Embed Size (px)
Citation preview

Vitamin D reduces deposition of advanced glycationend-products in the aortic wall and systemic oxidativestress in diabetic rats
Erik Salum a,b,c,*, Jaak Kals b,c,d, Priit Kampus a,b,c, Tiit Salum c, Kersti Zilmer c,Marina Aunapuu e,f, Andres Arend e, Jaan Eha a,b, Mihkel Zilmer b,c
aDepartment of Cardiology, University of Tartu, 8 Puusepa Street, Tartu 51014, EstoniabEndothelial Centre, University of Tartu, 8 Puusepa Street, Tartu 51014, EstoniacDepartment of Biochemistry, Centre of Excellence for Translational Medicine, University of Tartu, 19 Ravila Street, Tartu 50411, EstoniadDepartment of Vascular Surgery, Tartu University Hospital, 8 Puusepa Street, Tartu 51014, EstoniaeDepartment of Anatomy, University of Tartu, 19 Ravila Street, Tartu 50411, Estoniaf Institute of Veterinary Medicine and Animal Sciences, Estonian University of Life Sciences, 62 Fr. Kreutzwaldi Street, Tartu 51014, Estonia
d i a b e t e s r e s e a r c h a n d c l i n i c a l p r a c t i c e 1 0 0 ( 2 0 1 3 ) 2 4 3 – 2 4 9
a r t i c l e i n f o
Article history:
Received 5 September 2012
Received in revised form
17 January 2013
Accepted 1 March 2013
Published on line 21 March 2013
Keywords:
Streptozotocin
Total antioxidant capacity
Oxidative stress index
Ne-(carboxymethyl)lysine
Cholecalciferol
a b s t r a c t
Aims: Vitamin D may have an important role in reducing the risk of cardiovascular disease.
Advanced glycation end-products (AGEs) such as Ne-(carboxymethyl)lysine (CML), have
been implicated in diabetic vascular complications via oxidative stress-mediated pathways.
We investigated the potential protective effect of vitamin D on CML accumulation in the
diabetic aortic wall. To test the effects of vitamin D on systemic oxidative stress we also
assessed liver oxidative stress index (OSI) and serum total antioxidant capacity (TAC).
Methods: Male Wistar rats were assigned to three groups: control, untreated diabetes, and
diabetes + cholecalciferol. Diabetes was induced by streptozotocin, followed by oral admin-
istration of cholecalciferol (500 IU/kg) for 10 weeks in the treatment group. Aortic CML
accumulation was determined by ELISA and immunohistochemical assays. OSI was
assessed by measuring TAC and the level of total peroxides in the liver and serum using
colorimetric assays.
Results: Untreated diabetes was associated with significantly elevated CML levels in the
aortic wall (19.5 � 3.3 vs 10.2 � 4.7 ng/mL), increased liver OSI (6.8 � 1.9 vs 3.1 � 0.7), and
reduced serum TAC (0.4 � 0.1 vs 0.8 � 0.3 mmol Trolox/L), in comparison with the control
group. Cholecalciferol significantly blocked the accumulation of CML in the aortic wall
(10.4 � 8.4 vs 19.5 � 3.3 ng/mL), decreased liver OSI (4.2 � 1.4 vs 6.8 � 1.9), and improved
serum TAC (1.0 � 0.2 vs 0.4 � 0.1 mmol Trolox/L), compared with the untreated diabetic
group.
Conclusions: Streptozotocin-diabetes resulted in increased deposition of AGEs and increased
oxidative stress in the serum and liver. Vitamin D supplementation may provide significant
protection against oxidative stress-mediated vascular complications in diabetes.
# 2013 Elsevier Ireland Ltd. All rights reserved.
Contents available at Sciverse ScienceDirect
Diabetes Researchand Clinical Practice
journal homepage: www.elsevier.com/locate/diabres
* Corresponding author at: Department of Cardiology, University of Tartu, 8 Puusepa Street, Tartu 51014, Estonia. Tel.: +3727318317.E-mail address: [email protected] (E. Salum).
0168-8227/$ – see front matter # 2013 Elsevier Ireland Ltd. All rights reserved.http://dx.doi.org/10.1016/j.diabres.2013.03.008

d i a b e t e s r e s e a r c h a n d c l i n i c a l p r a c t i c e 1 0 0 ( 2 0 1 3 ) 2 4 3 – 2 4 9244
1. Introduction
Advanced glycation end-products (AGEs) are irreversibly
formed during glycation and oxidation of proteins [1]. Because
of its abundance in uncontrolled diabetes mellitus (DM), glucose
is assumed to be a major source of glycation and AGE formation.
AGEs can form covalent cross-links between the extracellular
matrix proteins in the vascular wall and alter their function,
particularly of collagen and elastin [2,3]. AGE cross-linking of
collagen has been associated with increased stiffness of the
arterial wall and impaired arterial haemodynamics [4].
Increases in the levels of Ne-(carboxymethyl)lysine (CML), a
major antigenic structure of AGEs [5], have been correlated with
the severity of diabetic complications [6,7], suggesting that CML
may be an important biomarker in the assessment of vascular
integrity in DM. Considering the emerging evidence about the
adverse effects of AGEs on the vascular properties, therapies
aimed at inhibiting AGEs may provide significant improvement
of diabetic vascular complications.
