
KOBIE (Serie Paleoantropología). Bilbao Bizkaiko Foru Aldundia-Diputación Foral de Bizkaia N.º XXVII, pp. 5-49, año 200317. ISSN 0214-7971
MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR)
(GAUTEGUIZ-ARTEAGA, BIZKAIA)
A microvertebrate assemblage from the Upper Pleistocene site of Antoliñako Koba (Gauteguiz-Arteaga, Biscay Province, Basque Country)
RESUMEN
H. Zubeldia ( 1) X. Murelaga (1)
S. Bailon (2) M. Aguirre (3)
X. Saez de Lafuente (1).
En este trabajo se describe la asociación de microvertebrados del yacimiento del Pleistoceno superior de Antoliñako Koba. Se han descrito cuatro unidades estratigráficas siendo la edad de la más joven de 10220 ± 40 B.P, mientras que, la más antigua se ha datado en 29.990 ± 230 B.P. La asociación de microvertebrados está representada por 13 especies de micromamíferos, 4 de reptiles y 3 de anfibios. La variación de la temperatura y humedad estimada en base a la asociación de micromamíferos presente en Antoliñako Koba es muy similar a las descritas en otros yacimientos del Pleistoceno superior. Los niveles que presentan una menor diversidad coinciden con los niveles más calidos mientras que en los niveles más fríos la diversidad taxonómica es mayor. Estas relaciones pueden estar condicionadas a la actividad del depredador que generó la acumulación de los restos de microvertebrados.
Palabras clave: Vertebrados, Pleistoceno superior, Cornisa Cantábrica.
ABSTRACT
In this work is described a microvertebrate assemblage from the Upper Pleistocene site of Antoliñako Koba. There are four stratigraphical units, the age of the younger one is 10220 ± 40 B.P. while the oldest one is 29.990 ± 230 B.P. The microvertebrate fossil assemblage is represented by 13 micromammal species, 4 reptilians and 3 amphibians. The temperature and the humidity variations estimated on the basis of the micromammal assemblage of Antoliñako Koba is very similar to that inferred from other sites of the Upper Pleistocene. The levels with low taxonomic diversity correspond to the most temperate periods, whereas higher diversity occurs in levels corresponding to cooler periods. This difference could be caused by the predator that produced the microvertebrate bone accumulation.
Key words: Vertebrates, Upper Pleistocene, Cantabrian Basin.
(1) Universidad del País Vasco/EHU, Facultad de Ciencia y Tecnología, Departamento de Estratigrafía y Paleontología, Apartado 644, E-48080 Bilbao. [email protected]; [email protected]; [email protected]
(2) Laboratoire départemental de Préhistoire du Lazaret, Département de Préhistoire du M.N.H.N. 33bis Bd Franck Pilatte, 06300 Nice, France. [email protected]
(3) UNED de Bergara. Plaza San Martín de Aguirre 4, Bergara, 20570, [email protected]

6 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
LABURPENA
Lan honetan Goi Pleistozenoko Antoliñako Koba aztarnategiko mikroomodunen elkarketa deskribatzen da. Lau unitate estratigrafico deskribatu dira, gazteenaren adina 10220 ± 40 B .P delarik eta zaharrenaren adina 29.990 ± 230 B.P delarik. Mikroomodunen elkarketa mikrougaztunen 13 espeziez, narrastien 4 espeziez eta anfibioen 3 espeziez dago osatuta. Antoliñako Kobako mikrougaztunen elkarketaren arabera estimatutako temperatura eta hezetasun bariazioak beste Goi Pleistozenoko aztarnategietan deskribatutakoen oso antzekoa da. Aniztasun baxueneko nibelak garairik beroenekin bat egiten dute, eta aniztazun taxonomiko handieneko nibelak garairik hotzenekin bat egiten dute. Erlazio hauek mikroomodun fosil akumulazioa sortu duen harrapariaren aktibitatera baldintzatuta egon daitezke.
Hitz gakoak: Ornodunak, Goi Pleistozenoa, Kantaurialdea.
INTRODUCCIÓN
La primera cita que se realiza sobre Antoliñako Koba fue realizada por J. M. Barandiaran en 1925, aunque es en un corto artículo de 194 7 donde da breve cuenta de un sondeo efectuado en 1923 (BARANDIARAN, 1947), en el que se identifica su yacimiento paleolítico. La primera identificación de material Solutrense fue realizada por J-C López-Quintana en la revisión del material arqueológico procedente de Antoliñako Koba existente en los locales del antiguo Grupo de Espeleología de Gemika (AGUIRRE RUIZ DE GOPEGUI, 2000). Se halla recogido en la Carta Arqueológica de Vizcaya (MARCOS MUÑOZ, 1982).
Desde 1995, se llevan realizando trabajos de excavación dirigidos por Mikel Aguirre. Durante la campaña de 2005 se recogió sedimento de los niveles excavados hasta esa fecha y en el presente trabajo se presenta el estudio de los restos de microvertebrados encontrados en ese muestreo (ZUBELDIA, 2006). Las dataciones se han realizado con C14 sobre todo en restos óseos y vegetales y han situado el registro estudiado entre 10220 ± 40 B.P. los más jóvenes, hasta 29.990 ± 230 B.P. los más antiguos. En la actualidad no se ha terminado de excavar todo el relleno por lo que se desconoce la serie que falta hasta el sustrato calizo del Cretácico inferior.
En el muestreo se han reconocido 20 taxones diferentes de micromamíferos, reptiles y anfibios. Los micromamíferos están representados por Eliomys quercinus, Clethrionomys glareolus, Arvicola terrestris, Arvicola cf. sapidus, Microtus oeconomus, Microtus arvalis-agrestis, Chionomys nivalis, Terrícola sp., Apodemus sylvaticus-flavicollis, Sorex coronatus-araneus, Neomysfodiens-anomalus, Crocidura sp. y Talpa sp., Los reptiles por Anguis fragilis, cf.
Podareis sp., Vipera sp. o Natrix sp. y Coronel/a austriaca y los anfibios por Salamandridae indet, Rana temporaria-iberica y Bufo sp. Aunque raros, también se han identificado restos de peces y de aves.
Los trabajos que han descrito restos de macrovertebrados en la Comisa Cantábrica son muy abundantes (ALTUNA, 1972; CASTAÑOS, 1986), mientras que los de microvertebrados en general, son aún muy puntuales. En los últimos años el estudio de los micromamíferos ha aumentado considerablemente debido sobre todo a su valor como indicadores paleoambientales (SESÉ, 2005). En el caso de los reptiles y anfibios son todavía más escasos los trabajos que se dedican a la descripción de estos grupos. Y ello a pesar de su gran valor en la reconstrucción climática y del paisaje del pasado (BAILON y RAGE, 1992).
En los alrededores de Antoliñako Koba se han realizado pocos estudios en los que se engloban diferentes grupos de microvertebrados: mamíferos, reptiles y anfibios. Entre los trabajos de este tipo se encuentran los realizados en los yacimientos de Kobeaga (Ispaster) (MUREL,AGA ET AL., 2006) y ATXAGAKOA (Forna) (LOPEZ-QUINTANA ET AL., 2005).
SITUACIÓN GEOGRÁFICA
Antoliñako Koba se encuentra situada en el Reserva de la Biosfera de Urdaibai, en el margen oriental del estuario que forma el río Oka, en el barrio Muruetagane de la localidad de Gautegiz-Arteaga y tan solo a dos kilómetros del clásico yacimiento de Santimamiñe (Kortezubi). Se encuentra localizado en la vertiente sur del monte Arlanburu, muy cerca de su cima y a unos 300 metros sobre el nivel del mar. Este monte se encuentra al norte del sistema cárstico Atxa-
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
7
rre. Las coordenadas UTM de la cavidad son 30T 0528684 4801191, y las coordenadas geodésicas angulares son N 43º 21' 46.9" W 002º 38'45.5" (Figura 1).
La entrada a la cueva es de muy pequeñas dimensiones, y esta orientada al S-SW. Dentro, la cavidad se divide en dos galerías. La de la izquierda mide unos 30 metros, y la de la derecha unos 1 O metros, donde se están centrando las campañas de excavación. Las dos galerías están casi colmatadas de material sedimentario. Seguramente, el origen de la cueva sea una galería meandriforme y tangencial respecto a la superficie del monte, que durante su evolución abrió la entrada actual (AGUIRRE RUIZ DE GOPEGUI, 2000).
CONTEXTO GEOLÓGICO
La cueva se localiza en sedimentos carbonatados Urgonianos del Cretácico Inferior. El entorno de la cueva está compuesto por calizas wackestone o packestone pararrecifales de color gris. Estos barros carbonatados se depositaron en un medio de baja energía y con poca contaminación de terrígenos, como un lagoon (GARROTE RUIZ ET AL., 1992) (Figura 2).
Figura 2. Contexto Geológico del entorno de Antoliñako Koba.
KOBIE (Paleoantropología n.º 27), año 2003/07
Figura l. Situación geográfica del yacimiento de Antoliñako Koba.
La cueva se encuentra casi colmatada de sedimento cuaternario. La superficie irregular del sustrato calizo ha condicionado el relleno sedimentario y la continuidad lateral de las diferentes unidades estratigráficas. Además, en algunos puntos la erosión generada por reactivaciones del sistema cárstico y/o la bioturbación realizada por organismos como el Tejón (Metes meles) ha alte
rado la estratigrafía complicando aún más su estudio. La estratigrafía que se describe en el presente trabajo se refiere únicamente a los dos primeros metros excavados, ya que, cuando se realizó el muestreo en el 2005 esta era la porción de la serie que afloraba. En general, todos los estratos buzan en la misma dirección, hacia el NW y parece que la galería profundiza en esa misma dirección, pero al estar esta rellena de sedimento no se puede confirmar este aspecto.
De techo a muro va aumentando la proporción de costras estalagmíticas. Algunas de estas se han interpretado como cuerpos sedimentarios cementados. Para
• Triásko. Cretácicolnferior 1 D Jurásico O \retácico Medio-Superior
Elanlxobe fll Cuaternario

8 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
+ 1A/1Z + 2A/2Z + 4A/4Z + 6A/6Z + BA/BZ +
-50cm
-100
• NivelA •Nivele 11 Nivel E O Bloques O NivelB • NlvelD 53 Sustrato calizo
Figura 3. Corte estratigráfico del yacimiento, entre los cuadros lA, 2A, 4A, 6A, 8A y lZ, 2Z, 4Z, 6Z, 8Z (Modificado de Aguirre Ruiz de Gopegui, 2000).
que estos cuerpos se formen es necesario que previamente existiera un cuerpo sedimentario y una corriente de agua carbonatada, Las aguas saturadas en carbonatos precipitan el carbonato en los huecos existentes entre los sedimentos, dando cohesión a los mismos.
Otras costras son producto de precipitación química, sin la presencia de sedimentos. Estas son las estalagmitas, estalactitas y coladas estalagmíticas, compuestas por mineral de calcita. Por debajo de las mismas suelen encontrarse los cuerpos sedimentarios cementados descritos anteriormente. Las costras estalagmíticas indican épocas sin sedimentación o épocas hídricamente húmedas.
En la secuencia sedimentaria excavada hasta el verano de 2006 se han diferenciado 8 niveles estratigráficos, A (Lanc), B (Lgc), C (Lmc+Lmb), D (Lab+Smb), E (Sm*), F (Sm), G (A-C) y H (Lmk). De estos niveles, el estudio de microfauna se ha realizado en los niveles A, B, C y D (Figura 3).
- Nivel A (Lanc): Estratigráficamente se trata del nivel más joven y
se encuentra constituido por arcilla y limo de color negro, y por clastos angulosos de caliza. Su potencia es muy pequeña, entre 3 y 8 centímetros. Contiene abundantes fragmentos de carbón, acumulados antrópicamente y en este nivel se han recuperado algunos restos líticos (Magdalenienses o Azilienses), óseos y conchíferos.
Se ha realizado una datación radiométrica con el método del C'4, para la cual se ha utilizado un resto óseo de la parte inferior del nivel y ha proporcionado una edad de 10.220 ± 40 B.P. (Beta-215544) (Figura 4).
- Nivel B (Lgc): Nivel de limo gris, con numerosos clastos y blo
ques angulosos. El nivel buza hacia el NE, y su
estructura interna es muy compleja (AGUIRRE RUIZ DE GOPEGUI, 2000). La potencia es de 50 centímetros como máximo.
Se han enviado dos restos óseos para datar radiométricamente por el método del C'4 • La primera muestra proviene de la parte superior de este nivel y ha proporcionado una edad de 10.800 ± 40 B.P. (Beta-215543). La segunda proviene de la parte inferior del nivel en el cuadro 2A y ha dado una edad de 14.680 ± 80 B.P. (GrN 23.783) (AGUIRRE RUIZ DE GOPEGUI, 2000). El contacto con el nivel superior es bastante neto. (Figura 4)
- Nivel C (Lmb+Lmc): Nivel de limo marrón, de 25 centímetros de poten
cia en el cuadro 6Z. Este nivel se encuentra a su vez subdividido en dos subniveles; el inferior, C2 (Lmc), y el superior, Cl (Lmb).
El subnivel Cl es más oscuro que el infrayacente y esta constituido por limos marrones. Contiene muchos bloques de gran tamaño, y pocos fragmentos de menor tamaño. Según los datos arqueológicos, el subnivel tendría una edad comprendida entre los 16.500-18.000 B.P. (AGUIRRE RUIZ DE GOPEGUI, 2000). El contacto de este subnivel con el nivel B es neto y erosivo (Figura 4).
El subnivel C2 se separa del infrayacente nivel D por medio de un límite erosivo. Está constituido por limos marrones, de color más claro que los suprayacentes. Contiene unos pocos bloques de gran tamaño, y muchos fragmentos de menor tamaño. Está datado aplicando el método del C'4 a un resto óseo recuperado en el cuadro 6A en 19.280 ± 120 B.P. (GrN 23.785) (AGUIRRE RUIZ DE GOPEGUI, 2000) (Figura 4).
- Nivel D (Lab-Sab+Lmb): Se trata de un nivel bastante potente de arenas y
KOBIE (Paleoantropología n.º 27), ano 2003/07
---------· ---
,. 1
r ~

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
9
Profundidad (cm.) Cuadro BZ Cuadro 6Z Cuadro lD Cuadro SD 56
C1$ Ak-M2
62 ---10800 -40 B.P Ak-Ml
14580 ±70 B.P. 68 Ak-M4 14630 ±70 B.P.
14680 ±80 B.P. 74
10220 -40 b.p.
80 --- -16500±18000 B.P.
86
92
98
104
110
116
122
128
134
140
l46
152
• Nivel A 156 O Nivel B
164
B Nivel
• Nivel
e D
26710 ±180 B.P.---+ 27390 ±320 B.P.-
19020±120 B.P.
19280 ± 120 B. P.
Ak-M19 +-- 29990 ± 230 E
Ak-M21
Figura 4. Columna estratigráfica con la situación de las muestras y de las dataciones realizadas.
limos. Dentro de este nivel se observan variaciones laterales y verticales de granulometría, compacidad y color. En los cuadros 3D y SD, la potencia es de 70 centímetros, pero el valor aumenta hacia el N. La compacidad es menor respecto al nivel anterior, y dentro del nivel disminuye a medida que subimos estratigráficamente.
Este Nivel se encuentra subdividido en tres subniveles; D3 (Lmbk inf. y Sm), D2 (Lmbk sup.) y Dl (Lab-Sab) (Figura 4).
El límite entre los niveles Dl y el C2 es erosivo, debido a que en toda la cueva aparecen muchas cubetas y canales entre ellos (AGUIRRE RUIZ DE GOPEGUI, 2000).
En el subnivel D2 se han encontrado abundantes restos fósiles, aunque estos, se encuentran muy fracturados y en mal estado de conservación. Se ha datado un resto óseo de este nivel proveniente del cuadro 8B utilizando el método del C14 y ha proporcionado una edad de 27.390 ± 320 B.P. (GrN 23.786) (AGUIRRE RUIZ DE GOPEGUI, 2000) (Figura 4).
KOBIE (Paleoantropología n.º 27), año 2003/07
El nivel D3 se ha datado en 29990 ± 230 b.p. (GrA-23898) aplicando el método C14 a un fragmento de carbón recogido en el cuadro 3D (Figura 4).
MATERIALES Y METODOLOGÍA
En este apartado se describen las técnicas utilizadas a la hora de estudiar los microvertebrados descritos en el presente trabajo.
En base al lugar donde se desarrollan las diferentes actividades se diferencian tres tipos de técnicas:
Técnicas de campo Dentro de estas técnicas se encuentran las utiliza
das a la hora de buscar los restos fósiles para su posterior estudio, la toma de muestras y su posterior lavado-tamizado.
En la mayoría de los casos, los restos de microvertebrados no se pueden ver sin la ayuda de aparatos ópticos, debido sobre todo a su pequeño tamaño. Por esta razón, hay que separar del sedimento los restos fósiles utilizando una lupa. Como no es posible revi-

