Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA DEPARTAMENTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA
VARIAÇÃO NA ESTRUTURA DA MADEIRA DE QUATRO ESPÉCIES DA CAATINGA NORDESTINA E SEU
POTENCIAL PARA O DESENVOLVIMENTO SUSTENTÁVEL
LAZARO BENEDITO DA SILVA
Feira de Santana, Bahia 2006

UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA DEPARTAMENTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA
VARIAÇÃO NA ESTRUTURA DA MADEIRA DE QUATRO ESPÉCIES DA CAATINGA NORDESTINA E SEU POTENCIAL PARA O
DESNVOLVIMENTO SUSTENTÁVEL
LAZARO BENEDITO DA SILVA
Tese apresentada ao Programa de Pós-Graduação em Botânica da Universidade Estadual de Feira de Santana, como parte dos requisitos necessários para obtenção do título de Doutor em Botânica.
Prof. Dr. Francisco de Assis R. dos Santos (UEFS) Orientador
Dr. Peter Erick Gasson (RBG/Kew)
Co-Orientador
Feira de Santana, Bahia 2006

BANCA EXAMINADORA
Prof. Dr. Francisco de Assis Ribeiro dos Santos (UEFS) Orientador e Presidente da Banca
Prof. Dr. Mário Tomazello-Filho (ESALQ/USP) Membro Titular
Prof. Dr. José Elias de Paula (UnB) Membro Titular
Profa. Dra. Ana Maria Giulietti-Harley (UEFS) Membro Titular
Profa. Dra. Cláudia Elena Carneiro (UEFS) Membro Titular
Tese defendida e aprovada em 03 de agosto de 2006

Ao único que é digno de receber a honra e a glória a força e o poder, ao Deus eterno imortal invisível mas real, a Ele todo o meu agradecimento.
A minha querida esposa, minha fiel amiga. Carmen, muito obrigado por ter cruzado o meu caminho e torná-lo, com sua presença, ele tão maravilhoso. Eu te amo!

“O sonho que você sonhou
ficou no coração
foi Deus quem colocou.
Ele quer muito lhe ajudar
e se você quiser
a força vai lhe dar.
Então prossiga pelo seu ideal
e va lutando pra alcançá-lo afinal
Tudo é possível
pois esse Deus de amor
move as montanhas
se preciso for”.
(Ronaldo Arco)

AGRADECIMENTOS
Àquele que deu a Sua própria vida por amor. Obrigado Jesus por tudo que tens feito
em minha vida e pela realização de mais este sonho.
Àquela que sempre me deu apoio geral e irrestrito e será eternamente lembrada dentro
do meu coração, e que espero vê-la novamente, para dar-lhe um grande abraço e dizer-lhe
muito obrigado, minha mãe Germana Aragão (in memorian). Você foi à mãe que todo filho
quer ter.
Ao Prof. Dr. Francisco de Assis Ribeiro dos Santos pela sua capacidade de trabalho e
objetividade e pela orientação e amizade dispensada.
Ao Dr. Peter Gasson pela co-orientação e apoio no Jodrell Laboratory.
Ao Departamento de Botânica do Instituto de Biologia da Universidade Federal da
Bahia que permitiu o meu afastamento para a realização do Doutorado.
À Universidade Estadual de Feira de Santana e ao Programa de Pós-Graduação em
Botânica por me receber como aluno.
À Comissão de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pelo apoio
financeiro, através do Programa Plano de Qualificação Institucional (PQI).
Ao Jodrell Laboratory do Royal Botanic Gardens pelo apoio dispensado no período
em que lá estive desenvolvendo parte do trabalho de laboratório.
À Kew Latin America Research Fellowships Programme (KLARF) e The
Clothworkers Guild. (através do Fuelwood project) pelo apoio financeiro no período em que
estive na Inglaterra no Jodrell Laboratory.
À Associação Plantas do Nordeste (APNE) pelo apoio logístico nas coletas em campo
e disponibilidade das amostras das madeiras do Projeto Madeiras.
Ao Núcleo de Tecnologia da Preservação e da Restauração (NTPR) da Escola
Politécnica da UFBA (NTPR), especialmente ao Prof. Dr. Mário Mendonça de Oliveira e ao
Sr. Allard Monteiro do Amaral, pelo auxílio nas análises de densidade.
Ao Laboratório de Anatomia de Madeiras e Dendrocronologia da Escola Superior
Luiz de Queiroz (ESALQ) por nos receber durante um mês para realização de análises de
amostras de madeira.
Ao Laboratório de Anatomia Vegetal do Instituto de Biociências da Universidade de
São Paulo (USP) onde preparamos algumas lâminas histológicas.
Ao Instituto de Pesquisa Tecnológica (IPT), especialmente a Antônio Carlos Barbosa,
pela forma amiga que me recebeu e pela ajuda na produção de algumas lâminas histológicas.

Ao Prof. Dr. José Pereira de Sousa com quem aprendi a ter respeito, dedicação e
honestidade pela Ciência, e me ensinou os primeiros passos a seguir no estudo da Anatomia
de Madeira.
Ao Dr. David Cutler pela ajuda na definição de caminhos a serem seguidos no
trabalho de tese
À Profa. Dra. Ana Maria Giulietti Harley pelo incentivo e apoio para a realização
deste trabalho.
Ao Curador do Herbário HUEFS, Prof. Dr. Luciano Paganucci, pela identificação da
Mimosa ophthalmocentra Mart. ex Benth.
Às Biólogas do HUEFS Teonildes Nunes, Maria José Sampaio e Elaine Miranda pela
forma sempre gentil em me atender quando solicitadas.
À Maísa Sales da PRPPG/UFBA pela dedicação, competência e apoio sempre presente
quando solicitados.
À Amélia e Ligia Baracat, Dr. Simon Mayo e família, e Dr. Raymond Harley pela
amizade e carinho dispensada em nossa estada no Royal Botanic Gardens.
A Frans Pareyn, Fernando Galindo, Jair Virgínio e toda equipe da Associação Plantas
do Nordeste (APNE) pelo apoio logístico nas coletas em campo e disponibilidade das
amostras das madeiras.
À Dra. Rita de Cássia Pereira e Dra. Maria Bernadete Silva do Herbário do IPA .
Ao Dr. Mário Tomazello Filho, Dr.Cláudio Lisi, Bióloga Maria Bermúdez e equipe do
Laboratório de Anatomia de Madeiras e Dendrocronologia da ESALQ/USP, pela
receptividade e dedicação a mim dispensada.
À Profa. MSc. Camila Pigozzo, Prof. Dr. Pedro Jacinto pelo apoio na estatística.
À Dra. Ana Maria Giulietti Harley pelo incentivo e apoio para a realização deste
trabalho.
Ao Prof. Dr. José Pereira de Sousa que me ensinou a ter dedicação e apreço pela
Ciência, especialmente pela anatomia de madeira.
À amiga Profa. MSc. Maria Lenise Guedes do IBIO/UFBA, que me direcionou para a
Anatomia de Madeiras.
À amiga Profa. MSc. Cláudia Dias de Santana que não poupou esforços na elaboração
do Projeto PQI.
Ao Prof. Dr. Gregório Ceccantini, Profa. Dra. Verônica Angyalossy, MSc. Érica
Amano e toda equipe por me receber, sempre tão bem, no Laboratório de Anatomia.
Aos amigos do LAMIV/UEFS pela demonstração de amizade verdadeira: Biólogo
MSc. Paulino Oliveira, Bióloga MSc. Kelly Leite, Profa. Dra. Cláudia Carneiro, Profa. MSc.

Marileide Saba, Profa. MSc. Luciene Lima, Prof. MSc. Danovan Novaes, Prof. MSc. Hilder
Magalhães, Profa. MSc. Bárbara Araújo, Biólogo Marcos Dórea, Profa. MSc. Rita de Cássia
Araújo, Profa. MSc. Rita Dória, Jailson Novais e Biólogo Ricardo Landin.
À Adriana Estrela e Gardênia Aires pelo apoio na Secretaria do PPGBot./UEFS.
À Janaína Alves pelas dicas das Mimosa e aos demais colegas do PPGBot./UEFS.
À Gisele Rocha pelo apoio total na obtenção das imagens no MEV (DCBio/UEFS).
Aos amigos da Igreja Adventista do Sétimo Dia do Cabula e Caminho das Árvores de
Salvador-Bahia e da IASD de expressão portuguesa em Londres, que muito oraram para que
este sonho se tornasse realidade.
Aos queridos Armando e Maria dos Mártires que nos acolheu com tanto carinho em
um momento tão especial em Londres, nosso muito obrigado.
A tia Margarida Aragão Andrade e sua linda família que me acolheu, na minha estada,
em Feira de Santana e me proporcionaram momentos tão felizes.
Aos meus irmãos Jorge Aragão, Antônio Carlos Santos, Valter Souza e irmãs Ivanice
Aragão, Jorge Aragão, Rosimeire Aragão, Jacira Aragão e Bárbara Cristina Sales e minhas
tias Nélia Silva e Nadinha Silva que sempre torceram pelo meu sucesso.
Às minhas mães queridas Antônia Anunciação, Conceição Sacramento, Iracema e
Gilca Macedo.
Aos meus demais amigos de longas jornadas Evandro Rego, Augusto Jorge Lima,
Gilberto Cafezeiro, Silvia Regina Pinheiro, Cláudia Leão, Isabel Santos, Rita Santos, Marta
Gonçalves, Nelsino e Guiomar Pastor, Álvaro Luiz Silva, Roque Alberto Silva, Abdias
Carvalho, Elzeni Gusmão, Juarez Jorge Santos, Hermínia Bastos e Ana Verena Madeira, que
aqui dispensei os títulos pela proximidade da amizade, aos quais reconheço como co-autores
da obra maior, a minha vida.
E finalmente, àqueles que de forma direta ou indireta contribuíram para a conclusão
desta obra, o meu sincero muito obrigado.

SUMÁRIO
AGRADECIMENTOS
RESUMO
ABSTRACT
INTRODUÇÃO ………………………………………………………………………...….. 1
CAPÍTULO 1. Anatomia e densidade básica da madeira de Croton sonderianus Müell.
Arg. (Euphorbiaceae), espécie da caatinga do Nordeste do Brasil ................. 8
CAPÍTULO 2. Influência da precipitação anual na formação dos anéis de crescimento de
Croton sonderianus Müell. Arg. (Euphorbiaceae) na caatinga do Nordeste
do Brasil .......................................................................................................... 30
CAPÍTULO 3. Anatomia e densidade básica da madeira de Caesalpinia pyramidalis
(Leguminosae - Caesalpinioideae), espécie endêmica da caatinga do
Nordeste do Brasil ......................................................................................... 61
CAPÍTULO 4. Estudo comparativo da madeira de Mimosa ophthalmocentra Mart. ex
Benth. e Mimosa tenuiflora (Willd.) Poir. (Leguminosae – Mimosoideae) 85
CONSIDERAÇÕES FINAIS …………………………………………………………...… 115

RESUMO

RESUMO
Este trabalho objetivou estudar a anatomia e a densidade básica da madeira de
Croton sonderianus Müell. Arg., Caesalpinia piramidalis Tul., Mimosa ophthalmocentra
Mart. ex Benth. e Mimosa tenuiflora (Willd.) Poir. utilizadas pelas comunidades locais
ocorrentes na caatinga de Pernambuco, nos municípios de Serra Talhada e Sertânia.
Descreveu-se a variação radial, sentido medula-câmbio, comparando-se a percentagem dos
elementos do lenho no tronco e nos galhos nos dois municípios e radialmente na própria
árvore, a fim de estabelecer o potencial total do lenho para produção de energia. Além disso,
objetivou-se também identificar caracteres anatômicos do lenho relacionando-os com o
ambiente da caatinga. O estudo anatômico e a determinação da densidade do lenho foram
realizados com amostras coletadas no tronco, ao nível do DAP e em dois galhos com
diferentes diâmetros. Para análise dos anéis de crescimento os métodos utilizados foram os da
densitometria de raios X e o das medidas dos anéis de crescimento através de mesa de
medição, conforme rotina de laboratório de dendrocronologia. A partir da aplicação de teste
estatístico para correlação entre a distância e a percentagem de células na análise da variação
radial do lenho das quatro espécies em estudo, pode-se concluir que existem diferenças
significativas na região próxima da medula em direção ao câmbio. Todas apresentaram grande
quantidade de celulose expressa na elevada densidade, no baixo percentual de parênquima e
alta freqüência de fibras possuindo, portanto, potencial para produção de carvão de alta
qualidade energética, desde a fase do lenho juvenil, visto que nesta região ocorre uma grande
concentração de fibras, vasos com menor diâmetro e menor quantidade de parênquima.
Assim, as áreas de caatinga estudadas nos municípios pernambucanos de Sertânia e Serra
Talhada, apresentam vegetação cujos indivíduos de Croton sonderianus, Caesalpinia
pyramidalis, Mimosa ophthalmocentra e M. tenuiflora produzem madeiras com potencial para
produção de álcool combustível, carvão vegetal e coque metalúrgico, conseqüentemente,
maior potencial energético para sustentar combustão duradoura. Croton sonderianus
apresentou a capacidade de formar em seu tronco, anéis de crescimento indicando seu grande
potencial para estudos dendrocronológicos nos trópicos. Anéis de crescimento mais largos são
formados durante os anos de maior disponibilidade de água e anéis mais estreitos, nos anos de
baixa precipitação considerados períodos de seca, com redução e/ou cessação da atividade do
câmbio (abaixo de 300 mm anual). Os galhos apresentaram potencial semelhante ao dos
troncos, expresso nas percentagens de fibras, vasos e parênquimas. Sendo assim, a utilização
dos mesmos pode ser útil porque evita o abate da árvore e facilita o manuseio. Observaram-se

diferenças marcantes entre as duas espécies de Mimosa, aqui estudadas. Mimosa
ophthalmocentra apresentou camadas de crescimento constituídas por linhas de parênquima
contendo cristais, parênquima escasso e menor percentagem de raios, enquanto a M.
tenuiflora apresentou camada de crescimento distinta, porém sem cristais, parênquima axial
vasicêntrico, em faixas e em confluência e maior percentagem de raios.
Palavras-chaves: Caesalpinia piramidalis, Croton sonderianus, Mimosa, lenha, anatomia de
madeira, caatinga, carvão, anéis de crescimento.

ABSTRACT

ABSTRACT
This research had the objective of studying the wood anatomy and basic density of Croton sonderianus Müell. Arg., Caesalpinia piramidalis Tul., Mimosa ophthalmocentra Mart. ex Benth. and Mimosa tenuiflora (Willd.) Poir., used caatinga of Pernambuco, Brazil, in the municipality of Serra Talhada and Sertânia. It was described the radial variation (from pith to bark), assessing the wood elements in the trunk and branches in both municipality and radially in the tree itself, in order to establish the total wood potential for energy production. Besides, it was the intention to identify wood anatomy characters relating them to the caatinga. Both the anatomic study and wood density determination were carried out with samples collected at DBH level and of two branches. In the analysis of the growth rings, the methods used were the X-ray densitometry and ring measurement by measurement table according to the dendrocronology laboratory routine. From the statistic test application in order to correlate the distance and percentage of cells, in the wood radial variation analysis of the four species being studied, it was possible to conclude that there are significant differences between the region closer to the pith towards the bark. They all presented a great amount of cellulose expressed in the high density and low parenchyma percentage, and the high frequency of fibers presenting. Therefore, the caatinga areas studied in the municipality of Sertânia and Serra Talhada (Pernambuco) present vegetation which individuals of Croton sonderianus, Caesalpinia pyramidalis, Mimosa ophtalmocentra and Mimosa tenuiflora produce wood with potentiality for the production of fuel alcohol, charcoal and metallurgic cock, and consequently, bigger energetic potential to provide enduring combustion. The Croton sonderianus presented the capability of forming growth rings in its trunk which indicated its great potential for dendrocronologic studies in the tropics. Larger rings are formed during the years with greater water abundance, and narrower rings in years of low rainfall considered drought periods, with reduction and/or ceasing of cambium activity (below 300 mm per year). The branches presented similar potentiality to the trunks, expressed in the percentage of fibers, vessels and parenchyma. Consequently, their use can be advisable since it avoids the killing of trees and facilitates handling. There were noticed significant differences between the two Mimosa studied. The Mimosa ophtalmocentra presented growth layers made of parenchyma lines containing crystals, scarce parenchyma and a smaller number of rays, whereas the Mimosa tenuiflora presented a distinct growth layer, but without crystals, axial parenchyma vessel centric, in zones and in confluence, and a larger percentage of rays. This study dealt with four species of northeaster caatinga will provide subside to governmental authorities to set environmental and conservations politcs.
Key Words: Caesalpinia piramidalis, Croton sonderianus, Mimosa, wood anatomy, wood densiy, caatinga, charcoal, growth ring.

INTRODUÇÃO

2
INTRODUÇÃO
A caatinga nordestina ocupa um espaço de aproximadamente um milhão de
quilômetros quadrados, compreendidos, segundo Lima (1996) e Rodal & Melo (1999) e
outros, pelo que se denomina o “Polígono das Secas”, correspondendo 54% da Região
Nordeste, estendendo-se do Estado do Piauí ao Norte do Estado de Minas Gerais. Apresenta
chuvas concentradas em um único período com duração de 3 a 5 meses, médias anuais de 400
a 800 mm e um regime pluvial com distribuição irregular no tempo e no espaço, tendo como
cobertura vegetal típica a caatinga que se caracteriza pela caducifolia da maioria das espécies.
A produção de lenha é a mais importante contribuição do extrativismo no Nordeste.
No Estado de Pernambuco são consumidos cerca de doze milhões de estéreos de lenha e
carvão anualmente, dos quais 70% são utilizados nos domicílios, demonstrando a importância
social e econômica desse recurso florestal (Sampaio 2002). Com enfoque conservacionista ou
da sustentabilidade das populações arbóreas da caatinga, as alternativas existentes relacionam-
se com a otimização do processo de produção e uso da biomassa, além da diminuição do
consumo de lenha e carvão vegetal. Como a atividade de reflorestamento tem limitações de
ordem climática e de solo, considera-se que o manejo florestal sustentado da caatinga é a
alternativa mais indicada para o Nordeste, permitindo o uso múltiplo dos recursos florestais,
com custos reduzidos e sua imediata exploração, além de preservar as características
ecológicas do meio ambiente. Para a aplicação de métodos de manejo florestal para fins
energéticos, como a produção de lenha comum nas comunidades rurais do Nordeste, são
necessárias análises da anatomia e das características físico-químicas das madeiras das
espécies florestais. De forma geral, determinam-se as dimensões e percentagem das fibras e
parâmetros de qualidade. Porém, segundo Sárkáni et al. (1957) e Nguyen (1977), outros
componentes anatômicos, como os vasos e os parênquimas axial e radial, podem também
influenciar na qualidade da madeira, requerendo maior investigação. Os caracteres
anatômicos das madeiras, tais como as variações nas dimensões, freqüência e arranjo dos
elementos estruturais, tem relação com as suas propriedades físico-mecânicas, sendo de
fundamental importância para a determinação do seu possível aproveitamento tecnológico
(Paula et al. 2000). Nesse aspecto, Prior & Cutler (1996) desenvolveram estudos anatômicos
da madeira de espécies arbóreas nativas do Zimbabwe, África, concluindo que são
potencialmente aptas como fonte de 90% da energia utilizada pelas comunidades rurais
daquele país.
Paula & Alves (1980) e Paula (1993) estudaram a anatomia da madeira de espécies da
caatinga, visando a sua caracterização para a produção de energia, especialmente carvão e

3
lenha. Ressaltaram a importância do conhecimento das espécies produtoras de madeira de boa
qualidade ocorrentes na caatinga para fins energéticos como alternativa para as populações do
semi-árido. Sendo as espécies adaptadas a períodos prolongados de estiagem, sua plantação
em grande escala evitaria a falta das mesmas, uma vez que os habitantes da caatinga obtêm
lenha de diferentes espécies para os fogões, padarias, olarias e fornos para secar farinha de
mandioca, produção de carvão e estacas para cercas. Esses mesmos autores recomendaram
um estudo pormenorizado da madeira do marmeleiro, da catingueira e das juremas entre
outras espécies ocorrentes na região.
Paula (1989) referiu que muitas madeiras têm sido estudadas como subsídio à
taxonomia e, em menor escala, na fisiologia e na ecologia. Contudo, esse autor tem
direcionado seus estudos à qualificação de madeiras para produção de energia e papel,
portanto relacionando à tecnologia de madeira. Também afirmou que milhares de espécies de
boa qualidade para estes fins fazem parte da composição de nossas florestas porém, a grande
maioria jaz à espera de estudos biotecnológicos. O referido autor citou as espécies Andira
nitida Mart. ex Benth. e Enterolobium timbouva Mart. como boas para a produção de papel
por possuírem fibras de baixa fração de parede, alto coeficiente de rigidez, baixo coeficiente
de flexibilidade, pouco parênquima, vasos de pequenos diâmetros, baixo peso específico e
índice de Runkel abaixo de dois; e Hymenaea stilbocarpa Mart. e Voucapoua americana
Aublet como aptas para produção de energia pois apresentam alta freqüência de fibras, alta
fração parede, vasos de pequeno diâmetro e pouco parênquima ou ausente.
Baas & Carlquist (1985), Carlquist & Hoekman (1985) abordaram as floras de regiões
de clima árido, semi-árido do Sul da Califórnia e Carlquist (1988) floras de outras regiões da
América do Norte. Esses trabalhos estabeleceram as tendências gerais de organização da
estrutura anatômica do lenho de plantas que habitam ambientes xéricos. Alves & Angyalossy-
Alfonso (2000) estudaram diversas espécies da flora brasileira relacionando-as com fatores
ecológicos, apresentando resultados concernentes a espécies de ambientes xéricos.
A densidade da madeira é em outro parâmetro importante a ser analisado na avaliação
da sua qualidade, sendo a razão entre o volume de parede celular e o dos espaços vazios
formado pelos lúmens das células. Assim, a densidade é um parâmetro fundamental em um
estudo da madeira sendo afetada por vários fatores: estrutura anatômica, composição e
dimensão das células e da parede celular, teor de goma, resina e extrativos, estando
relacionada às propriedades e características tecnológicas importantes na produção e
utilização dos produtos florestais. Madeiras de maior densidade geralmente apresentam maior
resistência mecânica e maior valor energético (Chimelo 1980; Fujiwara et al. 1991; Paula
1993; Angyalossy et al. 2005).

4
Considerando-se a importância da análise da qualidade da madeira de espécies nativas
da caatinga, utilizadas pela comunidade para obtenção de energia (lenha e carvão), e segundo
informações obtidas e pré-visita em campo, foram selecionadas quatro espécies para este
estudo: Caesalpinia piramidalis Tul., a catingueira (Leguminosae – Caesalpinioideae);
Croton sonderianus Müell. Arg. o marmeleiro (Euphorbiaceae); Mimosa ophthalmocentra
Mart. ex Benth. a jurema-de-imbira e Mimosa tenuiflora (Willd.) Poir., a jurema-preta (ambas
Leguminosae – Mimosoideae).
A família Euphorbiaceae compreende uma das maiores das dicotiledôneas com cerca
de 290 gêneros e aproximadamente 7.500 espécies distribuídas em todo o mundo,
principalmente nas regiões tropicais. No Brasil ocorrem 72 gêneros e cerca de 1.100 espécies
de hábitos e habitat diferentes, difundidos em todos os tipos de vegetação e composta por
árvores, arbustos, subarbustos ou ervas (Barroso et al. 1991). De acordo com Barbosa et al.
(2006), essa família está representada no Nordeste por 211 espécies.
A família Leguminosae é uma das que possui maior número de espécies na caatinga
do Nordeste com cerca de 1.027 espécies. A subfamília Caesalpinioideae compreende cerca
de 152 gêneros e aproximadamente 2.800 espécies tropicais e subtropicais. As 227 espécies
dessa subfamília que ocorrem no Nordeste têm hábito arbóreo, arbustivo ou herbáceo. A
subfamília Mimosoideae apresenta cerca de 50-60 gêneros com aproximadamente 2.800
espécies distribuídas nas regiões tropicais, subtropicais e cálido-temperadas do globo. São
desde ervas muito pequenas até árvores de médio a grande porte, mas em geral arbustivas ou
arvoretas e menos comumente lianas (Schrire et al. 2005). As Mimosoideae ocorrem no
Nordeste com 295 espécies (Barbosa et al. 2006).
Por considerar o momento oportuno para intervenção na frágil relação oferta x
demanda do recurso florestal na Região Nordeste, a Associação Plantas do Nordeste - PNE,
apresentou um projeto piloto para os Estados de Pernambuco e Paraíba, semelhante ao
realizado em Zimbabwe pelo Royal Botanic Gardens, visando contribuir com a definição de
ações para o aumento da oferta e ordenamento do uso do recurso florestal, tentando ajudar na
solução do problema que se apresenta na relação diária do homem do campo com suas
demandas por lenha e carvão, principalmente para cocção de seu alimento. As condições
climatológicas e antropológicas da região estudada em Zimbabwe assemelham-se a região da
caatinga nordestina. Por esta razão aplicaram-se os mesmos métodos de trabalho de campo ali
realizados. O referido projeto de cunho interinstitucional é intitulado “Manejo Sustentado da
Vegetação Lenhosa da Caatinga, com Ênfase à Produção de Lenha para uso Doméstico das
Comunidades do Nordeste do Brasil”, envolvendo as seguintes instituições: Centro
Nordestino de Informações sobre Plantas (CNIP); Conselho Nacional de Desenvolvimento

5
Científico e Tecnológico (CNPq); Royal Botanic Gardens, Kew (RBG Kew); Empresa
Pernambucana de Pesquisa Agropecuária (IPA), Universidade Estadual de Feira de Santana
(UEFS) e Universidade Federal da Bahia (UFBA). O presente estudo faz parte deste Projeto
interdisciplinar.
Objetivou-se estudar a anatomia e a densidade básica da madeira de Croton
sonderianus Müell. Arg. , Caesalpinia piramidalis Tul., Mimosa ophthalmocentra Mart.ex
Benth. e Mimosa tenuiflora (Willd.) Poir., espécies utilizadas pelas populações que vivem no
ambiente da caatinga nos municípios de Serra Talhada e Sertânia no Estado de Pernambuco,
assim como descrever a variação radial, sentido medula-câmbio, comparar a percentagem dos
elementos do lenho no tronco e nos galhos em indivíduos desses dois municípios e
radialmente na própria árvore a fim de estabelecer o potencial total do lenho para produção de
energia e identificar caracteres anatômicos do lenho relacionando-os com o ambiente de
caatinga.
Os resultados da pesquisa são apresentados em quatro capítulos que se seguem, sendo
os capítulos 1 e 2 sobre Croton sonderianus, o 3 sobre Caesalpinia pyramidalis e o 4 sobre as
duas espécies de Mimosa. Os capítulos serão posteriormente submetidos à publicação na Acta
Botanica Brasilica (1 e 3), na IAWA Journal (2) e no Kew Bulletin (4), após alguns ajustes de
ordem editorial e tradução para língua inglesa no caso dos capítulos 2 e 4.

6
REFERÊNCIAS BIBLIOGRÁFICAS
Alves, E.S. & Angyalossy-Alfonso, V. 2000. Ecological trends in the wood anatomy of some
Brazilian species. 1. Growth rings and vessels. IAWA Journal 21: 3-30.
Angyalossy, V.; Amano, E. & Alves, E.S. 2005. Madeiras utilizadas na fabricação de arcos
para instrumentos de corda: aspectos anatômicos. Acta Botanica Brasilica 19(4): 819-
834.
Baas, P. & Carlquist, S. 1985. A comparison of the ecological wood anatomy of the floras of
southern California and Israel. IAWA Bulletin 6(4): 349-354.
Barbosa, M.R. de V.; Sothers, C.; Mayo, S.; Gamarra-Rojas, C.F.L. & Mesquita, A.C. de.
2006. Checklist das plantas do Nordeste brasileiro: angiospermas e gymnospermas.
Brasília, Ministério de Ciência e Tecnologia.
Barroso, G.M.; Peixoto, A.L.; Ichaso, C.L.F.; Costa, C.G.; Guimarães, E.F. 1991. Sistemática
de Angiospermas do Brasil. V.2. Viçosa, UFV.
Carlquist, S. 1988. Comparative wood anatomy: systematic ecological and evolutionary
aspects of dicotyledonean wood. Berlin, Springer-Verlag.
Carlquist, S. & Hoekman, D.A. 1985. Ecological wood anatomy of the woody southern
California flora. IAWA Bulletin 6(4): 319-348.
Chimelo, J. 1980. Anotações sobre anatomia e identificações de madeiras. São Paulo, IPT.
Fujiwara, S.; Sameshima, K.; Kuroda, K. & Takamura, N. 1991. Anatomy and properties of
Japanese hardwoods I. Variation of fibre dimensions and tissue proportions and their
relation to basic density. IAWA Bulletin 12: 419-424.
Lima, J.L.S. 1996. Plantas forrageiras das caatingas: usos e potencialidades. Petrolina,
APNE.
Nguyen, T.V. 1977. Variation in vessel characteristics of Eucalyptus dalrympleana
Maiden associated with sample position growth rate and climate changes. Camberra,
Australian Nature University.
Paula, J.E. de. 1989. Estudo da estrutura da madeira de espécies nativas, visando o seu
aproveitamento na produção de energia e papel. Ciência e Cultura 41(4): 344-353.
Paula, J.E. de. 1993. Madeiras da caatinga úteis para produção de energia. Pesquisa
Agropecuária Brasileira 28(2): 153-165.
Paula, J.E. de & Alves, J.L.H. 1980. Estudo das estruturas anatômicas e de algumas
propriedades físicas da madeira de 14 espécies ocorrentes em áreas de caatinga. Brasil
Florestal 10(43): 47-58.

7
Paula, J.E de; Silva-Júnior, F.G. da & Silva, A.P.P. 2000. Caracterização anatômica de
madeiras nativas de matas ciliares do centro-oeste brasileiro. Scientia Forestalis 58: 73-
89.
Prior, J.A.B. & Cutler, D.F. 1996. Radial increments in four tropical, drought tolerant
firewood species. Commonwealth Forestry Review 75(3): 227-233.
Rodal, M.J.N. & Melo, A.L. de 1999. Levantamento preliminar das espécies lenhosas da
caatinga de Pernambuco . Pp. 53-62 In: F. D. de Araújo; H. D. Prendergast & S. J. Mayo
(eds.). Plantas do Nordeste – Anais do I Workshop Geral, RBG,KEW, PNE, CNPq.
Recife, APNE.
Sampaio, E.V.S.B. 2002. Uso das plantas da caatinga. Pp. 49-68. In: Everardo V.S.B.
Sampaio; A.M. Giulietti; J. Virgílio & C.F.L. Gamarra-Rojas (ed.). Vegetação e flora da
caatinga. Recife, APNE & CNIP.
Sárkáni, S.; Stieber, J. & Filló, Z. 1957. Investigation on the wood of Hungarian Populus
species by means of quantitative xylotomy. Annals University Science Budapest 1: 219-
229.
Schrire, B.D.; Lewis, G.P.; Lavin, M. 2005. Biogeography of the Leguminosae. Pp. 21-54. In:
G.P. Lewis; B. Schrire; B. Mackinder; M. Lock (eds.). Legumes of the World. Kew,
Royal Botanic Gardens.

CAPÍTULO 1
ANATOMIA E DENSIDADE BÁSICA DA MADEIRA DE CROTON
SONDERIANUS MÜELL. ARG. (EUPHORBIACEAE), ESPÉCIE DA
CAATINGA DO NORDESTE DO BRASIL

9
RESUMO
O presente trabalho teve como objetivos estudar a anatomia do lenho do tronco e dos
ramos de Croton sonderianus Müell. Arg. e determinar a sua densidade básica, visando
subsidiar a produção de carvão. As amostras foram coletadas em populações naturais nos
municípios de Serra Talhada e Sertânia, Pernambuco, Brasil, em 2002, com amostras
coletadas no tronco, ao nível do diâmetro a altura do peito (DAP = 1,30 m) e também em dois
galhos com diferentes diâmetros. As amostras foram analisadas conforme metodologia usual
para anatomia de madeiras. A espécie revelou grande quantidade de matéria energética
expressa pela elevada densidade, no baixo percentual de parênquima e na alta freqüência de
fibras, portanto, apresentando potencial para a produção de carvão de alta qualidade desde a
fase do lenho juvenil. Os galhos apresentaram potencial energético semelhante ao do tronco,
sendo possível o seu uso como lenha ou sob a forma de carvão, evitando o sacrifício dos
indivíduos adultos, além de facilitar o manuseio.
Palavras-chave: carvão, lenha, potencial energético, anatomia da madeira, densidade.
ABSTRACT
This work aimed to study the anatomical structure of wood of the trunk and branches
of Croton sonderianus Müell. Arg. trees and determine their density, in order to subside the
production of charcoal. Samples were collected in natural populations in the municipal
districts of Serra Talhada and Sertania, PE – Brazil, in 2002, at DBH and branches. The
samples were analyzed according to usual methodology for wood anatomy. The species
revealed an enormous amount of energetic matter expressed in the high density and low
percentage of parenchyma, and in the high frequency of fibers. Thus, these characters point to
its potentiality to produce high quality charcoal, since juvenile wood phase. The branches also
presented a similar energetic potential to the trunk one, because of that it is possible their use
as firewood or as charcoal, avoiding the sacrificing of adult shrubs, besides facilitating the
handling.
Key Words: Croton, charcoal, wood anatomy, wood density

10
INTRODUÇÃO
No Estado de Pernambuco são consumidos cerca de doze milhões de estéreos de lenha
e carvão anualmente, dos quais 70% são utilizados nos domicílios, demonstrando a
importância social e econômica dos recursos florestais (Sampaio 2002). Com enfoque
conservacionista ou da sustentabilidade das populações de plantas da caatinga, as alternativas
existentes relacionam-se com a otimização do processo de produção e uso da biomassa, além
da diminuição de lenha e carvão vegetal. Como a atividade de reflorestamento tem limitações
de ordem climática e de solo, considera-se que o manejo florestal sustentado da caatinga é a
alternativa mais indicada, permitindo o uso múltiplo dos recursos florestais com custos
reduzidos e seu imediato extrativismo, além de preservar as características ecológicas do meio
ambiente pois a exploração descontrolada contribui para o desmatamento e conseqüentemente
para o desequilíbrio ecológico. Para a aplicação de métodos de manejo florestal para fins
energéticos, como a produção de lenha comum nas comunidades rurais do Nordeste, são
necessárias análises da anatomia e das características físico-químicas das madeiras das
espécies a serem utilizadas.
De forma geral, determinam-se as dimensões e percentagem das fibras e parâmetros de
qualidade. Porém, segundo Sárkáni et al. (1957), outros componentes anatômicos, como a
quantidade e a forma dos vasos e os parênquimas axial e radial podem também influenciar na
qualidade da madeira, requerendo maior investigação. A estrutura anatômica das madeiras, tal
como as variações das dimensões, freqüência e o arranjo dos elementos constituintes da
madeira, tem relação com suas propriedades físico-mecânicas, sendo de fundamental
importância para a determinação do seu possível aproveitamento tecnológico (Paula et al.
2000). Nesse aspecto, Prior & Cutler (1996) desenvolveram estudos anatômicos da madeira
de espécies arbóreas nativas do Zimbabwe, África, concluindo que são potencialmente aptas
como fonte de 90% da energia utilizada pelas comunidades rurais daquele país.
A densidade da madeira é outro parâmetro importante a ser analisado na avaliação da
sua qualidade, sendo o resultado da razão entre o volume de parede celular e dos espaços
vazios formados pelos lúmens das células. Desta forma, a densidade da madeira é afetada pela
estrutura anatômica, com respeito à composição e dimensão das células e da parede celular, os
teores de goma e resina, estando relacionada às propriedades e características tecnológicas,
importantes na produção e utilização dos produtos florestais. Madeiras de maior densidade
geralmente apresentam maior resistência mecânica e maior valor energético (Paula & Alves
1980).
Paula & Alves (1980) e Paula (1993), estudiosos da anatomia da madeira de espécies

11
da caatinga visando a sua caracterização para a produção de energia, especialmente carvão e
lenha, recomendaram um estudo pormenorizado da madeira dos marmeleiros (Croton spp.),
entre outras espécies ocorrentes na região. Tertuliano et al. (2003) desenvolveram equações
de massa e determinação da massa específica básica da madeira de Croton sonderianus Müell
Arg.
A espécie Croton sonderianus Müell. Arg. pertence à família Euphorbiaceae, sendo
conhecida entre a população regional como marmeleiro. Croton L. é o segundo maior gênero
da família, com cerca de 1.200 espécies e distribuição pantropical (Govaerts et al. 2000), das
quais cerca de 1000 ocorrem no Novo Mundo (Webster 1944). Na América do Sul, o centro
de diversidade do gênero localiza-se no Brasil, onde são referidas mais de 300 espécies
(Allem 1979). Croton sonderianus ocorre em toda a região semi-árida entre as bacias dos rios
Parnaíba e São Francisco, nos Estados do Piauí, Ceará, Rio Grande do Norte, Paraíba,
Pernambuco, Alagoas, Sergipe e Bahia (Maia 2004), sendo utilizado para a produção de lenha
e carvão juntamente com outras espécies do gênero que são produtoras de excelente madeira.
Record & Hess (1949) salientaram o uso da madeira de espécies de Croton para a
fabricação de palitos de fósforo e caixas ou na carpintaria e construção. Croton urucurana
Baill. é utilizada na carpintaria e construção civil (Mello 1950); Croton nitens Sw. para cercas
e casas (Barajas-Morales et al. 1997) e Croton sp. para carpintaria e construções rurais
(Govaerts et al. 2000), palitos de dentes e pisos (Ramos & Lagos 1988).
Nesse contexto, a Associação Plantas do Nordeste (APNE) apresentou para os Estados
de Pernambuco e Paraíba um projeto interinstitucional no Programa Plantas do Nordeste,
Subprograma Botânica Econômica intitulado “Manejo sustentado da vegetação lenhosa da
caatinga, com ênfase à produção de lenha para uso doméstico das comunidades do Nordeste
do Brasil”. Foi identificada uma frágil relação entre a oferta e a demanda dos recursos
florestais e a necessidade de ações para o aumento da sua oferta e ordenamento. Assim, esse
projeto foi proposto visando minimizar o problema diário do homem do campo por lenha e
carvão para o cozimento do seu alimento.
Pelo exposto, o presente trabalho teve como objetivos descrever a estrutura anatômica
e determinar a densidade básica da madeira de árvores de Croton sonderianus, ocorrentes nos
municípios pernambucanos de Sertânia e Serra Talhada, para avaliar o seu potencial para a
produção de energia e contribuir com subsídios para a preservação da espécie através da
implementação futura de um programa de manejo sustentado.

