Embed Size (px)
Citation preview

Development of high-efficiency plant genome editing toolkit and its use in
gene functional study
Yao-Guang Liu
刘耀光
South China Agricultural University
华南农业大学

Our objectives
I. Development of a robust, high-efficiency
CRISPR/Cas9 vector and software toolkit
for multiplex genome editing in plants
II. Use of the genome editing toolkit to study
gene function and genetic improvement in
plants

A plant CRISPR/Cas9 multiplex genome editing system
CRISPR/Cas9binary vectors(Ma et al. MP 2015)

sgRNA intermediate vectors
5’-GGCGCTCGCCAAGTTCGACAGGG-3’
5’-GCCGGCGCTCGCCAAGTTCGACA-3’(FP)3’-CGCGAGCGGTTCAAGCTGTCAAA-5’(RP)
Target sequence:
Target adaptor: 5’-GTTGTGTGTGCTTACAGCCATGGC-3’3’-ACACACGAATGTCGGTACCGCAAA-5’
5’-TGTGTGCTTACAGCCATGGCAGG-3’PAM PAMIrregular targetRegular target
19 bp 20 bp
D
Transcript (5’ part): 5’-GGCGCTCGCCAAGTTCGACAGTTT-- 5’-GTGTGTGCTTACAGCCATGGCGTTT--
Extra nucleotide
3’-NCCGCGAGCGGTTCAAGCTGTCCCN-5’Genomic binding site: 3’-NACACACGAATGTCGGTACCGTCCN-5’
NGGTCTCNNNNNN
NCCAGAGNNNNNN
BsaI

(1) Golden Gate ligation
ctcggact
ctgattctaaga
ctgagRNAT1
gRNAT2
gRNAT3
gact gRNA
T4
gagc(B-L )
(B1’) (B2’) (B3’) (B4’)
(B2) (B3) (B4)
Mlu IMlu I
U6b prU6a prU3 prU6c pr
(BL)E.coli Pr
LacZ cggtgcca
Spe I
(B-R )
Assembly of multiple sgRNA expression cassettes into a CRISPR/Cas9 binary vector by single round of cloning
BsaI(B-L)
BsaI(B-R)
pYLCRISPR/Cas9Pubi-H (16419 bp)
RB
Pu
bi
LB
2xP35S HPT/Bar
Cas9p
NLS
pBR322 oriKanR
NLS
pVS1 replicon
pYLCRISPR/Cas9Pubi-B (15889 bp)
T35S
Tnos

U#U#
U#
Ptac promoter F primer
Sp6 promoter primer
(2) Gibson Assembly
Sp6 universal primerT2
T3T4
T1U3
LacZ
Mlu I
Mlu I
GAS-L
Spe I
BsaI(B-L)
BsaI(B-R)
pYLCRISPR/Cas9Pubi-H (16419 bp)
RB
Pu
bi
LB
2xP35S HPT/Bar
Cas9p
NLS
pBR322 oriKanR
NLS
pVS1 replicon
pYLCRISPR/Cas9Pubi-B (15889 bp)
T35S
Tnos
Assembly of multiple sgRNA expression cassettes into a CRISPR/Cas9 binary vector by single round of cloning

Construction of CRISPR/Cas9 binary constructs for targeting multiple genes
Mlu I-digestion
Vector Linked sgRNA cassettes

Simultaneous editing of seven genes

Features of CRISPR/Cas9-based genome editing in rice
53.3
20.0
8.9
64.3
100
80
60
40
20
OsU6a(18, 140)
40.7
39.025.6
OsU6b(9, 59)
OsU6c(5, 39)
OsU3(14, 90)
46.2 54.9
Sum (46, 328)
10.3
90.0
1.781.4 82.1 82,2 85.4
Biallelic Homozygous Heterozygous
21.4
4.35.8
24.7
Edi
ting
rate
(%
)
59.8
26.2
2.5
51.5
16.8
8.9
100
80
60
40
20
30-50 (14, 101)
36.0
51-60(15, 105)
61-85(17, 122)
52.4
77.2
89.6 88.56.7
Biallelic Homozygous Heterozygous
GC%
Edi
ting
rate
(%)
Efficiencies of different U3 & U6 promotersEfficiencies of targets with different GC contents
No. targets No. events

