnitude of loco-motor suppression.
LighT aCTS ThROUgh ThE SCN TO mOdify ThE aCTiViTy Of mULTipLE SySTEmS
Photic regulation of circadian rhythm phase and locomotor suppression appear to be regulated by similar activating and maintaining mechanisms, but the details of such responses are not identical. For example, subjective night is divided into delay and advance portions of the light-type phase response curve; there is no equivalent for locomotor suppres-sion (Redlin and Mrosovsky, 1999b). It is also possi-ble that melatonin suppression by light in the early night is regulated by an entirely different mechanism
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

102 JOURNAL OF BIOLOGICAL RHYTHMS / April 2013
than is active during the later night (Illnerova and Vanacek, 1982). At the very least, there are likely to be multiple regulatory circuits involved in the most basic light-initiated responses—a pathway intrinsic to the SCN for resetting circadian clock phase; SCN efferent pathways for suppressing locomotion and pineal function and for inducing sleep—and there should be an SCN-afferent route by which locomotor suppression/sleep enables light-induced phase shifts.
The SCN, as master circadian clock, dictates the harmony of many bodily oscillations through its efferent distribution of rhythm phase. It is more sur-prising that photic input to the SCN initiates addi-tional forms of physiological response, including suppression of pineal activity and melatonin produc-tion, as well as photosomnolence. The list of direct and indirect light-induced actions is expected to lengthen with additional research, especially given the presence of multiple pathways by which the SCN can modulate peripheral organ function (Bartness et al., 2001; Gerendai et al., 2009) and their role in regu-lation of rhythmicity in certain tissues but not others (Guo et al., 2006). A candidate for inclusion on this list is the adrenal corticosterone surge induced by nocturnal light (Ishida et al., 2005; Mohawk et al., 2007). The level of response is dependent upon stim-ulus irradiance, abolished by SCN lesions and medi-ated through SCN efferent activity in the splanchnic nerve, which provides input to the adrenal (Ueyama et al., 1999). Elevated ACTH does not mediate the effect. In addition, light pulses as short as 1 min are known to increase splanchnic nerve activity, which is also irradiance-dependent. A 10-min light pulse induces a steady increase in splanchnic nerve activity over at least 2 h which, once initiated by light, persists in the dark (Niijima et al., 1992, 1993). It is not yet known whether adrenal hormone synthesis responds to brief light exposure in the same manner as locomo-tor suppression or splanchnic nerve activity.
A new physiological event on the list of changes induced by nocturnal light is a rapid drop in mouse core body temperature (Tc; Fig. 4; see Studholme et al., 2013). The large, abrupt drop in Tc occurs in paral-lel with locomotor suppression and recovery. The pattern of Tc change, its induction by millisecond light flashes, the fact that it endures well beyond the presence of the stimulus, and that it is prolonged by long-duration light stimuli strongly suggest that the Tc response is regulated by the same mecha-nistic sequence triggering the other events. The
light-induced drop in Tc, along with photic control of corticosterone, must be considered central to a multi-faceted, whole body response involving, at the very least, the circadian visual, sleep, neuroendocrine, thermoregulatory, and autonomic nervous systems.
The suggestion that light initiates a time-limited event sequence common to its suppression of pineal function, induction of photosomnolence, phase shift-ing, augmented adrenal activity, and thermoregula-tory change is interesting but says little about how the brain might be organized to enable those responses. Figure 5 is offered as a guide for thought experiments regarding how photic information might act through the SCN to elicit the patterns of physiol-ogy and behavior discussed here. The figure does not imply any causal relation between presently known intrinsic SCN anatomy and efferent functional con-trol. In fact, recent research has revealed the SCN to be a plastic structure capable of organizing its own rhythmic internal activities to reflect the needs imposed by the prevailing photic environment (De la Iglesia et al., 2004; Karatsoreos et al., 2011; Lee et al., 2009; Liu et al., 2007; Watanabe et al., 2006). This being the case, what is considered “normal” SCN activity and efferent function must be accepted with caution. Perhaps more important is the fact that
-120 -100 -80 -60 -40 -20 0 20 40 60 80 100 120
35
36
37
38
39
0
10
20
30
40 10 flashes
Wh
eel R
evo
lutio
ns
Bo
dy T
em
pera
ture
Minute from First Flash
figure 4. Core body temperature (Tc; broken line) drops acutely in response to brief light exposure and remains reduced well beyond the presence of light. in this example, the stimulus con-sisted of 10 light flashes, each 2 msec long, distributed equally across a 5-min interval (gray vertical bar beginning at minute 0). The Tc change is temporally associated with suppression of loco-motion (solid line) but is not caused by it. The behavioral and physiological changes attain full recovery at approximately the same time, although Tc rises from its nadir well in advance of locomotion. morin and Studholme, unpublished data.
