Embed Size (px)
Citation preview

A Broad Transition from Cloud Forest to PáramoCharacterizes an Undisturbed Treeline in ParqueNacional Llanganates, Ecuador
Authors: Hofstede , Robert G. M., Dickinson, Katharine J. M., Mark,Alan F., and Narváez, Edwin
Source: Arctic, Antarctic, and Alpine Research, 46(4) : 975-986
Published By: Institute of Arctic and Alpine Research (INSTAAR),University of Colorado
URL: https://doi.org/10.1657/1938-4246-46.4.975
BioOne Complete (complete.BioOne.org) is a full-text database of 200 subscribed and open-access titlesin the biological, ecological, and environmental sciences published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Complete website, and all posted and associated content indicates youracceptance of BioOne’s Terms of Use, available at www.bioone.org/terms-of-use.
Usage of BioOne Complete content is strictly limited to personal, educational, and non - commercial use.Commercial inquiries or rights and permissions requests should be directed to the individual publisher ascopyright holder.
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

© 2014 Regents of the University of Colorado ROBERT G. M. HOFSTEDE ET AL. / 9751523-0430/17 $7.00
Arctic, Antarctic, and Alpine Research, Vol. 46, No. 4, 2014, pp. 975–986
A broad transition from cloud forest to páramo characterizes an undisturbed treeline in Parque Nacional Llanganates, Ecuador
Robert G. M. Hofstede*§ Katharine J. M. Dickinson† Alan F. Mark† and Edwin Narváez‡#*Institute for Biodiversity and Ecosystem Dynamics (IBED), University of Amsterdam, Kruislaan 318, 1098 SM Amsterdam, The Netherlands†Department of Botany, University of Otago, PO Box 56, Dunedin, New Zealand‡Escuela de Biología y Química, Universidad Central del Ecuador, Ciudadela Universitaria, Av. América s/n, Quito, Ecuador§Corresponding author, present address: Ecopar, Casilla Postal 17-11-6706, Quito, Ecuador, [email protected]#Present address: Geoplades, Rumania E5-87 y Hungría, Quito, Ecuador
DOI: http://dx.doi.org/10.1657/1938-4246-46.4.975
Introduction
The highest parts of the wet perhumid northern tropical Andes consist of fragile landscapes characterized by a high biodiversity (Körner and Spehn, 2002; Price, 2006). These ecosystems include alpine páramo grasslands and humid montane cloud forests, which originally covered large tracts in Venezuela, Colombia, Ecuador, and Peru (Luteyn, 1999; Hofstede et al., 2003). Inhabitants of these re-gions reflect complex cultural and socio-economic circumstances. Most communities are made up of indigenous peoples and mestizo farmers that rely almost entirely on subsistence use of the natural resources, principally for agricultural production (Ange Jaramillo et al., 2002; Hofstede, 2011; Cuesta et al., 2012). Additionally, these landscapes provide important ecosystem services, particularly pota-ble water for drinking and irrigation (Buytaert et al., 2006; Buytaert and de Bièvre, 2012), as well as carbon storage (Farley et al., 2005; Cabaneiro et al., 2008; Chimner and Karberg, 2008).
Over recent decades, population pressure on the ecosystems of the tropical high Andes has rapidly increased and agricultural land use has greatly expanded, often using inappropriate tech-niques and leading to serious land degradation (Podwojewski et al., 2002; López Sandoval, 2004). Natural montane cloud forests have been widely replaced by either potato cultivation, pastures for dairy cattle, or exotic conifer forest plantations, with native páramo grasslands being over-exploited by grazing combined with frequent burning (Hofstede, 1995; Verweij, 1995; Hofstede et al., 2002; Farley et al., 2004; González et al., 2011). In many situa-tions, these developments have led to a depression of the upper
forest limit (Laegaard, 1992; Sarmiento, 2002; Wille et al., 2002). If the uppermost forest is destroyed by human activities, páramo grassland from above the natural forest limit tends to colonize the cleared areas and thus over time can extend to lower altitudes than would otherwise be the case. By contrast, the upward movement of forest, attributed to the effects of contemporary global warming, has also been reported (Innes, 1991; Grabherr et al., 2010; Kessler et al., 2011; Jantz and Behling, 2012). These two factors, when combined, have potentially opposing effects, leading to complica-tions when formulating predictive scenarios (Castaño, 2002; Grace et al., 2002; Tonneijck et al., 2006; Bader et al., 2007).
What limits tree growth at alpine treelines has been explained in various ways, mostly with low and variable daily temperatures as principal factors, with precipitation and wind stress as additional considerations (Acosta Solis, 1984; Körner, 1998, 2012), locally or regionally. However, the exact position and structure of the treeline may be determined by low average and extreme temperatures as well as other microclimatic factors, such as strong irradiance (Wardle, 1971; Miehe and Miehe, 1994; Germino and Smith, 1999; Körner and Paulsen, 2004; Körner, 2012), along with the ecology of the par-ticular species involved (Smith, 1994; Bader, 2007; Llambí et al., 2013). The páramo itself is a tall tussock grass–dominated treeless tropical-alpine vegetation belt that extends from approximately 3400 m to over 4500 m altitude. Often a shrub-dominated (subpáramo) transition from upland forest to páramo is recognized, spanning an altitudinal range of ~300 m (Salamanca, 1992), but from ~3600 m to 4100 m, the vegetation is dominated by tussock grasslands (grass páramo). Above these altitudes from ~4100 m to 4800 m, a more
AbstractThis study aimed at investigating the vegetation pattern associated with the transition from upper northern Andean cloud forest to páramo grassland in a relatively unmodified area in Ecuador’s Eastern Cordillera. Single 20 × 10 m plots were randomly placed in representative sites along an altitudinal gradient from continuously wet Andean montane cloud forest at 3400–3700 m to páramo grassland at 3900–4000 m. Also sampled was a disjunct stand of the stem-rosette Espeletia pycnophylla var. llanganatensis, some 200 km south of the limit of the otherwise continuous distribution of this genus. Several physi-cal and chemical soil factors, including temperature at –30 cm, were determined. Visual estimates of cover in four life-form classes (trees, shrubs, herbs, ground cover) at each site revealed similarities related to their relative proximity along an elevation gradient. Stands with a tree cover were more species-rich than those without, due largely to the differential distribution of the woody flora. No abrupt discontinuity occurred in the mix of species along this gradient. Soil temperature and altitude were the most important factors of those measured to explain the variability in plant community structure and composition. There was no clear zonation evident along the altitudinal gradient, with vegetation structure and floristics displaying continuous variation. No clear upper forest limit was differentiated. The isolated presence of Espeletia in the area is discussed, but the theory about its origin remains speculative.
Note: with this study, Emeritus Professor Alan Mark, ΦΒΚ, FRSNZ, KNZM, has the milestone of 200 published research papers, mostly dedicated to alpine research.
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

976 / ARCTIC, ANTARCTIC, AND ALPINE RESEARCH
open, shorter cover occurs, the so-called superpáramo vegetation (Cuatrecasas, 1958; Cleef, 1981; Sklenár and Balslev, 2004).