A Web-based CRISPR-GE Toolkit(Web-site:http://skl.scau.edu.cn/)(Xie et al. MP 2017)

targetDesign/offTarget
Submission pageSelect PAM type (NGG, TTN, TTTN)
Select a reference genome (28 plant genomes and 5 animal genomes)
past a genomic sequence for editting
Or define a PAM type by user
Or input a gene locus

Result page
Potential off-target sites and scores
Goto primerDesign

Allelic types of targeted mutations in rice and other diploid organisms
GTCTGAATCTTTTTCACTaGCAGGttgct-3 Allele 1GTCTGAATCTTTTTCACTaGCAGGttgct-3 Allele 2GTCTGAATCTTTTTCACT-GCAGGttgct-3 Reference
Homozygous
mutation
Heterozygous/
biallelic
mutations
Chimeric
Mutations
(very few) Sequencing chromatogram with overlapped peaks

• Cloning of the PCR amplicons and sequencing multiple clones (>10) for each target site are time-consuming and high-cost !
• To resolve this problem, we developed a web-
based tool “DSDecode” and its updated version
“DSDecodeM”

Submission page of DSDecodeM
Past a wild-type reference sequence
Upload up to 20 sequencing files (ab1 file)

Results outputDownload results in a txt file

Worldwide decoded sequencing file number of targeted editing per month by DSDecode/DSDeceodeM
3661241
16141418
1174
1251
597 5111757
2508
2223
2247
2239
5003
5987
4645
7542
5870
3594 3768
4562
4747
57128268
13517
18588
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000

Use of the genome editing toolkit to
study gene function and genetic
improvement in plants

Strategies for targeted editing of genes
CRE: cis-regulatory element
Targeted editing in CDS of a gene
Targeted editing in one, two, or multiple CREs of a gene
*
Qualitative or quantitative variation of the trait
Quantitative variation of the trait
A
B
Cis-regulatory mutant alleles with altered expression level/pattern
Null-mutant alleles
Targeted editing in duplicated genes
Quantitative variation of
the trait
Gene dosage-reduced mutant alleles
*
A a-1
A1 A2 A1 a2
A1 A2 A2
CREsC
A
A
A
A
A-1
A-2
A-3
A-4
A-n
A1
a-2

Study on rice hybrid sterility by genome editing
栽培稻Cultivated rice
亚洲稻
非洲稻
粳稻
籼稻
(Oryza genus)
(Species)
(Subspecies)(Indica)
(Japonica)
(O. sativa)
(O. glaberrima)
野生稻Wild rice
Hybrid sterility (HS)
(male, female, male/female)

Cloned hybrid sterility genes in rice
Locus Sterility Gene structure Gene product Reference
Sa Male Two adjacent F-box, Long et al., 2008;genes SaF/SaM SUMO-ligase Xie et al., 2017a
S5 Female Three adjacent Aspartic Chen et al; 2008
genes protease & ? Yang et al., 2012
S27/S28 Male Duplicated Ribosomal protein Yamagata et al., 2010
genes DPL1/ Male Duplicated Small unknown Mizuta et al., 2010
DPL2 genes proteins
Sc Male Copy number DUF1618 protein Shen et al., 2017
variation
HSA1a/ Male Two adjacent DUF1618 protein, Kubo et al., 2015
HSA1b genes unkown proteins
S7 Female Single gene TPR protein Yu et al; 2016
S1 Female/male Multiple genes TPR protein/unkown p Xie et al; 2017b

S1 locus-mediated hybrid sterilityin Asian-African rice hybrids
SS: Semi-Sterility
FF: Full Fertility
RP(S1-s) NIL(S1-g) F1(S1-s/S1-g)
SSFFFF
FF FF SS

Map-based cloning of S1
No. recombinant
Centromere(Chr.6)
*
1 2 3 4 5 6 7OsORF
1 5 10 15 17Exon
TCA TAA (stop codon)
*
2 000 0
Chromosomal position (kb)
2
OgTPR1
OsTP1
IF2 IF31 3 4 6 7OgORF a2 a3 a4 a5 a6a1
IF1 IF4 IF5
Trypsin-like peptidase RRS
Trypsin-like peptidase
(RRS, ribosome biogenesis regulatory protein domain)
S1(Xie et al., Mol Plant 2017)

