Embed Size (px)
Citation preview

University of Groningen
Mechanisms involved in malabsorption of dietary lipidsKalivianakis, Mini
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:1998
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Kalivianakis, M. (1998). Mechanisms involved in malabsorption of dietary lipids. s.n.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
The publication may also be distributed here under the terms of Article 25fa of the Dutch Copyright Act, indicated by the “Taverne” license.More information can be found on the University of Groningen website: https://www.rug.nl/library/open-access/self-archiving-pure/taverne-amendment.
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 14-10-2021

RIJKSUNIVERSITEIT GRONINGEN
MECHANISMS INVOLVED INMALABSORPTION OF
DIETARY LIPIDS
PROEFSCHRIFT
ter verkrijging van het doctoraat in deMedische Wetenschappen
aan de Rijksuniversiteit Groningenop gezag van de
Rector Magnificus, dr. D.F.J. Bosscher,in het openbaar te verdedigen op
woensdag 23 september 1998om 14.45 uur
door
MINI KALIVIANAKIS
geboren op 25 februari 1970te Utrecht

Promotor: Prof. dr. R.J. VonkReferent: Dr. H.J. Verkade
ISBN: 90 367 0941 5

Promotiecommissie: Prof. dr. P.J.J. SauerProf. dr. G.L. ScherphofProf. dr. L.T. Weaver
The studies presented in this thesis were performed within the program of the GUIDEresearch school and were made possible by grants from the BIOMED Research Program(BMH1-CT93-1239, SIGN), Numico BV, Zoetermeer, and a starting grant from UniversityHospital Groningen.
The printing of this thesis was financially supported by ARC Laboratories, Amsterdam,Campro Scientific BV, Veenendaal, Glaxo Wellcome BV, Zeist, Hope Farms BV, Woerden.
Printer: Ponsen & Looijen BV, Wageningen, The Netherlands.

Contents
Chapter 1 General introduction1.1 Dietary lipids1.2 Intestinal absorption and digestion of dietary lipids1.3 Fate of lipids in the colon1.4 Lipid malabsorption1.5 Methods to measure lipid malabsorption1.6 Scope of this thesis
78
1014151721
Chapter 2 Detection of intestinal fat malabsorption due to impairedlipolysis by the 13C-mixed triglyceride breath test in ratsSubmitted
31
Chapter 3 The 13C-mixed triglyceride breath test in healthy adults:determinants of the 13CO2 responseEur J Clin Invest 1997;27:434-442
45
Chapter 4 The 13C-palmitic acid test with plasma sampling detects fatmalabsorption in bile-diverted ratsSubmitted
61
Chapter 5 The 13C-palmitic acid test for detection of fat malabsorptionin healthy adults on calcium supplementation
77
Chapter 6 Fat malabsorption in cystic fibrosis patients on enzymereplacement therapy is due to impaired intestinal uptakeof long-chain fatty acidsAm J Clin Nutr, in press 1998
89
Chapter 7 Increased fecal bile salt excretion is independent of thepresence of dietary fat malabsorption in two mouse modelsfor cystic fibrosis
105
Chapter 8 General discussion 119
Samenvatting 129
Nawoord 135

CHAPTER 1
General introduction

Chapter 1
8
CHAPTER 1
General introduction
1.1 Dietary lipids
On average, adult Western diets contain approximately 100 g of lipids per day, of which 92%to 96% are long-chain triacylglycerols [1,2]. Triacylglycerols (also referred to as triglycerides)are fatty acid triesters of glycerol (Figure 1.1). Triacylglycerols differ according to the identityand position of their three fatty acid residues. Most triacylglycerols in nature contain long-chain free fatty acids [1], although for example, triacylglycerols in human milk are mixturescontaining both medium- and long-chain fatty acids [3]. Fatty acids in biological systemsusually contain an even number of carbon atoms, typically between 14 and 24. The alkyl chainmay either be saturated or it may contain one or more double bonds. The predominant fattyacid residues in nature are those of the C16 and C18 species palmitic, oleic, linoleic, andstearic acids (Figure 1.2) [4]. Fatty acids can be divided into three main classes according totheir chain length: 1. short-chain fatty acids, less than 6 carbon atoms; 2. medium-chain fattyacids, from 6 to 12 carbon atoms; 3. long-chain fatty acids, 14 or more carbon atoms [5]. Theproperties of fatty acids are markedly dependent on their chain length and degree ofsaturation. Unsaturated fatty acids are more fluid than saturated fatty acids of the same length.By virtue of their smaller molecular size, medium-chain fatty acids are relatively soluble inwater [6].

General introduction
9
C
C
C
OH
OH
OH
H
H
H
H
H
1
2
3
C
C
C
O
O
O
O C
O
H
H
H
H
H
C
O
C
1
2
3
(A) (B)
Figure 1.1 The structural formulas of (a) glycerol and (b) a triacylglycerol.
An adequate intake of dietary lipid is essential for life and well-being. Lipids serveseveral important functions in the human body. Firstly, they represent the major source ofenergy (9 kcal g-1), double that of sugars and protein [4]. In the average Western diet, lipidsprovide approximately 40% of the caloric energy [1], which can be stored in the human bodyfor more than several days, in contrast to carbohydrates and proteins. The lipid content ofnormal humans (21% for men, 26% for women) enables them to survive energy starvation for2 to 3 months. Secondly, lipids are the major constituent of cell membranes in the form ofphospholipids, sphingolipids and cholesterol. Furthermore, lipids are the only source ofessential fatty acids, the precursors of eicosanoids such as prostaglandins, thromboxanes, andleucotrienes [7,8]. Finally, lipids are necessary for the solubilization and uptake of the fat-soluble vitamins A, D, E, and K.
OH
O
C1
CH3
18
OH
O
C1
CH3
18
9
OH
O
C1
9
12
CH3
18
OH
O
C1
9
12
15
CH3
18
Stearic acid Oleic acid Linoleic acid α−Linolenic acid
Figure 1.2 The structural formulas of some C18 fatty acids; stearic acid, oleic acid, linoleic acid, α-linolenic acid.
Lipids are particularly needed during periods of growth and development. In light ofthe important physiological roles of lipids, an efficient high-capacity absorption mechanism is

Chapter 1
10
required. Impaired lipid absorption has been associated with physical complications, such asdiarrhea, retarded growth, and essential fatty acid deficiency. The aim of this thesis is to obtainmechanistic information on the various pathophysiological processes involved in fatmalabsorption, with the purpose to increase diagnostic and eventually therapeutic possibilitiesin patients with fat malabsorption. The mechanisms by which lipids are taken up will bediscussed in paragraph 1.2, and subsequently lipid malabsorption with special emphasis on thedisease cystic fibrosis will be discussed in paragraph 1.3. The distinct methods to diagnose andquantify lipid malabsorption will be discussed in paragraph 1.4.
1.2 Intestinal absorption and digestion of dietary lipids
In order for intestinal lipid absorption to take place, lipids must undergo a number of physico-chemical changes to enable transport from the intestinal lumen to the plasma compartment.This is achieved by both mechanical and chemical means [5]. The overall process of intestinallipid absorption and digestion can be classified as a chain of events, including:1. Emulsification; the dispersion of bulk fat globules into finely divided emulsion particle.2. Lipolysis; the enzymatic hydrolysis of fatty acid esters at the emulsion-water interface.3. Micellar solubilization; the desorption and dispersion of insoluble lipid products into an
absorbable form.4. Membrane translocation; the transport of a lipid from the intestinal lumen across the
membrane of the intestinal mucosa cell.5. Intracellular events; this intracellular phase of lipid absorption involves re-esterification of
fatty acids and monoacylglycerols into triacylglycerols, the packaging of the lipids intochylomicrons and secretion of these chylomicrons at the basolateral side of the enterocyte[1,2,5,9-13]. The overall process of lipid absorption is shown in Figure 1.3.
1. Emulsification
2. Lipolysis
3. Solubilization
4. Translocation
5. Intracellularevents Lymphe
+
+
Figure 1.3 Schematic of intestinal lipid absorption.

General introduction
11
EmulsificationProcessing of lipids starts in the mouth with emulsification. The purpose of emulsification is toincrease the surface area of the lipid droplets, thereby increasing the area on which thedigestive enzymes can act effectively. Chewing breaks down large pieces of fat into smallersizes. Following ingestion, food enters the stomach, the major site for emulsification of dietarylipids. Muscle contraction of the stomach - particularly peristalsis against a closed pylorus andthe squirting of lipid through a partially opened pyloric canal - produces the shear forcessufficient for emulsification [1]. These peristaltic movements further grind the smaller piecesof lipid into a fine emulsion [14], which together with other emulsified foodstuffs is referred toas chyme. In addition to emulsifying food, the grinding action of the antrum mixes food withvarious digestive enzymes derived from the mouth and the stomach. Similarly, intestinalperistalsis continuously mixes luminal contents with digestive enzymes to ensure completedigestion [5].
LipolysisThe main objective of lipolysis is to convert triacylglycerols, which are virtually insoluble inthe aqueous phase of the gastrointestinal tract, into other forms of lipid with an increasedability to interact with water. Enzymatic hydrolysis of dietary triacylglycerols in humansbeyond the breast feeding period is mainly catalyzed by preduodenal and pancreatic lipases,and therefore in this thesis only these enzymes will be discussed. Preduodenal lipase is secretedfrom different tissues depending on species [15-17] and has therefore been assigned differentnames, e.g. gastric lipase [18], pharyngeal lipase [19], and lingual lipase [20]. In humans thelipase is entirely a product of the chief cells of the gastric mucosa, and is therefore calledgastric lipase [2]. Regardless of species or tissue origin, the preduodenal lipases sharemolecular and kinetic properties and it is assumed that they all have a common physiologicalfunction, i.e. to initiate triacylglycerol digestion in the stomach. Gastric lipase preferentiallyacts on the sn-3 position of the triacylglycerol molecule to release diacylglycerols and freefatty acids [21,22]. The level of hydrolysis in the stomach by gastric lipase in humans underphysiological conditions accounts for approximately 10 to 30% of total lipid ingested [23].
The lipid emulsion enters the small intestine as fine lipid droplets less than 0.5 µm indiameter [1]. Pancreatic colipase-dependent lipase, secreted by pancreatic acinar cells,completes dietary lipid digestion in the proximal small intestine [24]. Pancreatic lipase actsmainly on the sn-1 and sn-3 positions of the triacylglycerol molecule to release 2-monoacylglycerol and free fatty acids [21,22]. Pancreatic lipase is one of the most studied andbest characterized lipases, and is considered to be responsible for quantitative digestion of alltriacylglycerols in the adult. In healthy human adults, the level of enzyme secreted into theintestinal lumen was calculated to be in 1000-fold excess of what would be required forhydrolysis of daily lipid intake [2]. Pancreatic lipase is clearly essential for efficient dietary lipiddigestion as evidenced by the steatorrhoea present in patients with for example cystic fibrosis[25] or congenital pancreatic lipase deficiency [26,27].
The recent description of the primary and tertiary structures of pancreatic lipase hasprovided insight into the molecular detail of pancreatic lipase-catalyzed lipolysis [28,29].Pancreatic lipase is an enzyme with a marked substrate preference for triacylglycerols.

Chapter 1
12
Enzymatic hydrolysis of lipids can occur only on the surface of a lipid droplet, that is, at theinterface between the lipid droplet and the surrounding aqueous solution. When an oil-waterinterface is encountered, pancreatic lipase activity increases markedly, a property termedinterfacial activation [29]. Although pancreatic lipase is secreted into the duodenum along withbile salts, the enzyme is inhibited by physiological concentrations of bile salts and is dependenton another pancreatic protein, colipase, for activity in the presence of bile salts [30].
The action of both gastric and pancreatic lipase is facilitated for medium-chaintriacylglycerols when compared with long-chain triacylglycerols due to their more expandedsurface films in water [1]. Consequently, medium-chain triacylglycerols are hydrolyzed bothfaster and more completely than long-chain triacylglycerols [6]. In the case of mixedtriacylglycerols the medium-chain fatty acids are liberated preferentially [6].
Micellar solubilizationBile is secreted by the liver and enters the intestine through the biliary tract. One of theimportant properties of bile is its ability to increase the solubility of lipolytic products (i.e. 2-monoacylglycerols and free fatty acids) in the aqueous intestinal lumen by the formation ofmixed micelles. Micelles are structures in which the polar group projects into the aqueousphase while the nonpolar hydrocarbon chain forms the center. This macromolecular structurehas a high water solubility. Micellar solubilization increases the aqueous concentration of fattyacids and monoacylglycerols 100 to 1000 times [9].
Much of our current understanding on the uptake of dietary lipids was derived fromthe work of Hofmann and Borgström [31,32] and subsequent studies [33-35], who describethe importance of micellar solubilization of lipids in the uptake of lipid digestion products byenterocytes. To understand the importance of micellar solubilization, it is important to discussthe unstirred water layer, a concept introduced by Westergaard and Dietschy [36] (Figure1.4). According to this concept, the brush border membrane of the enterocytes is separatedfrom the bulk fluid phase in the intestinal lumen by an unstirred water layer, which is relativelyimpermeable for the lipolytic products, especially the long-chain fatty acids. The rate of long-chain fatty acid monomer diffusion in water is greater than that of aggregates of mixedmicelles [12]. The increased concentration of fatty acids by micellar solubilization overcomesthe slower diffusion rate, so that the net effect of micelle formation is an increase in thetransfer of lipolytic products across the unstirred water layer [36]. Thus, mixed micelles wouldact as lipid shuttles to overcome the unstirred water layer [36].
The validity of this concept was later challenged by Carey and his associates, whodiscovered the coexistence of unilamellar liposomes with bile salt-lipid mixed micelles in thesmall intestine [37]. They proposed that when the bile salt concentration in the lumen exceedsthe critical micellar concentration, the lipids in the intestinal lumen will be incorporated intomixed micelles [1]. When the amount of lipids in the aqueous phase increases further and theamount of bile salts does not increase, this eventually results in the formation of liquidcrystalline vesicles (liposomes) [1]. However, so far the relative roles of the micelle and theliquid crystalline vesicle in the uptake of fatty acids and monoacylglycerols have not beenresolved [9].

General introduction
13
The lipolytic products of medium-chain triacylglycerols are absorbed faster than thoseof long-chain triacylglycerols. As lipolysis of medium-chain triacylglycerols is more completethan that of long-chain triacylglycerols, the medium-chain triacylglycerols (unlike long-chaintriacylglycerols) are absorbed mainly as free fatty acids, and only rarely as mono- anddiacylglycerols [6]. Because of the increased water solubility of medium-chain fatty acids,absorption of medium-chain fatty acids is not dependent on micellar solubilization [38-40].Thus, for long-chain fatty acids passage across the unstirred water layer is rate limiting,whereas passage of medium-chain fatty acids is only limited by the brush border membrane[41].
Bulk solution inintestinal lumen
Diffusion barrieroverlying microvilli
Cytosoliccompartment of
intestinal cell
1
2
Figure 1.4 Diagrammatic representation of the effect of bile salt micelles (or vesicles) in overcoming the diffusion barrier
resistance offered by the unstirred water layer. In the absence of bile acids, individual lipid molecules must diffuse across the
barriers overlying the microvillus border of the intestinal epithelial cells (arrow 1). Hence, uptake of these molecules is largely
diffusion limited. In the presence of bile acids (arrow 2), large amounts of these lipid molecules are delivered directly to the
aqueous-membrane interface so that the rate of uptake is greatly enhanced [9].
TranslocationThe mechanism by which lipids are taken up by the enterocyte across its apical membraneremains unresolved. Previously, it has been accepted that the uptake of free fatty acids andmonoacylglycerols by the enterocytes is a passive diffusion process [10,42]. Recently, thepossibility has been raised that some lipids may be taken up by enterocytes by carrier-mediatedprocesses [43-47]. It was shown that fatty acid binding proteins and/or fatty acid translocase,associated with the brush border membrane, seem to play a role in the uptake of fatty acids byenterocytes [43,47]. However, the exact role of the protein has not been resolved yet, and theissue of whether fatty acids are taken up by passive diffusion or by a carrier-mediated processneeds further investigation.

Chapter 1
14
Intracellular eventsIn the intestinal cell the various absorbed lipids migrate from the site of absorption to theendoplasmic reticulum. It has been suggested that the migration of the lipids is mediated viafatty-acid-binding proteins (FABP) located in the intestine (intestinal FABP and liver FABP)[48]. Re-esterification of free fatty acids and monoacylglycerols into triacylglycerols takesplace at the cytoplasmic surface of the endoplasmic reticulum [49] mainly via themonoacylglycerol pathway [50,51]. This involves reacylation to diacylglycerols andtriacylglycerols by monoacylglycerol-acyltransferase and diacylglycerol-acyltransferase,respectively [9]. The other route of triacylglycerol synthesis, the alpha-glycerophosphatepathway, involves conversion of glycerol-3-phosphate via phosphatidic acid to diacylglycerolsand, subsequently, to triacylglycerols by various enzymes [9]. Under physiologicalcircumstances, the monoacylglycerol pathway predominates relative to the alpha-glycerophosphate pathway [9].
Triacylglycerols are then transferred by a transfer protein to the inside of theendoplasmic reticulum [52] and packaged into lipoprotein particles called chylomicrons.Chylomicrons are made exclusively by the small intestine, and consist mainly of phospholipids,dietary triacylglycerols and apolipoproteins apo A-I, apo A-IV, and apo B-48 [9]. Data fromboth animals and humans indicate that the fatty acid composition of the triacylglycerol ofchylomicrons closely resembles the dietary lipid fed [53,54]. The chylomicrons are releasedinto the bloodstream via the lymph system for delivery of triacylglycerols to the tissues.
1.3 Fate of lipids in the colon
The digested nutrients that enter the colon encounter a large population of bacteria capable ofa wide range of metabolic activities. For example, the colonic flora play a major part in thefermentation of carbohydrates to produce short-chain fatty acids. Although these short-chainfatty acids may play a role in the prevention of colonic inflammation, further discussion isbeyond the scope of this thesis and the interested reader is referred to [55,56].
The small amounts of long-chain fatty acids escaping absorption and entering thelarge bowel have been regarded as of trivial biological significance. However, there is evidencethat colonic bacteria can metabolize dietary fats: colonic bacteria secrete lipase enzymes [57],they have active transport mechanisms for medium- and long-chain fatty acids, and are capableof oxidation, desaturation and hydroxylation of fatty acids [58]. In the past several studiesshowed that the unabsorbed fraction of lipid may have important effects on bacterialmetabolism of the colon [59,60], and may even play a role in the etiology of colonic cancer[61].
Obviously, the daily input of lipids into the colon increases considerably in the case ofvarious lipid malabsorption syndromes. However, the role of large amounts of lipids in thecolon has only been partially resolved and further research is necessary [58,62].

General introduction
15
1.4 Lipid malabsorption
Fat malabsorption is characterized by increased fecal excretion of mostly dietary lipids.Increased fat content of the feces is also known as steatorrhoea, which may be a first symptomof underlying diseases affecting fat absorption. It has been convenient to divide fatmalabsorption into those disorders with an impaired digestion of triacylglycerols from thosedisorders with impaired intestinal uptake, which includes impaired mixed micelle formationand translocation of fatty acids over the intestinal mucosa.
Impaired lipolysisUnder physiological conditions, pancreatic lipase is present in pancreatic juice in abundance.Its high concentration in pancreatic secretions and its high catalytic efficiency ensure theefficient digestion of dietary lipid. However, impaired lipolysis of dietary triacylglycerols,caused by a lack of sufficient pancreatic lipases, is a well-recognized cause of steatorrhoea.Pancreatic lipase deficiency can either be due to the (relative) absence of the enzymes involvedor due to inactivity of these enzymes [63]. Steatorrhoea in lipase deficiency is usually notsevere, unless lipase concentration in the upper intestinal tract is less than 10% of normal.Such impaired lipolysis may be secondary to cystic fibrosis, chronic pancreatitis, pancreaticresection, or pancreatic carcinoma [38,63].
The most effective treatment of lipase deficiency is to restore lipase activity. This isaccomplished either by eliminating causes of lipase inactivation, such as correcting gastric acidhypersecretion, or to supply exogenous lipase [38,64].
Impaired uptake of long-chain fatty acidsPatients with a biliary fistula, biliary obstruction, chronic liver disease, or an interruption of thebile salt enterohepatic circulation by ileal resection or disease have a decreased bile saltsecretion rate. With less bile salt present in the intestinal lumen, fewer mixed micelles form,impairing solubilization of ingested lipids. However, total absence of bile in the intestine doesnot completely inhibit fat absorption. Even up to 80% of dietary lipids were found to beabsorbed in a study in adults with biliary fistula [33]. An explanation could be the observationof liquid crystalline vesicles by Carey et al. [1]. They suggested that when the amount of fat inthe aqueous intestinal phase is high compared with the amount of bile, liquid crystallinevesicles are formed. These vesicles may play an important role in the uptake of fats byenterocytes in disease states [33]. The finding of liquid crystalline vesicles may have importantpathophysiological implications. Because patients with low intraluminal bile saltconcentrations or with bile fistulae can have reasonably good lipid absorption, it was proposedthat the liquid crystalline vesicles may play an important role in the uptake of fatty acids andmonoacylglycerols by enterocytes in these disease states [1]. In addition, in the absence or atlow concentrations of bile salts, the absorption of fatty acids occurs to a relatively lower andslower extent [65]. Brand and Morgan [66] showed that fat absorption occurs largely from theproximal small intestine in control rats, whereas, in the absence of bile distal small intestine isalso involved. Presumably, the absorptive reserve of the distal small intestine is called upon in

Chapter 1
16
the case of bile diversion and much of the fat which failed to enter the proximal intestinalmucosa is absorbed more distally [67].
Therapy is directed toward either restoring the enterohepatic circulation of bile saltsor by substituting medium-chain triacylglycerols in the diet [38]. Because of the increasedwater solubility of medium-chain fatty acids, bile salts are not as necessary for efficientabsorption of medium-chain fatty acids.
Cystic fibrosisA frequently encountered genetic disorder associated with fat malabsorption is cystic fibrosis[68,69]. The pathophysiology of fat malabsorption in cystic fibrosis patients involves bothpancreatic insufficiency and deficient intestinal uptake of long chain fatty acids. Cystic fibrosisis an autosomal recessive disorder in which defective transepithelial chloride transport resultsin the production of mucus with increased viscosity in various organs. Among the organscommonly affected, the lungs and the pancreas frequently are involved in serious symptoms atyoung age [70]. The basic defect is the cystic fibrosis transmembrane regulator (CFTR), aprotein responsible for chloride ion transport. Both pancreatic insufficiency and high energyexpenditure due to increased respiratory work are thought to contribute to the frequentlyobserved poor nutritional status of these patients [68,71]. The positive correlation between agood nutritional status and long-term survival or well-being of cystic fibrosis patients is welldocumented [72]. This observation has led to increased attention for optimization of nutrientintake and absorption in cystic fibrosis patients [68]. Recommendations for treatment of cysticfibrosis patients include consumption of 120-150% of the recommended daily allowance ofenergy for healthy individuals [73], with a normal to high lipid (40 energy %) intake to offsetincreased energy requirements [74].
Despite recent improvements in the pharmacokinetics of the supplementarypancreatic enzymes, many patients continue to experience a certain degree of steatorrhoea[75-77], with lipid absorption reaching 80 to 90% of their dietary lipid intake. It has not beenelucidated if the remaining lipid malabsorption is due to an insufficient dosage of pancreaticenzyme replacement therapy. This possibility is not unlikely because a decreased pancreaticbicarbonate secretion may negatively affect enzyme activity by sustaining a low pH in theduodenum [64]. At a low duodenal pH, the release of the enzymes from the (micro)capsules isinhibited and the denaturation of the enzymes is stimulated [64,78]. However, it has beendemonstrated that increasing the pancreatic enzyme dosages does not completely correct lipidmalabsorption [79]. In addition, attempts to increase lipolysis by high-strength pancreaticenzyme supplements has led to the reported association with fibrosing colonopathy [80-82].
An alternative explanation for the continuing fat malabsorption in CF patients onpancreatic enzyme replacement therapy may involve inefficient intestinal uptake of fatty acids[75,83]. Impaired uptake in CF patients can be due to an altered bile composition, decreasedbile salt secretion by the liver, bile salt precipitation, a decreased bile salt pool size, and/or bilesalt inactivation at low intestinal pH [77,83-86]. Furthermore, small bowel mucosaldysfunction or alterations in the mucus layer may contribute to inefficient intestinal uptake oflong chain fatty acids in CF patients [68,87].

General introduction
17
Although it is known that the pathophysiology of fat malabsorption in cystic fibrosispatients involves both pancreatic insufficiency and deficient intestinal uptake of long chainfatty acids, the relative contribution of these two processes frequently remains unclear. Insightinto the contribution of either of these processes would benefit cystic fibrosis patients,however, it is difficult to obtain mechanistic information in patients. In an attempt to furtherelucidate the pathophysiology of cystic fibrosis, several mouse models of cystic fibrosis weredeveloped [88,89].
1.5 Methods to measure lipid malabsorption
The efficiency of intestinal lipid absorption in patients is routinely determined by means of alipid balance, requiring detailed analysis of daily lipid intake and the complete recovery offeces for 72 h. However, in the case of lipid malabsorption, this method does not discriminatebetween the potential causes, such as impaired intestinal lipolysis or disturbed micellarsolubilization of long-chain fatty acids. Since different therapies are selected for the differentcauses, it is important to know the etiology behind the fat malabsorption. In the developmentof novel diagnostic strategies, stable isotope techniques have been introduced. In this chapterthe several aspects regarding the fecal fat balance will be discussed first. Thereafter, attentionwill be paid to stable isotope tests measuring impaired lipolysis and/or disturbed uptake oflong-chain fatty acids.
Fecal fat balanceThe conventional method by which lipid absorption is evaluated is the 3-day fecal fat balance.Estimation of a fat balance is carried out as follows: the patient is kept on a diet containing aknown amount of lipid and dietary intake is recorded for a period of 3 days. The fecesexcreted during the same period is collected accurately, and lipid is determined quantitatively.Since its first description in 1949, the titrimetric procedure of Van de Kamer [90] has beenused as a reference method for the measurement of lipid in the feces. The percentage of totaldietary lipid absorption is calculated from the amount of lipid ingested and the amount of lipidexcreted via the feces by the following equation:
Percentage of total fat absorptionFat intake (g day ) Fecal fat output (g day )
Fat intake (g day )
-1 -1
-1=−
×100%
The 3-day fecal fat balance in Western adult humans shows that intake of majordietary lipids, principally triacylglycerols, constitutes approximately 100 g day-1. In addition,substantial amounts of endogenous lipids are delivered to the intestinal lumen from bile [91],desquamated cells [92], and dead bacteria [93,94]. Intestinal epithelial cells are being sloughedoff into the lumen continuously [92] and it can be estimated to amount to about 450 g of cellsper day of which 2 to 6 g are membrane lipids that are mostly digested and absorbed [1,91].
Although measurement of fecal lipid excretion during a standard lipid intake isgenerally considered to be the most accurate screening test for detecting lipid malabsorption,

Chapter 1
18
the test is not widely used because of its poor acceptability by patient, physician, and clinicalchemist. For the patient, the test involves the inconvenience of eating a defined diet and themechanical and esthetic problems of collecting, storing, and transporting stools. For thephysician, the test involves scientific uncertainty as to the completeness of the fecal collectionand also may involve storage and transport problems. For the clinical chemist, the test involvesthe storage of bulky specimens and the unpleasant task of sample homogenization andsampling [95]. Finally, in the case of lipid malabsorption, the lipid balance method does notdiscriminate between the underlying mechanisms, such as impaired intestinal lipolysis ordisturbed intestinal solubilization of long chain fatty acids. In order to investigate theunderlying mechanisms, stable isotope techniques have been introduced, which will bediscussed in the next paragraph.
Stable isotopesThe renaissance of interest in stable isotopes in the last ten years is based upon thedevelopment of new instrumentation, such as the availability of the quadruple massspectrometers interfaced with the gas chromatograph (GC/MS) and the development ofisotope ratio mass spectrometers (IRMS), which made possible the convenient use of selectiveion monitoring for the quantification of isotope enrichment. An increased awareness of thehealth hazards of radioactivity, as well as greater availability of stable isotopes, also stimulatedthe use of stable isotopes.
The most obvious advantage of stable isotopes is that they are nonradioactive andpresent little or no risk to human subjects and they are even suitable for the study of infants,children, and pregnant women [96]. Carbon 13 is a naturally occurring isotope present to theextent of approximately 1.1% of the major isotopic species, carbon 12 [97]. Since carbon 13naturally contributes 1.1% of the carbon pool, and since it has not been possible todemonstrate more than trivial in vitro isotopic effects on chemical reactions with carbon 13-labeled substrates [98,99], significant side effects in vivo are not expected from administrationof tracer doses of carbon 13.
Among the numerous applications of stable isotopes in physiology and medicine, theinvestigation of lipid absorption and metabolism poses considerable challenges because of thecomplexity of the subject, the multitude of influencing factors and the demanding analyticalrequirements [100]. Various labeled fatty acids and labeled triacylglycerols are available andcan be given orally. When a 13C-labeled fat is ingested, the substrate may be digested,absorbed and enters metabolic pathways leading to enrichment of bicarbonate, protein, lipidand carbohydrate within the body. Unabsorbed amounts of the 13C-labeled fat are excreted viathe feces. After absorption, the 13C-labeled fat enters the oxidative pathways and is excreted as13CO2 via the breath.
When stable isotopes are used to measure fat digestion and absorption, betweeningestion of the labeled fat and appearance of 13C in plasma and excretion of 13CO2 in thebreath, many factors can influence the outcome of the test and expression of the results, suchas gastric emptying rate, absorption rate, hepatic clearance etc. [101]. Choice of substrate andchoice of sampling compartment are the first factors in determining the sensitivity and

General introduction
19
specificity of the test. The rate-limiting step of interest in the handling of substrates by thebody determines the selection of a substrate.
13C-TRIOLEIN
Since triolein is a long-chain triacylglycerol, its efficient absorption depends upon theoverall process of fat absorption, thus, adequacy of lipolysis, bile salt solubilization andintact mucosal surface. Hence, this substrate is a sensitive indicator of steatorrhoea, but willnot distinguish between the underlying mechanisms [102-104]. It has been proposed thatthe triolein test is preferred when compared to the trioctanoin or the tripalmitin test for thescreening of total lipid malabsorption arising from a broad spectrum of gastrointestinaldisorders because of its higher sensitivity and specificity [95,105,106]. However, it hasbeen shown that the test has not the ability to predict the severity of malabsorption [107].So far 13C-triolein has only been used with collection of breath and analysis of 13CO2, but itsradioactive form, 14C-triolein, has also been used with measurements of postprandial serum[108,109].
13C-HIOLEIN
Naturally occurring hiolein provides a new tracer for lipid absorption studies. Hiolein is along chain triacylglycerols mixture obtained from algae which is uniformly labeled with 13Cand enriched by 98%. The major fatty acid composition of hiolein is oleic acid (51%),palmitic acid (17%) and linoleic acid (20%) [110]. Since hiolein consists mainly oftriacylglycerols, efficient absorption depends on the same processes as triolein, i.e. lipolysis,bile salts solubilization and intact mucosa. Patients with significantly impaired lipolysis, bilesalt deficiency, or mucosal disorders, excrete the substance in their stool, and havedecreased amounts of 13CO2 in their breath [110-113].
13C-TRIOCTANOIN
Trioctanoin is a medium-chain triacylglycerol. Although both medium and long-chaintriacylglycerols require lipolysis by gastric and pancreatic lipase, medium-chaintriacylglycerols, being water soluble, do not depend critically on the presence of bile saltsfor their digestion and absorption. The 13C-trioctanoin test thus focuses on lipase activityand a reduction in trioctanoin absorption reflects the level of lipolytic activity present in thepatients digestive tract. Digestion and absorption of trioctanoin have been assessed by13CO2 excretion via the breath [105,114-119]. The choice of a medium-chain triglyceridehas an additional advantage in that the lipolytic products are rapidly absorbed and oxidized[120], thus shortening the overall study period [114]. The 13C-trioctanoin test distinguishespancreatic from non-pancreatic causes of steatorrhoea, and it has been applied formeasurements of lipid maldigestion in adults [119] and in children with cystic fibrosis[115,116], and for measurements of lipid utilization in preterm and full-term neonates[116,121]. A disadvantage of the 13C-trioctanoin breath test is that the rate of lipolysis isfacilitated for medium-chain triacylglycerols when compared with long-chaintriacylglycerols [1,6]. Hence, the test does not exactly reflect lipolytic rate of dietary fats,because they mainly consist of long-chain triacylglycerols.

