
CHARACTERIZATION AND MODULATION OF ELECTRICAL
COUPLING BETWEEN MOLLUSCAN NEUROENDOCRINE CELLS
by
Zahra Dargaei
A thesis submitted to the Physiology Graduate Program in the
Department of Biomedical and Molecular Sciences
In conformity with the requirements for
the degree of Master of Science
Queen’s University
Kingston, Ontario, Canada
July, 2014
Copyright © Zahra Dargaei 2014

ii
Abstract
The bag cell neurons of Aplysia initiate reproductive behaviour by secreting egg-laying
hormone during a prolonged period of synchronous and repetitive firing known as the
afterdischarge. Electrical coupling facilitate the functional syncytium between bag cell neurons.
We used dual tight-seal whole-cell recording to examine the biophysical properties and
modulation of electrical synapses between pairs of cultured bag cell neurons. Transjunctional
voltage did not influence junctional conductance. Presynaptically evoked action potentials
transferred through gap junctions to elicit postsynaptic electrotonic potentials (ETPs). Bag cell
neurons did not show dye coupling; hence, the permeability of electrical synapses was tested
using block of K+ current. Presynaptic neurons were whole-cell loaded with Cs
+ or TEA and
transfer measured by suppression of postsynaptic K+ current. Cs
+, but not TEA, reduced the K
+
current in electrically-coupled neighbours. In addition, the gap junction blockers meclofenamic
acid, niflumic acid, and nitrobenzoic acid, but not glycyrrhetinic acid and quinine, considerably
attenuated junctional current.
The afterdischarge is accompanied by an increase in intracellular Ca2+
and upregulation
of PKC. Elevating Ca2+
with a train of voltage steps, which mimics the onset of the
afterdischarge, decreased junctional conductance. The inhibition was most effective when Ca2+
entry occurred in both electrically-coupled neurons. Depletion of Ca2+
from the mitochondria,
but not the endoplasmic reticulum, also attenuated junctional communication. Buffering Ca2+
with high intracellular EGTA prevented uncoupling, as did inhibition of CaM kinase.
Application of a PKC activator modestly decreased junctional current, while elevating Ca2+
along with PKC inhibited electrical synapses to an even greater extent than Ca2+
alone. Our

iii
results suggest that PKC and Ca2+
-dependent activation of CaM kinase inhibit electrical
signalling. This may contribute to an enhancement of neuronal excitability leading to the
secretion of reproductive hormone. Ca2+
influx also decreased the amplitude and time course of
the ETP, which may improve synchrony by preventing lengthy ETPs and out-of-phase
postsynaptic action potentials. Thus, modulation of electrical coupling may act as a means to
enhance and sustain the release of reproductive hormone.

iv
Co-Authorship
The following thesis expands on preliminary discoveries made in our laboratory by Dr.
Neil Magoski and Phillip Colmers (undergraduate project student) regarding the basic
electrophysiological properties of bag cell neuron electrical synapses. Dr. Magoski provided me
with training in dual whole-cell current- and voltage-clamp recording. Dr. Magoski and I
discussed on a regular basis my results and observations, experimental procedures, and relevant
literature.
I performed all data recording and analyses, with the exception of sharp-electrode
intracellular biocytin staining (Figure 8C), which was done by Ms. Heather Hodgson, and the
ensemble, extracellular recording from the intact bag cell neuron cluster (Figure 18B), which
was performed by Dr. Magoski. I prepared most of my own solutions and drugs. However, Ms.
Hodgson also provided assistance in some solution preparation. In addition, Ms. Hodgson
provided animal care services. Although I prepared the large majority of the cell cultures that I
used, some pairs of neurons were prepared by Dr. Magoski and fellow lab members. Animal
dissections and removal of the abdominal ganglia were shared between myself and other
members of the lab.
I prepared this thesis and all of its contents with necessary editing, proofreading, and
constructive criticism from Dr. Magoski. This work will be summarized in two manuscripts.
The main research paper will be based entirely on data presented here and written together with
Dr. Magoski.

v
Acknowledgements
It is difficult to overstate my gratitude to my wonderful supervisor, Dr. Magoski. I would
like to thank him for his support, inspiration, patience, continuous encouragement, attention to
details, and his great efforts to explain things clearly and simply. I feel incredibly privileged to
have him as my supervisor. The joy and enthusiasm he has for his research was contagious and
motivational for me.
I would like to thank Heather Hodgson for being a great source of positive energy,
Christopher Groten for always willing to answer my questions, Michel Sturgeon for being so
kind and helpful, Christopher Beekharry, and Tony Zhu for being friendly. Thank you all for
creating a productive and fun atmosphere in the lab.
I would also like to give a special thanks to my husband, Reza, for his endless love,
always believing in me and encouraging me to follow my dreams and my parents for their
constant support during my entire education.

vi
Table of Contents
Abstract ........................................................................................................................................... ii
Co-Authorship................................................................................................................................ iv
Acknowledgements ......................................................................................................................... v
List of Figures .............................................................................................................................. viii
List of Abbreviations ...................................................................................................................... x
Chapter 1 Introduction .................................................................................................................... 1
Neuroendocrine systems and synchronized patterns of activity........................................ 1
Electrical coupling via gap junctions ................................................................................ 2
Biophysical properties of gap junction channels ............................................................... 5
Gap junction modulation ................................................................................................... 9
The bag cell neurons of Aplysia ...................................................................................... 12
Electrical coupling between bag cell neurons ................................................................. 15
Hypothesis and Significance ........................................................................................... 18
Chapter 2 Materials and Methods ................................................................................................. 20
Animals and cell culture .................................................................................................. 20
Whole-cell voltage- and current-clamp recordings from cultured bag cell neurons ....... 21
Intracellular dye staining and fluorescence microscopy of cultured bag cell neurons .... 23
Ensemble, extracellular recording from the intact bag cell neuron cluster ..................... 25
Drug application and reagents ......................................................................................... 26
Analysis ........................................................................................................................... 27
Chapter 3 Results .......................................................................................................................... 29
Bag cell neuron electrical synapses can be recapitulated in primary culture .................. 29

vii
Electrical coupling between cultured bag cell neurons is voltage-independent .............. 31
Bag cell neurons present large coupling coefficients ...................................................... 34
Coupling between bag cell neurons is electrotonic rather than chemical ....................... 36
Bag cell neuron gap junctions are not dye-permeable .................................................... 38
Bag cell neuron gap junctions selectively pass solutes ................................................... 40
Select gap junction blockers inhibit cultured bag cell neuron electrical synapses .......... 42
The afterdischarge is abated by gap junction blockers .................................................... 48
Increased intracellular Ca2+
inhibits electrical synapses .................................................. 49
Synchronous stimulation is most effective at inhibiting electrical coupling ................... 51
Depletion of mitochondrial Ca2+
markedly attenuates junctional current ....................... 51
Sustained inhibition of electrical synapses depends on intracellular Ca2+
...................... 53
CaM kinase is involved in Ca2+
-induced uncoupling ...................................................... 56
Activation of PKC reduces the junctional current ........................................................... 58
Elevation of intracellular Ca2+
decreases coupling coefficient ....................................... 58
Intracellular Ca2+
rise decreases the amplitude and time course of ETPs ....................... 60
Chapter 4 Discussion .................................................................................................................... 64
Future Directions .......................................................................................................................... 76
Literature cited .............................................................................................................................. 79

viii
List of Figures
Figure 1. Gap junctions provide a communication pathway between adjoining cells. ....... 3
Figure 2. The bag cell neurons of Aplysia californica control egg-laying behavior. ....... 13
Figure 3. Electrical coupling is present between bag cell neurons in intact cluster. ........ 16
Figure 4. Cultured pairs of bag cell neurons show electrical coupling. ............................ 30
Figure 5. Electrical synapses between bag cell neurons are voltage-insensitive. ............. 32
Figure 6. Bag cell neurons can be highly coupled. ........................................................... 35
Figure 7. Electrotonic transmission is not interrupted when Ca2+
is absent from the
extracellular solution. ........................................................................................................ 37
Figure 8. Bag cell neuron electrical synapses show no dye-coupling. ............................. 39
Figure 9. K+ channel blockers reduce outward current in single bag cell neurons. .......... 41
Figure 10. Cs+, but not TEA, passes through bag cell neuron gap junctions. ................... 43
Figure 11. Gap junction blockers inhibit bag cell neuron electrical synapses. ................. 44
Figure 12. A combination of NFA and NPPB blocks bag cell neuron gap junctions....... 46
Figure 13. Electrical coupling and afterdischrge are inhibited by blockers. .................... 47
Figure 14. Elevation of intracellular Ca2+
decreases junctional current. .......................... 50
Figure 15. Inhibition of junctional current is most effective when Ca2+
entry occurs in
both neurons. ..................................................................................................................... 52
Figure 16. Depletion of mitochondria Ca2+
, but not endoplasmic reticulum Ca2+
, inhibits
junctional communication. ................................................................................................ 54
Figure 17. Buffering Ca2+
with high intracellular EGTA prevents uncoupling. .............. 55

ix
Figure 18. CaM kinase inhibition prevents the Ca2+
-dependent decrease in electrical
coupling............................................................................................................................. 57
Figure 19. PKC activation inhibits gap junctions and synergizes with Ca2+
influx. ........ 59
Figure 20. Coupling coefficient decreases after an intracellular Ca2+
rise. ...................... 61
Figure 21. Elevated intracellular Ca2+
reduces the size and time course of the ETP. ....... 62

x
List of Abbreviations
ºC degree Celcius
ANOVA analysis of variance
ATP adenosine triphosphate
ATPase adenosine triphosphatase
C cysteine
cAMP cyclic adenosine monophosphate
Ca2+
calcium ion
CaCl2 calcium chloride
CaM calmodulin
CaMKII calmodulin kinase II
CC current clamp
cfhmASW calcium-free high-magnesium artificial sea water
CICR calcium-induced calcium-release
Cl- chloride ion
cm centimeter
CPA cyclopiazonic acid
CO2 carbon dioxide
Cs+ cesium ion
Cx connexin
d day
DAG diacylglycerol
DMSO dimethyl sulfoxide
EGTA ethyleneglycol bis (aminoethylether) tetraacetic acid
ELH egg-laying hormone
ETP electrotonic potential
FCCP carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone
g gram
GTP guanosine triphosphate
HEPES N-2-hydroxyethlpiperazine-N’-2-ethanesulphonic acid
hr hour
H+ hydrogen ion
Hz hertz (1/s)
IP3 inositol 1,4,5-triphosphate
K+ potassium ion
KCl potassium chloride
KDa kilo daltons
kHz kilo hertz
KN-62 1-N-O-bis (5-isoquinolinesulfonyl)-N-methyl-1-4- phenyl-piperazine
KOH potassium hydroxide
l liter
M molar
MΩ megaohms

xi
McFA meclofenamic acid
Mg2+
magnesium ion
mg milligram
MgCl2 magnesium chloride
min minute
ml milliliter
mM millimolar
mm millimeter
ms millisecond
mV millivolt
MW molecular weight
n sample number
Na+ sodium ion
nA nanoamperes
nASW normal artificial salt water
NaCl sodium chloride
NaOH sodium hydroxide
NFA niflumic acid
nM nanomolar
nm nanometer
NMDG N-methyl- D-aspartic acid D-glucamine
NPPB nitrobenzoic acid
nS nano siemens
pA picoampere
pH negative logarithm of hydrogen ion concentration
PKA protein kinase A
PKC protein kinase C
PMA phorbol 12-myristate 13-acetate
sec second
TEA tetraethylammonium
tcASW tissue culture artificial salt water
TMA tetramethylammonium
VC voltage clamp
µM micromolar
µl microlitre
W watt

1
Chapter 1
Introduction
Neuroendocrine systems and synchronized patterns of activity
In neuroendocrine systems, hormone secretion evoked by action potentials can
control or influence many fundamental physiological behaviors, such as sleeping
(Steriade 2005), food intake (Kalra et al. 2003a), lactation (Lincoln and Wakerley 1974),
parturition (Summerlee 1981), and ovulation (Wayne 2001; Kalra et al. 2003b). Often,
these behaviors require the activation of a synchronized network of neurons that
generates regular or rhythmic firing. Such patterns have been identified in both the
vertebrate and invertebrate neurons system, where rapid communication between distinct
neurons or neuronal groups results in rhythmic, alternating, or even multiphasic output.
For example, in the pulmonate pond snail, Lymnaea stagnalis, neurosecretory
caudo-dorsal cells release ovulation hormone through a synchronous firing of action
potentials (De Vlieger et al. 1980). Similarly, five pairs of neurosecretory cells in the
subesophageal ganglion of the silkmoth, Bombyx mori, discharge action potentials in
synchrony to release a pheromone known as biosynthesis-activating neuropeptide
(Ichikawa 2002).
Periodic bursts of action potentials can also secrete neuropeptides into the
circulatory system to regulate diverse bodily functions. The mammalian hypothalamus
contains populations of neurons that organize fundamental behaviors through endocrine
or neuroendocrine function (Bear et al. 2007). The secretion of oxytocin during

2
parturition (Summerlee 1981) and milk injection during lactation (Lincoln and Wakerley
1974) from magnocellular neurosecretory cells in the supraoptic and paraventricular
nuclei of the hypothalamus have been the primary models of peptide release during a
synchronous discharge (Belin and Moos 1986). In addition, a different group of
hypothalamic neurons secretes gonadotropin-releasing hormone in an episodic manner,
which results in the release of luteinizing hormone and follicle-stimulating hormone to
control ovulation (Kalra et al. 2003b).
Electrical coupling via gap junctions
Neuroendocrine cell populations often require synchronous action potential firing
for en mass neuropeptide release (Sandoz and Dreifuss 1978; Ichikawa et al. 1999).
Synchrony can occur if neurons communicate with each other at specialized junctions
called electrical synapses. The most common type of synaptic transmission is chemical,
when one neuron releases a neurotransmitter, which is detected by an adjacent cell (Katz
and Miledi 1967; Jessell and Kandel 1993). However, a second and perhaps more
efficient means to transmit signals is through gap junctions (Bennett 1977; Bennett and
Zukin 2004). Gap junctions provide a communication pathway between the cytoplasm of
adjoining cells, and allow the transfer of ions or small molecules, such as metabolites and
signalling molecules (Figure 1A). They form the basis of an electrical synapse, where
current flows rapidly and directly between neurons, and thus is able to either promote
synchronized activity or reliably transfer information (Bennett and Goodenough 1978;
Bennett and Zukin 2004).
Using transmission electron microscopy, the morphology of gap junctions was

3
Figure 1. Gap junctions provide a communication pathway between adjoining cells.
A) Two adjacent cells connected via a gap junction array. The channels pass not only
ions, but potentially nutrients, metabolites and second messengers.
B) Structure of a gap junction. Two connexons from each adjacent cell create a single gap
junction. Each connexon is composed of six connexin protein subunits that are oriented
perpendicular to the membrane to form a central pore. Inset, a single connexin has four
transmembrane domains with two extracellular loops, containing highly conserved
cysteine residues (C) and one cytoplasmic loop, the amino and carboxyl termini are
exposed to the cytoplasm. The cysteines form di-sulfide bridges to dock to the opposite
connexin.

4
first described in the giant fibers of crayfish and squid as structures between two, closely-
opposed cell membranes with a potential to transmit electrical signals (Robertson 1953).
Shortly thereafter, electrical coupling was identified physiologically in the giant nerve of
crayfish (Furshpan and Potter 1959) and the Mauthner neurons of goldfish (Robertson et
al. 1963). Subsequently, gap junctions have been found to play a variety of roles in many
different cell types. In the heart, for example, synchronized contractions of cardiac
myocytes via gap junctions leads to rhythmic pumping, as well as the electrical
syncytium through which action potentials propagate (Rohr 2004). Gap junctions in the
respiratory epithelium coordinate the unidirectional motion of beating cilia (Evans and
Martin 2002). In the liver, hepatocytes are interconnected by gap junctions that allow for
the movement of metabolites and second messengers (Dermietzel and Spray 1993).
Fibroblast networks found in tissues such as skin, intestine, and kidney, are also formed
by gap junctions (de Roos et al. 1996; Langevin et al. 2004; Zeng et al. 2009).
Not surprisingly, gap junctions play important roles in physiology and
development, as various genetic-based diseases are associated with their mutation. The
loss or mutation of gap junction genes causes certain forms of hearing deficits, female
infertility, cataracts, ventricular arrhythmias, and embryonic lethality (Kelsell et al. 1997;
Peters et al. 1997; Xia et al. 1998; White 2002; Dermietzel and Spray 2013).
Gap junctions are critical for neuronal output in both the vertebrate and
invertebrate nervous system. For example, pharyngeal contraction in the marine mollusc,
Navanax (Levitan et al. 1970), swimming in the jellyfish, Polyorchis (Anderson 1979),
periodic waves in the olfactory lobe of the garden slug, Limax (Ermentrout et al. 2004),

5
as well as synchronous spiking activity in mouse suprachiasmatic nucleus (Long et al.
2004) and hypothalamus (Vazquez-Martinez et al. 2001), are controlled by gap junction-
connected neurons.
Biophysical properties of gap junction channels
A single gap junction channel is composed of twelve protein subunits termed
connexins (in vertebrates) and innexins (in invertebrates) (Figure 1B). Six connexins or
innexins form a hemichannel (connexon or innexon). Hemichannels composed of a single
connexin isoform are called homomeric, while those with at least two different connexin
isoforms are heteromeric. Two connexons, from adjacent cells, create a gap junction,
while multiple gap junctions, also known as an array, form an electrical synapse (Bennett
1977; Bennett and Goodenough 1978; Bennett and Zukin 2004) The connexin protein has
four transmembrane domains (M1-M4) with two extracellular loops, one cytoplasmic
loop, and the N- and C-termini exposed to the cytoplasm (Figure 1B, inset). The regions
between the transmembrane domains M2 and M3, as well as the C-terminus are
important for connexin regulation (Kumar and Gilula 1996). Phosphorylation of serine
and tyrosine residues in the C-terminal has been shown to control gap junction assembly
(Solan and Lampe 2005). Two to three cysteine residues found in each extracellular loop
appear to be responsible for the docking of one connexin to another through the
formation of di-sulfide bridges (Goodenough et al. 1996). A third family of gap junction-
like channels, the pannexins, has been described with some amino acid similarities to
innexins, but are present in both vertebrates and invertebrates (Panchina et al. 2000).

6
Pannexins almost exclusively form channels to the extracellular space, rather than
between cells (Mammano 2000; Dermietzel and Spray 2013).
The family of connexin (Cx) genes comprises at least 21 members in the human
genome (Sohl and Willecke 2004), while in Caenorhabditis elegans, 24 innexin genes are
expressed either in the CNS or peripherally (Phelan and Starich 2001) and comparative
sequence analysis of innexins from the leech genome provides evidence for 21 innexin
genes (Kandarian et al. 2012). Connexin proteins are named according to their estimated
molecular mass, e.g., Cx36 is the connexin protein of 36 kDa weight (Bennett et al.
1991). The pores they form are typically permeable to molecules of up to 1 kDa (Spray
and Bennett 1985); however, depending on spatial patterns of expression, the
combination of connexin expression, as well as the charge of the solutes, different
properties in permeability, conductance and gating can be achieved (Račkauskas et al.
2010). For example, Cx40 and Cx26 show higher permeability to cationic than to anionic
probes with a similar molecular weight, while Cx43 and Cx45 tend to pass solutes based
on their size without significant charge selectivity (Kanaporis et al. 2011), while, in the
vertebrate heart, the junctional conductance of mouse Cx30.2 and human Cx31.9
hemichannels is higher than Cx43 and Cx45, which has been suggested to influence
electrical propagation between cardiac myocytes (Bukauskas et al. 2006). The gap
junction conductance properties in Purkinje fibers and ventricular muscle depend on the
pattern of Cx40 and Cx43 mRNA expression (Kanter et al. 1993). Genetic replacement
of Cx45 with Cx36 results in functional compensation in retina, but not in the heart
(Frank et al. 2010).