Knocked out of OgTPR1 and OgORF7 did not affect male and female fertility
reference
Allele 1Allele 1
T0
(ogtpr1
)
D
CCATAG-CGATGCCTGAGCATG
CCATAGGCGATGCCTGAGCATGCCATAGGCGATGCCTGAGCATG
Aogtpr1 ogorf7B
reference
T0(ogorf7-1)
Allele 1Allele 1
TGGGGAACTTGGAC-TGGCGGTGGGGAACTTGGAC-TGGCGG
TGGGGAACTTGGACTTGGCGG

Knockout of OgTPR1 could rescue fertility in the interspecific hybrids
F1
NIL×RP-s
mF1
ogtpr1×RP-s
mF1
ogorf7×RP-s
PopulationGeno-type
No. plant
Pheno-type
χ2
(1:2:1)
g/gg/ss/s
FFFFFF
7015575
0.50mF2 of
(ogtpr1×RP-s)
mF2 of(ogorf7×RP-s)
g/gg/ss/s
FFSS
-
180150
472**
F2 of(NIL-g×RP-s)
g/gg/ss/s
FFSSFF
1194136
52694**

The S1-hybrid sterility involves at lest three linked genes
IF2 IF3
1 3 4 OgTPR1 OgORF7OgORF a2 a3 a4a1
IF1 IF4 IF5
* * * *
mF1-ogorfa2 mF1-ogorfa4 mF1-ogorfa5 mF1-ogorfa6
hypothetical
proteinhypothetical
protein
hypothetical
proteinAP2
TF
Typsin-like
proteinTypsin-like
protein
*Fertility-rescued
a5 a6
Mutant hybrids
Fertility-rescued

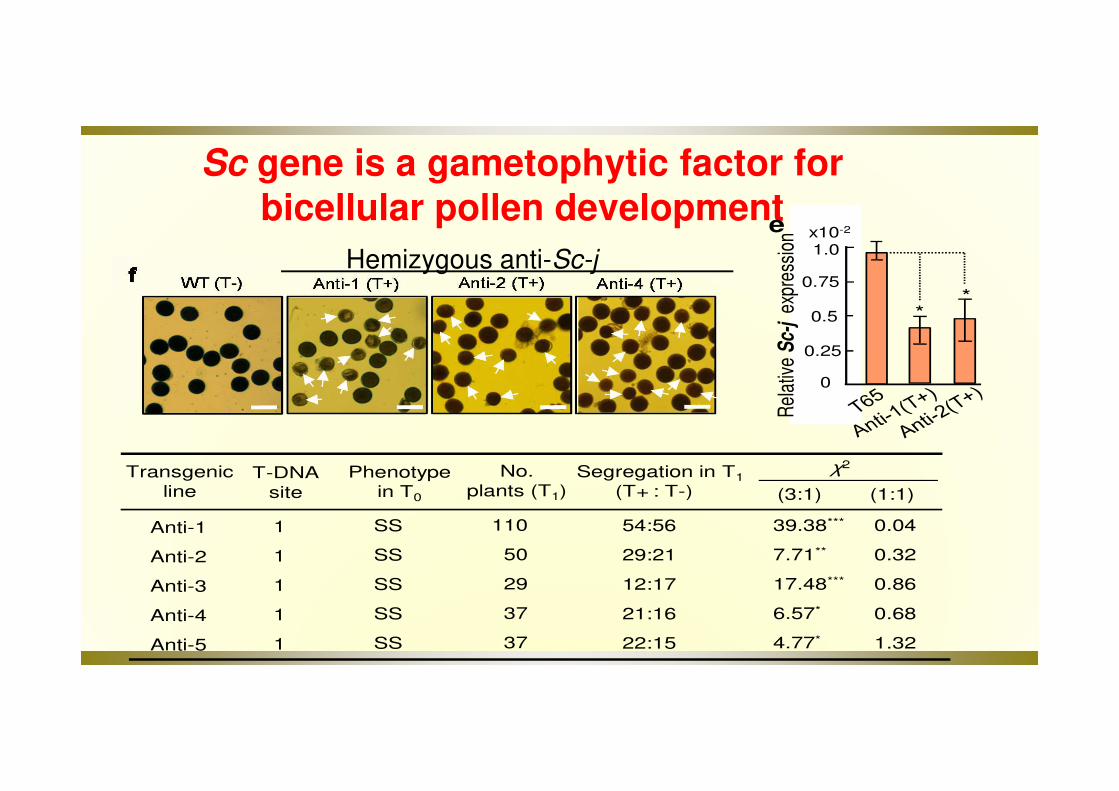
Sc-hybrid showed semi-male sterility and produced severe allele transmission ratio distortion (TDR)
FF: Full fertility;SS: Semi Sterility
• T65 (Sc-j/Sc-j): a japonica cultivar Taichung 65
• E5 (Sc-i/Sc-i): a near-isogenic line of T65 with Sc-i
Pollen grains carrying the Sc-j allele were selectively aborted

