Embed Size (px)
Citation preview

452
BBA 45 129
UPTAKE OF MAGNESIUM AND CALCIUM
EXCHANGE FOR HYDROGEN IONS
J. D. JUDAH, K. AHMED, A. E. M. McLEAN AND G. S. CHRISTIE*
Division of Metabolic Research, The Chicago Medical School, Institute for Medical Research, Chicago, Ill. (U.S.A.)
(Received July Ioth, i964)
BIOCHIMICA ET BIOPHYSICA ACTA
BY MITOCHONDRIA IN
SUMMARY
I. Rat-liver mitochondria take up Mg 2+ in exchange for H +. 2. Inorganic orthophosphate is not essential for Mg ~+ uptake. 3. Inorganic orthophosphate uptake can take place in the absence of Mg ~+. 4- Rat-liver mitochondria take up Ca 2+, in part at least, by the same route as
Mg~+. H+ is exchanged for Ca 2+. 5. The divalent cations appear to share a common transport mechanism
with K+. 6. I t is concluded that mitochondrial cation-transport mechanism is a system
which extrudes H+. Any one of several cations may be taken up in exchange. 7. An explanation is advanced for the opposing effects of 2,4-dinitrophenol and
Ca ~÷ on mitochondrial swelling. 8. The sensitivity and specificity of atomic absorption spectrophotometry fa-
cilitated analysis of Ca 2+ and Mg *+.
INTRODUCTION
In previous papers 1,2 we described conditions for obtaining net inward transport of K+ by rat-liver mitochondria, K + uptake was found to be associated with H ÷ efflux in i : I ratio. In the present work, transport of the divalent cations Mg 2+ and Ca ~+ has been investigated. Mg *+ appeared to be transported by the same mechanism as K +, and Mg ~+ uptake was associated with H + efflux in the ratio of I equiv, of H + to each equiv, of Mg 2+ (2 H÷ to each mole of Mg*+). Evidence is presented indicating that Mg 2+ and K+ occupy the same sites. Inward transport of inorganic phosphate could occur independently of Mg *+ although the two ions may interact indirectly. Some experiments with Ca ~+ are included for comparison with Mg ~+ transport.
METHODS
Animals, mitochondrial preparations and depletion procedure, and methods of analysis for K+ and Pt have been previously described1, 2.
p H changes following addition of cations
These changes were measured by titrating to constant pH by means of a Radio- meter recording automatic titrator. The mitochondria were usually added last, the
* Present address: Department of Pathology, University of Queensland, Brisbane, Australia.
Biochim. Biophys. Acta, 94 (1965) 452-46o

MAGNESIUM AND CALCIUM IN MITOCHONDRIA 453
pH was then rapidly adjusted to the desired value and the recorder started. When it was necessary to make a final addition of cation to mitochondria suspended in a relatively complex medium (containing substrate o r Pt or both), it was necessary to correct any subsequent fall in pH for that produced in the medium in the absence of mitochondria. This correction was only important with Mg *+ at the concentration used (IO mM), in the presence of Pi. Ca ~+ was used at a low concentration (0.2 mM), which produced only a small blank due to the medium.
Estimation of Ca 2+ and Mg 2+
These elements were determined by atomic absorption spectrophotometry z. The Techtron light sources, power supply, lenses, burner and gas regulator for acetylene and air were used in coniunction with the Zeiss M4QIII monochromator, PMQII detector and 75-cm optical bench. In estimating Ca z+ we used a regulated, unmodu- lated lamp current of 15 mA, a wavelength of 422.7 mt~, a 4o-~ slit and convenient photomultiplier settings. The flame characteristics were important, we found that a slightly luminous flame rich in acetylene was necessary; the burner position was less critical. When the settings were determined for the instrument, steady and reproducible readings were obtained. No interference by Na + or K + was observed. Phosphate levels in our samples were insufficient to cause interference, so that Sr 2+ or La 3+ addition4, 5 was not required. Trichloroacetic acid extracts of mitochondria were directly aspirated into the atomiser; the standards were made up in the same concentration of trichloroacetic acid as the test samples. In addition to absorption, some emis~,,ion was also observed, but the emission signal was eliminated by ap- propriate settings of the lamp and slit. Readings were linear at Ca ~+ concentrations between 0.o 5 and 0. 5 mM. Mg 2+ was estimated at 285.2 m~, using a lamp current of 8 mA; other conditions were the same as for Ca 2+. Readings were linear at Mg ~+ concentrations between 5 t ~M and o.o5 mM. The atomic absorption equipment was manufactured by Techtron Pty. Ltd., South Melbourne, Australia, and is available in the United States from Aztec Instruments Inc., Westport, Conn.
