
BIOMEMBRANAS: Membrana plasmática
1. Estructura de las biomembranas. Estructura y propiedades de la
bicapa lipídica. Estructura, composición y propiedades de la
membrana plasmática
2. Modelos de membrana, mosaico fluido.
3. Funciones de la membrana plasmática.
4. La matriz extracelular o glicocalix de células animales
5. La pared celular. Estructura, composición y función. No formación
ESTRUCTURA DE LAS BIOMEMBRANAS
1. Estructura de las biomembranas
La membrana plasmática es una barrera que limita la célula, la
separa de su entorno pero también la comunica, regulando el
transporte de sustancias entre la célula y el exterior, de
manera que ejerce una importante función en el intercambio
de sustancias. También tiene otras funciones importantes,
que veremos.
Además de la membrana plasmática, todos los orgánulos membranosos de la célula están
formados por biomembranas semejantes a aquella y que al MET aparecen con estructura
trilaminar: dos bandas oscuras separadas por una clara que llamamos unidad de membrana o
M. unitaria.
Tiene un espesor de 7,5 nm o 75 Aº y está compuesta, básicamente, por una bicapa lipídica
y proteínas. Un 40% son lípidos (fosfolípidos , glucolípidos, esteroles ) y un
60% proteínas (integrales y periféricas ).
ESTRUCTURA Y PROPIEDADES DE LA BICAPA LIPÍDICA.
(recordar fosfolípidos-el carácter anfipático que les hace muy apropiados para formar membranas, etc). http://www.youtube.com/watch?v=rmANPjbufrY&feature=related membrana celular
2- ESTRUCTURA, COMPOSICIÓN Y PROPIEDADES DE LA MEMBRANA PLASMÁTICA
Componentes de la membrana:
Como hemos dicho, en la membrana encontramos PROTEÍNAS Y LÍPIDOS. Dado que una
parte de ellos son glucolípidos y glucoproteínas, aparece un componente glucídico con
propiedades importantes.
1- LOS LÍPIDOS DE MEMBRANA 1.1 Fosfolípidos – Son los lípidos más abundantes de la membrana y son moléculas
anfipáticas. Presentan una zona hidrófila y (cabeza polar de la glicerina) y una zona
hidrófoba (cola apolar de los ácidos grasos). Forman la bicapa fosfolipídica ya que
el exterior y el interior de la célula son disoluciones acuosas, presentando los
extremos hidrófilos orientados hacia el exterior y las cadenas hidrófobas
enfrentadas y dirigidas hacia el interior.
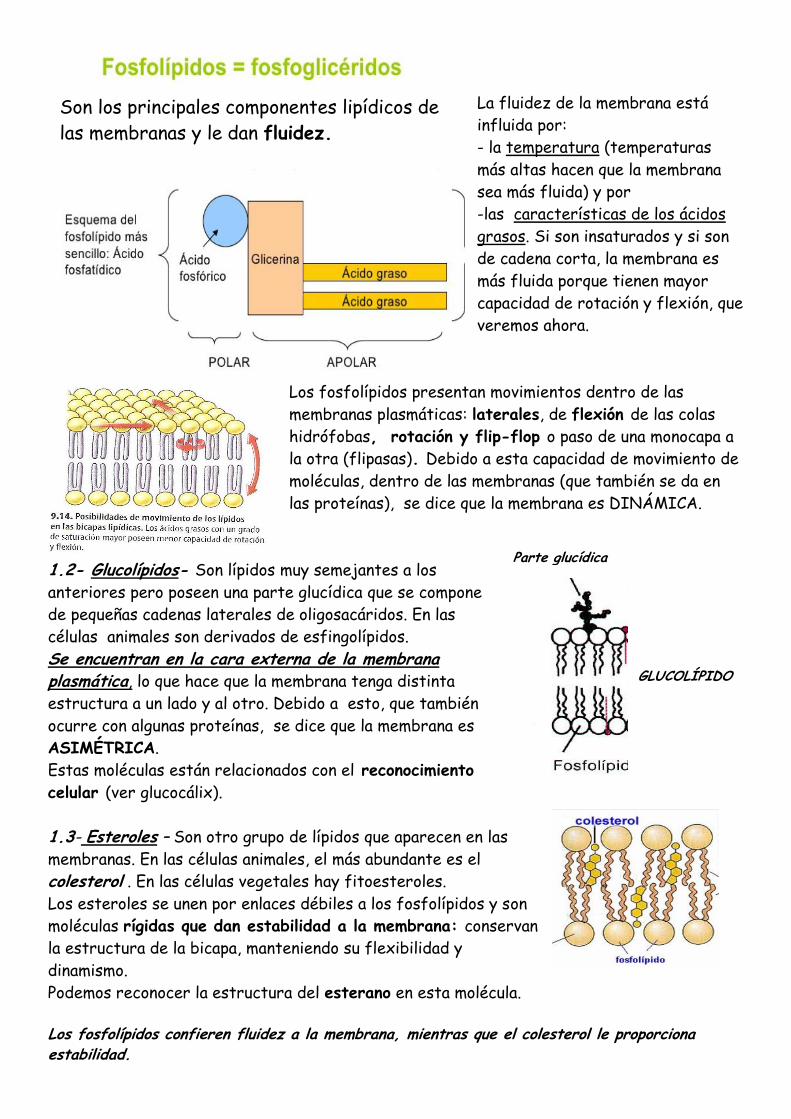