12
MATERIAL E MÉTODOS
Áreas de estudo - Foram selecionadas duas áreas de caatinga em Pernambuco, Brasil, nas
Estações Experimentais da Empresa Pernambucana de Pesquisa Agropecuária (IPA-PE), em
Serra Talhada (Fazenda Saco – 3.200 ha) e em Sertânia (Fazenda Cachoeira – 630 ha). Tais
áreas foram escolhidas por não sofrerem intervenção humana durante a execução do trabalho,
apresentarem grande número de indivíduos de Croton sonderianus Müell. Arg., por
encontrarem-se em um raio de 200 km e apresentarem semelhanças florísticas e também por
estarem próximas de comunidades locais com necessidade de carvão e de lenha, necessitando
de um programa de uso sustentável da vegetação,
Serra Talhada localiza-se na microrregião do Pajeú, a 418 km da capital (Recife), com
latitude de 07o53’57”S e 38o18’09”W e altitude de 500 msnm, apresentando solo maciço de
gnais superficial, marrom claro, ácido, franco argiloso com algum cascalho. A precipitação
varia de 450-750 mm de março a maio e a temperatura média anual é de 37oC. Trata-se de
uma extensa área de caatinga semi-densa, madura, pouco perturbada apresentando extrato
arbóreo, com algumas árvores medindo até 12 m, arbustivo e herbáceo, bastante
desenvolvidos. A área experimental foi de 150 ha, na região denominada Pimenteira
(PROBIO 2000).
Sertânia localiza-se na microrregião de Moxotó, a 316 km da capital, com latitude de
08o04’25”S e 37o12’24”W e altitude de 611 msnm, apresentando solo superficial, marrom
claro, ácido, franco argiloso com algum cascalho. A precipitação varia de 350-600 mm de
março a maio e a temperatura média anual é de 35o C. Trata-se de uma área extensa de
caatinga, com vegetação mais fechada na encosta, árvores e arbustos maduros, pouco
perturbada. A área experimental foi de 38 ha, conhecida como Manga do Cerecé (PROBIO
2000).
Estudo anatômico e densidade do lenho - Para o estudo anatômico do lenho foram coletadas
nas duas áreas de estudo, amostras do tronco de três indivíduos ao nível do DAP (1,30 m) e de
dois galhos (galho 01 e galho 2) em diferentes alturas e diferentes diâmetros. Foram
preparadas lâminas permanentes contendo uma seção transversal e duas longitudinais
tangencial e radial, com 18-25 µm de espessura utilizando-se micrótomo de deslize Spencer.
As seções do lenho foram clarificadas com hipoclorito de sódio (30%) e coradas com azul de
Alcião e safranina alcoólica (50%) 1:1, tratadas em série alcoólica de 50 a 100%, a cada 10%,
e montadas em euparral (Johansen 1940). Nas seções transversais do lenho, em áreas de 1 a 1
mm contíguas, ininterruptas, no sentido medula-câmbio, foi traçado um transecto ao longo de
um raio da madeira, na região de maior diâmetro para análise morfométrica. Foram

13
mensurados os elementos de vaso, as fibras e os parênquimas radial e longitudinal, utilizando-
se o programa de análise de imagem KS 300 da Carl Zeiss, no Jodrell Laboratory do Royal
Botanic Gardens Kew, Inglaterra. Na descrição dos parâmetros anatômicos, foram aplicadas
as normas da IAWA Committee (1989). As lâminas histológicas permanentes foram
depositadas na Xiloteca do Instituto de Biologia da Universidade Federal da Bahia, no
Laboratório de Micromorfologia Vegetal da Universidade Estadual de Feira de Santana e do
Laboratório de Micromorfologia do Jodrell Laboratory, RBG Kew.
Para a determinação da densidade foram coletadas amostras do tronco (DAP = 1,30m)
e amostras de galhos de maior e de menor diâmetros de dez árvores. As análises foram feitas
ao nível do cerne (próximo da medula) e ao nível do alburno (próximo ao câmbio). A
densidade do lenho foi determinada pela razão da massa seca do lenho (seco a 105o C) e o
volume saturado, de acordo com o método descrito por Vital (1984). Para a comparação das
médias e análises de correlação, utilizaram-se os testes t de Student e o Spearman Rank
Correlation, respectivamente.

14
RESULTADOS
A média do diâmetro dos troncos dos espécimes estudados foi 4,4 cm em Sertânia e
3,2 cm em Serra Talhada. Os ramos apresentaram 2,3 cm de diâmetro, os mais espessos
(Galhos 1), e 1,7 cm, os mais finos (Galhos 2) em Sertânia; em Serra Talhada, os galhos de
ambos os grupos (Galhos 1 e 2) apresentaram igual diâmetro médio, 1,7 cm.
Descrição anatômica - O lenho de Croton sonderianus apresentou anéis de crescimento
semiporosos com vasos de maiores diâmetros no lenho inicial e de menores diâmetros no
tardio (Fig. 1-3). No disco em seção transversal da madeira, às vezes são observados falsos
anéis, que não completam a circunferência totalmente. Os vasos são solitários, geminados,
múltiplos, radiais de até cinco elementos, com formato circular ou oval (Fig. 1-3), placa de
perfuração simples (Fig. 4) e pontoações areoladas intervasculares alternas, poligonais (Fig.
4). Presença de traqueídes observados na seção longitudinal tangencial (Fig. 5). O parênquima
axial é extremamente raro, quase ausente, às vezes, formado por células grandes e alongadas
(idioblastos) (Fig. 6), com ocorrência de cristais prismáticos em seu interior. Raios não
estratificados, heterogêneos, extremamente finos, medindo de 8–20,8 µm (15,40 µm) de
largura com 1-2 células, extremamente baixos, 121,66–262,57 µm (220,37 µm) de altura com
8-11 células (Fig. 6-7), contendo células procumbentes com 2-4 fileiras no centro e células
quadradas e eretas nas margens (Fig. 8). Presença de células bijugadas (Fig. 9). Fibras
libriformes com parede espessada e delgada (Fig. 10), sendo estas semelhantes às células de
parênquima axial na seção transversal, e em forma de fuso na seção longitudinal tangencial,
com pontoações areoladas. Fibras gelatinosas também foram observadas (Fig. 11).
Análise quantitativa - O teste t para a comparação de médias das percentagens de vasos não
evidenciou diferenças significativas entre as amostras do lenho do tronco procedentes de
Sertânia e de Serra Talhada. Nos galhos detectou-se maior percentagem de vasos nas amostras
de Sertânia em relação aos de Serra Talhada. Quanto à percentagem de fibras nas amostras do
tronco e do galho das árvores, detectaram-se menores valores para Sertânia em relação a Serra
Talhada. A percentagem de parênquima longitudinal foi menor nas amostras do tronco e
maior nas amostras do galho de Sertânia do que em Serra Talhada. Desta forma, verificou-se
uma maior ocorrência de parênquima no tronco e maior nos galhos das amostras do lenho de
Serra Talhada; enquanto que a percentagem de raios foi maior nas amostras do tronco e menor
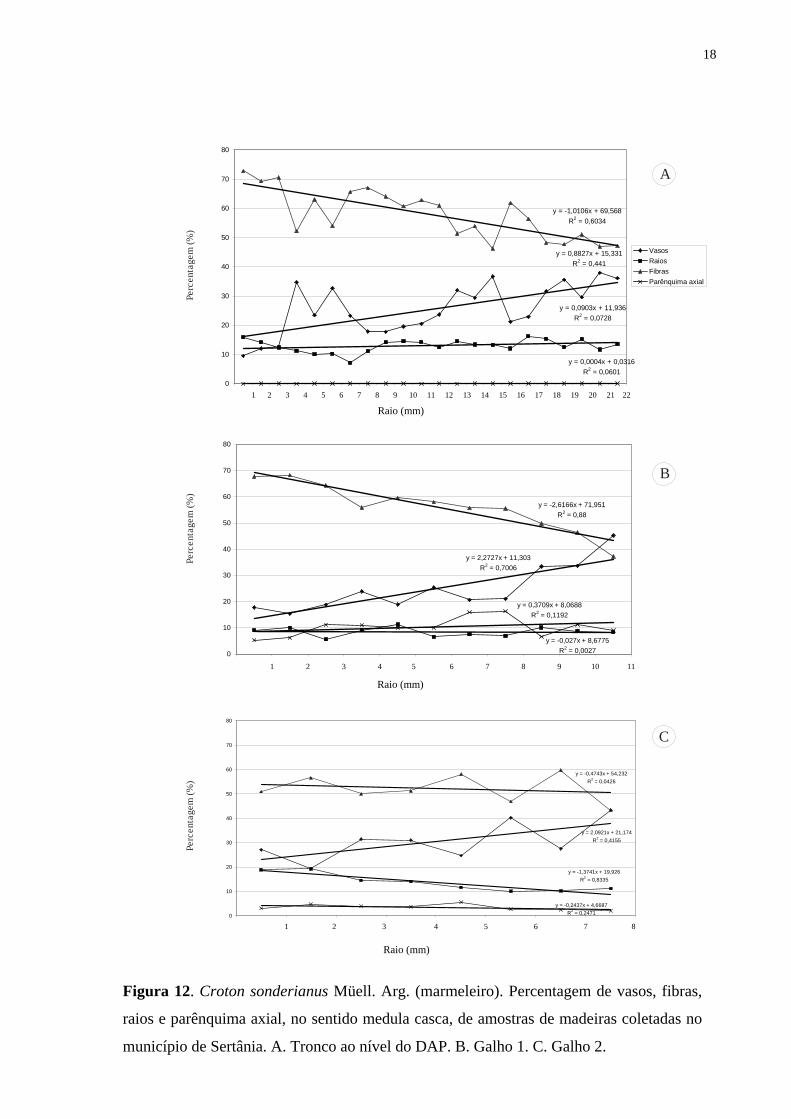
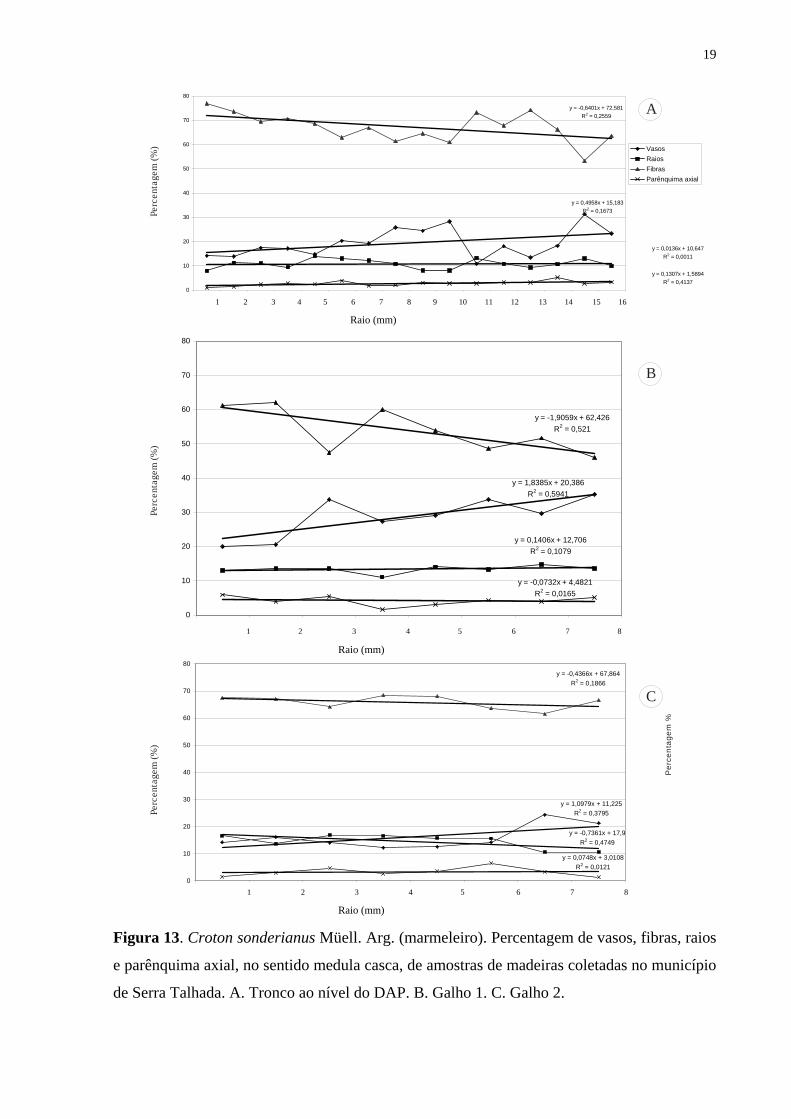
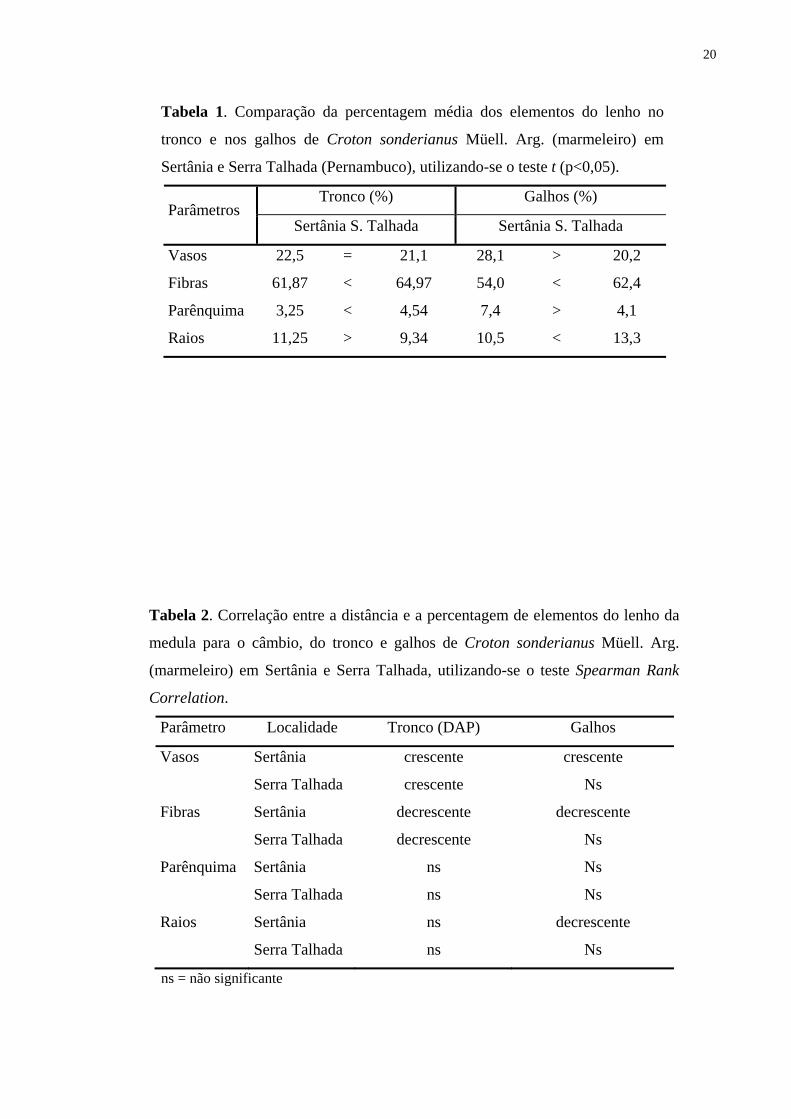
nos galhos em Sertânia (Tab. 1).
Na análise da variação medula-câmbio nas seções transversais do lenho do caule não
houve diferença significativa em relação à percentagem dos vasos (Tab. 1), procedeu-se o

15
agrupamento dos espécimes das duas regiões e aplicou-se o teste de correlação (Spearman
Rank Correlation). Detectou-se uma correlação significativa (r = 0,48 e P = 0,0001) indicando
que com o aumento da distância da medula ocorre um aumento da percentagem de vasos
(Tab. 2, Fig. 12-13). Na análise do lenho nos galhos, os indivíduos de Sertânia e Serra
Talhada não foram agrupados por apresentarem diferenças significativas entre as regiões
(Tab. 1). Na análise da Tab. 2, pode-se observar ainda que o lenho nos galhos e no tronco das
árvores procedentes de Sertânia apresentaram maior percentagem de vasos distante da medula
(Fig. 12). Todavia, não foram detectadas diferenças significativas para as amostras do lenho
de Serra Talhada (Fig. 13), exceto para o galho 01 (Fig. 13 B).
Em relação às fibras, os resultados da análise do teste de Spearman Rank Correlation
para as amostras procedentes de Sertânia e de Serra Talhada apresentaram correlações
negativas indicando que com a distância da medula ocorre uma redução da percentagem de
fibras, sendo, portanto, maior no lenho juvenil, exceto no galho 02 dos indivíduos nas duas
regiões (Tab. 2, Fig. 12 C; 13 C). Variações para vasos e fibras ocorreram nas amostras do
lenho dos galhos das árvores de Croton sonderianus em Sertânia, enquanto que em Serra
Talhada não houve diferença significativa (Tab. 2, Fig. 12 A, B).
Não foi observada correlação entre a distância da medula e a quantidade de
parênquima axial nas amostras de lenho do tronco e dos galhos das árvores de Croton
sonderianus procedentes de Sertânia e de Serra Talhada, indicando semelhanças nos lenhos
juvenil e adulto das plantas (Tab. 2, Fig. 12 A, B).
Da mesma forma, para o parênquima radial não ocorreram correlações significativas
em relação à distância da medula das amostras do lenho das árvores de Sertânia e de Serra
Talhada, com percentagem semelhante nos lenhos juvenil e no adulto (Tab. 2, Fig. 12-13).
Densidade básica - Os valores de densidade básica do lenho do tronco e dos galhos mostraram
uma amplitude de variação de 0,75–0,87 e de 0,75–0,81 g/cm3, em Serra Talhada e Sertânia
respectivamente (Tab. 3). Não foram observadas diferenças significativas para os valores de
densidade do lenho em relação à posição (lenho jovem e lenho adulto).

16
Figuras 1-9. Croton sonderianus Muell. Arg. Seções da madeira. 1-3: Transversais: 1.
Próximo da medula; 2. Região mediana; 3. Próximo do câmbio. 4-7 Tangenciais: 4.
Elementos de vaso com placas de perfuração simples e pontoações intervasculares areoladas;
5. Traqueídes com pontoações areoladas; 6. Parênquima axial em forma de idioblasto; 7.
Raios, vasos e fibras. 8-9 Radiais: 8.Raio heterocelular com células procumbentes no centro e
células erectas e quadradas nas margens; 9. Células bijugadas no raio. Barras = 100 µm (1, 3,
7); 20 µm (4-6, 8); 10 µm (9).
7 8 9
54 6
1 32
RV
F

17
1110Pd
Pe
Figuras 10-11. Croton sonderianus Muell. Arg. Seções transversais. 10. Ocorrência de fibras
libriformes de paredes espessadas (Pe) e de paredes delgadas (Pd). 11. Fibras gelatinosas
(setas). Barra = 100 µm.
18
y = -2,6166x + 71,951R2 = 0,88
y = 2,2727x + 11,303R2 = 0,7006
y = 0,3709x + 8,0688R2 = 0,1192
y = -0,027x + 8,6775R2 = 0,00270
10
20
30
40
50
60
70
80
Raio (mm)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
Raio (mm)
1 2 3 4 5 6 7 8 9 10 11
Raio (mm)
C
B
A
y = -1,0106x + 69,568R2 = 0,6034
y = 0,8827x + 15,331R2 = 0,441
y = 0,0903x + 11,936R2 = 0,0728
y = 0,0004x + 0,0316R2 = 0,0601
0
10
20
30
40
50
60
70
80
VasosRaiosFibrasParênquima axial
y = -0,4743x + 54,232R2 = 0,0426
y = 2,0921x + 21,174R2 = 0,4155
y = -1,3741x + 19,926R2 = 0,8335
y = -0,2437x + 4,6687R2 = 0,24710
10
20
30
40
50
60
70
80
1 2 3 4 5 6 7 8
Perc
enta
gem
(%)
Perc
enta
gem
(%)
Perc
enta
gem
(%)
Figura 12. Croton sonderianus Müell. Arg. (marmeleiro). Percentagem de vasos, fibras,
raios e parênquima axial, no sentido medula casca, de amostras de madeiras coletadas no
município de Sertânia. A. Tronco ao nível do DAP. B. Galho 1. C. Galho 2.
19
A
B
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Raio (mm)
y = 0,0136x + 10,647R2 = 0,0011
y = 0,4958x + 15,183R2 = 0,1673
y = 0,1307x + 1,5894R2 = 0,4137
y = -0,6401x + 72,581R2 = 0,2559
0
10
20
30
40
50
60
70
80
VasosRaiosFibrasParênquima axial
y = -0,0732x + 4,4821R2 = 0,0165
y = 0,1406x + 12,706R2 = 0,1079
y = 1,8385x + 20,386R2 = 0,5941
y = -1,9059x + 62,426R2 = 0,521
0
10
20
30
40
50
60
70
80
Perc
enta
gem
(%)
Perc
enta
gem
(%)
C
y = -0,4366x + 67,864R2 = 0,1866
y = 1,0979x + 11,225R2 = 0,3795
y = -0,7361x + 17,9R2 = 0,4749
y = 0,0748x + 3,0108R2 = 0,0121
0
10
20
30
40
50
60
70
80
Raio (mm)
1 2 3 4 5 6 7 8
Raio (mm)
1 2 3 4 5 6 7 8
Per
cent
agem
%
Perc
enta
gem
(%)
Figura 13. Croton sonderianus Müell. Arg. (marmeleiro). Percentagem de vasos, fibras, raios
e parênquima axial, no sentido medula casca, de amostras de madeiras coletadas no município
de Serra Talhada. A. Tronco ao nível do DAP. B. Galho 1. C. Galho 2.
20
Tabela 1. Comparação da percentagem média dos elementos do lenho no
tronco e nos galhos de Croton sonderianus Müell. Arg. (marmeleiro) em
Sertânia e Serra Talhada (Pernambuco), utilizando-se o teste t (p<0,05).
Tronco (%) Galhos (%) Parâmetros
Sertânia S. Talhada Sertânia S. Talhada
Vasos 22,5 = 21,1 28,1 > 20,2
Fibras 61,87 < 64,97 54,0 < 62,4
Parênquima 3,25 < 4,54 7,4 > 4,1
Raios 11,25 > 9,34 10,5 < 13,3
Tabela 2. Correlação entre a distância e a percentagem de elementos do lenho da
medula para o câmbio, do tronco e galhos de Croton sonderianus Müell. Arg.
(marmeleiro) em Sertânia e Serra Talhada, utilizando-se o teste Spearman Rank
Correlation.
Parâmetro Localidade Tronco (DAP) Galhos
Vasos Sertânia crescente crescente
Serra Talhada crescente Ns
Fibras Sertânia decrescente decrescente
Serra Talhada decrescente Ns
Parênquima Sertânia ns Ns
Serra Talhada ns Ns
Raios Sertânia ns decrescente
Serra Talhada ns Ns
ns = não significante

21
Tabela 3. Variação da densidade básica do lenho no tronco e galhos de Croton sonderianus
Müell. Arg. ocorrentes nos municípios de Sertânia e Serra Talhada, Pernambuco - Brasil.
Densidade Básica g/cm3 Tronco Galho 1 Galho 2
LJ LA LJ LA LJ LA Sertânia 0,81 0,79 0,76 0,75 0,81 0,79
Serra Talhada 0,79 0,78 0,76 0,87 0,75 0,75
LJ = lenho jovem LA = lenho adulto

22
DISCUSSÃO
A literatura especializada tem apresentado dados a respeito dos anéis de crescimento
no lenho de árvores de espécies de Croton. Em algumas espécies de Croton L. não foram
observados anéis de crescimento, dentre elas: Croton urucurana Baill., Croton lanjouwensis
Jablonski, Croton matourensis Aubl. e Croton argyratus Blume (Mello 1950; Loureiro 1968;
Phengklai 1998). Por outro lado, C.L. Dias-Leme (dados não publicados) identificou camadas
de crescimento demarcadas pelo achatamento e espessamento das fibras no lenho de árvores
de Croton floribundus Spreng. da mata atlântica; Barajas-Morales et al. (1997) fizeram igual
indicação para Croton nitens Sw.
Em determinadas regiões tropicais a disponibilidade de água, nutrientes, luz e
temperatura afetam a atividade do câmbio, induzindo a formação de zonas de crescimento
distintas no lenho (Détienne 1989; Jacoby 1989; Vetter & Botosso 1989; Worbes 1989). Os
efeitos desses fatores climáticos, relacionados com a formação dos anéis de crescimento em
árvores, parecem estar presentes em indivíduos analisados de Croton sonderianus Müell. Arg.
dos municípios de Sertânia e de Serra Talhada.
Com respeito à estrutura anatômica do lenho nas espécies do gênero Croton, tem sido
descrita a presença de vasos múltiplos e radiais, com placas de perfuração simples. Loureiro
(1968) e Phengklai (1998), analisando o lenho de C. lanjouwensis, C. matourensis e Croton
argyratus, observaram a presença de vasos moderadamente pequenos, distribuindo-se em
múltiplos e radiais (com séries de dois a sete vasos), semelhantes aos de Croton sonderianus.
Baas et al. (1983), Lindorf (1994) e Alves & Angyalossy-Alfonso (2000) encontraram vasos
agrupados predominantes no lenho de árvores de várias espécies em ambientes secos.
Wheeler & Baas (1991) consideraram que as placas de perfuração simples dos elementos de
vaso, como as de Croton sonderianus, são mais eficientes no momento do fluxo de água
intenso imposto pelas altas taxas de transpiração em regiões secas. No estudo de Alves &
Angyalossy-Alfonso (2000), 95% das espécies apresentaram lenho cujos vasos tinham placas
de perfuração simples e estavam relacionadas com ambientes quentes e secos, como o da
caatinga.
No que se refere ao parênquima axial, no lenho das árvores do gênero Croton,
Metcalfe & Chalk (1965) observaram parênquima apotraqueal, abundante, contínuo em
faixas, às vezes ocorrendo pequena quantidade do paratraqueal, além de raios maiores que 1
mm de altura, heterocelulares, estando em desacordo com os resultados apresentados aqui
para a espécie estudada, a qual apresenta pouco parênquima. Nesse aspecto, o lenho de
Croton sonderianus assemelha-se ao de Croton floribundus pela presença de parênquima

23
axial escasso, além de possuir duas células de largura e fibras com parede espessada e delgada
(Mainieri 1978). Essa característica anatômica – menor quantidade de parênquima - indica a
potencialidade da madeira da espécie como produtora de energia.
Os diferentes tipos de fibras encontrados no lenho das árvores de Croton sonderianus
indicam outra característica que a qualifica como espécie de madeira de alto potencial
energético. Hughes (1965) comentou que as fibras gelatinosas apresentam um espessamento
adicional de celulose na parede secundária e, conseqüentemente, maior espessura da parede
celular, estando relacionadas com lenho de reação e também com a função de resistência ao
estresse hídrico. Essas fibras são caracterizadas pela alteração da região interna da parede
secundária pela formação da camada G, gelatinosa ou mucilaginosa, pobre em lignina e rica
em celulose. Paviani (1978) observou fibras gelatinosas nos sistemas subterrâneos de espécies
do cerradão e concluiu que esses elementos do lenho possuem a função de reserva de água,
devido à abundância de celulose que é hidrófila. Luchi (2004), estudando árvores de Croton
urucurana Baill. procedentes de três regiões distintas (encharcada, úmida e seca), observou
também a presença de fibras gelatinosas no lenho dos espécimes de área seca, indicando-as
como mecanismo de resistência das plantas à baixa disponibilidade de água. Nas árvores de
Croton sonderianus analisadas, as fibras gelatinosas não estão relacionadas com o lenho de
reação, pois as amostras de madeiras foram coletadas de árvores com fustes retos. Desta
forma, o maior teor de celulose e a escassez de parênquima podem representar não somente
uma adaptação mecânica, mas, também, uma estratégia de sobrevivência dos indivíduos aos
períodos de baixa disponibilidade hídrica de ocorrência comum nas condições semi-áridas da
caatinga.
A análise comparativa do lenho do tronco e dos galhos das árvores de Croton
sonderianus em Serra Talhada e Sertânia indicou um comportamento diferenciado, no que se
refere a seus componentes anatômicos, ou seja, vasos, fibras, parênquimas axial e radial.
Verificou-se uma maior percentagem de fibras no lenho do tronco e dos galhos nas árvores de
Serra Talhada, sendo que os demais componentes anatômicos diferiram no tronco e nos
galhos. A precipitação pluviométrica e a temperatura são maiores e a altitude menor em Serra
Talhada, em relação a Sertânia (EMBRAPA 2001). Apesar da maior precipitação
pluviomética em Serra Talhada, lá a evapotranspiração tende a ser maior devido a alta
temperatura. Dessa forma, sugere-se que as árvores da espécie nessa região apresentam uma
maior taxa de esclerificação, representada pelo teor de fibras libriformes no seu lenho.
Os diferentes componentes anatômicos do lenho, resultantes da atividade do câmbio
vascular, podem apresentar diferenças quantitativas em função das adaptações ao ambiente e
situação fisiológica no fluxo da água, conforme afirmaram Webber (1936), Baas (1982),

24
Carlquist (1988) e Wheeler & Baas (1991). Um dos aspectos mais importantes no presente
estudo é a constatação de que o processo de esclerificação gera a formação de células de
paredes espessadas no lenho do tronco e dos galhos das árvores da espécie, indicando o seu
potencial para uso energético nas duas regiões.
É importante ressaltar que as percentagens dos elementos anatômicos do lenho podem
interferir em sua qualidade. Quanto maior a percentagem de vasos por mm2, maior será a área
de espaços vazios encontrada na madeira, conseqüentemente, menor será a proporção de
parede celular. Assim, a região da madeira onde há menor quantidade de vasos tende a
apresentar um maior potencial energético. No caso das fibras, como os lúmens são
proporcionalmente menores que os dos vasos e as paredes em geral mais espessadas, onde há
maior freqüência delas menor será a área de espaços vazios, conseqüentemente neste local o
potencial energético da madeira será maior. Croton sonderianus apresentou, no sentido
medula-casca, correlação positiva em relação aos vasos, isto é, menor percentagem próxima
da medula, correlação negativa em relação as fibras indicando uma maior percentagem
próxima da medula, e além disso, observou-se que os valores de densidade foram maiores que
0,70g/cm3, tanto próximo da medula quanto próximo da casca. Portanto, pode-se inferir que a
madeira de Croton sonderianus apresenta potencial energético para produção de carvão,
desde a fase de lenho juvenil.
Segundo Chimelo (1980), a densidade básica da madeira é considerada a sua
propriedade física mais importante, relacionando-se com outras propriedades e seus usos:
madeira de maior densidade corresponde, geralmente, a uma maior resistência mecânica e alto
valor energético. Com respeito à densidade básica do lenho, a literatura menciona que a sua
amplitude de variação em árvores de espécies florestais é de 0,13 g/cm3 a 1,4 g/cm3, sendo de
0,45 g/cm3 em Eucalyptus grandis W. Hill. (Mendes et al. 1999), espécie cuja madeira tem
alto valor comercial. O lenho das árvores de Croton glabellus L., cuja densidade básica é de
0,48 g/cm3, é considerada como moderadamente pesada (Ramos & Lagos 1988). O lenho de
Croton sonderianus procedente de Sertânia e de Serra Talhada, apresentou, tanto no tronco
como nos galhos, densidades que caracterizaram sua madeira como pesada e potencialmente
energética. Observou-se que uma maior variação da densidade em Serra Talhada ocorreu
devido ao valor de 0,87 g/cm3 no lenho adulto do galho 1 podendo estar relacionado a maior
quantidade de fibras gelatinosas nessa região deste galho (Tab. 3).
A densidade da madeira pode variar em relação à posição da amostra no tronco, à
idade fisiológica das árvores e/ou à sua distância da medula. Nas angiospermas, o aumento na
densidade da madeira na direção radial constitui-se em um modelo de variação comum,
devendo-se, também, considerar a influência dos teores das substâncias que estão presentes no

25
cerne em relação ao alburno. Para as árvores de Croton sonderianus, a diferença entre cerne e
alburno quanto à densidade não foi considerada significativa.
Considerando os valores de densidade encontrados nas amostras do lenho das árvores
de Croton sonderianus próximas da medula e do câmbio, nas duas áreas de estudo, pode-se
afirmar que desde a fase de formação do lenho juvenil sua madeira pode ser aproveitada para
fins energéticos. Esse fato torna-se significativo, especialmente na atual tendência de
utilização da madeira de árvores em ciclos mais curtos, com elevada percentagem de lenho
juvenil. Essa espécie apresentou um lenho juvenil com mais fibras e menos vasos,
influenciando positivamente no potencial energético da madeira.
Na produção de lenha e na geração de energia (carvão), o lenho das árvores deve
apresentar características propícias à combustão. Dessas características, as mais consideráveis
são: alto percentual de fibras; baixa percentagem de parênquima radial e longitudinal; baixa
percentagem de vasos; e elevada densidade. Essas características físico-anatômicas indicam
altos teores de celulose, lignina e de hemicelulose e, conseqüentemente, possui biomassa
necessária para a sustentação de uma combustão eficiente. As árvores da espécie estudada
mostraram essas características, exceto a quantidade de vasos, conforme discutido
anteriormente.
Ainda com respeito ao potencial energético apresentado pelo Croton sonderianus,
pode-se inferir que, apesar de apresentar em seu lenho os anéis semiporosos com vasos de
maior diâmetro no lenho inicial, o que poderia ocasionar uma menor densidade pelas células
largas e paredes delgadas, existiu uma compensação pela presença do lenho tardio no qual
ocorreram vasos mais estreitos com paredes mais espessadas, além da alta percentagem de
fibras e baixa percentagem de parênquima, resultando em uma maior densidade no lenho.

