Embed Size (px)
Citation preview

VODNI BALANS
BILJAKA

Da bi biljka opstala, neophodno je da postigne balans između primanja i gubljenja
vode.
Vodene biljke nemaju problem sa postizanjem vodnog balansa, jer celom
svojom površinom mogu da usvajaju vodu.
Neki kopneni organizmi (alge, lišajevi i mahovine) mogu da usvajaju vodu iz
vlažnog vazduha.
Međutim, većina kopnenih viših biljaka mora da reši dva problema: kako da do
vode dođe i kako da usvojenu vodu zadrži u meri koja omogućava normalno
funkcionisanje

Proučavanje vodnog balansa biljaka
obuhvata:
•Primanje vode iz zemljišta
•Transport vode kroz biljku
•Odavanje vode u atmosferu

PRIMANJE VODE PUTEM
KORENA

Osobine korenovog sistema
• Kod kopnenih biljaka obično je samo korenov sistem u dodiru sa vodom iz zemljišta. Stoga se koren tokom evolucije posebno prilagodio za funkciju usvajanja vode.
• Pošto iscrpi vodu iz svoje neposredne blizine, koren samo rastenjem može da dopre do delova zemljišta koji su još dovoljno vlažni. Zbog toga on raste relativno brzo i grana se vrlo bujno, tako da bočni korenovi prožimaju sve delove tla na kome biljka živi.
• Prema nekim podacima ukupna površina korenskih dlačica samo jedne jedinke može da iznosi i nekoliko stotina m2, što predstavlja ogromnu funkcionalnu površinu.
• Potreba za ovako velikim korenovim sistemom može se lakše shvatiti kad se zna da koren ne može da uzima vodu celom svojom površinom.

Uzdužni presek kroz apikalni
region korena
• Na vrhovima svih korenova, ispod korenske kape, nalazi se meristemska zona, u kojoj su ćelije bez krupnih vakuola.
• Meristemska zona se nastavlja u zonu izduživanja u kojoj se u ćelijama formira centralna vakuola i u kojoj ćelije dostižu svoju konačnu veličinu. Tek iznad tog dela korena nalazi se zona u kojoj su ćelije morfološki diferencirane.
• Epidermalne ćelije u ovoj zoni razvijaju korenske dlačice, a u centralnom cilindru se javljaju prvi ksilemski elementi. Ta zona korena je jedino aktivna u primanju vode za potrebe cele biljke.
• Simbioza sa gljivama (mikoriza)


Apsorpcija vode u korenu
• Koren prima vodu iz zemljišta osmotskim putem
• To znači da vodni potencijal zemljišta mora biti viši od vodnog
potencijala ćelija korena, a posebno korenskih dlačica.
• Procesi pomoću kojih biljke regulišu svoj osmotski, a time i
vodni potencijal obuhvaćeni su nazivom osmoregulacija.
• Primanje vode korenom može se prikazati sledećom formulom:
korena - tla
Wa = A x -------------------
Σ r
gde je Wa količina vode koju apsorbuje korenov sistem u
jedinici vremena po jedinici volumena tla, A – površina
korenskih dlačica, korena - tla razlika vodnih potencijala
korena i tla, i Σ r – zbir transportnih otpora za vodu u tlu i pri
prelasku iz tla u biljku.

Spoljašnji činioci koji utiču na primanje vode su:
1. vodni potencijal rastvora u zemljištu. Kada je zemljište zasićeno vodom, zemljišni rastvor ima visok vodni potencijal i biljke lako iz njega primaju vodu. Biljke koje žive na podlozi sa visokom koncentracijom soli su na poseban način prilagođene da iz nje apsorbuju vodu (halofite).
2. temperatura. Niska temperatura zemljišta može dvojako da utiče na primanje vode:
• povećava se broj vodoničnih veza među molekulima vode, čime je difuzija usporena
• koren na niskoj temperaturi sporije raste i tako ne dopire do delova zemljišta koji su bogati vodom.

3. aeracija zemljišta ima takođe značaja za rastenje korena. Osim izvesnih izuzetaka (npr. pirinač), koreni većine biljaka sporo rastu pri sniženom parcijalnom pritisku kiseonika i to onemogućava primanje vode. Osim rastenja, niska temperatura i slaba aeracija snižavaju i druge funkcije korena, kao što je disanje. U tom slučaju je smanjena apsorpcija mineralnih soli, što sprečava da koren primi vodu putem osmoze. Tako se može dogoditi da biljka vene, iako u zemljištu ima dovoljno vode, jer zbog niske temperature ili slabe aeracije ne može da je primi. Uslovi koji onemogućavaju primanje vode, iako je ona prisutna u zemljištu, označeni su kao stanje fiziološke suše.

Osobine zemljišta koje utiču na
primanje vode • Vodni potencijal () zemljišta obuhvata dve
komponente:
1. osmotski potencijal () vodenog rastvora u njemu i
2. hidrostatički pritisak (p)
• Osmotski potencijal zemljišta je veliki i retko ima
vrednost ispod – 0,01 MPa, jer voda u zemljištu obično
ne sadrži veću količinu rastvorenih soli. Prema tome, na
zemljištu koje nije slano, osmotski potencijal ne
predstavlja silu koja može da spreči primanje vode.
Izuzetak čine slana zemljišta, gde on može opasti do
– 0,2 MPa.