La fluidez de la membrana está
influida por:
- la temperatura (temperaturas
más altas hacen que la membrana
sea más fluida) y por
-las características de los ácidos
grasos. Si son insaturados y si son
de cadena corta, la membrana es
más fluida porque tienen mayor
capacidad de rotación y flexión, que
veremos ahora.
Los fosfolípidos presentan movimientos dentro de las
membranas plasmáticas: laterales, de flexión de las colas
hidrófobas, rotación y flip-flop o paso de una monocapa a
la otra (flipasas). Debido a esta capacidad de movimiento de
moléculas, dentro de las membranas (que también se da en
las proteínas), se dice que la membrana es DINÁMICA.
1.2- Glucolípidos- Son lípidos muy semejantes a los
anteriores pero poseen una parte glucídica que se compone
de pequeñas cadenas laterales de oligosacáridos. En las
células animales son derivados de esfingolípidos.
Se encuentran en la cara externa de la membrana plasmática, lo que hace que la membrana tenga distinta
estructura a un lado y al otro. Debido a esto, que también
ocurre con algunas proteínas, se dice que la membrana es
ASIMÉTRICA.
Estas moléculas están relacionados con el reconocimiento
celular (ver glucocálix).
1.3- Esteroles – Son otro grupo de lípidos que aparecen en las
membranas. En las células animales, el más abundante es el colesterol . En las células vegetales hay fitoesteroles.
Los esteroles se unen por enlaces débiles a los fosfolípidos y son
moléculas rígidas que dan estabilidad a la membrana: conservan
la estructura de la bicapa, manteniendo su flexibilidad y
dinamismo.
Podemos reconocer la estructura del esterano en esta molécula. Los fosfolípidos confieren fluidez a la membrana, mientras que el colesterol le proporciona estabilidad.
Parte glucídica
GLUCOLÍPIDO
Son los principales componentes lipídicos de
las membranas y le dan fluidez.

2- LAS PROTEÍNAS DE LA MEMBRANA PLASMÁTICA
Un 60% de la membrana son PROTEÍNAS. Según su colocación en la membrana distinguimos:
2.1- Integrales: las que están
inmersas en la bicapa lipídica y que
pueden ser:
-Transmembranales- son las
que atraviesan toda la membrana
debido a que poseen regiones
hidrófobas que se sitúan hacia el
interior e hidrófilas, hacia el
exterior. Sólo se separan de la
membrana si ésta se destruye (con
detergentes neutros, por ejemplo).