10 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
Sigla Nivel Lito logia Talla (cm) Cuadro Sector PO (kg) Plavado (kg)
Ak-Ml A Lanc 67-74 8Z 8y9 2 0,56
Ak-M2 B Clg 56-62 6Z 3 2,1 0,51
Ak-M4 B Lgc 67-72 6Z 3 3,25 0,82
Ak-M5 B Lgc 72-76 6Z 3 3,21 0,79
Ak-M6 e Lmb 76-81 6Z 3 3,86 1,10
Ak-M9 e Lmc 89-94 6Z 3 3,77 0,91
Ak-Mll D Sab+Smc 107-110 1D 6 3,8 0,45
AK-M12 D Sab 110-115 1D 6 5,6 0,594
Ak-M14 D Sgb 120-125 1D 6 4,2 0,80
Ak-M16 D Sab 130-135 1D 6 5 0,85
Ak-M18 D Lmbk 142-147 lD 6 5,4 1,52
Ak-Ml9 D Smt 139-147 5D 7 6,98 0,72
Ak-M21 D Sat 151-156 5D 7 5,93 1,42
Tabla 1. Muestras estudiadas en Antoliñako Koba con indicación de la cantidad de sedimento que se ha recogido en cada una y la cantidad de concentrado resultante del cribado.
sar todo el sedimento con la lupa, lo que se hace es pasar por una criba el sedimento de manera que todas aquellas partículas de menor tamaño que las que estamos buscando serán arrastradas de la muestra por el
O Zona excavada hasta 2006
Bloques
~--=-:-=~~m ~ Sustrato rocoso
Figura 5. Plano general de Antoliñako Koba (Modificado de Aguirre Ruiz de Gopegui, 2000).
agua. De esta manera reducimos la cantidad de muestra que tendremos que revisar con la lupa.
La excavación del relleno sedimentario se realiza de arriba abajo, empezando por los niveles más recientes y hasta encontrar el sustrato rocoso. En el caso de Antoliñako Koba, todavía no se ha encontrado el sustrato rocoso. Este trabajo se basa en el estudio de la columna estratigráfica excavada hasta el año 2005. En el año 2006 aparecieron niveles más antiguos, en los cuadros 5D y 3D y todavía no se ha llegado al sustrato calizo por lo que, en este estudio solamente se presentan los resultados obtenidos de la revisión de los niveles superiores, que fueron el tema de la tesis de licenciatura presentada por uno de los firmantes de este trabajo (ZUBELDIA, 2006).
Junto a lo restos de micromamíferos, aparecen restos de macromamíferos, reptiles, anfibios y humanos. Por esa razón, se están realizando excavaciones arqueológicas siguiendo la metodología clásica. Toda la cueva se encuentra dividida en cuadros y sectores (Figura 5). Los cuadros ocupan un área de 1 m2
, y se identifican con una letra y un número. A su vez, cada cuadro está dividido en 9 sectores, siendo cada sector de 33 cm2, e identificado con un número. La excavación se va desarrollando de sector en sector, realizando tallas de unos 5 centímetros de profundidad. Todos los restos que se encuentran se sitúan en el espacio mediante un sistema de coordenadas X, Y, Z.
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
11
La recogida de sedimento para la extracción de microvertebrados se ha realizado en aquellos sectores que están más alejados de las paredes de la cavidad, ya que entre el sedimento y las paredes pueden existir circulaciones de agua que pueden remover los sedimentos y fósiles, causando contaminaciones entre los niveles. El sedimento se ha recogido a lo largo de toda la columna estratigráfica, para que esté representada toda la serie estratigráfica. El muestreo se ha realizado en un sector (33 cm2
), tomando tallas de unos 5 centímetros de espesor y 2-7 kg. de peso, empezando por los niveles más recientes estratigráficamente, y se continúa hacia los más antiguos. Para que toda la serie estratigráfica esté representada, se han tomado muestras en cuatro cuadros; 8Z, 6Z, ID y 5D (Figura 5). En el cuadro 8Z, las muestras se han tomado en los sectores 8 y 9, en el cuadro 6Z en el sector 3, en cuadro ID en el sector 6, y en el cuadro 5D en el sector 7.
Las partículas de arcilla suelen estar unidas entre ellas creando partículas de diferente tamaño que entorpecen el lavado de las muestras. Para que esto no ocurra, es muy importante que el sedimento recogido esté bien seco antes de lavarlo. De esta manera, las partículas de arcilla desfloculan con mayor facilidad a la hora del lavado.
Una vez que las muestras están bien secas estas se pesan para poder calcular el número de restos por kg. de sedimento que contiene cada una de ellas (Tabla 1).
Figura 6. Proceso de lavado del sedimento.
KOBIE (Paleoantropología n.º 27), año 2003/07
................................. " .............................. - ... ·---------
El sedimento secado previamente se introduce en cubos llenos de agua durante unas horas, para que los limos se disuelvan y los posibles bloques se disgreguen. Después se procede a su lavado, con el fin de eliminar la mayor cantidad de material estéril reduciendo así la cantidad de material a revisar con la lupa (Figura 6) (Tabla 1). En el estudio de los micromarníferos se suele utilizar como criba superior la de 4 mm de luz de malla ya que los elementos de talla superior a esta se pueden separar del sedimento sin tener que utilizar aparato óptico alguno. Las muestras de microvertebrados identificables más pequeñas suelen ser mayores de 0.5 mm por lo que la última criba por la que haremos pasar el sedimento tendrá una luz de malla de este tamaño. Una vez elegidas las cribas, se pone encima la de 4 mm de luz de malla y por debajo la de 0.5 mm. Posteriormente se vierte el sedimento, que previamente habíamos introducido en baldes con agua, encima de las cribas y se les aplica un chorro de agua para facilitar la eliminación de todas aquellas partículas menores de O. 5 mm. Después del lavado, el concentrado obtenido se extiende encima de plásticos, para proceder a su secado y posterior pesado (Tabla 1 ).
Técnicas de laboratorio Estas técnicas son las utilizadas desde que las
muestras llegan al laboratorio, hasta que están preparadas para su estudio.
Una vez que el concentrado lavado llega al laboratorio, se procede a pesarlo. Para facilitar la separación de los restos fósiles d~l estéril se divide el concentrado en cuatro fracciones diferentes; >4 mm., 4-2 mm., 2-1 mm. y 1-0, 5 mm. Para fraccionar el material se ha utilizado una tarnizadora electromagnética, modelo CISA RP.10. Luego se ha procedido al pesado de cada fracción.
Durante la campaña de 2005 se recogieron un total de 22 muestras que posteriormente fueron lavado-tamizadas. Para el presente estudio no se han estudiado todas las muestras recogidas, si no· que se han elegido las muestras más representativas de cada nivel. De las 22 muestras recogidas, se han estudiado 13; la 1, 2, 4, 5, 6, 9, 11, 12, 14, 16, 1~, 19 y 21, que se han denominado como AK-Ml, AK-M2, AK-M4, AK-M5, AK-M6, AK-M9, AK-Mll, AK-Ml2, AKM14, AK-M16, AK-M18, AK-M19 y AK-M21 (Figura 4).
El concentrado obtenido del lavado-tamizado está compuesto por fósiles y numeroso material estéril. Para separar los fósiles del material estéril en las fracciones más finas (2-1 mm., 1-0, 5 mm.) se ha utilizado una lupa binocular marca LEITZ WETZLAR del
----~---~---~--~~----~- ----------------

12 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
Mandíbula
Milxilar
Cráneo
!nc.!mvo
Molar
Fémur
Pelvis
Calcénoo
As!r.iga!o
Húmero
Radio
CUbllo
Escapula
Costi!la
Vertebra
Mata podio
Fa!ange
Otrcs
lndel
Yacimiento: Anto!iña Koba Nivel: Cuadro: Sección: Talla:
Total Enteres Fragmentos
Proximal
Central
Dista!
A
B
e
D
A
B
e
D
Departamento de Estratigrafía y Paleontología de la Facultad de Ciencia y Tecnología de la UPV/EHU. En las fracciones más gruesas no hace falta la lupa binocular. En este caso (>4 mm., 4-2 mm.), los fósiles se han separado del material estéril a simple vista. En total se han triado 11, 44 kg. de concentrado, los cuales se han obtenido del lavado de 55, 1 kg. de sedimento. Los fósiles recogidos se han cla-sificado en base a la muestra a la que per- 1 tenece, su fracción granulométrica, y su ubicación anatómica. En las muestras AK-M18 y AK-M21 no se han triado en su totalidad la fracción de 1-0.5 mm debido a la gran cantidad de concentrado existente. En estas fracciones se han recuperado los dientes de micromamíferos de la mitad de la muestra, y ningún resto óseo más.
Figura 8. Mediciones realizadas en los distintos tipos de molares; l. Glíridos.; 2. Arvicólidos.; 3. Múridos.; 4. Insectívoros.
Figura 7. Plantilla utilizada en el recuento de los diferentes elementos anatómicos recuperados. Se puede observar los tipos de fracturas de mandíbulas, maxilares, tibia-fíbula, cúbito, húmeros y fémur (Modificado de ANDREWS 1990).
Técnicas de gabinete: Estas técnicas son las utilizadas a la hora de estu
diar los fósiles una vez separados del material estéril. También se tienen en cuenta los parámetros utilizados en la nomenclatura, a la hora de tomar medidas y de realizar las fotografías y dibujos.
Las muestras recogidas en este yacimiento se han nominado con la sigla AK y el número que les corresponde. Al final se hace referencia a la fracción granulométrica de la muestra.
Los restos fósiles obtenidos en el triado han sido clasificados en base a su determinación taxonómica y posteriormente a su ubicación anatómica. La mayoría de los restos recuperados son molares y huesos postcraneales.
En el caso de los micromamíferos, con la excepción del género Talpidae, los restos óseos postcraneales tienen poco valor sistemático ya que son difíciles de clasificar taxonómicamente. De todas maneras, en el caso de los micromamíferos, se ha realizado el contaje de todos los elementos anatómicos ya que la proporción en la que se encuentran estos y su estado de conservación puede ser indicativo de la historia tafonómica de la tafocenosis (ANDREWS, 1990). En la figura 7 se puede observar la plantilla utilizada para realizar el recuento de los diferentes elementos anató-
L
t 1
IA 1
1
'
L
A
t M/1 ~.,--------.....,.--...,
:A 4 ~S·~ l'r ;A ~ ,,~ -:::J \ 1 ~-+--~
-----L
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
13
micos, En esta misma plantilla se indican los diferentes estadios de integridad de los maxilares y las mandíbulas (ANDREWS, 1990),
Los principales elementos anatómicos utilizados en la identificación de los micromamíferos han sido los molares, en el caso de los anfibios y reptiles, los dentarios, osteodermos, vértebras y pelvis, Los restos de aves y peces se han guardado sin clasificarlos sistemáticamente.
Las mediciones realizadas a los diferentes elementos recuperados son parámetros muy importantes a la hora de estudiar los microvertebrados. El estudio de los micromamíferos se basa en el tamaño y la morfología de la corona dentaria, y el tamaño de los elementos dentarios es un factor sistemático muy importante. Para realizar las mediciones se ha utilizado una lupa NIKON SMZ-U con una cámara digital Nikon DIGITAL SIGHT DS-Ll perteneciente al Departamento de Estratigrafía y Paleontología de la Facultad de Ciencia y Tecnología de la UPV/EHU.
En el caso de los molares pertenecientes a la familia Arvicolidae, las mediciones se han realizado únicamente en los Mil (VAN DER MEULEN, 1973). En estos molares se han medido la longitud, anchura y ACC máximas de la superficie oclusal (Figura 8). La longitud máxima corresponde a la distancia entre la parte posterior del primer lóbulo, hasta la parte anterior del molar. La anchura máxima corresponde a la distancia existente entre las dos líneas imaginarias que forman las puntas de los lóbulos de cada lado del molar. La ACC máxima corresponde a la distancia existente entre la parte posterior del cuarto lóbulo y la parte anterior del molar (Figura 8).
Para diferenciar las especies Arvicola sapidus y A. terrestris también se ha calculado el índice SDQ (HEINRICH, 1978). Para calcular este índice hay que medir el grosor que tiene el esmalte en el lado distal y mesial de cada triangulo y en al menos cinco triángulos del mismo molar. La fórmula que se utiliza para calcular este índice es la siguiente (HEINRICH, 1978);
SDQ = (LP!I:A)*JOO
A, corresponde al grosor del esmalte en el lado mesial o anterior de cada triangulo y P del lado distal o posterior.
Si el resultado da un valor superior a 100, el molar corresponde a la especie A. sapidus, y si es menor de 100, el molar corresponde a la especie A. terrestris (HEINRICH, 1978).
KOBIE (Paleoantropología n.º 27), año 2003/07
Figura 9. Esquema donde se representan donde se han realizado las mediciones para calcular el índice SDQ (Modificado de HEINRICH, 1982).
En los restos dentarios recuperados de los Muridae, Gliridae (DAAMS, 1981) y de los insectívoros (REUMER, 1984) se han medido la longitud (L) y anchura (A) máximas de la superficie oclusal en todos los molares identificados (Figura 8).
Para la nomenclatura utilizada en las descripciones de los dientes de arvicólidos se han seguido los trabajos de VAN DER MEULEN (1973) y de HEINRICH (1978), para los insectívoros el de REUMER (1984) y para los glíridos el de DAAMS (1981).
Para designar las piezas dentarias se han utilizado las siguientes abreviaturas: U: unicúspide, 1: incisivo, D: premolar deciduo, P: premolar definitivo, M: molar. Las piezas inferiores se hacen destacar intercalando una barra inclinada (/) entre el tipo de pieza y su número (p. e. Mil). Las piezas superiores se indican dejando el número de la pieza intercalado entre el tipo de pieza y la barra inclinada (p. e. Ml/).
En la descripción sedimentológica se ha utilizado la nomenclatura usada habitualmente en las campañas arqueológicas (SAÉNZ DE BURUAGA, ET AL., 1998). Normalmente se suelen utilizar tres letras.
La primera, en mayúsculas, define la fracción dominante; La fracción fina se define con las letras A (Arcilla), L (Limo) y S (Arena), la fracción intermedia se define con la letra G (Gravas), y la fracción gruesa con las letras C (Clastos) y B (Bloques).
La segunda, en minúsculas, define el color de la fracción fina; n (negro), m (marrón), a (amarillo) y g (gris).
La tercera letra, también en minúsculas, define otro tipo de características de los materiales; k (compactado) y t (suelto).