There is a distinction between the limit of closed forest and that of tree growth; the two are not necessarily the same (Körner and Paulsen, 2004; Körner, 2012). Thus, although the páramo belt is essentially a treeless vegetation, patches of high-Andean dwarf for-est and shrubland, up to 10 m tall, may occupy more sheltered sites between 3400 and 4300 m elevation (Ulloa Ulloa and Jørgensen, 1993; Young and Keating, 2001; Ramírez et al., 2009; Contreras, 2010; Llambí et al., 2013). The significance of these patches of dwarf forest above the present upper forest limit is debated (Wille et al., 2002). According to some authors (Cuatrecasas, 1958; Ellenberg, 1958, 1979; Laegaard, 1992; Sarmiento, 2002), these fragments are remnants of previous extensive forests destroyed by people. Further, by this reasoning, the uppermost limit of closed forest has been esti-mated to be as high as 4100 m in the Ecuadorian Eastern Cordillera, and 4350 m in the Western Cordillera (Laegaard, 1992), and is rela-tively abrupt. However, other studies have proposed natural causes for the occurrence of at least some forest fragments and individual trees that occur above the present-day continuous forest limit (Wille et al., 2002; Bader, 2007; Ramírez et al., 2009), thus by implication inferring a more gradual transition from closed forest to grassland. The occurrence of these forest patches has been variously explained: in relation to such factors as locally favorable microenvironmental conditions (Walter and Medina, 1969; Smith, 1978; Lauer, 1981; Bader et al., 2007), and/or specific physiological adaptations or com-plex species-specific local spatial interactions that allow certain tree species at relatively high altitude to establish and compete in the tall grass matrix (Vélez et al., 1998; Llambí et al., 2013).
The present vegetation pattern in the tropical high Andes is gen-erally determined by human impact, particularly through the use of fire, leading to the destruction of woody vegetation usually on all but the most sheltered sites (Hofstede, 1995; Verweij, 1995; Hofstede et al., 2002; Young, 2009; Cuesta et al., 2012). Thus, few examples of what could be construed as a natural upper forest boundary now exist. Further, with the differing interpretations of past forest extent and altitudinal vegetation patterns, the degree to which human activ-ity is the overriding cause of an apparently low upper forest limit and the present situation of widespread páramo, remains unclear (Bader et al., 2007; Young and León, 2007). The nature of the upper forest limit and the associated distribution of woody vegetation where there has been negligible human activity is thus open to question, and there is a clear need to identify reference sites (Bader et al., 2007; Ramírez et al., 2009). Such sites by implication, inevitably would be in remote locations with difficult access, thus presenting consid-erable logistical and practical problems for research. These factors have no doubt contributed to the current lack of knowledge of natu-ral altitudinal vegetation patterns in the tropical high Andes (Miehe and Miehe, 2000). Notwithstanding these constraints, we aimed to identify a suitable location where, in the apparent absence of human activity, a natural altitudinal vegetation pattern covering a forest to páramo transition could be described and quantified both structur-ally and floristically. As such, we aimed to provide much-needed reference data enabling us to contribute to the debate on upper forest limits in the tropical high Andes.
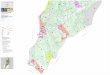
Our criteria were met in the heart of Parque Nacional Llanga-nates, in the Ecuadorian central Eastern Cordillera, which includes one of the least disturbed areas of forest and páramo remaining in the northern Andes. Covering 2197 km2, the park has an altitudinal range from 1200 m on the Andean footslopes to the east (Amazon side), up to 4638 m at its highest point (Cerro Hermoso), near the western boundary (Fig. 1). Road access to, and agricultural use of, the páramo occurs only in the northern portion of the park, more
than 30 km from the study site. The rest is without human use; indeed, few people enter and there is no discernible evidence of fire or domestic animals. Although there are indications of some pre-Hispanic use of certain peripheral parts (Anhalzer, 1998), it is highly unlikely that human impact, including fire, has determined any significant aspect of the present landscape. Importantly, Parque Nacional Llanganates is also home to a single stand of the emblem-atic páramo endemic stem-rosette of the genus Espeletia, located in a remote part of the park. This population is some 200 km south of the limit of continuous distribution of this taxon from Venezuela to northern Ecuador (Monasterio and Sarmiento, 1991; Cuatreca-sas, 2013). This biogeographical anomaly is still awaiting a satis-factory explanation. Being situated just below the forest-páramo ecotone, it is a prominent feature in our study area, and we will also discuss potential mechanisms explaining its presence here.
Study AreaParque Nacional Llanganates occupies the Llanganates Cor-
dillera, which is dominated by Paleozoic metamorphic outcrops consisting of gneiss and quartzite (Llanganates Group). As such, the geology of the park contrasts with that of the surrounding ar-eas. Though this formation constitutes the basis of the major part of the Ecuadorian Eastern Cordillera, elsewhere these rocks have been covered by volcanic formations of Tertiary and Quaternary age (Jørgensen and Ulloa Ulloa, 1994; Vázquez et al., 2000). Also, the higher parts of the Llanganates Cordillera have been heavily eroded by glacial activity during the Pleistocene, resulting in a roll-ing landscape with U-shaped valleys, many glacial lakes and mires, along with rocky outcrops in cirques and lateral moraines. Further-more, it is likely that the metamorphic rocks are less permeable than those of volcanic origin, leading to more ponds and lagoons on the valley floors in the Llanganates Cordillera when compared with other parts of the Eastern Cordillera. Soils in the region are influenced by volcanic ash deposits from neighboring active vol-canoes, such as Cotopaxi and Tungurahua, that have covered the entire Llanganates Cordillera (Mothes, 1991; Vázquez et al., 2000).
The park encompasses montane tropical rainforest and mon-tane cloud forest (1200 to ~3500 m a.s.l.), as well as extensive páramo vegetation above ~3500 m (approximately 25% of the park). There are no climatic data available for the region, but it is generally assumed that the Llanganates Cordillera is a continu-ously wet perhumid area, under the influence of humid air from the Amazon basin. Data from other páramo areas on the Amazonian side of the Andes indicate that annual precipitation generally ex-ceeds 3000 mm (Rangel, 2000).
MethodsWe studied an elevation gradient from 3400 to 4000 m in a re-
mote area, approximately 27 km from the park boundary (1°10′00″S; 78°15′00″W). The gradient was within the range of what could be as-sumed to include the natural forest limit, extending from montane cloud forest, including the Espeletia stand within this forest, to the highest ex-tent of the grass páramo. The latter limit was evidenced by the loss of continuous vegetation cover and the abundance of typical superpáramo species such as Loricaria ilinissae and Werneria pygmaea (Luteyn, 1999; Sklenár and Balslev, 2004). The direct distance between the low-ermost and the uppermost point along the gradient was approximately 5 km (Fig. 1), from the U-shaped valley (Valle de los Frailejones), over the lateral moraines that contained Laguna Soguillos, to an exposed rocky outcrop (Cerro Pan de Azucar) at just over 4000 m altitude.