• Hidrostatički pritisak zavisi od količine vode u zemljištu. Na zemljištu koje sadrži vodu u izobilju on je blizu nule i biljke se vrlo dobro snabdevaju vodom, jer korenovi imaju vrednost vodnog potencijala sa negativnim predznakom.
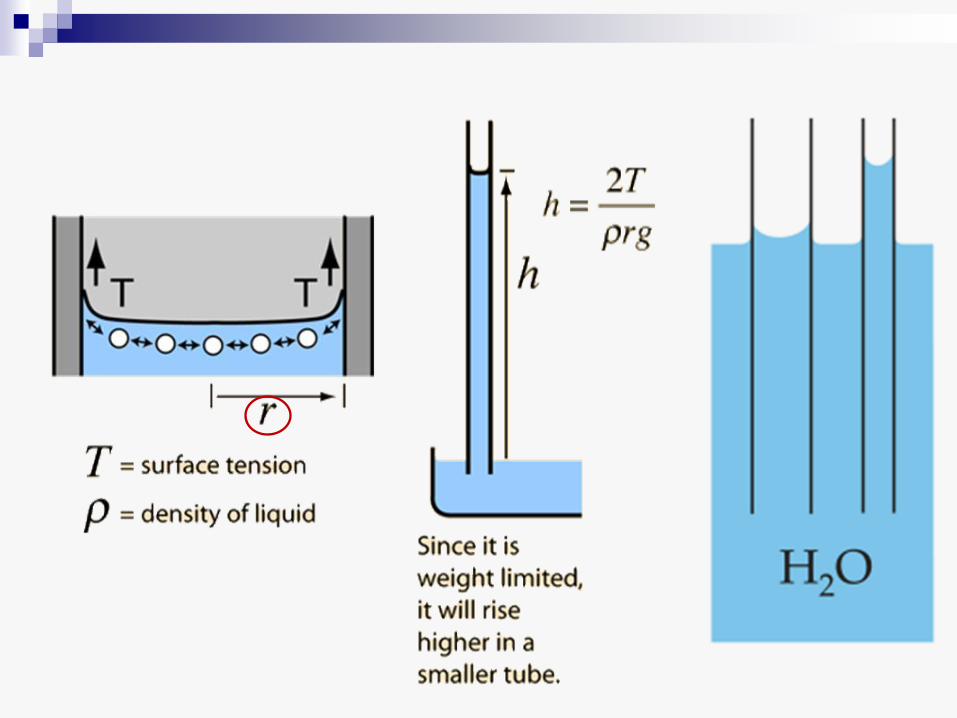
Sa sušenjem zemljišta voda iz kapilara isparava i zadržava se oko čestica tla (zbog adhezije), gradeći konkavne sferne površine. Pri tom se razvija tenzija, koja je utoliko veća ukoliko je radijus sferne površine manji. Ova tenzija može dostići vrednost od – 1 MPa, a na aridnim zemljištima i do – 3 MPa.
• Negativni hidrostatički pritisak (tenzija) koji se razvija sa povećanjem konkavne dodirne površine između vode i vazduha u kapilarima može da se proceni na osnovu sledeće formule:
-2T
p=--------
r
gde je T površinski napon vode (7,28 x 10-8 MPa m), a r radijus
krivine meniskusa
• Negativan hidrostatički pritisak, prema tome, predstavlja glavnu prepreku za apsorpciju vode iz zemljišta





• Voda može kroz zemljište da se kreće:
1. difuzijom usled razlika u osmotskom
potencijalu (što je spor proces i nema značaja za transport na veće distance), ili
2. kao struja cele vodene mase, usled
razlika u hidrostatičkom pritisku.
• Gradijent u hidrostatičkom pritisku se uspostavlja između neposredne i šire okoline korena, usled apsorpcije vode iz zemljišta.
• To izaziva strujanje vode ka korenu, a sa vodom se kreću i sve rastvorene supstance.
• Brzina ovog strujanja, ili hidraulička provodljivost zemljišta zavisi od veličine čestica, njihovog sastava i količine vode.
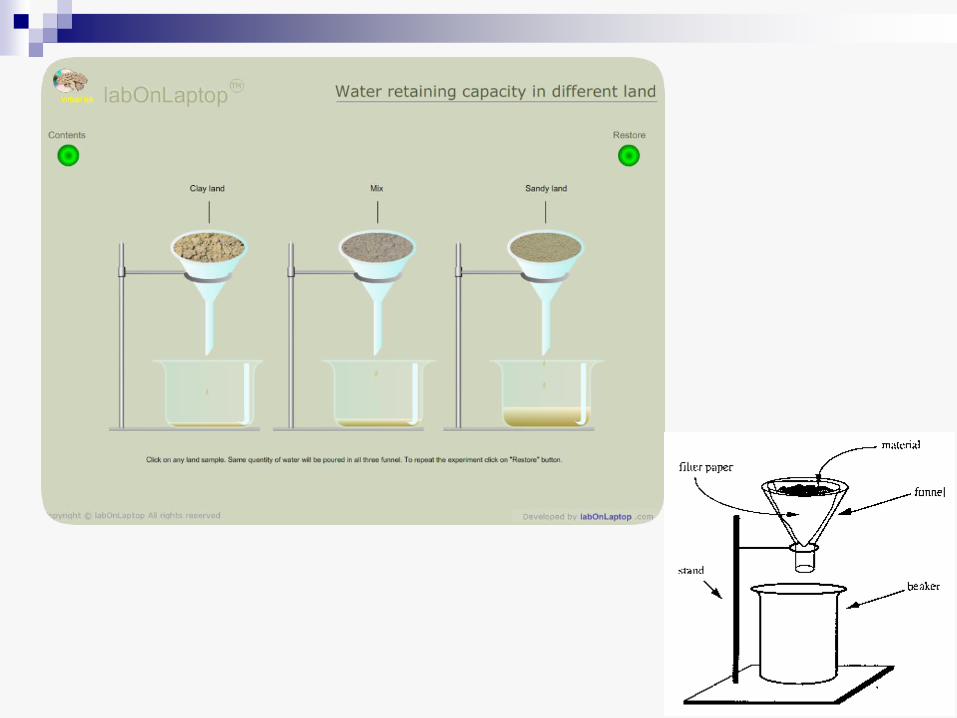

