
Lu
Aa
b
a
ARRA
KCHLS
1
iiPat
0d
Veterinary Parasitology 174 (2010) 305–312
Contents lists available at ScienceDirect
Veterinary Parasitology
journa l homepage: www.e lsev ier .com/ locate /vetpar
ife cycle of the camel tick Hyalomma dromedarii (Acari: Ixodidae)nder field conditions in Northern Sudan
. ELGhali a,∗, S.M. Hassanb
Central Veterinary Research Laboratories, P.O. Box 8067, Amarat, Khartoum, SudanDepartment of Parasitology, Faculty of Veterinary Medicine, University of Khartoum, Sudan
r t i c l e i n f o
rticle history:eceived 12 April 2010eceived in revised form 10 August 2010ccepted 23 August 2010
eywords:amel tickyalomma dromedariiife cycleudan
a b s t r a c t
Engorged Hyalomma dromedarii females were placed for development in shade and sun(open non-shaded site) in April, August and December for two successive years. Engorgednymphs were also placed at the same sites in January, March, May, June, July and August.Preoviposition periods ranged between 9.8 and 11.7 days in the shade but longer in the sunin December (14.7 days). Egg production index was higher in August (0.72) than in Apriland December (0.46 and 0.39, respectively) in shade and in August compared to Decemberin the sun (0.65 and 0.29, respectively). In shade and in sun, the prehatching periods werelonger in December (50.7 and 48.2 days) than in April and August (25.6 and 29.0 days).Percentage hatchability ranged between 82 to 94.1% and 56.2 and 58.9% in the shade andsun, respectively. Number of eggs laid and eggs hatched were positively correlated withfemales’ engorgement weights. There were high mortality rates of females in the sun andmost of the eggs desiccated. Slight increased ambient and soil temperatures increased eggproduction, shortened preoviposition periods and decreased hatchability while increasedhumidity shortened preoviposition and prehatching periods and increased hatchability.
Nymphal–adult moulting periods were long in January (20.7 days in the shade and 14.4days in the sun) and short in August in the shade (7.8 days) while they were only 6 days in
May and June in the sun. High numbers moulted in all months in the shade (ranges 23.3–29.5nymphs). The mean mortality of engorged nymphs in the shade was very low. However,in the sun mortality was high (12.9–30 nymphs). Premoulting periods were negativelycorrelated with ambient temperature but positively correlated with humidity. In the shade,mean number of nymphs moulting increased with increased mean ambient temperature,all.
humidity and rainf. Introduction
The entire life cycle or developmental period of tickss mainly affected by temperature and/or relative humid-
ty (Branagan, 1973a,b; Tukahirwa, 1976; King et al., 1988;egram and Banda, 1990). Egg production index, hatchingnd moulting percentages are also temperature and rela-ive humidity dependant (Norval, 1977; Zahler and Gothe,∗ Corresponding author. Tel.: +00 249 09 12805583.E-mail address: [email protected] (A. ELGhali).
304-4017/$ – see front matter. Published by Elsevier B.V.oi:10.1016/j.vetpar.2010.08.028
Published by Elsevier B.V.
1995; ELGhali et al., 2003). Life cycle of the camel tickHyalomma dromedarii has only been studied under labo-ratory conditions (Hagras and Khalil, 1988; Alahmed andKheir, 2003). This tick species follows different types oflife cycles to avoid unfavourable conditions. Delpy andGouchey (1937 cited in Hoogstraal, 1956) stated that H.dromedarii is a three-host tick, changes to two-host under
stress of heat to avoid larval desiccation. Alfeev (1951, citedin Hoogstraal, 1956), reported that H. dromedarii behavedas a single host on rabbits with adults moving to a new posi-tion shortly after moulting and remaining unattached fora day or two. Hoogstraal (1956) reported that field obser-
nary Par
306 A. ELGhali, S.M. Hassan / Veterivations suggested that change of host usually occurs afternymphal–adult moulting and rarely after larval–nymphalmoulting. Das and Subramanian (1972) reported that H.dromedarii on sheep and cattle is a three-host tick and 60%changed to two-host when fed on rabbits. Ouheli (1994)pointed out that this tick is a three-host changing to two-host when their density on the host becomes high. Alahmedand Kheir (2003) reported that this tick behaved as a two-host tick when fed on rabbits. Recently, ELGhali and Hassan(2010) found that this tick behaved as a two-host tickwhen fed on camels in Northern Sudan. This tick speciesis distributed throughout the world wherever camels arepresent, in southern Russia, in the Far, Middle and NearEast, in North Africa and south of the great northern desertsdown to Somalilands and northeastern Kenya (Hoogstraal,1956). It represents nearly 90% of ticks infesting camels(Karrar et al., 1963; Van straren and Jonjegan, 1993; Diabet al., 2001; ELGhali and Hassan, 2009). The economicimportance of this tick species is considered as the camelsare important exporting animals in the Sudan and theirhealth and production are affected by high tick infestation.The present work was conducted to study development H.dromedarii under natural conditions aiming to identify thebest periods of the tick development in Northern Sudan.The effects of temperature, relative humidity, shade andsun on the developmental periods are studied.