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

Morin / EFFECTS OF NOCTURNAL LIGHT 103
unexpected or seemingly outlandish light-induced responses (or lack thereof) are likely to provide invaluable clues as to ordinary function (Barakat et al., 2004; de Groot and Rusak, 2002; Erkert et al., 2006; Ruby et al., 2000).
aCkNOwLEdgmENT
Supported by NIH grant NS061804.
CONfLiCT Of iNTEREST STaTEmENT
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publica-tion of this article.
REfERENCES
Antle MC, Sterniczuk R, Smith VM, and Hagel K (2007) Non-photic modu-lation of phase shifts to long light pulses. J Biol Rhythms 22:524-533.
Aronin N, Sagar SM, Sharp FR, and Schwartz WJ (1990) Light regulates expression of a Fos-related protein in rat suprachias-matic nuclei. Proc Natl Acad Sci U S A 87:5959-5962.
Aschoff J (1960) Exogenous and endogenous compo-nents in circadian rhythms. Cold Spring Harb Symp Quant Biol 25:11-27.
Aschoff J (1981) Freerunning and entrained circadian rhythms. In Handbook of Behavioral Neurobiology, Vol 4, Aschoff J, ed, pp 81-95. New York: Plenum.
Barakat MT, O’Hara BF, Cao VH, Larkin JE, Heller HC, and Ruby NF (2004) Light pulses do not induce c-fos or per1 in the SCN of hamsters that fail to reen-train to the photocycle. J Biol Rhythms 19:287- 297.
Bartness TJ, Song CK, and Demas GE (2001) SCN efferents to peripheral tissues: implications for biological rhythms. J Biol Rhythms 16:196-204.
Berson DM, Dunn FA, and Takao M (2002) Phototransduction by retinal ganglion cells that set the circadian clock. Science 295:1070-1073.
de Groot MH and Rusak B (2002) Entrainment impaired, masking spared: an apparent genetic abnormality that prevents circadian rhythm entrainment to 24-h lighting cycles in California mice. Neurosci Lett 327:203-207.
De la Iglesia HO, Cambras T, Schwartz WJ, and ez-Noguera A (2004) Forced desynchronization of dual circadian oscillators within the rat suprachiasmatic nucleus. Curr Biol 14:796-800.
Dkhissi-Benyahya O, Sicard B, and Cooper HM (2000) Effects of irradiance and stimulus duration on early gene expression (Fos) in the suprachiasmatic nucleus: temporal summation and reciprocity. J Neurosci 20:7790-7797.
Ecker JL, Dumitrescu ON, Wong KY, Alam NM, Chen SK, LeGates T, Renna JM, Prusky GT, Berson DM, and
daynight
Light-Triggered
Event Sequence
light
RHT
SCN Mechanisms
Locomotion
Output Rhythms
Additive
Maintenance
Effects
IGL
Rhythm Phase
Adjustment
Core Temperature
Other
Autonomic?
NPY
Light Effects
2 hr
Increase
Sharp
Drop &
Sleep
?