Map-based cloning revealed a large structural change and copy number variation (CNV) at Sc
Sc-i is a recombinant new gene with CNV, and the promoter regions between Sc-j and Sc-i differ completely
Sc locus90.5
0 BAC: OSJNBb0078P24
80.4
5
84.485.792.093.0100.7
2 0 24No. recombinant:(from 8689 F2 plants)
38.0114.0Marker (kb):
0
E14-E2RG227
Chr. 3
Marker
8.6 kb
Sc-j (Os03g0247300)
japonica cv: T65, Nip, Koshihikari,
Hitomebore, HEG4, A123
Sc-ib2Sc-is
28.4 kb
T1 T2
E5
Sc-ib1MH63
93-11
14.9 kb
Sc-iaSc-ib2 Sc-ib1Sc-is
28.2 kb28.4 kb
indica cv:
23.9 kb 23.9 kb
23.9 kb

Sc alleles encode diverged DUF1618 proteins
VHTFPFPVSGVVWVGRAAVLRHAGGGGGGGDGTTASYVIAELLRPFHGSLPDATLVMWLS 180
VHTFPFPVSGVVWVGTAAVLRHAGGGGGG-DGTTA-YVIAELLRPFHGSLPDATLVMWLS 177
PIISRFSMLISRKAIRVVKSVRVRCADKSLVLFYAGTGFPGFSSHGCHLIYDAIDGSLTA 120
PIVSRFSMLISRKAIRVVKSIHVECADKSLVLFYAGTGFPGFS-HGCHLIYDAIDGSLTA 119
MAPTWVLLDRFVKPTIFDEEESKGKGESTGAPVKYLPARLRQEVPAGMRDVKPYPEVADP 60
MAPAWVLLDRVVKPAVFDEEESKGKGESTGAPVKYLPARLRQEVPAGMRDVKPYPEVADP 60
MAPAWVLLDRVVKPAVFDEEESKGKGESTGAPVKYLPARLRQEVPAGMRDVKPYPEVADP 60
Sc-jSc-ia
Sc-ib1/ib2
PIVSRFSMLISRKAIRVVESIHVECADKSLVLFYAGTGFPGFS-HGCHLIYDAIDGSLTA 119
Sc-jSc-ia
Sc-ib1/ib2
VHTFPFPVSGVVWVGTAAVLRHAGGGGGG-DGTTA-YVIAELLRPFHGSLPDATLVMWLS 177
Sc-jSc-ia
Sc-ib1/ib2
NSPASTSGSNGQWVKEDVRLPGEVCTGTDPFTTDLVFSFGESCLCWADLFMGILFCDLAT 240
NSPASTSGSNGQWVKEDVRLPGEVCTGTDPFTTDLVFSFGESCLCWADLFMGILFCDLAT 237
NSPASTSGSNGQWVKEDVRLPGEVCTGTDPFTTDLVFSFGESCLCWADLFMGILFCDLAT 237
LRAPRFRFIPLPKACSFDPVGKYGRPHMPEFRSMGRVNGVIRLIDMEGFTNEYLAVDEVK 300
LRAPRFRFIPLPKACSFDPVGKYGRPHMPEFRSMGRVNGVIRLIDMEGFTNEYLAVDEVK 297
LRAPRFRFIPLPKACSFDPVGKYGRPHMPEFRSMGRVNGVIRLIDMEGFTNEYLAVDEVK 297
Sc-jSc-ia
Sc-ib1/ib2
Sc-j
Sc-iaSc-ib1/ib2
LTIWTLSADLSEWEKGPVCTVGDIWASEEFVAMGLPQLRPMCPVLSMVDEDVVCVVMTEV 360
LTIWTLSADLSEWEKGPVCTVGDIWASEEFVAMGLPQLRPMCPILSMVDEDVVCVVMTEV 357
LTIWTLSADLSEWEKGPVCTVGDIWASEEFVAMGLPQLRPMCPVLSMVDEDVVCVVMTEV 357
Sc-j
Sc-iaSc-ib1/ib2
EIEESDVTDFDDEGNKLKFKAQYVLDIDVRRKRVLSITQHHIESMGDLIPDLIACEFTAY 420
EIEESDVTDFDDEGNKLKFKAQYVLDIDVRRKRVLSITQHHIESMGDLIPDLIACEFTAY 417
EIEESDVTDFDDEGNKLKFKAQYVLDIDVRRKRVLSITQHHIESMGDLIPDLIACEFTAY 417
Sc-jSc-ia
Sc-ib1/ib2
SELSKGMQAMVEGNEGEESTKRMKVK 446
SELSKGMQAMVEGNEGEESTKRMKVK 443
LELSKGMQAMVEGNEGEESTKRMKVK 443
Sc-j
Sc-iaSc-ib1/ib2

