Embed Size (px)
Citation preview

Developmental Brain Research, 24 (1986) 21-29 21 Elsevier
BRD 50305
Visual Behavior of Monocularly Deprived Kittens Treated with 6-Hydroxydopamine
BARBARA GORDON, JEFFREY MORAN*, PAUL TROMBLEY and JENNIFER SOYKE
Department of Psychology and Institute of Neuroscience, University of Oregon, Eugene, OR 97403 (U. S. A.)
(Accepted June 4th, 1985)
Key words: visual development - - visual behavior - - monocular deprivation - - 6-hydroxydopamine
Several investigators have reported that treating the visual cortex with 6-hydroxydopamine (6-OHDA) preserves the ability of a monocularly deprived eye to drive cells in the visual cortex. If 6-OHDA provides useful protection from the effects of monocular de- privation, it should also prevent the behavioral blindness that normally accompanies monocular deprivation. To test this prediction we compared the visual behavior of monocularly deprived kittens pretreated with 6-OHDA with that of kittens similarly deprived, but not drug-treated. Kittens were trained on a visual discrimination task before drug treatment or suture. Starting at about 5 weeks of age the kittens were given 6-OHDA via ventricular cannula, given vehicle solution, or given no treatment at all. At about 6 weeks of age all kittens were monocularly deprived for one week. When the deprived eye was opened at 7 weeks of age, most kittens not receiving 6- OHDA were blind when tested with the deprived eye. In contrast, none of the kittens receiving 6-OHDA intraventricularly were blind when tested with the deprived eye. 6-OHDA had no effect on performance with the non-deprived eye. We conclude that 6-OHDA protects vision through the monocularly deprived eye without impairing vision through the non-deprived eye.
INTRODUCTION
Modern work on plasticity in the mammal ian visu-
al system began with Wiesel and Hubel ' s paper in
1963 on monocular deprivation31: if one eye of a kit-
ten is deprived of vision for more than a few days dur-
ing the first weeks of life, subsequent st imulation of
that eye fails to activate cells in the pr imary visual
cortex. Al though the environmenta l condit ions con-
trolling this ocular dominance shift have subsequent-
ly been described in some detail (for reviews see
refs. 24 and 28), little research has been directed to-
wards the underlying cellular mechanisms until re-
cently.
In 1979, Kasamatsu and colleagues~7,18 found
that monocular depr ivat ion did not cause an ocular
dominance shift if the visual cortex was t rea ted with
6-hydroxydopamine ( 6 - O H D A ) , a neurotoxin that
selectively destroys catecholaminergic cells a n d
terminals5.6. They also found that exogenous norepi-
nephrine (NE) could prevent the effects of 6-
O H D A t6-18. They suggested that NE acts as a neuro-
modula tor controll ing plasticity and that destroying
NE terminals prevents the plastic changes that nor-
mally result from monocular deprivat ion.
Kasamatsu and colleagues ' results and interpreta-
tions have been subject to several criticisms. Two of
these criticisms provided the motivat ion for the cur-
rent study. First, al though some of these findings
have been replicated2.3.s, failures to replicate have
also been repor ted 1. Second, 6 - O H D A itself may
damage geniculocort ical or intracort ical connec-
tions. In this case it might preserve responses from
the deprived eye not by el iminating plasticity but by
eliminating the binocular compet i t ion that normally
accompanies monocular deprivat ion. Unde r some
conditions el iminating binocular compet i t ion by enu-
cleating the exper ienced eye greatly a t tenuates the
effects of monocular deprivat ion 4.20.
In order to address these criticisms we measured
the visual acuity of monocular ly depr ived kittens
treated with 6 - O H D A . We chose to measure acuity
because several groups of investigators have found
that the acuity of a depr ived eye is roughly related to
* Present address: Laboratory of Neuropsychology, NIMH, Bethesda, MD 20205, U.S.A. Correspondence: B. Gordon, Department of Psychology, University of Oregon, Eugene, OR 97403, U.S.A.
0165-3806/86/$03.50 O 1986 Elsevier Science Publishers B.V. (Biomedical Division)

its ability to drive cells in area 1715-21,3°. Finding that a
monocularly deprived animal t rea ted with 6 - O H D A
had good acuity in its depr ived eve would provide
corroborat ion for the results of Kasamatsu and col-
leagues using a method very different from theirs, A
negative result would not, of course, prove Kasamat-
su and colleagues wrong because some cells in the
primary visual cortex may retain input from the de-
prived eye and yet be unable to influence motor cen-
ters used in performing the visual task.
If 6 - O H D A damages geniculocort ical or intracor-
tical connect ions serving both eyes. it should reduce
the quality of vision through both eyes These deficits
might be detectable by measures of visual acuity,
even though abnormal cell responses have not been
repor ted in area 17 of monocular ly depr ived kit tens
t rea ted with 6-OHDA 2,~,~,~- ~. On the other h and. if
6 - O H D A provides genuine protect ion from the ef-
fects of monocular deprivat ion, it should prevent the
blindness that normal ly accompanies monocular dep-
rivation t2,23,:4,2<27 without decreasing the acuity of
the non-depr ived eye.