Corticosterone
Melatonin
Sharp
Drop
Sharp
Drop &
Loss
GHT
NPY
K
PRC
figure 5. Theoretical relationships between the retinorecipient SCN, the master circadian clock in the SCN (the two not necessarily being different), and efferent circuits controlling the rhythms of locomotion/sleep, core temperature, pineal melatonin, and adrenal corticosterone. “Other auto-nomic” rhythmic outputs are predicted (see text). in this diagram, “rhythm phase adjustment” is also a rhythm as defined by the light-type phase response curve (pRC). The generation and photic control of the pRC are fully intrinsic to the SCN. The retinorecipient mechanism underlying the “light-triggered event sequence” (a presumed interval timer with duration = k) resides in the SCN and mediates light effects on all rhythms. additional light can prolong k via “additive mainte-nance effects.” These will be seen as larger phase shifts or longer intervals of photosomnolence. The diagram presumes that the irradiance-sensitive, fixed-interval response sequence is activated for all light effects indicated. The immediate effect of nocturnal light is to inhibit locomotion, cause an abrupt drop in core temperature, induce a phase shift, cause a rapid decline in melatonin, and facilitate adrenal corticosterone synthesis. The drop in nocturnal locomotion (or correspond-ing sleep induction), and conceivably the drop in core temperature, may act directly or indirectly through the igL to inhibit release of Npy in the SCN. Only in the absence of such release will light-induced phase shift and locomotor suppression responses be normal. The intra-SCN release of Npy may also alter other listed responses to light (see text).
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

104 JOURNAL OF BIOLOGICAL RHYTHMS / April 2013
Hattar S (2010) Melanopsin-expressing retinal ganglion-cell photoreceptors: cellular diversity and role in pattern vision. Neuron 67:49-60.
Edelstein K, De la Iglesia HO, Schwartz WJ, and Mrosovsky N (2003) Behavioral arousal blocks light-induced phase advances in locomotor rhythmicity but not light-induced Per1 and fos expression in the hamster supra-chiasmatic nucleus. Neuroscience 118:253-261.
Edelstein K and Mrosovsky N (2001) Behavioral responses to light in mice with dorsal lateral geniculate lesions. Brain Res 918:107-112.
Erkert HG, Gburek V, and Scheideler A (2006) Photic entrainment and masking of prosimian circadian rhythms (Otolemur garnettii, Primates). Physiol Behav 88:39-46.
Gamble KL, Ehlen JC, and Albers HE (2005) Circadian con-trol during the day and night: role of neuropeptide Y Y5 receptors in the suprachiasmatic nucleus. Brain Res Bull 65:513-519.
Gamble KL, Paul KN, Karom MC, Tosini G, and Albers HE (2006) Paradoxical effects of NPY in the suprachiasmatic nucleus. Eur J Neurosci 23:2488-2494.
Gerendai I, Toth IE, Boldogkoi Z, and Halasz B (2009) Recent findings on the organization of central nervous system structures involved in the innervation of endo-crine glands and other organs; observations obtained by the transneuronal viral double-labeling technique. Endocrine 36:179-188.
Glass JD, Guinn J, Kaur G, and Francl JM (2010) On the intrinsic regulation of neuropeptide Y release in the mammalian suprachiasmatic nucleus circadian clock. Eur J Neurosci 31:1117-1126.
Gooley JJ and Saper CB (2003) A broad role for melanopsin in non-visual photoreception based on neuroanatomical evidence in rats. J Neurosci 23:7093-7106.
Goz D, Studholme K, Lappi DA, Rollag MD, Provencio I, and Morin LP (2008) Targeted destruction of photosen-sitive retinal ganglion cells with a saporin conjugate alters the effects of light on mouse circadian rhythms. PLoS One 3:e3153.
Guler AD, Ecker JL, Lall GS, Haq S, Altimus CM, Liao HW, Barnard AR, Cahill H, Badea TC, Zhao H, et al. (2008) Melanopsin cells are the principal conduits for rod-cone input to non-image-forming vision. Nature 453:102- 105.
Guo HG, Brewer JM, Lehman MN, and Bittman EL (2006) Suprachiasmatic regulation of circadian rhythms of gene expression in hamster peripheral organs: effects of transplanting the pacemaker. J Neurosci 26:6406-6412.