• Voda koju zemljište sadrži može biti u različitoj meri dostupna za biljku
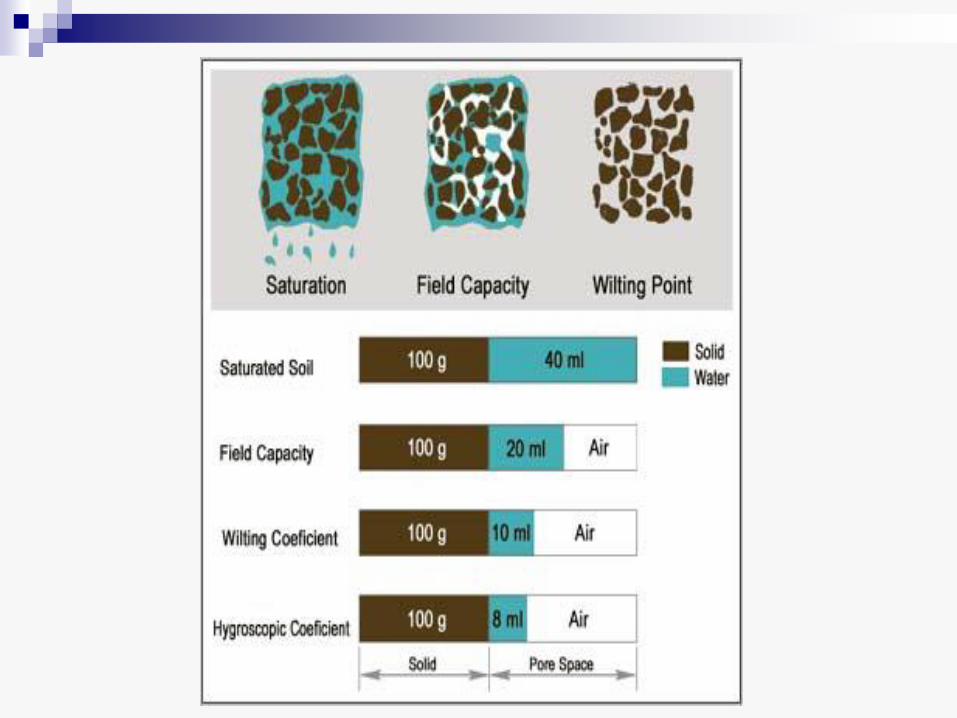
• Posle obilne kiše ili navodnjavanja, zemljište je zasićeno vodom, ali ona usled gravitacije brzo ponire u dublje slojeve. To je gravitaciona voda, koja ne koristi mnogo biljkama, u prvom redu zato što ispunjava sve pore u zemljištu i sprečava njegovu aeraciju.
• Kada se ovaj višak ocedi, zemljište još uvek sadrži znatnu količinu vode koja se zadržava u kapilarnim prostorima između čestica zemljišta. To je kapilarna voda, koja je biljci lako dostupna. Količina kapilarne vode je različita, zavisno od tipa zemljišta - ona je visoka u humusnom zemljištu i glini, a vrlo mala u pesku.

• Količina vode koju zemlja sadrži pošto se ocedi gravitaciona voda označava se kao kapilarni kapacitet zemljišta (poljski kapacitet) za tu vrstu zemljišta.
• Pri punom poljskom kapacitetu vode u zemljištu ne iznosi više od - 0,03 MPa, i u takvim uslovima sve biljke mogu da primaju vodu.
• Količina vode koju zemljište sadrži kada se primeti uvelost listova, zove se koeficijent venjenja i on označava vodu koja je za biljku nedostupna (mrtva rezerva). Ova voda delimično pripada i vodenom omotaču oko čestica zemljišta i u tom slučaju se zove imbibiciona voda.
• Venjenje može da bude toliko jako da biljka ne može da se oporavi kada ponovo primi vodu. Količina vode koju zemljište u tom trenutku još sadrži zove se procenat trajnog venjenja.



