
ORIGINAL PAPER
Fossil mesostigmatid mites (Mesostigmata: Gamasina,Microgyniina, Uropodina), associated with longhorn beetles(Coleoptera: Cerambycidae) in Baltic amber
Jason A. Dunlop & Jenő Kontschán & Michael Zwanzig
Received: 14 January 2013 /Revised: 25 February 2013 /Accepted: 1 March 2013 /Published online: 2 April 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Fossil mesostigmatid mites are extremely rare.Inclusions assignable to the tortoise mites (Mesostigmata,Uropodina) are described here for the first time from Eo-cene (ca. 44–49 Ma) Baltic amber. This is the oldest recordof Uropodina and documents the first unequivocal amberexamples potentially assignable to the extant genusUroobovella Berlese, 1903 (Uropodoidea: Urodinychidae). Further mites in the same amber pieces are tentativelyassigned to Microgynioidea (Microgyniina) and Ascidae(Gamasina), both potentially representing the oldest re-cords of their respective superfamily and family groups.This new material also preserves behavioural ecology in theform of phoretic deutonymphs attached to their carriers viaa characteristic anal pedicel. These deutonymphs in amberare intimately associated with longhorn beetles (Coleop-tera: Cerambycidae), probably belonging to the extinct
species Nothorhina granulicollis Zang, 1905. Modernuropodines have been recorded phoretic on species belong-ing to several beetle families, including records of livingUroobovella spp. occurring on longhorn beetles. Throughthese amber inclusions, a uropodine–cerambycid associationcan now be dated back to at least the Eocene.
Keywords Parasitiformes . Eocene . Deutonymph .
Anal pedicel . Phoresy
Introduction
Mites can be divided into two principal branches: Acariformesand Parasitiformes. The latter is further subdivided into fourclades (systematics after Krantz and Walter 2009), the mostspecies rich of which is the Mesostigmata (=Gamasida insome schemes) for whom Beaulieu et al. (2011) recognised11,424 living species. Despite this modern diversity and theirubiquity in soil habitats in particular, mesostigmatid miteshave a poor fossil record. Only four species have been for-mally described, plus some material often tentatively assignedto genera or families (Table 1; see also ‘Discussion’). Thisapparent rarity of fossil mesostigmatids remains a puzzle inarachnid palaeontology. It could be an artefact of ambers andother localities, not sampling enough of the soil fauna. Alter-natively, as we demonstrate here, specimens do exist but havenot been recognised or assigned correctly. Three principalclades of mesostigmatid mites are currently accepted: Sejida,Trigynaspida and Monogynaspida. The monogynaspidscan be further subdivided into various cohorts, includingMicrogyniina, Gamasina and Uropodina.
Uropodines are sometimes referred to as ‘tortoise mites’,although it should be stressed that this is a morphologicalcharacterisation referring to their often dome-like dorsalsurface, rather than a biological association with tortoises
Communicated by: Sven Thatje
J. A. Dunlop (*)Museum für Naturkunde, Leibniz Institute for Researchon Evolution and Biodiversity at the HumboldtUniversity Berlin, Invalidenstrasse 43,10115 Berlin, Germanye-mail: [email protected]
J. KontschánPlant Protection Institute, Centre for Agricultural Research,Hungarian Academy of Sciences, P.O. Box 102,1525 Budapest, Hungarye-mail: [email protected]
J. KontschánDepartment of Zoology and Animal Ecology, Szent IstvánUniversity, Gödöllő, Páter Károly str. 1,2100 Győr, Hungary