2. Materials and methods
The study was conducted under field conditions inAtbara town (Long. 33◦58′E and Lat. 17◦40′N) in RiverNile State, Northern Sudan (at the junction of River Nileand River Atbara) where camels are reared and marketed(Fig. 1). The mean maximum ambient temperature is about45 ◦C in summer (March to June) and the mean minimumambient temperature in winter (October to February) fallsto less than 10 ◦C. The mean ambient relative humidityis generally low while the mean soil temperature rangesbetween 30 and 55 ◦C in summer and ranges between 15and 30 ◦C in winter.
Two areas were divided into two sites shaded and open.Minimum and maximum ambient temperatures, relativehumidity and soil temperature at ground level were dailyrecorded at 09:00 and 15:00 h and total daily rainfall wasalso recorded. In this aspect all these meteorological factorswere measured closely to the tested ticks.
2.1. Eggs laying and hatching
Fully engorged H. dromedarii females were collectedfrom camels and immediately weighed and individuallyplaced in 7.5 cm × 2.5 cm glass vials, closed using cottonwool and fixed with adhesive tape. The vials were imme-diately placed in the field at a depth of 3–5 cm and coveredwith an approximately uniform thick layer of litter. A totalof 15–20 replicates were placed in the shade and a simi-
lar number in the sun at each area. This study was carriedout in December 2001, April 2002, August 2002, December2002, April 2003 and August 2003. Preoviposition, preeclo-sion periods were daily recoded and number of eggs laidand hatched for each female was estimated. For estimationasitology 174 (2010) 305–312
of total eggs produced by each female small batches of eggswere weighed and counted and accordingly the total eggswere estimated. By the end of hatching, larvae or the non-hatched eggs were counted and calculated as percentagefrom the total eggs.
2.2. Moulting periods of nymphs to adults
Flat larvae of H. dromedarii were fed on camels’ earsaccording to Bailey (1960) method. From the droppedengorged nymphs, 30 nymphs were immediately put invials, closed by cotton wool and wrapped in a piece of gauzeand placed in the field at depth of 3–5 cm then covered witha thick layer of litter in shade and in sun (16 vials/each site/area). The vials were daily monitored for moulting or mor-tality. This study was conducted in January 2002, March2003, May, June, July and August 2004.
2.3. Statistical analysis
In preoviposion and hatching study the data of the sameseasons and sites (shade or sun) in the 2 years were com-bined before evaluation.
Data were then subjected to appropriate general linearmodel (GLM) procedure of the statistical analyses usingthe SAS package. The SAS was used to perform analysisof variance (ANOVA) and mean separations were per-formed using Ryan–Einot–Gabriel–Welsch multiple rangetest (REGWQ) (Day and Quinn, 1989). The data, when-ever required, were transformed to (
√log10(x + 1) + 0.5)
scale before being subjected to ANOVA and percentages(e.g. hatchability) were transformed to arcsine scale. Cor-relation analysis was carried out to relate H. dromedariidevelopment with meteorological data and to correlate thedevelopmental periods with each other.
3. Results
Mean daily meteorological factors recorded during theoviposion and hatching experiments in two successiveyears are shown in Table 1. The lowest temperatureand highest humidity were recorded during the periodfrom December to March (winter). The rainfall was verylow and of short duration during August and Septem-ber. Table 2 presented the abiotic factors recorded duringnymphal–adult moulting and similarly January which rep-resents winter showed the lowest temperature degreesand highest humidity levels followed by March (transientbetween winter and summer). The rest of the months weresimilar except that August showed higher relative humid-ity in the morning due to light rainfall.