Harrington ME and Rusak B (1986) Lesions of the thalamic intergeniculate leaflet alter hamster circadian rhythms. J Biol Rhythms 1:309-325.
Hatori M, Le H, Vollmers C, Keding SR, Tanaka N, Schmedt C, Jegla T, and Panda S (2008) Inducible ablation of melanopsin-expressing retinal ganglion cells reveals their central role in non-image forming visual responses. PLoS One 3:e2451.
Hattar S, Kumar M, Park A, Tong P, Tung J, Yau KW, and Berson DM (2006) Central projections of melanopsin-expressing retinal ganglion cells in the mouse. J Comp Neurol 497:326-349.
Hattar S, Liao HW, Takao M, Berson DM, and Yau KW (2002) Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity. Science 295:1065-1070.
Hattar S, Lucas RJ, Mrosovsky N, Thompson S, Douglas RH, Hankins MW, Lem J, Biel M, Hofmann F, Foster RG, et al. (2003) Melanopsin and rod-cone photoreceptive systems account for all major accessory visual functions in mice. Nature 424:76-81.
Illnerova H and Vanacek J (1982) Complex control of the circadian rhythm in N-acetyltransferase activity in the rat pineal gland. In Vertebrate Circadian Systems, Aschoff J, Daan S, and Groos G, eds, pp 285-296. New York: Springer-Verlag.
Illnerova H and Vanecek J (1979) Response of rat pineal serotonin N-acetyltransferase to one min light pulse at different night times. Brain Res 167:431-434.
Illnerova H, Vanecek J, Krecek J, Wetterberg L, and Saaf J (1979) Effect of one minute exposure to light at night on rat pineal serotonin N-acetyltransferase and melatonin. J Neurochem 32:673-675.
Ishida A, Mutoh T, Ueyama T, Bando H, Masubuchi S, Nakahara D, Tsujimoto G, and Okamura H (2005) Light activates the adrenal gland: timing of gene expression and glucocorticoid release. Cell Metab 2:297-307.
Johnson RF, Moore RY, and Morin LP (1988a) Loss of entrainment and anatomical plasticity after lesions of the hamster retinohypothalamic tract. Brain Res 460:297-313.
Johnson RF, Morin LP, and Moore RY (1988b) Retinohypothalamic projections in the hamster and rat demonstrated using cholera toxin. Brain Res 462:301-312.
Karatsoreos IN, Butler MP, LeSauter J, and Silver R (2011) Androgens modulate structure and function of the suprachiasmatic nucleus brain clock. Endocrinology 152:1970-1978.
Klein DC and Moore RY (1979) Pineal N-acetyltransferase and hydroxyindole-O-methyltransferase: control by the retinohypothalamic tract and the suprachiasmatic nucleus. Brain Res 174:245-262.
Kornhauser JM, Nelson DE, Mayo KE, and Takahashi JS (1990) Photic and circadian regulation of c-fos gene expression in the hamster suprachiasmatic nucleus. Neuron 5:127-134.
Kuhlman SJ, Silver R, Le Sauter J, Bult-Ito A, and McMahon DG (2003) Phase resetting light pulses induce Per1 and persistent spike activity in a subpopulation of biological clock neurons. J Neurosci 23:1441-1450.
Lall GS and Biello SM (2002) Attenuation of phase shifts to light by activity or neuropeptide Y: a time course study. Brain Res 957:109-116.
Lall GS and Biello SM (2003a) Attenuation of circadian light induced phase advances and delays by neuropeptide Y and a neuropeptide Y Y1/Y5 receptor agonist. Neuroscience 119:611-618.
Lall GS and Biello SM (2003b) Neuropeptide Y, GABA and circadian phase shifts to photic stimuli. Neuroscience 120:915-921.
Lee ML, Swanson BE, and De la Iglesia HO (2009) Circadian timing of REM sleep is coupled to an oscillator within
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

Morin / EFFECTS OF NOCTURNAL LIGHT 105
the dorsomedial suprachiasmatic nucleus. Curr Biol 19:848-852.