RESULTS
Uptake of Mg 2+
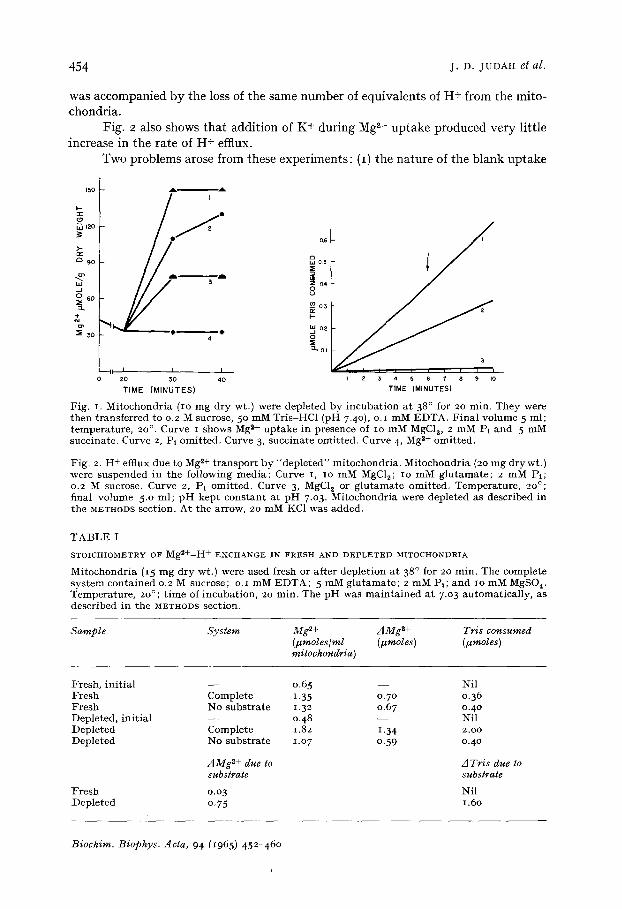
The Mg 2+ content of our freshly prepared mitochondria was 41.5 + 3.o mmole/kg dry wt. When the mitochondria were incubated at 38 ° for 20 min (depleted prepa- ration), the level fell by approx. 25 %. This contrasts with the greater relative loss of K + (ref. I). Fig. I shows the uptake of Mg z+ by a depleted preparation. The salient features were a large initial uptake of Mg 2+ even in the absence of substrate; this uptake was not materially influenced by the presence or absence of PI or of EDTA (o.I mM). In the presence of substrate, there was a further Mg 2+ uptake, more rapid in the presence of Pt, but tending to the same upper limit even in its absence.
Fig. 2 shows H + efflux in the same system. In the presence of substrate the rates of Mg z+ uptake and H+ efflux lay between a i : I and a 2:1 ratio (expressed in mequiv.). In the absence of substrate, however, this ratio was less than I. Table I shows the stoichiometry of the Mg 2+ uptake and H + efflux. After subtraction of the relevant blanks, it was apparent that the extra Mg 2+ uptake induced by substrate
Biochim. Biophys. Acta, 94 (1965) 452-460
454 j .D. JUDAH et al.
was accompanied by the loss of the same number of equivalents of H+ from the mito- chondria.
Fig. 2 also shows that addition of K+ during Mg ~÷ uptake produced very little increase in the rate of H+ efflux.
Two problems arose from these experiments: (i) the nature of the blank uptake
$- 15o ~b. I A iI ;T~ 120 2
C} 90 j ~
50 ~ 4 ~
t ~ I p I 510 I 0 20 40
TIME {MINUTES)
0.6
o.5
o4 o
u) o.5 p. i~, o2 o :1-o. I
i
' ' ' , , i i I
I 2 3 4 5 6 7 8 9 10
TIME (MINUTES)
Fig. i. Mi tochondr i a ( io mg d ry wt.) were deple ted by i n c u b a t i o n a t 380 for 2o min . They were then t r ans fe r red to o.2 M sucrose, 5 ° mM Tris-HC1 (pH 7.4o), o. z mM EDTA. F i n a l vo lume 5 ml ; t empe ra tu r e , 2o °. Curve I shows Mg z+ u p t a k e in presence of IO mM MgCI~, 2 mM Pt and 5 mM succ ina te . Curve 2, Pt omi t t ed . Curve 3, succ ina te omi t t ed . Curve 4, Mg ~+ omi t t ed .
