Embed Size (px)
Citation preview

IAWA Journal 41 (3) 2020 278ndash300
Wood anatomy of the neotropical liana lineage Paullinia L(Sapindaceae)
Joyce G Chery12 Israel L da Cunha Neto3Marcelo R Pace4Pedro Acevedo-Rodriacuteguez5 Chelsea D Specht2 and Carl J Rothfels1
1University Herbarium and Department of Integrative Biology University of CaliforniaBerkeley CA 94720 USA
2School of Integrative Plant Sciences and LH Bailey Hortorium Cornell University Ithaca NY 14853 USA3Laboratory of Plant Anatomy Department of Botany Instituto de BiociecircnciasUniversidade de Satildeo Paulo Rua do Matatildeo 277 Satildeo Paulo SP 05508-090 Brazil
4Department of Botany Instituto de Biologiacutea Universidad Nacional Autoacutenoma de MeacutexicoTercer Circuito sn de Ciudad Universitaria Mexico City 04510 Mexico
5Department of Botany Smithsonian National Museum of Natural History 10th Street andConstitution Avenue NWWashington DC 20560 USA
Present address Department of biology The Pennsylvania State University State College PA 16803 USACorresponding authors email joycegcherygmailcom israellopescngmailcom
Accepted for publication 12 May 2020
ABSTRACT
The liana genus Paullinia L is one of the most speciose in the neotropics and isunusual in its diversity of stem macromorphologies and cambial conformationsThese so-called ldquovascular cambial variantsrdquo are morphologically disparate evolu-tionarily labile and are implicated in injury repair and flexibility In this study weexplore at the finer scale how wood anatomy translates into functions related tothe climbing habitWepresent thewood anatomyof Paullinia anddiscuss the func-tional implications of key anatomical featuresWood anatomy characters were sur-veyed for 21 Paullinia species through detailed anatomical study Paullinia woodshave dimorphic vessels rays of two size classes and both septate and non-septatefibers Fibriformvessels fusiformaxial parenchyma andelementsmorphologicallyintermediate between fibers and axial parenchyma were observed Prismatic crys-tals are common in the axial andor ray parenchyma and laticifers are present inthe cortex andor the early-formed secondary phloem Some features appear asunique to Paullinia or the Sapindaceae such as the paucity of axial parenchymaand the abundance of starch storing fibers Although many features are conservedacross the genus the Paullinia wood anatomy converges on several features of theliana-specific functional anatomy expressed across distantly related lianas demon-strating an example of convergent evolution Hence the conservation of wood
copy International Association of Wood Anatomists 2020 DOI 10116322941932-bja10027Published by Koninklijke Brill NV Leiden
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 279
anatomy in Paullinia suggests a combination of phylogenetic constraint as a mem-ber of Sapindaceae and functional constraint from the liana habit
Keywords Convergent evolution cambial variants Paullinieae Sapindales vines
INTRODUCTION
The stemmorphology andwood anatomy of lianas are critical to understanding the climb-ing habit as herein lies distinctive features that separate this woody growth form fromtrees and shrubs In comparison to erect plantsmdashat the cellular scalemdashlianas have fewerfibers and allocate more cross-sectional area to water conduits wide vessels andor tra-cheids (Carlquist 1991 Angyalossy et al 2012 2015) Additionally non-lignified ray and axialparenchyma are more commonly found in lianas than in self-sustaining plants (Carlquist1991 Angyalossy et al 2012 2015) This combinationof anatomical features referred to as theldquoliana syndrome anatomyrdquo translates into eco-physiological functions that facilitate opti-mization of conductance flexibility and strength (Carlquist 1985 1991 FisherampEwers 1989RoweampSpeck 2004 Angyalossy et al 2012 2015) Consistent across lianas sensu lato includ-ing ferns gymnosperms monocots andmagnoliids the liana syndrome is even diagnosticfor identifying lianas in the fossil record (Krings et al 2006 Burnham 2009 Rozefelds ampPace 2018) At the macro-scale the gross morphology of liana stems is often unusual theseatypical configurations are called ldquocambial variantsrdquo (Carlquist 2001)Vascular cambial vari-ants arise as non-circular stem outlines aberrant tissue conformations andor shifts in therelative abundance of secondary xylem (wood) and phloem (inner bark) produced by thevascular cambiumTaken together both the cellular scale (micromorphology) and the stemmorphology (macromorphology) seem to be altered in lianas as a result of climbing adap-tations
Paullinia L (Sapindaceae) is a genus of ca 220 species of mostly neotropical lianas(Acevedo-Rodriacuteguez et al 2017 Chery et al 2019) Together with five other genera oflianas (Serjania L Cardiospermum Kunth Urvillea Kunth Lophostigma Radlk and Thi-nouiaTrianaampPlanch) Paullinia constitutes themonophyletic Paullinieae tribe (Acevedo-Rodriacuteguez et al 2017 Chery et al 2019) which contains approximately one-third of thespecies diversity of Sapindaceae (Buerki et al 2009) Paullinieae display an impressive as-sortment of different modes of fruit dispersal (Acevedo-Rodriacuteguez et al 2011 2017 Chery etal 2019) and diversity of secondary growth ontogenies (Schenk 1893 Radlkofer 1895 Pfeif-fer 1926 Obaton 1960 Van der Walt et al 1973 Klaassen 1999 Tamaio amp Angyalossy 2009Tamaio amp Somner 2010 Tamaio 2011 Bastos et al 2016 Cunha Neto et al 2018 Chery et al2020)
As in many liana lineages several species of Paullinia have vascular cambial variantsIndividual Paullinia species exhibit one or more of the following types of cambial variants(i) phloem wedges produced by elevated production of secondary phloem relative to sec-ondary xylem by the interfascicular cambium (ii) lobed xylem produced by the elevatedproduction of secondary xylem by the fascicular cambium (iii) compound stems gener-ated by the presence of multiple procambial strands forming multiple vascular cylinders
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
280 IAWA Journal 41 (3) 2020
and (iv) successive cambia generated by de novo cambial strands formed by the pericycle(Radlkofer 1895 Schenk 1893 Pfeiffer 1926 Obaton 1960 Van der Walt et al 1973 Klaassen1999 Tamaio amp Angyalossy 2009 Tamaio amp Somner 2010 Tamaio et al 2011 Tamaio 2011Bastos et al 2016 Cunha Neto et al 2018 Chery et al 2020) As cambial variants are asso-ciated with the repeated evolution of lianas across vascular plants these forms have beenimplicated in overcoming challenges related to the stress of climbing stems such functionsinclude injury repair as a regenerative source of essential vascular tissues (ie successivecambia Carlquist 2007) and stem flexibility (Dobbins amp Fisher 1986 Fisher amp Ewers 1989Carlquist 1991 Rowe et al 2006 Isnard amp Silk 2009)
Our current understanding of the distinctiveness of liana wood anatomy is the resultof the tremendous efforts by numerous authors who have detailed the anatomy of lianasfrom different lineages (Schenk 1893 Pfeiffer 1926 Carlquist 1991 Caballeacute 1993 Pace et al2009 2015 2018a bWagner et al 2012 Pace ampAngyalossy 2013) To contribute to this effortof linking wood anatomy to plant form and function we here present the wood anatomy ofPaullinia and discuss our anatomical results in the context of broad patterns of convergentevolution among lianas
MATERIALS ANDMETHODS
Extant species sampling tissue embedding sectioning and stainingWe studied the wood anatomy of 21 Paullinia species (one is as yet unidentified) that
broadly span the breadth of the Paullinia phylogeny (Chery et al 2019) Material was ob-tained from field collections in Central and South American localities (Table 1) Additionalsamples were obtained by sampling mature stems from herbarium vouchers and rehydrat-ing them by boiling them in 1 ml of glycerin + 100 ml of water for approximately 1 hourField-collected sampleswere immediately fixed in formaldehyde-acetic acid-alcohol (25ml95 ethanol 25 ml glacial acetic acid 5 ml of formalin (37 formaldehyde) + 175 ml wa-ter in a 50 ml falcon tube) then permanently stored in 70 ethanol following Johansen(1940)
Stems were gradually embedded in polyethylene glycol (PEG) 1500 (Rupp 1964) as de-scribed in Barbosa et al (2010) Stems were placed in a 1-liter beaker melted PEG waspoured to cover the specimen and the beaker was filled to 1 liter with water Beakers withstems + PEG + water were placed in a paraffin oven to allow the water to slowly evapo-rate and for the PEG to gradually infiltrate the stems (approxmately 9 days) Stems andPEG were poured into rectangular paper molds to generate PEG blocks Steel knives weresharpened with sandpaper (in a progressive series of grades 600 1200 1500 2000 follow-ing Barbosa et al 2018) Stems in PEG blocks were then sectioned with sharpened steelknives on a sliding microtome each section was coated with a polystyrene foam solution(Barbosa et al 2010) Sections were stained with Safrablau (nine parts 1 Astra Blue (1 g ofAstra Blue + 100 ml of 50 ethanol) to one part 1 Safranin O (1 g of Safranin-O in 100 mlof 50 ethanol) Bukatsch 1972modified by Kraus ampArduin 1997) washed with an ethanolseries (50 60 70 80 95 then 100 EtOH) followed by a xylene series (11 100EtOH100xylene then 100xylene) then finallymountedwith Eukittreg quick-hardening
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
CheryetalndashWoodanatom
yofPaullinia281
Table 1Material obtained from field collections in Central and South American localities
Species Collector Collection number (Herbarium code) Locality
Paullinia alata (Ruiz amp Pav) GDon Brandbyge 33347 (US) Pace 694 (US) San Pablo de los Secoyas Napo Ecuador Reserva Bioloacutegia La SelvaCosta Rica
Paullinia baileyi Standl Chery 37 (UC) Barro Colorado Island PanamaPaullinia bracteosa Radlk Chery 26 (UC) Chery 39 (UC) Barro Colorado Island PanamaPaullinia dasystachya Radlk Acevedo 14295 (US) Madre de Dios PeruPaullinia echinata Huber Schunke 2310 (US) Huaacutenuco PeruPaullinia exalata Radlk Couvreur 207 (US) Ixiamas La Paz BoliviaPaullinia fibrigera Radlk Chery 16 (UC) Chery 21 (UC) Barro Colorado Island PanamaPaullinia fimbriata Radlk Day 9005 (US) Acre BrazilPaullinia latifolia Benth ex Radlk Acevedo 5835 (US) Brasil 528 (US) Tumuc Humac Mts Talouakem Suriname Acre BrazilPaullinia micrantha Cambess Somner 1597 Somner 1562 Tinguaacute Brazil PARNASO BrazilPaullinia obovata (Ruiz amp Pav) Pers Mathias 3585 (US) San Martin PeruPaullinia pinnata L Chery 30 (UC) Chery 41 (UC) Barro Colorado Island PanamaPaullinia pseudota Radlk Bastos 1 Bona 5 Bona 15 Usina Satildeo Joseacute Brazil Jacarepiaacute Brazil Cagarras BrazilPaullinia rubiginosa Cambess Acevedo 14366 (US) Madre de Dios PeruPaullinia rugosa Benth ex Radlk Chery 10 (UC) Barro Colorado Island PanamaPaullinia sp Chery 46 (UC) Barro Colorado Island PanamaPaullinia spicata Benth Oldeman 1248 (US) Riviegravere Comtegrave French GuianaPaullinia sprucei JF Macbr Acevedo 7519 (US) Schunke 15407 (US) Napo Province Yasuni Forest Reserve Ecuador Sacarita del Riacuteo
Utiquinia Cornoel Portillo PeruPaullinia trigonia Vell Somner 1449 Somner 1609 Somner 1635 PARNAI Brazil Tinguaacute Brazil PARNAI BrazilPaullinia turbacensis Kunth Acevedo 15176 (US) Chery 13 (UC) Barro Colorado Island PanamaPaullinia weinmanniifoliaMart Acevedo 3698 Somner 1610 amp 1497 Espirito Santo Reserva Valle do Rio Doce Brazil APAMaricaacute Brazil
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
282 IAWA Journal 41 (3) 2020
mounting medium to prepare permanent slides Stems were sectioned in transverse tan-gential and radial planes
Macerations were performed to identify the different cell types of the secondary xylemSamples were incubated for 48 hours in equal part solutions of hydrogen peroxide + glacialacetic acid (Franklin 1945) then stained with Safranin-O
Slides were analyzed under (i) Zeiss Stemi 508 stereomicroscope (Carl Zeiss AGOberkochen Germany) and imaged with a Canon EOS M3 (Canon Ōta Tokyo Japan)(ii) Zeiss AxioImager light microscope (Carl Zeiss Oberkochen Germany) and imagedwith a QImaging 5MPix MicroPublisher camera (Teledyne QImaging Surrey BC Canada)and (iii) Olympus BX50 (Olympus Hatagaya Japan) and imaged with a Media Cybernet-ics CoolSNAP-Pro digital camera (Media Cybernetics Silver Springs MD USA) Slides areavailable upon request from the corresponding authors
Character codingAnatomical descriptions followed the IAWA Hardwood List (IAWA Committee 1989) as
a starting point adjusting for the Paullinia-specific features that we observed Due to thepresenceof vessel dimorphismwe report ldquonarrowvessel diameterrdquo and ldquowide vessel diame-terrdquo as two separate charactersTo this aim for each specieswemeasuredall vessels (at least125 vesselsspecies) by first thresholding the image then using the ldquoanalyze particlesrdquo func-tion in Image J 152a (Schneider et al 2012)We generated histograms of the log-transformeddistribution of vessel diameters to identify the peaks in the distribution that correspondto the narrow and wide vessel classes (see Distributions in the Appendix) The mean andstandard deviations of these respective classes were calculated (Tables 2 and 3) and boxand whisker plots are presented in Fig 3 For species with compound stems all charactersare reported from the central vascular cylinder (Tamaio et al 2011) Also for species withsuccessive cambia we measured the vessel element diameters in the central cylinder andthe peripheral vascular strands to test if there is a difference between this feature in the reg-ular versus variant xylemwithin a species Statistical analyses were performed in R (R CoreTeam 2018) and all measurements were performed in Image J 152a (Schneider et al 2012)
RESULTS
Cambial variantsSix different stem ontogenies are recognized in Paullinia (Chery et al 2020) regular I
regular II phloemwedges lobedxylem compound stems and successive cambia (Fig 1 andexplored in detail in Chery et al 2020) Regular I is observed in one species (P fibrigera)regular II in three species (P echinata P sprucei and P sp) phloem wedges in 10 speciesmdash including in the lobed xylem of P obovata and in the central cylinder of the compoundstemsof P alata andP spicata lobed xylem inone species compound stems in four speciesand successive cambia is observed in P weinmanniifolia P micrantha P pseudota and Ptrigonia
Growth ringsAll species of Paullinia have diffuse-porous woods and indistinct growth rings (Fig 2A)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
CheryetalndashWoodanatom
yofPaullinia283
Table 2Wood anatomy characters in Paullinia spp vessels
Species Cambialvariantpresent
Cambial varianttypea
Vessel groupingsb Vessel diameter(μm)
Vesselsmm2 Tracheidspresent
Intervessel pitswith slit-likecoalescentapertures
Gum-likesubstance in
vesselspresent
Alternatingthin-walled andthick-walled
fibers
Narrow Wide Narrow Wide
Paullinia alata + Comp with 3 PCPW in the CC
RM of 2ndash4 and 9ndash13 S and TM of 2 30 plusmn12 117 plusmn52 97 plusmn21 + + ndash +
Paullinia baileyi + PW S and RM of 2ndash4 S 21 plusmn 7 62 plusmn12 37 plusmn14 + + +Paullinia bracteosa + PW RM of 6ndash8 and 9ndash15 S and TM of 2 24 plusmn 5 93 plusmn33 91 plusmn25 + + ndash +Paullinia dasystachya + PW RM of 2ndash4 and 5ndash9 S TM and RM of 2 26 plusmn 5 84 plusmn31 37 plusmn 4 ndash + ndash +Paullinia echinata ndash NA RM of 2ndash4 S TM and RM of 2 31 plusmn10 163 plusmn50 63 plusmn19 + ndash + ndashPaullinia exalata + Comp with 3 PC RM of 2ndash4 and 5ndash9 S and TM of 2 17 plusmn 8 89 plusmn38 64 plusmn12 + + +Paullinia fibrigera ndash NA S and RM of 2ndash5 S and TM of 2 14 plusmn 3 106 plusmn52 74 plusmn22 + + ndash +Paullinia fimbriata + PW RM of 6ndash9 S and TM of 2 24 plusmn 4 83 plusmn28 75 plusmn15 + + + +Paullinia latifolia Very shallow PW RM of 3ndash4 S and TM of 2 40 plusmn11 165 plusmn68 129 plusmn26 + ndash ndashPaullinia micrantha + SC RM of 6ndash9 S and TM of 2 34 plusmn11 171 plusmn60 85 plusmn20 + + +Paullinia obovata + LX PW RM of 3ndash4 S TM and RM of 2 13 plusmn 2 44 plusmn21 80 plusmn13 + + Paullinia pinnata + Comp with 3 PC RM of 3ndash4 S and TM of 2 28 plusmn11 101 plusmn36 70 plusmn10 + + +Paullinia pseudota + SC RM of 6ndash12 S and TM of 2 19 plusmn 5 109 plusmn37 115 plusmn46 + + +Paullinia rubiginosa + PW RM of 3ndash4 S and TM of 2 31 plusmn11 147 plusmn51 162 plusmn19 + + +Paullinia rugosa + PW RM of (2) 4ndash13 S and TM of 2 28 plusmn 6 105 plusmn48 79 plusmn21 + + +Paullinia spicata + Comp with 3 PC
PW in the CCRM of 2ndash4 and 5ndash9 S and TM of 2 33 plusmn 9 94 plusmn32 40 plusmn10 + + ndash +
Paullinia sprucei minus NA S and RM of 2ndash4 S and TM of 2 21 plusmn 8 70 plusmn22 39 plusmn 8 ndash ndash +Paullinia trigonia + SC RM of 6ndash9 S and TM of 2 28 plusmn 6 132 plusmn60 116 plusmn39 + + + +Paullinia turbacensis + PW RM of 2ndash4 and 5ndash9 S and TM of 2 30 plusmn 7 99 plusmn32 44 plusmn 6 ndash + + +Paullinia weinmanniifolia + SC RM of 6ndash20 S and RM of 2 26 plusmn 6 98 plusmn41 132 plusmn26 + + + +Paullinia sp ndash NA RM of 2ndash4 and 5ndash9 S and TM of 2 27 plusmn 6 58 plusmn14 35 plusmn15 + + + +
aCambial variant type Comp compound PC peripheral cylinder of the compound cambial variant CC central cylinder of the compound cambial variant PW phloem wedges LX lobed xylem SCsuccessive cambia
bS solitary vessels TM tangential multiples RD radial multiplesThe thin-walled bands of this species consist of more axial parenchyma than others
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
284IAWA
Journal41(3)2020
Table 3Wood anatomy characters in Paullinia spp rays axial parenchyma and cell contents
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia alata + + + 1ndash2 (4) 6ndash10 Mixed + + + ndash + +Paullinia baileyi + 1ndash2 6 Mixed mostly
procumbentand square
+
Paullinia bracteosa ndash + + 1ndash2 5 Mixed mostlyupright
+ + + + ndash +
Paullinia dasystachya + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia echinata + ndash 2 + 1ndash2 3ndash5 Mixed + ndash + + + ndashPaullinia exalata ndash + + 1ndash2 3 Mixed + + ndash ndash +Paullinia fibrigera + ndash 2 + 1ndash2 5 Mixed + + + + ndash +Paullinia fimbriata ndash + 2 + 1ndash2 3ndash5 Mixed mostly
upright andsquare
+ + + + ndash
Paullinia latifolia + ndash 2 + 1 6ndash10 Mixed mostlyprocumbentand upright
+ + + +
Paullinia micrantha + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + ndash + +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
CheryetalndashWoodanatom
yofPaullinia285
Table 3(Continued)
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia obovata + ndash + 1ndash2 5 + + + + + (few) +Paullinia pinnata + ndash Mixed mostly
upright andsquare
+ ndash +
Paullinia pseudota + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + + ndash + +
Paullinia rubiginosa + ndash Mixed + + ndash ndash ndashPaullinia rugosa ndash + + 1ndash2 5 Mixed + ndash + + ndash ndashPaullinia spicata ndash + 2 + 1ndash2 6 Mixed + + + ndash + +Paullinia sprucei + ndash 2 ndash 1ndash2 1ndash2 Mixed ndash ndash + ndash + +Paullinia trigonia + + 2ndash4 + 1ndash4 3ndash7 Mixed mostly
procumbentand upright
+ + ndash + +
Paullinia turbacensis + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia weinmanniifolia + ndash NA + 1ndash2 3ndash7 Mixed + + + ndash + +Paullinia sp ndash + + 1ndash2 3ndash4 Mixed mostly
upright andsquare
+ ndash + ndash ndash +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
286 IAWA Journal 41 (3) 2020
Figure 1 Mature growth stage of the six stem ontogenies in Paullinia spp sensu Chery et al (2020)(A) Regular I in P fibrigera Note the latex in the phloem and cortex (B) Regular II in P latifolia(C) Phloemwedges inP rubiginosa (D) Lobedxylem inP caloptera (E)Compound stem inP pinnata(F) Successive cambia in P pseudota (image modified with permission from Cunha Neto et al 2018)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

