
Chemical biology –the biologist’s perspective
Drug development 2007
Per Sunnerhagen

What are the goals of chemical biology?
• To develop chemical tools and applying them to answer questions in cell biology
• The object is not to develop drugs, although resultsobtained with chemical biology may well becomeuseful in drug development

The roles of chemical biology in drug development

Workflow of chemical biology at the Broad Institute, MIT

What arguments can be used for chemical, ratherthan genetic, intervention as a tool for investigationsin cell biology?
1) Permanent shut-off (or overexpression) of a gene will cause secondaryadaptations in the mutant cell, leading to changes in e.g. expression levels of other genes. By contrast, short-term inhibition using a chemical will only produce primary effects
3) Conditional mutants (e.g. temperature-sensitive alleles) require conditions (e.g. temperature shift) that cause major changes in cell state,making appropriate controls difficult to evaluate
2) Certain cellular pathways and function (e.g. cytoskeleton, translation)have a large proportion of essential genes, making genetic analysis difficult.Chemical inhibitors open two possibilities for analysis of such pathways:a) Partial continuous inhibitionb) Complete but transient inhibition,in both cases preserving viability

4) Continuous variability - by applying different amounts of inhibitor, it ispossible to get a graded response from an enzyme or a pathway.
This is essential for a systems biology analysis of a pathway, with theintention of identifying the components, the activity of which are criticalfor the overall output of the pathway.Such components are potential drug targets.
5) Speed – it is possible to shut off the activity of a target (protein) withinseconds – a few minutes using small molecules as inhibitors.Cf: transcriptional shut-off 30 – 60 minutes; RNAi hours – days.Necessary to follow cellular events with fast kinetics – signal transduction,cell cycle etc.
6) Selective inhibition of the interaction between a specific protein pair,targeting one interaction domain, leaving other interactions intact.This allows shutdown of only one branch of a pathway
cont’d
7) Chemicals acting as fluorescent tags and e.g. ”caged substrates” can be developed, adding further versatility

What about drawbacks?
• side effects (small molecules are never entirely specific; but gene disruptions are)
• limited bioavailability: penetrating different cell types from different organismswill vary; a small molecule developed will to some degree be species-specific

Protein kinases constitute a large protein family
The selectivity of kinase inhibitors can be evaluated usingpanels of immobilised recombinant kinase enzyme

”Orthogonal chemical genetics” compared with ordinary genetics

Selective inhibition ofa genetically engineeredprotein kinase allele usinga designed ”orthogonal”inhibitor that can be accommodated only bythe modified kinase.All other ”wild-type”kinases are unaffected

Analogue-sensitive kinase alleles can be used for fingerprinting theresponse (e.g. transcript profile) to inhibition of a kinase. This can beutilised in the search for new wild-type kinase inhibitor lead compounds
Analog sensitive kinase allele (ASKA)-based gene expression ‘blueprints’ for kinase inhibitor lead profiling. Using the ASKA system, changes in gene expression resulting from potent, specific kinase inhibition can be measured by DNA microarrays. The open circles represent cells with either ASKA or wildtype kinases; the compounds are either the ASKA-specific orthogonal inhibitor or a lead compound of a wildtype kinase (lead). The checkerboards represent gene expression patterns. The far-left treatment shows the pattern that results from inhibition of the ASKA. The middle two treatments show the small changes that result from either the ASKA or the analog inhibitor that can be ‘subtracted’ from the ASKA + inhibitor pattern. The far-right treatment shows the pattern obtained by profiling a lead compound. By comparing gene expression patterns, such ASKA-based blueprints can be used to determine the possible off-target effects of kinase inhibitor lead compounds. In the example shown here, the lead compound gives the same profile as the ASKA blueprint; thus, the lead has a similar potency and specificity to the ASKA.