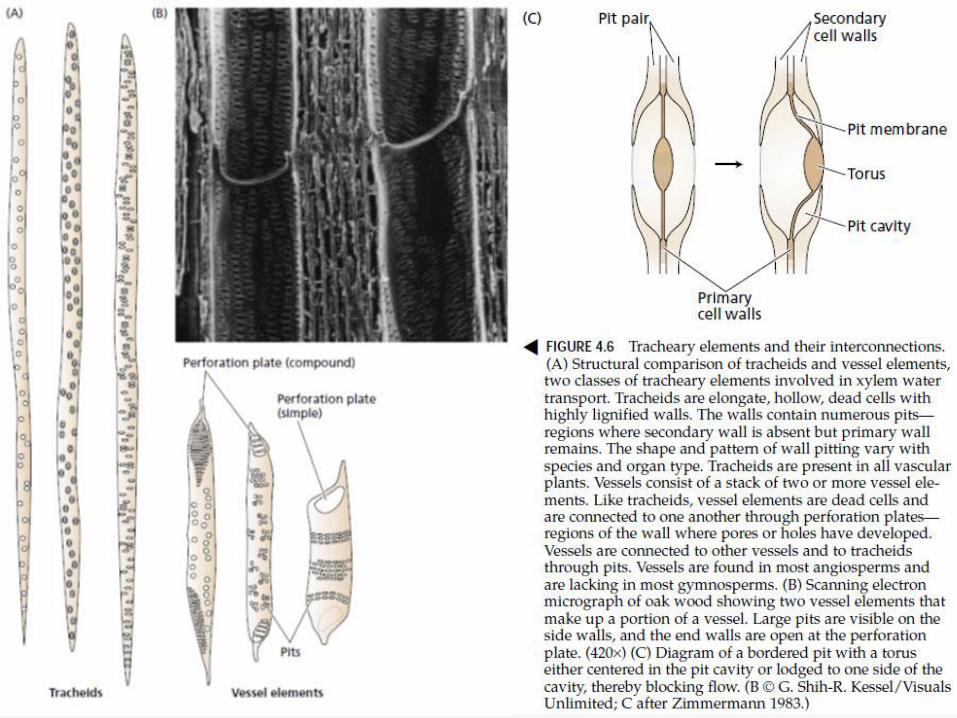
Put vode od korenske dlačice do
ksilema


KORENOV PRITISAK
• Tri pojave pokazuju da postoji sila koja vodu
potiskuje naviše i koja potiče iz korenovog
sistema:
1. Eksudacija
2. Suzenje (plač vinove loze)
3. Gutacija
• Ova sila se naziva korenov pritisak.

Kod mladih zeljastih biljaka se često
može primetiti da se voda izlučuje u
kapima sa oboda listova. Ova pojava
se naziva gutacija i do nje dolazi
kada biljka ima dosta vode u podlozi
i kada je transpiracija mala usled
velike vlažnosti vazduha
Kada se stablo neke biljke
preseče odmah iznad korena, na
presečenoj površini će se pojaviti
kapljice tečnosti, koja se iz korena
potiskuje naviše. Ova pojava se
zove eksudacija.


• Utvrđeno je da svi činioci koji deluju na uzimanje vode imaju uticaja i na korenov pritisak (pogodna temperatura, aeracija i visok vodni potencijal zemljišta).
• U najpovoljnijem slučaju, kod većine biljaka korenov pritisak ne prelazi vrednost od 0,1 MPa (1 atmosfera), a najčešće se kreće između 0,02 i 0,05 MPa (0,2 i 0,5 atmosfera).
• U retkim slučajevima su izmerene veće vrednosti korenovog pritiska.
• Najpoznatiji izuzetak predstavljaju izolovani korenovi paradajza, gajeni u sterilnoj kulturi in vitro, kod kojih je izmeren pritisak od 0,606 MPa.

Dnevna variranja
• Karakteristično je za korenov pritisak da njegova veličina znatno varira u toku dana kod iste biljke, čak i pod konstantnim spoljašnjim uslovima.
• Korenov pritisak dostiže najveće vrednosti u jutarnjim časovima, a zatim se smanjuje, tako da u podne predstavlja sasvim nemerljivu veličinu.
• Zbog toga je značaj korenovog pritiska za snabdevanje biljke vodom u toku dana ograničen. Međutim, on ima presudnu ulogu u nadoknađivanju deficita vode (nastalog usled intenzivne transpiracije) tokom noći i pred zoru.

Uporedno merenje korenovog pritiska, količine
eksudata i koncentracije jona u eksudatu
Postoji endogena dinamika u veličini svih merenih parametara i promene
su paralelne, što ukazuje na to da su sva tri procesa međusobno
povezana.