Chapter 1
20
13C-MIXED TRIGLYCERIDE
A substrate that has the advantages of the 13C-trioctanoin breath test (short study period)and avoids the disadvantages (facilitated lipolysis for medium-chain triacylglycerols) is theso-called 13C-mixed triglyceride breath test [122,123]. The mixed triglyceride used is 1,3-distearoyl, 2[carboxyl-13C]octanoyl glycerol. This molecule contains a 13C-labeled mediumchain fatty acid (octanoic acid) at the sn-2 position, and long-chain fatty acids (stearic acid)at the sn-1 and sn-3 positions of the glycerol backbone of the triacylglycerol [123]. The twostearoyl chains have to be hydrolyzed by lipolytic enzymes in the intestine before 13C-octanoate can be absorbed, thereby avoiding the disadvantage of the 13C-octanoin substratewhich contains only medium-chain fatty acids. Thus, the principle of the mixed triglyceridebreath test is based on lipolysis-dependent 13CO2 excretion via the breath. The applicabilityof the mixed triglyceride breath test has been demonstrated in healthy adults [124,125],pancreatic insufficiency patients [123,126], and preliminary data on the potentialapplicability in children are available [127]. The mixed triglyceride breath test has asensitivity of 89% and a specificity of 81% compared with direct measures of lipase inpatients with pancreatic and non-pancreatic causes of steatorrhoea [123].
CHOLESTERYL-[1-13C]OCTANOATE
The utilization of cholesteryl-[1-13C]octanoate is another attractive substrate for measuringpancreatic exocrine insufficiency. This substrate differs little from the cholesteryl estersnaturally present in food. It undergoes hydrolysis by pancreatic cholesteryl esterase, and thelabeled octanoate molecule is rapidly absorbed and oxidized [114,120]. Because cholesteryloctanoate is not hydrolyzed by gastric lipase it may be used to assess pancreatic exocrinefunction alone [128]. However, pancreatic carboxyl ester lipase activity requires thepresence of bile salts and therefore, this test will not only measure pancreatic function butalso solubilization by bile [128]. The test has been successfully used to diagnose exocrinepancreatic insufficiency [129] and to monitor pancreatic enzyme replacement therapy inpatients with pancreatic insufficiency [128,130,131].
[1-13C]PALMITIC ACID
Efficient absorption of long-chain fatty acids, e.g. palmitic acid, is not dependent onlipolysis since free fatty acids are already hydrolyzed substrates. Thus, application of thistest to patients with gastrointestinal complaints would identify individuals with inadequatesolubilization of long chain fatty acids or intestinal mucosa disease [105,132,133]. It isdifficult to discriminate between the two processes, which may be due to the fact thatimpaired solubilization is rarely an isolated event [105]. The test has been performed inhealthy controls [134,135] and in patients with gastrointestinal diseases [105,136]. Theadvantage of using palmitic acid as a substrate instead of other fatty acids is that palmiticacid is a saturated fatty acid and both solubilization and translocation across the intestinalmucosa of saturated fatty acids are more difficult when compared to unsaturated fattyacids. In addition, palmitic acid is the most predominant fatty acid in the Western diet, andtherefore experiments mimic dietary fat absorption as much as possible.

General introduction
21
[U-13C]LINOLEIC ACID
[U-13C]linoleic acid is a long-chain fatty acid and, thus depends on the same absorptiveprocesses as [1-13C]palmitic acid does, i.e. adequate solubilization by bile components andintact mucosa of the intestine. In addition, linoleic acid is an essential fatty acid andtherefore may be used for studies with respect to the essential fatty acid status.
1.6 Scope of this thesis
As discussed before, the process of lipid absorption can be viewed as a chain of eventsoccurring after lipid ingestion, including emulgation, lipolysis, solubilization, uptake in theenterocyte, and chylomicron assembly. Under physiological conditions, the efficacy of lipidabsorption ranges from 96 to 98% [1,2]. Until now, most attention has been paid towards theefficacy of the overall process of lipid absorption, yet, insight into the individual mechanismscausing fat malabsorption has remained rather incomplete. A detailed insight into theunderlying mechanisms would enable not only improvements in diagnostic methodologies, butalso treatment in individual patients by modulating diet therapy, pancreatic enzymereplacement therapy and supplementation of antacids and/or bile salts. Thereby, it is areasonable expectation that the prognosis of (pediatric) patients with impaired lipid absorptioncan be improved, given the positive correlation between a good nutritional status and long-term survival or well-being [72-74].
The aim of the thesis is to obtain mechanistic information on the various processesinvolved in fat malabsorption, with the purpose to increase diagnostic and eventuallytherapeutic possibilities in patients with fat malabsorption. The approach to achieve this aiminvolves studies in experimental animals, in human volunteers and in patients. The studies werechosen to investigate in detail the two most frequently occurring pathophysiological processesinvolved in human fat malabsorption, namely, impaired lipolysis and disordered bile formation,as well as the most frequently encountered disease in children associated with fatmalabsorption, cystic fibrosis. Since our purpose was to increase diagnostic and eventuallytherapeutic possibilities in patients, we applied stable isotopes in our experiments, allowingphysiological studies in humans in a non-harmful way. The applicability of stable isotopelabeled lipids for quantitative studies on lipid absorption has only been investigated to a verylimited extent.
A non-invasive test that has been described to characterize pancreatic insufficiency ina functional way is the 13C-MTG breath test [123]. However, widely variable results have beenobtained in children, healthy adults, and in cystic fibrosis patients with and without pancreaticenzyme replacement therapy [125,126]. The origin of this variability has not been elucidated.In fact, a quantitative relationship between the extent of fat malabsorption due to impairedlipolysis and the corresponding result of the 13C-MTG breath test has never been demonstratedin humans or in defined animal models. Therefore, in this thesis the efficiency and repeatabilityof the 13C-MTG breath test were investigated in rats treated with the lipase inhibitor orlistat(chapter 2) and in healthy adults (chapter 3), respectively.

Chapter 1
22
Few attempts to develop a specific test for the detection of impaired intestinal uptakeof long chain fatty acids have been reported [105]. Intestinal uptake involves solubilization oflipolytic products by the formation of mixed micelles composed of bile components andlipolytic products, followed by the translocation of the lipolytic products across the intestinalepithelium [2,9,34,35]. Potential substrates for the detection of impaired intestinal uptake are13C-labeled long chain fatty acids. In this thesis, the potency of 13C-labeled palmitic acid todetect impaired intestinal uptake was determined in rats with long-term diversion of the biliarytract (chapter 4). In addition, the sensitivity of the 13C-labeled palmitic acid test wasinvestigated in healthy adults supplemented with calcium in order to achieve mild fatmalabsorption due to decreased amounts of bile in the intestine (chapter 5).
A relatively frequently encountered disorder in Caucasian populations associated withfat malabsorption is cystic fibrosis. Although it is known that the pathophysiology of fatmalabsorption in cystic fibrosis patients involves both pancreatic insufficiency [68,69] anddeficient intestinal uptake of long chain fatty acids [75,83], the relative contribution of thesetwo processes frequently remains unclear. In order to obtain more insight into the impairedprocesses of fat malabsorption in cystic fibrosis we performed a study in pediatric cysticfibrosis patients treated with their usual pancreatic enzyme replacement therapy (chapter 6).The substrates 13C-MTG and uniformly labeled 13C-linoleic acid were both applied todetermine whether the rate-limiting step behind their remaining fat malabsorption was eitherimpaired lipolysis or impaired intestinal uptake of long chain fatty acids, respectively. Based onthe results of this study, we further explored the mechanisms involved in deficient intestinaluptake of long chain fatty acids in further detail in two recently generated cystic fibrosis mousemodels (chapter 7). 1. Mice with the ∆F508 mutation in the cftr gene, ∆F508/∆F508 mice.The ∆F508 mutation is the most frequently observed mutation in cystic fibrosis patients. 2.Mice with complete inactivation of the cftr gene, cftr -/- mice [137,138].
References
1. Carey MC, Small DM, Bliss CM. Lipid digestion and absorption. Ann Rev Physiol1983;45:651-677.
2. Carey MC, Hernell O. Digestion and absorption of fat. Sem Gastrointest Dis1992;3:189-208.
3. Van der Steege G, Muskiet FAJ, Martini IA, Hutter NH, Boersma ER. Simultaneousquantification of total medium- and long-chain fatty acids in human milk by capillary gaschromatography with split injection. J Chromatogr 1987;415:1-11.
4. Stryer L. Biochemistry. 2nd Ed. San Francisco, CA: Freeman, 1981:383-406.5. Shiau Y-F. Lipid digestion and absorption. In: Johnson LR, ed. Physiology of the
Gastrointestinal Tract. 2nd Ed. New York: Raven Press, 1987:1527-1556.6. Bach AC, Babayan VK. Medium-chain triglycerides: an update. Am J Clin Nutr
1982;36:950-962.7. Panossian A, Hamberg M, Samuelsson B. On the mechanism of biosynthesis of
leukotrienes and related compunds. FEBS Lett 1982;150:511-513.

General introduction
23
8. Samuelsson B, Dahlen SE, Lindgren JA, Rouzer CA, Serhan CN. Leukotrienes andlipoxins: structures, biosynthesis, and biological effects. Science 1987;237:1171-1176.
9. Tso P. Intestinal lipid absorption. In: Johnson LR, ed. Physiology of the gastrointestinaltract. 3rd Ed. New York: Raven Press, 1994:1867-1907.
10. Thomson ABR, Keelan M, Garg ML, Clandinin MT. Intestinal aspects of lipidabsorption: in review. Can J Physiol Pharmacol 1989;67:179-191.
11. Thomson ABR, Schoeller C, Keelan M, Smith L, Clandinin MT. Lipid absorption:passing through the unstirred layers, brush-border membrane, and beyond. Can J PhysiolPharmacol 1993;71:531-555.
12. Shiau Y-F. Mechanisms of intestinal fat absorption. Am J Physiol 1981;240:G1-G9.13. Minich DM, Vonk RJ, Verkade HJ. Intestinal absorption of essential fatty acids under
physiological and essential fatty acid-deficient conditions. J Lipid Res 1997;38:1709-1721.
14. Kelly KA. Motility of the stomach and gastroduodenal junction. In: Johnson LR, ed.Physiology of the gastrointestinal tract. New York: Raven Press, 1981:393-410.
15. Roy CC, Roulet M, Lefebvre D, Chartrand L, Lepage G, Fournier LA. The role ofgastric lipolysis on fat absorption and bile acid metabolism in the rat. Lipids1979;14:811-815.
16. Hamosh M, Klaeveman HL, Wolf RO, Scow RO. Pharyngeal lipase and digestion ofdietary triglyceride in man. J Clin Invest 1975;55:908-913.
17. Moreau H, Gargouri Y, Lecat D, Junien JL, Verger R. Screening of preduodenal lipasesin several mammals. Biochim Biophys Acta 1988;959:247-252.
18. Bernbäck S, Bläckberg L. Human gastric lipase; The N-terminal tetrapeptide is essentialfor lipid binding and lipase activity. Eur J Biochem 1989;1825:495-499.
19. Bernbäck S, Hernell O, Bläckberg L. Purification and molecular characterization ofbovine pregastric lipase. Eur J Biochem 1985;148:233-238.
20. Hamosh M, Scow RO. Lingual lipase and its role in the digestion of dietary lipid. J ClinInvest 1973;52:88-95.
21. Staggers JE, Fernando-Warnakulasuriya GJP, Wells MA. Studies on fat digestion,absorption, and transport in the suckling rat. II. Triacylglycerol molecular species,stereo-specific analysis and specificity of hydrolysis by lingual lipase. J Lipid Res1981;22:675-679.
22. Paltauf F, Esfandi F, Holasek A. Stereo-specificity of lipases. Enzymatic hydrolysis ofenantiomeric alkyl diglyceride by lipoprotein lipase, lingual lipase and pancreatic lipase.FEBS Lett 1974;40:119-123.
23. Liao TH, Hamosh P, Hamosh M. Fat digestion by lingual lipase: mechanism of lipolysisin the stomach and upper small intestine. Pediatr Res 1984;18:402-409.
24. Christensen MS, Müllertz A, Hoy C. Absorption of triglycerides with defined or randomstructure by rats with biliary and pancreatic diversion. Lipids 1995;30:521-526.
25. Fernandes J, Van de Kamer JH, Weijers HA. Differences in absorption of the variousfatty acids studied in children with steatorrhea. J Clin Invest 1962;41:488-494.
26. Figarella C, De Caro A, Leupoid D, Poley JR. Congenital pancreatic lipase deficiency.Pediatrics 1980;96:412-416.

Chapter 1
24
27. Ghishan FK, Moran JR, Durie PR, Greene HL. Isolated congenital lipase-colipasedeficiency. Gastroenterology 1984;86:1580-1582.
28. Lowe ME. Pancreatic triglyceride lipase and colipase: insights into dietary fat digestion.Gastroenterology 1994;107:1524-1536.
29. Lowe ME. Molecular mechanisms of rat and human pancreatic triglyceride lipases. JNutr 1997;127:549-557.
30. Labourdenne S, Brass O, Ivanova M, Cagna A, Verger R. Effects of colipase and bilesalts on the catalytic activity of human pancreatic lipase. A study using the oil droptensiometer. Biochemistry 1997;36:3423-3429.
31. Hofmann AF, Borgström B. The intraluminal phase of fat digestion in man: the lipidcontent of the micellar and oil phases on intestinal content obtained during fat digestionand absorption. J Clin Invest 1964;43:247-257.
32. Hofmann AF, Borgström B. Physico-chemical state of lipid in intestinal contents duringtheir digestion and absorption. Fed Proc 1962;21:43-50.
33. Porter HP, Saunders DR, Tytgat G, Brunser O, Rubin CE. Fat absorption in bile fistulaman; A morphological and biochemical study. Gastroenterology 1971;60:1008-1019.
34. Staggers JE, Hernell O, Stafford RJ, Carey MC. Physical-chemical behavior of dietaryand biliary lipids during intestinal digestion and absorption. 1. Phase behavior andaggregation states of model lipid systems patterned after aqueous duodenal contents ofhealthy adult human beings. Biochemistry 1990;29:2028-2040.
35. Hernell O, Staggers JE, Carey MC. Physical-chemical behavior of dietary and biliarylipids during intestinal digestion and absorption. 2. Phase analysis and aggregation statesof luminal lipids during duodenal fat digestion in healthy adult human beings.Biochemistry 1990;29:2041-2056.
36. Westergaard H, Dietschy JM. The mechanism whereby bile acid micelles increase therate of fatty acid and cholesterol uptake into the intestinal mucosal cell. J Clin Invest1976;58:97-108.
37. Patton JS, Carey MC. Watching fat digestion. Science 1979;204:145-148.38. Bliss CM. Fat absorption and malabsorption. Arch Intern Med 1981;141:1213-1315.39. Greenberger NJ, Rodgers JB, Isselbacher KJ. Absorption of medium and long chain
triglycerides: factors influencing their hydrolysis and transport. J Clin Invest1966;45:217-227.
40. Mabayo RT, Furuse M, Murai A, Okumura J. Cholestyramine alters the lipid and energymetabolism of chicks fed dietary medium- or long-chain triacylglycerol. Lipids1995;30:839-845.
41. Westergaard H, Dietschy JM. Delineation of the dimension and permeabilitycharacteristics of the two major diffusion barriers to passive mucosal uptake in the rabbitintestine. J Clin Invest 1974;174:718-732.
42. Ling KY, Lee HY, Hollander D. Mechanisms of linoleic acid uptake by rabbit smallintestinal brush border membrane vesicles. Lipids 1989;24:51-55.
43. Stremmel W. Uptake of fatty acids by jejunal mucosal cells is mediated by a fatty acidbinding membrane protein. J Clin Invest 1988;82:2001-2010.

General introduction
25
44. Chow SL, Hollander D. A dual, concentration-dependent absorption mechanism oflinoleic acid by rat jejunum in vitro. J Lipid Res 1979;20:349-356.
45. Hollander D, Chow SL, Dadufalza VD. Intestinal absorption of free oleic acid in theunanesthetized rat: evidence for a saturable component? Can J Physiol Pharmacol1984;62:1136-1140.
46. Chow SL, Hollander D. Linoleic acid absorption in the unanesthetized rat: mechanism oftransport and influence of luminal factors on absorption. Lipids 1979;14:378-385.
47. Diede HE, Rodilla-Sala E, Gunawan J, Manns M, Stremmel W. Identification andcharacterization of a monoclonal antibody to the membrane fatty acid binding protein.Biochim Biophys Acta 1992;1125:13-20.
48. Ockner RK, Manning JA. Fatty acid binding protein in small intestine. Identification,isolation and evidence for its role in cellular fatty acid transport. J Clin Invest1974;54:326-338.
49. Bell RM, Ballas LM, Coleman RA. Lipid topogenesis. J Lipid Res 1981;22:391-403.50. Lehner R, Kuksis A. Biosynthesis of triacylglycerols. Prog Lipid Res 1996;35:169-201.51. Johnston JM. Triglyceride biosynthesis in the intestinal mucosa. In: Rommel K, Goebel
H, Böhmer R, eds. Lipid absorption: Biochemical and clinical aspects. Lancaster: MTPPress, 1976:85-94.
52. Wetterau JR, Aggerbeck LP, Bouma M, Eisenberg C, Munck A, Hermier M, Schmitz J,Gay G, Rader DJ, Gregg RE. Absence of microsomal triglyceride transfer protein inindividuals with abetalipoproteinemia. Science 1992;258:999-1001.
53. Blomstrand R, Dahlbäck O. The fatty acid composition of human thoracic duct lymphlipids. J Clin Invest 1960;39:1185-1191.
54. Zilversmit DB. The composition and structure of lymph chylomicrons in dog, rat, andman. J Clin Invest 1965;44:1610-1622.
55. Cummings JH. Production and metabolism of short-chain fatty acids in humans. In:Roche AF, ed. Short-chain fatty acids: Metabolism and clinical importance. Report ofthe tenth Ross conference on medical research. Columbus, OH: Ross Laboratories,1991:11-17.
56. Scheppach W, Christl SU, Bartram H-P, Richter F, Kasper H. Effects of short-chainfatty acids on the inflamed colonic mucosa. Scand J Gastroenterol 1997;32 (suppl222):53-57.
57. Kouker G, Jaeger KE. Specific and sensitive plate assay for bacterial lipases. ApplEnviron Microbiol 1987;53:211-213.
58. Ling SC, Weaver LT. The fate of fat in the infant's colon. Q J Med 1997;90:553-555.59. Thompson L, Spiller RC. Impact of polyunsaturated fatty acids on human colonic
bacterial metabolism: an in vitro and in vivo study. Br J Nutr 1996;74:733-741.60. Spiller RC, Brown ML, Phillips SF. Decreased fluid tolerance, accelerated transit, and
abnormal motility of the human colon induced by oleic acid. Gastroenterology1986;91:100-107.
61. Bull AW, Nigro ND, Golembieski WA, Crissman JD, Marnett LJ. In vivo stimulation ofDNA synthesis and induction of ornithine decarboxylase in rat colon by fatty acidhydroperoxides, autoxidation products of unsaturated fatty acids. Cancer Res1984;44:4924-4926.

Chapter 1
26
62. Vonk RJ, Kalivianakis M, Minich DM, Bijleveld CMA, Verkade HJ. The metabolicimportance of unabsorbed dietary lipids in the colon. Scand J Gastroenterol 1997;32(suppl 222):65-67.
63. Poley JR. Fat digestion and absorption in lipase and bile acid deficiency. In: Rommel K,Goebel H, Böhmer R, eds. Lipid absorption: Biochemical and clinical aspects. Lancaster:MTP Press, 1976:151-202.
64. Weber AM, Roy CC. Intraduodenal events in cystic fibrosis. J Pediatr GastroenterolNutr 1984;3(suppl. 1):S113-S119.
65. Narayanan VS, Storch J. Fatty acid transfer in taurodeoxycholate mixed micelles.Biochemistry 1996;35:7466-7473.
66. Brand SJ, Morgan RG. The movement of an unemulsified oil test meal and aqueous- andoil-phase markers through the intestine of normal and bile-diverted rats. Q J Exp PhysiolCogn Med Sci 1975;60:1-13.
67. Lin HC, Zhao X-T, Wang L. Fat absorption is not complete by midgut but is dependenton load of fat. Am J Physiol 1996;271:G62-G67.
68. Pencharz PB, Durie PR. Nutritional management of cystic fibrosis. Annu Rev Nutr1993;13:111-136.
69. Shalon LB, Adelson JW. Cystic fibrosis. Gastrointestinal complications and genetherapy. In: Lebenthal E, ed. The pediatric clinics of North America. Pediatricgastroenterology I. Philadelphia: W.B. Saunders Company, 1996:157-196.
70. Welsh MJ, Smith AE. Molecular mechanisms of CFTR chloride channel dysfunction incystic fibrosis. Cell 1993;73:1251-1254.
71. Zemel BS, Kawchak DA, Cnaan A, Zhao H, Scanlin TF, Stallings VA. Prospectiveevaluation of resting energy expenditure, nutritional status, pulmonary function, andgenotype in children with cystic fibrosis. Pediatr Res 1996;40:578-586.
72. Corey M, McLaughlin FJ, Williams M, Levison H. A comparison of survival, growth,and pulmonary function in patients with cystic fibrosis in Boston and Toronto. J ClinEpidemiol 1988;41:583-591.
73. Dodge JA. Nutritional requirements in cystic fibrosis: a review. J Pediatr GastroenterolNutr 1988;7:S8-S11.
74. Ramsey BW, Farrell PM, Pencharz PB, the Consensus Committee. Nutritionalassessment and management in cystic fibrosis: a consensus report. Am J Clin Nutr1992;55:108-116.
75. Zentler-Munro PL, Fine DR, Batten JC, Northfield TC. Effect of cimetidine on enzymeinactivation, bile acid precipitation, and lipid solubilisation in pancreatic steatorrhoea dueto cystic fibrosis. Gut 1985;26:892-901.
76. Regan PT, Malagelada J-R, DiMagno EP, Go VLW. Reduced intraluminal bile acidconcentrations and fat maldigestion in pancreatic insufficiency: correction by treatment.Gastroenterology 1979;77:285-289.
77. Carroccio A, Pardo F, Montalto G, Iapichino L, Soreso M, Averna MR, Iacono G,Notarbartolo A. Use of famotidine in severe exocrine pancreatic insufficiency withpersistent maldigestion on enzymatic replacement therapy. A long-term study in cysticfibrosis. Dig Dis Sci 1992;37:1441-1446.

General introduction
27
78. Robinson PJ, Smith AL, Sly PD. Duodenal pH in cystic fibrosis and its relationship to fatmalabsorption. Dig Dis Sci 1990;35:1299-1304.
79. Robinson P, Sly P. High dose pancreatic enzymes in cystic fibrosis. Arch Dis Child1990;65:311-312.
80. Smyth R, Van Velzen D, Smyth A, Lloyd D, Heaf D. Strictures of ascending colon incystic fibrosis and high strength pancreatic enzymes. Lancet 1994;343:85-86.
81. MacSweeney E, Oades PJ, Buchdahl RM, Phelan M, Bush A. Relationship of thickeningof colon wall to pancreatic enzyme treatment in cystic fibrosis. Lancet 1995;345:752-756.
82. FitzSimmons SC, Burkhart GA, Borowitz D, Grand RJ, Hammerstrom T, Durie PR,Lloydstill JD, Lowenfels AB. High-dose pancreatic-enzyme supplements and fibrosingcolonopathy in children with cystic fibrosis. N Engl J Med 1997;336:1283-1289.
83. Zentler-Munro PL, FitzPatrick WJF, Batten JC, Northfield TC. Effect of intrajejunalacidity on aqueous phase bile acid and lipid concentrations in pancreatic steatorrhoeadue to cystic fibrosis. Gut 1984;25:500-507.
84. Watkins JB, Tercyak AM, Szczepanik P, Klein PD. Bile salt kinetics in cystic fibrosis:influence of pancreatic enzyme replacement. Gastroenterology 1977;73:1023-1028.
85. Roy CC, Weber EA, Morin CL, Combes J-C, Nusslé D, Mégevand A, Lasalle R.Abnormal biliary lipid composition in cystic fibrosis. Effect of pancreatic enzymes. NEng J Med 1977;297:1301-1305.
86. Belli DC, Levy E, Darling P, Leroy C, Lepage G. Taurine improves the absorption of afat meal in patients with cystic fibrosis. Pediatrics 1987;80:517-523.
87. Eggermont E, De Boeck K. Small intestinal abnormalities in patients with cystic fibrosis.Eur J Pediatr 1991;150:824-828.
88. Dorin JR. Development of mouse models for cystic fibrosis. J Inher Metab Dis1995;18:495-500.
89. Dickinson P, Dorin JR, Porteous DJ. Modelling cystic fibrosis in the mouse. Mol MedToday 1995;1:140-148.
90. Van de Kamer JH, Ten Bokkel Huinink H, Weyers HA. Rapid method for thedetermination of fat in feces. J Biol Chem 1949;177:347-355.
91. Carey MC, Cahalane MJ. Enterohepatic circulation. In: Arias FM, Jacoby WB, PopperH, eds. The liver: Biology and pathobiology. 2nd Ed. New York: Raven, 1988:573-616.
92. Madara JL, Trier JS. Functional morphology of the mucosa of the small intestine. In:Johnson LR, ed. Physiology of the gastrointestinal tract. 2nd Ed. New York: Raven,1987:1209-1249.
93. Lewis GT, Partin HC. Fecal fat on an essentially fat free diet. J Lab Clin Med1954;44:91-93.
94. Wiggins HS, Howell KE, Kellock TD. The origin of fecal fat. Gut 1969;10:400-403.95. Newcomer AD, Hofmann AF, DiMagno EP, Thomas PJ, Carlson GL. Triolein breath
test; A sensitive and specific test for fat malabsorption. Gastroenterology 1979;76:6-13.96. Koletzko B, Sauerwald T, Demmelmair H. Safety of stable isotope use. Eur J Pediatr
1997;156 (suppl 1):S12-S17.

Chapter 1
28
97. Dole M. Relative atomic weights of O in water and air. J Am Chem Soc 1935;57:2731-3735.
98. O'Leary MD, Richard DT, Hendrickson J. Carbon isotope effects on the enzymaticdecarboxylation of glutamic acid. J Am Chem Soc 1970;92:4435-4440.
99. Gregg CT, Huston JY, Prine JR, Ott DG, Furchner JE. Substantial replacement ofmammalian body carbon with carbon-13. Life Sci 1973;13:775-782.
100. Demmelmair H, Sauerwald T, Koletzko B, Richter T. New insights into lipid and fattyacid metabolism via stable isotopes. Eur J Pediatr 1997;156 (Suppl 1):S70-S74.
101. Amarri S, Weaver LT. 13C-breath tests to measure fat and carbohydrate digestion inclinical practice. Clin Nutr 1995;14:149-154.
102. Pedersen NT. Estimation of assimilation of simultaneously ingested 14C-triolein and 3H-oleic acid as a test of pancreatic digestive function. Scand J Gastroenterol 1984;19:161-166.
103. Absalom SR, Saverymuttu SH, Maxwell JD, Levin GE. Triolein breath test of fatabsorption in patients with chronic liver disease. Dig Dis Sci 1988;33:565-569.
104. Mylvaganam K, Hudson PR, Ross A, Williams CP. 14C triolein breath test: a routine testin the gastroenterology clinic? Gut 1986;27:1347-1352.
105. Watkins JB, Klein PD, Schoeller DA, Kirschner BS, Park R, Perman JA. Diagnosis anddifferentiation of fat malabsorption in children using 13C-labeled lipids: trioctanoin,triolein an palmitic acid breath tests. Gastroenterology 1982;82:911-917.
106. Einarsson K, Björkhem I, Eklöf R, Blomstrand R. 14C-triolein breath test as a rapid andconvenient screening test for fat malabsorption. Scand J Gastroenterol 1983;18:9-12.
107. Duncan A, Cameron A, Stewart MJ, Russell RI. Limitations of the triolein breath test.Clin Chim Acta 1992;205:51-64.
108. Pedersen NT, Andersen BN, Marqversen J. Estimation of 14C-triolein assimilation as atest of lipid assimilation. Scand J Gastroenterol 1982;17:309-316.
109. Pedersen NT. Estimation of 14C-triolein assimilation from the postprandial serumradioactivity of 14C. Scand J Clin Lab Invest 1983;43:415-420.
110. Lembcke B, Braden B, Caspary WF. Exocrine pancreatic insufficiency: accuracy andclinical value of the uniformly labelled 13C-hiolein breath test. Gut 1996;39:668-674.
111. Odeka EB, Miller V. Evaluation of fat utilization in paediatric Crohn's disease using 13C-labelled fat. J Pediatr Gastroenterol Nutr 1995;21:430-434.
112. Hsu HW, Butte NF, Wong WW, Moon JK, Ellis KJ, Klein PD, Moise KJ. Oxidativemetabolism in insulin-treated gestational diabetes mellitus. Am J Physiol1997;272:E1099-E1107.
113. Braden B, Picard H, Caspary WF, Posselt HG, Lembcke B. Monitoring pancreatinsupplementation in cystic fibrosis patients with the 13C-Hiolein breath test: evidence fornormalized fat assimilation with high dose pancreatin therapy. Z Gastroenterol1997;35:123-129.
114. Watkins JB, Schoeller DA, Klein PD, Ott DG, Newcomer AD, Hofmann AF. 13C-trioctanoin: a nonradioactive breath test to detect fat malabsorption. J Lab Clin Med1977;90:422-430.