M. ZwanzigScheiblerstrasse 26,12437 Berlin, Germanye-mail: [email protected]
Naturwissenschaften (2013) 100:337–344DOI 10.1007/s00114-013-1031-8
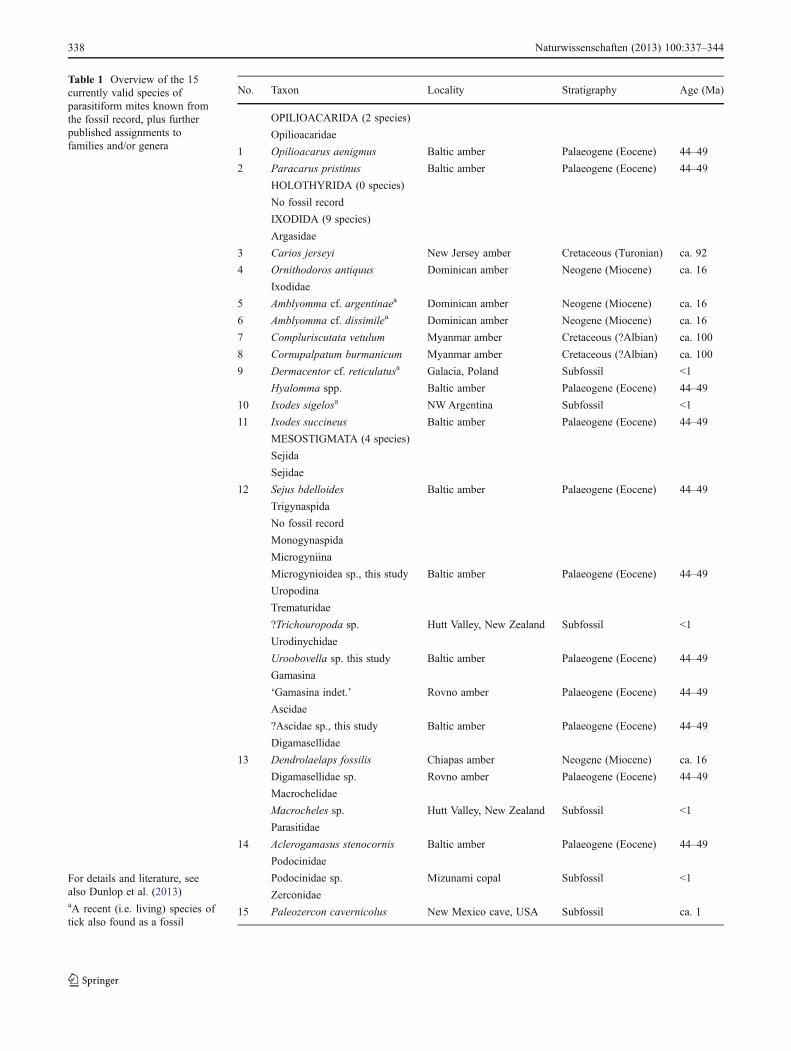
Table 1 Overview of the 15currently valid species ofparasitiform mites known fromthe fossil record, plus furtherpublished assignments tofamilies and/or genera
For details and literature, seealso Dunlop et al. (2013)aA recent (i.e. living) species oftick also found as a fossil
No. Taxon Locality Stratigraphy Age (Ma)
OPILIOACARIDA (2 species)
Opilioacaridae
1 Opilioacarus aenigmus Baltic amber Palaeogene (Eocene) 44–49
2 Paracarus pristinus Baltic amber Palaeogene (Eocene) 44–49
HOLOTHYRIDA (0 species)
No fossil record
IXODIDA (9 species)
Argasidae
3 Carios jerseyi New Jersey amber Cretaceous (Turonian) ca. 92
4 Ornithodoros antiquus Dominican amber Neogene (Miocene) ca. 16
Ixodidae
5 Amblyomma cf. argentinaea Dominican amber Neogene (Miocene) ca. 16
6 Amblyomma cf. dissimilea Dominican amber Neogene (Miocene) ca. 16
7 Compluriscutata vetulum Myanmar amber Cretaceous (?Albian) ca. 100
8 Cornupalpatum burmanicum Myanmar amber Cretaceous (?Albian) ca. 100
9 Dermacentor cf. reticulatusa Galacia, Poland Subfossil <1
Hyalomma spp. Baltic amber Palaeogene (Eocene) 44–49
10 Ixodes sigelosa NWArgentina Subfossil <1
11 Ixodes succineus Baltic amber Palaeogene (Eocene) 44–49
MESOSTIGMATA (4 species)
Sejida
Sejidae
12 Sejus bdelloides Baltic amber Palaeogene (Eocene) 44–49
Trigynaspida
No fossil record
Monogynaspida
Microgyniina
Microgynioidea sp., this study Baltic amber Palaeogene (Eocene) 44–49
Uropodina
Trematuridae
?Trichouropoda sp. Hutt Valley, New Zealand Subfossil <1
Urodinychidae
Uroobovella sp. this study Baltic amber Palaeogene (Eocene) 44–49
Gamasina
‘Gamasina indet.’ Rovno amber Palaeogene (Eocene) 44–49
Ascidae
?Ascidae sp., this study Baltic amber Palaeogene (Eocene) 44–49
Digamasellidae
13 Dendrolaelaps fossilis Chiapas amber Neogene (Miocene) ca. 16
Digamasellidae sp. Rovno amber Palaeogene (Eocene) 44–49
Macrochelidae
Macrocheles sp. Hutt Valley, New Zealand Subfossil <1
Parasitidae
14 Aclerogamasus stenocornis Baltic amber Palaeogene (Eocene) 44–49
Podocinidae
Podocinidae sp. Mizunami copal Subfossil <1
Zerconidae
15 Paleozercon cavernicolus New Mexico cave, USA Subfossil ca. 1
338 Naturwissenschaften (2013) 100:337–344

or turtles (Testudines). Uropodina are recognisable by areduction in the amount of setae on the legs and palps, aswell as modifications of the mouthparts and tritosternum(Lindquist et al. 2009). While the ecology of only a fewliving species is known in detail, a diversity of lifestyles hasbeen observed across the group (see ‘Discussion’). Manyoccur in dung or decaying organic matter where the adultsmay feed on nematodes and/or insect larvae. Numerousmesostigmatids show intimate associations with other ar-thropods (reviewed by Hunter and Rosario 1988). Amonguropodines, this manifests itself as phoresy; the mitehitching a ride on another arthropod in order to facilitateits distribution over a wider area and/or to reach a morefavourable habitat. Specifically, in uropodines, the immaturedeutonymph stage can secrete a stalk-like pedicel from itsanal pedicellar glands and uses this to attach itself to acarrier (Faasch and Schaller 1966; Faasch 1967; Athias-Binche and Evans 1981; Bajerlein and Błoszyk 2004). Fora detailed modern account of this attachment mechanismand the associated gland structures, see Bajerlein andWitaliński (2012). A general summary of uropodine biologyand systematics can be found in Karg (1989).