Oxidative stress is regarded as an important factor in the
pathogenesis and progression of DM and its associated
cardiovascular complications [8]. In diabetes, persistent
hyperglycemia is associated with increased production of
reactive oxygen species (ROS) [9] with concomitant depletion
of intrinsic antioxidant defense mechanisms [10], rendering
the organism more susceptible to oxidative damage. The
imbalance between oxidants and antioxidants could be largely
responsible for the functional and morphological damages to
the blood vessels associated with uncontrolled hyperglycemia
[11]. Oxidative reactions also enhance the formation of AGEs
and their accumulation in cardiovascular tissue [12], raising a
potential link between diabetic vascular complications and
deposition of AGEs.
Agents that enhance the antioxidant defense and reduce
the damage caused by oxidative stress to the blood vessels
may be beneficial in the treatment of diabetic vascular
complications. Several studies have demonstrated that vita-
min D may reduce lipid peroxidation [13,14] and maintain a
steady level of glutathione (GSH), a potent intracellular
antioxidant [15]. Calcitriol, the active form of vitamin D, has
been shown to improve vascular endothelial function by
modulating the expression of radical generating and scaveng-
ing enzymes, thus preventing the overproduction of ROS [16].
In our previous study, we demonstrated that chronic
supplementation of vitamin D improves aortic wall remodel-
ling in streptozotocin-induced diabetes [17]. To test the
possible ways how vitamin D may have protective effects
concerning diabetes-induced vascular abnormalities we in-
vestigated in the current study the effect of vitamin D on CML
deposition in the aortic wall, and its potential antioxidative
influence by assessing systemic oxidative stress-related
parameters in the serum and liver in a rat model of type 1 DM.
2. Methods
2.1. Animals
Male Wistar rats (n = 24, age 4 months), purchased from Harlan
Laboratories (Harlan Laboratories, Inc., The Netherlands),
were used in the experiments. The animals were housed in
a room maintained at 21 � 2 8C under a standard 12-h light/12-
h dark cycle and were given normal rat chow and tap water
ad libitum. All experimental procedures were approved by the
Estonian National Board of Animal Experiments and were
conducted in accordance with the European Communities
Directive (86/609/EEC).
2.2. Experimental protocol
Rats were randomly assigned to one of three groups: control,
diabetic, and cholecalciferol-treated diabetic group. Diabetes
was induced by an intraperitoneal administration of strepto-
zotocin (STZ) 50 mg/kg (Sigma–Aldrich, St. Louis, MO, USA),
freshly dissolved in 0.9% saline solution. Blood samples from
the tail vein were obtained 48 h later and glucose levels were
measured with a glucometer (Glucocard X-meter, Arkray Inc.,
Japan). All rats in the diabetic groups had blood glucose level
>15 mmol/L. Supplementation of cholecalciferol (Sigma–
Aldrich, St. Louis, MO, USA) 12.5 mg (500 IU) kg�1 body weight,
dissolved in 0.3 ml olive oil, was started immediately after
confirmation of diabetes. Cholecalciferol was administered
orally every other day for 10 weeks.
2.3. Laboratory parameters
At 10 weeks, the animals were anaesthetised with a mixture of
fentanyl (0.07 mg/kg, Gedeon-Richter Plc., Hungary), midazo-
lam (5 mg/kg, Roche Pharma AG, Germany), and ketamine
(75 mg/kg, Vetoquinol Biowet Sp. z.o.o., Poland) administered
subcutaneously. Blood samples were taken from the tail vein
for assessment of glucose levels. Anaesthetised animals were
subjected to cardiac puncture for blood withdrawal, followed
by cervical dislocation. One part of blood sample was used for
assessment of glycated haemoglobin (HbA1c) and glucose
levels, and the remaining portion was centrifuged at 3000 rpm
for 15 min to obtain serum. Samples of liver and aortic tissue
were collected, soaked in ice-cold 0.9% NaCl, snap-frozen in
liquid nitrogen, and stored at �60 8C until use.
Calcium (Ca) concentration in the serum was determined by
a colorimetric test (Calcium liquicolor, HUMAN Gesellschaft fur
Biochemica und Diagnostica mbH, Germany). Serum albumin
levels were measured using a colorimetric test (Albumin
liquicolor, HUMAN Gesellschaft fur Biochemica und Diagnos-
tica mbH, Germany). Serum Ca levels were corrected for
albumin concentrations using the following formula: corrected
Ca = serum Ca + 0.02 � (40 � serum albumin) [18].
Serum 25-hydroxyvitamin D [25(OH)D] levels were mea-
sured using a radioimmune assay (25-Hydroxyvitamin D, 125I
Ria Kit, Diasorin Corporation, USA).
2.4. Oxidative stress markers
Liver samples were homogenised in ice-cold 0.9% NaCl
solution and centrifuged at 15,000 � g for 10 min at 4 8C.
The insoluble pellets were discarded and the supernatants
were used for analysis. Total peroxide concentrations (TPX)
were measured with an OxyStat colorimetric assay kit
(Biomedica Gruppe, Austria), which assesses the colour

d i a b e t e s r e s e a r c h a n d c l i n i c a l p r a c t i c e 1 0 0 ( 2 0 1 3 ) 2 4 3 – 2 4 9 245
change produced by the reaction of total peroxides in the
sample with peroxidase. The data were expressed as nmol/
mg protein, with protein concentration measured spectro-
photometrically using the Lowry method [19].