-Parcialmente integradas- se
encuentran en la cara externa y
llevan glúcidos unidos a ellas
covalentemente. Se trata por tanto de glucoproteínas que originan la asimetría de la
membrana que hemos mencionado.
2.2-Perifericas- Son proteínas que no penetran en la bicapa porque no tienen zona hidrófoba
que les permita hacerlo. Se unen a la membrana por enlace iónico y se separan con facilidad.
Aparecen, sobre todo, en la cara interna (asimetría).
Las proteínas de la membrana pueden desplazarse lateralmente, tienen movilidad, al igual que
los lípidos.
3- LOS GLÚCIDOS DE LA MEMBRANA: EL GLICOCÁLIX
Las partes glucídicas de glucolípidos y glucoproteínas son
oligosacáridos que se encuentran en la cara externa de la
membrana formando una matriz extracelular o estructura
fibrosa que une las células, llamada glicocálix.
No se encuentra en las membranas de los orgánulos celulares y
a veces tiene mayor grosor que toda la membrana. Funciones:
-Soporte- mantiene la forma de las células y la estructura
del tejido. Suele formar la sustancia intercelular.
-Es importante en el reconocimiento celular: reconocimiento
entre óvulo y espermatozoide en la fecundación. Es
responsable de la identidad celular.
-Tiene propiedades inmunitarias como la identificación de
antígenos
Está muy desarrollado en células epiteliales y del conjuntivo.
Es una estructura que no se encuentra en las células vegetales (no le llamamos orgánulo).
Parte glucídica de

CARACTERÍSTICAS O PROPIEDADES DE LA MEMBRANA:
Carácter dinámico- La membrana tiene
mucha FLUIDEZ Y MOVILIDAD debido a que los
lípidos y las proteina pueden desplazarse
lateralmente por ella. No es estática.
Asimetría- La composición de los lípidos y
proteínas es diferente en sus dos caras, externa
e interna.
Permeabilidad selectiva- No es una barrera
pasiva sino muy activa y consume mucha energía
(veremos)
MODELOS DE MEMBRANA
1- Modelo de Davson y Danielli (modelo de sándwich), según el cual la bicapa lipídica
estaría recubierta por proteínas en una estructura rígida, sin movilidad, formando poros.
2- Modelo de Singer y Nicholson, (modelo del mosaico fluido), según el cual la membrana
es una estructura fluida, no rígida, que permite el movimiento de las proteínas. Es la teoría
que hoy día se acepta y que hemos explicado.
ESTRUCTURA DE LA MEMBRANA PLASMÁTICA

FUNCIONES DE LA MEMBRANA PLASMÁTICA
Las funciones derivadas de la presencia de la bicapa lipídica son:
1- Mantener separados el medio interno y externo
2- Realizar los procesos de endocitosis y exocitosis
Las funciones que derivan de las proteínas son:
Regular la entrada y salida de moléculas- La membrana es una barrera selectiva.
Mantiene la estructura del tejido, protegiendo a la célula de lesiones
Une las células gracias a estructuras protéicas.
Posibilita el reconocimiento y comunicación celular. Recibe señales químicas y
mensajeros del medio externo, los reconoce y responde a ellos.
GRUPOS SANGUÍNEOS
Las diferencias entre los grupos sanguíneos se
hallan determinadas por ciertos oligosacáridos
muy cortos, presentes en las membranas
plasmáticas de los glóbulos rojos o
eritrocitos. Estos oligosacáridos sólo difieren
en sus monómeros terminales y están ligados a
una proteína transmembranal o a una ceramida
de la membrana plasmática. Por ejemplo, los
eritrocitos pertenecientes al grupo sanguíneo
A, presentan como monosacárido terminal una
N-acetilgalactosamina y los del grupo B una
galactosa. Cuando ambos monosacáridos
terminales están ausentes estamos en
presencia del grupo 0.