7
Although, the properties and function of connexin-based gap junctions have been
studied extensively, innexins have been examined to a lesser degree. For example, the
expression of innexin, UNC-9, in body-wall muscle cells of C. elegans plays an
important role in locomotion; mutant UNC-9 inhibits junctional conductance and reduces
locomotor activity (Liu et al. 2006). In Drosophila, mutant innexin-2 results in improper
embryonic morphogenesis (Bauer et al. 2004), while in C.elegans an innexin-3 mutation
causes inappropriate hypodermal cell contacts during development (Starich et al. 2003).
Measurement of junctional current (the current that passes through gap junctions)
is a common approach to study electrical synapses. This can be achieved by dual voltage-
clamp, where the membrane potential of one cell is clamped and a command pulse to the
second cell causes transjunctional current corresponding to the current flowing through
gap junctions. The driving-force voltage and the junctional current measured in the
passive cell can be used to calculate the junctional conductance (Bennett 1966).
In addition to passively allow ionic current and solutes to flow between cells,
some gap junctions are dependent on transjunctional voltage and/or show rectification
(when current only flows one way). This was first observed in the landmark study of
electrical transmission in crayfish between giant pre- and postsynaptic neurons (Furshpan
and Potter 1959). Specifically, junctional conductance was found to be highly related to
transjunctional voltage, where presynaptic depolarization or postsynaptic
hyperpolarization increases the strength of the electrical synapse. Since then, voltage
sensitivity has been reported in other tissues, such as the giant fibers of the hatchetfish,
Gasteropelecus (Auerbach and Bennett 1969), early embryonic cells of the amphibians,

8
Ambystoma mexicanum, Xenopus laevis, and Rana pipiens (Spray et al. 1981),
blastomeres of the killifish, Fundulus (Spray et al. 1984), salivary glands of Drosophila
(Verselis et al. 1991), cell lines expressing human Cx32, Cx37, and Cx43 (Reed et al.
1993; Revilla et al. 1999), and in lumbar spinal motor neurons of neonatal rat (Chang et
al. 1999). However, in earthworm septum (Verselis and Brink 1984), ventricular
myocytes of mammalian heart, (White et al. 1985; Wang et al. 1992), or mouse
pancreatic β-cells (Perez-Armendariz et al. 1991), electrical synapses are insensitive to
transjunctional voltage.
Gap junction-mediated coupling can also be examined by testing for dye-coupling
between electrically-connected cells (Stewart 1978). For example, injection of the
fluorescent dye, Lucifer yellow, into rat hypothalamic magnocellular neurosecretory cells
or neocortical neurons showed dye coupling (Andrew et al. 1981). Similarly, neurobiotin
diffuses from injected to adjacent cells within neocortex of rat (Peinado et al. 1993);
however; the extent of dye travel depends on the molecular weight and contact location.
Li et al. (1996) showed that different dyes, including carboxyfluorescein, fura-2, and
fluorescein-dextran display a wide variation in the degree to which they pass between
HeLa cells (~30%, ~10% and <1%, respectively). The morphology of cell-to-cell
connectivity can also affect the extent of dye-coupling; case in point, Ewadinger et al.
(1994) found that the electrotonic distance and the contact area of the electrical synapse
profoundly influence the diffusion of dyes between neurons in Lymnaea.
A number of chemically-distinct molecules that act as gap junction blockers have
proven useful to disrupt electrical coupling. For example, Srinivas et al. (2001) showed

9
that quinine blocks junctional communication between transfected mammalian cells in a
dose- and connexin-dependent manner, with a maximal blocking concentration of ~30
and ~70 μM for Cx36 and Cx50, respectively. Alternatively, glycyrrhetinic acid inhibits
gap junctions in mesenteric small artery smooth muscle, with a maximal effect at 10 μM
(Matchkov et al. 2004). Fenamates (meclofenamic, niflumic and flufenamic acid), best
known as nonsteroidal anti-infammatory drugs (Juszczak and Swiergiel 2009), and also
5-nitro-2-(3-phenylpropylamino) benzoic acid, inhibit Cx50 hemichannels (Srinivas and
Spray 2003). In vivo electrophysiology experiments showed that application of
carbenoxolone and mefloquine within the rat dorsal hippocampus impairs fear memory
and prevents context fear learning (Bissiere et al. 2011).
Gap junction modulation
Gap junctions may pass second messengers, such as cyclic adenosine
monophosphate (cAMP), Ca2+
and inositol 1, 4, 5-triphosphate (IP3) (Loewenstein et al.
1967; Saez et al. 1989), the latter of which can trigger the release of Ca2+
from
intracellular stores (Saez et al. 1989; Krysko et al. 2005). Increased intracellular Ca2+
has
long been implicated in inhibition of gap junctions (Spray and Bennett 1985), which was
first reported by Loewenstein et al. (1967) in salivary glands from the midge,
Chironomus. Subsequently, this phenomenon has been investigated in many non-
neuronal and neuronal cells, either by intracellular injection of Ca2+
(De Mello 1975;
Schirrmacher et al. 1996) or by liberating Ca2+
either with pharmacological agents (Baux
et al. 1978; Dahl and Isenberg 1980; Mustonen et al. 2005) or with cytoplasmic
acidification (Spray and Bennett 1985; Lazrak and Peracchia 1993). For example,

10
junctional communication between dog Purkinje cells substantially decreases after
injection of Ca2+
(De Mello 1975). Similarly, lowering pH by application of CO2 causes a
cytosolic Ca2+
rise and uncouples Novikoff hepatoma cells (Lazrak and Peracchia 1993),
while inhibition of intracellular Ca2+
release in rat gastric mucosal cells decreases the
uncoupling effect of ethanol-induced Ca2+
increase (Mustonen et al. 2005).
The activation of intracellular signal transduction pathways, such as calmodulin
(CaM) (Peracchia et al. 1983) and CaM-kinases (Pereda et al. 1998) are likely involved
in Ca2+
-dependent inhibition of gap junctions. Application of trifluoperazine, a CaM
antagonist, to Rana and Xenopus embryos, prevents the uncoupling effect of CO2-
induced Ca2+
increase (Peracchia et al. 1983). Similarly, lowering CaM mRNA with
antisense oligonucleotides in Xenopus oocytes expressing Cx38 reduces the inhibitory
effect of released intracellular Ca2+
(Peracchia et al. 1996). Also, W7, another CaM
blocker, decreases the uncoupling effect of Ca2+
in ventricular myocytes isolated from
guinea pig heart (Toyama et al. 1994). Conversely, in Mauthner neurons, CaM-kinase II
increases the junctional current (Pereda et al. 1998).
In addition to releasing Ca2+
, the production of IP3 is accompanied by the
liberation of diacylglycerol (DAG) which in turn activates protein kinase C (PKC)
(Newton 2010). PKC largely inhibits gap junctions in many cell types. For example, the
PKC activator, phorbol 12-myristate 13-acetate (PMA), results in phosphorylation of
Cx43 and suppresses junctional conductance in rat epithelial and some fibroblastic cells
(Lampe et al. 2000; Cruciani et al. 2001). Similarly, in rat hippocampus astrocytes,
activation of PKC weakens Lucifer yellow, but not neurobiotin, transfer to neighboring

11
cells (Konietzko and Müller 1994), while exposure of pancreatic acinar cells to the PKC
inhibitor, polymyxin B, lowers diacyglycerol-induced uncoupling (Somogyi et al. 1989).
It is also suggested that PKC is involved in the both acetylcholine- and serotonin-induced
uncoupling in rat lacrimal glands and neocortex, respectively (Randriamampita et al.
1988; Rörig and Sutor 1996). Alternatively, the phosphorylation of mouse Cx45 (van
Veen et al. 2000) and gap junctions in rat cardiac myocytes (Spray and Burt 1990) by
PKC elevates junctional conductance.
cAMP and cAMP-dependent protein kinase A (PKA) pathway are also typically
reported to decrease junctional communication. Intracellular injection of cAMP into
HeLa cells expressing Cx40, Cx43, or Cx26 showed that cAMP is rapidly delivered to
neighboring cells, with Cx43 gap junctions being most permeable (Kanaporis et al.
2008). Similarly, cAMP causes a slight decrease in electrical coupling between canine
Purkinje cells (De Mello 1984). Recording from electrically-coupled hippocampal
neurons shows that the activation of β- adrenergic receptors reduces electrical synapses
through the cAMP/PKA pathway (Zsiros and Maccaferri 2008). To a lesser degree, there
is evidence suggesting that cAMP/PKA enhances junctional communication. For
example, PKA controls endothelium-derived hyperpolarizing factor-type responses in
smooth muscle cells by strengthening electrical coupling (Griffith et al. 2002). Also, in
isolated hepatocytes, cAMP increases junctional communication (Saez et al. 1986).
The present study aims to characterize the biophysical properties and examine the
role of second messengers in regulating electrical synapses between neuroendocrine cells
from the marine mollusc, Aplysia californica.

12
The bag cell neurons of Aplysia
The nervous system of Aplysia contains peripheral sensory structures (such as
rhinophores, tentacles, lips, and eyes) and nine major ganglia: the paired buccal, pedal,
pleural, and cerebral ganglia as well as a single abdominal ganglion (Figure 2A) (Kandel
et al. 1980). Two clusters of 200-400 neuroendocrine cells, known as the bag cell
neurons, are found at the rostral end of the abdominal ganglion. These neurons initiate
egg-laying behavior by secreting egg-laying hormone (ELH), a 36-amino acid peptide,
into the blood stream during a prolonged period of synchronous and repetitive firing
called the afterdischarge (Figure 2B) (Kupfermann 1967; Arch 1972; Kaczmarek et al.
1978; Rothman et al. 1983; Conn and Kaczmarek 1989).
The afterdischarge is induced by brief peptidergic and cholinergic afferent input
descending from the pleural ganglia (Brown et al. 1989; White and Magoski 2012); most
prominently, acetylcholine gates an ionotropic receptor that strongly depolarizes the bag
cell neurons. The response presents as a fast phase of high-frequency firing (~5 Hz for ~1
min), followed by a slow phase of low-frequency firing (~1 Hz for ~30 min), and ends
with a prolonged refractory period of ~18 hr, during which further stimulation fails to
trigger long-lasting afterdischarges or neuropeptide secretion (Figure 2C) (Arch and
Smock 1977; Pinsker and Dudek 1977; Kaczmarek et al. 1982). The transition from fast
to slow phase of the afterdischarge as well as refractoriness appears to be mediated by
intracellular Ca2+
(Kaczmarek and Kauer 1983; Magoski et al. 2000).
ELH is secreted into the blood stream at a neural-hemal area in the pleuro-
abdominal connective and acts on nervous system targets as well as peripheral

13
Figure 2. The bag cell neurons of Aplysia californica control egg-laying behavior.
A) Dorsal view of the Aplysia nervous system. Two clusters of bag cell neurons are found
at the anterior of the abdominal ganglion (courtesy of Dr. N.S. Magoski).
B) The animal is typically ~15 cm long and ~5 cm wide. As a part of its reproductive
behavior, it releases a string-like mass of fertilized eggs (modified from Kandel, 1976).
C) A brief stimulation of the synaptic input (stim) to the bag cell neurons induces an
afterdischarge. Extracellular recording from the intact cluster shows a fast and a slow
phase of action potential firing that ends with a refractory period, during which further
stimuli fails to elicit a second afterdischarge. The afterdischarge is truncated for clarity
(courtesy of Dr. N.S. Magoski).

14
organs to produce a series of behaviors including: increased respiratory pumping, reduced
locomotion, suppressed defensive responses, redistribution of blood flow, and head
waving, before culminating in egg deposition behaviors (Arch and Smock 1977; Mayeri
et al. 1979; Stuart et al. 1980; Mackey and Carew 1983; Rothman et al. 1983; Ligman
and Brownell 1985; Goldsmith and Byrne 1993).
The generation of an afterdischarge is accompanied by an increase in intracellular
Ca2+
, due to both influx and release (Fisher et al. 1994; Geiger and Magoski 2008). Ca2+
influx occurs through action potentials triggering voltage-gated Ca2+
channels (Acosta‐
Urquidi and Dudek 1981; Woolum and Strumwasser 1988). During the slow phase, Ca2+
-
induced Ca2+
release (CICR) from the mitochondria and endoplasmic reticulum also
takes place (Geiger and Magoski 2008). Finally, the production of IP3 and activation of
IP3 receptors liberates additional Ca2+
from the endoplasmic reticulum (Fink et al. 1988).
Along with Ca2+
and IP3, other second messengers modulate the excitability of
the bag cell neurons to maintain the afterdischarge. At the beginning of the burst, cAMP
levels rise rapidly (Kaczmarek et al. 1978) and activate PKA to regulate various voltage-
gated ion channels that increase excitability and broaden the action potential (Kaczmarek
et al. 1980; Kaczmarek and Strumwasser 1984; Strong 1984; Strong and Kaczmarek
1986). Application of cAMP analogs, or the adenylate cyclase activator, forskolin,
inhibits both the delayed rectifier and transient K+ current to promote spike broadening.
Increasing cAMP levels in the intact bag cell neuron cluster with dopamine or
methylxanthine, a phosphodiesterase inhibitor (Kaczmarek et al. 1978), as well as adding

15
cAMP analogs to cultured bag cell neurons (Kaczmarek and Strumwasser 1981), can
increase excitability and initiate an afterdischarge-like response.
Another key second messenger controlling bag cell neuron excitability is PKC,
which appears to turn on by two min and stays elevated throughout the afterdischarge
(Conn et al. 1989a; Wayne et al. 1999). PKC potentiates voltage-dependent Ca2+
channels and enhances the Ca2+
-dependent prolonged depolarization (Conn et al. 1989a;
Knox et al. 1992; Tam et al. 2009). Application of phorbol esters to activate PKC
enhances the amplitude of action potentials by increasing Ca2+
current, whereas PKC
inhibitors prevent this effect (Conn et al. 1989b).
PKC is also a regulator of the non-selective cation channel that maintains the
afterdischarge (Wilson and Kaczmarek 1993; Magoski and Kaczmarek 2005). The
activation of this channel requires the association of both CaM and PKC (Lupinsky and
Magoski 2006; Gardam and Magoski 2009). The cation channel-PKC association is
mediated through src homology 3 domain protein-protein interactions (Magoski et al.
2002), and src tyrosine kinase is required for this link (Magoski and Kaczmarek 2005).
The transition to the refractory period appears to depend on the dissociation of PKC from
the channel (Magoski 2004; Magoski and Kaczmarek 2005).
Electrical coupling between bag cell neurons
Bag cell neuron firing is synchronous, which is likely achieved through electrical
coupling within and between the clusters (Figure 3A) (Kupfermann and Kandel 1970;
Blankenship and Haskins 1979; Kaczmarek et al. 1979; Brown et al. 1989). Freeze
fracture studies of the intact cluster indicate the presence of gap junctions between bag

16
Figure 3. Electrical coupling is present between bag cell neurons in intact cluster.
A) A schematic of the abdominal ganglion and bag cell neuron clusters showing
electrical coupling (only a few neurons from each cluster are shown for clarity). They are
electrically-coupled within a cluster and to the neurons in the contralateral cluster.
B) Simultaneous recording from two, randomly selected bag cell neurons in the same
cluster shows that depolarizing the presynaptic neuron, V1 (+I, at bar), elicits a small
electrotonic potential in the postsynaptic cell, V2 (upper traces). Negative current
injection in the presynaptic cell (-I, at bar) induces a hyperpolarization in the pulsed cell
and a small response in the postsynaptic neuron (lower traces) (modified from
Blankenship and Haskin, 1979).
C) Sharp-electrode current-clamp recording of two neurons in the intact bag cell neuron
cluster. Shortly after extracellular stimulation of the synaptic input in the connective (syn
stim), both neurons depolarize and undergo an afterdischarge. Spiking during the
afterdischarge is essentially synchronous, with the initial fast phase followed by the slow
phase. This afterdischarge lasts just over 20 min, but is truncated for display. Scale bars
apply to both traces (courtesy of Dr. N.S. Magoski).

17
cell neuron processes (Kaczmarek et al. 1979). Intracellular injection of Lucifer yellow
into bag cell neuron clusters of A. californica show that dye can move from injected to
adjacent cell(s) (Kaczmarek et al. 1979). Blankenship and Haskin (1979) demonstrated
that stimulating one bag cell neuron in the intact cluster of A. dactylomela induces a very
small electrotonic response in another cell (Figure 3B). They also showed that this
response has a very long latency and prolonged time course, suggesting that coupling is
distant from the somata. Pleural neurons in A. californica are also found to be
electrically-connected, with similar electrophysiological characteristics to bag cell
neurons. Synchronous firing in this pleural cluster can initiate a prolonged burst of action
potentials in the bag cell neurons (Brown et al. 1989).
The best characterized example of electrical coupling in Aplysia is between paired
L14 neurons in the abdominal ganglion (Bodmer et al. 1988). L14 neurons are
motoneurons that elicit inking when animal is disturbed or attacked by a predator (Carew
and Kandel 1977). These gap junctions show voltage-independence as well as non-
rectifying behaviour, i.e., current flows equally from one neuron to the other. This
synapse is also weakly sensitive to intracellular acidification, which has been generally
reported to reduce junctional conductance and coupling in different cells (Spray and
Bennett 1985; Lazrak and Peracchia 1993). Despite the fact that injection of fluorescent
tracer molecules into one neuron often passes to adjacent cells in other preparations
(Andrew et al. 1981; Peinado et al. 1993; Ewadinger et al. 1994), Lucifer yellow, 6-
carboxyfluorescein and dichlorofluorescein (1.2 to 1.4 nm maximal diameter), are not

18
appreciably permeant between L14 neurons; however, tetramethylammonium (TMA) and
tetraethylammonium (TEA) are transferred (Bodmer et al. 1988).
Hypothesis and Significance
The coupling characteristics between neurons in the abdominal ganglion of
Aplysia exhibit some differences compared to electrical synapse in other systems, and it
appears that the study of bag cell neuron gap junctions in situ is hindered by the remote
site of the electrical synapses. However, the permeability, pharmacology, and modulation
of these channels are poorly understood. Thus, I will test the following hypotheses in
electrically-coupled bag cell neurons, largely in primary culture, with the goal of
understanding how coupling may coordinate rhythmic output.
1) Electrical signaling is voltage-independent and size-restrictive.
2) Electrical coupling affects spike synchrony during a bona fide afterdischarge.
3) PKC and Ca2+
suppress electrical coupling.
If only small molecules pass, it would suggest that despite electrical continuity,
bag cell neurons are not a broad biochemical syncytium. Given that even single bag cell
neurons can generate long-term bursts of activity (Hung and Magoski 2007), the
application of gap junction blockers may reveal if blocking electrical coupling will
prevent the afterdischarge or simply result in de-synchronous action potential firing.
During the afterdischarge, cytosolic Ca2+
rises due to voltage-gated influx and
release from intracellular stores (Fisher et al. 1994; Geiger and Magoski 2008). Similarly,
PKC activity is increased shortly after the onset of the afterdischarge (Wayne et al.
1999). If Ca2+
and PKC decrease junctional conductance between bag cell neurons, this

19
may increase excitability by reducing overall leakiness, making the neurons more
responsive to the depolarizing currents driving the afterdicharge. Alternatively, less
coupling may promote precise information transfer by reducing prolonged postsynaptic
responses.

20
Chapter 2
Materials and Methods
Animals and cell culture
Adult Aplysia californica (a hermaphrodite) weighing 150-500 g were obtained
from Marinus (Long Beach, CA, USA), and housed in an ∼300 l aquarium containing
continuously circulating, aerated sea water (Instant Ocean; Aquarium Systems, Mentor,
OH, USA) at 14-16°C on a 12:12-h light/dark cycle, and fed Romaine lettuce five times
per week. All experiments were approved by the Queen’s University Animal Care
Committee (protocols Magoski-100323 or Magoski-100845).
For primary cultures of isolated bag cell neurons, animals were anaesthetized by
an injection of isotonic MgCl2 (50% of body weight), the abdominal ganglion removed
and treated with dispase II (13.3 mg/ml; 165859; Roche Diagnostics, Indianapolis, IN,
USA) dissolved in tissue culture artificial sea water (tcASW) (composition in mM: 460
NaCl, 10.4 KCl, 11 CaCl2, 55 MgCl2, 15 HEPES, 1 mg/ml glucose, 100 U/ml penicillin,
and 0.1 mg/ml streptomycin, pH 7.8 with NaOH) for 18 h at 22°C. The ganglion was
then rinsed in tcASW for 1 h, and the bag cell neuron clusters dissected from their
surrounding connective tissue. Using a fire-polished glass Pasteur pipette and gentle
trituration, dissociated neurons were dispersed in tcASW onto 35 x 10 mm polystyrene
tissue culture dishes (353001; Falcon Becton-Dickinson, Franklin Lakes, NJ, USA). In a
small number of experiments, cells were plated onto glass coverslips (#1; 12-542-B;
Fisher Scientific, Ottawa, ON, Canada) coated with 20 μg/ml poly-L-lysine
hydrobromide, MW=300,000 (P1524; Sigma-Aldrich, Oakville, ON, Canada) and glued

21
with Sylgard (SYLG184; World Precision Instruments, Sarasota, FL, USA) over holes
drilled out of the bottom of the tissue culture dish. Neurons were paired by bringing a
free neuron into contact with a neuron already adhered to the dish. This was achieved by
triturating fluid to either push or pull the free neuron and place it next to the adhered
neuron. Neurons were plated either with their somata (soma-soma) or primary neurites
(neurite-neurite) in contact; in some cases, to act as a control, neurons were separated by
a maximum distance of one soma diameter, without neurites touching. Cultures were
maintained in a 14°C incubator in tcASW and used for experimentation within 2-4 d.
Salts were obtained from Fisher, ICN (Aurora, OH, USA), or Sigma-Aldrich.
Whole-cell voltage- and current-clamp recordings from cultured bag cell neurons
Recordings of membrane potential or current were made from cultured bag cell
neurons using EPC-8 amplifiers (HEKA Electronics; Mahone Bay, NS, Canada), and the
tight-seal, whole-cell method. Microelectrodes were pulled from 1.5 mm external
diameter/1.12 mm internal diameter, borosilicate glass capillaries (TW150F-4; World
Precision Instruments) and had a resistance of 1-3 MΩ when filled with our standard
intracellular saline (composition in mM: 500 K+-aspartate, 70 KCl, 1.25 MgCl2, 10
HEPES, 11 glucose, 10 glutathione, 5 ethylene glycol bis(aminoethyl ether) tetraacetic
acid (EGTA), 5 ATP (grade 2, disodium salt; A3377; Sigma-Aldrich), and 0.1 GTP (type
3, disodium salt; G8877; Sigma-Aldrich); pH 7.3 with KOH). The free Ca2+
concentration was set at 300 nM by adding 3.75 mM CaCl2, as calculated by WebMaxC
(http://www.stanford.edu/cpatton/webmaxcS.htm). Neuronal pairs were usually dialyzed
for 15-30 min under voltage-clamp before experimentation.