14 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
Después de estas tres letras, y en caso de ser necesario y separados con un guión, se definen las características antrópicas o diagenéticas de los materiales, en mayúsculas; H (Hogares), B (Brechas) ...
En el caso de que los materiales tiendan a cambiar a otros lateralmente, se refleja entre paréntesis (SAÉNZ DE BURUAGA, ET AL., 1998).
Todos los taxones de microvertebrados identificados en las muestras de Antoliñako Koba, tienen representantes vivos en la actualidad. Basándonos en el principio del actualismo, se presupone que las especies actuales viven en las mismas o parecidas condiciones ecológicas que sus antepasados. Para conocer lo requisitos ambientales de los micromamíferos se han utilizado los trabajos de PEMÁN (1985), SESÉ (2005), POKINES (1998) y CUENCA BESCÓS y GARCÍA PIMIENTA (en prensa). En el caso de los reptiles y anfibios los de HOLMAN (1998), BAILÓN (1999) y bases de datos provenientes de Internet.
Para las estimaciones sinecológicas se ha utilizado el índice de diversidad de Shannon-Weaver y curvas de Rarefracción Estos dos índices se han calculado con el programa "Past 3.1".
Las dataciones se han realizado utilizando el método C14 aplicado a restos óseos y carbón encontrados en diferentes niveles de la secuencia estratigráfica. Estas dataciones se han realizado en el laboratorio de Groningen de Holanda (GrA para dataciones AMS, y GrN para dataciones normales) y Beta Analytic de Florida.
El material estudiado en este trabajo procede de Antoliñako Koba y una vez se termine el estudio se depositarán en el Museo Etnográfico de Bilbao.
CONSIDERACIONES TAFONÓMICAS
La formación de un depósito de fósiles está estrechamente relacionado con los procesos físicos, químicos y orgánicos que en ella ocurren (ANDREWS, 1990). Estos procesos actúan desde que el organismo vive y muere, hasta que lo recuperamos como fósil. A la hora de interpretar la información que proporciona una asociación fósil de un determinado yacimiento, es necesario estudiar los procesos físicos, químicos y orgánicos que han actuado en el yacimiento y sobre los restos.
En las muestras recogidas se han estudiado los restos de microfauna, para las cuales es imprescindible aplicar técnicas de microscopía a la hora de su estudio. Por esta razón, las consideraciones tafonómicas se basan en el estudio de los microrestos (LÓPEZ-MARTÍNEZ, 1992).
Los depósitos de restos microscópicos pueden ser causados por procesos fluviales u orgánicos entre otros. Las acumulaciones de microvertebrados más habituales son las generadas por procesos orgánicos como las egagrópilas y coprolitos (MELLET, 1974; ANDREWS, 1990).
Las acumulaciones de restos fósiles de microfauna pueden ser consecuencia de diversos procesos, y
con el estudio del estado físico de los restos óseos, composición de la fauna y el estado de conservación de los distintos restos se pueden determinar esos procesos (PRATT, 1989).
El transporte sufrido por los restos acumulados en un sistema kárstico, puede ser estimado con la proporción de los diferentes elementos anatómicos conservados y el estudio de la fracturación que presentan estos (ANDREWS, 1990; ANDREWS & EVANS, 1983).
En las tanatocenosis creadas por agentes biológicos, los restos óseos y denta-
Figura 10. Aspecto general de los restos óseos recuperados en la muestra AK-M4.
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
15
-AK-M1 AK-M2
45% ~ AK-M4
40 -AK-M5
AK-M6
35 AK-M9
- AK-M11
30 -AK-M12 -AK-M14
25 -AK-M16 -AK-M18
20 -AK-M19 -AK-M21
15
10
5
o e<:! ~ o ta ti e<:! "' o o o o e<:! e<:!
~ > ~ :E (!) e;; ..... :.a ..E ~ 'i< ·¡¡:; o E ¡;: (!)
'9 ~ ..o (l.J •e<:! OI) E o:! ::::> o..
o:! -¡:; •(l.J ¡.L; p.. ..S:l o:! o::: e<:! 'O µ... ..... ·::l (J
e<:! ~ ¡;: 1 e<:! - ::e: "' Q ...... e<:! u "' u.:¡ ~ :E <r:
¡:::
rios suelen presentar marcas físicas y químicas producidas por el organismo creador del depósito. Las marcas físicas son las creadas por el depredador a la hora de cazar y comer su presa. Las químicas son las creadas principalmente por los jugos gástricos. En este tipo de acumulaciones, la proporción de algunas especies suele ser mayor, y también la proporción de diversos elementos anatómicos. Esto ocurre por la caza selectiva del depredador y las preferencias tróficas del mismo (MELLET, op. cit.; ANDREWS, 1990). La tanatocenosis creada por fluidos de agua presenta una mejor representación de la fauna del entorno donde se formo el depósito.
Las acumulaciones de microfósiles de los sistemas kársticos suelen tener origen
Figura 12. Porcentaje relativo de los diferentes ele-mentos anatómicos. La zona con sombreado indica las variaciones observadas en acumulaciones genera-das por varios depredadores actuales. Con línea con-tinua se indica la proporción en la que se encuentran las mismas piezas anatómicas en un yacimiento cárs-tico (Modificado de ANDREWS y EVANS, 1983).
KOBIE (Paleoantropologfa n.º 27), año 2003/07
ctl ..... ::l ~ ..e ·x "O ctl e ~ ctl ~
Figura 11. Riqueza relativa de los restos de microvertebrados en las diferentes muestras estudiadas.
mixto. Los restos se acumulan en la entrada o alrededores de la cueva por un organismo depredador, y más tarde, esos restos sufren un transporte al interior de la cueva por procesos físicos.
Los sedimentos que rellenan la cueva han sido transportados desde el exterior de la cavidad. En general, estos sedimentos son de granulometría fina, arcillas, limos y arenas finas, pero de vez en cuando, también aparece algún canto de tamaño con-glomerado. También se han
observado estructuras creadas por el agua, como canales, y superficies de erosión.
~ o o ctl (/) ..... ctl ~ o ..... Q.) .Q ..... :a e ·::; ::l :o > ctl Cl ::l Q) E .e o "O o. E ctl :::> Q5 ¡.= Q.) ·e:;; e o
ctl cr: •Q) t ·5 ~ ctl o. •::l o.. LL ctl (..)
~ e ctl
(/) I LL ....... UJ Q.)
~
---~------------
16 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
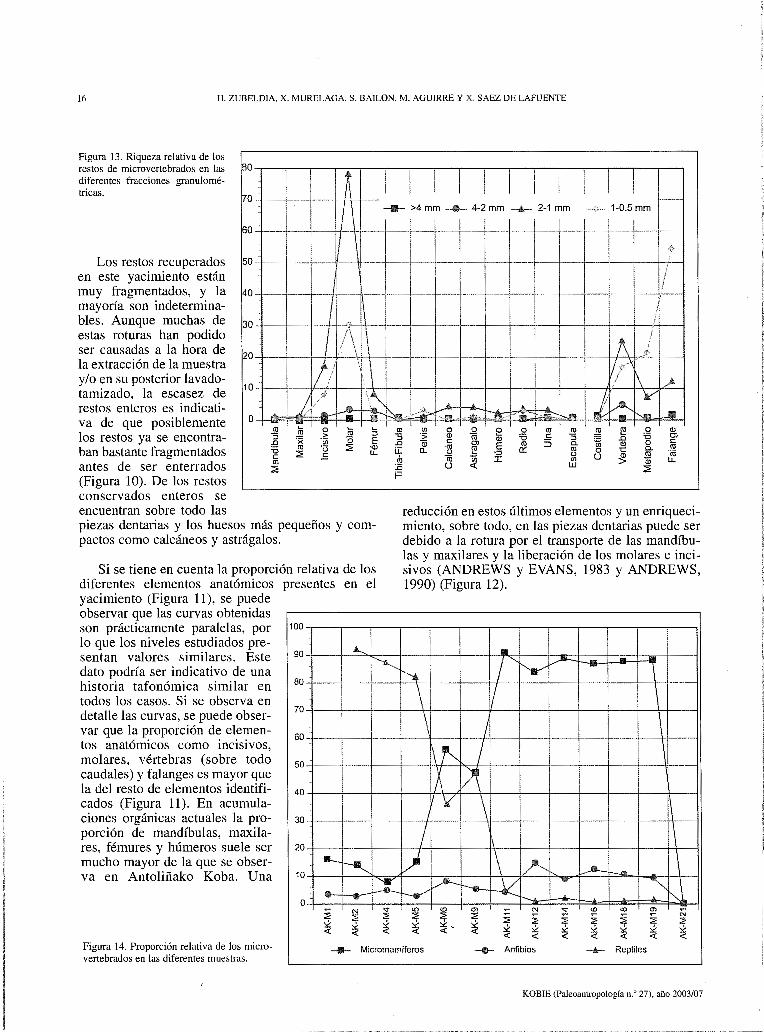
Figura 13. Riqueza relativa de los restos de microvertebrados en las diferentes fracciones granulométricas.
80
70
o
o
Los restos recuperados 50
en este yacimiento están muy fragmentados, y la mayoría son indeterminables. Aunque muchas de 30 estas roturas han podido ser causadas a la hora de 20 la extracción de la muestra
~
I I
1
I .. 1 l\
\
1//
1 1 1 1 1 1 1 1 1 1 1 ~
--it- >4 mm __._. 4-2 mm -ilr- 2-1 mm 1-0.5 mm
1 1
./-
!
! !
f\ j / "·'.
1\ > y/o en su posterior lavadotamizado, la escasez de 10
restos enteros es indicati j .ti 11 \~
"" ..
/b 1 A ~ . .. • .
va de que posiblemente los restos ya se encontra-ban bastante fragmentados antes de ser enterrados (Figura 10). De los restos conservados enteros se encuentran sobre todo las
o . - -- - .. ,¡;,
.... :;¡
E •QJ lL
piezas dentarias y los huesos más pequeños y compactos como calcáneos y astrágalos.
Si se tiene en cuenta la proporción relativa de los diferentes elementos anatómicos presentes en el
100
90 J...
~ - ""'··· -.¡,- - ... """ llllil ·1
Cll <J) o o e o Cll Cll ª ~ o QJ
::; ·:;; QJ ro (J) 'O e: ::; .n 'O Ol .n a; e: Ol Cll 5 o. ~ QJ o e: u: a.. ·ro ~ E o:: ro o t:'. c. Cll
u •:J ro ro ni ro 1ií u (.) :r: <J) lL ~ Qí :o ü <( UJ 2 F
reducción en estos últimos elementos y un enriquecimiento, sobre todo, en las piezas dentarias puede ser debido a la rotura por el transporte de las mandíbulas y maxilares y la liberación de los molares e incisivos (ANDREWS y EVANS, 1983 y ANDREWS, 1990) (Figura 12).
...
yacimiento (Figura 11), se puede observar que las curvas obtenidas son prácticamente paralelas, por lo que los niveles estudiados presentan valores similares. Este dato podría ser indicativo de una historia tafonómica similar en todos los casos. Si se observa en detalle las curvas, se puede observar que la proporción de elementos anatómicos como incisivos, molares, vértebras (sobre todo caudales) y falanges es mayor que la del resto de elementos identificados (Figura 11). En acumulaciones orgánicas actuales la proporción de mandíbulas, maxilares, fémures y húmeros suele ser mucho mayor de la que se observa en Antoliñako Koba. Una
~ /" ~..-- --- l
Figura 14. Proporción relativa de los microvertebrados en las diferentes muestras.
80
70
60
50
40
30
20
10
o
~
\ \
\"' X \(
/ 11.._
~ ) -:: v-9--~ -- -
1 CD :¡¡ N .... LO ::¡; ::;; ::¡; ::;;
'-"= '-"= '-"= '-"= '-"=' <( <( <( <( <(
-111- Micromamíferos
/ \ J \ / \
)\ \ 1
\ \ ~ \ ¡____._ -\/ -
'"""' ---- ----- . . ""-i "' ~ N .... <D "' "' ;;:¡ ::¡; i :¡¡ :¡¡ :¡¡ '-"=
::¡; ::¡; ::¡; <( '-"= '-"= '-"= '-"= '-"= '-"= '-"= <( <( <( <( <( <( <(
__._ Anfibios -4-- Reptiles
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLJÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
17
1400
1200
1000
Restos/kg sedimento
800
600
400
200
o T""
~ 1 ~ <(
N ~
1 ~ <(
~ l.()
~ ~ 1 1 ~ ~ <( <(
<O O) T"" N ~ ~ T"" T""
1 1 ~ ~ ~ ~ 1 1 <( <( ~ ~ <( <(
Si se observan las curvas de las distintas fracciones de cada muestra, se observa que las fracciones con más restos son las de 2-1 mm. y 1-0.5 mm. En la fracción 2-1 mm. los restos más abundantes son los molares e incisivos, y en la fracción 1-0.5 mm. los metapodios y falanges (Figura 13).
Si se comparan las proporciones de los distintos grupos de microvertebrados presentes en las muestras estudiadas, se puede observar que estas varían a lo
Sigla Nivel Litología Talla( cm)
Ak-Ml A Lanc 67-74
Ak-M2 B Clg 56-62
Ak-M4 B Lgc 67-72
Ak-M5 B Lgc 72-76
Ak-M6 e Lmb 76-81
Ak-M9 e Lmc 89-94
Ak-Mll D Sab+Smc 107-110
AK-Ml2 D Sab 110-115
Ak-Ml4 D Sgb 120-125
Ak-Ml6 D Sab 130-135
Ak-Ml8 D Lmbk 142-147
Ak-Ml9 D Smt 139-147
Ak-M21 D Sat 151-156
T""
T"" T"" T"" T"" N ~ ~ ~ ~ ~ Figura 15. Número de restos 1 1 1 1 1 ~ ~ ~ ~ ~ de microvertebrados recu-<( <( <( <( <( perados en cada muestra por
cada kg de sedimento.
largo del registro estratigráfico (Figura 14). En el caso de los micromamíferos, su proporción es muy pequeña en las primeras muestras, entre el 7.5 y 16%, pero a partir de la muestra AK-M6 la proporción aumenta considerablemente, llegando a ser entre el 80 y el 90% en las últimas muestras. En el caso de los anfibios, la proporción de los restos se mantiene constante en casi todas las muestras, entre el 2.7 y 14.67%, menos en las últimas muestras, donde la proporción aumenta un poco. En el caso de los reptiles,
PO (kg) Plavado (kg) Numero restos Restos/kg
2 0,56 1872 936,00
2,1 0,51 2183 1039,52
3,25 0,82 4203 1293,23
3,21 0,79 1810 563,86
3,86 1,10 714 184,97
3,77 0,91 463 122,81
3,8 0,45 574 151,05
5,6 0.594 1070 191,07
4,2 0,80 829 197,38
5 0,85 632 126,40
5,4 1,52 198 36,67
6,98 0,72 1847 264,61
5,93 1,42 589 99,33
Tabla 2. Número de restos de microvertebrados recuperados en cada muestra.
KOBIE (Paleoantropología n.º 27), año 2003/07

18 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
su proporción es grande en las primeras cuatro muestras, donde llega a ser 80 o 90%, pero a partir de la muestra AK-M6, esta proporción disminuye hasta ser el 1 o 2%. Se puede observar que entre los cambios de proporciones de micromamíferos y reptiles están relacionados. Cuando disminuye la proporción de micromamíferos, aumenta la proporción de reptiles, y viceversa (Figura 14).
Los restos de reptiles son principalmente osteodermos de la especie Anguis fragilis, y aparecen principalmente en las primeras cuatro muestras. Este tipo de restos aparecen en grandes cantidades, se han contado hasta 2000 osteodermos en una misma muestra. Cuando disminuye la cantidad de osteodermos, la proporción de restos de reptiles disminuye considerablemente, y en consecuencia aumenta la proporción de restos de micromamíferos.
Este cambio en las proporciones puede haber sido debido a varias razones. La primera, sería que el cambio se ha producido debido a variaciones de los procesos tafonómicos. Pero como se ha visto anterior-mente, las curvas en las que se representan las proporciones entre los distintos elementos anatómicos recuperados son muy similares en todas las muestras, lo que hace pensar en una historia tafonómica similar.
La segunda razón, sería la caza selectiva. Puede darse la posibilidad de que cuando se sedimentaron los materiales que forman las primeras muestras, existiera un depredador y que en su dieta esta especie tuviera un papel importante, aumentando así su representación en la tanatocenosis.
Otra posibilidad podría ser que estas variaciones sean debidas a cambios en el ambiente. Cuando se sedimentaron los materiales que forman las primeras muestras, debido a las condiciones ambientales reinantes, la cantidad de individuos de la especie Anguis fragilis sería abundante, y en
AK·M195'.J
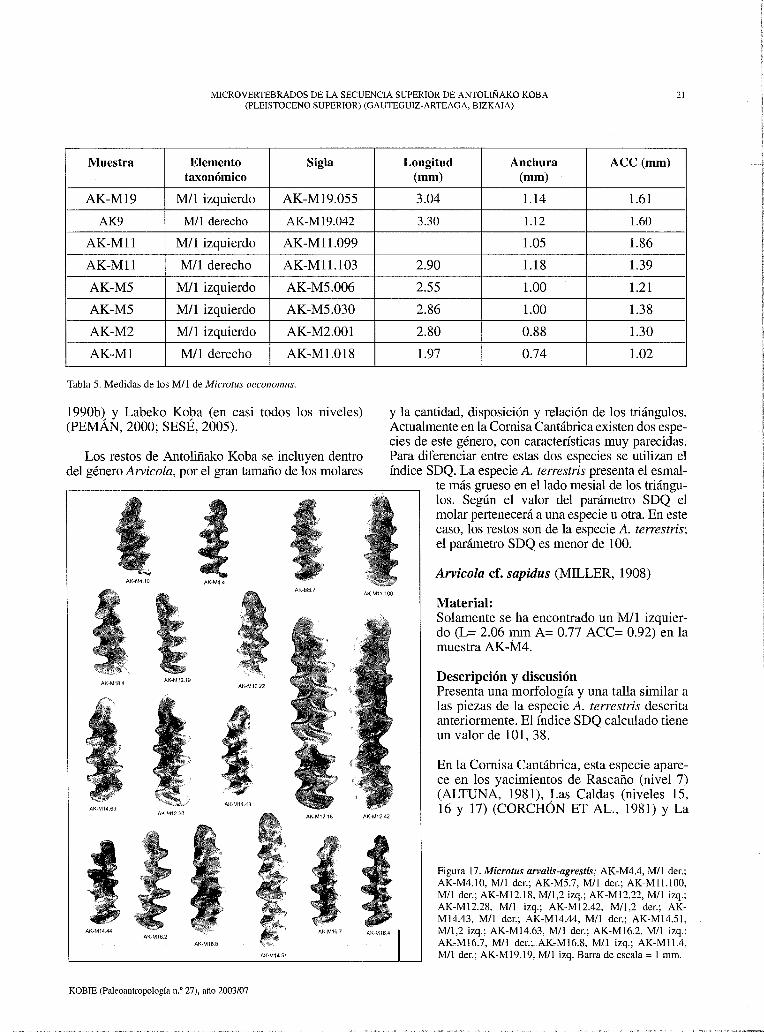
Figura 16. Arvicola terrestris; AK-Ml.20, Mil der.; AK-Ml2.12, M/1 der.; AK-Ml2.41, Mil y M/2 der.; AK-Ml4.55, M/l izq.; AK-Ml6.6, Mii der.; AKMl9.59, M/1 der.; AK-Ml9.67, M/l der.; AK-Ml9.72, Mil der.; AK-Ml9.69, M/l der.; AK-Ml9.74, M/1,2 der. Microtus oeco11omus; AK-Ml.18, M/l der.; AKM5.6, Mil izq.; AK-MS.30, Mil izq.; AK-Mll.103, M/l der.; AK-Ml4.42, M/l der.; AK-Ml9.55, Mil izq. Eliomys querci11us; AK-Ml9.34, M/1-2 der. Clethrio-11omys glareolus; AK-M21.39, M/2 izq. Barra de escala= 1 mm.
AK·M5.6
AK-M1.1B
consecuencia, aumentan los restos de esta especie en el registro. En las muestras que representan sedimentos más antiguos, estos restos no serían tan numerosos, debido a un empeoramiento de las condiciones ambientales para esta especie.
También se ha tenido en cuenta la riqueza en restos en las distintas muestras. Esta abundancia de restos se representa con la relación entre el número de restos recuperados en la muestra y su peso antes de ser lavado. La muestra con más restos es la muestra AK-M4, con 1293, 23 restos por Kg. de sedimento y la más pobre es AK-Ml8, con 36, 67 restos por Kg. de sedimento (Figura 15). En la curva en la que se representa la riqueza de restos, se puede observar un cambio bastante brusco. Las primeras tres muestras contienen numerosos restos en comparación con las otras, aumentando la proporción de restos desde la primera, AK-Ml, hasta la AK-M4, para luego disminuir considerablemente hasta la muestra AK-M18. En la AK-M19 aumenta un poco, para disminuir de nuevo en la AK-M21 (Tabla 2 y Figura 15).
AKM166
AK-M1972 AK~19t9
AK·M11.103
AK·M1934 AK·M2139
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
19
Nivel Talla( cm) PO (kg)
A 67-74 2
B 56-76 8,56
c 76-94 7,63
D 107-156 36,91
Tabla 3. Número de restos de microvertebrados recuperados en cada nivel.
Esta curva es similar a las curvas en las que se representan las proporciones de reptiles, anfibios y micromamíferos, ya que los dos cambios más significativos se producen en parecidas muestras (Figuras 14 y 15).
Existen algunas piezas que presentan alteraciones químicas producidas por los jugos gástricos de sus depredadores, que podrían haber sido algún pequeño mamífero carnívoro o alguna ave como el Cárabo o Búho.
Teniendo en cuenta que las muestras se han obtenido del interior de la cavidad, se puede afirmar que los taxones identificados en este yacimiento son todos adémicos. La mayoría de estos restos han sido transportados junto a los sedimentos desde el exterior de la cueva al interior, sufriendo un menor o mayor transporte.
SISTEMÁTICA
En este apartado se describen los diferentes taxones identificados en las muestras. Entre los distintos restos de microfauna identificados, se encuentran los micromamíferos (Clase Mammalia), anfibios (Clase Amphibia), reptiles (Clase "Reptilia"), peces (Clase Osteichthyes) y aves (Clase Aves). Para realizar la clasificación se han tenido en cuenta los distintos tipos de molares y elementos óseos.
Clase Mammalia Orden Rodentia Familia Gliridae Género Eliomys (WAGNER, 1840) Eliomys quercinus (LINNAEUS, 1766)
Material: 1 M/1-2 derecho (L= 1.86 mm A=l.49 mm) (Figura 16. AK-Ml9.34)
Descripción y discusión El molar presenta un contorno subcuadrangular.
La superficie oclusal es ligeramente cóncava. Presenta 4 crestas transversales bien marcadas, el anteroló-
KOBIE (Paleoantropología n.º 27), año 2003/07
Plavado (kg) Numero restos Restos/kg
0,56 1872 936
2,12 8096 945,79
2,01 1177 154,25
6,354 5739 155,48
fido, el metalófido, el mesolófido y el posterolófido. Entre las crestas principales no se desarrolla cresta accesoria alguna.
Los primeros restos del Pleistoceno superior de esta especie de la zona Cantábrica se encontraron en los niveles 8 y 11 de este periodo del yacimiento de Esquilleu (BAENA et al., 2004; SESÉ, 2005).
La pieza de Antoliñako Koba se incluye dentro de la especie E. quercinus por su morfología dentaria simple, sin crestas accesorias. Los molares de la especie Glis glis presentan una morfología más compleja con crestas accesorias.
Familia Arvicolidae Género Clethrionomys (TILESIUS, 1850) Clethrionomys glareolus (SCHREBER, 1780)
Material: (Figura 16. AK-M21-39) Los únicos restos que se han recuperado pertene
cen a la muestra AK-M21. Se han encontrado dos molares, un M/2 izquierdo (L= 1.8 mm A= 1.1 mm) y un fragmento de molar no identificable.
Descripción y discusión El M/2 presenta dos raíces muy desarrolladas.
También presenta cemento en los entrantes entre los triángulos. El esmalte es grueso y mantiene el grosor en todo el perímetro del molar de forma continua. Los triángulos se encuentran todos abiertos.
Esta especie se ha encontrado en los yacimientos de Covalejos (niveles B, J y K) (SESÉ, 2005), Erralla (nivel II) (PEMÁN, 1985), y Laminak II (niveles II y III) (PEMÁN, 1994) del Pleistoceno superior. También aparece en los yacimientos Holocenos de Amalda (PEMÁN, 1990) y Laminak II (PEMÁN, 1994) (SESÉ, 2005).
En la familia Arvicolidae existen dos géneros con raíces, Clethrionomys y Pliomys. El primero de ellos tiene cemento entre los triángulos y generalmente el segundo no. En los triángulos de los molares de Pliomys el grosor del esmalte es mayor en el lado