Here, we describe the first formal records of the Uropodinafamily Urodinychidae (superfamily Uropodoidea) from Balticamber, based on deutonymphs in phoretic association withfossil longhorn beetles (Coleoptera: Cerambycidae). Othermites are found in the same amber pieces and are thus poten-tially associated with these beetles, too. They probably belongto two different cohorts: the superfamily Microgynioidea in theMicrogyniina and the family Ascidae in the Gamasina. In allcases, these are the oldest and, in the last two cases, the onlypalaeontological records of their respective (super) families.
Material and methods
Three pieces of Baltic amber were available for studycontaining mites associated with beetles. One of these (Fig. 1)stems is from the collection of one of the authors (MZ) andaccording to the original dealer originated from the Yantarnymine in the Kaliningrad province of the Russian Federation.This specimen has now been deposited in the palaeontologicalamber collections of theMuseum für Naturkunde, Berlin, underthe repository number MB.A. 1879 (for Museum BerlinArthropoda). The second specimen (Fig. 2) was made availablefrom the private collection of Wolfgang Weitschat. It has alsonow been donated to the Berlin museum under the repositorynumber MB.A. 1900. A third specimen (not figured) camefrom the Geologische-Pälaontologisches Institut und Museumin Hamburg (GPIH), no. 0425. For the latter two specimens,exact details of the original locality are not available. Fossilswere examined under a Leica MZ12s stereomicroscope andphotographed using a Nikon D700 digital camera with a series
of lenses. Multiple image stacks were combined into finalimages using the software package Combine ZP. All measure-ments are in micrometres.
Amber is difficult to date accurately, but Baltic am-ber is usually assigned to an Eocene (Lutetian) age ofabout 44–49 Ma. The original amber forest has beeninterpreted as a warm temperate to subtropical environ-ment covering much of modern Fennoscandia and be-yond. For a recent overview and geological setting, seeWeitschat and Wichard (2010). Mite nomenclature andhigher taxon names are derived from Lindquist et al. (2009)and Beaulieu et al. (2011).
Systematic palaeontology
Order Parasitiformes Reuter, 1909Suborder Mesostigmata G. Canestrini, 1891Cohort Uropodina Kramer, 1881Superfamily Uropodoidea Kramer, 1881Family Urodinychidae Berlese, 1917Genus ?Uroobovella Berlese, 1903 sensu lato (Figs. 1 and 2)
Material, locality and horizon MB.A. 1879 (ex coll.Zwanzig) and 1900 (ex coll. Weitschat); GPIH 0425. Balticamber forest; Palaeogene, Eocene, Lutetian.
Description Small, shield-shaped deutonymphs have a max-imum length 460 and maximum width 300. Idiosoma is thusoval, ca. 1.5 times longer than being wide; colour, reddishbrown. Dorsal part of the body is covered by slightly domedholodorsal shield; ventral part, with sternal and ventral shields.Pedofossae are deep and well developed; legs are short andplaced on their respective pedofossae. Individual mites oftenattached to the carrier via a short, sometimes curved, analpedicel; maximum length is 220. MB.A. 1879 hosts the larg-est number of mites (Fig. 1); isolated deutonymphs hangventrally from the thorax or sit on the femur of the secondleg, with much greater concentrations of at least 20 individualsattached around the coxae of the second leg (near the thorax–abdomen junction) and at least 14 individuals on the thirdfrom the last sternite. In MB.A. 1900, a few deutonymphs areattached to the distal articles of the hindlegs. In GPIH 0425(not figured), three to four deutonymphs are attached to thetrochanter of leg 1 and the femur of leg 3.
