
IS IT POSSIBLE TO INFLUENCE METABOLIC PROCESS OF EUROPEAN SEA BASS JUVENILES BY A PROCESS OF EUROPEAN SEA BASS JUVENILES BY A
NUTRITIONAL CONDITIONING DURING LARVAL STAGE ?
• M Vagner• M. Vagner• J.L. Zambonino Infante • J.H. Robin • J. Person-Le Ruyet
larvi 20095th fish & shellfish larviculture symposium
ghent university, belgium7 - 10 september 2009

INTRODUCTION
N dNatural fish stocks Nowadays
Carnivorous marine fish are mainly fed with fish meals and oils
Natural fish stocks
meals and oils
BUT fish aquaculture and natural fish stocks
Substitution of fish lipid sources with vegetable lipid sources
• Not adapted to the requirement in essential fatty acids (HUFA):
Disadvantages of vegetable products:
Not adapted to the requirement in essential fatty acids (HUFA):
rich in C18 fatty acids (i.e. HUFA precursors) BUT poor in HUFA (EPA, DHA)
• Low capacity for marine fish to convert C18 FA into HUFA, in particular because p y , pof the low expression of the Delta-6 desaturase (D6D)

*
EPAAA EPAAA
DHADHA

Hi h i i d i fi h hi h
INTRODUCTION
High interest in producing fish, which
could better incorporate vegetable p g
products.
BUT HOW ?
Could it be possible using the concept of p g p
metabolic programming ?

INTRODUCTION
Adaptive process at a cellular, molecular and biochemical levels occuring during young stages, and then maintained in further developmental stages during young stages, and then maintained in further developmental stages
and potentially transmissible to the offspring (Lucas, 1998)
I it ibl i b t i t h i l gi l f j il b l l Is it possible in sea bass to orient physiological process of juveniles by a larval
conditioning to dietary vegetable products, characterized by a low HUFA
content?

1. Experimental design
I- Nutritional conditioning 1
HH22HH16LH22LH16d-5-45
Mouth openning
stag
e
LH Low HUFA EPA+DHA= 0.8% DM HH High HUFA EPA+DHA=2.2% DM
16°C 22°C
2 diet
2 T°C
C1
Larv
al s
1 commercial IP
90 days
ex-LH22ex-LH16 ex-HH16 ex-HH22
ex LH22LH16 ex-HH16 ex-HH221 HUFA-d-151-21160 days
R1
diet
nile
sta
ge EPA+DHA= 2.7% DM 19°C
EPA+DHA= 0.5% DM
ex-LH22ex-LH16 ex-HH16 ex HH22
19°C
1 HUFArestricted
diet
R1
Juve
n

2. Growth performances
I- Nutritional conditioning 1 s
tage
100
120gh
t (m
g)
No effect of diet on T***d-45
C1Larv
al
40
60
80
vidu
al m
ean
wei
g
the survival ratediet ***
0
20
LH16 HH16 LH22 HH22
Indi
v
exHH2218
nile
sta
ge
R1
100% survival rate in all groups
exLH16exHH16
exLH22
14
18
an w
eigh
t (g)
Juve
n R1No significant effect of
nutritional conditionning10
Indi
vidu
al m
ea
J 211
6-10 0 10 20 30 40 50 60
time (days)

5. Lipid metabolism in larvae
I- Nutritional conditioning 1
D6D mRNA level at d-45 D6D Desaturation product at d-45 :
18:3n-6 in PL
0 4R l ti i i
***
***
0.3
0.418:3n-6 in % FAME***
6
8 Relative expression in relation to an
housekeeping gene and to a referential condition
HH22
0 1
0.2
NS
***
2
4
HH22
0
0.1
LH16 HH16 LH22 HH220
2
LH16 HH16 LH22
Stimulation of desaturation pathways for HUFA synthesis, without any effect of temperature

5. Lipid metabolism in larvae
I- Nutritional conditioning 1
******0.8
1.2
1.6
2
AA % FAME PL
10
0
0.4
LH16 HH16 LH22 HH22
EPA % FAME PL
******
4
6
8
HUFA deficiency
0
2
LH16 HH16 LH22 HH22
30DHA % FAME PL
******
5
10
15
20
25DHA % FAME PL
0
5
LH16 HH16 LH22 HH22LH16 HH16 LH22 HH22

6. Lipid metabolism in juveniles
I- Nutritional conditioning 1
D6 mRNA level – 1st part of the challenge PL final composition in DHA
*24% FAME
***6
Relative expression in
*22
% FAME
***3
4
5 relation to an house-keeping gene
20NS
NS
1
2
3
• Significant effect of larval conditionning
18ex-LH16 ex-HH16 ex-LH22 ex-HH22
0ex-LH16 ex-HH16 ex-LH22 ex-HH22
• Better capacity of ex-LH groups to Significant effect of larval conditionning
• BUT transient (30 days)
Better capacity of ex LH groups to adapt to a low-HUFA diet
• BUT with a low amplitude
• NS for EPA

7. Conclusions
I- Nutritional conditioning 1
POSSIBLE to influence fatty acid desaturation pathways for HUFA synthesis in
juveniles, using a nutritional conditioning during larval stage, without any effect
of temperature on the capacity of FA desaturation (- 30% of HUFA requirement)
BUT this modulation seems to be limited
Is it possible to obtain a response with a larger magnitude?