Fig. 2. H + efflux due to Mg 2+ t r a n s p o r t b y " d e p l e t e d " mi tochondr i a . Mi tochondr i a (2o mg dry wt.) were suspended in the fo l lowing med ia : Curve i, IO mM MgC12; IO mM g l u t a m a t e ; 2 mM Fl; 0.2 M sucrose. Curve 2, Pt omi t t ed . Curve 3, MgCI~ or g l u t a m a t e omi t t ed . Tempera tu re , 20°; f inal vo lume 5.o ml ; pH k e p t c o n s t a n t a t p H 7.03. Mi tochondr i a were deple ted as descr ibed in the METHODS sect ion. A t the arrow, 2o mM KC1 was added.
T A B L E I
S T O I C H I O M E T R Y OF Mg2+-H + E X C H A N G E I N F R E S H A N D D E P L E T E D M I T O C H O N D R I A
Mitochondr i a (I 5 m g d ry wt.) were used fresh or a f te r dep le t ion a t 38~ for 2o min . The comple te s y s t e m con ta ined o.2 M sucrose; o.I mM E D T A ; 5 mM g l u t a m a t e ; 2 mM Pt; and io mM MgSO a. Tempera tu re , 2o ° ; t i m e of incuba t ion , 20 rain. The p H was m a i n t a i n e d a t 7.o3 au t oma t i ca l l y , as descr ibed in the METHODS sect ion.
Sample System Mg 2+ AMg ~+ Tris consumed (l~moles/ml (#,moles) (l~moles) mitochondria)
Fresh, i n i t i a l - - 0.65 - - Nil Fresh Complete 1.35 o.7o o.36 Fresh No subs t r a t e 1.32 0.67 0.40 Depleted, i n i t i a l - - 0.48 - - Ni l Dep le ted Complete 1.82 1.34 2.oo Dep le ted No subs t r a t e 1.o 7 0.59 0.40
A Mg 2+ due to ATris due to substrate substvate
F r e s h 0.o 3 Ni l Dep le ted o.75 1.6o
Biochim. Biophys. Aeta, 94 (1965) 452-460
M A G N E S I U M A N D CALCIUM IN M I T O C H O N D R I A 4 5 5
of Mg 2+ and (2) the reason why the substrate-induced Mg ~+ uptake was shown only by depleted mitochondria. The first problem will be considered later, in connection with a similar effect shown by Ca ~+.
Relation of r~'citochondrial K + level to Mg ~+ uptake
Mitochondria subjected to the standard depletion procedure lost 50-7 ° % of their K+ content, but only 20-25 % of their Mg 2+. Furthermore, in the presence of substrate the Mg ~+ levels rose far above the value for the freshly prepared particles. These findings suggested that the Mg 2+ taken up was occupying sites vacated by K +. This interpretation was supported by the experiment shown in Table II, in which the presence of Mg ~+ depressed K + uptake by depleted mitochondria.
TABLE II
EFFECT OF IV[I~2+ ON K + UPTAKE BY DEPLETED MITOCHONDRIA
Mitochondria (8.2 mg dry wt.) were depleted at 38 ° for 2o rain. They were then transferred to a medium containing o.I 7 M sucrose; 2o mM IiCI; io mM glutamate; I mM Pt; and Mg z+ as shown. Temperature, 38°; incubation time, io min.
Sample Medium K + content A K % inhibition Mg ~+ (raM) (mequiv./hg
dry wt.)
I n i t i a l - - 8 6 - - - -
I n c u b a t e d o .o 128 42 - - I n c u b a t e d i . o 118 32 23 I n c u b a t e d 3 .o 112 26 38 I n c u b a t e d 5 .0 9 6 IO 76
T A B L E I I I
STABILIZING :EFFECT OF M g ~+ ON MITOCHONDRIAL K + LEVEL
M i t o c h o n d r i a (6 m g d r y w t . ) w e r e s u s p e n d e d i n 0 .22 M s u c r o s e ; o . i m M E D T A ; 5o m M T r i s - H C 1 ( p H 7.40) . T e m p e r a t u r e , 2 0 ° ; f i n a l v o l u m e 5 ml . O t h e r d e t a i l s a s i n t h e METHODS s e c t i o n . I n - c u b a t i o n t i m e i o m i l l .
Sample Addition (raM) K + content A K + (mequiv./kg dry wt.)