Lehman MN, Silver R, Gladstone WR, Kahn RM, Gibson M, and Bittman EL (1987) Circadian rhythmicity restored by neural transplant: immunocytochemical character-ization of the graft and its integration with the host brain. J Neurosci 7:1626-1638.
Lerchl A (1995) Sustained response of pineal melatonin synthesis to a single one-minute light pulse during night in Djungarian hamsters (Phodopus sungorus). Neurosci Lett 198:65-67.
Li X, Gilbert J and Davis FC (2005) Disruption of masking by hypothalamic lesions in Syrian hamsters. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 191:23-30.
Liu AC, Welsh DK, Ko CH, Tran HG, Zhang EE, Priest AA, Buhr ED, Singer O, Meeker K, Verma IM, et al. (2007) Intercellular coupling confers robustness against muta-tions in the SCN circadian clock network. Cell 129:605-616.
Marchant EG and Morin LP (1999) The hamster circadian rhythm system includes nuclei of the subcortical visual shell. J Neurosci 19:10482-10493.
Maywood ES, Mrosovsky N, Field MD, and Hastings MH (1999) Rapid down-regulation of mammalian Period genes during behavioral resetting of the circadian clock. Proc Natl Acad Sci U S A 96:15211-15216.
Minors DS and Waterhouse J (1989) Masking in humans: the problem and some attempts to solve it. Chronobiol Int 6:29-53.
Mistlberger RE and Antle MC (1998) Behavioral inhibition of light-induced circadian phase resetting is phase and serotonin dependent. Brain Res 786:31-38.
Mistlberger RE, Antle MC, Webb IC, Jones M, Weinberg J, and Pollock MS (2003) Circadian clock resetting by arousal in Syrian hamsters: the role of stress and activ-ity. Am J Physiol Regul Integr Comp Physiol 285:R917-R925.
Mistlberger RE and Holmes MM (1999) Morphine-induced activity attenuates phase shifts to light in C57BL/6J mice. Brain Res 829:113-119.
Mohawk JA, Pargament JM, and Lee TM (2007) Circadian dependence of corticosterone release to light exposure in the rat. Physiol Behav 92:800-806.
Moore RY and Eichler VB (1972) Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res 42:201-206.
Morin LP and Blanchard JH (1999) Forebrain connections of the hamster intergeniculate leaflet: comparison with those of ventral lateral geniculate nucleus and retina. Vis Neurosci 16:1037-1054.
Morin LP, Blanchard JH, and Provencio I (2003) Retinal ganglion cell projections to the hamster suprachiasmatic nucleus, intergeniculate leaflet and visual midbrain: bifurcation and melanopsin immunoreactivity. J Comp Neurol 465:401-416.
Morin LP, Lituma PJ, and Studholme KM (2010) Two com-ponents of nocturnal locomotor suppression by light. J Biol Rhythms 25:197-207.
Morin LP and Pace L (2002) The intergeniculate leaflet, but not the visual midbrain, mediates hamster circadian
rhythm response to constant light. J Biol Rhythms 17:217-226.
Morin LP, Shivers KY, Blanchard JH, and Muscat L (2006) Complex organization of mouse and rat suprachias-matic nucleus. Neuroscience 137:1285-1297.
Morin LP and Studholme KM (2009) Millisecond light pulses make mice stop running, then display prolonged sleep-like behavior in the absence of light. J Biol Rhythms 24:497-508.
Morin LP and Studholme KM (2011) Separation of function for classical and ganglion cell photoreceptors with respect to circadian rhythm entrainment and induction of photosomnolence. Neuroscience 199:213-224.
Mrosovsky N (1999) Masking: history, definitions, and measurement. Chronobiol Int 16:415-429.
Mrosovsky N, Foster RG and Salmon PA (1999) Thresholds for masking responses to light in three strains of reti-nally degenerate mice. J Comp Physiol A Sens Neural Behav Physiol 184:423-428.
Mrosovsky N and Hattar S (2003) Impaired masking responses to light in melanopsin-knockout mice. Chronobiol Int 20:989-999.