26
CONCLUSÕES
A madeira de Croton sonderianus apresentou camadas de crescimento distintas
delimitadas por parênquima apotraqueal marginal associado aos vasos com diâmetros
maiores.
Na análise da variação radial do lenho (sentido medula-câmbio), pode-se concluir que
existem diferenças significativas na constituição anatômica da região mais próxima da medula
em relação à região próxima do câmbio.
A espécie revelou grande quantidade de matéria energética expressa na elevada
densidade, no baixo percentual de parênquima e na alta freqüência de fibras, apresentando,
portanto, potencial para produção de carvão de alta qualidade desde a fase do lenho juvenil,
visto que nesta região ocorre uma grande concentração de fibras, vasos com menor diâmetro e
menor quantidade de parênquima.
Os galhos apresentaram potencial energético semelhante ao do tronco expresso nas
percentagens de fibras, vasos, parênquimas e densidade básica. Sendo assim, a utilização dos
mesmos pode ser útil porque evita o sacrifício dos indivíduos adultos e facilita o manuseio.

27
REFERÊNCIAS BIBLIOGRÁFICAS
Allem, A.C. 1979. Quatro novas espécies de Croton L. (Euphorbiaceae) do Rio Grande do
Sul, Brasil. Boletin Sociedad Argentina Botánica 18(3/4): 63-81
Alves, E.S. & Angyalossy-Alfonso, V. 2000. Ecological trends in the wood anatomy of some
Brazilian species. 1. Growth rings and vessels. IAWA Journal 21: 3-30.
Baas, P. 1982. Systematics, phylogenetic and ecological wood anatomy – History and
perspectives. Pp. 23-58. In: P. Baas (ed.): New perspectives in wood anatomy. Leiden,
The Hague Martinus Nijhoff Publishers.
Baas, P.; Werker, E.E. & Fahn, A. 1983. Some ecological trends in vessel characters. IAWA
Bulletin 4: 141-159.
Barajas-Morales, J.; Álvarez, G.A. & Sánchez, P.S. 1997. Anatomía de maderas de México:
especies de una selva alta perennifolia I. Publicaciones especiales de Universidad
Nacional Autonoma del Mexico 16: 46-47.
Carlquist, S. 1988. Comparative wood anatomy: systematic, ecological and evolutionary
aspects of dicotyledon wood. Berlin, Springer-Verlag.
Chimelo, J. 1980. Anotações sobre anatomia e identificações de madeiras. São Paulo, IPT.
Détienne, P. 1989. Appearance and periodicity of growth rings in some tropical woods.
IAWA Bulletin 10: 123-132.
EMBRAPA 2001. Monitoramento por Satélite. Brasil visto do espaço: Nordeste.
http://www.cdbrasil.cnpm.embrapa/ne/index. html. Acesso em 20 jun 2005.
Govaerts, R; Frodin, D.G. & Radcliffe-Smith, A. 2000. A world checklist and bibliography
of Euphorbiaceae (and Pandaceae). Kew, Royal Botanic Gardens.
Hughes, J.F. 1965. Tension wood: a review of literature. Forestry Abstracts 26(1): 1-15
IAWA Committee. 1989. List of microscopic features for hardwood identification. IAWA
Bulletin 10(3): 219-332.
Jacoby, G.C. 1989. Overview of tree-ring analysis in tropical regions. IAWA Bulletin 10(3):
219-332.
Johansen, D.A. 1940. Plant microtechnique. New York, McGraw-Hill.
Lindorf , H. 1994. Eco-anatomical wood features of species from a very dry tropical forest.
IAWA Journal 15: 361-376.

28
Loureiro, A.A. 1968. Contribuição ao estudo anatômico das madeiras de Croton lanjouwensis
(Müell. Arg.) Jablonski e Croton matourensis Aublet (Euphorbiaceae). CNP/INPA
Botânica 24: 2-17.
Luchi, A.E. 2004. Anatomia do lenho de Croton urucurana Baill. (Euphorbiaceae) de solos
com diferentes níveis de umidade. Revista Brasileira de Botânica 27(2): 271-280.
Maia, G.N. 2004. Caatinga, árvores e arbustos e suas utilidades. Fortaleza, Leitura & Arte.
Mainieri, C. 1978. Fichas de características das madeiras brasileiras. São Paulo,
Publicação IPT 966.
Mello, E.C. 1950. Estudo dendrológico de essências florestais do Parque Nacional de Itatiaia
e os caracteres anatômicos de seus lenhos. Boletim do Parque Nacional de Itatiaia 2: 1-
20.
Mendes, L.M.; Silva, J.R.M. da; Truguilho, P.F. & Lopes, G. de A. 1999. Anatomia e
características físico-químicas da madeira de Zanthoxylum tingoassuiba St. Hil. de
ocorrência na região de Lavras/MG. Cerne 5(1): 1-11.
Metcalfe, C.R. & Chalk, L. 1965. Anatomy of the dicotyledons. Oxford, Clarendon Press
Paula, J.E. de. 1993. Madeiras da caatinga úteis para produção de energia. Pesquisa
Agropecuária Brasileira 28(2): 153-165.
Paula, J.E. de & Alves, J.L.H. 1980. Estudo das estruturas anatômicas e de algumas
propriedades físicas da madeira de 14 espécies ocorrentes em áreas de caatinga. Brasil
Florestal 10(43): 47-58.
Paula, J.E de; Silva-Júnior, F.G. da & Silva, A.P.P. 2000. Caracterização anatômica de
madeiras nativas de matas ciliares do centro-oeste brasileiro. Scientia Forestalis 58: 73-
89.
Paviani, T.I. 1978. Anatomia vegetal e cerrado. Ciência e Cultura 30: 1076-1086.
Phengklai, C. 1998. Croton L. Pp. 168-169. In: M.S.M.Sosef; L.T. Hong; S. Prawirohatmodjo
(eds.). Plant resources of south-east Asia 5(3). Timber trees: lesser-known timbers.
Bogor, Prosea Foundation.
Prior, J.A.B. & Cutler, D.F. 1996. Radial increments in four tropical, drought tolerant
firewood species. Commonwealth Forestry Review 75(3): 227-233.
PROBIO, 2000. Seminário sobre avaliação e identificação de ações prioritárias para a
conservação, utilização sustentável e repartição de benefícios da biodiversidade do bioma

29
caatinga. Disponível em: http://www.biodiversitas.org/caatinga. Acesso em 20 jul.
2001.
Ramos, R.F.B. & Lagos, J.L.M. 1988. Catalogo de cien espécies forestales de Honduras:
distribuitión, propiedades y usos. Siguatepeque, Escuela Nacional de Ciencias
Forestales.
Raven, P.H.; Evert, R.F. & Eichhorn, S.E. 2001. Biologia Vegetal. 6a. ed. Rio de Janeiro,
Guanabara Koogan.
Record, S.J. & Hess, R.W. 1949. Timbers of the New World. New Haven, Yale University
Press.
Sampaio, E.V.S.B. 2002. Uso das plantas da caatinga. Pp. 49-68. In: Everardo V.S.B.
Sampaio; A.M. Giulietti; J. Virgílio & C.F.L. Gamarra-Rojas (eds.). Vegetação e flora da
caatinga. Recife, APNE & CNIP.
Tertuliano, S.S.X.; Silva, J.A.da; Paes, J.B.; Araújo, L.V.C. Equações de massa e
determinação da massa específica básica da madeira de Croton sonderianus Müell. Arg.
Brasil Florestal 22(76): 15-22.
Sarkani, S.; Stieber, J. & Fillo, Z. 1957. Investigation on the wood of Hungarian Populus
species by means of quantitative xylotomy. Annals University Science Buda 1: 219-229.
Vetter, R.E. & Botosso, P.C. 1989. Remarks on age and periodicity of growth rings in some
tropical woods. IAWA Bulletin 10(2): 133-146.
Vital, B.R. 1984. Métodos de determinação de densidade da Madeira. Viçosa, UFV.
Webber, I.E. 1936. The woods of sclerophyllous and desert shrubs of California. American
Journal of Botany 23: 181-188.
Webster, G.L. 1944. A synopsis of the genera and suprageneric taxa of Euphorbiaceae.
Annals Missouri Botanical Garden 81(1): 33-144.
Wheeler, E.A. & Baas, P. 1991. A survey of the fossil record for dicotyledon wood and its
significance for evolutionary and ecological wood anatomy. IAWA Bulletin 12(3): 275-
332.
Worbes, M. 1989. Growth rings, increment and age of trees in inundation forests, savannas
and a mountain forest in the neotropics. IAWA Bulletin 10(2): 109-122

CAPÍTULO 2
INFLUÊNCIA DA PRECIPITAÇÃO PLUVIOMÉTRICA ANUAL NA
FORMAÇÃO DOS ANÉIS DE CRESCIMENTO DE CROTON
SONDERIANUS MÜELL. ARG. (EUPHORBACEAE) PROVENIENTE
DA CAATINGA DO NORDESTE DO BRASIL

31
RESUMO
O presente trabalho objetivou apresentar a descrição anatômica dos anéis de
crescimento do lenho de Croton sonderianus, relacionando-a com a precipitação anual, a
densidade e o incremento anual no tronco, a fim de se obter a dinâmica de crescimento e
qualidade da madeira. Utilizaram-se os métodos da densitometria de raios X e o das medidas
dos anéis através de mesa de medição conforme rotina de laboratório de dendrocronologia. As
amostras do lenho de seis árvores da espécie foram coletadas nos municípios de Serra Talhada
e Sertania, Pernambuco, Brasil, em 2002. Observou-se que esta espécie na caatinga nordestina
apresenta a capacidade de formar anéis de crescimento indicando seu grande potencial para
estudos dendrocronológicos nos trópicos. Anéis mais largos foram formados durante os anos
de maior disponibilidade de água e anéis mais estreitos, nos anos de baixa precipitação, com
redução e/ou suspensão da atividade do câmbio (abaixo de 300 mm anual). Finalmente, as
correlações obtidas entre incremento anual e o histórico de precipitações demonstraram tratar-
se de anéis anuais de crescimento.
Palavras-chave: Croton, anel de crescimento, caatinga, carvão.
ABSTRACT
This study had the objective of presenting the anatomic description of the growth rings
of the Croton sonderianus wood, relating it to the annual rainfall, the density and the annual
increment of the trunk, in order to obtain growth dynamics and quality of the wood. Both
were used, the X-ray densitometry and the ring measurement methods via measuring table
according to dendrocronology laboratory routine. Wood samples of six trees of this species
were collected in the municipal districts of Serra Talhada and Sertania, PE – Brazil, in 2002.
There was noticed that this species in the northeastern caatinga presents the ability to form
growth rings indicating its great potential for dendrocronology studies in the tropic regions.
Larger rings were formed during the years of greater abundance of water, and narrower rings
in years of low precipitation, with reduction and/or ceasing of cambium activity (below 300
mm per year). Finally, the correlations obtained between the annual increment and the
precipitation records demonstrated that those were indeed annual growth rings.
Kew wood: Croton, growth ring, wood anatomy, charcoal.

32
INTRODUÇÃO
Localizada na região Nordeste, ocupando a segunda posição no balanço energético
regional com aproximadamente 25% de suas madeiras voltada basicamente para cocção e
geração de vapor, a caatinga encontra-se exposta à intensa exploração antrópica,
especialmente as espécies vegetais arbustivas e arbóreas que têm suas madeiras consumidas
pela população local para obtenção de energia (Sampaio 2002). Assim, este Bioma funciona
como grande fonte de energia primária. Pouco se sabe sobre a importância, a dinâmica de
crescimento e regeneração natural dessas espécies da caatinga.
Para Rodal & Melo (1999), o Nordeste semi-árido brasileiro ocupa uma área de
aproximadamente 900.000 km2 correspondendo a 54% da região Nordeste. As chuvas estão
concentradas em um único período com duração de três a cinco meses, valores anuais de 400
a 800 mm e um regime pluvial com distribuição irregular no tempo e no espaço, tendo como
cobertura vegetal típica a caatinga. Essa vegetação estende-se do Estado do Piauí ao Norte do
Estado de Minas Gerais e caracteriza-se pela caducifolia da maioria das espécies.
A caatinga apresenta uma riqueza em espécies e clima característico de ambiente
semi-árido. Isso favorece um sítio para pesquisas em dendrocronologia pela reação sensitiva a
variáveis ambientais, especialmente a precipitação pluviométrica.
Dentre as diversas espécies endêmicas da caatinga utilizadas pela comunidade para
obtenção de energia, Croton sonderianus Müell. Arg., conhecida pelo nome vulgar de
marmeleiro e pertencente à família Euphorbiaceae, é utilizada para a produção de lenha e
carvão. O gênero Croton L. é o segundo maior da família, com distribuição pantropical e
cerca de 1.200 espécies (Govaerts et al. 2000), sendo a maioria americana e apenas 200
distribuídas pelo Velho Mundo (Webster 1944). Na América do Sul, o centro de diversidade
do gênero localiza-se no Brasil, no qual são referidas mais de 300 espécies (Allem 1979),
dessas, cerca de 1/3 ocorre no bioma das caatingas. Croton sonderianus ocorre em toda a
região semi-árida entre as bacias dos rios Parnaíba e São Francisco, ou seja, nos Estados do
Piauí, Ceará, Rio Grande do Norte, Paraíba, Pernambuco, Alagoas, Sergipe e Bahia (Lima
1996). Nos municípios pernambucanos de Sertânia e Serra Talhada, a madeira da espécie
também é utilizada para produção de lenha e carvão. Algumas espécies do gênero são
mencionadas como produtoras de excelentes madeiras. Croton urucurana Baill. é utilizada na
carpintaria e construção civil (Mello 1950). A madeira de Croton nitens Sw. é indicada para o
uso de cercas e casas (Barajas-Morales et al. 1997). Record & Hess (1949), Ramos & Lagos
(1988) e Govaerts et al. (2000) indicaram a madeira de espécies de Croton para carpintaria,
construções rurais, fabricação de caixas, palitos de dentes e palitos de fósforos.

33
A necessidade de obtenção de informações sobre a taxa de crescimento e a idade de
árvores de espécies tropicais levou ao desenvolvimento de métodos de análise de amostras de
madeiras extraídas das árvores, caracterizando o método estático, ou que utilizam as árvores
vivas, caracterizando o método dinâmico (Fahn et al. 1981). A densitometria de raios X é um
método estático, que possibilita a determinação da periodicidade da atividade cambial e da
taxa de crescimento das árvores, e comprova a formação de anéis de crescimento. As análises
de anéis de crescimento atingiram grande avanço após o desenvolvimento das análises de
raios X na França por Polge (1963), permitindo a determinação da densidade da madeira intra
e interanéis de crescimento através da leitura de radiografias de amostras de madeira em um
microdensitômetro. Atualmente tornou-se rotina em laboratórios de dendrocronologia,
inclusive no Brasil (Amaral & Tomazello-Filho 1998).
Tomazello-Filho et al. (2001) afirmaram que existe um princípio estabelecido que “a
seleção de um sítio para pesquisas em dendrocronologia deve ser conduzida com critérios de
forma que as árvores mostrem reação sensitiva à variável ambiental a ser estudada” ou “sítios
selecionados devem maximizar o sinal climático que está sendo analisado pelos anéis de
crescimento das árvores”. Por exemplo, as árvores que respondem especificamente às
condições de seca são usualmente encontradas em locais onde as chuvas são limitantes, como
nos terrenos com exposição de rochas. Isto é uma característica que ocorre na caatinga
nordestina tornando esta região apta para estudos dendrocronológicos.
Apesar da discussão que ainda persiste quanto à questão dos anéis anuais de
crescimento em regiões tropicais, desde a década de 20, no século XX, autores têm
apresentado resultados quanto à presença desses anéis: Coster (1927, 1928); Schulz (1960);
Alvim (1964) e Alvim & Alvim (1976), Botosso & Vetter (1991), Vetter & Botosso (1993),
Worbes (1995), Wehr & Tomazello-Filho (2000) e Ferreira-Fedele et al. (2004). Nas últimas
décadas a União Internacional das Organizações de Pesquisas Florestais (IUFRO) tem
realizado sessões específicas relativas à ocorrência de anéis de crescimento em espécies
tropicais. Em 1996, o tema foi reconhecido pela comunidade científica como importante a ser
discutido (Baas & Vetter 1989). Segundo Tomazello-Filho et al. (2001), os resultados
publicados são suficientes para convencer à comunidade científica acerca da sazonalidade e
do crescimento rítmico de inúmeras espécies tropicais em diversas condições ecológicas,
resultando na formação de anéis de crescimento. Contudo, ainda são poucas as informações
concernentes à periodicidade e sazonalidade da atividade cambial e a formação de anéis de
crescimento no lenho do tronco de árvores tropicais, especialmente de plantas de caatinga.
Assim, o presente trabalho teve como objetivo apresentar a descrição anatômica dos
anéis de crescimento ocorrentes em Croton sonderianus, relacionando a formação destes

34
anéis com a precipitação anual, a densidade e o incremento anual no tronco, a fim de se obter
a dinâmica de crescimento e qualidade da madeira, bem como para servir de estímulo na
preservação e o aproveitamento da espécie em plantios de manejo sustentável na caatinga
nordestina; utilizando-se, para tanto, os métodos da densitometria de raios X e o das medidas
dos anéis através de mesa de medição.

35
MATERIAL E MÉTODOS
Áreas de estudo - Foram selecionadas duas áreas de caatinga em Pernambuco, Brasil, nas
Estações Experimentais da Empresa Pernambucana de Pesquisa Agropecuária (IPA-PE), em
Serra Talhada (Fazenda Saco – 3.200 ha) e em Sertânia (Fazenda Cachoeira – 630 ha). Tais
áreas foram escolhidas por não sofrerem intervenção humana durante a execução do trabalho,
apresentarem grande número de indivíduos de Croton sonderianus Müell. Arg., por
encontrarem-se em um raio de 200 km e apresentarem semelhanças florísticas e também por
estarem próximas de comunidades locais com necessidade de carvão e de lenha, necessitando
de um programa de uso sustentável da vegetação.
Serra Talhada, localiza-se na microrregião do Pajeú, a 418 km da capital (Recife), com
latitude de 07o53’57”S e 38o18’09”W e altitude de 500 msnm, apresentando solo maciço de
gnais superficial, marrom claro, ácido, franco argiloso com algum cascalho. A precipitação
varia de 450-750 mm, de março a maio e a temperatura média anual é de 37o C. Trata-se de
extensa área de caatinga semi-densa, pouco perturbada, apresentando extrato arbóreo, com
árvores de até 12m, arbustivo e herbáceo, bastante desenvolvidos. A área experimental foi de
150 ha em uma região denominada Pimenteira (PROBIO 2000).
Sertânia localiza-se na microrregião de Moxotó, a 316 km da capital, com latitude de
08o04’25”S e 37o12’24”W e altitude de 611 msnm, apresentando solo superficial, marrom
claro, ácido, franco argiloso com algum cascalho. A precipitação varia de 350-600 mm, de
março a maio e a temperatura média anual é de 35o C. Trata-se de uma área extensa de
caatinga, com vegetação mais fechada na encosta, árvores e arbustos bem desenvolvidos,
pouco perturbada. A área experimental foi de 38 ha em uma região denominada Manga do
Cerecé (PROBIO 2000).
Coleta e preparo das amostras - Para a avaliação do incremento anual e do incremento
acumulado do tronco de Croton sonderianus, foram coletados em março de 2002, na altura do
diâmetro a altura do peito (DAP = 1,30 m), discos dos troncos de três árvores de cada local de
estudo, totalizando seis árvores. Os planos transversais dos discos de madeira foram polidos
com lixas de diferentes granulometrias (80-600) para possibilitar o exame visual dos
elementos anatômicos sob aumento de 10x em estereomicroscópio.
Foram analisadas três amostras de cada município , sendo SE1, SE2 e SE3 as amostras
de Sertânia e ST1, ST2 e ST3 as de Serra Talhada (Fig. 1).
Análise do lenho por densitometria de raios X - Foram realizadas análises de densitometria de
raios X da madeira para a determinação da massa específica aparente (m.e.a.) ou densidade
básica em seis amostras de Croton sonderianus, bem como para melhor definir os limites das

36
camadas de crescimento. Essas análises foram realizadas de acordo com procedimentos
descritos por Schweingruber (1988) e Amaral & Tomazello-Filho (1998). Nas amostras dos
discos de madeira, foram selecionados raios representativos em cada amostra, que foram
cortados com serra em dimensões de aproximadamente 1x1 cm largura/altura e comprimento
variável de acordo com a extensão dos raios. Essas sub-amostras foram fixadas em suportes
de madeira e serradas novamente em equipamento de dupla serra paralela, resultando em
amostras com 2x10 mm de espessura/largura e comprimento igual ao do raio, de forma a se
realizar a radiografia do corte transversal da madeira. As amostras foram então mantidas em
sala de climatização juntamente com um padrão de acetato de celulose até atingir a umidade
de equilíbrio (12%, 8-12 h), para serem irradiadas em equipamento de raios X Hewlett
Packard, Faxitron 43805N. Utilizou-se filme de raios X Kodak modelo t-mat G/RA (18-24
cm) que foi revelado em equipamento automático Macrotec MX-2. A radiografia foi
digitalizada em scanner Hewlett Packard ScanJet 6100C/T com resolução de 1000 dpi, em
escala de cinza de 256 graus. A imagem do padrão de acetato de celulose que apresentava
densidade conhecida semelhante ao da madeira foi comparada com a densidade óptica das
imagens das amostras, corrigindo-se os valores para massa específica aparente (kg/m3) a cada
10 µm do raio. Para isso, foram utilizados softwares específicos, como o CRAD que
correlacionou as densidades ópticas do padrão e das amostras, e o CERD que possibilitou
discriminar as camadas de crescimento e a determinação da massa específica aparente das
amostras (Mothe et al. 1998). Os resultados foram analisados utilizando-se o EXCEL para o
preparo de gráficos, interpretações dos resultados e correlações entre as amostras.
Análise da largura dos anéis (incremento anual) - Selecionaram-se dois raios (medula-câmbio)
de cada amostra, representativos do crescimento médio de cada árvore. A observação
anatômica das amostras sob estereomicroscópio foi realizada para identificar e demarcar as
camadas de crescimento. Nesse procedimento, cada camada de crescimento foi acompanhada
ao longo de toda sua circunferência no disco de madeira para ter certeza de não tratar-se de
falsa camada de crescimento e também para conferir se coincidiam sucessivamente nos dois
raios analisados. A mensuração das camadas de crescimento foi realizada utilizando-se de
uma mesa de medição Velmex, no Laboratório de Anatomia e Identificação de Madeiras da
Escola Superior de Agricultura “Luiz de Queiroz” – Universidade de São Paulo (Piracicaba,
SP). Esse procedimento consistiu em posicionar o início do raio selecionado na amostra
(limite da medula) sobre a mesa de medição em coincidência com o retículo da lente ocular
do estereomicroscópio, que estava acoplado à mesa de medição. O deslocamento vertical da
mesa moveu a amostra permitindo a medição precisa de cada camada de crescimento (em
milímetros).

37
As medidas obtidas pela mesa de medição foram transferidas para uma planilha
eletrônica EXCEL, na qual foram conferidas e analisadas. Gráficos dos incrementos anuais
das camadas de crescimento foram utilizados primeiramente para conferir a coincidência nos
valores dos incrementos entre os raios de cada amostra e, a seguir, entre as médias das
diversas amostras. Estas análises permitiram verificar as semelhanças entre os aumentos e os
decréscimos do incremento anual entre cada raio de uma árvore e entre árvores.
Posteriormente, os incrementos anuais médios de cada amostra foram comparados com as
precipitações anuais registradas para cada local de estudo. Por fim, as médias dos incrementos
das três árvores de Sertânia e das três árvores de Serra Talhada foram comparadas com os
registros das precipitações, estabelecendo-se curvas de incremento anuais representativas para
a espécie e para cada local de estudo. A partir da soma dos incrementos anuais de cada raio
analisado, determinou-se o incremento acumulado de cada amostra, representando o ritmo de
crescimento das árvores desde o nascimento até a data da coleta. Vale ressaltar que o último
anel foi formado em 2001, sendo assim no sentido câmbio-medula analisou-se ano a ano
considerando-se a cronologia anual regressiva.

38
RESULTADOS
Os valores dos diâmetros dos troncos dos espécimes estudados foram 3,4; 5,1; 5,8 cm
em Sertânia e 2,6; 3,3; 3,8 cm em Serra Talhada.
A quantidade de camadas de crescimento foi a mesma em todos os raios analisados
correspondentes a cada amostra. Porém, devido à formação de madeira de reação no tronco, a
medula de cada amostra encontrava-se deslocada do centro dos discos, resultando em
diferenças nas larguras de cada camada de crescimento (Fig. 1).
Descrição anatômica - Os anéis de crescimento são perfeitamente demarcados no lenho de
Croton sonderianus e apresentam o lenho inicial formado por vasos em disposição difusa e
fibras com paredes delgadas, e o lenho tardio também com vasos difusos, porém as fibras
possuem paredes mais espessadas. Camadas de crescimento distintas delimitadas por
parênquima apotraqueal marginal associado aos vasos com diâmetros maiores. Observou-se
ainda, vasos agrupados em faixas tangenciais com maior diâmetro em comparação com os
demais vasos do anel (Fig. 2).
Densitometria de raios X - Nas Fig. 3 e 4, são apresentadas as oscilações da m.e.a. das
amostras de Croton analisadas. Nessas figuras, são apresentadas também as imagens das
amostras retiradas das radiografias das mesmas, bem como, indicações dos limites das
camadas de crescimento, demonstrados através de traços verticais na parte superior dos
gráficos. Os resultados mostraram variações da m.e.a. ao longo dos raios analisados com
características próprias da espécie em estudo. Observou-se como constante, valores de m.e.a.
menores no início das camadas de crescimento, seguidos de aumentos gradativos até o final
da mesma camada de crescimento (Fig. 3, 4). No entanto, algumas camadas de crescimento
apresentaram m.e.a. maiores que a média em quase toda a extensão da camada de
crescimento, como entre 8-10 mm do raio da amostra ST2 (Fig. 3B).
Outra característica observada foi a pequena oscilação na massa específica aparente
(m.e.a.) em algumas camadas de crescimento sucessivas e de pouca largura, como entre 6-7
mm do raio da amostra SE1 (Fig. 4A). Verificaram-se ainda alguns casos em que os maiores
valores da m.e.a., ocorreram no início da camada de crescimento, como entre 10-12 mm da
amostra ST2 (Fig. 3B).
Estas características nas oscilações da m.e.a. demonstram a dificuldade de se utilizar
apenas análises de densitometria para a definição dos limites das camadas de crescimento, o
que também foi verificado para a análise anatômica das camadas de crescimento, que muitas
vezes não mostraram os limites das camadas de crescimento, como por exemplo, para os anos
de seca. No entanto, a análise conjunta da anatomia do lenho com as análises de densitometria

39
de raios X mostrou-se muito adequada para a definição dos limites das camadas de
crescimento.
Larguras das camadas de crescimento - As larguras das camadas de crescimento registradas
para cada raio, de cada amostra são apresentadas na Tabela 1. A partir destas medidas,
verificaram-se diferenças nas quantidades de camadas de crescimento com relação a cada
árvore analisada, sendo que a amostra ST2 foi a que apresentou a menor quantidade de
camadas de crescimento com 12 camadas, e a amostra SE3 a que apresentou a maior
quantidade de camadas de crescimento, com 30 camadas.
Pela Tabela 1 verifica-se a menor medida de incremento anual de 0,158 mm na
amostra ST3 em Serra Talhada e SE1 em Sertânia, e a maior medida de 4,121 mm na amostra
SE2 determinadas em mesa de medição. Pode-se também verificar, pela densitometria de
raios X, a menor medida de incremento anual de 0,11 mm e a maior medida de 4,46 mm,
ambas na amostra SE2. O incremento médio dos três raios analisados para cada amostra foi de
0,96; 1,06; 0,62; 0,99; 1,11 e 0,96 mm para as amostras ST1, ST2, ST3, SE1, SE2 e SE3,
respectivamente.
Verificou-se para os três raios analisados em todas as árvores, as mesmas tendências
de variação do incremento, com pequenas alterações entre os raios, devido ao deslocamento
da medula do centro do tronco e também da formação de madeira de reação (Fig. 5, 6.).
As árvores apresentaram no início do desenvolvimento, altos índices de incremento
em diâmetro do tronco, que se alternaram com períodos de menor incremento, como pode ser
observado nas Fig. 5 e 6. Com exceção da amostra ST2, as demais árvores apresentaram
diminuição progressiva do incremento nas últimas 8 camadas de crescimento. Para todas
estas, foram ainda observadas baixas taxas de incremento nos anos de 1988 e 1989, seguidos
de outros anos também com baixo incremento, como na amostra SE2 (Fig. 6B). No entanto,
no período anterior, de 1984 a 1987, todas apresentaram altos índices de incremento.
Relação entre as larguras das camadas de incremento e a precipitação - Para verificar qual
fator ambiental induziu a formação das camadas de crescimento nas árvores de Croton
sonderianus, foram realizadas correlações entre as larguras das camadas de crescimento com
as precipitações dos locais de estudo. Para isso, considerou-se a princípio que as camadas de
crescimento foram formadas em períodos anuais de incremento, obedecendo ao ritmo
climático sazonal.
Os resultados mostraram que todas as amostras apresentaram correlação significativa
entre as larguras das camadas de crescimento e as médias anuais de precipitações, como
mostram as Fig. 7 e 8.
Os registros de precipitações para a região de Serra Talhada mostraram a ocorrência

40
de dois períodos de seca intensa sem precipitações durante todo o ano de 1978 e nos anos de
1988 e 1989. Outros períodos de menores precipitações, entre 200-300 mm anuais, foram
verificados nos anos de 1982, 1993 e 1998. Nos intervalos destes períodos de seca intensa
verificou-se a ocorrência de anos de maiores precipitações, entre 300-600 mm anuais,
destacando-se o ano de 1985 com precipitação acima de 1.600 mm.
Tais alterações no regime de precipitações ocasionaram respostas das plantas com
maiores incrementos anuais durante os anos de regime normal de precipitação, destacando-se
os anos entre 1984 e 1986 quando ocorreu a precipitação acima da média em 1985,
registrando 1.600 mm. Para os anos de menores precipitações verificou-se uma resposta das
árvores através da diminuição expressiva de seus incrementos, ou seja, com a formação de
camadas de crescimento bastante estreitas. Destacaram-se as camadas formadas nos anos de
1988 e 1989, como se verificou para a amostra ST3, que apresentou camadas de crescimento
com larguras entre 0,2-0,3 mm (Fig. 7C), e diferenciada anatomicamente das demais, com
vasos de diâmetros reduzidos e concentrados em pequena porção do lenho.
Para outras amostras, como por exemplo a ST1, as camadas de crescimento
apresentaram-se de difícil distinção e não demarcadas por parênquima marginal ou por anéis
semi-porosos. Observaram-se alterações anatômicas como ligeiro aumento do lume dos vasos
no início destas camadas de crescimento. No entanto, através das análises por densitometria
de raios X, verificaram-se camadas demarcadas com mudanças abruptas de densidade nos
limites das camadas de crescimento, devido à presença de maior número de fibras com
paredes celulares espessas na parte final da camada de crescimento e queda brusca desta
densidade no início da camada seguinte, como normalmente observado para as camadas de
crescimento distintas (Fig. 3A, 7A).
Determinações do incremento acumulado - As amostras SE1 e SE2 de Sertânia apresentaram
as maiores taxas de crescimento determinadas para o início de desenvolvimento das plantas,
com relação às demais árvores analisadas. A amostra SE3, também de Sertânia, apresentou no
início do desenvolvimento taxas de crescimento semelhantes às observadas para as amostras
de Serra Talhada (ST1, ST2 e ST3). Após os 15 mm de raio a amostra SE3 apresentou
tendência de incremento do tronco semelhante às demais do mesmo local de estudo (Fig. 9).
Verificou-se a presença de falsos anéis de crescimento, como na amostra SE2, que
apresentou uma camada de crescimento distinta quando observada sob microscopia, porém
que não permitia correlação com os registros de precipitação para o período. Assim, quando
eliminada esta camada e considerada a soma da mesma com a camada sucessiva, a correlação
com a precipitação demonstrou-se melhor.
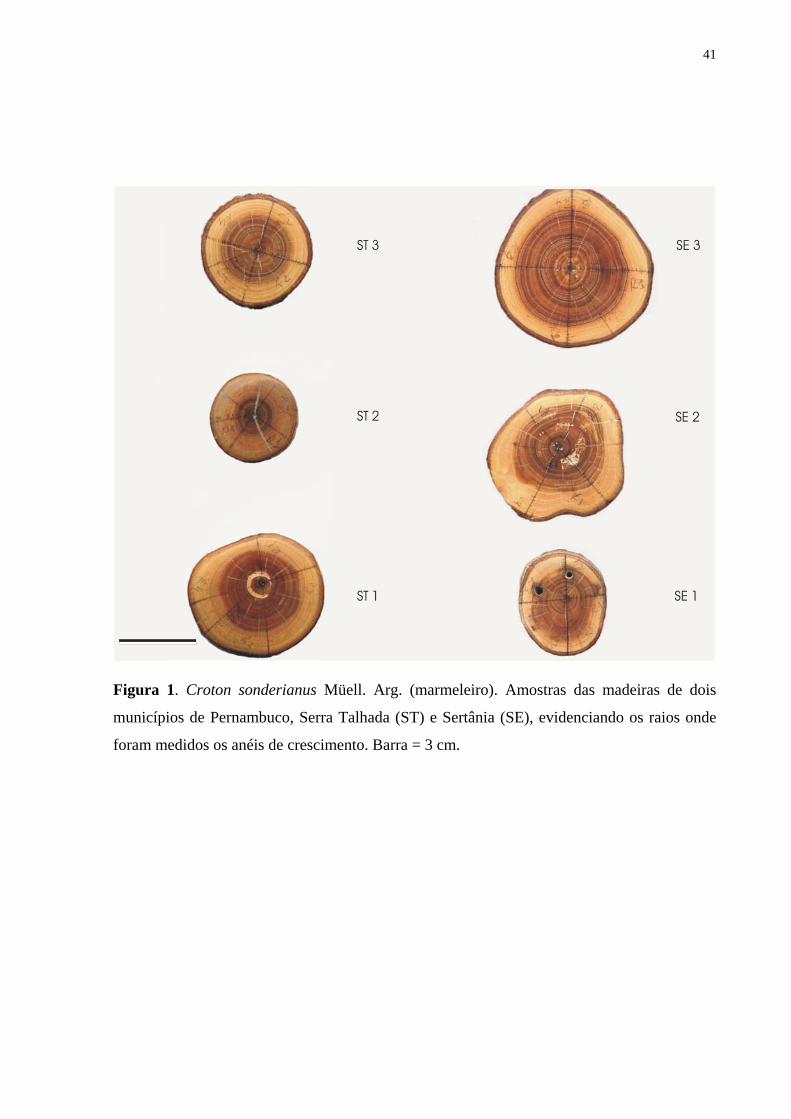
41
SE 3
SE 1
SE 2
ST 3
ST 2
ST 1
Figura 1. Croton sonderianus Müell. Arg. (marmeleiro). Amostras das madeiras de dois
municípios de Pernambuco, Serra Talhada (ST) e Sertânia (SE), evidenciando os raios onde
foram medidos os anéis de crescimento. Barra = 3 cm.