I n i t i a l - - 138 - - I n c u b a t e d - - 121 - - i 7 I n c u b a t e d M g 2+, IO 129 - - 9 I n c u b a t e d 2 , 4 - D i n i t r o p h e n o l , o . i 63 - - 7 5 I n c u b a t e d 2 . 4 - D i n i t r o p h e n o l p l u s M g ~+ 117 - - 2 1
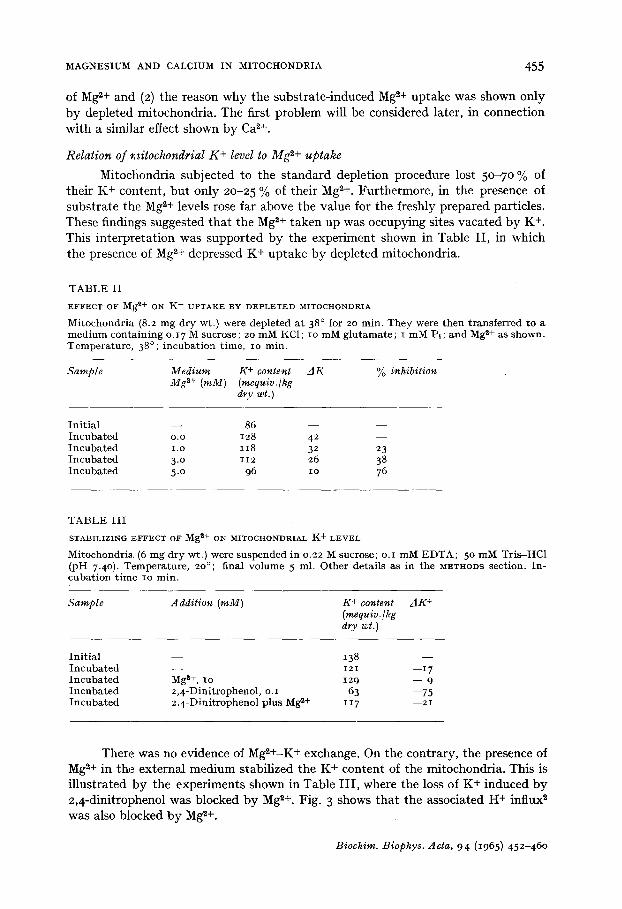
There was no evidence of MgZ+-K + exchange. On the contrary, the presence of Mg ~+ in the external medium stabilized the K + content of the mitochondria. This is illustrated by the experiments shown in Table III , where the loss of K+ induced by 2,4-dinitrophenol was blocked by Mg ~+. Fig. 3 shows that the associated H+ influx 2 was also blocked by Mg z+.
Bioehim. Biophys. Acta, 9 4 (1965) 4 5 2 - 4 6 0
456 J.D. JUDAH et al.
Relation of P~ to Mg ~+ uptake
The preceding experiments (Fig. I) showed that Mg ~+ uptake could occur in the absence of Pl. Table IV shows that Pi uptake could occur in the absence of Mg 2+ from the medium.TableVshows that if the mitochondria were only partially depleted, the requirement for substrate was not apparent. In these experiments a very rapid uptake o f P l occurred, even in the absence of added Mg 2+. Table V also shows that 2,4-dinitro- phenol (o.I mM) blocked Pi uptake, and Table VI shows that 2,4-dinitrophenol released Pi from freshly prepared mitochondria.