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

ROBERT G. M. HOFSTEDE ET AL. / 977
FIG
UR
E 1
. T
he lo
cati
on o
f th
e st
udy
area
wit
hin
Par
que
Nac
iona
l Lla
ngan
ates
, and
its
loca
tion
wit
hin
Ecu
ador
.
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

978 / ARCTIC, ANTARCTIC, AND ALPINE RESEARCH
Field work was conducted in May 2000. We used a method that has been found suitable to characterize a wide range of vegetation types, including mixed rainforest and tall tussock grasslands (Dick-inson et al., 1998). Six 20 × 10 m plots were randomly located at ap-proximately 100 m intervals of altitude, where terrain permitted (time and logistical limitations unfortunately prevented plot replication). An additional plot was randomly placed in the disjunct Espeletia stand, situated on a colluvial slope at 3430 m, within the upper montane for-est, the latter being recorded in the nearby (~300 m distant) plot at 3400 m (Fig. 2). All plots were aligned with the long axis across the contour, on similar slopes (20–35°), with a generally southerly aspect. Percent cover of all vascular plant species was visually estimated in four height-life-form categories: tree (single woody stem >2 m); shrub (multi-stemmed); herb (non-woody); and ground layer (<0.5 m), apart from the stem-rosettes of Espeletia pycnophylla var. llanganatensis and Blechnum occidentale, which were recorded separately because of their distinctive and contrasting life-forms. Where species required further identification, collections were made. For the ground layer, the general cover of Sphagnum sp., and “other bryophytes,” “lichens,” lit-ter, bare ground, and rotting stems were noted. Nomenclature follows Luteyn (1999) and Vázquez et al. (2000).
Species abundance data were analyzed using the multivari-ate statistical package PRIMER 5.0 (Clarke and Warwick, 2001) for community structure and composition. Cluster analyses us-ing the group average algorithm were performed on Bray-Curtis similarity matrices derived from presence-absence and square-root transformed percent cover data to test for similarity between plant composition in the seven plots.
Soils were collected to 10 cm depth from each of the seven sites for later analyses of bulk density (dry weight per standard volume of 100 cm3), moisture content, and macronutrient composition. Two samples,
each composed of five subsamples were collected systematically from each plot. On return to Quito, samples were refrigerated and later ana-lyzed for bulk density and moisture content by means of weight differ-ence between the field and oven-dried samples: 24 hr, 105 °C. Soil sam-ples were visually examined for charcoal remains, indicating fire history (Di Pasquale et al., 2008). Chemical analyses were done at the Labo-ratoire du Sols, Institute de Réchèrche pour le Developpement (IRD), France. These samples were dried for 48 hr at 30 °C and passed through a 2 mm sieve. Organic carbon and nitrogen were obtained using a CHN Carlo-Erba auto-analyser. Al, Fe, and Si were extracted with 0.2 M am-monium oxalate at pH 3 (Blakemore et al., 1981), and 0.1 M sodium pyrophosphate (Wada and Higashi, 1976), and the extracts measured by inductively coupled plasma atomic emission spectroscopy (ICP-AES). In each plot, soil temperature was also recorded at –30 cm at the time of sampling; values at this depth are likely to be stable through the day and reflect the mean annual value in these relatively thermo-stable environ-ments (Thouret and Faivre, 1989).
To indicate microclimatic conditions at the time of the study, two data loggers were installed at 3845 m altitude on a southerly aspect from 11:30 on 8 May until 15:30 on 13 May 2000. Air tem-perature and humidity were recorded using a shielded Hobo H8 Pro-model, 1.5 m above ground level on the canopy fringe of a small Gynoxys acostae tree, and soil temperature at –10 cm, using an Optic Stowaway data logger.
ResultsThe seven plots studied along the 600 m altitudinal gradient in-
cluded a total of 105 vascular plant species and one identified bryo-phyte, Sphagnum magellanicum. Of the vascular plant species, there were 63 herbs, 37 erect woody species, 3 creeping shrubs, and 2 stem-
FIGURE 2. The Espeletia pychnophylla var. llanganatensis stand within the upper Andean forest (P1) at 3430 m a.s.l. in Parque Nacional Llanganates, Ecuador.
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

ROBERT G. M. HOFSTEDE ET AL. / 979
Species names P1 P2 P3 P4 P5 P6 P7
Valeriana microphylla
<1 <1 <1 <1 <1 2
Geranium sibbaldioides
<1 <1 <1 <1 <1 1 <1
Neurolepis asymmetrica
30
Equisetum bogotense
4
Asteraceae sp. 1 (RH 1703)
2
Peperomia saligna 1
Rorippa bonariensis
1
Urticaceae sp. 1 (RGMH 1731)
1
Calceolaria sp.1 (RH1717)
<1
Indet. 2 (RGMH 1718)
<1
Monnina obtusifolia
<1
Pilea vegesana <1
Solanaceae sp. 1 (RGMH 1743)
<1
Cerastostema alatum
2 <1
Poaceae sp. 2 (RGMH 1723)
<1 <1
Ribes lehmannii <1 <1
Desfontainia spinosa
6 1 <1
Miconia corimbiformis
<1 2 1
Greigia mulfordii <1 1 <1
Oxalis phaeotricha <1 <1 <1
Ranunculus peruvianus
<1 <1 <1
Rubus adenotrichos <1 <1 <1
Nertera granadensis
1 <1 <1 <1
Gynoxys acostae <1 3 4 4
Calamagrostis brevifolia
5
Luzula gigantea 4
Diplostephium rupestre
3
Myrsine dependens 2
Indet. 1 (RGMH 1742)
1
Baccharis latifolia <1
TABLE 1
Plant species cover in the seven quadrats sorted by occurrence along the altitudinal gradient. Shaded cells are woody species, and nonshaded cells are herbaceous species. Nonvascular categories are shown in bold. Accession numbers are shown for unidentified
specimens deposited in the author’s private herbarium.