Mrosovsky N, Lucas RJ, and Foster RG (2001) Persistence of masking responses to light in mice lacking rods and cones. J Biol Rhythms 16:585-588.
Mrosovsky N, Salmon PA, Foster RG, and McCall MA (2000) Responses to light after retinal degeneration. Vis Res 40:575-578.
Muscat L, Huberman AD, Jordan CL, and Morin LP (2003) Crossed and uncrossed retinal projections to the ham-ster circadian system. J Comp Neurol 466:513-524.
Muscat L and Morin LP (2005) Binocular contributions to the responsiveness and integrative capacity of the circa-dian rhythm system to light. J Biol Rhythms 20:513-525.
Muscat L and Morin LP (2006) Intergeniculate leaflet: con-tributions to photic and non-photic responsiveness of the hamster circadian system. Neuroscience 140:305-320.
Nelson DE and Takahashi JS (1991a) Comparison of visual sensitivity for suppression of pineal melatonin and cir-cadian phase-shifting in the golden hamster. Brain Res 554:272-277.
Nelson DE and Takahashi JS (1991b) Sensitivity and inte-gration in a visual pathway for circadian entrainment in the hamster (Mesocricetus auratus). J Physiol (Lond) 439:115-145.
Niijima A, Nagai K, Nagai N, and Akagawa H (1993) Effects of light stimulation on the activity of the autonomic nerves in anesthetized rats. Physiol Behav 54:555-561.
Niijima A, Nagai K, Nagai N, and Nakagawa H (1992) Light enhances sympathetic and suppresses vagal out-flows and lesions including the suprachiasmatic nucleus eliminate these changes in rats. J Auton Nerv Syst 40:155-160.
Panda S, Nayak SK, Campo B, Walker JR, Hogenesch JB, and Jegla T (2005) Illumination of the melanopsin sig-naling pathway. Science 307:600-604.
Panda S, Provencio I, Tu DC, Pires SS, Rollag MD, Castrucci AM, Pletcher MT, Sato TK, Wiltshire T, Andahazy M, et al (2003) Melanopsin is required for non-image-forming photic responses in blind mice. Science 301:525-527.
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from

106 JOURNAL OF BIOLOGICAL RHYTHMS / April 2013
Pickard GE, Ralph MR, and Menaker M (1987) The interge-niculate leaflet partially mediates effects of light on cir-cadian rhythms. J Biol Rhythms 2:35-56.
Pickard GE and Turek FW (1982) Splitting of the circadian rhythm of activity is abolished by unilateral lesions of the suprachiasmatic nuclei. Science 215:1119-1121.
Qiu X, Kumbalasiri T, Carlson SM, Wong KY, Krishna V, Provencio I, and Berson DM (2005) Induction of photo-sensitivity by heterologous expression of melanopsin. Nature 433:745-749.
Ralph MR and Mrosovsky N (1992) Behavioral inhibition of circadian responses to light. J Biol Rhythms 7:353-359.
Redfern P, Minors D, and Waterhouse J (1994) Circadian rhythms, jet lag, and chronobiotics: an overview. Chronobiol Int 11:253-265.
Redlin U (2001) Neural basis and biological function of masking by light in mammals: suppression of melatonin and locomotor activity. Chronobiol Int 18:737-758.
Redlin U, Cooper HM, and Mrosovsky N (2003) Increased masking response to light after ablation of the visual cortex in mice. Brain Res 965:1-8.
Redlin U and Mrosovsky N (1999a) Masking by light in hamsters with SCN lesions. J Comp Physiol A Sens Neural Behav Physiol 184:439-448.
Redlin U and Mrosovsky N (1999b) Masking of locomotor activity in hamsters. J Comp Physiol A Sens Neural Behav Physiol 184:429-437.
Redlin U, Vrang N, and Mrosovsky N (1999) Enhanced masking response to light in hamsters with IGL lesions. J Comp Physiol A Sens Neural Behav Physiol 184:449-456.