=¢~ 0.6
c~5
O~ O.4
IJJ Ck3
~: o.2 :::L
TIME (MINUTES)
1,4
~:~ 1.2
I.O
0.8
0 0 . 6 =E ::L
0.4
0.2 ~ . ~
I
l l
I I I I I 3 4 5 6 7
TIME (MINUTES)
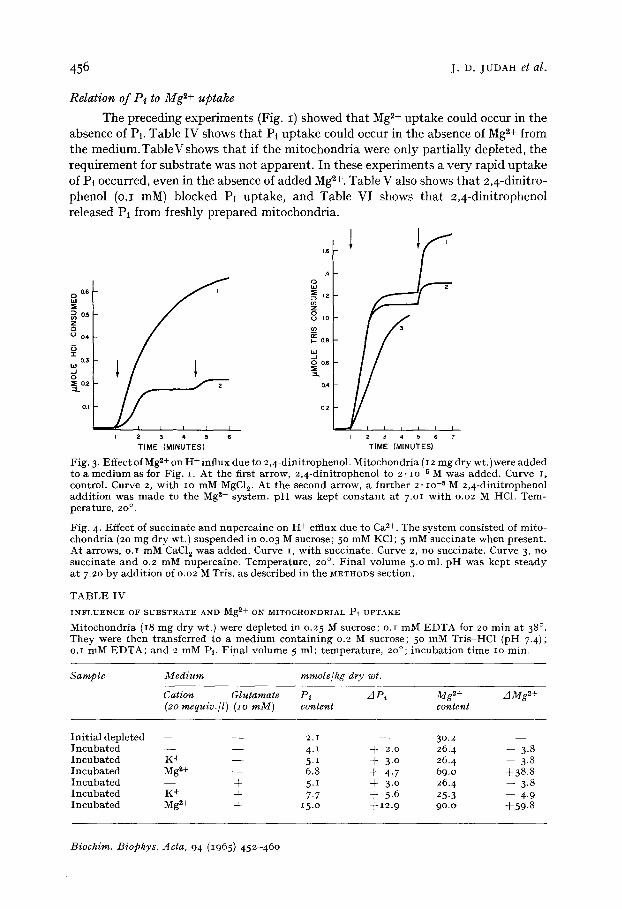
F i g . 3- E f f e c t o f M g ~+ o n H + i n f l u x d u e t o 2 , 4 - d i n i t r o p h e n o l . M i t o c h o n d r i a (I 2 m g d r y w t . ) w e r e a d d e d t o a m e d i u m a s f o r F i g . i . A t t h e f i r s t a r r o w , 2 , 4 - d i n i t r o p h e n o l t o 2 . lO -5 M w a s a d d e d . C u r v e I , c o n t r o l . C u r v e 2, w i t h IO m M MgC1 v A t t h e s e c o n d a r r o w , a f u r t h e r 2 . lO -5 M 2 , 4 - d i n i t r o p h e n o l a d d i t i o n w a s m a d e t o t h e M g 2+ s y s t e m , p H w a s k e p t c o n s t a n t a t 7.Ol w i t h o .o2 M HC1. T e m - p e r a t u r e , 2 0 ° .
F i g . 4. E f f e c t of s u c c i n a t e a n d n u p e r c a i n e o n H + e f f lux d u e t o C a 2+. T h e s y s t e m c o n s i s t e d o f m i t o - c h o n d r i a (20 m g d r y w t . ) s u s p e n d e d i n 0 .03 M s u c r o s e ; 5 ° m M KC1; 5 m M s u c c i n a t e w h e n p r e s e n t . A t a r r o w s , o . I m M CaCI~ w a s a d d e d . C u r v e i , w i t h s u c c i n a t e . C u r v e 2, n o s u c c i n a t e . C u r v e 3, n o s u c c i n a t e a n d 0 .2 m M n u p e r c a i n e . T e m p e r a t u r e , 20 °. F i n a l v o l u m e 5 .o ml . p H w a s k e p t s t e a d y a t 7 .20 b y a d d i t i o n of 0 .02 M T r i s , a s d e s c r i b e d i n t h e METHODS s e c t i o n .
T A B L E I V
I N F L U E N C E OF SUBSTRATE AND M g 2+ ON MITOCHONDRIAL P i UPTAKE
M i t o c h o n d r i a ( I8 m g d r y w t . ) w e r e d e p l e t e d i n 0 .25 M s u c r o s e ; o . i m M E D T A f o r 2o m i n a t 38° . T h e y w e r e t h e n t r a n s f e r r e d t o a m e d i u m c o n t a i n i n g o .2 M s u c r o s e ; 50 m M T r i s - H C l ( p H 7 . 4 ) ; o . i m M E D T A ; a n d 2 m M Pt . F i n a l v o l u m e 5 m l ; t e m p e r a t u r e , 2o° ; i n c u b a t i o n t i m e i o m i n .
Sample Medium mmole/kg dry wt.
Cation Glutamate Pt AP~ Mg 2+ AMg ~+ (2o mequiv./l) (io raM) content content
I n i t i a l d e p l e t e d - - - - 2. i - - 3 o. 2 - - I n c u b a t e d - - - - 4.1 + 2 .0 26. 4 - - 3 .8 I n c u b a t e d K + - - 5 . I + 3 .0 26 .4 - - 3.8 I n c u b a t e d M g ~+ - - 6 .8 + 4 .7 69 .0 - 7 3 8 . 8 I n c u b a t e d - - + 5.1 + 3 .0 26 .4 - - 3 .8 I n c u b a t e d K + + 7.7 + 5 .6 25.3 - - 4 .9 I n c u b a t e d M g 2+ + 15 .o + 12.9 9 o . 0 + 5 9 . 8
Biochim. Biophys. Acta, 94 (1965) 4 5 2 - 4 6 o

M A G N E S I U M A N D CALCIUM IN M I T O C H O N D R I A 457
Uptake of Ca 2+
Table VII shows that Ca z+ was taken up by fresh mitochondria, and that sub- strate increased the uptake. Fig. 4 shows the accompanying H + efflux. The data indicate that uptake of Ca *+ was associated with H + efflux in the ratio of I equiv.