DNA fiber-FISH verified the Sc-i CNV

indica cultivars have different copies (2 or 3) of Sc-i


Structures of Sc-j and the duplicated Sc-i paralogs
Sc-ia
Sc-ib1
Sc-j
Sc-ib2
Exon1(1380 bp)
Exon2(62 bp)
P6
P727-bp deletion
P1
P2
P5
P7
P7
P5
243-bp insertion
3x3-bp deletion
243-bp insertion
Promoter PSc-j
Promoter PSc-ia
PSc-ib
PSc-ib
A G G C C
A G G C C
G A A T T
G A A T T
P7
P6 P5
P5P6
P2
P2
P2
1363 bp
451 bp
ATG
Intron (1509 bp)
P3
P3
P3
P3
P8
P8
P8
P8
P4
P4
P4
P4
TGA

(RiceXPro,
http://ricexpro.dna.affrc.go.jp/)
Sc-j and Sc-i have different expression patterns and levels
OsActin 1 (25 cycles)
Sc
(28 cycles)
M
T65 E5a
kb
1.00.75
0.5
1.00.75
0.5
b
OsActin 1
Sc-j
Mean value = 2413 in 1.6-2.0 mm anthers
Mean value = 58620 in 1.6-2.0 mm anthers
Sc-j : Actin 1 = 0.04