Remarks The deutonymph fossils with the anal pedicel aretentatively assigned here to the Recent genusUroobovella basedlargely on the shape of the idiosoma. Specifically, their bodiesare more elliptical than in species of the similar looking andclosely related genus Uropoda Latreille, 1806 (Uropodidae).Unfortunately, the most important difference betweenUropodidae/Uropoda and Urodinychidae/Uroobovella is thepresence or absence of an internal sclerotised node associated
Naturwissenschaften (2013) 100:337–344 339

with the levator tendon on the chelicerae (mouthparts). Thischaracter is not visible in our amber specimens and would behard to resolve even using tomographic methods. Note thatseveral Trichouropoda (Trematuridae) nymphs also occur inassociation with longhorn beetles. Recent examples of thisgenus are easy to recognise from the shape of the corniculi,tritosternum and hypostomal setae; all of which are unfortunate-ly not visible in our fossils. We favour an assignment toUroobovella because the dorsal shield seems to be smooth inour specimens—something typical for Uroobovelladeutonymphs—as opposed to being ornamented, as is moreusual for Trichouropoda. Admittedly, some livingTrichouropoda species have been described as deutonymphswithout dorsal sculpture (e.g. Wiśniewski and Hirschmann1993; pp. 140–141), but these taxa are not associated withlonghorn beetles and were found instead on species ofScarabeidae, Brenthidae and Passalidae. In addition to the dorsalornament character, marginal scallops are not observed in ourspecimens. These would also be important characters forrecognising the fossils as Trichouropoda deutonymphs.
Irrespective of their generic affinities, it is worth noting thatin MB.A. 1879, the bodies of the mites attached to the beetle
thorax appear to be ca. 1.5 times shorter than the mites on theventral part of the beetle abdomen. In Uropodina, only thisdeutonymph stage undergoes phoresy, and in general,deutonymph body size is taxon specific; thus, we suspect thatat least two uropodine species attaching to different parts ofthe carrier's body may be present. The fossils are only knownfrom immature instars, and it would be poor practice in therecent uropodine taxonomy to name species based ondeutonymphs only. Since these fossils do not preserve mean-ingful characters appropriate for modern species descriptions,we simply record the presence of Uropodina (probably ?Uroobovella) in Eocene Baltic amber and refrain from creat-ing any species name(s) at this stage.
Cohort Microgyniina Trägårdh, 1942Superfamily ?Microgynioidea Trägårdh, 1942 (Fig. 2, inset left)
Material, locality and horizon. MB.A. 1900 (see above).
Description There are three small and narrow adult mites;idiosoma is oblong with parallel lateral margins, length 390and width 190. Ventral side of body is not visible; dorsal
Fig. 1 MB.A. 1879. Fossil longhorn beetle (Coleoptera: Cerambycidae)in Eocene (ca. 44–49 Ma) Baltic amber, probably assignable to N.granulicollis Zhang, 1905. This specimen carries multiple phoreticdeutonymphs of tortoise mites (Mesostigmata: Uropodina:Urodinychidae: ?Uroobovella). Lower left and right insets (arrowed)
show details of some of these mites, attached to the carrier by a distinctanal pedicel. These findings represent the oldest published record ofUropodina. Upper left inset shows a putative member of anothermesostigmatid family, Ascidae (note the pro- and opisthonotal shield);potentially the oldest record of the group
340 Naturwissenschaften (2013) 100:337–344

side with three dorsal plates is interpreted as the pronotal,mesonotal and pygidial shields with respective lengths 230,80 and 80. Division between pronotal and mesonotal shieldsis clearer (accentuated by slight lateral indents) than thatbetween mesonotal and pygidial shields. Pedipalps are smalland pediform; legs are short and robust, but details and/orsetation is equivocal. All three mites apparently sit as agroup on the posterolateral sides of the carrier's thorax, butno attachment organ or pedicel is evident.