T A B L E V
EFFECT OF 2,4-DINITROPHENOL ON P I UPTAKE BY MITOCHONDRIA
M i t o c h o n d r i a (5 m g d r y w t . ) w e r e i n c u b a t e d f o r IO r a i n a t 20 ° i n o . 1 3 4 M K C I ; o .o3 M s u c r o s e ; 5 ° m M T r i s - H C 1 ( p H 7.4) ; o. I m M E D T A ; f i n a l v o l u m e 5 ml , t o l o w e r t h e i r P t l eve l . A d d i t i o n s of P t a n d 2 , 4 - d i n i t r o p h e n o l w e r e t h e n m a d e a s s h o w n . I n c u b a t i o n w a s c o n t i n u e d f o r a f u r t h e r i o r a i n , w h e n t h e m i t o c h o n d r i a w e r e s e p a r a t e d f o r a n a l y s i s of PI, w e t w t . a n d d r y w t . , a s d e s c r i b e d i n t h e METHODS s e c t i o n .
Sample Additions Pt content P, content z]Pt (mmole/kg (mmole/l (mmole/kg dry tot.) pellet roarer) dry tot.)
I n i t i a l N i l 14 .o 6. i - - I n c u b a t e d 2 m M P t 6 7 . 0 29.1 53 .0 I n c u b a t e d o . i m M 2 , 4 - d i n i t r o p h e n o l 14 .o 6 .6 - - I n c u b a t e d 2 , 4 - D i n i t r o p h e n o l p l u s P t 26. 7 i 1.6 12.7
T A B L E V I
EFFECT OF 2,4-DINITROPHENOL ON LEVEL OF MITOCHONDRIAL P t
M i t o c h o n d r i a (5 m g d r y w t . ) w e r e i n c u b a t e d a t 2o o f o r i o r a i n a s i n T a b l e V, w i t h a d d i t i o n s a s s h o w n .
Sample A dditions Pi content AP (mmole/kg (mmole/kg dry tot.) dry tot.)
I n i t i a l N i l 32 .0 - - I n c u b a t e d N i l 33 .5 + 1.5 I n c u b a t e d o . i m M 2 , 4 - d i n i t r o p h e n o l 12. 5 - - 1 5 . 5
T A B L E V I I
U P T A K E OF C a 2+ B Y MITOCHONDRIA
F r e s h m i t o c h o n d r i a (12 m g d r y w t . ) w e r e i n c u b a t e d i n a m e d i u m c o n t a i n i n g 0 . 0 6 M s u c r o s e ; 5 ° m M KC1; 5 ° m M T r i s - H C 1 ( p H 7.4) ; a n d a d d i t i o n s a s s h o w n . F i n a l v o l u m e 5 m l ; t e m p e r a t u r e , 2o ° ; t i m e 3 r a i n . C a ~+ w a s m e a s u r e d b y a t o m i c a b s o r p t i o n s p e c t r o p h o t o m e t r y a s d e s c r i b e d i n t h e METHODS s e c t i o n .
Sample Medium
C a 2+ Succinate (o.2 raM) (5 raM)
Ca ~+ content ACa+ (mmoles/kg dry wt.)
I n i t i a l - - - - lO. 7 - - I n c u b a t e d - - - - 9 .5 - - 1.2 I n c u b a t e d + - - 49 .5 + 38 .8 I n c u b a t e d - - + 8. 4 - - 2. 3 I n c u b a t e d + + 58. 5 + 4 7 . 8
Biochim. Biophys. Acta, 94 (1965) 4 5 2 - 4 6 0
458 J.D. JUDAH et aI.
of Ca 2+ to I equiv, of H +, in conformity with previous work 6. Part of the Ca 2+ uptake may be a surface attachment. Fig. 4 (curve 3) shows that nupercaine decreased the rate of H + release substantially. Nupercaine has been shown in other systems to act as a stabiliser of natural membranes, and we have found that nupercaine has no effect on mitochondrial K+-H+ exchange.