20 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
Muestra Elemento Sigla Longitud Anchura ACC SDQ taxonómico (mm)
AK-M19 M/1 derecho AK-M19.059 4.17
AK-Ml9 Mil izquierdo AK-Ml9.063 3.49
AK-Ml9 Mil derecho AK-Ml9.067 3.92
AK-Ml9 Mil derecho AK-Ml9.069 3.83
AK-Ml9 Mil derecho AK-Ml9.072 3.69
AK-Ml9 Mil izquierdo AK-Ml9.075 4.32
AK-Ml9 Mil derecho AK-Ml9.079 3.89
AK-Ml6 Mil derecho AK-Ml6.006 3.82
AK-Ml4 Mil izquierdo AK-Ml4.055 3.83
AK-Ml2 Mil derecho AK-Ml2.012 4.27
AK-Ml2 Mil izquierdo AK-Ml2.016 3.99
AK-Ml2 Mil izquierdo AK-Ml2.017 4.02
AK-Ml2 Mil derecho AK-Ml2.040 3.01
AK-Ml Mil derecho AK-Ml.020 2.75
Tabla 4. Medidas de los Mil de Arvicola terrestris.
mesial que en el distal. Debido a la continuidad en el grosor del esmalte y a la presencia de entrantes en los cementos se ha atribuido el material de Antoliñako Koba al género Clethrionomys. En la península Ibérica, solo vive una especie de este género, Cletrhionomys glareolus, por esa razón, estos molares se incluyen dentro de esta especie.
Aunque los dos molares sean de la misma especie, tienen un aspecto muy diferente. Las raíces de uno de los molares están más desarrolladas que los del otro molar. Las raíces aumentan de tamaño desde que nace el individuo. Los individuos jóvenes poseen raíces pequeñas, y a medida que pasa el tiempo, estas aumentan de tamaño y grosor (POKINES, 1998). Por esa razón, se puede decir que los dos molares pertenecen a distintos individuos de distinta edad.
Género Arvicola (LACÉPEDE, 1799) Arvicola terrestris (LINNAEUS, 1758)
Material: (Figura 16 y Tabla 4) El material que corresponde a esta especie se ha
recuperado en las muestras AK-Ml, AK-Ml2, AKMl4, AK-Ml6 y AK-Ml9. En general las muestras son muy pobres en restos de esta especie, menos en la muestra AK-Ml9, donde se han recuperado siete Mil. En las demás muestras se ha recuperado solo un Mil por muestra.
(mm) (mm)
1.74 1.81 70,8
1.46 1.38 85,68
1.73 1.75 76,23
1.49 1.74 82,2
1.43 1.72 83,76
1.50 2.07 76,46
1.65 1.64 84,29
1.49 1.67 68,21
1.72 1.74 69,37
1.72 2.01 58,55
1.63 1.93 49,96
1.77 1.90 69
1.19 1.38 47,82
1.09 1.28 61,86
Descripción y discusión Los molares de las especies de este género son de
gran tamaño si se comparan con otros molares, y por tanto, se diferencian fácilmente de los demás géneros de la familia Arvicolidae. Si se observa el lado oclusal de los molares, se puede ver que están constituidos por cuatro triángulos linguales y tres triángulos labiales. Los molares no tienen raíces y desarrollan cemento en los entrantes que hay entre los triángulos. El complejo posterior de los molares está constituido por tres triángulos no comunicados o débilmente comunicados entre ellos. El complejo anterior está compuesto por dos triángulos comunicados entre ellos, los triángulos t4-t5, y estos se juntan con el lóbulo anterior, quedando los tres comunicados entre ellos. El esmalte tiene continuidad en todo el perímetro del molar.
En la Comisa Cantábrica, esta especie aparece en el Pleistoceno superior, y es muy abundante en los yacimientos de esta época; Covalejos (en casi todos los niveles) (SESÉ, 2005); Esquilleu (en los niveles 8 y 11) (BAENA ET AL., 2005); Ermittia (en todos los niveles), Aitzbitarte IV (en muchos niveles); Urtiaga (en los niveles F y 1) (ALTUNA, 1972); Tito Bustillo (en los subniveles la, by c) (ALTUNA, 1976); Rascaño (en los niveles 1, 3, 4, 5 y 7) (ALTUNA, 1981); La Riera (en casi todos los niveles) (ALTUNA, 1986); Amalda (en muchos niveles) (PEMÁN, 1990b), Laminak 11 (en todos los niveles) (PEMÁN,
KOBIE (Paleoantropología n.º 27), año 2003/07
MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
21
Muestra Elemento Sigla taxonómico
AK-Ml9 Mil izquierdo AK-Ml9.055
AK9 Mil derecho AK-Ml9.042
AK-Mll Mil izquierdo AK-Mll.099
AK-Mll M/1 derecho AK-Mll.103
AK-M5 Mil izquierdo AK-M5.006
AK-M5 Mil izquierdo AK-M5.030
AK-M2 Mil izquierdo AK-M2.001
AK-Ml M/l derecho AK-Ml.018
Tabla 5. Medidas de los Mil de Microtus oeconomus.
1990b) y Labeko Koba (en casi todos los niveles) (PEMÁN, 2000; SESÉ, 2005).
Los restos de Antoliñako Koba se incluyen dentro del género Arvicola, por el gran tamaño de los molares
AK-M4.10 AK-M4.4 AK-M5.7
AK-M19.19 AK-11.112 22
AK-M14fü AK-M1228
AK-M12.18
AK-M14.44 AK-M16.2
AK-M16.8
AK-M1451
KOBIE (Paleoantropología n.º 27), año 2003/07
Longitud Anchura ACC (mm) (mm) (mm)
3.04 1.14 1.61
3.30 1.12 1.60
1.05 1.86
2,90 1.18 1.39
2.55 1.00 1.21
2.86 1.00 1.38
2.80 0.88 1.30
1.97 0.74 1.02
y la cantidad, disposición y relación de los triángulos, Actualmente en la Cornisa Cantábrica existen dos especies de este género, con características muy parecidas, Para diferenciar entre estas dos especies se utilizan el índice SDQ, La especie A terrestris presenta el esmal
AK-~.111.100
AM,112.42
AKM164
te más grueso en el lado mesial de los triángulos, Según el valor del parámetro SDQ el molar pertenecerá a una especie u otra, En este caso, los restos son de la especie A terrestris; el parámetro SDQ es menor de 100,
Arvicola cf. sapidus (MILLER, 1908)
Material: Solamente se ha encontrado un Mil izquierdo (L= 2.06 mm A= 0.77 ACC= 0.92) en la muestra AK-M4.
Descripción y discusión Presenta una morfología y una talla similar a las piezas de la especie A terrestris descrita anteriormente. El índice SDQ calculado tiene un valor de 101, 38.
En la Cornisa Cantábrica, esta especie aparece en los yacimientos de Rascaño (nivel 7) (ALTUNA, 1981), Las Caldas (niveles 15, 16 y 17) (CORCHÓN ET AL., 1981) y La
Figura 17. Microtus arvalis-agrestis; AK-M4.4, Mil der.; AK-M4.10, Mil der.; AK-MS.7, Mil der.; AK-Mll.100, Mil der.; AK-Ml2.18, M/1,2 izq.; AK-M12.22, Mil izq.; AK-Ml2.28, Mil izq.; AK-Ml2.42, Mll,2 der.; AKM14.43, Mil der.; AK-M14.44, Mil der.; AK-Ml4.51, Mll,2 izq.; AK-Ml4.63, Mil der.; AK-Ml6.2, Mil izq.; AKcM16.7, Mil der.; AK-M16.8, Mil izq.; AK-Mll.4, Mil der.; AK-Ml9.19, Mil izq. Barra de escala= 1 mm.

22 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
Muestra Elemento Sigla taxonómico
AK-Ml Mil izquierdo AK-Ml.019
AK-M4 Mil derecho AK-M4.004
AK-M5 Mil derecho AK-M5.007
AK-Mll Mil izquierdo AK-Mll.100
AK-Mll M/l izquierdo AK-Mll.102
AK-Mll Mil izquierdo AK-Mll.110
AK-Ml2 Mil izquierdo AK-Ml2.019
AK-Ml2 Mil derecho AK-Ml2.021
AK-Ml2 Mil izquierdo AK-Ml2.022
AK-Ml2 M/l izquierdo AK-Ml2.028
AK-Ml2 Mil izquierdo AK-Ml2.029
AK-Ml2 Mil izquierdo AK-Ml2.030
AK-Ml2 Mil derecho AK-Ml2.031
AK-Ml2 Mil derecho AK-Ml2.032
AK-Ml2 Mil derecho AK-Ml2.043
AK-Ml4 Mil izquierdo AK-Ml4.041
AK-Ml4 Mil derecho AK-Ml4.043
AK-Ml4 Mil izquierdo AK-Ml4.044
AK-Ml4 Mil izquierdo AK-Ml4.051
AK-Ml4 Mil derecho AK-Ml4.053
AK-Ml4 Mil izquierdo AK-Ml4.056
AK-Ml4 Mil derecho AK-Ml4.63
AK-Ml6 M/l derecho AK-Ml6.001
AK-Ml6 M/l izquierdo AK-Ml6.002
AK-Ml6 M/1 derecho AK-Ml6.003
AK-Ml6 Mil derecho AK-Ml6.004
AK-Ml6 M/l derecho AK-Ml6.005
Tabla 6. Medidas de los M/1 de Microtus an1alis-agrestis.
Riera (nivel 9) (ALTUNA, 1986) del Pleistoceno superior (SESÉ, 2005).
Este material se ha incluido en la especie A. sapidus y no en A. terrestris por el índice SDQ, que es mayor de 100. De todas maneras, solamente disponemos de un ejemplar y por esa razón la atribución taxonómica final se realiza con reservas como A. cf. sapidus
Longitud Anchura ACC (mm) (mm) (mm)
1.95 0.60 1.09
2.60 0.88 1.36
2.59 0.97 1.24
2.91 1.07 1.59
2.75 1.01 1.46
2.69 0.94 1.43
3.00 1.10 1.72
2.73 0.94 1.60
2.58 0.91 1.47
3.36 1.14 1.84
3.00 1.03 1.68
2.86 1.08 1.60
2.72 1.02 1.53
2.99 1.06 1.64
3.04 0.99 1.77
2.78 0.98 1.50
2.84 0.98 1.59
2.61 0.97 1.47
2.79 0.97 1.51
2.77 1.01 1.50
2.79 0.94 1.68
3.18 1.18 1.88
2.97 1.01 1.75
3.09 0.97 1.82
2.56 0.88 1.46
2.85 0.97 1.56
2.74 1.02 1.57
Género Microtus (SCHRANK, 1798) Microtus oeconomus (PALLAS, 1776) (=Micro
tus ratticeps KEYSERLING & BLASIUS, 1841)
Material: (Figura 16 y Tabla 5) Los restos de esta especie se han encontrado en las
muestras AK-Ml, AK-M2, AK-M5, AK-Mll, AKMl4 y AK-Ml9.
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
23
Muestra Elemento Sigla taxonómico
AK-Ml6 Mil derecho AK-Ml6.007
AK-Ml6 Mil izquierdo AK-Ml6.008
AK-Ml6 Mil derecho AK-Ml6.013
AK-Ml8 Mil derecho AK-Ml8.006
AK-Ml9 Mil izquierdo AK-Ml9.002
AK-Ml9 M/1 izquierdo AK-Ml9.019
AK-Ml9 Mil izquierdo AK-Ml9.022
AK-Ml9 Mil derecho AK-Ml9.057
AK-Ml9 Mil derecho AK-Ml9.058
AK-Ml9 Mil derecho AK-Ml9.061
AK-Ml9 Mil izquierdo AK-Ml9.064
AK-Ml9 Mil derecho AK-Ml9.065
AK-Ml9 Mil derecho AK-Ml9.066
AK-Ml9 M/l izquierdo AK-Ml9.070
AK-Ml9 Mil derecho AK-Ml9.071
AK-Ml9 Mil derecho AK-Ml9.073
AK-Ml9 Mil izquierdo AK-Ml9.077
AK-Ml9 Mil izquierdo AK-Ml9.078
AK-M21 M/l izquierdo AK-M21.007
AK-M21 Mil izquierdo AK-M21.009
AK-M21 Mil derecho AK-M21.010
AK-M21 Mil derecho AK-M21.012
AK-M21 Mil derecho AK-M21.033
Tabla 7. Medidas de los Mil de Microtus arvalis-agrestis.
Descripción y discusión Al no desarrollarse el triangulo 6 la morfología de
la superficie oclusal presenta cinco triángulos en el lado labial y tres en el lingual. Los triángulos t5-t7 se encuentran comunicados entre ellos y el triángulo t5 con el lóbulo anterior. Al no existir el triángulo t6, el lóbulo anterior presenta una marcada asimetría. No tienen raíces y desarrollan cemento entre los triángulos.
Esta especie también se conoce como Microtus ratticeps, pero tiene preferencia la denominación de Microtus oeconomus. La ausencia del triangulo t6 es un carácter diagnóstico propio de M. oeconomus. Esta especie aparece en numerosos yacimientos del Pleistoceno superior de la Comisa Cantábrica: Cova-
KOB!E (Paleoantropología n.º 27), año 2003/07
Longitud Anchura ACC (mm) (mm) (mm)
2.90 0.90 1.68
3.47 1.10 1.99
2.27 0.91 1.18
2.90 1.06 1.61
2.90 1.06 1,59
2.57 0.87 1.43
2.53 0.99 1.37
3.17 1.14 1.80
2.99 1.03 1.58
3.03 1.06 1.73
2.17 0.89 1.58
2.78 0.89 1.56
2.62 0.98 1.38
3.04 1.03 1.62
2.48 1.05 1.36
3.02 0.99 1.66
3.08 1.09 1.70
3.01 0.80 1.71
2.86 1.04 1.55
2.60 1.04 1.43
2.72 1.48 0.95
3.08 1.06 1.72
2.26 0.72 1.22
lejos (niveles A, B, I, J y K) (SESÉ, 2005); Lezetxiki (nivel I) (CHALINE, 1970); Aitzbitarte IV (en todos los niveles); Ermittia (en el Solutrense) (ALTUNA, 1972); Tito Bustillo (en los subniveles 1 a, b y c) (ALTUNA, 1976); Rascaño (en los niveles 2, 4 y 5) (ALTUNA, 1981); Ekain ( en los niveles VI, VII y VIII) (ZABALA, 1984); Erralla (el taxón más numeroso en todos los niveles) (PEMÁN 1985); La Riera (nivel 7) (ALTUNA, 1986); Amalda (PEMÁN 1990b); Laminak II (en los niveles II y III) (PEMÁN, 1994). También aparece en los yacimientos ,Holocenos de La Riera, Amalda y Laminak II (SESE, 2005).
Microtus arvalis-agrestis (PALLAS, 1779 y LINNAEUS, 1761)

24 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
AK·M16.16 AK-M16.14 AK-M19,65
AK-M21.12 AK-M21.33 AI0.1119.77
AK~M21.11
AK-M19.21 AK-M2.2
AK-M11.104
AK-M19 22
AK·M1961
AK-M19.66 AK-M19.73
AK-MS.3 AK-M9.1
Figura 18. Microtus arvalis-agrestis: AKMl6.14, Mil izq.; AK-M16.16, Mil der.; AK-Ml 1.5, Mll,2 der.; AK-M19.22, Mil izq.; AK-M19.61, Mil der.; AK-M19.65, Mil der.; AK-Ml9.66, Mil der.; AK-M19.73, Mil der.; AK-Ml9.77, Mil izq.; AK-M21.l l, Mll,2 der.; AK-M21.12, Mil der.; AKM21.33, Mil der. Terricola sp.: AK-M2.2, Mil der.; AK-M6.3, Mil izq.; AK-M9.l, Mil der.; AK-Mll.104, Mil der.; AK-M19.21, Mil izq.; AK-M19.60, Mil izq.; AK-M21.35, Mil der. Chionomys nivalis: AK-M9.9, Mil izq.; AK-Mll.44, Mil der.; AK-Ml4.54, Mil der. Barra de escala = 1 mm.
que más restos contiene. Se han recuperado 56 Mil y se han contabilizado 36 NMI.
Descripción y discusión Los M/1 presentan cinco triángulos linguales y cuatro labiales. Los entrantes entre los triángulos tienen cemento y no desarrollan raíces. Los triángulos t4-t5 están cerrados. El esmalte es más grueso en el lado mesial que en el distal. El complejo posterior está constituido por tres triángulos cerrados, y el complejo anterior por cuatro triángulos cerrados o casi cerrados. Todos los triángulos son muy asimétricos entre ellos.
AK-M21.35 AK·M19 60 AK·M11.44
Las dos especies de este grupo, Microtus arvalis y Microtus agrestis, son muy difíciles de diferenciar entre ellos. Por esta razón, no se ha
AK-M1454
Material: (Figuras 17 y 18 y Tablas 6 y 7) Los restos atribuibles a esta especie se han recu
perado en muchas muestras; AK-Ml, AK-M4, AKM5, AK-Mll, AK-M12, AK-M14, AK-M16, AKM18, AK-M19 y AK-M21. En general su representación en las diferentes muestras aumentan a partir de la muestra AK-Mll, siendo la muestra AK-M19 la
Muestra Elemento Sigla taxonómico
AK-M9 M/1 izquierdo AK-M9.009
AK-Mll M/1 derecho AK-Mll.044
AK-M14 M/1 derecho AK-M14.054
Tabla 8. Medidas de los Mil de Chionomys nivalis.
especificado a cual de las dos especies pertenece el material descrito en este trabajo.
En los yacimientos del Pleistoceno superior de la Cornisa Can,tábrica este grupo es muy habitual; Covalejos (SESE, 2005); Esquilleu (BAENA ET AL., 2005); Lezetxiki, Aitzbitarte IV y Ermittia (ALTUNA, 1972); Tito Bustilo (ALTUNA, 1976); Rascaño (ALTUNA, 1981); Las Caldas (CORCHÓN ET AL.,
Longitud Anchura ACC (mm) (mm) (mm)
1.85 0.83 0.74
2.33 0.91 0.90
3.00 1.05 1.55
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
25
Muestra Elemento Sigla taxonómico
AK-M2 Mil derecho AK-M2.002
AK-M5 Mil izquierdo AK-M5.005
AK-M5 M/l derecho AK-M5.023
AK-M6 Mil izquierdo AK-M6.003
AK-M9 Mil derecho AK-M9.001
AK-Mll Mil derecho AK-Mll.104
AK-Ml9 Mil izquierdo AK-Ml9.021
AK-Ml9 Mil izquierdo AK-Ml9.060
AK-Ml9 Mil derecho AK-Ml9.068
AK-M21 Mil derecho AK-M21.035
Tabla 9. Medidas de los M/l de Terrícola sp.
1981); La Riera (ALTUNA, 1986); Ekain (ZABALA, 1984); Erralla (PEMÁN, 1985); Amalda (PEMÁN, 1990b); Laminak II (Pemán, 1994) y Labeko Koba (PEMÁN, 2000). En estos yacimientos aparecen en todos o casi todos los niveles y en algunos llega a ser el taxón más numeroso.
Género Chionomys (MILLER, 1908) Chionomys nivalis (MARTINS, 1842)
Material: (Figura 18 y Tabla 8) El material adscrito a esta especie se ha recupera
do en las muestras AK-M9, AK-Mll y AK-Ml4.
Descripción y discusión Presenta cuatro triángulos y entrantes en el lado lin
gual, y tres triángulos y entrantes en el lado labial. Este molar no tiene raíces y desarrolla cemento en los entrantes. El lóbulo anterior es muy característico, con una morfología de hongo inclinado hacia el lado labial.
En la Cornisa Cantábrica esta especie aparece en muchos yacimientos del Pleistoceno superior: Covalejos (en el nivel J) (SESÉ, 2005), Esquilleu (nive18) (BAENA ET AL., 2005), Aitzbitarte (en los cuatro niveles) (ALTUNA, 1972), Ekain (en los niveles VII y VIII) (ZABALA, 1984), Erralla (en todos los niveles) (PEMÁN, 1985), La Riera (en los niveles 18 y 24) (ALTUNA, 1986), Amalda (en los niveles IV, VI y X) (PEMÁN, 1990b) y Laminak II (en los niveles II y III) (PEMÁN, 1994). También aparece en los yacimientos Holocenos de Amalda (PEMÁN, 1990b) y Laminak II (PEMÁN, 1994) (SESÉ, 2005).
KOBIE (Paleoantropología n.º 27). año 2003/07
Longitud Anchura ACC (mm) (mm) (mm)
2.35 0.72 1.24
2.47 0.78 1.29
2.56 0.90 1.36
2.40 0.82 1.25
2.59 0.91 1.36
2.71 1.42 0.96
2.35 0.85 1.13
2.92 0.96 1.61
2.84 1.01 1.50
2.54 0.91 1.33
Esta especie se ha incluido dentro del género Microtus y algunas veces dentro del subgénero Chionomys. Por esta razón se puede encontrar en la literatura científica como Microtus nivalis o Microtus (Chionomys) nivalis. NADACHOWSKI (1990) diferenció el género Chionomys (MILLER, 1990) del género Microtus (SCHRANCK, 1798) debido a diferencias morfológicas y bioquímicas, por lo que en la actualidad se denomina como C. nivalis.
Los restos descritos anteriormente se incluyen dentro de este género y especie por su lóbulo anterior de morfología de hongo y número de triángulos. Los dos tipos de molares pertenecen a la misma especie. Las diferencias que existen entre ellos son debido a diferencias de edad entre ellos: a los individuos más jóvenes les falta el cuarto entrante y el triángulo t5 del lado lingual. Con el tiempo, el triángulo t5 aumenta de tamaño y se genera el cuarto entrante.
Género Terrícola (CHALINE, 1988) Terrícola sp.
Material: (Figura 18 y Tabla 9) Molares de esta especie se han recuperado en las
muestras AK-M2, AK-M5, AK-M6, AK-M9, AKMll, AK-Ml9 y AK-M21.
Descripción y discusión Estos molares tienen cuatro triángulos en el lado
lingual y cinco triángulos en el lado labial. No presentan raíces, pero si cemento en los entrantes. Los triángulos t4-t5 se encuentran totalmente comunica-