Number of eggs laid and egg production index (EPI)(egg batch weight/female engorgement weight) in theshade, were significantly higher in August than in Apriland December (Table 3). However, in the sun, althoughthere was no significant difference in number of eggs laid
between August and December but EPI was significantly(P < 0.001) higher in August than in December. In April, allthe females died in the sun. In the shade, preovipositionperiods ranged between 9.8 ± 0.8 and 11.7 ± 0.6 days withno significant differences (Table 3) but, in the sun, these
A. ELGhali, S.M. Hassan / Veterinary Parasitology 174 (2010) 305–312 307
location
pcicwel(
Fig. 1. Sudan map showing Atbara (study area)
eriods were significantly (P < 0.01) longer in Decemberompared to August. Prehatching periods, in the shade andn the sun, were significantly (P < 0.01) longer in Decemberompared to other months. Per cent hatchability of eggs
as higher in the shade than in the sun. In the shade, femalengorgement weights were significantly positively corre-ated with the number of eggs laid and number hatchedTable 4).
and a part of camel grazing areas in the Sudan.
The number of eggs laid was significantly negativelycorrelated with preoviposition periods and significantlynegatively correlated with prehatching periods. The num-ber of eggs, in the shade, was also positively correlated
with number of eggs hatched. Prehatching periods in theshade were significantly negatively correlated with num-ber of eggs hatched and significantly positively correlatedwith preoviposition periods. Similarly, egg batches laid in
308A
.ELGhali,S.M
.Hassan
/Veterinary
Parasitology174 (2010) 305–312
Table 1Averages of daily recorded meteorological factors during H. dromedarii oviposition and hatching experiments.
Periods Min. temperature (◦C) Max. temperature (◦C) Soil temp. (◦C) (9:00 h) Soil temp. (◦C) (15:00 h) Rh. (%) (9:00 h) Rh. (%) (15:00 h) Rain (mm)
Year 1 Year 2 Year 1 Year 2 Year 1 Year 2 Year 1 Year 2 Year 1 Year 2 Year 1 Year 2 Year 1 Year 2
December–March 18.1 16.1 38 37.4 27.7 26.1 43.8 43.8 56.3 35.7 49.3 30.8 0 0April–June 26 30.3 46.7 45.2 33.6 32.2 50.8 49.5 38.3 25.5 31.4 22 0 0August–September 28.5 29 45.7 42.3 35.4 32.7 49.3 42.7 49.1 53.6 31.2 38.4 0.04 1.4
Rh, relative humidity.
Table 2Averages of daily recorded meteorological factors during nymphal–adult moulting experiments.
Month Min. temperature (◦C) Max. temperature (◦C) Soil temp. (◦C) (9:00 h) Soil temp. (◦C) (15:00 h) Rh. (%) (9:00 h) Rh. (%) (15:00 h) Rain (mm)
January 2002 12.3 30.5 20.2 36.7 58.7 40.9 0March 2003 16.1 37.3 26.1 43.8 35.7 30.8 0May 2004 25 48.2 35.9 51.6 24.2 14.2 0June 2004 27.4 46.3 35.2 51.3 32.9 18.3 0July 2004 27.2 45.6 35.6 51.3 36.6 21.9 0August 2004 28.5 45.5 35.8 51.1 43.6 23.1 0.9
Rh, relative humidity.

A. ELGhali, S.M. Hassan / Veterinary Par
Tab
le3
Mea
ns
(±SE
)of
dev
elop
men
talp
erio
ds
ofH
.dro
med
arii
engo
rged
fem
ales
un
der
fiel
dco
nd
itio
ns
inA
tbar
a,N
orth
ern
Sud
an.
Site
Mon
thN
um
ber
ofeg
gsEn
g.fe
m.
mor
tali
ty(%
)
Preo
vip
osit
ion
per
iod
(day
s)Eg
gp
rod
uct
ion
ind
exPr
ehat
chin
gp
erio
d(d
ays)
Nu
mbe
rof
eggs
hat
ched
Hat
chab
ilit
y(%
)
Shad
eA
pri
l49
72.4
±30
0.0b
(79)
1.3
11.7
±0.
6a(7
9)0.
46±
0.03
b(7
9)29
.9±
0.4b
(51)
4226
.2±
365.
1a(5
1)85
.1±
2.7a
(51)
Au
gust
6932
.6±
383.
3a(7
6)5.
09.
8±
0.8a
(76)
0.72
±0.
03a
(76)
28.8
±0.
8b(6
8)65
48.2
±48
6.2a
(68)
94.1
±4.
1a(6
8)D
ecem
ber
3872
.7±
284.
8c(6
5)18
.611
.2±
0.6a
(76)
0.39
±0.
03b
(65)
50.7
±0.
8a(5
9)32
33.2
±33
2.9b
(59)
82.0
±4.