Reiter RJ, Joshi BN, Heinzeller T, and Nurnberger F (1986) A single 1- or 5-second light pulse at night inhibits ham-ster pineal melatonin. Endocrinology 118:1906-1909.
Rensing L (1989) Is “masking” an appropriate term? Chronobiol Int 6:297-300.
Ruby NF, Dubocovich ML and Heller HC (2000) Siberian hamsters that fail to reentrain to the photocycle have suppressed melatonin levels. Am J Physiol Regul Integr Comp Physiol 278:R757-R762.
Rusak B (1977) The role of the suprachiasmatic nuclei in the generation of circadian rhythms in the golden hamster, Mesocricetus auratus. J Comp Physiol 118:145-164.
Rusak B and Morin LP (1976) Testicular responses to pho-toperiod are blocked by lesions of the suprachiasmatic nuclei in golden hamsters. Biol Reprod 15:366-374.
Schaap J and Meijer JH (2001) Opposing effects of behav-ioural activity and light on neurons of the suprachias-matic nucleus. Eur J Neurosci 13:1955-1962.
Stephan FK and Zucker I (1972) Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proc Natl Acad Sci U S A 69:1583-1586.
Stetson MH and Watson-Whitmyre M (1976) Nucleus suprachiasmaticus: the biological clock in the hamster? Science 191:197-199.
Studholme KM, Gompf HS, and Morin LP (2013) Brief light stimulation during the mouse nocturnal activity phase simultaneously induces a decline in core temperature and locomotor activity followed by EEG-determined sleep. Am J Physiol Regul Integr Comp Physiol Published online 30 January 2013.
Thompson S, Foster RG, Stone EM, Sheffield VC, and Mrosovsky N (2008) Classical and melanopsin photore-ception in irradiance detection: negative masking of locomotor activity by light. Eur J Neurosci 27:1973- 1979.
Ueyama T, Krout KE, Van Nguyen X, Karpitskiy V, Kollert A, Mettenleiter TC, and Loewy AD (1999) Suprachias-matic nucleus: a central autonomic clock. Nat Neurosci 2:1051-1053.
van den Pol AN, Cao V, and Heller HC (1998) Circadian system of mice integrates brief light stimuli. Am J Physiol Regul Integr Comp Physiol 275:R654-R657.
Vidal L and Morin LP (2007) Absence of normal photic integration in the circadian visual system: response to millisecond light flashes. J Neurosci 27:3375-3382.
Vollrath L, Seidel A, Huesgen A, Manz B, Pollow K, and Leiderer P (1989) One millisecond of light suffices to suppress nighttime pineal melatonin synthesis in rats. Neurosci Lett 98:297-298.
Watanabe T, Naito E, Nakao N, Tei H, Yoshimura T, and Ebihara S (2006) Bimodal clock gene expression in mouse suprachiasmatic nucleus and peripheral tissues under a 7-hour light and 5-hour dark condition. J Biol Rhythms 22:58-68.
Yannielli P and Harrington ME (2004) Let there be “more” light: enhancement of light actions on the circadian sys-tem through non-photic pathways. Prog Neurobiol 74:59-76.
Yannielli PC and Harrington ME (2000) Neuropeptide Y applied in vitro can block the phase shifts induced by light in vivo. Neuroreport 11:1587-1591.
Yannielli PC and Harrington ME (2001a) Neuropeptide Y in the mammalian circadian system: effects on light-induced circadian responses. Peptides 22:547- 556.
Yannielli PC and Harrington ME (2001b) The neuropeptide YY5 receptor mediates the blockade of “photic-like” NMDA-induced phase shifts in the golden hamster. J Neurosci 21:5367-5373.
Yokota S, Horikawa K, Akiyama M, Moriya T, Ebihara S, Komura G, Ohta T, and Shibata S (2000) Inhibitory action of brotizolam on circadian and light-induced Per1 and Per1 expression in the hamster suprachias-matic nucleus. Br J Pharmacol 131:1739-1747.
at CARLETON UNIV on July 6, 2014jbr.sagepub.comDownloaded from
Recommended

