Effect of Ca 2+ on mitochondria
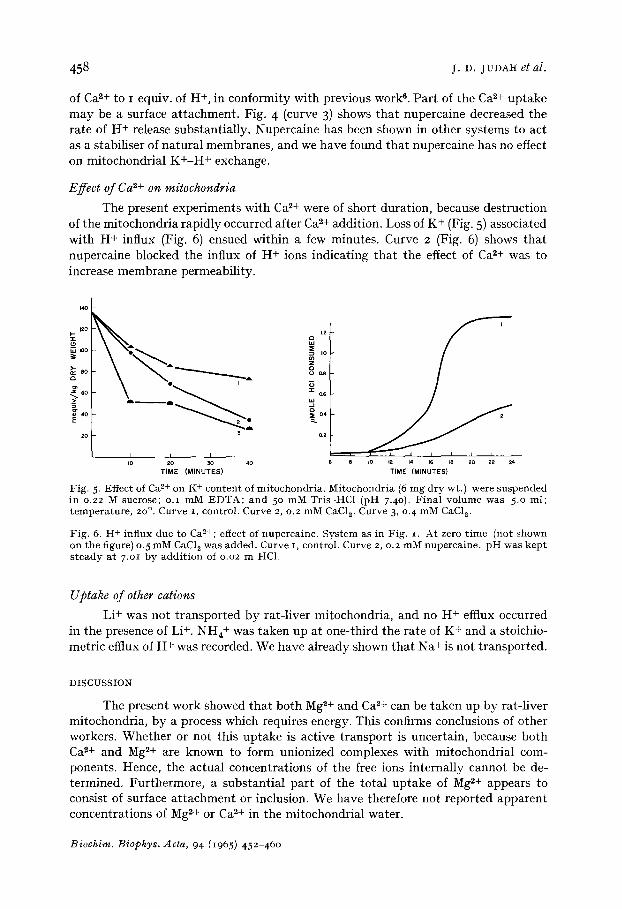
The present experiments with Ca ~+ were of short duration, because destruction of the mitochondria rapidly occurred after Ca 2+ addition. Loss of K + (Fig. 5) associated with H+ influx (Fig. 6) ensued within a few minutes. Curve 2 (Fig. 6) shows that nupercaine blocked the influx of H+ ions indicating that the effect of Ca*+ was to increase membrane permeability.
r4o
I - 120
~ I00
~ so 0
"~ 4O E
,'o 2" ;o 'o T IME (M INUTES)
1.2
I.O
o a4
0.2
, i t I I I I I I S Io 12 14 16 18 2O 22 24
T IME (M INUTES)
Fig. 5- Effect of Ca 2+ on K + conten t of mi tochondr i a . Mi tochondr i a (6 mg d ry wt.) were suspended in 0.22 M sucrose; o . i mM E D T A ; a nd 5 ° mM Tris-HC1 (pH 7.40). F i n a l vo lume was 5.0 ml ; t empe ra tu r e , 2o °. Curve i , control . Curve 2, 0.2 mM CaC12. Curve 3, o.4 mM CaC12.
Fig. 6. H + inf lux due to Ca l+ ; effect of nuperca ine . Sys tem as in Fig. i . At zero t i m e (not shown on the figure) 0. 5 mM CaC12 was added. Curve i, control . Curve 2, 0.2 mM nuperca ine , p H was k e p t s t e a d y a t 7.Ol by a d d i t i o n of 0.02 m HC1.
Uptake of other cations
Li+ was not transported by rat-liver mitochondria, and no H+ efflux occurred in the presence of Li÷. NH4+ was taken up at one-third the rate of K+ and a stoichio- metric efflux of H + was recorded. We have already shown that Na + is not transported.
DISCUSSION
The present work showed that both Mg 2+ and Ca 2+ can be taken up by rat-liver mitochondria, by a process which requires energy. This confirms conclusions of other workers. Whether or not this uptake is active transport is uncertain, because both Ca 2+ and Mg ~+ are known to form unionized complexes with mitochondrial com- ponents. Hence, the actual concentrations of the free ions internally cannot be de- termined. Furthermore, a substantial part of the total uptake of Mg ~+ appears to consist of surface attachment or inclusion. We have therefore not reported apparent concentrations of Mg 2+ or Ca 2+ in the mitochondrial water.