22
Recordings were performed in normal ASW (nASW; composition as per tcASW,
but with glucose and antibiotics omitted). The standard intracellular saline had a
calculated liquid junction potential of 15 mV vs. ASW, which was corrected by off-line
subtraction. Pipette junction potentials were nulled immediately before seal formation.
Following membrane rupture, pipette and neuronal capacitive currents were cancelled,
the series resistance (3-5 MΩ) compensated to 70-80%, and monitored throughout the
experiment. Input signals were filtered at 1 kHz (for current) and 5 kHz (for voltage) by
the EPC-8 Bessel filter, and sampled at 2 kHz using an IBM-compatible personal
computer, a Digidata 1322A analogue-to-digital converter (Molecular Devices;
Sunnyvale, CA, USA) and the Clampex acquisition program of pCLAMP 8.1 (Molecular
Devices). Clampex was also used to control the membrane potential under voltage-clamp
and inject current steps under current-clamp; in addition, neurons were also manually
current-clamped to set membrane potentials by delivering constant current with the EPC-
8 V-hold.
In a small number of experiments, voltage-gated K+ current was recorded. In
single cells, neurons were voltage-clamped at -40 mV, and a voltage step to +30 mV for
200 ms elicited K+ current. In pairs, K
+ current was examined only in neuron 2 by
holding neuron 1 continuously at -40 mV and delivering a 200-ms step in neuron 2 to
+30 mV. In these cases, a Na+-free/Ca
2+-free ASW (composition in mM: 471 NMDG,
10.4 KCl, 66 MgCl2, 15 HEPES; pH 7.8 with KOH) was used as the extracellular
solution. Whole-cell pipettes were filled with either standard intracellular saline (junction
potential of 23 mV vs. Na+-free/Ca
2+-free ASW), or saline where the K
+-aspartate was

23
replaced equimolar with either Cs+-aspartate (BP210100; Fisher) (junction potential of 23
mV) or tetraethylammonium-Cl (TEA; AC15090; Acros Organics, Morris Plains, NJ,
USA) (junction potential of 3 mV). Off-line subtraction was again used to correct for
junction potentials, whereas leak current was subtracted on-line using a P/4 protocol from
-40 mV, with sub-pulses of opposite polarity and one-fourth the magnitude, an inter-sub-
pulse interval of 500 ms, and 100 ms before actual test pulses.
Some pairs were bathed in Ca2+
-free ASW (composition as per nASW but with no
added Ca2+
, 66 mM Mg2+
and 0.5 mM ethylene glycol bis (aminoethyl ether) tetraacetic
acid (EGTA)). A number of experiments related to Ca2+
influx used an intracellular saline
with either 0 mM EGTA and no added Ca2+
or 20 mM EGTA and 35 nM Ca2+
or 40 mM
EGTA and 15 nM Ca2+
.
Intracellular dye staining and fluorescence microscopy of cultured bag cell neurons
Dye-coupling between electrically-coupled bag cell neurons was investigated by
staining one cell of a given pair with either sulforhodamine 101 (S-359; Invitrogen,
Burlington, ON, Canada), carboxyfluorescein (C194; Invitrogen), or biocytin (B-1592;
Invitrogen). For sulforhodamine or carboxyfluorescein, individual dyes were dissolved in
standard intracellular saline at 0.5% or 0.1% (w/v) and loaded under whole-cell voltage-
clamp at -60 mV. In some cases, the second neuron was also voltage-clamped at -60 mV,
but using a pipette containing with no added dye. Neurons were dye-loaded for 30 min,
by which time the soma of the cell was essentially the same colour as the electrode under
appropriate excitation light (see below). Photomicroscopy (see below) of neurons stained
with sulforhodamine or carboxyfluorescein was usually carried out at the end of the

24
recording period, with the electrode(s) still in place. However, in a few instances, the
electrode(s) were withdrawn, the dish returned to the 14°C incubator overnight, and
documentation undertaken the next day.
For biocytin, the tips of sharp microelectrodes pulled from 1.2 mm external
diameter/0.9 mm internal diameter borosilicate glass capillaries (TW120F-4; World
Precision Instruments) were filled with dye dissolved in water at 4% (w/v). When
backfilled with 0.5 M KCl, these electrodes had a resistance of 2-7 MΩ. One neuron from
a given pair, plated on glass coverslips (see Animals and cell culture for details), was
impaled and the membrane potential (typically -50 mV to -60 mV) monitored with an
Axoclamp 2B amplifier (Molecular Devices). Microelectrodes were bridge-balanced with
a Grass S88 stimulator (Astro-Med, Longueuil, QC, Canada) and the dye injected for 20
min. Injection used the Axoclamp DC current command to depolarize the neuron to -30
mV and then, with the stimulator, simultaneously inject 500-ms negative current steps at
0.5 Hz to periodically hyperpolarize the cell to -90 mV. The electrode was then
withdrawn, the dish returned to the incubator, and the neurons fixed the next day for 25
min in 4% (w/v) paraformaldehyde (04042; Fisher) in 400 mM sucrose dissolved in
nASW (pH 7.5 with NaOH). They were then permeabilized for 5 min with 0.3% (w/v)
Triton X-100 (BP151; Fisher) in fix and washed four times with phosphate-buffered
saline (PBS; composition in mM: 137 NaCl, 2.7 KCl, 4.3 Na2HPO4, 1.5 KH2PO4; pH 7.0
with NaOH). Neurons were then incubated overnight in the dark at 4°C in streptavidin-
Alexa Fluor 488 conjugate (S-32354; Invitrogen) at 1:100 in PBS, washed four times the
next day with PBS, the wells filled with mounting solution (26% w/v glycerol (BP2291;

25
Fisher), 11% w/v Mowiol 4-88 (17951; Polysciences, Warrington, PA, USA), and 110
mM TRIS (pH 8.5)), and covered with a glass coverslip.
Stained neurons were imaged with a Nikon TS100-F inverted microscope (Nikon,
Mississauga, ON, Canada), equipped with a Nikon Plan Fluor 20X (numerical aperture
0.50) objective and a 50-W mercury lamp. For carboxyfluorescein or biocytin-
streptavidin Alexa Fluor 488, excitation was provided by a 480/15-nm band pass filter,
and the fluorescence emitted to the eyepiece or camera through a 505-nm dichroic mirror
and 520-nm barrier filter. Sulforhodamine was excited with a 535/25-nm band pass filter,
and the emitted light passed through a 575 nm dichroic mirror and 590-nm barrier filter.
Images (1392 x 1040 pixels) were acquired at the mid-level of the soma on the vertical
axis using a Pixelfly USB camera (PCO-TECH, Romulus, MI, USA/Photon Technology
International, London, ON, Canada) and the Micro-Manager 1.4.5 plugin
(http://valelab.ucsf.edu/~MM/MMwiki/index.php/Micro-Manager) for ImageJ 1.43
(http://rsbweb.nih.gov/ij/) with 100-500 ms exposure times.
Ensemble, extracellular recording from the intact bag cell neuron cluster
To record afterdischarges from the intact bag cell neuron cluster, the abdominal
ganglion was removed as per Animals and cell culture. Ganglia were pinned ventral
surface up in nASW to the bottom of a Sylgard-lined 35 x 10 mm tissue culture dish,
which served as the bath, and maintained at 14°C using an Isotemp circulating chiller
(3016; Fisher). A wide-bore, fire-polished glass suction recording electrode (containing
nASW) was placed over the right bag cell neuron cluster, while a similar stimulating
electrode was placed at the rostral end of the right pleuroabdominal connective.

26
Stimulating current pulses were delivered with a Grass SD9 stimulator while voltage was
monitored using a Warner DP-301 differential amplifier (Warner Instruments; Hamden,
CT, USA). Voltage was high-pass filtered at 0.1 Hz and low-pass filtered at 1 kHz using
the DP-301 filters and acquired at a sampling rate of 2 kHz using the Digidata 1322A and
Axoscope 9.0 (Molecular Devices).
Drug application and reagents
Solution changes were accomplished using a calibrated transfer pipette to
exchange the bath (tissue culture dish) solution. Drugs were introduced by initially
removing a small volume (∼75 µl) of saline from the bath, combining that with an even
smaller volume (<10 µl) of drug stock solution, then reintroducing that mixture back into
the bath. Care was taken to pipette near the side of the dish and as far away as possible
from the neurons. Niflumic acid (N0630; Sigma-Aldrich), meclofenamic acid (M4531;
Sigma-Aldrich), 5-nitro-2-(3-phenylpropylamino), benzoic acid (N4779; Sigma-Aldrich),
18-α-glycyrrhetinic acid (G8503; Sigma-Aldrich), phorbol 12-myristate 13-acetate
(PMA; P8139; Sigma-Aldrich), cyclopiazonic acid (CPA; C1530; Sigma-Aldrich or
239805; Calbiochem), carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP;
21857; Sigma-Aldrich) and KN-62 (I-2142; Sigma-Aldrich) were dissolved as stocks in
dimethyl sulfoxide (DMSO; BP231-1; Fisher). Quinine (Q1250; Sigma-Aldrich) stock
was dissolved in ethanol. The maximal final concentration of DMSO or ethanol ranged
from 0.05% to 0.5% (v/v), which in control experiments had no effect on holding current,
membrane conductance, or junctional current (Magoski et al. 2000; Magoski and
Kaczmarek 2005; Kachoei et al. 2006; Lupinsky and Magoski 2006; Hung and Magoski

27
2007; Geiger and Magoski 2008; Gardam and Magoski 2009; Hickey et al. 2010; Tam et
al. 2011; Groten et al. 2013; Hickey et al. 2013).
Analysis
Most analysis involved cell pairs designated as neuron 1 and neuron 2, with
membrane potentials and membrane currents specified as V1, V2, I1, and I2,
respectively. The Clampfit (8.1 or 10.2) analysis program of pCLAMP was used to
determine the amplitude and time course of membrane current or voltage. For current,
cursors were placed at an average point along the baseline, prior to any voltage step, and
at steady-state during the response to a voltage step (usually within 20 ms of the end of
the step). Similarly, for voltage, cursors were placed at the baseline, prior to a current
step, and at the steady-state membrane potential seen during a hyperpolarizing pulse or
the peak voltage of an action potential or ETP. In both cases, the difference between the
two cursors values was taken as the amplitude of the response.
Membrane conductance was determined by taking the difference between the
holding potential and a given test voltage step delivered to a neuron (V1- -60 mV), and
then dividing that into the current (I1) evoked by the test voltage (i.e., I1/V1 - -60 mV),
and vice versa (I2/V2- -60 mV). Junctional conductance was calculated by dividing the
current in the second neuron (V2) by the junctional voltage (V2-V1) (i.e., I2/V2 -V1),
and vice versa (I1/V1 -V2). Both membrane and junctional conductance were plotted
against the test voltage using Prism 6.0 (GraphPad Software; La Jolla, CA, USA).
Coupling coefficient was ascertained by the ratio of V2/V1 or V1/V2, at the steady state
membrane potential after the hyperpolarizing current pulse to neuron 1 or neuron 2,

28
respectively. Times of peak voltage change were determined with Clampfit by placing
cursors before and after an action potential in one neuron as well as the corresponding
ETP in the second neuron. The peak-to-peak latency between action potential and ETP
was calculated as the difference between these times. The percent change in junctional
current following Ca2+
influx due to stimulation or the addition of a drug was calculated
by comparing the subsequent current to control. Finally, ETP area was measured by
placing cursors at the beginning of the ETP and at the steady-state voltage after the ETP,
i.e., the time at which the postsynaptic membrane potential had fully returned to baseline.
The difference between the two cursors was taken as the region of the ETP, above which
the ETP area was calculated. Time to recovery of the ETP was achieved by placing
cursors at peak of the ETP and steady-state voltage after the ETP, then taking the
difference.
Statistics were performed using Prism or InStat 3.1 (version 3.0; GraphPad
Software Inc, San Diego, CA, USA). Data are presented as the mean ± standard error of
the mean. The Kolmogorov-Smirnov method was used to test data sets for normality. For
normality distributed data, Student’s paired or unpaired t-test was used to test for
differences between two means, while a standard one-way analysis of variance
(ANOVA) with Dunnett or Tukey-Kramer multiple comparisons test was used to test for
differences between multiple means. For not-normal distributed data, one-tailed Mann-
Whitney U-test was used to test for differences between two means. Difference in
frequency was studied using Fisher's exact test. Data was considered statistically different
if the one- or two-tailed p-value was <0.05.

29
Chapter 3
Results
Bag cell neuron electrical synapses can be recapitulated in primary culture
Previous morphological and electrophysiological studies of bag cell neurons in
the intact cluster indicate that the synchronous firing during the afterdischarge is achieved
through electrical coupling (Kupfermann and Kandel 1970; Blankenship and Haskins
1979; Kaczmarek et al. 1979; Brown et al. 1989). However, because the neurons are
coupled to more than one partner, perhaps many more, the ability to characterize the
electrical synapse in the intact cluster is limited. Thus, to study these synapses under
more controlled conditions, bag cell neurons were examined in primary culture. Neurons
were disassociated and brought into contact at either the somata (Figure 4A) or neurites
(Figure 4B). After 2-4 d in culture, pairs usually exhibited electrical coupling (~ 80% of
pairs). In the case of opposition between somata, pairs showed stronger coupling than via
neurites. Thus, most experiments were done on soma-soma pairs. Whole-cell recording,
either in voltage- or current-clamp was used in almost all cases. Unless stated otherwise,
recordings were made in nASW as the extracellular solution and with a K+-Asp-based
intracellular saline in the recording pipettes (see Materials and Methods for details).
Neurons were identified as neuron 1 and neuron 2, with the protocol run using neuron 1
as presynaptic (receiving test pulses) and neuron 2 as postsynaptic (held at -60 mV), and
vice versa.
To confirm the existence of electrical synapses between bag cell neurons in
culture, a 1-nA, 5-sec depolarizing current was injected into one neuron under current-

30
Figure 4. Cultured pairs of bag cell neurons show electrical coupling.
A, B) Phase-contrast photomicrographs of paired bag cell neurons. The soma-soma
configuration (A) is between cells cultured for 2 d, while the neurons making neurite-to-
neurite contact (B) are in vitro for 3 d. The scale bar applies to both pictures.
C) Whole-cell current-clamp recording from a pair of coupled bag cell neurons dialyzed
in nASW with standard K+-Asp-based intracellular saline. A 1-nA depolarizing current
injection for 5 sec into neuron 1 (I1) evokes action potentials in neuron 1 (V1), resulting
steady-state depolarization and electrotonic potentials (ETP) in neuron 2 (V2) that reach
threshold to generate action potentials (AP).
D) Under whole-cell voltage-clamp, a pair of coupled bag cell neurons is held at -60 mV.
Changing the voltage of one cell to -90 mV (V1) for 200 ms elicits both inward
membrane current in neuron 1 (I1) and junctional current in neuron 2 (I2). The outward
current of I2 is the current required to keep the second cell voltage clamped at -60 mV
(V2) in the face of a potential difference across the gap junction, and thus represents the
junctional current.

31
clamp, which usually elicited a train of non-accommodating action potentials throughout
the duration of the stimulus (Figure 4C). Both the steady-state depolarization and the
action potentials in the stimulated neuron generated ETPs in the second neuron. This
electrotonic transmission was in some cases able to recruit the coupled cell and elicit full
action potentials.
Junctional current was measured under voltage-clamp by applying a potential
difference to one neuron and measuring the evoked current in the second cell (Bennett
1966). From a holding potential of -60 mV for both cells, a 200-ms voltage step to -90
mV, while continuing to hold the second cell at -60 mV, induced current in both neurons
(Figure 4D). This hyperpolarizing pulse elicited an inward current in the stimulated
neuron, and when cells were electrically-coupled, evoked an outward current in the
unstimulated neuron. Because the second neuron was maintained at -60 mV, the outward
current corresponds to current flowing across the electrical synapse.
Electrical coupling between cultured bag cell neurons is voltage-independent
Prior work has shown the gating of electrical synapses and gap junctions from a
number of species to be either voltage-dependent or -independent (Furshpan and Potter
1959; Spray et al. 1981; Verselis et al. 1991). To investigate this in bag cell neurons,
pairs of electrically-coupled cultured cells were whole-cell voltage-clamped at -60 mV,
then one cell was stepped through a series of 10-mV square pulses from -90 mV through
to 0 mV in 200-ms intervals, while maintaining the other cell at -60 mV (Figure 5A, top
and lower middle). In response to the step commands, the presynaptic neuron displayed
membrane current that was linear near -90 mV, but then rectified outwardly in a time-

32
Figure 5. Electrical synapses between bag cell neurons are voltage-insensitive.
A) Under voltage-clamp, a pair of neurons is in nASW dialyzed with standard
intracellular saline. While holding both neurons at -60 mV, a series of 10-mV (V1)
voltage steps from -90 to 0 mV in 200-ms intervals in neuron 1 elicits both voltage-
dependent membrane current (I1) in neuron 1 and voltage-independent junctional current
(I2) in neuron 2.
B) When current in neuron 2 (I2) (junctional current) is plotted against the presynaptic
voltage (V1), it shows a linear and non-rectifying relationship. The current flows both
ways (from neuron 1 to neuron 2 and vice versa) and the difference between the two is
not significant. Inset shows recording configuration. VC, voltage clamp.
C, D) The voltage steps and corresponding current in the stepped neuron are used to
determine the membrane conductance (G-membrane). The driving force voltage (the
difference between the test voltage in the pulsed neuron and holding voltage in the non-
pulsed neuron) and junctional current are used to determine junctional conductance (G-
junction). The membrane conductance is voltage-dependent, while the junctional
conductance is insensitive to transjunctional voltage.

33
and voltage-dependent manner closer to 0 mV, while the postsynaptic neuron presented a
nearly instantaneous and completely linear current to all test pulses (Figure 5A, upper
middle and bottom). When the average steady-state postsynaptic current (junctional
current) was plotted against the presynaptic voltage step, a linear current-voltage
relationship was observed (Figure 5B, 1→2 n=11; 2→1 n=11). This was the case
regardless if either neuron 1 or neuron 2 received the voltage protocol as the presynaptic
cell. Furthermore, in all 11 pairs examined, the junctional current did not rectify, and was
the same magnitude when either neuron 1 or neuron 2 served in a presynaptic role.
Membrane conductance (G-membrane) and junctional conductance (G-junction)
were obtained by dividing the steady-state presynaptic membrane current and junctional
current (both measured within 20 ms of the end of the step) by the membrane driving
force (test voltage - -60 mV) and the transjunctional voltage (-60 mV - test voltage),
respectively. As such, the -60 mV holding potential was treated as the reversal potential
for both the membrane and junctional conductance. Plotting either of these derived
variables on the ordinate and the test voltage on the abscissa provided G-membrane vs.
voltage (n=11) (Figure 5C) and G-junction vs. voltage (n=11) (Figure 5D) curves for
neuron 1 and neuron 2. As expected from the membrane current data, the presynaptic
membrane conductance had a linear portion, composed of leak channels, from -90 mV to
-50 mV, and a non-linear portion, composed ostensibly of voltage-gated K+ channels,
from -40 mV to 0 mV. The membrane conductance was largely the same for both cells in
the pairs, with neither neuron 1 nor neuron 2 yielding significantly different conductances
at a given command potential. Conversely, junctional conductance showed no sensitivity

34
to transjunctional voltage, and was approximately equal (8-10 nS) at all command
potentials, with no significant difference at any of the test voltages. Consistent with the
junctional current not rectifying, the junctional conductance was not different when the
presynaptic cell was neuron 1 or neuron 2.
Bag cell neurons present large coupling coefficients
While junctional current and conductance provide an exact measure of electrical
synapse performance, under physiological conditions, coupled neurons function to
transfer changes in membrane potential. Thus, coupling coefficient, defined as the ratio
of membrane voltage changes between postjunctional and prejunctional cells, was
determined using whole-cell current-clamp of cultured bag cell neuron pairs (Figure 6A,
inset). For coupling coefficient, both neurons were set at -60 mV and a 1500-ms
hyperpolarization current step (-50 to -250 pA) was delivered to one cell, resulting in
near simultaneous hyperpolarization of the second cell (Figure 6A). If the presynaptic
cell was either neuron 1 (n=8) or neuron 2 (n=8), the steady-state hyperpolarization was
equally prominent, resulting in similar coupling coefficient of ∼0.6 in both directions
(Figure 6C). For neurons that were not coupled, either because they failed to form an
electrical synapse despite being in contact or were close but not touching, changing the
presynaptic membrane potential had little postsynaptic impact (Figure 6B). In some cases
there were modest alterations in postsynaptic voltage, but these were likely due to
membrane noise. As such, the apparent coupling coefficient between not-coupled bag cell
neurons was very low and similar regardless if neuron 1 (n=7) or neuron 2 (n=6) served
in a presynaptic capacity (Figure 6B). The difference between coupling coefficient for

35
Figure 6. Bag cell neurons can be highly coupled.
A) Neurons are bathed in nASW, dialyzed with standard intracellular saline and current-
clamped at -60 mV. Neuron 1 is injected with a 1500-ms step of -250 pA (I1), producing
hyperpolarization in both neuron 1 (V1) and neuron 2 (V2). Inset shows recording
configuration. CC, current clamp. The scale bars apply to both traces.
B) In not-coupled pairs (no electrical coupling or no contact), hyperpolarizing neuron 1
(V1) does not alter the membrane potential in neuron 2 (V2).
C) Summary graph comparing the degree of coupling between coupled and not-coupled
pairs (two-tailed unpaired Student’s t-test). Electrical transmission is reciprocal with no
appreciable difference between directions (from neuron 1 to 2 or vice versa). For this and
subsequent bar graphs, n-value (typically the number of pairs of cultured bag cell
neurons) is shown in brackets within, just above, or below the individual bars.
D) The relationship between junctional conductance and coupling coefficient is weak.

36
coupled pairs vs. not-coupled pairs was significant (Figure 6C). For some coupled pairs,
junctional conductance was also measured with voltage-clamp prior to assaying coupling
coefficient under current-clamp (1→2 n=8; 2→1 n=8). Plotting junctional conductance
on the abscissa as the independent variable and coupling coefficient on the ordinate as the
dependent variable yielded a linear and modestly positive correlation (Figure 6D). The
higher the junctional conductance between a pair of neurons, the greater the coupling
coefficients.
Coupling between bag cell neurons is electrotonic rather than chemical
To evaluate electrotonic signaling, current-clamped pairs were set to -60 mV and
presynaptic action potentials elicited with five 100-ms, 750-pA depolarizing current steps
at 1 Hz. A given presynaptic spike consistently produced an ~10 mV postsynaptic
depolarizing ETPs in a one-for-one relationship (Figure 7A) (n=5). A full recovery of
postsynaptic voltage after each spike was seen. Latency was rapid, with postsynaptic
depolarization beginning early in the rising phase of the presynaptic action potential.
Because of the difficulty in precisely ascertaining when the postsynaptic response began,
latency was quantitated by taking the time from the peak of the fifth action potential to
the peak of the corresponding evoked ETP and found to be ~15 ms (Figure 7D).
Communication between pairs of bag cell neurons is consistent with electrical
signalling; even so, there is evidence that, in addition to egg-laying hormone, the cells
secrete so-called α-, β-, and γ-bag cell peptides, which can depolarize bag cell neurons
when applied exogenously (Loechner and Kaczmarek 1994; Hatcher and Sweedler 2008).
To address the possibility that the ETP contained a chemical component, paired bag cell
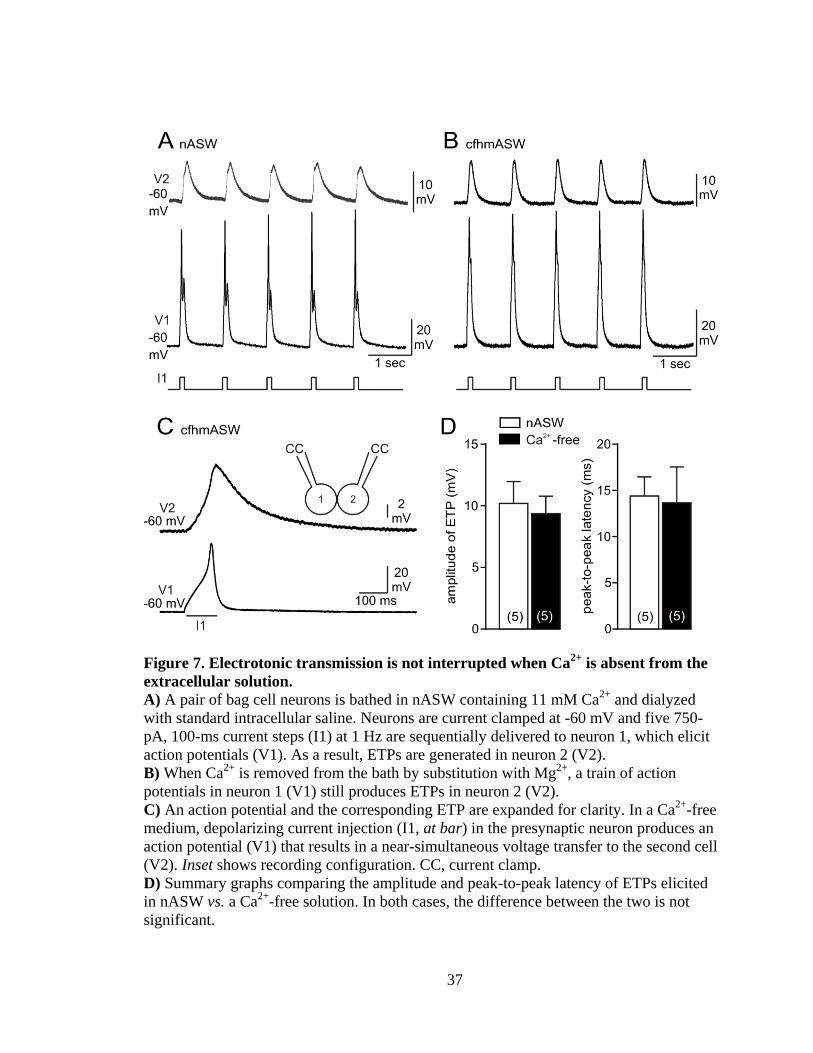
37
Figure 7. Electrotonic transmission is not interrupted when Ca2+
is absent from the
extracellular solution.
A) A pair of bag cell neurons is bathed in nASW containing 11 mM Ca2+
and dialyzed
with standard intracellular saline. Neurons are current clamped at -60 mV and five 750-
pA, 100-ms current steps (I1) at 1 Hz are sequentially delivered to neuron 1, which elicit
action potentials (V1). As a result, ETPs are generated in neuron 2 (V2).
B) When Ca2+
is removed from the bath by substitution with Mg2+
, a train of action
potentials in neuron 1 (V1) still produces ETPs in neuron 2 (V2).
C) An action potential and the corresponding ETP are expanded for clarity. In a Ca2+
-free
medium, depolarizing current injection (I1, at bar) in the presynaptic neuron produces an
action potential (V1) that results in a near-simultaneous voltage transfer to the second cell
(V2). Inset shows recording configuration. CC, current clamp.
D) Summary graphs comparing the amplitude and peak-to-peak latency of ETPs elicited
in nASW vs. a Ca2+
-free solution. In both cases, the difference between the two is not
significant.