Species names P1 P2 P3 P4 P5 P6 P7
Espeletia pychnophylla ssp. llanganatensis
7
Ericaceae sp.1 (RGMH 1712)
<1
Huperzia crassa <1
Poaceae sp. 3 (RGMH 1722)
<1
Weinmannia pinnata
<1
Baccharis genistelloides
3 <1
Gaiadendron punctatum
<1 3
Blechnum occidentale
<1 <1
Rubus coreaceus <1 <1
Hesperomeles heterophylla
<1 3
Brachyotum ledifolium
<1 <1
Hymenophyllum sp.1 (RGMH 1733)
<1 1
Sphagnum magellanicum
3 2 <1
Elaphoglossum sp. 1 (RGMH 1724)
<1 <1 <1
Elaphoglossum sp. 2 (RGMH 1738)
<1 <1 <1
Ranunculus praemorsus
<1 <1 <1
Rhynchospora macrochaeta
8 4 5 4
Escallonia myrtilloides
<1 30 25 12
Pteridophyta sp. 1 (RGMH 1686)
<1 <1 <1 <1
Disterigma empetrifolium
20 4 3 1 <1
Galium hypocarpium
<1 2 <1 <1 <1
Blechnum loxense <1 <1 <1 <1 <1
Pteridophyta sp. 3 (RGMH 1730)
<1 <1 <1 <1 <1
Vaccinium floribundum
<1 <1 1 <1 <1
Lichens <1 <1 <1 <1 1
Neurolepis aristata 40 12 7 30 25 5
TABLE 1
Continued
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

980 / ARCTIC, ANTARCTIC, AND ALPINE RESEARCH
Species names P1 P2 P3 P4 P5 P6 P7
Oreomyrrhis andicola
<1 <1 <1
Baccharis brachylaecnoides
<1
Baccharis parviflora
<1
Rosaceae sp. 2 (RGMH 1735)
<1
Valeriana plantaginea
5 2
Loricaria ilinissae 2 3
Gunnera magellanica
1 <1
Lasiocephalus ovatus
<1 <1
Geraniaceae sp (RGMH 1704)
<1 <1
Lachemilla nivalis <1 <1
Oritrophium peruvianum
<1 1
Ranunculus sp. 1 (RGMH 1748)
<1 <1
Azorella pedunculata
<1
Berberis sp. (RGMH 1682)
3
Carex bonplandii <1
Gnaphalium linearifolium
<1
Hypochaeris sessiliflora
4
Oreobolus goeppingerii
1
Satureja nubigena <1
Werneria pygmaea 5
Species names P1 P2 P3 P4 P5 P6 P7
Gaultheria sclerophylla
<1
Luzula vulcanica 2 <1
Luzula racemosa 1 1
Pernettya prostrata <1 <1
Cortaderia nitida <1 30 2
Indet. sp. 3 (RGMH 1695)
<1 <1 <1
Macleania rupestris <1 <1 <1
Gynoxys sodiroi <1 4 6 3
Lachemilla pectinata
<1 <1 <1 <1
Agrostis tolucensis <1 <1 4 4 <1
Bomarea glaucescens
<1
Gaultheria amoena <1
Hypericum laricifolium
<1
Lupinus prostratus <1
Monnina sp. 2 (RGMH 1693)
<1
Poaceae sp. (RGMH 1744)
<1
Ranunculus geranoides
<1
Diplostephium glandulosum
3 2
Gynoxys buxifolia 1 <1
Bartsia laticrenata <1 <1
Pteridophyta sp. 2 (RGMH 1690)
<1 <1
Baccharis caespitosa
<1 4
Calamagrostis intermedia
<1 10 10
Miconia theaezans <1
Pinguicula calyptrata
<1
Sisyrinchium palustre
<1
Stellaria leptopetala
<1
Grammitis cf. sodoroi
<1 <1
Puya hamata <1 <1
Calamagrostis macrophylla
<1 <1
Diplostephium aff. ericoides
1 <1 <1
TABLE 1
Continued
TABLE 1
Continued
rosettes (Espeletia pycnophylla var. llanganatensis and Blechnum oc-cidentale) (Table 1). The plots with a tree cover (P2, P3, and P4) had more plant species (37–41) than those without a tree layer (P1, P5, P6, and P7; 26–30: Fig. 3). This difference in species number was mostly associated with the richer woody flora in the plots with a tree layer (14–15 woody species) compared to those without (5–9 woody species). There was no clear difference in herbaceous species richness between the plots with (20–25) and without (16–23) a tree layer. The plot with the highest overall plant richness (P4; 41species) had the greatest number of both woody and herbaceous species.
There was only one plant species that occurred in all seven quadrats (Geranium sibbaldioides), and two that occurred in six plots (Neurolepis aristata and Valeriana microphylla; Table 1). On the other hand, 45 vascular species occurred in only one plot. In all plots, there was a dominance of tall, wet-condition–indicating graminoids, such as the bamboo genus Neurolepis (present in all
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

ROBERT G. M. HOFSTEDE ET AL. / 981
FIGURE 3. Number of vascular plant species according to general life form category, for seven plots along an altitudinal transect between 3400 and 4000 m a.s.l. in an undisturbed treeline ecotone in Parque Nacional Llanganates.
FIGURE 4. Cover of tree layer and shrub layer (%) for the seven plots along an altitudinal transect between 3400 and 4000 m a.s.l. in an undisturbed treeline ecotone in Parque Nacional Llanganates.
plots as N. aristata or N. asymmetrica), Cortaderia nitida (P4, P7), Rhynchospora macrochaeta (P1, P2, P4, P5), and Luzula gigantea (P3). The characteristic páramo tussock grass, Calamagrostis in-termedia, was abundant only in the two highest altitude plots (P6 and P7) above 3850 m. Dicotylous herbs attaining more than 1% cover (Valeriana plantaginae, Werneria pygmaea, and Hypocha-eris sessiliflora) were also found only in the two highest plots.
The tree layer of the lower plots (P2, P3, and P4; Fig. 4) was composed principally of Escallonia myrtilloides and Gynoxys spe-cies. Less abundant were Myrsine dependens and Desfontainia spi-nosa, the latter a shrub reaching the tree layer. The shrub layer was more diverse, with Diplostephium sp., Baccharis sp., Disterigma em-petrifolium, and Gaiadendron punctatum attaining considerable cov-er, especially in the lower plots. In the higher altitude plots, woody species either had a creeping growth form (Pernettya prostrata) or are known to tolerate (Baccharis spp., Valeriana microphylla), or be indicative of (Loricaria ilinissae), conditions associated with higher altitudes such as freezing and high irradiation (Sklenár and Balslev, 2004; Kessler et al., 2011). The change in vegetation structure along the altitudinal gradient is most clearly indicated by the decrease of the summed cover of the tree and shrub layers (Fig. 4).
The greatest floristic similarity (58%) was shown between the two highest altitude plots without a tree layer (P6 and P7). These were also the least similar to the rest (23%). Among the latter, the greatest floristic similarities were shown between P3 and P4 (at 56%) and P1 (Espeletia dominated) and P2 (closed Escallonia forest; 48%); P5 was more similar (44%) to P3 and P4 than to the other plots.
Cluster analysis of the square-root transformed percent cover data confirmed the greatest similarity to be between P6 and P7 (48%), and their differentiation from the rest of the plots (only 19% similarity). Of these, P2 (closed Escallonia forest) was least similar to the others (26%) (Fig. 5).
At an intermediate altitude along the gradient (3845 m), air temperatures over the six-day period of recording in May 2000 ranged from 0.3 to 11.5 °C, soil temperatures from 5.8 to 9.1 °C (Fig. 6), and relative humidity from 47% to 100%, but was >90% for 63% of the time. Typically, the warmest time of the day was mid-afternoon, while the coldest was variable over the period of darkness. Soil temperatures at this depth (–10 cm) were relatively stable over this period (Fig. 6). Spot soil temperatures at the more stable depth of –30 cm decreased significantly with altitude from 8.5 to 6.2 °C (r = –0.74, p < 0.01). The site at the highest elevation (P7) with a relatively open grassland was relatively warm, but this statistical relationship between temperature and altitude is driven by the temperature recorded at the Espeletia site (P7: 8.5 °C).