Chery et al ndashWood anatomy of Paullinia 279
anatomy in Paullinia suggests a combination of phylogenetic constraint as a mem-ber of Sapindaceae and functional constraint from the liana habit
Keywords Convergent evolution cambial variants Paullinieae Sapindales vines
INTRODUCTION
The stemmorphology andwood anatomy of lianas are critical to understanding the climb-ing habit as herein lies distinctive features that separate this woody growth form fromtrees and shrubs In comparison to erect plantsmdashat the cellular scalemdashlianas have fewerfibers and allocate more cross-sectional area to water conduits wide vessels andor tra-cheids (Carlquist 1991 Angyalossy et al 2012 2015) Additionally non-lignified ray and axialparenchyma are more commonly found in lianas than in self-sustaining plants (Carlquist1991 Angyalossy et al 2012 2015) This combinationof anatomical features referred to as theldquoliana syndrome anatomyrdquo translates into eco-physiological functions that facilitate opti-mization of conductance flexibility and strength (Carlquist 1985 1991 FisherampEwers 1989RoweampSpeck 2004 Angyalossy et al 2012 2015) Consistent across lianas sensu lato includ-ing ferns gymnosperms monocots andmagnoliids the liana syndrome is even diagnosticfor identifying lianas in the fossil record (Krings et al 2006 Burnham 2009 Rozefelds ampPace 2018) At the macro-scale the gross morphology of liana stems is often unusual theseatypical configurations are called ldquocambial variantsrdquo (Carlquist 2001)Vascular cambial vari-ants arise as non-circular stem outlines aberrant tissue conformations andor shifts in therelative abundance of secondary xylem (wood) and phloem (inner bark) produced by thevascular cambiumTaken together both the cellular scale (micromorphology) and the stemmorphology (macromorphology) seem to be altered in lianas as a result of climbing adap-tations
Paullinia L (Sapindaceae) is a genus of ca 220 species of mostly neotropical lianas(Acevedo-Rodriacuteguez et al 2017 Chery et al 2019) Together with five other genera oflianas (Serjania L Cardiospermum Kunth Urvillea Kunth Lophostigma Radlk and Thi-nouiaTrianaampPlanch) Paullinia constitutes themonophyletic Paullinieae tribe (Acevedo-Rodriacuteguez et al 2017 Chery et al 2019) which contains approximately one-third of thespecies diversity of Sapindaceae (Buerki et al 2009) Paullinieae display an impressive as-sortment of different modes of fruit dispersal (Acevedo-Rodriacuteguez et al 2011 2017 Chery etal 2019) and diversity of secondary growth ontogenies (Schenk 1893 Radlkofer 1895 Pfeif-fer 1926 Obaton 1960 Van der Walt et al 1973 Klaassen 1999 Tamaio amp Angyalossy 2009Tamaio amp Somner 2010 Tamaio 2011 Bastos et al 2016 Cunha Neto et al 2018 Chery et al2020)
As in many liana lineages several species of Paullinia have vascular cambial variantsIndividual Paullinia species exhibit one or more of the following types of cambial variants(i) phloem wedges produced by elevated production of secondary phloem relative to sec-ondary xylem by the interfascicular cambium (ii) lobed xylem produced by the elevatedproduction of secondary xylem by the fascicular cambium (iii) compound stems gener-ated by the presence of multiple procambial strands forming multiple vascular cylinders
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
280 IAWA Journal 41 (3) 2020
and (iv) successive cambia generated by de novo cambial strands formed by the pericycle(Radlkofer 1895 Schenk 1893 Pfeiffer 1926 Obaton 1960 Van der Walt et al 1973 Klaassen1999 Tamaio amp Angyalossy 2009 Tamaio amp Somner 2010 Tamaio et al 2011 Tamaio 2011Bastos et al 2016 Cunha Neto et al 2018 Chery et al 2020) As cambial variants are asso-ciated with the repeated evolution of lianas across vascular plants these forms have beenimplicated in overcoming challenges related to the stress of climbing stems such functionsinclude injury repair as a regenerative source of essential vascular tissues (ie successivecambia Carlquist 2007) and stem flexibility (Dobbins amp Fisher 1986 Fisher amp Ewers 1989Carlquist 1991 Rowe et al 2006 Isnard amp Silk 2009)
Our current understanding of the distinctiveness of liana wood anatomy is the resultof the tremendous efforts by numerous authors who have detailed the anatomy of lianasfrom different lineages (Schenk 1893 Pfeiffer 1926 Carlquist 1991 Caballeacute 1993 Pace et al2009 2015 2018a bWagner et al 2012 Pace ampAngyalossy 2013) To contribute to this effortof linking wood anatomy to plant form and function we here present the wood anatomy ofPaullinia and discuss our anatomical results in the context of broad patterns of convergentevolution among lianas
MATERIALS ANDMETHODS
Extant species sampling tissue embedding sectioning and stainingWe studied the wood anatomy of 21 Paullinia species (one is as yet unidentified) that
broadly span the breadth of the Paullinia phylogeny (Chery et al 2019) Material was ob-tained from field collections in Central and South American localities (Table 1) Additionalsamples were obtained by sampling mature stems from herbarium vouchers and rehydrat-ing them by boiling them in 1 ml of glycerin + 100 ml of water for approximately 1 hourField-collected sampleswere immediately fixed in formaldehyde-acetic acid-alcohol (25ml95 ethanol 25 ml glacial acetic acid 5 ml of formalin (37 formaldehyde) + 175 ml wa-ter in a 50 ml falcon tube) then permanently stored in 70 ethanol following Johansen(1940)
Stems were gradually embedded in polyethylene glycol (PEG) 1500 (Rupp 1964) as de-scribed in Barbosa et al (2010) Stems were placed in a 1-liter beaker melted PEG waspoured to cover the specimen and the beaker was filled to 1 liter with water Beakers withstems + PEG + water were placed in a paraffin oven to allow the water to slowly evapo-rate and for the PEG to gradually infiltrate the stems (approxmately 9 days) Stems andPEG were poured into rectangular paper molds to generate PEG blocks Steel knives weresharpened with sandpaper (in a progressive series of grades 600 1200 1500 2000 follow-ing Barbosa et al 2018) Stems in PEG blocks were then sectioned with sharpened steelknives on a sliding microtome each section was coated with a polystyrene foam solution(Barbosa et al 2010) Sections were stained with Safrablau (nine parts 1 Astra Blue (1 g ofAstra Blue + 100 ml of 50 ethanol) to one part 1 Safranin O (1 g of Safranin-O in 100 mlof 50 ethanol) Bukatsch 1972modified by Kraus ampArduin 1997) washed with an ethanolseries (50 60 70 80 95 then 100 EtOH) followed by a xylene series (11 100EtOH100xylene then 100xylene) then finallymountedwith Eukittreg quick-hardening
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
CheryetalndashWoodanatom
yofPaullinia281
Table 1Material obtained from field collections in Central and South American localities
Species Collector Collection number (Herbarium code) Locality
Paullinia alata (Ruiz amp Pav) GDon Brandbyge 33347 (US) Pace 694 (US) San Pablo de los Secoyas Napo Ecuador Reserva Bioloacutegia La SelvaCosta Rica
Paullinia baileyi Standl Chery 37 (UC) Barro Colorado Island PanamaPaullinia bracteosa Radlk Chery 26 (UC) Chery 39 (UC) Barro Colorado Island PanamaPaullinia dasystachya Radlk Acevedo 14295 (US) Madre de Dios PeruPaullinia echinata Huber Schunke 2310 (US) Huaacutenuco PeruPaullinia exalata Radlk Couvreur 207 (US) Ixiamas La Paz BoliviaPaullinia fibrigera Radlk Chery 16 (UC) Chery 21 (UC) Barro Colorado Island PanamaPaullinia fimbriata Radlk Day 9005 (US) Acre BrazilPaullinia latifolia Benth ex Radlk Acevedo 5835 (US) Brasil 528 (US) Tumuc Humac Mts Talouakem Suriname Acre BrazilPaullinia micrantha Cambess Somner 1597 Somner 1562 Tinguaacute Brazil PARNASO BrazilPaullinia obovata (Ruiz amp Pav) Pers Mathias 3585 (US) San Martin PeruPaullinia pinnata L Chery 30 (UC) Chery 41 (UC) Barro Colorado Island PanamaPaullinia pseudota Radlk Bastos 1 Bona 5 Bona 15 Usina Satildeo Joseacute Brazil Jacarepiaacute Brazil Cagarras BrazilPaullinia rubiginosa Cambess Acevedo 14366 (US) Madre de Dios PeruPaullinia rugosa Benth ex Radlk Chery 10 (UC) Barro Colorado Island PanamaPaullinia sp Chery 46 (UC) Barro Colorado Island PanamaPaullinia spicata Benth Oldeman 1248 (US) Riviegravere Comtegrave French GuianaPaullinia sprucei JF Macbr Acevedo 7519 (US) Schunke 15407 (US) Napo Province Yasuni Forest Reserve Ecuador Sacarita del Riacuteo
Utiquinia Cornoel Portillo PeruPaullinia trigonia Vell Somner 1449 Somner 1609 Somner 1635 PARNAI Brazil Tinguaacute Brazil PARNAI BrazilPaullinia turbacensis Kunth Acevedo 15176 (US) Chery 13 (UC) Barro Colorado Island PanamaPaullinia weinmanniifoliaMart Acevedo 3698 Somner 1610 amp 1497 Espirito Santo Reserva Valle do Rio Doce Brazil APAMaricaacute Brazil
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
282 IAWA Journal 41 (3) 2020
mounting medium to prepare permanent slides Stems were sectioned in transverse tan-gential and radial planes
Macerations were performed to identify the different cell types of the secondary xylemSamples were incubated for 48 hours in equal part solutions of hydrogen peroxide + glacialacetic acid (Franklin 1945) then stained with Safranin-O
Slides were analyzed under (i) Zeiss Stemi 508 stereomicroscope (Carl Zeiss AGOberkochen Germany) and imaged with a Canon EOS M3 (Canon Ōta Tokyo Japan)(ii) Zeiss AxioImager light microscope (Carl Zeiss Oberkochen Germany) and imagedwith a QImaging 5MPix MicroPublisher camera (Teledyne QImaging Surrey BC Canada)and (iii) Olympus BX50 (Olympus Hatagaya Japan) and imaged with a Media Cybernet-ics CoolSNAP-Pro digital camera (Media Cybernetics Silver Springs MD USA) Slides areavailable upon request from the corresponding authors
Character codingAnatomical descriptions followed the IAWA Hardwood List (IAWA Committee 1989) as
a starting point adjusting for the Paullinia-specific features that we observed Due to thepresenceof vessel dimorphismwe report ldquonarrowvessel diameterrdquo and ldquowide vessel diame-terrdquo as two separate charactersTo this aim for each specieswemeasuredall vessels (at least125 vesselsspecies) by first thresholding the image then using the ldquoanalyze particlesrdquo func-tion in Image J 152a (Schneider et al 2012)We generated histograms of the log-transformeddistribution of vessel diameters to identify the peaks in the distribution that correspondto the narrow and wide vessel classes (see Distributions in the Appendix) The mean andstandard deviations of these respective classes were calculated (Tables 2 and 3) and boxand whisker plots are presented in Fig 3 For species with compound stems all charactersare reported from the central vascular cylinder (Tamaio et al 2011) Also for species withsuccessive cambia we measured the vessel element diameters in the central cylinder andthe peripheral vascular strands to test if there is a difference between this feature in the reg-ular versus variant xylemwithin a species Statistical analyses were performed in R (R CoreTeam 2018) and all measurements were performed in Image J 152a (Schneider et al 2012)
RESULTS
Cambial variantsSix different stem ontogenies are recognized in Paullinia (Chery et al 2020) regular I
regular II phloemwedges lobedxylem compound stems and successive cambia (Fig 1 andexplored in detail in Chery et al 2020) Regular I is observed in one species (P fibrigera)regular II in three species (P echinata P sprucei and P sp) phloem wedges in 10 speciesmdash including in the lobed xylem of P obovata and in the central cylinder of the compoundstemsof P alata andP spicata lobed xylem inone species compound stems in four speciesand successive cambia is observed in P weinmanniifolia P micrantha P pseudota and Ptrigonia
Growth ringsAll species of Paullinia have diffuse-porous woods and indistinct growth rings (Fig 2A)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
CheryetalndashWoodanatom
yofPaullinia283
Table 2Wood anatomy characters in Paullinia spp vessels
Species Cambialvariantpresent
Cambial varianttypea
Vessel groupingsb Vessel diameter(μm)
Vesselsmm2 Tracheidspresent
Intervessel pitswith slit-likecoalescentapertures
Gum-likesubstance in
vesselspresent
Alternatingthin-walled andthick-walled
fibers
Narrow Wide Narrow Wide
Paullinia alata + Comp with 3 PCPW in the CC
RM of 2ndash4 and 9ndash13 S and TM of 2 30 plusmn12 117 plusmn52 97 plusmn21 + + ndash +
Paullinia baileyi + PW S and RM of 2ndash4 S 21 plusmn 7 62 plusmn12 37 plusmn14 + + +Paullinia bracteosa + PW RM of 6ndash8 and 9ndash15 S and TM of 2 24 plusmn 5 93 plusmn33 91 plusmn25 + + ndash +Paullinia dasystachya + PW RM of 2ndash4 and 5ndash9 S TM and RM of 2 26 plusmn 5 84 plusmn31 37 plusmn 4 ndash + ndash +Paullinia echinata ndash NA RM of 2ndash4 S TM and RM of 2 31 plusmn10 163 plusmn50 63 plusmn19 + ndash + ndashPaullinia exalata + Comp with 3 PC RM of 2ndash4 and 5ndash9 S and TM of 2 17 plusmn 8 89 plusmn38 64 plusmn12 + + +Paullinia fibrigera ndash NA S and RM of 2ndash5 S and TM of 2 14 plusmn 3 106 plusmn52 74 plusmn22 + + ndash +Paullinia fimbriata + PW RM of 6ndash9 S and TM of 2 24 plusmn 4 83 plusmn28 75 plusmn15 + + + +Paullinia latifolia Very shallow PW RM of 3ndash4 S and TM of 2 40 plusmn11 165 plusmn68 129 plusmn26 + ndash ndashPaullinia micrantha + SC RM of 6ndash9 S and TM of 2 34 plusmn11 171 plusmn60 85 plusmn20 + + +Paullinia obovata + LX PW RM of 3ndash4 S TM and RM of 2 13 plusmn 2 44 plusmn21 80 plusmn13 + + Paullinia pinnata + Comp with 3 PC RM of 3ndash4 S and TM of 2 28 plusmn11 101 plusmn36 70 plusmn10 + + +Paullinia pseudota + SC RM of 6ndash12 S and TM of 2 19 plusmn 5 109 plusmn37 115 plusmn46 + + +Paullinia rubiginosa + PW RM of 3ndash4 S and TM of 2 31 plusmn11 147 plusmn51 162 plusmn19 + + +Paullinia rugosa + PW RM of (2) 4ndash13 S and TM of 2 28 plusmn 6 105 plusmn48 79 plusmn21 + + +Paullinia spicata + Comp with 3 PC
PW in the CCRM of 2ndash4 and 5ndash9 S and TM of 2 33 plusmn 9 94 plusmn32 40 plusmn10 + + ndash +
Paullinia sprucei minus NA S and RM of 2ndash4 S and TM of 2 21 plusmn 8 70 plusmn22 39 plusmn 8 ndash ndash +Paullinia trigonia + SC RM of 6ndash9 S and TM of 2 28 plusmn 6 132 plusmn60 116 plusmn39 + + + +Paullinia turbacensis + PW RM of 2ndash4 and 5ndash9 S and TM of 2 30 plusmn 7 99 plusmn32 44 plusmn 6 ndash + + +Paullinia weinmanniifolia + SC RM of 6ndash20 S and RM of 2 26 plusmn 6 98 plusmn41 132 plusmn26 + + + +Paullinia sp ndash NA RM of 2ndash4 and 5ndash9 S and TM of 2 27 plusmn 6 58 plusmn14 35 plusmn15 + + + +
aCambial variant type Comp compound PC peripheral cylinder of the compound cambial variant CC central cylinder of the compound cambial variant PW phloem wedges LX lobed xylem SCsuccessive cambia
bS solitary vessels TM tangential multiples RD radial multiplesThe thin-walled bands of this species consist of more axial parenchyma than others
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
284IAWA
Journal41(3)2020
Table 3Wood anatomy characters in Paullinia spp rays axial parenchyma and cell contents
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia alata + + + 1ndash2 (4) 6ndash10 Mixed + + + ndash + +Paullinia baileyi + 1ndash2 6 Mixed mostly
procumbentand square
+
Paullinia bracteosa ndash + + 1ndash2 5 Mixed mostlyupright
+ + + + ndash +
Paullinia dasystachya + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia echinata + ndash 2 + 1ndash2 3ndash5 Mixed + ndash + + + ndashPaullinia exalata ndash + + 1ndash2 3 Mixed + + ndash ndash +Paullinia fibrigera + ndash 2 + 1ndash2 5 Mixed + + + + ndash +Paullinia fimbriata ndash + 2 + 1ndash2 3ndash5 Mixed mostly
upright andsquare
+ + + + ndash
Paullinia latifolia + ndash 2 + 1 6ndash10 Mixed mostlyprocumbentand upright
+ + + +
Paullinia micrantha + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + ndash + +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
CheryetalndashWoodanatom
yofPaullinia285
Table 3(Continued)
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia obovata + ndash + 1ndash2 5 + + + + + (few) +Paullinia pinnata + ndash Mixed mostly
upright andsquare
+ ndash +
Paullinia pseudota + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + + ndash + +
Paullinia rubiginosa + ndash Mixed + + ndash ndash ndashPaullinia rugosa ndash + + 1ndash2 5 Mixed + ndash + + ndash ndashPaullinia spicata ndash + 2 + 1ndash2 6 Mixed + + + ndash + +Paullinia sprucei + ndash 2 ndash 1ndash2 1ndash2 Mixed ndash ndash + ndash + +Paullinia trigonia + + 2ndash4 + 1ndash4 3ndash7 Mixed mostly
procumbentand upright
+ + ndash + +
Paullinia turbacensis + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia weinmanniifolia + ndash NA + 1ndash2 3ndash7 Mixed + + + ndash + +Paullinia sp ndash + + 1ndash2 3ndash4 Mixed mostly
upright andsquare
+ ndash + ndash ndash +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
286 IAWA Journal 41 (3) 2020
Figure 1 Mature growth stage of the six stem ontogenies in Paullinia spp sensu Chery et al (2020)(A) Regular I in P fibrigera Note the latex in the phloem and cortex (B) Regular II in P latifolia(C) Phloemwedges inP rubiginosa (D) Lobedxylem inP caloptera (E)Compound stem inP pinnata(F) Successive cambia in P pseudota (image modified with permission from Cunha Neto et al 2018)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