42
A
B
Figura 2. Croton sonderianus Müell. Arg. A. Seções transversais montadas desde
a região da medula até a região próxima da casca (sem escala). B. Seção
transversal do tronco (DAP) evidenciando os anéis de crescimento. Barra = 100
µm.

43
ST 1
0
200
400
600
800
1000
1200
1400
0 2 4 6 8 10 12 14 16 18 20 22
m.e
.a. (
kg/m
3 )
A
B
ST 2
0
200
400
600
800
1000
1200
1400
0 2 4 6 8 10 12 1
m.e
.a. (
kg/m
3 )
Raio (mm)
4
ST 3
0
200
400
600
800
1000
1200
1400
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
m.e
.a. (
kg/m
3 )
Raio (mm)
Raio (mm)
C Figura 3. Croton sonderianus Müell. Arg. Massa específica aparente
(m.e.a.) das três amostras de Serra Talhada, determinada por densitometria
de raios X.
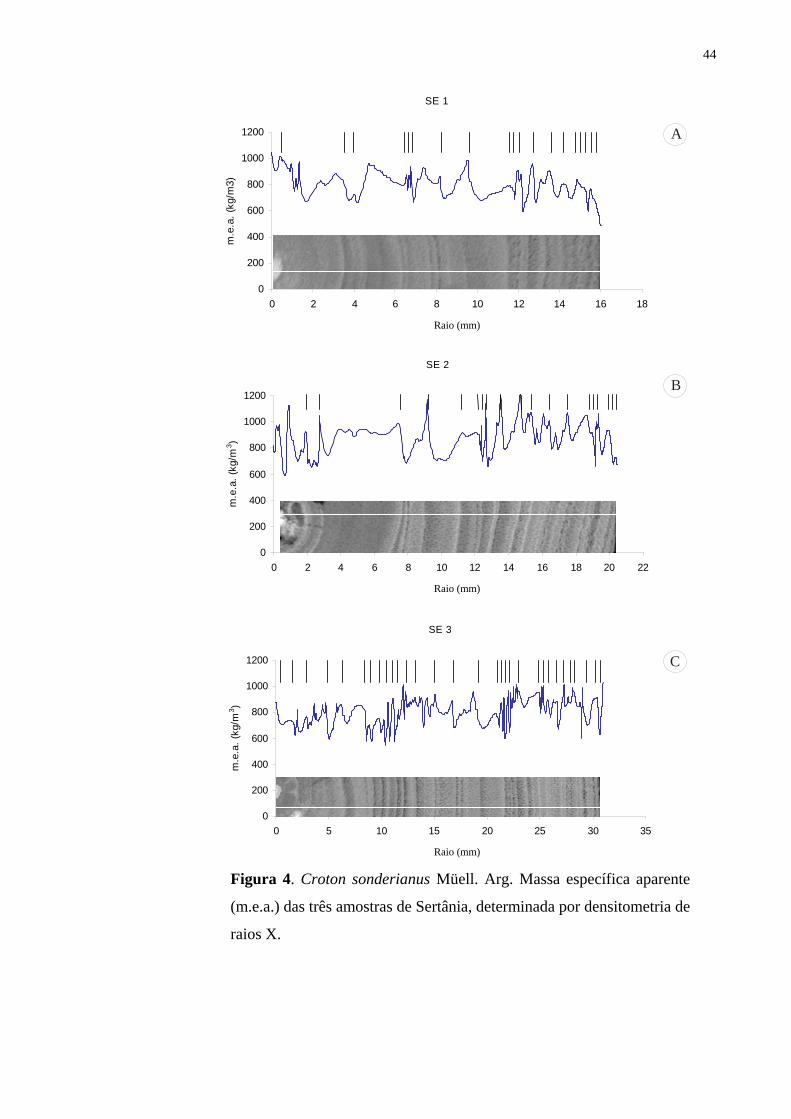
44
SE 1
0
200
400
600
800
1000
1200
0 2 4 6 8 10 12 14 16 18
m.e
.a. (
kg/m
3)
Raio (mm)
A
B SE 2
0
200
400
600
800
1000
1200
0 2 4 6 8 10 12 14 16 18 20 22
m.e
.a. (
kg/m
3 )
Raio (mm)
SE 3
0
200
400
600
800
1000
1200
0 5 10 15 20 25 30 35
m.e
.a. (
kg/m
3 )
Raio (mm)
C
Figura 4. Croton sonderianus Müell. Arg. Massa específica aparente
(m.e.a.) das três amostras de Sertânia, determinada por densitometria de
raios X.
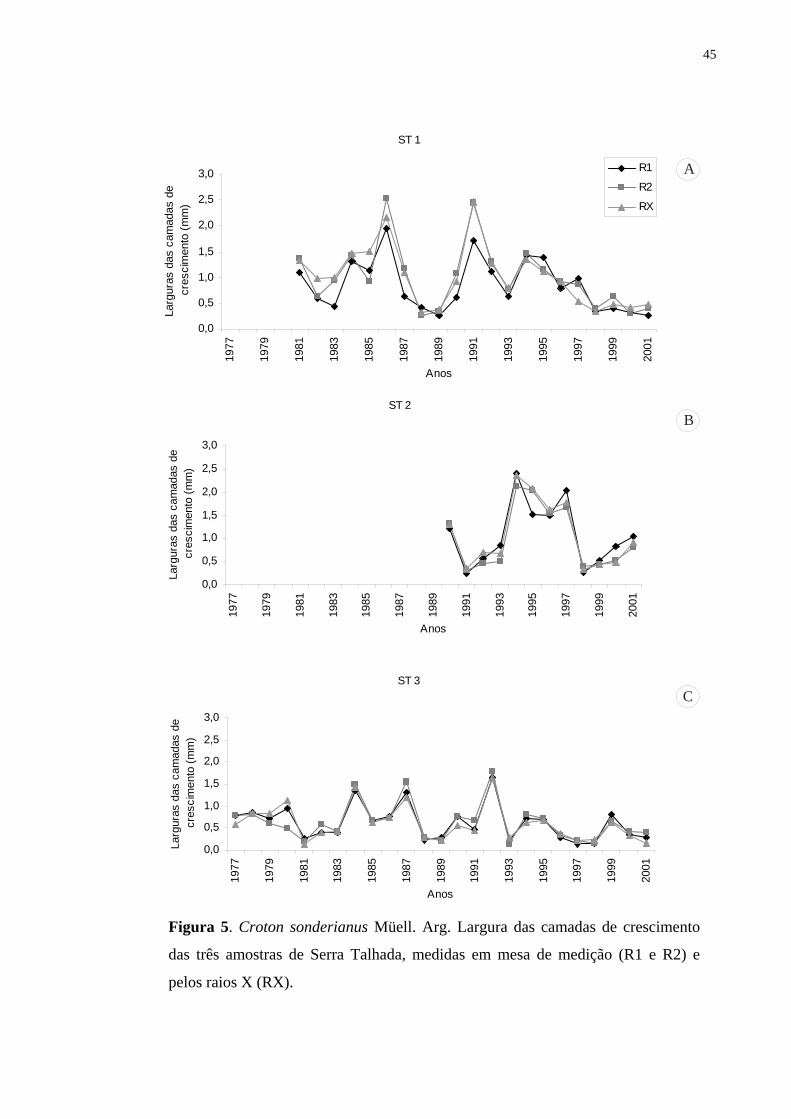
45
ST 1
0,0
0,5
1,0
1,5
2,0
2,5
3,0
1977
1979
1981
1983
1985
1987
1989
1991
1993
1995
1997
1999
2001
Anos
Larg
uras
das
cam
adas
de
cres
cim
ento
(mm
)R1
R2
RX
A
ST 2
0,0
0,5
1,0
1,5
2,0
2,5
3,0
1977
1979
1981
1983
1985
1987
1989
1991
1993
1995
1997
1999
2001
Anos
Larg
uras
das
cam
adas
de
cres
cim
ento
(mm
)
B
C
Figura 5. Croton sonderianus Müell. Arg. Largura das camadas de crescimento
das três amostras de Serra Talhada, medidas em mesa de medição (R1 e R2) e
pelos raios X (RX).
ST 3
0,0
0,5
1,0
1,5
2,0
2,5
3,0
1977
1979
1981
1983
1985
1987
1989
1991
1993
1995
1997
1999
2001
Anos
Larg
uras
das
cam
adas
de
cres
cim
ento
(mm
)
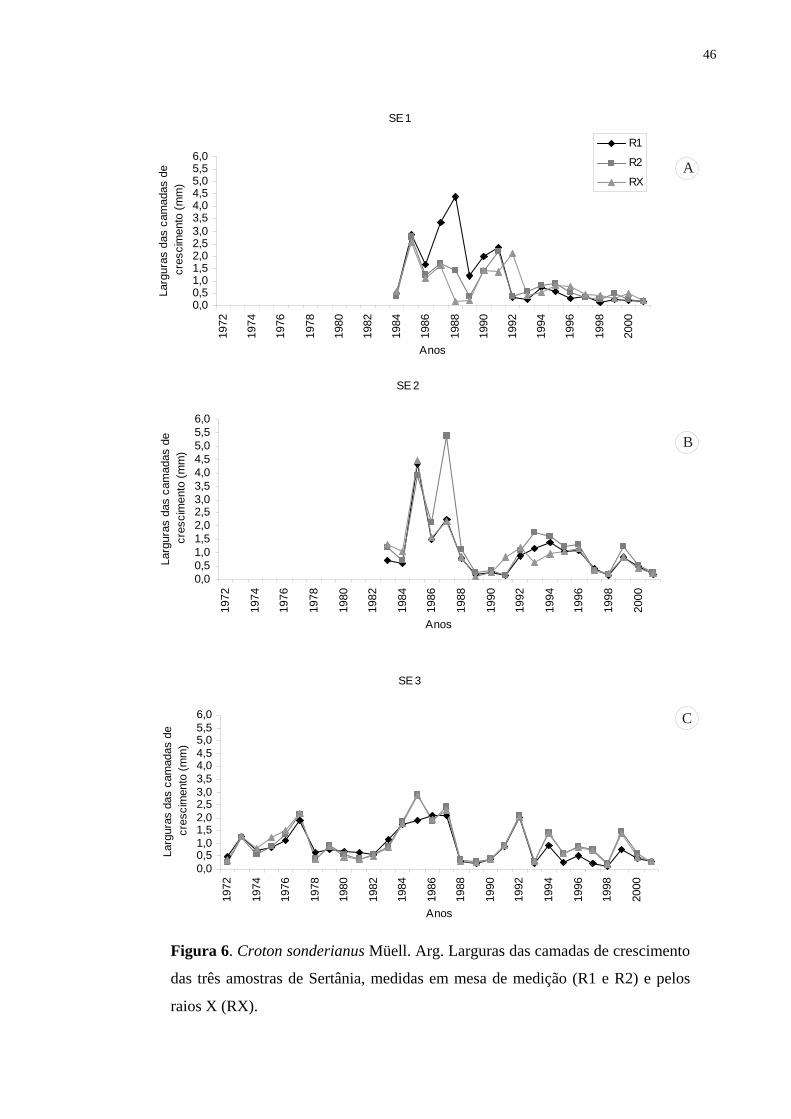
46
SE 1
0,00,51,01,52,02,53,03,54,04,55,05,56,0
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
Anos
Larg
uras
das
cam
adas
de
cres
cim
ento
(mm
)
R1
R2
RX
A SE 2
0,00,51,01,52,02,53,03,54,04,55,05,56,0
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
Anos
Larg
uras
das
cam
adas
de
cres
cim
ento
(mm
)
B
SE 3
0,00,51,01,52,02,53,03,54,04,55,05,56,0
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
Anos
Larg
uras
das
cam
adas
de
cres
cim
ento
(mm
)
C
Figura 6. Croton sonderianus Müell. Arg. Larguras das camadas de crescimento
das três amostras de Sertânia, medidas em mesa de medição (R1 e R2) e pelos
raios X (RX).
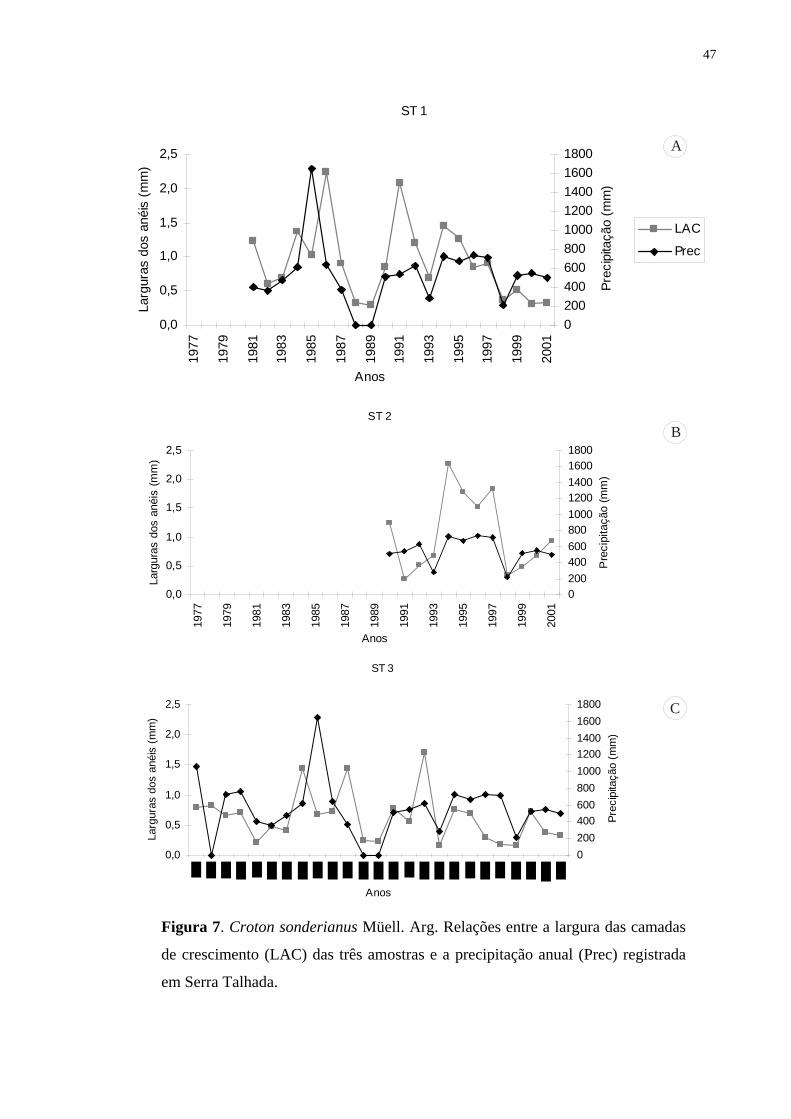
47
ST 1
0,0
0,5
1,0
1,5
2,0
2,5
1977
1979
1981
1983
1985
1987
1989
1991
1993
1995
1997
1999
2001
Anos
Larg
uras
dos
ané
is (m
m)
020040060080010001200140016001800
Prec
ipita
ção
(mm
)
LAC
Prec
A
C
Figura 7. Croton sonderianus Müell. Arg. Relações entre a largura das camadas
de crescimento (LAC) das três amostras e a precipitação anual (Prec) registrada
em Serra Talhada.
ST 2
0,0
0,5
1,0
1,5
2,0
2,5
1977
1979
1981
1983
1985
1987
1989
1991
1993
1995
1997
1999
2001
Anos
Larg
uras
dos
ané
is (m
m)
020040060080010001200140016001800
Pre
cipi
taçã
o (m
m)
ST 3
0,0
0,5
1,0
1,5
2,0
2,5
Anos
Larg
uras
dos
ané
is (m
m)
020040060080010001200140016001800
Prec
ipita
ção
(mm
)
B
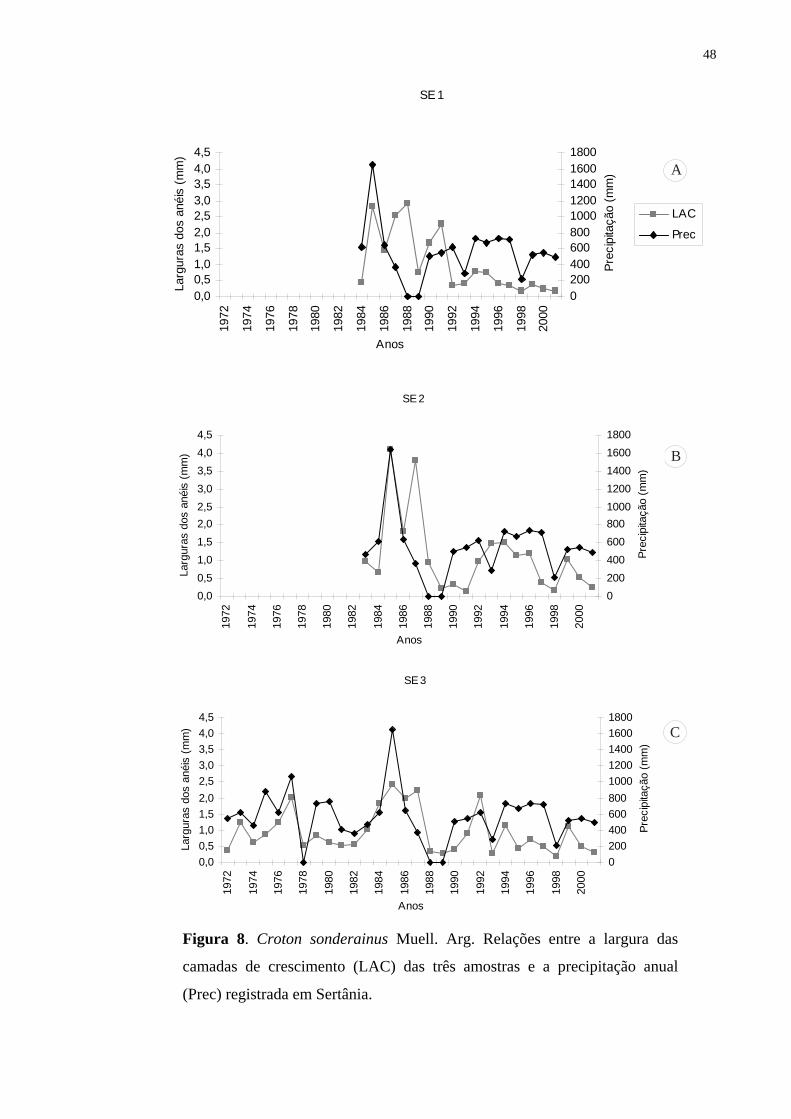
48
SE 1
0,00,51,01,52,02,53,03,54,04,5
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
Anos
Larg
uras
dos
ané
is (m
m)
020040060080010001200140016001800
Prec
ipita
ção
(mm
)
LAC
Prec
A
B
SE 2
0,00,51,01,52,02,53,03,54,04,5
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
Anos
Larg
uras
dos
ané
is (m
m)
020040060080010001200140016001800
Prec
ipita
ção
(mm
)
C
SE 3
0,00,51,01,52,02,53,03,54,04,5
1972
1974
1976
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
Anos
Larg
uras
dos
ané
is (m
m)
020040060080010001200140016001800
Prec
ipita
ção
(mm
)
Figura 8. Croton sonderainus Muell. Arg. Relações entre a largura das
camadas de crescimento (LAC) das três amostras e a precipitação anual
(Prec) registrada em Sertânia.
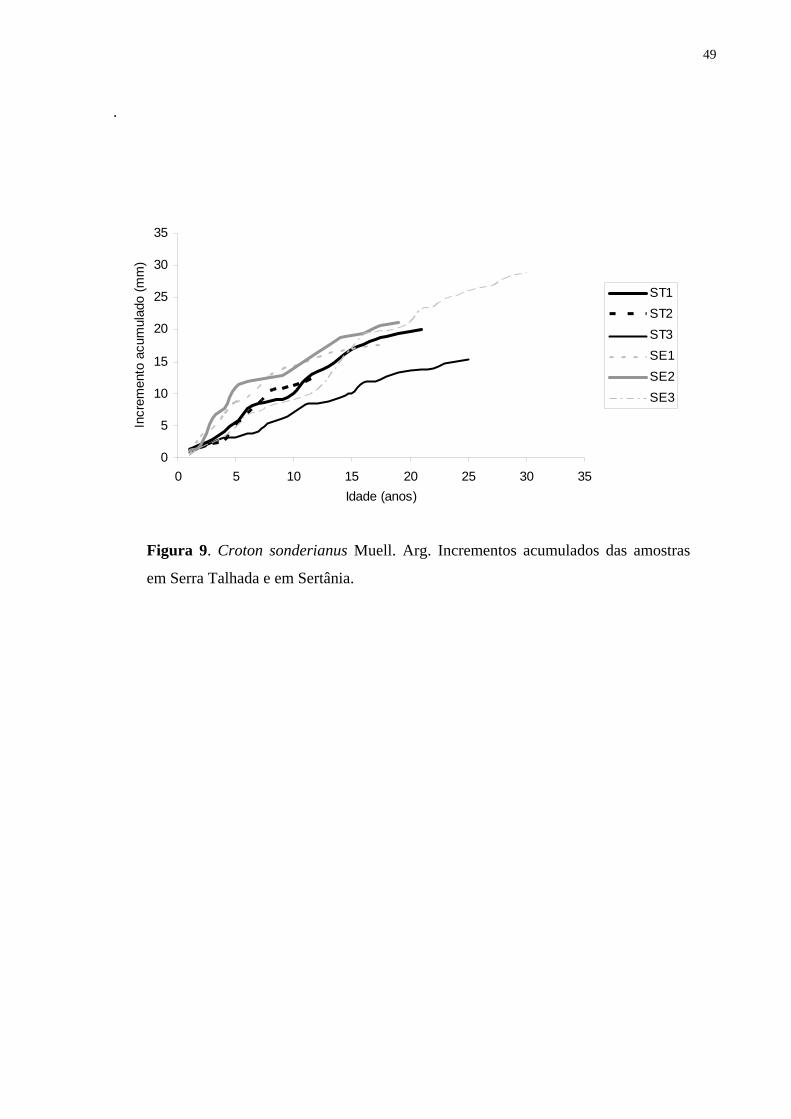
49
.
0
5
10
15
20
25
30
35
0 5 10 15 20 25 30 35Idade (anos)
Incr
emen
to a
cum
ulad
o (m
m)
ST1ST2ST3SE1SE2SE3
Figura 9. Croton sonderianus Muell. Arg. Incrementos acumulados das amostras
em Serra Talhada e em Sertânia.
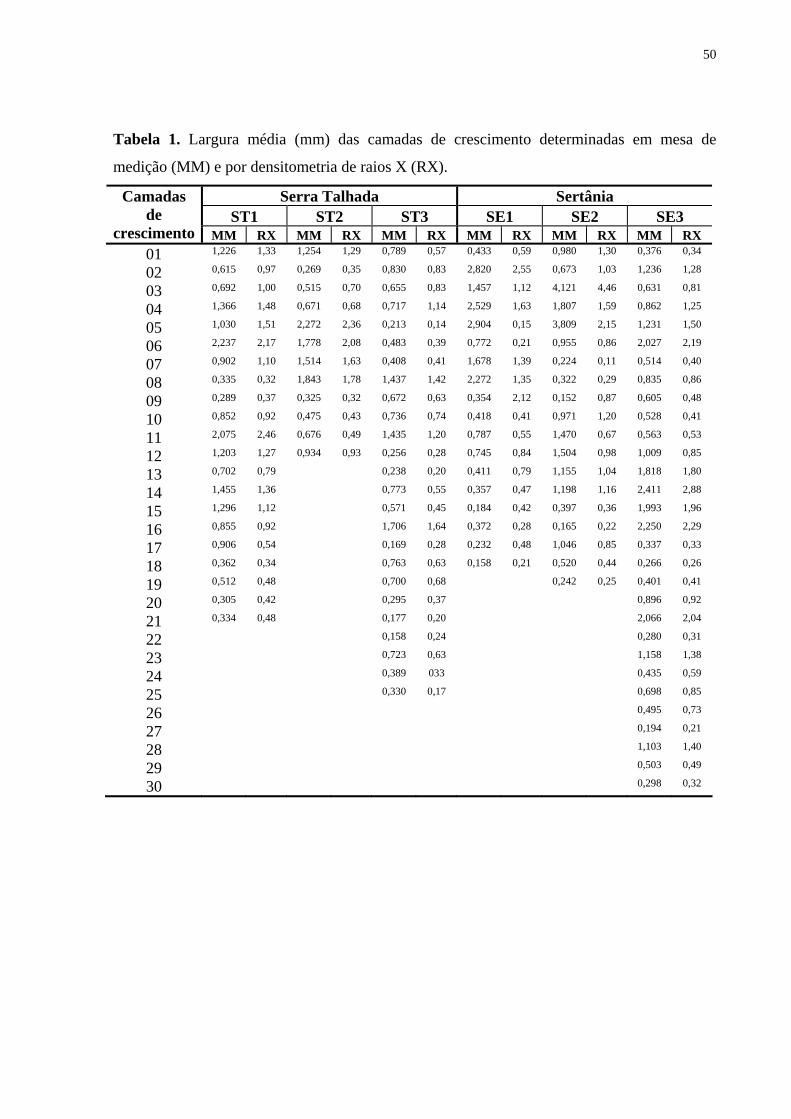
50
Tabela 1. Largura média (mm) das camadas de crescimento determinadas em mesa de
medição (MM) e por densitometria de raios X (RX).
Serra Talhada Sertânia ST1 ST2 ST3 SE1 SE2 SE3
Camadas de
crescimento MM RX MM RX MM RX MM RX MM RX MM RX 01 1,226 1,33 1,254 1,29 0,789 0,57 0,433 0,59 0,980 1,30 0,376 0,34
02 0,615 0,97 0,269 0,35 0,830 0,83 2,820 2,55 0,673 1,03 1,236 1,28
03 0,692 1,00 0,515 0,70 0,655 0,83 1,457 1,12 4,121 4,46 0,631 0,81
04 1,366 1,48 0,671 0,68 0,717 1,14 2,529 1,63 1,807 1,59 0,862 1,25
05 1,030 1,51 2,272 2,36 0,213 0,14 2,904 0,15 3,809 2,15 1,231 1,50
06 2,237 2,17 1,778 2,08 0,483 0,39 0,772 0,21 0,955 0,86 2,027 2,19
07 0,902 1,10 1,514 1,63 0,408 0,41 1,678 1,39 0,224 0,11 0,514 0,40
08 0,335 0,32 1,843 1,78 1,437 1,42 2,272 1,35 0,322 0,29 0,835 0,86
09 0,289 0,37 0,325 0,32 0,672 0,63 0,354 2,12 0,152 0,87 0,605 0,48
10 0,852 0,92 0,475 0,43 0,736 0,74 0,418 0,41 0,971 1,20 0,528 0,41
11 2,075 2,46 0,676 0,49 1,435 1,20 0,787 0,55 1,470 0,67 0,563 0,53
12 1,203 1,27 0,934 0,93 0,256 0,28 0,745 0,84 1,504 0,98 1,009 0,85
13 0,702 0,79 0,238 0,20 0,411 0,79 1,155 1,04 1,818 1,80
14 1,455 1,36 0,773 0,55 0,357 0,47 1,198 1,16 2,411 2,88
15 1,296 1,12 0,571 0,45 0,184 0,42 0,397 0,36 1,993 1,96
16 0,855 0,92 1,706 1,64 0,372 0,28 0,165 0,22 2,250 2,29
17 0,906 0,54 0,169 0,28 0,232 0,48 1,046 0,85 0,337 0,33
18 0,362 0,34 0,763 0,63 0,158 0,21 0,520 0,44 0,266 0,26
19 0,512 0,48 0,700 0,68 0,242 0,25 0,401 0,41
20 0,305 0,42 0,295 0,37 0,896 0,92
21 0,334 0,48 0,177 0,20 2,066 2,04
22 0,158 0,24 0,280 0,31
23 0,723 0,63 1,158 1,38
24 0,389 033 0,435 0,59
25 0,330 0,17 0,698 0,85
26 0,495 0,73
27 0,194 0,21
28 1,103 1,40
29 0,503 0,49
30 0,298 0,32

51
DISCUSSÃO
Massa específica aparente - Em regiões de clima tropical seco ocorre uma menor, e às vezes,
muito baixa precipitação pluviométrica. Assim, as árvores adaptam seus processos
fisiológicos para condições de menor disponibilidade de água no solo, com reflexo na
atividade do câmbio e conseqüentemente na anatomia do lenho. Entre estas adaptações
existem espécies que perdem suas folhas. Dessa maneira, pode-se relacionar esses fatos à
marcação de anéis com menores larguras, pois o câmbio diminui ou, algumas vezes, até cessa
a produção de lenho, podendo ainda produzir vasos de pequenos diâmetros. A área ocupada
por estes vasos e sua freqüência afetam diretamente a densidade do lenho de algumas espécies
(Guilley & Nepveu 2003). Croton sonderianus da caatinga do nordeste brasileiro, perde todas
suas folhas no período de deficit hídrico. Além disso, apresenta vasos de pequenos diâmetros
associados a grandes quantidades de fibras e baixa quantidade de parênquima, resultando em
altas taxas de densidade aparente variando de 700 kg/m3 a 1.250 kg/m3 (Fig. 3, 4). Em ambos
os municípios, Sertânia e Serra Talhada, a madeira da espécie em estudo apresentou anéis
com alta densidade, independente da posição radial, no sentido medula-câmbio.
A irregularidade das precipitações afeta o crescimento e o desenvolvimento das
árvores e, especificamente, a atividade cambial, induzindo a formação de regiões de alta e
baixa densidades em um anel de crescimento. Espécies de Gmelina arborea
Roxb.(Verbenaceae) de um local da Costa Rica com 1.781 mm/ano de precipitação
pluviométrica e com 12 semanas de estação seca evidencia a demarcação dos anéis de
crescimento pelo maior valor da densidade da madeira como resultado do aumento da
espessura da parede celular e pela presença de vasos de menor diâmetro (R. Roque dados não
publicados). Na caatinga, Croton sonderianus apresentou anéis mais largos quando a
precipitação foi acima de 300 mm anuais, enquanto nos anos de precipitação abaixo desse
valor verificou-se resposta no lenho através da diminuição expressiva de seus incrementos, ou
seja, anéis estreitos.
Panshin & De Zeeuw (1970) e Panshin (1980) afirmaram que a densidade é uma
propriedade física importante da madeira e que aquelas de árvores de clima tropical seco
possuem maior resistência e propriedades estruturais, recomendando-as para utilização na
construção civil devido a essas características. Segundo Paula et al. (2000), madeiras com alta
densidade são recomendadas para a produção de energia (carvão), situação esta encontrada
em Croton sonderianus.
Relação com a precipitação pluviométrica - Já está perfeitamente estabelecido que a formação
dos anéis de crescimento nas espécies de clima temperado ocorre pelas variações climáticas,

52
mais especificamente a queda de temperatura e a quantidade de luminosidade/dia durante o
outono. Porém, nas espécies tropicais a marcação dos anéis de crescimento ocorre pela
intermitência da precipitação pluviométrica durante o ano, sendo demarcado no final do anel
de crescimento o efeito do deficit hídrico no solo (Alvim 1964; Worbes 1990). Luchi (2004),
analisando Croton urucurana Baill. à margem do Rio Moji-Guaçu, São Paulo, crescendo em
diferentes níveis de umidade, observou que apenas os indivíduos da área de terreno elevado
considerada seca apresentaram delimitação de camadas de crescimento.
A literatura especializada tem apresentado dados a respeito dos anéis de crescimento
no lenho de árvores de espécies de Croton. Dentre os trabalhos mencionam-se os que não
trazem citação da presença dos anéis de crescimento em Croton sp., Croton urucurana Baill.,
Croton lanjouwensis Jablonski, Croton matourensis Aubl. e Croton argyratus Blume
(Tomazello-Filho et al. 2004; Mello 1950; Loureiro 1968; Sosef et al. 1998). Por outro lado,
Dias-Leme (dados não publicados) e Barajas-Morales et al. (1997) identificaram camadas de
crescimento demarcadas pelo achatamento e espessamento das fibras no lenho de árvores de
Croton floribundus Spreng. e em Croton nitens Sw., respectivamente. Em ambientes de clima
semi-árido as condições climáticas, a disponibilidade de água, nutrientes, luz e temperatura
afetam a atividade do câmbio, induzindo a formação de zonas de crescimento distintas no
lenho (Détienne 1989; Jacoby 1989; Vetter & Botosso 1989; Worbes 1989).
Além da precipitação, existem outros fatores relacionados e que também influenciam
na delimitação dos anéis de crescimento nas regiões tropicais como a fenologia da espécie, o
que inclui a queda de folhas durante o ano. Algumas plantas são consideradas como “sempre-
verdes” por não apresentarem queda de folhas durante o ano ou em períodos muito curtos,
induzindo a formação de anéis pouco nítidos ou até mesmo inconspícuos (Tomlinson &
Craighead 1972). Alvim (1964) observou que a fenologia das árvores mostra relação com a
atividade cambial de muitas espécies, sendo que os eventos fenológicos podem ser anuais e
em ciclos não anuais. Croton sonderianus na caatinga perde todas as folhas na estação seca,
que ocorre no período de março a outubro, característica esta que deve contribuir com a
atividade do câmbio e conseqüentemente na formação das camadas de crescimento.
Em climas tropicais com longas e severas condições de seca, a estacionalidade da
precipitação constitui o fator determinante na fenologia de árvores decíduas. Nos
semidesertos tropicais, a alternância de períodos secos e úmidos induzem a mudanças
importantes na fisionomia das florestas. Borchert (1999), na Costa Rica, observou uma
correlação entre a quantidade de água, a fenologia e a atividade cambial em árvores de
espécies decíduas, todavia menos intensa que em florestas temperadas.
Segundo Fahn et al. (1981), nos casos em que os anéis de crescimento são formados,

53
três interpretações são possíveis: (1) as zonas representam incrementos anuais; (2) as zonas
refletem periodicidades diversas de mudanças externas ou internas ocorrentes durante o ano,
ou (3) as zonas são devido a ciclos não-anuais. Os anéis de Croton sonderianus aqui
estudados parecem refletir mudanças externas da precipitação pluviométrica que podem
interferir diretamente na atividade do câmbio, resultando em camadas de crescimento mais
largas ou mais estreitas.
Algumas zonas podem ser formadas por falsos anéis (Studhalter et al. 1963). As
causas do aparecimento de falsos anéis podem ser inundações, secas, geadas, fogo, queda de
folhas ou durante a formação de ramos.
Mariaux (1967, 1969) e Détienne & Mariaux (1977) constataram que o crescimento
em diâmetro do tronco de espécies de árvores tropicais do continente africano depende
diretamente da disponibilidade de água no período de chuvas, ocorrendo na estação seca uma
redução da atividade cambial, refletindo na formação das camadas de crescimento. Détienne
(1989), Vetter & Botosso (1988) e Botosso & Vetter (1991) observaram que o ritmo de
crescimento e a atividade cambial de espécies tropicais da Amazônia reagiram ao estresse de
água do período de baixa precipitação. Eles observaram que no período da estação seca (junho
a outubro) houve redução ou ausência de incremento do tronco das árvores, com a sua
atividade cambial diminuindo ou até cessando.
Coster (1927, 1928) observou que espécies tropicais com um período de ausência de
folhas tendem a apresentar anéis de crescimento distintos. Amburana cearensis (Allem) A. C.,
perde as folhas anualmente e isto está relacionado com a presença de anéis de crescimento
visíveis e anuais (Schwyzer 1988). Tomazello-Filho & Cardoso (1999) observaram em
Tectona grandis L. f. que o câmbio manifesta períodos de máxima e mínima atividade em
fase de mudanças fenológicas e variações climáticas estacionais. No semi-árido do nordeste
do Brasil, a maioria das plantas apresenta queda de folhas e diminuição do ritmo de
crescimento durante o período de seca de maio à outubro (Ratisbona 1976).
Espécies da caatinga nordestina, como Caesalpinia pyramidalis Tul. e Cnidoscolus
phyllacanthus (M. Arg.) Pax et K. Hoffin, reiniciam sua atividade vegetativa ao começar o
período de chuvas e evidenciam uma grande variabilidade anual e na largura de seus anéis,
relacionado a intensidade e extensão temporal da época seca (A. Tsuchiya & M. Nemoto
dados não publicados). Mimosa acutistipula Benth. e Astronium urundeuva Engl. formam
anéis de crescimento mais largos e com maior número de vasos de condução quando o ano se
apresenta com um elevado conteúdo de água nos solos, enquanto que em anos secos, tanto a
largura do anel como o número de vasos diminuem (Tsuchiya 1995). Este autor, analisando a
relação entre o crescimento dos vasos em arbustos e o balanço hídrico no semi-árido do

54
nordeste brasileiro, observou que o número de vasos foi maior nos anos úmidos e menor nos
anos secos, refletindo-se na largura dos anéis. Aspidosperma pyrifolium Mart. e Mimosa
hostilis Benth. também apresentaram uma forte correlação entre a largura dos anéis de
crescimento e a precipitação que se produz durante os meses de maior déficit de água (maio a
outubro) na região semi-árida do nordeste do Brasil (Tsuchiya 1988). Assim, pode-se
observar que na caatinga o desenvolvimento vegetativo de diversas espécies é inversamente
proporcional a variação do déficit anual de água (Tsuchiya 1990). Isto prova que o
crescimento está regulado pela intensidade de períodos de seca na região do sertão brasileiro
(Roig 2000).
Nos bosques chuvosos do Brasil, 35% das árvores apresentam anéis de crescimento
presumivelmente relacionados com a ocorrência de secas estacionais (Mainieri 1959,1978).
Croton sonderianus aqui estudada demonstrou claramente a formação dos anéis de
crescimento inversamente proporcional à disponibilidade de água, a qual foi baseada na
precipitação pluviométrica anual. Nos anos de mais chuva os anéis formados foram mais
largos. Rao & Rajput (1999) observaram que há uma relação entre a atividade cambial e os
fatores climáticos em árvores de Tectona grandis. Nesta espécie, a temperatura do ar e a
precipitação agem como fatores interdependentes sobre o câmbio, o qual terá sua máxima
divisão celular quando a precipitação for intensa e as temperaturas estiverem em torno de
35oC, ficando dormente durante a estação seca. Assim, podemos verificar que os fatores
climáticos provocam variações na estrutura anatômica do lenho. A espécie Croton
sonderianus, conforme observado nos resultados aqui obtidos, apresentou comportamento
semelhante em relação à precipitação anual, no qual o câmbio vascular apresentou dormência
no período seco e atividade durante o período chuvoso.
Incremento – L. Ferreira-Fedele (dados não publicados), analisando árvores de diferentes
famílias, incluindo Euphorbiaceae, observou que no período chuvoso o incremento foi maior
quando comparado com o período de seca, em que houve diminuição e/ou a cessação da
atividade cambial. Détienne (1989) na Guiana Francesa, Vetter & Botosso (1988, 1989) e
Botosso & Vetter (1991) na Amazônia e Worbes (1999) na Venezuela, encontraram este
mesmo tipo de comportamento nas árvores, nas quais no período de baixos índices de
precipitação pluviométrica ocorreu a diminuição ou até mesmo a cessação da atividade
cambial. Croton sonderianus, tanto em Serra Talhada quanto em Sertânia, manifestou
comportamento semelhante evidenciando a estreita relação da variação da precipitação
pluviométrica e a atividade cambial expressa na formação de diferentes larguras dos seus
anéis de crescimento.

