Embed Size (px)
Citation preview

i
Utilisation and Managementof Medicinal Plants

Utilisation and Managementof Medicinal Plants
– Editor-in-Chief –V.K. Gupta
– Editors –
Anil K. VermaSushma Koul
2010DAYA PUBLISHING HOUSE
Delhi - 110 035
iii

iv
© 2010 EDITORSISBN 81-7035-625-3ISBN 978-81-7035-625-7
All rights reserved. Including the right to translate or to reproduce this book or parts thereof except for briefquotations in critical reviews.
Published by : Daya Publishing House1123/74, Deva Ram ParkTri Nagar, Delhi - 110 035Phone: 27383999Fax: (011) 23260116e-mail : [email protected] : www.dayabooks.com
Showroom : 4760-61/23, Ansari Road, Darya Ganj,New Delhi - 110 002Phone: 23245578, 23244987
Laser Typesetting : Classic Computer ServicesDelhi - 110 035
Printed at : Chawla Offset PrintersDelhi - 110 052
PRINTED IN INDIA

Dedicated to
Prof. C. K. AtalFormer Director, Indian Institute of Integrative Medicine, Jammu
(Erstwhile, Regional Research Laboratory, Jammu)
v

AMITY INSTITUTE FOR HERBALAND BIOTECH PRODUCTS DEVELOPMENT
– An Institution of Ritnand Balved Education Foundation –Thiruvananthapuram
Prof. (Dr.) P. PUSHPANGADANM.Sc, M.Phil, Ph.D., FBRS, FES, FNRS, FNSE, FNESA, FNAASc, FNASc., (UN Equator Initiative Laureate)Director General & Senior Vice President, RBEF(Former Director, NBRI, Lucknow)
No. 3, Ravi Nagar, Ambalamukku Tel: (0471) 3269772, 2432138, 2432148Peroorkada P.O Mob: 0-98950 66816Thiruvananthapuram - 695 005 E-mail: [email protected]
Foreword
With the growing demand and popularity of herbal drugs and the wide use of plant derivedphyto-molecules in modern medicine, in recent times, there has been great demand for medicinalplants in the drug industry.
The gap between demand and supply are widening in the herbal drug industry. Until recently,the demands for raw materials were mostly met from wild naturally growing plants. Owing to variousreasons, particularly because of habitat destruction and shrinking of forest area, it is no longer possibleto meet the requirement of medicinal plants from wild or natural sources. This book–Utilisation andManagement of Medicinal Plants edited by Dr. Anil K. Verma and Dr. Sushma Koul with Dr. V.K. Guptaas Editor-in-chief is a valuable and timely attempt to bring together expert opinions and ideas for theUtilisation and Management of Medicinal Plants in a single volume.
There are twenty one chapters in this book. The editors have invited eminent scientists from allover the world to contribute chapters for the book. The contributed chapters cover a wide range ofsubjects related to the management and utilisation of medicinal plants such as biotechnology,phytochemistry, plant-physiology, fungal metabolites, bioactivity of secondary metabolites, anti-bacterial and anti-fungal activity of secondary metabolites, bio-prospecting etc. Each chapter in thebook is contributed by eminent scientists who have been working in the relevant areas for a considerableperiod.
vii

This book, I am glad to see, is dedicated to Dr. C.K. Atal, a towering figure in the study of medicinalplants and their bioactive secondary metabolites. As Director of Regional Research Laboratory, Jammuwhich is now known as Indian Institute of Integrative Medicine, Dr. Atal inspired a generation ofyoung scientists to devote their life in the study of medicinal and aromatic plants. Under his leadershipand guidance, several scientific discoveries in natural sciences, especially discovery of a large numberof bioactive molecules, novel mechanisms of drug action etc were carried out. He instilled confidenceand inspiration in his colleagues, co-workers and students. Being a visionary with a brilliant, intuitiveand analytical mind, Dr. Atal, encouraged his co-workers to tread untravelled paths which yieldedgreat dividends in terms of research publications, discovery of novel bioactivities, molecules, newdrugs and perfumery products, patents and recognition. Prof. Atal has been known to me from 1969onwards. I had the privilege of working in his team from 1969 to 1989 at IIIM. All these years, I hadvery close association with Prof. (Dr.) C.K. Atal. It is with his blessings and good wishes that I havegrown up in the scientific ladder. Hence I have immense pleasure in writing this foreword to apublication dedicated to Dr. C.K.Atal.
I complement the editors as well as the authors for their valuable contribution. This book will beof immense value and use to students, researchers and professionals engaged in the study andutilisation of medicinal and aromatic plants.
P. Pushpangadan
iviii
No. 3, Ravi Nagar, Ambalamukku Tel: (0471) 3269772, 2432138, 2432148Peroorkada P.O Mob: 0-98950 66816Thiruvananthapuram - 695 005 E-mail: [email protected]

Preface
Nature has provided mankind with products for good health since the beginning of time. Forthousands of years medicine and natural products have been closely linked through the use oftraditional medicines and natural poisons (Newman et al., 2000). Plants hold a prominent position inthe available sources of natural bioactive molecules. Clinical, pharmacological, and chemical studiesof traditional medicines, derived predominantly from plants, were the basis of most early medicinessuch as aspirin, digitoxin, morphine, quinine, emetine, and pilocarpine. Approximately 25 per cent ofthe drugs prescribed worldwide come from plants, whereas 11 per cent of the 252 drugs considered asbasic and essential by the World Health Organization (WHO) derive their origin exclusively fromplants. Prescription drugs containing phytochemicals were valued at more than US$ 30 billion in2002 in the US alone.
Eighty percent of the people in developing countries rely on traditional medicines for primaryhealth care. Development of phyto-therapeutic during last few decades has evolved into a scienceitself and is undergoing further change. The traditional, complementary and the alternative medicineis attracting more and more attention within the context of health care provision and health sectorreforms. Many of the Botanicals as ingredients have histories of prior human. These components arealready being used in food as dietary ingredients, in traditional system of medicine for differentailments or as food and medicine both. Obliviously the food substances are considered to be safe andnourishing, for those which are having medicinal properties and are described for treatment of specificalignments, proper procedure to eliminate adulteration, contamination and toxic side effects are needed.Worldwide need of alternative medicine and growth of natural product inserts the traditional systemsof medicine into markets. The traditional medicines are slowly being integrated into modern medicinein the form of dietary and nutritional supplements and the scientific community is challenged toaddress the issues of utility, safety, quality efficacy and standardization.
ix

During the last few decades, public interest in natural therapies has increased tremendouslyaround the world and natural bioresource as therapeutic has evolved into a science and is undergoingfaster changes. The challenge today is transforming the long history of experience-based medicine inevidence based medicine. Indeed, the lack of readily accessible information on how much of whatmedicinal plant biodiversity occurs where has become a limiting factor in conservation planning andconverting our biowealth into economic wealth. Over harvesting from forests, deforestation and otheranthropogenic factors have already placed several species at risk.
Since new diseases as well as drug-resistant strains of known pathogens continue to emerge, thesearch for novel compounds in combating many diseases and conditions is continuing globally. Thescientific integration of herbal medicinal into modern medical practices must take into account theinterrelated issues of quantitative and qualitative assessment of bioresource, mass production and itsappropriate use. The current worldwide trend towards the utilisation of natural remedied has createdan enormous need for information about the properties and uses of natural resources.
In the present book “Utilisation and Management of Medicinal Plants” an attempt has been made toencapsulate scientific information pertaining to modern-day drug research and this effort is likely toserve as a catalyst for the development of innovative methodologies and approaches for further studies,paving the way for discovery of novel drug for various human ailments.
The topics have been contributed by the experts in the field with exhaustive, relevant and up-to-date information. It is hoped the book be a useful resource of knowledge for ethnobotanists,pharmacologists, biochemists, physiologists, agronomists, medicinal researchers, pharmaceuticalscientists and people of allied disciplines engaged in research new drugs of plant origin. We aregreatly indebted to the scientists and researchers who have contributed for this volume.
V. K. Gupta
Anil K. Verma
Sushma Koul
x

xi
Contents
Foreword vii
Preface ix
1. Biotechnological Interventions for Enhancing the Availability ofHigh Value Medicinal Plants 1
Nishritha Bopana and Sanjay Saxena
2. Environmental Regulation of Secondary Metabolite Accumulation inAromatic Crops: A Review 32
K. Ramesh and Virendra Singh
3. Search for Useful Substances in Papyrus Cyperus papyrus L. 48
Nariaki Wakiuchi, Hajime Tamaki, Takashi Nishino and Yukihiro Sugimoto
4 Secondary Plant Products and their Biopotentials 55
S.C. Jain, R. Singhl and R. Jain
5. Bioactive Compounds from Fungi: A Review 107
Nilanjana Das and Lazar Mathew
6. Elevated Carbon Dioxide Levels Enhance Rosmarinic Acid Productionin Spearmint Plantlets 124
Brent Tisserat, Mark Berhow and Steven F. Vaughn
7. Approaches for Enhancing Yield of Artemisinin: A Novel Antimalarial Compound,in Artemisia annua L. Plants 136
M.Z. Abdin, Mauji Ram, Usha Kiran and M.A. Khan