5a(5
9)A
pri
l0
100
00
00
0Su
nA
ugu
st52
48.0
±63
4.3a
(32)
609.
0±
0.7b
(32)
0.65
±0.
06a
(32)
25.6
±0.
6b(5
)44
57.0
±23
41.8
a(5
)58
.9±
22.9
a(5
)D
ecem
ber
2569
.3±
264.
2a(6
5)18
.614
.7±
1.0a
(65)
0.29
±00
3b(6
5)48
.2±
1.7a
(14)
1587
.0±
387.
8a(1
4)56
.2±
13.1
a(1
4)
Mea
ns
(±SE
)fo
llow
edby
the
sam
ele
tter
inea
chco
lum
nfo
rea
chsi
tear
en
otsi
gnifi
can
tly
dif
fere
nt
at5%
leve
lbas
edon
Rya
n’s
Qte
st(R
EWG
Q),
figu
res
inp
aren
thes
is=
nu
mbe
rof
tick
s.
asitology 174 (2010) 305–312 309
the shade significantly increased with the increase in meanminimum temperature and rainfall (Table 5) and there wasa significant positive correlation with increase in mean soiltemperature at 09:00 h. Although preoviposition periodsin the shade were negatively correlated with meteorologicdata with the exception of maximum temperature and soiltemperature at 15:00 h but in the sun other meteorologicaldata were negatively correlated. Mean number of eggs laidin the sun was significantly positively correlated with allmeteorological data with the exception of relative humid-ity at 15:00 h.
Premoulting periods of nymphs in the shade were sig-nificantly longer in January followed by March and shorterfrom May to August (Table 6). Premoulting in the sun waslonger in January but was not significantly different inMarch, July and August. The shortest period was recordedin May and June. Number of nymphs moulted in the shadewas not significantly different in all months with the excep-tion of March. Mean mortality of nymphs in the shade wasvery low but was very high in the sun.
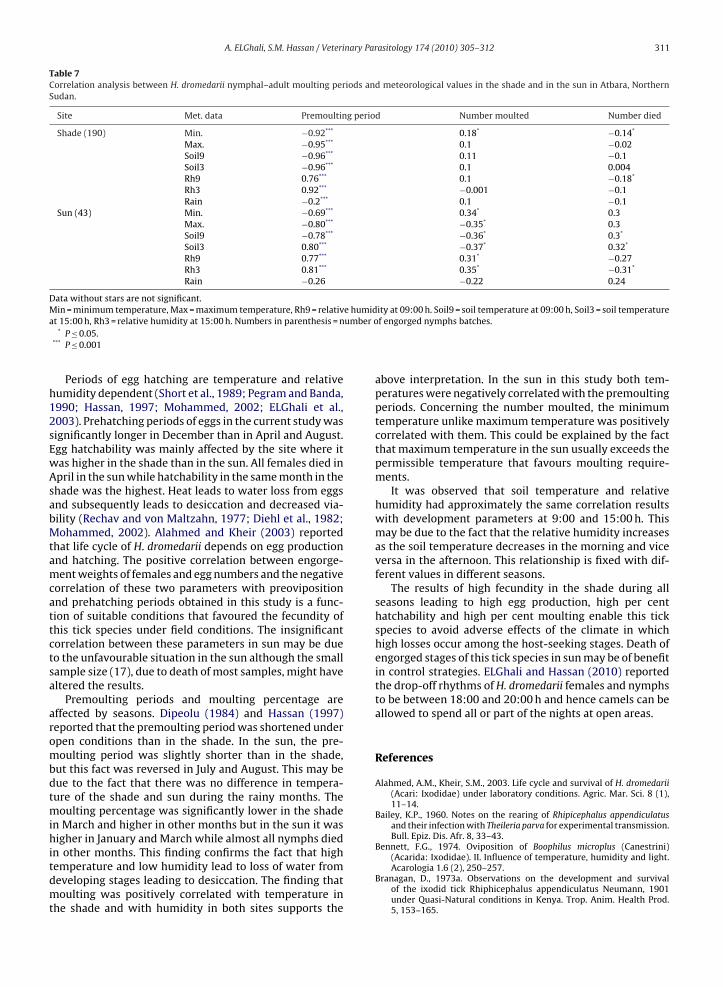
The premoulting periods in the shade and in the sunwere significantly negatively correlated with mean max-imum, minimum and soil temperatures (Table 7) butsignificantly positively correlated with relative humidity at09:00 and 1500 h. With increased temperature and relativehumidity in the shade, there was an increase in moultingof nymphs and decrease in mortality rate. Increase in meanambient and soil temperatures in the sun led to a decreasedmoulting rate and an increased mortality rate.