55
CONCLUSÕES
Croton sonderianus apresentou a capacidade de formar, em seu tronco, anéis de
crescimento indicando seu grande potencial para estudos dendrocronológicos nos trópicos.
Tais estudos são indispensáveis para o conhecimento da dinâmica do ecossistema, dos
processos de regeneração, e para a aplicação de planos de manejo sustentado em áreas de
florestas tropicais.
A presença de delimitação de camadas de crescimento em Croton sonderianus refletiu
o período de seca pelo qual a região passa.
A formação e o tamanho dos anéis de crescimento nessa espécie na caatinga estão
relacionados à precipitação pluviométrica e variam no mesmo sentido, conforme os resultados
apresentados. Assim, anéis mais largos são formados durante os anos de maior
disponibilidade de água e anéis mais estreitos, nos anos de baixa precipitação considerados
períodos de seca (abaixo de 300 mm anual). Inclusive esse fato foi corroborado pelas
correlações obtidas entre as medidas de incremento anual do lenho e o histórico de
precipitações que demonstrou se tratarem de anéis anuais de crescimento.
Croton sonderianus da caatinga apresentou anéis anuais de crescimento.
Recomendam-se que estudos semelhantes sejam realizados com outras espécies da
caatinga nordestina mais longevas, a fim de se obter outros resultados com diferentes espécies
da caatinga e assim uma discussão mais ampla quanto a relação da pluviosidade e os anéis de
crescimento anuais nesse ecossistema.

56
REFERÊNCIAS BIBLIOGRÁFICAS
Allem, A.C. 1979. Quatro novas espécies de Croton L. (Euphorbiaceae) do Rio Grande do
Sul, Brasil. Boletim da Sociedade Argentina de Botánica 18(3-4): 63-81.
Alvim, P.T. 1964. Tree growth peridicity in tropical climates. In: M.H. Zimmermann (ed.).
The formation of wood in forest trees. New York, Academic Press.
Alvim, P.T. & Alvim, R. 1976. Relation of climate to growth periodicity in tropical trees.
Pp.80-102. In: P.B. Tomlinson e M.H. Zimmermann (eds.). Tropical trees as living
systems. Cambridge, Cambridge University Press.
Amaral, A.C.B. & Tomazello-Filho, M. 1998. Avaliação das características dos anéis de
crescimento de Pinus taeda pela técnica de microdensitometria de raios X. Revista
Ciência e Tecnologia 6(11-12): 17-23.
Baas, P. & Vetter, R.E. 1989. Growth rings in tropical trees. IAWA Bulletin new series
10(2): 95-184.
Barajas-Morales, J.; Álvarez, G.A. & Sánchez, P.S. 1997. Anatomía de maderas de México:
espécies de una selva alta perennifolia I. Publicaciones especiales Instituto de Biología
de la Universidad Nacional Autonoma de Mexico 16: 46-47.
Borchert, R. 1999. Climatic periodicity, phenology and cambium activity in tropical dry forest
trees. IAWA Journal 20(3): 239-247.
Botosso, P.C. & Vetter, R.E. 1991. Alguns aspectos sobre a periodicidade e taxa de
crescimento em oito espécies arbóreas tropicais de floresta de terra firme (Amazônia).
Revista do Instituto Florestal 3(2): 163-180.
Coster, C. 1927. Zur anatomie und physiologie der zuwachszonen und jahrringbildung in den
tropen. Annals der Botanik Garde der Buitenzorg 37: 49-160.
Coster, C. 1928. Zur anatomie und physiologie der zuwachszonen und jahrringbildung in den
tropen. Annais do Jardim Botanik Buitenzorg 38: 1-44.
Détienne, P. 1989. Appearance and periodicity of growth rings in some tropical woods.
IAWA Bulletin new series 10(2): 123-132.
Détienne, P. & Mariaux, A. 1977. Nature et periodicité des cernes dans le bois rouges de
Meliacées africanes. Bois et Forests des Tropiques, Nogent-sur-Marne 175: 53-61.
Fahn, A.; Burley, J.; Longman, K.A.; Mariaux, A. & Tomlinson, P.B. 1981. Possible

57
contributions of wood anatomy to the determination of the age of tropical trees. Pp. 31-
54.In: Bormann, F. H. & Berlyn, G. Age and growth rate of tropical trees: new
directions for research. New Haven, Yale University. Bulletin 94.
Ferreira-Fedele, L; Tomazello-Filho, M.; Botosso, P.C. & Giannotti, E. 2004. Periodicidade
do crescimento de Esenbeckia leiocarpa Engl. (guarantã) em duas áreas da região Sudeste
do Estado de São Paulo. Scientia Forestalis 65: 141-149.
Govaerts, R; Frodin, D.G. & Radclif-Smith, A. 2000. A world checklist and bibliography of
Euphorbiaceae (and Pandaceae). Kew, Royal Botanic Gardens.
Guilley, E. & Nepveu, G. 2003. Interprétation anatomique des composantes d'un modèle
mixte de densité du bois chez le Chêne sessile (Quercus petraea Liebl.): âge du cerne
compté depuis la moelle, largeur de cerne, arbre, variabilité interannuelle, duraminisation.
Annales du Science Forst 60: 331-346.
Jacoby, G.C. 1989. Overview of tree-ring analysis in tropical regions. IAWA Bulletin new
series 10(3): 219-332.
Lima, J.L.S. 1996. Plantas forrageiras das caatingas: usos e potencialidades. Petrolina,
EMBRAPA, CPATSA, PNE & RBG-KEW.
Loureiro, A.A. 1968. Contribuição ao estudo anatômico das madeiras de Croton lanjouwensis
(Müell. Arg.) Jablonski e Croton matourensis Aublet (Euphorbiaceae). CNP/INPA
Botânica 24: 2-17.
Luchi, A.E. 2004. Anatomia do lenho de Croton urucurana Baill. (Euphorbiaceae) de solos
com diferentes níveis de umidade. Revista Brasileira de Botânica 27(2): 271-280.
Mainieri, C. 1959. Identificação das principais madeiras comerciais no Brasil. Instituto de
Pesquisas Tecnológicas, São Paulo, Boletim. 46: 1-189.
Mainieri, C. 1978. Fichas de características das madeiras brasileiras série: estado de São
Paulo. Instituto de Pesquisa Tecnológica nº 966.
Mariaux, A. 1967. Les cernes dans les bois tropicaux africains, nature et periodicité, Nogent-
sur-Marne. Bois et Forets dês Tropiques 113: 3-14.
Mariaux, A. 1969. La periodicité des cernes dans le bois de Limba, Nogent-sur-Marne. Bois
et Forets dês Tropiques 128: 39-53.
Mello, E.C. 1950. Estudo dendrológico de essências florestais do parque nacional de Itatiaia e
os caracteres anatômicos de seus lenhos. Boletim do Parque Nacional de Itatiaia 2: 1-
60.

58
Mothe, F.; Duchanois, G.; Zannier, B.; Leban, J.M. 1998. Microdensitometric analysis of
wood samples: data computation method used at Inra-ERQB (CERD program). Annals
des Foresters Sciences 55(3): 301-313.
Panshin, A.J. & De Zeeuw, C. 1970. Text book of wood technology: Structure,
identification, uses and proprieties of the commercial woods of the United States and
Canada. Boston, McGraw-Hill Book v. 1.
Panshin, A.J. 1980. Text book of wood technology. New York, McGraw-Hill Book.
Paula, J.E.; Silva-Júnior, F.G. da; Silva, A.P.P. 2000. Caracterização anatômica de madeiras
nativas de matas ciliares do centro-oeste brasileiro. Scientia Forestalis 58: 73-89.
Polge, H. 1963. Une nouvelle méthode de détermination de la texture du bois: I’analyse
densitométrique de clichês radiographiques. Annals of Science Forest 20(1): 533-580.
PROBIO, 2000. Seminário sobre avaliação e identificação de ações prioritárias para a
conservação, utilização sustentável e repartição de benefícios da biodiversidade do bioma
caatinga. Disponível em: http://www.biodiversitas.org/caatinga. Acesso em: 20 jul.
2001.
Ramos, R.F.B. & Lagos, J.L.M. 1988. Catalogo de espécies forestales de Honduras:
distribuitión, propriedades y usos. Siguatepeque, Escuela Nacional de Ciencias
Forestales.
Rao, K.S. & Rajput, K.S. 1999. Seasonal behaviour of vascular cambium in teak (Tectona
grandis) growing in moist deciduous and dry deciduous forests. IAWA Journal 20(1): 85
– 93.
Ratisbona, L.R. 1976. The climate of Brazil. Pp. 219-293. In: W. Schwerdtfeger (ed.),
Climates of Central and South América. Oxford, Elsevier.
Raven, P.H., Evert, R.F. & Eichhorn, S.E. 2001. Biologia Vegetal. 6a ed. Rio de Janeiro, Ed.
Guanabara Koogan.
Record, S.J. & Hess, R.W. 1949. Timbers of the New World. New Haven, Yale University
Press.
Rodal, M.J.N. & Melo, A.L. de 1999. Levantamento preliminar das espécies lenhosas da
caatinga de Pernambuco . Pp. 53-62 In: F. D. de Araújo; H. D. Prendergast & S. J. Mayo
(eds.). Plantas do Nordeste – Anais do I Workshop Geral, RBG, KEW, PNE, CNPq.
Recife, APNE.
Roig, F. 2000. Dendrocronología en los bosques del Neotrópico revisión y prospección futura.

59
Pp. 307-355. In: F.A. Roig. Dendrocronologia em America Latina. Mendonza,
EDIUNC.
Sampaio, E.V.S.B. 2002. Uso das plantas da caatinga. Pp. 49-68. In: Everardo V.S.B.
Sampaio; A.M. Giulietti; J. Virgílio & C.F.L. Gamarra-Rojas (ed.). Vegetação e flora da
caatinga. Recife, APNE & CNIP.
Schulz, J.P. 1960. The vegetation of Surinam. II. Ecological studies on rain forest in
northern Surinam. Amsterdam, van Eedenfonds.
Schwyzer, A. 1988. Anillos de crecimiento em cuatro árboles tropicales. Boletin de Lima 58:
83-86.
Schweingruber, F.H. 1988 Tree rings, basics and applications of dendrochronology.
Dordrecht, Reidel.
Sosef, M.S.M.; Hong, L.T. & Prawirohatmodjo, S. 1998. Plant resources of south-east Asia:
5(3): Timber trees: Lesser-known timbers. Leiden, Backhuys Publishers.
Studhalter, R.A.; Glock, W.S. & Agerter, S.R. 1963. Tree growth: some historical chapters.
Botanical Review 29: 245-365.
Tomazello-Filho, M. & Cardoso, N.S. 1999. Seasonal variations of the vacular cambium of
Teak (Tectona grandis L. f.) in Brazil. Pp. 147-154. In: R. Wimmer & R. E. Vetter (ed.).
Tree-ring analysis: biological, methodological and environmental aspects. London,
CABI.
Tomazello-Filho, M.; Botosso, P.C. & Lisi, C.S. 2001. Análise e aplicação dos anéis de
crescimento das árvores como indicadores ambientais: dendrocronologia e
dendroclimatologia. Pp. 117-143. In: N. B. Maia; H. L. Martos; W. Barrella (ed.).
Indicadores ambientais: conceitos e aplicações. São Paulo, EDUC/COMPED/INEP.
Tomazello-Filho, M; Lisi, C.S.; Hansen, N.; Cury, G. 2004. Anatomical features of increment
zones in different tree species in the state of São Paulo, Brazil. Scientia Forestalis 66: 46-
55.
Tomlinson, P.B. & Craighead, F.C. 1972. Growth-ring studies on native trees of subtropical
Florida. Pp. 39-51. In: A. K. M. Ghouse & M. Yunus (eds.). Research trends in plant
anatomy. New Delhi, McGraw-Hill Publishing Co.
Tsuchiya, A.1988. Water balance in northeast Brazil. Latin American Studies 10: 47-59.
Tsuchiya, A.1990. Hypertrophic growth of trees of the caatinga plant community and water
balance. Latin American Studies 11: 51-70.

60
Tsuchiya, A. 1995. Preliminary study on the relationship between vessel growth of thorny
shrubs and water balance in the semi-arid region, northeastern Brazil. Geographical
Sciences 50: 123-131.
Vetter, R.E. & Botosso, P.C. 1988. Observações preliminares sobre a periodicidade e taxa de
crescimento em árvores tropicais. Acta Amazonica 18(1-2): 189-195.
Vetter, R.E. & Botosso, P.C. 1989. Remarks on age and periodicity of growth rings in some
tropical woods. IAWA Bulletin new series 10(2): 133-146.
Vetter, R.E. & Botosso, P.C. 1993. A idade de árvores da Amazônia. Pp. 405-417. In. E.J.G.
Ferreira; G.M. Santos; E.L. Leão & L.A. Oliveira (eds.). Bases científicas para
estratégias de preservação e desenvolvimento da Amazônia (2). Manaus, INPA.
Webster, G.L. 1944. A synopsis of the genera and suprageneric taxa of Euphorbiaceae.
Annals Missouri Botanical Garden 81(1): 33-144.
Wehr, N.J. & Tomazello-Filho, M. 2000. Caracterização dos anéis de crescimento de árvores
de Araucaria angustifolia (Bert.) O. Ktze, através da microdensitometria de raios X.
Scientia Forestalis 58: 161-170.
Worbes, M. 1989. Growth rings, increment and age of trees in inundation forests, savannas
and mountain forest in Neotropics. IAWA Bulletin 10(2): 109-122.
Worbes, M. 1990 Dendrochronology studies on tropical trees. Plant Research and
Development 32: 86-98.
Worbes, M. 1995. How to measure growth dynamics in tropical trees – a review. IAWA
Journal 16(4): 337-351.
Worbes, M. 1999. Annual growth rings, rainfall-dependent growth and long-term growth
pattern of tropical trees from Caparo Forest Reserve in Venezuela. Journal of Ecology
87: 391-403.

CAPÍTULO 3
ANATOMIA E DENSIDADE BÁSICA DA MADEIRA DE CAESALPINIA
PYRAMIDALIS TUL. (LEGUMINOSAE-CAESALPINIOIDEAE),
ESPÉCIE ENDÊMICA DA CAATINGA DO NORDESTE DO BRASIL

62
RESUMO
Este trabalho objetivou estudar a anatomia e a densidade básica da madeira de
Caesalpinia pyramidalis ocorrente na caatinga de Pernambuco, nos municípios de Serra
Talhada e Sertânia, utilizada pelas comunidades locais, assim como comparar a percentagem
dos elementos do lenho no tronco e nos galhos nos dois municípios, e radialmente na própria
árvore, a fim de estabelecer o potencial total do lenho para produção de energia. As amostras
do lenho do tronco (DAP) e de ramos de seis árvores da espécie foram coletadas nos
municípios acima citados, em 2002. As análises das amostras seguiram os métodos
usualmente empregados em estudo de anatomia de madeiras. Pelos parâmetros anatômicos do
lenho e a elevada densidade básica (>0,84 g/cm3), concluiu-se que Caesalpinia pyramidalis
tem potencial para geração de energia, notadamente, álcool combustível, carvão vegetal e
lenha como fonte primária de energia em fornos. Pode-se propor o uso como combustível do
lenho do tronco e dos galhos, desde a fase do lenho juvenil, pela grande concentração de
fibras, menor concentração de vasos e menor quantidade de parênquima nesta fase, nos dois
municípios.
Palavras-chaves: Caesalpinia, anatomia e densidade da madeira, caatinga, carvão.
ABSTRACT
This work aimed to study the anatomy and basic density of Caesalpinia pyramidalis wood,
used by local populations in the municipality of Serra Talhada and Sertânia (Pernambuco), to
assess the percentage of wood elements in the trunk and branches in both locality, and radially
in the tree itself, in order to establish the total wood potential for energy production. Samples
of the trunk (DBH) and of branches from six trees of the species were collected in 2002. The
analyses of the samples dealt with the usual methods in wood anatomy study. Through the
anatomic parameters of the trunk and the high basic density (>0,84 g/cm3), so the wood of
Caesalpinia pyramidalis presents an enormous amount of cellulose and lignin. Due to these
features, this species has shown to have sure perspectives for the production of alcohol and
charcoal. It’s possible to propose the use of its trunk and branches as fuel since juvenile wood
because of the great concentration of fibers, smaller concentration of vessels and smaller
quantity of parenchyma at this stage in both municipalities.
Kew words: Caesalpinia, anatomy and density wood, caatinga, charcoal.

63
INTRODUÇÃO
A exploração predatória de madeira aumenta continuamente no mundo, urgindo
medidas para a implementação de programas para a conservação e manejo sustentável dessas
espécies. Cerca de 200.000 km2 de matas naturais foram substituídas por culturas agrícolas
e/ou pastagens, além da exploração de madeiras de espécies nativas utilizadas como carvão
para uso doméstico e industrial (Sampaio 2002). Juntamente a isto, iniciou-se no País a
introdução de espécies de rápido crescimento visando à substituição da madeira de espécies
nativas. Existe uma gama de pesquisas com essas espécies, especialmente as dos gêneros
Eucalyptus L´Hér. e Pinus L., inclusive a análise da qualidade de suas madeiras. Entretanto,
as pesquisas com as espécies nativas não têm tido o mesmo desenvolvimento, faltando dados
especialmente das espécies de menor porte, como as plantas da caatinga.
Uma grande preocupação que vem sendo colocada como fundamental no setor
florestal é a chamada qualidade intrínseca da madeira destinada à fabricação de carvão
vegetal. Isso tem feito surgir a possibilidade de que as propriedades anatômicas, físicas e
químicas da madeira também possam ser trabalhadas e conseqüentemente melhoradas (Brito
& Barichelo 1979). Em geral, a relação das fibras e a qualidade da madeira são os destaques
no estudo da madeira, porém, segundo Sárkáni et al. (1957) e Nguyen (1977) outros
componentes estruturais das angiospermas podem influenciar na qualidade da madeira, tais
como as dimensões, proporções e distribuição dos vasos, parênquima axial e radial,
requerendo uma maior investigação. Prior & Cutler (1996) desenvolveram estudos
anatômicos com espécies nativas do Zimbabwe, sul da África, onde 90% da energia
consumida pela comunidade rural provem dessas madeiras.
O presente trabalho faz parte de um dos resultados do projeto interinstitucional do
Programa Plantas do Nordeste, Subprograma Botânica Econômica intitulado “Manejo
Sustentado da Vegetação Lenhosa da Caatinga, com Ênfase à Produção de Lenha para uso
Doméstico das Comunidades do Nordeste do Brasil”, envolvendo as seguintes instituições:
Associação Plantas do Nordeste (APNE); Conselho Nacional de Desenvolvimento Científico
e Tecnológico (CNPq); Royal Botanic Gardens, Kew (RBG Kew, Inglaterra); Empresa
Pernambucana de Pesquisa Agropecuária (IPA), Universidade Estadual de Feira de Santana
(UEFS) e Universidade Federal da Bahia (UFBA). Nesse contexto, incluiu-se o estudo da
descrição anatômica da madeira de Caesalpinia pyramidalis Tul., conhecida popularmente
como catingueira, utilizada pelas comunidades pernambucanas de Serra Talhada e Sertânia
para a obtenção de lenha e carvão.
Segundo Polhill (1994), a subfamília Caesalpinioideae compreende cerca de 158

64
gêneros, e segundo Schrire et al. (2005), são 171 gêneros e cerca de 2.250 espécies tropicais e
subtropicais. A espécie Caesalpinia pyramidalis tem hábito arbóreo de porte médio, sem
espinhos, com 4-6 m de altura, podendo atingir até 12 m. A casca das árvores adultas é de cor
cinza-claro, às vezes castanho, com manchas de cor amarelo, verde e branco. Libera a camada
superficial em lâminas pouco alongadas. A madeira é branco-amarelada com cerne escuro,
muito pesada, com densidade de 0,99 g/cm3 de madeira seca, contendo grandes quantidades
de celulose e lignina. A cinza da madeira tem elevado teor de potássio e é usada para
fabricação de sabão (Maia 2004).
É uma das espécies de mais ampla dispersão no nordeste semi-árido podendo ser
encontrada em diversas associações vegetais. Ocorre nos Estados do Piauí, Ceará, Rio Grande
do Norte, Paraíba, Pernambuco, Alagoas, Sergipe e Bahia, sendo considerada endêmica na
caatinga. A madeira da catingueira é usada como lenha, carvão, estacas, mourões, na
construção de casas de taipas e pode ser utilizada para produção de álcool combustível e
coque metalúrgico. O chá da casca é utilizado para tratamento de hepatite e anemia. Além
disso, a espécie é indicada para a primeira e a segunda fase de recomposição florestal mista de
áreas degradadas (Maia 2004).
Este trabalho objetivou estudar a anatomia e a densidade básica da madeira de
Caesalpinia pyramidalis utilizada pelas comunidades locais ocorrente na caatinga de
Pernambuco, nos municípios de Serra Talhada e Sertânia, descrever a variação radial, sentido
medula-câmbio, comparar a percentagem dos elementos do lenho no tronco e nos galhos nos
dois municípios e radialmente na própria árvore, a fim de estabelecer o potencial total do
lenho para produção de energia.

65
MATERIAL E MÉTODOS
Áreas de estudo - Foram selecionadas áreas de caatinga nas Estações Experimentais da
Empresa Pernambucana de Pesquisa Agropecuária (IPA-PE), em Serra Talhada (Fazenda
Saco – 3.200 ha) e em Sertânia (Fazenda Cachoeira – 630 ha), com vasta ocorrência de
árvores de Croton sonderianus, por não sofrerem intervenção humana durante a execução do
trabalho, pela necessidade do uso sustentável da vegetação, por encontrarem-se em um raio de
200 km e apresentarem semelhanças florísticas e por estarem próximas de comunidades locais
com necessidade de carvão e de lenha.
Serra Talhada localiza-se na microrregião do Pajeú, a 418 km da capital (Recife), com
latitude de 07o53’57”S e 38o18’09”W e altitude de 500 msnm, apresentando solo maciço de
gnais superficial, marrom claro, ácido, franco argiloso com algum cascalho. A precipitação
varia de 450-750 mm, de março a maio e a temperatura média anual é de 37o C. Trata-se de
extensa área de caatinga semi-densa, madura, pouco perturbada, apresentando extrato arbóreo
com algumas árvores de até 12 m de altura, arbustivo e herbáceo bastante desenvolvidos. A
área experimental foi de 150 ha, região denominada Pimenteira (PROBIO 2000).
Sertânia localiza-se na microrregião de Moxotó, a 316 km da capital, com latitude de
08o 04’25”S e 37o12’24”W e altitude de 611 msnm, apresentando solo superficial, marrom
claro, ácido, franco argiloso com algum cascalho. A precipitação varia de 350-600 mm, de
março a maio e a temperatura média anual é de 35o C. Trata-se de uma área extensa de
caatinga, com vegetação mais fechada na encosta, árvores e arbustos maduros, pouco
perturbada. A área experimental foi de 38 ha, conhecida como Manga do Cerecé (PROBIO
2000).
Estudo anatômico e densidade do lenho - Para o estudo anatômico da madeira foram coletadas
amostras circulares do tronco de três árvores ao nível do diâmetro a altura do peito (DAP =
1,30 m) do tronco e de dois galhos de cada uma das áreas de estudo, com diferentes
diâmetros.
Foram preparadas lâminas histológicas permanentes contendo uma seção transversal e
duas longitudinais tangencial e radial do lenho, com 18-25 µm de espessura utilizando-se
micrótomo de deslize Spencer. As seções do lenho foram clarificadas com hipoclorito de
sódio (50%) e coradas com azul de alcião e safranina alcoólica (50%) 1:1, tratadas em série
alcoólica de 50 a 100%, a cada 10% e montadas em euparol (Johansen 1940). Nas seções
transversais do lenho, em raio contíguo ininterrupto, no sentido da medula para o câmbio,
foram mensurados os elementos de vaso, fibras e parênquima radial e longitudinal, utilizando-
se o programa de análise de imagem KS 300 da Carl Zeiss, no Jodrell Laboratory do Royal

66
Botanic Gardens Kew. Na descrição dos parâmetros anatômicos, foram aplicadas as normas
da IAWA Committee (1989). As lâminas histológicas permanentes foram depositadas na
Xiloteca do Instituto de Biologia da UFBA, laboratório de micromorfologia da UEFS e do
Laboratório de Micromorfologia do Jodrell Laboratory, RBG Kew.
Para a análise da ultraestrutura dos elementos de vasos em microscopia eletrônica de
varredura (MEV) foram utilizadas amostras de madeiras de 1 x 1cm, no sentido longitudinal à
altura da região mediana entre o alburno e o cerne. Estes cortes foram montados nos porta-
espécimes com fita adesiva de grafite e posteriormente recobertos com ouro em sputter
Balzers SCD 050. As amostras assim preparadas foram analisadas e elétron-
fotomicrografadas em microscópio eletrônico de varredura LEO 1430VP.
Na determinação da densidade do lenho, se utilizou amostras da região próxima da
medula e da região próxima do câmbio do DAP do tronco e de dois galhos. Essa densidade foi
determinada pela razão da massa seca do lenho (seco a 105o C)/volume saturado, de acordo
com o método descrito por Vital (1984). Para a comparação das médias e análises de
correlação, utilizaram-se os testes t de Student e o Spearman Rank Correlation,
respectivamente.

67
RESULTADOS
A média do diâmetro dos troncos dos espécimes estudados foi 4,8 cm em Sertânia e
8,4 cm em Serra Talhada. Os galhos com menor diâmetro médio (Galhos 1) apresentaram
com 3,2 cm, enquanto os mais espessos (Galhos 2) apresentaram-se com 4,0 cm nos
espécimes amostrais da área de Sertânia; em Serra Talhada, os galhos mais finos (1) e os mais
grossos (2) apresentaram, respectivamente, 2,4 cm e 2,8 cm de diâmetro médio.
Descrição anatômica – O lenho apresenta camadas de crescimento delimitadas por finas
linhas de parênquima marginal descontínuas com uma a duas células de largura que
tangenciam faixas de parênquima. Vasos solitários (maioria), múltiplos e em arranjos radiais
de 2-4 elementos, porosidade difusa (Fig. 1-4); pontoações intervasculares alternas,
guarnecidas (Fig. 5-7); placa de perfuração simples (Fig. 6). Pontoações raio-vasculares
semelhantes às intervasculares em forma e tamanho. Fibras de paredes muito espessas com
lúmen às vezes quase colapsado (Fig. 1-4); pontoações simples oblíquas; presença de fibras
gelatinosas (Fig. 4). Parênquima axial aliforme e confluente às vezes em confluência (Fig. 1-
4). Raios estratificados, homogêneos, unisseriados e bisseriados 1-2 células de largura com
(11-)15,7(-20) µm e 9-10 células de altura com (124,7-)136,4(-147,8) µm (Fig. 8, 9). Cristais
prismáticos em células do parênquima axial (Fig. 5, 10).
Tronco - Aplicando-se o teste t (P<0,05) para comparação de médias, não se observou
diferença significativa entre as amostras de Sertânia e Serra Talhada em relação à
percentagem de vasos. Já em relação às fibras, observou-se diferença extremamente
significativa (P = 0,0003), conseqüentemente a percentagem de fibras em Serra Talhada foi
maior que em Sertânia. Já a percentagem de parênquima, ao contrário do que aconteceu com
as fibras, foi maior em Sertânia do que em Serra Talhada. Semelhante às fibras, a
percentagem de raios foi maior nas amostras de Serra Talhada que nas de Sertânia (Tab. 1).
Na análise da variação medula-câmbio nas seções transversais do lenho do caule, não
houve diferença significativa entre os dois municípios quanto à percentagem de vasos,
portanto agruparam-se os espécimes desses dois locais e aplicou-se o teste Spearman Rank
Correlation observando-se correlação extremamente significativa (r = 0,47 e P = 0,0001)
entre a distância da medula e a percentagem de vasos, ou seja, à medida que se afasta da
medula a percentagem de vasos aumenta. Em relação às fibras, como houve diferença
significativa entre os municípios aplicou-se o teste nas amostras tanto de Sertânia como de
Serra Talhada, observando-se correlações negativas, isto é, quanto mais distante da medula
menor a percentagem de fibras. Assim como os vasos, a percentagem de parênquima também
tendeu a aumentar à proporção que se distanciava da medula nos indivíduos dos dois
municípios. Em relação aos raios, não houve correlação nas amostras de Sertânia, nem nas de

68
Serra Talhada (Tab. 2; Fig. 11, 12)
As médias de densidade foram altas nos troncos tanto próximo da medula como
próximo do câmbio, não havendo diferenças significativas em relação a essas posições,
tampouco em relação aos municípios (Tab. 3).
Galhos - Aplicando-se o teste t, comparando-se Serra Talhada com Sertânia foram observadas
diferenças significativas em relação a proporção de todos os elementos analisados, sendo as
maiores percentagens de vasos e parênquima encontradas nas amostras de Serra Talhada e as
maiores percentagens de fibras e raios nas amostras de Sertânia (Tab. 1).
Nas amostras de Sertânia as proporções de vasos e parênquima não apresentaram
correlação significativa em um dos dois galhos analisados, porém uma tendência à correlação
positiva. Todavia, quanto às fibras houve correlação negativa significativa, isto é, quanto mais
distante do centro menor a percentagem. Houve, portanto, uma maior concentração de fibras e
tendência a menor concentração de vasos e de parênquima no início do crescimento (Tab. 2).
No município de Serra Talhada, os resultados foram semelhantes aos ocorridos em
Sertânia. Apenas a percentagem de fibras apresentou correlação negativa com a distância no
sentido medula-câmbio (Tab. 2).
As médias de densidade nos galhos também se mostraram altas, tanto próxima da
medula quanto próxima ao câmbio. Assim como ocorreu ao nível do tronco, não houve
diferenças significativas em relação a essas posições, tampouco em relação aos municípios
(Tab. 3).

69
Figuras 1-4. Caesalpinia pyramidalis Tul. (catingueira). Seção transversal. 1. Vasos (poros) de
distribuição difusa, solitários, geminados e múltiplos radiais; parênquima marginal em linhas finas
(seta) e parênquima aliforme e confluente. 2. Região próxima da casca e casca (seta). 3. Detalhe da
seção visualizando parênquima marginal em linha (seta). 4. Parênquima confluente formando faixas e
fibras gelatinosas (seta). Barras = 200 µm (1); 50 µm (3); 100µm (4).