MATERIALS AND METHODS
Behavioral apparatus and procedure
Eighteen kittens were tested in an appara tus simi-
lar to that descr ibed by Mitchell et al. 22. The appara-
tus consisted of a small box and a pla t form The box
stood a short distance (13.5-20 cm~ above the plat-
form. The pla t form was divided into two sides by a
low part i t ion. The floor of each side of the pla t form
was actually a t rapdoor . Both t rapdoors were nor-
mally kept closed and locked.
A photograph of a square-wave grating (light and
dark phases = 40 cd/m 2 and 6.8 cd/m 2. respectively)
was placed on one t rapdoor . The other t r apdoor con-
ta ined a gray photograph of luminance equivalent to
the mean of the grating (23 cd/m2}. The posi t ions of
the two photographs could be interchanged.
The ki t ten 's task was to jump out of the box and
onto the side of the pla t form that held the grating.
Correct responses were rewarded with a taste of
baby food; incorrect responses ( jumping to the grayl
were punished with a loud " N o ! " from the experi-
menter . The kittens ate their rewards avidly, even
during and after 6 - O H D A t rea tment . Each kitten
first learned this discrimination with the t r apdoor
open on the side containing the uniform gray stimu-
lus. The other side of the pla t form contained a very
low spatial f requency grating (0. t l cycles/degree~.
Af ter learning this simple d iscr iminauon, training
was continued with both doors closed and locked. All
kittens rapidly learned the discrimination with both
t rapdoors closed: we could then es thnate the kittens'
acuity by increasing the spatial f requency of the grat-
ing until the kittens could no longer discriminate be-
tween stripes and the uniform gray.
After learning the discriminatinn, kittens were
trained 7 days a week. 0 .5 -2 h/da~, They were de-
prived of food by taking them awax from their moth-
ers or removing food from their cages for a few hours
before training or tesnng.
Every session began with an easily discr iminated
grating. If the kitten scored 5 out of the first 7 trials
correct, the frequency of the grating was increased
by 1/3 octave and 7 more trials were run. If the kit ten
gave fewer than 5 correct responses, the spatial fre- quency was decreased by l/3 octave and 7 more trials
were run. On the days when an acmty measurement
was required I see Schedule of exper iments , be lowl
this procedure was repea ted until 2i trials were col-
lected at two grating frequencies t/3 octave apart
such that the kitten gave 15 or more correct re-
sponses at the lower f requency and fewer than 15 cor-
rect responses at the higher frequency. The mean of
these two frequencies was defined as the ki t ten 's acu-
ity for that session, The probabi l i ty of 15 correct re-
sponses occurring by chance is less than 0.05 {binomi-
al test}. On days when no acuitx measure was re-
quired, each kit ten was t rained for 0 . 5 - 2 h in order to
maintain its familiari ty with the task,
When testing an eye that had been monocular ly
deprived, testing began with the t r apdoor which he ld
the gray photograph p ropped open about 45 ° and
with the lowest spatial frequency {0.11 cvcles/de-
greel on the other (closed} t rapdoor . If the kit ten
scored 5 out of 7 trials correct , the t r apdoor holding
the gray stimulus was closed and the sequence of tri-
als continued as descr ibed above. If the kit ten failed
to score 5 trials correct , the exper iment cont inued
with the t r apdoor open and the (I.11 cycles/degree
grating. If a kit ten failed to achieve 15 out of 2t trials
correct with the t rapdoor open. we considered it
blind Distinguishing the open t r apdoor was an easier
discrimination than distinguishing the lowest fre-

quency from a uniform gray. This ' trapdoor open'
test allowed the kitten to demonstrate some visual ability when its acuity could not be measured.
In order to decrease the likelihood that the kittens
would develop a preference for jumping to one par- ticular side, the positions of the grating and the gray photographs were interchanged pseudorandomly; i.e., the stimulus positions were random except that
the photographs were left in one position for no more than two trials in a row. In the event that a kitten did develop a side preference, as evidenced by 3 consec- utive errors to one side, several trials were given in
which the grating always appeared on the side ig- nored by the kitten. These trials did not contribute to
the acuity measure.
Groups The experimental group received 6-OHDA. The
control group did not. The right eye of every kitten in both groups was sutured shut at about 43 days of age.
The left eye was untouched. The number of animals in each group at each point of the experiment is given in Table 1.