Remarks As noted by Lindquist et al. (2009; p. 131), thedorsal surface of microgynioids is typically divided intothree distinct plates (Fig. 2, inset left, plates numbered). Athree-part-divided dorsal surface also occurs in certain othermesostigmatids, such as Sejidae from the Sejida clade.However, sejiids tend to be larger mites with convex lateralmargins of the body (cf. Lindquist et al. 2009, Fig. 12.6–7).We favour assigning these three inclusions to themicrogynioids, based on their small size and parallel sidedbody. In particular, there are strong hints that a further(posterior) element may also be present on the dorsal sur-face, representing the typical third (pygidial) shield ofmicrogynioids. This observation is towards the limits ofresolution in the amber, and we thus assign the fossilstentatively to Microgynioidea, but if correct, it would
represent the first fossil record of this superfamily. Note thata phoretic deutonymph with a peduncle is also known fromat least one extant genus, Microsejus Trägårdh, 1942, and itis conceivable that some of the deutonymphs associatedwith these amber beetles originated from microgynioids aswell as urodinychids.
Cohort Gamasina Kramer, 1881Subcohort Dermanyssiae Evans and Till, 1979Superfamily Ascoidea Voigts and Oudemans, 1905Family ?Ascidae Voigts and Oudemans, 1905 (Fig. 1, insetleft above)
Material, locality and horizon MB.A. 1879 (see above).
Description There is a medium-sized mite (length 300),with dorsum covered with two shields. Idiosoma is oval,caudally rounded, on anterolateral areas of margins shal-low, with shoulder-like bulge. Ventral aspect is not vis-ible; dorsal side is divided into pronotal and opisthonotalshields with respective lengths 160 and 140. Caudaldepression is not visible. Legs are short and stout, anddetails are equivocal. This mite sits on the anterior leftcorner of the hosts' thorax, but no attachment organ orpedicel is evident.
Fig. 2 MB.A. 1900. Second example of N. granulicollis Zhang, 1905from Baltic amber-carrying mites. Left inset shows three putativeexamples of microgynioid mites (Mesostigmata: Microgyniina:
Microgynioidea) attached to the thorax; the characteristic tripartitedivision of the dorsal shield is numbered. Right inset again showsdeutonymphs of uropodine mites, here attached to the beetle's hindleg
Naturwissenschaften (2013) 100:337–344 341

Remarks Ascidae is quite a variable group, but we tentativelyrefer the fossil to this gamasine family based on the division ofthe dorsal surface into a pronotal and opisthonotal shield (cf.the tripartite shield of the fossils above). This two-part divi-sion (Fig. 1, inset above left) characterises at least some generaand species of this family (Lindquist et al. 2009, Fig. 12.51),although others have an entire dorsal shield. Based on theshape of the idiosoma, this inclusion is reminiscent of therecent ascid genera Zerconopsis Hull, 1918 and AnntenoseiusBerlese, 1916. Species of Zercon Koch, 1836 (familyZerconidae) have similarly divided dorsal plates, but the ab-sence of dorsal cavities in the fossil suggests that it does notbelong to the zerconids. We provisionally refer this amberinclusion to Ascidae. Again, this would be the oldest—andonly—fossil record of the family, and if the ventral part ofbody was accessible, it may be possible to determine itsaffinities more precisely.
Discussion
Fossil Mesostigmata
Compared to a relatively rich fossil record for acariformmites, almost 300 species going back at least 410 millionyears, the record of parasitiform mites (Table 1) is surpris-ingly sparse. Fifteen species have been formally described,including four subfossil ticks (Ixodida) effectively assign-able to living species. Despite being a derived (i.e. ectopar-asitic) clade, ticks also represent the oldest parasitiformscourtesy of Cretaceous (ca. 100 Ma) records from Myanmar(Burmese) amber. Of the other major parasitiform clades,Holothyrida have no fossil record. Opilioacarida are repre-sented by two species, both from Eocene (44–49 Ma) Balticamber, and potentially assignable to living genera.
Mesostigmata makes up the fourth major parasitiform mitegroup. The oldest examples are contemporary findings fromBaltic amber including a putative member of the Sejida: Sejusbdelloides Koch and Berendt, 1854 (Sejidae). The originaldrawing is somewhat idealised, and the fossil merits restudy todetermine whether it really belongs in this group. The secondclade, Trigynaspida, does not have any fossil representativesat all. Most of the (very few) fossil mesostigmatids thus comefrom the third clade Monogynaspida and, in particular, thecohort Gamasina (Table 1). They include a second, morereliable, record from Baltic amber in the subcohort ParasitaeAclerogamasus stenocornis Witaliński, 2000 (Parasitidae).From the probably contemporary Rovno amber of theUkraine, there are reports of another gamasine subcohortDermanyssiae, specifically the family Digamasellidae, plus‘Gamasina indet.’ (Perkovsky et al. 2007, Table 2). Neitherrecord has been formally described. Somewhat youngeris another example of Dermanyssiae, this time from Miocene
(ca. 16 Ma) Chiapas amber of Mexico described asDendrolaelaps fossilis Hirschmann, 1971 (Digamasellidae).A putative tortoise mite was also figured from thestratigraphically contemporary Dominican Republic amberin a popular science article by Poinar (1982), here attachedto a wood-boring beetle. A formal description enabling theaffinities of this fossil mite to be confirmed is unfortunatelylacking, and the specimen can no longer be traced (Poinar,pers. comm., 2012).