38
neurons were bathed in Ca2+
-free/high-Mg2+
ASW (0 mM Ca2+
, 66 mM Mg2+
), and
spikes initiated in the presynaptic neuron from -60 mV with a 100-ms, 750-pA
depolarizing current step (n=5 pairs). Without extracellular Ca2+
, the onset of the ETP
remained before the peak of the presynaptic action potential, and the shape of the ETP
was not dramatically altered compared to that produced in the presence of Ca2+
(Figure
7B, C). The amplitude and latency of the ETP in Ca2+
-free/high-Mg2+
ASW were not
statistically different from that evoked in nASW (Figure 7D).
Bag cell neuron gap junctions are not dye-permeable
Passage of dyes between coupled cells has often been used to define the
permeability characteristics of electrical synapses (Andrew et al. 1981; Peinado et al.
1993; Ewadinger et al. 1994; Li et al. 1996). Kaczmarek et al. (1979) reported occasional
dye-coupling using Lucifer yellow injections both in the intact cluster and between
cultured bag cell neurons. This was investigated here by introducing either
sulforhodamine or carboxyfluorescein (both dissolved in the standard intracellular saline)
through the whole-cell pipette under voltage-clamp at -60 mV into one of the two cells
from a pair of cultured bag cell neurons that were in physical contact. In some cases, only
the one neuron was recorded using the dye-filled pipette, while in other instances the
second cell was also whole-cell voltage-clamped at -60 mV, but with a dye-free pipette.
In the latter situations, coupling was confirmed under voltage-clamp. Staining of one
neuron for ∼30 min did not result in appreciable dye transfer when either sulforhodamine
(n=4 at 0.1% w/v and n=3 at 0.5%) or carboxyfluorescein (n=5 at 0.1% and n=3 at 0.5%)
was used (Figure 8A, B). This was observed both at the end of the recording period or if

39
Figure 8. Bag cell neuron electrical synapses show no dye-coupling.
A, B) Fluorescence-emission photomicrographs of different dye-stained coupled paired
cultured bag cell neurons. In the upper-right of each picture, the phase image taken prior
to recording shows somata and associated neurites (full field of view of inset=256 μm).
Either sulforhodamine-101 (Mr=606.71) or carboxyfluorescein (Mr=376.32) are whole-
cell loaded at 0.5% (w/v) in K+-aspartate-based intracellular saline under voltage-clamp
at -60 mV in nASW for 30 min. Dye-containing pipettes can be seen in the middle-right
and upper-left of A and B, respectively.
C) Biocytin (4% w/v in water), introduced by sharp-electrode and allowed to diffuse over
night, then labelled with streptavidin Alexa Fluor 488 (courtesy of Ms. H. Hodgson).
Although all dyes clearly stain individual neurons, none transfers appreciably to the non-
stained cell.

40
the electrode(s) were taken off the neuron(s) and the dish kept overnight in the incubator.
As an alternative to whole-cell dye-loading, we also used sharp-electrode current-clamp
to inject biocytin (n=6) into one of the cells from a pair of cultured bag cell neurons.
Following an overnight period in the incubator, pairs were exposed to streptavidin
conjugated with Alexa Fluor 488 (see Material and Methods, Intracellular dye staining
and fluorescence microscopy of cultured bag cell neurons for details). The high affinity
binding of streptavidin to biocytin allows for fluorescence-based localization of the tracer
subsequent to any diffusion (Heitzmann and Richards 1974). However, biocytin was also
not detected in non-injected neurons of the pair (Figure 8C).
Bag cell neuron gap junctions selectively pass solutes
As an alternative to dye-coupling, the permeability of the electrical synapse was
examined by whole-cell loading the presynaptic neuron with either of the K+ channel
blockers, TEA or Cs+, and then testing the postsynaptic neuron for changes in K
+ current.
The effectiveness of the K+ channel blockers was first examined in single neurons.
Specifically, neurons were voltage-clamped in Na+-free/Ca
2+-free ASW at -40 mV while
dialyzing for 10 min with an intracellular saline where the K+ was equimolar substituted
with either TEA or Cs+, and a 200-ms voltage step to +30 mV elicited K
+ current (Strong
and Kaczmarek 1986). K+ channel blockers, TEA (n=3) and Cs
+ (n=5), inhibited the
outward current in single bag cell neurons by ~70% at 10 min vs. 5 min after
breakthrough (Figure 9B, C). In control neurons (n=4) where the pipette contained the
K+-Asp based intracellular saline, an ~10% increase in K
+ current was seen (Figure 9A).
TEA and Cs+ significantly reduced K
+ current as compared to K
+-Asp (Figure 9D).

41
Figure 9. K+ channel blockers reduce outward current in single bag cell neurons.
A) A bag cell neuron is bathed in Na+-free/Ca
2+-free ASW and dialyzed with standard
intracellular saline. Insets represent single neurons with a pipette containing different
intracellular salines. From a holding potential of -40 mV, a voltage step to +30 mV is
given to the neuron. K+ current elicited by the 200-ms voltage step (I) (black trace) does
not change over time (gray trace).
B, C) When K+-Asp in the standard intracellular saline is replaced with either Cs
+-Asp or
TEA-Cl, the K+
current (black traces) is blocked after 10 min (gray traces).
D) The summary graph showing the mean change in K+ current with intracellular K
+-
Asp, Cs+-Asp or TEA-Cl. K
+ current significantly decreases when Cs
+-Asp or TEA-Cl is
in the recording pipette (ANOVA; Dunnett multiple comparisons test).

42
To test whether the K+ channel blockers are able to pass between bag cell neurons
via gap junctions, K+ current was examined only in the postsynaptic neuron of an
electrically-coupled pair. Neurons were bathed in Na+-free/Ca
2+-free ASW, the
presynaptic neuron was voltage-clamped at -40 mV while dialyzing with intracellular
saline where the K+ was replaced equimolar with TEA or Cs
+. The postsynaptic cell was
also voltage-clamped at -40 mV, but using standard K+-containing intracellular saline,
and at both 5 and 10 min of dialysis stepped to +30 mV for 200 ms to generate K+ current
(Figure 10A, inset). In controls (n=8), where both neurons were recorded using K+-Asp in
the pipette, the steady-state K+ current in the postsynaptic neuron showed an ∼5%
increase at 10 min vs. 5 min (Figure 10A). A similar run-up was seen when neuron 1 was
dialyzed with TEA-containing intracellular saline (n=7) (Figure 10B); however,
introducing Cs+ into neuron 1 (n=7) caused the K
+ current in neuron 2 to decrease by
∼10%, which was significantly different from control (Figure 10C, D).
Select gap junction blockers inhibit cultured bag cell neuron electrical synapses
Gap junctions can be blocked by a variety of compounds, including various
arylaminobenzoates, the triterpenoid, glycyrrhetinic acid, and the anti-malarial drug,
quinine (Davidson and Baumgarten 1988; Harks et al. 2001; Srinivas et al. 2001;
Srinivas and Spray 2003; Juszczak and Swiergiel 2009). The ability of these reagents to
inhibit the cultured bag cell neuron junctional current was tested under whole-cell
recording. Both neurons were voltage-clamped at -60 mV and a 200-ms step to -90 mV
delivered to neuron 1 while maintaining neuron 2 at -60 mV (Figure 11A, inset). The
junctional current in neuron 2, arising from the step in neuron 1, was measured before

43
Figure 10. Cs+, but not TEA, passes through bag cell neuron gap junctions.
A) Insets, a voltage change from -40 to +30 mV is given to neuron 2 (V2), while the
membrane potential in neuron 1 (V1) is kept at -40 mV. The diagram shows a pair of
neurons with pipettes containing K+-Asp-based intracellular saline. In Na
+-free/Ca
2+-free
ASW, a 200-ms voltage change elicits K+ current in neuron 2 (I2) (black trace) that
shows a small increase after 10 min (gray trace).
B, C) Insets, the K+-Asp in the standard intracellular saline in neuron 1 is replaced with
either TEA-Cl or Cs+-Asp and K
+ current is recorded in neuron 2 (I2) (black traces).
After 10 min, Cs+, but not TEA, travels to neuron 2 and decreases the K
+ current (gray
traces).
D) The summary graph compares the mean K+ current change in neuron 2. When the
standard intracellular saline in neuron 1 is substituted with Cs+-Asp or TEA-Cl, dialysis
results in Cs+-Asp significantly reducing K
+ current in neuron 2 as compared to K
+-Asp
(ANOVA; Dunnett multiple comparisons test).
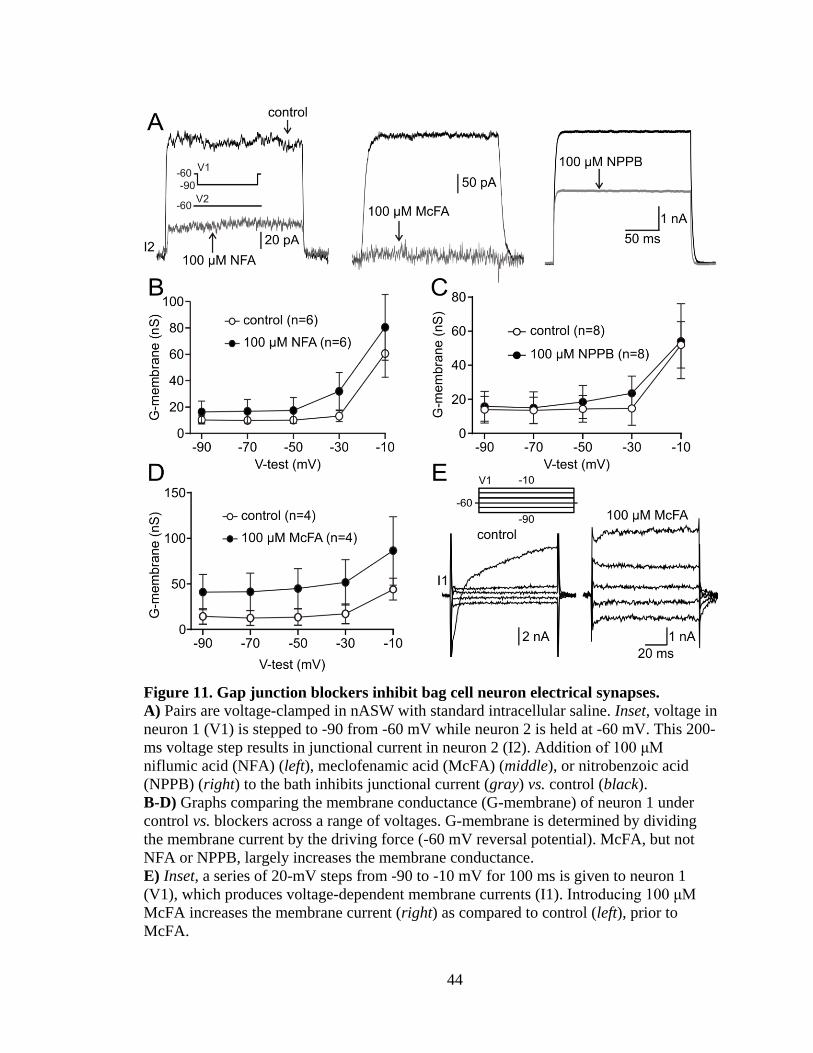
44
Figure 11. Gap junction blockers inhibit bag cell neuron electrical synapses.
A) Pairs are voltage-clamped in nASW with standard intracellular saline. Inset, voltage in
neuron 1 (V1) is stepped to -90 from -60 mV while neuron 2 is held at -60 mV. This 200-
ms voltage step results in junctional current in neuron 2 (I2). Addition of 100 μM
niflumic acid (NFA) (left), meclofenamic acid (McFA) (middle), or nitrobenzoic acid
(NPPB) (right) to the bath inhibits junctional current (gray) vs. control (black).
B-D) Graphs comparing the membrane conductance (G-membrane) of neuron 1 under
control vs. blockers across a range of voltages. G-membrane is determined by dividing
the membrane current by the driving force (-60 mV reversal potential). McFA, but not
NFA or NPPB, largely increases the membrane conductance.
E) Inset, a series of 20-mV steps from -90 to -10 mV for 100 ms is given to neuron 1
(V1), which produces voltage-dependent membrane currents (I1). Introducing 100 μM
McFA increases the membrane current (right) as compared to control (left), prior to
McFA.

45
and 10 min after introducing either DMSO (the vehicle) or one or more of the putative
blockers. Electrical transmission was markedly lowered by all of the arylaminobenzoates
tested. For example, 100 μM of either niflumic acid (NFA) (n=14) or 5-nitro-2-(3-
phenylpropylamino) benzoic acid (NPPB) (n=7) resulted in an ∼50% decrease in
junctional current (Figure 11A, left and right), while 100 μM of meclofenamic acid
(McFA) (n=6) produced an up to ~80% inhibition (Figure 11A, middle). Neither NFA
(n=6) nor NPPB (n=8) at 100 μM had any obvious consequence on membrane
conductance (Figure 11B, C), while McFA (n=4) often increased the membrane
conductance (Figure 11D, E); as such, despite being an effective blocker, this drug was
not pursued further.
A cocktail of 100 μM each of NFA and NPPB (n=7) was found to be most
effective, as it caused an ∼75% decrease in junctional current (Figure 12A) and had no
apparent effect on membrane conductance (n=8) (Figure 12B, C). This inhibitory effect
was also seen under current clamp where the hyperpolarizing response to negative current
injection into the presynaptic neuron transferred poorly to the coupled partner (n=10)
(Figure 12D, left). Application of 100 μM NFA and 100 μM NPPB together strongly
attenuated coupling as compared to DMSO (n=5) (the vehicle) (Figure 12D, right).
The effects of both NFA and NPPB, as well as another arylaminobenzoate, 30-
200 μM McFA, were concentration dependent, with 30 or 60 μM having less or no
impact, while DMSO (the vehicle) caused only a small increase in junctional current
(Figure 13A). Two other gap junction blockers, 50 μM 18-α-glycyrrhetinic acid (n=6)
and 300 μM quinine (n=12), did not change the junctional current between cultured bag
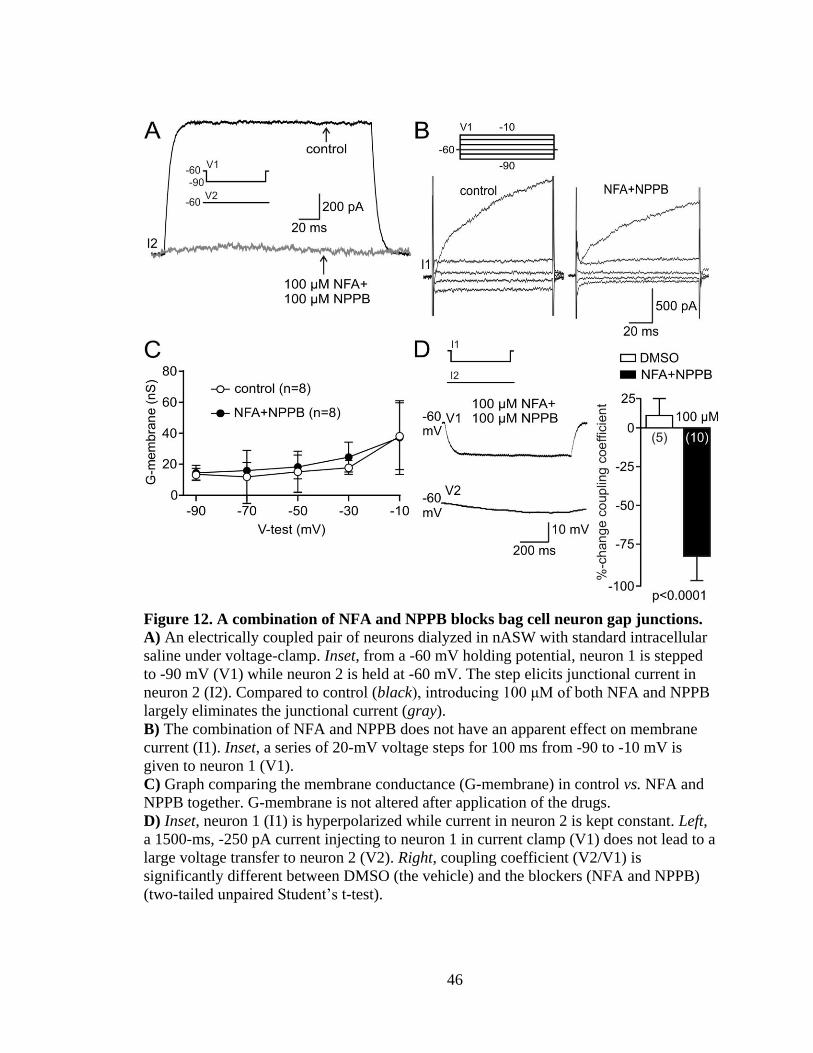
46
Figure 12. A combination of NFA and NPPB blocks bag cell neuron gap junctions.
A) An electrically coupled pair of neurons dialyzed in nASW with standard intracellular
saline under voltage-clamp. Inset, from a -60 mV holding potential, neuron 1 is stepped
to -90 mV (V1) while neuron 2 is held at -60 mV. The step elicits junctional current in
neuron 2 (I2). Compared to control (black), introducing 100 μM of both NFA and NPPB
largely eliminates the junctional current (gray).
B) The combination of NFA and NPPB does not have an apparent effect on membrane
current (I1). Inset, a series of 20-mV voltage steps for 100 ms from -90 to -10 mV is
given to neuron 1 (V1).
C) Graph comparing the membrane conductance (G-membrane) in control vs. NFA and
NPPB together. G-membrane is not altered after application of the drugs.
D) Inset, neuron 1 (I1) is hyperpolarized while current in neuron 2 is kept constant. Left,
a 1500-ms, -250 pA current injecting to neuron 1 in current clamp (V1) does not lead to a
large voltage transfer to neuron 2 (V2). Right, coupling coefficient (V2/V1) is
significantly different between DMSO (the vehicle) and the blockers (NFA and NPPB)
(two-tailed unpaired Student’s t-test).

47
Figure 13. Electrical coupling and afterdischrge are inhibited by blockers.
A) Summary graph of percent-change from control in junctional current subsequent to
adding vehicle or drug. Compared to DMSO, NFA (60 to 200 μM), McFA (60 and 100
μM), and NPPB (30 and 100 μM) significantly inhibit junctional current in a dose-
dependent manner. However, 50 μM α-GRA and 300 μM quinine (quin) do not; in fact,
α-GRA may enhance coupling. In addition, a cocktail of NFA and NPPB (100 μM each)
results in a block that is more effective than either agent alone (ANOVA; * P<0.05, **
P<0.01, Dunnett multiple comparisons test).
B) Ensemble extracellular recordings (ext) from the intact bag cell neuron cluster
following stimulation of the corresponding pleuroabdominal connective (stim) in the
abdominal ganglia (inset) bathed with nASW. Left, exciting the synaptic input (stim; at
bar; 5-ms, 20-V at 5 Hz for 10 sec) under control conditions elicits a typical
afterdischarge with a fast- and slow-phase. Afterdischarge lasts 24 min, but is truncated
at 9 min for display. Right, a 10-min incubation of a different preparation with NFA and
NPPB renders stimulation of the connective ineffective at provoking an afterdischarge
(courtesy of Dr. N.S. Magoski).

48
cell neurons (Figure 13A).
The afterdischarge is abated by gap junction blockers
The effect of gap junction blockers was tested on the afterdischarge itself by using
suction electrodes to make both extracellular recordings from the intact bag cell neuron
cluster in the abdominal ganglion and stimulate the synaptic input descending via the
pleuroabdominal connective. These recordings were ensemble and represented the
activity of the entire cluster (Kupfermann and Kandel 1970; Dudek and Blankenship
1977). In all five control preparations, afferent activation triggered an afterdischarge,
with the characteristic 1-2 min fast-phase of firing at the start, followed by the 20-30 min
slow-phase, and an average duration of 24.5 ± 2.3 min (Figure 13B, left). Clusters from
ganglia treated with 100 μM of each of NFA and NPPB usually did not display an
afterdischarge (Figure 13B, right), with only one of five presenting a normal bursting
pattern subsequent to synaptic stimulation. Hence, the average duration of the
afterdischarge under gap junction block was 6.1 ± 6.1 min, which was significantly
different from control (p<0.05, one-tailed Mann-Whitney U-test). Moreover, the
difference in frequency of afterdischarge inhibition of control vs. block was significant
(p<0.03, Fisher’s exact test).

49
Increased intracellular Ca2+
inhibits electrical synapses
The afterdischarge is accompanied by an increase in intracellular Ca2+
due to both
voltage-gated influx and release (Arch 1972; Pinsker and Dudek 1977; Kaczmarek et al.
1982; Fisher et al. 1994). Elevated intracellular Ca2+
has been shown to inhibit gap
junctions in both non-neuronal (Loewenstein et al. 1967; Spray et al. 1982) and neuronal
(Baux et al. 1978; Rao et al. 1987) cells; however, it is rare to find cases of Ca2+
channels
provoking the intracellular Ca2+
rise. Thus, we examined whether Ca2+
influx, produced
by a train of 75-ms pulses from -60 to 0 mV at 5 Hz for 1 min and 1 Hz for 5 min, which
mimics the fast- and early slow-phase of the afterdischarge, respectively, suppresses
junctional conductance.
Electrically-coupled cultured bag cell neurons were voltage-clamped in nASW
(11 mM Ca2+
) and dialyzed with standard intracellular saline (300 nM free Ca2+
and 5
mM EGTA) in the pipettes. The membrane potential of both neurons was initially held at
-60 mV. In response to a 200-ms voltage command to -90 mV in one neuron, while
keeping the voltage of the second cell at -60 mV, the junctional current produced in the
second neuron was measured. Following a 30-min dialysis period, when neurons were
not stimulated (n=6), this junctional current showed an ~10% increase over time (Figure
14A). However, delivering the train-stimulus caused an ~30% decrease in junctional
current (Figure 14B) (n=7). To initially confirm that Ca2+
was inhibiting electrical
communication, extracellular Ca2+
was removed by substituting with Mg2+
. With no Ca2+
present in the bath (n=7), the train-stimulus failed to alter the junctional current (Figure
14C). The results were then quantified by measuring the percent change in junctional

50
Figure 14. Elevation of intracellular Ca2+
decreases junctional current.
A) Neurons are voltage-clamped in nASW (11 mM Ca2+
) and dialyzed with standard
intracellular saline containing 5 mM EGTA and 300 nM free Ca2+
. Inset, neuron 1 is
stepped from -60 to -90 mV (V1), while neuron 2 is clamped at -60 mV (V2). The 200-
ms voltage change in neuron 1 produces membrane current in neuron 1 (I1) and
junctional current in neuron 2 (I2). The scale bars apply to both traces.
B) Ca2+
influx, produced by simultaneously delivering to both cells (stim) a train of 75-
ms pulses from -60 to 0 mV, at 5 Hz for 1 min followed by 1 Hz for 5 min, decreases the
junctional current (after) compared to the initial response (before). The altered junctional
current (right) is recorded immediately after stimulation. For panels B and C only the
junctional current (I2) is shown. Inset shows a pair of neurons with stimulation to both
recording pipettes.
C) In a Ca2+
-free extracellular medium, where Mg2+
is substituted for Ca2+
, the train-
stimulus does not change the junctional current.
D) Summary graph showing that the percent change in junctional current significantly
decreases after stimulation in the presence of extracellular Ca2+
vs. both the unstimulated
control and those neurons stimulated in Ca2+
-free solution (ANOVA; Tukey-Kramer
multiple comparisons test).