4. Discussion
Effect of shade and sun on oviposition was signifi-cantly different. While all engorged females under shadeoviposited and most eggs hatched, most of the femalesdied in few days before laying of eggs in the sun duringthe hot months. It was noticed that although all ticks usedin this study were from the same population, few indi-viduals could escape the adverse conditions in the sun.This might be controlled by some intrinsic factors allowedthese individuals to resist the unfavourable climatic con-ditions. Eggs laid in the sun in December and August 2003were found desiccated due to low relative humidity butin the shade hatchability rate was high even during hotdry months. This implies that tick population in nature issustained despite adverse climatic conditions. This find-ing, also, helps to explain the observations of ELGhaliand Hassan (2009), at the same study area, in that ticks(89.9% H. dromedarii) were found on camels throughoutthe year with the highest infestation during March to Octo-ber. Patrick and Hair (1979) and Punyua (1984) found thatpreoviposition periods of ticks within the permissible tem-perature were found to shorten as ambient temperatureelevates. Hagras and Khalil (1988) reported that preovipo-sition of H. dromedarii was prolonged as the temperaturedecreased from 34 to 21 ◦C. In the current study, preovipo-
sition period was longer in December and April than inAugust, but the difference was not significant in the shadewhich seems to minimize the effect of heat. This differ-ence may be attributed to the fact that in August 2003,rainfall of about 22 mm was recorded leading to a rise in
310 A. ELGhali, S.M. Hassan / Veterinary Parasitology 174 (2010) 305–312
Table 4Correlation analysis among H. dromedarii engorged female developmental periods in the shade and in the sun in Atbara, Northern Sudan.
Site Parameters Number of eggs Preoviposition periods Prehatch. Period Hatchability (%)
Shade (174) Engorged weight 0.43*** −0.07NS −0.04NS 0.36***
Number of eggs −0.18* −0.45*** 0.88***
Preoviposition Period 0.63*** −0.26***
Prehatching period −0.45***
Sun (17) Engorgement weight 0.32NS −0.43NS 0.1NS 0.13NS
Number of eggs 0.2NS −0.4NS 0.02NS
Preoviposition Period 0.15NS −0.22NS
Prehatching period 0.11NS
NS = not significant, figures in parenthesis = number of ticks.* P < 0.05.
*** P < 0.001.
Table 5Correlation analysis between H. dromedarii engorged female developmental periods and meteorological values in the shade and in the sun in Atbara,Northern Sudan.
Site Met. data Number of eggs Preoviposition period Prehatch. periods Hatching (%)
Shade (221) Min. 0.32*** −0.12 0.45*** −0.27***
Max. 0.06 0.01 0.39*** −0.33***
Soil9 0.16* −0.03 0.42*** −0.31***
Soil3 −0.11 0.12 0.38*** −0.31***
Rh9 0.06 −0.11 −0.35*** 0.24**
Rh3 −0.1 −0.16* −0.48*** 0.34***
Rain 0.7*** −042*** 0.34*** 0.17*
Sun (100) Min. 0.52*** −0.38*** 0.55*** −0.34*
Max. 0.39*** −0.38*** 0.56*** −0.44**
Soil9 0.45*** −0.39*** 0.58*** −0.40**
Soil3 0.21* −0.35*** 0.54*** −0.37**
Rh9 0.32** −0.2* −0.51*** 0.48***
Rh3 −0.12 −0.01 −0.63*** 0.53***
Rain 0.64*** −0.33*** 0.62*** −0.27
Data without stars are not significant. Min. = minimum temperature, Max. = maximum temperature, Rh9 = relative humidity at 09:00 h. Soil9 = Soil temper-ty at 15
ature at 09:00 h, Soil3 = soil temperature at 15:00 h, Rh3 = relative humidi* P < 0.05.** P < 0.01.
*** P < 0.001.
relative humidity for some days. This observation was inagreement with Sweatman (1967) who pointed out thathigh saturation deficit shortened preoviposition period attemperatures between 20 and 30 ◦C.