26 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
AK-M1.6 AK-M1.1
AK-M1.13 AK-M1.9
AK-M1 .12
Ak-M2.37
Ak-M2.8
Ak-M2.10 Ak-M2.9 Ak-M2.6 Ak-M2.32
AK-M4.34 AK-M4.42 AK-M4.33
AK-M4.41 AK-M4.64 AK-M4.36
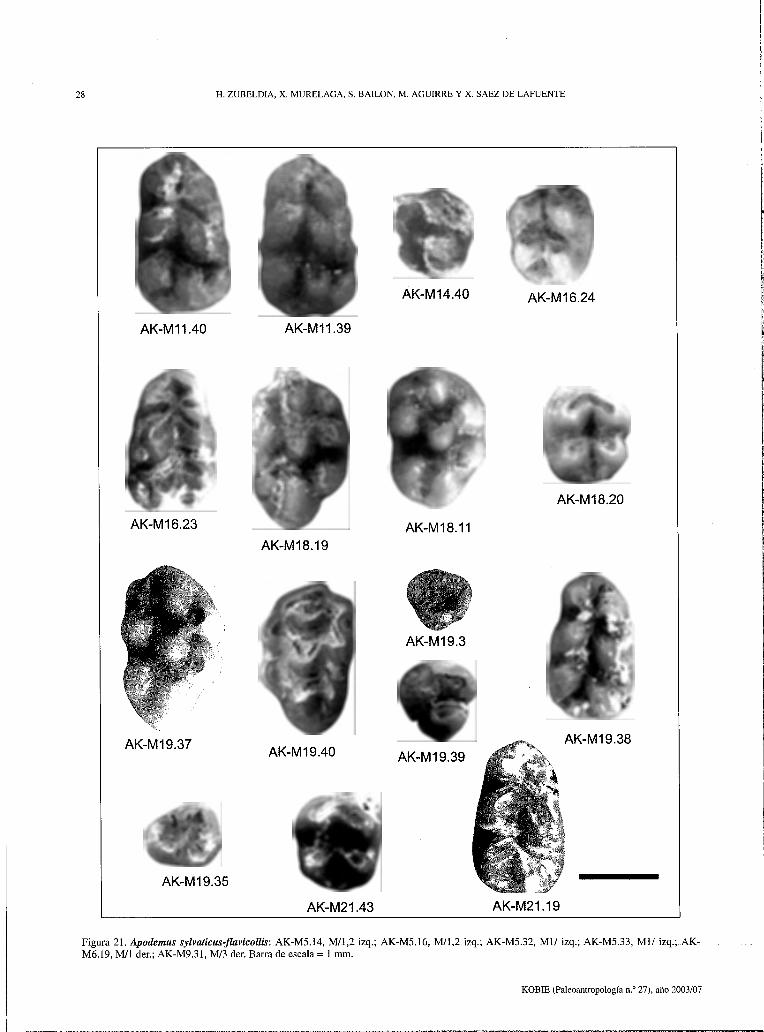
Figura 19. Apodemus sylvaticus-flavicollis: AK-Ml.1, M/3 der.; AK-Ml.6, M3/ der.; AK-Ml.9, Mil izq.; AK-Ml.12, MI/ der.; AK-Ml.13, Ml/ der.; AK-M2.6, Mil der.; AK-M2.8, M/2 der.; AK-M2.9, Mil der.; AK-M2.10, Ml/ der.; AK-M2.32, M3/ der.; AK-M2.33, M/3 der.; AK-M2.37, M3/ der.; AK-M4.33, Ml/ der.; AK-M4.34, Ml/ izq.; AK-M4.36, Mil der.; AK-M4.41, M/2 izq.; AK-M4.42, Ml/ izq.; AK-M4.64, M/3 izq. Barra de escala = 1 mm.
KOBIE (Paleoantropologfa n.º 27), año 2003/07
.... ········--------·- -----

AK-MS.33
MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
AK-MS.16
AK-MS.14
AK-MS.32
AK-M9.31 AK-M9.31
AK-M6.19
27
Figura 20. Apodemus sylvaticus-flavicollis: AK-Mll.39, M3/ der.; AK-Ml 1.40, M/l der.; AK-Ml4.40, M3/ der.; AK-Ml6.23, Mil izq.; AKMl6.24, M/2 der.; AK-Mll.11, M2/ izq.; AK-Mll.19, Ml/ der.; AK-Mll.20, M/2 izq.; AK-M19.3, M3/ izq.; AK-Ml9.35, M/3 der.; AKMl9.37, Ml/ der.; AK-Ml9.38, Mil der.; AK-Ml9.39, M/3 der.; AK-Ml9.40, Ml/ izq.; AK-M21.19, Mil der.; AK-M21.43, M/2 der. Barra de escala = 1 mm.
KOBIE (Paleoantropologfa n.º 27), año 2003/07
28 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
AK-M14.40 AK-M16.24
AK-M11.40 AK-M11.39
AK-M18.20
AK-M16.23 AK-M18.11 AK-M18.19
AK-M19.3
AK-M19.40 AK-M19.37 AK-M19.38
AK-M19.39
AK-M19.35
AK-M21.43 AK-M21.19
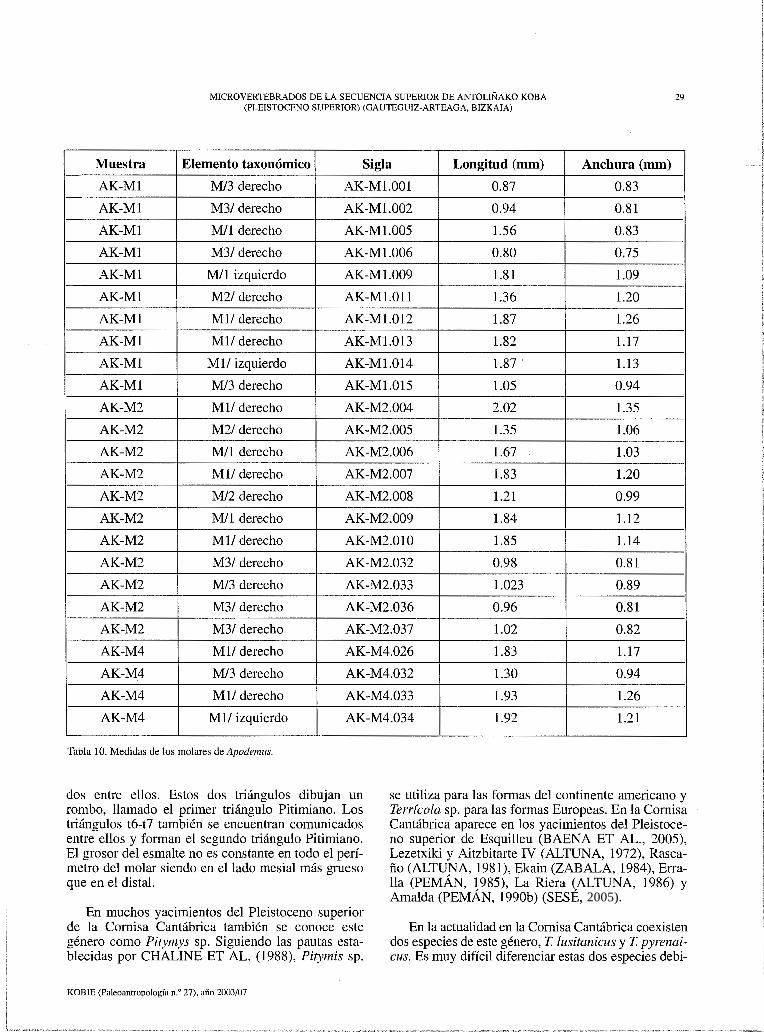
Figura 21. Apodemus sylvaticus-jlavicollis: AK-MS.14, M/1,2 izq.; AK-MS.16, M/1,2 izq.; AK-MS.32, Ml/ izq.; AK-MS.33, Ml/ izq.; AKc M6.19, Mil der.; AK-M9.31, M/3 der. Barra de escala= 1 mm.
KOBIE (Paleoantropologfa n.º 27), año 2003/07
Muestra
AK-Ml
AK-Ml
AK-Ml
AK-Ml
AK-Ml
AK-Ml
AK-Ml
AK-Ml
AK-Ml
AK-Ml
AK-M2
AK-M2
AK-M2
AK-M2
AK-M2
AK-M2
AK-M2
AK-M2
AK-M2
AK-M2
AK-M2
AK-M4
AK-M4
AK-M4
AK-M4
MICRO VERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
Elemento taxonómico Sigla Longitud (mm)
M/3 derecho AK-Ml.001 0.87
M3/ derecho AK-Ml.002 0.94
Mil derecho AK-Ml.005 1.56
M3/ derecho AK-Ml.006 0.80
Mil izquierdo AK-Ml.009 1.81
M2/ derecho AK-Ml.011 1.36
Ml/ derecho AK-Ml.012 1.87
Ml/ derecho AK-Ml.013 1.82
Ml/ izquierdo AK-Ml.014 1.87
M/3 derecho AK-Ml.015 1.05
Ml/ derecho AK-M2.004 2.02
M2/ derecho AK-M2.005 1.35
Mil derecho AK-M2.006 1.67
Ml/ derecho AK-M2.007 1.83
M/2 derecho AK-M2.008 1.21
Mil derecho AK-M2.009 1.84
Ml/ derecho AK-M2.010 1.85
M3/ derecho AK-M2.032 0.98
M/3 derecho AK-M2.033 1.023
M3/ derecho AK-M2.036 0.96
M3/ derecho AK-M2.037 1.02
Ml/ derecho AK-M4.026 1.83
M/3 derecho AK-M4.032 1.30
Ml/ derecho AK-M4.033 1.93
Ml/ izquierdo AK-M4.034 1.92
Tabla 1 O. Medidas de los molares de Apodemus.
29
Anchura (mm)
0.83
0.81
0.83
0.75
1.09
1.20
1.26
1.17
1.13
0.94
1.35
1.06
1.03
1.20
0.99
1.12
1.14
0.81
0.89
0.81
0.82
1.17
0.94
1.26
1.21
dos entre ellos. Estos dos triángulos dibujan un rombo, llamado el primer triángulo Pitimiano. Los triángulos t6-t7 también se encuentran comunicados entre ellos y forman el segundo triángulo Pitimiano. El grosor del esmalte no es constante en todo el perímetro del molar siendo en el lado mesial más grueso que en el distal.
se utiliza para las formas del continente americano y Terrícola sp. para las formas Europeas. En la Comisa Cantábrica aparece en los yacimientos del Pleistoceno superior de Esquilleu (BAENA ET AL., 2005), Lezetxiki y Aitzbitarte IV (ALTUNA, 1972), Rascaño (ALTUNA, 1981), Ekain (ZABALA, 1984), Erralla (PEMÁN, !985), La Riera (ALTUNA, 1986) y Amalda (PEMAN, 1990b) (SESE, 2005).
En muchos yacimientos del Pleistoceno superior de la Comisa Cantábrica también se conoce este género como Pitymys sp. Siguiendo las pautas establecidas por CHALINE ET AL. (1988), Pitymis sp.
KOBIE (Paleoantropología n.º 27), año 2003/07
En la actualidad en la Comisa Cantábrica coexisten dos especies de este género, T. lusitanicus y T. pyrenaicus. Es muy difícil diferenciar estas dos especies debí-

30 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
Muestra Elemento taxonómico Sigla Longitud (mm) Anchura (mm)
AK-M4 AK-M4.036 1.73 1.12
AK-M4 M/2 izquierdo AK-M4.041 1.11 0.93
AK-M4 M 1/ izquierdo AK-M4.042 1.75 1.09
AK-M4 M/3 izquierdo AK-M4.064 0.98 0.87
AK-M5 Mil izquierdo AK-M5.014 1.74 1.05
AK-M5 M/2 izquierdo AK-M5.014 1.17 1.05
AK-M5 Ml/ derecho AK-M5.019 1.78 1.16
AK-M5 M 1/ izquierdo AK-M5.033 2.05 1.31
AK-M5 M3/ izquierdo AK-M5.034 1.28 1.23
AK-M5 M3/ derecho AK-M5.036 1.30 1.12
AK-M5 M2/ izquierdo AK-M5.051 1.18 1.05
AK-M5 M2/ izquierdo AK-M5.054 1.16 0.98
AK-M6 Mil derecho AK-M6.019 1.64 0.94
AK-M6 M3/ izquierdo AK-M6.029 1.17 0.98
AK-M9 M/2 izquierdo AK-M9.019 1.16 1.11
AK-M9 M/3 derecho AK-M9.031 0.99 0.94
AK-Mll M3/ derecho AK-Mll.019 1.03 0.85
AK-Mll M/1 derecho AK-Mll.040 2.00 1.14
AK-Ml4 M3/ derecho AK-Ml4-040 1.15 1.05
AK-Ml6 Mil izquierdo AK-Ml6-023 1.78 1.05
AK-Ml6 M/2 derecho AK-Ml6-024 1.26 1.10
AK-Ml8 M2/ izquierdo AK-M18-011 1.81 1.30
AK-Ml8 Ml/ derecho AK-Ml8-019 2.02 1.26
AK-M18 M/2 izquierdo AK-M18-020 1.22 1.07
AK-Ml9 M3/ izquierdo AK-M19.003 0.82 0.78
AK-Ml9 M/3 derecho AK-Ml9.035 0.97 0.81
AK-Ml9 Mil derecho AK-Ml9.036 1.65 1.04
AK-Ml9 Ml/ derecho AK-Ml9.037 1.85 1.32
AK-Ml9 M/1 derecho AK-Ml9.038 1.85 1.06
AK-Ml9 M/3 derecho AK-M19.039 0.96 1.01
AK-Ml9 Ml/ izquierdo AK-Ml9.040 1.99 1.24
AK-M21 Mil derecho AK-M21.019 1.92 1.11
AK-M21 M/2 derecho AK-M21.043 1.20 1.09
Tabla 10. Medidas de los molares de Apodemus.
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
31
AK-M12.23
AK-M21.41
AK-M18.9
AK-M16.25
Figura 22. Sorex coronatus-araneus: AKMl 1.9, I inf. izq.; AK-M21.41, I inf. der. AKM12.23, M/1,2,3 der.; AK-M14.25, I inf., A/1,2 izq.; AK-M16.25, M/2,3 izq.; AKM19.45, Ml/ der.; AK-M21.24, Nl,2,3 der. Neomys fodiens-anomalus: AK-Ml 1.24, 1 inf. der. Crocidura sp.: AK-Ml.4, I sup. izq.; AK-M2.38, M/1 der.; AK-M6.26, M2/ izq. Barra de escala = 1 mm.
AK·M19.45 AK·M21.24
AK-M11.24
AK-M2.38 AK·M2.38
AK·M6.26 AK-M1.4
do a su gran parecido morfológico. Por esa razón, se agrupan estas dos especies en el grupo Terrícola sp.
Familia Muridae Género Apodemus (KAUP, 1829) Apodemus sylvaticus-flavicollis (LINNAEUS,
1758; MELCHIOR, 1834)
Material: (Figuras 19, 20 y 21 y Tablas 10 y 11). Molares atribuibles a este grupo se han recupera
do en todas las muestras estudiadas, siendo este uno de los grupos más numerosos del yacimiento.
Descripción y discusión Este género se caracteriza por tener dientes bra
quiodontos con raíces. La superficie oclusal esta for-
KOBIE (Paleoantropología n.º 27), año 2003/07
mada por seis cúspides en los primeros molares (tanto superior como inferior) y cuatro en los molares segundo y tercero. En la cara labial el Ml tiene tres cúspides accesorias, el M2 de dos a una y el M3 son reducidos Y, con la parte,posterior acuminada (CUENCA BESCOS y GARCIA PIMIENTA, en prensa).
Este grupo aparece en los yacimientos del Pleistoceno superior de Esquilleu (BAENA ET AL., 2005), Lezetxiki y Aitzbitarte IV (ALTUNA, 1972), Tito Bustillo (en el subnivel le) (ALTUNA, 1976), Erralla (PEMÁN, 1985) y Amalda (PEMÁN, 1990b) (SESÉ, 2005).
Los restos recuperados se incluyen dentro del grupo debido a su corona dentaria de gran tamaño y