Grupos sanguíneos

TRANSPORTE DE SUSTANCIAS A TRAVÉS DE LA MEMBRANA
Las células requieren nutrientes del exterior y deben eliminar sustancias de desecho
procedentes del metabolismo y mantener su medio interno estable. La membrana es
semipermeable pero presenta una permeabilidad selectiva, ya que permite el paso de
pequeñas moléculas, siempre que sean lipófilas, pero regula el paso de no lipófilas. La
membrana presenta gran facilidad de penetración para sustancias liposolubles.
Abordaremos dos casos: El paso de sustancias disueltas y el de grandes partículas
A-EL PASO DE SUSTANCIAS DISUELTAS puede ser de dos tipos, según se consuma o no
energía: Pasivo si no consume ATP y activo si consume energía.
1- EL TRANSPORTE PASIVO: Es un proceso de difusión de sustancias a través de la
membrana que se produce siempre con dos características:
- Se da a favor del gradiente,
es decir, de donde hay más
concentración hacia el medio
donde hay menos.
- No hay gasto de energía.
Mediante transporte pasivo
entran en la célula O2, CO2, agua,
sales, monosacáridos, ac. Grasos,
glicerina y aminoácidos.
Dentro de éste se dan otros dos tipos:
1.1- Difusión simple- Es el paso de pequeñas moléculas y lípidos a favor del gradiente; puede
realizarse a través de la bicapa lipídica o a través de canales proteicos.
-Difusión de gases como el oxígeno, el nitrógeno atmosférico y el CO2
-Moléculas lipídicas como las hormonas esteroideas, fármacos liposolubles....
Algunas moléculas polares de
muy pequeño tamaño, como el
agua, el etanol y la glicerina,
también atraviesan la
membrana por difusión
simple. (recordamos que la
difusión del agua recibe el
nombre de ósmosis).

1.2- Difusión simple a través de canales (2).
Es utilizado por moléculas polares pequeñas
como los iones (sodio Na+, potasio K+, calcio Ca2-)
Se realiza mediante las denominadas proteínas
de canal que tienen un orificio o canal interno.
Los iones tienen carga eléctrica, y se necesita
en la membrana una proteína que le sirva de
pasaje. La proteína es un canal, y tiene una
puerta que se puede abrir o cerrar para
permitir el paso. Estos canales no son
específicos, es decir que iones pequeños con la
misma carga pueden entrar. Es a favor de
gradiente y no hay gasto de energía.
1.3- Transporte pasivo facilitado (difusión facilitada). Las
moléculas polares o hidrófilas (glúcidos, aminoácidos...) no
pueden atravesar la doble capa lipídica por difusión a favor
del gradiente de concentración. Requieren que proteínas
trasmembranosas faciliten su paso. Éstas son llamadas
permeasas y actúan como "barcas" para que estas
sustancias puedan salvar el obstáculo que supone la doble
capa lipídica. Las permeasas, all unirse a la molécula a
transportar sufren un cambio en su estructura que arrastra
a dicha molécula hacia el interior de la célula.
Este tipo de transporte tampoco requiere un consumo de
energía, pues se realiza a favor de gradiente.
2-TRANSPORTE ACTIVO Características:
1- Se da en contra gradiente (de concentracióno o eléctrico).
2- Se da mediante proteinas transportadoras específicas instaladas en la membrana.
3- Se da con gasto de energía (ATP).
Se transportan, además de pequeñas partículas, moléculas orgánicas de mayor tamaño.
Son ejemplos de transporte activo la bomba de Na/K, y la bomba de Ca.
1-Difusión 2-poro 3-dif. Facilitada 4-Transp.activo. En todos ellos, se distingue uniporte
cuando se transporta una sola sustancia y cotransporte cuando son dos simultáneamente, pudiendo darse
sinporte o antiporte si el transporte es en el mismo sentido o en sentido contrario. 1-

LA BOMBA DE NA+/K+ se encarga de mantener el equilibrio de iones en la membrana, pues
el sodio Na+ trata de entrar a la célula a favor de gradiente y el potasio K+ de salir.
Hay que evitar esto en todas las células pues es necesario mantener una gran diferencia,
contra gradiente de estos iones. La bomba de Na+/K+ es encarga de ello y requiere una
proteína transmembranosa que bombea Na+ hacia el exterior de la membrana y K+ hacia
el interior.