8. Pieris brassicae as Laboratory of Synthesis of New Compounds with BiologicalPotential: Interaction with Brassica oleracea var. costata and Brassica rapa var. rapa. 157
David M. Pereira, Patrícia Valentão, Federico Ferreres, Rosa Seabra and Paula B. Andrade
9. Medicinal Properties of Mistletoe: A Review 174
P.O. Osadebe, I.C. Uzochukwu and E.O. Omege
10. Comparative Assessment of Growth Productivity and Chemical Profiling ofWild and Cultivated Populations of Withania somnifera (L.) Dunal. 183
Arun Kumar, M.K. Kaul, Punit Kumar Khanna, Sushma Koul and K.A. Suri
11. Bioprospecting: A Move from Land to the Seas 202
Kushal Qanungo
12. The Genus Mentha: A Versatile Plant 217Amrita Chakraborty, Krittika Sasmal and Sharmila Chattopadhyay
13. Medicinal and Aromatic Potentialities of Ocimums and their Efficacy inTraditional Systems of Medicine 228
M.K. Khosla and V.K.Gupta
14. Herbs that can Compact Noise Stress: A Mini Review 244
R. Sheeladevi
15. Bioactive Compounds from Annona Species 278
Beena Joy
16. Saffron: A High Value Crop 294
Sushma Koul, Esha Abrol, Bilal A. Mir and A.K. Koul
17. Antibacterial Activity of Acacia daviesii (M. Bartolome) sp. nov.:A New and Rare Species Identified in North-Eastern Victoria, Australia 306Hilde Lie Kjaerstad, Allison McGill and Enzo A. Palombo
18. A Review on Some Potential Anthelmintic Herbal Drugs 319
Ravindra G. Mali
19. Variations in Tannin and Oxalic Acid Content in Terminalia arjuna (Arjuna) Bark 340
A.K. Pandey and D.C. Kori
20. Natural Sweetening Agents: A Review 348
Annie Shirwaikar, Arun Shirwaikar, Richard Lobo and Kirti S. Prabhu
21. Pseudomonas fluorescens as Microbial Pesticides in Black Pepper:Their Mechanisms of Action 361
Diby Paul and Y.R. Sarma
Index 377
xii

Utilisation and Management of Medicinal Plants402
Medicinal plants have extensive past and present use in treatment of diverse diseases and serveas compounds of interest both in their natural form and as templates for synthetic modifications.According to an estimate of the World Health Organization (WHO), approximately 80 per cent of thepeople in developing countries rely chiefly on traditional medicines for primary health care needs; amajor portion of these involves the use of medicinal plants.
The present book “Utilisation and Management of Medicinal Plants” encompass 21 research andreview articles contributed by eminent researchers/scientists from various parts of the globe andcovers the importance of scientific research on medicinal plants in the areas of botany, chemistry,biotechnology, toxicology and pharmacology. Following are some of the lead articles included in thisbook:
v Biotechnological Interventions for Enhancing the Availability of High Value Medicinal Plants
v Environmental Regulation of Secondary Metabolite Accumulation in Aromatic Crops
v Search for Useful Substances in Papyrus Cyperus papyrus
v Secondary Plant Products and their Biopotentials
v Bioactive Compounds from Fungi
v Approaches for Enhancing Yield of Artemisinin
v Pieris brassicae as Laboratory of Synthesis
v Bioprospecting: A Move from Land to the Seas
v Medicinal and Aromatic Potentialities of Ocimums
v Medicinal Properties of Mistletoe
v Saffron: A High Value Crop
v Some Potential Anthelminitic Herbal Drugs
v Variations in Tannin and Oxalic acid content in Terminalia arjuna
v Natural Sweetening Agents
It is hoped that this book with wide spectrum of themes shall generate further interest and databasefor the benefit of academicians, researchers/scientists working on medicinal plants who shall find itvery useful and indispensable in their relevant research pursuits.

403
Dr. Vijay Kumar Gupta (born 1953-) obtained his Masters (1975) and Doctorate (1979) fromUniversity of Jammu, Jammu-India and is serving as Deputy Director and Head, Animal House,Indian Institute of Integrative Medicine (CSIR), Jammu, India. His research capabilities are substantiatedby his excellent work on histopathology, ecology and reproductive biology of fishes, turtles, birds andmammals, which has already got recognition in India and abroad.
Dr. Gupta has to his credit more than 75 scientific publications and review articles which haveappeared in internationally recognized Indian and foreign journals. Founder fellow, life member andoffice bearer of many national societies, academies and associations. He has successfully completed anumber of research/consultancy projects funded by various governments, private and multinationalagencies. His current areas of interest are histopathology, toxicology, pre-clinical safety pharmacologyand reproductive efficacy studies of laboratory animals.
He is also Editor-in-chief of the book series “Perspectives in Animal Ecology and Reproduction”a Daya Publications, New Delhi, India. The Editor-in-chief of the American Biographical Institute,USA, has appointed him as Consulting Editor of The Contemporary Who’s Who. Dr. Gupta recently alsoappointed as Nominee for the Committee for the Purpose of Control and Supervision of Experiments onAnimals (CPCSEA, Govt. of India).
Dr. Anil K. Verma, Ph.D., M.N.A.Sc., FLS, London (born 1963-) Sr. Grade Lecturer, Department ofZoology, Govt. College for Women (P.G.), Gandhi Nagar, Jammu, J&K State, did his M.Sc. in Zoology(1986) from University of Jammu, Jammu. He has undergone his M.Phil. (1988) and awarded first rankand Ph.D.(1993) in the field of animal reproduction at the same University and has published about50 research papers and review articles in reputed journals and books. He is also a member EditorialBoard of the book series “Advances in Fish and Wildlife: Ecology and Biology” a Daya Publications,New Delhi. In recognition of his standing in greater scientific community, the Board of Directors of theAmerican Association for the advancement of science (AAAS) New York, Washington, has awardedmembership to him. Recently the Linnaean Society of London, U.K . has awarded fellowship to him inOctober 2006 in recognition of his contribution towards the cultivation of Science in Natural History.
Dr Sushma Koul (born 1954-) obtained her Masters and Doctorate from University of Jammu,Jammu-India and is serving as Deputy Director in Indian Institute of Integrative Medicine (CSIR),Jammu. She has contributed more than 50 research papers and review articles, guided 4 Ph.D. studentsduring her 30 years of Scientific career in the field of Medicinal Plant Biotechnology. She has visitedUniversity of Tubingen (Germany) under INSA-DAD exchange programme and has worked with ProfE.Reinhard in Pharmazeutisches Institute of Tubingen University. Her main areas of research aremicropropagation, bioreactor cultivation, conservation biology, adaptive biology and bioprospectingof bioactive molecules.

Chapter 8
Pieris brassicae as Laboratory ofSynthesis of New Compounds with
Biological Potential: Interaction withBrassica oleracea var. costata and
Brassica rapa var. rapa.David M. Pereira1, Patrícia Valentão1, Federico Ferreres2,
Rosa Seabra1 and Paula B. Andrade1*1REQUIMTE/Serviço de Farmacognosia, Faculdade de Farmácia, Universidade do
Porto, R. Aníbal Cunha, 164, 4050-047 Porto, Portugal2Research Group on Quality, Safety and Bioactivity of Plant Foods,
Department of Food Science and Technology, CEBAS (CSIC), P.O. Box 164, 30100Campus University Espinardo, Murcia, Spain
ABSTRACT
Interactions between insects and plants have been studied for a long time, showing beneficialand deleterious effects for both organisms. A general overview about the relationship betweenglucosinolates, flavonoids and insects will be presented. Several aspects of the complex interactionsestablished by cabbage white butterfly (Pieris brassicae L.; Lepidoptera: Pieridae) are alsoconsidered. The larvae of this species constitute a frequent pest of some Brassica species, whichare an important way of subsistence in several countries. As insects are unable to synthesize
————————* Corresponding Author: E-mail: [email protected].
Utilisation and Management of Medicinal Plants (2010) Pages 157–173Editors: V.K. Gupta, Anil K. Verma and Sushma KoulPublished by: DAYA PUBLISHING HOUSE, NEW DELHI