Total antioxidant capacity (TAC) was measured in the
serum and liver samples using a colorimetric assay (Randox
Laboratories Ltd., UK). This assay is based on the decolorisa-
tion of 2,20-azinobis(3-ethylbenzothiazoline 6-sulfonate)
(ABTS) radical action. The TAC value was expressed as an
equivalent of the millimolar concentration of Trolox (a soluble
vitamin E analogue) solution. Oxidative stress index (OSI), an
indicator of the redox balance between oxidation and
antioxidation, was expressed as the percent ratio of TPX to
TAC [20].
Aortic strips (10 mm) were homogenised in ice-cold 0.9%
NaCl solution, using a blade type homogeniser (Tekmar
Tissumizer, Cincinnati, OH, USA) and centrifuged at
10,000 � g for 10 min at 4 8C. The insoluble pellets were
discarded and the supernatants were used for analysis. The
concentration of CML was assessed using a commercial
enzyme-linked immunosorbent (ELISA) test OxiSelectTM N-
epsilon-(Carboxymethyl) Lysine ELISA Kit (Cell Biolabs, Inc.,
San Diego, CA, USA). Ten mg/ml of total protein extracts were
adsorbed onto a 96-well plate at 4 8C overnight. The absor-
bance of each well was read using a microplate reader (Tecan
Sunrise, Tecan GmbH, Austria) using 450 nm as the primary
wavelength. The concentration of CML was calculated by
comparison to a standard curve consisting of known con-
centrations of CML-BSA. Results are presented as nanogram of
CML per millilitre of solution.
2.5. Immunohistochemistry
Three-mm thick paraffin sections mounted on poly-L-lysine
coated SuperFrost slides (Menzel-Glaser, Germany) were
deparaffinised and rehydrated. Peroxidase activity was
blocked by 0.6% H2O2 (Merck, Germany) in methanol (Merck,
Germany). Then the sections were washed in tap water and in
PBS (pH = 7.4; Gibco, Invitrogen, USA) for 10 min, treated with
normal 1.5% goat serum (Gibco, Invitrogen Corporation, USA)
for 20 min at room temperature and incubated with the first
antibody: anti-carboxymethyl lysine (mouse monoclonal
antibody [CML26], abcam, UK) diluted 1:50 overnight at 4 8C
in the humidity chamber. On the next day the sections were
incubated with the biotinylated horse anti-mouse secondary
Table 1 – Body weights and serum laboratory values at the en
Group Body weight(g)
Blood glucose(mmol/L)
HbA1c(%)
Control 456 � 26 6.3 � 1.6 4.0 � 0.1
Diabetes 374 � 52** 28.3 � 3.9** 10.3 � 0.7**
Diabetes +
vitamin D
348 � 43** 28.5 � 5.9** 9.5 � 1.3**
Data are expressed as mean � SD (n = 8 per group).
HbA1c, glycated haemoglobin; 25(OH)D, 25-hydroxyvitamin D; Corrected
using the following formula: corrected calcium = serum calcium + 0.02 �* p < 0.05 vs control.** p < 0.01 vs control.*** p < 0.001 vs diabetes + vitamin D.
antibody for 30 min at room temperature. After a wash step
the sections were incubated with the avidin-biotin peroxidase
complex ELITE system (Vectastain Elite ABC Kit, Vector
Laboratories Inc., Burlingame, USA) for 30 min. Peroxidatic
activity was detected with 3,30-diaminobenzidine [DAB]
(Vector Laboratories Inc., USA) and the sections were counter-
stained with hemalaun, dehydrated and mounted with DPX
(Fluka, Switzerland). The labelling was expressed on a semi-
quantitative scale ranging from 0 to 4 (0 – no staining, 1 – weak
staining, 2 – moderate staining, 3 – strong staining, 4 – very
strong staining). Two independent observers in a blinded
fashion performed the evaluation. Immunohistochemistry
negative controls were performed by omitting the primary
antibody (mouse IgG was used in place of the primary
antibody).
2.6. Statistical analysis
Results are expressed as means � standard deviation (SD).
Differences between the groups were evaluated using the one-
way analysis of variance (ANOVA) followed by Tukey’s post
hoc analysis for multiple comparisons of group means. Semi-
quantitative data were compared by the Kruskal–Wallis one-
way ANOVA followed by Mann–Whitney U test. Differences
were considered to be statistically significant when p was
<0.05. All statistical comparisons were performed with the
Statistica software (version 8; StatSoft, USA).
3. Results
The basic parameters and biochemical results are presented
in Table 1. Rats in both diabetic groups presented with
significantly lower body weights and increased levels of
blood glucose and HbA1c. Serum 25(OH)D levels were
significantly decreased in untreated diabetic rats and
completely restored by vitamin D supplementation. Serum
Ca and albumin-corrected Ca levels were not different
between all groups, indicating that calcium homeostasis
was not affected by short-term changes in circulating
vitamin D levels.