The soil organic C content correlated negatively with bulk density (–0.97, p < 0.01) and positively with its moisture content (0.87, p < 0.01). This intercorrelation is a common feature in mineral soils with high or-ganic content, since organic matter is of low density and has a high water retention capacity. There was also a highly significant positive correla-tion (0.97, p < 0.01) between the silica content and the aluminum/iron factor (Al + 0.5Fe). There was a weaker negative correlation between altitude and %C content (and hence with soil bulk density and moisture content; r = –0.73, p < 0.01). This indicates a trend of less organic mate-rial, higher bulk density, and lower soil moisture content with increasing altitude. Soil moisture varied, but all plots were more than 60% saturated. The Al/Fe factor varied from 0.9 (P2) to 2.6 (P4), but no particular trend was obvious (Table 2). Silica was also at a maximum in P4 (6.0%) and minimum in P2 (0.6%). No charcoal was found in any sample.
DiscussionThis study provides a rare contemporary ecological data set
for an area in the tropical high Andes that has experienced negli-
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

982 / ARCTIC, ANTARCTIC, AND ALPINE RESEARCH
gible human influence. Few other such studies exist (Yánez, 1998; Ramírez et al., 2009). Based on our study and wider observations of the surrounding slopes, it appears that no distinct upper for-est limit occurs in this area. There was some decline in tree spe-cies richness and cover between the obvious forest plots (P1–P3: 3430–3630 m) and shrub-dominated plot 4 (subpáramo: 3700 m), with a more obvious floristic dissimilarity between this plot and plots 5–7 (open páramo: >3800 m). Thus, there was a closed forest up to ~3600 m elevation, which graded through a narrow grass-shrub-tree dominated belt (subpáramo) of variable width but ~200 m elevation, into grass páramo above ~3800 m. We did not find any abrupt changes in species richness or a significant increase or de-crease in species turnover at a particular altitude. Rather, turnover appeared continuous, plots displaying an essentially pair-wise pat-tern: the closest plots geographically, were the most similar, floris-tically and structurally. The lack of plot replication, however, limits interpretation. The inference of continuous turnover contrasts with findings in other transect studies in the forest-páramo ecotone by Wille et al. (2002) and Moscol (2010), who found strong species turnover around what was assumed as the original upper forest limit. The difference is most likely explained by the greater human intervention in the páramo landscape as compared to the (relatively
intact) Andean forests in their study areas, while our entire study area was without apparent intervention.
These findings lend support to the hypothesis that, in the ab-sence of human activity, tree patches and tree individuals may occur naturally above 3500 m without having to infer a previous history of clearance, as also described by Ramírez et al. (2009). Indeed, we found no evidence of a fire history in our study area, supported by the absence of charcoal in our soil samples. The highest altitude of the closed forest (~3600 m) in our study area is similar to that reported by Wille et al. (2002) and Jantz and Behling (2012) for other páramo sites in Ecuador, based on paleoecological studies. The observed wide gradient from forest to páramo differs from the observations of Bader et al. (2007), who described rather abrupt treelines in a comparative study on various treelines in the Andes and in Hawaii. They studied upper forest limits in mountain areas that had light to moderate anthropogenic influence, and related the abrupt upper forest limit to fire occurrence since the Holocene and positive feedback mechanisms limiting tree establishment in open páramo vegetation.
Altitude reflects a composite of environmental factors, par-ticularly in relation to temperature and usually also precipitation. The reductions in soil organic material and moisture content, and
FIGURE 5. Cluster analysis of square-root transformed abundance data using the Bray-Curtis similarity coefficient, from the seven plots along an altitudinal transect between 3400 and 4000 m a.s.l. in an undisturbed treeline ecotone in Parque Nacional Llanganates.
FIGURE 6. Air (1.5 m) and soil (–10 cm) temperatures (°C) recorded from 11:30 on 8 May 2000 to 15:30 on 13 May 2000 at 3845 m elevation on a southerly aspect in Parque Nacional Llanganates.
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

ROBERT G. M. HOFSTEDE ET AL. / 983
the increase in bulk density with increasing altitude, for example, could be explained by lower primary productivity and lower lit-ter input as temperature decreases with altitude. Of the other soil factors measured, both the silica content and the Al/Fe factor are good indicators of recent volcanic ash in the soil (Zehetner et al., 2003). There was variation along the gradient in the Al/Fe factor: at the lower altitudes (especially P2 and P3) this value was approxi-mately 1, whereas in the higher altitude plots values were about 2, suggesting a greater influence of volcanic ash in the soil formation of the latter (Table 2). Based on the Al/Fe factor, only P4 (2.6) can technically be considered as an Andisol (FAO/ISRIC/ISSS, 1998). Thus, it could be inferred that in the lower altitude plots there is a higher proportion of weathered metamorphic rock and less pyro-clastic material in the soil formation. Such an interpretation could imply a lesser influence of volcanic activity on the lower altitude systems. Our study area was also extremely wet and perhumid. The interaction between climate and soil parent material can lead to local topographic variation, which in turn influences plant commu-nity structure and composition. In our case, the most widespread taxa were of bamboo, which clearly favored the wet conditions.
Our results can be interpreted in the general context of the wet páramo, but to what extent such patterns would be evident in drier conditions remains debatable (Stevens and Fox, 1991). Obviously, climatic conditions and plant ecophysiology are interrelated, but to what extent tree dynamics toward their upper limit are affected by wet versus dry conditions and the interactive effect with human activity and fire is open to debate (Bader, 2007; Sarmiento, 2002). Climatic variability over time also should be taken into account given environmental change and particularly the moisture fluctua-tions that are evident through the Holocene (Baker et al., 2001; Marchant and Hooghiemstra, 2004).
Climatic variability, combined with the vagaries of volcanic activity, lends itself to producing some anomalous species distri-butions. Such may be the underlying reason behind the presence of the stand of Espeletia pychnophylla var. llanganatensis and other páramo-like vegetation on the colluvial slope within the forest in our study area. Floristically, the Espeletia stand (P1) was most similar to the nearby closed forest (P2), but only at 48%. Nevertheless, their floristic composition was more similar than to any of the other five plots. When abundance is taken into account,
TABLE 2
Environmental data for each of the seven plots recorded in May 2000 in Parque Nacional Llanganates.
Plot number P1 P2 P3 P4 P5 P6 P7
Vegetation type
Espeletia pycnophylla stand (to 5 m tall) in forest gap. Bamboo herb layer (Neurolepis aristata).