Utilisation and Management of Medicinal Plants158
phenolic compounds or their precursors, their presence in the different stages of P. brassicae lifecycle can only arise from the food it has ingested. Deacylation, deglycosylation and sulphatingsteps are known to be involved in the metabolic process. Thus, with this study we intend todemonstrate the possible use of P. brassicae as a source of compounds with interest for thehealth, unusual in nature and hard to be synthesized in the laboratory. Two examples will bepresented: the sequestration of phenolics by P. brassicae larvae fed with tronchuda cabbageleaves (Brassica oleracea L. var. costata DC) and kept without food for one hour, and the phenolicprofiles of P. brassicae at different development stages (larvae, exuviae and butterfly), itsexcrements and its host plant Brassica rapa var. rapa L.
Keywords: Brassica oleracea var. costata, Brassica oleracea var. rapa, Flavonoids, Glucosinolates, Insect-plant interactions,Pieris brassicae.
IntroductionThe role of plant chemistry in shaping plant-insect relationships is well recognized, with a close
association of certain oligophagous insects with specific chemicals of their host plants (Renwick,2002).
Large white butterfly Pieris brassicae L. (Lepidoptera: Pieridae), an insect whose larvae constitutesa frequent pest of some Brassica species, has a life cycle that lasts about 45 days from egg to adult. Thelarvae feed exclusively on crucifers (namely, cauliflower, cabbage, turnip, nasturtium and, more rarely,on red cabbage and radish), while adults feed on the nectar of several plants (Renwick, 2002; Murieland Grez, 2002).
Herein, it will be presented a general overview about the relationship between glucosinolates,flavonoids and insects, with special emphasis on P. brassicae. Additionally, two examples of theinteraction between this insect with two different host plants (Brassica oleraceae L. var. costata DC andBrassica rapa var. rapa L.) will be presented. These examples reveal that those interactions producedinteresting bioactive compounds.
Interactions Insect-Plant
GlucosinolatesGlucosinolates (Figure 8.1) are β-thioglucoside N-
hydroxysulfates [also known as (Z)-(or cis )-N-hydroximinosulfate esters or S-glucopyranosylthiohydroximates], with a side chain (R) and a sulfur-linked β-d-glucopyranose moiety. The side chain (R) ischaracterized by a wide variety of chemical structures,which depends on the aminoacid precursor. So, they aresubdivided into three classes: aliphatic, indolic andaromatic (Fahey et al., 2001).
Glucosinolates have frequently been quoted as both a defense against generalist herbivores andan agent for host choice by specialist herbivores (Moyes et al., 2000).
In what concerns to plant-insect interactions, the most important reaction of glucosinolates istheir hydrolysis by the enzyme myrosinase, with the production of compounds, including
O S
N
O
S
OO
O-
HO
OH
HO OH
R
Figure 8.1: Chemical Structure ofGlucosinolates

159
isothiocyanates, referred as the mustard oils. Glucosinolates and these volatile compounds are largelyresponsible for the close association between crucifers and their specialist insect invaders.Isothiocyanates may serve to attract specialist insects to their hosts, whereas glucosinolates oftentrigger oviposition or feeding after an insect lands on the plant (Renwick, 2002). Once glucosinolates,or their hydrolysis products, are generally toxic to non-adapted insects, specialists have ways toprevent the potential toxicity of these compounds. This may occur by rapid excretion, glucosideshydrolysis, inhibition of hydrolysis, the action of protective enzymes, or by sequestering theglucosinolates (Schoonhoven et al., 1998). This association was already demonstrated in the specificcase of Pieris sp. larvae (Renwick, 2002).
In a general way, sequestration of plant toxic compounds in herbivores is correlated withaposematic coloration and gregarious behaviour. Once larvae of P. brassicae present these characteristics,it was suggested that it sequester glucosinolates of their host plants. In opposition, Pieris rapae L.(Lepidoptera: Pieridae) are camouflaged and solitary, so there‘s no expectation of occurrence of thatsequestration. To test this hypothesis and to check the repeatability of a study that did report thepresence of the glucosinolate sinigrin (Figure 8.2A) in P. brassicae, Müller et al. (2003) analysed theglucosinolate composition of larvae reared on three species of Brassicaceae (Sinapis alba, Brassica nigraand Barbarea stricta). Host plant glucosinolates were found only in traces or not at all in the larvae of P.rapae and of P. brassicae reared on S. alba, B. nigra or B. stricta. Thus, the larvae of both species do notsequester these secondary metabolites from their host plants. The results obtained by Aplin et al.(1975) indicating the accumulation of sinigrin in P. brassicae pupae wasn‘t confirmed. Also, the authorsdidn‘t find a correlation between glucosinolate sequestration and aposematism or gregariousness inthe two Pieris spp.
The explanation for the presence of glucosinolates only in vestigial traces in larvae and faeces ofboth Pieris species, was their hydrolysis by myrosinase present in the ingested leaf tissue and/ormetabolisation by the insect. Larvae feeding on S. alba excreted a progenitor of 4-hydroxybenzylcyanide(HBC) in the faeces. One possible justification for the detection of HBC after sulfatase treatment of
O S
N
O
S
OO
O-
HO
OH
HO OH
OH
O S
N
O
S
OO
O-
HO
OH
HO OH
(A) (B)
Figure 8.2: Chemical Structures of Sinigrin (A) and Sinalbin (B)
Pieris brassicae as Laboratory of Synthesis of New Compounds with Biological Potential

Utilisation and Management of Medicinal Plants160
faecal extracts is that sinalbin, in the Pieris digestive tract or body, is hydrolysed into HBC and thatenzymatic conversion of HBC into HBC-sulfate occurs subsequently. As Müller et al. (2003) referred,conjugation of phenols (case of sinalbin, Figure 8.2A) with sulfate is a detoxification path, increasingwater solubility, which allows the excretion of a dietary compound, an example of the so called ‘Phase2 Metabolism’ (Brattsten 1992).
Later, it was demonstrate by Agerbirk et al . (2006) that caterpillars of P. rapae convert4-hydroxybenzylglucosinolate (sinalbin, Figure 8.2B) of brassicaceous plants into4-hydroxybenzylcyanide sulfate (HBC sulfate), having 4-hydroxybenzylcyanide (HBC) asintermediate. This apparently serves as a detoxification process, as alternative formation of a mustardoil is avoided.
It would be interesting to assess the biological activity of these products of detoxification ofglucosinolates by insects, namely by P. brassicae.
FlavonoidsLike glucosinolates, flavonoids can influence the feeding behaviour of larvae and oviposition of
adult insects (van Loon et al., 2002).
Flavonoids uptake is relatively diffused in the Lepidoptera, namely in butterfly families asPapilionidae, Nymphalidae and Lycaenidae, in which they participate in wing pigmentation(Burghardt et al., 1997, 2001; Schittko et al., 1999). Actually, although most pigments are likely to besynthesised de novo during scale development in the pupa, others are secondary plant metabolitesobtained from the larval diet once insects are incapable to synthesise flavonoids or their precursors(Knüttel and Fiedler, 2001). Several studies confirmed that flavonoids in insects arise from the diet(Harborne and Grayer, 1994; Burghardt et al., 1997; Schittko et al., 1999; Knüttel and Fiedler, 2001). So,flavonoid uptake and metabolization is effectively depending on the specific flavonoid pattern of hostplants (Burghardt et al., 1997, 2001; Geuder et al., 1997; Schittko et al., 1999).
Flavonoids sequestered by the larvae are later biotransformed, retained and shifted to the wingsduring the late pupal stage (Geuder et al., 1997). The antioxidant capacity of flavonoids is wellrecognized (Ferreres et al., 2006; Vrchovská et al., 2006) and can play different roles in insects, workingas antibiotic and antiviral (Harborne and Grayer, 1994).
Regarding flavonoid patterns, it was demonstrated that only part of the flavonoidic compoundsof host plant are sequestered by larvae, with the uptaken flavonoids being object of several glycosylationreactions. In addition, butterflies belonging to the same species can show distinct flavonoid composition(related to the host plant used in the larval phases). Also, there is a tendency for female butterflies to bemore rich in flavonoids than males (Burghardt et al., 2001).
According to the above mentioned, flavonoids in insects are effectively allied to the contents offlavonoids in their food (Burghardt et al., 2000). Larvae revealed a preference for sequestering andmetabolising quercetin and kaempferol derivatives, the main flavonoids in the analysed plants. Otherflavonoids like myricetin derivatives, flavones and isoflavonoids were mostly excreted (Burghardt etal., 2001).
As far as we know, there are only two studies concerning the sequestration of phenolic compoundsby P. brassicae from Brassica leaves: one about flavonoids uptake by P. brassicae from B. oleracea var.costata (tronchuda cabbage) and another involving the phenolic compounds in P. brassicae reared on B.rapa var rapa (turnip leaves).