Untreated diabetes produced significantly higher levels of
CML in the aortic wall homogenates and OSI in the liver (Figs. 1
and 2). The levels of serum TAC were significantly decreased in
untreated diabetic rats compared to non-diabetic rats (Fig. 3).
d of the study.
25(OH)D(nmol/L)
Calcium(mmol/L)
Albumin(g/L)
Corrected calcium(mmol/L)
140 � 21*** 2.6 � 0.2 39.3 � 2.7 2.6 � 0.3
108 � 38*,*** 2.5 � 0.2 30.8 � 3.2** 2.6 � 0.3
494 � 125 2.6 � 0.3 33.4 � 5.1** 2.7 � 0.2
calcium, calcium concentrations adjusted to albumin concentrations
(40 � serum albumin).
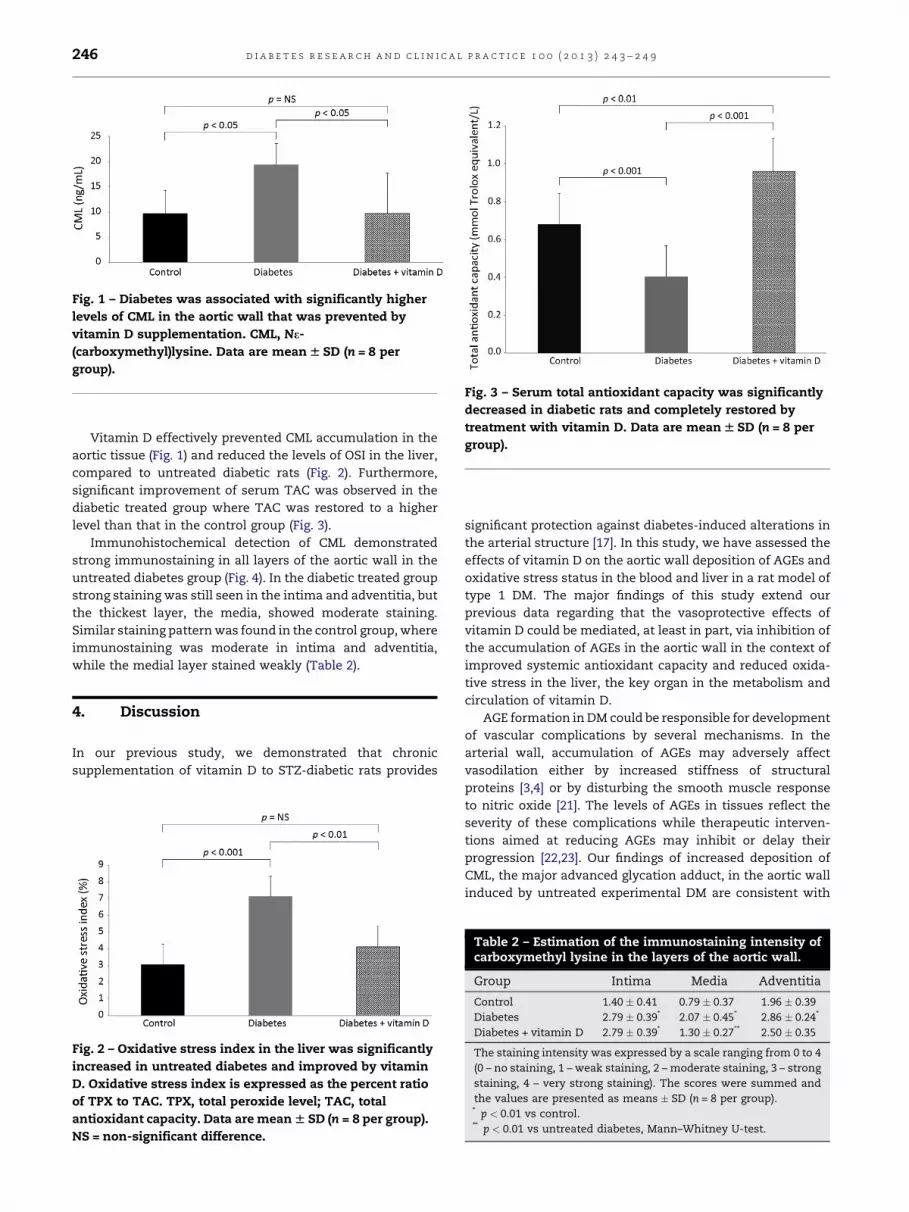
Fig. 1 – Diabetes was associated with significantly higher
levels of CML in the aortic wall that was prevented by
vitamin D supplementation. CML, Ne-
(carboxymethyl)lysine. Data are mean W SD (n = 8 per
group).
Fig. 3 – Serum total antioxidant capacity was significantly
decreased in diabetic rats and completely restored by
treatment with vitamin D. Data are mean W SD (n = 8 per
group).
d i a b e t e s r e s e a r c h a n d c l i n i c a l p r a c t i c e 1 0 0 ( 2 0 1 3 ) 2 4 3 – 2 4 9246
Vitamin D effectively prevented CML accumulation in the
aortic tissue (Fig. 1) and reduced the levels of OSI in the liver,
compared to untreated diabetic rats (Fig. 2). Furthermore,
significant improvement of serum TAC was observed in the
diabetic treated group where TAC was restored to a higher
level than that in the control group (Fig. 3).