Egg production index (EPI) was highly affected by ambi-ent temperature and relative humidity. Nearly all femalesdied within a few days in the sun in April and August giv-ing rise to zero and 0.65% EPI, respectively. Bennett (1974)reported that temperature of 40.5 ◦C was lethal to engorged
Table 6Means (±SE) of H. dromedarii nymphal–adult moulting periods in the shade and i
Site Month Mean premoulting period (days) Mean numb
Shade January 20.7 ± 0.4a(38) 29.9 ± 0.7aMarch 14.3 ± 0.2b(26) 23.3 ± 1.8bMay 8.4 ± 0.2c(31) 28.1 ± 0.6aJune 8.1 ± 0.2c(32) 29.5 ± 0.2aJuly 8.1 ± 0.1c(38) 29.1 ± 0.2aAugust 7.8 ± 0.1c(26) 29.4 ± 0.3a
Sun January 14.4 ± 0.2a(29) 16.9 ± 2.0aMarch 12.0 ± 1.3ab(9) 5.6 ± 1.9bMay 6.0 ± 0.0b(2) 0.6 ± 0.6cJune 6.0 ± 0.0b(1) 0.03 ± 0.03July 10.0 ± 0.0ab(1) 0.4 ± 0.4cAugust 10.5 ± 0.5ab(1) 0.7 ± 0.7c
Means (±SE) followed by the same letter in each column are not significantly diffsis = number of nymph batches.
:00 h. Numbers in parenthesis = number of ticks.
females. Similarly, ELGhali et al. (2003) found that EPI istemperature and relative humidity dependent. It shouldbe noticed that EPI of H. dromedarii was the highest dur-ing August due to probably high humidity recorded during
this month. Eggs and the engorged stages cannot take waterfrom the sub-saturated atmosphere and loss of water is afunction of temperature and saturation deficit (Diehl et al.,1982). They reported that when eggs lose up to 35% of waterembryonic development terminates.n the sun in Atbara, Northern Sudan.
er moulted Mean number died Eng. nymphs mortality (%)
(38) 0.1 ± 0.1b(38) 0.3(26) 6.7 ± 1.8a(26) 22.3(31) 1.9 ± 0.6b(31) 6.3(32) 0.5 ± 0.2b(32) 1.7(38) 0.9 ± 0.2b(34) 3.0(26) 0.6 ± 0.3b(26) 2.0(37) 12.9 ± 2.0c(37) 43(25) 23.8 ± 2.0b(25) 79.7(34) 28.9 ± 0.7a(34) 96.3c(32) 30.0 ± 0.03a(32) 99.9(40) 29.6 ± 0.4a(40) 98.7(26) 29.3 ± 0.7a(26) 97.7
erent at 5% level based on Ryan’s Q test (REWGQ). Numbers in parenthe-
A. ELGhali, S.M. Hassan / Veterinary Parasitology 174 (2010) 305–312 311
Table 7Correlation analysis between H. dromedarii nymphal–adult moulting periods and meteorological values in the shade and in the sun in Atbara, NorthernSudan.
Site Met. data Premoulting period Number moulted Number died
Shade (190) Min. −0.92*** 0.18* −0.14*
Max. −0.95*** 0.1 −0.02Soil9 −0.96*** 0.11 −0.1Soil3 −0.96*** 0.1 0.004Rh9 0.76*** 0.1 −0.18*
Rh3 0.92*** −0.001 −0.1Rain −0.2*** 0.1 −0.1
Sun (43) Min. −0.69*** 0.34* 0.3Max. −0.80*** −0.35* 0.3Soil9 −0.78*** −0.36* 0.3*
Soil3 0.80*** −0.37* 0.32*
Rh9 0.77*** 0.31* −0.27Rh3 0.81*** 0.35* −0.31*
Rain −0.26 −0.22 0.24
Data without stars are not significant.M e humida umber o
h12sEwAsabMtamcattctsa
arombdtmihitdmt
in = minimum temperature, Max = maximum temperature, Rh9 = relativt 15:00 h, Rh3 = relative humidity at 15:00 h. Numbers in parenthesis = n
* P ≤ 0.05.*** P ≤ 0.001
Periods of egg hatching are temperature and relativeumidity dependent (Short et al., 1989; Pegram and Banda,990; Hassan, 1997; Mohammed, 2002; ELGhali et al.,003). Prehatching periods of eggs in the current study wasignificantly longer in December than in April and August.gg hatchability was mainly affected by the site where itas higher in the shade than in the sun. All females died inpril in the sun while hatchability in the same month in thehade was the highest. Heat leads to water loss from eggsnd subsequently leads to desiccation and decreased via-ility (Rechav and von Maltzahn, 1977; Diehl et al., 1982;ohammed, 2002). Alahmed and Kheir (2003) reported
hat life cycle of H. dromedarii depends on egg productionnd hatching. The positive correlation between engorge-ent weights of females and egg numbers and the negative
orrelation of these two parameters with preovipositionnd prehatching periods obtained in this study is a func-ion of suitable conditions that favoured the fecundity ofhis tick species under field conditions. The insignificantorrelation between these parameters in sun may be dueo the unfavourable situation in the sun although the smallample size (17), due to death of most samples, might haveltered the results.