280 IAWA Journal 41 (3) 2020
and (iv) successive cambia generated by de novo cambial strands formed by the pericycle(Radlkofer 1895 Schenk 1893 Pfeiffer 1926 Obaton 1960 Van der Walt et al 1973 Klaassen1999 Tamaio amp Angyalossy 2009 Tamaio amp Somner 2010 Tamaio et al 2011 Tamaio 2011Bastos et al 2016 Cunha Neto et al 2018 Chery et al 2020) As cambial variants are asso-ciated with the repeated evolution of lianas across vascular plants these forms have beenimplicated in overcoming challenges related to the stress of climbing stems such functionsinclude injury repair as a regenerative source of essential vascular tissues (ie successivecambia Carlquist 2007) and stem flexibility (Dobbins amp Fisher 1986 Fisher amp Ewers 1989Carlquist 1991 Rowe et al 2006 Isnard amp Silk 2009)
Our current understanding of the distinctiveness of liana wood anatomy is the resultof the tremendous efforts by numerous authors who have detailed the anatomy of lianasfrom different lineages (Schenk 1893 Pfeiffer 1926 Carlquist 1991 Caballeacute 1993 Pace et al2009 2015 2018a bWagner et al 2012 Pace ampAngyalossy 2013) To contribute to this effortof linking wood anatomy to plant form and function we here present the wood anatomy ofPaullinia and discuss our anatomical results in the context of broad patterns of convergentevolution among lianas
MATERIALS ANDMETHODS
Extant species sampling tissue embedding sectioning and stainingWe studied the wood anatomy of 21 Paullinia species (one is as yet unidentified) that
broadly span the breadth of the Paullinia phylogeny (Chery et al 2019) Material was ob-tained from field collections in Central and South American localities (Table 1) Additionalsamples were obtained by sampling mature stems from herbarium vouchers and rehydrat-ing them by boiling them in 1 ml of glycerin + 100 ml of water for approximately 1 hourField-collected sampleswere immediately fixed in formaldehyde-acetic acid-alcohol (25ml95 ethanol 25 ml glacial acetic acid 5 ml of formalin (37 formaldehyde) + 175 ml wa-ter in a 50 ml falcon tube) then permanently stored in 70 ethanol following Johansen(1940)
Stems were gradually embedded in polyethylene glycol (PEG) 1500 (Rupp 1964) as de-scribed in Barbosa et al (2010) Stems were placed in a 1-liter beaker melted PEG waspoured to cover the specimen and the beaker was filled to 1 liter with water Beakers withstems + PEG + water were placed in a paraffin oven to allow the water to slowly evapo-rate and for the PEG to gradually infiltrate the stems (approxmately 9 days) Stems andPEG were poured into rectangular paper molds to generate PEG blocks Steel knives weresharpened with sandpaper (in a progressive series of grades 600 1200 1500 2000 follow-ing Barbosa et al 2018) Stems in PEG blocks were then sectioned with sharpened steelknives on a sliding microtome each section was coated with a polystyrene foam solution(Barbosa et al 2010) Sections were stained with Safrablau (nine parts 1 Astra Blue (1 g ofAstra Blue + 100 ml of 50 ethanol) to one part 1 Safranin O (1 g of Safranin-O in 100 mlof 50 ethanol) Bukatsch 1972modified by Kraus ampArduin 1997) washed with an ethanolseries (50 60 70 80 95 then 100 EtOH) followed by a xylene series (11 100EtOH100xylene then 100xylene) then finallymountedwith Eukittreg quick-hardening
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
CheryetalndashWoodanatom
yofPaullinia281
Table 1Material obtained from field collections in Central and South American localities
Species Collector Collection number (Herbarium code) Locality
Paullinia alata (Ruiz amp Pav) GDon Brandbyge 33347 (US) Pace 694 (US) San Pablo de los Secoyas Napo Ecuador Reserva Bioloacutegia La SelvaCosta Rica
Paullinia baileyi Standl Chery 37 (UC) Barro Colorado Island PanamaPaullinia bracteosa Radlk Chery 26 (UC) Chery 39 (UC) Barro Colorado Island PanamaPaullinia dasystachya Radlk Acevedo 14295 (US) Madre de Dios PeruPaullinia echinata Huber Schunke 2310 (US) Huaacutenuco PeruPaullinia exalata Radlk Couvreur 207 (US) Ixiamas La Paz BoliviaPaullinia fibrigera Radlk Chery 16 (UC) Chery 21 (UC) Barro Colorado Island PanamaPaullinia fimbriata Radlk Day 9005 (US) Acre BrazilPaullinia latifolia Benth ex Radlk Acevedo 5835 (US) Brasil 528 (US) Tumuc Humac Mts Talouakem Suriname Acre BrazilPaullinia micrantha Cambess Somner 1597 Somner 1562 Tinguaacute Brazil PARNASO BrazilPaullinia obovata (Ruiz amp Pav) Pers Mathias 3585 (US) San Martin PeruPaullinia pinnata L Chery 30 (UC) Chery 41 (UC) Barro Colorado Island PanamaPaullinia pseudota Radlk Bastos 1 Bona 5 Bona 15 Usina Satildeo Joseacute Brazil Jacarepiaacute Brazil Cagarras BrazilPaullinia rubiginosa Cambess Acevedo 14366 (US) Madre de Dios PeruPaullinia rugosa Benth ex Radlk Chery 10 (UC) Barro Colorado Island PanamaPaullinia sp Chery 46 (UC) Barro Colorado Island PanamaPaullinia spicata Benth Oldeman 1248 (US) Riviegravere Comtegrave French GuianaPaullinia sprucei JF Macbr Acevedo 7519 (US) Schunke 15407 (US) Napo Province Yasuni Forest Reserve Ecuador Sacarita del Riacuteo
Utiquinia Cornoel Portillo PeruPaullinia trigonia Vell Somner 1449 Somner 1609 Somner 1635 PARNAI Brazil Tinguaacute Brazil PARNAI BrazilPaullinia turbacensis Kunth Acevedo 15176 (US) Chery 13 (UC) Barro Colorado Island PanamaPaullinia weinmanniifoliaMart Acevedo 3698 Somner 1610 amp 1497 Espirito Santo Reserva Valle do Rio Doce Brazil APAMaricaacute Brazil
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
282 IAWA Journal 41 (3) 2020
mounting medium to prepare permanent slides Stems were sectioned in transverse tan-gential and radial planes
Macerations were performed to identify the different cell types of the secondary xylemSamples were incubated for 48 hours in equal part solutions of hydrogen peroxide + glacialacetic acid (Franklin 1945) then stained with Safranin-O
Slides were analyzed under (i) Zeiss Stemi 508 stereomicroscope (Carl Zeiss AGOberkochen Germany) and imaged with a Canon EOS M3 (Canon Ōta Tokyo Japan)(ii) Zeiss AxioImager light microscope (Carl Zeiss Oberkochen Germany) and imagedwith a QImaging 5MPix MicroPublisher camera (Teledyne QImaging Surrey BC Canada)and (iii) Olympus BX50 (Olympus Hatagaya Japan) and imaged with a Media Cybernet-ics CoolSNAP-Pro digital camera (Media Cybernetics Silver Springs MD USA) Slides areavailable upon request from the corresponding authors
Character codingAnatomical descriptions followed the IAWA Hardwood List (IAWA Committee 1989) as
a starting point adjusting for the Paullinia-specific features that we observed Due to thepresenceof vessel dimorphismwe report ldquonarrowvessel diameterrdquo and ldquowide vessel diame-terrdquo as two separate charactersTo this aim for each specieswemeasuredall vessels (at least125 vesselsspecies) by first thresholding the image then using the ldquoanalyze particlesrdquo func-tion in Image J 152a (Schneider et al 2012)We generated histograms of the log-transformeddistribution of vessel diameters to identify the peaks in the distribution that correspondto the narrow and wide vessel classes (see Distributions in the Appendix) The mean andstandard deviations of these respective classes were calculated (Tables 2 and 3) and boxand whisker plots are presented in Fig 3 For species with compound stems all charactersare reported from the central vascular cylinder (Tamaio et al 2011) Also for species withsuccessive cambia we measured the vessel element diameters in the central cylinder andthe peripheral vascular strands to test if there is a difference between this feature in the reg-ular versus variant xylemwithin a species Statistical analyses were performed in R (R CoreTeam 2018) and all measurements were performed in Image J 152a (Schneider et al 2012)
RESULTS
Cambial variantsSix different stem ontogenies are recognized in Paullinia (Chery et al 2020) regular I
regular II phloemwedges lobedxylem compound stems and successive cambia (Fig 1 andexplored in detail in Chery et al 2020) Regular I is observed in one species (P fibrigera)regular II in three species (P echinata P sprucei and P sp) phloem wedges in 10 speciesmdash including in the lobed xylem of P obovata and in the central cylinder of the compoundstemsof P alata andP spicata lobed xylem inone species compound stems in four speciesand successive cambia is observed in P weinmanniifolia P micrantha P pseudota and Ptrigonia
Growth ringsAll species of Paullinia have diffuse-porous woods and indistinct growth rings (Fig 2A)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
CheryetalndashWoodanatom
yofPaullinia283
Table 2Wood anatomy characters in Paullinia spp vessels
Species Cambialvariantpresent
Cambial varianttypea
Vessel groupingsb Vessel diameter(μm)
Vesselsmm2 Tracheidspresent
Intervessel pitswith slit-likecoalescentapertures
Gum-likesubstance in
vesselspresent
Alternatingthin-walled andthick-walled
fibers
Narrow Wide Narrow Wide
Paullinia alata + Comp with 3 PCPW in the CC
RM of 2ndash4 and 9ndash13 S and TM of 2 30 plusmn12 117 plusmn52 97 plusmn21 + + ndash +
Paullinia baileyi + PW S and RM of 2ndash4 S 21 plusmn 7 62 plusmn12 37 plusmn14 + + +Paullinia bracteosa + PW RM of 6ndash8 and 9ndash15 S and TM of 2 24 plusmn 5 93 plusmn33 91 plusmn25 + + ndash +Paullinia dasystachya + PW RM of 2ndash4 and 5ndash9 S TM and RM of 2 26 plusmn 5 84 plusmn31 37 plusmn 4 ndash + ndash +Paullinia echinata ndash NA RM of 2ndash4 S TM and RM of 2 31 plusmn10 163 plusmn50 63 plusmn19 + ndash + ndashPaullinia exalata + Comp with 3 PC RM of 2ndash4 and 5ndash9 S and TM of 2 17 plusmn 8 89 plusmn38 64 plusmn12 + + +Paullinia fibrigera ndash NA S and RM of 2ndash5 S and TM of 2 14 plusmn 3 106 plusmn52 74 plusmn22 + + ndash +Paullinia fimbriata + PW RM of 6ndash9 S and TM of 2 24 plusmn 4 83 plusmn28 75 plusmn15 + + + +Paullinia latifolia Very shallow PW RM of 3ndash4 S and TM of 2 40 plusmn11 165 plusmn68 129 plusmn26 + ndash ndashPaullinia micrantha + SC RM of 6ndash9 S and TM of 2 34 plusmn11 171 plusmn60 85 plusmn20 + + +Paullinia obovata + LX PW RM of 3ndash4 S TM and RM of 2 13 plusmn 2 44 plusmn21 80 plusmn13 + + Paullinia pinnata + Comp with 3 PC RM of 3ndash4 S and TM of 2 28 plusmn11 101 plusmn36 70 plusmn10 + + +Paullinia pseudota + SC RM of 6ndash12 S and TM of 2 19 plusmn 5 109 plusmn37 115 plusmn46 + + +Paullinia rubiginosa + PW RM of 3ndash4 S and TM of 2 31 plusmn11 147 plusmn51 162 plusmn19 + + +Paullinia rugosa + PW RM of (2) 4ndash13 S and TM of 2 28 plusmn 6 105 plusmn48 79 plusmn21 + + +Paullinia spicata + Comp with 3 PC
PW in the CCRM of 2ndash4 and 5ndash9 S and TM of 2 33 plusmn 9 94 plusmn32 40 plusmn10 + + ndash +
Paullinia sprucei minus NA S and RM of 2ndash4 S and TM of 2 21 plusmn 8 70 plusmn22 39 plusmn 8 ndash ndash +Paullinia trigonia + SC RM of 6ndash9 S and TM of 2 28 plusmn 6 132 plusmn60 116 plusmn39 + + + +Paullinia turbacensis + PW RM of 2ndash4 and 5ndash9 S and TM of 2 30 plusmn 7 99 plusmn32 44 plusmn 6 ndash + + +Paullinia weinmanniifolia + SC RM of 6ndash20 S and RM of 2 26 plusmn 6 98 plusmn41 132 plusmn26 + + + +Paullinia sp ndash NA RM of 2ndash4 and 5ndash9 S and TM of 2 27 plusmn 6 58 plusmn14 35 plusmn15 + + + +
aCambial variant type Comp compound PC peripheral cylinder of the compound cambial variant CC central cylinder of the compound cambial variant PW phloem wedges LX lobed xylem SCsuccessive cambia
bS solitary vessels TM tangential multiples RD radial multiplesThe thin-walled bands of this species consist of more axial parenchyma than others
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
284IAWA
Journal41(3)2020
Table 3Wood anatomy characters in Paullinia spp rays axial parenchyma and cell contents
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia alata + + + 1ndash2 (4) 6ndash10 Mixed + + + ndash + +Paullinia baileyi + 1ndash2 6 Mixed mostly
procumbentand square
+
Paullinia bracteosa ndash + + 1ndash2 5 Mixed mostlyupright
+ + + + ndash +
Paullinia dasystachya + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia echinata + ndash 2 + 1ndash2 3ndash5 Mixed + ndash + + + ndashPaullinia exalata ndash + + 1ndash2 3 Mixed + + ndash ndash +Paullinia fibrigera + ndash 2 + 1ndash2 5 Mixed + + + + ndash +Paullinia fimbriata ndash + 2 + 1ndash2 3ndash5 Mixed mostly
upright andsquare
+ + + + ndash
Paullinia latifolia + ndash 2 + 1 6ndash10 Mixed mostlyprocumbentand upright
+ + + +
Paullinia micrantha + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + ndash + +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
CheryetalndashWoodanatom
yofPaullinia285
Table 3(Continued)
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia obovata + ndash + 1ndash2 5 + + + + + (few) +Paullinia pinnata + ndash Mixed mostly
upright andsquare
+ ndash +
Paullinia pseudota + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + + ndash + +
Paullinia rubiginosa + ndash Mixed + + ndash ndash ndashPaullinia rugosa ndash + + 1ndash2 5 Mixed + ndash + + ndash ndashPaullinia spicata ndash + 2 + 1ndash2 6 Mixed + + + ndash + +Paullinia sprucei + ndash 2 ndash 1ndash2 1ndash2 Mixed ndash ndash + ndash + +Paullinia trigonia + + 2ndash4 + 1ndash4 3ndash7 Mixed mostly
procumbentand upright
+ + ndash + +
Paullinia turbacensis + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia weinmanniifolia + ndash NA + 1ndash2 3ndash7 Mixed + + + ndash + +Paullinia sp ndash + + 1ndash2 3ndash4 Mixed mostly
upright andsquare
+ ndash + ndash ndash +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
286 IAWA Journal 41 (3) 2020
Figure 1 Mature growth stage of the six stem ontogenies in Paullinia spp sensu Chery et al (2020)(A) Regular I in P fibrigera Note the latex in the phloem and cortex (B) Regular II in P latifolia(C) Phloemwedges inP rubiginosa (D) Lobedxylem inP caloptera (E)Compound stem inP pinnata(F) Successive cambia in P pseudota (image modified with permission from Cunha Neto et al 2018)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
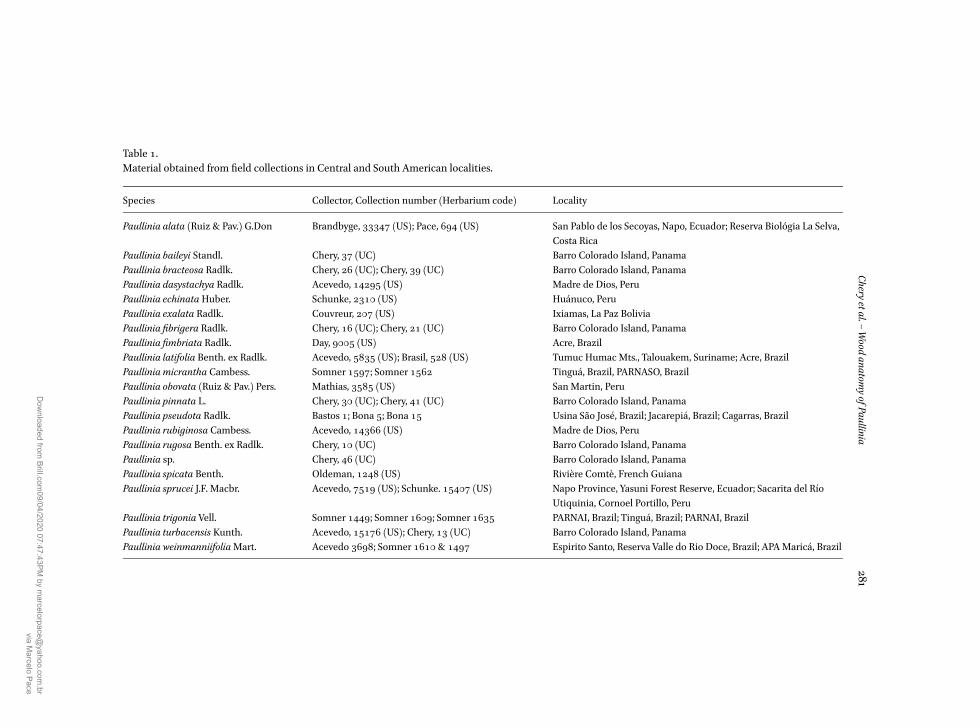
CheryetalndashWoodanatom
yofPaullinia281
Table 1Material obtained from field collections in Central and South American localities
Species Collector Collection number (Herbarium code) Locality
Paullinia alata (Ruiz amp Pav) GDon Brandbyge 33347 (US) Pace 694 (US) San Pablo de los Secoyas Napo Ecuador Reserva Bioloacutegia La SelvaCosta Rica
Paullinia baileyi Standl Chery 37 (UC) Barro Colorado Island PanamaPaullinia bracteosa Radlk Chery 26 (UC) Chery 39 (UC) Barro Colorado Island PanamaPaullinia dasystachya Radlk Acevedo 14295 (US) Madre de Dios PeruPaullinia echinata Huber Schunke 2310 (US) Huaacutenuco PeruPaullinia exalata Radlk Couvreur 207 (US) Ixiamas La Paz BoliviaPaullinia fibrigera Radlk Chery 16 (UC) Chery 21 (UC) Barro Colorado Island PanamaPaullinia fimbriata Radlk Day 9005 (US) Acre BrazilPaullinia latifolia Benth ex Radlk Acevedo 5835 (US) Brasil 528 (US) Tumuc Humac Mts Talouakem Suriname Acre BrazilPaullinia micrantha Cambess Somner 1597 Somner 1562 Tinguaacute Brazil PARNASO BrazilPaullinia obovata (Ruiz amp Pav) Pers Mathias 3585 (US) San Martin PeruPaullinia pinnata L Chery 30 (UC) Chery 41 (UC) Barro Colorado Island PanamaPaullinia pseudota Radlk Bastos 1 Bona 5 Bona 15 Usina Satildeo Joseacute Brazil Jacarepiaacute Brazil Cagarras BrazilPaullinia rubiginosa Cambess Acevedo 14366 (US) Madre de Dios PeruPaullinia rugosa Benth ex Radlk Chery 10 (UC) Barro Colorado Island PanamaPaullinia sp Chery 46 (UC) Barro Colorado Island PanamaPaullinia spicata Benth Oldeman 1248 (US) Riviegravere Comtegrave French GuianaPaullinia sprucei JF Macbr Acevedo 7519 (US) Schunke 15407 (US) Napo Province Yasuni Forest Reserve Ecuador Sacarita del Riacuteo
Utiquinia Cornoel Portillo PeruPaullinia trigonia Vell Somner 1449 Somner 1609 Somner 1635 PARNAI Brazil Tinguaacute Brazil PARNAI BrazilPaullinia turbacensis Kunth Acevedo 15176 (US) Chery 13 (UC) Barro Colorado Island PanamaPaullinia weinmanniifoliaMart Acevedo 3698 Somner 1610 amp 1497 Espirito Santo Reserva Valle do Rio Doce Brazil APAMaricaacute Brazil
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
282 IAWA Journal 41 (3) 2020
mounting medium to prepare permanent slides Stems were sectioned in transverse tan-gential and radial planes
Macerations were performed to identify the different cell types of the secondary xylemSamples were incubated for 48 hours in equal part solutions of hydrogen peroxide + glacialacetic acid (Franklin 1945) then stained with Safranin-O
Slides were analyzed under (i) Zeiss Stemi 508 stereomicroscope (Carl Zeiss AGOberkochen Germany) and imaged with a Canon EOS M3 (Canon Ōta Tokyo Japan)(ii) Zeiss AxioImager light microscope (Carl Zeiss Oberkochen Germany) and imagedwith a QImaging 5MPix MicroPublisher camera (Teledyne QImaging Surrey BC Canada)and (iii) Olympus BX50 (Olympus Hatagaya Japan) and imaged with a Media Cybernet-ics CoolSNAP-Pro digital camera (Media Cybernetics Silver Springs MD USA) Slides areavailable upon request from the corresponding authors
Character codingAnatomical descriptions followed the IAWA Hardwood List (IAWA Committee 1989) as
a starting point adjusting for the Paullinia-specific features that we observed Due to thepresenceof vessel dimorphismwe report ldquonarrowvessel diameterrdquo and ldquowide vessel diame-terrdquo as two separate charactersTo this aim for each specieswemeasuredall vessels (at least125 vesselsspecies) by first thresholding the image then using the ldquoanalyze particlesrdquo func-tion in Image J 152a (Schneider et al 2012)We generated histograms of the log-transformeddistribution of vessel diameters to identify the peaks in the distribution that correspondto the narrow and wide vessel classes (see Distributions in the Appendix) The mean andstandard deviations of these respective classes were calculated (Tables 2 and 3) and boxand whisker plots are presented in Fig 3 For species with compound stems all charactersare reported from the central vascular cylinder (Tamaio et al 2011) Also for species withsuccessive cambia we measured the vessel element diameters in the central cylinder andthe peripheral vascular strands to test if there is a difference between this feature in the reg-ular versus variant xylemwithin a species Statistical analyses were performed in R (R CoreTeam 2018) and all measurements were performed in Image J 152a (Schneider et al 2012)
RESULTS
Cambial variantsSix different stem ontogenies are recognized in Paullinia (Chery et al 2020) regular I
regular II phloemwedges lobedxylem compound stems and successive cambia (Fig 1 andexplored in detail in Chery et al 2020) Regular I is observed in one species (P fibrigera)regular II in three species (P echinata P sprucei and P sp) phloem wedges in 10 speciesmdash including in the lobed xylem of P obovata and in the central cylinder of the compoundstemsof P alata andP spicata lobed xylem inone species compound stems in four speciesand successive cambia is observed in P weinmanniifolia P micrantha P pseudota and Ptrigonia
Growth ringsAll species of Paullinia have diffuse-porous woods and indistinct growth rings (Fig 2A)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
CheryetalndashWoodanatom
yofPaullinia283
Table 2Wood anatomy characters in Paullinia spp vessels
Species Cambialvariantpresent
Cambial varianttypea
Vessel groupingsb Vessel diameter(μm)
Vesselsmm2 Tracheidspresent
Intervessel pitswith slit-likecoalescentapertures
Gum-likesubstance in
vesselspresent
Alternatingthin-walled andthick-walled
fibers
Narrow Wide Narrow Wide
Paullinia alata + Comp with 3 PCPW in the CC
RM of 2ndash4 and 9ndash13 S and TM of 2 30 plusmn12 117 plusmn52 97 plusmn21 + + ndash +
Paullinia baileyi + PW S and RM of 2ndash4 S 21 plusmn 7 62 plusmn12 37 plusmn14 + + +Paullinia bracteosa + PW RM of 6ndash8 and 9ndash15 S and TM of 2 24 plusmn 5 93 plusmn33 91 plusmn25 + + ndash +Paullinia dasystachya + PW RM of 2ndash4 and 5ndash9 S TM and RM of 2 26 plusmn 5 84 plusmn31 37 plusmn 4 ndash + ndash +Paullinia echinata ndash NA RM of 2ndash4 S TM and RM of 2 31 plusmn10 163 plusmn50 63 plusmn19 + ndash + ndashPaullinia exalata + Comp with 3 PC RM of 2ndash4 and 5ndash9 S and TM of 2 17 plusmn 8 89 plusmn38 64 plusmn12 + + +Paullinia fibrigera ndash NA S and RM of 2ndash5 S and TM of 2 14 plusmn 3 106 plusmn52 74 plusmn22 + + ndash +Paullinia fimbriata + PW RM of 6ndash9 S and TM of 2 24 plusmn 4 83 plusmn28 75 plusmn15 + + + +Paullinia latifolia Very shallow PW RM of 3ndash4 S and TM of 2 40 plusmn11 165 plusmn68 129 plusmn26 + ndash ndashPaullinia micrantha + SC RM of 6ndash9 S and TM of 2 34 plusmn11 171 plusmn60 85 plusmn20 + + +Paullinia obovata + LX PW RM of 3ndash4 S TM and RM of 2 13 plusmn 2 44 plusmn21 80 plusmn13 + + Paullinia pinnata + Comp with 3 PC RM of 3ndash4 S and TM of 2 28 plusmn11 101 plusmn36 70 plusmn10 + + +Paullinia pseudota + SC RM of 6ndash12 S and TM of 2 19 plusmn 5 109 plusmn37 115 plusmn46 + + +Paullinia rubiginosa + PW RM of 3ndash4 S and TM of 2 31 plusmn11 147 plusmn51 162 plusmn19 + + +Paullinia rugosa + PW RM of (2) 4ndash13 S and TM of 2 28 plusmn 6 105 plusmn48 79 plusmn21 + + +Paullinia spicata + Comp with 3 PC
PW in the CCRM of 2ndash4 and 5ndash9 S and TM of 2 33 plusmn 9 94 plusmn32 40 plusmn10 + + ndash +
Paullinia sprucei minus NA S and RM of 2ndash4 S and TM of 2 21 plusmn 8 70 plusmn22 39 plusmn 8 ndash ndash +Paullinia trigonia + SC RM of 6ndash9 S and TM of 2 28 plusmn 6 132 plusmn60 116 plusmn39 + + + +Paullinia turbacensis + PW RM of 2ndash4 and 5ndash9 S and TM of 2 30 plusmn 7 99 plusmn32 44 plusmn 6 ndash + + +Paullinia weinmanniifolia + SC RM of 6ndash20 S and RM of 2 26 plusmn 6 98 plusmn41 132 plusmn26 + + + +Paullinia sp ndash NA RM of 2ndash4 and 5ndash9 S and TM of 2 27 plusmn 6 58 plusmn14 35 plusmn15 + + + +
aCambial variant type Comp compound PC peripheral cylinder of the compound cambial variant CC central cylinder of the compound cambial variant PW phloem wedges LX lobed xylem SCsuccessive cambia
bS solitary vessels TM tangential multiples RD radial multiplesThe thin-walled bands of this species consist of more axial parenchyma than others
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
284IAWA
Journal41(3)2020
Table 3Wood anatomy characters in Paullinia spp rays axial parenchyma and cell contents
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia alata + + + 1ndash2 (4) 6ndash10 Mixed + + + ndash + +Paullinia baileyi + 1ndash2 6 Mixed mostly
procumbentand square
+
Paullinia bracteosa ndash + + 1ndash2 5 Mixed mostlyupright
+ + + + ndash +
Paullinia dasystachya + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia echinata + ndash 2 + 1ndash2 3ndash5 Mixed + ndash + + + ndashPaullinia exalata ndash + + 1ndash2 3 Mixed + + ndash ndash +Paullinia fibrigera + ndash 2 + 1ndash2 5 Mixed + + + + ndash +Paullinia fimbriata ndash + 2 + 1ndash2 3ndash5 Mixed mostly
upright andsquare
+ + + + ndash
Paullinia latifolia + ndash 2 + 1 6ndash10 Mixed mostlyprocumbentand upright
+ + + +
Paullinia micrantha + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + ndash + +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
CheryetalndashWoodanatom
yofPaullinia285
Table 3(Continued)
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia obovata + ndash + 1ndash2 5 + + + + + (few) +Paullinia pinnata + ndash Mixed mostly
upright andsquare
+ ndash +
Paullinia pseudota + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + + ndash + +
Paullinia rubiginosa + ndash Mixed + + ndash ndash ndashPaullinia rugosa ndash + + 1ndash2 5 Mixed + ndash + + ndash ndashPaullinia spicata ndash + 2 + 1ndash2 6 Mixed + + + ndash + +Paullinia sprucei + ndash 2 ndash 1ndash2 1ndash2 Mixed ndash ndash + ndash + +Paullinia trigonia + + 2ndash4 + 1ndash4 3ndash7 Mixed mostly
procumbentand upright
+ + ndash + +
Paullinia turbacensis + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia weinmanniifolia + ndash NA + 1ndash2 3ndash7 Mixed + + + ndash + +Paullinia sp ndash + + 1ndash2 3ndash4 Mixed mostly
upright andsquare
+ ndash + ndash ndash +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
286 IAWA Journal 41 (3) 2020
Figure 1 Mature growth stage of the six stem ontogenies in Paullinia spp sensu Chery et al (2020)(A) Regular I in P fibrigera Note the latex in the phloem and cortex (B) Regular II in P latifolia(C) Phloemwedges inP rubiginosa (D) Lobedxylem inP caloptera (E)Compound stem inP pinnata(F) Successive cambia in P pseudota (image modified with permission from Cunha Neto et al 2018)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