51
current. The uncoupling effect of Ca2+
influx was statistically significant when compared
to control and Ca2+
-free groups (Figure 14D).
Synchronous stimulation is most effective at inhibiting electrical coupling
Closure of gap junctions often requires increasing cytosolic Ca2+
to a specific
threshold (Rose and Loewenstein 1976). We examined whether a Ca2+
rise in only one
cell of a pair is sufficient to inhibit the electrical synapses between bag cell neurons.
Following a 30-min dialysis under voltage-clamp, a 200-ms voltage step to -90 from -60
mV was first given to the presynaptic cell, while maintaining the postsynaptic cell at -60
mV, to induce junctional current. As control (n=5), the train-stimulus was then
simultaneously delivered to both neurons, which resulted in an ~25% decrease in
junctional current that lasted for up to 30 min (Figure 15A). However, the same train-
stimulus in only one neuron (n=9), while keeping the second cell at -60 mV, caused a
smaller inhibition of ~15% that decayed to baseline over time (Figure 15B). The Ca2+
-
induced uncoupling was significantly greater and lasted longer when both neurons were
stimulated as compared to one neuron (Figure 15C).
Depletion of mitochondrial Ca2+
markedly attenuates junctional current
During the slow phase of the afterdischarge, CICR occurs from intracellular stores
(Fisher et al. 1994), which requires an interaction between mitochondria and endoplasmic
reticulum (Geiger and Magoski 2008). As such, EGTA and Ca2+
were removed from the
recording pipette solutions (to avoid influencing store-derived Ca2+
) and the impact of the
Ca2+
liberated from either the endoplasmic reticulum or mitochondria on electrical
coupling examined. Using pairs of cultured bag cell neurons, both cells were held at -60

52
Figure 15. Inhibition of junctional current is most effective when Ca2+
entry occurs
in both neurons.
A) Intracellular Ca2+
rise in a pair of neurons by delivering to both neurons (stim) a train
of 75-ms depolarizing pulses from -60 to 0 mV at 5 Hz for 1 min then 1 Hz for 5 min,
decreases the junctional current (I2) for up to 30 min. Insets, neuron 1 is clamped at -60
mV while neuron 2 is stepped to -90 mV. The diagram represents a pair of neurons with
stimulation to both pipettes. The scale bars apply to all time points.
B) When the train-stimulus is delivered to only one neuron (inset), lowering of the
junctional current is seen only immediately after stimulation. The gap junctions do not
stay closed over time, and the junctional current recovers by 30 min.
C) Summary graph comparing the percent change in junctional current. A Ca2+
rise in
both neurons reduces the junctional current. When Ca2+
is elevated in only one cell, the
inhibitory effect is smaller and returns to baseline by 15 min. The difference between the
stimulation conditions is significant immediately and 4 min after the train (two-tailed
unpaired Student’s t-test).

53
mV, the presynaptic neuron was then stepped to -90 mV, while clamping the second
neuron at -60 mV. The resulting junctional current was recorded before and after the
addition of Ca2+
-liberating drugs or DMSO (the vehicle). Introduction of 20 μM CPA
(n=6), an inhibitor of the endoplasmic reticulum Ca2+
-ATPase that depletes Ca2+
(Seidler
et al. 1989), produced no appreciable change in junctional current (Figure 16A, gray) as
compared to control, prior to CPA (Figure 16A, black). FCCP, a protonophore that
collapses the mitochondrial membrane potential and causes Ca2+
to exit (Heytler and
Prichard 1962), was applied at 100 nM (n=4), 1 (n=6) or 3 μM (n=4). Junctional current
was inhibited in a concentration-dependent fashion (Figure 16B) and was significant at
the two higher doses of FCCP in comparison to DMSO (Figure 16C).
Sustained inhibition of electrical synapses depends on intracellular Ca2+
To confirm whether the prolonged decrease in coupling after the train-stimulus is
due to changes in intracellular Ca2+
, the Ca2+
buffering capacity of the pipette solution
was enhanced and junctional current studied before and after the train-stimulus. This was
achieved by replacing standard intracellular saline containing 5 mM EGTA and 300 nM
free Ca2+
to one with either 20 mM EGTA and 35 nM free Ca2+
or 40 mM EGTA and 15
nM free Ca2+
. Neurons were voltage-clamped at -60 mV and dialyzed for 30 min. A
voltage step to -90 was given to the presynaptic neuron which in turn resulted in
junctional current in the coupled partner. In control, where standard intracellular saline
was in the pipettes (n=10), the train-stimulus of 75-ms depolarizing pulses at 5 Hz for 1
min followed by 1 Hz for 5 min, produced an ~15% decrease in junctional current that
lasted for at least 15 min (Figure 17A). Subsequent to a 30-min dialysis with high-EGTA

54
Figure 16. Depletion of mitochondria Ca2+
, but not endoplasmic reticulum Ca2+
,
inhibits junctional communication.
A) Neurons are voltage-clamped at -60 mV and dialyzed for 30 min with 0 mM EGTA
and no added Ca2+
in the intracellular saline. Inset, a voltage step to -90 mV is given to
neuron 1 (V1) to evoke junctional current in neuron 2 (I2). Liberating endoplasmic
reticulum Ca2+
with 20 μM CPA, an inhibitor of the endoplasmic reticulum Ca2+
-ATPase,
does not appreciably alter the junctional current (gray) as compared to control, prior to
CPA (black).
B) Addition of 3 μM FCCP to the bath collapses the mitochondrial membrane potential
and results in a Ca2+
leak into the cytosol from this organelle. This inhibits junctional
current 10 min after FCCP is applied. The FCCP-induced inhibition is dose-dependent, as
lower concentrations (1 μM and 100 nM) have a progressively weaker effect on the
electrical synapse. The time base applies to all three sets of traces.
C) Summary data of the mean change from control in junctional current following
DMSO (the vehicle), CPA or FCCP. The junctional current is significantly reduced by
higher concentrations of FCCP, but not CPA, vs. DMSO (ANOVA; Dunnett multiple
comparisons test).

55
Figure 17. Buffering Ca2+
with high intracellular EGTA prevents uncoupling.
A) Recording of junctional current between a pair of electrically-coupled bag cell
neurons dialyzed for 30 min with intracellular saline containing 5 mM EGTA and 300
nM free Ca2+
. Insets, both neurons are held at -60 mV and a voltage step to -90 mV is
given to neuron 1. The diagram shows a pair of neurons with stimulation to both pipettes.
The resulting junctional current (I2) shows a decrease after Ca2+
influx induced by
delivering the 5-Hz, 1-min then 1-Hz, 5-min train in both cells (stim). This inhibition of
junctional communication lasts for up to 15 min. The scale bars apply to both traces.
B) When the standard intracellular saline in the recording pipettes is replaced with one
containing 40 mM EGTA and 15 nM free Ca2+
, elevation of intracellular Ca2+
with the
train-stimulus fails to inhibit junctional current.
C) Summary graph comparing the junctional current after Ca2+
influx in pairs dialyzed
with a high (40 mM or 20 mM) vs. standard (5 mM) concentration of EGTA. The
prevention of the Ca2+
-induced decrease in junctional current with 40 mM EGTA is
significant compared to standard EGTA (ANOVA; Dunnett multiple comparisons test).

56
intracellular saline (40 mM EGTA), the same stimulus failed to induce a reduction in
junctional current and showed an ~5% increase after 15 min (Figure 17B, n=6). To a
lesser degree, 20 mM EGTA (n=5) prevented the uncoupling effect of stimulation, with
only an ~5% reduction in junctional current (Figure 17C). As summarized in Figure 17C,
eliminating the change in intracellular Ca2+
by adding more EGTA, prevented Ca2+
-
induced uncoupling in a statistically significant manner.
CaM kinase is involved in Ca2+
-induced uncoupling
A previous description of bag cell neurons shows that CaM-kinase is activated
during the afterdischarge (DeRiemer et al. 1984). Activation of CaM-kinase II in mouse
astrocyte (De Pina-Benabou et al. 2001), and goldfish Mauthner cells (Pereda et al. 1998)
has been reported to increase junctional conductance. To determine whether CaM-kinase
may participate in the regulation of bag cell neuron electrical synapses, KN-62, a CaM-
kinase II antagonist (Tokumitsu et al. 1990) was employed. Pairs were voltage-clamped
and dialyzed with standard intracellular saline for 30 min. From a holding potential of -60
mV, a voltage change to -90 mV given to the presynaptic neuron, while keeping the
postsynaptic cell at -60 mV, elicited junctional current. As expected, the train-stimulus in
neurons treated with DMSO (n=9) (the vehicle) for 15 min prior to recording (n=9),
produced an ~20% decrease in junctional current (Figure 18A). A similar inhibition of
junctional current after the train-stimulus was observed when neurons were stimulated in
10 μM concentration of KN-62 (n=5) (Figure 18B). However, 50 μM of KN-62 (n=6)
prevented the uncoupling effect of stimulation and showed an ~5% increase (Figure
18C). This lowering of the Ca2+
-induced uncoupling effect was statistically significant in
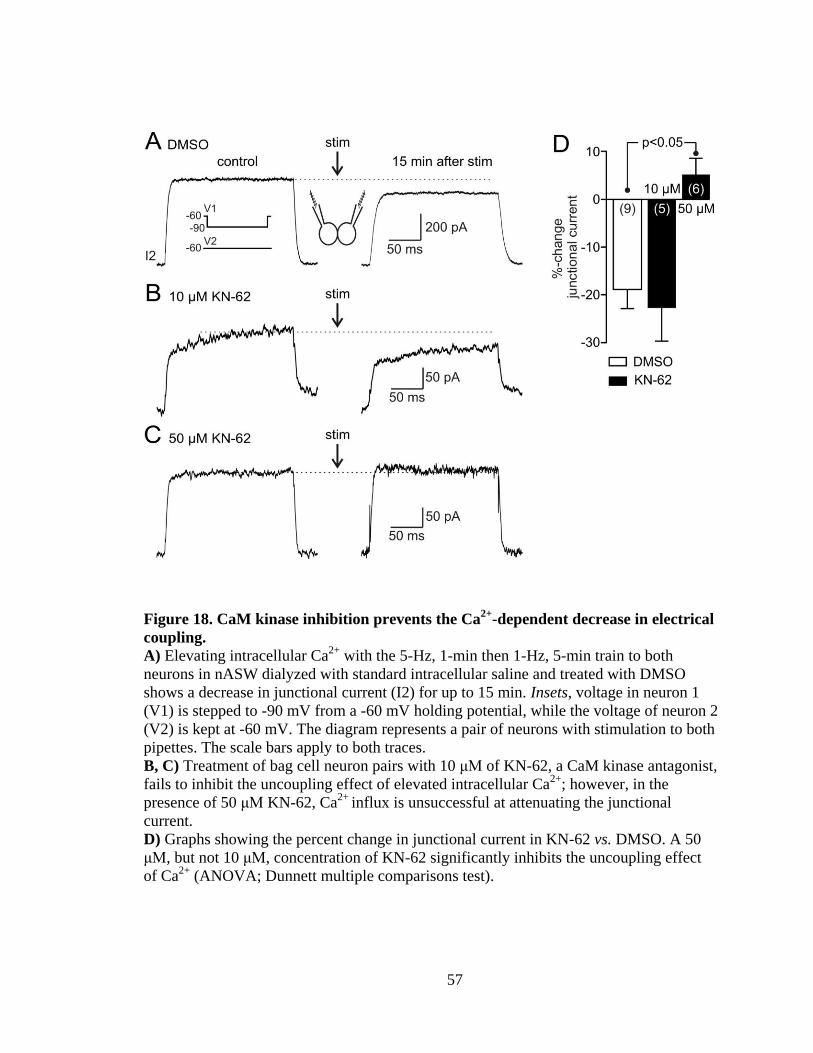
57
Figure 18. CaM kinase inhibition prevents the Ca2+
-dependent decrease in electrical
coupling.
A) Elevating intracellular Ca2+
with the 5-Hz, 1-min then 1-Hz, 5-min train to both
neurons in nASW dialyzed with standard intracellular saline and treated with DMSO
shows a decrease in junctional current (I2) for up to 15 min. Insets, voltage in neuron 1
(V1) is stepped to -90 mV from a -60 mV holding potential, while the voltage of neuron 2
(V2) is kept at -60 mV. The diagram represents a pair of neurons with stimulation to both
pipettes. The scale bars apply to both traces.
B, C) Treatment of bag cell neuron pairs with 10 μM of KN-62, a CaM kinase antagonist,
fails to inhibit the uncoupling effect of elevated intracellular Ca2+
; however, in the
presence of 50 μM KN-62, Ca2+
influx is unsuccessful at attenuating the junctional
current.
D) Graphs showing the percent change in junctional current in KN-62 vs. DMSO. A 50
μM, but not 10 μM, concentration of KN-62 significantly inhibits the uncoupling effect
of Ca2+
(ANOVA; Dunnett multiple comparisons test).

58
comparison to control (Figure 18D).
Activation of PKC reduces the junctional current
Shortly after the onset of the afterdischarge, PKC activity is upregulated and stays
elevated throughout (Conn et al. 1989a; Wayne et al. 1999). PKC activation has been
shown to regulate electrical coupling in different cells (Peracchia 1987; Saez et al. 1990;
Spray and Burt 1990; Lampe et al. 2000). The effect of the phorbol ester, PMA, an
effective PKC activator in bag cell neurons (Castagna et al. 1982; Sossin et al. 1994) was
tested on electrical coupling. Neurons were voltage-clamped at -60 mV for 30 min and a
voltage step to -90 in one neuron generated junctional current in the second cell. Addition
of DMSO (the vehicle) to pairs of coupled neurons (n=4) did not appreciably alter the
junctional current (Figure 19A, middle) as compared to the initial response (Figure 19A,
left). Subsequent delivery of the train-stimulus to these neurons resulted in an ~25%
inhibition (Figure 19A, right). In contrast, introducing 100 nM PMA to electrically-
coupled neurons (n=13) slightly decreased the junctional current by ~15% (Figure 19B,
middle, left). Moreover, the combination of PMA and the train-stimulus was more
effective at lowering electrical coupling than PMA alone, and caused an ~35% decrease
in junctional current (Figure 19B, right). Summary data showed a significant difference
in junctional current between control pairs and those treated with PMA. PMA-induced
uncoupling appeared to synergize with Ca2+
influx (Figure 19C).
Elevation of intracellular Ca2+
decreases coupling coefficient
To test the effects of Ca2+
influx on membrane potential, current clamp recording
was made from electrically-coupled bag cell neurons. The coupling strength was

59
Figure 19. PKC activation inhibits gap junctions and synergizes with Ca2+
influx.
A) Coupled bag cell neurons in nASW dialyzed for 30 min with standard intracellular
saline. Inset, neurons are voltage-clamped at -60 mV and a voltage step to -90 is
delivered to one cell (V1). Exposure of a control pair to DMSO for 40 min (left) does not
cause an appreciable change in junctional current (I2) (middle). Ca2+
influx (stim,
diagram represents a pair of neurons with stimulation to both pipettes) produced by a 5
Hz, 1 min then 1 Hz, 5 min train of stimulation to both cells reduces the junctional
current (right). The scale bars apply to all traces.
B) Introduction of 100 nM of the PKC activator, PMA, modestly suppresses the
junctional current. Elevating intracellular Ca2+
in these neurons with stimulation inhibits
the gap junction to an even greater extent.
C) Summary data compares the mean percent change in junctional current following
DMSO or PMA. There is a significant difference between DMSO and neurons in which
PKC is triggered. The inhibitory effect of PMA is also significantly larger when Ca2+
influx occurs (two-tailed unpaired Student’s t-test).

60
assessed by measurement of coupling coefficient. Neurons were dialyzed in nASW for 30
min with standard intracellular saline. They next were current clamped at -60 mV and a
1500-ms current step of -250 pA given to neuron 1. This induced a large
hyperpolarization in neuron 1 and a voltage transfer to neuron 2 (Figure 20A, left). In
control pairs (n=6), where neurons were not given the train-stimulus, a small ~10%
increase in coupling coefficient was observed (Figure 20A, right). In contrast, the
experimental group, which was first switched to voltage clamp and given the train-
stimulus (n=9), showed an ~15% decrease in electrical coupling (Figure 20B) upon return
to current clamp. The difference between the two groups was statistically significant
(Figure 20C).
Intracellular Ca2+
rise decreases the amplitude and time course of ETPs
Electrical coupling appears to mediate synchronous action potential firing of bag
cell neurons throughout the afterdischarge (Blankenship and Haskins 1979; Brown et al.
1989). We sought to test the impact of Ca2+
influx on the ability of presynaptic action
potentials to evoke ETPs. Action potential-like waveforms were generated in one neuron
under voltage-clamp and the ETP was recorded in the second cell in current-clamp.
Idealized action potentials, based on spikes recorded in prior work from our lab, were
favoured over actual spikes, so to avoid changes to the waveform that can occur
following repetitive stimulation and Ca2+
influx (unpublished observation). In control
pairs (n=5), a series of voltage ramps from -60, mimicking the action potential, was
delivered at 1 Hz to the presynaptic neuron (Figure 21A, lower), which in turn elicited an
ETP in the coupled partner current-clamped at -60 mV (Figure 21A, upper-black).

61
Figure 20. Coupling coefficient decreases after an intracellular Ca2+
rise.
A) In current clamp, neurons are dialyzed in nASW with standard intracellular saline.
Inset, negative current is injected to neuron 1 (I1) while neuron 2 current (I2) is kept
constant. A -250 pA current injected in neuron 1 for 1500 ms induces a large
hyperpolarization (V1) that transfers to the coupled neuron (V2). The hyperpolarizing
response in both neurons does not appreciably change over time (right traces) in
comparison to control (left traces). The scale bars apply to both left and right tarces.
B) When neurons are given the train-stimulus (75-ms depolarizing pulses at 5 Hz for 1
min, then 1 Hz for 5 min), current injection causes a smaller hyperpolarizing response
(right traces) as compared to prior to stimulation (left traces).
C) Coupling coefficient, determined by the ratio of V2/V1 at the steady state membrane
potential after hyperpolarizing neuron 1, shows a significant decrease in pairs given the
train-stimulus (two-tailed unpaired Student’s t-test).

62
Figure 21. Elevated intracellular Ca2+
reduces the size and time course of the ETP.
A) Recording from a pair of electrically-coupled neurons dialyzed for 30 min with
standard intracellular saline in nASW. From a holding potential of -60 mV, an idealized
action potential waveform (lower) (composed of four ramps to +10 (20 ms), -10 (15 ms),
-45 (15 ms), and then -60 mV (20 ms)) delivered to neuron 1 (V1) at 1 Hz elicits an ETP
in neuron 2 (V2) (black trace - 30 min dialysis) recorded in current-clamp. The amplitude
of the ETP does not appreciably change over time (gray trace - 35 min). Inset, neuron 1
is voltage clamped and the second neuron is recorded under current clamp. The scale bars
apply to both traces.
B) The action potential waveform in neuron 1 (lower) produces an ETP in neuron 2
(black trace - before). Simultaneously delivering to both cells the train-stimulus (5 Hz, 1
min then 1 Hz, 5 min) causes a smaller ETP that recovers faster (gray trace - after)
recorded immediately after the train-stimulus.
C, D) Summary graphs comparing the mean change in ETP area and time-to-recovery in
neuron 2 evoked by an action potential waveform in neuron 1 shows significant decreases
after Ca2+
influx as compared to control groups with no stimulation (two-tailed unpaired
Student’s t-test).

63
The ETP did not change over time (Figure 21A, upper-gray); however, when neurons
were given the train-stimulus (n=5), under voltage-clamp to increase the intracellular
Ca2+
, the same action potential waveform in the presynaptic neuron (Figure 21B, lower)
produced a smaller and faster ETP in the postsynaptic neuron, recorded immediately after
stimulation (Figure 21B, upper-gray) as compared to the initial ETP (Figure 21B, upper-
black). The area of the ETP as well as the time required to recover from the peak
response to steady-state were quantified and showed a significant decrease in stimulated
vs. the unstimulated control (Figure 21C, D).

64
Chapter 4
Discussion
Electrical synapses allow for current to pass directly from cell to cell, thus
providing a means for the membrane potential of a neuron to immediately and reliably
influence that of neighbouring neurons (Bennett and Zukin 2004; McCracken and
Roberts 2006). The characteristics of bag cell neuron communication are consistent with
an electrical synapse - presumably mediated by gap junctions. In the intact cluster, this
would promote firing synchrony during the afterdischarge, leading to en masse secretion
of egg-laying hormone and the initiation of reproduction (Dudek et al. 1980; Stuart et al.
1980; Michel and Wayne 2002; Hatcher and Sweedler 2008).
Cultured bag cell neurons appear to form electrical, rather than chemical,
synapses. In voltage-clamp, presynaptic depolarization or hyperpolarization results in a
near-simultaneous flow of current from pre- to postsynaptic neuron. In current-clamp,
presynaptic depolarization or hyperpolarization causes a concomitant change in
postsynaptic membrane potential. Chemical transmission would present Ca2+
-sensitivity,
and the change in postsynaptic voltage or current would occur at a fixed latency
following the presynaptic stimulus (Katz and Miledi 1967; Berry and Pentreath 1976;
Llinas et al. 1981). However, as expected for electrical transmission, the amplitude and
time course of the ETP is not altered when extracellular Ca2+
is removed; furthermore,
the onset of the ETP occurs during the rising phase of the presynaptic action potential,
rather than after the peak (Bennett 1977, 2000). There are two prior studies on cultured
bag cell neuron electrical synapses (Kaczmarek et al. 1979; Lin and Levitan 1987).