General introduction
29
115. Murphy MS, Eastham EJ, Nelson R, Aynsley-Green A. Non-invasive assessment ofintraluminal lipolysis using a 13CO2 breath test. Arch Dis Child 1990;65:574-578.
116. McClean P, Harding M, Coward WA, Green MR, Weaver LT. Measurement of fatdigestion in early life using a stable isotope breath test. Arch Dis Child 1993;69:366-370.
117. McClean P, Harding M, Coward WA, Prentice A, Austin S, Weaver LT. Bile salt-stimulated lipase and digestion of non-breast milk fat. J Pediatr Gastroenterol Nutr1998;26:39-42.
118. Miyakawa S, Hayakawa M, Horiguchi A, Mizuno K, Ishihara S, Niwamoto N, Miura K.Estimation of fat absorption with the 13-trioctanoin breath test afterpancreatoduodenectomy or pancreatic head resection. World J Surg 1996;20:1024-1028.
119. Kato H, Nakao A, Kishimoto W, Nonami T, Harada A, Hayakawa T, Takagi H. 13C-labeled trioctanoin breath test for exocrine pancreatic function test in patients afterpancreatoduodenectomy. Am J Gastroenterol 1993;88:64-69.
120. Schwabe AD, Bennett LR, Bowman LP. Octanoic acid absorption and oxidation inhumans. J Appl Physiol 1964;19:335-337.
121. Hoshi J, Nishida H, Yasui M, Ohishi M, Takahashi M. [13C]breath test of medium-chaintriglycerides and oligosaccharides in neonates. Acta Paediatr Jpn 1992;34:674-677.
122. Ghoos YF, Vantrappen GR, Rutgeerts PJ, Schurmans PC. A mixed-triglyceride breathtest for intraluminal fat digestive activity. Digestion 1981;22:239-247.
123. Vantrappen GR, Rutgeerts PJ, Ghoos YF, Hiele MI. Mixed triglyceride breath test: Anoninvasive test of pancreatic lipase activity in the duodenum. Gastroenterology1989;96:1126-1134.
124. Kalivianakis M, Verkade HJ, Stellaard F, Van der Werf M, Elzinga H, Vonk RJ. The13C-mixed triglyceride breath test in healthy adults: determinants of the 13CO2 response.Eur J Clin Invest 1997;27:434-442.
125. Swart GR, Baartman EA, Wattimena JL, Rietveld T, Overbeek SE, Van den Berg JW.Evaluation studies of the 13C-mixed triglyceride breath test in healthy controls and adultcystic fibrosis patients with exocrine pancreatic insufficiency. Digestion 1997;58:415-420.
126. Amarri S, Harding M, Coward WA, Evans TJ, Weaver LT. 13Carbon mixed triglyceridebreath test and pancreatic enzyme supplementation in cystic fibrosis. Arch Dis Child1997;76:349-351.
127. Van Aalst K, Veereman-Wauters G, Ghoos YF, Schiffelers S, Van 't Westeinde T,Eggermont E. The 13C mixed triglyceride breath test in children. Gastroenterology1995;108:A759(Abstract).
128. Cole SG, Rossi S, Stern A, Hofmann AF. Cholesteryl octanoate breath test; Preliminarystudies on a new noninvasive test of human pancreatic exocrine function.Gastroenterology 1987;93:1372-1380.
129. Ventrucci M, Cipolla A, Ubalducci GM, Roda A, Roda E. 13C labelled cholesteryloctanoate breath test for assessing pancreatic exocrine insufficiency. Gut 1998;42:81-87.

Chapter 1
30
130. Adler G, Mundlos S, Kühnelt P, Dreyer E. New Methods for assessment of enzymeactivity: do they help to optimize enzyme treatment? Digestion 1993;54:3-9.
131. Mundlos S, Kühnelt P, Adler G. Monitoring enzyme replacement treatment in exocrinepancreatic insufficiency using the cholesteryl octanoate breath test. Gut 1990;31:1324-1328.
132. Hamilton JD, Webb JPW, Dawson AM. The absorption of tristearin and stearic acid andtripalmitin and palmitic acid. Studies on the rate-limiting steps in rats. Biochim BiophysActa 1969;176:27-36.
133. Mills PR, Horton PW, Watkinson G. The value of the 14C breath test in the assessmentof fat absorption. Scand J Gastroenterol 1979;14:913-921.
134. Murphy JL, Jones A, Brookes S, Wootton SA. The gastrointestinal handling andmetabolism of [1-13C]palmitic acid in healthy women. Lipids 1995;30:291-298.
135. Stolinski M, Murphy JL, Jones AE, Jackson AA, Wootton SA. Stable-isotope methodfor determining the gastrointestinal handling of [1-13C]palmitic acid. Lipids1997;32:337-340.
136. Murphy JL, Jones AE, Stolinski M, Wootton SA. Gastrointestinal handling of [1-13C]palmitic acid in healthy controls and patients with cystic fibrosis. Arch Dis Child1997;76:425-427.
137. Van Doorninck JH, French PJ, Verbeek E, Peters RHPC, Morreau H, Bijman J, ScholteBJ. A mouse model for the cystic fibrosis delta-F508 mutation. EMBO J 1995;14:4403-4411.
138. Trezise AEO, Ratcliff R, Hawkins TE, Evans MJ, Freeman TC, Romano PR, HigginsCF, Colledge WH. Co-ordinate regulation of the cystic fibrosis and multidrug resistancegenes in cystic fibrosis knockout mice. Hum Mol Genet 1997;6:527-537.

CHAPTER 2
Detection of intestinal fat malabsorption due toimpaired lipolysis by the 13C-mixed triglyceride
breath test in rats
M. Kalivianakis, J. Elstrodt, R. Havinga, F. Kuipers,F. Stellaard, R.J. Vonk, H.J. Verkade

Chapter 2
32
CHAPTER 2
Detection of intestinal fat malabsorption due toimpaired lipolysis by the 13C-mixed triglyceride
breath test in rats
Abstract
Background & Aim: The 13C-mixed triglyceride (13C-MTG) breath test has become popularfor the detection of impaired intestinal lipolysis as a cause for fat malabsorption. However, thediagnostic value has been questioned because the relation between the extent of fatmalabsorption and the corresponding result of the 13C-MTG breath test has not beenestablished. We characterized the 13C-MTG breath test in rats with variable degrees of fatmalabsorption, achieved by feeding the lipase inhibitor orlistat. Methods: Rats were fed highfat chow (35 en% fat) to which orlistat was added in amounts of 0, 50, 200, and 800 mg kg-1
chow for 5 days. Breath 13CO2 recovery was determined for 6 h after oral administration of13C-MTG (13 mg kg-1 BW). Total dietary fat absorption was measured by means of a 3-dayfecal fat balance. Results: Upon orlistat administration, total fat absorption decreased in adose-dependent way from 80.2 ± 2.2% to 32.8 ± 3.7% (mean ± SEM; 0 mg and 800 mgorlistat kg-1 chow, respectively; P<0.001). Correspondingly, breath 13CO2 recovery from 13C-MTG at 6 h decreased from 84.5 ± 7.8% to 42.0 ± 1.5% of the dose (P<0.001). The 6-hrecovery of breath 13CO2 appeared highly correlated with total fat absorption for the differentdosages of orlistat (r=0.88, P<0.001). However, in rats with fat absorption higher than 70%,the coefficient of variation of cumulative breath 13CO2 excretion was large (15%) comparedwith that of fat absorption (5%). Conclusion: The 13C-MTG breath test correlates significantlywith the extent of fat malabsorption in a rat model of impaired intestinal lipolysis. However,the considerable interindividual variation of the 13C-MTG breath test does not support itsapplication for diagnostic purposes in individual patients.

The 13C-MTG breath test in rats fed orlistat
33
Introduction
Reduced secretion of pancreatic lipase into the intestine is a common feature of pancreaticinsufficiency. This condition may lead to fat malabsorption due to incomplete intestinalhydrolysis of dietary triacylglycerols [1,2]. Intestinal fat malabsorption in patients can bequantified by means of a fat balance, but this method does not discriminate between thepotential causes, such as impaired intestinal lipolysis, disturbed intestinal solubilization of long-chain fatty acids, or decreased chylomicron formation. Measurement of maximal pancreaticlipase output by means of an invasive, marker-corrected perfusion technique is considered tobe the gold standard for pancreatic insufficiency tests [3,4]. A non-invasive test has beendescribed to characterize pancreatic insufficiency in a functional way. In this test, a 13C-labeledmixed triglyceride (13C-MTG; 1,3-distearoyl, 2[carboxyl-13C]octanoyl glycerol) is orallyingested and the amount of 13C in expired air is determined [5]. 13C-MTG contains a 13C-labeled medium-chain fatty acid (octanoic acid) at its sn-2 position, and long-chain fatty acids(stearic acid) at the sn-1 and sn-3 positions of the glycerol backbone. The two stearoylacylchains have to be hydrolyzed by the pancreatic enzyme lipase before 13C-octanoate can beabsorbed, either in the form of a free fatty acid or as a mono-acylglycerol [6]. After itsabsorption, octanoate is rapidly oxidized [6,7]. Thus, the principle of the 13C-MTG test isbased on lipolysis-dependent 13CO2 excretion via the breath.
Since the original description of the 13C-MTG breath test, the test has becomepopular in clinical practice [5,8-11]. However, widely variable results have been obtained inchildren [9], healthy adults [12], and in cystic fibrosis patients with or without pancreaticenzyme replacement therapy [10,11]. The reason for this variability has not been elucidated: infact, quantitative relationship between the extent of fat malabsorption due to impaired lipolysisand the corresponding result of the 13C-MTG breath test has never been demonstrated inhumans or in defined animal models.
A reliable way to decrease the lipolysis activity dose-dependently is with the use oforlistat, an inhibitor of pancreatic lipase [13,14]. Orlistat, the chemically synthesized derivativeof the natural product lipstatin, is a selective and potent inhibitor of lipases, among which,pancreatic lipase [15-23]. Orlistat inactivates pancreatic lipase by reacting covalently withserine (Ser-152) in the active site of the catalytic domain [24,25].
In the present study we aimed to determine the relationship between the extent of fatmalabsorption and the results of the 13C-MTG breath test in a defined controlled animalmodel. We applied the dietary supplementation of orlistat as a reproducible inducer of variousdegrees of fat malabsorption in rats, in analogy to previous studies in mice and humans [26-28]. To ensure that orlistat-induced fat malabsorption was exclusively due to impairedlipolysis, we performed control experiments in which the absorption of the fatty acid[1-13C]palmitic acid was determined, a substrate independent of lipolysis.

Chapter 2
34
Materials and Methods
RatsMale Wistar rats (Harlan, Zeist, The Netherlands), weighing approximately 400 g, werehoused in an environmentally controlled facility with diurnal light cycling and free access totap water and chow. Experimental protocols were approved by the Ethical Committee forAnimal Experiments, Faculty of Medical Sciences, University of Groningen.
MaterialsThe mixed triglyceride (1,3-distearoyl, 2[1-13C]octanoyl glycerol) was purchased from Euriso-Top (Saint Aubin Cedex, France) and was 99% 13C-enriched. In previous articles [5,10,12],the breath test performed with the use of this compound has been denominated as the mixed-triglyceride breath test or as the 13C-MTG breath test. For reasons of consistency, we adhereto this nomenclature. [1-13C]palmitic acid was purchased from Isotec Inc. (Matheson, USA)and was 99% 13C-enriched. Orlistat (previously known as tetrahydrolipstatin, THL, Ro 18-0647) is a synthetic product and was kindly provided by Hoffmann-La Roche (Basel,Switzerland).
Study protocol13C-MTG breath test. Rats were fed ground high-fat chow (35 en% fat; 4.538 kcal kg-1 food;fatty acid composition measured by GC analysis: C8-C12, 4.4 mol%; C16:0, 28.5%; C18:0,3.9%; C18:1n-9, 33.2%; C18:2n-6, 29.3%; C18:3n-3, 0.2%) (Hope Farms BV, Woerden, TheNetherlands) mixed with water (3:2, w/w) to form a homogenous paste. After 2 weeks on thediet, rats were divided into a control group (no orlistat added to the diet) and 3 orlistat groups(50, 200 or 800 mg orlistat per kg chow). There were 4 rats in each experimental group.Orlistat was ground together with the high-fat chow and mixed with water. Administration oforlistat started 2 days prior to the fat balance experiments. Food intake was recorded and feceswas collected for 3 days, in order to perform a fat balance. Feces was stored at -20°C prior toanalysis. After the fat balance, rats were fasted overnight. The following morning they wereplaced in an airtight container (volume ~ 4.5 L) through which CO2-free air was passed at acontinuous flow of 750 mL min-1. The air leaving the metabolic cage was partly diverted (50mL min-1) to a CO2 monitor (Capnograph IV, Gould Medical BV, Bilthoven, TheNetherlands) for measuring percentage of total CO2 in the breath, and to 10 mL test tubes(Exetainers; Labco Limited, High Wycombe, United Kingdom) for collection of breathsamples. The rats were placed in the container at least 30 min before administration of the testmeal containing the label, to have the rats adapted to the cage and to obtain backgroundbreath samples. The test meal consisted of 13C-MTG (13 mg kg-1 body weight) mixed withhigh fat chow (6 g kg-1 body weight), orlistat and water. All rats ingested the test meal within5 min. After ingestion of the test meal, 1-min breath samples were collected in duplicates at30-min intervals for a period of 6 hours.
[1-13C]palmitic acid test. After 1 week on high fat chow, rats were equipped withpermanent catheters in jugular vein, and duodenum as described by Kuipers et al. [29]. Thisexperimental model allows to obtain multiple blood samples in unanesthetized rats without the

The 13C-MTG breath test in rats fed orlistat
35
interference of stress or restraint. Animals were allowed to recover from surgery for 6 daysand were subsequently divided into 2 groups: 1 control group receiving no orlistat and anexperimental group receiving 200 mg orlistat per kg chow. On day 7, 1.67 mL liquid fat kg-1
body weight was slowly administered as a bolus via the duodenal catheter. The fat bolus wascomposed of olive oil (25% v/v; fatty acid composition: C16:0, 14%; C18:1n-9, 79%; C18:2n-6, 8%) and medium-chain triglyceride oil (75% v/v; composed of extracted coconut oil andsynthetic triacylglycerols; fatty acid composition: C6:0, 2% max.; C8:0, 50-65% max.; C10:0,30-45%; C12:0, 3% max.) and contained 33 mg kg-1 body weight [1-13C]palmitic acid and0.47 mg kg-1 body weight orlistat for the experimental group. The fat bolus representedapproximately 15% of the daily fat intake. Blood samples (0.2 mL) were taken from thejugular cannula at baseline, 1, 2, 3, 4, 5, 6 and 24 h after administration of the label and werecollected into tubes containing heparin. Plasma was separated by centrifugation (10 min, 5000rpm, 4°C) and stored at -20°C until further analysis. Feces was collected in 24-h fractionsstarting 1 day before administration of the label and ending 2 days afterwards. Feces sampleswere stored at -20°C prior to analysis. Food intake was determined for 3 days.
Analytical techniquesBreath sample analysis. 13C-enrichment in aliquots of breath samples was determined bymeans of continuous flow isotope ratio mass spectrometry (Finnigan Breath MAT, FinniganMAT GmbH, Bremen, Germany). The 13C-abundance of breath CO2 was expressed as thedifference per mil from the reference standard Pee Dee Belemnite limestone (δ13CPDB, ‰). Theproportion of 13C-label excreted in breath CO2 was expressed as the percentage ofadministered 13C-label recovered per hour (% 13C dose h-1), and as the cumulative percentageof administered 13C-label recovered over the 6-h study period (cum % 13C).
Plasma fats. Total plasma fats (triacylglycerols, phospholipids, etc.) were extracted,hydrolyzed and methylated according to Lepage and Roy [30]. Resulting fatty acid methylesters were analyzed by gas chromatography to measure the total amount of palmitic acid andby gas chromatography combustion isotope ratio mass spectrometry to measure the 13C-enrichment of palmitic acid, as detailed below. The concentration of 13C-palmitic acid inplasma was expressed as the percentage of the dose administered per liter plasma (% dose/L).
Rat chow and fecal fats. Rat chow and feces were freeze-dried and mechanicallyhomogenized, after which aliquots were extracted, hydrolyzed and methylated according tothe method of Lepage and Roy [30]. Resulting fatty acid methyl esters were analyzed by gaschromatography to allow calculation of total fat intake, total fecal fat excretion, and totalpalmitic acid concentration in food and feces. Total fecal fat excretion of rats was expressed asg fat day-1 and percentage of total fat absorption was calculated from the daily fat intake andthe daily fecal fat excretion and expressed as a percentage of the daily fat intake.
Total fat absorptionFat intake (g day ) Fecal fat excretion (g day )
Fat intake (g day )
-1 -1
-1=−
×100%
A similar calculation was performed to measure the absorption of [1-13C]palmitic acid. Valueswere expressed as percentage of the dose administered (% dose).

Chapter 2
36
Gas liquid chromatography. Fatty acid methyl esters were separated and quantifiedby gas liquid chromatography on a Hewlett Packard gas chromatograph Model 6890 equippedwith a CP-SIL 88 capillary column (50 m x 0.32 mm; Chrompack, Middelburg, TheNetherlands) and an FID detector. The gas chromatograph oven was programmed from aninitial temperature of 150°C to 240°C in 2 temperature steps (150°C held 5 min; 150-200°C,ramp 3°C min-1, held 1 min; 200-240°C, ramp 20°C min-1, held 10 min). Quantification of thefatty acid methyl esters was done by adding heptadecanoic acid (C17:0) as internal standard.
Gas chromatography combustion isotope ratio mass spectrometry. 13C-enrichment ofthe palmitic acid methyl esters was determined on a gas chromatography combustion isotoperatio mass spectrometer (Delta S/GC Finnigan MAT, Bremen, Germany). Separation of themethyl esters was achieved on a CP-SIL 88 capillary column (Chrompack; 50 m x 0.32 mm).The gas chromatograph oven was programmed from an initial temperature of 80°C to 225°Cin 3 temperature steps (80°C held 1 min; 80-150°C, ramp 30°C min-1; 150-190°C, ramp 5°Cmin-1; 190-225°C, ramp 10°C min-1, held 5 min).
Calculations and statisticsThe experimental data are reported as means ± SEM. Differences between sample means werecalculated with the use of Student t-test or ANOVA followed by post-hoc analysis (Student-Newman-Keuls). For correlating two variables, regression lines were fitted by the method ofleast squares and expressed as the Pearson correlation coefficient r. Differences betweenmeans were considered statistically significant at the level of P<0.05. Analysis was performedusing SPSS for Windows software (SPSS, Chicago, IL, USA).
Table 2.1 Nutritional data and breath 13CO2 data obtained from control and orlistat-fed rats during 13C-MTG experiments (mean ±
SEM).
Orlistat Fat intake Fecal fat Fat uptake Fat absorption Cum breath 13Cmg kg-1 chow (g day-1) (g day-1) (g day-1) (% intake) (% dose)
0 2.7 ± 0.2a 0.5 ± 0.1a 2.1 ± 0.1a 80.2 ± 2.2a 84.5 ± 7.8a
50 2.1 ± 0.2b 0.3 ± 0.0a 1.8 ± 0.2a 85.2 ± 0.8a 82.0 ± 4.9a
200 2.9 ± 0.1a 1.2 ± 0.1b 1.7 ± 0.1a 59.2 ± 2.1b 58.5 ± 5.3b
800 3.0 ± 0.2a 2.0 ± 0.2c 1.0 ± 0.1b 32.8 ± 3.7c 42.0 ± 1.5c
Unlike letters indicate a significant difference (P<0.05).
Results
13C-MTG testFecal fat balance. Nutritional data of the control and orlistat-fed rats are shown in Table 2.1.Rats fed 50 mg orlistat kg-1 chow showed significantly lower food intake than the othergroups. Administration of 50 mg orlistat kg-1 chow did not lead to a change in fecal fatexcretion. Fecal fat excretion in rats fed 200 and 800 mg orlistat kg-1 chow, however, wassignificantly increased when compared with rats fed 0 or 50 mg orlistat kg-1. In addition, fecalfat excretion in rats fed 800 mg orlistat kg-1 chow was significantly higher compared with rats
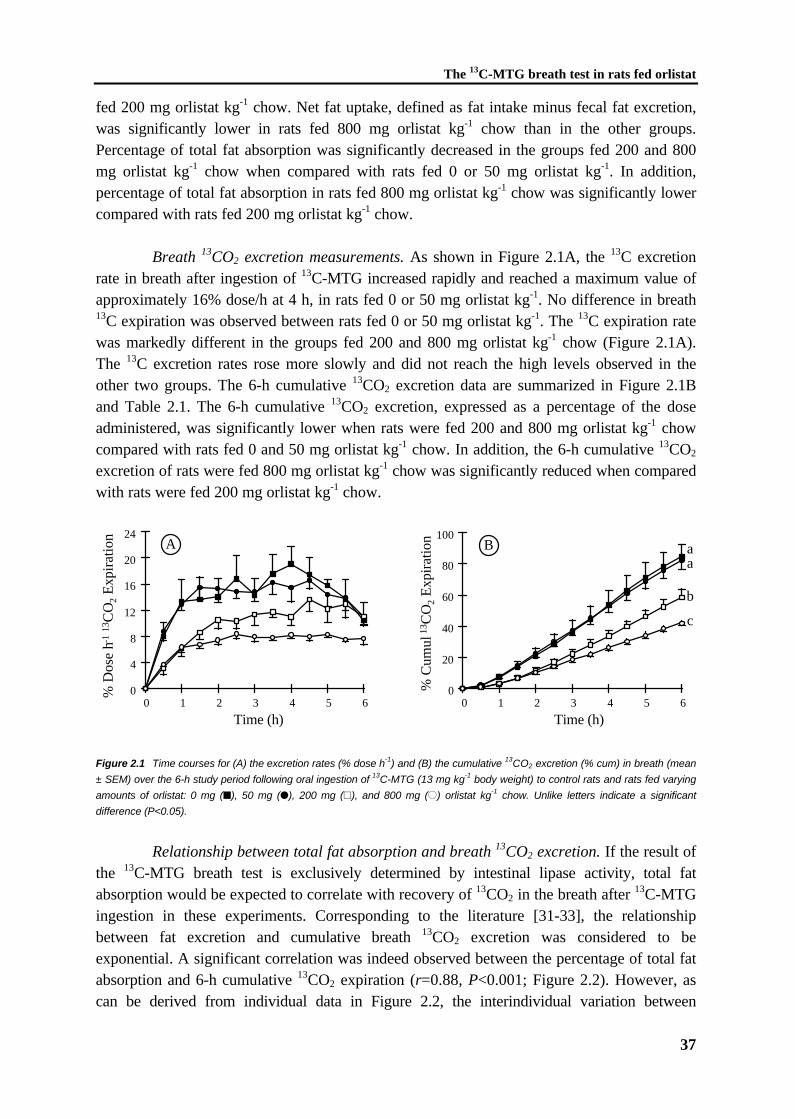
The 13C-MTG breath test in rats fed orlistat
37
fed 200 mg orlistat kg-1 chow. Net fat uptake, defined as fat intake minus fecal fat excretion,was significantly lower in rats fed 800 mg orlistat kg-1 chow than in the other groups.Percentage of total fat absorption was significantly decreased in the groups fed 200 and 800mg orlistat kg-1 chow when compared with rats fed 0 or 50 mg orlistat kg-1. In addition,percentage of total fat absorption in rats fed 800 mg orlistat kg-1 chow was significantly lowercompared with rats fed 200 mg orlistat kg-1 chow.
Breath 13CO2 excretion measurements. As shown in Figure 2.1A, the 13C excretionrate in breath after ingestion of 13C-MTG increased rapidly and reached a maximum value ofapproximately 16% dose/h at 4 h, in rats fed 0 or 50 mg orlistat kg-1. No difference in breath13C expiration was observed between rats fed 0 or 50 mg orlistat kg-1. The 13C expiration ratewas markedly different in the groups fed 200 and 800 mg orlistat kg-1 chow (Figure 2.1A).The 13C excretion rates rose more slowly and did not reach the high levels observed in theother two groups. The 6-h cumulative 13CO2 excretion data are summarized in Figure 2.1Band Table 2.1. The 6-h cumulative 13CO2 excretion, expressed as a percentage of the doseadministered, was significantly lower when rats were fed 200 and 800 mg orlistat kg-1 chowcompared with rats fed 0 and 50 mg orlistat kg-1 chow. In addition, the 6-h cumulative 13CO2
excretion of rats were fed 800 mg orlistat kg-1 chow was significantly reduced when comparedwith rats were fed 200 mg orlistat kg-1 chow.
% D
ose
h-1 13
CO
2 E
xpir
atio
n
0
4
8
12
16
20
24A
Time (h)
0
20
40
60
80
100
0 1 2 3 4 5 6 0 1 2 3 4 5 6
% C
umul
13C
O2
Exp
irat
ion B a
a
b
c
Time (h)
Figure 2.1 Time courses for (A) the excretion rates (% dose h-1) and (B) the cumulative 13CO2 excretion (% cum) in breath (mean
± SEM) over the 6-h study period following oral ingestion of 13C-MTG (13 mg kg-1 body weight) to control rats and rats fed varying
amounts of orlistat: 0 mg (�), 50 mg (�), 200 mg (~), and 800 mg (±) orlistat kg-1 chow. Unlike letters indicate a significant
difference (P<0.05).
Relationship between total fat absorption and breath 13CO2 excretion. If the result ofthe 13C-MTG breath test is exclusively determined by intestinal lipase activity, total fatabsorption would be expected to correlate with recovery of 13CO2 in the breath after 13C-MTGingestion in these experiments. Corresponding to the literature [31-33], the relationshipbetween fat excretion and cumulative breath 13CO2 excretion was considered to beexponential. A significant correlation was indeed observed between the percentage of total fatabsorption and 6-h cumulative 13CO2 expiration (r=0.88, P<0.001; Figure 2.2). However, ascan be derived from individual data in Figure 2.2, the interindividual variation between

Chapter 2
38
recovery of 13CO2 excretion was large. Especially the individual 13C-results in rats with adietary fat absorption higher than 60% showed strong overlap. In these rats, the coefficient ofvariation for percentage of total dietary fat absorption was only 5%, whereas coefficient ofvariation for cumulative breath 13CO2 excretion was 15%.
% C
umul
13C
O2
Exp
irat
ion r = 0.88, P<0.001
0
20
40
60
80
100
0 20 40 60 80 100
% Total Fat Absorption
Figure 2.2 Relationship between the percentage of total fat absorption and breath 13CO2 excretion after oral administration of 13C-
MTG (13 mg kg-1 body weight) in rats fed varying amounts of orlistat (r=0.88, P<0.001); 0 mg (�), 50 mg (�), 200 mg (~), and 800
mg (±) orlistat kg-1 chow.
13C-palmitic acid testData of the 13C-palmitic acid experiment are shown in Table 2.2. No significant difference inmean fat intake was observed between control rats and rats fed 200 mg orlistat kg-1 chow(P=0.36). Orlistat-fed rats excreted significantly more fat into the feces when compared withcontrol rats (P<0.01). The percentage of total fat absorption was significantly decreased inorlistat-fed rats when compared with controls (46.7 ± 5.4% and 74.6 ± 1.3%, respectively,P<0.01).
Table 2.2 Nutritional data of control and orlistat-fed rats during [1-13C]-palmitic acid experiment (mean ± SEM).
Orlistat Fat intake Fecal fat Fat absorption 13C16:0 absorptionmg kg-1 chow (g day-1) (g day-1) (% intake) (% dose)
0 2.2 ± 0.2 0.6 ± 0.0 74.6 ± 0.3 83.7 ± 2.0200 2.5 ± 0.2 1.3 ± 0.2* 46.7 ± 5.4** 87.0 ± 1.0A symbol indicates a significant difference from the control group (0 mg orlistat kg-1 chow). * P<0.05; ** P<0.01.
The amount of 13C-palmitic acid excreted into the feces was calculated for the 48-hperiod following administration of [1-13C]palmitic acid. No significant difference in absorptionof [1-13C]palmitic acid over the 48-h period studied was observed between control andorlistat-fed rats (P=0.71, Table II), demonstrating that administration of orlistat does notaffect the absorption of [1-13C]palmitic acid. This is supported by the fact that 13C-palmiticacid concentrations in plasma after intraduodenal administration of [1-13C]palmitic acid weresimilar in control and orlistat-fed rats (Figure 2.3). The data of the [1-13C]palmitic acidexperiment indicate that fat malabsorption in orlistat treated rats is solely due to impairedlipolysis.
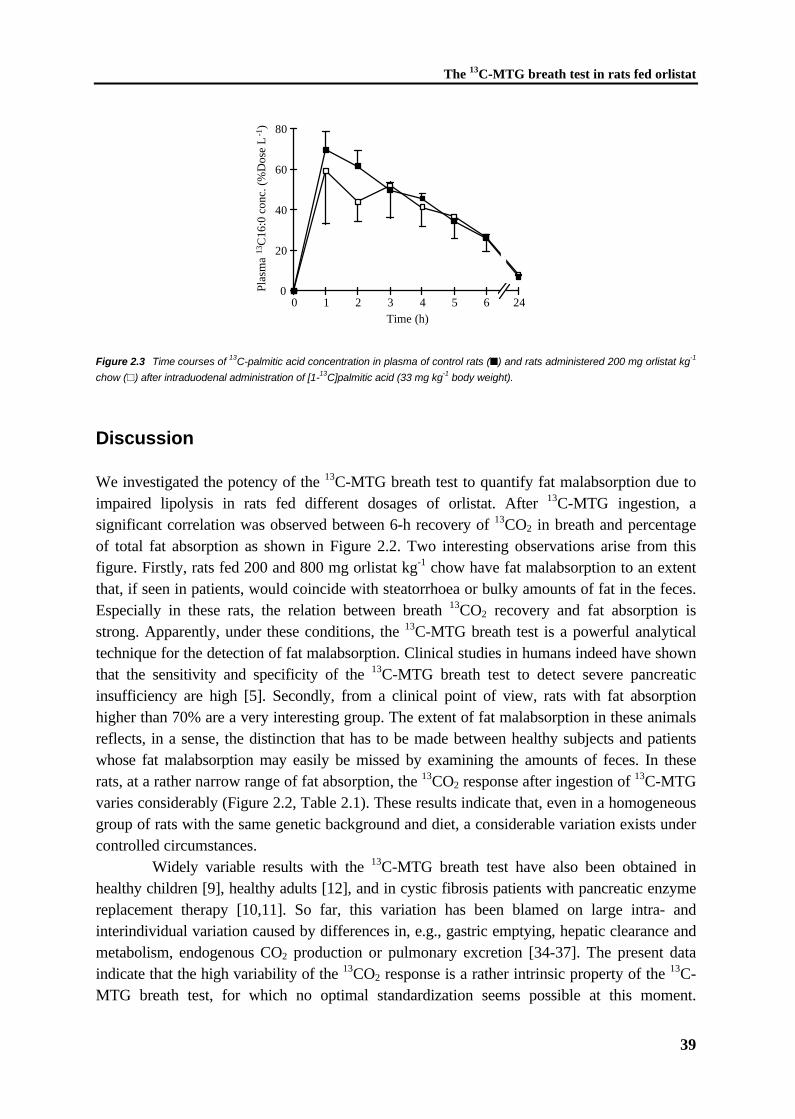
The 13C-MTG breath test in rats fed orlistat
39
0
20
40
60
80
0 1 2 3 4 5 6 24Time (h)
Plas
ma
13C
16:0
con
c. (
%D
ose
L-1
)
Figure 2.3 Time courses of 13C-palmitic acid concentration in plasma of control rats (�) and rats administered 200 mg orlistat kg-1
chow (~) after intraduodenal administration of [1-13C]palmitic acid (33 mg kg-1 body weight).
Discussion
We investigated the potency of the 13C-MTG breath test to quantify fat malabsorption due toimpaired lipolysis in rats fed different dosages of orlistat. After 13C-MTG ingestion, asignificant correlation was observed between 6-h recovery of 13CO2 in breath and percentageof total fat absorption as shown in Figure 2.2. Two interesting observations arise from thisfigure. Firstly, rats fed 200 and 800 mg orlistat kg-1 chow have fat malabsorption to an extentthat, if seen in patients, would coincide with steatorrhoea or bulky amounts of fat in the feces.Especially in these rats, the relation between breath 13CO2 recovery and fat absorption isstrong. Apparently, under these conditions, the 13C-MTG breath test is a powerful analyticaltechnique for the detection of fat malabsorption. Clinical studies in humans indeed have shownthat the sensitivity and specificity of the 13C-MTG breath test to detect severe pancreaticinsufficiency are high [5]. Secondly, from a clinical point of view, rats with fat absorptionhigher than 70% are a very interesting group. The extent of fat malabsorption in these animalsreflects, in a sense, the distinction that has to be made between healthy subjects and patientswhose fat malabsorption may easily be missed by examining the amounts of feces. In theserats, at a rather narrow range of fat absorption, the 13CO2 response after ingestion of 13C-MTGvaries considerably (Figure 2.2, Table 2.1). These results indicate that, even in a homogeneousgroup of rats with the same genetic background and diet, a considerable variation exists undercontrolled circumstances.
Widely variable results with the 13C-MTG breath test have also been obtained inhealthy children [9], healthy adults [12], and in cystic fibrosis patients with pancreatic enzymereplacement therapy [10,11]. So far, this variation has been blamed on large intra- andinterindividual variation caused by differences in, e.g., gastric emptying, hepatic clearance andmetabolism, endogenous CO2 production or pulmonary excretion [34-37]. The present dataindicate that the high variability of the 13CO2 response is a rather intrinsic property of the 13C-MTG breath test, for which no optimal standardization seems possible at this moment.