Kako nastaje korenov pritisak?
• Za objašnjenje o tome kako nastaje korenov pritisak od velikog je značaja činjenica da je eksudacija povezana sa aktivnim transportom jona u korenu.
• Zapaženo je da je osmotski potencijal eksudata uvek niži (sadrži veću koncentraciju soli) od osmotskog potencijala rastvora u zemljištu.
• To znači da parenhimske ćelije korena prenose jone do ksilemskih elemenata uz gradijent koncentracije.

• Smatra se da žive ćelije korena aktivno transportuju jone u unutrašnjost ksilemskih sudova.
• Ceo korenov sistem tada postaje analog osmometru. Sok u ksilemu odgovara rastvoru u unutrašnjem sudu osmometra, a membrane svih ćelija predstavljaju jednu složenu diferencijalno propustljivu membranu, koja razdvaja dva rastvora različitih koncentracija.
• Voda osmotskim putem ulazi spolja, iz rastvora niže koncentracije soli, u unutrašnjost sudova i nivo rastvora u njima raste, kao što raste i u cevi osmometra.
• Korenov pritisak, je, prema tome, analog pritisku koji vrši rastvor u kapilarnoj cevi osmometra (deluje naviše).

Pošto je korenov pritisak povezan
sa aktivnim transportom jona,
jasno je da je i za eksudaciju
potrebna energija. Ćelije korena
dobijaju ovu energiju iz disanja i
stoga je razumljivo što niska
temperatura ili slaba aeracija
zemljišta ometaju i pojavu
korenovog pritiska.


• Zbog dnevnih variranja i nesposobnosti da podigne vodu na veliku visinu korenovom pritisku se danas ne pridaje presudan značaj u transportu vode kroz biljku i snabdevanju nadzemnih delova biljke vodom.
• Presudan značaj za kretanje vode kroz biljku ima
transpiracija (odavanje vode, u obliku vodene pare, u atmosferu, preko nadzemnih delova biljke).
• Transpiracija ispunjava dva osnovna uslova neophodna za ulogu ključne sile koja pokreće vodu kroz biljku:
1. da je sila koja izaziva difuziju vodene pare iz listova dovoljno velika da savlada silu Zemljine teže i trenje o zidove suda
2. da stub vode u kapilarnim sudovima može da izdrži velika istezanja tako da se struja vode ne prekine

• Transpiracija generiše gradijent vodnog potencijala celom dužinom biljke.
• Kretanje vode kroz biljku, prema tome, moglo bi da se obavi difuzijom niz dovoljan gradijent u vodnom potencijalu. Međutim, difuzija je spor proces i nije efikasna u transportu na veća rastojanja.
• Vodni potencijal atmosfere (koncentracija vodene pare u vazduhu) postaje dominantan za transport kontinualne vodene mase kroz ksilem.
• Vodni potencijal atmosfere je veoma nizak, čak i pri zasićenju. Razlika u vodnom potencijalu na graničnoj površini listova dovodi do isparavanja vode sa listova. Gubitak vode iz intercelulara lista generiše negativan hidrostatički pritisak – tenziju u intercelularima lista (isto kao u zemljištu), što predstavlja pokretačku silu za kretanje struje vode niz gradijent hidrostatičkog pritiska duž ksilema.
• Usled velike kohezije molekula vode, tanke niti vode u kapilarnim cevima ksilema ostaju neprekinute i pored velikog istezanja. U ksilemskim sudovima, vodeni rastvor skoro da ima čvrstinu metalne žice.
DIKSONOVA TENZIONO-KOHEZIONA
TEORIJA


• Obrazovanje gasnih mehurova u ksilemu (usled tenzije vode nastale usisavanjem), koji prekidaju vodenu nit, naziva se embolija ili kavitacija. Ova pojava nestaje kad transpiraciona tenzija opadne (u toku noći) a gasovi se ponovo rastvore u vodi.
• Brzina kretanja vodenog rastvora kroz ksilem izražava se kao količina vode koja se transportuje kroz stablo u jedinici vremena, kroz jedinicu površine sudova kojima se transport obavlja. Ona zavisi u najvećoj meri od spoljašnjih uslova, intenziteta transpiracije, od vrste i od površine poprečnog preseka sudova. Može da dostigne i do 45 m h-1.


TRANSPIRACIJA

DEFINICIJA:
Pod transpiracijom se podrazumeva
odavanje vode, u obliku vodene pare, u
atmosferu, preko nadzemnih delova biljke.
Voda se odaje u obliku vodene pare, ali se
transpiracija bitno razlikuje od isparavanja sa
površine slobodne vode ili drugih tela.
Tokom evolucije, kopnene biljke su stekle
morfološke i fiziološke osobine pomoću kojih
mogu da regulišu transpiraciju i da se zaštite
od njenog štetnog efekta

Korist i šteta od transpiracije
• Proračunato je da samo jedan kukuruz transpiriše oko 200 L vode u toku svoje vegetacione periode. Jedno razgranato drvo može toliko vode da izgubi za samo jedan letnji dan.
• Ovako velike količine vode biljke moraju da apsorbuju iz zemljišta, iako koriste najviše 1-5% vode koja prođe kroz njih u atmosferu.
• Ono što biljka samo propusti zove se tranzitna voda.
• Transpiracija je, u stvari, proces koji je za biljke neizbežan.