2. Experimental design
II. Nutritional conditioning 2
LH1 HH1d-5-45= C1
Mouth opening
sta
ge
C2XLH1 XH1
19°C
LH1 = 0.7% HH1 = 1.7%
30 days
Larv
al C24 diets XH1 =3.7%XLH1= 0.5%
30 days≠ 90 (C1)
PI
ge
1 commercial
diet EPA+DHA= 2.7% DM 19°C
XLH2 XH2LH2 HH2
d-83- 118≠d151-211 en
ile s
tag
R2
diet
1 more HUFA XLH2 XH2LH2 HH2
EPA DHA 2.7% DM
(R1)
35 daysJuve R2restricted
diet EPA+DHA= 0.3% DM 19°C

3. Growth performances
II. Nutritional conditioning 2ge a a
b b
***
No effect of diet on 50
60
ght (
mg)
Larv
al s
tag
C2
the survival rate
30
40
n in
divi
dual
wei
g
Idem C110
20
XLH1 LH1 HH1 XH1
Mea
nile
sta
ge
R2
100% of survival rate in all groups3.5
4.5
ivid
ual w
eigh
t (g
)
Juve
n R2 No significant effect of larval conditionning
Idem R11.5
2.5
Mea
n in
d
Idem R10.5
0 5 10 15 20 25 30 35

4. Lipid metabolism in larvae
II. Nutritional conditioning 2
0 6%18:3n-6 in % FAME
D6D mRNA level at d-45 D6D desaturation product at d-45 in PL
6 Relative expression in relation to an house keeping gene
a
a0.4%
0.5%
0.6%***
a
4
5 ***
house-keeping gene
b0.2%
0.3%
cc
b
2
3
c0.0%
0.1%
XLH LH HH XH0
1
XLH1 LH1 HH1 XH1
Stimulation of desaturation pathways for HUFA synthesis
Idem C1Idem C1

4. Lipid metabolism in larvaeII. Nutritional conditioning 2
d3.5%
d
c
b1.5%
2.0%
2.5%
3.0%
***
AA (% FAME in PL)Idem C1
d10%
12%
b
a
0.0%
0.5%
1.0%
XLH1 LH1 HH1 XH1
EPA (% FAME in PL)c
ba4%
6%
8%
10%
***EPA (% FAME in PL)
HUFA deficiency
0%
2%
XLH1 LH1 HH1 XH1d
30%
35%
40%
***DHA (% FAME in PL)
c
b
a10%
15%
20%
25%
30% ***
0%
5%
XLH1 LH1 HH1 XH1XLH1 LH1 HH1 XH1

5. Lipid metabolism in juveniles
II. Nutritional conditioning 2
D6D mRNA level PL final composition in DHA
NS18
% FAMEa
6
7XLH2 LH2 HH2 XH2
Relative expression in relation to an house-keeping gene
16
18
aa
ab a
b b4
5
6Diet ***
14
a
aa
bb
bc
cab
b
c1
2
3 IDEM for EPA
• Significant effect of larval
12ex-XLH ex-LH ex-HH ex-XH
bc c
0
1
J-83 J-90 J-107 J-118
• Non significant effect of larval conditioning
≠ R1
• D6D stimulation NON transient
gconditionning
≠ R1 ≠ R1≠ R1

Summary
Although no beneficial effect on growth performances and no clear higher HUFA content in PL, POSSIBLE to influence FA desaturation pathways for HUFA synthesis for a better incorporation of vegetable products, using a nutritional
Possible to use the concept of metabolic programming in sea bass
y p g p , gconditioning during larval stage (-60% of the HUFA requirement)
Possible to use the concept of metabolic programming in sea bass
BUT is it a:Long lasting adaptation?Long-lasting adaptation?
Transient acclimatation of juveniles to their nutritional environment ?
Need to better investigate mechanisms involved in desaturation pathways in response to a low HUFA dietary content in larvae and in juveniles
PPAR