Younger still is a representative of the gamasine subcohortEpicriiae from a ca. 1Ma cave system described asPaleozerconcavernicolus Błaszak, Cokendolpher and Polyak, 1995(Zerconidae). This is the only fossil mesostigmatid to beassigned to an extinct genus; all other described taxa have beenreferred to Recent genera. Probably of similar age, there is acitation to Podocinidae sp. (Dermanyssiae) from the JapaneseMizunami copal (Aoki 1974). Finally and barely counting asfossils at all, Ramsay (1960) described mesostigmatids fromboreholes in the Hutt Valley, Wellington, New Zealand. Thesemicropalaeontological subfossils from the ca. 7–8,000-year-oldPetone Marine Beds include the living genus Macrocheles sp.(Dermanyssiae: Macrochelidae). Significantly, the Hutt Valleymaterial also includes the only other (sub)fossil record ofUropodina—a fossil questionably assigned to Oodinychus sp.Note that this genus is now a junior synonym of TrichouropodaBerlese, 1916 (cf. Karg 1989, p. 88). Thus, the fossil shouldnow be formally listed as ?Trichouropoda sp. in the uropodinefamily Trematuridae (cf. Table 1).
Mite associations in amber
The Baltic amber specimens described here extend the fossilrecord of Uropodina back to ca. 49 million years via two of itsfour superfamilies: Uropodoidea and Microgynioidea. In addi-tion to their systematic value, these findings are also significantfor preserving biological associations and behavioural ecology.Like living uropodines (Bajerlein and Witaliński 2012; andreferences therein), the fossil ?Uroobovella deutonymphs areattached to their carriers via a short, stalk-like anal pedicel(Fig. 1, insets). Our material thus dates this specific mode ofphoretic behaviour in uropodines back to the Eocene. It is afurther example of associations between mites and other ar-thropods in amber, as reviewed by Eichmann (2002). The mostcommon records are of parasitism by larval parasitengonidmites, particularly Erythraeidae, which are found attached bytheir mouthparts to insects across a range of ambers. RarerCretaceous findings include the Heterostigmata mite familiesAcarophenacidae associated with coccids (scale insects) inSiberian amber (Magowski 1994) andResinacaridae associatedwith mantis flies in Myanmar (Burmese) amber (Khaustovand Poinar 2010). There is also an example of phoresy by?Histostomatidae (Astigmata) on a spider in Baltic amber(Dunlop et al. 2012). Note that all of these records are
342 Naturwissenschaften (2013) 100:337–344

based on acariform mites. The fossils described here arethe oldest parasitiform mites found phoretically attachedto other arthropods.
The carrier
Of particular interest is the identity of the carrier. Modernuropodines are sometimes found in dung and may rely oncoprophilous beetles such as dung (Aphodiidae), scarab(Scarabaeidae) or dor beetles (Geotrupidae) for transportingtheir deutonymphs to new habitats (Bajerlein and Błoszyk2004). By contrast, the fossils described here are associatedwith longhorn beetles (Cerambycidae). This makes a certaindegree of ecological sense in that amber is a fossilised treeresin. It is thus more likely to trap insects associated withtree trunks—longhorn beetle larvae typically burrow intowood—as compared to taxa associated with ephemeral hab-itats on the forest floor such as animal faeces. Cerambycidaehave long been known from Baltic amber. Based on theproportions of the antennal segments and the hindleg tarsi,our fossils closely match the description of Palaeoasemumduffyi Abdullah, 1966. This species was subsequentlyregarded as a junior synonym of Nothorhina granulicollisZhang, 1905 by Vitali (2006).