Premoulting periods and moulting percentage areffected by seasons. Dipeolu (1984) and Hassan (1997)eported that the premoulting period was shortened underpen conditions than in the shade. In the sun, the pre-oulting period was slightly shorter than in the shade,
ut this fact was reversed in July and August. This may beue to the fact that there was no difference in tempera-ure of the shade and sun during the rainy months. The
oulting percentage was significantly lower in the shaden March and higher in other months but in the sun it wasigher in January and March while almost all nymphs died
n other months. This finding confirms the fact that highemperature and low humidity lead to loss of water fromeveloping stages leading to desiccation. The finding thatoulting was positively correlated with temperature in
he shade and with humidity in both sites supports the
ity at 09:00 h. Soil9 = soil temperature at 09:00 h, Soil3 = soil temperaturef engorged nymphs batches.
above interpretation. In the sun in this study both tem-peratures were negatively correlated with the premoultingperiods. Concerning the number moulted, the minimumtemperature unlike maximum temperature was positivelycorrelated with them. This could be explained by the factthat maximum temperature in the sun usually exceeds thepermissible temperature that favours moulting require-ments.
It was observed that soil temperature and relativehumidity had approximately the same correlation resultswith development parameters at 9:00 and 15:00 h. Thismay be due to the fact that the relative humidity increasesas the soil temperature decreases in the morning and viceversa in the afternoon. This relationship is fixed with dif-ferent values in different seasons.
The results of high fecundity in the shade during allseasons leading to high egg production, high per centhatchability and high per cent moulting enable this tickspecies to avoid adverse effects of the climate in whichhigh losses occur among the host-seeking stages. Death ofengorged stages of this tick species in sun may be of benefitin control strategies. ELGhali and Hassan (2010) reportedthe drop-off rhythms of H. dromedarii females and nymphsto be between 18:00 and 20:00 h and hence camels can beallowed to spend all or part of the nights at open areas.
References
Alahmed, A.M., Kheir, S.M., 2003. Life cycle and survival of H. dromedarii(Acari: Ixodidae) under laboratory conditions. Agric. Mar. Sci. 8 (1),11–14.
Bailey, K.P., 1960. Notes on the rearing of Rhipicephalus appendiculatusand their infection with Theileria parva for experimental transmission.Bull. Epiz. Dis. Afr. 8, 33–43.
Bennett, F.G., 1974. Oviposition of Boophilus microplus (Canestrini)
(Acarida: Ixodidae). II. Influence of temperature, humidity and light.Acarologia 1.6 (2), 250–257.Branagan, D., 1973a. Observations on the development and survivalof the ixodid tick Rhiphicephalus appendiculatus Neumann, 1901under Quasi-Natural conditions in Kenya. Trop. Anim. Health Prod.5, 153–165.

nary Par
312 A. ELGhali, S.M. Hassan / VeteriBranagan, D., 1973b. The developmental periods of the ixodid tick Rhipi-cephalus appendiculatus Neum. under laboratory conditions. Bull.Entomol. Res. 63, 155–168.
Das, H.I., Subramanian, G., 1972. Biology of Hyalomma dromedarii Koch,1844 (Acari: Ixodidae). Indian J. Anim. Sci. 42, 285–289.
Day, R.W., Quinn, G.P., 1989. Comparison of treatments after analysis ofvariance in ecology. Ecol. Monogr. 59 (4), 433–463.
Diab, F.M., ELkady, G.A., Shouky, A., 2001. Bionomics of ticks collected fromSinai. 2. Abundance, attachment sites and density of ticks infestingArabian camels. J. Egypt. Soc. Parasitol. 31, 479–489.
Diehl, P.A, Aeschlimann, A., Obenchain, F.D., 1982. Tick reproduction,oogenesis and oviposition. In: Obenchain, F.D., Galun, R. (Eds.), Phys-iology of Ticks. Pergamon Press, New York, pp. 277–350.
Dipeolu, O.O., 1984. Development of ixodid ticks under natural conditionsin Nigeria. Trop. Anim. Health Prod. 16, 13–20.
ELGhali, A., Hassan, S.M., 2009. Ticks (Acari: Ixodidae) infesting camels(Camelus dromedarius) in Northern Sudan. Onderstepoort J. Vet. Res.76, 177–185.