Escallonia myrtillioides dominant Andean forest to 8 m tall. Under-growth: Bamboo (Neurolepis asymmetrica)
Escallonia myrtillioides, Myrsine dependens Andean forest to 8 m tall. Under-growth: Bamboo (N. aristata) and other graminoids
E. myrtillioides, Gynoxys open forest 8 m tall. Shrubby bamboo grassland (N. aristata)
Bamboo grassland (N. aristata) with shrubs and scattered small trees (Gynoxys, 4 m tall)
Bamboo (N. aristata) tussock (Calamagrostis intermedia) grassland with scattered shrubs (3 m tall)
Open bamboo (N. aristata) tussock (C. intermedia) grassland with small shrubs (1.2 m tall)
Altitude
(m a.s.l)
3430 3400 3630 3700 3810 3900 4000
Soil temp.
(–30 cm; °C)
8.5 7.4 6.8 6.8 6.6 6.2 6.9
Soil C (%) 11.6 9.2 11.4 9.0 8.1 7.7 7.8
Soil N (%) 1.2 0.4 0.8 0.4 0.5 0.8 0.6
Soil S (%) 0.08 0.06 0.05 0.07 0.07 0.04 0.06
Soil Fe
(mg kg–1)
8.5 10.6 5.0 8.0 7.6 7.8 6.8
Soil Al
(mg kg–1)
11.6 3.8 7.8 21.9 15.6 14.1 13.4
Soil Si
(%)
1.6 0.6 1.0 6.0 3.5 3.0 2.7
Soil Al + (0.5 Fe/10)
1.6 0.9 1.0 2.6 1.9 1.8 1.7
Soil bulk density (10 cm; kg m–3)
598 700 624 722 712 740 732
Soil moisture
(% ODWt)
76.5 63.3 72.4 67.3 59.9 65.3 64.4
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

984 / ARCTIC, ANTARCTIC, AND ALPINE RESEARCH
however, their similarity lessens and greater affinities are shown by the Espeletia stand with the mid-altitude plots (P3, P4, P5; Fig. 5) in relation to their woody/non-woody mix of vegetation. The soil temperature in the Espeletia stand indicated a relatively warm site, but the ecological significance can only be speculated from our results.
Site favorability may explain persistence of the stand at the site but not the presence of this outlier Espeletia population, 200 km south of the southernmost limit of the species’ present-day continuous natural distribution between Venezuela and northern Ecuador (Monasterio and Sarmiento, 1991). We hypothesize that the past continuous distribution of this species probably extended to central Ecuador, but perhaps through volcanic activity in the Quaternary, the species was rendered extinct between northern and central Ecuador. This single Espeletia population in our study may have survived through a combination of the generally lower volcanic activity in the Llanganates area (the only non-volcanic area between northern and central Ecuador), and local site factors. Potential establishment of the Espeletia pycnophylla stand, following long-distance dispersal from the main popula-tion of the species ~200 km to the north has little support, given the absence of a pappus or any other dispersal adaptation in the fruits of this genus.
ConclusionsOur study is one of the first to provide quantitative ecologi-
cal data from a remote, continuously moist region of the tropical high Andes, apparently without the influence of human activity. The area is difficult of access and presents considerable logistical challenges, no doubt contributing to the relative lack of ecologi-cal information on such ecosystems. Our results provide baseline reference data with which to compare the impact of human activity elsewhere in the páramo region. At the least, our data give little support to there being a clear and discernible treeline under such climatic conditions. In this relatively natural setting we observed that there is a broad ecotone from montane Andean cloud forest at 3400–3600 m altitude up to open grass alpine páramo at 3800 m and above, and that within this subpáramo ecotone, there is a mosaic of scattered trees, shrubland, and grassland. This pattern is consistent with the views of Körner and Paulsen (2004), Bader et al. (2007), and Ramírez et al. (2009). Moreover, the most remote parts of Parque Nacional Llanganates present one of the last re-maining reference sites in Ecuador on which to gauge the impacts of human activity. Thus, from a range of perspectives, the Parque Nacional’s ecosystems are highly significant and we encourage further research to test the hypotheses that we have raised in this paper.
AcknowledgmentsThe authors would like to thank Galo Medina for his great
help during the field work, and Maaike Bader and Luis Daniel Llambí for reviewing our manuscript. Pascal Podwojewski ar-ranged the soils analysis in the Institut de Recherche pour le Développement (IRD) laboratory in France, while Pascale Michel and Jacqueline Nielsen provided helpful discussion and comments on the manuscript. The field work for this study was done within the scope of the Ecuadorian Páramo Conservation Project, executed by the University of Amsterdam, Ecociencia, and the Mountain Institute, with financial support from the Royal Netherlands Embassy in Ecuador.
References CitedAcosta-Solis, M., 1984: Los páramos andinos del Ecuador. Quito:
Publicaciones MAS, 220 pp.Ange Jaramillo, C., Castaño Uribe, C., Arjona Hincapie, F., Rodriguez,
J. V., and Durán, C. L., 2002: Congreso Mundial de Páramos. Memorias. Bogotá: Ministerio del Medio Ambiente, CAR, IDEAM, Conservation International, 1185 pp.
Anhalzer, J. J., 1998: Llanganati. Quito: Libri Mundi, 97 pp.Bader, M. Y., 2007: Tropical Alpine Treelines: How Ecological
Processes Control Vegetation Patterns and Dynamics. Ph.D. thesis, Wageningen, Wageningen University, 193 pp.
Bader, M. Y., Rietkerk, M., and Bregt, A. K., 2007: Vegetation structure and temperature regimes of tropical alpine treelines. Arctic, Antarctic, and Alpine Research 39: 353–364.
Baker, P. A., Seltzer, G. O., Fritz, S. C., Dunbar, R. B., Grove, M. J., Tapia, P. M., Cross, S. L., Brown, H. D., and Broda, J. P., 2001: The history of South American tropical precipitation for the past 25,000 years. Science, 291: 640–643.
Blakemore, L. C., Searle, P L., and Daly, B. K., 1981: Methods for Chemical Analysis of Soils. Wellington: New Zealand Soil Bureau, Scientific Report 80, 103 pp.
Buytaert, W., Célleri, R., De Bièvre, B., Cisneros, F., Wyseure, G., Deckers, J., and Hofstede, R., 2006: Human impact on the hydrology of the Andean páramos. Earth-Science Reviews, 79: 53–72.
Buytaert, W., and De Bièvre, B., 2012: Water for cities: the impact of climate change and demographic growth in the tropical Andes. Water Resources Research, 48: W08503.
Cabaneiro, A., Fernandez, I., Pérez-Ventura, L., and Carballas, T., 2008: Soil CO
2 emissions from Northern Andean páramo
ecosystems: Effects of fallow agriculture. Environmental Science and Technology, 42: 1408–1415.
Castaño, C. 2002: Páramos y ecosistemas alto andinos de Colombia en condición hotspot y global climatic tensor. Bogotá: IDEAM.
Chimner, R. A., and Karberg, J. M., 2008: Long-term carbon accumulation in two tropical mountain peatlands, Andes Mountains, Ecuador. Mires and Peat, 3: art 4.