Biochim. Biophys. Acta, 94 (1965) 452-46o

MAGNESIUM AND CALCIUM IN MITOCI-IONDRIA 459
The data obtained for Mg 2+ suggest that this ion can be transported by the same mechanism as K +. The evidence is: (I) H + efflux occurred with both cations in a I : I ratio of equiv. (2) Addition of either cation in the presence of the other did not accelerate HI+ efflux. (3) The presence of Mg *+ in the system during substrate-induced K + uptake decreased K + transport. (4) Substrate-dependent Mg *+ uptake did not occur unless the mitochondria were depleted of K +.
This view is in conflict with that of BRIERLEY et al. ~, who implied that Mg *+ uptake is coupled to Pt uptake. Our experiments show that Mg *+ or PI may be taken up independently of each other. There is no doubt that the presence of both ions together results in faster rates of uptake, but this could be due to several possible mechanisms. One possibility is precipitation of magnesium phosphate within the mitochondria, thus lowering the concentration gradient of each ion across the mem- brane, and facilitating their entry.
In a previous paper, we suggested that our observation that K+-depleted mito- chondria are capable of oxidative phosphorylation, although they display no H ÷ gradient, cast doubt on MITCHELL'S hypothesis that concentration gradients of H + and O H - across the membrane control oxidative phosphorylation. Our present finding that Mg *÷ can replace K ÷ (thus restoring the H + gradient) removes this objection.
Pi uptake may represent true active transport. We have therefore expressed (Table V) the probable concentrations of Pi in the mitochondrial pellet water. In the absence of external Mg 2÷ an apparent concentration of 29 mM was attained, against an external concentration of 2 mM.
Ca*+ ~Lptake resembled Mg ~÷ uptake, but was rapidly followed by mitochondrial destruction, release of K + and uptake of H ÷.
The known effect of Ca 2+ in causing mitochondrial swelling may be explained by supposing that Ca *+ disturbs the permeability of the membrane to allow entry of external sol,ire and water more rapidly than internal solutes escape. This suggestion is supported by the lag period before K ÷ efflux and H ÷ influx occur. In contrast, 2,4-dinitrophenol "inhibits" mitochondrial swelling by permitting almost in- stantaneous equilibration of internal and external phases (Fig. 3)- The apparent protection against swelling produced by 2,4-dinitrophenol would then be merely a reflection of the severity of its action on the mitochondrial membrane. I t is possible that many other "anti-swelling" agents might act in a similar way to 2,4- dinitrophenol.
We have concluded that mitochondrial cation transport is performed by a mechanism which pumps out H +. Ca *÷, Mg 2÷, K ÷ or NH4÷ may be taken up in ex- change for H ÷, in this order of preference; Na ÷ and Lt ÷ are not actively taken up.
ACKNOWLEDGEMENTS
We are indebted to Miss S. ELSEY and Mr. W. GARRETT for skilled technical assistance. This work was supported by grants from the U.S. Public Health Service (AM 7226-02); the Life Insurance Medical Research Fund, the Otho S.A. Sprague Memorial hlst i tute and the Burroughs-Wellcome Fund. G. S. C. was in receipt of a travel grant from the Anti Cancer Council, Victoria, Australia. A. E.M. McL. was in receipt of a U.S. Public Health Service International Fellowship (No. FF 492).
Biochim. Biophys. Acta, 94 (1965) 452-46o

4 6 0 j . D . JUDAH el al.
REFERENCES
i G. S. CHRISTIE, K. AHMED, A. E. M. MCLEAN AND J. D. JUDAH, Biochim. Biophys. Acta, 94 (1965) 432.
2 J. D. JUDAH, A. E. M. MCLEAN, I£. AHMED AND G. S. CHRISTIE, Biochim. Biophys. Acta, 94 (1965) 441 .
3 J. B. WILLIS, Spectrochim. Acta, 16 (196o) 259. 4 D. J. DAVID, Analyst, 85 (196o) 495. 5 J. B. WILLIS, Anal. Chem., 34 (1962) 614. 6 N.-E. L. SARIS, The Calcium Pump in Mitochondria, Comm. Phys. Math. X X V I I I Soc. Scient.
Fenn. HeIsinki, 1963, p. 42. 7 G. BRIERLEY, E. MURER, E. BACHMANN AND D. E. GREEN, J. Biol. Chem., 238 (1963) 3482. 8 P. MITCHELL, Biochem. Soc. Syrup. (Cambridge, Engl.), 22 (1963) 142.
Biochim. Biophys. Acta, 94 (1965) 452-46o





