ELGhali, A., Hassan, S.M., 2010. Drop-off rhythms and survival periodsof Hyalomma dromedarii (Acari: Ixodidae) fed on camels (Camelusdromedarius) in the Sudan. Vet. Parasitol. 170, 302–306.
ELGhali, A., Osman, O.M., ELRabaa, F.M.A., 2003. Some studies on biolog-ical behaviour of H. a. anatolicum (Acari: Ixodidae) under laboratoryconditions. I. Development. Sud. J. Vet. Sci. Anim. Husb. 42 (1 and 2),72–88.
Hagras, A.E., Khalil, G.A.M., 1988. Effect of temperature on Hyalommadromedarii Koch (Acari: Ixodidae). J. Med. Entomol. 25 (5), 354–359.
Hassan, S. M., 1997. Ecological studies on R. appendiculatus and A. var-iegatum (Acari: Ixodidae): drop-off rhythms, development, survivaland seasonal population dynamics. Ph.D thesis, Kenyatta University,Kenya. 206 pp.
Hoogstraal, H. (1956). African Ixodoidea. 1. Ticks of the Sudan with specialreference to Equatoria Province and with preliminary reviews of the
genera Boophilus, Margaropus and Hyalomma. U.S. Navy WashingtonD.C. 1101 pp.Karrar, G., Kaiser, M.N., Hoogstraal, H., 1963. Ecol ogy and host-relationship of ticks (Ixodoidea) infesting domestic animals in KassalaProvince, Sudan with special reference to Amblyomma lepidum Donitz.Bull. Entomol. Res. 54, 509–522.
asitology 174 (2010) 305–312
King, D., Getting, Newson, R.M., 1988. A climate-based model for the devel-opment of the ixodid tick, Rhipicephalus appendiculatus in East Coastfever zones. Vet. Parasite 29, 441–451.
Mohammed, A.S., 2002. Bioecological studies on the tick A. lepidum(Donitz, 1909) under natural field conditions in Blue Nile State, Sudan.Ph.D thesis, U. of K, Sudan.
Norval, R.A.I., 1977. Studies on the ecology of the tick Amblyommahebraeum Koch in Eastern Cape Province of South Africa. II. Survivaland development. J. Parasitol. 63, 740–747.
Ouheli, H., 1994. Comparative development of Hyalomma marginatum(Koch, 1844), H. detritum (Schulze, 19919), H. lusitanicum (Koch, 1844)and H. dromedarii (Koch, 1844) under laboratory conditions. Acta Par-asitol. 39 (3), 153–157.
Patrick, C.D., Hair, J.A., 1979. Ovipostion behaviour and larval longevityof the lone star tick, (A. americanum) (Acarina: Ixodidae) in differenthabitats. Entomol. Soc. Am. 72 (2), 308–312.
Pegram, R.G., Banda, D.S., 1990. Ecology and phenology of cattle ticks inZambia: development and survival of free living stages. Exp. Appl.Acarol. 8, 291–301.
Punyua, D.K., 1984. Development periods of Rhipicephalus appendiculatusNeumann (Acarina: Ixodidae) under field conditions. Insect Sci. Appl.5 (4), 247–250.
Rechav, Y., von Maltzahn, H., 1977. Hatching and weight changes in eggsof two species of ticks in relation to saturation deficit. Ann. Entomol.Soc. Am. 70 (5), 768–770.
Short, N.J., Floyd, R.B., Norval, R.A.I., Sutherst, R.W., 1989. Developmentrate, fecundity and survival of developmental stages of the ticks R.appendiculatus, B. decoloratus and B. microplus under field conditionsin Zimbabwe. Exp. Appl. Acarol. 6, 123–141.
Sweatman, G.K., 1967. Longevity and oviposition of engorged Rhipi-cephalus sanguineus female ticks. J. Parasitol. 53 (2), 432–445.
Tukahirwa, E.M., 1976. The effect of temperature and relative humidity onthe development of R. appendiculatus Neumann (Acarina: Ixodidae).Bull. Entomol. Res. 66, 303–312.
Van straren, M., Jonjegan, F., 1993. Ticks (Acari: Ixodidae) infesting theArabian camel (Camelus dromedarius) in Sinai, Egypt with a note onacaricidal efficacy of ivermectin. Exp. Appl. Acarol. 17, 605–612.
Zahler, M., Gothe, R., 1995. Effect of temperature and humidity on egghatching, moulting and longevity of larvae and nymphs of Dermacen-tor reticulatus (Ixodidae). Appl. Parasitol. 36, 53–65.
Recommended

