32 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
Figura 23. Talpa sp.: AK-Ml4.33, M/1,2 der.; AK-Ml6.19, M2/ der.; AK-Ml9.29, M3/ izq.; AK-Ml9.62, NI y P/1,2 der.; AK-Ml9.88, M2/ der.; AK-M21.16, M/2 der.; AK-M21.13, M2/ izq.; AK-MS.106, Falange; AK-Ml2.37, humero izq. Barra de escala = 1 mm (b para AK-Ml2.37 y a para el resto).
su morfología, y por la disposición y cantidad de las cúspides del lado oclusal del molar.
En la Península Ibérica coexisten dos especies del género Apodemus, A. sylvaticus y A. flavicollis. Diferenciar estas dos especies entre ellas utilizando las características de los molares es muy difícil y por esa razón se han agrupado y descrito dentro del grupo A. sylvaticus-flavicollis. Hacen falta análisis morfométricos muy específicos para diferenciar las dos especies. En general, el tamaño del A. flavicollis es mayor que el del A. sylvaticus. Cuando las dos especies viven en simpatría en un mismo entorno, su tamaño suele ser muy parecido y es muy difícil diferenciarlos. La diferencia de tamaño es muy notable en Europa central y Norte, disminuyendo la diferencia de tamaño hacia el sur. En los territorios cercanos al Mediterráneo el tamaño suele ser muy parecido (NORES, 1988).
Subfamilia Soricinae Género Sorex (LINNAEUS, 1758)
AK-M14.33
AK-M19 62
Sorex coronatus-araneus (MILLET, 1882-LINNAEUS, 1758)
Material: (Figura 22) Se han recuperado numerosos restos de este grupo
en todas las muestras.
Descripción y discusión El color rojo de las cúspides de los dientes es un
claro indicativo de la subfamilia Soricinae. La pequeña talla y los incisivos inferiores con tres lóbulos en la superficie oclusal permite incluir este material dentro del género Sorex. Hoy en día coexisten en la Península Ibérica las especies S. araneus, S. granarius, S coronatus, S. minutus y S. minutissimus (CUENCA
AK-M21.16
BESCÓS y GARCÍA PIMIENTA, en prensa). Diferenciar entre estas especies utilizando elementos dentarios y óseos sueltos es muy difícil. Por esa razón estas especies se agrupan en dos grupos, Sorex coronatus-araneus y Sorex minutus-minutissimus. Los molares del segundo grupo son de pequeño tamaño, característica que los diferencia del otro grupo. El primer grupo se parece mucho al grupo Neomys fodiensanomalus, pero se diferencian entre ellos en el tamaño de los molares, en la cantidad de cúspides del lado oclusal de los incisivos inferiores y en la cantidad de unicuspides del lado superior.
Los restos encontrados en Antoliñako Koba se incluyen dentro del grupo S. coronatus-araneus, debido a que los incisivos inferiores tienen tres lóbulos en la superficie oclusal y por su talla, inferior a Neomys pero superior al grupo Sorex minutus-minutissimus.
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
33
Figura 24. Anguis fragilis: AK-M2.22, maxilar derecho, vista medial; AK-M2.118, Vértebra troncal, a: vista ventral, b: vista anterior; AK-M2.97; AK-M2.98; AK-M2.101; AKM2.105; AK-M2.102 y AK-M2.107, osteodermos, vistas externas. Rana sp.: AKM21.29, premaxilar, vista medial; AKMll.73, fragmento de maxilar, vista medial; AK-Ml4.38, fragmento de maxilar; vista medial. Rana temporaria-iberica (Grupo de ranas pardas): AK-Mll.102, Húmero derecho de un macho, vista ventral; AK-M21.57, escápula derecha, vista ventral. Amphibia sp.: AK-M4.139, falange. Barra de escala= 1 mm.
AK-M222
Género Neomys (KAUP, 1829) Neomys fodiens-anomalus
(PENNANT, 1771-CABRERA, 1907)
AK-M21.29
Material: (Figura 22) El material que corresponde a
este grupo se ha recuperado en la muestra AK-Mll y se trata de un incisivo inferior.
Descripción y discusión El incisivo es alargado y fino,
con punta de color rojo y con un solo lóbulo en la superficie oclusal.
AK-M11.73
Neomys sp. aparece en los yaci- AK-M2.11aa
mientos de la Cornisa Cantábrica del Pleistoceno Superior de An:Jalda (niveles N y VI) (PEMAN, l 990b) y Larninak II (en el nivel II) (PEMÁN, 1994) (SESÉ, 2005). En la Península Cantábrica coexisten dos especies de este género, N. fodiens y N. anomalus.
Las dos especies que constitu-yen este grupo son muy difíciles de diferenciar utilizando las características de los restos sueltos, y por esa razón se agrupan y describen como grupo.
Subfamilia Crocidurinae Género Crocidura (WAGLER, 1832) Crocidura sp.
Material: (Figura 22) Se han recuperado restos de este género en las mues
tras AK-Ml, AK-M2, AK-M6, AK-Ml6 y AK-Ml9.
Descripción y discusión Los restos de este género en morfología y en talla
son muy parecidos a los del género Sorex. La principal diferencia entre estos dos géneros es gue ffilentras Sorex presenta una coloracion rojiza característica en
KOBIE (Paleoantropología n.º 27), año 2003/07
AK-M11.102
AK·M14.-:l8
AK-M2.107
AK-M2.97 AK-M2.101
AK-M4.139
AK·M2.105 -sus molares Crocidura carece de ella. El material del que se disp~ne no permite realizar una determinación más precisa que a nivel de género.
Familia Talpidae Género Talpa (LINNAEUS, 1758) Talpa sp.
Material: (Figura 23) Los restos de Talpa sp. se han recuperado en las
muestras AK-Ml, AK-M2, AK-M5, AK-M6, AK-Ml l, AK-Ml4, AK-Ml6, AK-Ml8, AK-Ml9 y AK-M21.
Descripción y discusión El material atribuido a este género esta compues
to por varios molares sueltos, fragmentos de mandíbula, húmeros y terceras falanges. El tamaño de l?s molares y lo característico del postcraneal de los tál-
---· ·--------------··---- --------··------------------------------------~---·~-~ .... ·-~~~---~~---=--=· ~-

34 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
pidas permite su adscripción al género Talpa. Para poder precisar a nivel de especie no se dispone de suficiente material.
Clase ''Reptilia" Orden Squamata Suborden Lacertilia Familia Anguidae Género Anguis (LINNAEUS, 1758) Anguis fragilis (LINNAEUS, 1758)
Material y descripción: (Figura 24) El material de esta especie es muy abundante en la
mayoría de las muestras, sobre todo en las más jóvenes. Se han recuperado osteodermos, maxilares, manchbulas, vértebras troncales, caudales y cervicales y costillas. De todo el material recuperado en las muestras más jóvenes los más abundantes son los osteodermos.
Desde el punto de vista morfológico, el material no difiere de la especie actual A. fragilis, único representante de la familia en la Península Ibérica. Para conocer los caracteres morfológicos de la especie consultar, entre otros, los trabajos de HOFFSTETTER (1962) o BAILON (1991).
Familia Lacertidae Género Podareis (WAGLER,
1830) cf. Podareis
Material y descripción: (Figura 24)
El material identificado de este grupo es muy escaso. El material atribuido a este grupo aparece en las muestras AK-M2, AK-M4, AK-M6 y AKMl9 y esta compuesto por premaxilares, maxilares y dentarios portadores de dientes bien característicos de la fami-
Figura 25. Vipera o Namx: AK-M2.122, vértebra troncal, a: vista anterior, b: vista posterior, c: vista dorsal, d: vista ventral. Coronel/a austriaca: AK-M4.155, vértebra lumbar, a: vista ventral (invertir la figura de 180º), b: vista anterior. Rana tempora-ria-iberica (Grupo de ranas pardas): AK-Ml 1.105, Vértebra sacra, a: vista posterior, b: vista ventral; AK-Mll.108, Esfenetrnoideo, vista ventral; AKMl 1.123, vértebra, a: vista anterior, b: vista posterior. Rana sp.: AK-Mll.113, 8. vértebra, a: vista dorsal, b: vista posterior, c: vista anterior. Barra de escala = 1 mm.
lia. Las dimensiones de los elementos pueden adscribirse, con reservas, a un representante del género Podareis.
Orden Serpentes Familia Viperidae o Familia Colubridae Género Vipera (LAURENTI, 1768) o género
Namx LAURENTI, 1768
Material y descripción (Figura 25) El único resto identificable es una vértebra troncal
recuperada en la muestra AK-M2.
Esta vértebra se caracteriza por la presencia de hipapófisis. Las superficies articulares de las zigapófisis se inclinan dorsalmente, el centro vertebral posee márgenes laterales difusos y el cotilo y el condilo están bastante desarrollados. Estos caracteres se encuentran en los representantes del género Vipera.
AK·M4155b AK-M4.155a
AK-M11.113a
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BlZKAIA)
35
Sin embargo; esta vértebra posee, en vista posterior, un arco neural anormalmente abombado, más característico del género Natrix que de Vipera. Una revisión del material, estudiado a partir de una fotografía, se impone abtes de avanzar una atribución más precisa.
Familia Colubridae Subfamilia Colubrinae Género Coronella (LAURENTI, 1768) Coronella austriaca (LAURENTI, 1768)
Material y descripción: (Figura 25) El material identificado de esta especie aparece en
las muestras AK-M4 y AK-M5. Los restos recuperados son vértebras troncales.
La ausencia de hipapófisis permite adscribir estas vértebras a un colúbrido no natricino y la atribución a C. austriaca está basada, entre otros caracteres, en la presencia de un arco neural aplastado dorso-ventralmente y de parapófisis más desarrolladas que las diapófisis (SZYNDALR, 1984).
Clase Amphibia Familia Salamandridae Salamandridae indet.
Material y descripción: El único resto identificable posiblemente atribui
ble a esta familia, es una vértebra bastante fragmentada que se ha encontrado en la muestra AK-M4 y un atlas y una vértebra recuperados en las muestras AKMl6 y AK-Ml8 que también podrían pertenecer a esta familia.
Familia Ranidae Subfamilia Raninae Género Rana (LINNAEUS, 1758) Rana temporaria-iberica (LINNAEUS, 1758-
BOULENGER, 1879)
Material y descripción: (Figura 24) Restos de este taxón son bastante numerosos y
aparecen en las muestras AK-M6, AK-Mll, AKMl4, AK-Ml6, AK-Ml9 y AK-M21. Entre el material recuperado se encuentran iliones, escápulas, vértebras, esfenetmoideos y húmeros cuya morfología concuerda con la de este grupo de ranas pardas [para los caracteres distintivos de los diferentes representantes del género Rana consultar por ejemplo, HOLMAN (1998), SANCHIZ (1998) y BAILON (1999)].
Aparte del material perteneciente a este grupo, son muy numerosos los restos atribuibles al género Rana en general (radio-ulnas, coracoides, tibio-fíbu-
KOBIE (Paleoantropología n.º 27), año 2003/07
las, fémures, vértebras, falanges y fragmentos cranianos ). Estos aparecen en casi todas las muestras, y son bastante abundantes. Algunos de estos restos parecen pertenecer al grupo de las ranas verdes.
Familia Bufonidae (GRAY, 1825) Subfamilia Bufoninae (GRAY, 1825) Género Bufo (LAURENTI, 1768) Bufo sp.
Material y descripción El material de este género se ha recuperado en las
muestras AK-Ml8, AK-Ml9 y AK-M21, y está constituido por varias vértebras, dos fragmentos de húmeros y un urostilo. La morfología general de estos elementos concuerda bien con la establecida para los representantes de este género (SANCHIZ, 1977, BAILON, 1999) aunque su atribución específica no puede ser establecida.
Clase Osteicthyes Osteicthyes indet.
Material y descripción: Los restos de este grupo aparecen en casi todos las
muestras, y están constituidos principalmente por vértebras. Se han recuperado en las muestras AK-M2, AK-M4, AK-M5, AK-M6, AK-M9, AK-Mll, AKMl2 y AK-Ml9.
Clase Aves Aves indet.
Material y descripción: Elementos de este grupo de vertebrados se han
recuperado en las muestras AK-M2, AK-M4, AKMll, AK-Ml4, AK-Ml8, AK-Ml9 y AK-M21. El material está constituido por una falange proximal y fragmentos de falanges, metacarpos y cuadrados.
PALEO ECOLOGÍA
La Paleoecología es la ciencia que estudia la relación existente entre los organismos y el entorno en el que vivieron en el pasado (BRENCHLEY, 1998). Esta relación es muy estrecha, y un pequeño cambio de las condiciones del entorno acarrea cambios de la fauna y flora. Gracias a los fósiles, se puede conocer la fauna y flora de un lugar concreto, y con los cambios que sufren en el tiempo, se pueden conocer los cambios ambientales que acontecieron en el lugar durante el tiempo. Para realizar los análisis paleoecológicos hay que tener en cuenta la historia sedimentológica del lugar y la historia tafonómica de la asociación fósil a estudiar.

36 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
Características geológicas y tafonómicas La columna sedimentaria de la cueva está consti
tuida principalmente por arenas y limos, pero de vez en cuando también aparecen sedimentos más gruesos. Estos son brechas, bloques y algún canto rodado. Entre capas de sedimento aparecen estructuras creadas por circulación de agua, como canales y cubetas. Los fósiles recuperados están muy fracturados, y apenas se han recuperado restos de microfauna en estado completo. Como se ha visto en el apartado de consideraciones tafonómicas, los elementos anatómicos más numerosos son las falanges y dentarios sueltos, debido al transporte sufrido (ANDREWS, 1990; ANDREWS & EVANS, 1983).
La información recuperada indica que la mayoría de los sedimentos y fósiles han sido transportados por procesos físicos naturales. Los fósiles acumulados serán adémicos, acumulados fuera del entorno en el que vivían. Pero también pueden existir fósiles no acumulados por procesos físicos naturales. Estos fósiles pueden haber sido transportados por un depredador al interior de la cueva.
Muchos restos recuperados presentan disoluciones químicas producidas por jugos gástricos. Estas disoluciones se aprecian alrededor de los restos óseos y dentarios, y afectan el resto desde el exterior hacia el interior. Estas disoluciones indican que el individuo ha sido cazado por un depredador.
Autoecología Los distintos taxones tienen necesidades ecológi
cas muy específicas. Las condiciones de un entorno pueden cambiar con mucha facilidad en el tiempo, y los taxones que allí vivían no podían más que cambiar de entorno o adaptarse a las nuevas condiciones.
A continuación se describen las afinidades ecológicas de los distintos géneros, grupos y especies descritos en este trabajo. Se han tomado en cuenta los micromamíferos, anfibios y reptiles.
-Eliomys quercinus Esta especie vive en la Europa templada y medi
terránea, Norte de África y Levante.
En la Península Ibérica, vive en bosque y zonas de matorrales (CUENCA BESCÓS y GARCÍA PIMIENTA, en prensa). Se adapta a distintos hábitat, y es muy numeroso en los bosques caducifolios y de coníferas de los Pirineos y los Alpes de hasta 2000 metros. También aparece en zonas más bajas y pedregosas de matorrales (SESÉ, 2005). Se considera como especie termófila.
Durante el Pleistoceno, su presencia está relacionada con bosques de clima templado (SESÉ, 2005), y normalmente no aparece en épocas de climas fríos.
- Clethrionomys glareolus: Esta especie vive en la Eurasia templada, desde el
lago Baikal hasta las Islas Británicas. En la Península Ibérica vive sobre todo en la zona Norte (VENTURA et al., 1993; POKINES, 1998), pero también aparece en las zonas montañosas de Levante, debido a su clima Eurosiberiano (ARRIZABALAGA ET AL., 1986).
Es de los únicos arvicólidos que vive en bosques caducifolios. Vive en zonas de más de 800 milímetros de precipitación anual (SESÉ, 2005). Esos bosques suelen ser bastante abiertos, con vegetación arbustiva y herbácea (CUENCA BESCÓS y GARCÍA PIMIENTA, en prensa). Se considera especie termófila.
Durante el Pleistoceno su presencia se relaciona con zonas boscosas y zonas con mucha vegetación en climas templados y húmedos (PEMÁN, 1985, 1990a).
- Microtus arvalis-agrestis: Es muy difícil diferenciar entre estas dos especies,
e imposible cuando la población y el material recuperado son escasos.
La especie M. arvalis vive en muchas zonas de Europa, menos en Gran Bretaña (excepto en las islas Orkney), Irlanda y en la Península Escandinava. Hacia el Norte aparece hasta el paralelo 60º, en el sur hasta los montes Ural y Altai y en el Este hasta Asia central (SESÉ, 2005).
Es más oportunista que M. agrestis, y aparece más repartido (CUENCA BESCÓS y GARCÍA PIMIENTA, en prensa). En la Península Ibérica aparece en el Sistema Ibérico, Sistema Central, Meseta Norte y Norte de la Península, pero nunca en la zona mediterránea. Vive en zonas abiertas, en zonas con numerosa vegetación arbustiva y herbácea (SESÉ, 2005).
La especie M. agrestis tiene gran distribución en Europa. Vive más al Norte que la especie M. arvalis. En el Oeste de Asia y Europa aparece entre los paralelos 40º y 70º. En la Península Ibérica aparece en la zona Norte y no en la zona mediterránea.
Aparece en las zonas donde predomina el clima centroeuropeo, desde bosques hasta zonas abiertas de poca vegetación; praderas húmedas, juncales ... (SESÉ, 2005). En la Península Ibérica es buen indicador del clima Atlántico, y aparece desde encinares hasta hayedos (ARRIZABALAGA ET AL., 1986).
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
37
Estas dos especies no tienen determinaciones ambientales muy específicas, y no son muy utilizados a la hora de definir la paleoecología (PEMÁN, 1985).
- Microtus oeconomus Actualmente, la especie se encuentra en las zonas
Nortes de Eurasia; Holanda, Alemania, Polonia, Escandinavia y toda Siberia, pero no en la Península Ibérica.
Vive en zonas muy húmedas como praderas, juncales y zonas pantanosas (SESÉ, 2005). Es una especie muy común en zonas de Taiga y Tundra. Debido al reparto periártico que tiene la especie, algunos autores (CHALINE, 1972; PEMÁN, 1985), relacionan las fluctuaciones que tiene la población durante el Pleistoceno con cambios climáticos; en las etapas climáticas más frías será más numeroso, constituyendo casi la mitad de la población total (PEMÁN, 1985). Pero también aparece en las etapas climáticas más templadas, aunque en proporciones mucho más pequeñas. Por esa razón, no hay que tomar la mera presencia de esta especie en una asociación faunística determinada como indicador climático, sino más bien su proporción respecto a los demás taxones de rnicromamíferos de la asociación faunística. Vive en competencia con la especie M. arvalis, y ante la presencia de esta especie, M. oeconomus se adapta a condiciones más frías y húmedas (CUENCA BESCÓS y GARCÍA PIMIENTA, en prensa).
- Terricola sp. Es muy difícil diferenciar las especies de este
género, y por esa razón se describen las afinidades ecológicas del género.
Las especies de este género viven en zonas de suelos profundos y húmedos y zonas de pradera con mucha vegetación.
- Chionomys nivalis Viven en populaciones aisladas de Europa y Asia;
en los Alpes, montes Tatra, Carpatos, Balcanes y en el Caucaso. En la Península Ibérica vive en los montes de Sierra Nevada y montes del Norte. Esta especie necesita zonas pedregosas y soleadas.
Es una especie bien adaptada a zonas de alta montaña, pero también aparece cerca del nivel del mar, en Cantabria, en la localidad de Ramales de la Victoria. Normalmente vive en laderas orientadas al Sur, en zonas abiertas y pedregosas de poca vegetación.
Con la distribución que presenta esta especie en la actualidad y su distribución del Cuaternario, KOWALSKI (1992) la relaciona con zonas abiertas más que con zonas de alta montaña.
KOBIE (Paleoantropología n.º 27), año 2003/07
- Arvicola terrestris Esta especie tiene una distribución paleártica
encontrándose en casi toda Eurasia, empezando desde la costa mediterránea hasta el Océano Glacial Ártico, atraviesa parte de Siberia y casi llega hasta el Pacífico. En Asia su distribución llega hasta Palestina, el Sur de Irán y noroeste de China (SESÉ, 2005).
En la Península Ibérica viven en populaciones aisladas en el Norte; en los Pirineos y el Norte de Gipuzkoa, empezando desde el Norte de Bizkaia hasta la Sierre de los Aneares de Lugo, y el Norte de Portugal.
Esta especie necesita suelos profundos y húmedos para sobrevivir. Vive en alta montaña y en praderas, a distintas altitudes. También aparece cerca de los ríos y arroyos, pero nunca en bosques densos (SESÉ, 2005). Con la presencia de la especie Arvicola sapidus, ocupa zonas más lejanas de los ríos y arroyos (CUENCA BESCÓS y GARCÍA PIMIENTA en prensa).
- Arvicola sapidus Aparece en la mayor parte de Francia y toda la
Península Ibérica.
Vive en zonas húmedas, cerca de circulaciones constantes de agua, en zonas de vegetación herbácea y matorrales.
-Apodemus sylvaticus-flavicollis Es difícil diferenciar las especies de este género, y
sobre todo las especies de este grupo. Por esa razón es normal que se describa como grupo (SESÉ, 2005). Este grupo está muy distribuido en Europa y la Península Ibérica, pero A. flavicollis tiene menor distribución en Europa, y en la Península Ibérica aparece sólo en la zona Norte, desde los Pirineos hasta Asturias. A. sylvaticus aparece en toda la península, y es más numeroso en la zona mediterránea.
Durante el Pleistoceno, las especies de este grupo son indicadores de climas cálidos, son termófilos, y buenos indicadores de zonas boscosas. Las variaciones que sufre la población a lo largo del tiempo son utilizadas como indicadores climáticos. La población suele aumentar cuando el clima es más templado y disminuye cuando el clima es más frío. Estas variaciones de la población también son utilizadas como indicadores de cambios sufridos por los bosques (PEMÁN, 1990).
- Sorex coronatus-araneus Las tres especies del género Sorex viven en la
zona norte de la Península Ibérica; la especie S. araneus vive al Norte de Cataluña, desde los Piri-