Immunohistochemical detection of CML demonstrated
strong immunostaining in all layers of the aortic wall in the
untreated diabetes group (Fig. 4). In the diabetic treated group
strong staining was still seen in the intima and adventitia, but
the thickest layer, the media, showed moderate staining.
Similar staining pattern was found in the control group, where
immunostaining was moderate in intima and adventitia,
while the medial layer stained weakly (Table 2).
4. Discussion
In our previous study, we demonstrated that chronic
supplementation of vitamin D to STZ-diabetic rats provides
Fig. 2 – Oxidative stress index in the liver was significantly
increased in untreated diabetes and improved by vitamin
D. Oxidative stress index is expressed as the percent ratio
of TPX to TAC. TPX, total peroxide level; TAC, total
antioxidant capacity. Data are mean W SD (n = 8 per group).
NS = non-significant difference.
significant protection against diabetes-induced alterations in
the arterial structure [17]. In this study, we have assessed the
effects of vitamin D on the aortic wall deposition of AGEs and
oxidative stress status in the blood and liver in a rat model of
type 1 DM. The major findings of this study extend our
previous data regarding that the vasoprotective effects of
vitamin D could be mediated, at least in part, via inhibition of
the accumulation of AGEs in the aortic wall in the context of
improved systemic antioxidant capacity and reduced oxida-
tive stress in the liver, the key organ in the metabolism and
circulation of vitamin D.
AGE formation in DM could be responsible for development
of vascular complications by several mechanisms. In the
arterial wall, accumulation of AGEs may adversely affect
vasodilation either by increased stiffness of structural
proteins [3,4] or by disturbing the smooth muscle response
to nitric oxide [21]. The levels of AGEs in tissues reflect the
severity of these complications while therapeutic interven-
tions aimed at reducing AGEs may inhibit or delay their
progression [22,23]. Our findings of increased deposition of
CML, the major advanced glycation adduct, in the aortic wall
induced by untreated experimental DM are consistent with
Table 2 – Estimation of the immunostaining intensity ofcarboxymethyl lysine in the layers of the aortic wall.
Group Intima Media Adventitia
Control 1.40 � 0.41 0.79 � 0.37 1.96 � 0.39
Diabetes 2.79 � 0.39* 2.07 � 0.45* 2.86 � 0.24*
Diabetes + vitamin D 2.79 � 0.39* 1.30 � 0.27** 2.50 � 0.35
The staining intensity was expressed by a scale ranging from 0 to 4
(0 – no staining, 1 – weak staining, 2 – moderate staining, 3 – strong
staining, 4 – very strong staining). The scores were summed and
the values are presented as means � SD (n = 8 per group).* p < 0.01 vs control.** p < 0.01 vs untreated diabetes, Mann–Whitney U-test.
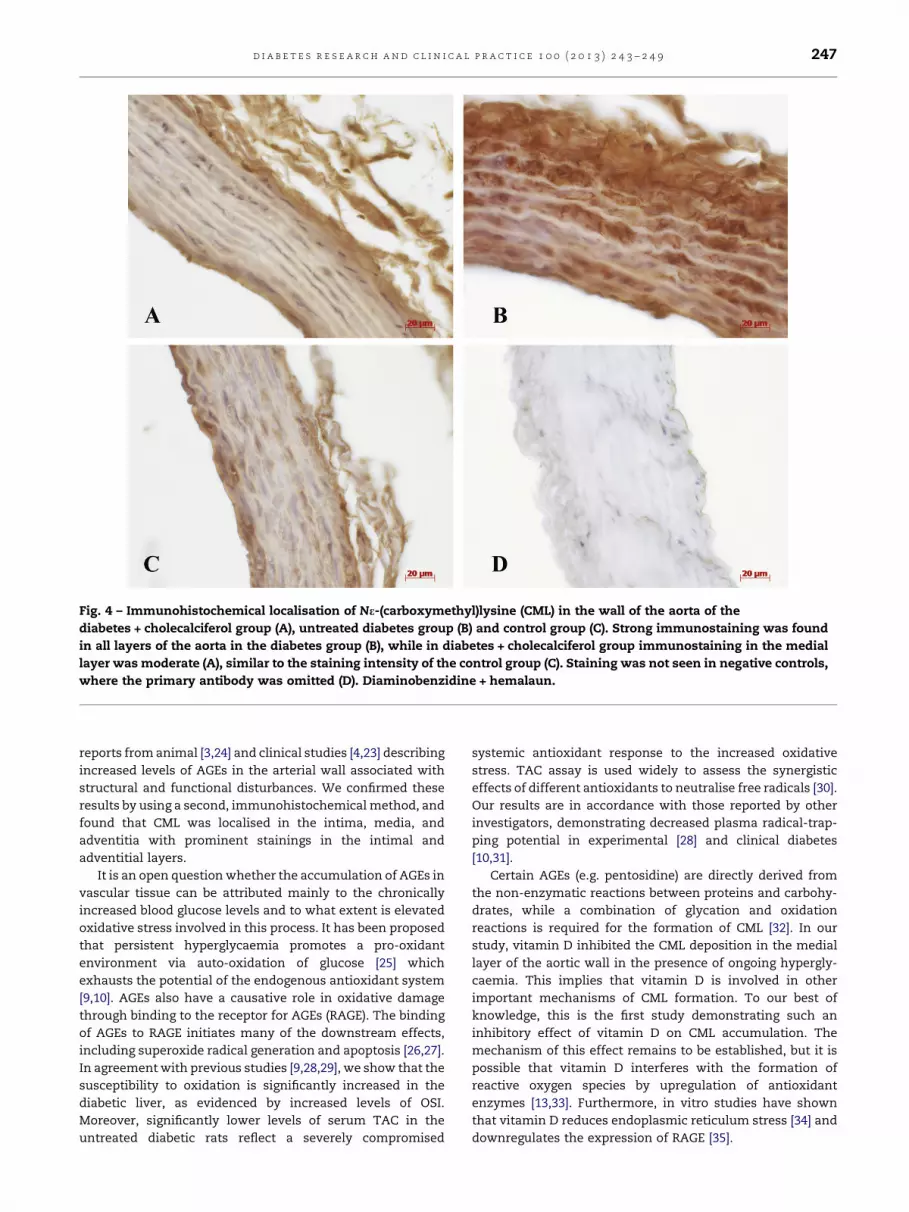
Fig. 4 – Immunohistochemical localisation of Ne-(carboxymethyl)lysine (CML) in the wall of the aorta of the
diabetes + cholecalciferol group (A), untreated diabetes group (B) and control group (C). Strong immunostaining was found
in all layers of the aorta in the diabetes group (B), while in diabetes + cholecalciferol group immunostaining in the medial
layer was moderate (A), similar to the staining intensity of the control group (C). Staining was not seen in negative controls,
where the primary antibody was omitted (D). Diaminobenzidine + hemalaun.
d i a b e t e s r e s e a r c h a n d c l i n i c a l p r a c t i c e 1 0 0 ( 2 0 1 3 ) 2 4 3 – 2 4 9 247
reports from animal [3,24] and clinical studies [4,23] describing
increased levels of AGEs in the arterial wall associated with
structural and functional disturbances. We confirmed these