Figuras 5-10. Caesalpinia pyramidalis Tul. (Catingueira). 5-8. Seção tangencial. 5. Vaso com pontoações guarnecidas (seta). 6. Placa de perfuração simples (seta branca) e pontoações guarnecidas (seta preta). 7. Detalhe das pontoações guarnecidas. 8. Raios estratificados, unisseriados (seta) e bisseriados. 9-10. Seção radial. 9. Raio homogêneo. 10. Cristais prismáticos em células do parênquima axial (seta). Barras = 20 µm (5); 10 µm (6); 5 µm (7); 100 µm ( 8, 9); 50 µm (10).
y = -3,6151x + 58,779R2 = 0,8161
y = 2,5287x + 24,571R2 = 0,7688
y = 1,0663x + 9,62R2 = 0,6286
y = 0,0202x + 7,0303R2 = 0,0031
0
10
20
30
40
50
60
VasosRaiosFibrasParênquima
1 2 3 4 5 6 7 8 9 10 11 12
Raio (mm)
Perc
enta
gem
(%
)
1 2 3 4 5 6 7 8 0
y = -0,2306x + 42,047R2 = 0,02
y = -0,1639x + 29,5R2 = 0,0123
y = 0,3095x + 15,705R2 = 0,216
y = 0,085x + 12,748R2 = 0,0151
0
10
20
30
40
50
60
Raio (mm)
1 2 3 4 5 6 7 8 9 10
Raio (mm)
Figura 11. Caesalpinia pyramidalis Tul. (catingueira). Percentagem de vasos, raios, fibras e parênquima axial, no sentido medula casca, de amostras de madeiras coletadas no município de Sertânia. A. Tronco ao nível do DAP (1,30 m). B. Galho 1. C. Galho 2
y = -0,9683x + 37,629R2 = 0,3104
y = -0,4869x + 37,276R2 = 0,1188
y = 1,091x + 13,963R2 = 0,6718
y = 0,3642x + 11,132R2 = 0,144
10
20
30
40
50
60
Perc
enta
gem
(%
)Pe
rcen
tage
m (
%)
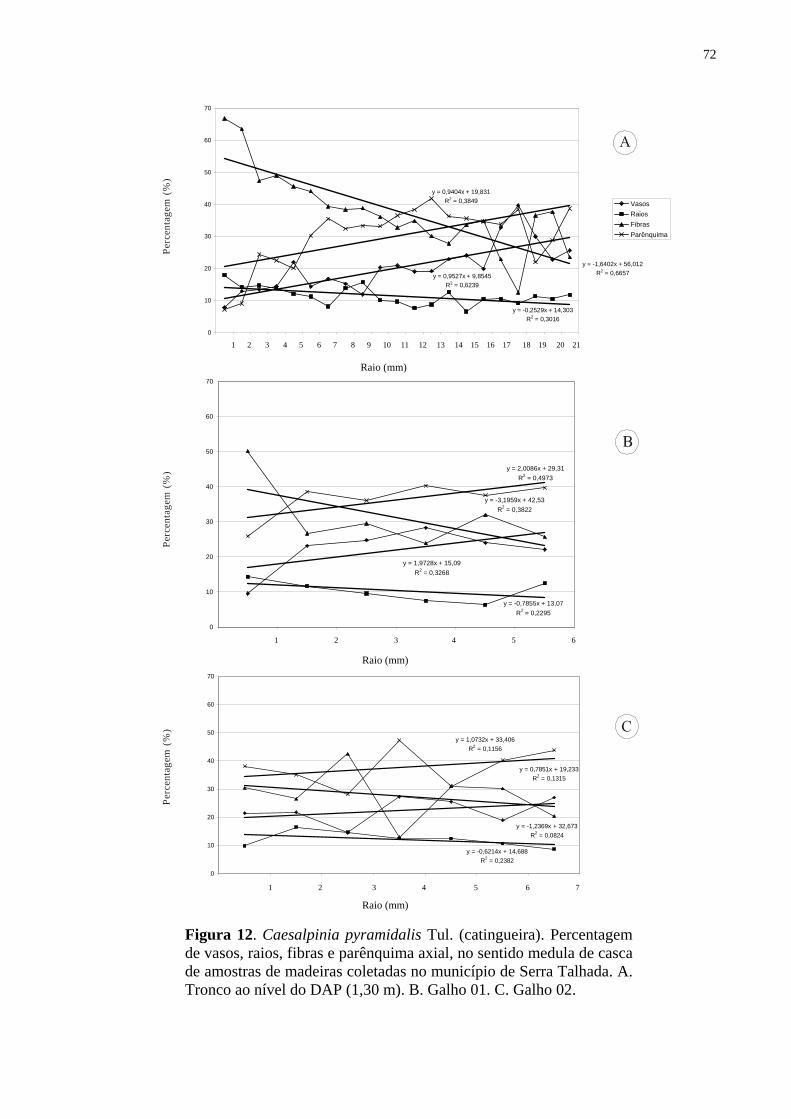
72
y = -1,6402x + 56,012R2 = 0,6657
y = -0,2529x + 14,303R2 = 0,3016
y = 0,9527x + 9,8545R2 = 0,6239
y = 0,9404x + 19,831R2 = 0,3849
0
10
20
30
40
50
60
70
VasosRaiosFibrasParênquima
Perc
enta
gem
(%
)
y = 2,0086x + 29,31R2 = 0,4973
y = -3,1959x + 42,53R2 = 0,3822
y = -0,7855x + 13,07R2 = 0,2295
y = 1,9728x + 15,09R2 = 0,3268
0
10
20
30
40
50
60
70
Raio (mm)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Perc
enta
gem
(%
)
Raio (mm)
1 2 3 4 5 6
y = 1,0732x + 33,406R2 = 0,1156
y = -1,2369x + 32,673R2 = 0,0824
y = -0,6214x + 14,688R2 = 0,2382
y = 0,7851x + 19,233R2 = 0,1315
0
10
20
30
40
50
60
70
Perc
enta
gem
(%
)
Raio (mm)
1 2 3 4 5 6 7
Figura 12. Caesalpinia pyramidalis Tul. (catingueira). Percentagem de vasos, raios, fibras e parênquima axial, no sentido medula de casca de amostras de madeiras coletadas no município de Serra Talhada. A. Tronco ao nível do DAP (1,30 m). B. Galho 01. C. Galho 02.
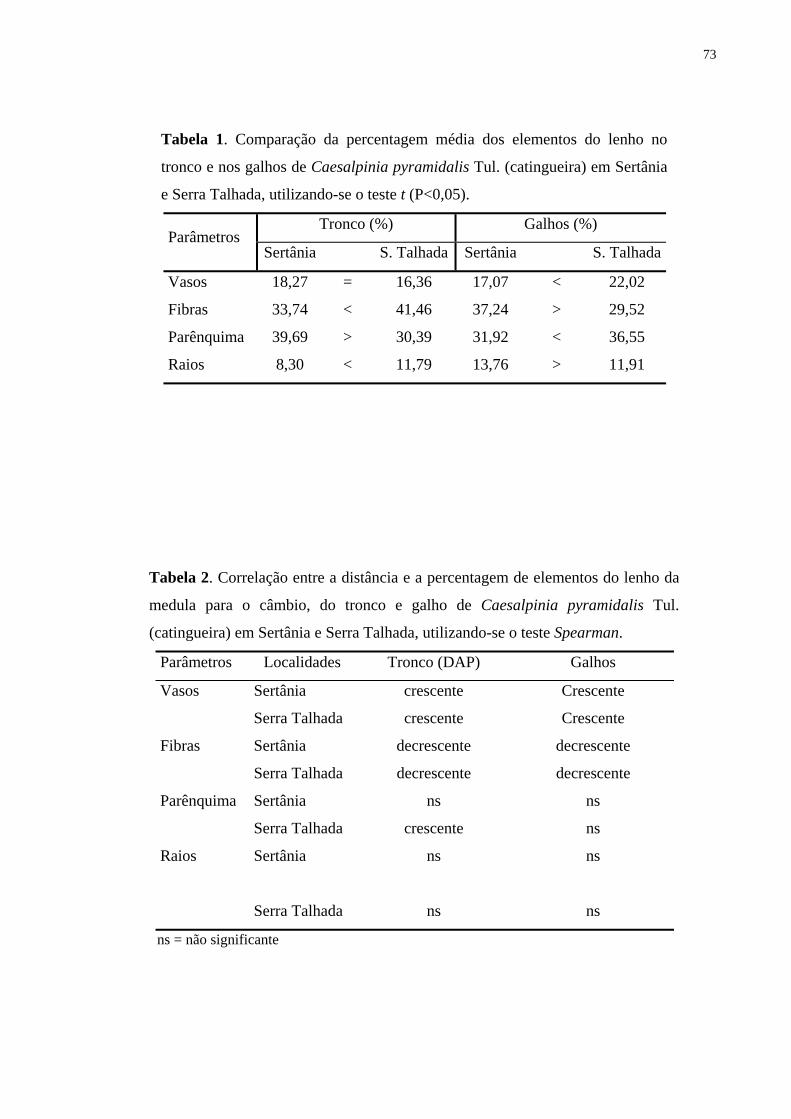
73
Tabela 1. Comparação da percentagem média dos elementos do lenho no
tronco e nos galhos de Caesalpinia pyramidalis Tul. (catingueira) em Sertânia
e Serra Talhada, utilizando-se o teste t (P<0,05).
Tronco (%) Galhos (%) Parâmetros
Sertânia S. Talhada Sertânia S. Talhada
Vasos 18,27 = 16,36 17,07 < 22,02
Fibras 33,74 < 41,46 37,24 > 29,52
Parênquima 39,69 > 30,39 31,92 < 36,55
Raios 8,30 < 11,79 13,76 > 11,91
Tabela 2. Correlação entre a distância e a percentagem de elementos do lenho da
medula para o câmbio, do tronco e galho de Caesalpinia pyramidalis Tul.
(catingueira) em Sertânia e Serra Talhada, utilizando-se o teste Spearman.
Parâmetros Localidades Tronco (DAP) Galhos
Vasos Sertânia crescente Crescente
Serra Talhada crescente Crescente
Fibras Sertânia decrescente decrescente
Serra Talhada decrescente decrescente
Parênquima Sertânia ns ns
Serra Talhada crescente ns
Raios Sertânia ns ns
Serra Talhada ns ns
ns = não significante
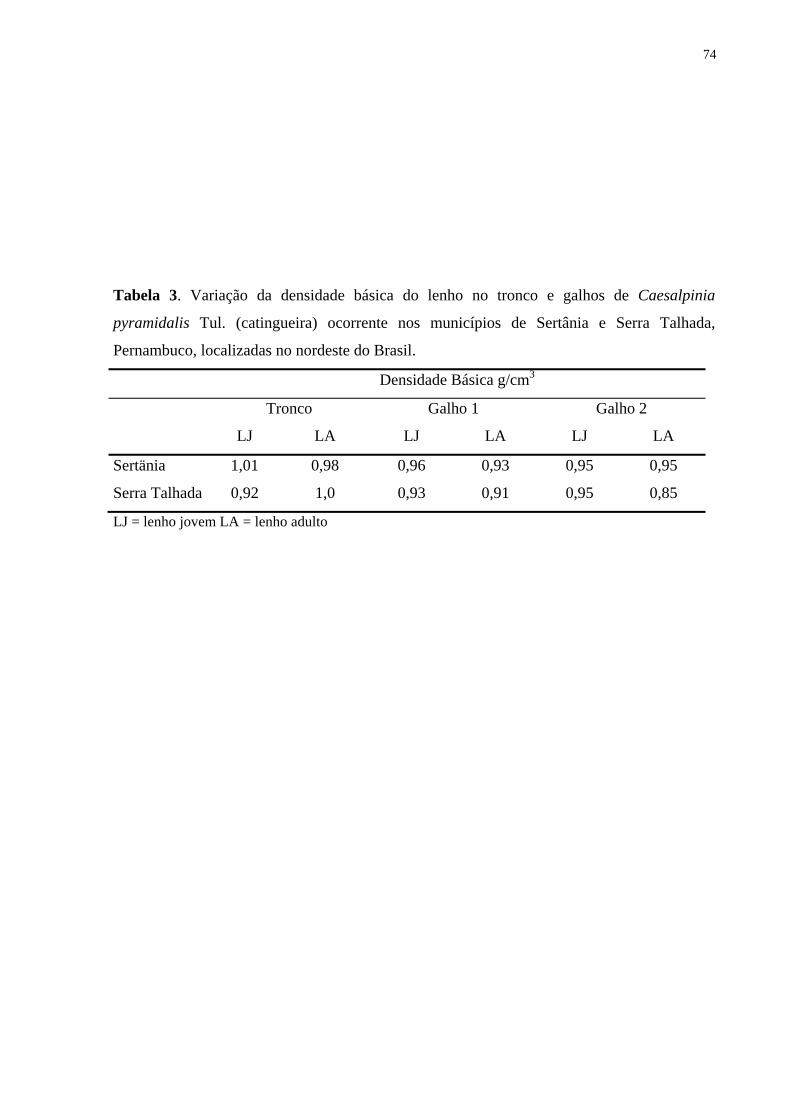
74
Tabela 3. Variação da densidade básica do lenho no tronco e galhos de Caesalpinia
pyramidalis Tul. (catingueira) ocorrente nos municípios de Sertânia e Serra Talhada,
Pernambuco, localizadas no nordeste do Brasil.
Densidade Básica g/cm3
Tronco Galho 1 Galho 2
LJ LA LJ LA LJ LA
Sertänia 1,01 0,98 0,96 0,93 0,95 0,95
Serra Talhada 0,92 1,0 0,93 0,91 0,95 0,85
LJ = lenho jovem LA = lenho adulto

75
DISCUSSÃO
Os caracteres anatômicos observados em Caesalpinia pyramidalis estão de acordo
com aqueles referidos para o gênero por Tortorelli (1956), Latorre (1983), Ranjani &
Krishnamurthy (1991) e Gasson et al. (2003), e para a própria espécie por Paula & Alves
(1980).
A presença de vasos solitários e múltiplos com arranjo difuso é um padrão comum no
lenho da maioria das espécies madeireiras. Sendo assim, Caesalpinia pyramidalis inclui-se
nesta categoria.
A pontoação guarnecida é uma característica marcante entre as Leguminosas (Jansen
et al. 2003). Todos os gêneros das subfamílias Mimosoideae e Papilionoideae apresentam este
tipo de pontoação. Apesar de alguns gêneros de Caesalpinioideae não apresentar esta
característica, o gênero Caesalpinia L. apresenta este tipo de pontoação. Este caráter pode ser
uma informação importante para a filogenia do grupo e para a definição de um largo clado
próximo à base da família (Herendeen 2000; Gasson et al. 2003).
Além da pontoação guarnecida ser um traço filogenético, ela pode também representar
uma estratégia adaptativa de sucesso para a sobrevivência e competição de plantas sujeitas a
contínuos ou sazonais períodos de seca em regiões de clima quente (Jansen et al. 2004),.
Assim, Caesalpinia pyramidalis, espécie endêmica da caatinga, apresentou este tipo de
pontoação que caracteriza o traço filogenético do gênero e também contribui para a
sobrevivência da espécie neste tipo de ecossistema caracterizado por períodos de seca e com
altas temperaturas. As pontoações guarnecidas funcionam como um eficiente sistema
hidráulico, prevenindo contra embolias dos vasos (Carlquist 1983, 1988).
Parece existir uma forte correlação negativa entre pontoações guarnecidas e placas de
perfuração escalariformes. Em geral, os táxons com pontoações guarnecidas têm placas de
perfuração simples. Vasos com pontoações e perfurações destes tipos evitam a embolia em
plantas crescendo em ambientes quentes com sazonalidade de estresses hídricos. Este tipo de
sistema é mais eficiente quanto à demanda do intenso fluxo de água imposta pelas altas taxas
de transpiração em plantas de regiões quentes e secas (Wheeler & Baas 1991; Jansen et al.
2004).
As fibras gelatinosas podem estar relacionadas ao lenho de tração quando ocorre
tortuosidade no caule ou nos galhos, ou terem a função de reserva devido a grande quantidade
de água em suas paredes (Paviani 1978). Na espécie em estudo, as fibras gelatinosas parecem
estar relacionadas com o armazenamento de água por duas razões: (i) a amostra do lenho
coletada não apresentava nenhuma tortuosidade e (ii) pelas características da região da

76
caatinga, onde a baixa precipitação média anual condiciona naturalmente o estabelecimento
de uma vegetação caracteristicamente de zona semi-árida, necessitando, portanto, de
adaptações para a sobrevivência. Alves & Angyalossy-Alfonso (2000), analisando a madeira
de várias regiões brasileiras, e Marcati et al. (2001), analisando Caesalpinia echinata Lam.
comprovaram esta evidência. Neste estudo de Caesalpinia pyramidalis, pode-se também
identificar essa característica.
Diversos gêneros da subfamília Caesalpinioideae apresentam uma tendência para
parênquima axial aliforme e vasicêntrico, usualmente em bandas, raios estreitos estratificados
com até três células de largura e presença de cristais nas células parenquimáticas axiais e
radiais e às vezes nas fibras (Gasson et al. 2003). Estes autores afirmaram ainda que estrutura
estratificada, composição e tamanho dos raios, presença de cristais nas células radiais e
pontoações guarnecidas são caracteres diagnósticos em potencial para análises filogenéticas
na subfamília Caesalpinioideae. Assim, os resultados aqui obtidos contribuem com mais
dados para estudos taxonômicos e filogenéticos dentro do gênero Caesalpinia.
Baas (1973), Wheeler & Baas (1991) e Zimmermann (1983) observaram que em
ambientes mais quentes a quantidade de parênquima é maior, especialmente em plantas
decíduas, como é o caso da espécie aqui estudada da caatinga.
Grisi (1976), citando trabalhos realizados sobre a anatomia de folhas de Caesalpinia
pyramidalis, informou que não foram encontrados elementos estruturais que comumente
aparecem nos vegetais adaptados à seca, afirmando, portanto, que a importância das
características fisiológicas é maior do que as anatômicas na adaptação da planta de caatinga à
seca. Concordamos que as características fisiológicas são fundamentais e importantes, porém
neste trabalho no qual se demonstra a estrutura do lenho de Caesalpinia pyramidalis, pode-se
observar grande concentração de fibras gelatinosas, percentagem considerável de parênquima
e distribuição difusa dos vasos, demonstrando que ocorrem variações nos elementos
estruturais do lenho que podem estar relacionadas com a questão da seca.
As camadas de crescimento distintas se destacaram pelas diferentes larguras que
apresentaram na Caesalpinia pyramidalis. Essas podem estar relacionadas com períodos
marcantes de seca e de chuva na caatinga, conforme observado para outras espécies da região.
A disponibilidade de água, nutrientes, luz e temperatura afetam a atividade do câmbio,
induzindo à formação de camadas de crescimento distintas no lenho (Détienne 1989, Vetter &
Botosso 1989 e Worbes 1989). Raven et al. (2001) afirmaram que em regiões semi-áridas
com baixa precipitação, as árvores são sensíveis aos níveis de pluviosidade, afetando a
atividade do câmbio, o que resulta na espessura da camada de crescimento, constituindo-se
em um indicador preciso da quantidade de chuvas de um determinado ano.

77
Além dos fatores climáticos, um outro fato que deve ser levado em consideração é a
queda das folhas. No período seco, observa-se que a catingueira perde suas folhas e é uma das
primeiras árvores a rebrotar com o início das chuvas. Alvim (1964) observou que a fenologia
das árvores mostra relação com a atividade cambial de muitas espécies, sendo esses eventos
fenológicos em ciclos anuais ou não anuais. Porém, somente um estudo amplo de
dendrocronologia, levando-se em consideração os fatores climáticos e fenológicos, poderá
fornecer dados mais precisos sobre essa questão.
Na comparação do lenho dos galhos das duas áreas de estudo, a maior percentagem de
fibras ocorreu em Sertânia. Assim, um aspecto importante aqui constatado é que quando a
proporção de parede celular das fibras é menos intensa no tronco, como observado nas
amostras de Serra Talhada, ocorre a compensação nos galhos como visto em Sertânia e vice-
versa. Portanto, ambos os municípios apresentam indivíduos de catingueira com potencial
para produção de energia (carvão), e Sertânia pode funcionar como região de manutenção da
espécie pelo fato de os galhos apresentarem potencial energético maior. Assim, pelo potencial
dos galhos, reservando as proporções, o corte dos troncos e sacrifício dos indivíduos poderá
ser evitado.
Aplicando-se o teste de correlação e analisando-se a proporção dos elementos no
sentido medula-câmbio no tronco, observou-se uma maior percentagem de fibras, menor
proporção de vasos e menor percentagem de parênquima próximo da medula em Serra
Talhada. O mesmo ocorreu em Sertânia. A percentagem de cada elemento anatômico da
madeira interfere na qualidade da mesma. Quanto menor a percentagem de vasos por mm2,
menor será a área de espaços vazios encontrada na madeira, conseqüentemente, maior será a
proporção de parede celular. Portanto, a região da madeira onde há menor quantidade de
vasos tende a apresentar um maior potencial energético. Nas fibras, como os lúmens são
proporcionalmente menores que os lumens dos vasos e as paredes, em geral, são mais
espessadas, a maior freqüência delas resultará em menores áreas de espaços vazios,
conseqüentemente neste local o potencial energético da madeira será maior. Caesalpinia
pyramidalis apresentou, no sentido medula-casca, menor percentagem de vasos próxima da
medula, maior percentagem de fibras e, além disso, observou-se que os valores de densidade
foram maiores que 0,90g/cm3, tanto próximo da medula quanto próximo da casca. Portanto,
pode-se inferir que a madeira de Caesalpinia pyramidalis apresenta potencial energético para
produção de carvão, desde a sua1 fase de lenho juvenil.
Quando se aplicou o teste de correlação, analisando-se a proporção dos elementos no
sentido medula-câmbio nos galhos, observou-se que houve diferença significativa na
percentagem de vasos, fibras e parênqima, semelhante ao ocorrido no tronco e no galho 1 nos

78
dois municípios . Portanto, nesses galhos o lenho apresentou-se apto para produção de carvão
e lenha.
Densidade básica – esse é um fator relevante na qualificação da madeira. Segundo Panshin &
De Zeeuw (1980), a densidade é uma propriedade física importante da madeira, pois é um
parâmetro que pode afetar outras de suas propriedades. Segundo Chimello (1980), a
densidade da madeira é considerada a propriedade física mais importante, relacionando-se
com outras propriedades e a sua utilização, assim, madeira de maior densidade corresponde,
em geral, a uma maior resistência mecânica e alto valor energético. Isto porque a densidade
está diretamente relacionada à quantidade de celulose, lignina e hemicelulose que a constitui.
Gasson (1987) e Rao et al. (1997) encontraram relação entre densidade do lenho e os
vasos. Eles observaram que madeiras com baixa freqüência de vasos apresentaram alta
densidade. Fujiwara et al. (1991) observaram em madeiras do Japão, que a espessura da
parede das fibras e a percentagem de material da parede têm influência direta na densidade da
madeira. Paula (1993) afirmou que quanto mais alta for a massa específica (densidade) da
madeira maior será o rendimento de energia em virtude do maior teor de celulose e lignina.
Além disso, altas densidades estão relacionadas também com a redução do tamanho das
células que compõem a madeira (Angyalossy et al. 2005). Prior & Gasson (1993) observaram
que o comportamento de seis madeiras da África submetidas à carbonização foi influenciado
pela natureza de suas fibras. Portanto, pode-se inferir que Caesalpinia pyramidalis, por
possuir uma grande percentagem de fibras em seu lenho e isto, conseqüentemente, está
relacionado com a alta densidade dessa madeira, quando submetida à carbonização produzirá
matéria prima de qualidade para obtenção de energia.
Como Caesalpinia pyramidalis, aqui estudada, apresentou no alburno e no cerne do
tronco e dos galhos, tanto em Serra Talhada quanto em Sertânia, densidade próxima ou maior
que 1,0 g/cm3, podemos afirmar, levando-se em consideração os parâmetros anatômicos
analisados anteriormente, que o lenho dessa espécie revela grande quantidade de celulose e
lignina, podendo ser utilizado para carvão, álcool combustível e coque metalúrgico.
Inúmeras pesquisas vêm sendo desenvolvidas com o objetivo de atenuar o efeito
negativo dos ciclos de curta rotação na qualidade da madeira, através da seleção e do
melhoramento genético, aumentando a densidade do lenho juvenil e a uniformidade da
densidade ao longo do tronco (Louzada et al. 1994). Os valores de densidade da madeira
encontrados no lenho das árvores de Caesalpinia pyramidalis, nas amostras analisadas,
próximas da medula e do câmbio em Sertânia e em Serra Talhada, indicam que desde a fase
de formação do lenho juvenil a madeira pode ser aproveitada para fins energéticos. Esse fato
torna-se significativo, na atual tendência de utilização da madeira de árvores em ciclos mais

79
curtos, com elevada proporção de lenho juvenil. A espécie apresentou um lenho juvenil
influenciando positivamente no potencial da madeira, devido à presença de maior proporção
de fibras e menor proporção de vasos.
Para a geração de energia (carvão) e produção de lenha, o lenho das árvores deve
apresentar alto percentual de fibras de paredes espessas, baixa proporção de parênquima
radial, longitudinal e de vasos e elevada densidade. Estas características físico-anatômicas
indicam altos teores de celulose e lignina e, em conseqüência, a biomassa necessária para a
sustentação de uma combustão eficiente. As árvores da espécie estudada mostraram essas
características.

80
CONCLUSÕES
Caesalpinia pyramidalis, em Serra Talhada e em Sertânia, apresenta demarcações
evidentes de camadas de crescimento que devem estar associadas a pluviosidade e a
fenologia, especialmente porque essa espécie é decídua, havendo momentos especiais em que
necessita de adaptações para sobrevivência às intemperes do ambiente.
Pelos parâmetros anatômicos do lenho aqui analisados (proporção dos vasos, raios,
parênquima e proporção e espessura das paredes das fibras) e a elevada densidade básica
(0,84-1,01g/cm3), conclui-se que Caesalpinia pyramidalis, espécie endêmica da caatinga,
revela grande quantidade de celulose e lignina, portanto apresenta potencial para produção de
álcool combustível, carvão vegetal e coque metalúrgico, conseqüentemente, maior potencial
energético.
Pela análise da variação radial pode-se propor o uso do lenho do tronco e dos galhos,
da madeira estudada, desde a fase do lenho juvenil, visto que nesta região ocorre uma grande
concentração de fibras, menor concentração de vasos e menor quantidade de parênquima,
tanto em Serra Talhada quanto em Sertânia.
Em ambos os municípios, Serra Talhada e Sertânia, a Caesalpinia pyramidalis
apresentou-se como espécie potencialmente apta para produção de energia pois quando o seu
potencial de celulose e lignina foi menor no tronco no município de Serra Talhada houve uma
compensação nos galhos e vice-versa. Os galhos apresentam potencial semelhante ao do
tronco expresso nas percentagens de fibras, vasos e parênquimas e densidade básica. Sendo
assim, a utilização dos mesmos pode ser útil porque evita o abate da árvore e facilita o
manuseio.

81
REFERÊNCIAS BIBLIOGRÁFICAS
Alves, E.S. & Angyalossy-Alfonso, V. 2000. Ecological trends in the wood anatomy of some
Brazilian species. IAWA Journal 21(1): 3-20.
Angyalossy, V.; Amano, E. & Alves, E.S. 2005. Madeiras utilizadas na fabricação de arcos
para instrumentos de corda: aspectos anatômicos. Acta Botánica Brasilica 19(4): 819-
834.
Alvim, P.T. 1964. Periodicidade do crescimento das árvores em climas tropicais. Pp.405-422.
In: Anais do Congresso Nacional de Botânica. 1964. Porto Alegre, Universidade
Federal do Rio Grande do Sul.
Baas, P. 1973. The anatomy of Ilex (Aquifoliaceae) and its ecological and phylogenetic
significance. Blumea 21: 193-258.
Brito, J.O. & Barrichelo, L.E.G. 1979. Aspectos florestais e tecnológicos da matéria-prima
para carvão vegetal. Circular técnica do Instituto de Pesquisas e Estudos
Florestais/ESALQ-USP 67: 1-20.
Carlquist, S. 1983. Wood anatomy of Onagraceae: further species; root anatomy; significance
of vestured pits and allied structures in dicotyledons. Annals of the Missouri Botanical
Garden 69(4): 755-769.
Carlquist, S. 1988. Comparative wood anatomy: systematic ecological and evolutionary
aspects of dicotyledonean wood. Berlin, Springer-Verlag.
Chimelo, J. 1980. Anotações sobre anatomia e identificações de madeiras. São Paulo, IPT.
Détienne, P. 1989. Appearance and periodicity of growth rings in some tropical woods.
IAWA Bulletin 10:123-132.
Fujiwara, S.; Sameshima, K.; Kuroda, K. & Takamura, N. 1991. Anatomy and properties of
Japanese hardwoods I. Variation of fibre dimensions and tissue proportions and their
relation to basic density. IAWA Bulletin 12: 419-424.
Gasson, P. 1987. Some implications of anatomical variations in the wood of pedunculate oak
(Quercus robur L.) including comparisons with common beech (Fagus sylvatica L.)
IAWA Bulletin 8: 149-166.
Gasson, P.; Trafford, C. & Matthews, B. 2003. Wood anatomy of Caesalpinioideae. Pp. 63-
93. In: B.B. Klitgaard and A. Bruneau (eds.). Advances in Legume Systematics, part 10,
Higher Level Systematics. Kew, Royal Botanic Gardens.
Grisi, B.M. 1976. Ecofisiologia da caatinga: comportamento hídrico de Caesalpinia

82
pyramidalis Tul. e Schinopsis brasiliensis Engl. Ciência e Cultura 28(4): 417-425.
Herendeen, P.S. 2000. Structural evolution in the Caesalpinioideae (Leguminosae).45-64. In:
P.S. Herendeen & A. Bruneau (eds.). Advances in Legume Systematics, part 9. Kew,
Royal Botanic Gardens.
IAWA Committee.1989. IAWA list of microscopic features for hardwood identification. E.
A. Wheeler; P. Baas & P. Gasson (eds.). IAWA Bulletin 10: 219-332.
Jansen, S.; Baas, P.; Gasson, P.; Lens, F.; Smets, E. 2004. Variation in xylem structure from
tropics to tundra: evidence from vestured pits. Proceedings of the National Academy of
Sciences 101(23): 8833-8837.
Johansen, D.A. 1940. Plant microtechnique. New York, MacGraw-Hill Book Company.
Latorre, F. 1983. Wood anatomy of eleven legumes from southeast of Ecuador. Ciencia y
Naturaleza 10: 84-92.
Louzada, J.L.P.C.; Fonseca, F.M.A. & Silva, M.E.C.M. 1994. Relações entre componentes da
densidade da madeira no lenho juvenil e no lenho adulto em Pinus pinaster Ait. Pp. 393-
405. In: Anais do III Congresso Florestal Nacional. Figueira da Foz, 1994. Figueira da
Foz , SFN..
Maia, G.N. 2004. Caatinga, árvores e arbustos e suas utilidades. Fortaleza, Leitura & Arte.
Marcati, C.R.; Angyalossy-Alfonso, V. & Benetati, L. 2001. Anatomia comparada do lenho
de Copaifera langsdorffii Desf. (Leguminosae-Caesalpinioideae) de floresta e cerradão.
Revista Brasileira de Botânica 24(3): 311-320.
Nguyen, T.V. 1977. Variation in vessel characteristics of Eucalyptus dalrympleana
Maiden associated with sample position growth rate and climate changes. Canberra,
Australian Nature University.
Panshin, A.J. & De Zeeuw, C. 1980. Textbook of wood technology. 4a ed. New York,
McGraw-Hill Book Co.
Paula, J.E. 1993. Madeiras da caatinga úteis para produção de energia. Pesquisa
Agropecuária Brasileira 28(2): 153-165.
Paula, J.E. & Alves, J.L.H. 1980. Estudo das estruturas anatômicas e de algumas propriedades
físicas da madeira de 14 espécies ocorrentes em áreas de caatinga. Brasil Florestal
10(43): 47-58.
Paviani, T.I. 1978.. Anatomia vegetal e cerrado. Ciência e Cultura 30: 1076-1086.
Polhill, R.M. 1994. Classification of the Leguminosae. Pp. 45-54. In: F.A. Bisby, J.
Buckingham & J.B. Harbone (eds.). Phytochemistry dictionary of the Leguminosae.

83
New York, Chapman and Hall.
Prior, J.A.B. & Cutler, D.F. 1996. Radial increments in four tropical, drought tolerant
firewood species. Commonwealth Forestry Review 75(3): 227-233.
Prior, J.A.B. & Gasson, P.E. 1993. Anatomical changes on charring six african hardwoods.
IAWA Journal 14(1): 77-86.
PROBIO 2000. Seminário sobre avaliação e identificação de ações prioritárias para a
conservação, utilização sustentável e repartição de benefícios da biodiversidade do bioma
caatinga. Disponível em: http://www.biodiversitas.org/caatinga. Acesso em: 20 jul.
2001.
Ranjani, K. & Krishnamurthy, K.V. 1991. Chambered crystal strands in the wood of
Caesalpiniaceae. Feddes Repertorium 102(1-2): 57-62.
Rao, R.V.; Aebischer, D.P. & Denne, M.P. 1997. Latewood density in relation to wood fibre
diameter, wall thickness, and fibre and vessel percentages in Quercus robur. IAWA
Journal 18: 127-138.
Raven, P.H., Evert, R.F. & Eichhorn, S.E. 2001. Biologia Vegetal 6a ed. Rio de Janeiro, Ed.
Guanabara Koogan.
Sampaio, E.V.S.B. 2002. Uso das plantas da caatinga. Pp. 49-68. In: Everardo V.S.B.
Sampaio; A.M. Giulietti; J. Virgílio & C.F.L. Gamarra-Rojas (ed.). Vegetação e flora da
caatinga. Recife, APNE & CNIP.
Sarkani, S.; Stieber, J. & Fillo, Z. 1957. Investigation on the wood of Hungarian Populus
species by means of quantitative xylotomy. Annals University Science Buda (Section
Bio) 1: 219-229.
Schrire, B.D.; Lewis, G.P.; Lavin, M. 2005. Biogeography of the Leguminosae. Pp. 21-54. In:
G.P. Lewis; B. Schrire; B. Mackinder; M. Lock (eds.). Legumes of the World. Kew,
Royal Botanic Gardens.
Tortorelli, L.A. 1956. Maderas y bosques argentinos. Buenos Aires, ACME, S. A. C. I.
Vetter, R.E. & Botosso, P.C. 1989. Remarks on age and periodicity of growth rings in some
tropical woods. IAWA Bulletin 10(2): 133-146.
Vital, B.R. 1984. Métodos de determinação de densidade da Madeira. Viçosa, UFV.
Wheeler, E.A. & Baas, P. 1991. A survey of the fossil record for dicotyledonous wood and its
significance for evolutionary and ecological wood anatomy. IAWA Bulletin 12(3): 275-
332.
Worbes, M. 1989. Growth rings, increment and age of trees in inundation forests, savannas

84
and a mountain forest in the neotropics. IAWA Bulletin 10(2): 109-122.
Zimmermann, M. H. 1983. Xylem structure and the ascent of sap. Berlin, Springer Verlag.

CAPÍTULO 4
ESTUDO COMPARATIVO DA MADEIRA DE MIMOSA
OPHTHALMOCENTRA MART. EX BENTH E MIMOSA TENUIFLORA
(WILLD.) POIR. (LEGUMINOSAE-MIMOSOIDEAE)

86
RESUMO
Pela importância econômica da madeira das espécies de Mimosa L. na caatinga
nordestina e pelo fato de, às vezes, Mimosa ophthalmocentra Mart. ex Benth. (jurema-branca)
ser confundida com Mimosa tenuiflora (Willd.) Poir. (jurema-preta) ou vice-versa, este
trabalho objetivou apresentar novos dados referentes à anatomia e densidade básica da
madeira das duas espécies, contribuir para a delimitação taxonômica das mesmas, identificar
caracteres anatômicos do lenho relacionando-os com o ambiente caatinga, e apresentar o
potencial energético que elas possuem. O estudo anatômico e a determinação da densidade do
lenho foram realizados com amostras ao nível do (1,30 m) e em dois galhos com diferentes
diâmetros, de indivíduos dos municípios de Serra Talhada e Sertânia, Pernambuco, Brasil. As
espécies apresentaram distinção expressa através do tipo de casca, coloração do cerne e do
alburno, além das características diferentes nos seus lenhos. Mimosa ophthalmocentra
apresentou camadas de crescimento distintas, constituídas por linhas de parênquima contendo
cristais, parênquima escasso e menor quantidade de raios. Já Mimosa tenuiflora apresentou
camada de crescimento distinta, porém sem cristais, parênquima axial vasicêntrico, em faixas
e em confluência, e maior percentagem de raios. Ambas exibiram alguns elementos do lenho
com as características anatômicas comuns às diversas espécies do gênero Mimosa.
contribuindo assim para a taxonomia do gênero. Pelos parâmetros anatômicos do lenho e pela
elevada densidade básica (>0,84 g/cm3), concluiu-se que as duas espécies apresentaram
perspectivas seguras para a produção de álcool combustível e carvão desde a fase de lenho
juvenil. Mimosa ophthalmocentra apresentou maior potencial energético, por possuir
parênquima escasso e menor percentagem de raios.
Palavras-chaves: Mimosa, anatomia e densidade de madeira, carvão, caatinga.