Chapter 2
40
Therefore, we propose that the 13C-MTG breath test is useful for the detection of severe fatmalabsorption due to low lipase activity in groups of patients. However, the large variation inthe 13CO2 response at a mild degree of fat malabsorption limits the diagnostic possibilities ofthe 13C-MTG breath test in humans [10,11].
No data concerning the 13C-MTG breath test in rats have been published so far. If wecompare the present results on the 13C-MTG breath test in control rats with data obtained inhealthy humans, the 6-h cumulative percentage of breath 13CO2 appears to be much higher inrats: 85% compared with 30% in humans [5,10,12]. We speculate that the extended fastingperiod of our rats directs the absorbed 13C-octanoic acid directly into the oxidation pathway.However, it can not be excluded that part of the difference is based on species specificity.
In vitro studies with orlistat have shown that orlistat is insoluble in aqueous buffers,very poorly soluble in micellar lipid phases, but exhibits good solubility in emulsified lipids[13,38-40]. Therefore, in studies of fat absorption in mice, rats, and humans, inhibition byorlistat was mainly determined by the concentration of the drug in the lipid phase [26,27,40].In contrast, when the dose of orlistat was not pre-dissolved in the dietary fat, but simplyadmixed to the diet or administered as suspension or in capsules in a meal-contingent manner,the inhibitory effect on fat absorption was reduced to a variable extent [26,27]. Therefore, ameaningful comparison between our results on inhibition of fat absorption by orlistat andpreviously published studies is only possible if the experimental design and the mode of drugadministration are taken into account. Since in the present study, orlistat was admixed to thediet, the major part of it was likely dissolved in the target dietary fat upon preparation andmixing of the diet. Except for the 50 mg kg-1 experiment, the effect of orlistat on fatabsorption was dose-dependent up to the largest dose tested. Whether with dose escalationthe effect could be intensified or would level out is unknown. Previously, a similar dose-response relationship has been described in mice to which orlistat was administered eitherdissolved in the fat component of the meal or administered as suspension immediately after themeal [26]. In these mice, excretion of fat in the feces increased exponentially when orlistatdose was increased, until a plateau of 80% of the ingested amount [26]. The orlistat doserequired for half maximal elimination of fat (ID50) reported for mice was approximately 3.3 mgorlistat per g of fat ingested [26]. In our study, the ID50 for rats was approximately 500 mgorlistat per kg chow, corresponding to roughly 2 mg orlistat per g of dietary fat. Thus, despitethe different design of our study, the potency of orlistat expressed as dose per dietary fatingested was rather similar, indicating that the mode of action of orlistat in our study was veryefficient.
To investigate whether the orlistat-induced fat malabsorption was not partially due toother intestinal effects of orlistat resulting in fat malabsorption, control experiments with [1-13C]palmitic acid were performed. The [1-13C]palmitic acid absorption test detects fatmalabsorption due to impaired intestinal uptake of long-chain fatty acids [41]. If fatmalabsorption in rats fed with orlistat were not solely due to the inhibition of intestinallipolysis, an impaired absorption of intraduodenal administered [1-13C]palmitic acid would beexpected. However, fecal 13C-palmitic acid excretion and plasma 13C-palmitic acidconcentrations were not affected at a dosage of 200 mg orlistat kg-1 chow, despite significantlyreduced absorption of dietary fats. These data indicate that the orlistat-fed rat model indeed is

The 13C-MTG breath test in rats fed orlistat
41
specific for impaired lipolysis as cause of fat malabsorption, as has been shown before [26,42-45]. In addition, these data show that in this rat model lipolytic and non-lipolytic processesregarding fat malabsorption can be dissected and measured separately with the use of differentstable isotope tests.
In summary, dietary orlistat administration to rats provides a model for fatmalabsorption, specifically due to impaired intestinal lipolysis. The 13C-MTG breath test in thisanimal model correlates significantly with the extent of induced fat malabsorption. However,variation in 13CO2 results between individual rats was large, especially in rats with dietary fatabsorption higher than 70%. The present data do not support the application of the 13C-MTGbreath test for diagnostic purposes in individual patients.
References
1. Carey MC, Hernell O. Digestion and absorption of fat. Sem Gastrointest Dis 1992;3:189-208.
2. Shiau Y-F. Lipid digestion and absorption. In: Johnson LR, ed. Physiology of theGastrointestinal Tract. 2nd Ed. New York: Raven Press, 1987:1527-1556.
3. Schmidt E, Schmidt FW. Advances in the enzyme diagnosis of pancreatic diseases. ClinBiochem 1990;23:383-394.
4. Goldberg DM, Durie PR. Biochemical tests in the diagnosis of chronic pancreatitis and inthe evaluation of pancreatic insufficiency. Clin Biochem 1993;26:253-275.
5. Vantrappen GR, Rutgeerts PJ, Ghoos YF, Hiele MI. Mixed triglyceride breath test: Anoninvasive test of pancreatic lipase activity in the duodenum. Gastroenterology1989;96:1126-1134.
6. Bach AC, Babayan VK. Medium-chain triglycerides: an update. Am J Clin Nutr1982;36:950-962.
7. Schwabe AD, Bennett LR, Bowman LP. Octanoic acid absorption and oxidation inhumans. J Appl Physiol 1964;19:335-337.
8. Ghoos YF, Vantrappen GR, Rutgeerts PJ, Schurmans PC. A mixed-triglyceride breathtest for intraluminal fat digestive activity. Digestion 1981;22:239-247.
9. Van Aalst K, Veereman-Wauters G, Ghoos YF, Schiffelers S, Van 't Westeinde T,Eggermont E. The 13C mixed triglyceride breath test in children. Gastroenterology1995;108:A759(Abstract).
10. Amarri S, Harding M, Coward WA, Evans TJ, Weaver LT. 13Carbon mixed triglyceridebreath test and pancreatic enzyme supplementation in cystic fibrosis. Arch Dis Child1997;76:349-351.
11. Swart GR, Baartman EA, Wattimena JL, Rietveld T, Overbeek SE, Van den Berg JW.Evaluation studies of the 13C-mixed triglyceride breath test in healthy controls and adultcystic fibrosis patients with exocrine pancreatic insufficiency. Digestion 1997;58:415-420.
12. Kalivianakis M, Verkade HJ, Stellaard F, Van der Werf M, Elzinga H, Vonk RJ. The 13C-mixed triglyceride breath test in healthy adults: determinants of the 13CO2 response. Eur JClin Invest 1997;27:434-442.

Chapter 2
42
13. Borgström B. Mode of action of tetrahydrolipstatin: a derivative of the naturallyoccurring lipase inhibitor lipstatin. Biochim Biophys Acta 1988;962:308-316.
14. Hadváry P, Lengsfeld H, Wolfer H. Inhibition of pancreatic lipase in vitro by the covalentinhibitor tetrahydrolipstatin. Biochem J 1988;256:357-361.
15. Gargouri Y, Chahini H, Moreau H, Ransac S, Verger R. Inactivation of pancreatic andgastric lipases by THL and C12:0-TNB: a kinetic study with emulsified tributyrin. BiochimBiophys Acta 1991;1085:322-328.
16. Ransac S, Gargouri Y, Moreau H, Verger R. Inactivation of pancreatic and gastric lipasesby tetrahydrolipstatin and alkyl-dithio-5-(2-nitrobenzoic acid). A kinetic study with 1,2-didecanoyl-sn-glycerol monolayers. Eur J Biochem 1991;202:395-400.
17. Cudrey C, Van Tilbeurgh H, Gargouri Y, Verger R. Inactivation of pancreatic lipases byamphiphilic reagents 5-(dodecyldithio)-2-nitrobenzoic acid and tetrahydrolipstatin.Dependence upon partitioning between micellar and oil phases. Biochemistry1998;32:13800-13808.
18. Lookene A, Skottova N, Olivecrona G. Interactions of lipoprotein lipase with the active-site inhibitor tetrahydrolipstatin (orlistat). Eur J Biochem 1994;222:395-403.
19. Lee MW, Kraemer FB, Severson DL. Characterization of a partially purifieddiacylglycerol lipase from bovine aorta. Biochim Biophys Acta 1995;1254:311-318.
20. Sheriff S, Du H, Grabowski GA. Characterization of lysosomal acid lipase by site-directedmutagenesis and heterologous expression. J Biol Chem 1995;270:27766-27772.
21. Smith GM, Garton AJ, Aitken A, Yeaman SJ. Evidence for a multi-domain structure forhormone-sensitive lipase. FEBS Lett 1996;396:90-94.
22. Guzelhan C, Odink J, Niestijl Jansen-Zuidema JJ, Hartmann D. Influence of dietarycomposition on inhibition of fat absorption by orlistat. J Int Med Res 1994;22:255-265.
23. Hartmann D, Guzelhan C, Crijns HJMJ, Peeters PAM, Persson P, Jonkman JHG.Comparison of galenical formulations of orlistat (tetrahydrolipstatin). A pharmacologicalapproach. Drug Invest 1993;5:44-50.
24. Hadváry P, Sidler W, Meister W, Vetter W, Wolfer H. The lipase inhibitortetrahydrolipstatin binds covalently to the putative active site serine of pancreatic lipase. JBiol Chem 1991;266:2021-2027.
25. Lüthi-Peng Q, Marki HP, Hadváry P. Identification of the active-site serine in humanpancreatic lipase by chemical modification with tetrahydrolipstatin. FEBS Lett1992;299:111-115.
26. Isler D, Moeglen C, Gains N, Meier MK. Effect of the lipase inhibitor orlistat and ofdietary lipid on the absorption of radiolabelled triolein, tri-gamma-linolenin and tripalmitinin mice. Br J Nutr 1995;73:851-862.
27. Zhi J, Melia AT, Guerciolini R, Chung J, Kinberg J, Hauptman JB, Patel IH.Retrospective population-based analysis of the dose-response (fecal fat excretion)relationship of orlistat in normal and obese volunteers. Clin Pharmacol Ther 1994;56:82-85.
28. Hauptman JB, Jeunet FS, Hartmann D. Initial studies in humans with the novelgastrointestinal lipase inhibitor Ro 18-0647 (tetrahydrolipstatin). Am J Clin Nutr1992;55:309S-313S.

The 13C-MTG breath test in rats fed orlistat
43
29. Kuipers F, Havinga R, Bosschieter H, Toorop GP, Hindriks FR, Vonk RJ. Enterohepaticcirculation in the rat. Gastroenterology 1985;88:403-411.
30. Lepage G, Roy CC. Direct transesterification of all classes of lipids in a one-step reaction.J Lipid Res 1986;27:114-120.
31. Newcomer AD, Hofmann AF, DiMagno EP, Thomas PJ, Carlson GL. Triolein breathtest; A sensitive and specific test for fat malabsorption. Gastroenterology 1979;76:6-13.
32. Einarsson K, Björkhem I, Eklöf R, Blomstrand R. 14C-triolein breath test as a rapid andconvenient screening test for fat malabsorption. Scand J Gastroenterol 1983;18:9-12.
33. Mills PR, Horton PW, Watkinson G. The value of the 14C breath test in the assessment offat absorption. Scand J Gastroenterol 1979;14:913-921.
34. Watkins JB, Schoeller DA, Klein PD, Ott DG, Newcomer AD, Hofmann AF. 13C-trioctanoin: a nonradioactive breath test to detect fat malabsorption. J Lab Clin Med1977;90:422-430.
35. Watkins JB, Tercyak AM, Szczepanik P, Klein PD. Bile salt kinetics in cystic fibrosis:influence of pancreatic enzyme replacement. Gastroenterology 1977;73:1023-1028.
36. Murphy MS, Eastham EJ, Nelson R, Aynsley-Green A. Non-invasive assessment ofintraluminal lipolysis using a 13CO2 breath test. Arch Dis Child 1990;65:574-578.
37. Amarri S, Coward WA, Harding M, Weaver LT. Importance of measuring CO2
production rate in 13C breath tests. Proc Nutr Soc 1994;54:111A(Abstract).38. Guerciolini R. Mode of action of orlistat. Int J Obes 1997;21:S12-S23.39. Fernández E, Borgström B. Effects of tetrahydrolipstatin, a lipase inhibitor, on absorption
of fat from the intestine of the rat. Biochim Biophys Acta 1989;1001:249-255.40. Fernández E, Borgström B. Intestinal absorption of retinol and retinyl palmitate in the rat.
Effects of tetrahydrolipstatin. Lipids 1990;25:549-552.41. Minich DM, Kalivianakis M, Havinga R, Kuipers F, Stellaard F, Vonk RJ, Verkade HJ. A
novel 13C-linoleic acid absorption test detects lipid malabsorption due to impairedsolubilization in rats. Gastroenterology 1997;112 (suppl.):A894(Abstract).
42. Fernandez E, Borgstrom B. Effects of tetrahydrolipstatin, a lipase inhibitor, on absorptionof fat from the intestine of the rat. Biochim Biophys Acta 1989;1001:249-255.
43. Hogan S, Fleury A, Hadváry P, Lengsfeld H, Meier MK, Triscari J, Sullivan AC. Studieson the antiobesity activity of tetrahydrolipstatin, a potent and selective inhibitor ofpancreatic lipase. Int J Obes 1987;11(Suppl 3):35-42.
44. Froehlich F, Hartmann D, Guezelhan C, Gonvers JJ, Jansen JBMJ, Fried M. Influence oforlistat on the regulation of gallbladder contraction in man; A randomized double-blindplacebo-controlled crossover study. Dig Dis Sci 1996;41:2404-2408.
45. Schwizer W, Asal K, Kreiss C, Mettraux C, Borovicka J, Remy B, Guzelhan C, HartmannD, Fried M. Role of lipase in the regulation of upper gastrointestinal function in humans.Am J Physiol 1997;273:G612-G620.


CHAPTER 3
The 13C-mixed triglyceride breath test in healthyadults: determinants of the 13CO2 response
M. Kalivianakis, H.J. Verkade, F. Stellaard,M. van der Werf, H. Elzinga, R.J. Vonk
Eur J Clin Invest (1997) 27, 434-442

Chapter 3
46
CHAPTER 3
The 13C-mixed triglyceride breath test in healthyadults: determinants of the 13CO2 response
Abstract
Background & Aim: Defects in lipolysis due to pancreatic insufficiency can be diagnosed bythe mixed triglyceride 13CO2 breath test. However, the effects of various test conditions on the13CO2 response have only partially been elucidated. Methods: In healthy adults we performedthe 13CO2 mixed triglyceride breath test and we compared a) the inter- and intra-individualvariation in the 13CO2 response, b) the effect of two different test meals, c) the effect of anadditional meal during the test, and d) the effect of physical exercise during the test. Results:Upon repeating the test in the same individual (test meal cream), repeatability coefficientswere large, either with respect to time to maximum 13C excretion rate (3.8 h), maximum 13Cexcretion rate (4.9% 13C dose/h), or cumulative recovery of 13C over the 9-h study period(22.7% 13C dose). The cumulative 13C expiration over 9 h obtained with the test mealcomposed of cream was quantitatively similar to that obtained with bread and butter: 42.2 ±8.4%, and 47.7 ± 6.3%, respectively. Fasting for 9 h during the test resulted in similar 13Cexpiration rates and cumulative 13C expiration (43.4% ± 7.2%), when compared toconsumption of an additional meal at 3 h after the start of the test (38.3 ± 5.3%). The 13CO2
response increased in 5 out of 7 subjects, but decreased in the other 2, when moderateexercise was performed (bicycle ergometer, 50W for 5 h). Conclusion: Repeatability of theMTG test in healthy adults is low. The present results indicate that a solid and a liquid testmeal, containing similar amount of fats, give similar cumulative 13CO2 responses, and thatstringent prolonged fasting during the test is unnecessary. Standardization of physical activityseems preferable, since unequivocal effects of moderate exercise on the 13CO2 response wereobserved in the individuals studied.

The 13C-MTG breath test in healthy adults
47
Introduction
A common feature of pancreatic insufficiency is a reduced output of pancreatic lipase. Thiscondition may lead to lipid malabsorption due to reduced intestinal hydrolysis oftriacylglycerols [1,2]. Measurement of maximal pancreatic lipase output by means of aninvasive, marker-corrected perfusion technique is considered to be the gold standard ofpancreatic insufficiency tests [3,4]. A non-invasive test has recently been described in which a13C-labeled mixed triglyceride (MTG) was ingested together with a test meal, after which theamount of 13C in expired air was determined [5]. The MTG used is 1,3-distearoyl, 2[carboxyl-13C]octanoyl glycerol. This molecule contains a 13C-labeled medium-chain fatty acid (octanoicacid) at the sn-2 position, and long-chain fatty acids (stearic acid) at the sn-1 and sn-3positions of the glycerol backbone of the triacylglycerol [5]. The two stearoyl chains have tobe hydrolyzed by lipolytic enzymes in the intestine (mainly of pancreatic origin) before[13C]octanoate can be absorbed, either in the form of a free fatty acid or of a mono-acylglycerol [6]. It has been known that, after absorption, octanoate is rapidly oxidized to aconsiderable extent [6,7]. Thus, the principle of the MTG test is based on lipolysis-dependent13CO2 excretion via the breath. The applicability of the MTG test in pancreatic insufficiencyhas been demonstrated in adults [5,8,9], and preliminary data on the potential applicability inchildren are available [10].
A general problem of breath tests using labeled lipids for the diagnosis of pancreaticinsufficiency or fat malabsorption in general, is a relatively poor sensitivity and specificity,probably due to the numerous steps involved in the metabolism of the tracer compound [11].Differences in gastric emptying, solubilization by bile acids, mucosal absorption, hepaticclearance and metabolism, endogenous CO2 production and pulmonary excretion may obscurethe relationship between the quantity of label expired and the aim of the study, for examplehydrolysis in the intestine [12-15]. Up to now, none of these breath tests has been clinicallyvalidated in different disease states. Presently, the discriminating test parameters are onlypoorly defined, which limits the application of these tests in clinical studies.
The aim of the present study was to further characterize the MTG breath test and toidentify some factors apart from pancreatic insufficiency that may influence the quantitativerecovery of 13CO2 in breath. We examined the variation of the 13CO2 response within andbetween healthy human adults, in which no rate-limiting variation in pancreatic exocrinefunction was expected. In addition, we evaluated various determinants of the 13CO2 response:
1. The test meal. A variety of test meals has been described for breath tests with a diversity ofsubstrates [5,14,16,17]. So far, no standardized test meal for clinical purpose of thesebreath tests has been proposed. A disadvantage of a test meal such as bread and butter forchildren could be the extended time it would take for consumption, and the risk of notconsuming it quantitatively. Furthermore, such a test meal is not applicable to small infants.Therefore, we examined whether the 13CO2 response of a liquid test meal is similar to the13CO2 response of the mentioned solid test meal.

Chapter 3
48
2. The fasting condition during the test. Since it may be cumbersome to keep patients, inparticular infants, fasted for at least six hours, we determined to what extent consumptionof an extra meal during the test influenced the 13CO2 response.
3. Physical activity. It is known that physical activity considerably affects the production rateof CO2 and nutrient oxidation [18-20], however it is not established to what extent itinfluences the results of the 13C-MTG test.
Materials and methods
SubjectsThe studies were conducted with four male and seven female volunteers with a mean age of 23± 1 (SEM) years and a mean body mass index of 20.9 ± 0.3 kg/m2. The volunteers werehealthy according to medical histories and did not have symptoms of lipid malabsorption, suchas diarrhea or gastrointestinal complaints. Informed consent was obtained, and the studyprotocol was approved by the Medical Ethics Committee of the University HospitalGroningen.
13C-labeled substrateMTG (mixed triglyceride, 1,3-distearoyl, 2[carboxyl-13C]octanoyl glycerol) was purchasedfrom the Belgian Institute of Isotopes (IRE, Fleurus, Belgium) and from Euriso-Top, SaintAubin Cedex, France, and was 99% 13C-enriched. The chemical purity exceeded 98%. Boththe isotopic and chemical purity were checked by NMR.
Study protocolThe subjects were instructed to avoid consumption of naturally 13C-enriched foods (e.g. cornor corn products, pineapple, cane sugar) for at least two days prior to the study. After anovernight fast (approximately 10 hours), each subject consumed a test meal consisting ofeither 75 ml cream or 2 slices of bread and 25 g butter, each mixed with 13C labeled MTG (4mg/kg body weight). Breath samples were collected in duplicates before consumption of thetest meal to provide a value of baseline 13C-excretion in expired CO2, and were subsequentlycollected at 30-min intervals for a period of 9 hours after the ingestion of the test meal. Unlessstated otherwise, all experiments were performed under standard conditions, which impliedthat: 1. the test meal consisted of 75 ml cream, 2. no additional food or liquids were permittedduring the 9-h period except for water, tea and coffee without sugar and milk, and 3. thesubjects only performed light office tasks during the tests.
Analytical techniquesBreath was collected by expiration via a straw into a 10 ml tube (Exetainers; Labco Limited,High Wycombe, United Kingdom), from which aliquots were taken to determine 13C-enrichment by means of continuous flow isotope ratio mass spectrometry (Finnigan TracerMAT, Finnigan MAT GmbH, Bremen, Germany). The 13C-abundance of breath CO2 wasexpressed as the difference per mil from the reference standard Pee Dee Belemnite limestone

The 13C-MTG breath test in healthy adults
49
(δ13CPDB, ‰). The proportion of 13C-label excreted in breath CO2 was expressed as thepercentage of administered 13C-label recovered per hour, and as the percentage ofadministered 13C-label recovered over the 9-h study period.
Mean values of whole body CO2 excretion were measured by indirect calorimetry(Oxycon, model ox-4, Dräger, Breda, The Netherlands) at 3 separate periods of 5 minutesduring the 9-h study period.
Intra- and inter-individual variationThe intra-individual variation was examined in eleven subjects by repeating the study underidentical (standard) conditions within four weeks after the first test. The individualrepeatability of the test was examined with the use of repeatability coefficients according toBland and Altman [21]. Coefficients of repeatability are based on the mean of the differencesbetween repeated measurements on a series of subjects and the standard deviation of thedifferences. The definition of a repeatability coefficient adopted by the British StandardsInstitution is the expectation that 95% of differences is within two standard deviations of themean difference [22]. Repeatability coefficients were calculated with respect to threeparameters: time to maximum 13C excretion rate, tmax, maximum 13C excretion rate, andcumulative recovery of 13C after 9 h [23].
Influence of two different test meals on 13CO2 expirationIn six subjects the influence of two different easily applicable test meals on the 13CO2 responsein breath after oral ingestion of MTG was examined: 1. 75 ml cream (1040 kJ, 26 g fat, 2 gcarbohydrate, 2 g protein), and 2. two slices of bread with 25 g butter (1550 kJ, 22 g fat, 32 gcarbohydrate, 6 g protein). In addition, the influence of either test meal in itself on 13C-enrichment of breath was examined in each subject, by repeating the test with a test meal towhich no 13C-MTG was added.
Prolonged fasting during the test versus consumption of an additional mealIn seven subjects the effect of prolonged fasting on the 13C expiration rate was examined.Fasting represented the standard condition mentioned above, whereas in the non-fastingcondition an additional meal was consumed at 3 hours after the start of the experiment. Theadditional meal consisted of 2 slices of bread and 30 g strawberry jam (960 kJ, 2 g fat, 49 gcarbohydrate, 6 g protein). The experiments were performed with the initial test meal (time 0h) consisting of 75 ml cream. In five subjects a control study was performed, in which the 13C-labeled MTG was omitted from the test meal.
Influence of physical exercise on 13CO2 expirationIn seven subjects, the influence of physical exercise on the 13CO2 response was investigated.The results from the tests done under standard conditions were compared to those obtainedduring moderate exercise on a bicycle ergometer. The physical exercise started at 10 minutesbefore the consumption of the test meal (75 ml cream). The energy performance was 50 wattfor 5 hours, which represents an intensity of approximately 25 to 35% of the subjects’maximal aerobic capacity (VO2max). Drinking of water was allowed ad libitum during the

Chapter 3
50
bicycle test for the whole 9-h period. The last 4 hours of the experiment (time 5-9 h) thesubjects were in rest. Five subjects underwent a control study during which the influence ofthe test meal and exercise as mentioned above was examined under background conditions,i.e. without the addition of the label.
Statistical methodsThe experimental data are reported as means ± SEM. Statistical comparisons between the datawere performed with the use of the two-tailed non-parametric Wilcoxon signed-rank test forpairs. Differences between means were considered statistically significant at the level ofP<0.05. For statistical analysis three different characteristics of the 13CO2 expiration wereanalyzed according to Matthews et al. [23]: 1. the time to the maximum 13C excretion rate,tmax, 2. the maximum 13C excretion rate (expressed as % 13C dose per hour), and 3. therecovery of 13C over the 9-h study period (expressed as % cumulative 13C excreted).
Results
Background variation of 13CO2 expiration after an unlabeled test mealThe background variation of 13C in breath was examined in several subjects by performing thetest without administration of the label under 4 different experimental conditions: standardcondition, test meal consisting of bread and butter, additional meal at 3 h after the start of thetest, and moderate exercise during the test for 5 hours. There was no detectable change in theaverage expiration of 13CO2 over the 9-h study period, in any of the experimental settings. Theinter-individual background 13C-variation in breath CO2 at the various time points was small(average SEM 0.12‰), as was the intra-individual variation (average SEM 0.21‰, data notshown).
Intra- and inter-individual variationThe baseline 13C-abundance in breath prior to consumption of the test meal was -26.2 ± 0.2‰in test 1. The standard error of the analysis at this enrichment level was 0.03‰ (n=10). Afteringestion of the 13C-MTG containing test meal at time 0, different time-course patterns wereobserved for the excretion of 13C-label in breath over the 9-h study period, with maximumexcretion rates varying between 3 and 8 h after administration of the 13C-labeled test meal(Figure 3.1, closed squares). At the maximum excretion rate, the enrichment of 13C in breathwas -21.6 ± 0.5‰ and at the end of the 9-h study period, the enrichment of 13C in breath hadnot yet returned to the level of baseline 13C-abundance (-24.6 ± 0.3‰). When expressed as aproportion of administered 13C, the peak excretion rate of label in breath in the first test was9.3 ± 1.3% 13C per hour of the administered dose, varying between 5.0 and 16.3%. Over the9-h study period the excretion of 13C in breath was 40.3 ± 5.0% of that administered, rangingbetween 16.7 and 66.4% (Table 3.1; the subject numbers in Table 3.1 are corresponding to thesubject numbers in the figures).