InhibitionStimulation of desaturation pathways for HUFA synthesis
0.8 1.7%0.5 0.7XLH1 LH1 LH HH1 XH1
3.7%HH2.2
D6D+ + D6D - D6DPPAR+ PPAR- PPAR-
Implementation of metabolic programming using a nutritional conditioning during larval stage
InhibitionStimulation of desaturation pathways for HUFA synthesis
D6D+ + D6D - D6DPPAR+ PPAR- PPAR-
Possible to influence the FA desaturation pathways for HUFA synthesis for a better incorporation of vegetable products
BUT
NO transient stimulation Transient stimulation
0,3 0,5HUFA
requirement at 0,7%
R2 R1
Additional factors: HUFA n-3 from the juvenile environment
DHA content in PL
NO transient stimulation Transient stimulationDHA content in PL
Control of desaturation mechanisms previously gained
Control of the fish lipid composition

IV- Perspectives
Was the metabolic programming really present ?
Transmission to Intermediate Transmission to the offspring ?period: Stop
or Non stop
?Challenge extension:
what will happen during
?adulthood ?
IF YES
Application to other carnivorous species with high commercial value in aquaculture

Thanks to the financially support: Ifremer, INRA, RAQ, Larvi.
Th k h h i l M M L G ll A Sé è H
y pp , , Q,
Thanks to the technical support: M.M Le Gall, A. Sévère, H. Le Delliou, P. Quazuguel, N. Le Bayon, J. Moriceau, C.
Huelvan & E. Desbruyères.
Thanks for your attention
y
Thanks for your attention
larvi 20095th fish & shellfish larviculture symposium
ghent university, belgium7 - 10 september 2009

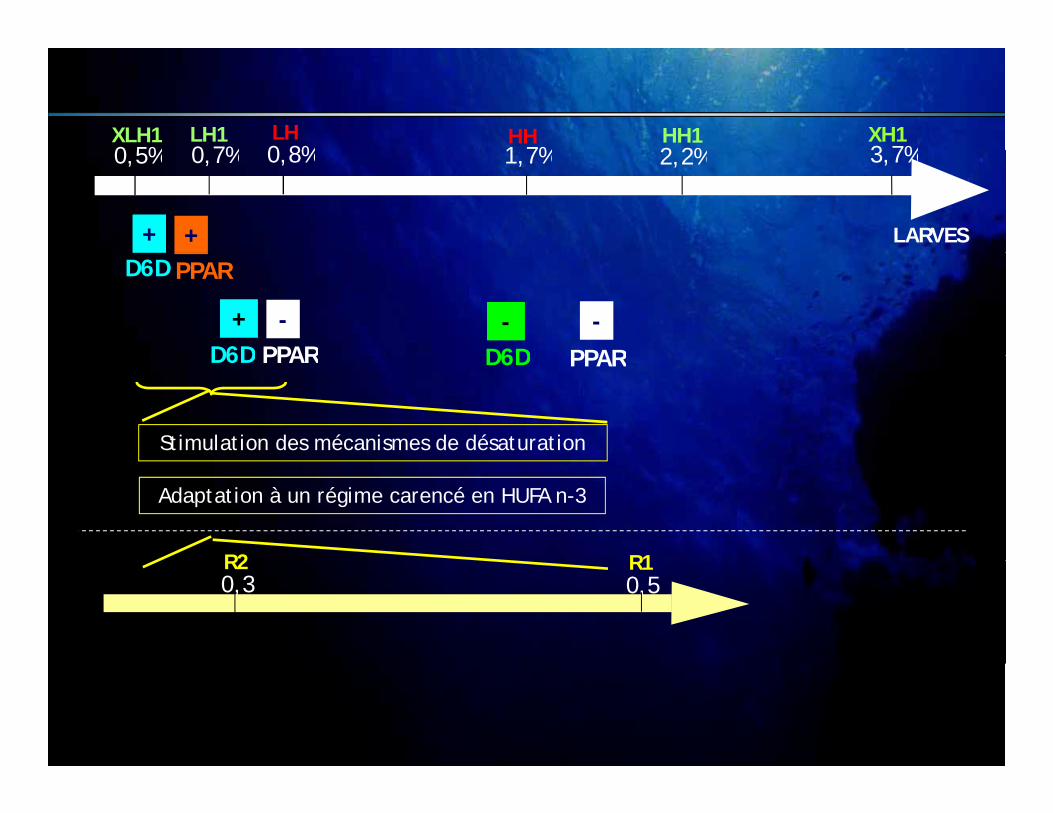
Récapitulatif
XLH1 LH1 LH HH1HH XH10 5% 0 7% 0 8% 1 7% 2 2% 3 7%
LARVES
0,5% 0,7% 0,8% 1,7% 2,2% 3,7%
Adaptation à un régime carencé en HUFA n-3
Stimulation des mécanismes de désaturation
Niveau ARNm D6 continue Niveau ARNm D6 transitoire
R2 R10,3 0,5
JUVENILES
Niveau ARNm D6 continue Niveau ARNm D6 transitoire
D6 non fonctionnelle ?D6 fonctionnelle ?
R1 trop proche du besoin (0,7% EPA+DHA)Synthèse d’HUFA-
dé i é
teneur en DHA Faible en DHA
dérivées
XLH1 LH1 LH HH1HH XH10 5% 0 7% 0 8% 1 7% 2 2% 3 7%
LARVES
0,5% 0,7% 0,8% 1,7% 2,2% 3,7%
+ +D6D
+D6D
-D6D
PPAR
PPAR-
PPAR-
D6D D6DPPAR PPAR
Stimulation des mécanismes de désaturation
Adaptation à un régime carencé en HUFA n-3
R2 R1R2 R10,3 0,5