161
These studies can be important, attending to the fact that the larvae may accumulate and/ormetabolize host plant constituents, namely complex flavonol glycosides (Ferreres et al., 2005, 2006),constituting a source of potential bioactive compounds not available in nature.
Flavonoid Pattern of Larvae ofPieris brassicae Reared on Brassicaoleraceae var. costata
The flavonoid pattern of larvae ofcabbage white butterfly (P. brassicae )(Figure 8.3) reared on the leaves oftronchuda cabbage was analyzed byHPLC-DAD-MS/MS-ESI, a highlyadvanced and valuable technique for thecharacterization of complex phenolicmolecules (Ferreres et al., 2007a).
Wild P. brassicae larvae (fourth instar)and respective tronchuda cabbageexternal leaves (from three individualswith 45 days-old) host plants werecollected on fields located in Samil,Bragança, northeastern Portugal.
The phenolic composition of thetronchuda cabbage external leaves hadalready been studied in a previous work(Ferreres et al., 2005). The composition ofthe host leaves, from which the larvaefeed, revealed to be similar to thatdescribed before, being detected thirteenkaempferol derivatives (Table 8.1). Theflavonoid profile obtained with P. brassicae(Figures 8.4 and 8.5) was then comparedwith that of the cabbage. Kaempferol-3-O - s o p h o r o s i d e - 7 -O - g l u c o s i d e ,kaempferol-3-O-sophoroside-7-O-sophoroside and kaempferol-3-O-sophoroside were the onlycompounds that the larvae and cabbage had in common.
Although the glycosylation pattern of the flavonols was the same in both extracts, it was observedthat the flavonol 3-O-glycosides represented more than ca. 50 per cent in the larvae extract (Table 8.2),while they corresponded to ca.12 per cent of their food plant (Table 8.1). This was ascribed to themetabolism of the flavonols glycosylated at 3 and 7 positions present on tronchuda cabbage, or to ahigher efficiency of sequestration of flavonol-3-O-glycosides (Ferreres et al., 2007a).
Kaempferol-3-O-sophoroside was the most abundant flavonol glycoside derivative withoutacylation, corresponding to ca. 16 per cent of the total amount of phenolic compounds of the larvae(Table 8.2), while in tronchuda cabbage leaves it only represented ca. 5 per cent (Table 8.1). Thisdifference can result from the metabolism of kaempferol-3-O-sophoroside-7-O-glucoside and its acylatedderivatives, which are the most abundant compounds of tronchuda cabbage external leaves.
Figure 8.3: Brassica oleraceae var. costataPlant Material (A), Infested with Pieris brassicae
Larvae (B), Pieris brassicae Larvae
(A)
(B)
Pieris brassicae as Laboratory of Synthesis of New Compounds with Biological Potential

Utilisation and Management of Medicinal Plants162
Table 8.1: Phenolic Composition of Brassica oleracea var. costata External Leaves(Ferreres et al., 2007a)
Compound Per cent
21 + Kaempferol-3-O-sophorotrioside-7-O-glucoside 7.6
22 Kaempferol-3-O-(methoxycaffeoyl/caffeoyl)-sophoroside-7-O-glucoside
2 Kaempferol-3-O-sophoroside-7-O-glucoside 22.9
23 Kaempferol-3-O-sophorotrioside-7-O-sophoroside 1.4
3 + Kaempferol-3-O-sophoroside-7-O-sophoroside 11.4
24 Kaempferol-3-O-tetraglucoside-7-O-sophoroside
25 Kaempferol-3-O-(sinapoyl/caffeoyl)-sophoroside-7-O-glucoside 17.1
26 Kaempferol-3-O–(feruloyl/caffeoyl)-sophoroside-7-O-glucoside 27.8
27 + Kaempferol-3-O-sophorotrioside 5.1
28 Kaempferol-3-O-(sinapoyl)-sophoroside
29 Kaempferol-3-O-(feruloyl)-sophorotrioside 0.4
30 Kaempferol-3-O-(feruloyl)-sophoroside 1.1
15 Kaempferol-3-O-sophoroside 5.2
Table 8.2: Phenolic Composition of P. brassicae (Ferreres et al., 2007a)
Compound Per cent
1 Quercetin-3-O-sophoroside-7-O-glucoside 8.7
2 Kaempferol-3-O-sophoroside-7-O-glucoside 10.0
3 Kaempferol-3-O-sophoroside-7-O-sophoroside 6.6
4 Quercetin-3-O-(feruloyl)-triglucoside-7-O-glucoside 4.5
5 Kaempferol-3-O-(sinapoyl)-triglucoside-7-O-glucoside 5.0
6 Kaempferol-3-O-(feruloyl)-triglucoside-7-O-glucoside 5.6
7 Kaempferol-3-O-(p-coumaroyl)-triglucoside-7-O-glucoside 2.6
8 Kaempferol-3-O-(methoxycaffeoyl)-sophoroside-7-O-glucoside 0.5
9 Kaempferol-3-O-(caffeoyl)-sophoroside-7-O-glucoside 1.8
10 Quercetin-3-O-(p-coumaroyl)-sophoroside 3.4
11 Kaempferol-3-O-(p-coumaroyl)–triglucoside 3.3
12 Kaempferol-3-O-(p-coumaroyl)-sophoroside 13.4
13 + Kaempferol-3-O-(methoxycaffeoyl)-sophoroside 9.2
14 Quercetin-3-O-sophoroside
15 Kaempferol-3-O-sophoroside 15.8
16 Kaempferol-3-O-(p-coumaroyl)-sophoroside (isomer) 2.4
17 Kaempferol-3-O-(disinapoyl)-triglucoside-7-O-glucoside 2.1
18 Kaempferol-3-O-(feruloyl/sinapoyl)-triglucoside-7-O-glucoside 1.3
19 Quercetin-3-O-(feruloyl)-triglucoside 1.9
20 Kaempferol-3-O-glucoside 1.9

163
In what concerns the presence of quercetin derivatives, the authors mention that they have re-analysed the composition of the external leaves of tronchuda cabbage (Ferreres et al., 2005) and theyhad detected these compounds in vestigial amounts, which also happened with the tronchuda cabbageexternal leaves eaten by P. brassicae. The larvae contained high amounts of quercetin derivatives (ca. 18per cent of the total amount of phenolic compounds) (Table 8.2), while in tronchuda cabbage thesecompounds were present only in trace amounts, which suggests that P. brassicae selectively sequestersthese flavonoids or that the kaempferol glycosides are metabolised into quercetin glycosides by thelarvae.
The presence of p-coumaroyl derivatives (kaempferol-3-O-(p-coumaroyl)-triglucoside-7-O-glucoside, quercetin-3-O-(p-coumaroyl)-sophoroside, kaempferol-3-O-(p-coumaroyl)–triglucoside,kaempferol-3-O-(p-coumaroyl)-sophoroside and respective isomer) in larvae aqueous extract (Figure8.2), which had not been found on either the internal or external leaves of tronchuda cabbage (Ferrereset al., 2005; Sousa et al., 2005) were explained by the demethoxylation of the sinapoyl and/or feruloyl
Figure 8.4: HPLC-DAD Phenolic Profile of Pieris brassicae Larvae HydromethanolicExtract, Reared with B. oleracea var. costata.
Detection at 330 nm. Peaks: (1) quercetin-3-O-sophoroside-7-O-glucoside; (2) kaempferol-3-O-sophoroside-7-O-glucoside; (3) kaempferol-3-O-sophoroside-7-O-sophoroside; (4) quercetin-3-O-(feruloyl)-triglucoside-7-O-glucoside; (5) kaempferol-3-O-(sinapoyl)-triglucoside-7-O-glucoside; (6)kaempferol-3-O-(feruloyl)-triglucoside-7-O-glucoside; (7) kaempferol-3-O-(p-coumaroyl)-triglucoside-7-O-glucoside; (8) kaempferol-3-O-(methoxycaffeoyl)-sophoroside-7-O-glucoside; (9) kaempferol-3-O-(caffeoyl)-sophoroside-7-O-glucoside; (10) quercetin-3-O-(p-coumaroyl)-sophoroside; (11)kaempferol-3-O-(p-coumaroyl)–triglucoside; (12) kaempferol-3-O-(p-coumaroyl)-sophoroside; (13)kaempferol-3-O-(methoxycaffeoyl)-sophoroside; (14) quercetin-3-O-sophoroside; (15) kaempferol-3-O-sophoroside; (16) kaempferol-3-O-(p-coumaroyl)-sophoroside (isomer); (17) kaempferol-3-O-(disinapoyl)-triglucoside-7-O-glucoside; (18) kaempferol-3-O-(feruloyl/sinapoyl)-triglucoside-7-O-glucoside; (19) quercetin-3-O-(feruloyl)-triglucoside; (20) kaempferol-3-O-glucoside (Ferreres et al.,2007a).
Pieris brassicae as Laboratory of Synthesis of New Compounds with Biological Potential