The 13C-MTG breath test in healthy adults
51
12
16
0
4
8
0 3 6 9
8
0
4
8
12
16
0 3 6 9
7
0
4
8
12
16
0 3 6 9
2
0
4
8
12
16
0 3 6 9
6
0
4
8
12
16
0 3 6 9
4
0
4
8
12
16
0 3 6 9
3
0
4
8
12
16
0 3 6 9
5
0
4
8
12
16
0 3 6 9
912
16
0
4
8
0 3 6 9
10
0
4
8
12
16
0 3 6 9
11
0
4
8
12
16
0 3 6 9
1
Time (h)
% 13
C d
ose
h-1
Figure 3.1 Time courses for the excretion of 13C in breath over 9 h following oral ingestion of MTG (4 mg per kg body weight) at
time 0 in eleven healthy adults in test 1 (� ) and test 2 (�). The tests were done under standard conditions with the test meal 75 ml
cream.
On repeating the test under identical circumstances, the time-course patterns of mostindividuals appeared rather similar to the first test with maximum excretion rates varyingbetween 3 and 9 h after administration of the 13C-labeled test meal (Figure 3.1, open squares).However, in one subject (Figure 3.1, subject 1) a strikingly different time course of labelexpiration was observed when compared to the first test: tmax of 3 h in the first study and a tmax
probably after 9 h. The mean results on 13CO2 expiration were not significantly different whencompared to the first test: the baseline 13C-abundance was -26.0 ± 0.1‰, the enrichment of13C in breath at peak excretion was -21.7 ± 0.5‰, and the enrichment of 13C in breath at theend of the 9-h study period was, again, not yet at baseline 13C-abundance (-24.8 ± 0.6‰).When expressed as a proportion of administered 13C, the peak excretion rate of label was 8.3 ±1.0%, ranging from 4.5 to 14.5%, and excretion of label in breath over the 9-h study periodwas 33.2 ± 3.6%, ranging from 18.8 to 48.3% (Table 3.1).
For the 3 parameters studied, the repeatability coefficients [21] of time to maximum13C excretion tmax, maximum 13C excretion rate, and cumulative recovery of 13C after 9 h were3.8 h, 4.9% 13C dose/h, and 22.7% 13C cumulative excreted, respectively (Figure 3.2). Thus,for example, since the repeatability coefficient of the cumulative percentage 13C excreted is22.7%, the cumulative percentage 13C excreted in a second experiment performed underidentical circumstances may be 22.7% above or below the cumulative percentage 13C excreted
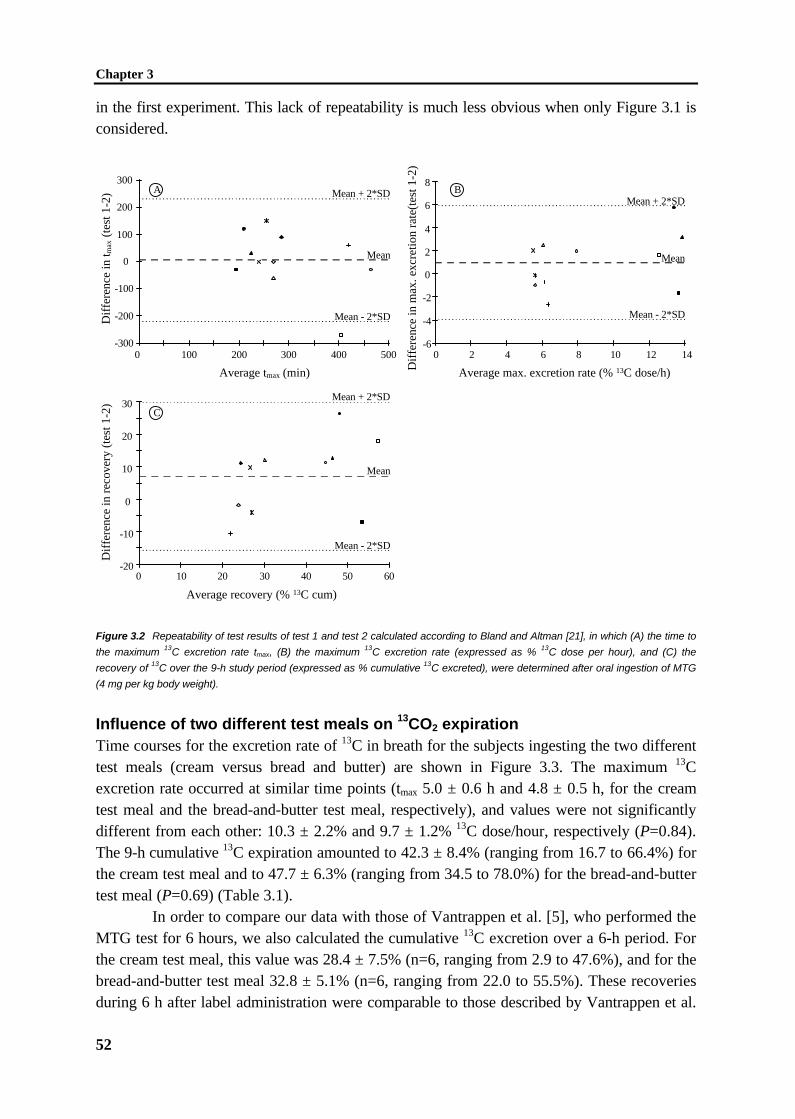
Chapter 3
52
in the first experiment. This lack of repeatability is much less obvious when only Figure 3.1 isconsidered.
-300
-200
-100
0
100
200
300
0 100 200 300 400 500
Mean
Mean - 2*SD
Mean + 2*SD
Average tmax (min)
Dif
fere
nce
in t m
ax (
test
1-2
)
-6
-4
-2
0
2
4
6
8
0 2 4 6 8 10 12 14
Mean
Mean - 2*SD
Mean + 2*SD
Average max. excretion rate (% 13C dose/h)Dif
fere
nce
in m
ax. e
xcre
tion
rate
(tes
t 1-2
)
-20
-10
0
10
20
30
0 10 20 30 40 50 60
Mean
Mean - 2*SD
Mean + 2*SD
Average recovery (% 13C cum)
Dif
fere
nce
in r
ecov
ery
(tes
t 1-2
)
A B
C
Figure 3.2 Repeatability of test results of test 1 and test 2 calculated according to Bland and Altman [21], in which (A) the time to
the maximum 13C excretion rate tmax, (B) the maximum 13C excretion rate (expressed as % 13C dose per hour), and (C) the
recovery of 13C over the 9-h study period (expressed as % cumulative 13C excreted), were determined after oral ingestion of MTG
(4 mg per kg body weight).
Influence of two different test meals on 13CO2 expirationTime courses for the excretion rate of 13C in breath for the subjects ingesting the two differenttest meals (cream versus bread and butter) are shown in Figure 3.3. The maximum 13Cexcretion rate occurred at similar time points (tmax 5.0 ± 0.6 h and 4.8 ± 0.5 h, for the creamtest meal and the bread-and-butter test meal, respectively), and values were not significantlydifferent from each other: 10.3 ± 2.2% and 9.7 ± 1.2% 13C dose/hour, respectively (P=0.84).The 9-h cumulative 13C expiration amounted to 42.3 ± 8.4% (ranging from 16.7 to 66.4%) forthe cream test meal and to 47.7 ± 6.3% (ranging from 34.5 to 78.0%) for the bread-and-buttertest meal (P=0.69) (Table 3.1).
In order to compare our data with those of Vantrappen et al. [5], who performed theMTG test for 6 hours, we also calculated the cumulative 13C excretion over a 6-h period. Forthe cream test meal, this value was 28.4 ± 7.5% (n=6, ranging from 2.9 to 47.6%), and for thebread-and-butter test meal 32.8 ± 5.1% (n=6, ranging from 22.0 to 55.5%). These recoveriesduring 6 h after label administration were comparable to those described by Vantrappen et al.
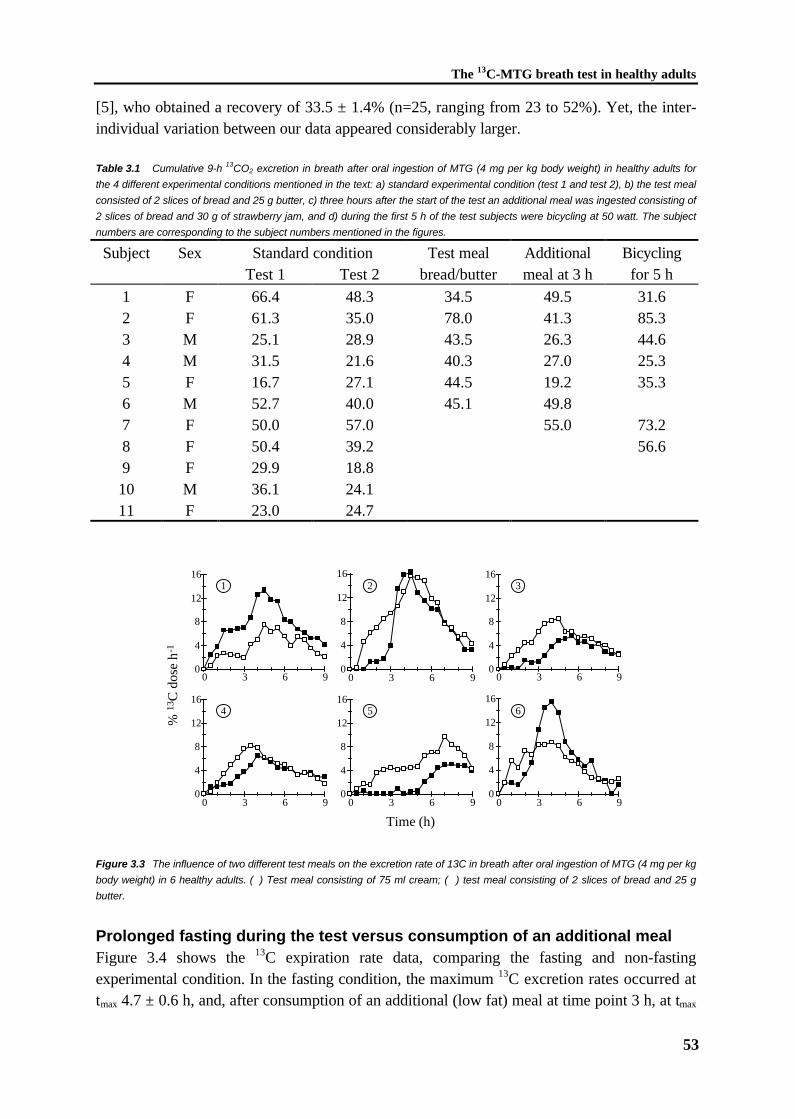
The 13C-MTG breath test in healthy adults
53
[5], who obtained a recovery of 33.5 ± 1.4% (n=25, ranging from 23 to 52%). Yet, the inter-individual variation between our data appeared considerably larger.
Table 3.1 Cumulative 9-h 13CO2 excretion in breath after oral ingestion of MTG (4 mg per kg body weight) in healthy adults for
the 4 different experimental conditions mentioned in the text: a) standard experimental condition (test 1 and test 2), b) the test meal
consisted of 2 slices of bread and 25 g butter, c) three hours after the start of the test an additional meal was ingested consisting of
2 slices of bread and 30 g of strawberry jam, and d) during the first 5 h of the test subjects were bicycling at 50 watt. The subject
numbers are corresponding to the subject numbers mentioned in the figures.
Subject Sex Standard condition Test meal Additional BicyclingTest 1 Test 2 bread/butter meal at 3 h for 5 h
1 F 66.4 48.3 34.5 49.5 31.62 F 61.3 35.0 78.0 41.3 85.33 M 25.1 28.9 43.5 26.3 44.64 M 31.5 21.6 40.3 27.0 25.35 F 16.7 27.1 44.5 19.2 35.36 M 52.7 40.0 45.1 49.87 F 50.0 57.0 55.0 73.28 F 50.4 39.2 56.69 F 29.9 18.810 M 36.1 24.111 F 23.0 24.7
0 3 6 9 0 3 6 9
0 3 6 9 0 3 6 90
4
8
12
16
0
4
8
12
16
0
4
8
12
16
0
4
8
12
16
0
4
8
12
16
0 3 6 9
4
1
5
2 3
0
4
8
12
16
0 3 6 9
6
Time (h)
% 1
3 C d
ose
h-1
Figure 3.3 The influence of two different test meals on the excretion rate of 13C in breath after oral ingestion of MTG (4 mg per kg
body weight) in 6 healthy adults. (� ) Test meal consisting of 75 ml cream; (�) test meal consisting of 2 slices of bread and 25 g
butter.
Prolonged fasting during the test versus consumption of an additional mealFigure 3.4 shows the 13C expiration rate data, comparing the fasting and non-fastingexperimental condition. In the fasting condition, the maximum 13C excretion rates occurred attmax 4.7 ± 0.6 h, and, after consumption of an additional (low fat) meal at time point 3 h, at tmax

Chapter 3
54
4.4 ± 0.4 h, and did not differ between the two groups: 10.7 ± 1.9% and 9.4 ± 1.4% 13Cdose/hour, respectively (P=0.38). The 9-h cumulative 13C expiration amounted to 43.4 ± 7.2%(ranging from 16.7 to 66.4%) for the fasting condition and to 38.3 ± 5.3% (ranging from 19.2to 55.0%) for the non-fasting condition (not significantly different, P=0.38) (Table 3.1). Theconsumption of an additional low-fat meal at 3 h after the start of the experiment neitheraltered the form nor the height of the mean curve for all subjects.
0
4
8
12
16
0 3 6 9
5
0
4
8
12
16
0 3 6 9
4
0
4
8
12
16
0 3 6 9
3
0
4
8
12
16
0 3 6 9
7
0
4
8
12
16
0 3 6 9
6
0
4
8
12
16
0 3 6 9
2
0
4
8
12
16
0 3 6 9
1
Time (h)
% 13
C d
ose
h-1
Figure 3.4 The influence of the consumption of an additional meal 3 h after the start of the experiment on the excretion rate of 13C
in breath after oral ingestion of MTG (4 mg per kg body weight) in 7 healthy adults. (� ) Prolonged fasting during the test; (�)
Consumption of an additional carbohydrate rich meal 3 hours after the start of the experiment.
Influence of physical exercise on 13CO2 expirationThe effects of physical exercise on the 13C excretion were not similar in the seven subjectsstudied (Figure 3.5). In five of the seven subjects, physical exercise during the test induced anincrease in the maximal 13C excretion rate and cumulative 13C excretion when compared to thetest under standard conditions (i.e. resting during the entire experiment). However, one of theseven subjects responded in the opposite direction (Figure 3.5, Table 3.1; subject 1), as noincrease of the peak excretion rate was observed after physical exercise, and an actualdecrease in the cumulative 13C excretion was noticed. The remaining subject (Figure 3.5, Table3.1; subject 4) showed a small increase in the maximal 13C excretion rate, but still thecumulative 13C excretion was decreased. The peak excretion of 13C-label in breath during therest session in the subjects occurred at tmax 5.2 ± 0.7 h after administration of the 13C-labeledtest meal and amounted to 9.8 ± 1.7% 13C dose per hour (n=7). The total excretion of 13Cover the 9-h study period was 43.1 ± 7.1%. Bicycling for 5 h at moderate intensity decreasedthe time to maximum 13C excretion (tmax 3.9 ± 0.9 h), however, this effect was not significant(P=0.22). Furthermore, the maximum 13C excretion rate in the seven subjects was significantlyincreased (16.3 ± 2.6% versus 9.8 ± 1.7%, P=0.03), although total excretion of 13C over the9-h study period was not significantly increased (50.3 ± 8.5% versus 43.1 ± 7.1%, P=0.47).

The 13C-MTG breath test in healthy adults
55
0
10
20
30
0 3 6 9
5
0
10
20
30
0 3 6 9
4
0
10
20
30
0 3 6 9
3
8
0 3 6 90
10
20
307
0 3 6 90
10
20
30
2
0 3 6 90
10
20
30
0
10
20
30
0 3 6 9
1
Time (h)
% 13
C d
ose
h-1
Figure 3.5 The influence of physical exercise on the cumulative excretion of 13C in breath after oral ingestion of MTG (4 mg per kg
body weight) in 7 healthy adults. (� ) Prolonged resting during the test; (�) Performance of physical exercise during the test on an
bicycle ergometer at 50W for 5 hours. During last 4 hours of the experiment subjects were in rest.
Discussion
The present investigations in healthy adults were designed to further characterize thedeterminants of the 13C-MTG test in healthy volunteers. Intra- and inter-individual variationswere examined by repeating the study in the same individuals on a separate occasion. We alsodetermined to which extent the 13CO2 response was influenced by different test meals, byprolonged fasting, and by physical exercise.
The variability of measurements in different subjects (i.e. the inter-individualvariation) is usually considerably larger than the variability between measurements on the samesubject (i.e. the intra-individual variation) [24]. Obviously, both kinds of variability areimportant to assess the potential applicability of a diagnostic test. Only in one recent study therepeatability of breath tests was investigated based on the ingestion of 13C-labeled lipid,involving [1-13C]palmitic acid in healthy volunteers [17]. Also in this study a poor repeatabilitywas reported. No information is available on the repeatability of the MTG test underphysiological or pathological conditions. We examined the repeatability of the MTG test bydetermining coefficients of repeatability [21]. The calculated repeatability coefficients fromdata of 11 healthy adults for the 3 parameters studied are considerable (see Results section).The values of the repeatability coefficients reflect the range of outcomes in which a repeatedtest in the same individual will result with a 95% likelihood. Thus, for example, since therepeatability coefficient of the cumulative percentage 13C excreted is 22.7%, the cumulativepercentage 13C excreted in a second experiment is predicted to be (with a 95% likelihood)22.7% above or below the cumulative percentage 13C excreted in the first experiment. Theselarge repeatability coefficients are predominantly due to a considerable intra-individualvariation. The acceptability of a certain diagnostic test is based on the actual values obtainedand their repeatability coefficients in patients, compared to healthy controls. At present, only

Chapter 3
56
information on the mean values of 13CO2 expiration of pancreatic insufficiency patients isavailable from the literature. It remains to be established whether the means and repeatabilitycoefficients of diseased individuals allow a clear discrimination from unaffected individualsusing the MTG-test.
Various test meals have been described for the use in breath tests with a diversity oflipid substrates [5,14,16,17]. So far, no standardized test meal for clinical purpose of thesebreath tests has been proposed. Theoretically, however, the test meal might influence the timecourse of the 13CO2 response, for example by affecting the rate of gastric emptying [25-29]. Inour present study a liquid and a solid test meal with a comparable amount of fat were appliedto investigate the effect of different test meals on the 13CO2 response. The cumulativepercentage 13C of the dose excreted was comparable for the two test meals. This particularfeature is valuable for the potential application of the MTG test in children, since a liquid testmeal is more convenient to administer.
The originally described MTG breath test [5] lasted for 6 hours. Cumulative excretionmight be the most discriminative parameter when MTG is used as a clinical test. However,also the cumulative excretion of 13CO2 depends on the end point (e.g. 6, 9 or more hours),which has not yet been defined by validation studies. In order to evaluate possible diagnosticadvantages we extended the collection period of breath sampling up to 9 hours. In subjects,who reached their maximum excretion rate before 6 h, an increase in oxidation byapproximately 50% was observed when the cumulative percentage 13C excreted after 9 hourswas compared to that after 6 hours. However, in 2 subjects (Figure 3.1, subjects 5 and 8), thecumulative percentage 13C excreted after 9 hours was more than twice the amount excretedafter 6 hours. It is tempting to speculate that these particular subjects have a decreased gastricemptying rate, compared to the others. If so, especially subjects with slow gastric emptyingwill be diagnosed incorrectly when the time period of the breath test is not sufficiently long.
The originally described MTG-test [5] involved prolonged fasting for 6 hours. Thisfeature of the test would probably limit its applicability in (very young) pediatric patients. Toallow eating during the test would alleviate the applicability. However, the results of the 13CO2
response should then not be influenced by eating. Present data indicate that the consumptionof an additional low-fat meal 3 hours after the start of the experiment does neither change theform of the 13C expiration curve nor the height of the curve. These observations suggest thateating an additional meal during the test does not influence the results, which is in agreementwith the findings of Schwabe et al. [7]. They concluded that oral administration of glucose didnot seem to inhibit 14C-labeled octanoic acid oxidation [7].
In five out of seven individuals studied, the performance of physical exercise duringthe test increased the cumulative recovery of 13C in breath, when compared to performance ofthe test under resting conditions. Exercise enhances the oxidation of carbohydrates and lipids,which is associated with a higher CO2 and 13CO2 response. Available data suggest that gastricemptying during exercise is subject to a number of factors including calorie count, mealosmolality, meal temperature and exercise conditions [30]. There are several indicationssuggesting that light to moderate exercise accelerates gastric emptying of either liquid andsolid meals [30-33]. Intestinal absorption per se has not been evaluated in great detail duringexercise, but probably changes little [34]. All together, this explains the results of the five

The 13C-MTG breath test in healthy adults
57
mentioned individuals quite well. Unexpectedly, two subjects responded differently. Atpresent, no clear explanation could be obtained for the particular results in these two subjects;both were healthy according to medical histories and, like the others, showed no symptoms oflipid malabsorption, such as diarrhea or gastrointestinal complaints. This observation doesunderline, however, the importance of standardizing the resting conditions during the MTGbreath test.
In summary, we have found that the repeatability of the mixed triglyceride test inhealthy adults is low. The results also suggest that two distinct test meals (cream versus breadand butter) give a similar cumulative 13CO2 response and that stringency on continuous fastingduring the test is unnecessary, which is in favor of the applicability of the test in pediatricpatients. Standardization of resting conditions still seems preferable. In future patient studies itremains to be established whether MTG-test results obtained under conditions of an impairedintestinal lipolysis allow the sensitive and specific discrimination of affected from unaffectedindividuals.
References
1. Carey MC, Hernell O. Digestion and absorption of fat. Sem Gastrointest Dis 1992;3:189-208.
2. Shiau Y-F. Lipid digestion and absorption. In: Johnson LR, ed. Physiology of theGastrointestinal Tract. 2nd Ed. New York: Raven Press, 1987:1527-1556.
3. Schmidt E, Schmidt FW. Advances in the enzyme diagnosis of pancreatic diseases. ClinBiochem 1990;23:383-394.
4. Goldberg DM, Durie PR. Biochemical tests in the diagnosis of chronic pancreatitis and inthe evaluation of pancreatic insufficiency. Clin Biochem 1993;26:253-275.
5. Vantrappen GR, Rutgeerts PJ, Ghoos YF, Hiele MI. Mixed triglyceride breath test: Anoninvasive test of pancreatic lipase activity in the duodenum. Gastroenterology1989;96:1126-1134.
6. Bach AC, Babayan VK. Medium-chain triglycerides: an update. Am J Clin Nutr1982;36:950-962.
7. Schwabe AD, Bennett LR, Bowman LP. Octanoic acid absorption and oxidation inhumans. J Appl Physiol 1964;19:335-337.
8. Ghoos YF, Vantrappen GR, Rutgeerts PJ, Schurmans PC. A mixed-triglyceride breathtest for intraluminal fat digestive activity. Digestion 1981;22:239-247.
9. Maes BD, Ghoos YF, Geypens BJ, Hiele MI, Rutgeerts PJ. Relation between gastricemptying rate and rate of intraluminal lipolysis. Gut 1996;38:23-27.
10. Van Aalst K, Veereman-Wauters G, Ghoos YF, Schiffelers S, Van 't Westeinde T,Eggermont E. The 13C mixed triglyceride breath test in children. Gastroenterology1995;108:A759(Abstract).
11. Pedersen NT, Jorgensen BB, Rannem T. The [14C]-triolein breath test is not valid as a testof fat absorption. Scand J Clin Lab Invest 1991;51:699-703.

Chapter 3
58
12. Watkins JB, Schoeller DA, Klein PD, Ott DG, Newcomer AD, Hofmann AF. 13C-trioctanoin: a nonradioactive breath test to detect fat malabsorption. J Lab Clin Med1977;90:422-430.
13. Watkins JB, Tercyak AM, Szczepanik P, Klein PD. Bile salt kinetics in cystic fibrosis:influence of pancreatic enzyme replacement. Gastroenterology 1977;73:1023-1028.
14. Murphy MS, Eastham EJ, Nelson R, Aynsley-Green A. Non-invasive assessment ofintraluminal lipolysis using a 13CO2 breath test. Arch Dis Child 1990;65:574-578.
15. Amarri S, Coward WA, Harding M, Weaver LT. Importance of measuring CO2
production rate in 13C breath tests. Proc Nutr Soc 1994;54:111A(Abstract).16. Watkins JB, Klein PD, Schoeller DA, Kirschner BS, Park R, Perman JA. Diagnosis and
differentiation of fat malabsorption in children using 13C-labeled lipids: trioctanoin, trioleinan palmitic acid breath tests. Gastroenterology 1982;82:911-917.
17. Murphy JL, Jones A, Brookes S, Wootton SA. The gastrointestinal handling andmetabolism of [1-13C]palmitic acid in healthy women. Lipids 1995;30:291-298.
18. Brooks GA, Mercier J. Balance of carbohydrate and lipid utilization during exercise: the"crossover" concept. J Appl Physiol 1994;76:2253-2261.
19. Coyle EF. Substrate utilization during exercise in active people. Am J Clin Nutr1995;61:968S-979S.
20. Romijn JA, Coyle EF, Sidossis LS, Gastaldelli A, Horowitz JF, Endert E, Wolfe RR.Regulation of endogenous fat and carbohydrate metabolism in relation to exerciseintensity and duration. Am J Physiol 1993;265:E380-E391.
21. Bland JM, Altman DG. Statistical methods for assessing agreement between two methodsof clinical measurement. Lancet 1986;307-310.
22. British Standards Institution. Precision of test methods I: Guide for the determination andreproducibility for a standard test method (BS 5497, part I), 1979. (Abstract).
23. Matthews JNS, Altman DG, Campbell MJ, Royston P. Analysis of serial measurements inmedical research. Br Med J 1990;300:230-235.
24. Bland JM, Altman DG. Correlation, regression, and repeated data. Br Med J1994;308:896.
25. Maes BD, Ghoos YF, Geypens BJ, Hiele MI, Rutgeerts PJ. Relation between gastricemptying rate and energy intake in children compared with adults. Gut 1995;36:183-188.
26. Hölzer HH, Turkelson CM, Solomon TE, Raybould HE. Intestinal lipid inhibits gastricemptying via CCK and a vagal capsaicin-sensitive afferent pathway in rats. Am J Physiol1994;267:G625-G629.
27. Edelbroek M, Horowitz M, Maddox A, Bellen J. Gastric emptying and intragastricdistribution of oil in the presence of a liquid or a solid meal. J Nucl Med 1992;33:1283-1290.
28. Hunt JN, Stubbs DF. The volume and energy content of meals as determinants of gastricemptying. J Physiol London 1975;245:209-225.
29. Hunt JN, Knox MT. A relation between the chain length of fatty acids and the slowing ofgastric emptying. J Physiol 1968;194:327-336.
30. Moses FM. The effect of exercise on the gastrointestinal tract. Sports Med 1990;9:159-172.

The 13C-MTG breath test in healthy adults
59
31. Moore JG, Datz FL, Christian PE. Exercise increases solid meal gastric emptying rates inmen. Dig Dis Sci 1990;35:428-432.
32. Cammack J, Read NW, Cann PA, Greenwood B, Holgate AM. Effect of prolongedexercise on the passage of a solid meal through the stomach and small intestine. Gut1982;23:957-961.
33. Marzio L, Formica P, Fabiani F, LaPenna D, Vecchiett L, Cuccurullo F. Influence ofphysical activity on gastric emptying of liquids in normal human subjects. Am JGastroenterol 1991;86:1433-1436.
34. Feldman M, Nixon JV. Effect of exercise on postprandial gastric secretion and emptyingin humans. J Appl Physiol 1982;53:851-854.


CHAPTER 4
The 13C-palmitic acid test with plasma samplingdetects fat malabsorption in bile-diverted rats
M. Kalivianakis, D.M. Minich, R. Havinga, F. Kuipers,F. Stellaard, R.J. Vonk , H.J. Verkade

Chapter 4
62
CHAPTER 4
The 13C-palmitic acid test with plasma samplingdetects fat malabsorption in bile-diverted rats
Abstract
Background & Aims: The fecal fat balance does not discriminate between the distinct intestinalprocesses, such as lipolysis, solubilization, chylomicron formation, as causes of fatmalabsorption. In the present study we characterized a rat model with fat malabsorption dueto bile diversion and we investigated the diagnostic potency of the [1-13C]palmitic acid test.Methods: Bile-diverted and control rats were fed standard (14 en% fat) or high-fat chow (35en% fat) for 2 weeks. After intraduodenal administration of [1-13C]palmitic acid (33 mg kg-1
BW) blood samples were taken for measurements of 13C-palmitic acid enrichment. Foodintake was quantified and feces was collected for balance studies. Intestinal histology wasdetermined. Results: Total fat absorption was highly efficient in control rats (mean ± SEM:standard chow 96.7 ± 0.2%; high-fat chow 93.2 ± 0.4%), but was significantly decreased inbile-diverted rats (standard chow 87.2 ± 0.9%, P<0.001; high-fat chow 53.9 ± 3.9%,P<0.001). Plasma 13C-palmitate concentrations allowed discrimination between normal(>91%) and decreased fat absorption due to bile diversion. Conclusion: The [1-13C]palmiticacid absorption test detects fat malabsorption due to bile diversion in rats. Application of thistest in clinical states with fat malabsorption may allow design of specific treatment strategies.