65
Although less quantitative than the present study, they do show presynaptic
hyperpolarization and action potentials eliciting postsynaptic hyperpolarization and
ETPs/spikes, respectively.
The pharmacological profile of the bag cell neuron junctional current also points
to an electrical synapse. All of the arylaminobenzoates inhibited the junctional current,
with a combination of NFA and NPPB being the most effective, including blocking
electronic transmission in the intact cluster. These drugs inhibit electrical synapses
between hepatoma or neuroblastoma cells transfected with various connexin genes, as
well as horizontal cells in the retina and connexin hemichannels in oocytes (Harks et al.
2001; Eskandari et al. 2002; Srinivas and Spray 2003; Pan et al. 2007). Certain long-
chain alcohols, like heptanol or octanol, also block electrical transmission and gap
junctions in many species (Johnston et al. 1980; Juszczak and Swiergiel 2009). However,
we did not test these alcohols because earlier reports indicated they were ineffective on
coupled L14 Aplysia ink motor neurons in situ as well as pairs of coupled Aplysia buccal,
cerebral, or pleural neurons in vitro (Carew and Kandel 1977; Bodmer et al. 1988;
Carrow and Levitan 1989). Those results, along with our finding that neither
glycyrrhetinic acid nor quinine block the bag cell neuron electrical synapse, may suggest
that Aplysia gap juction proteins (Apl-innexins) are distinct from innexins in Drosophila,
crayfish, or Limax, which are inhibited by alcohols or glycyrrhetinic acid (Johnston et al.
1980; Ermentrout et al. 2004; Cao and Nitabach 2008). The original Aplysia neuronal
transcriptome yielded nine predicted Apl-innexin genes (Moroz et al. 2006), while our
PCR-based cloning indicates 20 full-length Apl-innexins (Carter and Magoski 2014).

66
Thus, the molecular substrate for gap junctions is present, and differences in protein
structure or assembly could account for the distinct pharmacological and biophysical
profile of the bag cell neuron electrical synapse.
Although dye-coupling is often used as a criterion for the presence of electrical
synapses (Stewart 1978; Peinado et al. 1993), we did not detect transfer of either of the
polar dyes, sulforhodamine 101 or carboxyfluorescein, as well as the uncharged tracer,
biocytin. The latter is somewhat unexpected, given that biocytin usually crosses gap
junctions when other tracers fail (Ewadinger et al. 1994). Dye-coupling is also not
observed between Aplysia L14 neurons (Bodmer et al. 1988) or Caenorhabditis myocytes
(Liu et al. 2006), although other invertebrate neurons will dye-couple, including those
from the pulmontate molluscs, Lymnaea and Helisoma (Murphy et al. 1983; Ewadinger
et al. 1994), as well as crayfish (Payton et al. 1969) and leech (Fan et al. 2005). This
notwithstanding, our postsynaptic K+ channel block experiments imply that Cs
+, but not
TEA, permeates the bag cell neuron gap junction. The ionic radius of TEA is two to three
times that of Cs+, which in turn is only slightly larger than K
+, the likely dominant
endogenous charge carrier (Hille 2001). Compared to other connexins and innexins
(Descheˆnes and Bennett 1974; Verselis et al. 1986; Beblo and Veenstra 1997), bag cell
neuron junctional channels appear to be more restrictive.
The junctional current between cultured bag cell neurons is independent of both
junctional voltage and the postsynaptic membrane potential. This is also the case for
coupled Aplysia L14 neurons and paired buccal, cerebral, or pleural neurons (Bodmer et
al. 1988; Carrow and Levitan 1989). Many vertebrate gap junctions have some form of

67
voltage-dependence (Spray et al. 1979; Bukauskas and Verselis 2004), as do some leech
innexins (Dykes et al. 2004) and electrical synapses in Chironomus (Obaid et al. 1983).
However, closure of certain connexins, such as Cx26, does not occur until the junctional
voltage is altered by >75 mV. The presynaptic voltage changes used in the present study
are at the physiological limits of resting potential (-90 mV through to 0 mV); still, it is
conceivable that if we extended the range to non-physiological voltages, the junctional
current may display voltage-dependence. The bag cell neuron electrical synapse also does
not rectify, as all assays consistently show current or voltage transmitting equally well
from neuron 1 to neuron 2, and vice versa. Rectification is absent from most coupled
molluscan neurons, including Aplysia (Bodmer et al. 1988; Carrow and Levitan 1989),
Lymnaea (Wildering et al. 1991) and the nudibranch, Tritonia (Getting and Willows
1974).
Although the bag cell neuron electrical synapse is very reliable, with no observed
failures, the low-pass filter function does limit transmission. This is inherent to most gap
junctions, and manifests as a reduction in any presynaptic voltage change as it passes to
the postsynaptic neuron, particularly for rapid or higher-frequency signals (Bennett 1977,
2000). In bag cell neurons, the time needed for charging the postsynaptic capacitance
draws out and dampens the ETP; hence, a 70-90 mV magnitude presynaptic action
potential renders an ∼10 mV ETP with a peak-to-peak latency of ∼15 ms. However, at
steady-state, low-pass filtering is less consequential, and the 20-30 mV chronic
depolarization associated with the afterdischarge likely transfers effectively between
neurons (Kupfermann and Kandel 1970; Brown et al. 1989). From these depolarized

68
membrane potentials, spiking in one neuron has a greater tendency to evoke spiking in a
coupled partner.
Firing synchrony is a key feature of the afterdischarge, and since early
descriptions of bag cell neuron function, it has been asserted that electrical coupling
underlies this phenomenon (Kupfermann and Kandel 1970; Blankenship and Haskins
1979). The gap junction blockers, NFA and NPPB (Harks et al. 2001; Srinivas and Spray
2003), largely prevent afterdischarge generation in the intact bag cell neuron cluster.
However, despite our observing little-to-no side-effects for NFA and NPPB on whole-cell
conductance, there are reports of arylaminobenzoates, more so for flufenamic acid (which
we did not employ here) and McFA, rather than NFA or NPPB, opening potassium
channels, releasing intracellular Ca2+
, or blocking cation and Cl- channels (Gögelein and
Pfannmüller 1989; White and Aylwin 1990; Poronnik et al. 1992; Takahira et al. 2005;
Gardam et al. 2008). While we find no evidence of such alterations by NFA and/or
NPPB, these reagents could have undefined effects in addition to acting on gap junctions.
The voltage-independent nature of the bag cell neuron electrical synapse
translates into the junction being open at rest, which would allow current to spread
through the network from neuron(s) receiving excitatory acetylcholine synapses (White
and Magoski 2012). Spatial spread of excitation within the cluster was postulated by
Brown and Mayeri (1989), who envisaged electrical coupling acting as part of a positive-
feedback mechanism promoting the afterdischarge. Similarly, electrical synapses are
required to distribute synaptic depolarization and drive firing during synchronous
afterdischarges in prey-capture motor neurons of the gymnosomatan mollusc, Clione

69
(Norekian 1999), while electrical synapses mediate regenerative excitation of trigger-
group neurons, that burst simultaneously to control swimming in Tritonia (Getting and
Willows 1974). If the cholinergic input that initiates the afterdischarge (White and
Magoski 2012) is limited to a select few bag cell neurons, then blocking gap junctions
may occlude the spread of excitation and, as we saw for most preparations, result in a
paucity of firing.
Coupling in the intact bag cell neuron cluster is quite low, as we find coupling
coefficients of ∼0.015, which is only twice that reported by Blankenship and Haskins
(1979) in A. dactylomela. The difference between culture and the intact cluster likely
reflects electrotonically-remote gap junctions (Blankenship and Haskins 1979), and that
bag cells are coupled to more than one partner in situ (Kaczmarek et al. 1979).
Nevertheless, strong coupling is not necessarily required for synchrony; similar to what
we report, various networks of rat cortical or thalamic neurons show synchrony despite
presenting very low coupling coefficients (Gibson et al. 1999; Traub et al. 2001;
Landisman et al. 2002). Bag cell neuron secretion requires relatively lengthy periods of
activity (Michel and Wayne 2002; Hickey et al. 2013), and the synchrony promoted by
electrical coupling may ensure that all neurons spike sufficiently for optimal release of
reproductive hormone.
Cytosolic Ca2+
plays a vital role in the regulation of fundamental cellular
processes; such as channel gating, excitability and secretion (Hille 2001). Elevated
intracellular Ca2+
decreases coupling in many cell types, including excitable and non-
excitable cells (Spray and Bennett 1985; Saez et al. 1989; Peracchia 2004; Rohr 2004;

70
Sohl and Willecke 2004). For neurons, Ca2+
entry from voltage-gated Ca2+
channels, as a
function of action potentials magnitude and duration, is the primary means to increase
cytosolic Ca2+
(Klein and Kandel 1978; Hung and Magoski 2007; Catterall and Few
2008). In bag cell neurons, delivery of the 5 Hz, 1 min then 1 Hz, 5 min stimulus mimics
the fast and early slow phases of the afterdischarge, and leads to an immediate and
prominent voltage-gated Ca2+
influx (Geiger and Magoski 2008). This Ca2+
rise inhibits
the bag cell neuron electrical synapse, and is ineffective in a Ca2+
-free medium,
suggesting Ca2+
entry as the cause of uncoupling. This is consistent with Chanson et al.
(1999), who showed that Ca2+
influx associated with a reduction in junctional current is
inhibited in acinar cells after removing external Ca2+
. Similarly, activation of Ca2+
channels after exposure of hepatoma cell pairs to arachidonic acid causes a rapid
inhibition of junctional current (Lazrak et al. 1994). Furthermore, we showed that a high-
EGTA intracellular saline eliminates the uncoupling, supporting the role of intracellular
Ca2+
at inhibiting bag cell neuron electrical synapses. Similarly, a Ca2+
increase-induced
decrease in junctional conductance between hepatoma cells is prevented by the Ca2+
buffer, BAPTA (Lazrak et al. 1994).
While it is clear that Ca2+
modulates gap junctions, there are contradictory
findings on whether a Ca2+
rise in only one cell is able to travel to the other side of the
junction. In coupled rat hepatocytes, Ca2+
imaging shows that injection of either IP3 or
Ca2+
in one cell increases Ca2+
in the adjacent cell (Saez et al. 1989). Conversely, Rose
and Loewenstein (1976) found that injection of Ca2+
in a salivary gland cell of
Chironomus does not diffuse to the adjacent cell, as imaged using aequorin. Moreover, a

71
Ca2+
increase in only one cell did not induce detectable uncoupling (Rose and
Loewenstein 1976). This has parallels with our findings that a synchronous train-stimulus
in both neurons of a pair of bag cell neurons is needed to chronically inhibit junctional
communication. In those neurons where we stimulated just one cell, it is likely that Ca2+
reaches the junctional region but induces only partial uncoupling; however, it is not clear
whether Ca2+
diffuses through the junction.
Ca2+
release from intracellular stores can also inhibit electrical coupling. In
Chironomus salivary gland cells, cyanide, a mitochondrial toxin that blocks oxidative
metabolism and releases Ca2+
(Carafoli 1967), suppresses junctional communication
(Rose and Loewenstein 1975). Also, blocking the ability of the endoplasmic reticulum to
pump Ca2+
with thapsigargin (Thastrup et al. 1990) substantially reduces gap junction
communication in pancreatic acinar cells (Chanson et al. 1999). In bag cell neurons,
CICR also contributes to Ca2+
dynamics, particularly during the slow phase of the
afterdischarge (Fisher et al. 1994; Geiger and Magoski 2008). On entry, Ca2+
is taken up
by the mitochondria through the Ca2+
uniporter and is then released though cation
exchangers. This released Ca2+
stimulates ryanodine receptors on the endoplasmic
reticulum to produce further Ca2+
release (Geiger and Magoski 2008). We directly
investigated the role of mitochondrial and endoplasmic reticulum Ca2+
on bag cell neuron
electrical synapses. Eliminating the ability of mitochondrial to clear intracellular Ca2+
with the protonophore, FCCP, caused a substantial and dose-dependent decrease in
junctional current. However, endoplasmic reticulum Ca2+
depletion with the pump
inhibitor, CPA, did not appreciably reduce the junctional current. This is consistent with

72
prior work in our lab showing that mitochondrial, but not endoplasmic reticulum, Ca2+
has preferential access to both gating a plasma membrane cation channel and evoking
neuropeptide secretion (Hickey et al. 2010, 2013). That stated, depletion of endoplasmic
reticulum Ca2+
does exclusively activate a Ca2+
influx pathway in cultured bag cell
neurons (Kachoei et al. 2006).
The possibility that the Ca2+
-induced reduction in coupling we observed is a
secondary effect of cytoplasmic acidification or that elevated Ca2+
might synergize with
acidosis has been suggested for several cell types. For example, Ca2+
and H+ by
themselves do not block electrical coupling between rat myocardial cells, but coupling
significantly decreases when acidosis is accompanied by Ca2+
elevation (Burt 1987).
Furthermore, CO2-induced cytosolic acidification decreases electrical coupling in the
presence of a normal concentration of Ca2+
, whereas removing Ca2+
from the
extracellular solution prevents this effect (White et al. 1990). In contrast, intracellular
acidification reduces electrical coupling in Xenopus embryos, with no change in the
intracellular Ca2+
concentration (Rink et al. 1980). Similarly, the Ca2+
-dependent decrease
in junctional communication between crayfish septate axons is related to an intracellular
Ca2+
increase rather than low pH (Peracchia 1990). Also, in astrocytes, Ca2+
decreases the
permeability of gap junctions, but is not potentiated by intracellular acidosis (Cotrina et
al. 1998). For bag cell neurons, there is evidence to rule out the possibility of H+
participation in uncoupling: First, Ca2+
influx induced by pharmacological activation of a
store-operating pathway does not change intracellular pH in these cells (Knox et al.
2004). Second, in the present study, intracellular dialysis with the Ca2+
chelator, EGTA,

73
eliminates the uncoupling effect of Ca2+
in a concentration-dependent fashion. Third, we
also showed here that a Ca2+
/CaM-dependent protein kinase blocker prevents Ca2+
-
induced uncoupling.
Ca2+
might not always interact directly with gap junctions, but rather via a
cytoplasmic intermediate. CaM, a ubiquitous Ca2+
-binding protein known to regulate ion
channels (Levitan 1999), is found to be a mediator of gap junction regulation in many
systems. This was first suggested in crayfish axons (Johnston and Ramon 1981), and
subsequently, demonstrated at both vertebrate (Peracchia et al. 1983; Peracchia 1984;
Toyama et al. 1994) and invertebrate (Peracchia 1987) electrical synapses. CaM can
activate CaM kinase which is known to regulate, though phosphorylation, the activity of
proteins involved in many cellular process (Hanson and Schulman 1992; Enslen and
Soderling 1994; Lledo et al. 1995; Pereda et al. 1998). However, CaM kinase as a
regulator of gap junctions is less well-defined. In fact, CaM kinase phosphorylation
increases junctional conductance in goldfish Mauthner neurons (Pereda et al. 1998),
mouse astrocytes (De Pina-Benabou et al. 2001), and neuroblasma cells expressing rat
Cx36 (Del Corsso et al. 2012). DeRiemer et al. (1984) found CaM kinase in Aplysia is
homologous to mammals, and may mediate the long lasting effect of intracellular Ca2+
entry during the afterdischarge. Application of the CaM blockers calmidazolium,
trifluoperazine, or chlorpromazine, inhibits both Aplysia CNS Ca2+
-CaM-dependent
kinase and the ability to fire an afterdischarge (DeRiemer et al. 1984. 1985). The CaM
kinase antagonist, KN-62, also attenuates a prolonged depolarization in cultured bag cell
neurons evoked by a train-stimulus (Hung and Magoski 2007).

74
We have shown here that CaM kinase may have a role in the Ca2+
-modulation of
the bag cell neuron electrical synapse, based on KN-62 inhibiting the uncoupling effect of
Ca2+
. Electrical uncoupling induced by Ca2+
is believed to be associated with a decrease
in junctional thickness and a reduction in gap junctional particle size (Peracchia and
Dulhunty 1976; Bernardini and Peracchia 1981). These conformational changes may
occur in a phosphorylation-dependent fashion (Moreno et al. 1994). Potential
phosphorylation sites for various kinases have been identified in many connexins
(Cruciani and Mikalsen 2002; Giepmans 2004; Laird 2005; Solan and Lampe 2005) and
to a lesser extent in innexins (Potenza et al. 2002; Bauer et al. 2005). Phosphorylation
can change the unitary conductance, open probability, or the number of gap junction
channels (Moreno and Lau 2007). CaM kinase phosphorylates Cx32 on serine residues
(Saez et al. 1990) and Cx36 specifically on serine 110 and threonine 111 (Alev et al.
2008). We propose that a combination of free intracellular Ca2+
and protein
phosphorylation contributes in regulating bag cell neuron intercellular communication.
During the slow phase of the afterdischarge, PKC turns on and stays elevated
throughout (Conn et al. 1989a; Wayne et al. 1999). Tam et al. (2009) showed that the
intracellular Ca2+
rises through PKC-dependent upregulation of Ca2+
current, which
promotes bag cell neuron depolarization to sustain the afterdischarge. PKC activation
enhances voltage-gated Ca2+
currents and increases the overall amount of Ca2+
during the
afterdischarge (DeRiemer et al. 1985). Application of phorbol esters, such as PMA, to
activate PKC, enhances the amplitude of action potentials by increasing Ca2+
current in
bag cell neurons, whereas PKC inhibitors prevent this effect (Conn et al. 1989b). PMA

75
has been used to activate Aplysia PKC, including in biochemical assays of nervous
system ganglia, as well as bag cell neurons in the intact cluster or primary culture
(DeRiemer et al. 1985; Sossin and Schwartz 1992; Sossin et al. 1994; Tam et al. 2011).
In this study, PMA inhibited electrical coupling, supporting the involvement of PKC in
the regulation of bag cell neuron gap junctions. The ability of PKC to close gap junctions
has been shown extensively in a number of cell types (Spray and Bennett 1985; Lampe et
al. 2000; Cruciani et al. 2001). However, there are cases where PMA causes a gradual
increase in junctional conductance (van Veen et al. 2000) or no change (Spray and Burt
1990). The inhibitory effect in bag cell neurons was even greater when PMA-treated cells
were given the train-stimulus to increase intracellular Ca2+
. The synergy between Ca2+
and PKC to decrease electrical coupling may serve to elevate the overall excitability by
increasing system input resistance in the intact cluster.
Alternatively, a reduction in coupling may, somewhat paradoxically, promote
synchrony during the afterdischarge. Ca2+
influx decreases the amplitude and time course
of ETPs produced by idealized presynaptic action potentials. The low-pass filtering effect
of the electrical synapse sustains the ETP well beyond the presynaptic spike. A modest
decrease in junctional coupling may improve synchrony by preventing lengthy ETPs and
out-of-phase postsynaptic action potentials. In acinar pancreatic cells, Ca2+
-induced
uncoupling leads to enhanced hormone secretion (Chanson et al. 1998, 1999).
Furthermore, an increased ETP between electrically-coupled neuroendocrine cells of
cockroach is correlated to reduced juvenile hormone secretion (Lococo et al. 1986).
Asynchrony may be essential for ELH secretion from bag cell neurons, and a modest

76
decrease in electrotonic transmission could help to ensure in-phase spiking and
consistently-timed hormone release necessary for reproduction.
Burst synchrony is fundamental to the output of many different neurons. The
present study offers the prospect of examining burst coordination in neurons that trigger a
behaviour fundamental to animal survival; moreover, determining how bag cell neurons
synchronize action potentials has consequences for the general understanding of nervous
system.
Future Directions
The majority of the data for this study was gathered using cultured neurons paired
in the soma-soma configuration. To examine if the strength of electrical synapse is
sensitive to the contact location, I would plate neurons to make contact between somata,
short or long neurites and quantatively compare junctional current. Lesser junctional
conductance and weaker electrical coupling in neurite-to-neurite vs. soma-soma pairs
would support smaller gap junctions on distal bag cell processes in the intact cluster than
larger ones in somata reported by Kaczmarek et al. (1979).
As mentioned, there are limitations to studying electrical coupling between bag
cell neurons in culture. The coupling ratio between cultured bag cell neurons, on average,
is much higher than reported for the intact cluster (Blankenship and Haskin, 1979). In
addition, changes in Aplysia innexin expression as well as lack of endogenous growth
factors may alter the nature of electrical transmission in culture. More studies of bag cell
neuron electrical synapses in intact cluster would help to overcome these limitations.

77
To see if Ca2+
diffuses to the adjacent cell in a pair of bag cell neurons, Ca2+
imaging from a pair can be performed, but when the train-stimulus is applied to only one
neuron. This would reveal whether partial inhibition of electrical coupling between bag
cell neurons, when only one neuron is stimulated, is due to failure of Ca2+
passing
through the junction.
Delivery of just a 5-Hz, 1-min train-stimulus results in Ca2+
influx in cultured bag
cell neurons (Geiger and Magoski 2008), similar to what we used here, although less
lengthy. This can be tested in coupled neurons to show whether Ca2+
influx during only
the fast phase of the afterdischarge is sufficient to tonically inhibit junctional
communication.
At the onset of the afterdischarge, there is an increase in cAMP levels which is
involved in changes to excitability and broadening of action potentials (Kaczmarek et al.
1978; Conn and Kaczmarek 1989). Applying agents such as methylxanthine to inhibit
cAMP phosphodiesterases or cAMP activator, forskolin, to coupled bag cell neurons
could be used to examine the involvement of cAMP in electrical coupling. Given that
injection of cAMP analogs decreases K+ current in bag cell neurons and enhances action
potentials (Kaczmarek et al. 1980; Kaczmarek and Strumwasser 1984), cAMP inhibition
may increase electrical coupling.
The neuropeptide, FMRFamide, is known as a modulator of Aplysia nervous
system and is expressed in abdominal ganglion (Schaefer et al. 1985). FMRFamide
decreases the Ca2+
current and supresses the afterdischarge in bag cell neurons (Brezina
et al. 1987; Fisher et al. 1993). Considering the decreased Ca2+
current induced by

78
FMRFamide, addition of FMRFamide to electrically coupled neurons may enhance
junctional communication.

79
Literature cited
Acosta‐Urquidi J & Dudek FE (1981). Soma spike of neuroendocrine bag cells of Aplysia
californica. J Neurobiol 12, 367-378.
Alev C, Urschel S, Sonntag S, Zoidl G, Fort AG, Hoher T, Matsubara M, Willecke K,
Spray DC & Dermietzel R (2008). The neuronal connexin36 interacts with and is
phosphorylated by CaMKII in a way similar to CaMKII interaction with glutamate
receptors. Proc Natl Acad Sci USA 105, 20964-20969.
Anderson PA (1979). Ionic basis of action potentials and bursting activity in the
hydromedusan jellyfish Polyorchis penicillatus. J Exp Biol 78, 299-302.
Andrew RD, MacVicar BA, Dudek FE & Hatton GI (1981). Dye transfer through gap
junctions between neuroendocrine cells of rat hypothalamus. Science 211, 1187-1189.
Arch S & Smock T (1977). Egg-laying behavior in Aplysia californica. Behav Biol 19,
45-54.
Arch S (1972). Biosynthesis of the egg-laying hormone (ELH) in the bag cell neurons of
Aplysia californica. J Gen Physiol 60, 102-119.
Auerbach AA & Bennett MV (1969). A rectifying electrotonic synapse in the central
nervous system of a vertebrate. J Gen Physiol 53, 211-237.
Bauer R, Löer B, Ostrowski K, Martini J, Weimbs A, Lechner H & Hoch M (2005).
Intercellular communication: the Drosophila innexin multiprotein family of gap junction
proteins. Chem Biol 12, 515-526.
Bauer R, Lehmann C, Martini J, Eckardt F & Hoch M (2004). Gap junction channel
protein innexin 2 is essential for epithelial morphogenesis in the Drosophila embryo. Mol
Biol Cell 15, 2992-3004.
Baux G, Simonneau M, Tauc L & Segundo JP (1978). Uncoupling of electrotonic
synapses by calcium. Proc Natl Acad Sci USA 75, 4577-4581.
Bear MF, Connors BW & Paradiso MA (2007). Neuroscience: Exploring the brain. 3rd
edition. Lippincott Williams & Wilkins. Chapter 15, 481-509.
Beblo DA & Veenstra RD (1997). Monovalent cation permeation through the connexin40
gap junction channel. Cs, Rb, K, Na, Li, TEA, TMA, TBA, and effects of anions Br, Cl,
F, acetate, aspartate, glutamate, and NO3. J Gen Physiol 109, 509-522.