Sc-j and Sc-i have different expression patterns and levels
x10-2
1
7
6
0Expr
essi
on (S
c-j/A
ctin
1)
5
Sc gene is a gametophytic factor for bicellular pollen development
e
0
0.5
0.25
x10-2
0.75
1.0
Rel
ativ
e S
c-j
expr
essi
on
**
Transgenic
lineT-DNA
site
Phenotype in T0
No.
plants (T1)Segregation in T1
(T+ : T-)
χ2
(3:1) (1:1)
Anti-1
Anti-2
Anti-3
Anti-4
Anti-5
1
1
1
1
1
SS
SS
SS
SS
SS
110
50
29
37
37
54:56
29:21
12:17
21:16
22:15
39.38***
7.71**
17.48***
6.57*
4.77*
0.04
0.32
0.86
0.68
1.32
Hemizygous anti-Sc-j
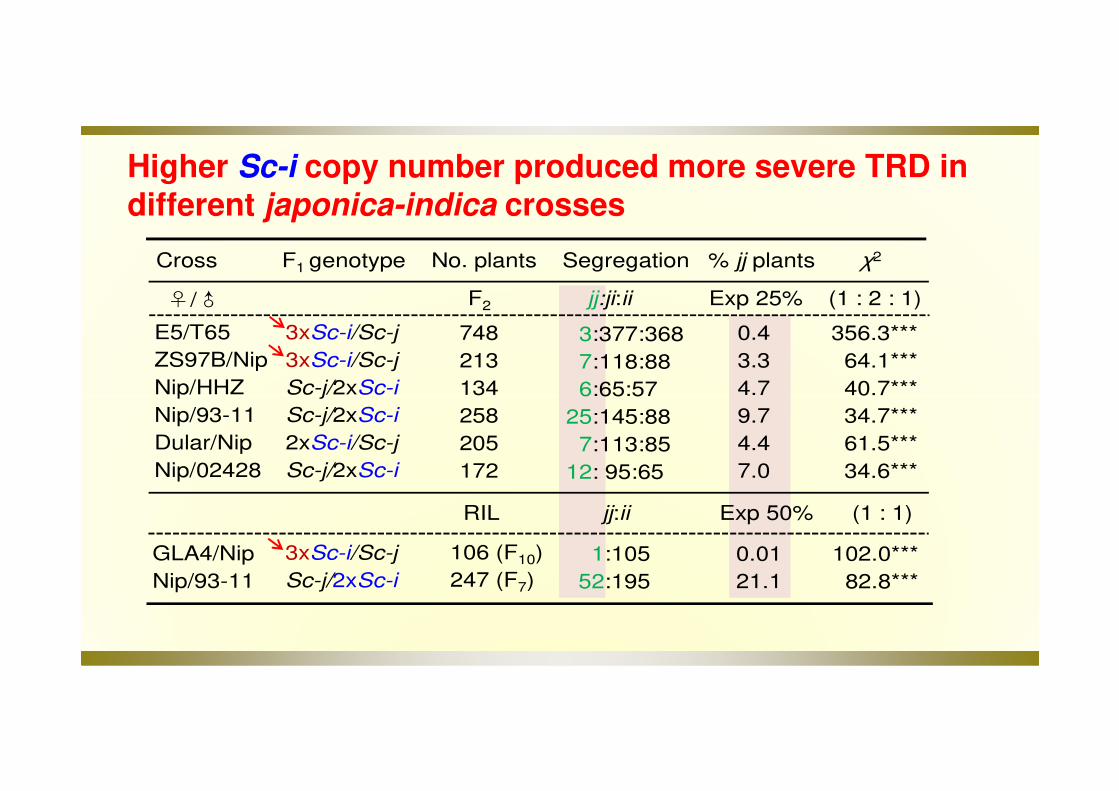
Higher Sc-i copy number produced more severe TRD in different japonica-indica crosses
Cross F1 genotype No. plants Segregation % jj plants χ2
E5/T65
ZS97B/Nip
Nip/HHZ
Nip/93-11
Dular/Nip
Nip/02428
F2 jj:ji:ii Exp 25% (1 : 2 : 1)
3xSc-i/Sc-j
3xSc-i/Sc-j
Sc-j/2xSc-i
Sc-j/2xSc-i
2xSc-i/Sc-j
Sc-j/2xSc-i
748
213
134
258
205
172
3:377:368
7:118:88
6:65:57
25:145:88
7:113:85
12: 95:65
356.3***
64.1***
40.7***
34.7***
61.5***
34.6***
0.4
3.3
4.7
9.7
4.4
7.0
RIL jj:ii Exp 50% (1 : 1)
106 (F10)
247 (F7)
1:105
52:195
0.01
21.1
102.0***
82.8***
GLA4/Nip
Nip/93-11
3xSc-i/Sc-j
Sc-j/2xSc-i
♀/♂

The Sc-j expression in the hybrid was largely suppressed
Early-bicellular anthers
Purified early-bicellular pollen
T65 F1
0.25
0.5
0.75
1.25
1.0
x10-2
Rel
ativ
e S
c-j
expr
essi
on
0
x10-2
0.5
1.0
1.5
2.5
2.0
T65 F1
0
***
(The expression levels were calculated as per Sc-j copy)