The 13C-palmitic acid test in bile-diverted rats
63
Introduction
Adequate absorption of dietary fats by the intestine is required for supply of energy, membraneconstituents, and precursors for the formation of hormones and inflammatory mediators [1,2].In Western diets, triacylglycerols composed of long-chain fatty acids constitute 92 to 96% ofdietary fats [1]. The absorption of these fats involves several specific processes. Firstly,lipolysis, by lipolytic enzymes originating predominantly from the pancreas, leads to hydrolysisof triacylglycerols into fatty acids and 2-monoacylglycerols. Secondly, during the process ofsolubilization, mixed micelles are formed consisting of bile acids, phospholipids andcholesterol [3]. The mixed micelles are thought to act as the physiological transport vehicles oflipolytic products from dietary fats in the intestinal lumen. Finally, the fatty acids andmonoacylglycerols are translocated across the intestinal epithelium, converted back totriacylglycerols, assembled into chylomicrons and secreted into the lymph [1,4,5].
The efficiency of intestinal fat absorption in patients is routinely determined by meansof a fat balance, requiring detailed analysis of daily fat intake and the complete recovery offeces for 72 h. However, in the case of fat malabsorption, this method does not discriminatebetween the potential causes, such as impaired intestinal lipolysis, disturbed intestinalsolubilization of long-chain fatty acids or chylomicron formation. Stable isotope techniqueshave been introduced in the development of novel diagnostic strategies. Several 13C-labeledfats, such as tri-[1-13C]octanoin and 1,3-distearoyl, 2[1-13C]octanoyl glycerol, have beensuccessfully applied for the rather specific detection of impaired lipolysis [6-8]. Attempts todevelop a specific test for the detection of impaired solubilization have been less successful.Watkins et al. [9] administered [1-13C]palmitic acid, tri-[1-13C]octanoin or tri-[1-13C]olein topatients with impaired lipolysis, bile salt deficiency or mucosal disease and measured breath13CO2 excretion.
So far, the use of 13C-labeled fats for quantitative studies on defective fat absorptionhas been limited to breath and feces analysis [9-11]. The excretion rate of 13C in the form ofexhaled 13CO2, however, does not necessarily reflect quantitative differences in the absorptionof the 13C-labeled parent compound, e.g., due to variations in the post-absorptive metabolism[12,13]. The availability of gas chromatography isotope ratio mass spectrometry allows foraccurate determination of 13C enrichments in plasma fatty acids [14]. The determination ofplasma concentrations of absorbed 13C-labeled fats as a measure of their absorption offers atheoretical advantage over breath 13CO2 analysis, since numerous steps are involved in the(post-absorptive) metabolism of the tracer prior to exhalation of 13CO2 [15].
In order to determine the potency of a novel test, the availability of an animal modelis almost essential. Manipulation of the enterohepatic circulation of bile components has beenrepeatedly and successfully used in rat studies on fat (mal)absorption and metabolism [16-19].During bile diversion, no bile components are available in the intestine and fat malabsorptionappears to be mainly due to impaired solubilization of long-chain fatty acids [20]. However,recent observations have suggested that bile is also essential for efficient chylomicronformation [21-23]. Bile-diverted rats regain normal feeding behavior and normal growthcurves within several days after surgical interruption of the enterohepatic circulation [24].

Chapter 4
64
In the present study, firstly, we aimed to characterize the rat model of fatmalabsorption due to bile diversion. For this purpose, total fat absorption and [1-13C]palmiticacid absorption was determined in rats that received either standard (14 en% fat) or high-fatchow (35 en% fat). Secondly, we investigated the potency of the [1-13C]palmitic acid test withplasma 13C-palmitic acid concentrations to detect fat malabsorption due to bile diversion.Control experiments, in which tri-[1-13C]palmitoylglycerol was administered, were performedto check whether lipolysis is unaffected in our animal model.
Materials and Methods
AnimalsMale Wistar rats (Harlan, Zeist, The Netherlands), weighing 300 to 400 g, were housed in anenvironmentally controlled facility with diurnal light cycling and free access to food and tapwater (and additional saline, 0.9% NaCl w/v, in the case of bile-diverted rats). Experimentalprotocols were approved by the Ethical Committee for Animal Experiments, Faculty ofMedical Sciences, University of Groningen.
13C-labeled substrates[1-13C]-labeled palmitic acid was purchased from Isotec Inc. (Matheson, USA) and was 99%13C-enriched. Tri-[1-13C]palmitoylglycerol was purchased from ICN Biomedicals Inc.(Cambridge, United Kingdom) and was 99% 13C-enriched.
Study protocolRats were assigned to either standard chow (14 en% fat; 4.575 kcal kg-1 food; fatty acidcomposition measured by GC analysis: C8-C12, 0.9 mol%; C16:0, 25.2%; C18:0, 5.5%;C18:1n-9, 30.3%; C18:2n-6, 33.9%; C18:3n-3, 3.6%) or to high-fat chow (35 en% fat; 4.538kcal kg-1 food; fatty acid composition measured by GC analysis: C8-C12, 4.4 mol%; C16:0,28.5%; C18:0, 3.9%; C18:1n-9, 33.2%; C18:2n-6, 29.3%; C18:3n-3, 0.2%) (Hope Farms BV,Woerden, The Netherlands). After 1 week, rats were equipped with permanent catheters injugular vein, bile duct and duodenum as described by Kuipers et al. [24]. This experimentalmodel allows for physiological studies in unanesthetized rats with long term bile diversionwithout the interference of stress or restraint. One day after surgery, catheters in bile duct andduodenum were either connected at time of surgery to restore the enterohepatic circulation(control rats) or catheters were chronically interrupted (bile-diverted rats). Animals wereallowed to recover from surgery for 6 days.
On day 7, 1.67 mL fat kg-1 body weight was slowly administered as a bolus via theduodenal catheter. The fat bolus was composed of olive oil (25% v/v; fatty acid composition:C16:0, 14 mol%; C18:1n-9, 79%; C18:2n-6, 8%) and medium-chain triglyceride oil (75% v/v;composed of extracted coconut oil and synthetic triacylglycerols; fatty acid composition: C6:0,2 mol% ; C8:0, 50-65% max.; C10:0, 30-45%; C12:0, 3% max.) and contained either 33 mgkg-1 body weight [1-13C]palmitic acid or 33 mg kg-1 body weight tri-[1-13C]palmitoylglycerol.The fat bolus represented approximately 25 and 15% of the daily fat intake in the standard and

The 13C-palmitic acid test in bile-diverted rats
65
the high-fat group, respectively. Blood samples (0.2 mL) were taken from the jugular cannulaat baseline, 1, 2, 3, 4, 5, 6 and 24 h after administration of the label and were collected intotubes containing heparin. Plasma was separated by centrifugation (10 min, 2000 rpm, 4°C) andstored at -20°C until further analysis. Feces was collected in 24-h fractions starting 1 daybefore administration of the fat bolus and ending 2 days afterwards. Feces samples were storedat -20°C prior to analysis. Food intake was determined for 3 days by daily weighing of thefood container.
Analytical techniquesPlasma fats. Total plasma fats (triacylglycerols, phospholipids, etc.) were extracted,hydrolyzed and methylated according to Lepage and Roy [25]. Resulting fatty acid methylesters were analyzed by gas chromatography to measure the total amount of palmitic acid andby gas chromatography combustion isotope ratio mass spectrometry to measure the 13C-enrichment of palmitic acid. The concentration of 13C-palmitic acid in plasma was expressed asthe percentage of the dose administered per liter plasma (% dose L-1).
Rat chow and fecal fats. Feces was freeze-dried and mechanically homogenized.Aliquots of rat chow and freeze-dried feces were extracted, hydrolyzed and methylated [25].Resulting fatty acid methyl esters were analyzed by gas chromatography to calculate total fatintake, total fecal fat excretion, and total palmitic acid concentration in food and feces. Fattyacid methyl esters were analyzed by gas chromatography combustion isotope ratio massspectrometry to calculate the 13C-enrichment of palmitic acid. Total fecal fat excretion of ratswas expressed as g fat day-1 and percentage of total fat absorption was calculated from thedaily fat intake and the daily fecal fat excretion and expressed as a percentage of the daily fatintake.
Percentage of total fat absorptionFat intake (g day ) Fecal fat output (g day )
Fat intake (g day )
-1 -1
-1=−
×100%
A similar calculation was performed to measure the absorption of [1-13C]palmitic acid and tri-[1-13C]palmitoylglycerol. The absorption of the label was determined from the intake andexcretion of 13C-palmitic acid. Values were expressed as percentage of the dose administered(% dose).
Gas liquid chromatography. Fatty acid methyl esters were separated and quantifiedby gas liquid chromatography on a Hewlett Packard gas chromatograph Model 5880 equippedwith a CP-SIL 88 capillary column (Chrompack; 50 m x 0.32 mm) and an FID detector[26,27]. The gas chromatograph oven was programmed from an initial temperature of 150°Cto 240°C in 2 temperature steps (150°C held 5 min; 150-200°C, ramp 3°C min-1, held 1 min;200-240°C, ramp 20°C min-1, held 10 min). Quantification of the fatty acid methyl esters wasachieved by adding heptadecanoic acid (C17:0) as internal standard.
Gas chromatography combustion isotope ratio mass spectrometry. 13C-enrichment ofthe palmitic acid methyl esters was determined on a gas chromatography combustion isotoperatio mass spectrometer (Delta S/GC Finnigan MAT, Bremen, Germany) [28]. Separation ofthe methyl esters was achieved on a CP-SIL 88 capillary column (Chrompack; 50 m x 0.32

Chapter 4
66
mm). The gas chromatograph oven was programmed from an initial temperature of 80°C to225°C in 3 temperature steps (80°C held 1 min; 80-150°C, ramp 30°C min-1; 150-190°C, ramp5°C min-1; 190-225°C, ramp 10°C min-1, held 5 min).
Calculations and statisticsThe experimental data are reported as means ± SEM. Significance of differences wascalculated with the use of the two-tailed Student’s t-test for unpaired data. For correlating twovariables, linear regression lines were fitted by the method of least squares and expressed asthe Pearson correlation coefficient r. Differences between means were considered statisticallysignificant at the level of P<0.05.
Results
Fecal fat balanceIn Table 4.1 nutritional data of control and bile-diverted rats on standard chow (14 en% fat)and high-fat chow (35 en% fat) are shown.
Standard chow (14 en% fat). Mean food intake, and thus fat intake, over the 3-dayperiod was significantly increased in bile-diverted rats on standard chow when compared tocontrol rats (P<0.05). Bile-diverted rats excreted significantly more fat into the feces whencompared to control rats (P<0.001). Although the percentage of total fat absorption waslower in bile-diverted rats when compared to control rats (87.2 ± 0.9% vs. 96.7 ± 0.2%,respectively, P<0.001), net fat uptake in both control and bile-diverted rats was similar(P=0.91).
High-fat chow (35 en% fat). Both food and fat intake were significantly increased inbile-diverted rats when compared to control rats (P<0.01). Similarly, fecal fat excretion wassignificantly increased in bile-diverted rats compared to control rats (P<0.001). Again, therewas no significant difference in the net fat uptake of either control and bile-diverted rats(P=0.10), although percentage of total fat absorption was considerably decreased in bile-diverted rats when compared to control rats (53.9 ± 3.9% vs. 93.2 ± 0.4%, respectively,P<0.001).
Table 4.1 Nutritional data (mean ± SEM) of control and bile-diverted rats on standard chow (14 en% fat) and high-fat chow (35
en% fat).
Category n Food intake(g day-1)
Fat intake(g day-1)
Fecal fat(g day-1)
Net fat uptake(g day-1)
Fat absorption(% intake)
Standard chowControl rats 9 18.9 ± 0.7 1.04 ± 0.04 0.04 ± 0.00 1.00 ± 0.04 96.7 ± 0.2Bile-diverted 11 21.2 ± 0.7# 1.17 ± 0.03# 0.15 ± 0.01* 1.01 ± 0.04 87.2 ± 0.9*
High-fat chowControl rats 9 15.7 ± 0.7 1.92 ± 0.10 0.13 ± 0.01 1.80 ± 0.11 93.2 ± 0.4Bile-diverted 6 21.9 ± 1.6§ 2.67 ± 0.20§ 1.15 ± 0.12* 1.51 ± 0.10 53.9 ± 3.9*
Symbols indicate significant difference within the same dietary group: #, P<0.05; §, P<0.01; * P<0.001.
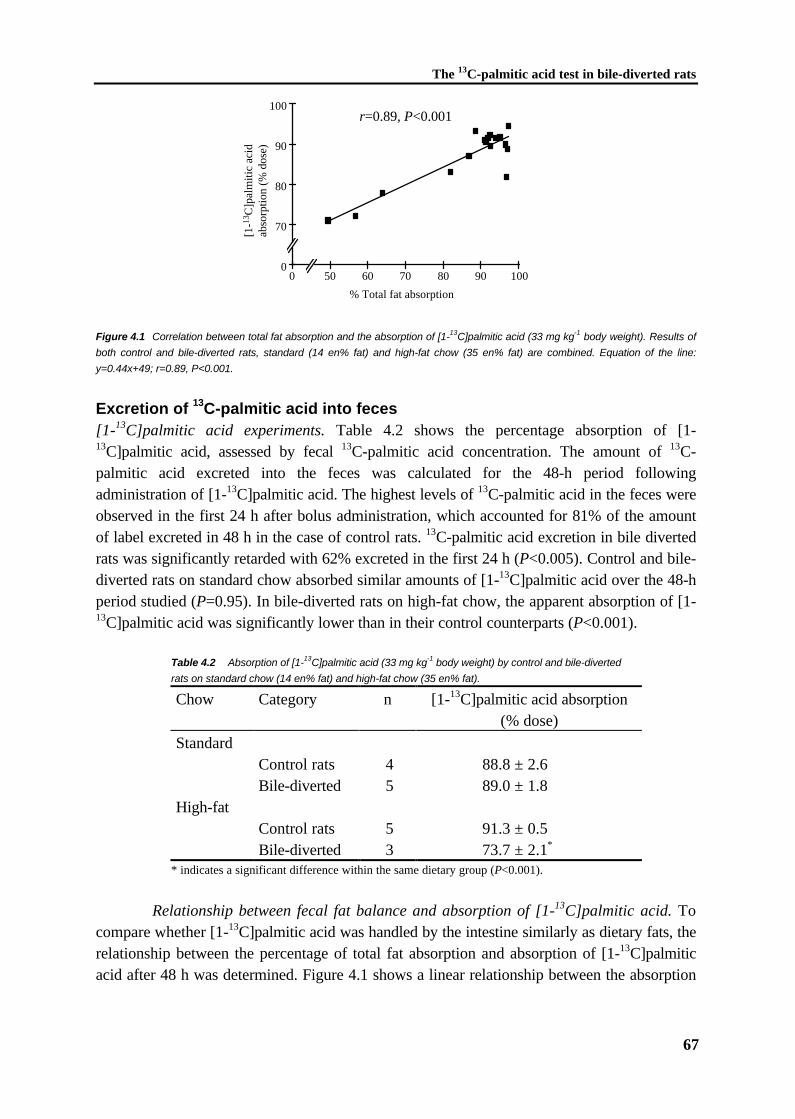
The 13C-palmitic acid test in bile-diverted rats
67
r=0.89, P<0.001
0
70
80
90
100
0 50 60 70 80 90 100
% Total fat absorption
[1-13
C]p
alm
itic
acid
abso
rptio
n (%
dos
e)
Figure 4.1 Correlation between total fat absorption and the absorption of [1-13C]palmitic acid (33 mg kg-1 body weight). Results of
both control and bile-diverted rats, standard (14 en% fat) and high-fat chow (35 en% fat) are combined. Equation of the line:
y=0.44x+49; r=0.89, P<0.001.
Excretion of 13C-palmitic acid into feces[1-13C]palmitic acid experiments. Table 4.2 shows the percentage absorption of [1-13C]palmitic acid, assessed by fecal 13C-palmitic acid concentration. The amount of 13C-palmitic acid excreted into the feces was calculated for the 48-h period followingadministration of [1-13C]palmitic acid. The highest levels of 13C-palmitic acid in the feces wereobserved in the first 24 h after bolus administration, which accounted for 81% of the amountof label excreted in 48 h in the case of control rats. 13C-palmitic acid excretion in bile divertedrats was significantly retarded with 62% excreted in the first 24 h (P<0.005). Control and bile-diverted rats on standard chow absorbed similar amounts of [1-13C]palmitic acid over the 48-hperiod studied (P=0.95). In bile-diverted rats on high-fat chow, the apparent absorption of [1-13C]palmitic acid was significantly lower than in their control counterparts (P<0.001).
Table 4.2 Absorption of [1-13C]palmitic acid (33 mg kg-1 body weight) by control and bile-diverted
rats on standard chow (14 en% fat) and high-fat chow (35 en% fat).
Chow Category n [1-13C]palmitic acid absorption(% dose)
StandardControl rats 4 88.8 ± 2.6Bile-diverted 5 89.0 ± 1.8
High-fatControl rats 5 91.3 ± 0.5Bile-diverted 3 73.7 ± 2.1*
* indicates a significant difference within the same dietary group (P<0.001).
Relationship between fecal fat balance and absorption of [1-13C]palmitic acid. Tocompare whether [1-13C]palmitic acid was handled by the intestine similarly as dietary fats, therelationship between the percentage of total fat absorption and absorption of [1-13C]palmiticacid after 48 h was determined. Figure 4.1 shows a linear relationship between the absorption

Chapter 4
68
of total fat and the absorption of [1-13C]palmitic acid (r=0.89, P<0.001; equation of the line: y= 0.44 x + 49).
Tri-[1-13C]palmitoylglycerol experiments. In control experiments with tri-[1-13C]palmitoylglycerol we verified if lipolyis in bile-diverted rats was not impaired. In bile-diverted rats on standard chow, the absorption of tri-[1-13C]palmitoylglycerol was virtuallyidentical to that of [1-13C]palmitic acid (88.3 ± 1.7% and 89.0 ± 1.8%, respectively). In bile-diverted rats on high fat chow, again, the absorption of tri-[1-13C]palmitoylglycerol was notsignificantly lower than the absorption of [1-13C]palmitic acid (80.8 ± 5.1% and 93.6 ± 6.4%,respectively). These results indicate that lipolysis per se is not impaired in the bile-diverted rat.
Plasma 13C-palmitic acid concentrations[1-13C]palmitic acid experiments. Figure 4.2A shows the time course patterns of 13C-palmiticacid appearance in plasma after intraduodenal administration of [1-13C]palmitic acid to rats onstardard chow (14 en% fat). After administration of [1-13C]palmitic acid to control rats,plasma 13C-palmitic acid concentrations increased within 1 h, reaching a maximum value of 58± 21% dose L-1 plasma at 2 h after bolus administration (Figure 4.2A). Upon bile diversion,plasma 13C-palmitic acid concentrations were significantly lower than in controls (P<0.05). Amaximum value of 10 ± 2% dose L-1 plasma was obtained at 6 h (Figure 4.2A). Figure 4.2Bshows the time course patterns of 13C-palmitic acid appearance in plasma after intraduodenaladministration of [1-13C]palmitic acid to rats on high-fat chow (35 en% fat). Afteradministration of [1-13C]palmitic acid to control rats on high-fat chow, plasma 13C-palmiticacid concentrations increased within 1 h after administration of the bolus and reached amaximum value of 55 ± 7% dose L-1 plasma at 3 h (Figure 4.2B). Upon bile diversion, plasma13C-palmitic acid concentrations were significantly lower (P<0.05) when compared to thevalues obtained in the controls rats (Figure 4.2B) and a maximum value of 16 ± 6% dose L-1
plasma was obtained after 6 h.
Time (h)
A B
Time (h)
0
20
40
60
80
0 1 2 3 4 5 6 24
Time (h)
Plas
ma
13C
16:0
(%
dos
e L
-1)
* **
*#
#
*
0
20
40
60
80
0 1 2 3 4 5 6 24
Time (h)
Plas
ma
13C
16:0
(%
dos
e L
-1)
* *# # #
Figure 4.2 Time courses of 13C-palmitic acid concentration in plasma of rats fed (A) standard chow (14 en% fat) and (B) high-fat
chow (35 en% fat) after intraduodenal administration of [1-13C]palmitic acid (33 mg kg-1 body weight). (�) Control rats, (±) bile-
diverted rats. Symbols indicate significant difference between control and bile-diverted rats after [1-13C]palmitic acid administration
(* P<0.05, # P<0.01).
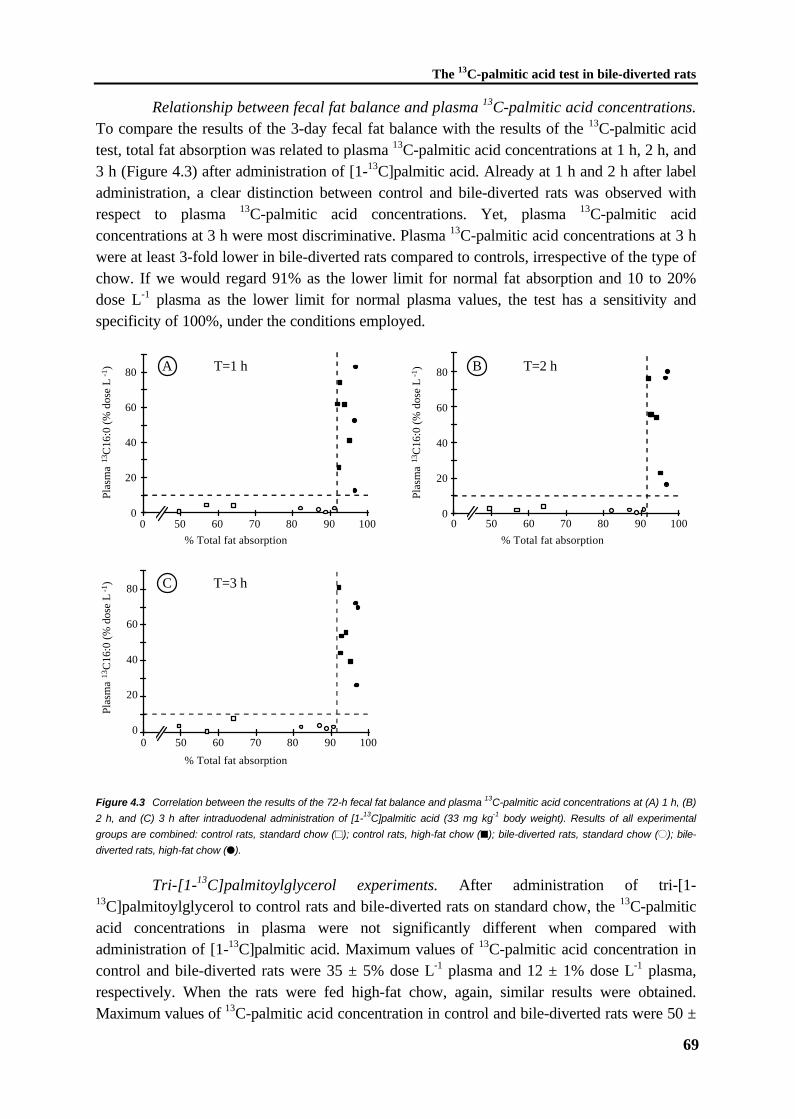
The 13C-palmitic acid test in bile-diverted rats
69
Relationship between fecal fat balance and plasma 13C-palmitic acid concentrations.To compare the results of the 3-day fecal fat balance with the results of the 13C-palmitic acidtest, total fat absorption was related to plasma 13C-palmitic acid concentrations at 1 h, 2 h, and3 h (Figure 4.3) after administration of [1-13C]palmitic acid. Already at 1 h and 2 h after labeladministration, a clear distinction between control and bile-diverted rats was observed withrespect to plasma 13C-palmitic acid concentrations. Yet, plasma 13C-palmitic acidconcentrations at 3 h were most discriminative. Plasma 13C-palmitic acid concentrations at 3 hwere at least 3-fold lower in bile-diverted rats compared to controls, irrespective of the type ofchow. If we would regard 91% as the lower limit for normal fat absorption and 10 to 20%dose L-1 plasma as the lower limit for normal plasma values, the test has a sensitivity andspecificity of 100%, under the conditions employed.
0
20
40
60
80
0 50 60 70 80 90 100
Plas
ma
13C
16:0
(%
dos
e L
-1) A T=1 h
0
20
40
60
80
0 50 60 70 80 90 100
Plas
ma
13C
16:0
(%
dos
e L
-1) B T=2 h
0
20
40
60
80
0 50 60 70 80 90 100
Plas
ma
13C
16:0
(%
dos
e L
-1) C T=3 h
% Total fat absorption
% Total fat absorption % Total fat absorption
Figure 4.3 Correlation between the results of the 72-h fecal fat balance and plasma 13C-palmitic acid concentrations at (A) 1 h, (B)
2 h, and (C) 3 h after intraduodenal administration of [1-13C]palmitic acid (33 mg kg-1 body weight). Results of all experimental
groups are combined: control rats, standard chow (~); control rats, high-fat chow (�); bile-diverted rats, standard chow (±); bile-
diverted rats, high-fat chow (�).
Tri-[1-13C]palmitoylglycerol experiments. After administration of tri-[1-13C]palmitoylglycerol to control rats and bile-diverted rats on standard chow, the 13C-palmiticacid concentrations in plasma were not significantly different when compared withadministration of [1-13C]palmitic acid. Maximum values of 13C-palmitic acid concentration incontrol and bile-diverted rats were 35 ± 5% dose L-1 plasma and 12 ± 1% dose L-1 plasma,respectively. When the rats were fed high-fat chow, again, similar results were obtained.Maximum values of 13C-palmitic acid concentration in control and bile-diverted rats were 50 ±

Chapter 4
70
20% dose L-1 plasma and 6 ± 4% dose L-1 plasma, respectively. These results are inaccordance with results on fecal excretion of 13C-palmitic acid reported above, and furtherunderline that lipolysis is not impaired in the bile-diverted rat.
Discussion
In the present study we characterized a rat model with a defined cause of fat malabsorptiondue to bile diversion and we investigated the potency of the [1-13C]palmitic acid test to detectthis fat malabsorption. As no bile is available in the intestinal tract of bile-diverted rats, thisanimal model is useful for determining the contribution of bile (components) to the process ofintestinal fat absorption. Although the role of bile for efficient fat absorption is well-established, its quantitative importance has been a matter of debate and is likely dependent onthe amount and composition of dietary fats [16-18].
Total dietary fat absorption was examined in chronically bile-diverted rats onstandard chow (14 en% fat) and on high-fat chow (35 en% fat). In control rats on eitherstandard or high-fat chow, the absorption of dietary fats was very efficient and varied from92% to 97%. This percentage fat absorption is in accordance to that found in healthy humans[20]. Total fat absorption was significantly decreased in bile-diverted rats. Bile-diverted ratson standard chow still absorbed 87% of their dietary fats. An explanation could be theformation of liquid crystalline vesicles in the intestinal lumen, as proposed by Carey et al. [20].They suggested that liquid crystalline vesicles are formed, when the amount of fat in theaqueous intestinal phase is relatively high compared to the amount of bile. These vesicles mayplay an important role in in the uptake of fats by enterocytes in certain disease states [29].However, bile-diverted rats on high-fat chow absorbed only 54% of their dietary fats,indicating that at relatively high dietary fat intake, surface increase and formation of vesiclesare not sufficient to restore fat absorption completely.
Yet, in spite of the decrease in percentage of fat absorption in bile-diverted rats, therats managed to maintain a similar net fat uptake by increasing their food intake. This effectwas observed on either of the two diets. It has been observed previously that after bilediversion rats increase their food intake in order to maintain an adequate energy balance [24].It is intriguing to speculate which physiological stimulus mediates the adaptation of foodintake. A possible candidate is apolipoprotein A-IV. Synthesis of apo A-IV is stimulated upontransport of absorbed lipid via chylomicrons in lymph [30-32]. Evidence of decreasedchylomicron assembly due to interference of biliary phospholipid availability by manipulationwith cholestyramine or dietary zinc was presented previously [21,22]. Impaired chylomicronassembly in bile-diverted rats would decrease concentrations of apo A-IV in plasma, resultingin enhanced food intake [33].
Using this animal model, we found that at 1 h, 2 h, and 3 h after administration of [1-13C]palmitic acid, plasma 13C-palmitic acid concentrations clearly differentiated betweencontrol rats and chronically bile-diverted rats (Figure 4.3). Using 10% dose L-1 plasma as thelower limit of normal plasma values and 91% as the lower limit of normal fat absorption, thetest had a sensitivity and specificity of 100% under the test conditions used. The results were

The 13C-palmitic acid test in bile-diverted rats
71
essentially similar on standard and high-fat chow, emphasizing the potency of the 13C-palmiticacid absorption test. It is interesting to hypothesize on the low plasma 13C-palmitic acidconcentrations in bile-diverted rats when compared to control rats. It is not likely that thewhole effect is solely due to impaired solubilization, because fat absorption in bile-divertedrats on standard chow is still rather efficient. However, the effect of dimished solubilizationcould be enhanced by the absence of stimulatory effects of biliary phospholipids on assemblyof intestinal chylomicrons [21-23]. If chylomicron assembly is impaired, 13C-palmitic acid willappear in plasma to a lesser extent and on a slower time scale.
Time course patterns of plasma 13C-palmitic acid concentrations in control and bile-diverted rats were considerably different (Figure 4.2). In control rats plasma 13C-palmitic acidconcentrations increased rapidly and peak values were observed after 1 to 3 h. In bile-divertedrats plasma 13C-palmitic acid concentrations continuously increased up to 5 or 6 h but did notreach the high values obtained in the control rats. Brand & Morgan [34] showed that fatabsorption occurs largely from the upper small intestine in control rats, whereas, in theabsence of bile lower small intestine is also involved. Presumably, the absorptive reserve of thedistal small intestine is called upon in the case of bile diversion and much of the fat whichfailed to enter the proximal intestinal mucosa is absorbed more distally [35]. The delayed timecourse patterns of plasma 13C-palmitate concentrations upon bile diversion could also be dueto decreased intestinal motility. In support of this explanation, fecal 13C-palmitate excretionwas significantly retarded in bile-diverted rats compared to controls in the first 24 h afteradministration of [1-13C]palmitate. A cyclic pattern of motor activity known as the migratingmotor complex occurs in dogs, humans, and most other mammals during fasting [36-38].Feeding interrupts the migrating motor complex and induces a different pattern of intermittentcontractile activity. However, the migrating motor complex activity does not seem to beaffected by bile diversion in rats [34] and dogs [39].
The fat bolus containing the [1-13C]palmitic acid was administered to the ratsintraduodenally. Although oral administration would have been more physiological,intraduodenal administration has the benefit that the results are not influenced by gastricemptying. Intra-individual and inter-individual variation of gastric emptying have been shownto vary widely in humans [40,41] and would probably affect the outcomes of the study. This isalso a likely explanation for the high specificity and sensitivity of the test in our experiments.An adventitious circumstance is that experiments run for a shorter period when the bolus isadministered intraduodenally, as gastric emptying of oils delays the process of fat absorptionwith approximately 2 to 3 h [42,43].
In order to determine if stable isotopically-labeled fats are handled within the bodysimilar as those of dietary origin, the percentage of total fat absorption was compared to theabsorption of [1-13C]palmitic acid. Under physiological circumstances, palmitic acid isconsumed in the diet in the form of mixed triacylglycerols predominantly esterified at the sn-1and sn-2 positions [44]. Hydrolysis of dietary triacylglycerols in the intestinal lumen bypancreatic lipase and other enzymes, such as carboxyl ester hydrolase, results in the generationof 2-monoacylglycerols and fatty acids. After interaction with bile components and theformation of mixed micelles, the hydrolyzed fats can be absorbed. The absorption of dietaryfats was significantly correlated with the absorption of [1-13C]palmitic acid, assessed by fecal