38 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
neos hasta el Prepirineo Catalán. S. coronatus vive desde los Pirineos hasta Galicia, y al sur hasta el Sistema Ibérico. S. granarius vive desde el Sistema Central hasta la desembocadura del Tajo, y hasta Galicia (LÓPEZ-FUSTER & VENTURA, 1996).
Las tres especies viven en zonas húmedas con abundante vegetación tipo herbácea, arbustiva o arborea (POKINES, 1998; CUENCA BESCÓS y GARCÍA PIMIENTA, en prensa).
- Neomys fodiens-anomalus: La especie N. fodiens está muy repartida en Euro
pa, y en la Península Ibérica vive en la zona norte. La especie N. anomalus está relacionada con las zonas montañosas de Europa central y este, y de la Península Ibérica (BLANCO, 1998).
Las dos especies están relacionadas a masas de agua, lagos, ríos y arroyos, pero en menor medida la
lfoi;cá~ Hmquc füt"jUC >
\iutonul Pra<km
f:bt.>m }S <ttll!ICll'.IUS X
Clcllm1m1>mJi~ ¡¡la""~us X
\!icmll1~ .. , , "' . -,., ..
\lkmlll~ ~iernnt1rnos
Tcnlcula sp.
CJ11ui11.1m~.,; nind1~
,\f\'~~~.a tc.~lri~
Ar't1cola llllp«lu~
A¡xidernns sylvullcu~·ll.A\'Írnlli* X
Suric:m~~ X
Surex cuninal111> Mallcll::! X
S(Offi)l' r1:..1k-llii-iU!t>malu~
Cl'IXi<lu;;i.~ X
'folpasp.
especie N. anomalus. También puede estar relacionado con bosques y praderas húmedas (CUENCA BESCÓS y GARCÍA PIMIENTA, en prensa).
Las especies de este género son más numerosas en climas templados y húmedos, pero han tenido que adaptarse a climas más fríos, debido al cambio climático que se dio a partir del Villafranquiense. (SESÉ, 2005).
Este grupo es indicador de zonas húmedas, y vive en zonas de praderas y bosques (SESÉ, 2005).
- Crocidura sp. Las especies del género Crocidura que viven en la
Península Ibérica son C. suaveolens y C. russula. La primera vive más al norte, en un clima Atlántico, más húmedo (Cuenca Bescós y García Pimienta, en prensa). La segunda es muy abundante en la Península Ibérica, incluso en zonas con influencia humana. En
l'lud.:141 lonas /.unus f'radcrn Semi·
~blumll ,\cu:~~ de
Ando Hllmc!b i\cmitica~ M1.mtaim ht;U
X
X
X
X
X
X
X
X
Tabla 12. Afinidades ecológicas de los diferentes micromamíferos recuperados.
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
39
la zona mediterránea es bastante abundante y disminuye la proporción en zonas montañosas, donde se adapta a zonas de poca altura. Se considera especie termófila con necesidades mediterráneas para sobrevivir. Está relacionada con zonas boscosas no densas, praderas y zonas arbustivas (RZEBIK-KOWALSKA, 1995; BLANCO, 1998; PALOMO y GISBERT, 2002; SESÉ, 2005). Necesita biotopos abiertos y secos, pero también aparece en zonas boscosas y arbustivas.
Crocidura sp. tiene necesidades ambientales muy parecidas. Según RZEBIK-KOWALSKA (1995), el género Crocidura es indicativo de un clima templado. Este género está adaptado a zonas más secas y cálidas que los sorícidos (RZEBIK-KOWALSKA, 1995; SESÉ, 2005).
-Talpa sp. Es un género muy extendido en Europa, menos
desde Irlanda y la Península de Escandinavia, hasta el oeste de Siberia. En la Península Ibérica aparece en el noreste del territorio, empezando desde Asturias hasta ,Cataluña, y hasta el Sistema Central en el sur (SESE, 2005).
Las especies del género Talpa son insectívoros y cavadores, y por eso necesitan suelos profundos para sobrevivir (CUENCA BESCÓS y GARCÍA PIMIENTA, en prensa). Son euritermos, y suelen estar relacio-
80%
60%
40%
20%
0%
A B
Figura 26. Variaciones relativas de temperatura en los distintos niveles.
KOBIE (Paleoantropología n.º 27), año 2003/07
nados con grandes praderas húmedas. Por esa razón, la presencia del género puede tomarse como indicativo de humedad (MURELAGA et al., 2005). En un principio, estaban relacionados con zonas boscosas, pero han tenido que adaptarse a las praderas (KOLFSCHOTEN, 1995).
- Anguis fragilis Especie ampliamente distribuida en Europa y que
penetra en Asia hasta el nosoeste de Irán y el oeste de Siberia. En la Penísula Ibérica, se encuentra en su mitad Norte. Es un taxón hygrófilo, distribuido en diversos hábitat de la región Eurosiberiana ibérica. Vive en zonas húmedas y sombrías con abundante vegetación con estrato herbáceo más o menos denso, bosques caducifolios y mixtos y matorrales (BARBADILLO y SÁNCHEZ-HERRÁIZ, 1997; MURELAGA ET AL., 2007).
- cf. Podareis Las especies de este género en la Cornisa Cantá
brica viven en zonas rocosas. También aparecen en bosques, praderas y zonas de matorral. Es un género de clima templado, y suele huir de las temperaturas frías, por eso no aparecen en altas altitudes.
- Corone/la austriaca Especie con amplia distribución europea que
hacia el este, alcanza Asia. En la Penísula Ibérica se distribuye principalmente en la región Eurosiberiana
e o

40 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
0% .,.... N -.:;!" l() <O m .,.... N ::;=! ::;=! ::;=! ::;=! ::;=! ::;=! ..- ..-
~ ~ 1
~ ~ 1 ::;=! ::;=!
~ ~ ~ ~ <( <( <( <( <( <( <( <(
y en algunas cordilleras de centro y del sur, prueba de una distribución más amplia en el pasado. Habita zonas con precipitaciones superiores a 600-800 milímetros y con una gran variedad de hábitat con cobertera vegetal abundante. En la Cornisa Cantábrica vive en zonas con densa vegetación o matorrales; en praderas, límites de bosques caducifolios y mixtos, zonas litorales arenosas.
- Rana temporaria-ibérica Grupo de especies de la región Eurosiberiana y
características de la Iberia húmeda que habitan prin-
A B
Figura 28. Variación del NMI del género Apodemus en cada nivel.
-.:;!" ..-::;=! ~ <(
<O 00 .,.... .,.... ::;=! ::;=!
1
~ ~ <( <(
m ..-::;=!
1
~ <(
..-N ::;=!
1
~ <(
Figura 27. Variaciones relativas de temperatura en las distintas muestras.
cipalmente las zonas húmedas con abundante vegetación.
-Bufo sp. Las especies de este género viven en toda la
Península Ibérica, pero sobre todo aparece en la mitad norte y asociado a circulaciones de agua. Estas especies se han adaptado a distintas condiciones.
Las distintas especies y géneros de micromamíferos se han agrupado en ocho grupos según sus preferencia ecológicas (Tabla 12).
e D
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
41
4
3
2
o ..- N "'1" l.(") e.o (j)
::.'.? ::.'.? ::.'.? ::.'.? ::.'.? ::.'.? ' ' ' ~ ' ' ::!:'.'. ::!:'.'. ::!:'.'. ::!:'.'. ::!:'.'.
<( <( <( <( <( <(
Figura 29. Variación del NMI del género Apodemus en cada muestra.
Figura 30. Variaciones relativas de humedad en los distintos niveles.
A
- 1. Especies termófilas asociadas a bosques: Clethrionomys glareolus Género Apodemus
- 2. Especies asociadas a matorrales y bosques: Crocidura sp (género de clima seco mediterráneo
o termófilo) Eliomys quercinus (durante el Pleistoceno se aso
cia a bosques y climas templados)
- 3. Especies asociadas a bosques y praderas: Sorex coronatus-araneus (en climas centroeuro
peos o Atlánticos húmedos) Soricinae sp.
KOBIE (Paleoantropología n.º 27), año 2003/07
..-::.'.? ' ::!:'.'.
<(
B
N "'1" e.o co (j) ..-...-- ..- ...-- ...-- ...-- N ::.'.? ::.'.? ::.'.? ::.'.? ::.'.? ::.'.? ~ ~ ' ' ' ' ::!:'.'. ::!:'.'. ::!:'.'. ::!:'.'. <( <( <( <( <( <(
e D
- 4. Especies asociadas a praderas y matorrales de climas secos:
Microtus arvalis-agrestis (M. agrestis también aparece en bosques)
- 5. Especies asociadas a praderas húmedas: Talpa sp. (En un principio se relaciona con zonas
boscosas) Terrícola sp. Arvicola terrestris
- 6. Especies asociadas a praderas húmedas, juncales y masas acuosas de climas fríos:
Microtus oeconomus
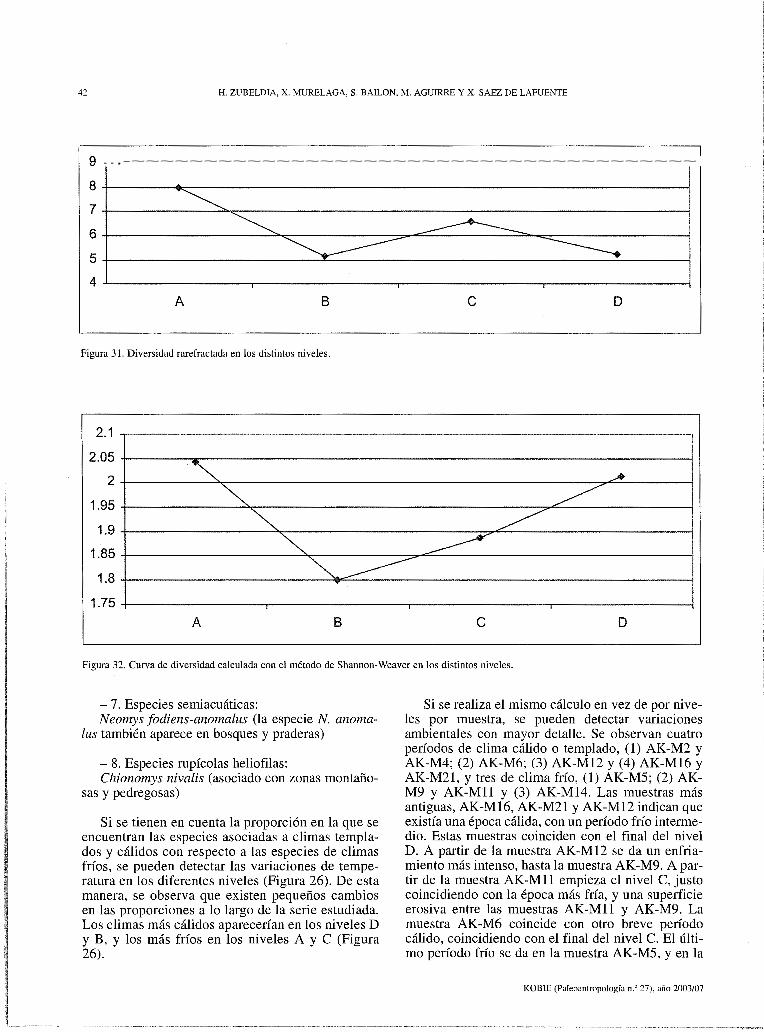
42 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
9 ..,-~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
8 -r-~~~-.o:::-~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~--;
7 -t-~~~~~~-""~~~~~~~~~~~~~~~~~~~~~~~~~~~~--l
6 +-~~~~~~~~~--""-o::-~~~~~~-=--::::;:_~~~~__;::::::.........,,,~~~~~~~~--l
5 -r-~~~~~~~~~~~~~<--~~~~~~~~~~~~~~~~~-'-~~~--l
4 -'-~~~~~~~~--.-~~~~~~~~-.-~~~~~~~~-.-~~~~~~~~--l
A B e D
Figura 31. Diversidad rarefractada en los distintos niveles.
2.1
2.05
2
1.95
1.9
1.85
1.8
1.75 A B e D
Figura 32. Curva de diversidad calculada con el método de Shannon-Weaver en los distintos niveles.
- 7. Especies semiacuáticas: Neomys fodiens-anomalus (la especie N. anoma
lus también aparece en bosques y praderas)
- 8. Especies rupícolas heliofilas: Chionomys nivalis (asociado con zonas montaño
sas y pedregosas)
Si se tienen en cuenta la proporción en la que se encuentran las especies asociadas a climas templados y cálidos con respecto a las especies de climas fríos, se pueden detectar las variaciones de temperatura en los diferentes niveles (Figura 26). De esta manera, se observa que existen pequeños cambios en las proporciones a lo largo de la serie estudiada. Los climas más cálidos aparecerían en los niveles D y B, y los más fríos en los niveles A y C (Figura 26).
Si se realiza el mismo cálculo en vez de por niveles por muestra, se pueden detectar variaciones ambientales con mayor detalle. Se observan cuatro períodos de clima cálido o templado, (1) AK-M2 y AK-M4; (2) AK-M6; (3) AK-Ml2 y (4) AK-Ml6 y AK-M21, y tres de clima frío, (1) AK-M5; (2) AKM9 y AK-Mll y (3) AK-Ml4. Las muestras más antiguas, AK-Ml6, AK-M21 y AK-Ml2 indican que existía una época cálida, con un período frío intermedio. Estas muestras coinciden con el final del nivel D. A partir de la muestra AK-Ml2 se da un enfriamiento más intenso, hasta la muestra AK-M9. A partir de la muestra AK-Mll empieza el nivel C, justo coincidiendo con la época más fría, y una superficie erosiva entre las muestras AK-Mll y AK-M9. La muestra AK-M6 coincide con otro breve período cálido, coincidiendo con el final del nivel C. El último período frío se da en la muestra AK-M5, y en la
KOBIE (Paleoantropología n.º 27), año 2003/07
MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
43
muestra AK-M4 es de nuevo cálido. Estas dos muestras forman el nivel B. En las muestras AK-M2 y AK-Ml se da otro enfriamiento, pero menor que las anteriores. Estas dos muestras coinciden con el nivel A (Figura 27).
Las variaciones en el NMI de Apodemus sylvaticus-flavicollis es un indicativo de cambios en el entorno. En las épocas más cálidas aumenta la cantidad de individuos de este grupo, y en las épocas más frías disminuye. La mayor cantidad de individuos se encuentran en los niveles B y D, coincidiendo con las épocas más cálidas, y la menor en los niveles A y C, las épocas más frías (Figura 28).
El mayor número de individuos de este grupo aparece en las muestras AK-M2-AK-M4 y AK-M19,
Zonas
Polen
Preboreal
Dryasm
AllerOd
Dryas ll
Bolllng
Dryas Je
Pre-Bolllng
Dryas lb Anglas
Dryas la
Lascaux
Laugerle
Tursac
Kesselt
(POKINES, 1998)
Variación relativa del nivel del mar C14 BP
Cilldo Fr1o Cálido
OIS '!',;
10.000
12.000
14.000
16.000
18.000
20.000
22.000
24.000
26.000
28.000
30.000
coincidiendo con las muestras más cálidas (Figura 29) y con la presencia de representantes del género Podareis.
También se han estudiado los cambios de humedad del entorno en los distintos niveles y muestras, relacionando las especies y grupos asociados a climas húmedos y secos. Se puede observar que del nivel D al A disminuye el grado de humedad levemente, pero en general el clima es muy húmedo en todos los niveles. Los niveles más húmedos son el B y D, y los menos húmedos el A y C (Figura 30).
Sinecología Normalmente cuando se da un cambio importante
del entorno que rompe el equilibrio del ecosistema, como un cambio climático, el índice de diversidad
(ZUBELDIA, 2006)
Frlo
~ ~ 10.220
~ 10.800
14.580 ~ 14.630 É 14.680
: ¡ 16.500
. ¡ 19.020 . 19.280
~ ;:;
~ ~ ¡;; 26.710
~ 27.390 ~ 27.520
~ ~ 29.990
suele disminuir considerablemente. Cuando el clima y el equilibrio se recuperan, la diversidad aumenta y se recupera.
Para estudiar las diversidades de las distintas muestras y niveles, se han calculado los índices de diversidad ShannonWeaver y curvas de rarefracción, por niveles y muestras. Los valores conseguidos de los cálculos de índice de diversidad por muestras no son fiables, debido a la poca cantidad de restos e individuos. La cantidad de restos e individuos de algunos niveles sigue siendo escaso, sobre todo en los niveles A, B y C.
Las curvas de rarefracción indican un aumento de diversidad del nivel D
Arcy
32.000 ~·--------- ------· -~---~--.-·----- ; 34.000
Cottés
36.000
38.000 Hengelo
KOBIE (Paleoantropología n.º 27), año 2003/07
3 Figura 33. Variaciones relativas del nivel del mar durante el Pleistoceno superior comparados con las variaciones de temperatura calculados en Antoliñako Koba (modificado de Pokines, 1998 y de Zubeldia 2006).