Transpiracija je “nužno zlo” za biljku.

Osnovne funkcije transpiracije su:
1. omogućavanje toka vode i u njoj rastvorenih materija (transpiracioni tok) kroz biljku - bez utroška energije sa transpiracionim tokom prenose se kroz biljku i mineralne soli.
2. hlađenje površine sa koje voda isparava (sa lista). Pošto voda ima vrlo visoku temperaturu isparavanja, jasno je da se pri transpiraciji gube i znatne količine toplotne energije. Ako je temperatura toliko visoka, da ćelije lista mogu biti oštećene, ili njihove funkcije smanjene, onda sniženje temperature lista, makar i za nekoliko stepeni, može biti vrlo značajno za preživljavanje.

Vrste transpiracije • Kutikularna transpiracija obavlja
se sa površine epidermalnih ćelija koje su prevučene kutikulom.
• Lenticelarna transpiracija obavlja se kroz lenticele, ali je ona obično vrlo mala u odnosu na ukupnu transpiraciju. Izuzetno, u toku zime kod listopadnog drveća jedini gubitak vode može biti preko lenticela.
• Stomaterna transpiracija je transpiracija kojom se gubi najviše vode iz biljaka, ali i jedini tip transpiracije koji podleže fiziološkoj kontroli.


Merenje transpiracije
• Transpiracija se određuje po količini vode koju biljka izgubi u nekom vremenskom periodu.
• Najčešće se izračunava merenjem težine biljke, težine jedne grane ili pojedinačnih listova u vrlo kratkim razmacima vremena (1-5 min), kada promene u težini mogu da znače samo promenu u količini vode.
• Intenzitet transpiracije se dobija kada se gubitak u težini sračuna na jedinicu površine lista, u jedinici vremena. On se izražava kao gram izgubljene vode, na 1 dm2, za sat .



• Druge veličine kojima se izražava
transpiracija predstavljaju u stvari odnos
između transpiracije i fotosinteze.
• Produktivnost transpiracije pokazuje koliko
se grama suve supstance izgradi u listu za
vreme za koje se transpiracijom izgubi 1 kg
vode.
• Recipročna vrednost od toga je koeficijent
transpiracije, koji pokazuje koliko vode
ispari iz lista, dok se izgradi 1 g suve
supstance.

Osobine biljaka koje utiču na
transpiraciju
• spoljašnja površina lista
• površina intercelulara u listu
• zaštitne tvorevine na listu, kao što su
kutikula, vosak, dlake i dr.
• položaj stoma na listu
• razvijenost provodnog sistema

Adaptacije
• Mnoge kserofite imaju manje ili više redukovanu lisnu površinu, što je izraz njihove adaptacije na sušne uslove
• Mnoge mezofite zbacuju sve, ili deo listova kada su izložene suši
• Unutrašnja površina isparavanja može biti veća od spoljašnje površine lista 7 do 30 puta
• Ukupna zapremina intercelulara je različita. U iglicama bora ona iznosi 5%, u listu kukuruza 10%, a duvana 40% ukupne zapremine lista
• Mnogi listovi imaju na epidermisu dlake, koje smanjuju transpiraciju time što zadržavaju sloj vlažnog vazduha uz list, a takođe reflektuju svetlost i smanjuju zagrevanje lista
• Kod većine biljaka stome se nalaze u ravni epidermisa, ali kod mnogih kserofita one su uvučene ispod ravni lisne ploče i time su zaštićene od strujanja vazduha
• Dobro razvijen provodni sistem u listu i stablu utiče na transpiraciju, time što omogućava dobro snabdevanje vodom iz zemljišta

Difuzija gasova kroz stome • Transpiracija se zasniva na difuziji vodene pare kroz
stome, a niz gradijent njenog parcijalnog pritiska.
• Razmena svih gasova između biljke i spoljašnje
sredine odvija se skoro isključivo preko listova, kroz
otvore stoma. U spoljašnju sredinu se izbacuju
vodena para i O2 (proizveden u fotosintezi), a iz
spoljašnje sredine se usvaja CO2 (neophodan za
fotosintezu).
• Kretanje gasova kroz stome, kao i njihovo kretanje kroz intercelulare obavlja se difuzijom. Brzina difuzije, tj. provodljivost stoma za gasove, direktno je proporcionalna gradijentu parcijalnih pritisaka (odnosno koncentracija) gasova između lista i atmosfere, a obrnuto proporcionalna otporu prema difuziji.

• Koncentracija vodene pare u intercelularima zavisi od:
1. vodnog potencijala lista i
2. temperature
• Stvarna koncentracija vodene pare pri ravnoteži zavisi od temperature i obično se izražava ne kao apsolutna, nego kao relativna vlažnost vazduha.
• Ovaj izraz podrazumeva količinu vode koju sadrži jedinica zapremine vazduha (1 m3), u odnosu na 100% zasićenja.
• List čiji je vodni potencijal v = 0, što znači da je potpuno zasićen vodom, ima u svojim intercelularima 100% relativnu vlažnost.
• Sa povišenjem temperature povećava se kapacitet vazduha da primi vodenu paru, i to sa porastom od 12 0C, koncentracija vodene pare raste dva puta.