results by using a second, immunohistochemical method, and
found that CML was localised in the intima, media, and
adventitia with prominent stainings in the intimal and
adventitial layers.
It is an open question whether the accumulation of AGEs in
vascular tissue can be attributed mainly to the chronically
increased blood glucose levels and to what extent is elevated
oxidative stress involved in this process. It has been proposed
that persistent hyperglycaemia promotes a pro-oxidant
environment via auto-oxidation of glucose [25] which
exhausts the potential of the endogenous antioxidant system
[9,10]. AGEs also have a causative role in oxidative damage
through binding to the receptor for AGEs (RAGE). The binding
of AGEs to RAGE initiates many of the downstream effects,
including superoxide radical generation and apoptosis [26,27].
In agreement with previous studies [9,28,29], we show that the
susceptibility to oxidation is significantly increased in the
diabetic liver, as evidenced by increased levels of OSI.
Moreover, significantly lower levels of serum TAC in the
untreated diabetic rats reflect a severely compromised
systemic antioxidant response to the increased oxidative
stress. TAC assay is used widely to assess the synergistic
effects of different antioxidants to neutralise free radicals [30].
Our results are in accordance with those reported by other
investigators, demonstrating decreased plasma radical-trap-
ping potential in experimental [28] and clinical diabetes
[10,31].
Certain AGEs (e.g. pentosidine) are directly derived from
the non-enzymatic reactions between proteins and carbohy-
drates, while a combination of glycation and oxidation
reactions is required for the formation of CML [32]. In our
study, vitamin D inhibited the CML deposition in the medial
layer of the aortic wall in the presence of ongoing hypergly-
caemia. This implies that vitamin D is involved in other
important mechanisms of CML formation. To our best of
knowledge, this is the first study demonstrating such an
inhibitory effect of vitamin D on CML accumulation. The
mechanism of this effect remains to be established, but it is
possible that vitamin D interferes with the formation of
reactive oxygen species by upregulation of antioxidant
enzymes [13,33]. Furthermore, in vitro studies have shown
that vitamin D reduces endoplasmic reticulum stress [34] and
downregulates the expression of RAGE [35].

d i a b e t e s r e s e a r c h a n d c l i n i c a l p r a c t i c e 1 0 0 ( 2 0 1 3 ) 2 4 3 – 2 4 9248
The liver is central to the vitamin D metabolism and
circulation in that liver converts absorbed vitamin D into
25(OH)D, the circulating form vitamin D, which is the best
indicator of vitamin D status in the organism [36]. Thus, we
aimed to assess the impact of vitamin D pooled in the liver on
the increased oxidative stress, as seen in the diabetic rats. We
found that supplementation of vitamin D was able to reduce
OSI to the level that of normal rats, clearly demonstrating the
antioxidative potential of vitamin D. These results are in line
with those by Hamden et al. [13] who showed that oxidative
stress in the diabetic liver may be improved by calcitriol, the
active form of vitamin D. Moreover, we found that treatment
of diabetic rats with vitamin D restored the serum TAC to a
level that was significantly higher than that found in control
rats. Detailed explanation to the antioxidative effects of
vitamin D cannot be provided, but these may include
stabilisation of the plasma membrane against lipid peroxida-
tion [12] or upregulation of antioxidant systems, including
GSH, GSH peroxidase, and superoxide dismutase, via its
nuclear receptors [13,33].