Utilisation and Management of Medicinal Plants164
OH
OH O
OOH
OR1
R2O
OH
OH O
O
OR1
R2O
1 R1 = sophoroside; R2 = glucoside4 R1 = (feruloyl)triglucoside; R2 = glucoside10 R1 = (p-coumaroyl)sophorosido; R2 = H14 R1 = sophoroside; R2 = H19 R1 = (feruloyl)triglucoside; R2 = H
2 R1 = sophoroside; R2 = glucoside3 R1 = sophoroside; R2 = sophoroside5 R1 = (sinapoyl)triglucoside; R2 = glucoside6 R1 = (feruloyl)triglucoside; R2 = glucoside7 R1 = (p-coumaroil)triglucoside; R2 = glucoside8 R1 = (methoxycaffeoyl)sophoroside; R2 = glucoside9 R1 = (caffeoyl)sophoroside; R2 = glucoside11 R1 = (p-coumaroyl)triglucoside; R2 = H12, 16 R1 = (p-coumaroyl)sophoroside; R2 = H13 R1 = (methoxycaffeoyl)sophoroside; R2 = H15 R1 = sophoroside; R2 = H17 R1 = (disinapoyl)triglucoside; R2 = glucoside18 R1 = (feruloyl/sinapoyl)triglucoside; R2 = glucoside20 R1 = glucoside; R2 = H21 R1 = sophorotrioside; R2 = glucoside22 R1 = (methoxycaffeoyl/caffeoyl)sophoroside; R2 = glucoside23 R1 = sophorotrioside; R2 = sophoroside24 R1 = tetraglucoside; R2 = sophoroside25 R1 = (sinapoyl/caffeoyl)sophoroside; R2 = glucoside26 R1 = (feruloyl/caffeoyl)sophoroside; R2 = glucoside27 R1 = sophorotrioside; R2 = H28 R1 = (sinapoyl)sophoroside; R2 = H29 R1 = (feruloyl)sophorotrioside; R2 = H30 R1 = (feruloyl)sophoroside; R2 = H
Figure 8.5: Structures of the Phenolic Compounds Identified in Pieris brassicae andBrassica oleracea var. costata. Identity of compounds as in Tables 8.1 and 8.2.
derivatives during the metabolism process in the larvae. On the other hand, Ferreres et al. (2007)referred that the absorbance of the peaks observed in Figure 4 for acylated flavonoid derivatives, couldnot be taken as proportional to their abundance, as some of them co-eluted with other unidentifiedcinnamoyl acids’ derivatives, presenting a similar UV spectrum and contributing to the overallabsorbance of those peaks. Another advanced explanation was that, despite their existence intronchuda cabbage leaves in concentrations below the detection limits, P. brassicae selective uptakesand accumulates them.

165
The existence of two methoxylated flavonol derivatives in P. brassicae (kaempferol-3-O-(methoxycaffeoyl)-sophoroside-7-O-glucoside and kaempferol-3-O-(methoxycaffeoyl)-sophoroside)(Figure 8.4) were explained as a result from the metabolism of kaempferol-3-O-(methoxycaffeoyl/caffeoyl)-sophoroside-7-O-glucoside present on the external leaves of the tronchuda cabbage.
Phenolics in Pieris brassicae Related on Brassica rapa var. rapaThe aim of the study of Ferreres et al. (2008) was to characterize the phenolic compounds of B. rapa
var. rapa leaves and to establish possible relations with their ingestion, metabolism and accumulationby P. brassicae in the different stages of its life cycle (Figure 8.6). For this purpose, turnip leaves, P.brassicae larvae reared on this leaves and deprived of food for 12 hours, their excrements, exuviae andbutterflies were analysed by HPLC-DAD-MS/MS-ESI.
In B. rapa var. rapa leaves it were characterized for all the acylated derivatives (methoxycaffeic,caffeic, sinapic, ferulic and p-coumaric acids) of quercetin-3-O-sophoroside-7-O-glucoside, kaempferol-3-O-sophoroside-7-O-glucoside and kaempferol-3-O-sophoroside (Figure 8.7). The kaempferol-3-O-sophoroside-7-O-glucoside derivatives are the most abundant ones (Figure 8.7B), while those ofkaempferol-3-O-sophoroside were present in trace amounts. Quercetin-3-O-sophoroside-7-O-glucosidederivatives are found in considerable contents (Figure 8.7B). However, the deacylated glycosides weredetected only in vestigial amounts in the saponificated hydromethanolic extract (Figure 8.7A). Thiscan be due to the alkaline decomposition, during the saponification process, of phenolic compoundswith an o-dihydroxy group, resulting in the presence of quercetin derivatives in trace amounts whilecaffeic acid is not observed.
In general, this kind of acylated derivatives (Figures 8.7B and 8.10) are very common in Brassicacea(Llorach et al., 2003, Vallejo et al., 2004, Ferreres et al., 2005, 2006 and 2007b), and particularly indistinct B. rapa subspecies (Romani et al., 2006; Rochfort et al., 2006). On the other hand, the presenceof flavonol-3,7-di-O-glucosides, namely non-acylated isorhamnetin-3,7-di-O-glucoside characterizesB. rapa (Fernandes et al, 2007; Romani et al., 2006; Rochfort et al., 2006) relatively to other Brassicaspecies. Despite contributing to the organoleptic characteristics of the plant, these compounds mayparticipate in the defence against external aggressions.
The two main phenolic compounds in excrements’ hydromethanolic extract (Figure 8.8) wereferulic and sinapic acids (Figure 8.10). Other compounds present in considerable amounts, and thathave been found before in the native hydromethanolic extract of B. rapa var. rapa leaves, were kaempferol-3-O-sophoroside, isorhamnetin-3,7-di-O-glucoside and isorhamnetin-3-O-glucoside. Some of theacylated derivatives already described were also detected, in low or trace amounts: kaempferol-3-O-(caffeoyl)sophoroside-7-O-glucoside, quercetin-3-O-(sinapoyl)sophoroside-7-O-glucoside, quercetin-3-O-(feruloyl)sophoroside-7-O-glucoside, quercetin-3-O-(p -coumaroyl)sophoroside-7-O-glucoside),kaempferol-3-O-(feruloyl)sophoroside-7-O-glucoside, kaempferol-3-O-(p-coumaroyl)sophoroside-7-O-glucoside and kaempferol-3-O-(p-coumaroyl)sophoroside. Other compounds that were not found inthe native extract of the leaves were kaempferol-3-O-sophoroside-7-O-glucoside, described in thesaponificated extract, quercetin-3-O-sophoroside, kaempferol-3-O-sophorotrioside and three isomersof kaempferol-3-O-(p-coumaroyl)sophoroside. In the first part of the chromatogram (Figure 8.8) severalflavonoids derivatives were detected and were identified as sulphate flavonoids: isorhamnetin-3,7-di-O-glucoside sulphate and monoglucosides (kaempferol-3-O-glucoside sulphate isomers andisorhamnetin-3-O-glucoside sulphate isomers). This kind of compounds is very usual in animals’metabolic process. Considering the obtained results it was inferred that during the metabolic processof P. brassicae it occurs the deacylation of flavonoids, leading to the disappearance or decrease of
Pieris brassicae as Laboratory of Synthesis of New Compounds with Biological Potential

Utilisation and Management of Medicinal Plants166
Figure 8.6: Pieris brassicae Material Rearedon B. rapa var. rapa Leaves
A: B. rapa var. rapa leaves;B: P. brassicae larvae;C: P. brassicae excrements;D: P. brassicae exuviae;E: P. brassicae butterfly.
(A) (B)
(C) (D)
(E)