Ventricular injection group (n = 7). Animals in this group received injections of 6-OHDA via ventricular cannulas. Cannulas were constructed of 19 mm lengths of 20-gauge hypodermic stock fitted with a removable cap and implanted in the left lateral ven-
tricle under ketamine (30 mg/kg) and acepromazine (4 mg/kg) anesthesia. The cannula was placed stereo- taxically at AP +11.0, L 3.0 and its tip was lowered
TABLE I
Number of animals in each group tested for acuity at each stage of the experiment
6-OHDA yen- Controls tricular cannula
Initial binocular testing 7 5 Non-deprived eye prior to
opening deprived eye 7 11 Non-deprived eye after deprived
eye measures 7 11 Deprived eye, first day of testing 7 11 Deprived eye, second day of
testing 7 10' Both eyes occluded 7 10"
* One animal was in a 'dark' cage with a light leak. We used the deprived eye data collected on the day of eye opening, but discarded the remainder of the data from this animal.
23
11-11.5 mm below the surface of the brain. The can- nula was cemented to the skull with dental acrylic. Prior to applying the acrylic, the skull was coated with cyanoacrylate glue to increase adhesion of the acrylic.
Controls (n = 11). All kittens in this group re- ceived right eye suture. Three animals received no other surgery or treatment. Two animals were im- planted with 2 ml osmotic minipumps containing only vehicle Solution (0.4% ascorbic acid in physiological
saline). Six animals were implanted with ventricular cannulas. These animals received injections on the same schedule as the 6-OHDA ventricular injection group, but received vehicle solution rather than 6- OHDA. The results from all control animals were similar and have been combined.
Statistical analyses All statistical tests comparing the groups on their
performance with the deprived eye were non-para-
metric because we could not assign a number to an
animal's performance if it was blind or if it could per- form only the trapdoor-open discrimination. We made 3 types of comparisons. First, we used the Mann-Whitney U-test (MWU) to find which groups were different 29. This test uses rank order. We as- signed the lowest ranks to those animals which were
blind and assigned the next higher rank to those ani- mals which could only perform the discrimination with the trapdoor open. Second, we used Fisher's ex- act probability test (FEP) 29 to find out whether the
groups differed in the proportion of animals that were blind. Third, we used the sign test 29 to deter- mine whether the animals performed better with their deprived or with their non-deprived eyes.
Schedule of experiments A typical experimental schedule for one kitten is
shown in Table II. For all kittens, training was begun
at 28 days of age. By 33-37 days of age all kittens had learned the discrimination. At this time we deter- mined the binocular acuity of 12 animals (7 experi- mental and 5 control animals); 6 animals were not tested at this time. All animals were then assigned to either the 6-OHDA or the control group, the appro- priate surgery performed, and drug administration begun. At 42-44 days of age, the right eye of each animal was sutured under ketamine anesthesia (35

24
TABLE 11
Typical experimental schedule. animal V. C. 3
Age Event ~daysl
10 Eyes open 28 Begin training on task 33 Animal can perform task; implant cannula; begin
6-OHDA administration: continue training 42 Suture right eye; continue training 44 Discontinue 6-OHDA administration:
continue training 48 Acuity measure of left, non-deprived eye (start with
easily discriminated grating) 49 Reopen sutured, right eye; acuity measure of right,
deprived eye (start with trapdoor open): place in dark room
50 Acuity measure of right, deprived eye (start with trapdoor open); place in dark room
51 Acuity measure of left, non-deprived eye (start with easily discriminated grating); place in dark room
52 'Acuity measure' with both eyes occluded
mg/kg). Exper imenta l animals received at least 83%
of their total dose of 6 - O H D A pr ior to suture.
The acuity of the left. unsutured eye was deter-
mined 6 days after suture, the day before the right,
sutured eye was reopened.
The sutured eye of 9 animals (2 ventr icular injec-
tion, 7 controls) was opened under ke tamine anes-
thesia. Vision through this eye was tested the follow-
ing day. In 9 animals (5 ventr icular inject ion, 4 con-
trols) the right, sutured eye was opened under halo-
thane anesthesia and vision through this eye was test-
ed half an hour later. There were no differences be-
tween the results obta ined with these two procedures
and the results have been combined. To minimize re-
covery after the depr ived eye was opened , all ani-
mals lived in l ightproof cages, coming out only for
testing.
To assess changes in vision over t ime and variabili-
ty in per formance , the acuity of the depr ived eye was
retested on the day after its initial test. Then, on the
next day, the acuity of the non-depr ived eye was test-
ed a second time. Finally, to be certain that occlusion
of the untested eye was adequate , we tested the kit-
tens with both eyes occluded.
Drug doses and schedules Two animals received a total of l0 mg of 6 - O H D A
(16 mg/ml, in 0.1% ascorbic acid in Ringer ' s solu-
tion) over a 7-day per iod following the adminis tra-
tion schedule descr ibed by Kasamatsu and Petti-
grewl7 Because this dose made the animals eat poor-
ly and lose weight, the remaming 5 animals received
a total of 4.8 mg 6 - O H D A (16 mg/ml, in 0. lq~ ascor-
bic acid in Ringer 's) in 6 equal doses over a 6-day pe-
riod. These la t ter animals were considerably health-
ier. The behavioral results were similar regardless of
total 6 - O H D A dose.