Clarke, K. R., and Warwick, R. M., 2001: Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd edition. Plymouth: Primer-E, 172 pp.
Cleef, A. M., 1981: The Vegetation of the Páramos of the Colombian Cordillera Oriental. Vaduz, Liechtenstein: J. Cramer, 321 pp.
Contreras, D. A., 2010: Landscape and environment: insights from the prehispanic Central Andes. Journal of Archaeological Research, 18: 241–288.
Cuatrecasas, J., 1958: Aspectos de la vegetación natural de Colombia. Revista de la Academia Colombiana de Ciencias Exactas y Físicas, 10: 221–264.
Cuatrecasas, J., 2013: A Systematic Study of the Subtribe Espeletiinae (Heliantaceae, Asteracea). New York: NYBG Press, Memoirs of the New York Botanical Garden, Volume 107, 704 pp.
Cuesta, F., Bustamante, M., Becerra, M. T., Postigo, J., and Peralvo M., 2012: Panorama andino de cambio climático: vulnerabilidad y adaptación en los Andes Tropicales. Lima: Condesan, SG-CAN, 329 pp.
Di Pasquale, G., Marziano, M., Impagliazzo, S., Lubritto, S., De Natale, A., and Bader, M. Y., 2008: The Holocene treeline in the northern Andes (Ecuador): First evidence from soil charcoal. Palaeogeography, Palaeoclimatology, Palaeoecology, 259: 17–34.
Dickinson, K. J. M., Mark, A. F., Barratt, B. I. P., and Patrick, B. H., 1998: Rapid ecological survey, inventory and implementation: a case study from Waikaia Ecological Region, New Zealand. Journal of the Royal Society of New Zealand, 28: 83–156.
Ellenberg, H., 1958: Wald oder Steppe. Die natüraliche Planzendecke der Anden Perus (I, II). Die Umschau im Wissenschaft und Technik, 21: 679–681, 22: 679–681.
Ellenberg, H., 1979: Man’s influence on tropical mountain ecosystems in South America. Journal of Ecology, 67: 401–416.
FAO/ISRIC/ISSS, 1998: World Reference Base for Soil Resources. Rome: FAO, World Soil Resources Report, Volume 84, 88 pp.
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

ROBERT G. M. HOFSTEDE ET AL. / 985
Farley, K. A., Jobbágy, E. G., and Jackson, R. B., 2005: Effects of afforestation on water yield: a global synthesis with implications for policy. Global Change Biology, 11: 1565–1576.
Farley, K. A., Kelly, E. F., and Hofstede, R. G. M., 2004: Soil organic carbon and water retention after conversion of grasslands to pine plantations in the Ecuadorian Andes. Ecosystems, 7: 729–739.
Germino, M. J., and Smith, W. K., 1999: Sky exposure, crown architecture, and low-temperature photoinhibition in conifer seedlings at alpine treeline. Plant, Cell, and Environment, 22: 407–415.
González, W., Llambí, D. L., and Smith, J. K., 2011: Dinámica sucesional del componente arbóreo en la transición entre el bosque-páramo en los Andes Tropicales. Ecotrópicos, 24: 60–79.
Grabherr, G., Gottfried, M., and Pauli, H., 2010: Climate change impacts in alpine environments. Geography Compass, 4: 1133–1153.
Grace, J., Berninger, F., and Nagy, L., 2002: Impacts of climate change on the tree line. Annals of Botany, 90: 537–544.
Hofstede, R. G. M., 1995: Effects of livestock farming and recommendations for management and conservation of páramo grasslands (Colombia). Land Degradation and Rehabilitation, 6: 133–147.
Hofstede, R. G. M., Groenendijk, J. P., Coppus, R., Fehse J., and Sevink, J., 2002: Impact of pine plantations on soils and vegetation in the Ecuadorian high Andes. Mountain Research and Development, 22: 159–167.
Hofstede, R., Segarra, P., and Mena Vásconez, P., 2003: Los páramos del mundo. Quito: IUCN, Global Peatland Initiative, Ecociencia, 299 pp.
Hofstede, R., 2011: Historia, tendencias y perspectivas para el manejo de los páramos. In Maldonado, G., and de Bievre, B. (eds.), PARAMUNDI, 2do Congreso Mundial de Páramos. Memorias. Lima: CONDESAN, 34–45.
Innes, J. L., 1991: High-altitude and high-latitude tree growth in relation to past, present and future global climate change. Holocene, 1: 168–173.
Jantz, N., and Behling, H., 2012: A Holocene environmental record reflecting vegetation, climate, and fire variability at the Páramo of Quimsacocha, southwestern Ecuadorian Andes. Vegetation History and Archaeobotany, 21: 169–185.
Jørgensen, P. M., and Ulloa Ulloa, C., 1994: Seed Plants of the High Andes of Ecuador: A Checklist. Aarhus: Department of Systematic Botany, Aarhus University, AAU Report 34, 453 pp.
Kessler, M., Grytnes, J. A., Halloy, S. R. P., Kluge, J., Krömer, T., León, B., Macía, M. J., and Young, K. R., 2011: Gradients of plant diversity: local patterns and processes. In Herzog, S. K., Martínez, R., Jørgensen, P. M., and Tiessen, H. (eds.), Climate Change and Biodiversity in the Tropical Andes. São José dos Campos, Brazil: IAI-SCOPE, 204–219.
Körner, C., 1998: A re-assessment of high elevation treeline positions and their explanation. Oecologia, 115: 445–459.
Körner, Ch., and Spehn, E., 2002: Mountain Biodiversity: A Global Assessment. New York: Parthenon, 341 pp.
Körner, C., and Paulsen, J., 2004: A world-wide study of high altitude treeline temperatures. Journal of Biogeography, 31: 713–732.
Körner, C., 2012: Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits. Basel, Switzerland: Springer, 220 pp.
Laegaard, S., 1992: Influence of fire in the grass páramo vegetation of Ecuador. In Balslev, H., and Luteyn, J. L. (eds.), Páramo: An Andean Ecosystem under Human Influence. London: Academic Press, 151–170.
Lauer, W., 1981: Ecoclimatological conditions of the pàramo belt in the tropical high mountains. Mountain Research and Development, 1: 209–221.
Llambí, L. D., Puentes, J. A., and Garcia-Núñez, C., 2013: Spatial relations and population structure of a dominant tree along a treeline ecotone in the Tropical Andes: interactions at gradient and plant-neighbourhood scales. Plant Ecology & Diversity, 6: 343–353.
López Sandoval, M. F., 2004: Agricultural and Settlement Frontiers in the Tropical Andes: The Páramo Belt of Northern Ecuador, 1960–1990. Regensburg, Germany: University of Regensburg, Regensburger Geographische Schriften, Heft 37, 180 pp.
Luteyn, J. L., 1999: Páramos. A Checklist of Plant Diversity, Geographical Distribution and Botanical Literature. New York: The New York Botanical Garden Press, Memoirs of the New York Botanical Garden, Volume 84, 278 pp.