80
Belin V & Moos F (1986). Paired recordings from supraoptic and paraventricular
oxytocin cells in suckled rats: recruitment and synchronization. J Physiol 377, 369-390.
Bennett MVL & Zukin RS (2004). Electrical coupling and neuronal synchronization in
the mammalian brain. Neuron 41, 495-511.
Bennett MVL (2000). Seeing is relieving: electrical synapses between visualized neurons.
Nat Neurosci 3, 7-8.
Bennett MVL (1977). Electrical transmission: a functional analysis and comparison to
chemical transmission. Handbook of Physiology, The Nervous System, Cellular Biology
of Neurons. Chapter 11, 359-395.
Bennett MVL (1966). Physiology of electrotonic junctions. Ann N Y Acad Sci 137, 509-
539.
Bennett MVL, Barrio L, Bargiello T, Spray D, Hertzberg E & Saez J (1991). Gap
junctions: new tools, new answers, new questions. Neuron 6, 305-320.
Bennett MVL & Goodenough DA (1978). Gap junctions, electrotonic coupling, and
intercellular communication. Neurosci Res Program Bull 16, 1-486.
Bernardini G & Peracchia C (1981). Gap junction crystallization in lens fibers after an
increase in cell calcium. Invest Ophthalmol Vis Sci 21, 291-299.
Berry M & Pentreath V (1976). Criteria for distinguishing between monosynaptic and
polysynaptic transmission. Brain Res 105, 1-20.
Bissiere S, Zelikowsky M, Ponnusamy R, Jacobs NS, Blair HT & Fanselow MS (2011).
Electrical synapses control hippocampal contributions to fear learning and memory.
Science 331, 87-91.
Blankenship JE & Haskins JT (1979). Electrotonic coupling among neuroendocrine cells
in Aplysia. J Neurophysiol 42, 347-355.
Bodmer R, Verselis V, Levitan I & Spray D (1988). Electrotonic synapses between
Aplysia neurons in situ and in culture: aspects of regulation and measurements of
permeability. J Neurosci 8, 1656-1670.
Brezina V, Eckert R & Erxleben C (1987). Suppression of calcium current by an
endogenous neuropeptide in neurones of Aplysia californica. J Physiol 388, 565-595.

81
Brown R, Pulst S & Mayeri E (1989). Neuroendocrine bag cells of Aplysia are activated
by bag cell peptide-containing neurons in the pleural ganglion. J Neurophysiol 61, 1142-
1152.
Bukauskas FF, Kreuzberg MM, Rackauskas M, Bukauskiene A, Bennett MVL, Verselis
VK & Willecke K (2006). Properties of mouse connexin 30.2 and human connexin 31.9
hemichannels: implications for atrioventricular conduction in the heart. Proc Natl Acad
Sci USA 103, 9726-9731.
Bukauskas FF & Verselis VK (2004). Gap junction channel gating. Biochim Biophys
Acta 1662, 42-60.
Burt JM (1987). Block of intercellular communication: interaction of intracellular H+ and
Ca2+
. Am J Physiol 253, C607-12.
Cao G & Nitabach MN (2008). Circadian control of membrane excitability in Drosophila
melanogaster lateral ventral clock neurons. J Neurosci 28, 6493-6501.
Carafoli E (1967). In vivo effect of uncoupling agents on the incorporation of calcium
and strontium into mitochondria and other subcellular fractions of rat liver. J Gen Physiol
50, 1849-1864.
Carew TJ & Kandel ER (1977). Inking in Aplysia californica. III. Two different synaptic
conductance mechanisms for triggering central program for inking. J Neurophysiol 40,
721-734.
Carrow GM & Levitan IB (1989). Selective formation and modulation of electrical
synapses between cultured Aplysia neurons. J Neurosci 9, 3657-3664.
Carter CJ and Magoski NS. Electrical coupling in Aplysia: identification of innexin
genes. Can Assoc Neurosci Abstr 8: 1-B-24, 2014.
Castagna M, Takai Y, Kaibuchi K, Sano K, Kikkawa U & Nishizuka Y (1982). Direct
activation of calcium-activated, phospholipid-dependent protein kinase by tumor-
promoting phorbol esters. J Biol Chem 257, 7847-7851.
Catterall WA & Few AP (2008). Calcium channel regulation and presynaptic plasticity.
Neuron 59, 882-901.
Chang Q, Gonzalez M, Pinter MJ & Balice-Gordon RJ (1999). Gap junctional coupling
and patterns of connexin expression among neonatal rat lumbar spinal motor neurons. J
Neurosci 19, 10813-10828.

82
Chanson M, Fanjul M, Bosco D, Nelles E, Suter S, Willecke K & Meda P (1998).
Enhanced secretion of amylase from exocrine pancreas of connexin32-deficient mice. J
Cell Biol 141, 1267-1275.
Chanson M, Mollard P, Meda P, Suter S & Jongsma HJ (1999). Modulation of pancreatic
acinar cell to cell coupling during ACh-evoked changes in cytosolic Ca2+
. J Biol Chem
274, 282-287.
Conn PJ & Kaczmarek LK (1989). The bag cell neurons of Aplysia. Mol Neurobiol 3,
237-273.
Conn P, Strong J, Azhderian E, Nairn A, Greengard P & Kaczmarek LK (1989a). Protein
kinase inhibitors selectively block phorbol ester-or forskolin-induced changes in
excitability of Aplysia neurons. J Neurosci 9, 473-479.
Conn P, Strong J & Kaczmarek LK (1989b). Inhibitors of protein kinase C prevent
enhancement of calcium current and action potentials in peptidergic neurons of Aplysia. J
Neurosci 9, 480-487.
Cotrina ML, Kang J, Lin JH, Bueno E, Hansen TW, He L, Liu Y & Nedergaard M
(1998). Astrocytic gap junctions remain open during ischemic conditions. J Neurosci 18,
2520-2537.
Cruciani V, Husøy T & Mikalsen S (2001). Pharmacological evidence for system-
dependent involvement of protein kinase C isoenzymes in phorbol ester-suppressed gap
junctional communication. Exp Cell Res 268, 150-161.
Cruciani V & Mikalsen S (2002). Connexins, gap junctional intercellular communication
and kinases. Biol Cell 94, 433-443.
Dahl G & Isenberg G (1980). Decoupling of heart muscle cells: correlation with
increased cytoplasmic calcium activity and with changes of nexus ultrastructure. J
Membr Biol 53, 63-75.
Davidson JS & Baumgarten IM (1988). Glycyrrhetinic acid derivatives: a novel class of
inhibitors of gap-junctional intercellular communication. Structure-activity relationships.
J Pharmacol Exp Ther 246, 1104-1107.
De Mello WC (1984). Effect of intracellular injection of cAMP on the electrical coupling
of mammalian cardiac cells. Biochem Biophys Res Commun 119, 1001-1007.
De Mello WC (1975). Effect of intracellular injection of calcium and strontium on cell
communication in heart. J Physiol 250, 231-245.

83
De Pina-Benabou MH, Srinivas M, Spray DC & Scemes E (2001). Calmodulin kinase
pathway mediates the K+-induced increase in Gap junctional communication between
mouse spinal cord astrocytes. J Neurosci 21, 6635-6643.
de Roos ADG, Zoelen EJJ & Theuvenet APR (1996). Determination of gap junctional
intercellular communication by capacitance measurements. Pflugers Arch 431, 556-563.
De Vlieger T, Kits K, Ter Maat A & Lodder J (1980). Morphology and electrophysiology
of the ovulation hormone producing neuro-endocrine cells of the freshwater snail
Lymnaea stagnalis (L.). J Exp Biol 84, 259-271.
Del Corsso C, Iglesias R, Zoidl G, Dermietzel R & Spray DC (2012). Calmodulin
dependent protein kinase increases conductance at gap junctions formed by the neuronal
gap junction protein connexin36. Brain Res 1487, 69-77.
DeRiemer S, Strong J, Albert K, Greengard P & Kaczmarek L (1985). Enhancement of
calcium current in Aplysia neurones by phorbol ester and protein kinase C. Nature 313,
313-316.
DeRiemer SA, Kaczmarek LK, Lai Y, McGuinness TL & Greengard P (1984).
Calcium/calmodulin-dependent protein phosphorylation in the nervous system of Aplysia.
J Neurosci 4, 1618-1625.
Dermietzel R & Spray DC (1993). Gap junctions in the brain: where, what type, how
many and why? Trends Neurosci 16, 186-192.
Dermietzel R & Spray DC (2013). Gap Junctions, Electric Synapses. Neuroscience in the
21st Century: From Basic to Clinical, pp 439-473.
Descheˆnes M & Bennett MVL (1974). A qualification to the use of TEA as a tracer for
monosynaptic pathways. Brain Res 77, 169-172.
Dudek FE, Weir G, Acosta-Urquidi J & Tobe SS (1980). A secretion from
neuroendocrine bag cells evokes egg release in vitro from ovotestis of Aplysia
californica. Gen Comp Endocrinol 40, 241-244.
Dudek FE & Blankenship JE (1977). Neuroendocrine cells of Aplysia brasiliana. I. Bag
cell action potentials and afterdischarge. J Neurophysiol 40, 1301-1311.
Dykes IM, Freeman FM, Bacon JP & Davies JA (2004). Molecular basis of gap
junctional communication in the CNS of the leech Hirudo medicinalis. J Neurosci 24,
886-894.

84
Enslen H & Soderling TR (1994). Roles of calmodulin-dependent protein kinases and
phosphatase in calcium-dependent transcription of immediate early genes. J Biol Chem
269, 20872-20877.
Ermentrout B, Wang JW, Flores J & Gelperin A (2004). Model for transition from waves
to synchrony in the olfactory lobe of Limax. J Comput Neurosci 17, 365-383.
Eskandari S, Zampighi G, Leung D, Wright E & Loo D (2002). Inhibition of gap junction
hemichannels by chloride channel blockers. J Membr Biol 185, 93-102.
Evans WH & Martin PEM (2002). Gap junctions: structure and function (Review). Mol
Membr Biol 19, 121-136.
Ewadinger N, Syed N, Lukowiak K & Bulloch A (1994). Differential tracer coupling
between pairs of identified neurones of the mollusc Lymnaea stagnalis. J Exp Biol 192,
291-297.
Fan R, Marin-Burgin A, French KA & Friesen WO (2005). A dye mixture (Neurobiotin
and Alexa 488) reveals extensive dye-coupling among neurons in leeches; physiology
confirms the connections. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 191,
1157-1171.
Fink L, Connor J & Kaczmarek L (1988). Inositol trisphosphate releases intracellularly
stored calcium and modulates ion channels in molluscan neurons. J Neurosci 8, 2544-
2555.
Fisher T, Levy S & Kaczmarek L (1994). Transient changes in intracellular calcium
associated with a prolonged increase in excitability in neurons of Aplysia californica. J
Neurophysiol 71, 1254-1257.
Fisher T, Lin C & Kaczmarek LK (1993). The peptide FMRFa terminates a discharge in
Aplysia bag cell neurons by modulating calcium, potassium, and chloride conductances. J
Neurophysiol 69, 2164-2164.
Frank M, Eiberger B, Janssen-Bienhold U, de Sevilla Muller LP, Tjarks A, Kim JS,
Maschke S, Dobrowolski R, Sasse P, Weiler R, Fleischmann BK & Willecke K (2010).
Neuronal connexin-36 can functionally replace connexin-45 in mouse retina but not in
the developing heart. J Cell Sci 123, 3605-3615.
Furshpan E & Potter D (1959). Transmission at the giant motor synapses of the crayfish.
J Physiol 145, 289-325.
Gardam KE & Magoski NS (2009). Regulation of cation channel voltage and Ca2+
dependence by multiple modulators. J Neurophysiol 102, 259-271.

85
Gardam KE, Geiger JE, Hickey CM, Hung AY & Magoski NS (2008). Flufenamic acid
affects multiple currents and causes intracellular Ca2+
release in Aplysia bag cell neurons.
J Neurophysiol 100, 38-49.
Geiger JE & Magoski NS (2008). Ca2+
induced Ca2+
release in Aplysia bag cell neurons
requires interaction between mitochondrial and endoplasmic reticulum stores. J
Neurophysiol 100, 24-37.
Getting PA & Willows AO (1974). Modification of neuron properties by electrotonic
synapses. II. Burst formation by electrotonic synapses. J Neurophysiol 37, 858-868.
Gibson JR, Beierlein M & Connors BW (1999). Two networks of electrically coupled
inhibitory neurons in neocortex. Nature 402, 75-79.
Giepmans BN (2004). Gap junctions and connexin-interacting proteins. Cardiovasc Res
62, 233-245.
Gögelein H & Pfannmüller B (1989). The nonselective cation channel in the basolateral
membrane of rat exocrine pancreas. Pflügers Archiv 413, 287-298.
Goldsmith J & Byrne J (1993). Bag cell extract inhibits tail-siphon withdrawal reflex,
suppresses long-term but not short-term sensitization, and attenuates sensory-to-motor
neuron synapses in Aplysia. J Neurosci 13, 1688-1700.
Goodenough DA, Goliger JA & Paul DL (1996). Connexins, connexons, and intercellular
communication. Annu Rev Biochem 65, 475-502.
Griffith TM, Chaytor AT, Taylor HJ, Giddings BD & Edwards DH (2002). cAMP
facilitates EDHF-type relaxations in conduit arteries by enhancing electrotonic
conduction via gap junctions. Proc Natl Acad Sci USA 99, 6392-6397.
Groten CJ, Rebane JT, Blohm G & Magoski NS (2013). Separate Ca2+
sources are
buffered by distinct Ca2+
handling systems in Aplysia neuroendocrine cells. J Neurosci
33, 6476-6491.
Hanson PI & Schulman H (1992). Neuronal Ca2+
/calmodulin-dependent protein kinases.
Annu Rev Biochem 61, 559-601.
Harks EGA, de Roos ADG, Peters PHJ, de Haan LH, Brouwer A, Ypey DL, van Zoelen
EJJ & Theuvenet APR (2001). Fenamates: a novel class of reversible gap junction
blockers. J Pharmacol Exp Ther 298, 1033-1041.
Hatcher NG & Sweedler JV (2008). Aplysia bag cells function as a distributed
neurosecretory network. J Neurophysiol 99, 333-343.

86
Heitzmann H & Richards FM (1974). Use of the avidin-biotin complex for specific
staining of biological membranes in electron microscopy. Proc Natl Acad Sci USA 71,
3537-3541.
Heytler P & Prichard W (1962). A new class of uncoupling agents carbonyl cyanide
phenylhydrazones. Biochem Biophys Res Commun 7, 272-275.
Hickey CM, Geiger JE, Groten CJ & Magoski NS (2010). Mitochondrial Ca2+
activates a
cation current in Aplysia bag cell neurons. J Neurophysiol 103, 1543-1556.
Hickey C, Groten C, Sham L, Carter C & Magoski N (2013). Voltage-gated Ca2+
influx
and mitochondrial Ca2+
initiate secretion from Aplysia neuroendocrine cells.
Neuroscience 250, 755-772.
Hille B (2001). Ion channels of excitable membranes. Sinauer Sunderland, MA.Chapter
16 pp 504-536.
Hung AY & Magoski NS (2007). Activity-dependent initiation of a prolonged
depolarization in Aplysia bag cell neurons: role for a cation channel. J Neurophysiol 97,
2465-2479.
Ichikawa T (2002). Synchronous firing dynamics in a heterogeneous neurosecretory-cell
population in an insect. Brain Res 929, 156-165.
Ichikawa T, Imafuku Y & Tawada K (1999). Synchronous firing patterns of a set of
insect neurosecretory cells. Neurosci Lett 264, 85-88.
Jessell TM & Kandel ER (1993). Synaptic transmission: A bidirectional and self-
modifiable form of cell-cell communication. Cell 72, 1-30.
Johnston M, Simon S & Ramon F (1980). Interaction of anaesthetics with electrical
synapses. Nature 286, 498-500.
Johnston MF & Ramon F (1981). Electrotonic coupling in internally perfused crayfish
segmented axons. J Physiol 317, 509-518.
Juszczak GR & Swiergiel AH (2009). Properties of gap junction blockers and their
behavioural, cognitive and electrophysiological effects: animal and human studies. Prog
Neuropsychopharmacol Biol Psychiatry 33, 181-198.
Kachoei BA, Knox RJ, Uthuza D, Levy S, Kaczmarek LK & Magoski NS (2006). A
store-operated Ca2+
influx pathway in the bag cell neurons of Aplysia. J Neurophysiol 96,
2688-2698.

87
Kaczmarek L, Finbow M, Revel J & Strumwasser F (1979). The morphology and
coupling of Aplysia bag cells within the abdominal ganglion and in cell culture. J
Neurobiol 10, 535-550.
Kaczmarek L, Jennings K & Strumwasser F (1978). Neurotransmitter modulation,
phosphodiesterase inhibitor effects, and cyclic AMP correlates of afterdischarge in
peptidergic neurites. Proc Natl Acad Sci USA 75, 5200-5204.
Kaczmarek LK, Jennings K & Strumwasser F (1982). An early sodium and a late calcium
phase in the afterdischarge of peptide-secreting neurons of Aplysia. Brain Res 238, 105-
115.
Kaczmarek LK & Kauer J (1983). Calcium entry causes a prolonged refractory period in
peptidergic neurons of Aplysia. J Neurosci 3, 2230-2239.
Kaczmarek LK & Strumwasser F (1981). The expression of long lasting afterdischarge
by isolated Aplysia bag cell neurons. J Neurosci 1, 626-634.
Kaczmarek LK & Strumwasser F (1984). A voltage-clamp analysis of currents
underlying cyclic AMP-induced membrane modulation in isolated peptidergic neurons of
Aplysia. J Neurophysiol 52, 340-349.
Kaczmarek LK, Jennings KR, Strumwasser F, Nairn AC, Walter U, Wilson FD &
Greengard P (1980). Microinjection of catalytic subunit of cyclic AMP-dependent protein
kinase enhances calcium action potentials of bag cell neurons in cell culture. Proc Natl
Acad Sci USA 77, 7487-7491.
Kalra SP, Bagnasco M, Otukonyong EE, Dube MG & Kalra PS (2003a). Rhythmic,
reciprocal ghrelin and leptin signaling: new insight in the development of obesity. Regul
Pept 111, 1-11.
Kalra SP, Horvath T, Naftolin F, Xu B, Pu S & Kalra PS (2003b). The interactive
language of the hypothalamus for the gonadotropin releasing hormone (GnRH) system. J
Neuroendocrinol 9, 569-576.
Kanaporis G, Mese G, Valiuniene L, White TW, Brink PR & Valiunas V (2008). Gap
junction channels exhibit connexin-specific permeability to cyclic nucleotides. J Gen
Physiol 131, 293-305.
Kanaporis G, Brink PR & Valiunas V (2011). Gap junction permeability: selectivity for
anionic and cationic probes. Am J Physiol Cell Physiol 300, C600-C609.
Kandarian B, Sethi J, Wu A, Baker M, Yazdani N, Kym E, Sanchez A, Edsall L,
Gaasterland T & Macagno E (2012). The medicinal leech genome encodes 21 innexin

88
genes: different combinations are expressed by identified central neurons. Dev Genes
Evol 222, 29-44.
Kandel E, Kriegstein A & Schacher S (1980). Development of the central nervous system
of Aplysia in terms of the differentiation of its specific identifiable cells. Neuroscience 5,
2033-2063.
Kanter HL, Laing JG, Beau SL, Beyer EC & Saffitz JE (1993). Distinct patterns of
connexin expression in canine Purkinje fibers and ventricular muscle. Circ Res 72, 1124-
1131.
Katz B & Miledi R (1967). The timing of calcium action during neuromuscular
transmission. J Physiol 189, 535-544.
Kelsell D, Dunlop J, Stevens H, Lench N, Liang J, Parry G, Mueller R & Leigh I (1997).
Connexin 26 mutations in hereditary non-syndromic sensorineural deafness. Nature 387,
80-83.
Klein M & Kandel ER (1978). Presynaptic modulation of voltage-dependent Ca2+
current: mechanism for behavioral sensitization in Aplysia californica. Proc Natl Acad
Sci USA 75, 3512-3516.
Knox RJ, Quattrocki EA, Connor JA & Kaczmarek LK (1992). Recruitment of Ca2+
channels by protein kinase C during rapid formation of putative neuropeptide release sites
in isolated Aplysia neurons. Neuron 8, 883-889.
Knox RJ, Magoski NS, Wing D, Barbee SJ & Kaczmarek LK (2004). Activation of a
calcium entry pathway by sodium pyrithione in the bag cell neurons of Aplysia. J
Neurobiol 60, 411-423.
Konietzko U, Müller CM (1994). Astrocytic dye coupling in rat hippocampus:
topography, developmental onset, and modulation by protein kinase C. Hippocampus 4,
297-306.
Krysko D, Leybaert L, Vandenabeele P & D’Herde K (2005). Gap junctions and the
propagation of cell survival and cell death signals. Apoptosis 10, 459-469.
Kumar NM & Gilula NB (1996). The gap junction communication channel. Cell 84, 381-
388.
Kupfermann I (1967). Stimulation of egg laying: possible neuroendocrine function of bag
cells of abdominal ganglion of Aplysia californica. Nature 216, 814-815.