Assays o f drug effectiveness
For the animals receiving 6 - O H D A . the presence
of side effects (e.g. urination, defecat ion, circling,
hissing) indicated that the drug had reached the ven-
tricles. In order to get a quant i tauve est imate of de-
pletion of NE we used high-performance liquid chro-
matography with electrochemical detect ion (HPL( ' -
EC) to measure the amount of NE in the cortices of 2
control and 5 ventr icular injection animals (4.8 mg
total dose J. Samples were taken from frontal , parie-
tal. and occipital cortex on both sides of the midl ine
as well as from the lateral geniculate nuclei.
We used the procedure descr ibed by Cassone el
al, v except that our mobile phase consisted ol 0.1 M
sodium acelate . 0,02 M citric acid monohydra te . |00
mg/l octvl sodium sulfate. 50 mg/1 E D T A , 10('4- meth-
anol.
RESULI'S
Bilateral distribution o f 6 - O H D A in cortex
We have two pieces of evidence that 6 - O H D A in-
jected unilateral ly into a lateral ventricle effects cor-
tex in both hemispheres equally, First. at the end ol
the exper iment . 0.1 cc ot radio opaque dye (sodium
diatr izoate , Winthrop) was injected into the cannulas
of several kittens and an X-ray taken immediate ly
thereafter . These X-rays showed that the dye dif-
fused throughout the ventricles on both sides of the
brain. Second. injecnon of 6 - O H D A into the left lat-
eral ventricle depletes monoamines to similar levels
in the cortex of both hemispheres (Trombley , Allen,
Sovke, Blaha. Lane and Gordon . J. Neurosci.. in
press).
Initial acuity measure Prior to any drugs or surgeries, a subset of the ant~
reals (7 exper imenta l and 5 control ~ was tes ted binoc-
ularly and found able to perform the acuity task. The
o
1.0
0 .5
0 D
P r e - d r u g
D = d r u g C = c o n t r o l
/ / / / / A
VIllA <ILIA '/Ill.z4
D C
P o s t - d r u g , P r e - e y e o p e n i n g
l
19 C
P o s t - e y e o p e n i n g
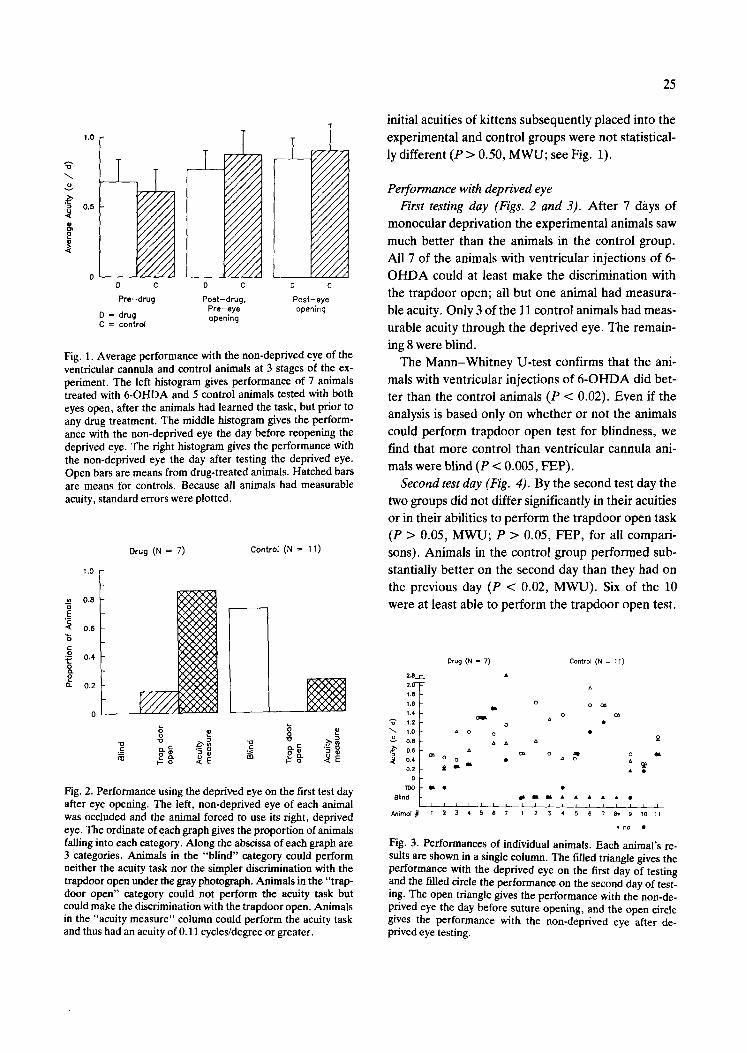
Fig. 1, Average performance with the non-deprived eye of the ventricular cannula and control animals at 3 stages of the ex- periment. The left histogram gives performance of 7 animals treated with 6-OHDA and 5 control animals tested with both eyes open, after the animals had learned the task, but prior to any drug treatment. The middle histogram gives the perform- ance with the non-deprived eye the day before reopening the deprived eye. The right histogram gives the performance with the non-deprived eye the day after testing the deprived eye. Open bars are means from drug-treated animals. Hatched bars are means for controls. Because all animals had measurable acuity, standard errors were plotted.