87
ABSTRACT
Due to the economic importance of the wood from Mimosa L. species in the
Northeastern Brazil’s caatinga and to the fact that the Mimosa ophtalmocentra Mart. ex
Benth. (jurema-branca) is confused with the Mimosa tenuiflora (Willd.) Poir. (jurema-preta)
and vice versa, this research had the objective to present new data related to the anatomy and
basic density of the wood of both species; to contribute to the taxonomic study in
differentiating them; to identify anatomic characters of the wood relating them to the caatinga
environment; and to present the energetic potential that they possess. The anatomic study and
wood density determination were carried out with samples at DAP level (DBH) and with two
branches of individuals within the municipalitys of Serra Talhada and Sertânia (Pernambuco).
The species both differed by their bark type, coloring of heartwood and sapwood, besides the
different anatomic characteristics found in their wood. Mimosa ophtalmocentra presented
distinct growth layers constituted of parenchyma lines containing crystals, scarce parenchyma
and smaller quantity of rays. The Mimosa tenuiflora, on the other hand, also presented growth
layers, but without crystals, axial parenchyma vasicentric, confluent or in bands, and greater
percentage of rays. Both species presented some wood elements with anatomic characteristics
typical of the representatives of the Mimosa, contributing therefore to a better taxonomic
study of the genus. Through the wood anatomic parameters and the elevated basic density (>
0,84 g/cm3), it was reached the conclusion that the both species presented great perspectives
to the production of fuel alcohol and charcoal since juvenile wood age. Because the Mimosa
ophthalmocentra had scarce parenchyma and a smaller number of rays, it showed a greater
energetic potential thanj Mimosa tenuiflora.
Palavras-chaves: Mimosa, anatomy and density wood, charcoal, caatinga.

88
INTRODUÇÃO
A anatomia da madeira tem contribuído para a solução de questões taxonômicas,
evolutivas e ecológicas. Barreta-Kuipers (1981) estudou a anatomia de madeira de algumas
Leguminosae com relevância à taxonomia. Loureiro & Silva (1968) apresentaram métodos
para identificação macroscópica de madeiras, dados gerais sobre as espécies estudadas e
respectivas descrições anatômicas das mesmas. Fedalto et al. (1986) afirmaram que a
anatomia da madeira, como pesquisa subsidiária, muitas vezes tem se prestado à solução de
problemas taxonômicos e como fonte de dados complementares, sendo utilizada em trabalhos
de inventários florestais, nos quais é comum deparar-se com material estéril ou incompleto.
Atualmente, o interesse maior nesses estudos se deve, geralmente, ao auxílio na identificação
e separação de espécies (Soffiati & Angyalossy-Alfonso 1999). Gasson et al. (2003)
afirmaram que a anatomia da madeira representa uma importante ferramenta na identificação
de amostras desconhecidas e uma fonte potencial de caracteres para o estudo em sistemática
das Leguminosae.
Barreta-Kuipers (1981) apresentou um abrangente estudo sobre a anatomia da madeira
de Leguminosae, no qual examinou 83 gêneros de Caesalpinioideae arbóreas, 35 de
Mimosoideae e 68 de Papilionoideae, considerando caracteres relacionados ao tamanho e
estratificação dos raios para discussão de semelhanças e diferenças dentro destas subfamílias.
Gasson et al. (2003) descreveram os caracteres da madeira de 131 dos 158 gêneros de
Caesalpinioideae, levando em consideração a organização dos dados no contexto filogenético.
A subfamília Mimosoideae (Leguminosae) apresenta cerca de 82 gêneros com
aproximadamente 3.271 espécies, distribuídas nas regiões tropicais, subtropicais e
temperadas-quente do globo (Schrire et al. 2005). O gênero Mimosa L. apresenta cerca de 350
espécies endêmicas da América do Sul (Luckow 2005). Entre as espécies do gênero que
ocorrem na caatinga pernambucana, estão as espécies Mimosa ophthalmocentra Mart. ex
Benth. (jurema-branca ou jurema-de-imbira) e Mimosa tenuiflora (Willd.) Poir. (jurema-
preta), simpátricas, pertencentes à série Leiocarpae Benth., que possui 28 espécies das quais
nove ocorrem na caatinga do Nordeste (Barneby 1991).
Mimosa ophthalmocentra, espécie endêmica da caatinga (Giulietti et al. 2002), é um
arboreto medindo de 3 a 6 m, de caule rugoso com espinhos retos, folhas bipinadas com dois
a quatro pares de pinas e 15 a 22 pares de folíolos por pina, inflorescência em forma de
espiga, flor branca a creme e fruto seco. É usada como forrageira e as suas folhas e vagens são
consumidas por cabras, ovelhas e bovinos; a madeira é utilizada como lenha e estacas; a
resina é comestível, saborosa e recomendada no combate à gripe. Mimosa tenuiflora é um

89
arboreto que varia de 4 a 7 m de altura, possui caule reto ou inclinado, com espinhos retos ou
ligeiramente curvos, ramos verdes, e folhas viscosas pela presença de glândulas minúsculas,
bipinadas, com cinco a sete pares de pinas e 18 a 36 pares de folíolos por pina, inflorescência
em forma de espiga, flor branca a creme e fruto seco. Esta espécie tem folhas e vagens
consumidas pelas cabras, ovelhas e bovinos, alta capacidade de produção de lenha e estacas, e
seu carvão é considerado de boa qualidade. A casca é utilizada como cicatrizante e
antiinflamatório. Além disso, é uma planta melífera cujas flores são boas fornecedoras de
pólen (Costa et al. 2002). Segundo Maia (2004), a madeira de Mimosa tenuiflora é muito
resistente e empregada em construções como estacas e pontes, além de fornecer excelente
lenha e carvão de alto valor energético.
Mimosa ophthalmocentra e Mimosa tenuiflora são espécies de elevado potencial
madeireiro para o nordeste (Figueirôa et al. 2005). Mimosa tenuiflora apresenta-se com
grande potencial como elemento fundamental dentro de um sistema agroflorestal em Serra
Talhada (Sampaio 1993). Esta espécie é oportunista e secundária, estabelecendo-se em áreas
antropizadas além de apresentar uma grande amplitude de tolerância aos diferentes
parâmetros físico-químicos do solo (Camargo-Ricalde 2000).
O reconhecimento das espécies de Mimosa não é muito fácil, sem que seja preciso
recorrer a caracteres que para serem visíveis necessitam do auxílio de lupa com aumento de
10x. Tenta-se, quando possível, usar caracteres macroscópicos que, mesmo não
diagnosticando esses táxons em toda a sua área de distribuição, possam auxiliar no
reconhecimento das espécies no âmbito da caatinga. Estes caracteres podem servir para
formar grandes grupos taxonômicos de espécies, mas deve-se estar atento que para separar
espécies muito parecidas eles não são suficientes. A utilização de caracteres anatômicos do
lenho pode auxiliar conjuntamente para este fim. Atualmente, vários trabalhos de taxonomia
estão associados a caracteres da anatomia do xilema, principalmente quando emprega-se a
cladística. Alguns autores recentes conferiram à anatomia da madeira grande valor em estudos
evolutivos os quais provêm marcantes avanços nas filogenias apontadas (Herendeen & Miller
2000; Olson 2005).
Além de contribuir para a taxonomia, análises da anatomia do lenho e das
características físicas e químicas das madeiras são fundamentais em um estudo de manejo que
vise o conhecimento de espécies florestais a serem utilizadas para fins energéticos,
especialmente para a produção de lenha, prática comum nas comunidades rurais do Nordeste.
Segundo Paula et al. (2000), a descrição anatômica das madeiras é de fundamental
importância para a determinação de seu possível aproveitamento. Esta afirmação está baseada
no fato de que as dimensões, a freqüência e o arranjo dos elementos anatômicos têm grande

90
influência sobre as propriedades físicas e mecânicas da madeira.
Zimmermann (1983), Baas et al. (1983), Baas & Carlquist (1985), Carlquist &
Hoekman (1985) e Lindorf (1994), abordando as floras de regiões de clima árido, semi-árido
e mediterrâneo, estabeleceram as tendências gerais de organização da estrutura anatômica do
lenho de plantas que habitam ambientes xéricos.
Considerando-se a importância econômica da madeira das espécies do gênero Mimosa
para as comunidades da caatinga nordestina, e pelo fato de, às vezes, Mimosa
ophthalmocentra ser confundida com Mimosa tenuiflora ou vice-versa, este trabalho
objetivou: (i) apresentar novos dados referentes à anatomia e a densidade básica da madeira
dessas duas espécies; (ii) contribuir para o estudo taxonômico auxiliando para a diferenciação
das mesmas; (iii) identificar caracteres anatômicos do lenho relacionando-os com o ambiente
da caatinga; e (iv) estimar a potencialidade de produção de energia (carvão) a partir das
características anatômicas e da densidade do lenho dessas espécies.

91
MATERIAL E MÉTODOS
Foram selecionadas áreas de caatinga nas Estações Experimentais da Empresa
Pernambucana de Pesquisa Agropecuária (IPA-PE), em Serra Talhada (Fazenda Saco – 3.200
ha) e em Sertânia (Fazenda Cachoeira – 630 ha), com vasta ocorrência de Mimosa
ophthalmocentra Mart. ex Benth. (Fig. 1-3) e Mimosa tenuiflora (Willd.) Poir. (Fig. 4-6), por
se encontrarem em um raio de 200 km e apresentarem semelhanças florísticas. Serra Talhada
localiza-se na microrregião do Pajeú, a 418 km da capital (Recife), com latitude de 07o53’
57”S e 38o18’09”W e altitude de 500 msnm, apresentando solo maciço de gnais superficial,
marrom claro, ácido, franco argiloso com algum cascalho. A precipitação varia de 450-750
mm, de março a maio e a temperatura média anual é de 37o C. Trata-se de uma extensa área
de caatinga semi-densa, madura, pouco perturbada, apresentando extrato arbóreo com
algumas árvores de até 12 m, e arbustivo e herbáceo bastante desenvolvidos. A área
experimental foi de 150 ha, região denominada de Pimenteira (PROBIO 2000). Sertânia
localiza-se na microrregião de Moxotó, a 316 km da capital, com latitude de 08o04’25”S e
37o12’24”W e altitude de 611 msnm, apresentando solo superficial, marrom claro, ácido,
franco argiloso com algum cascalho. A precipitação varia de 350-600 mm, de março a maio e
a temperatura média anual é de 35o C. Trata-se de uma área extensa de caatinga, com
vegetação mais fechada na encosta, árvores e arbustos maduros pouco perturbada. A área
experimental foi de 38 ha, conhecida como Manga do Cerecé. (PROBIO 2000).
Para o estudo anatômico do lenho das duas espécies foram coletadas amostras
circulares do tronco de três árvores ao nível do diâmetro a altura do peito (DAP = 1,30 m) e
de dois ramos em cada uma das áreas de estudo. Foram preparadas lâminas permanentes
contendo uma seção transversal e duas longitudinais tangencial e radial do lenho, com 18-25
µm de espessura, utilizando-se micrótomo de deslize Spencer. As seções do lenho foram
clarificadas com hipoclorito de sódio e coradas com azul de alcião e safranina alcoólica (50%)
1:1, tratadas em série alcoólica de 50 a 100%, a cada 10% e montadas em euparol (Johansen
1940). Nas seções transversais do lenho, em áreas de 1 a 1 mm contíguas, ininterruptas, no
sentido da medula para o câmbio, foi traçado um transecto ao longo de um raio da madeira, na
região de maior diâmetro, para análise morfométrica. Foram mensurados os elementos de
vaso, fibras e parênquima radial e longitudinal, utilizando-se o programa de análise de
imagem KS 300 da Carl Zeiss, no Jodrell Laboratory do RBG Kew, Inglaterra. Para a
descrição dos parâmetros anatômicos, foram aplicadas as normas da IAWA Committee
(1989). As exsicatas encontram-se depositadas no Herbário da Empresa Pernambucana de
Pesquisa Agropecuária (IPA-PE). As lâminas histológicas permanentes foram depositadas na

92
Xiloteca do Instituto de Biologia da UFBA, laboratório de micromorfologia da Universidade
Estadual de Feira de Santana e do Laboratório de Micromorfologia do Jodrell Laboratory,
Royal Botanic Gargens, Kew.
Na análise ultraestrutural dos elementos de vasos em microscopia eletrônica de
varredura (MEV), foram utilizadas amostras de madeiras de 1 x 1cm, no sentido longitudinal
à altura da região mediana entre o alburno e o cerne. Estes cortes foram montados em porta-
espécimes com fita adesiva de grafite sendo em seguida recobertos com ouro em sputter
Balzers SCD 050. As amostras preparadas foram analisadas e elétronfotomicrografadas em
microscópio eletrônico de varredura LEO 1430VP.
Na determinação da densidade do lenho foram utilizadas amostras de regiões próximas
da medula e próximas do câmbio, coletadas do tronco (DAP) e de dois galhos. A densidade
do lenho foi determinada pela razão da massa seca do lenho (seco a 105o C) pelo volume
saturado, de acordo com o método descrito por Vital (1984). Para a comparação das médias e
análises de correlação, utilizaram-se os testes t de Student e o Spearman Rank Correlation,
respectivamente.

93
RESULTADOS
Mimosa ophthalmocentra e Mimosa tenuiflora apresentam inflorescência em forma de
espiga com flores brancas a creme e frutos secos (Fig. 1, 4)
Foi evidente a diferença de coloração da casca das duas espécies. Mimosa
ophthalmocentra (jurema-de-imbira) apresentou casca fina, praticamente lisa e de coloração
cinza-amarronzada com lenticelas amareladas (Fig. 3). Já Mimosa tenuiflora (jurema-preta)
apresentou casca grossa e rugosa, com grandes lenticelas e sulcos longitudinais liberando
facilmente partes deste ritidoma, e de coloração escura, característica importante que
provavelmente lhe conferiu o nome popular de jurema-preta (Fig. 6). Ambas as espécies
apresentaram distinção entre alburno e cerne. O alburno de Mimosa ophthalmocentra é de
coloração clara amarelada e o cerne amarronzado, enquanto Mimosa tenuiflora apresenta
alburno amarelo-avermelhado e cerne escuro, também avermelhado (Fig. 2, 5).
Descrição anatômica do lenho – Qualitativamente, as espécies Mimosa ophthalmocentra e
Mimosa tenuiflora variaram quanto à anatomia da madeira em alguns parâmetros, como o
agrupamento dos vasos e o tipo de parênquima axial (Fig. 7, 11, 12). Ambas as espécies
apresentaram camadas de crescimento delimitadas por linhas de parênquima, sendo que na
primeira essa camada é rica em cristais (Fig. 8), o que não foi observado na outra espécie
(Fig. 12). Em ambas as espécies os vasos apresentaram-se em arranjo difuso, entretanto, na
primeira distribuíram-se em solitários, múltiplos de 2-6 em disposição radial e, às vezes,
formando agrupamentos de até oito vasos (Fig. 7). Já em Mimosa tenuiflora distribuíram-se
predominantemente em solitários e múltiplos de dois, sendo algumas vezes de três (Fig. 11,
12). Nas duas espécies as placas de perfuração são simples e as pontoações intervasculares
alternas guarnecidas (Fig. 9, 10, 13, 14). Fibras libriformes e gelatinosas, com paredes muito
espessadas, ocorreram também nas duas espécies (Fig. 8, 12). O parênquima axial em Mimosa
ophthalmocentra apresentou-se bastante escasso, enquanto em Mimosa tenuiflora foi
paratraqueal aliforme, vasicêntrico e confluente (Fig. 7, 11). Os raios das duas espécies
assemelharam-se estruturalmente apresentando estratificação irregular, homogêneos,
bisseriados e compostos de células procumbentes (Fig. 15-20). Cristais prismáticos em células
subdivididas do parênquima axial estão presentes no lenho das duas espécies (Fig. 16, 19).
A Tabela 1 apresenta os dados quantitativos das características anatômicas da madeira
das duas espécies. O comprimento e a largura dos vasos e fibras foram maiores em Mimosa
tenuiflora, assim como a altura e a largura dos raios. Já essas medidas nos raios diferiram de
uma espécie para outra. Mimosa ophthalmocentra apresentou raios mais baixos e mais
estreitos, enquanto em Mimosa tenuiflora os raios foram mais altos e mais largos.

94
Analisando o tronco das duas espécies nas duas áreas experimentais, observou-se que
a madeira delas apresentou maior percentagem de vasos em Sertânia. Já em relação às fibras,
ao parênquima e aos raios, as espécies apresentaram diferenças a depender do município onde
foram analisadas. As fibras do tronco de Mimosa ophthalmocentra, que são abundantes em
relação aos demais elementos chegando a mais de 50%, não diferiram significativamente em
ambas as estações experimentais (Tab. 2). Já em Mimosa tenuiflora, a percentagem das fibras
foi maior em Serra Talhada (Tab. 3). A percentagem de parênquima em Serra Talhada foi
maior nos espécimes de Mimosa ophthamocentra e menor naqueles de Mimosa tenuiflora
(Tab. 2, 3). Nessa última espécie, a percentagem de raios também foi maior em Sertânia (Tab.
3). Os raios na madeira dos espécimes de Mimosa ophthalmocentra de Sertânia não diferiram
significativamente daqueles de Serra Talhada.
Nas Tabelas 2 e 3, pode-se observar também a comparação de médias do lenho nos
galhos entre Serra Talhada e Sertânia. Mimosa ophthalmocentra não apresentou diferenças
significativas entre os dois municípios em relação à percentagem de vasos. Porém, em
Mimosa tenuiflora houve uma maior percentagem de vaso em Sertânia. As fibras da primeira
apresentaram maior percentagem em Sertânia, enquanto Mimosa tenuiflora foi maior em
Serra Talhada. A percentagem de parênquima foi menor em Sertânia em Mimosa
ophthalmocentra e em Mimosa tenuiflora esta percentagem, diferente do que aconteceu com
as fibras, foi maior nesse município. A percentagem dos raios de ambas as espécies foi maior
em Serra Talhada.
Quanto à análise dos vasos no tronco, em ambas as espécies não ocorreram
correlações significativas em relação à distância da medula, tanto em Serra Talhada quanto
em Sertânia (Tab. 4, 5). Em relação às fibras de Mimosa ophthalmocentra, como não houve
diferença significativa entre os dados dos dois municípios, agruparam-se os espécimes das
duas regiões e aplicou-se o teste (Spearman), que indicou correlação extremamente
significativa negativa, ou seja, à medida que afasta-se da medula a percentagem de vasos
diminui em ambos os municípios (Tab. 4). O mesmo ocorrendo em Mimosa tenuiflora nos
dois municípios (Tab. 5).
Não ocorreu correlação entre a distância da medula e a percentagem de parênquima
tanto em Serra Talhada quanto em Sertânia, nos indivíduos de Mimosa ophthalmocentra (Tab.
4). Porém em Mimosa tenuiflora, à proporção que se afasta da medula há uma crescente
percentagem de parênquima (Tab. 5). Em ambas as espécies, os raios não apresentaram
correlações significativas em relação à distância da medula.
Para a análise da variação no sentido medula-câmbio nas seções transversais do lenho
dos galhos, observou-se que os vasos de Mimosa ophthalmocentra apresentaram correlação

95
extremamente significativa, tanto em Sertânia quanto em Serra Talhada, isto é, quanto mais
longe da medula maior a percentagem de vasos (Tab. 4). Em Mimosa tenuiflora não houve
diferenças significativas em nenhum dos dois municípios (Tab. 5). Já a percentagem de fibras,
em ambas às espécies, foi diminuindo à medida que a distância da medula aumenta.
Em relação ao parênquima e aos raios de Mimosa ophthalmocentra não houve
correlações significativas em nenhum dos dois municípios (Tab. 4). A percentagem de
parênquima de Mimosa tenuiflora foi aumentando à medida que se distanciava da medula, o
mesmo ocorrendo em relação aos raios, apenas em Sertânia. Já em Serra Talhada, os raios não
apresentaram diferença significativa (Tab. 5).
Densidade – Os dados quantitativos da densidade básica em Sertânia apresentaram uma
variação no lenho incluindo tronco e galhos entre 0,82-1,06 g/cm3 em Mimosa
ophthalmocentra e 0,86-1,03 g/cm3 em Mimosa tenuiflora (Tab. 6). Em Serra Talhada
apresentaram uma variação entre 0,93-1,0 g/cm3 em Mimosa ophthalmocentra e 0,88-1,0
g/cm3 em Mimosa tenuiflora (Tab. 6). Não foram observadas diferenças significativas em
relação às posições próximas da medula e próximas do câmbio.

11
4
1 3
65
2
Figuras 1-6. Caracteres macromorfológicos de duas espécies do gênero Mimosa que ocorrem na caatinga nordestina 1-3. Mimosa ophthalmocentra Mart. Ex Benth. 1. Inflorescência e frutos. 2. Tronco evidenciando alburno e cerne. 3. Casca. 4-6. Mimosa tenuiflora (Willd.) Poir. 4. Inflorescência. 5. Tronco demonstrando alburno, cerne e casca. 6. Casca. Barras = 2 cm (2, 5); 1 cm (3, 6). (Figuras 1 e 4 de autoria de Peter E. Gasson).
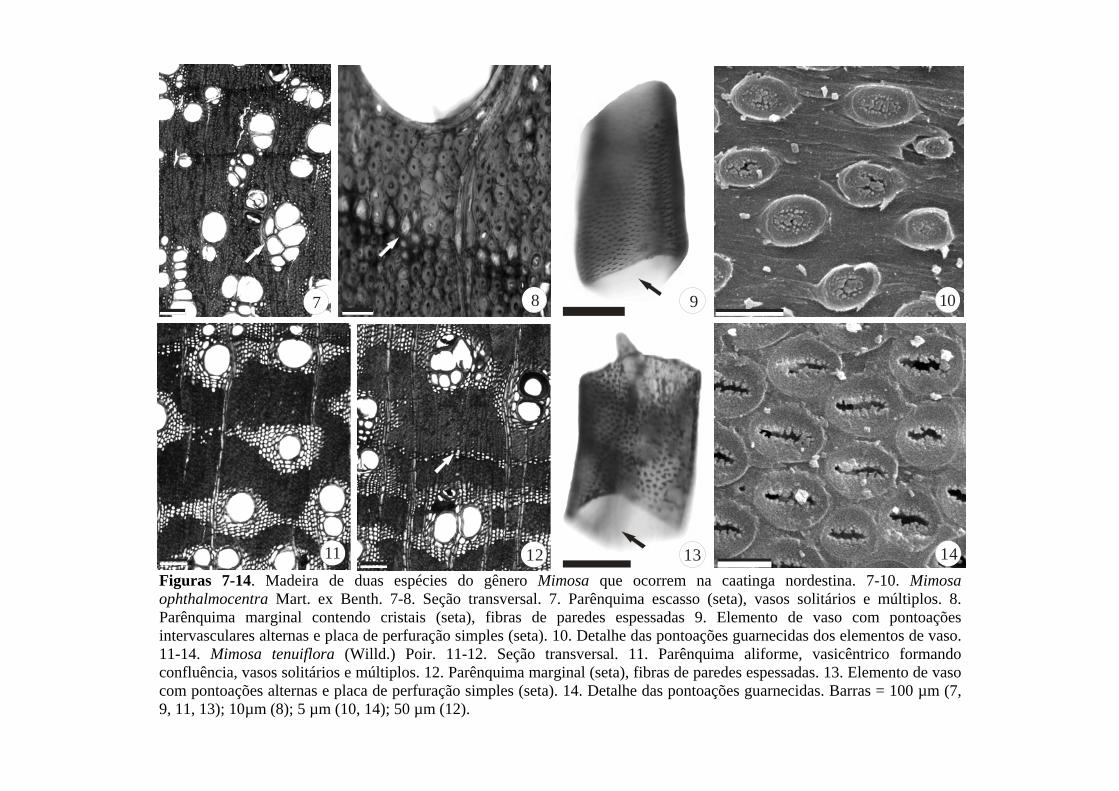
87 8 1097
11 12 13 14
Figuras 7-14. Madeira de duas espécies do gênero Mimosa que ocorrem na caatinga nordestina. 7-10. Mimosa ophthalmocentra Mart. ex Benth. 7-8. Seção transversal. 7. Parênquima escasso (seta), vasos solitários e múltiplos. 8. Parênquima marginal contendo cristais (seta), fibras de paredes espessadas 9. Elemento de vaso com pontoações intervasculares alternas e placa de perfuração simples (seta). 10. Detalhe das pontoações guarnecidas dos elementos de vaso. 11-14. Mimosa tenuiflora (Willd.) Poir. 11-12. Seção transversal. 11. Parênquima aliforme, vasicêntrico formando confluência, vasos solitários e múltiplos. 12. Parênquima marginal (seta), fibras de paredes espessadas. 13. Elemento de vaso com pontoações alternas e placa de perfuração simples (seta). 14. Detalhe das pontoações guarnecidas. Barras = 100 µm (7, 9, 11, 13); 10µm (8); 5 µm (10, 14); 50 µm (12).

Figuras 15-17. Madeira de duas espécies do gênero Mimosa que ocorrem na caatinga nordestina. 15-17. Mimosa ophthalmocentra Mart. ex Benth. 15-16. Seção tangencial. 15. Raios bisseriados (setas), não estratificados. 16. Cristais nas células do parênquima axial (seta). 17. Seção radial evidenciando raio homogêneo composto de células procumbentes e pontoações radiovasculares (seta). 18-20. Mimosa tenuiflora (Willd.) Poir. 18-19. Seção tangencial. 18. Raios uni e bisseriados (setas), não estratificados. 19. Cristais nas células do parênquima (seta). 20. Seção radial com raio homogêneo composto de células procumbentes. Barras = 100 µm (15, 18); 20 µm (16, 20);10 µm (17); 50 µm (19).
17
19
16
18 20
15
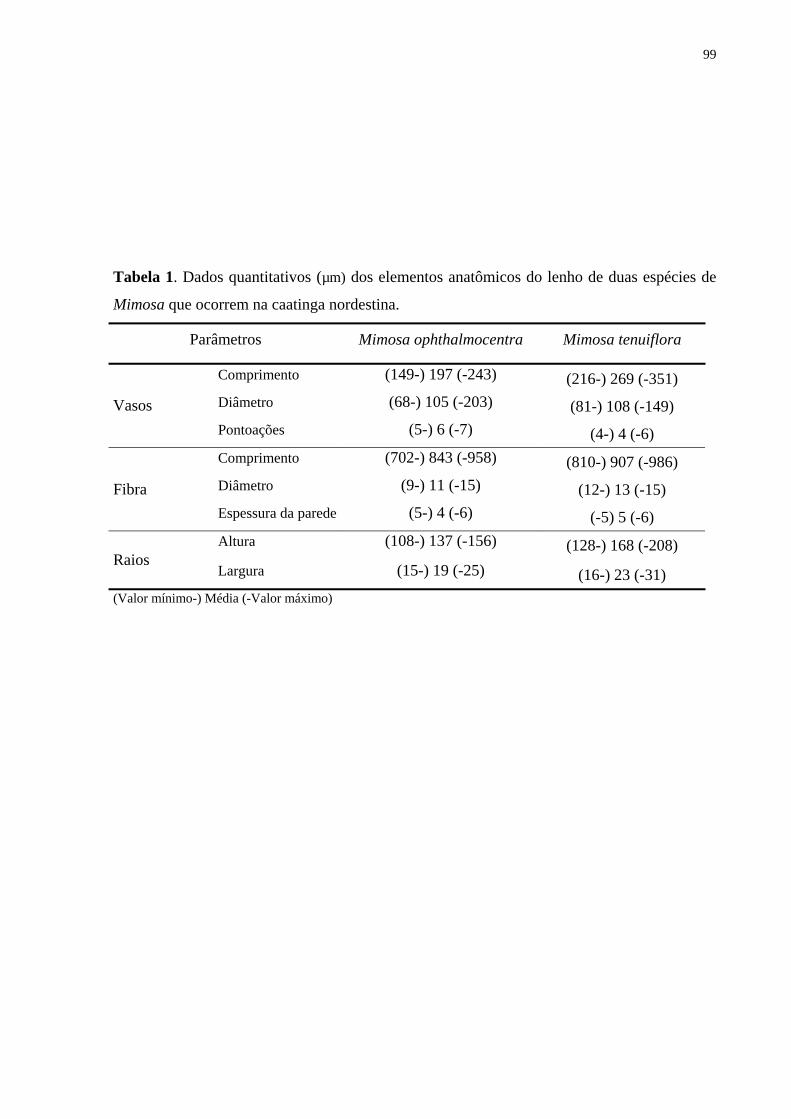
99
Tabela 1. Dados quantitativos (µm) dos elementos anatômicos do lenho de duas espécies de
Mimosa que ocorrem na caatinga nordestina.
Parâmetros Mimosa ophthalmocentra Mimosa tenuiflora
Comprimento (149-) 197 (-243) (216-) 269 (-351) Diâmetro (68-) 105 (-203) (81-) 108 (-149) Vasos Pontoações (5-) 6 (-7) (4-) 4 (-6) Comprimento (702-) 843 (-958) (810-) 907 (-986) Diâmetro (9-) 11 (-15) (12-) 13 (-15) Fibra Espessura da parede (5-) 4 (-6) (-5) 5 (-6) Altura (108-) 137 (-156) (128-) 168 (-208)
Raios Largura (15-) 19 (-25) (16-) 23 (-31)
(Valor mínimo-) Média (-Valor máximo)
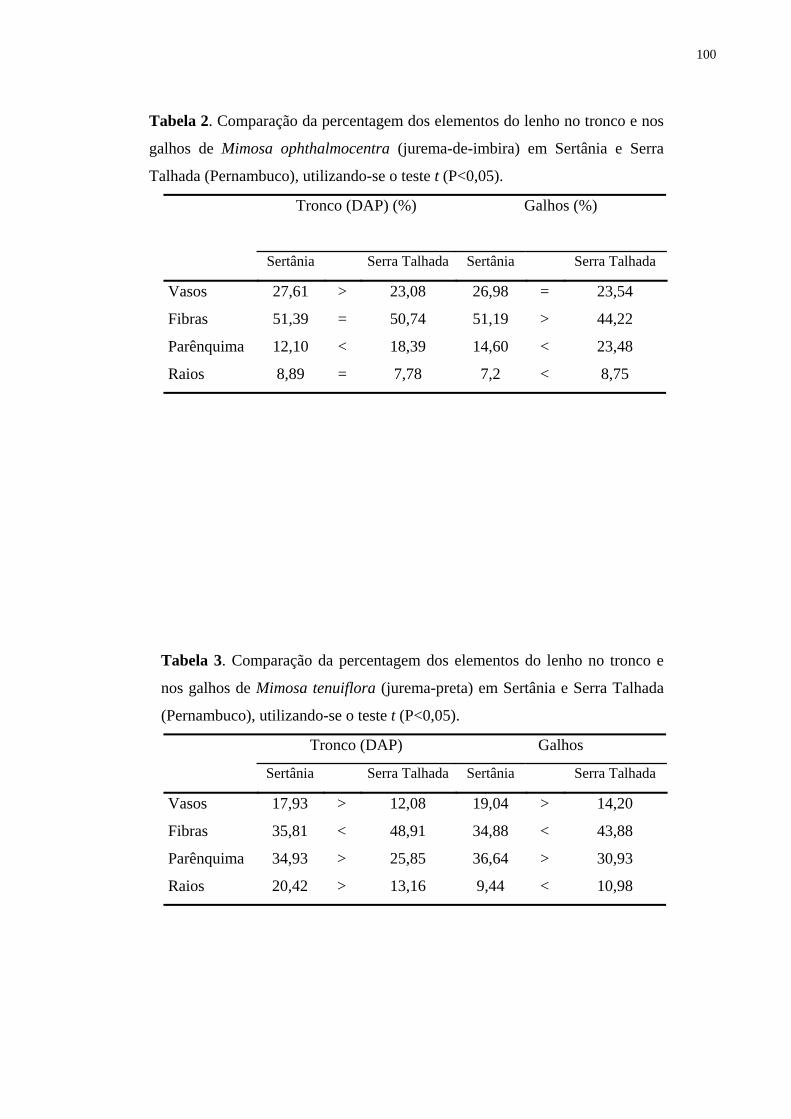
100
Tabela 2. Comparação da percentagem dos elementos do lenho no tronco e nos
galhos de Mimosa ophthalmocentra (jurema-de-imbira) em Sertânia e Serra
Talhada (Pernambuco), utilizando-se o teste t (P<0,05).
Tronco (DAP) (%)
Galhos (%)
Sertânia Serra Talhada Sertânia Serra Talhada
Vasos 27,61 > 23,08 26,98 = 23,54
Fibras 51,39 = 50,74 51,19 > 44,22
Parênquima 12,10 < 18,39 14,60 < 23,48
Raios 8,89 = 7,78 7,2 < 8,75
Tabela 3. Comparação da percentagem dos elementos do lenho no tronco e
nos galhos de Mimosa tenuiflora (jurema-preta) em Sertânia e Serra Talhada
(Pernambuco), utilizando-se o teste t (P<0,05).
Tronco (DAP) Galhos
Sertânia Serra Talhada Sertânia Serra Talhada
Vasos 17,93 > 12,08 19,04 > 14,20
Fibras 35,81 < 48,91 34,88 < 43,88
Parênquima 34,93 > 25,85 36,64 > 30,93
Raios 20,42 > 13,16 9,44 < 10,98

101
Tabela 4. Correlação entre a distância e a percentagem de elementos do lenho da
medula para o câmbio, do tronco e galho de Mimosa ophthalmocentra (jurema-de-
imbira) em Sertânia e Serra Talhada (Pernambuco), utilizando-se o teste Spearman.
Tronco (DAP) Galhos
Sertânia ns crescente Vasos
Serra Talhada ns crescente
Sertânia decrescente decrescente Fibras
Serra Talhada decrescente decrescente
Sertânia ns ns Parênquima
Serra Talhada ns ns
Sertânia ns ns Raios
Serra Talhada ns ns
ns = não significante
Tabela 5. Correlação entre a distância e a proporção de elementos do lenho da
medula para o câmbio, do tronco e galho de Mimosa tenuiflora (jurema-preta) em
Sertânia e Serra Talhada (Pernambuco), utilizando-se o teste Spearman.
Parâmetros Localidades Tronco (DAP) Galhos
Sertânia ns ns Vasos
Serra Talhada ns ns
Sertânia decrescente decrescente Fibras
Serra Talhada decrescente decrescente
Sertânia crescente crescente Parênquima
Serra Talhada crescente crescente
Sertânia ns crescente Raios
Serra Talhada ns ns

102
Tabela 6. Variação da densidade básica do lenho no tronco e galhos de Mimosa
ophthalmocentra (jurema-de-imbira) e Mimosa tenuiflora (jurema-preta) ocorrentes nos
município de Sertânia e Serra Talhada, Pernambuco.
Densidade básica g/cm3 Tronco Galho 1 Galho 2
LJ LA LJ LA LJ LA Sertânia
Mimosa ophthalmocentra
1,06
0,92
0,95
0,94
0,92
0,82
Mimosa tenuiflora 1,03 0,91 0,94 0,91 0,92 0,86
Serra Talhada
Mimosa ophthalmocentra
0,94
0,95
1,0
0,93
0,99
0,94
Mimosa tenuiflora 0,98 0,99 1,0 0,92 1,0 0,88
LJ = lenho jovem LA = lenho adulto