• Listovi koji su izloženi svetlosti imaju uvek višu temperaturu od okolnog vazduha, pa je vodni potencijal vazduha niži od vodnog potencijala lista.
• Tome doprinosi i strujanje vazduha, kojim se odnosi vodena para. Tako se između lista i atmosfere uspostavlja manje ili više oštar gradijent u koncentraciji vodene pare; relativna vlažnost od 100% u listu do 50% i manje u okolini, predstavlja razliku usled koje para uvek difunduje iz lista u atmosferu.

• Ugljen dioksid prolazi takođe difuzijom kroz stome,
samo u suprotnom smeru od vodene pare.
• Gradijent u količini CO2 između atmosfere (0,03%
CO2) i lista (koji sadrži samo 0,01% CO2) je znatno
manji od gradijenta vodene pare.
• Mezofilne ćelije apsorbuju CO2 iz intercelulara,
čime se uspostavlja ova razlika u koncentraciji,
odnosno parcijalnom pritisku CO2.
• Zbog manje oštrog gradijenta, kao i zbog razlike u
veličini molekula, 1 molekul CO2 difunduje za isto
vreme kao približno 100 molekula vode.

• Drugi faktor koji utiče na brzinu difuzije je otpor (rezistencija), koji se javlja na putu vodene pare.
• Prvi otpor pružaju stome, koje mogu biti manje ili više otvorene. Tako se govori o provodljivosti stoma koja je regulisana fiziološkim procesima.
• Drugi otpor je fizičke prirode i on potiče od nepokretnog, mirnog sloja vazduha koji leži uz površinu lista, a koji sadrži veliku količinu vodene pare.
• To je označeno kao rezistencija graničnog sloja i takođe može biti varijabilna veličina.
• Takav sloj se obrazuje iznad svake površine koja isparava. Iznad vode u nekom otvorenom sudu taj sloj se nalazi kao stub pare koji može imati znatnu debljinu. Jasno je da on ograničava isparavanje i da od njegove debljine zavisi kojom brzinom će molekuli vode dospeti do slobodnih slojeva vazduha.


• Ako je voda prekrivena nepropustljivom folijom koja ima perforacije, jasno je da je ukupna površina isparavanja
znatno manja, ali je difuzija brža, jer je i granični sloj tanji.
Tako se dolazi do paradoksa malih otvora koji važi za stome i
kojim se objašnjava njihova visoka efikasnost u transpiraciji.
• Ukupna površina stoma na listu nije velika i obično ne prelazi 1% od površine lista. Kada se uporedi količina transpirisane vode sa količinom koja je isparila sa iste tolike slobodne površine, može se konstatovati da je transpiracija za oko 50% veća.
• Slični podaci su dobijeni i za ugljen dioksid; računajući na
jedinicu površine otvorenih stoma, apsorpcija je znatno veća
nego u sudu koji sadrži jak apsorbent za CO2, kao što je NaOH. Ova se pojava može objasniti time, što je difuzija kroz
male otvore srazmerna njihovom obimu, a ne površini.

• Veći broj malih otvora ima veći
ukupan obim u odnosu na površinu,
nego što je to slučaj kod velikog
otvora, pošto je obim srazmeran
poluprečniku (2r), a površina
njegovom kvadratu (r2).
• Zato je jasno zašto veći broj manjih
otvora ima veću efikasnost nego jedan veliki iste površine.

Isparavanje kroz male otvore

Otpor graničnog sloja zavisi od
strujanja vazduha. Na mirnom
vazduhu otpor je znatan i merenja
pokazuju da ima čak i veći značaj
za transpiraciju od otvorenosti
stoma.

Transpiracija lista Zebrina pendula merena na
mirnom vazduhu i na vetru, pri jednakoj otvorenosti
stoma

Uticaj spoljašnjih činilaca na
transpiraciju
Pri nepromenjenoj otvorenosti stoma, na transpiraciju utiču svi faktori koji
povećavaju gradijent u koncentraciji
vodene pare i koji smanjuju otpor
prema difuziji. To su:
1. Temperatura
2. Relativna vlažnost vazduha i
3. Strujanje vazduha (vetar)

• Sa povećanjem temperature povećava se
brzina difuzije.
• Temperatura deluje i na vlažnost vazduha.
Sa njenim povećanjem smanjuje se
relativna vlažnost, tj. određena zapremina
vazduha pri datoj temperaturi može da
primi veće količine vodene pare.
• Strujanje vazduha, tj. vetar, brže odnosi sa
površine lista sloj vazduha koji je zasićen
vodenom parom. Međutim, jak vetar (brži
od 15 km h-1) dovodi do hlađenja lista i time
se transpiracija ipak smanjuje.