Drug (N = 7) Control (N = 11)
1.0
E
g
2
0 . 8
0 .6
0 . 4
0 .2
0
7
C¢¢C~
, ~ O / C X X
~Ox 'XXX x~CxQ, C'X K ~ X X
Fig. 2. Performance using the deprived eye on the first test day after eye opening. The left, non-deprived eye of each animal was occluded and the animal forced to use its right, deprived eye. The ordinate of each graph gives the proportion of animals failing into each category. Along the abscissa of each graph are 3 categories. Animals in the "blind" category could perform neither the acuity task nor the simpler discrimination with the trapdoor open under the gray photograph. Animals in the "trap- door open" category could not perform the acuity task but could make the discrimination with the trapdoor open. Animals in the "acuity measure" column could perform the acuity task and thus had an acuity of 0.11 cycles/degree or greater.
25
initial acuities of kittens subsequently placed into the experimental and control groups were not statistical- ly different (P > 0.50, MWU; see Fig. 1).
Performance with deprived eye First testing day (Figs. 2 and 3). After 7 days of
monocular deprivation the experimental animals saw much better than the animals in the control group. All 7 of the animals with ventrieular injections of 6- OHDA could at least make the discrimination with the trapdoor open; all but one animal had measura- ble acuity. Only 3 of the 11 control animals had meas- urable acuity through the deprived eye. The remain- ing 8 were blind.
The Mann-Whitney U-test confirms that the ani- mals with ventricular injections of 6-OHDA did bet- ter than the control animals (P < 0.02). Even if the analysis is based only on whether or not the animals could perform trapdoor open test for blindness, we find that more control than ventricular cannula ani- mals were blind (P < 0.005, FEP).
Second test day (Fig. 4). By the second test day the two groups did not differ significantly in their acuities or in their abilities to perform the trapdoor open task (P > 0.05, MWU; P > 0.05, FEP, for all compari- sons). Animals in the control group performed sub- stantially better on the second day than they had on the previous day (P < 0.02, MWU). Six of the 10 were at least able to perform the trapdoor open test.
2.8_E_ 2.0- 1.8 1.6 1,4
%" 1.2
"~ 1.0 o 0.8 ~" o.s "5
0.4 0.2
0 TOO
8lind
Animol
Drug (N = 7) Control (N = ~1)
o D
o
o o
o ~
I I I i i i i ~ 1 i a I I 2 3 4 5 6 7 1 2 3 4 5
A
o o~
o C~
o ~x
6 7 8. 9 10 111
* no •
Fig. 3. Performances of individual animals. Each animal's re- sults are shown in a single column. The filled triangle gives the performance with the deprived eye on the first day of testing and the filled circle the performance on the second day of test- ing. The open triangle gives the performance with the non-de- prived eye the day before suture opening, and the open circle gives the performance with the non-deprived eye after de- prived eye testing.

26
Drug (N = 7 ) C o n t r o l (N = 10)
1.0
0.8 o E
~ 0.6 "d g
~: o.4
P ~- 0.2
0
( X X ~ X ~
"o
~-o < E
Fig. 4. Performance through the deprived eye on second testing day. Conventions as in Fig. 2.
We presume that this improvement resulted from the
visual experience they obtained in the first testing
day.
All animals with ventricular injection of 6-OHDA
continued to make the trapdoor open discrimination.
Five of the 7 animals in this group had measurable
acuity (Figs. 3 and 4). Although the 6-OHDA ani-
mals appeared to perform slightly worse on the sec-
ond day than the first, their performance was not sta-
tistically different on the two days (P > 0.4, MWU).
Furthermore, even the poorer performance of the 6-
OHDA animals on the second day of testing was bet- ter than the performance of the control animals on
the first day (P < 0.02. MWU).
Performance with non-deprived eye When using the non-deprived eye, all animals
were able to perform the acuity task and their acui-
ttes did not deteriorate from their pretreatment lev-
els (Figs. I and 3). If there is any trend at all, the ani-
mals' acuity with the non-deprived eye seems to im-
prove with age. There was no difference, on any day of testing, between the acuities of the two groups
when using this eye (P > 0.05. MWU).