167
H o j a R . D : U V C h r o m a t o g r a m , 3 3 0 . 1 0 0 n m
0
1 0 0
2 0 0
3 0 0
I n t e n s .
0 5 1 0 1 5 2 0 2 5 3 0 3 5 T i m e [ m i n ]
h o j a s a p . D : U V C h r o m a t o g r a m , 3 3 0 . 1 0 0 n m
0
1 0 0
2 0 0
3 0 0
4 0 0
I n t e n s .
0 5 1 0 1 5 2 0 2 5 3 0 3 5 T i m e [ m i n ]
1 2
3
456
87 9
1011 12 13
14 15
16
17
18
20
1921
2223
24
76
89
25+2627
2829
10+30
31
12 13
A
B
Figure 8.7: HPLC-DAD Phenolic Profile of Brassica rapa var. rapa Leaves.(A) saponified hydromethanolic extract and (B) native hydromethanolic extract.
Detection at 330 nm. Peaks: (1) quercetin-3-O-sophoroside-7-O-glucoside; (2) kaempferol-3-O-sophorotrioside-7-O-glucoside; (3) kaempferol-3-O-sophoroside-7-O-glucoside; (4) quercetin-3,7-di-O-glucoside; (5) p-coumaric acid; (6) kaempferol-3,7-di-O-glucoside; (7) isorhamnetin-3,7-di-O-glucoside; (8) ferulic acid; (9) sinapic acid; (10) kaempferol-3-O-sophoroside; (11) kaempferol-7-O-glucoside; (12) kaempferol-3-O-glucoside; (13) isorhamnetin-3-O-glucoside; (14) quercetin-3-O-(methoxycaffeoyl)sophoroside-7-O-glucoside; (15) quercetin-3-O-(caffeoyl)sophoroside-7-O-glucoside; (16) kaempferol-3-O-(methoxycaffeoyl)sophoroside-7-O-glucoside; (17) kaempferol-3-O-(caffeoyl)sophoroside-7-O-glucoside; (18) quercetin-3-O-(sinapoyl)sophoroside-7-O-glucoside; (19)quercetin-3-O-(feruloyl)sophoroside-7-O-glucoside; (20) quercetin-3-O-(p-coumaroyl)sophoroside-7-O-glucoside; (21) kaempferol-3-O-(sinapoyl)sophoroside-7-O-glucoside; (22) kaempferol-3-O-(feruloyl)sophoroside-7-O-glucoside; (23) kaempferol-3-O-(p-coumaroyl)sophoroside-7-O-glucoside;(24) quercetin-3- O-(caffeoyl)sophoroside-7-O-glucoside (isomer); (25) kaempferol-3- O-(feruloyl)sophoroside-7-O-glucoside (isomer); (26) kaempferol-3-O-(methoxycaffeoyl)sophoroside;(27) kaempferol-3-O-(p-coumaroyl)sophoroside-7-O-glucoside (isomer); (28) kaempferol-3-O-(caffeoyl)sophoroside; (29) kaempferol-3-O-(sinapoyl)sophoroside; (30) kaempferol-3-O-(feruloyl)sophoroside; (31) kaempferol-3-O-(p-coumaroyl)sophoroside (Ferreres et al., 2008).
acylated derivatives. Besides this, the absence of glycosilation in the 7 position in the majority offlavonoid sulphates, as well as in the remaining compounds, indicates the loss of the sugar in thisposition, together with the above mentioned deacylation, in the derivatives of kaempferol-3-O-sophoroside-7-O-glucoside and of quercetin-3-O-sophoroside-7-O-glucoside and in kaempferol-3-O-
Pieris brassicae as Laboratory of Synthesis of New Compounds with Biological Potential
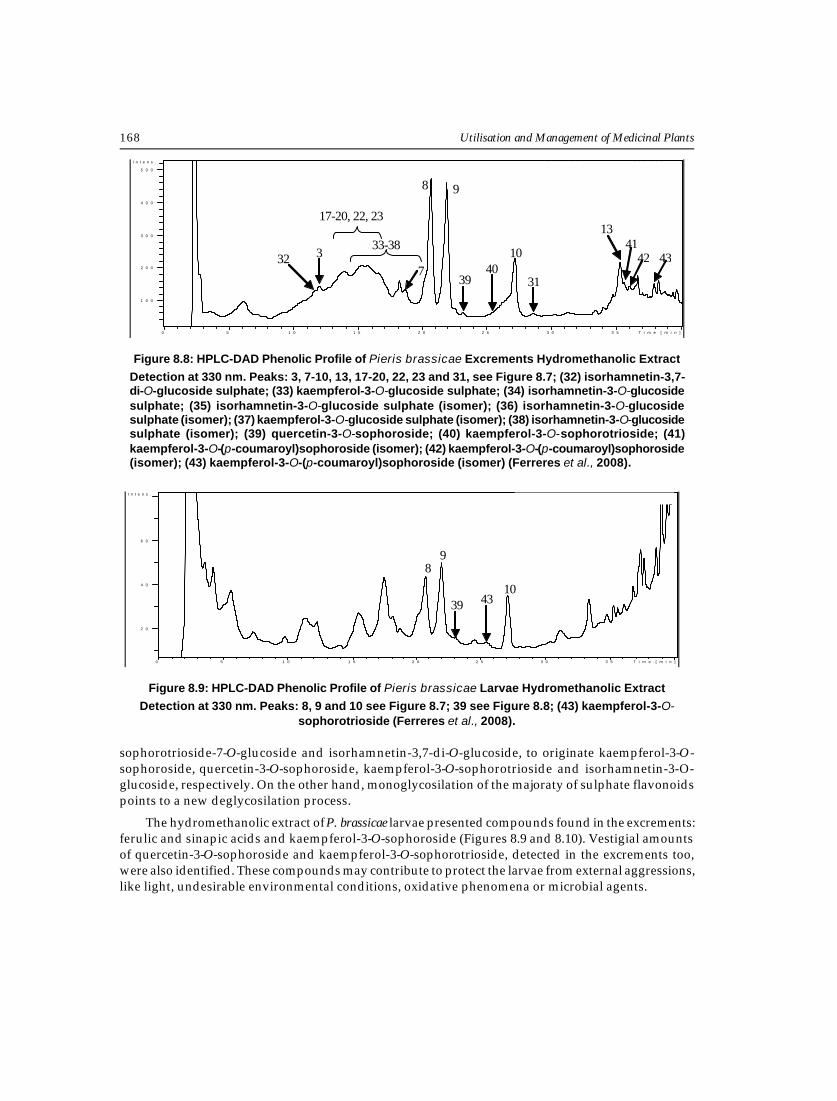
Utilisation and Management of Medicinal Plants168
sophorotrioside-7-O-glucoside and isorhamnetin-3,7-di-O-glucoside, to originate kaempferol-3-O-sophoroside, quercetin-3-O-sophoroside, kaempferol-3-O-sophorotrioside and isorhamnetin-3-O-glucoside, respectively. On the other hand, monoglycosilation of the majoraty of sulphate flavonoidspoints to a new deglycosilation process.
The hydromethanolic extract of P. brassicae larvae presented compounds found in the excrements:ferulic and sinapic acids and kaempferol-3-O-sophoroside (Figures 8.9 and 8.10). Vestigial amountsof quercetin-3-O-sophoroside and kaempferol-3-O-sophorotrioside, detected in the excrements too,were also identified. These compounds may contribute to protect the larvae from external aggressions,like light, undesirable environmental conditions, oxidative phenomena or microbial agents.
e x c r e m e n t o s . D : U V C h r o m a t o g r a m , 3 3 0 . 1 0 0 n m
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
I n t e n s .
0 5 1 0 1 5 2 0 2 5 3 0 3 5 T i m e [ m i n ]
3
8 9
10
31
13
3233-38
39
17-20, 22, 23
7
4142 43
40
Figure 8.8: HPLC-DAD Phenolic Profile of Pieris brassicae Excrements Hydromethanolic Extract
Detection at 330 nm. Peaks: 3, 7-10, 13, 17-20, 22, 23 and 31, see Figure 8.7; (32) isorhamnetin-3,7-di-O-glucoside sulphate; (33) kaempferol-3-O-glucoside sulphate; (34) isorhamnetin-3-O-glucosidesulphate; (35) isorhamnetin-3-O-glucoside sulphate (isomer); (36) isorhamnetin-3-O-glucosidesulphate (isomer); (37) kaempferol-3-O-glucoside sulphate (isomer); (38) isorhamnetin-3-O-glucosidesulphate (isomer); (39) quercetin-3-O-sophoroside; (40) kaempferol-3-O-sophorotrioside; (41)kaempferol-3-O-(p-coumaroyl)sophoroside (isomer); (42) kaempferol-3-O-(p-coumaroyl)sophoroside(isomer); (43) kaempferol-3-O-(p-coumaroyl)sophoroside (isomer) (Ferreres et al., 2008).
l a r v a . D : U V C h r o m a t o g r a m , 3 3 0 . 1 0 0 n m
2 0
4 0
6 0
I n t e n s .
0 5 1 0 1 5 2 0 2 5 3 0 3 5 T i m e [ m i n ]
89
1039 43
Figure 8.9: HPLC-DAD Phenolic Profile of Pieris brassicae Larvae Hydromethanolic Extract
Detection at 330 nm. Peaks: 8, 9 and 10 see Figure 8.7; 39 see Figure 8.8; (43) kaempferol-3-O-sophorotrioside (Ferreres et al., 2008).
169
O
OOH
R2O
OR1
OH
OH
1 R1 = sophorose; R2 = glucose4 R1 = R2 = glucose14 R1 = (methoxycaf feoyl)sophorose; R2 = glucose15, 24 R1 = (caff eoyl)sophorose; R2 = glucose18 R1 = (sinapoyl)sophorose; R2 = glucose19 R1 = (feruloyl)sophorose; R2 = glucose20 R1 = (p -coumaroyl)sophorose; R2 = glucose39 R1 = sophorose; R2 = H
OH
OHO
5 R1 = R2 = H8 R1 = OCH3; R2 = H9 R1 = R2 = OCH3 O
OOH
R2O
OR1
OH
OCH3
7 R1 = R2 = glucose13 R1 = glucose; R2 = H
O
OOH
R2O
OR1
OH
2 R1 = sophorotriose; R2 = glucose3 R1 = sophorose; R2 = glucose6 R1 = R2 = glucose10 R1 = sophorose; R2 = H11 R1 = H; R2 = glucose12 R1 = glucose; R2 = H16 R1 = (methoxycaffeoyl)sophorose; R2 = glucose17 R1 = (caffeoyl)sophorose; R2 = glucose21 R1 = (sinapoyl)sophorose; R2 = glucose22, 25 R1 = (feruloyl)sophorose; R2 = glucose23, 27 R1 = (p -coumaroyl)sophorose; R2 = glucose26 R1 = (methoxycaffeoyl)sophorose; R2 = H28 R1 = (caffeoyl)sophorose; R2 = H29 R1 = (sinapoyl)sophorose; R2 = H30 R1 = (feruloyl)sophorose; R2 = H31, 41, 42, 43 R1 = (p -coumaroyl)sophorose; R2 = H40 R1 = sophorotriose; R2 = H
R1 R2
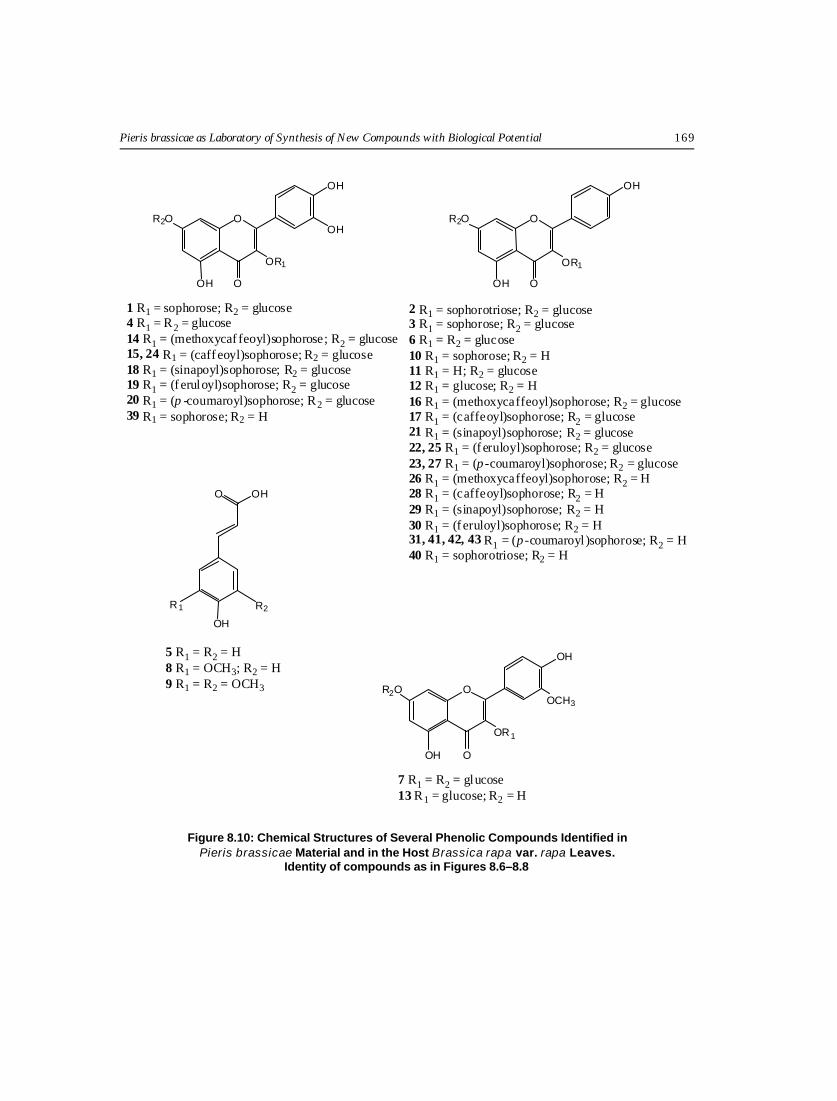
Figure 8.10: Chemical Structures of Several Phenolic Compounds Identified inPieris brassicae Material and in the Host Brassica rapa var. rapa Leaves.
Identity of compounds as in Figures 8.6–8.8
Pieris brassicae as Laboratory of Synthesis of New Compounds with Biological Potential