Deletion and mutation of one or two of the Sc-i copies by CRISPR/Cas9 editing
E5-d1Sc-iaSc-ib1Sc-is 28.4-kb deletion
ATTTCGCGGAAGGCGAT------AGTGGAATCCA
60 bp
ATTTCGCGGAAGGCGATCAGGGTTAGTGGAATCCA
E5-ed1Sc-ib1ed1
Sc-ib2ed1
E5-ed2Sc-ib1ed2
Sc-ib2ed2
P9c
P9d
GATTTCGCGGAAGGC-------------GAATCCA
AAGCCCTACCCAGA GGTAGTGGAATCCA--- ---
P9b
P9e
13 bp
ATTTCGCGGAAGGCGATCAGGGTAGTGGAATCCA
ATTTCGCGGAAGGCGATCAGGGTAGTGAAATCCG
PAM
Sc-ib1/ib2
Sc-iaE5
exon 1
P10
P9a

The edited Sc-i alleles are hybrid-compatible, overcoming male fertility in the hybrids
(1 copy deleted) (2 copies mutated) (2 copies mutated)

D E
0.25
0.5
0.75
1.25
1.0
x10-2
0
*
****
Exp
ress
ion
pe
r co
py
(Sc-j
/Act
in 1
)
0.1
0.05
0.025
0
0.075Sc-ia
Sc-ib1ed1
Sc-ib1ed2
Sc-ib2ed1
Sc-ib2ed2
Sc-ia/ib1/ib2
Exp
ress
ion
(S
c-i
/Act
in 1
)
The edited hybrids had normal male fertility and genotypic segregation

Indica-japonica hybrid male sterility at the Sc locus is caused by the duplications of the indica allele gene Sc-i with a gene-dosage effect
(Shen et al., Nat Commun 2017)
Sc-i acts as both the Sc-j-pollen killer and the self-protector
This molecular model well explains the classical “one-locus sporo-gametophytic interaction” model for hybrid sterility
Allelic suppression

SaF SaM
T860 (E4) AG(E4) TGA (T65)
TGA (E4)ATGATG TAGC860 (T65) AT(T65)
P0013G02 (163.4 kb)
A10-2 G02-148G02-45Centromere
~5.4 Mb
63
Recombinant Phenotype
E4
T65
G392 MSS
MSS
MFFA408
H70 MFF
MFFC513
MFFH70T65
MFFMFFG735MFFA408T65
H122
MSS
MSSC513H70
370 kb
69 79.6 (kb)78.576.374.6737269.8G02-
H71
MFFLS38
G02-84
(Chr. 1)
*
*
*
*
SaF SaM
T860 (E4) AG(E4) TGA (T65)
TGA (E4)ATGATG TAGC860 (T65) AT(T65)
P0013G02 (163.4 kb)
A10-2 G02-148G02-45Centromere
~5.4 Mb
63
Recombinant Phenotype
E4
T65
G392 MSS
MSS
MFFA408
H70 MFF
MFFC513
MFFH70T65
MFFMFFG735MFFA408T65
H122
MSS
MSSC513H70
370 kb
69 79.6 (kb)78.576.374.6737269.8G02-
H71
MFFLS38
G02-84
(Chr. 1)
*
*
*
*
The heterozygous SaM and at least one E4 (indica) SaF allele are required for the hybrid male sterility.
Male Semi-Sterility
Male Full Fertility
The Sa complex locus for japonica-indica hybrid male sterility (Long et al., PNAS 2008)

Knockout of SaF or SaM of the Sa locus could rescue fertility in japonica-indica hybrids
(Xie et al. JIPB 2017)
Knock out of SaF or SaM
did not affect male fertility
The mutant japonica-indicahybrids with mutant saf or samshowed normal male fertility

Rapid breeding of glutinous rice by editing Waxy
(Ma et al., MP 2015)
WT T0-1 T0-2 T0-3 T0-4
Knockout of the transporter genes (OsGSTU, OsMRP15, and OsAnP) in a purple-leaf rice destroyed the anthocyanin biosynthesis pathway

Summary
• The hybrid sterility loci often include multiple functionally-related genes
• The CRISPR/Cas9 toolkit is powerful for genome editing in plants
• The genome editing technology is very useful for study of gene function and genetic improvement, especially for breaking down of the reproductive barrier in hybrid breeding

ACKNOWLEDGMENTS
• Ma Xingliang, Xie Xianrong, Liu Weizhi, Shen Rongxing, other members of our lab
• MOST of China





