Comparison between deprived and non-deprived eye There was no difference between the performance
of the drug-treated animals with the deprived eye and their performance with the non-deprived eye (P > 0.05, sign test). In contrast, animals in the control group showed evidence of visual deprivation: i.e.. they performed more poorly with the deprived eye
the day it was opened than they had with the non-de-
prived eye on the previous day (P < 0.01, sign test).
Performance with both eyes covered With both eyes occluded, none of the animals
could even detect the open trapdoor.
Assays of drug effectiveness All the animals receiving 6-OHDA via ventricular
injection showed side effects (loud meowing, cir-
cling, hiding in corners, etc.). The average concen-
tration of NE in two undepleted control brains was 81
_+ 14 ng/g per tissue. In 5 animals receiving 6- O H D A via ventricular injection, the average con-
centration was 8.3 + 2.7 ng of NE/g per tissue. When
each of the animals with ventricular injection was
compared to the mean normal concentration, deple-
tion varied from 81 to 98%. Fig. 5 shows sample
HPLC records.
DISCUSSION
Our results indicate that intraventricular 6-OHDA
largely prevents the behaviorally measured blindness
that usually results from monocular deprivation. Im-
mediately after reopening the deprived eye, animals
©
n ~
A mlr~
B
Je !
l ~ j
0 5 n&
4 r r t l n
Fig. 5. Sample HPLC records. Arrowspoint to the NE peak. A: record from a kitten depleted by intraventricular 6-OHDA. The peak represents 4.7 ng/g of NE. B: record from a contro t kitten. Note that the sensitivity of themaehine is only one-fifth that in record A. The peak represents 62 ng/g of NE. The amount of NE in the sample cannot be calculated directly from the height of the peaks. The final calculations must take ac- count of the peaks produced by standards and the weight of the tissue in the sampte.

treated with 6-OHDA perform as well with this eye as they do with the non-deprived eye. In contrast,
control animals perform much more poorly with the deprived than with the non-deprived eye.
Alternative explanations Because all animals were blind when tested with
both eyes occluded, it is unlikely that the occlusion of the untested eye was inadequate or that the kittens had developed an effective strategy to win at our pseudorandom task. Because all animals underwent similar surgeries for reopening the deprived eye, it is unlikely that control animals performed poorly sim- ply because they did not feel well after surgery.
We do not think that 6-OHDA causes non-specific brain damage that results in a decrease in plasticity. Using the deprived eye, the animals receiving 6- OHDA actually performed better than the controls; it is difficult to explain how a toxic reaction would ac- tually improve performance. Furthermore, when using the non-deprived eye, performance was similar for 6-OHDA animals and controls. These results are not consistent with the view that the effects are due to non-specific brain damage but are consistent with the view that 6-OHDA prevents plasticity 17,18.
Similarly, we do not think that 6-OHDA prevents the effects of monocular deprivation by creating a pseudo-binocular deprivation that prevents the de- velopment of competition between synapses serving the deprived eye and synapses serving the non-de- iprived eye. Although we could find no behavioral studies of short-term binocular suture or dark-rear- ing, animals dark-reared for several months are blind when their eyes are first opened 30. Dark-rearing for only 3-6 days causes a marked decrease in the re- sponsiveness of the visual cortex 14. These results pre- dict that if 6-OHDA causes an effect akin to binocu- lar deprivation, it should decrease the acuity of both eyes. Instead, we found that both the deprived and non-deprived eye acuities of kittens treated with 6- OHDA were similar before and after treatment. In contrast, the performance of the control animals was extremely poor on the first day the deprived eye was tested. Because our animals never performed as well as those studied by Mitchell 22 (see below) and were somewhat variable from day to day, we cannot rule out the possibility that the 6-OHDA animals had small decreases in acuity that we did not detect.
27
Comparison with other measures of acuity Our measured acuities are lower than those of
Mitchell.et al. 22. At 5 weeks of age his animals could discriminate 1.2 cycles/degree while our controls could only discriminate 0.62 cycles/degree. By 7 weeks his animals could discriminate 2.5 cycles/de- gree while our controls could still only discriminate 0.78. On the other hand, Elberger 13 has also meas- ured the acuity of young kittens and obtained results similar to ours. At about 7 weeks of age her kittens' acuity averaged slightly more than 0.5 cycles/degree. We do not know how to account for these differences among laboratories.
Comparison with physiology Several groups of investigators have provided sup-
port for the assumption that the acuity of an eye roughly reflects the number of cortical cells driven by that eye 15,2~,30. Therefore, it is fair to say that our be- havioral results are qualitatively in agreement with the physiological results of Kasamatsu and collea- gues 16117 and in disagreement with those of Adrien et al.l. Like Kasamatsu and colleagues, we find that the visual system is less responsive to monocular depriva- tion if the brain is treated with 6-OHDA. They re- ported that the ocular dominance histograms from monocularly deprived animals treated with 6-OHDA were close to normal. We found that the performance of the drug-treated animals was better than the per- formance of the controls on the first day of deprived eye testing. In addition, the deprived eye perform- ance on the first day of testing was not statistically different from non-deprived eye performance on the previous day.