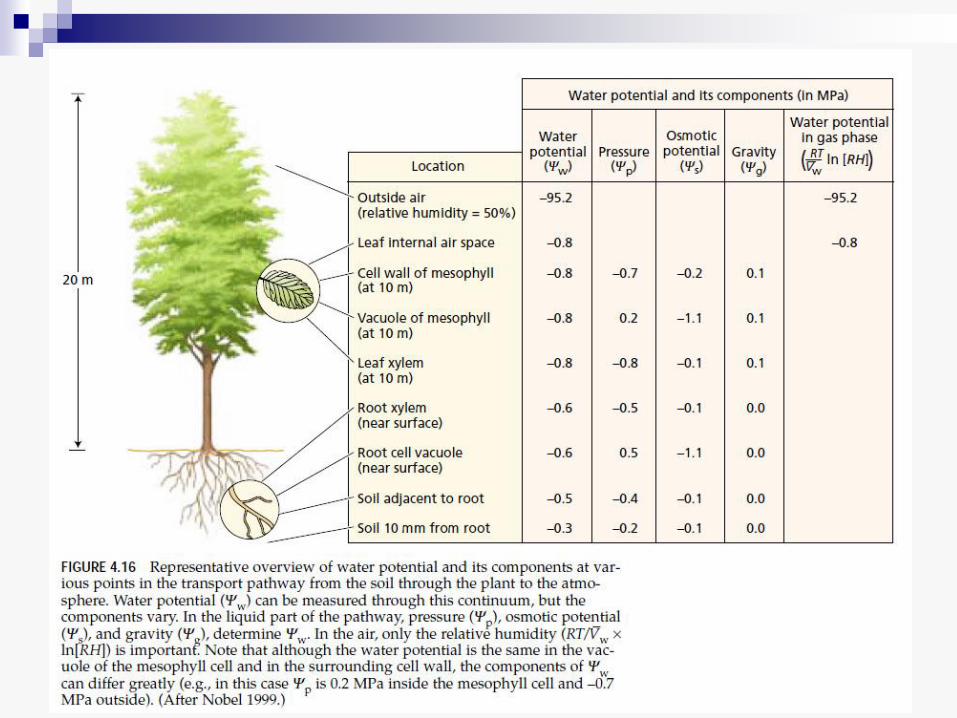
KRETANJE VODE KROZ
BILJKU

• Sistem „zemljište - biljka – atmosfera“
predstavlja kontinuum kad je reč o
kretanju vode.
• Na putu od korena do listova voda se
kreće kroz 2 tipa ćelija:
žive – kretanje od zemljišta do ksilema (kroz
apoplast ili simplast ćelija korena) i od kraja
provodnih snopića do površine lista (tj. do
ćelija koje se graniče sa intercelularima i sa
čije površine voda isparava)
mrtve – kretanje kroz ksilem, od korena do
listova (kroz stablo)

Postoje 3 načina kretanja vode kroz sistem „zemljište –
biljka – atmosfera“:
1. Osmoza (kretanje kroz ćelijske membrane u živim ćelijama)
2. Strujanje vodene mase (u zemljištu i u ksilemu) 3. Difuzija (kretanje vodene pare iz lista u atmosferu i u atmosferi)
Pokretačka sila za kretanje vode (u sva tri načina kretanja) zavisi od dve komponente:
a. osmotskog potencijala b. hidrostatičkog pritiska
Koja od ovih komponenti ima odlučujuću, glavnu ulogu u kretanju vode, zavisi od načina kretanja.
Gradijent osmotskog potencijala reguliše kretanje vode osmozom kroz žive ćelije i difuzijom prilikom transpiracije (ovde se koriste termini vodni potencijal, parcijalni pritisak vodene pare ili relativna vlažnost vazduha). Gradijent hidrostatičkog pritiska ima odlučujuću ulogu u kretanju vode kroz ksilem.

Provodljivost sistema „zemljište – biljka – atmosfera“ je obrnuto proporcionalna zbiru otpora na koje nailazi voda prilikom kretanja. Ti otpori su:
u kretanju osmozom kroz žive ćelije: otpor
prolasku kroz membrane (što više membrana
voda prođe, veći je otpor)
u kretanju kroz ksilem: trenje o zidove
ksilemskih elemenata i Zemljina teža
u transpiraciji: otpor stoma, otpor kutikule i
otpor graničnog sloja vazduha
• Najmanji otpor u sistemu je u ksilemu, a najveći je otpor zatvorenih stoma.

KRETANJE VODE KROZ ŽIVE ĆELIJE KORENA I LISTA
• Odvija se apoplastom ili simplastom, osmozom ili difuzijom, niz gradijent vodnog potencijala, tj. od zemljišta ka centralnom cilindru u korenu, odnosno od provodnih snopića ka površini lista u listovima.
KRETANJE VODE KROZ PROVODNE SNOPIĆE (KSILEM)
• Odvija se strujanjem mase vode niz gradijent hidrostatičkog pritiska.
• Na kretanje vode ksilemom utiču dve sile: korenov pritisak (potiskuje vodu naviše) i transpiracija (“vuče” vodu naviše)


















![Fiziologija 2 Vodni režim [Compatibility Mode]](https://img.dokumen.tips/doc/110x75/5572106b497959fc0b8d2337/fiziologija-2-vodni-rezim-compatibility-mode.jpg)