Chapter 4
72
13C-palmitic acid concentration (Figure 4.1). Thus, it seems that the fate of [1-13C]palmiticacid with respect to absorption reflects the fate of total mass of dietary fats under theexperimental circumstances of this study. It has to be noted that the line of correlation doesnot cut the origin of the plot, and that the slope of the line is smaller than unity. Apparently,[1-13C]palmitic acid is preferentially absorbed when compared to dietary fats, which may bedue to the fact that only tracer amounts of [1-13C]palmitic acid were administered to the rats ina specific, soluble form.
Absorption of tri-[1-13C]palmitoylglycerol and plasma 13C-palmitic acidconcentrations were determined to ascertain that lipolysis was not affected in the bile-divertedrat model. In case of impaired lipolysis, tri-[1-13C]palmitoylglycerol could only be hydrolyzedpartially, resulting in decreased plasma 13C-palmitic acid concentrations when compared toadministration of [1-13C]palmitic acid. In addition, fecal 13C-palmitic acid concentrationswould be expected to be increased in analogy to studies involving drug-induced impairment inlipolysis [45]. In both control and bile-diverted rats on standard chow, the plasma 13C-palmiticacid concentration curves after administration of either [1-13C]palmitic acid or tri-[1-13C]palmitoylglycerol were not significantly different (Figure 4.2). After administration of tri-[1-13C]palmitoylglycerol, fecal excretion of 13C-palmitic acid was not significantly increasedcompared with administration of [1-13C]palmitic acid. Based on these observations, weconclude that lipolysis is not a rate-limiting step in this experimental model, in accordance toprevious observations [18,29]. Hamilton et al. [18] reported that in bile-diverted ratsabsorption of free fatty acids was equal to the absorption of triacylglycerols with respect toboth palmitic acid and stearic acid. Similarly, Porter et al. [29] reported that lipolysis wasunimpaired in bile fistula man.
In summary, we show in a rat model of fat malabsorption due to bile deficiency thatpercentage of dietary fat absorption depends on the presence of bile in the intestinal lumen.With the use of this rat model, the [1-13C]palmitic acid absorption test, based on thequantification of plasma 13C-palmitic acid concentrations, was sensitive enough to discriminatebetween total fat absorption above 91% (control rats) and below 91% (bile-diverted rats).These observations underline the potency of the 13C-palmitic acid absorption test incombination with the technique of gas chromatography combustion isotope ratio massspectrometry to detect disorders in intestinal solubilization. Application of the test in clinicalabsorption studies may allow a differentiated diagnosis and subsequent specific treatment.
References
1. Carey MC, Hernell O. Digestion and absorption of fat. Sem Gastrointest Dis 1992;3:189-208.
2. Sardesai VM. The essential fatty acids. Nutr Clin Pract 1992;7:179-186.3. Shiau Y-F. Lipid digestion and absorption. In: Johnson LR, ed. Physiology of the
Gastrointestinal Tract. 2nd Ed. New York: Raven Press, 1987:1527-1556.4. Staggers JE, Hernell O, Stafford RJ, Carey MC. Physical-chemical behavior of dietary and
biliary lipids during intestinal digestion and absorption. 1. Phase behavior and aggregation

The 13C-palmitic acid test in bile-diverted rats
73
states of model lipid systems patterned after aqueous duodenal contents of healthy adulthuman beings. Biochemistry 1990;29:2028-2040.
5. Hernell O, Staggers JE, Carey MC. Physical-chemical behavior of dietary and biliary lipidsduring intestinal digestion and absorption. 2. Phase analysis and aggregation states ofluminal lipids during duodenal fat digestion in healthy adult human beings. Biochemistry1990;29:2041-2056.
6. McClean P, Harding M, Coward WA, Green MR, Weaver LT. Measurement of fatdigestion in early life using a stable isotope breath test. Arch Dis Child 1993;69:366-370.
7. Vantrappen GR, Rutgeerts PJ, Ghoos YF, Hiele MI. Mixed triglyceride breath test: Anoninvasive test of pancreatic lipase activity in the duodenum. Gastroenterology1989;96:1126-1134.
8. Lembcke B, Braden B, Caspary WF. Exocrine pancreatic insufficiency: accuracy andclinical value of the uniformly labelled 13C-hiolein breath test. Gut 1996;39:668-674.
9. Watkins JB, Klein PD, Schoeller DA, Kirschner BS, Park R, Perman JA. Diagnosis anddifferentiation of fat malabsorption in children using 13C-labeled lipids: trioctanoin, trioleinan palmitic acid breath tests. Gastroenterology 1982;82:911-917.
10. Murphy JL, Jones AE, Stolinski M, Wootton SA. Gastrointestinal handling of [1-13C]palmitic acid in healthy controls and patients with cystic fibrosis. Arch Dis Child1997;76:425-427.
11. Odeka EB, Miller V. Evaluation of fat utilization in paediatric Crohn's disease using 13C-labelled fat. J Pediatr Gastroenterol Nutr 1995;21:430-434.
12. Murphy JL, Jones A, Brookes S, Wootton SA. The gastrointestinal handling andmetabolism of [1-13C]palmitic acid in healthy women. Lipids 1995;30:291-298.
13. Kalivianakis M, Verkade HJ, Stellaard F, Van der Werf M, Elzinga H, Vonk RJ. The 13C-mixed triglyceride breath test in healthy adults: determinants of the 13CO2 response. Eur JClin Invest 1997;27:434-442.
14. Goodman KJ, Brenna JT. High sensitivity tracer detection using high-precision gaschromatography-combustion isotope ratio mass spectrometry and highly enriched [U-13C]-labeled precursors. Anal Chem 1992;64:1088-1095.
15. Pedersen NT, Jorgensen BB, Rannem T. The [14C]-triolein breath test is not valid as a testof fat absorption. Scand J Clin Lab Invest 1991;51:699-703.
16. Christensen MS, Müllertz A, Hoy C. Absorption of triglycerides with defined or randomstructure by rats with biliary and pancreatic diversion. Lipids 1995;30:521-526.
17. Gallagher N, Webb J, Dawson AM. The absorption of 14C oleic acid and 14C triolein inbile fistula rats. Clin Sci 1965;29:73-82.
18. Hamilton JD, Webb JPW, Dawson AM. The absorption of tristearin and stearic acid andtripalmitin and palmitic acid. Studies on the rate-limiting steps in rats. Biochim BiophysActa 1969;176:27-36.
19. Graham DY, Sackman JW. Mechanism of increase in steatorrhea with calcium andmagnesium in exocrine pancreatic insufficiency: an animal model. Gastroenterology1982;83:638-644.
20. Carey MC, Small DM, Bliss CM. Lipid digestion and absorption. Ann Rev Physiol1983;45:651-677.

Chapter 4
74
21. Ahn J, Koo SI. Intraduodenal phosphatidylcholine infusion restores the lymphaticabsorption of vitamin A and oleic acid in zinc-deficient rats. J Nutr Biochem 1995;6:604-612.
22. Cassidy MM, Lightfoot FG, Grau L, Satchitanandum S, Vahouny GV. Lipidaccumulation in jejunal and colonic mucosa following chronic cholestyramine (Questran)feeding. Dig Dis Sci 1985;30:468-476.
23. Minich DM, Vonk RJ, Verkade HJ. Intestinal absorption of essential fatty acids underphysiological and essential fatty acid-deficient conditions. J Lipid Res 1997;38:1709-1721.
24. Kuipers F, Havinga R, Bosschieter H, Toorop GP, Hindriks FR, Vonk RJ. Enterohepaticcirculation in the rat. Gastroenterology 1985;88:403-411.
25. Lepage G, Roy CC. Direct transesterification of all classes of lipids in a one-step reaction.J Lipid Res 1986;27:114-120.
26. Eder K. Gas chromatography analysis of fatty acid methyl esters. J Chromatogr B1995;671:113-131.
27. Gutnikov G. Fatty acid profiles of lipid samples. J Chromatogr B 1995;671:71-89.28. Guo Z, Nielsen S, Burguera B, Jensen MD. Free fatty acid turnover measured using
ultralow doses of [U-13C]palmitate. J Lipid Res 1997;38:1888-1895.29. Porter HP, Saunders DR, Tytgat G, Brunser O, Rubin CE. Fat absorption in bile fistula
man; A morphological and biochemical study. Gastroenterology 1971;60:1008-1019.30. Kalogeris TJ, Rodriguez MD, Tso P. Control of synthesis and secretion of intestinal
apolipoprotein A-IV by lipid. J Nutr 1997;127:S537-S543.31. Rodriguez MD, Kalogeris TJ, Wang XL, Wolf R, Tso P. Rapid synthesis and secretion of
intestinal apolipoprotein A-IV after gastric fat loading in rats. Amer J Physiol-RegulIntegr C 1997;41:R1170-R1177.
32. Kalogeris TJ, Monroe F, Demichele SJ, Tso P. Intestinal synthesis and lymphaticsecretion of apolipoprotein A-IV vary with chain length of intestinally infused fatty acidsin rats. J Nutr 1996;126:2720-2729.
33. Tso P, Chen Q, Fujimoto K, Fukagawa K, Sakata T. Apolipoprotein A-IV: A circulatingsatiety signal produced by the small intestine. Obes Res 1995;3(suppl 5):689S-695S.
34. Brand SJ, Morgan RG. The movement of an unemulsified oil test meal and aqueous- andoil-phase markers through the intestine of normal and bile-diverted rats. Q J Exp PhysiolCogn Med Sci 1975;60:1-13.
35. Lin HC, Zhao X-T, Wang L. Fat absorption is not complete by midgut but is dependenton load of fat. Am J Physiol 1996;271:G62-G67.
36. Szurszewski JH. A migrating electric complex of the canine small intestine. Am J Physiol1969;217:1757-1763.
37. Sarna SK. Cyclic motor activity: Migrating motor complex: 1985. Gastroenterology1985;89:894-913.
38. Zenilman ME, Parodi JE, Becker JM. Preservation and propagation of cyclic myoelectricactivity after feeding in rat small intestine. Am J Physiol 1992;263:G248-G253.
39. Hughes SJ, Behrns KE, Sarr MG. Chronic bile diversion does not alter canineinterdigestive myoelectric activity. Dig Dis Sci 1993;38:1055-1061.

The 13C-palmitic acid test in bile-diverted rats
75
40. Maes BD. Measurement of gastric emptying using dynamic breath analysis. 1994;1-133.(Abstract)
41. Brophy CM, Moore JG, Christian PE, Egger MJ, Taylor AT. Variability of gastricemptying measurements in man employing standardized radiolabeled meals. Dig Dis Sci1986;31:799-806.
42. Carney BI, Jones KL, Horowitz M, Sun WM, Penagini R, Meyer JH. Gastric emptying ofoil and aqueous meal components in pancreatic insufficiency: effects of posture and onappetite. Am J Physiol 1995;268:G925-G932.
43. Meyer JH, Hlinka M, Kao D, Lake R, MacLauglin E, Graham LS, Elashoff JD. Gastricemptying of oil from solid and liquid meals: Effect of human pancreatic insufficiency. DigDis Sci 1996;41:1691-1699.
44. Padley FB, Gunstone FD, Harwood JL. Gunstone FD, Harwood JL, Padley FB, eds. Thelipid handbook. London: Chapman Hall, 1986:49-170.
45. Isler D, Moeglen C, Gains N, Meier MK. Effect of the lipase inhibitor orlistat and ofdietary lipid on the absorption of radiolabelled triolein, tri-gamma-linolenin and tripalmitinin mice. Br J Nutr 1995;73:851-862.


CHAPTER 5
The 13C-palmitic acid test for detection of mild fatmalabsorption in healthy adults on calcium
supplementation
M. Kalivianakis, F. Stellaard, R. Havinga, H. Elzinga,R.J. Vonk, E.T.H.G.J. Oremus, H.J. Verkade

Chapter 5
78
CHAPTER 5
The 13C-palmitic acid test for detection of mild fatmalabsorption in healthy adults on calcium
supplementation
Abstract
Background & Aims: Recently we developed a [1-13C]palmitic acid absorption test for thedetection of fat malabsorption in rats with chronic bile diversion. In the present study inhealthy human adults we investigated whether this test was sensitive enough to detect mild fatmalabsorption induced by dietary supplementation of calcium carbonate. Methods: After oralsupplementation of [1-13C]palmitic acid (10 mg kg-1) to 10 healthy adults, breath and plasmasamples were obtained for 8 h and feces was collected for 72 h. Dietary fat intake wasassessed on the basis of a 4-day dietary record. After collection of feces, volunteers weresupplemented with 1000 mg calcium twice daily for 1 week, after which the [1-13C]palmiticacid experiment was repeated. Results: Percentage of total fat absorption in healthy volunteerson their habitual diets was (mean ± SEM) 96.6 ± 0.6%. Daily calcium supplementation led to aslight but significant decrease in total fat absorption (94.9 ± 0.9%, P<0.05). The 8-hcumulative percentage of 13CO2 expiration decreased from 11.4 ± 1.2% under controlconditions to 10.3 ± 1.1% upon calcium supplementation (P<0.05). Yet, plasma 13C-palmiticacid concentrations were significantly higher after calcium supplementation when compared tothe control experiment. Conclusion: Dietary calcium supplementation to healthy adults leadsto a slight impairment of fat absorption. Although calcium supplementation clearly affects theoutcomes of the [1-13C]palmitic acid test, present data do not indicate that the test is sensitiveenough to reliably quantitate this degree of fat malabsorption in human adults.

The 13C-palmitic acid test in humans on calcium supplementation
79
Introduction
Adequate absorption of dietary fats by the intestine is required for supply of energy, membraneconstituents, and precursors for the formation of hormones or inflammatory mediators [1-4].In Western diets, triacylglycerols composed of long-chain fatty acids constitute 92 to 96% ofdietary fats [2]. The absorption of these fats involves several processes. Firstly, lipolysis, bylipolytic enzymes originating predominantly from the pancreas, leads to hydrolysis oftriacylglycerols into fatty acids and 2-monoacylglycerols. Secondly, during the process ofsolubilization, mixed micelles are formed, consisting of bile acids, phospholipids, cholesteroland the products of lipolysis (free fatty acids and monoacylglycerol) [5]. The mixed micellesare thought to act as the physiological transport vehicles of lipolytic products from dietary fatsin the intestinal lumen. Finally, the fatty acids and monoacylglycerols are translocated acrossthe intestinal epithelium, reassembled into chylomicrons and secreted into the lymph [2,6,7].
The efficiency of intestinal fat absorption in patients is routinely determined by meansof a fat balance, requiring detailed analysis of daily fat intake and the complete recovery offeces for 72 h. However, the fat balance does not provide information on the etiology of fatmalabsorption, i.e. impaired lipolysis or solubilization. This feature limits its use for definingoptimal treatments for patients. One approach to address specifically the causes of fatmalabsorption involves the determination of (mal)absorption of stable isotopically-labeled fats.Recently, we described the results of the application of 13C-labeled linoleic acid [8] and ofpalmitic acid [9] for detection of fat malabsorption due to chronic bile diversion in rats. Inthese studies we observed that, after intraduodenal administration of the label, plasma 13C-lipidconcentrations reflect the absorption efficiency of dietary lipids. The present study wasdesigned to determine whether the [1-13C]palmitic acid test is sensitive enough to detect aslight fat malabsorption in healthy adults.
Under physiological conditions, healthy individuals excrete approximately 4-6 g day-1
of fat via the feces [10], which generally means that over 96% of the dietary fats entering theintestinal lumen is absorbed [10]. Even under physiological circumstances, non-lipidcomponents in the diet, such as calcium, have been demonstrated to interfere with the efficientabsorption of the lipids [11-14]. Oral calcium supplementation in healthy subjects has beenreported to increase fat excretion via the feces in a dose-dependent fashion, presumably due tointestinal precipitation of bile salts and/or formation of insoluble calcium-fatty acid complexes,leading to impaired solubilization [13-17]. Thus, oral calcium supplementation in humansseems to be a reproducible method to induce a slight fat malabsorption due to impairedsolubilization of long-chain fatty acids.
The aim of the present study was to investigate whether the [1-13C]palmitic acidabsorption test could detect a mild degree of fat malabsorption in humans. Fat malabsorptionwas achieved by supplementation of calcium. We investigated whether the absorptionefficiency of dietary fats was mildly decreased upon calcium supplementation for 1 week tohealthy adults and whether this possible effect could be quantified after oral ingestion ofadministered [1-13C]palmitic acid. Quantification was achieved by determination of plasma13C-palmitic acid concentrations and breath 13CO2 concentrations.

Chapter 5
80
Materials and methods
Human volunteers10 healthy students (7 females, 3 males) with a mean ± SEM age of 22 ± 0.3 y and a bodymass index of 20.5 ± 0.1 kg m-2 participated in the studies. The volunteers were healthyaccording to medical histories and showed no symptoms of diarrhea, fat malabsorption, orgastrointestinal complaints. The study protocol was approved by the Medical EthicsCommittee of the University Hospital Groningen.
Study protocolEach subject completed two tests with [1-13C]-labeled palmitic acid separated by an interval ofone week. [1-13C]palmitic acid was purchased from Isotec Inc. (Matheson, USA) and was99% 13C-enriched. The subjects were asked to maintain their usual dietary habits during thetotal experimental period of 11 days. The subjects were instructed to avoid consumingnaturally 13C-enriched foods (e.g. corn products, pineapple, cane sugar) for at least two daysprior to and during each three-day study period. The same pre-selected test meal wasconsumed throughout both tests to limit any effect that diet may have on the metabolism of [1-13C]palmitic acid. During each test, intake of nutrients was calculated from 4-day consecutivefood diaries by a clinical dietitian using The Netherlands Nutrients Table “NEVO” 1993.
Subjects started with a control experiment during which they had their habitualdietary calcium intake. After an overnight fast, the subjects consumed [1-13C]palmitic acid at adose of 10 mg kg-1 body weight as part of a controlled standard test meal consisting of 2 slicesof wheat bread, 20 g butter, 1 boiled chicken egg, 25 g cheese or liver-pie according to theirpersonal preference, 150 ml orange juice, and 150 g full fat yogurt (3500 kJ; 37 g fats, 94 gcarbohydrates, 32 g proteins). Butter was used as a vehicle to administer the [1-13C]palmiticacid. Before consuming the test meal, breath samples were collected in duplicates to provide ameasure of baseline 13C-excretion in expired CO2. After ingestion of the test meal, breathsamples were collected periodically at 30-min intervals for a period of 8 hours. A baselineblood sample (5 ml, EDTA) was collected before consuming the test meal, and was thencollected hourly during the 8-h study period. Plasma was isolated and stored frozen (-20ºC)until further analysis. A baseline feces sample was collected on the day before administrationof [1-13C]palmitic acid. Thereafter, all stools passed were collected for three days,homogenized, and frozen at -20ºC until further analysis. All the subjects were rested for theduration of the test. No additional food or liquids were permitted during the test except fornon-calorie drinks such as water and tea.
From 3 days after the first study, calcium intake was increased for 7 days by oralsupplementation of 2000 mg calcium per day in the form of calcium carbonate, divided overtwo doses (before breakfast and before dinner). At day 5 after the start of the calciumsupplementation, the [1-13C]palmitic acid test was repeated identical to the proceduresdescribed above. Before ingestion of the test meal (breakfast), calcium supplementation wasadministered.

The 13C-palmitic acid test in humans on calcium supplementation
81
The influence of the test meal on 13C-enrichment in breath 13CO2 was examined byhaving one subject completing the study, using unlabeled palmitic acid instead of [1-13C]palmitic acid.
Analytical techniquesBreath sample analysis. End expiratory breath was collected via a straw into a 10 mL tube(Exetainers; Labco Limited, High Wycombe, United Kingdom), from which aliquots weretaken to determine 13C-enrichment by means of continuous flow isotope ratio massspectrometry (Finnigan Breath MAT, Finnigan MAT GmbH, Bremen, Germany). The 13C-abundance of breath CO2 was expressed as the difference per mil from the reference standardPee Dee Belemnite limestone (δ13CPDB, ‰). The proportion of 13C-label excreted in breathCO2 was expressed as the percentage of administered 13C-label recovered per hour (% 13Cdose h-1), and as the cumulative percentage of administered 13C-label recovered over the studyperiod (cum % 13C).
Mean values of whole body CO2 excretion were measured by indirect calorimetry(Oxycon, model ox-4, Dräger, Breda, The Netherlands) at 2 separate periods of 5 minutesduring both test days. As a control, this sampling method was compared to sampling every 30min (results not shown). These results indicated that, under the test conditions chosen, themean values of the CO2 production obtained from 2 randomly chosen periods were within the95% confidence interval of the mean values obtained when sampling occurred every 30 min.
Plasma fats. Plasma fats were extracted, hydrolyzed and methylated according toLepage and Roy [18]. Resulting fatty acid methyl esters were analyzed both by gaschromatography to measure the total amount of palmitic acid and by gas chromatographycombustion isotope ratio mass spectrometry to measure the enrichment of palmitic acid. Theconcentration of 13C-palmitate in plasma was expressed as the molar percentage of the doseper liter plasma (% dose L-1).
Fecal fats. Total fecal fat excretion in human subjects was measured according to themethod of Van de Kamer et al. [19]. Feces was partly freeze-dried and mechanicallyhomogenized. Aliquots of freeze-dried feces were extracted according to the method of Blighand Dyer [20], and subsequently hydrolyzed and methylated [18]. Resulting fatty acid methylesters were analyzed by gas chromatography to calculate both total fecal fat excretion andtotal palmitic acid concentration. Fatty acid methyl esters were analyzed by gaschromatography combustion isotope ratio mass spectrometry to calculate the isotopicenrichment of palmitic acid. Total fecal fat excretion was expressed as g fat day-1 and thepercentage of total fat absorption was calculated from the daily dietary intake and the dailyfecal fat output and expressed as a percentage of the daily fat intake.
Total fat absorptionFat intake (g day ) Fat output (g day )
Fat intake (g day )
-1 -1
-1=−
×100%
A similar calculation was performed to measure the absorption of [1-13C]palmitic acid.Gas liquid chromatography. Fatty acid methyl esters were separated and quantified
by gas liquid chromatography on a Hewlett Packard gas chromatograph Model 5880 equipped
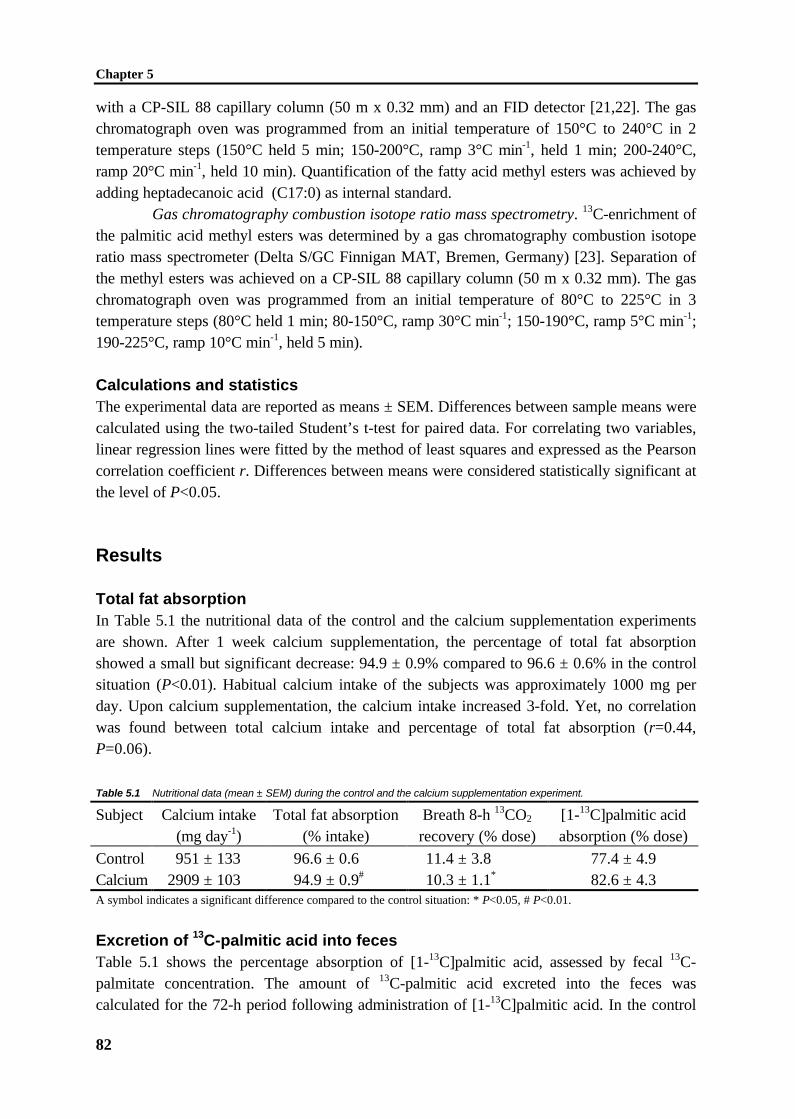
Chapter 5
82
with a CP-SIL 88 capillary column (50 m x 0.32 mm) and an FID detector [21,22]. The gaschromatograph oven was programmed from an initial temperature of 150°C to 240°C in 2temperature steps (150°C held 5 min; 150-200°C, ramp 3°C min-1, held 1 min; 200-240°C,ramp 20°C min-1, held 10 min). Quantification of the fatty acid methyl esters was achieved byadding heptadecanoic acid (C17:0) as internal standard.
Gas chromatography combustion isotope ratio mass spectrometry. 13C-enrichment ofthe palmitic acid methyl esters was determined by a gas chromatography combustion isotoperatio mass spectrometer (Delta S/GC Finnigan MAT, Bremen, Germany) [23]. Separation ofthe methyl esters was achieved on a CP-SIL 88 capillary column (50 m x 0.32 mm). The gaschromatograph oven was programmed from an initial temperature of 80°C to 225°C in 3temperature steps (80°C held 1 min; 80-150°C, ramp 30°C min-1; 150-190°C, ramp 5°C min-1;190-225°C, ramp 10°C min-1, held 5 min).
Calculations and statisticsThe experimental data are reported as means ± SEM. Differences between sample means werecalculated using the two-tailed Student’s t-test for paired data. For correlating two variables,linear regression lines were fitted by the method of least squares and expressed as the Pearsoncorrelation coefficient r. Differences between means were considered statistically significant atthe level of P<0.05.
Results
Total fat absorptionIn Table 5.1 the nutritional data of the control and the calcium supplementation experimentsare shown. After 1 week calcium supplementation, the percentage of total fat absorptionshowed a small but significant decrease: 94.9 ± 0.9% compared to 96.6 ± 0.6% in the controlsituation (P<0.01). Habitual calcium intake of the subjects was approximately 1000 mg perday. Upon calcium supplementation, the calcium intake increased 3-fold. Yet, no correlationwas found between total calcium intake and percentage of total fat absorption (r=0.44,P=0.06).
Table 5.1 Nutritional data (mean ± SEM) during the control and the calcium supplementation experiment.
Subject Calcium intake(mg day-1)
Total fat absorption(% intake)
Breath 8-h 13CO2
recovery (% dose)[1-13C]palmitic acidabsorption (% dose)
Control 951 ± 133 96.6 ± 0.6 11.4 ± 3.8 77.4 ± 4.9Calcium 2909 ± 103 94.9 ± 0.9# 10.3 ± 1.1* 82.6 ± 4.3A symbol indicates a significant difference compared to the control situation: * P<0.05, # P<0.01.
Excretion of 13C-palmitic acid into fecesTable 5.1 shows the percentage absorption of [1-13C]palmitic acid, assessed by fecal 13C-palmitate concentration. The amount of 13C-palmitic acid excreted into the feces wascalculated for the 72-h period following administration of [1-13C]palmitic acid. In the control