On the other hand, it is probably not possible to make detailed quantitative comparisons between acuity and ocular dominance histograms. The acuity of the deprived eye may be controlled, at least par- tially, by the spatial frequency sensitivity of the cells driven by that eye.
Our results do not provide information about the anatomic site where 6-OHDA protects visual acuity. Because intraventricular injection allows the drug to work diffusely throughout the brain, we are unable to pinpoint the critical site or sites of action.
Our results suggest that 6-OHDA does, indeed, decrease plasticity. Our finding that 6-OHDA also depletes NE does not, however, prove that NE is

necessary for plasticity. The best ev idence for this in-
t e rp re ta t ion comes f rom Kasamat su and col leagues"
demons t r a t ion that exogenous ly suppl ied N E re-
s tored plasticity to a 6 - O H D A t rea ted cor tex Is,l~.
Never the less , 3 l abora to r i e s have cha l lenged the
view that N E is necessary and suff icient for plasticity:
( l ) Daw et al. dep l e t ed cor t ical N E by cut t ing the
dorsal no rad rene rg ic bundle or by t rea t ing animals
with N - ( 2 - c h t o r o e t h y l ) - N - e t h y l - 2 - b r o m o b e n z y l a m i n e
(DSP-4) . Ne i the r of these man ipu la t ions dec reased
plast ici tyg-l l ; (2) B e a r and Danie l s 2 and Bear
et al.3 t r ea ted neona ta l ki t tens wi th in t r ape r i tonea l
6 - O H D A . This p r o c e d u r e p r o d u c e d last ing N E
deple t ion but fai led to p ro tec t the p r imary visual cor-
tex against the effects of m o n o c u l a r depr iva t ion ; (3)
Shaw and C y n a d e r ~5 have p rov ided ev idence that the
presence of N E is not suff ic ient for plasticity. They
ob ta ined a decrease in plast ici ty by in jec t ing gluta-
mate , not 6 - O H D A . They suggest that any disrup-
REFERENCES
1 Adrien, J., Buisseret, P.. Fregnac, Y., Gary-Bobo, E., hn- bert, M., Tassin, J.-P. and Trotter, Y., Noradrenaline et plasticit6 du cortex visuel du chaton: un reexamen, C.R. Acad. Sci., Ser. D., 295 (1982) 745-750,
2 Bear. M.F. and Daniels, J.D., The plastic response to mon- ocular deprivation persists in kitten visual cortex after chronic depletion of norepinephrine, J. Neurosci., 3 (1983) 407-416.
3 Bear, M.F.. Paradiso, M.A,. Schwartz, M., Nelson, S.B.. Carnes, K,M. and Daniels, J.D., Two methods of catechol- amine depletion in kitten visual cortex yield different ef- fects on plasticity, Nature (Londont, 302 (1983) 245-247.
4 Blakemore, C. and Hawken. M.J., Rapid restoration ol functional input to the visual cortex of the cat after brief monocular deprivation. J. Physiol. (London), 327 (1982) 463-487.
5 Breese, G.R. and Cooper, B.R., Chemical lesioning: cate- cholamine pathways. In R.D. Myers lEd.), Methods in Psychobiology, Academic Press. New York, 1977, pp. 27-48.
6 Breese, G.R. and Traylor, l .D , , Depletion of brain nor- adrenaline and dopamine by 6-hydroxydopamine, Br. ,1, Pharmacol., 42 (1971) 88-99.
7 Cassonc, V.M., Lane, R.F. and Menaker, M., Daily rhythms of serotonin metabolism in the medial hypothal a- mus of the chicken: effects of pinealectomy and exogenous melatonin, Brain Res., 289 (1983) 129-134.
8 Daw, N.W., Rader, R.K., Robertson. T.W. and Ariel, M., The effects of 6-hydroxydopamine on visual deprivation in the kitten striate cortex, J. Neurosci., 3 (1983)907-914.
9 Daw, N,W.. Robertson, T.W., Rader, R.W. and Videen, Y.O., Effect of lesions of the dorsal adrenergic bundle on plasticity in the visual cortex, lnvest. Ophthalmol. (Vis. Sci., Suppl., 24 (1983) 140.
10 Daw, N.W., Robertson, T.W. , Rader, R.W., Videen,
tion of the normal s ignal - to-noise ra t io in the cor tex
can decrease plasticity. All of these results suggest
that 6 - O H D A probab ly does not exer t its effect via
NE deple t ion . This conc lus ion does not. howeve r ,
imply that the effect of 6 - O H D A is ~ 'non-specif ic" . It
does suggest, though , that a t t empts to unders tand
the cel lular basis of 6 - O H D A ' s act ion will have to
search for some addi t iona l m e c h a n i s m
ACKNOWLEDGEMENTS
We thank Rose Lane , Vincen t Cassone and
Char les Blaha for teaching us to do H P L C . W e t h a n k
Alvse Rail, M a t t h e w Har r i son Leah Tufane l l i ,
Mark Met t ler . Ma lco lm Manness and Steve Bylsma
for test ing some of the animals . I 'his research was
suppor ted by N I H G r a m 5 RO1 EY04050. J . M was
suppor ted by N I H Tra in ing Gran t G M 07257.