282 IAWA Journal 41 (3) 2020
mounting medium to prepare permanent slides Stems were sectioned in transverse tan-gential and radial planes
Macerations were performed to identify the different cell types of the secondary xylemSamples were incubated for 48 hours in equal part solutions of hydrogen peroxide + glacialacetic acid (Franklin 1945) then stained with Safranin-O
Slides were analyzed under (i) Zeiss Stemi 508 stereomicroscope (Carl Zeiss AGOberkochen Germany) and imaged with a Canon EOS M3 (Canon Ōta Tokyo Japan)(ii) Zeiss AxioImager light microscope (Carl Zeiss Oberkochen Germany) and imagedwith a QImaging 5MPix MicroPublisher camera (Teledyne QImaging Surrey BC Canada)and (iii) Olympus BX50 (Olympus Hatagaya Japan) and imaged with a Media Cybernet-ics CoolSNAP-Pro digital camera (Media Cybernetics Silver Springs MD USA) Slides areavailable upon request from the corresponding authors
Character codingAnatomical descriptions followed the IAWA Hardwood List (IAWA Committee 1989) as
a starting point adjusting for the Paullinia-specific features that we observed Due to thepresenceof vessel dimorphismwe report ldquonarrowvessel diameterrdquo and ldquowide vessel diame-terrdquo as two separate charactersTo this aim for each specieswemeasuredall vessels (at least125 vesselsspecies) by first thresholding the image then using the ldquoanalyze particlesrdquo func-tion in Image J 152a (Schneider et al 2012)We generated histograms of the log-transformeddistribution of vessel diameters to identify the peaks in the distribution that correspondto the narrow and wide vessel classes (see Distributions in the Appendix) The mean andstandard deviations of these respective classes were calculated (Tables 2 and 3) and boxand whisker plots are presented in Fig 3 For species with compound stems all charactersare reported from the central vascular cylinder (Tamaio et al 2011) Also for species withsuccessive cambia we measured the vessel element diameters in the central cylinder andthe peripheral vascular strands to test if there is a difference between this feature in the reg-ular versus variant xylemwithin a species Statistical analyses were performed in R (R CoreTeam 2018) and all measurements were performed in Image J 152a (Schneider et al 2012)
RESULTS
Cambial variantsSix different stem ontogenies are recognized in Paullinia (Chery et al 2020) regular I
regular II phloemwedges lobedxylem compound stems and successive cambia (Fig 1 andexplored in detail in Chery et al 2020) Regular I is observed in one species (P fibrigera)regular II in three species (P echinata P sprucei and P sp) phloem wedges in 10 speciesmdash including in the lobed xylem of P obovata and in the central cylinder of the compoundstemsof P alata andP spicata lobed xylem inone species compound stems in four speciesand successive cambia is observed in P weinmanniifolia P micrantha P pseudota and Ptrigonia
Growth ringsAll species of Paullinia have diffuse-porous woods and indistinct growth rings (Fig 2A)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
CheryetalndashWoodanatom
yofPaullinia283
Table 2Wood anatomy characters in Paullinia spp vessels
Species Cambialvariantpresent
Cambial varianttypea
Vessel groupingsb Vessel diameter(μm)
Vesselsmm2 Tracheidspresent
Intervessel pitswith slit-likecoalescentapertures
Gum-likesubstance in
vesselspresent
Alternatingthin-walled andthick-walled
fibers
Narrow Wide Narrow Wide
Paullinia alata + Comp with 3 PCPW in the CC
RM of 2ndash4 and 9ndash13 S and TM of 2 30 plusmn12 117 plusmn52 97 plusmn21 + + ndash +
Paullinia baileyi + PW S and RM of 2ndash4 S 21 plusmn 7 62 plusmn12 37 plusmn14 + + +Paullinia bracteosa + PW RM of 6ndash8 and 9ndash15 S and TM of 2 24 plusmn 5 93 plusmn33 91 plusmn25 + + ndash +Paullinia dasystachya + PW RM of 2ndash4 and 5ndash9 S TM and RM of 2 26 plusmn 5 84 plusmn31 37 plusmn 4 ndash + ndash +Paullinia echinata ndash NA RM of 2ndash4 S TM and RM of 2 31 plusmn10 163 plusmn50 63 plusmn19 + ndash + ndashPaullinia exalata + Comp with 3 PC RM of 2ndash4 and 5ndash9 S and TM of 2 17 plusmn 8 89 plusmn38 64 plusmn12 + + +Paullinia fibrigera ndash NA S and RM of 2ndash5 S and TM of 2 14 plusmn 3 106 plusmn52 74 plusmn22 + + ndash +Paullinia fimbriata + PW RM of 6ndash9 S and TM of 2 24 plusmn 4 83 plusmn28 75 plusmn15 + + + +Paullinia latifolia Very shallow PW RM of 3ndash4 S and TM of 2 40 plusmn11 165 plusmn68 129 plusmn26 + ndash ndashPaullinia micrantha + SC RM of 6ndash9 S and TM of 2 34 plusmn11 171 plusmn60 85 plusmn20 + + +Paullinia obovata + LX PW RM of 3ndash4 S TM and RM of 2 13 plusmn 2 44 plusmn21 80 plusmn13 + + Paullinia pinnata + Comp with 3 PC RM of 3ndash4 S and TM of 2 28 plusmn11 101 plusmn36 70 plusmn10 + + +Paullinia pseudota + SC RM of 6ndash12 S and TM of 2 19 plusmn 5 109 plusmn37 115 plusmn46 + + +Paullinia rubiginosa + PW RM of 3ndash4 S and TM of 2 31 plusmn11 147 plusmn51 162 plusmn19 + + +Paullinia rugosa + PW RM of (2) 4ndash13 S and TM of 2 28 plusmn 6 105 plusmn48 79 plusmn21 + + +Paullinia spicata + Comp with 3 PC
PW in the CCRM of 2ndash4 and 5ndash9 S and TM of 2 33 plusmn 9 94 plusmn32 40 plusmn10 + + ndash +
Paullinia sprucei minus NA S and RM of 2ndash4 S and TM of 2 21 plusmn 8 70 plusmn22 39 plusmn 8 ndash ndash +Paullinia trigonia + SC RM of 6ndash9 S and TM of 2 28 plusmn 6 132 plusmn60 116 plusmn39 + + + +Paullinia turbacensis + PW RM of 2ndash4 and 5ndash9 S and TM of 2 30 plusmn 7 99 plusmn32 44 plusmn 6 ndash + + +Paullinia weinmanniifolia + SC RM of 6ndash20 S and RM of 2 26 plusmn 6 98 plusmn41 132 plusmn26 + + + +Paullinia sp ndash NA RM of 2ndash4 and 5ndash9 S and TM of 2 27 plusmn 6 58 plusmn14 35 plusmn15 + + + +
aCambial variant type Comp compound PC peripheral cylinder of the compound cambial variant CC central cylinder of the compound cambial variant PW phloem wedges LX lobed xylem SCsuccessive cambia
bS solitary vessels TM tangential multiples RD radial multiplesThe thin-walled bands of this species consist of more axial parenchyma than others
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
284IAWA
Journal41(3)2020
Table 3Wood anatomy characters in Paullinia spp rays axial parenchyma and cell contents
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia alata + + + 1ndash2 (4) 6ndash10 Mixed + + + ndash + +Paullinia baileyi + 1ndash2 6 Mixed mostly
procumbentand square
+
Paullinia bracteosa ndash + + 1ndash2 5 Mixed mostlyupright
+ + + + ndash +
Paullinia dasystachya + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia echinata + ndash 2 + 1ndash2 3ndash5 Mixed + ndash + + + ndashPaullinia exalata ndash + + 1ndash2 3 Mixed + + ndash ndash +Paullinia fibrigera + ndash 2 + 1ndash2 5 Mixed + + + + ndash +Paullinia fimbriata ndash + 2 + 1ndash2 3ndash5 Mixed mostly
upright andsquare
+ + + + ndash
Paullinia latifolia + ndash 2 + 1 6ndash10 Mixed mostlyprocumbentand upright
+ + + +
Paullinia micrantha + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + ndash + +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
CheryetalndashWoodanatom
yofPaullinia285
Table 3(Continued)
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia obovata + ndash + 1ndash2 5 + + + + + (few) +Paullinia pinnata + ndash Mixed mostly
upright andsquare
+ ndash +
Paullinia pseudota + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + + ndash + +
Paullinia rubiginosa + ndash Mixed + + ndash ndash ndashPaullinia rugosa ndash + + 1ndash2 5 Mixed + ndash + + ndash ndashPaullinia spicata ndash + 2 + 1ndash2 6 Mixed + + + ndash + +Paullinia sprucei + ndash 2 ndash 1ndash2 1ndash2 Mixed ndash ndash + ndash + +Paullinia trigonia + + 2ndash4 + 1ndash4 3ndash7 Mixed mostly
procumbentand upright
+ + ndash + +
Paullinia turbacensis + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia weinmanniifolia + ndash NA + 1ndash2 3ndash7 Mixed + + + ndash + +Paullinia sp ndash + + 1ndash2 3ndash4 Mixed mostly
upright andsquare
+ ndash + ndash ndash +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
286 IAWA Journal 41 (3) 2020
Figure 1 Mature growth stage of the six stem ontogenies in Paullinia spp sensu Chery et al (2020)(A) Regular I in P fibrigera Note the latex in the phloem and cortex (B) Regular II in P latifolia(C) Phloemwedges inP rubiginosa (D) Lobedxylem inP caloptera (E)Compound stem inP pinnata(F) Successive cambia in P pseudota (image modified with permission from Cunha Neto et al 2018)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