Analogue-sensitive kinase allele (ASKA) mice can be used either directly (in various chemically
induced disease models) or by mating to appropriate disease model strains. Specific kinase inhibition can
be achieved by injection of an orthogonal inhibitor that inhibits only the ASKA and not any wildtype
kinases.
Use of analogue-sensitive kinase alleles in mice

Identification of protein kinase substrates using orthogonal mutations andATP analogues
Only the genetically modifiedprotein kinase – in this caseSrc – will be able to accept thebulky ATP analogues, which areradiolabeled
As a result, only the direct substrates of Src will beradiolabeled, allowing theiridentification

Second generation ATP analogues and orthogonal mutationswould be specific (excluding normal ATP from reaction)

Chemical Biology at Göteborg University

Current focus of the Chemical Biology Platform
Can we create several independent orthogonal pairs of sensitive kinase alleles and inhibitors?
This would allow simultaneous and independent manipulations in vivo of two or more protein kinases in a signalling pathway –approaching true systems biology analysis

A chemical genetics approach to direct proteolysis to selected targets in vivo (”chemical knockout”)
A heterofunctional synthetic molecule (PROTAC) includes ligand functionality for the target protein (white triangle), a linker moiety (grey hourglass shape) and a ligand for E3 ligase (black rectangle)

Schematic representation of orthogonal chemical genetic strategy for site specifically incorporating unnatural aminoacids. An orthogonal synthetase is evolved that specifically acylates a cognate orthogonal suppressor tRNA with a desiredunnatural amino acid. To introduce an unnatural amino acid site specifically in a target protein, an appropriate codon is mutated to an amber, opal or four base codon and transfected in the host cells. The orthogonal tRNA/synthetase pair in the host cells responds to the mutated codon by incorporating the desired unnatural amino acid at the site of interest in vivo.
Expansion of the genetic code through incorporation of unnaturalamino acids through chemical genetics

Biological uses of unnatural amino acids incorporated inspecific proteins
• a photo-crosslinking amino acid can be used to probe dynamicprotein-protein interactions at different stages of a signalling cascade;transient enzyme-substrate complexes can be covalently bonded andisolated
• fluorescent amino acids can be used to probe the microenvironmentcreated during the signalling processes (pH, redox etc.)
• amino acids bearing a photo-cleavable side chain provide atemporal switch to regulate activity in vivo using laser
• ”killer” functionalities (e.g. azido groups) can be used to inactivatea target enzyme at a specific time and place in the cell
• heavy atom-containing amino acids for protein structure studies

Incorporation of unnatural UV-crosslinkable amino acids intoprotein can be used to reveal protein-protein interactions,with high resolution in space and time

The bioorthogonal chemical strategy extended
A chemical reporter linked to a substrate is introduced into a target biomolecule through cellular
metabolism. In a second step, the reporter is covalently tagged with an exogenously delivered
probe. Both the chemical reporter and exogenous probe must avoid side reactions with nontarget
biomolecules.