Utilisation and Management of Medicinal Plants170
Hydromethanolic extracts of exuviae and butterflies analysed by HPLC-MS revealed peaks intrace amounts, none of them corresponding to the studied phenolic compounds or possibly relatedwith them. Excrements are produced only at the larval stage, being the material containing higherphenolics content (Figure 8.8). These results are not surprising, considering that phenolic compoundsare sequestered and undergo metabolization, regarding their detoxification and excretion. If thecompounds are excreted, then they won’t be present in the subsequent stages, and this maybe thereason for not finding them in the exuviae and butterflies.
In the previous work (Ferreres et al., 2007a) using B. oleracea var. costata as host plant, it wasalready possible to see that the larva can sequester and metabolize this type of compounds. Thephenolics of these two Brassica species are different, so, the phenolic profile found for the larvae is alsodistinct form that observed before. This fact confirms the strong dependency on the phenolic pattern ofthe host plant. In addition, the study involving P. brassicae excrements, exuviae and butterflies (Ferrereset al., 2008), allowed to accomplish that, besides deacylation and deglycosylation already reported(Ferreres et al., 2007a), sulphating reactions also occur in the metabolic process of the larvae, and thatphenolics are mainly excreted and not transferred into the wings.
As insects are unable to synthesize phenolic compounds or their precursors, their presence in thedifferent stages of P. brassicae life cycle can only arise from the food it has ingested, that is, from thecomplex flavonoid derivatives and free phenolic acids of B. rapa var. rapa leaves. So, the detection ofthis kind of compounds in the larvae indicates that it has the ability to sequester them. Additionally,the fact that both larvae and excrements exhibit phenolic compounds distinct from those of the hostplant evidences that the larvae has the capacity to metabolize these phytochemicals, and to excretethem by the faeces, which included sulphate derivatives, reported for the first time. As these kind offlavonoids are known for their antioxidant potential (Ferreres et al., 2006 and 2007b; Vrchovská et al.,2006), in what concerns the obtainment of potential health promoting compounds, unusual in natureand of difficult laboratorial synthesis, P. brassicae (larvae, exuviae and butterfly) and its excrementsmay constitute a promising source.
ConclusionThe two examples presented herein showed that the P. brassicae larvae reared on B. oleraceae var.
costata and B. rapa var. rapa leaves their excrements, exuviae and butterflies presented new compounds,with complex chemical structures, impossible to be synthesized in the laboratory. These compoundsare related with those from B. oleracea var. costata (Ferreres et al., 2005) and B. rapa var. rapa (Ferreres etal., 2008). These two species revealed high antioxidant capacity for which its phenolic compounds areresponsible (Ferreres et al., 2005, 2006, 2007b). So, there‘s a strong possibility that extracts of P. brassicaelarvae reared B. oleraceae var. costata and B. rapa var. rapa leaves, their excrements, exuviae and butterfliesprovide powerful natural antioxidants. If that happens, those extracts may be used by thepharmaceutical industry in antioxidative formulations for prevention of free radicals-mediateddiseases, or even as preservative of other oxidizable formulations. The same can be applied to cosmeticindustry, for which it can be further used in anti-ageing formulations. Food industry may employ it toprevent the oxidation of its products, maintaining their quality and safety and extending their shelf-life, or to improve their nutritional value, by incorporating the extract in foodstuffs, thus increasing thedietary supply of antioxidants. In addition, it may constitute an economical advantage for B. oleraceavar costata and B. rapa var. rapa producers, who have great losses caused by P. brassicae infestations.