Associations between Uropodoidea deutonymphs andCerambycidae are also known from modern warm ortemperate ecosystems. Polyaspis criocephali Wiśniewski,1980 (Polyaspidae) was reported by Karg (1989) fromCriocephalus rusticus L. in central Europe. Trichouropodaovalis (Koch, 1839) (Trematuridae) was found on Asemumstriatum (L.) by Kofler and Schmölzer (2000) in Austria.Several Trichouropoda deutonymphs have been collectedfrom Cerambycidae in Slovakia (cf. Mašán 2001);Trichouropoda livorniana Wiśniewski and Hirschmann,1988 was collected on Rhagium inquisitor L., Trichouropodapolysetosa Mašán, 1999 on Cerambyx cerdo L. andTrichouropoda shcherbakae Hirschmann, 1972 on Megopisscabricornis Scopoli. From Louisiana, USA, Moser andRoton (1971) reported Trichouropoda tuberosa Hirschmannand Zirngiebl-Nicol, 1961 on Xylotrechus rusticus L. InPoland, Trichouropoda australis Hirschmann, 1972 andTrichouropoda lamellosa Hirschmann, 1972 were found onMonochamus carolinensis (Olivier), and Trichouropodahirsuta Hirschmann, 1972 on Monochamus titillator(Farbicius) (cf. Wiśniewski 1982). Additionally, Wiśniewskiand Hirschmann's (1993) catalogue lists numerousUropodina species associated with Cerambycidae, butwithout details of the host. Of particular relevance toour fossil ?Uroobovella specimens, the extant speciesUroobovella californica Wiśniewski and Hirschmann, 1992and Uroobovella longitricha Wiśniewski and Hirschmann,1992 (Urodinychidae) were recorded by these authors fromundetermined Californian Cerambycidae. Our fossils imply
that such intimate relationships between uropodine mites andlonghorn beetles have a long history, datable to at least theEocene in north-central Europe.
The other mites
Of the other findings described here, modern microgynioidmites can be associated with rotting wood (Lindquist et al.2009), and this may explain their presence on a longhornbeetle, too. As noted above, at least one living microgynioidgenus has a phoretic deutonymph. Associations with a fewbeetle families (albeit not Cerambycidae) were reported forMicrogyniidae by Salmane and Telnov (2009). However, it isunclear how common such associations are in modern eco-systems, and without a clear Recent comparison, it is difficultto say whether the three (adult) amber microgynioids de-scribed here (Fig. 2) were also habitually phoretic. The thirdgroup, the ascids, is generally free, living and apart from sometaxa found in the nasal cavities of hummingbirds (Hunter1970) and two genera having associations with carabid beetles(e.g. Eidelberg 2000); they are not known to specificallyexhibit phoresy on beetles in modern ecosystems. In this case,the association of this ascid mite with a longhorn beetle(Fig. 1) may simply be fortuitous.
Conclusions
From only three pieces of amber, we could describe fossilmesostigmatid mites belonging to three different cohorts.All represent the oldest records of their respective familiesor superfamilies. In at least two cases, this was thanks totheir associations with other (larger) arthropods. We suggestthat the rarity of Mesostigmata in the fossil record is largelya sampling artefact and encourage further detailed study ofbeetles (and other insects) in amber in the hope that this willyield more examples of fossil uropodines and other miteswith phoretic or ectoparasitic instars.
Acknowledgments We thank Wolfgang Weitschat (Hamburg) forproviding material and Christian Neumann (MfN) for curatorial assis-tance. Johannes Frisch (MfN) helped identify the beetles, Stefan Wirth(MfN) and George Poinar Jr. (Oregon) provided helpful comments andAndreas Stark (Halle (Saale)) and Linnea Blase (Berlin) assisted withpreparing the figures. The reviewers are also thanked for their numeroushelpful suggestions. JK's visit to the MfN was funded by the EuropeanUnion's ‘Synthesys’ program.