Specific sequences of natural amino acids can work as a specificbase for chemical reporters
• Cys-Cys-X-X-Cys-Cys reacts selectively with biasenicalsThis amino acid sequence can be introduced genetically intoany protein. This works in instances where GFP tagging disruptsfunction of the protein.
HeLa cells expressing tetracysteine-fused connexin
were treated with FlAsH (green), incubated for 4
hours, then treated with ReAsH (red).
This two-color pulse-chase labeling experiment
shows that newly synthesized connexin is
incorporated at the outer edges of existing gap
junctions
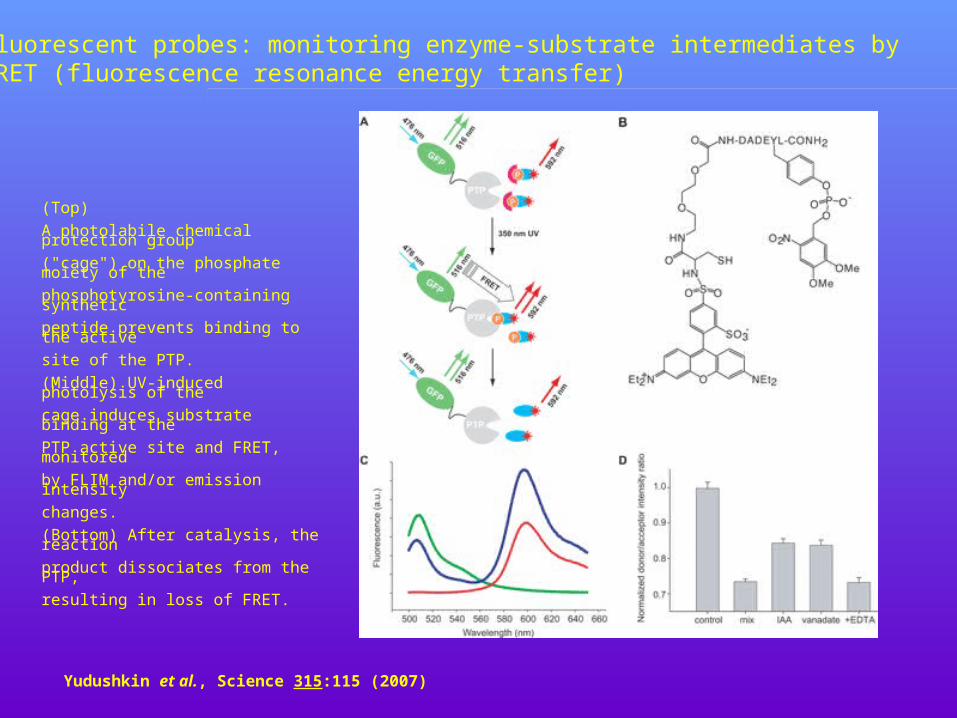
Yudushkin et al., Science 315:115 (2007)
Fluorescent probes: monitoring enzyme-substrate intermediates by FRET (fluorescence resonance energy transfer)
(Top)
A photolabile chemical protection group
("cage") on the phosphate moiety of the
phosphotyrosine-containing synthetic
peptide prevents binding to the active
site of the PTP.
(Middle) UV-induced photolysis of the
cage induces substrate binding at the
PTP active site and FRET, monitored
by FLIM and/or emission intensity
changes.
(Bottom) After catalysis, the reaction
product dissociates from the PTP,
resulting in loss of FRET.

Yudushkin et al., Science 315:115 (2007)
“A map of Km/S for the enzyme (PTP1B)reveals that the peripheral pool, near theplasma membrane, of PTP1B operates in a near-saturation regime (i.e., low Km/S)”

Combining genetics and chemical screening
A compound that causes cell death on its own can be used in agenetic suppressor screen that identifies cDNAs (or siRNAs) that prevent death caused by the compound. Such suppressor reagentsare likely to encode proteins involved in the response to the compound, including the direct protein target of the compound

Mutants
Chem
icals
Parsons et al., Cell 126:611 (2006)
Classification of crude extracts from nature containing bioactive compounds
Clustering of mutants and chemicals according to sensitivity profiles groups compoundswith similar mechanisms of action
(Data are cleaned through eliminating mutants in membranepumps and membrane lipid metabolism, which givemultidrug sensitivity)
DNA-damaging compounds
Crude extract clusters together with pure chemicals
Synthesis of ergosterol (membrane lipid)
Cell wall (staurosporin inhibits PKC;caspofungin inhibits synthesis of -glucan)

Factoring analysis finds correlations between compoundsbased on responses in only a small cluster of genes
Actinomycin, aDNA-damagingcompound, which alsoinhibits RNA polymerase, is groupedhere together withother DNA-damagingcompounds
Verrucarin and neomycinboth inhibittranslation.They are grouped together in afactoringanalysis butnot with hierarcical clustering.Mutants inthis group affect the40S sub-unit ofribosomes

Active compound of extract 00-192
Active compoundof extract 00-132
High correlationbetween sensitivity profiles forextracts 00-192 and 00-132
Spontaneouslymutated strains co-resistant toboth chemicals
Recommended

