Esta proteína actúa contra el gradiente gracias a su actividad como ATP-asa, ya que rompe el
ATP para obtener la energía necesaria para el transporte. En el lado interno de la célula, la
proteína se une al sodio Na+ y, la misma proteína, en el lado externo de la célula, se une al
potasio K+. A la bomba se la llama sodio potasio ATPasa
La proteína, al unirse al sodio Na+ del lado interno y al potasio K+ del lado externo, cambia su
forma, dejando al sodio Na+ en el lado externo, y dejando al potasio K+ en el lado interno. Es
decir que saca al sodio Na+ y mete el potasio K+. Se satura la bomba si hay exceso, trabaja a
velocidad máxima. Todas las células del cuerpo tienen esta bomba de sodio potasio, y
mantienen un gradiente. Este sistema se denomina contratransporte porque una sustancia
entra mientras la otra sustancia sale.
COTRANSPORTE: Por este mecanismo, se bombea 3 Na+ hacia el exterior y 2 K+ hacia
el interior, con la hidrólisis acoplada de ATP. El transporte activo de Na+ y K+
tiene una gran importancia
fisiológica. De hecho todas las células
animales gastan más del 30% del ATP
que producen ( y las células nerviosas
más del 70%) para bombear estos.
Un tipo de sinporte (activo) es el caso del transporte de glucosa en el intestino.
Es igual que el anterior pero la proteína transportadora transporta una molécula en contra
de gradiente (glucosa) y otra a favor(sodio), acopladas.
Difusión simple o
tendencia a favor de gradiente

B-TRANSPORTE DE GRANDES PARTÍCULAS:
Las macromoléculas o partículas de mayor tamaño penetran en la célula mediante procesos de
ENDOCITOSIS. El proceso opuesto o salida de estas grandes partículas se llama
EXOCITOSIS. Ambos procesos suponen un tránsito de sustancias envueltas en una membrana
que forma vesículas. Se trata de un mecanismo que sólo es utilizado por algunos tipos de
células, por ejemplo: amebas, macrófagos o las células del epitelio intestinal.
I) ENDOCITOSIS o entrada de grandes partículas que entran en la célula envueltas en vesículas
formadas a partir de la membrana plasmática. Distinguiremos dos tipos de endocitosis: la
fagocitosis y la pinocitosis
a-Fagocitosis: Se llama así cuando son
partículas sólidas (bacterias, restos celulares)
y se da por medio de seudópodos - grandes
evaginaciones de la membrana plasmática- que
envuelven a la partícula. Son un especie de
deformaciones de la membrana y la partícula
pasa al citoplasma de la célula en forma de
vacuola fagocítica. Este tipo de transporte
puede ser de gran importancia en ciertas
células, como por
ejemplo, en los
macrófagos y en las
amebas. Las amebas
son protozoos que se
alimentan por fagocitosis
b-Pinocitosis. Es la ingestión de
sustancias disueltas en forma de
pequeñas gotitas líquidas que atraviesan
la membrana al invaginarse ésta. Se
forman así pequeñas vacuolas llamadas
vacuolas pinocíticas que pueden reunirse
formando vacuolas mayores
II- EXOCITOSIS: Consiste en la 1.secreción o 2.excreción de sustancias por medio de
vacuolas, vesículas de exocitosis, que se fusionan con la membrana plasmática abriéndose al
exterior y expulsando su contenido. Las vacuolas provienen de los sistemas de membranas o
de la endocitosis. La membrana de la vacuola queda incluida en la membrana plasmática . En
todos los mecanismos de endocitosis hay una disminución de la membrana plasmática al
introducirse ésta en el citoplasma. Esta disminución es compensada por la formación de
membranas por exocitosis. La membrana plasmática está en estas células en un continuo
proceso de renovación. En un macrófago, toda su membrana es ingerida en 30 min.
Recommended