89
Kupfermann I & Kandel E (1970). Electrophysiological properties and functional
interconnections of two symmetrical neurosecretory clusters (bag cells) in abdominal
ganglion of Aplysia. J Neurophysiol 33, 865-876.
Laird DW (2005). Connexin phosphorylation as a regulatory event linked to gap junction
internalization and degradation. Biochimica et Biophysica Acta 1711, 172-182.
Lampe PD, TenBroek EM, Burt JM, Kurata WE, Johnson RG & Lau AF (2000).
Phosphorylation of connexin43 on serine368 by protein kinase C regulates gap junctional
communication. J Cell Biol 149, 1503-1512.
Landisman CE, Long MA, Beierlein M, Deans MR, Paul DL & Connors BW (2002).
Electrical synapses in the thalamic reticular nucleus. J Neurosci 22, 1002-1009.
Langevin HM, Cornbrooks CJ & Taatjes DJ (2004). Fibroblasts form a body-wide
cellular network. Histochem Cell Biol 122, 7-15.
Lazrak A & Peracchia C (1993). Gap junction gating sensitivity to physiological internal
calcium regardless of pH in Novikoff hepatoma cells. Biophys J 65, 2002-2012.
Lazrak A, Peres A, Giovannardi S & Peracchia C (1994). Ca2+
-mediated and independent
effects of arachidonic acid on gap junctions and Ca2+
-independent effects of oleic acid
and halothane. Biophys J 67, 1052-1059.
Levitan IB (1999). It is calmodulin after all! Mediator of the calcium modulation of
multiple ion channels. Neuron 22, 645-648.
Levitan H, Tauc L & Segundo JP (1970). Electrical transmission among neurons in the
buccal ganglion of a mollusc, Navanax inermis. J Gen Physiol 55, 484-496.
Li H, Liu T, Lazrak A, Peracchia C, Goldberg GS, Lampe PD & Johnson RG (1996).
Properties and regulation of gap junctional hemichannels in the plasma membranes of
cultured cells. J Cell Biol 134, 1019-1030.
Ligman SH & Brownell PH (1985). Differential hormonal action of the bag cell neurons
on the arterial system of Aplysia. J Comp Physiol A 157, 31-37.
Lin SS & Levitan IB (1987). Concanavalin A alters synaptic specificity between cultured
Aplysia neurons. Science 237, 648-650.
Lincoln D & Wakerley J (1974). Electrophysiological evidence for the activation of
supraoptic neurones during the release of oxytocin. J Physiol 242, 533-554.

90
Liu Q, Chen B, Gaier E, Joshi J & Wang Z (2006). Low conductance gap junctions
mediate specific electrical coupling in body-wall muscle cells of Caenorhabditis elegans.
J Biol Chem 281, 7881-7889.
Lledo PM, Hjelmstad GO, Mukherji S, Soderling TR, Malenka RC & Nicoll RA (1995).
Calcium/calmodulin-dependent kinase II and long-term potentiation enhance synaptic
transmission by the same mechanism. Proc Natl Acad Sci USA 92, 11175-11179.
Llinas R, Steinberg I & Walton K (1981). Relationship between presynaptic calcium
current and postsynaptic potential in squid giant synapse. Biophys J 33, 323-351.
Lococo D, Thompson C & Tobe, S (1986). Intercellular communication in an insect
endocrine gland. J Exp Biol 121, 407-419.
Loechner KJ & Kaczmarek LK (1994). Autoactive peptides act at three distinct receptors
to depolarize the bag cell neurons of Aplysia. J Neurophysiol 71, 195-203.
Loewenstein WR, Nakas M & Socolar SJ (1967). Junctional membrane uncoupling.
Permeability transformations at a cell membrane junction. J Gen Physiol 50, 1865-1891.
Long MA, Jutras MJ, Connors BW & Burwell RD (2004). Electrical synapses coordinate
activity in the suprachiasmatic nucleus. Nat Neurosci 8, 61-66.
Lupinsky DA & Magoski NS (2006). Ca2+‐dependent regulation of a non‐selective cation
channel from Aplysia bag cell neurones. J Physiol (Lond ) 575, 491-506.
Mackey S & Carew TJ (1983). Locomotion in Aplysia: triggering by serotonin and
modulation by bag cell extract. J Neurosci 3, 1469-1477.
Magoski NS & Kaczmarek LK (2005). Association/dissociation of a channel–kinase
complex underlies state-dependent modulation. J Neurosci 25, 8037-8047.
Magoski NS, Knox RJ & Kaczmarek LK (2000). Activation of a Ca2+
permeable cation
channel produces a prolonged attenuation of intracellular Ca2+
release in Aplysia bag cell
neurones. J Physiol 522, 271-283.
Magoski NS (2004). Regulation of an Aplysia bag cell neuron cation channel by closely
associated protein kinase A and a protein phosphatase. J Neurosci 24, 6833-6841.
Magoski NS, Wilson GF & Kaczmarek LK (2002). Protein kinase modulation of a
neuronal cation channel requires protein–protein interactions mediated by an Src
homology 3 domain. J Neurosci 22, 1-9.

91
Mammano F (2000). Gap Junctions: Cell-Cell Channels in Animals. Landes Bioscience.
pp 186-201.
Matchkov VV, Rahman A, Peng H, Nilsson H & Aalkjær C (2004). Junctional and
nonjunctional effects of heptanol and glycyrrhetinic acid derivates in rat mesenteric small
arteries. Br J Pharmacol 142, 961-972.
Mayeri E, Brownell P, Branton W & Simon S (1979). Multiple, prolonged actions of
neuroendocrine bag cells on neurons in Aplysia. I. Effects on bursting pacemaker
neurons. J Neurophysiol 42, 1165-1184.
McCracken CB & Roberts DC (2006). Neuronal gap junctions: expression, function, and
implications for behavior. Int Rev Neurobiol 73, 125.
Michel S & Wayne NL (2002). Neurohormone secretion persists after post-afterdischarge
membrane depolarization and cytosolic calcium elevation in peptidergic neurons in intact
nervous tissue. J Neurosci 22, 9063-9069.
Moreno AP & Lau AF (2007). Gap junction channel gating modulated through protein
phosphorylation. Prog Biophys Mol Biol 94, 107-119.
Moreno AP, Saez JC, Fishman GI & Spray DC (1994). Human connexin43 gap junction
channels. Regulation of unitary conductances by phosphorylation. Circ Res 74, 1050-
1057.
Moroz LL, Edwards JR, Puthanveettil SV, Kohn AB, Ha T, Heyland A, Knudsen B,
Sahni A, Yu F & Liu L (2006). Neuronal Transcriptome of Aplysia: Neuronal
Compartments and Circuitry. Cell 127, 1453-1467.
Murphy AD, Hadley RD & Kater SB (1983). Axotomy-induced parallel increases in
electrical and dye coupling between identified neurons of Helisoma. J Neurosci 3, 1422-
1429.
Mustonen H, Kiviluoto T, Paimela H, Puolakkainen P & Kivilaakso E (2005). Calcium
signaling is involved in ethanolinduced volume decrease and gap junction closure in
cultured rat gastric mucosal cells. Dig Dis Sci 50, 103-110.
Newton AC (2010). Protein kinase C: poised to signal. Am J Physiol Endocrinol Metab
298, E395-E402.
Norekian TP (1999). GABAergic excitatory synapses and electrical coupling sustain
prolonged discharges in the prey capture neural network of Clione limacina. J Neurosci
19, 1863-1875.

92
Obaid AL, Socolar SJ & Rose B (1983). Cell-to-cell channels with two independently
regulated gates in series: analysis of junctional conductance modulation by membrane
potential, calcium, and pH. J Membr Biol 73, 69-89.
Panchina Y, Kelmanson I, Matz M, Lukyanov K, Usman N, Lukyanov S (2000). A
ubiquitous family of putative gap junction molecules. Curr Biol 10, R473-R474.
Pan F, Mills SL & Massey SC (2007). Screening of gap junction antagonists on dye
coupling in the rabbit retina. Vis Neurosci 24, 609-618.
Payton BW, Bennett MVL & Pappas GD (1969). Permeability and structure of junctional
membranes at an electrotonic synapse. Science 166, 1641-1643.
Peinado A, Yuste R & Katz LC (1993). Extensive dye coupling between rat neocortical
neurons during the period of circuit formation. Neuron 10, 103-114.
Peracchia C (2004). Chemical gating of gap junction channels: Roles of calcium, pH and
calmodulin. Biochimica et Biophysica Acta 1662, 61-80.
Peracchia C (1990). Increase in gap junction resistance with acidification in crayfish
septate axons is closely related to changes in intracellular calcium but not hydrogen ion
concentration. J Membr Biol 113, 75-92.
Peracchia C (1987). Calmodulin-like proteins and communicating junctions. Pflügers
Archiv 408, 379-385.
Peracchia C (1984). Communicating junctions and calmodulin: Inhibition of electrical
uncoupling in Xenopus embryo by calmidazolium. J Membr Biol 81, 49-58.
Peracchia C, Bernardini G & Peracchia LL (1983). Is calmodulin involved in the
regulation of gap junction permeability? Pflügers Archiv 399, 152-154.
Peracchia C, Wang X, Li L & Peracchia LL (1996). Inhibition of calmodulin expression
prevents low-pH-induced gap junction uncoupling in Xenopus oocytes. Pflügers Archiv
431, 379-387.
Peracchia C & Dulhunty AF (1976). Low resistance junctions in crayfish. Structural
changes with functional uncoupling. J Cell Biol 70, 419-439.
Pereda AE, Bell TD, Chang BH, Czernik AJ, Nairn AC, Soderling TR & Faber DS
(1998). Ca2+
/calmodulin-dependent kinase II mediates simultaneous enhancement of gap-
junctional conductance and glutamatergic transmission. Proc Natl Acad Sci USA 95,
13272-13277.

93
Perez-Armendariz M, Roy C, Spray DC & Bennett MVL (1991). Biophysical properties
of gap junctions between freshly dispersed pairs of mouse pancreatic beta cells. Biophys J
59, 76-92.
Peters NS, Coromilas J, Severs NJ & Wit AL (1997). Disturbed connexin43 gap junction
distribution correlates with the location of reentrant circuits in the epicardial border zone
of healing canine infarcts that cause ventricular tachycardia. Circulation 95, 988-996.
Phelan P & Starich TA (2001). Innexins get into the gap. Bioessays 23, 388-396.
Pinsker HM & Dudek FE (1977). Bag cell control of egg laying in freely behaving
Aplysia. Science 197, 490-493.
Poronnik P, Ward M & Cook D (1992). Intracellular Ca2+
release by flufenamic acid and
other blockers of the non-selective cation channel. FEBS Lett 296, 245-248.
Potenza N, del Gaudio R, Rivieccio L, Russo GMR & Geraci G (2002). Cloning and
molecular characterization of the first innexin of the phylum annelida Expression of the
gene during development. J Mol Evol 54, 312-321.
Račkauskas M, Neverauskas V & Skeberdis VA (2010). Diversity and properties of
connexin gap junction channels. Medicina (Kaunas) 46, 1-12.
Randriamampita C, Giaume C, Neyton J & Trautmann A (1988). Acetylcholine-induced
closure of gap junction channels in rat lacrimal glands is probably mediated by protein
kinase C. Pflügers Archiv 412, 462-468.
Rao G, Barnes C & McNaughton B (1987). Occlusion of hippocampal electrical
junctions by intracellular calcium injection. Brain Res 408, 267-270.
Reed KE, Westphale EM, Larson DM, Wang HZ, Veenstra RD & Beyer EC (1993).
Molecular cloning and functional expression of human connexin37, an endothelial cell
gap junction protein. J Clin Invest 91, 997-1004.
Revilla A, Castro C & Barrio LC (1999). Molecular dissection of transjunctional voltage
dependence in the connexin-32 and connexin-43 junctions. Biophys J 77, 1374-1383.
Rink T, Tsien R & Warner A (1980). Free calcium in Xenopus embryos measured with
ion-selective microelectrodes. Nature 283, 658-660.
Robertson JD (1953). Ultrastructure of two invertebrate synapses. Proc Soc Exp Biol Med
82, 219-223.

94
Robertson JD, Bodenheimer TS & Stage DE (1963). The ultrastructure of Mauthner cell
synapses and nodes in goldfish brains. J Cell Biol 19, 159-199.
Rohr S (2004). Role of gap junctions in the propagation of the cardiac action potential.
Cardiovasc Res 62, 309-322.
Rörig B & Sutor B (1996). Serotonin regulates gap junction coupling in the developing
rat somatosensory cortex. Eur J Neurosci 8, 1685-1695.
Rose B & Loewenstein WR (1976). Permeability of a cell junction and the local
cytoplasmic free ionized calcium concentration: a study with aequorin. J Membr Biol 28,
87-119.
Rose B & Loewenstein WR (1975). Calcium ion distribution in cytoplasm visualised by
aequorin: diffusion in cytosol restricted by energized sequestering. Science 190, 1204-
1206.
Rothman BS, Weir G & Dudek FE (1983). Egg-laying hormone: Direct action on the
ovotestis of Aplysia. Gen Comp Endocrinol 52, 134-141.
Saez JC, Nairn, AC, Czernik, AJ, Spray, DC, Hertzberg, EL, Greengard, P & Bennett,
MVL (1990). Phosphorylation of connexin 32, a hepatocyte gap‐junction protein, by
cAMP‐dependent protein kinase, protein kinase C and Ca2+
/calmodulin‐dependent
protein kinase II. Eur. J. Biochem 192, 263-273.
Saez JC, Connor JA, Spray DC & Bennett MVL (1989). Hepatocyte gap junctions are
permeable to the second messenger, inositol 1,4,5-trisphosphate, and to calcium ions.
Proc Natl Acad Sci USA 86, 2708-2712.
Saez JC, Spray DC, Nairn AC, Hertzberg E, Greengard P & Bennett MVL (1986). cAMP
increases junctional conductance and stimulates phosphorylation of the 27-kDa principal
gap junction polypeptide. Proc Natl Acad Sci USA 83, 2473-2477.
Sandoz P & Dreifuss J (1978). Neurones with synchronous bursting discharges in organ
cultures of the hypothalamic supraoptic nucleus area. Brain Res 151, 245-253.
Schaefer M, Picciotto MR, Kreiner T, Kaldany R, Taussig R & Scheller RH (1985).
Aplysia neurons express a gene encoding multiple FMRFamide neuropeptides. Cell 41,
457-467.
Schirrmacher K, Nonhoff D, Wiemann M, Peterson-Grine E, Brink P & Bingmann D
(1996). Effects of calcium on gap junctions between osteoblast-like cells in culture.
Calcif Tissue Int 59, 259-264.

95
Seidler NW, Jona I, Vegh M & Martonosi A (1989). Cyclopiazonic acid is a specific
inhibitor of the Ca2+
-ATPase of sarcoplasmic reticulum. J Biol Chem 264, 17816-17823.
Sohl G & Willecke K (2004). Gap junctions and the connexin protein family. Cardiovasc
Res 62, 228-232.
Solan JL & Lampe PD (2005). Connexin phosphorylation as a regulatory event linked to
gap junction channel assembly. Biochimica et Biophysica Acta 1711, 154-163.
Somogyi R, Batzer A & Kolb H (1989). Inhibition of electrical coupling in pairs of
murine pancreatic acinar cells by OAG and isolated protein kinase C. J Membr Biol 108,
273-282.
Sossin WS & Schwartz JH (1992). Selective activation of Ca2+
-activated PKCs in Aplysia
neurons by 5-HT. J Neurosci 12, 1160-1168.
Sossin WS, Sacktor TC & Schwartz JH (1994). Persistent activation of protein kinase C
during the development of long-term facilitation in Aplysia. Learn Mem 1, 189-202.
Spray D & Bennett MVL (1985). Physiology and pharmacology of gap junctions. Annu
Rev Physiol 47, 281-303.
Spray D, Harris A & Bennett MVL (1981). Equilibrium properties of a voltage-
dependent junctional conductance. J Gen Physiol 77, 77-93.
Spray D, White R, De Carvalho AC, Harris A & Bennett MVL (1984). Gating of gap
junction channels. Biophys J 45, 219-230.
Spray DC & Burt JM (1990). Structure-activity relations of the cardiac gap junction
channel. Am J Physiol 258, C195-C205.
Spray DC, Harris AL & Bennett MVL (1979). Voltage dependence of junctional
conductance in early amphibian embryos. Science 204, 432-434.
Spray DC, Stern JH, Harris AL & Bennett MV (1982). Gap junctional conductance:
comparison of sensitivities to H and Ca ions. Proc Natl Acad Sci USA 79, 441-445.
Srinivas M, Hopperstad MG & Spray DC (2001). Quinine blocks specific gap junction
channel subtypes. Proc Natl Acad Sci USA 98, 10942-10947.
Srinivas M & Spray DC (2003). Closure of gap junction channels by arylaminobenzoates.
Mol Pharmacol 63, 1389-1397.

96
Starich TA, Miller A, Nguyen RL, Hall DH & Shaw JE (2003). The Caenorhabditis
elegans innexin INX-3 is localized to gap junctions and is essential for embryonic
development. Dev Biol 256, 403-417.
Steriade M (2005). Sleep, epilepsy and thalamic reticular inhibitory neurons. Trends
Neurosci 28, 317-324.
Stewart WW (1978). Functional connections between cells as revealed by dye-coupling
with a highly fluorescent naphthalimide tracer. Cell 14, 741-759.
Strong JA (1984). Modulation of potassium current kinetics in bag cell neurons of
Aplysia by an activator of adenylate cyclase. J Neurosci 4, 2772-2783.
Strong J & Kaczmarek L (1986). Multiple components of delayed potassium current in
peptidergic neurons of Aplysia: modulation by an activator of adenylate cyclase. J
Neurosci 6, 814-822.
Stuart DK, Chiu AY & Strumwasser F (1980). Neurosecretion of egg-laying hormone
and other peptides from electrically active bag cell neurons of Aplysia. J Neurophysiol
43, 488-498.
Summerlee A (1981). Extracellular recordings from oxytocin neurones during the
expulsive phase of birth in unanaesthetized rats. J Physiol 321, 1-9.
Takahira M, Sakurai M, Sakurada N & Sugiyama K (2005). Fenamates and diltiazem
modulate lipid-sensitive mechano-gated 2P domain K channels. Pflügers Archiv 451,
474-478.
Tam A, Gardam K, Lamb S, Kachoei B & Magoski NS (2011). Role for protein kinase C
in controlling Aplysia bag cell neuron excitability. J Neurosci 179, 41-55.
Tam AK, Geiger JE, Hung AY, Groten CJ & Magoski NS (2009). Persistent Ca2+
current
contributes to a prolonged depolarization in Aplysia bag cell neurons. J Neurophysiol
102, 3753-3765.
Thastrup O, Cullen PJ, DrØbaks KB, Hanleys MR & DAWSONt PK (1990).
Thapsigargin, a tumor promoter, discharges intracellular Ca2+
stores by specific inhibition
of the endoplasmic reticulum Ca2+
-ATPase. Proc Natl Acad Sci USA 87 2466-2470.
Tokumitsu H, Chijiwa T, Hagiwara M, Mizutani A, Terasawa M & Hidaka H (1990).
KN-62, 1-[N,O-bis(5-isoquinolinesulfonyl)-N-methyl-L-tyrosyl]-4-phenylpiperazi ne, a
specific inhibitor of Ca2+
/calmodulin-dependent protein kinase II. J Biol Chem 265,
4315-4320.

97
Toyama J, Sugiura H, Kamiya K, Kodama I, Terasawa M & Hidaka H (1994). Ca2+
-
calmodulin mediated modulation of the electrical coupling of ventricular myocytes
Isolated from Guinea Pig Heart. J Mol Cell Cardiol 26, 1007-1015.
Traub RD, Kopell N, Bibbig A, Buhl EH, LeBeau FE & Whittington MA (2001). Gap
junctions between interneuron dendrites can enhance synchrony of gamma oscillations in
distributed networks. J Neurosci 21, 9478-9486.
van Veen TA, van Rijen HV & Jongsma HJ (2000). Electrical conductance of mouse
connexin45 gap junction channels is modulated by phosphorylation. Cardiovasc Res 46,
496-510.
Vazquez-Martinez R, Shorte SL, Boockfor FR & Frawley LS (2001). Synchronized
exocytotic bursts from gonadotropin-releasing hormone-expressing cells: dual control by
intrinsic cellular pulsatility and gap junctional communication. Endocrinology 142, 2095-
2101.
Verselis V & Brink P (1984). Voltage clamp of the earthworm septum. Biophys J 45,
147-150.
Verselis VK, Bennett MVL & Bargiello TA (1991). A voltage-dependent gap junction in
Drosophila melanogaster. Biophys J 59, 114-126.
Verselis V, White RL, Spray DC & Bennett MVL (1986). Gap junctional conductance
and permeability are linearly related. Science 234, 461-464.
Wang HZ, Li J, Lemanski L & Veenstra RD (1992). Gating of mammalian cardiac gap
junction channels by transjunctional voltage. Biophys J 63, 139.
Wayne NL, Lee W & Kim YJ (1999). Persistent activation of calcium-activated and
calcium-independent protein kinase C in response to electrical afterdischarge from
peptidergic neurons of Aplysia. Brain Res 834, 211-213.
Wayne NL (2001). Regulation of seasonal reproduction in mollusks. J Biol Rhythms 16,
391-402.
White R, Spray D, De Carvalho ACC, Wittenberg B & Bennett MVL (1985). Some
electrical and pharmacological properties of gap junctions between adult ventricular
myocytes. Am J Physiol 249, C447-C455.
White SH & Magoski NS (2012). Acetylcholine-evoked afterdischarge in Aplysia bag
cell neurons. J Neurophysiol 107, 2672-2685.

98
White TW (2002). Unique and redundant connexin contributions to lens development.
Science 295, 319-320.
White MM & Aylwin M (1990). Niflumic and flufenamic acids are potent reversible
blockers of Ca2+
-activated Cl- channels in Xenopus oocytes. Mol Pharmacol 37, 720-724.
White RL, Doeller JE, Verselis VK & Wittenberg BA (1990). Gap junctional
conductance between pairs of ventricular myocytes is modulated synergistically by H+
and Ca2+
. J Gen Physiol 95, 1061-1075.
Wildering W, Janse C & De Vlieger T (1991). The role of pacemaker properties and
synaptic input in generation and modulation of spiking activity in a pair of electrically
coupled peptidergic neurons. Brain Res 556, 324-328.
Wilson GF & Kaczmarek LK (1993). Mode-switching of a voltage-gated cation channel
is mediated by a protein kinase A-regulated tyrosine phosphatase. Nature 366, 433-438.
Woolum JC & Strumwasser F (1988). Calcium changes in isolated peptidergic neurons
during activation by a cAMP analog. Brain Res 444, 1-9.
Xia J, Liu C, Tang B, Pan Q, Huang L, Dai H, Zhang B, Xie W, Hu D & Zheng D
(1998). Mutations in the gene encoding gap junction protein beta-3 associated with
autosomal dominant hearing impairment. Nat Genet 20, 370-373.
Zeng Y, Lv X, Zeng S & Shi J (2009). Activity-dependent neuronal control of gap-
junctional communication in fibroblasts. Brain Res 1280, 13-22.
Zsiros V & Maccaferri G (2008). Noradrenergic modulation of electrical coupling in
GABAergic networks of the hippocampus. J Neurosci 28, 1804-1815.
Recommended

