HUFA = AGLPI (n-9, n-6 et n-3) = Acides gras insaturés à 20-22 atomes de
carbone et au moins 4 doubles liaisons = Acides gras essentiels
C:X n-Y
g
Constituants principaux des PL de la bi-couche membranaire
O
OH -Acide docosahéxaénoique 22:6n-3 (DHA)
Chez les poissons, majoritairement les HUFA n-3:
1
2
3
45
6
-Acide écosapentaénoique 20:5n-3 (EPA)
Acide arachidonique 20:4n-6 (AA)
6
• Croissance et survie larvaire
• Développement de la vision et du cerveau
• Résistance au stress et aux maladies
• Qualité de la chair, …

PPARs
III- Hypothesis concerning the regulation of lipid metabolism in sea bass
XLH1 LH1 LH HH1HH XH10.5% 0.7% 0.8% 1.7% 2.2% 3.7%
stag
e
5 D6D
+ +
D6D
-
D6D
Larv
al s
d-4
PPAR+
PPAR-
PPAR-
tage
of
nge
++ -
uven
ile s
tTh
e en
d he
cha
lle
D6DD6D
+
D6D
-
J t
PPAR+
PPAR-
PPAR

Étude des PPAR
III- Hypothèses de régulation du métabolisme lipidique du bar
P i P lif t A ti t d R t é t lé iPeroxisome Proliferator-Activated Receptor, récepteurs nucléaires
Trois isoformes: α, β et γ
Décrits chez les mammifères et le bar
Facteur de transcription de la delta 6 chez les mammifères
Dans le contexte de programmation métabolique, les PPAR sont-ils impliqués dans la stimulation de la transcription du gène de la D6D chez le bar?

III- Hypothèses de régulation du métabolisme lipidique du bar
PPAR
D6D+
PPAR
D6D+
Mise en place d’une mémorisation au niveau moléculaire des pmécanismes de désaturation des AG

IV- Conclusions & Perspectives
1. General conclusion
1 Nutritional environment
2
POSSIBLE to influence the fatty acid desaturation pathways for
Nutritional conditionning acid desaturation pathways for
HUFA synthesis for a better incorporation of vegetable meals
conditionning during larval
stagePreviously gained
mechanisms
Fish lipid composition3
0 5 < HUFA di < 0 7
PPAR+
D6D transcription
0.5 < HUFA dietary content < 0.7

0,80,5 0,7 1,7Teneur en EPA+DHA du régime (%MS)
AAA
Inhibition
2,2 3,7
Stimulation des mécanismes de désaturation
-
HUFAHUFA HUFA
HUFA
HUFAHUFA
-
HUFAHUFA HUFA
HUFA
HUFAHUFA
HUFAHUFA HUFA
HUFA
HUFAHUFA
HUFAHUFA
+
HUFAHUFA
HUFAHUFA
HUFA
+
e
Gène de la delta 6
PPAR? ?
de la
rvai
re
-++
Stad
Traduction de l’ARNm en protéine
-+ +Transcription du gène de la Δ6-D
Synthèse d’AG précurseurs des HUFA-+ +
Mise en place d’une programmation métabolique par le conditionnement larvaire

Présence de cette programmation chez les juvéniles
Facteurs additionnels: teneur en HUFA n-3 de l’environnement nutritionnel
Teneur en EPA+DHA du régime (%MS)R2 = 0 3% R1 = 0 5% 0 7g ( )R2 = 0,3% R1 = 0,5% 0.7
Contrôle des mécanismes de désaturation préalablement acquis
e ju
véni
le +
Ni ARN D6 t it i
Transcription du gène de la D6D
-Début du challenge Fin du challenge+
Stad
e
D6 non fonctionnelle ?
D6 fonctionnelle ?
Niveau ARNm D6 continue Niveau ARNm D6 transitoire
+ -
dérivées
-Synthèse d’HUFA
+ -dérivées
teneur en DHA Faible en DHA
+ -
Recommended

