References
Aoki J (1974) On the fossil mites in Mizunami amber from GifuPrefecture, Central Japan. Bull Mizunami Fossil Mus 1:397–399, in Japanese with English summary
Naturwissenschaften (2013) 100:337–344 343

Athias-Binche F, Evans GO (1981) Observations on the genusProtodinychus Evans, 1957 (Acari: Mesostigmata) with descrip-tions of the male and phoretic deuteronymph. Proc R Irish Acad81B:25–36
Bajerlein D, Błoszyk J (2004) Phoresy of Uropoda orbicularis (Acari:Mesostigmata) by beetles (Coleoptera) associated with cattle dungin Poland. Eur J Entomol 101:185–188
Bajerlein D, Witaliński W (2012) Anatomy and fine structure ofpedicellar glands in phoretic deutonymphs of uropodid mites(Acari: Mesostigmata). Arth Strc Dev 41:245–257
Beaulieu F, Dowling APG, Klompen H, de Moraes GJ, Walter DE(2011) Superorder Parasitiformes Reuter, 1909. In: Zhang Z-Q(ed) Animal biodiversity: an outline of higher-level classificationand survey of taxonomic richness. Zootaxa 3148:123–128
Dunlop JA, Wirth S, Penney D, McNeil A, Bradley RS, Withers PJ,Preziosi RF (2012) A minute fossil phoretic mite recovered byphase-contrast X-ray computed tomography. Biol Lett 8:457–460
Dunlop JA, Penney D, Jekel D (2013) A summary list of fossil spidersand their relatives. In Platnick NI (ed) The world spider catalog,version 13.5 American Museum of Natural History. http://research.amnh.org/entomology/spiders/catalog/index.html. doi:10.5531/db.iz.0001
Eichmann F (2002) Palaosymbiosen im Bernstein. ArbeitskreisPalaont, Hannover 30:1–28
Eidelberg MM (2000) Three new species of carabidophilic mites of thefamily Antennoseiidae (Parasitiformes, Gamasina). Entomol Rev80:704–709
Faasch H (1967) Beitrag zur Biologie der einheimischen UropodidenUroobovella marginata (C. L. Koch 1839) und Uropodaorbicularis (O. F. Müller 1776) und experimentelle Analyse ihresPhoresieverhaltens. Zool Jb Syst 94:521–608
Faasch H, Schaller F (1966) Das Phoresie-Verhalten der MilbenUroobovella marginata Koch 1839 und Uropoda orbicularisMüller (Acari, Uropodina). Zool Anz 176:176–182
Hunter PE (1970) New Rhinoseius species (Mesostigmata: Ascidae)from Costa Rican hummingbirds. J Georgia Entomol Soc 7:26–35
Hunter PE, Rosario RMT (1988) Associations of Mesostigmata withother arthropods. Ann Rev Entomol 33:393–417
Karg W (1989) Uropodina Kramer, Schildkrötenmilben. TierweltDeutschlands 67:202
Khaustov AA, Poinar G Jr (2010) Protoresinacarus brevipedis gen. n.,sp. n. from early Cretaceous Burmese amber: the first fossil recordof mites of the family Resinacaridae (Acari: Heterostigmata:Pyemotoidea). Hist Biol 23:219–222
Kofler A, Schmölzer K (2000) Zur Kenntnis phoretischer Milben undihre Tragwirte in Österreich (Acarina: Gamasina, Uropodina). Bernat-med Verein Innsbruck 87:133–157
Krantz GW, Walter DE (2009) A Manual of Acarology; 3rd edn. TexasTech University Press, Lubbock, p 807
Lindquist EE, Krantz GW, Walter DE (2009) Order Mesostigmata. In:Krantz GW, Walter DE (eds) A manual of Acarology, 3rd edn.Texas Tech University Press, Lubbock, pp 124–231
Magowski WŁ (1994) Discovery of the first representative of the mitesubcohort Heterostigmata (Arachnida: Acari) in the MesozoicSiberian amber. Acarologia 35:229–241
Mašán P (2001) Mites of the cohort Uropodina (Acarina, Mesostigmata)in Slovakia. Ann Zool Botan 223:1–320
Moser JC, Roton LM (1971) Mites associated with southern pine barkbeetles in Allen Parish, Louisiana. Can Entomol 103:1775–1798
Perkovsky EE, Rasnitsyn AP, Vlaskin AP, Taraschuk MV (2007) Acomparative analysis of the Baltic and Rovno amber arthropodfaunas: representative samples. African Invert 48:229–245
Poinar G Jr (1982) Sealed in amber. Nat Hist 91(6):26–32Ramsay GW (1960) Sub-fossil mites from the Hutt Valley. Trans R Soc
New Zealand 88:575–576Salmane I, Telnov D (2009) Mesostigmata mites (Acari: Parasitiformes)
associated with beetles (Insecta: Coleoptera) in Latvia. Lat Entomol47:58–70
Vitali F (2006) Taxonomic, biological and evolutionistic notes on theSpondylidinae included in Baltic amber (Coleoptera, Cerambycidae).Entomapeiron (P S) 1(3):29–44
Weitschat W, Wichard W (2010) Baltic amber. In: Penney D (ed)Biodiversity of fossils in amber from the major world deposits.Siri Scientific Press, Manchester, pp 80–115
Wiśniewski J (1982) Für die Fauna Polens neue Uropodina (Acari:Parasitiformes). Teil. II. Fragm Faun 27:143–147
Wiśniewski J, Hirschmann W (1993) Gangsystematik der ParasitiformesTeil 548. Katalog der Ganggattungen, Untergattungen, Gruppenund Arten der Uropodiden der Erde. Acarol Schrift Vergl Milbenk40:1–220
344 Naturwissenschaften (2013) 100:337–344
Recommended

