CheryetalndashWoodanatom
yofPaullinia283
Table 2Wood anatomy characters in Paullinia spp vessels
Species Cambialvariantpresent
Cambial varianttypea
Vessel groupingsb Vessel diameter(μm)
Vesselsmm2 Tracheidspresent
Intervessel pitswith slit-likecoalescentapertures
Gum-likesubstance in
vesselspresent
Alternatingthin-walled andthick-walled
fibers
Narrow Wide Narrow Wide
Paullinia alata + Comp with 3 PCPW in the CC
RM of 2ndash4 and 9ndash13 S and TM of 2 30 plusmn12 117 plusmn52 97 plusmn21 + + ndash +
Paullinia baileyi + PW S and RM of 2ndash4 S 21 plusmn 7 62 plusmn12 37 plusmn14 + + +Paullinia bracteosa + PW RM of 6ndash8 and 9ndash15 S and TM of 2 24 plusmn 5 93 plusmn33 91 plusmn25 + + ndash +Paullinia dasystachya + PW RM of 2ndash4 and 5ndash9 S TM and RM of 2 26 plusmn 5 84 plusmn31 37 plusmn 4 ndash + ndash +Paullinia echinata ndash NA RM of 2ndash4 S TM and RM of 2 31 plusmn10 163 plusmn50 63 plusmn19 + ndash + ndashPaullinia exalata + Comp with 3 PC RM of 2ndash4 and 5ndash9 S and TM of 2 17 plusmn 8 89 plusmn38 64 plusmn12 + + +Paullinia fibrigera ndash NA S and RM of 2ndash5 S and TM of 2 14 plusmn 3 106 plusmn52 74 plusmn22 + + ndash +Paullinia fimbriata + PW RM of 6ndash9 S and TM of 2 24 plusmn 4 83 plusmn28 75 plusmn15 + + + +Paullinia latifolia Very shallow PW RM of 3ndash4 S and TM of 2 40 plusmn11 165 plusmn68 129 plusmn26 + ndash ndashPaullinia micrantha + SC RM of 6ndash9 S and TM of 2 34 plusmn11 171 plusmn60 85 plusmn20 + + +Paullinia obovata + LX PW RM of 3ndash4 S TM and RM of 2 13 plusmn 2 44 plusmn21 80 plusmn13 + + Paullinia pinnata + Comp with 3 PC RM of 3ndash4 S and TM of 2 28 plusmn11 101 plusmn36 70 plusmn10 + + +Paullinia pseudota + SC RM of 6ndash12 S and TM of 2 19 plusmn 5 109 plusmn37 115 plusmn46 + + +Paullinia rubiginosa + PW RM of 3ndash4 S and TM of 2 31 plusmn11 147 plusmn51 162 plusmn19 + + +Paullinia rugosa + PW RM of (2) 4ndash13 S and TM of 2 28 plusmn 6 105 plusmn48 79 plusmn21 + + +Paullinia spicata + Comp with 3 PC
PW in the CCRM of 2ndash4 and 5ndash9 S and TM of 2 33 plusmn 9 94 plusmn32 40 plusmn10 + + ndash +
Paullinia sprucei minus NA S and RM of 2ndash4 S and TM of 2 21 plusmn 8 70 plusmn22 39 plusmn 8 ndash ndash +Paullinia trigonia + SC RM of 6ndash9 S and TM of 2 28 plusmn 6 132 plusmn60 116 plusmn39 + + + +Paullinia turbacensis + PW RM of 2ndash4 and 5ndash9 S and TM of 2 30 plusmn 7 99 plusmn32 44 plusmn 6 ndash + + +Paullinia weinmanniifolia + SC RM of 6ndash20 S and RM of 2 26 plusmn 6 98 plusmn41 132 plusmn26 + + + +Paullinia sp ndash NA RM of 2ndash4 and 5ndash9 S and TM of 2 27 plusmn 6 58 plusmn14 35 plusmn15 + + + +
aCambial variant type Comp compound PC peripheral cylinder of the compound cambial variant CC central cylinder of the compound cambial variant PW phloem wedges LX lobed xylem SCsuccessive cambia
bS solitary vessels TM tangential multiples RD radial multiplesThe thin-walled bands of this species consist of more axial parenchyma than others
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
284IAWA
Journal41(3)2020
Table 3Wood anatomy characters in Paullinia spp rays axial parenchyma and cell contents
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia alata + + + 1ndash2 (4) 6ndash10 Mixed + + + ndash + +Paullinia baileyi + 1ndash2 6 Mixed mostly
procumbentand square
+
Paullinia bracteosa ndash + + 1ndash2 5 Mixed mostlyupright
+ + + + ndash +
Paullinia dasystachya + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia echinata + ndash 2 + 1ndash2 3ndash5 Mixed + ndash + + + ndashPaullinia exalata ndash + + 1ndash2 3 Mixed + + ndash ndash +Paullinia fibrigera + ndash 2 + 1ndash2 5 Mixed + + + + ndash +Paullinia fimbriata ndash + 2 + 1ndash2 3ndash5 Mixed mostly
upright andsquare
+ + + + ndash
Paullinia latifolia + ndash 2 + 1 6ndash10 Mixed mostlyprocumbentand upright
+ + + +
Paullinia micrantha + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + ndash + +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
CheryetalndashWoodanatom
yofPaullinia285
Table 3(Continued)
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia obovata + ndash + 1ndash2 5 + + + + + (few) +Paullinia pinnata + ndash Mixed mostly
upright andsquare
+ ndash +
Paullinia pseudota + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + + ndash + +
Paullinia rubiginosa + ndash Mixed + + ndash ndash ndashPaullinia rugosa ndash + + 1ndash2 5 Mixed + ndash + + ndash ndashPaullinia spicata ndash + 2 + 1ndash2 6 Mixed + + + ndash + +Paullinia sprucei + ndash 2 ndash 1ndash2 1ndash2 Mixed ndash ndash + ndash + +Paullinia trigonia + + 2ndash4 + 1ndash4 3ndash7 Mixed mostly
procumbentand upright
+ + ndash + +
Paullinia turbacensis + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia weinmanniifolia + ndash NA + 1ndash2 3ndash7 Mixed + + + ndash + +Paullinia sp ndash + + 1ndash2 3ndash4 Mixed mostly
upright andsquare
+ ndash + ndash ndash +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
286 IAWA Journal 41 (3) 2020
Figure 1 Mature growth stage of the six stem ontogenies in Paullinia spp sensu Chery et al (2020)(A) Regular I in P fibrigera Note the latex in the phloem and cortex (B) Regular II in P latifolia(C) Phloemwedges inP rubiginosa (D) Lobedxylem inP caloptera (E)Compound stem inP pinnata(F) Successive cambia in P pseudota (image modified with permission from Cunha Neto et al 2018)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
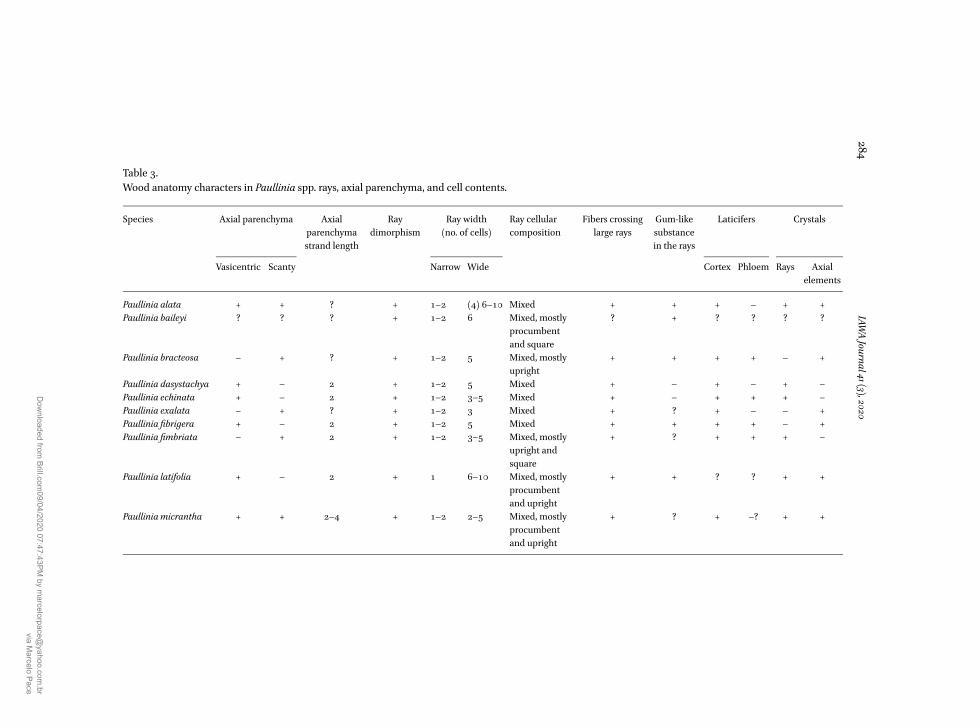
284IAWA
Journal41(3)2020
Table 3Wood anatomy characters in Paullinia spp rays axial parenchyma and cell contents
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia alata + + + 1ndash2 (4) 6ndash10 Mixed + + + ndash + +Paullinia baileyi + 1ndash2 6 Mixed mostly
procumbentand square
+
Paullinia bracteosa ndash + + 1ndash2 5 Mixed mostlyupright
+ + + + ndash +
Paullinia dasystachya + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia echinata + ndash 2 + 1ndash2 3ndash5 Mixed + ndash + + + ndashPaullinia exalata ndash + + 1ndash2 3 Mixed + + ndash ndash +Paullinia fibrigera + ndash 2 + 1ndash2 5 Mixed + + + + ndash +Paullinia fimbriata ndash + 2 + 1ndash2 3ndash5 Mixed mostly
upright andsquare
+ + + + ndash
Paullinia latifolia + ndash 2 + 1 6ndash10 Mixed mostlyprocumbentand upright
+ + + +
Paullinia micrantha + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + ndash + +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
CheryetalndashWoodanatom
yofPaullinia285
Table 3(Continued)
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia obovata + ndash + 1ndash2 5 + + + + + (few) +Paullinia pinnata + ndash Mixed mostly
upright andsquare
+ ndash +
Paullinia pseudota + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + + ndash + +
Paullinia rubiginosa + ndash Mixed + + ndash ndash ndashPaullinia rugosa ndash + + 1ndash2 5 Mixed + ndash + + ndash ndashPaullinia spicata ndash + 2 + 1ndash2 6 Mixed + + + ndash + +Paullinia sprucei + ndash 2 ndash 1ndash2 1ndash2 Mixed ndash ndash + ndash + +Paullinia trigonia + + 2ndash4 + 1ndash4 3ndash7 Mixed mostly
procumbentand upright
+ + ndash + +
Paullinia turbacensis + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia weinmanniifolia + ndash NA + 1ndash2 3ndash7 Mixed + + + ndash + +Paullinia sp ndash + + 1ndash2 3ndash4 Mixed mostly
upright andsquare
+ ndash + ndash ndash +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
286 IAWA Journal 41 (3) 2020
Figure 1 Mature growth stage of the six stem ontogenies in Paullinia spp sensu Chery et al (2020)(A) Regular I in P fibrigera Note the latex in the phloem and cortex (B) Regular II in P latifolia(C) Phloemwedges inP rubiginosa (D) Lobedxylem inP caloptera (E)Compound stem inP pinnata(F) Successive cambia in P pseudota (image modified with permission from Cunha Neto et al 2018)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