44 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
C14(BP)
10220t40
10800*40
14580t-70 1463Qt70 14680-t-80
19280t120
2671Qt-1B0
27390*320
27520*190
2999{h230
Nivel Mue5tra
A AK-M1
K-M2
B K-M4
AK-M5
K-M6
e AK-M9
K-M11
K-M12
K-M14
D
K-M16
K-M18
K-M19
Figura 34. Variación de las proporciones de los diferentes micromamíferos de Antoliñako Koba en las diferentes muestras y niveles.
al A. La diversidad aumenta del nivel D al C, para disminuir posteriormente del C al B, y aumentar considerablemente del nivel B al A. Los mayores
valores de diversidad se dan en el nivel A, y los valores de menor diversidad se dan en el nivel B (Figura 31).
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
45
Las curvas de diversidad Shannon-Weaver indican otra tendencia. Del nivel D al B disminuye considerablemente la diversidad, para aumentar luego del nivel B al A. Los mayores valores de diversidad se dan en el nivel A, y los valores menores se dan en el nivel B (Figura 32).
Si se comparan las dos curvas, se observa que existe una diferencia entre ellas. En las dos se observan parecidas tendencias, pero existen diferencias en el nivel D, En las curvas Shannon-Weaver, este nivel presenta un valor mayor que en los niveles B y C, y parecido al nivel A. En las curvas de rarefracción, el nivel D tiene valores menores que el nivel C, y parecido al nivel B. Estas diferencias se pueden explicar por la escasez de restos e individuos con los que se está trabajando. A la hora de elegir entre estos dos valores obtenidos con estos dos índices son más fiables las curvas de rarefracción para calcular la diversidad ya que estas se utilizan cuando la muestra es muy pequeña.
Conclusiones paleoecológicas Al comparar las curvas que relacionan la propor
ción de individuos de clima cálido y frío, y aquellas donde se refleja la cantidad de individuos del género Apodemus, se observa que son muy similares (Figuras 26 a 29). Las muestras y niveles más cálidos coinciden con los niveles y muestras con mayor número de individuos de este género.
Si se comparan las curvas que relacionan la proporción de individuos de climas húmedos y no húmedos con las curvas anteriores, se observa que también se parecen (Figuras 26 a 30). Las muestras y niveles más húmedos coinciden con las muestras y niveles más cálidos y con mayor número de individuos del género Apodemus. Normalmente, las épocas de clima cálido o templado suelen coincidir con épocas húmedas, y épocas de clima frío, con épocas más secas. La evolución ambiental que se puede observar desde el nivel más antiguo (D), es un comienzo con clima cálido y húmedo, cambia a un clima frío y más seco (C), posteriormente se pasa de nuevo a un clima cálido y húmedo (B) y por último cambia de nuevo a más frío y más seco (A).
La falta de concordancia entre los datos autoecológicos y sinecológicos puede ser debido a factores tafonómicos.
Entre estos factores, uno puede ser el tamaño de las muestras. Para conseguir valores fiables en los índices de diversidad, el tamaño de las muestras tiene que ser mayor que las estudiadas para este trabajo.
Otro factor puede ser la presencia e influencia de un agente biológico que ha generado la tanatocenosis.
KOBIE (Paleoantropología n.º 27), año 2003/07
Cuando las condiciones del entorno son benignas para alguna especie concreta cuya caza le sea más provechosa al depredador, esta especie estará sobrerepresentada en el yacimiento, disminuyendo la diversidad real. Cuando las condiciones no son tan favorables, y las presas no abundan, en el yacimiento que creará estará mejor representada la diversidad del entorno.
Por ejemplo, la especie Bubo hubo (Buho real) cuando vive en bosque, en sus egagropilas está bien representada la diversidad de la fauna de microvertebrados que vive en el bosque. Pero cuando vive en el desierto, aparecen sobrerepresentados aquellos micromamíferos de mayor biomasa (ANDREWS, 1990).
Las curvas obtenidas de los datos paleoecológicos en este trabajo se pueden correlacionar mediante las dataciones de C14 con las curvas climáticas del Pleistoceno superior obtenidas en otras localidades. Las variaciones observadas con la información autoecológica son similares a los cambios climáticos acaecidos en el Pleistoceno superior (Figura 33).
La asociación de micromamíferos encontrada en Antoliñako Koba muestra que al igual que en la actualidad, convivían especies y géneros con distintas preferencias ambientales. En general aparecen especies y géneros típico~ de Clima Atlántico o Centroeuropeo húmedo (SESE, 2005).
CONCLUSIONES
En la secuencia sedimentaria estudiada se han diferenciado cuatro niveles estratigráficos y sedimentarios que de más joven a más antiguo son: A, B, C y D.
De las 22 muestras obtenidas en la campaña de 2005 se han estudiado en su totalidad 13. Estas muestras han supuesto el lavado-tamizado de 55 kg. de sedimento, el triado con la lupa de 11.44 kg de concentrado y la recuperación de 16.884 restos óseos.
Se han identificado 22 especies; 14 micromamíferos, 4 anfibios, 4 reptiles, así como peces y aves no determinados.
Después de estudiar la conservación y proporción de los distintos elementos anatómicos recuperados, se ha llegado a la conclusión de que los restos fueron transportados al lugar donde se sedimentaron.
Con las dataciones C14 realizadas en distintas muestras, se han podido correlacionar los datos con los de otros yacimientos del entorno.

46 H. ZUBELDIA, X. MURELAGA, S. BAILON. M. AGUIRRE Y X. SAEZ DE LAFUENTE
Las disoluciones que presentan algunos restos indican la presencia de rapaces. Seguramente, los restos fueron acumulados en la entrada de la cueva por un ave rapaz, y luego, los agentes biológicos transportaron los restos al interior de la cueva.
Con el estudio de los micromamíferos y las dataciones realizadas, se han definido cambios climáticos en la serie estudiada, desde el final de la glaciación Würm hasta el Holoceno. En ella se observan cuatro secuencias de calentamiento y cinco secuencias de enfriamiento, que coinciden con los cambios climáticos del Pleistoceno superior.
AGRADECIMIENTOS
En la obtención y preparación del material fósil, así como en la realización del presente estudio se ha contado con la inestimable ayuda de las siguientes personas e instituciones:
Pedro Castaños, Eduardo Pemán (Aranzadi), Salvador García (Aula paleontológica de Cenicero), Gloria Cuenca (Universidad de Zaragoza) y Carmen Sesé (Museo Nacional de Ciencias Naturales de Madrid)
Departamento de Cultura, Sección de Patrimonio Histórico de la Diputación Foral de Bizkaia, los proyectos CGL2004-02987 del Ministerio de Educación y Ciencia, S-PE06UN30 (Gobierno Vasco) y UNESCO 05/01 y EHU06/84 (Universidad del País Vasco/EHU).
BIBLIOGRAFIA
Aguirre Ruiz de Gopegui, M.
2000 El yacimiento paleolítico de Antoliñako Koba (Gautegiz-Arteaga, Bizkaia): secuencia estratigráfica y dinámica industrial. Avance de las campañas de excavación 1995-2000. lllunzar. 4, 39-81.
Altuna, J.
1972 Fauna de mamíferos de los yac1m1entos prehistóricos de Gipúzcoa. Con catálogo de los Mamíferos Cuaternarios del Cantábrico y del Pirineo Occidental. Munibe, 1-4: 464 pp.
Altuna, J.
1976 Los Mamíferos del yacimiento prehistórico de Tito Bustillo (Asturias). En: MOURE,
J.A., CANO, M., et al. (Ed.): Excavaciones en la cueva de «Tito Bustillo» (Asturias). Trabajos de I975, Oviedo, Instituto de Estudios Asturianos: 146-194.
Altuna, J.
1981 Restos óseos del yacimiento prehistórico del Rascaño. En: GONZÁLEZ ECHEGARAY, J., BARANDIARÁN MAESTU, I. et al., (Ed): El Paleolítico Superior de la cueva de Rascaño (Santander). Monografía del Museo Nacional y Centro de Investigación de Altamira, 3: 221-269.
Altuna, J.
1986 The mammalian faunas from the prehistoric site ofLa Riera: En: STRAUS, L. G., CLARK, G. A. et al., (Ed.): La Riera cave. Stone age huntergatherer adaptations in northem Spain, Anthropological Research Papers, 36, Arizona State University: 237-273.
Andrews, P.J. y Evans, E.M. N.
1983 Small mammal bone accumulations produced by mammalian carnivores. Paleobiology: 9(3), 289-307.
Andrews, P. J.
1990 Owls, Caves and Fossils. Natural History Museum Publications, London. 231 pp.
Arrizabalaga, A.; Montagut, E. y Gosálbez, J.
1986 Introducció a la Biología i zoogeografia deis petits mamifers (insectívors i rosegadors) del Montseny (Catalunya). Barcelona: CIRJT, 1986, 113 p.
Baena, J.; Carrión, E.; Ruiz, B.; Sesé, C.; Yravedra, J.; Jordá, J.; Barez, S.; Uzquiano, P.; Velázquez, R. y Manzano, l.
2004 Paleoecología y comportamiento humano durante el Pleistoceno Superior en la Comarca de Liébana: La secuencia del a Cueva del Esquilleu (Occidente de Cantabria, España). Pre-actas de la Reunión Científica: Neandertales Cantábricos: Estado del a Cuestión. Museo Nacional y Centro de Investigación de Altamira, Santillana del Mar, Octubre 2004: 54-55.
Baena, J.; Carrión, E.; Ruiz, B.; Sesé, C.; Yravedra, J.; Jordá, J.; Ellwood, B.; Uzquiano, P.; Velázquez, R.; Manzano, l.; Sánchez, S. y Hernández, E.
2005b, en prensa: Paleoecología y comportamiento humano durante el Pelistoceno Superior en la
KOBIE (Paleoantropología n.º 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA, BIZKAIA)
47
Comarca de Liébana: La secuencia del a Cueva del Esquilleu (Occidente de Cantabria, España). Monografías del Museo Nacional y Centro de Investigación de Altamira.
Bailon S.
1991 Amphibiens et reptiles du Pliocene et du Quaternaire de France et d'Espagne: mise en place et évolution des faunes. These Université Paris VII, 499p., 89 fig.
Bailon, S.
1999 Fiches D'Ostéologie Animale Pour L' Archéologie. Série C: Varia. Différenciation Ostéologique des Anoures (Amphibia, Anura) de France.
Bailon S. y Rage J.C.
1992 Amphibiens et reptiles du Quaternaire. Relations avece l'homme. Mém. Soc. Géol. De France, 160: 95-100.
Barandiaran, J. M.
1925 Anuario de Eusko-Folklore, tomo V.
Barandiaran, J. M.
1947 Prehistoria de Vizcaya, un cuarto de siglo de investigaciónes. IKUSKA 4-5, , pp. 134-147.
Barbadillo, L. J. y Sánchez-Herráiz, M. J.
1997 Anguis fragilis (LINAAEUS, 1758). Lución, Licranc;o. Pp. 187-189. En: Pleguezuelos, J. M. (Ed.). Distribución y biogeografía de los Anfibios y Reptiles en España y Portugal. Monografías de Herpetología, nº 3. Editorial Universidad de Granada & Asociación Herpetológica Española, Granada.
Blanco, J.C.
1998 Mamíferos de España (I): Insectívoros, Quirópteros, Primates y Carnívoros de la Península Ibérica, Baleares y Canarias. Geoplaneta: 457pp.
Blanco, J.C.
1998 Mamíferos de España (II): Cetáceos, Artiodáctilos, Roedores y Lagomorfos de la Península Ibérica, Baleares y Canarias. Geoplaneta: 383 pp.
Brenchley, P. J.
1998 Paleoecology: ecosystems, environments and evolution. Edt. Chapman & Hall, 402 pp.
KOBIE (Paleoantropología n.º 27), año 2003/07
Castaños, P.
1986 Los macromamíferos del Pleistoceno y Holoceno de Vizcaya. Faunas asociadas a yacimientos arqueológicos. Tesis Doctoral. UPV/EHU.
Chaline, J. 1970 Pliomys Lenki, forme relique dans la
Microfaune du Würm ancien de la Grotte de Lezetxiki (Guipúzcoa-Espagne). Munibe, XXII, 1/2: 43-49.
Chaline, J. 1972 Les Rongeurs du Pléistocene Moyen et
Supérieur de France. Systématique, Biostratigraphie, Paléoclimatologie, C.N.R.S.: 410 pp.
Chaline, J., Brunet-Lecomte, P. y Graf, J. D.
1988 Les proies des rapaces. Petits mammiferes et leur environnement. Doin Ed., Paris. 141 pp.
Corchon, S.; Hoyos, M.; Soto, E. y Meléndez, G.
1981 Cueva de las Caldas. San Juan de Priorio (Oviedo ). Excavaciones Arqueológicas en España, 115, Madrid, Mimisterio de Cultura.
Cuenca-Bescós, G. y García Pimienta, J. C.
En prensa: Holocene biostratigraphy and climatic change in Cantabria: the micromammalian faunas of el Miron Cave (Ramales de la Victoria, Cantabria, Spain). Journal of anthropological research.
Daams, R.
1981 The dental pattern of the dormice Dryomys, Myomimus, Microdyromys and Peridyromys. Tesis doctoral. Univ. Utrech, 72 pp.
Garrote Ruiz, A.; García Portero, J.; Arriola Garrido, A.; Eguiguren Altuna, E.; García Pascual, l. y Garrote Ruiz, R.
1992 Euskal Herriko Mapa Geologikoa, 38-IV Elantxobe. E: 1125.000. E.V.E.
Heinrich, W.-D.
1978 Biometrischen Erfassung eines Evolutionstrends bei Arvicola (Rodentia, Mammalia) aus dem Pleistozan Thüringens. Siiugetierk. Inform: 2, 3-21.
Hoffstetter R.
1962 Observations sur les ostéodermes et la classification des Anguidés actuels et fossiles (Reptiles, Sauriens). Bull. Mus. Nat. Hist. Nat. (2e série), Paris, 34(2), 149-157.

48 H. ZUBELDIA, X. MURELAGA, S. BAILON, M. AGUIRRE Y X. SAEZ DE LAFUENTE
Holman, J.A.
1998 Pleistocene Amphibians and Reptiles in Britain and Europe. Oxford Monographs on Geology and Geophysics No. 38. 254 pp.
Kolfschoten, T.
1995 On the application of fossil mammals to the reconstruction of the paleoenvironment of northwestern Europe. Acta Zool. Cracov. 38, 225-274.
Kowalski, K.
1992 Remarks on the zoogeography of Quaternary Rodents in Europe. Courier Forsch-Inst. Senckenberg, 153: 197-203.
Lopez Quintana, J. C., Castaños Ugarte, P.; Guenaga lizaso, A.; Murelaga, X.; Areso, P. y Uriz, A.
2005 La cueva de Atxagakoa (Forua, Bizkaia): ocupación humana y guarida de carnívoros durante el Musteriense en Urdaibai. Illunzar 01/05, 5: 11-24.
López-Fuster & Ventura.
1996 A morphometrical review of the Sorex araneus-articus species group from the Iberian Pe nin sula (Insectívora, Soricidae ). Bonner Zoologische Beitrage, 46, 327-337.
López-Martínez, N.
1992 Técnicas de estudio de microvertebrados. Los micromamíferos y su interés bioestratigráfico. En: H. Astibia ed. Paleontología de vertebrados: faunas y filogenía, aplicación y sociedad, Servicio editorial de la Universidad del País Vasco/EHU, Bilbao, 345-365.
Muñoz,M.
1982 Carta Arqueológica de Vizcaya. Primera parte: yacimientos en cueva. Cuadernos de Arqueología de Deusto, 8, 244 pp.
Mellet, J. S.
1974 Scatological origin of microvertebrate fossil acumulations. Science. 185, 349-350.
Murelaga, X.; López Quintana J. C.; Castaños Ugarte, P.; Guenaga Lizasu, A. y Zubeldia, H.
2005 Micromamíferos del yacimiento Holoceno de Kobeaga 11 (lspaster, Bizkaia). /llunzar 01/05, 5: 49-55.
Murelaga, X.; López Quintana J. C.; Castaños Ugarte, P.; Guenaga Lizasu, A. Bailon, S. y Zubeldia, H.
2007 Vertebrados del yacimiento Holoceno de Kobeaga 11 (lspaster, Bizkaia). Geogaceta: 41, 143-146.
Nadachowski, A.
1990 Comments on vanat10n, evolution and Phylogeny of Chionomys (Arvicolidae). Int. Symp. Evol. Phyl. Biostr. Arvicolids. Praga: 353-368.
Nores, C.
1988 Diferenciación biométrica de Apodemus sylvaticus y Apodemus flavicollis en la Coordillera Cantábrica. Primeros resultados. Rev. Biol. Univ. Oviedo: 6, 109-116.
Palomo, J. y Gisbert, J. (Ed.)
2002 Atlas de los mamiferos terrestres de España. Organismo Autónomo de Parques Nacionales. DGCNA-MIMAM: 564pp.
Pemán,E.
1985 Aspectos climáticos y ecológicos de los micromamíferos del yacimiento de Erralla. Munibe: 37, 49-57.
Pemán,E.
1990a Los micromamíferos en el Pleistoceno superior del País Vasco. Munibe: 42, 259-262.
Pemán,E.
1990b Los micromamíferos de la cueva de Amalda y su significación. Comentarios sobre Pliomys lenki. En: Altuna, J., Baldeón, A., Mariezkurena, K., et al., La cueva de Amalda (Zestoa, País Vasco). Ocupaciones Paleolíticas y Postpaleolíticas. San Sebastián, Fundación Eusko lkastunza: 225-238.
Pemán,E.
1994 Los micromamíferos de Laminakll (Berriatua, Bizkaia). Kobie serie Paleoantropología, XXI: 225-233.
Pemán,E.
2000 Los micromamíferos de Labeko Koba (Arrasate, País Vasco). Arrizabalaga, A. y Altuna, J.-ed (2000). Labeko Koba (País Vasco). Hienas y Humanos en los albores del Paleolítico superior. Munibe, 52:183-185.
KOBIE (Paleoantropología n.° 27), año 2003/07

MICROVERTEBRADOS DE LA SECUENCIA SUPERIOR DE ANTOLIÑAKO KOBA (PLEISTOCENO SUPERIOR) (GAUTEGUIZ-ARTEAGA. BIZKAIA)
49
Pokines, J. T.
1998 The paleoecology of Lower Magdalenian Cantabrian Spain. Bar International series: 713, 1-189.
Pratt, A. E.
1989 Taphonomy of the microvertebrate fauna from the early Miaocene Thomas farm locality, Florida (U.S.A.). Palaeogeograph. Palaeoclim, Palaeoecol. 76, 125-151.
Reumer, J. W. F.
1984 Rusnician and early Pleistocene Soricidae (lnsectivora, Mammalia) from Tegelen (The Netherlands) and Hungary. Scripta Geologica: 73, 173.
Rzebik-Kowalska, B.
1995 Climate and history of European shrews (family Soricidae). Acta. Zoo! Cracov, 38 (1): 95-107.
Saénz de Buruaga, A; Aguirre, M.; Grima, C.; López-Quintana, J. C.; Ormazabal, A. y Pastor, B.
1998 Método y práctica de la Estratigrafía Analítica. Krei, 3. 7-41.
Sanchiz F.B.
1977 La familia Bufonidae (Amphibia, Anura) en el Terciario europeo. Trabajos Neógeno!Cuaternario, 8: 75-111.
Sanchiz F.B.
1998 Handbuch der Palaoherpetologie. Verlag Dr Friedrich Pfeil, München, 275 p.
Sesé, C.
2005 Aportación de los micromamíferos al conocimiento paleoambiental del Pleistoceno Superior de la Región Cantábrica: nuevos datos y síntesis. Monografía del Museo
KOBIE (Paleoantropología n." 27), año 2003/07
Nacional y Centro de investigación de Altamira, 20, pp. 167-200.
Szyndlar Z.
1984 Fossil Snakes from Poland. Acta Zoologica Cracoviensia, XXVIII (1), 1-156.
Van der Meulen, A.
1973 Middle Pleistocene smaller mammals from Monte Peglia (Orvieto, Italy) with special reference to the phylogeny of Microtus (Arvicolidae, Rodentia)» Quaternaria: 1-144.
Ventura, J.; López-Fuster, M.A. eta Gosálbez, J.
1993 A morphometric analysis of cranial variation in the bank vole, Clethrinimys glareolus (Schreber, 1780) (Rodentia, Arvicolidae) from the Iberian Peninsula. Zoo l. Anz., 231, 183-193.
Zabala, J. 1984 Los Micromamíferos del yacimiento de Ekain.
Altuna, J.; Merino, J. M.; et al en: El yacimiento prehistórico de la cueva de Ekain, San Sebastian, Sociedad de Estudios Vascos, Sociedad de Ciencias Aranzadi: 317-330.
Zubeldia, H.
2006 Goi Pleistoceno-Holozeno garaiko Antoliñako Koba aztarnategiko mikroordunen ikerketa. Tesis de Licenciatura.
Se han utilizado las siguientes direcciones de internet como referencia en el capítulo de Paleoecología.
- www.fauna-iberica.mncn.csic.es
- www.hispagua.cedex.es/datos/agua_espana/anfibios. htm
- www.mma.es/portal/secciones.es
- www.vertebradosibericos.org

Recommended

