Marchant, R., and Hooghiemstra, H., 2004: Rapid environmental change in African and South American tropics around 4000 years before present: a review. Earth Science Reviews, 66: 217–260.
Miehe, G., and Miehe, S., 1994: Zur oberen Waldgrenze in tropischen Gebirge. Phytocoenologia, 24: 53–110.
Miehe, G., and Miehe, S., 2000: Comparative high mountain research on the treeline ecotone under human impact. Carl Troll’s “asymmetrical zonation of the humid vegetation types of the world” of 1948 reconsidered. Erdkunde, 54: 34–50.
Monasterio, M., and Sarmiento, L., 1991: Adaptive radiation of Espeletia in the cold Andean tropics. Trends in Ecology and Evolution, 6: 387–391.
Moscol, M. C., 2010: Holocene Upper Forest Line Dynamics in the Ecuadorian Andes. Ph.D. dissertation. Amsterdam: University of Amsterdam, 228 pp.
Mothes, P., 1991: El paisaje volcánico de la Sierra Ecuatoriana. Geomorfología, fenómenos volcánicos y recursos asociados. Quito: Corporación Editora Nacional/Colegio de Geógrafos del Ecuador, Estudios de Geografía, 4, 92 pp.
Podwojewski, P., Poulenard, J., Zambrana, T., and Hofstede, R., 2002: Overgrazing effects on vegetation cover and volcanic ash soil properties in the páramo of Llangahua and La Esperanza (Tungurahua, Ecuador). Soil Use and Management, 18: 45–55.
Price, M., 2006: Global Change in Mountain Regions. Duncow, U.K.: Sapiens, 343 pp.
Ramírez, L., Llambí, L. D., Schwarzkopf, T., Gámez, L. E., and Márquez, N. J., 2009: Vegetation structure along the forest-páramo transition belt in the Sierra Nevada de Mérida: implications for understanding treeline dynamics. Ecotrópicos, 22: 83–98.
Rangel, C. J. O., 2000: Colombia Diversidad Biológica III. La región de vida paramuna. Bogotá: Universidad Nacional de Colombia, Instituto de Ciencias Naturales, Instituto Alexander von Humboldt, 902 pp.
Smith, A. P., 1994: Introduction to tropical alpine vegetation. In Rundel, P. W., Smith, A. P., and Meinzer, F. C. (eds.), Tropical Alpine Environments. Plant Form and Function. Cambridge: Cambridge University Press, 1–19.
Salamanca, S., 1992: La vegetación del páramo y su dinámica en la macizo volcanico Ruiz-Tolima (Cordillera Central, Colombia). Bogotá: Instituto Geográfico Augustin Codazzi, Analisis Geograficos, 21, 155 pp.
Sarmiento, F. O., 2002: Human drivers of landscape change: treeline dynamics in neotropical montology. Ecotropicos, 15: 129–146.
Sklenár, P., and Balslev, H., 2004: Superpáramo plant species diversity and phytogeography in Ecuador. Flora, 200: 416–433.
Smith, A. P., 1978: Establishment of seedlings of Polylepis sericea in the páramo (Alpine) zone of the Venezuelan Andes. Bartonia, 45: 11–14.
Stevens, G. C., and Fox, J. F., 1991: The causes of treeline. Annual Review of Ecology and Systematics, 22: 177–191.
Tonneijck, F. H., Van Der Plicht, J., Jansen, B., Verstraten, J. M., and Hooghiemstra, H., 2006: Radiocarbon dating of soil organic matter fractions in Andosols in northern Ecuador. Radiocarbon, 48: 337–353.
Thouret, J. C., and Faivre, P., 1989: Suelos de la Cordillera Central. In Van der Hammen, T., Diaz Piedrahita, S., and Alvarez, V. J. (eds.), La Cordillera Central Colombiana. Transecto Parque los Nevados (segunda parte). Estudios de Ecosistemas Tropandinos, 3. Berlin/Stuttgart: J. Cramer, 293–442.
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use

986 / ARCTIC, ANTARCTIC, AND ALPINE RESEARCH
Ulloa Ulloa, C., and Jørgensen, P., 1993: Árboles y arbustos de los Andes del Ecuador. AAU Reports 30. Aarhus, Denmark: Department of Systematic Botany, Aarhus University, 264 pp.
Vázquez, M. A., Larrea, M., and Suárez, L., 2000: Biodiversidad en el Parque Nacional Llanganates: un reporte de las evaluaciones y socioeconómicas rápidas. Quito: EcoCiencia, Ministerio del Ambiente, Herbario Nacional del Ecuador, Museo Ecuatoriana de Ciencias Naturales, Instituto Internacional de Reconstrucción Rural, 203 pp.
Vélez, V., Cavelier, J., and Devia, B., 1998: Ecological traits of the tropical treeline species Polylepis quadrijuga (Rosaceae) in the Andes of Colombia. Journal of Tropical Ecology, 15: 771–787.
Verweij, P. A., 1995: Spatial and Temporal Modelling of Vegetation Patterns. Burning and Grazing in the Páramo of Los Nevados National Park, Colombia. Ph.D. dissertation. Amsterdam: University of Amsterdam, 233 pp.
Wada, K., and Higashi, T., 1976: The categories of aluminium- and iron-humus complexes in ando soils determined by selective dissolution. Journal of Soil Science, 27: 357–368.
Walter, H., and Medina, E., 1969: La temperatura del suelo como factor determinante para la caracterización de los pisos subalpino y alpino en los Andes de Venezuela. Boletín de la Sociedad Venezolana de Ciencias Naturales, 28: 201–210.
Wardle, P., 1971: An explanation for alpine timberline. New Zealand Journal of Botany, 9: 371–402.
Wille, M., Hooghiemstra, H., Hofstede, R., Fehse, J., and Sevink, J., 2002: Upper forest line reconstruction in a deforested area in northern Ecuador based on pollen and vegetation analysis. Journal of Tropical Ecology, 18: 409–440.
Yánez, P., 1998: Caracterización florística en un sector de cambio páramo-selva nublada en el Parque Nacional Sierra Nevada, Venezuela. Revista Forestal Venezolana, 42: 51–62.
Young, K. R., and Keating, P. L., 2001: Remnant forests of Volcán Cotacachi, northern Ecuador. Arctic, Antarctic, and Alpine Research, 33: 165–172.
Young, K. R., and León, B., 2007: Tree-line changes along the Andes: implications of spatial patterns and dynamics. Philosophical Transactions of the Royal Society B, 362: 263–272.
Young, K. R., 2009: Andean land use and biodiversity: humanized landscapes in a time of change. Annals of the Missouri Botanical Garden, 96: 492–507.
Zehetner, F., Miller, W. P., and West, L. T., 2003: Pedogenesis of volcanic ash soils in Andean Ecuador. Journal of the Soil Science Society of America, 67: 1797–1809.
MS accepted September 2014
Downloaded From: https://bioone.org/journals/Arctic,-Antarctic,-and-Alpine-Research on 08 Oct 2020Terms of Use: https://bioone.org/terms-of-use