CheryetalndashWoodanatom
yofPaullinia285
Table 3(Continued)
Species Axial parenchyma Axialparenchymastrand length
Raydimorphism
Ray width(no of cells)
Ray cellularcomposition
Fibers crossinglarge rays
Gum-likesubstancein the rays
Laticifers Crystals
Vasicentric Scanty Narrow Wide Cortex Phloem Rays Axialelements
Paullinia obovata + ndash + 1ndash2 5 + + + + + (few) +Paullinia pinnata + ndash Mixed mostly
upright andsquare
+ ndash +
Paullinia pseudota + + 2ndash4 + 1ndash2 2ndash5 Mixed mostlyprocumbentand upright
+ + + ndash + +
Paullinia rubiginosa + ndash Mixed + + ndash ndash ndashPaullinia rugosa ndash + + 1ndash2 5 Mixed + ndash + + ndash ndashPaullinia spicata ndash + 2 + 1ndash2 6 Mixed + + + ndash + +Paullinia sprucei + ndash 2 ndash 1ndash2 1ndash2 Mixed ndash ndash + ndash + +Paullinia trigonia + + 2ndash4 + 1ndash4 3ndash7 Mixed mostly
procumbentand upright
+ + ndash + +
Paullinia turbacensis + ndash 2 + 1ndash2 5 Mixed + ndash + ndash + ndashPaullinia weinmanniifolia + ndash NA + 1ndash2 3ndash7 Mixed + + + ndash + +Paullinia sp ndash + + 1ndash2 3ndash4 Mixed mostly
upright andsquare
+ ndash + ndash ndash +
Dow
nloaded from Brillcom
09042020 074743PM by m
arcelorpaceyahoocom
brvia M
arcelo Pace
286 IAWA Journal 41 (3) 2020
Figure 1 Mature growth stage of the six stem ontogenies in Paullinia spp sensu Chery et al (2020)(A) Regular I in P fibrigera Note the latex in the phloem and cortex (B) Regular II in P latifolia(C) Phloemwedges inP rubiginosa (D) Lobedxylem inP caloptera (E)Compound stem inP pinnata(F) Successive cambia in P pseudota (image modified with permission from Cunha Neto et al 2018)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

286 IAWA Journal 41 (3) 2020
Figure 1 Mature growth stage of the six stem ontogenies in Paullinia spp sensu Chery et al (2020)(A) Regular I in P fibrigera Note the latex in the phloem and cortex (B) Regular II in P latifolia(C) Phloemwedges inP rubiginosa (D) Lobedxylem inP caloptera (E)Compound stem inP pinnata(F) Successive cambia in P pseudota (image modified with permission from Cunha Neto et al 2018)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
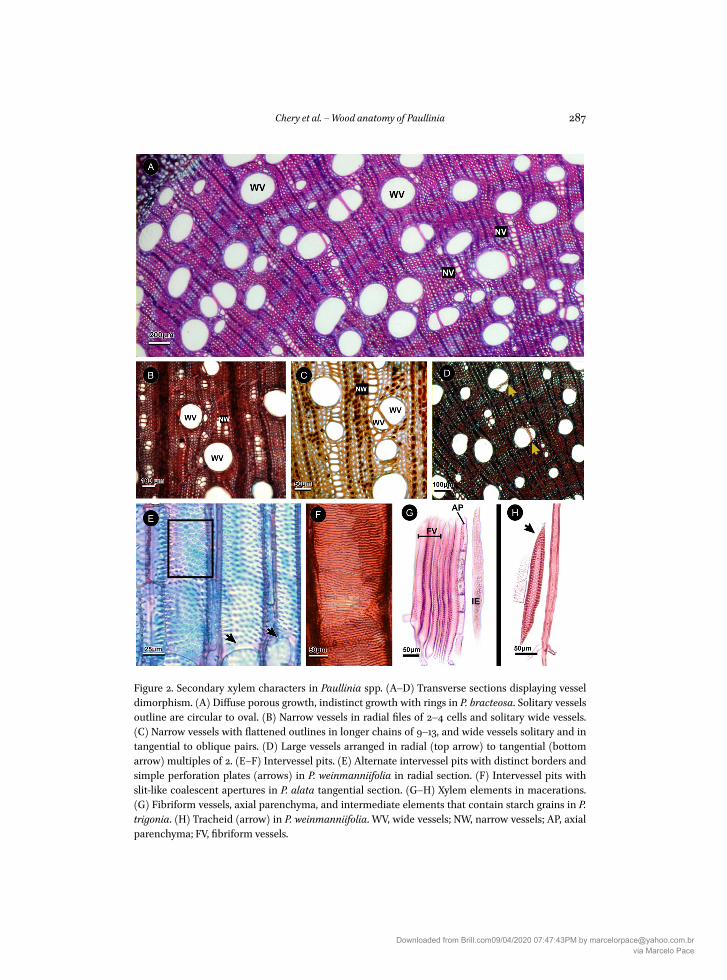
Chery et al ndashWood anatomy of Paullinia 287
Figure 2 Secondary xylem characters in Paullinia spp (AndashD) Transverse sections displaying vesseldimorphism (A) Diffuse porous growth indistinct growth with rings in P bracteosa Solitary vesselsoutline are circular to oval (B) Narrow vessels in radial files of 2ndash4 cells and solitary wide vessels(C) Narrow vessels with flattened outlines in longer chains of 9ndash13 and wide vessels solitary and intangential to oblique pairs (D) Large vessels arranged in radial (top arrow) to tangential (bottomarrow) multiples of 2 (EndashF) Intervessel pits (E) Alternate intervessel pits with distinct borders andsimple perforation plates (arrows) in P weinmanniifolia in radial section (F) Intervessel pits withslit-like coalescent apertures in P alata tangential section (GndashH) Xylem elements in macerations(G) Fibriform vessels axial parenchyma and intermediate elements that contain starch grains in Ptrigonia (H) Tracheid (arrow) in P weinmanniifolia WV wide vessels NW narrow vessels AP axialparenchyma FV fibriform vessels
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