171
AcknowledgementsThe authors are grateful to Fundação para a Ciência e a Tecnologia (PTDC/AGR-AAM/64150/
2006) for financial support of this work.
REFERENCESAgerbirk, N., Müller, Olse, C.E.,Chew, F.S. (2006). A common pathway for metabolism of 4-
hydroxybenzylglucosinolate in Pieris and Anthocaris (Lepidoptera: Pieridae). Biochemical Systematicsand Ecology, 34: 189–198.
Aplin, R.T, D’Arcy W.R, and Rothschild, M. (1975). Examination of the large white butterflies andsmall white butterflies (Pieris spp.) for the presence of mustard oils and mustard oil glycosides.Journal of Entomology A, 50:73–78.
Brattsten L.B. (1992). Metabolic defences against plant allelochemicals. In: Herbivores and theirInteractions with Secondary Plant Metabolites, Vol. 2. Ed. By Rosenthal, G.A., and Berenbaum,M.R., Academic Press, USA, pp. 175-242.
Burghardt, F., Fiedler, K., and Proksch, P. (1997). Uptake of flavonoids from Vicia villosa (Fabaceae) bythe lycaenid butterfly, Polyommatus icarus (Lepidoptera: Lycaenidae). Biochemical Systematics andEcology, 25: 527-536.
Burghardt, F., Knüttel, H., Becker, M., and Fiedler, K. (2000). Flavonoid wing pigments increaseattractiveness of female common blue (Polyommatus icarus) butterflies to mate-searching males.Naturwissenschaften, 87: 304-307.
Burghardt, F., Proksch, P., and Fiedler, K. (2001). Flavonoid sequestration by the common blue butterflyPolyommatus icarus: quantitative intraspecific variation in relation to larval hostplant, sex andbody size. Biochemical Systematics and Ecology, 29: 875-889.
Fahey, J.W., Zalcmann, A.T., and Talalay, P. (2001). The chemical diversity and distribution ofglucosinolates and isothiocyanates among plants. Phytochemistry, 56: 5–51.
Fernandes, F., Valentão, P., Sousa, C., Pereira, J.A., Seabra, R.M., AND Andrade, P.B. (2007). Chemicaland antioxidative assessment of dietary turnip (Brassica rapa var. rapa L.). Food Chemistry , 105:1003-1010.
Ferreres, F., Sousa, C., Valentão, P., Pereira, J.A., Seabra, R.M., and Andrade, P.B. (2007a). Tronchudacabbage flavonoids uptake by Pieris brassicae. Phytochemistry , 68: 361-367.
Ferreres, F., Sousa, C., Valentão, P., Seabra, R.M., Pereira, J.A., and Andrade, P.B. (2007b). Tronchudacabbage (Brassica oleracea L. var. costata DC) seeds: Phytochemical characterization and antioxidantpotential. Food Chemistry , 101, 549-558.
Ferreres, F., Sousa, C., Vrchovská, V., Valentão, P., Pereira, J.A., Seabra, R.M., and Andrade, P.B. (2006).Chemical composition and antioxidant activity of tronchuda cabbage internal leaves. EuropeanFood Research and Technology, 222: 88-98.
Ferreres, F., Valentão, P., Llorach, R., Pinheiro, C., Cardoso, L., Pereira, J.A., Sousa, C., Seabra, R.M., andAndrade, P.B. (2005). Phenolic compounds in external leaves of tronchuda cabbage (Brassicaoleracea L. var. costata DC). Journal of Agricultural and Food Chemistry, 53: 2901-2907.
Pieris brassicae as Laboratory of Synthesis of New Compounds with Biological Potential

Utilisation and Management of Medicinal Plants172
Ferreres, F., Valentão, P., Pereira, J.A., Bento, A., Noites, A., Seabra, R.M., and Andrade, P.B. (2008).HPLC-DAD-MS/MS-ESI Screening of phenolic compounds in Pieris brassicae L. reared on Brassicarapa var rapa L. Journal of Agricultural and Food Chemistry, 56: 844-853.
Geuder, M., Wray, V., Fiedler, K., and Proksch, P. (1997). Sequestration and metabolism of host-plantflavonóides by the lycaenid butterfly Polyommatus bellargus. Journal of Chemical Ecology, 23: 1361-1372.
Harborne, J.B., and Grayer, R.J. (1994). Flavonoids and insects. In: The Flavonoids–Advances in Researchsince 1986, Ed. By Harborne, J.B., Chapman and Hall, London, pp. 589-618.
Knüttel, H., and Fiedler, K. (2001). Host-plant-derived variation in ultraviolet wing patterns influencesmate selection by male butterflies. The Journal of Experimental Biology, 204: 2447-2459.
Llorach, R., Gil-Izquierdo, A., Ferreres, F., and Tomás-Barberán, F.A. (2003). HPLC-DAD-MS/MS ESIcharacterization of unusual highly glycosylated acylated flavonoids from cauliflower (Brassicaoleracea L. var. botrytis) agroindustrial byproducts. Journal of Agricultural and Food Chemistry, 51:3895-3899.
Moyes, C.L., Collin, H.A., Britton, G., and Raybould, A. (2000). Glucosinolates and differential herbivoryin wild populations of Brassica oleracea. Journal of Chemical Ecology, 26: 2625-2641.
Müller, C., Agerbirk, Niels, and Olsen, C.E. (2003). Lack of sequestration of host plant glucosinolatesin Pieris rapae and P. brassicae. Chemoecology, 13: 47–54.
Muriel, S. B., and Grez, A.A. (2002). Effect of plant patch shape on the distribution and abundance ofthree lepidopteran species associated with Brassica oleracea. Agricultural and Forest Entomology, 4:179-185.
Renwick, J.A.A. (2002). The chemical world of crucivores: lures, treats and traps. EntomologiaExperimentalis et Applicata, 104: 35-42.
Rochfort, S.J., Imsic, M., Jones, R., Trenerry, V.C., and Tomkins, B. (2006). Characterization of flavonolconjugates in immature leaves of pak choi (Brassica rapa L. ssp. chinensis L. (Hanelt.)) by HPLC-DAD and LC-MS/MS. Journal of Agricultural and Food Chemistry, 54: 4855-4860.
Romani, A., Vignolini, P., Isolani, L., Ieri, F., and Heimler, D. (2006). HPLC-DAD/MS characterizationof flavonoids and hydroxycinnamic derivatives in turnip tops (Brassica rapa L. Subsp. sylvestrisL.). Journal of Agricultural and Food Chemistry , 54: 1342-1346.
Schittko, U., Burghardt, F., Fiedler, K., Wray, V., and Proksch, P. (1999). Sequestration and distributionof flavonoids in the common blue butterfly Polyommatus icarus reared on Trifolium repens.Phytochemistry, 51: 609-614.
Schoonhoven, L.M., Jermy, T., and van Loon, J.J.A. (1998). Insect-Plant Biology: From Physiology toEvolution. Chapman and Hall, London.
Sousa, C., Valentão, P., Rangel, J., Lopes, G., Pereira, J.A., Ferreres, F., Seabra, R.M., and Andrade, P.B.(2005). Influence of two fertilization regimens on the amounts of organic acids and phenoliccompounds of tronchuda cabbage (Brassica oleracea L. var. costata DC). Journal of Agricultural andFood Chemistry , 53: 9128-9132.

173
Vallejo, F., Tomás-Barberán, F.A., and Ferreres, F. (2004). Characterisation of flavonols in broccoli(Brassica oleracea L. var. italica) by liquid chromatography–UV diode-array detection–electrosprayionisation mass spectrometry. Journal of Chromatography A., 1054: 181-193.
van Loon, J.J.A., Wang, C.Z., Nielsen, J.K., Gols, R., and Qiu, Y.T. (2002). Flavonoids from cabbage arefeeding stimulants for diamondback moth larvae additional to glucosinolates: Chemoreceptionand behaviour. Entomologia Experimentalis et Applicata , 104: 27-34.
Vrchovská, V., Sousa, C., Valentão, P., Ferreres, F., Pereira, J.A., Seabra, R.M., and Andrade, P.B. (2006).Antioxidative properties of tronchuda cabbage (Brassica oleracea L. var. costata DC) external leavesagainst DPPH, superoxide radical, hydroxyl radical and hypochlorous acid. Food Chemistry , 98:416-425.
Pieris brassicae as Laboratory of Synthesis of New Compounds with Biological Potential





