I .O and ('oscia. C.J. Substantial reduction oI cortical noradrenaline by lesions of the dorsal noradrenergic path- way does not prevent effects of monocular deprivation, J Neurosci. 4 (1984) 1354-1360.
17 Daw. N.W.. Videen. T.O., Robertson. T.W. and Rader K.. An evaluation of the hypothesis that noradrenaline af- fects plasticity in the developing visual cortex. In A. Fine led.) , The Visual System. Symposium to Honor E.F. Mac- Nichol. A.R. Liss, Philadelphia, 1985, pp. 133-144.
12 Dews. P.B, and Wiesel T.N.. Consequence of monocular de privation on visual behaviour in kittens. J Physiol. ,'Lon- don j, 206 (1970) 437-455.
13 Elberger. A.J., The corpus callosmn is a cmical factor fo~ developing maximum visual acmtv Dev. Brain Res. 5 11982~ 350-352.
14 Freeman. R.D.. Mallach. R and ttartley. S,. Responsivlty of normal kitten striate cortex deteriorates after brief bin- ocular deprivation, J. Neuroph.vsiol.. 45 (1981~ 1074-1084
15 Giffin, F. and Mitchell, D.E.. The rate of recovery of vision after early monocular deprivation in kittens, J. Physiol. cLondom 274 (1978] 511-537.
16 Kasamatsu. T.. Neuronal plasticity maintained by the cen- tral norepinephrine system in the cat visual cortex. In J.M Sprague and A.N. Epstein (Eds. ~, Progress in Psychobiolo- gy and Physiological Psychology, Vol. 10. Academic Press, New York. [983, pp. 1-111.
17 Kasamatsu T. and Pettigrew. J.D.. Preservation of binocu- larity after monocular deprivation in the striate cortex ol kittens treated with 6-hydroxydopamine, J. Comp. Neu-
rol. 5119791139-162 18 Kasamatsu. T.. Pettigrew. J.D. anti Ary. M., Restorauon
of visual cortical plasticity by local microperfusion of nor- epmephrine. J. Comp. Neurol. 185 (1979) 163-182,.
19 Kasamatsu. T., Pettigrew, J.D. and Ary. M.. Cortical rc- coverv from effects of monocular deprivation: acceleranon with norepinephrine and suppressmn with 6-hydroxydopa- mine J. Neurophysiol., 45 119811 254-266

20 Kratz, K.E. and Spear, P.D., Post-critical period reversal of effects of monocular deprivation in the striate cortex in cats, Neurophysiology, 39 (1976) 501-511.
21 Mitchell, D.E., Cynader, M. and Movshon, J.A., Recov- ery from the effects of monocular deprivation, J. Comp. Neurol., 176 (1977) 53-63.
22 Mitchell, D.E., Giffin, F., Wilkinson, F., Anderson, P. and Smith, M.L., Visual resolution in young kittens, Vision Res., 16 (1976) 363-366.
23 Movshon, J.A., Reversal of the behavioural effects of mo- nocular deprivation in the kitten, J. Physiol. (London), 261 (1976) 175-187.
24 Movshon, J.A. and Van Sluyters, R.C., Visual neural de- velopment, Ann. Rev. Psychol., 32 (1981) 477-522.
25 Shaw, C. and Cynader, M., Disruption of cortical activity prevents ocular dominance changes in monocularly de- prived kittens, Nature (London), 308 (1984) 731-734.
29
26 Sherman, S.M., Visual field defects in monocularly and binocularly deprived cats, Brain Res., 49 (1973) 25-45.
27 Sherman, S.M., Permanence of visual perimetry deficits in monocularly and binocularly deprived cats, Brain Res., 73 (1974) 491-501.
28 Sherman, S.M. and Spear, P.D., Organization of visual pathways in normal and visually deprived cats, Physiol. Rev., 62 (1982) 738-855.
29 Siegel, S., Nonparametric Statistics for the Behavioral Sci- ences, McGraw-Hill, New York, 1956.
30 Timney, B., Mitchell, D.E. and Cynader, M., Behavioral evidence of prolonged sensitivity to effects of monocular deprivation in dark-reared cats, J. Neurophysiol., 43 (1980) 1041-1054.
31 Wiesel, T.N. and Hubel, D.H., Single cell responses in striate cortex of kittens deprived of vision in one eye, J. Neurophysiol., 26 (1963) 1003-1007.