Our findings underscore the importance of oxidative and
antioxidative status in the development of diabetic vascular
complications. Indeed, among diabetic populations, a consid-
erable variance in the rates of AGE accumulation, despite
similar blood glucose and HbA1c levels, has been demonstrat-
ed which may be attributed to individual variations in
oxidative stress status [6]. Furthermore, antioxidants are
known to decrease CML formation [37] which, again, empha-
sises the essential role of oxidative stress in the advanced
glycation processes.
In conclusion, the findings of the present study indicate
that vitamin D may have a role in reducing different
diabetes-related complications. Particularly, we demon-
strate that treatment with vitamin D reduces the accumula-
tion of AGEs in the medial layer of the aortic wall and
oxidative stress at a systemic and end organ level. Thus, our
results support the understanding that vitamin D supple-
mentation provides an important protection from the
oxidative damage associated with the development of
diabetic vascular complications.
Conflict of interest
The authors declare that they have no conflict of interest.
Acknowledgement
This study was supported by the Estonian Science Foundation
grants Nos. 9094 and 8273 and by Target Financing (Nos.
0180001s07 and 0180012s11) by the European Union through
the European Regional Development Fund.
r e f e r e n c e s
[1] Brownlee M. Glycation products and the pathogenesis ofdiabetic complications. Diabetes Care 1992;15:1835–43.
[2] Meng J, Sakata N, Takebayashi S, Asano T, Futata T, ArakiN, et al. Advanced glycation end products of the Maillardreaction in aortic pepsin-insoluble and pepsin-solublecollagen from diabetic rats. Diabetes 1996;45:1037–43.
[3] Winlove CP, Parker KH, Avery NC, Bailey AJ. Interactions ofelastin and aorta with sugars in vitro and their effects onbiochemical and physical properties. Diabetologia1996;39:1131–9.
[4] Airaksinen KE, Salmela PI, Linnaluoto MK, Ikaheimo MJ,Ahola K, Ryhanen LJ. Diminished arterial elasticity indiabetes: association with fluorescent advancedglycosylation end products in collagen. Cardiovasc Res1993;27:942–5.
[5] Reddy S, Bichler J, Wells-Knecht KJ, Thorpe SR, Baynes JW.Ne-(carboxymethyl)lysine is a dominant advancedglycation end product (AGE) antigen in tissue proteins.Biochemistry 1995;34:10872–8.
[6] McCance DR, Dyer DG, Dunn JA, Bailie KE, Thorpe SR,Baynes JW, et al. Maillard reaction products and theirrelation to complications in insulin-dependent diabetesmellitus. J Clin Invest 1993;91:2470–8.
[7] Dyer DG, Dunn JA, Thorpe SR, Bailie KE, Lyons TJ, McCanceDR, et al. Accumulation of Maillard reaction products inskin collagen in diabetes and aging. J Clin Invest1993;91:2463–9.
[8] Ceriello A, Taboga C, Tonutti L, Quagliaro L, Piconi L, Bais B,et al. Evidence for an independent and cumulative effect ofpostprandial hypertriglyceridemia and hyperglycemia onendothelial dysfunction and oxidative stress generation:effects of short- and long-term simvastatin treatment.Circulation 2002;106:1211–8.
[9] Kakkar R, Mantha SV, Radhi J, Prasad K, Kalra J. Increasedoxidative stress in rat liver and pancreas duringprogression of streptozotocin-induced diabetes. Clin Sci1998;94:623–32.
[10] Tsai EC, Hirsch IB, Brunzell JD, Chait A. Reduced plasmaperoxyl radical trapping capacity and increasedsusceptibility of LDL to oxidation in poorly controlledIDDM. Diabetes 1994;43:1010–4.
[11] Baynes JW. Role of oxidative stress in development ofcomplications in diabetes. Diabetes 1991;40:405–12.
[12] Smith PR, Thornalley PJ. Mechanism of the degradation ofnon-enzymatically glycated proteins under physiologicalconditions. Studies with the model fructosamine, Ne-(1-deoxy-D-fructos-1-yl)hippuryl-lysine. Eur J Biochem1992;210:729–39.
[13] Hamden K, Carreau S, Jamoussi K, Miladi S, Lajmi S,Aloulou D, et al. 1a,25 dihydroxyvitamin D3: therapeuticand preventive effects against oxidative stress, hepatic,pancreatic and renal injury in alloxan-induced diabetes inrats. J Nutr Sci Vitaminol 2009;55:215–22.
[14] Wiseman H. Vitamin D is a membrane antioxidant, abilityto inhibit iron-dependent lipid peroxidation in liposomescompared to cholesterol, ergosterol and tamoxifen andrelevance to anticancer action. FEBS Lett 1993;326:285–8.
[15] Sardar S, Chakraborty A, Chatterjee M. Comparativeeffectiveness of vitamin D3 and dietary vitamin E onperoxidation of lipids and enzymes of the hepaticantioxidant system in Sprague–Dawley rats. Int J VitamNutr Res 1996;66:39–45.
[16] Dong J, Wong SL, Lau CW, Lee HK, Ng CF, Zhang L, et al.Calcitriol protects renovascular function in hypertensionby down-regulating angiotensin II type 1 receptors andreducing oxidative stress. Eur Heart J 2012;33:2980–90.