288 IAWA Journal 41 (3) 2020
Vessel charactersAll species of Paullinia present vessels of two size classes (Fig 2AndashD) ie vessel dimor-
phism The large vessels aremostly solitary (Fig 2AndashD) and some are in tangential (Fig 2AC D) or radial pairs of two (Fig 2D) or both The narrow vessels are typically in short ra-dial chains of 2ndash4 (Fig 2B) and long radial chains of 5ndash15 cells (Fig 2A C) and up to 20 inP weinmanniifolia Paullinia latifolia and P echinata (Fig 2B) notably have shorter chainsof 2ndash4 narrow vessels while some species for example P alata P weinmanniifolia and Pbracteosa (Fig 2A) have longer chains of nine or more vessels Solitary vessels are circu-lar to oval in outline (Fig 2A-D) Narrow vessels that are arranged in radial multiples haveflattened outlines (Fig 2B) Vessel dimorphism iswidespread and vessels were divided intotwo classes (Fig 3) The narrow vessel diameters (Fig 3A) range from 13 plusmn 3 μm in P obovatato 40 plusmn 11 μm in P latifolia and large vessel diameters (Fig 3B) range from 443 plusmn 20 μm in
Figure 3 Box and whisker plots of the vessels diameter (μm) in Paullinia spp (A) Narrow vessel ele-ment diameters (B)Wide vessel diameters bull = outliers
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

Chery et al ndashWood anatomy of Paullinia 289
P obovata to 171 plusmn 60 μm in P micrantha Fibriform vessels (sensu Carlquist 1985) were alsoobserved (Fig 2G) All vessels have simple perforation plates (Fig 2E) Intervessel pits areround in outline and alternate with distinct borders (Fig 2E) and often with slit-like coa-lescent apertures (Fig 2F) Slit-like coalescent apertures were not observed in thematerialsof P echinata and P sprucei Tracheids were observed in some species (Fig 2H) Gum-likedeposits are sometimes present in the vessels Neither vestured pits nor helical thickeningsor tyloses were observed Only two of the four species with successive cambia have largervessel elements in the regular xylem of the central cylinder relative to the variant xylemof the vascular arcs (Wilcox test the regular xylem vessels are significantly larger in P mi-crantha (p = 00001) and P trigona (p = 00001)) but are not different in P pseudota and Pweinmanniifolia
Fibers and parenchymaMost species present alternating bands of thin- and thick-walled regions (Fig 4AndashE)
representing parenchyma-like fiber bands sensu IAWA Committee (1989) with the thin-walled bands consisting of septate fibers non-septate fibers axial parenchyma strandsfusiform axial parenchyma and fusiform elements that are morphologically intermedi-ate between axial parenchyma and fibers (here referred to as ldquointermediate elementsrdquoFig 4CndashH) The intermediate elements can be starch storing or crystalliferous (Fig 4DF G) The thick-walled bands consist of septate and non-septate ordinary fibers (Fig 4C)All fibers have simple tominutely bordered pits (Fig 4F) Septate fibers often have an abun-dance of starch and additionally some thick-walled non-septate fibers also contain starchOnly one species P fibrigera has bands of thick-walled non-septate and septate fibers al-ternating with bands of axial parenchyma (Fig 4C) Parenchyma is apotracheal diffuse toscanty or vasicentric paratracheal Axial parenchyma strands are mostly composed of twocells (Fig 4H) however the strands can contain up to five cells
Ray charactersRay dimorphism is commonwith short and narrow rays 1ndash2 cells wide and tall andwide
rays 3ndash10 cells wide (Fig 5A) Ray composition is heterocellular (Fig 5A) comprising up-right square and procumbent cells mixed throughout the ray body Vessel-ray pitting hasreduced borders compared to intervessel pitting (Fig 5E)We frequently observed fusiformelements (which differentiate from ray initials) crossing over wide rays (Fig 5C D ie ldquoher-ringbonerdquo sensuKlaasen 1999) Rays are especiallywide at the sites of differential secondarygrowth in cambial variants for example at the juncture of a phloem wedge and regularphloem (Fig 1C) Occasionally there are axially fused multiseriate rays
Secretory cells and contents in the secondary xylem and phloemLaticifers are common and can be found either in the early-formed secondary phloem
the cortex or both (Fig 6) This feature is especially clear in freshly cut material in thefield (Fig 1A) Prismatic crystals in chambered axial andor ray parenchyma are common(Fig 5D) Silica perforated ray cells oil or mucilage cells and intercellular canals were
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

290 IAWA Journal 41 (3) 2020
Figure 4 (AndashE) Axial elements in the secondary xylem of Paullinia spp (A) P trigonia alternatingbands of thin-(yellowbracket) and thick-walled cells (black bracket) (B)P fibrigera alternating bandsof thin (yellow bracket) and thick-walled cells (black bracket) in transverse section (C) P fibrigera al-ternating bands of thin-(yellow bracket) and thick-walled cells (black bracket) in radial section Notethat the thin-walled regions are mostly composed of axial parenchyma in this species (D) P trigoniaalternating bands in radial section Note the crystalliferous elements in chambered axial parenchymacells (E) P weinmanniifolia alternating bands of ordinary fibers (far right) and thin-walled cells inradial section Note the presence of an axial parenchyma strand a septate fiber and two interme-diate elements in the thin-walled region (FndashH) Axial elements in macerations (F) P trigonia axialparenchyma with prismatic crystals non-septate fiber with starch non-septate fiber without starchnon-septate fiber with starch crystalliferous intermediate elements (from left to right) (G) P trigo-nia two fusiform parenchyma cells and ordinary fiber (from left to right) (H) P weinmanniifolia axialparenchyma and intermediate element CE crystalliferous elements ThickWR thick-walled regionThinWR thin-walled region V vessel AP axial parenchyma SF septate fiber IE intermediate ele-ments
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

Chery et al ndashWood anatomy of Paullinia 291
Figure 5 Ray characters in Paullinia spp (A) P fibrigera ray dimorphism Note the phenolic con-tents in the ray and axial parenchyma (B) P dasystachya heterocellular ray composition with squareupright and procumbent cells mixed throughout the rays (C) P micrantha developing a fusiformelement that crosses the multiseriate ray (arrow) (D) P trigonia detail of ray cell becoming an axi-ally fusiform element that crosses the ray (left arrow) and prismatic crystal in ray cell (right arrow)(E) P weinmanniifolia vesselndashray pits with reduced borders in the rectangle WR wide rays NR nar-row rays
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

292 IAWA Journal 41 (3) 2020
Figure 6 Secretory features in the xylem phloem andcortexof Paullinia spp (A)P bracteosa laticifersin the early formed secondary phloem in transverse section (arrows) (B) P echinata laticifers in theearly formed secondary phloem (lower arrow) and in the cortex in transverse section (upper arrow)(C) Cortical laticifers (arrows) in P weinmannifolia in transverse section (D) Cortical laticifer (arrow)in P weinmannifolia in tangential view Pe pericyclic fibers
not observed Contents are commonly found in axial and radial parenchyma of all species(Fig 5A)
DISCUSSION
Paulliniawood anatomy is conserved yet functionally convergent with other lianalineages
Paullinia species are distinct from the other Paullinieae genera by the following com-bination of characters septate fibers and crystals in both the rays and axial parenchyma(summarized in Table 615 in Klaasen 1999 Pellissari et al 2018) Additionally previous re-searchers have observed a ldquoherringbonerdquo arrangement of ray elements (Klaasen 1999) andldquoboomerangrdquo fibers in macerations (Klaasen 1999 Pellissari et al 2018) however in thisstudy we only observed the formerWithin the genus however we here find that Paulliniawood anatomy is largely conserved with few variable characters
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

Chery et al ndashWood anatomy of Paullinia 293
The wood anatomy of Paullinia shares many functional traits with distantly relatedlianas which are representative of the liana vascular syndrome For example vessel di-morphism has been reported in multiple liana lineages (Carlquist 1985 Klaassen 1999Angyalossy et al 2012 2015 Bastos et al 2016 Pellissari et al 2018) The narrow vesselshave been suggested to serve as a back-up in the event that the larger (and potentiallymore vulnerable) vessels embolize (Carlquist 1985) Recent results by Bouda et al (2019)utilizing both magnetic resonance imaging and X-ray μCT tomography however suggestsamore complex relationship between bulk flow and vessel width Bouda et al (2019) foundthat 15 of the bulk flow predicted to be achieved by wide vessels based on the idealizedHagen-Poiseuille model are transversally redirected to the narrow vessels in grapevine (Vi-tis) These authors do not cite embolism as the reason for this redirection instead theyfound the heterogeneity of the vessel network gives rise to transverse pressure gradientsthat redirect flow fromwide to narrow vessels These new findings highlight the importantrole of narrow vessel elements not only as an emergency route after large vessels embolizebut as a fundamental part of ordinary bulk flowThe importance of narrow vessels has beenpreviously emphasized by Carlquist (1985) who suggested that the total conductance areaof lianas is likely even higher than estimated if researchers accounted for both wide andnarrow vessels as well as tracheids
Our comparison of the vessel diameters between the variant and regular xylem ofspecies with successive cambia revealed that in only two of the four species the vessel el-ements are larger in the regular xylem of the central cylinder The decreasing size of axialelements in the variant xylem could be explained by their difference in origin While thevariant xylem comes from a cambium originated exclusively from the pericyclic parenchy-matic cells the tracheary elements and fibers that originate from the regular cambium areusually longer since they arise from elongated fusiform cells (Zumaya-Mendoza ampTerrazas2016) A variety of studies have analysed comparatively the regular and variant xylem inspecies with successive cambia (Horak 1981 Rajput amp Rao 2003 Tamaio amp Brandes 2010Rajput et al 2013 Zumaya-Mendoza amp Terrazas 2016) However similar to the results pre-sented here divergent results have been found in these studies concerning the increasingor decreasing size in tracheary elements
Another feature cited for the ldquolianescent vascular syndromerdquo is the presence of non-lignified parenchyma interspersed within the lignified tissues of the secondary xylem (iefibers and vessels) of some species which is suggestive of a balancing act tomaintain stemsthat can twist without breaking We did not observe this in Paullinia Instead we foundalternating bands of thin and thick-walled elements Metcalfe and Chalk (1950) and Pel-lissari et al (2018) also observed fiber dimorphism of this type across Paullinia Pellissariet al (2018) reported P ingifolia and we here report P fibrigera as the only species to havebands formed by axial parenchyma Our initial interpretation of this feature was that thealternation of parenchyma-like fiber bands and ordinary fibers could represent Paulliniarsquosstrategy to achieve the simultaneous functions of strength and flexibility that are commonacross lianas For example the mixing of soft and hard tissues is found in the prolifera-tion of disruptive parenchyma from the pith cortex and the secondary xylem and phloemin Callaeum (Malpighiaceae Cabanillas et al 2017) the production of secondary phloem
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

294 IAWA Journal 41 (3) 2020
strands within the secondary xylem (interxylary phloem Carlquist 2013 Angyalossy et al2016) and the conspicuously large rays of Aristolochia (Wagner et al 2012) However thefact that numerous self-supporting plants also display alternating bands of thin-and thick-walled elements such as the erect trees and shrubs of Lythraceae (Baas amp Zweypfenning1979) and Melastomataceae (Ter Welle amp Koek-Normaan 1978) refute our hypothesis thatthis feature is related to the climbing habit in Paullinia Lianas of other plant families mayalso lack axial parenchyma in their stems such as a number of Bignoniaceae lianas (Paceamp Angyalossy 2013 mdash except for Dolichandra and the Fridericia and allied clade) and Ru-biaceae lianas (Leal et al 2020) These cited examples demonstrate that the co-occurrenceof all aspects of the ldquolianescent vascular syndromerdquo sensu Angyalossy et al (2015) is notuniversal across all lianas
The abundance of septate fibers in species with a paucity of axial parenchyma has beennoted by numerous authors (summarized by Carlquist 2001 andWheeler et al 2007) Sep-tate fibers are both superficially similar and functionally analogous to axial parenchymaserving the functions of photosynthate and water storage Living fibers in Acer (also inSapindaceae) have been shown to store starch (Heimsch 1942) which upon starch hydrol-ysis produces sugars that are transported into vessels to drive sap flow (Sauter et al 1973)Therefore the starch-storing fibers in the thin-walled bands (and sometimes observed inthe thick-walled fibers) of Paullinia likely function in storage of water and non-structuralcarbohydrates and in themaintenance of turgor pressure and elasticity (Morris et al 2016)corroborating the idea that these cells assume a storage function as do the parenchyma inother taxa where it is more abundant (Patil et al 2011 Pace amp Angyalossy 2013 Angyalossyet al 2015) In light of Carlquistrsquos (2014) hypothesis that starch-storing living fibers may beimplicated in reversing embolisms this could be a critical strategy to maintain water con-ductance for these lianas given their large and vulnerable vessels (yet to be experimentallytested) Given the scarcity of axial parenchyma in Sapindaceae (Metcalfe amp Chalk 1950Klaassen 1999) the co-option of fibers for these functions in Paulliniamay reflect phyloge-netic constraint However this hypothesis is complicated by the fact that the sister genusSerjaniaMill has abundant axial parenchyma and no septate fibers (Klaassen 1999) indi-cating lability rather than constraint
As also observed in different families the splitting of multiseriate rays in Paullinia re-sults from the conversion of ray initials into fusiform initials (eg fiber-tracheids vesselelements or fusiform parenchyma cells Chattaway 1937 Tamaio 2006 Rajput amp Baijnath2016) This feature was not reported for most authors studying Paullinieae species (Arauacutejoamp Costa 2007 Tamaio 2011 Tamaio et al 2011 Pelissari et al 2018) but it was addressed byKlaassen (1999) who stated that ray cells may gradually differentiate into horizontal andoblique fusiform initials arranged in a herringbone pattern The opposite fusiform initialstransforming into ray initials is also observed In some cases multiseriate rays in Paulliniawere fused via their long uniseriate ray margins this is likely the result of an increase inray height by the transformation of fusiform initials into a series of ray initials (Chattaway1937)
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

Chery et al ndashWood anatomy of Paullinia 295
At present the functional interpretations of the liana vascular syndrome are largelyinferred directly from their anatomy Empirical studies that invoke experiments that de-termine the relative contributions of parenchyma septate and non-septate starch storingfibers and the interactions with vessel dimorphism are needed
Secretory structuresThe anatomy and histochemistry of laticifers and idioblasts in stems of Paullinieae
lianas have been studied by Cunha Neto et al (2017) who observed that in Paullinia lati-cifers occur in the cortex while the phloem only contains secretory idioblasts In this studythe observation of laticifers in the phloem of some Paullinia species extends the distri-bution of this secretory structure presenting a similar distribution of laticifers as Serjaniawhich has laticifers in the pith cortex early- and late-formed phloem including in the vari-ant phloem (CunhaNeto et al 2017) The secretory idioblasts foundwithin the ray and axialparenchyma are equivalent to tannin-containing cells (phenolic compounds) that occurthroughout the stems (Metcalfe amp Chalk 1950 Cunha Neto et al 2017)
Relationship betweenmacromorphology and wood anatomyBy examining thewood anatomy in the different types of cambial variants we identified
a relationship between stemmacromorphology and wood anatomy In the region betweentwo lobes of the lobed xylem cambial variant of P obovata we observed a phloem wedgeand large rays at the juncture of a phloem wedge and lobe a feature referred to as limitingrays by Schenck (1893) ldquobegrenzenden Markstrahlenrdquo in the original authorrsquos terminologyLimiting rays are widespread in stemswith phloemwedges (Dobbins 1971 Pace et al 2009)especially in thosewith discontinuous cambia ie where portions of cambia remain insideof the phloem wedges flanked by the wide limiting rays (Angyalossy et al 2015) In thecompound cambial variant we also consistently observe larger rays in the central cylinderin the regions just below each of the peripheral cylinders The expansion of rays at sitesof differential growth suggests that these wide rays function to accommodate growth andavoid stem fracture This mechanism suggests that the development of vascular cambialvariants not only changes the macromorphology of the stem but is also associated withanatomical changes such as the widening of rays
CONCLUSIONS
Paullinia wood anatomy is largely conserved with most species displaying vessel dimor-phism ray dimorphism and fiber dimorphism Many of the wood features in Paullinia areconsistent with what has been reported in other liana lineages including the presence ofcambial variants vessel dimorphism and heterocellular ray composition Other charactersare the opposite of what is expected from the liana syndrome such as the scarcity of axialparenchyma suggesting that not all features described under the liana vascular syndromeare necessarily present in every lianescent lineagePaulliniahas a diversity of cell types thatcontribute to storage including parenchyma intermediate elements non-septate and sep-tate fibersWe also observed that rays widen at sites of differential growth to accommodategrowth and avoid stem fractures
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