103
DISCUSSÃO
Comparação de espécies de Mimosa - A maioria dos caracteres anatômicos observados nas
espécies Mimosa ophthalmocentra e Mimosa tenuiflora está de acordo com aqueles referidos
para o gênero (Carnieletto & Marchiori 1993; Marchiori 1993, 1996; Maccari & Marchiori
1994; Paula 1995; Marchiori & Muniz 1996).
Segundo Barneby (1991), essas duas espécies pertencem ao gênero Mimosa L. seção
Batocaulon DC. e à série Leiocarpae Benth., e a principal diferença morfológica que as
separa é a presença de pontoações víscido-glandulares na face abaxial dos folíolos de Mimosa
tenuiflora. Neste estudo pode-se observar que existem outras diferenças marcantes no lenho
dessas duas espécies. Mimosa ophthalmocentra apresenta parênquima escasso, camada de
crescimento distinta delimitada por linha de parênquima marginal com as células ricas em
cristais, enquanto Mimosa tenuiflora tem parênquima vasicêntrico, em faixas e em
confluência, e camada de crescimento formada também por parênquima, porém, sem
ocorrência de cristais nessa região.
Marchiori (1993, 1996), Marchiori & Muniz (1996), Maccari & Marchiori (1994)
encontraram em três espécies da seção Calothamnus Barneby, Mimosa eriocarpa Benth.,
Mimosa incana (Spreng.) Benth. e Mimosa pilulifera Benth., e em uma outra espécie da seção
Mimosa L., Mimosa sparsa Benth., camadas de crescimento formadas por parênquima
marginal. Camargo-Ricalde (2000) observou que Mimosa tenuiflora, no México, também
apresenta camada de crescimento distinta. As duas espécies analisadas neste estudo também
apresentaram este tipo de camada de crescimento.
Marchiori (1993), estudando Mimosa bimucronata (DC.) O. Kuntze, e Paula (1995),
estudando Mimosa artemisiana Heringer & Paula, ambas espécies pertencentes à mesma
seção de Mimosa ophthalmocentra e Mimosa tenuiflora, observaram que essas espécies não
possuem camada de crescimento distinta. Assim, a presença de camada de crescimento parece
não ser um bom caráter para agrupar as espécies de Mimosa da seção Batocaulon. Porém, faz-
se necessário uma análise mais aprofundada em relação a essas duas espécies e a outras
pertencentes às referidas seções a fim de se chegar a conclusões mais consistentes.
Nas duas espécies aqui analisadas, a presença de cristais no parênquima que delimita a
camada de crescimento só foi observada em Mimosa ophthalmocentra, podendo ser
considerado um caráter diagnóstico na distinção de ambas. Segundo Fahn (1990), a forma, a
localização e o tipo de cristal são características úteis em estudos taxonômicos.
Especialistas do Oxford Forestry Institute, examinando o lenho de Acacia erioloba E.
Mey. (Leguminosae-Mimosoideae) da região sul da África, encontraram faixas estreitas

104
distintas de células especializadas contendo cristais de oxalato de cálcio e observaram que
essas faixas corresponderam à idade aproximada da respectiva árvore (Barnes 2004). Como
Mimosa ophthalmocentra pertence à mesma subfamília de Acacia erioloba que também
apresentou a camada de crescimento delimitada por células ricas em cristais, pode-se supor
que essas camadas possam revelar a idade aproximada de espécimes dessa espécie na
caatinga. Contudo, se faz necessário um estudo sobre o comportamento do câmbio da espécie
na formação das camadas de crescimento relacionando-o com os fatores ambientais tais como
temperatura, pluviosidade e luminosidade.
Como no período de seca na caatinga (junho a dezembro) ambas as espécies perdem
parte de suas folhas, ficando praticamente apenas os troncos e os galhos, a diferença das
camadas de crescimento pode corroborar na distinção entre ambas contribuindo assim na
taxonomia do gênero.
Em Mimosa artemisiana, M. eriocarpa, M. incana, M. pilulifera e M. sparsa,
ocorreram poros com padrão difuso (Carnieletto & Marchiori 1993; Maccari & Marchiori
1994; Paula 1995; Marchiori & Muñiz 1996; Marchiori 1996). Marchiori (1993) analisando
Mimosa bimucronata observou poros semidifusos. Camargo-Ricalde (2000) já tinha
observado em Mimosa tenuiflora do México, poros de distribuição difusa, alguns solitários,
porém a maioria múltipla radial de dois, vasos com pontoações areoladas alternas e placa de
perfuracão simples, semelhantes às espécies estudadas provenientes da caatinga. Pode-se
observar que o agrupamento solitário e a distribuição difusa dos poros presentes nas espécies
estudadas corroboram os dados de Alves & Angyalossy-Alfonso (2000), os quais afirmaram
ser essas características comuns dentro da flora brasileira.
As diferenças encontradas entre o comprimento e o diâmetro dos elementos do lenho
das duas espécies deste estudo podem ser utilizadas na separação das mesmas (Tab. 1).
Camargo-Ricalde (2000), descrevendo a madeira de Mimosa tenuiflora do México,
encontrou para os elementos de vasos, diâmetro tangencial moderadamente pequeno, com
(47)80(91) µm, e extremamente curtos, com (113)160(220) µm, diferindo do encontrado nos
indivíduos da espécie na caatinga (Tab. 1), e fibras libriformes muito curtas de (236)546(803)
µm, com diâmetro mediano de (9)14(22) µm e paredes grossas de (6)7(8) µm, raios com
(69)144(315) de altura e (8)10(18) de largura, diferindo do encontrado nos indivíduos da
espécie na caatinga (Tab. 1).
As diferenças observadas entre Mimosa tenuiflora de Sertânia e Serra Talhada (região
de caatinga do Brasil) aqui estudada e Mimosa tenuiflora de Oaxaca e Chiapas (região de
clima semiseco e quente do México) estudadas por Camargo-Ricalde (2000), podem ser
atribuídas às diferenças de precipitação pluviométrica anual, que é maior nesta região do

105
México (500-1200mm) que nos municípios da caatinga estudados (350-750mm), e à diferença
de altitude de 760-1250 msnm e 500-611 msnm, respectivamente. A menor disponibilidade de
chuva pode induzir o câmbio à formação de vasos com maiores comprimentos e maiores
diâmetros para aproveitar ao máximo a quantidade de água disponível no solo. Além disso,
como os raios são formados por células parenquimáticas, pode ser que sendo maiores e mais
largos representem uma estratégia de sobrevivência para reserva em períodos mais secos.
As fibras ocorrentes tanto em Mimosa tenuiflora quanto em Mimosa ophthalmocentra
apresentaram maiores comprimentos que as de Mimosa tenuiflora do México, porém a largura
e a grossa espessura da parede foram semelhantes (Tab. 1).
M. artemisiana, M. bimucronata, M. eriocarpa, M. incana, M. pilulifera e M. sparsa
apresentaram parênquima aliforme vasicêntrico, às vezes confluente (Marchiori 1993, 1996;
Carnieletto & Marchiori 1993; Maccari & Marchiori 1994; Paula 1995; Marchiori & Muniz
1996). Mimosa incana e Mimosa pilulifera apresentaram parênquima estratificado. Neste
estudo, o tipo de parênquima observado em Mimosa tenuiflora assemelhou-se ao observado
nas espécies citadas. Mimosa ophthalmocentra apresentou parênquima escasso, diferindo de
M. tenuiflora e das demais espécies estudadas pelos vários autores acima citados. Considera-
se, portanto, o parênquima como uma característica adicional do lenho na delimitação de
Mimosa ophthalmocentra.
Mimosa artemisiana e Mimosa bimucronata apresentaram raios homogêneos
(Marchiori 1993; Paula 1995), similares aos de Mimosa ophthalmocentra e Mimosa tenuiflora
aqui estudadas. Como já foi visto anteriormente, essas duas espécies pertencem à seção
Batocaulon DC. (Barneby 1991), e a mesma série de M. artemisiana e de M. bimucronata.
Ambas as espécies aqui analisadas seguem o padrão característico das Mimosoideae, de
acordo com Barreta-Kuipers (1981). Segundo esta autora, a homogeneidade dos raios é uma
das características mais notáveis na madeira desta subfamília, constituindo-se em importante
tendência evolutiva do grupo, a partir das Leguminosae-Caealpinioideae. Porém, raios
heterogêneos foram encontrados em M. daleoides Benth., M. eriocarpa, M. eriocarpa, M.
pilulifea, M. sparsa (Carnieletto & Marchiori 1993; Maccari & Marchiori 1994; Marchiori
1982; Marchiori 1996; Marchiori & Muniz 1996). Todas essas espécies citadas pertencem a
outras seções. Estudos referentes às espécies citadas na literatura, que apresentam raios
heterogêneos, devem ser analisados objetivando uma melhor caracterização da madeira do
gênero Mimosa quanto aos raios. Pode ser ainda, que nas análises de Barreta-Kuipers estas
espécies não tenham sido incluídas.
Camargo-Ricalde (2000) encontrou, em Mimosa tenuiflora do México, raios
unisseriados e parcialmente bisseriados, heterogêneos, extremadamente baixos de

106
(69)144(315) µm e extremadamente finos de (8)10(18) µm, diferindo dos resultados
encontrados em Mimosa tenuiflora da caatinga (Tab. 1). Assim, a variação dos raios em
relação à composição (homogêneos ou heterogêneos) e quanto ao número de células de
largura parece ser um caráter variável no gênero. Pelas diferenças de Mimosa tenuiflora do
Brasil e do México, pode-se até inferir um caráter variável na própria espécie. Propõe-se,
portanto, um estudo mais detalhado dessa espécie.
Todas as espécies apresentaram raios extremamente baixos, semelhantes às Mimosa
ophthalmocentra e M. tenuiflora, podendo representar uma característica predominante no
gênero, mas são necessários análises de uma freqüência maior de espécies para se confirmar
este fato.
Comparação em relação ao ambiente – A presença de elementos de vasos com pontoações
guarnecidas é uma característica geral da maioria das Leguminosae (Carlquist 1983; Jansen et
al. 2003). De acordo com Herendeen (2000) e Gasson et al. (2003), apesar de algumas
Caesalpinioideae não apresentarem elementos de vaso com esta característica, o gênero
Caesalpinia e todos os gêneros de Mimosoideae e Papilionoideae apresentam este tipo de
pontoação em seus elementos de vaso. Segundo estes autores, pontoações guarnecidas podem
ter evoluído dentro das Leguminosae e conseqüentemente esta característica pode ser uma
sinapomorfia para união da maioria dos gêneros e a segregação da tribo Cercideae e 15
gêneros da tribo Cassieae.
Geralmente os táxons que apresentam elementos de vaso com pontoações guarnecidas
têm placas de perfuração simples. Esta correlação juntamente com as tendências ecológicas,
contrastando com as placas de perfuração escalariforme, têm inspirado a hipótese funcional
que as pontuações guarnecidas contribuem para a segurança hidráulica. Este tipo de
pontuação parece facilitar a reversão do embolismo em regiões secas e quentes com altas
taxas de transpiração (Jansen et al. 2003; Jansen et al. 2004). Portanto, além de ser um traço
filogenético, as pontoações guarnecidas parecem representar uma estratégia adaptativa de
sucesso à sobrevivência e competição de plantas. M. ophthalmocentra e M. tenuiflora aqui
estudadas confirmam essas acepções, pois pertencem a família Mimosoideae e encontram-se
em ambiente quente e seco.
As guarnições nas pontoações são importantes na prevenção de embolias aumentando
a aderência das moléculas de água à parede das células (Carlquist 1983, 1988). Assim, essas
pontoações seriam adaptativas às plantas de ambientes que sofrem algum tipo de estresse
hídrico, como é o caso de M. ophthalmocentra e M. tenuiflora na caatinga, dos municípios de
Sertânia e Serra Talhada. Alves & Angyalossy-Alfonso (2000) corroborando os estudos
ecológicos em anatomia de madeira de Carlquist (1983, 1988), Wheeler & Baas (1991) e

107
outros, observaram fortes indícios da existência da relação entre pontoações guarnecidas,
placas de perfuração simples e ambientes secos.
Mimosa artemisiana, M. bimucronata, M. eriocarpa, M. incana, M. pilulifera e M.
sparsa possuem fibras libriformes e gelatinosas (Carnieletto & Marchiori 1993; Marchiori
1993, 1996; Maccari & Marchiori 1994; Paula 1995; Marchiori & Muniz 1996). Ambas as
espécies aqui analisadas apresentaram os mesmos tipos de fibras. As fibras gelatinosas podem
estar relacionadas ao lenho de tração quando ocorre tortuosidade no caule ou nos galhos ou
terem a função de reserva de água devido a grande quantidade em suas paredes da celulose
que é hidrófila (Esau 1974; Paviani 1978), especialmente em ambientes com grandes períodos
de seca como a caatinga.
Quanto ao parênquima paratraqueal em faixas, Wheeler & Baas (1991) afirmaram que
é mais comum nas regiões tropicais do que nas temperadas. Alves & Angyalossy-Alfonso
(2000), analisando a flora arbórea de várias regiões brasileiras, comprovaram esta evidência.
Esses resultados estão de acordo com os de Mimosa tenuiflora aqui encontrados que
apresentou grande quantidade de parênquima axial. Zimmermann (1983) afirmou que, durante
estações desfavoráveis, árvores decíduas baixam até zero a taxa de transpiração, necessitando
de fluxo de água condicionado à pressão osmótica. Para gerar esse fluxo é necessária a
liberação constante de açúcares para manter elevada a pressão osmótica dentro dos vasos.
Esse açúcar é disponibilizado pelo tecido parenquimático (Braun 1984).
Mimosa ophthalmocentra diferiu de Mimosa tenuiflora e das espécies do gênero
anteriormente citadas, pois apresentou parênquima axial bastante escasso. Essa espécie
apresenta outro tipo de adaptação para sobrevivência. A estratégia de sobrevivência pode estar
expressa em uma maior quantidade de células com paredes espessadas (esclerificadas). Arens
(1958) observou que no cerrado as espécies têm dificuldade para realização da fotossíntese
por conta da deficiência de minerais no solo. Assim, essas espécies tendem a apresentar um
alto grau de esclerificação que foi denominado escleromorfismo oligotrófico. Na caatinga,
existem problemas como alto índice de luminosidade, elevadas temperaturas e altos índices de
evapotranspiração, dificultando a sobrevivência dos vegetais. Todavia, essas plantas
apresentaram diferentes estratégias de sobrevivência das quais pode-se citar em M.
ophthalmocentra e M. tenuiflora queda de folhas na fase seca. Outra estratégia ocorre no
umbuzeiro (Spondias tuberosa Arruda, Anacardiaceae), que é a presença de parênquima
aquífero nas suas raízes. Pode-se então inferir que Mimosa ophthalmocentra utiliza também
como estratégia de sobrevivência na caatinga, pela dificuldade de obtenção de água, a
esclerificação de suas células, resultando, portanto, em uma maior percentagem de fibras.
Porém, estudos mais detalhados devem ser realizados para melhor esclarecimento dessa

108
questão na caatinga, comparando-se espécies de locais em diferentes altitudes relacionando a
estrutura do lenho com os fatores do meio.
Em Serra Talhada, a precipitação pluviométrica e a temperatura são mais elevadas,
porém a altitude é menor em relação a Sertânia. Apesar da maior pluviosidade, a temperatura
mais elevada leva a uma maior evapotranspiração. Considerando-se que a atividade cambial
está relacionada com os fatores climáticos (Rao & Rajput 1999; Salvador et al. 1997), pode-se
observar que os indivíduos estudados em Serra Talhada apresentaram uma maior taxa de
esclerificação representada pela maior percentagem de fibras no lenho, corroborando com
Webber (1936) e Baas (1982). Estes autores observaram que os diferentes componentes
anatômicos do lenho resultantes da atividade do câmbio vascular podem apresentar diferenças
quantitativas em função das adaptações ao ambiente e situação fisiológica no fluxo da água.
Comparação quanto ao potencial energético das espécies – Notou-se que Mimosa
ophthalmocentra apresenta parênquima escasso, enquanto a Mimosa tenuiflora, parênquima
vasicêntrico em faixas e em confluência. Além disso, o comprimento e diâmetro dos vasos e
dos raios foram maiores em Mimosa tenuiflora. Segundo Paula (1989), as madeiras com
potencial energético, ou seja, as ricas em celulose e lignina, são aquelas com alta freqüência
de fibras com paredes espessas, vasos de diâmetro grande escassos, parênquima axial e radial
pouco ou ausente, e densidade básica a partir de 0,60g/cm3. Assim, pode-se observar que M.
ophthalmocentra, por possuir escassez de parênquima, apresenta teor energético mais alto, em
virtude de obter maior teor de celulose e lignina. Portanto, pode-se identificar outra diferença
entre as espécies quanto ao potencial energético; a madeira de Mimosa ophthalmocentra por
possuir essas características é potencialmente mais energética.
O lenho dos galhos das duas espécies estudadas apresentou potencial para produção de
energia tanto em Sertânia quanto em Serra Talhada. Todavia, Sertânia pode funcionar como
região de manutenção de Mimosa tenuiflora pelo fato dos galhos apresentarem potencial
energético ainda maior que em Serra Talhada. Assim, reservando as proporções, poder-se-á
evitar o corte dos troncos desta espécie.
M. ophthalmocentra e M. tenuiflora revelaram que, pela grande percentagem de fibras
e pouca percentagem de vasos e raios, o lenho em sua fase juvenil apresentou-se
potencialmente apto para produção de carvão e lenha. Resultados semelhantes foram obtidos
nas análises dos galhos.
Densidade - segundo Chimello (1980), a densidade da madeira é considerada a sua
propriedade física mais importante, relacionando-se com outras propriedades e a sua
utilização. Dessa forma a madeira de maior densidade corresponde, em geral, a uma maior
resistência mecânica e alto valor energético. Isto porque a densidade está diretamente

109
relacionada à quantidade de celulose que a constitui (Angyalossy et al. 2005). M.
ophthalmocentra e M. tenuiflora apresentaram altos valores de densidade tanto no tronco
como nos galhos, demonstrando que essas madeiras possuem uma alta resistência. Gasson
(1987) e Rao et al. (1997) encontraram relação entre densidade do lenho e os vasos. Eles
observaram que a madeira com baixa freqüência de vasos apresenta alta densidade. Fujiwara
et al. (1991) observaram, em madeiras do Japão, que a espessura da parede das fibras e a
percentagem de material da parede tem influência direta na densidade da madeira. Paula
(1993) afirmou que quanto mais alta for a massa específica (densidade) da madeira maior será
o rendimento de energia em virtude do maior teor de celulose e lignina. Assim, pode-se
afirmar que os componentes do lenho têm uma influência direta na qualidade da sua madeira.
Portanto, as duas espécies aqui estudadas apresentaram, em seus lenhos, características que as
qualificam como espécies potencialmente energéticas.
A percentagem de fibras nas duas espécies apresentou valores acima de 35%, o que
determinou uma alta densidade em ambas. Mimosa tenuiflora, apesar de possuir alta
percentagem de parênquima em seu lenho, apresentou baixa percentagem de vasos, enquanto
Mimosa ophthalmocentra apresentou baixa percentagem de parênquima e maior percentagem
de vasos. Por esta razão a densidade das duas espécies foi semelhante, tornando-as aptas para
a obtenção de energia.
Pelas altas densidades encontradas no alburno e no cerne do tronco e dos galhos de M.
ophthalmocentra e M. tenuiflora, pode-se afirmar, levando-se em consideração os parâmetros
anatômicos aqui analisados radialmente, que o lenho dessas espécies revela grande quantidade
de celulose e lignina. Portanto, pode ser utilizado para carvão, álcool combustível e coque
metalúrgico, inclusive como já foi citado anteriormente, desde a fase de lenho juvenil.
Concordamos com Paula (1989) quando ressalta que madeira de boa qualidade para a
produção de energia, especialmente carvão e lenha, só será viável do ponto de vista
econômico e ecológico se incluídas na formação de grandes maciços florestais heterogêneos
sob planos de manejo ecológico de rendimento sustentado, a fim de evitar o extrativismo em
alta escala para fins energéticos. Portanto, jurema-de-imbira e jurema-preta que apresentaram
esse potencial energético, devem ser incluídas em projetos de manejo sustentável na caatinga
para estimular o seu uso de maneira racional, como um recurso renovável que é, para
populações atuais e as futuras gerações.

110
CONCLUSÕES
1. A observação dos troncos de Mimosa ophthalmocentra e de Mimosa tenuiflora
demonstrou distinção expressa através do tipo de casca e coloração do cerne e alburno,
contribuindo assim para um melhor diagnóstico taxonômico dessas espécies.
2. Com base na anatomia da madeira aqui analisada, ambas as espécies apresentaram
características diferentes nos seus lenhos que podem contribuir na distinção das mesmas. M.
ophthalmocentra apresentou camadas de crescimento constituída por linhas de parênquima
contendo cristais, parênquima escasso e menor percentagem de raios, enquanto a M.
tenuiflora apresentou camada de crescimento distinta porém sem cristais, parênquima axial
vasicêntrico, em faixas e em confluência e maior percentagem de raios.
3. M. ophthalmocentra e M. tenuiflora exibiram alguns elementos do lenho com as
características anatômicas comuns às diversas espécies do gênero Mimosa. Todavia, a análise
quali-quantitativa indicou que existem variações de algumas características entre as duas
espécies que podem estar relacionadas à sobrevivência dessas plantas ao ambiente caatinga,
onde a vegetação está sujeita a alta incidência luminosa e ao déficit hídrico.
4. Pelos parâmetros aqui analisados (percentagem dos vasos, raios, parênquima e
percentagem e espessura das paredes das fibras) concluiu-se que M. ophthalmocentra e M.
tenuiflora revelaram, nas respectivas madeiras, grande quantidade de celulose, portanto
podem ser utilizadas para produção de álcool combustível e carvão. Todavia, M.
ophthalmocentra por apresentar parênquima escasso e menor percentagem de raios, permitiu
maior concentração de parede rica em celulose, conseqüentemente maior potencial energético.
5. A partir da aplicação de teste estatístico, para correlação entre a distância e a percentagem
de células na análise da variação radial do lenho das espécies em estudo, pode-se concluir que
existem diferenças significativas da região próxima do centro da madeira para a periferia.
Propõe-se, portanto, o uso das madeiras das espécies estudadas desde a fase do lenho juvenil,
momento este em que, nas duas espécies estudadas, ocorreram grandes percentagens de fibras
e vasos com menor diâmetro. Em M. tenuiflora, nessa fase, ocorreu quantidade de parênquima
ainda menor.
6. Pela elevada densidade básica (0,82-1,06 g/cm3) no tronco e nos galhos, concluiu-se que
M. ophthalmocentra e M. tenuiflora da caatinga, revelaram grande quantidade de celulose e
lignina, portanto apresentam perspectivas seguras para produção de álcool combustível,
carvão vegetal e coque metalúrgico, conseqüentemente, maior potencial energético.

111
REFERÊNCIAS BIBLIOGRÁFICAS
Alves, S.P. de & Angyalossy-Alfonso, V. 2000. Ecological trends in the wood anatomy of
some Brazilian species. IAWA Journal 21(1): 3-20.
Angyalossy, V.; Amano, E. & Alves, E.S. 2005. Madeiras utilizadas na fabricação de arcos
para instrumentos de corda: aspectos anatômicos. Acta Botanica Brasilica 19(4): 819-
834.
Arens, K. 1958. O cerrado como vegetação oligotrófica. Boletim da Faculdade de Filosofia,
Ciências e Letras da Universidade de São Paulo, Botânica 15: 59-77.
Baas, P. 1982. Systematics, phylogenetic and ecological wood anatomy – History and
perspectives. Pp. 23-58. In: P. Baas (ed.). New perspectives in wood anatomy. Leiden,
The Hague Martinus Nijhoff Publishers.
Baas, P. & Carlquist, S. 1985. A comparison of the ecological wood anatomy of the floras of
southern California and Israel. IAWA Bulletin 6(4): 349-354.
Baas, P.; Werker, E. & Fahn, A. 1983. Some ecological trends in vessel characters. IAWA
Bulletin 4: 141-159.
Barneby, R.C. 1991. Sensitivae censitae. A description of the genus Mimosa Linnaeus
(Mimosaceae) in the New World. Memoirs of the New York Botanical Garden 65: 1-
835.
Barnes, R.D. 2004. The Acacia tree: a sustainable resource for Africa. Penzance, Rowes
the printers.
Barreta-Kuipers 1981. Wood anatomy of the Leguminosae: its relevance to taxonomy. Pp.
677-705. In: R. M. Polhill , P. H. Raven (eds.). Advances in legume systematics, Part 2.
Kew, Royal Botanic Gardens.
Braun, H.J. 1984. The significance of the accessory tissues of the hydrosystem for osmotic
water shifting as the second principle of water ascent, with some thoughts concerning the
evolution of trees. IAWA Bulletin 5(4): 275-294.
Camargo-Ricalde, S.L. 2000. Descripción, distribución, anatomía, composición química y
usos de Mimosa tenuiflora (Fabaceae-Mimosoideae) em México. Revista de Biología
Tropical 48(4): 1-23.
Carlquist, S. 1983. Wood anatomy of Onagraceae: further species; root anatomy; significance
of vestured pits and allied structures in dicotyledon wood. Annals of the Missouri
Botanical Garden 69(4): 755-769.
Carlquist, S. 1988. Comparative wood anatomy: systematic ecological and evolutionary
aspects of dicotyledonean wood. Berlin, Springer-Verlag.

112
Carlquist, S. & Hoekman, D.A. 1985. Ecological wood anatomy of the woody southern
California flora. IAWA Bulletin 6(4): 319-348.
Carnieletto, C. & Marchiori, J.N.C. 1993. Anatomia da madeira de Mimosa eriocarpa Benth.
Ciência Florestal 3(1): 67-75.
Chimelo, J. 1980. Anotações sobre anatomia e identificações de Madeiras. São Paulo, IPT.
Costa, J.A.S.; Nunes, T.S.; Ferreira, A.P.L.; Stradmann, M.T.S. & Queiroz, L.P. de. 2002.
Leguminosas forrageiras da caatinga: espécies importantes para as comunidades
rurais do sertão da Bahia. Feira de Santana, Universidade Estadual de Feira de Santana,
SASOP.
Esau, K. 1974. Anatomia das plantas com sementes. Trad.: Berta Lange de Morretes. São
Paulo, Edgard Blücher Ltda.
Fahn, A. 1990. Plant anatomy. 4a ed. Oxford, Pergamon Press.
Fedalto, L.C.; Mendes, I. da C.A. & Coradin, V.T.R. 1986. Madeiras da Amazônia.
Descrição do lenho de 40 espécies ocorrentes na floresta nacional de Tapajós.
Brasília, IBAMA.
Figueirôa, J.M.; Pareyn, F.G.C.; Drumond, M.; Araújo, E.L. 2005. Madeireiras. Pp. 101-
1330. In: E.V.S.B. Sampaio, F.G.C. Pareyn, J.M. de Figueirôa & A.G. Santos Jr. (eds.)
Espécies da flora nordestina de importância econômica potencial. Recife, Associação
Plantas do Nordeste (APNE).
Fujiwara, S.; Sameshima, K.; Kuroda, K. & Takamura, N. 1991. Anatomy and properties of
Japanese hardwoods I. Variation of fibre dimensions and tissue proportions and their
relation to basic density. IAWA Bulletin 12: 419-424.
Gasson, P. 1987. Some implications of anatomical variations in the wood of pedunculate oak
(Quercus robur L.) including comparisons with common beech (Fagus sylvatica L.).
IAWA Bulletin 8: 149-166.
Gasson, P.; Trafford, C.; Mathews, B. 2003. Wood anatomy of Caesalpinioideae. Pp. 63-93.
In: B.B. Klitgaard & A. Bruneau (eds.). Advances in Legumes Systematics, part 10.
Kew, Royal Botancic Gardens.
Giulietti, A.M.; Harley, R.M.; Queiroz, L.P. de; Barbosa, M.R. de V.; Neta, A.L.B. &
Figueiredo, M.A. 2002. Espécies endêmicas da caatinga. Pp. 103-176. In: E.V.S.
Sampaio; A.M. Giulietti; J. Virgínio; C.F.L. Gamarra-Rojas (eds.). Vegetação e flora da
caatinga. Recife, Associação Plantas do Nordeste.
Herendeen, P.S. 2000. Structural evolution in the Caesalpinioideae (Leguminosae).Pp. 45-64.
In: P.S. Herendeen & A. Bruneau (eds.). Advances in Legume Systematics, part 9. Kew,
Royal Botanic Gardens.

113
IAWA Committee.1989. IAWA list of microscopic features for hardwood identification. E.
A. Wheeler; P. Baas & P. Gasson (eds.). IAWA Bulletin 10: 219-332.
Jansen, S.; Baas, P.; Gasson, P.; Smets, E. 2003. Vestured pits: do they promote safer water
transport? International Journal of Palnt Sciences 164(3): 405-413.
Jansen, S.; Baas, P.; Gasson, P.; Lens, F.; Smets, E. 2004. Variation in xylem structure from
tropics to tundra: evidence from vestured pits. Proceedings of the National Academy of
Sciences 101(23): 8833-8837.
Johansen, D.A. 1940. Plant microtechnique. New York, McGraw -Hill Book Co.
Lindorf, H. 1994. Eco-anatomical wood features of species from a very dry tropical Forest.
IAWA Journal 15: 361-376.
Loureiro, A.A. & Silva, M.F. 1968. Catálogo das madeiras da Amazônia. Belém.
Ministério do Interior. Superintendência. Desenvolvimento da Amazônia (SUDAM). V.
2.
Luckow, M. 2005. Tribo Mimosaceae. Pp. 163-183. In: G.P. Lewis; B. Schrire; B.
Mackinder; M. Lock (eds.). Legumes of the World. Kew, Royal Botanic Gardens.
Maccari, A. & Marchiori, J.N.C. 1994. Estudo anatômico do xilema secundário de Mimosa
sparsa Benth. Ciência Florestal 4(1): 105-112.
Maia, G.N. 2004. Caatinga, árvores e arbustos e suas utilidades. São Paulo, Leitura &
Arte.
Marchiori, J.N.C. 1982. A estrutura do xilema secundário de Mimosa daleoides Benth.
(Leguminosae-Mimosoideae). Ciência e Natura 4: 107-113.
Marchiori, J.N.C. 1993. Anatomia da madeira e casca do maricá, Mimosa bimucronata (DC.)
O. Kuntze.. Ciência Florestal 3(1): 48-67.
Marchiori, J.N.C. 1996. Anatomia do xilema secundário de Mimosa incana (Spreng.) Benth.
Ciência Florestal 6(1): 53-63.
Marchiori, J.N.C. & Muñiz, G.I.B. de. 1996. Anatomia do xilema secundário de Mimosa
pilulifera Benth. Ciência Florestal 6(1): 65-75.
Olson, M. E. 2005. Commentary: typology, homology and homoplasy in comparative wood
anatomy. IAWA Journal 20(1): 85-93.
Paula, J.E. de. 1989. Estudo da estrutura da madeira de espécies nativas, visando o seu
aproveitamento na produção de energia e papel. Ciência e Cultura 41(4): 344-353.
Paula, J. E. 1993. Madeiras da caatinga úteis para produção de energia. Pesquisa
Agropecuária Brasileira 28(2): 153-165.
Paula, J.E. de. 1995. Anatomia e dendrometria de Mimosa artemisiana e Eucalyptus grandis.
Pesquisa Agropecuária Brasileira 30(6): 745-757.

114
Paula, J.E.; Silva-Júnior, F.G. da; Silva, A.P.P. 2000. Caracterização anatômica de madeiras
nativas de matas ciliares do centro-oeste brasileiro. Scientia Forestalis 58: 73-89.
Paviani, T.I. 1978. Anatomia vegetal e cerrado. Ciência e Cultura 30: 1076-1086.
PROBIO 2000. Seminário sobre avaliação e identificação de ações prioritárias para a
conservação, utilização sustentável e repartição de benefícios da biodiversidade do bioma
caatinga.Disponível em: http://www.biodiversitas.org/caatinga. Acesso em: 20 jul.
2001.
Rao, K.S. & Rajput, K.S. 1999. Seasonal behavior of vascular cambium in teak (Tectona
grandis) growing in moist deciduous and dry deciduous forests. IAWA Journal 20(1):
85-93.
Rao, R.V.; Aebischer, D.P. & Denne, M.P. 1997. Latewood density in relation to wood fibre
diameter, wall thickness, and fibre and vessel percentages in Quercus robur. IAWA
Journal 18: 127-138.
Salvador, P.V.; Diez, P.C.; Rontome, C.P.; Marti, G.M. 1997. Stem xylem features in three
Quercus (Fagaceae) species along a climatic gradient in NE Spain. Trees 12: 90-96.
Sampaio, E.V.S.B.; Salgado, I.H.; Kanffman, J.B. 1993. Effect of different fire severities on
coppicing of caatinga vegetation in Serra Talhada, PE, Brazil. Biotropica 25: 452-460.
Schrire, B.D.; Lewis, G.P.; Lavin, M. 2005. Biogeography of the Leguminosae. Pp. 21-54. In:
G.P. Lewis; B. Schrire; B. Mackinder; M. Lock (eds.). Legumes of the World. Kew,
Royal Botanic Gardens.
Soffiatti, P. & Angyalossy-Alfonso, V. 1999. Estudo anatômico comparativo do lenho e da
casca de duas espécies de Eugenia L. (Myrtaceae). Revista Brasileira de Botânica 22:
275-284.
Vital, B.R. 1984. Métodos de determinação de densidade da Madeira. Viçosa, UFV
Webber, I.E. 1936. The woods of sclerophyllous and desert shrubs of California. American
Journal of Botany 23: 181-188.
Wheeler, E.A. & Baas, P.1991. A survey of the fossil record for dicotyledons wood and its
significance for evolutionary and ecological wood anatomy. IAWA Bulletin 12(3): 275-
332.
Zimmermann, M.H. 1983. Xylem structure and the ascent of sap. Berlin, Springer Verlag.

115
CONCLUSÕES FINAIS

116
CONCLUSÕES FINAIS
Caesalpinia pyramidalis, Croton sonderianus, Mimosa ophthalmocentra na caatinga apresentam demarcações evidentes de camadas de crescimento que devem estar associadas a fatores ambientais, fotoperiodismo e pluviosidade, especialmente porque essas espécies são decíduas, havendo momentos especiais quando as mesmas necessitam de adaptações para sobrevivência.
A presença de delimitação de camadas de crescimento em Croton sonderianus reflete o período de seca pelo qual passa a espécie na região onde ocorre.
A formação e o tamanho dos anéis de crescimento em Croton sonderianus na caatinga estão estreitamente ligados à precipitação pluviométrica e variam na mesma proporção. Anéis mais largos são formados durante os anos de maior disponibilidade de água e anéis mais estreitos, nos anos de baixa precipitação considerados períodos de seca, com redução e/ou cessação da atividade do câmbio (abaixo de 300 mm anual).
Pelos parâmetros anatômicos do lenho do tronco e dos galhos aqui analisados (proporção dos vasos, raios, parênquima e proporção e espessura das paredes das fibras) e a elevada densidade básica (>0,84 g/cm3), conclui-se que Caesalpinia pyramidalis, Croton sonderianus, Mimosa ophthalmocentra e M. tenuiflora, espécies ocorrentes na caatinga, revelam grande quantidade de celulose e lignina, apresentando, portanto perspectivas seguras para produção de álcool combustível, carvão vegetal e coque metalúrgico, conseqüentemente, maior potencial energético, desde a fase do lenho juvenil, visto que nesta região ocorre uma grande concentração de fibras, vasos com menor diâmetro e menor quantidade de parênquima.
Todavia, Croton sonderianus e Mimosa ophthalmocentra, por apresentarem parênquima escasso, têm maior concentração de paredes ricas em celulose, conseqüentemente maior potencial energético.
As duas áreas estudadas de caatinga, Sertânia e Serra Talhada, apresentaram vegetação cujos indivíduos produzem madeiras com potencial energético para sustentar combustão duradoura. Todavia, devido às madeiras de Croton sonderianus de Serra Talhada possuíram maior proporção de fibras e menor proporção de parênquima e vasos do que as de Sertânia, essas apresentaram melhor qualidade para carvão e lenha.
M. ophthalmocentra e M. tenuiflora apresentaram diferenças que as distinguem, expressas através do tipo de casca, coloração do cerne e alburno, elementos anatômicos e densidade do lenho, contribuindo assim para a taxonomia do grupo. Este diagnóstico da madeira dessas quatro espécies fornece importantes subsídios para um plano de manejo e aproveitamento dessas espécies e demonstra a necessidade de se realizar estudos anatômicos e de densidade de outras essências florestais da caatinga nordestina.

ENDEREÇO ATUAL DO AUTOR:
Prof. Lazaro Benedito da Silva
Departamento de Botânica – IBIO
Universidade Federal da Bahia
Campus Universitário de Ondina s/n
40270-190 Salvador – BA
E-mail: [email protected]





