[17] Salum E, Kampus P, Zilmer M, Eha J, Butlin M, Avolio AP,et al. Effect of vitamin D on aortic remodeling instreptozotocin-induced diabetes. Cardiovasc Diabetol2012;11:58.

d i a b e t e s r e s e a r c h a n d c l i n i c a l p r a c t i c e 1 0 0 ( 2 0 1 3 ) 2 4 3 – 2 4 9 249
[18] Gardner MD, Dryburgh FJ, Fyffe JA, Jenkins AS. Predictivevalue of derived calcium figures based on the measurementof ionised calcium. Ann Clin Biochem 1981;18:106–9.
[19] Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Proteinmeasurement with the Folin phenol reagent. J Biol Chem1951;193:265–75.
[20] Harma M, Harma M, Erel O. Increased oxidative stress inpatients with hydatidiform mole. Swiss Med Wkly2003;133:563–6.
[21] Bucala R, Tracey KJ, Cerami A. Advanced glycosylationproducts quench nitric oxide and mediate defectiveendothelium-dependent vasodilatation in experimentaldiabetes. J Clin Invest 1991;87:432–8.
[22] Ono Y, Aoki S, Ohnishi K, Yasuda T, Kawano K, Tsukada Y.Increased serum levels of advanced glycation end-productsand diabetic complications. Diabetes Res Clin Pract1998;41:131–7.
[23] Monnier VM. Intervention against the Maillard reactionin vivo. Arch Biochem Biophys 2003;419:1–15.
[24] Soulis T, Thallas V, Youssef S, Gilbert RE, McWilliam BG,Murray-McIntosh RP, et al. Advanced glycation endproducts and their receptors co-localise in rat organssusceptible to diabetic microvascular injury. Diabetologia1997;40:619–28.
[25] Wolff SP. Diabetes mellitus and free radicals, Free radicals,transition metals and oxidative stress in the aetiology ofdiabetes mellitus and complications. Br Med Bull1993;49:642–52.
[26] Vincent AM, Perrone L, Sullivan KA, Backus C, Sastry AM,Lastoskie C, et al. Receptor for advanced glycation endproducts activation injures primary sensory neurons viaoxidative stress. Endocrinology 2007;148:548–58.
[27] Nitti M, d’Abramo C, Traverso N, Verzola D, Garibotto G,Poggi A, et al. Central role of PKCdelta in glycoxidation-dependent apoptosis of human neurons. Free Radic BiolMed 2005;38:846–56.
[28] Feillet-Coudray C, Rock E, Coudray C, Grzelkowska K,Azais-Braesco V, Dardevet D, et al. Lipid peroxidation andantioxidant status in experimental diabetes. Clin ChimActa 1999;284:31–43.
[29] Sun F, Iwaguchi K, Shudo R, Nagaki Y, Tanaka K, Ikeda K,et al. Change in tissue concentrations of lipidhydroperoxides, vitamin C and vitamin E in rats withstreptozotocin-induced diabetes. Clin Sci 1999;96:185–90.
[30] Valkonen M, Kuusi T. Spectrophotometric assay for totalperoxyl radical-trapping antioxidant potential in humanserum. J Lipid Res 1997;38:823–33.
[31] Santini SA, Marra G, Giardina B, Cotroneo P, Mordente A,Martorana GE, et al. Defective plasma antioxidant defensesand enhanced susceptibility to lipid peroxidation inuncomplicated IDDM. Diabetes 1997;46:1853–8.
[32] Fu MX, Requena JR, Jenkins AJ, Lyons TJ, Baynes JW, ThorpeSR. The advanced glycation end product. Ne-(carboxymethyl)lysine, is a product of both lipidperoxidation and glycoxidation reactions. J Biol Chem1996;271:9982–6.
[33] Garcion E, Sindji L, Leblondel G, Brachet P, Darcy F. 1,25-dihydroxyvitamin D3 regulates the synthesis of gamma-glutamyl transpeptidase and glutathione levels in ratprimary astrocytes. J Neurochem 1999;73:859–66.
[34] Riek AE, Oh J, Sprague JE, Timpson A, de Las Fuentes L,Bernal-Mizrachi L, et al. Vitamin D suppression ofendoplasmic reticulum stress promotes an antiatherogenicmonocyte/macrophage phenotype in type 2 diabeticpatients. J Biol Chem 2012;46:38482–94.
[35] Talmor Y, Golan E, Benchetrit S, Bernheim J, Klein O, GreenJ, et al. Calcitriol blunts the deleterious impact of advancedglycation end products on endothelial cells. Am J PhysiolRenal Physiol 2008;294:1059–64.
[36] Wagner CL, Greer FR, American Academy of PediatricsSection on Breastfeeding, American Academy of PediatricsCommittee on Nutrition. Prevention of rickets and vitaminD deficiency in infants, children, and adolescents.Pediatrics 2008;122:1142–52.
[37] Fu MX, Wells-Knecht KJ, Blackledge JA, Lyons TJ, Thorpe SR,Baynes JW. Glycation, glycoxidation, and cross-linking ofcollagen by glucose. Kinetics, mechanisms, and inhibitionof late stages of the Maillard reaction. Diabetes 1994;43:676–83.