296 IAWA Journal 41 (3) 2020
ACKNOWLEDGEMENTS
The corresponding authors contributed equally to this article The authors thank the American Society ofPlant Taxonomists Society of Systematic Biologists and a Smithsonian Predoctoral Research Fellowshipand Fundaccedilatildeo de Amparo agrave Pesquisa do Estado de Sao Paulo (FAPESP Proc 201717107-3) for funding Wealso thank Stanley Yankowski for access to the Plant Anatomy Lab at the Smithsonian Museum of NaturalHistory and Steve Ruzin and Denise Schichnes for access to the Berkeley Bioimaging Facility
REFERENCESAcevedo-Rodriacuteguez P vanWelzen PC AdemaF van derHamRWJM 2011 Sapindaceae In Kubitzki K
(ed) The families and genera of vascular plants Eudicots Sapindales Cucurbitales MyrtaceaeSpringer Berlin 357ndash407
Acevedo-Rodriacuteguez PWurdackKJ FerrucciMS JohnsonGDias P CoelhoRG SomnerGV SteinmannVW Zimmer EA Strong MT 2017 Generic relationships and classification of tribe Paullinieae(Sapindaceae) with a new concept of supertribe Paulliniodae Syst Bot 42 96ndash114 DOI 101600036364417X694926
Angyalossy V Angeles G Pace MR Lima AC Dias-Leme CL Lohmann LG Madero-Vega C 2012 Anoverview of the anatomy development and evolution of the vascular system of lianas Plant EcolDivers 5 167ndash182 DOI 101080175508742011615574
Angyalossy V Pace MR Evert RF Marcati CR Oskolski AA Terrazas T Kotina E Lens F Mazzoni SCAngeles G Machado SR Crivellaro A Rao KS Junikka L Nikolaeva N Baas P 2016 IAWA list ofmicroscopic bark features IAWA J 37 517ndash615 DOI 10116322941932-20160151
Angyalossy V PaceMR Lima AC 2015 Liana anatomy a broad perspective on structural evolution ofthe vascular system In Schnitzer SA Bongers F Burnham RJ Putz FE (eds) Ecology of lianas251ndash287 JohnWiley amp Sons West Sussex DOI 1010029781118392409ch19
Arauacutejo GUC Costa CG 2007 Anatomia do caule de Serjania corrugata Radlk (Sapindaceae) ActaBot Bras 21 489ndash497 DOI 101590S0102-33062007000200021
Baas P Zweypfenning RCVJ 1979 Wood antomy of Lythraceae Acta Bot Neerl 28 117ndash155 DOI 101111j1438-86771979tb00329x
Barbosa ACF Pace MR Witovisk L Angyalossy V 2010 A new method to obtain good anatomicalslides of heterogeneous plant parts IAWA J 31 373ndash383 DOI 103969jissn1673-1719201403011
Barbosa ACF Costa GRO Angyalossy V Dos Santos TC Pace MR 2018 A simple and inexpensivemethod for sharpening permanent steel knives with sandpaper IAWA J 39 497ndash503 DOI 10116322941932-20170212
Bastos CLTamaioN AngyalossyV 2016 Unravelling roots of lianas a case study in Sapindaceae AnnBot 118 733ndash746 DOI 101093aobmcw091
Bouda M Windt CW McElrone AJ Brodersen CR 2019 In vivo pressure gradient heterogeneity in-creases flow contribution of small diameter vessels in grapevine Nat Commun 10 5645 DOI 101038s41467-019-13673-6
Buerki S Forest F Acevedo-Rodriacuteguez P Callmander MW Nylander JAA HarringtonM Sanmartiacuten IKuumlpfer PAlvarezN 2009 Plastid andnuclearDNAmarkers reveal intricate relationships at subfa-milial and tribal levels in the soapberry family (Sapindaceae) Mol Phylogenet Evol 51 238ndash258DOI 101016jympev200901012
Bukatsch F 1972 Bermerkungen zur Doppelfaumlrbung Astrablau-Safranin Mikrokosmos 61 255Burnham R 2009 An overview of the fossil record of climbers bejucos sogas trepadoras lianas
cipoacutes and vines Rev Bras Paleontol 12 149ndash160 DOI 104072rbp2009205Caballeacute G 1993 Liana structure function and selection a comparative study of xylem cylinders of
tropical rainforest species in Africa and America Bot J Linn Soc 113 41ndash60 DOI 101111j1095-83391993tb00328x
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

Chery et al ndashWood anatomy of Paullinia 297
Cabanillas PA PaceMR AngyalossyV 2017 Structure and ontogeny of the fissured stems of Callaeum(Malpighiaceae) IAWA J 38 49ndash66 DOI 10116322941932-20170156
Carlquist S 1985 Observations on functional wood histology of vines and lianas vessel dimorphismtracheids vasicentric tracheids narrow vessels and parenchyma Aliso 11 139ndash157
Carlquist S 1991 Anatomy of vine and liana stems a review and synthesis In Putz FE Mooney HA(eds) The biology of vines 53ndash72 Cambridge University Press Cambridge DOI 101017CBO9780511897658004
Carlquist S 2001 Cambial variants (anomalous secondary growth) In Comparative wood anatomySpringer series in wood science 271ndash295 Springer Berlin DOI 101016jympev2019106577
Carlquist S 2007 Successive cambia revisited ontogeny histology diversity and functional signifi-cance J Torrey Bot Soc 134 301ndash332 DOI 1031591095-5674(2007)134
Carlquist S 2013 Interxylary phloem diversity and functions Brittonia 65 477ndash495 DOI 101007s12228-012-9298-1
Carlquist S 2014 Fiber dimorphism cell type diversification as an evolutionary strategy in an-giosperm woods Bot J Linn Soc 174 44ndash67 DOI 101111boj12107
Chery JG Acevedo-Rodriacuteguez P Rothfels CJ Specht CD 2019 Phylogeny of Paullinia L (PaullinieaeSapindaceae) a diverse genus of lianas with dynamic fruit evolution Mol Phylogenet Evol 140106577 DOI 101016JYMPEV2019106577
Chery JG Pace MR Acevedo-Rodriacuteguez P Specht CD Rothfels CJ 2020 Modifications during earlyplant development promote the evolution of naturersquos most complex woods Curr Biol 30 1ndash8DOI 102139ssrn3471317
Cunha Neto IL Martins FM Somner GV Tamaio N 2017 Secretory structures in stems of five lianasof Paullinieae (Sapindaceae) morphology and histochemistry Flora 235 29ndash40 DOI 101016jflora201709001
Cunha Neto IL Martins FM Somner GV Tamaio N 2018 Successive cambia in liana stems ofPaullinieae and their evolutionary significance in Sapindaceae Bot J Linn Soc 186 66ndash88 DOI101093botlinneanbox080
DobbinsDR 1971 Studies on the anomalous cambial activity inDoxanthaunguis-cati (Bignoniaceae)II A case of differential production of secondary tissues Am J Bot 58 697ndash705 DOI 101002j1537-21971971tb10022x
Dobbins DR Fisher JB 1986Wound responses in girdled stems of lianas Bot Gaz 147 278ndash289 DOI101086337595
Fisher JB Ewers FW 1989 Wound healing in stems of lianas after twisting and girdling injuries BotGaz 150 251ndash265 DOI 101086337770
Franklin GL 1945 Preparation of thin sections of synthetic resins and wood-resin composites and anewmacerating method for wood Nature 155 3924ndash3951 DOI 101038155051a0
HeimschC 1942Comparative anatomyof the secondary xylem in the ldquoGruinalesrdquo and ldquoTerebinthalesrdquoof Wettstein with reference to taxonomic grouping Lilloa 8 83ndash198
HorakKE 1981 Anomalous secondary thickening in Stegnosperma (Phytolaccaceae) Bull Torrey BotClub 108 189ndash197 DOI 1023072484898
IAWA Committee 1989 IAWA list of microscopic features for hardwood identification IAWA J 10221ndash332 DOI 10116322941932-20160151
Isnard S SilkWK 2009Movingwith climbing plants fromCharlesDarwinrsquos time into the 21st centuryAm J Bot 96 1205ndash1221 DOI 103732ajb0900045
Johansen D 1940 Plant microtechnique McGraw-Hill New YorkKlaassen R 1999 Wood anatomy of the Sapindaceae IAWA J suppl 2 International Association of
Wood Anatomist LeidenKraus JE Arduin M 1997 Manual baacutesico de meacutetodos em morfologia vegetal Ed EDUR Seropeacutedica
Rio de Janeiro
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

298 IAWA Journal 41 (3) 2020
Krings M Kerp H Taylor TN Taylor EL 2006 How Paleozoic vines and lianas got off the groundon scrambling and climbing CarboniferousndashEarly Permian Pteridosperms Bot Rev 69 204ndash224DOI 1016630006-8101(2003)069[0204hpvalg]20co2
Leal MOL Nascimento LB Coutinho AJ Tamaio N Brandes AFN 2020 Development of external vas-cular cylinders (neoformations) in stems and roots of Chiococca alba (L) Hitchc (Rubiaceae)Flora 264 151569 DOI 101016jflora2020151569
Metcalfe CR Chalk L 1950 Anatomy of the dicotyledons Clarendon Press OxfordMorris H Plavcovaacute L Cvecko P Fichtler E Gillingham MAF Martiacutenez-Cabrera HI McGlinn DJ
Wheeler E Zheng J Ziemińska K Jansen S 2016 A global analysis of parenchyma tissue fractionsin secondary xylem of seed plants New Phytol 209 1553ndash1565 DOI 101111nph13737
Obaton M 1960 Les lianes ligneuses a structure anormale des forecircts denses drsquoAfrique occidentaleAnn Sci Nat Bot Ser 12 1ndash220
Pace MR Lohmann LG Angyalossy V 2009 The rise and evolution of the cambial variant in Bignon-ieae (Bignoniaceae) Evol Dev 11 465ndash479 DOI 101111j1525-142X200900355x
Pace MR Lohmann LG Olmstead RG Angyalossy V 2015 Wood anatomy of major Bignoniaceaeclades Plant Syst Evol 301 967ndash995 DOI 101007s00606-014-1129-2
Pace MR Acevedo-Rodriacuteguez P Amorim A Angyalossy V 2018a Ontogeny structure and occurrenceof interxylary cambia in Malpighiaceae Flora 241 46ndash60 DOI 101016jflora201802004
Pace MR Angyalossy V Acevedo-Rodriacuteguez P Wen J 2018b Structure and ontogeny of successivecambia in Tetrastigma (Vitaceae) the host plants of Rafflesiaceae J Syst Evol 56 396ndash400 DOI101111jse12303
Patil VS Marcati CR Rajput KS 2011 Development of intra- and interxylary secondary phloem inCoccinia Indica (Cucurbitaceae) IAWA J 32 475ndash491 DOI 10116322941932-90000072
Pellissari LCO Barros CF Medeiros H Tamaio N 2018 Cambial patterns of Paullinia (Sapindaceae)in southwestern Amazonia Brazil Flora 246 71ndash82 DOI 101016jflora201807002
Pfeiffer H 1926 Das Abnorme Dickenwachstum ndash Handbuch der Pflanzenanatomie Band IX Ge-bruumlder Borntraaeger Berlin
R Core Team 2018 R A language and environment for statistical computing R Foundation for Statis-tical Computing Vienna Austria httpswwwR-projectorg
Radlkofer L 1895 Monographie der Sapindaceen-Gattung Paullinia K B Akademie Munich DOI105962bhltitle65993
RajputKS RaoKS 2003 Cambial variant andxylemstructure in the stemof Cocculus hirsutus (Menis-permaceae) IAWA J 24 411ndash420 DOI 10116322941932-90000345
Rajput KS Marcati CR 2013 Stem anatomy and development of successive cambia in Hebanthe eri-antha (Poir) Pedersen a neotropical climbing species of theAmaranthaceae Plant Syst Evo 2991449ndash1459 DOI 101007s00606-013-0807-9
Rowe N Speck T 2004 Plant growth forms an ecological and evolutionary perspective New Phytol166 61ndash72 DOI 101111j1469-8137200401309x
Rowe N Isnard S Gallenmuumlller F Speck T 2006 Diversity of mechanical architectures in climbingplants an ecological perspective In Herrel A Rowe NP Speck T (eds) Biomechanics and ecol-ogy Dekker 36ndash59
Rupp P 1964 Polyglykol als Einbettungsmedium zum Schneiden botanischer Praumlparate Mikrokos-mos 53 123ndash128
Sauter JJ Iten W Zimmermann MH 1973 Studies on the release of sugar into the vessels of sugarmaple (Acer saccharum) Can J Bot 51 1ndash8 DOI 101139b73-001
Schenk H 1893 Beitraumlge zur Biologie und Anatomie der Lianen im besonderen der in Brasilien ein-heimischen Arten In Schimper AFM (ed) Botanische Mittheilungen Aus Den Tropen 1ndash71Gustav Fisher Jena
Schneider CA RasbandWS Eliceiri KW 2012 NIH Image to ImageJ 25 years of image analysis NatMethods 9 671ndash675 DOI 101038nmeth2089
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

Chery et al ndashWood anatomy of Paullinia 299
Tamaio N 2006 Anatomia do caule e da raiz em Menispermaceae PhD thesis Universidade de SatildeoPaulo Satildeo Paulo Brasil
Tamaio N 2011Wood anatomy of lianas of Sapindaceae commercially used in Satildeo PaulomdashSP Cerne17 533ndash540
Tamaio N Angyalossy V 2009 Variaccedilatildeo cambial em Serjania caracasana (Sapindaceae) enfoque naadequaccedilatildeo terminoloacutegica Rodrigueacutesia 60 651ndash666 DOI 1015902175-7860200960311
Tamaio N Brandes AFN 2010 Xylem structure of successive rings in the stem of Abuta grandifolia(Menispermaceae) a statistical approach IAWA J 31 309ndash316 DOI 10116322941932-90000025
Tamaio N Neves MF Brandes AFN Vieira RC 2011 Quantitative analyses establish the central vas-cular cylinder as the standard for wood-anatomy studies in lianas having compound stems(Paullinieae Sapindaceae) Flora 206 987ndash996 DOI 101016jflora201107006
Tamaio N Somner GV 2010 Development of corded vascular cylinder in Thinouia restingae Ferruciamp Somner (Sapindaceae Paullinieae) J Torrey Bot Soc 137 319ndash326 DOI 10315910-RA-0471
TerWelle BJH Koek-Noorman J 1978 On fibers parenchyma and intermediate forms in the genusMi-conia (Melastomataceae) Acta Bot Neerl 27 1ndash9 DOI 101111j1438-86771978tb01142x
Van derWalt JJA Van der Schijff HP SchweickerdtHG 1973 Anomalous secondary growth in the stemof lianas Mikania cordata (Burm F) Robins (Compositae) and Paullinia pinnata Linn (Sapin-daceae) Kirkia 9 106ndash138
Wagner ST Isnard S Rowe NP Samain MS Neinhuis C Wanke S 2012 Escaping the lianoid habitEvolution of shrub-like growth forms in Aristolochia subgenus Isotrema (Aristolochiaceae) AmJ Bot 99 1609ndash1629 DOI 103732ajb1200244
Zumaya-Mendoza S Terrazas T 2016 Vessel element and fiber length variation in successive cambiaof Iresine latifolia (Amaranthaceae) Rev Mex Biodiv 87 1315ndash1320 DOI 101016jrmb201610006
Edited by Pieter Baas
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace
300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace

300 IAWA Journal 41 (3) 2020
APPENDIX
Histograms of log-transformed vessel element diameters for each species in the study Inall plots y-axis = count x-axis = log-transformed vessel element value
Downloaded from Brillcom09042020 074743PM by marcelorpaceyahoocombrvia Marcelo Pace





