The 13C-palmitic acid test in humans on calcium supplementation
83
experiment and in the calcium experiment the absorption of [1-13C]palmitic acid was similar:77.4 ± 4.9% and 82.6 ± 4.3% dose, respectively (P=0.39). No correlation was found betweencalcium intake and percentage absorption of [1-13C]palmitic acid (r=0.17). A significantrelationship was observed between percentage of total fat absorption and absorption of [1-13C]palmitic acid (Figure 5.1; r=0.47, P<0.05), indicating that [1-13C]palmitic acid washandled by the intestine in a similar fashion as the mass of unlabeled dietary fats.
0 1 2 3 4 5 6
% Total fat absorption
[1-13
C]p
alm
itic
acid
abso
rptio
n (%
dos
e)
0
50
60
70
80
90
100
85 90 95 1000
r=0.47, P<0.05
Figure 5.1 Correlation between total fat absorption and the absorption of [1-13C]palmitic acid (10 mg kg-1 body weight) of 10
healthy adults in the control situation (�) and upon calcium supplementation (~) (2000 mg day-1; CaCO3). r=0.47, P<0.05.
Plasma 13C-palmitate concentrationIn the control experiment, analysis of the 13C-palmitic acid concentration in plasma samplesshowed a slow increase over the time and a maximum of 0.44 ± 0.10% dose L-1 plasma wasobtained after 5 h in the control experiment (Figure 5.2). Upon calcium supplementation the13C-palmitate concentrations in plasma were initially similar, yet after 4 h significantlyincreased when compared with the controls (P<0.05, Figure 5.2), with a maximum of 0.81 ±0.21% dose L-1 plasma obtained after 6 h, respectively.
0 1 2 3 4 5 6
Plas
ma
13C
16:0
(%
dos
e L
-1)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Time (h)
*
0 2 4 6 81 3 5 7
* *
Figure 5.2 Time courses of 13C-palmitate concentration in plasma of 10 healthy adults in the control situation (�) and upon
calcium supplementation (~) (2000 mg day-1; CaCO3) after a single oral dose of [1-13C]palmitic acid (10 mg kg-1 body weight) (*
P<0.05).
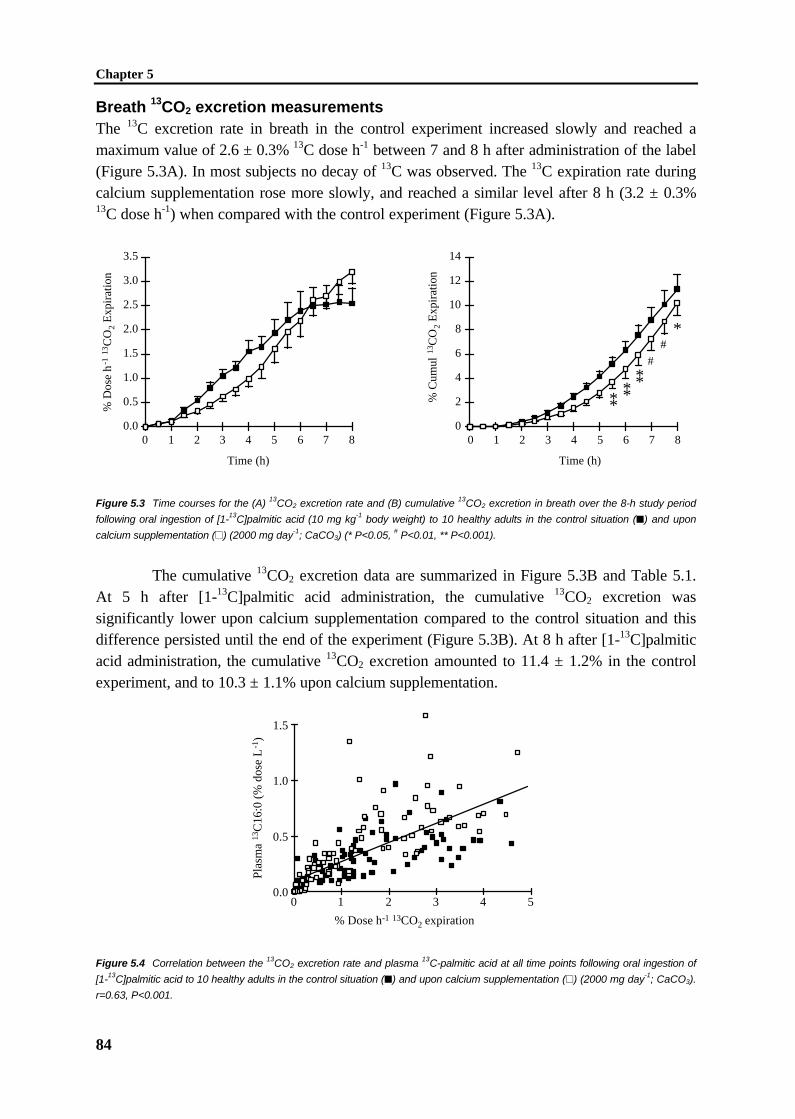
Chapter 5
84
Breath 13CO2 excretion measurementsThe 13C excretion rate in breath in the control experiment increased slowly and reached amaximum value of 2.6 ± 0.3% 13C dose h-1 between 7 and 8 h after administration of the label(Figure 5.3A). In most subjects no decay of 13C was observed. The 13C expiration rate duringcalcium supplementation rose more slowly, and reached a similar level after 8 h (3.2 ± 0.3%13C dose h-1) when compared with the control experiment (Figure 5.3A).
% D
ose
h-1
13C
O2
Exp
irat
ion
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
0 1 2 3 4 5 6 7 8
Time (h)
% C
umul
13C
O2
Exp
irat
ion
0
2
4
6
8
10
12
14
0 1 2 3 4 5 6 7 8
Time (h)
*#
#******
Figure 5.3 Time courses for the (A) 13CO2 excretion rate and (B) cumulative 13CO2 excretion in breath over the 8-h study period
following oral ingestion of [1-13C]palmitic acid (10 mg kg-1 body weight) to 10 healthy adults in the control situation (�) and upon
calcium supplementation (~) (2000 mg day-1; CaCO3) (* P<0.05, # P<0.01, ** P<0.001).
The cumulative 13CO2 excretion data are summarized in Figure 5.3B and Table 5.1.At 5 h after [1-13C]palmitic acid administration, the cumulative 13CO2 excretion wassignificantly lower upon calcium supplementation compared to the control situation and thisdifference persisted until the end of the experiment (Figure 5.3B). At 8 h after [1-13C]palmiticacid administration, the cumulative 13CO2 excretion amounted to 11.4 ± 1.2% in the controlexperiment, and to 10.3 ± 1.1% upon calcium supplementation.
0.0
0.5
1.0
1.5
0 1 2 3 4 5
Plas
ma
13C
16:0
(%
dos
e L
-1)
% Dose h-1 13CO2 expiration
Figure 5.4 Correlation between the 13CO2 excretion rate and plasma 13C-palmitic acid at all time points following oral ingestion of
[1-13C]palmitic acid to 10 healthy adults in the control situation (�) and upon calcium supplementation (~) (2000 mg day-1; CaCO3).
r=0.63, P<0.001.

The 13C-palmitic acid test in humans on calcium supplementation
85
In order to compare whether 13CO2 excretion in breath may be extrapolated directly toabsorption efficiency, the relationship between the breath 13CO2 excretion rates and plasma13C-palmitic acid concentrations was determined. A significant relationship was observedbetween breath 13CO2 excretion rates and plasma 13C-palmitic acid concentrations (Figure 5.4;r=0.63, P<0.001).
Background 13C-enrichment after an unlabeled test mealThe background enrichment of 13C was examined in breath, plasma and feces in 1 subject byperforming the control experiment and the experiment during the calcium supplementationwithout administration of the label. There was no detectable change in the plasma 13C-palmitate, breath 13CO2 and fecal 13C-palmitate over the 8-h study period in either of theexperimental settings (data not shown).
Discussion
Recently we characterized the [1-13C]palmitic acid absorption test for the detection of fatmalabsorption due to long-term bile diversion in rats [9]. In the present study we investigatedwhether this test was sensitive enough to detect mild fat malabsorption in healthy volunteers,induced by oral supplementation of 2000 mg calcium carbonate per day. It has beendemonstrated that an increased calcium intake leads to modestly increased amounts of fat inthe feces, leading to decreased percentages of total fat absorption [15,16]. Also in our study,percentage of total fat absorption was slightly but significantly decreased upon calciumsupplementation.
The absorption of dietary fats was significantly correlated with the absorption of [1-13C]palmitic acid, assessed by fecal 13C-palmitate concentrations (Figure 5.1), indicating thatthe fate of [1-13C]palmitic acid parallels the fate of total mass of dietary fats with respect toabsorption under the experimental circumstances of this study. Correlations between total fatabsorption and absorption of labeled fats have been reported before with outcomes varyingfrom a strong correlation [9,24] to absence of a correlation [25,26]. It could be that thepresentation of [1-13C]palmitic acid to the absorptive site may not always be the same, aspalmitic acid is normally hydrolyzed from dietary triacylglycerols in the gastrointestinal tract,indicating that analysis of fecal 13C-palmitate concentrations is not a representative method todetermine dietary fat absorption.
Theoretically, based on the positive correlation between total fat absorption andabsorption of [1-13C]palmitic acid, one would expect that, upon calcium supplementation, notonly total fat absorption would be decreased but also the absorption of [1-13C]palmitic acid,resulting in reduced amounts of 13C-palmitic acid in blood plasma and decreased amounts of13CO2 in breath. Indeed, in breath a small but significantly decreased amount of 13CO2 wasrecovered after 8 h upon calcium supplementation when compared with controls. Our resultswith respect to breath 13CO2 excretion during the control experiment are rather similar to whatother scientists report [25,27]: peak excretion rate of 13CO2 appears rather late (afterapproximately 6 h) and does not exceed 3% dose h-1. Only a few studies have appeared inwhich the [1-13C]palmitic acid breath test was studied in patients with disturbed fat absorption

Chapter 5
86
[26,27]. Watkins et al. [27] reported data on the [1-13C]palmitic acid breath test in patientswith known bile salt deficiency, and found a significantly lower 6-h cumulative 13CO2
expiration when compared with healthy controls.In contrast to the decrease observed in breath 13CO2 excretion upon calcium
supplementation, plasma 13C-palmitic acid concentrations after 4 h were significantly increasedupon calcium supplementation when compared with controls. The apparent contradictionbetween plasma 13C-palmitic acid concentrations and breath 13CO2 recovery suggests thatpost-absorptive metabolic changes take place. Previously, it has been reported that plasmatriacylglycerol concentrations were increased upon dietary calcium fortification of 1800 mgday-1 in humans [16]. However, it is not known whether this observation is related to theresults we obtained.
In summary, we show in healthy humans that percentage of total fat absorption canbe manipulated to a minor extent with the use of calcium administration. Calciumsupplementation resulted in a small but significant decrease of percentage of total fatabsorption due to impaired bile solubilization. After oral ingestion of [1-13C]palmitic acid, thecalcium-induced fat malabsorption was associated with a decreased cumulative expiration of13CO2 in breath but with increased 13C-palmitic acid concentrations in plasma. Present dataindicate that calcium supplementation does not only affect the overall quantity of fatabsorption, but also leads to alterations in post-absorptive metabolism. Finally, the presentdata indicate that the [1-13C]palmitic acid test is not sensitive enough to detect mild fatmalabsorption induced by calcium supplementation in human adults.
References
1. Beylot M. The use of stable isotopes and mass spectrometry in studying lipid metabolism.Proc Nutr Soc 1994;53:355-362.
2. Carey MC, Hernell O. Digestion and absorption of fat. Sem Gastrointest Dis 1992;3:189-208.
3. Sardesai VM. The essential fatty acids. Nutr Clin Pract 1992;7:179-186.4. Stryer L. Biochemistry. 2nd Ed. San Francisco, CA: Freeman, 1981:383-406.5. Shiau Y-F. Lipid digestion and absorption. In: Johnson LR, ed. Physiology of the
Gastrointestinal Tract. 2nd Ed. New York: Raven Press, 1987:1527-1556.6. Staggers JE, Hernell O, Stafford RJ, Carey MC. Physical-chemical behavior of dietary and
biliary lipids during intestinal digestion and absorption. 1. Phase behavior and aggregationstates of model lipid systems patterned after aqueous duodenal contents of healthy adulthuman beings. Biochemistry 1990;29:2028-2040.
7. Hernell O, Staggers JE, Carey MC. Physical-chemical behavior of dietary and biliary lipidsduring intestinal digestion and absorption. 2. Phase analysis and aggregation states ofluminal lipids during duodenal fat digestion in healthy adult human beings. Biochemistry1990;29:2041-2056.
8. Minich DM, Kalivianakis M, Havinga R, Kuipers F, Stellaard F, Vonk RJ, Verkade HJ. Anovel 13C-linoleic acid absorption test detects lipid malabsorption due to impairedsolubilization in rats. Gastroenterology 1997;112 (suppl.):A894(Abstract).

The 13C-palmitic acid test in humans on calcium supplementation
87
9. Kalivianakis M, Minich DM, Havinga R, Kuipers F, Stellaard F, Vonk RJ, Verkade HJ.[1-13C]palmitic acid absorption test detects fat malabsorption due to impaired intestinalsolubilization of dietary fats in rats. 1998;(Abstract).
10. Carey MC, Small DM, Bliss CM. Lipid digestion and absorption. Ann Rev Physiol1983;45:651-677.
11. Gallaher D, Olds Schneeman B. Intestinal interaction of bile acids, phospholipids, dietaryfibers, and cholestyramine. Am J Physiol 1986;250:G420-G426.
12. Lairon D, Lafont H, Vigne J-L, Nalbone G, Léonardi J, Hauton JC. Effects of dietaryfibers and cholestyramine on the activity of pancreatic lipase in vitro. Am J Clin Nutr1985;42:629-638.
13. Saunders D, Sillery J, Chapman R. Effect of calcium carbonate and aluminium hydroxideon human intestinal function. Dig Dis Sci 1988;33:409-413.
14. Govers MJAP, Termont DSML, Lapré JA, Kleibeuker JH, Vonk RJ, Meer Rv. Calcium inmilk products precipitates intestinal fatty acids and secondary bile acids and thus inhibitscolonic cytotoxicity in humans. Cancer Res 1996;56:3270-3275.
15. Welberg JWM, Monkelbaan JF, De Vries EGE, Muskiet FAJ, Cats A, Oremus ETHGJ,Boersma-van Ek W, Van Rijsbergen H, Meer Rv, Mulder NH, Kleibeuker JH. Effects ofsupplemental dietary calcium on quantitative and qualitative fecal fat excretion in man.Ann Nutr Metab 1994;38:185-191.
16. Denke MA, Fox MM, Schulte MC. Short-term dietary calcium fortification increases fecalsaturated fat content and reduces serum lipids in men. J Nutr 1996;123:1047-1053.
17. Potter SM, Kies CV, Rojhani A. Protein and fat utilization by humans as affected bycalcium phosphate, calcium carbonate, and manganese gluconate supplements. Nutrition1990;6:309-312.
18. Lepage G, Roy CC. Direct transesterification of all classes of lipids in a one-step reaction.J Lipid Res 1986;27:114-120.
19. Van de Kamer JH, Ten Bokkel Huinink H, Weyers HA. Rapid method for thedetermination of fat in feces. J Biol Chem 1949;177:347-355.
20. Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can JBiochem Physiol 1959;37:911-917.
21. Eder K. Gas chromatography analysis of fatty acid methyl esters. J Chromatogr B1995;671:113-131.
22. Gutnikov G. Fatty acid profiles of lipid samples. J Chromatogr B 1995;671:71-89.23. Guo Z, Nielsen S, Burguera B, Jensen MD. Free fatty acid turnover measured using
ultralow doses of [U-13C]palmitate. J Lipid Res 1997;38:1888-1895.24. Pedersen NT, Halgreen H. Simultaneous assessment of fat maldigestion and fat
malabsorption by a double-isotope method using fecal radioactivity. Gastroenterology1985;88:47-54.
25. Murphy JL, Jones A, Brookes S, Wootton SA. The gastrointestinal handling andmetabolism of [1-13C]palmitic acid in healthy women. Lipids 1995;30:291-298.
26. Murphy JL, Jones AE, Stolinski M, Wootton SA. Gastrointestinal handling of [1-13C]palmitic acid in healthy controls and patients with cystic fibrosis. Arch Dis Child1997;76:425-427.
27. Watkins JB, Klein PD, Schoeller DA, Kirschner BS, Park R, Perman JA. Diagnosis anddifferentiation of fat malabsorption in children using 13C-labeled lipids: trioctanoin, trioleinan palmitic acid breath tests. Gastroenterology 1982;82:911-917.


CHAPTER 6
Fat malabsorption in cystic fibrosis patients onenzyme replacement therapy is due to impaired
intestinal uptake of long-chain fatty acids
M. Kalivianakis, D.M. Minich, C.M.A. Bijleveld, W.M.C. van Aalderen,F. Stellaard, M. Laseur, R.J. Vonk, H.J. Verkade
Am J Clin Nutr (1998) in press

Chapter 6
90
CHAPTER 6
Fat malabsorption in cystic fibrosis patients onenzyme replacement therapy is due to impaired
intestinal uptake of long-chain fatty acids
Abstract
Background & Aim: Pancreatic enzyme replacement therapy frequently fails to correctintestinal fat malabsorption completely in cystic fibrosis (CF) patients. The reason behindtherapy failure in these patients is unknown. We investigated whether fat malabsorption in CFpatients treated with pancreatic enzymes is caused by insufficient lipolysis of triacylglycerolsor by defective intestinal uptake of long-chain fatty acids. Methods: In 10 CF patientsreceiving their habitual pancreatic enzymes, lipolysis was determined by analysis of breath13CO2 recovery after oral ingestion of 1,3-distearoyl, 2[1-13C]octanoyl glycerol (13C-MTG).Intestinal uptake of long-chain fatty acids was determined by analysis of plasma 13C-linoleicacid concentrations after oral ingestion of 13C-linoleic acid (13C-LA). For 3 days, dietary intakewas recorded and feces was collected. Results: Fecal fat excretion ranged from 5.1 to 27.8 gday-1 (mean ± SD: 11.1 ± 7.0 g day-1) and fat absorption ranged from 79 to 93% (89 ± 5%).After ingestion of 13C-MTG no relationship was observed between breath 13CO2 recovery anddietary fat absorption (r=0.04). In contrast, a strong relationship was observed between 8-hplasma 13C-LA concentrations and dietary fat absorption (r=0.88, P<0.001). Conclusion: Ourresults suggest that continuing fat malabsorption in CF patients on enzyme replacementtherapy is not likely due to insufficient lipolytic enzyme activity, but rather due to eitherincomplete intraluminal solubilization and/or reduced mucosal uptake of long-chain fatty acids.

Impaired uptake of fats in CF patients
91
Introduction
In humans, triacylglycerols composed of long-chain fatty acids constitute 92 to 96% of dietaryfats [1]. Absorption of these fats comprises two main processes. Firstly, lipolysis, by lipolyticenzymes originating predominantly from the pancreas, leads to hydrolysis of triacylglycerolsinto fatty acids and 2-monoacylglycerols. And secondly, intestinal uptake involves theformation of mixed micelles composed of bile components and lipolytic products, followed bythe desintegration of the mixed micelles in the unstirred water layer, and the translocation ofthe lipolytic products across the intestinal epithelium [1-4].
Most CF patients have a considerable malabsorption of dietary fats due to pancreaticinsufficiency leading to impaired lipolysis [5,6]. The symptoms of pancreatic insufficiency,such as steatorrhea and poor growth, can be alleviated by oral supplementation of pancreaticenzymes. However, despite recent improvements in the pharmacokinetics of the supplements,many patients continue to experience a certain degree of steatorrhea [7-9], with fat absorptionreaching 80 to 90% of their dietary fat intake. It has not been elucidated if the remaining fatmalabsorption is due to an insufficient dosage of pancreatic enzyme replacement therapy. Thispossibility is not unlikely because a decreased pancreatic bicarbonate secretion may negativelyaffect enzyme activity by sustaining a low pH in the duodenum [10,11]. At a low duodenalpH, the release of the enzymes from the (micro)capsules is inhibited and the denaturation ofthe enzymes is stimulated [11,12]. However, it has been demonstrated that increasing thepancreatic enzyme dosages does not completely correct fat malabsorption [13]. In addition,attempts to increase lipolysis by high-strength pancreatic enzyme supplements has led to thereported association with fibrosing colonopathy [14-16].
An alternative explanation for the continuing fat malabsorption in CF patients onpancreatic enzyme replacement therapy may involve inefficient intestinal uptake of fatty acids[7,17]. Impaired uptake in CF patients can be due to an altered bile composition, decreasedbile salt secretion by the liver, bile salt precipitation, a decreased bile salt pool size, and/or bilesalt inactivation at low intestinal pH [9,17-20]. Furthermore, small bowel mucosal dysfunctionor alterations in the mucus layer contribute to inefficient intestinal uptake of long-chain fattyacids in CF patients [5,21].
The gold standard for monitoring enzyme replacement therapy is the fat balance. Adrawback of the fat balance is that it does not provide insight into the pathophysiology of fatmalabsorption. Insight into the adequacy of these separate processes (lipolysis, intestinaluptake) would enable treatment in individual patients by modulating diet therapy, pancreaticenzyme replacement therapy and supplementation of antacids and bile salts. So far, it has notbeen possible to determine whether fat malabsorption in CF patients is due to impairedlipolysis or due to impaired uptake of long-chain fatty acids. Therapeutic improvements of fatabsorption may be of benefit for CF patients, as a positive correlation has been observedbetween a good nutritional status and long-term survival or well-being of CF patients [22].
The aim of the present study was to determine whether continued fat malabsorptionencountered in pediatric CF patients on their habitual pancreatic enzyme replacement therapyresults from either insufficient lipolysis or from defective intestinal uptake of long-chain fattyacids in the lumen. We choose to measure lipolysis and uptake by two indepedent tests in CF
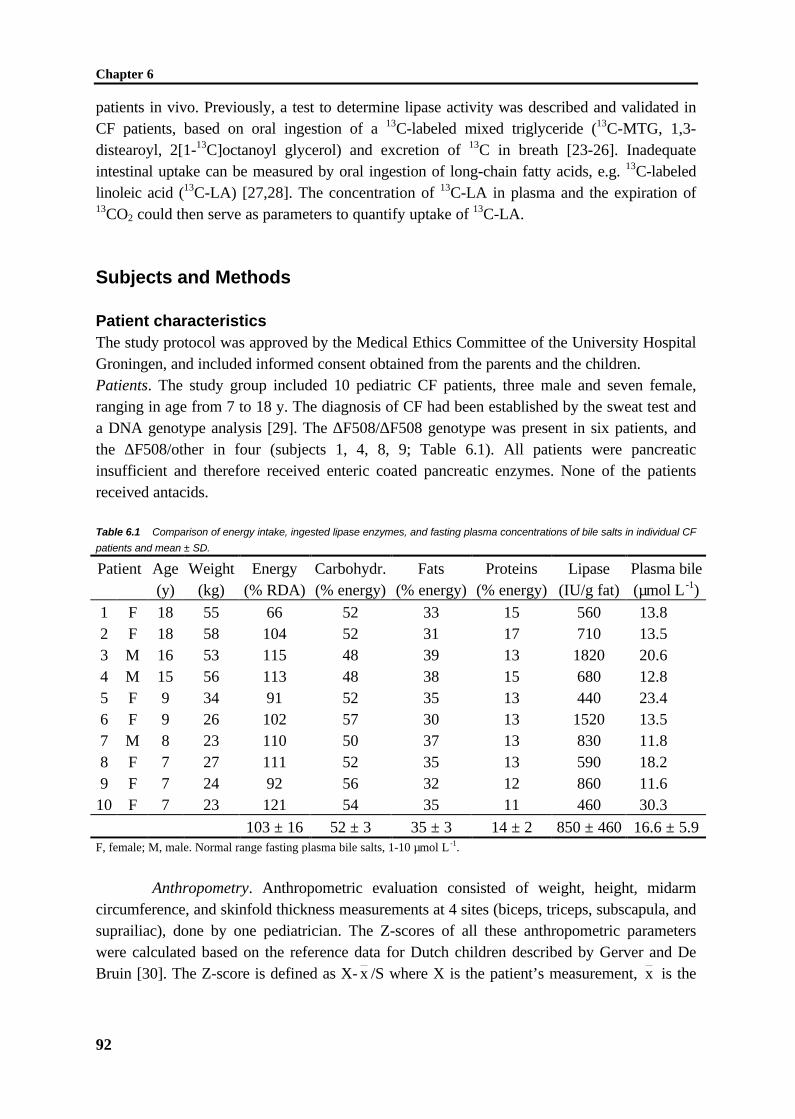
Chapter 6
92
patients in vivo. Previously, a test to determine lipase activity was described and validated inCF patients, based on oral ingestion of a 13C-labeled mixed triglyceride (13C-MTG, 1,3-distearoyl, 2[1-13C]octanoyl glycerol) and excretion of 13C in breath [23-26]. Inadequateintestinal uptake can be measured by oral ingestion of long-chain fatty acids, e.g. 13C-labeledlinoleic acid (13C-LA) [27,28]. The concentration of 13C-LA in plasma and the expiration of13CO2 could then serve as parameters to quantify uptake of 13C-LA.
Subjects and Methods
Patient characteristicsThe study protocol was approved by the Medical Ethics Committee of the University HospitalGroningen, and included informed consent obtained from the parents and the children.Patients. The study group included 10 pediatric CF patients, three male and seven female,ranging in age from 7 to 18 y. The diagnosis of CF had been established by the sweat test anda DNA genotype analysis [29]. The ∆F508/∆F508 genotype was present in six patients, andthe ∆F508/other in four (subjects 1, 4, 8, 9; Table 6.1). All patients were pancreaticinsufficient and therefore received enteric coated pancreatic enzymes. None of the patientsreceived antacids.
Table 6.1 Comparison of energy intake, ingested lipase enzymes, and fasting plasma concentrations of bile salts in individual CF
patients and mean ± SD.
Patient Age Weight Energy Carbohydr. Fats Proteins Lipase Plasma bile(y) (kg) (% RDA) (% energy) (% energy) (% energy) (IU/g fat) (µmol L -1)
1 F 18 55 66 52 33 15 560 13.82 F 18 58 104 52 31 17 710 13.53 M 16 53 115 48 39 13 1820 20.64 M 15 56 113 48 38 15 680 12.85 F 9 34 91 52 35 13 440 23.46 F 9 26 102 57 30 13 1520 13.57 M 8 23 110 50 37 13 830 11.88 F 7 27 111 52 35 13 590 18.29 F 7 24 92 56 32 12 860 11.6
10 F 7 23 121 54 35 11 460 30.3
103 ± 16 52 ± 3 35 ± 3 14 ± 2 850 ± 460 16.6 ± 5.9F, female; M, male. Normal range fasting plasma bile salts, 1-10 µmol L -1.
Anthropometry. Anthropometric evaluation consisted of weight, height, midarmcircumference, and skinfold thickness measurements at 4 sites (biceps, triceps, subscapula, andsuprailiac), done by one pediatrician. The Z-scores of all these anthropometric parameterswere calculated based on the reference data for Dutch children described by Gerver and DeBruin [30]. The Z-score is defined as X- x /S where X is the patient’s measurement, x is the

Impaired uptake of fats in CF patients
93
median value for age and sex, and S is the standard deviation of x . A negative value indicatesa value under the median reference value.
Pulmonary function. Pulmonary function was assessed by standard spirometrictechniques and was characterized by the parameters forced vital capacity, and forcedexpiratory volume in one second. For each patient, results were expressed as percentage ofpredicted (control) values for sex and height [31].
Liver function tests. Liver function had been screened during a standard routinecontrol at the time of the study using serum enzyme activities: g-glutamyl transpeptidase,aspartate transaminase, and alanine transaminase.
Diet evaluation. Intake of nutrients was calculated from 3-day consecutive fooddiaries by a clinical dietitian using The Netherlands Nutrients Table “NEVO” 1993. Intakeswere expressed as the recommended dietary allowance (RDA) for weight, age and sex (Table6.1).
13C-labeled substratesThe mixed triglyceride (1,3-distearoyl, 2[1-13C]octanoyl glycerol; S*OS) was purchased fromEuriso-Top (Saint Aubin Cedex, France) and was 99% 13C-enriched. In the original literature,the breath test performed with the use of this molecule has been named the mixed-triglyceridebreath test or the 13C-MTG breath test [23,25]. For reasons of consistency, we adhered to thisnomenclature. Uniformly labeled 13C-linoleic acid (13C-LA), obtained from Campro ScientificB.V. (Veenendaal, The Netherlands), had an enrichment exceeding 97%. 13C-LA was includedinto a gelatin capsule coated with an acid-resistant layer consisting of 4.8% cellulose acetatehydrogen phthalate in acetone.
Study protocolThe subjects were instructed to avoid consumption of naturally 13C-enriched foods (e.g. cornor corn products, pineapple, cane sugar) for at least two days prior to the study. The 13C-LAtest and the 13C-MTG test were performed on two subsequent days. On day 1, after anovernight fast, the patients received a capsule with 13C-LA (1 mg kg-1 BW), together withtheir habitual breakfast (bread, butter, ham, cheese, etc.) and pancreatic enzymes. A baselineblood sample (EDTA) was collected before consumption of breakfast, every 2 h for 8 h, and at24 h. Immediately after sampling, plasma was isolated and stored frozen (-20ºC) until furtheranalysis. Breath samples were collected in duplicate at baseline and every 30 min for 6 h. Onday 2, the patients received 13C-MTG (4 mg kg-1 BW) mixed with their habitual breakfast andpancreatic enzymes. Breath samples were collected in duplicate at baseline and every 30 minfor 6 h. The fecal fat balance and both breath tests were performed in the same 3-day period.On the day before the 13C-LA test, a feces sample was collected for baseline 13C-measurements. After consumption of the breakfast on the first day, all feces passed wascollected for three days (72 h) to determine the presence of fat malabsorption and the amountof 13C-LA excretion into the feces. Collected feces was stored at -20°C. During this period,intake of nutrients was determined from food diaries also. During the first six hours of bothtests, no additional food or liquids were permitted except for non-caloric drinks such as water

Chapter 6
94
and tea (without milk and sugar). After 6 hours, patients were allowed to have their habituallunch, including pancreatic enzymes.
Analytical techniquesBreath sample analysis. End expiratory breath was collected via a straw into a 10 ml tube(Exetainers; Labco Limited, High Wycombe, United Kingdom), from which aliquots weretaken to determine 13C-enrichment by means of continuous flow isotope ratio massspectrometry (Finnigan Breath MAT, Finnigan MAT GmbH, Bremen, Germany), conformprevious experiments [24]. The 13C-abundance of breath CO2 was expressed as the differenceper mil from the reference standard Pee Dee Belemnite limestone (δ13CPDB, ‰).
Mean values of whole body CO2 excretion were measured by indirect calorimetry(Oxycon, model ox-4, Dräger, Breda, The Netherlands) at 2 separate periods of 5 minutesduring both test days. This sampling method was compared to sampling every 30 min (resultsnot shown). The results indicated that, under the test conditions chosen, the mean values ofthe CO2 production obtained from 2 randomly chosen periods were within the 95% confidenceinterval of the mean values obtained when sampling occurred every 30 min.
Plasma fats. Plasma fats were extracted, hydrolyzed and methylated according toLepage and Roy [32]. Resulting fatty acid methyl esters were analyzed both by gaschromatography and by gas chromatography combustion isotope ratio mass spectrometry.Quantification of the resulting fatty acid methyl esters was performed with the use ofheptadecanoic acid (C17:0) as an internal standard.
Fecal fats. After thawing, feces was weighed and homogenized. Fecal fat wasdetermined according to the method of Van de Kamer et al. [33] and expressed as g fat day-1.The percentage of total fat absorption was calculated from the daily dietary fat intake and thedaily fecal fat output and expressed as a percentage of the daily fat intake.
Percentage of total fat absorptionFat intake (g day ) Fecal fat output (g day )
Fat intake (g day )
-1 -1
-1=−
×100%
Aliquots of freeze-dried feces were extracted according to the method of Bligh and Dyer [34],and subsequently hydrolyzed and methylated [32]. Resulting fatty acid methyl esters wereanalyzed by both gas chromatography and gas chromatography combustion isotope ratio massspectrometry.
Plasma and fecal bile salts. Fasting and postprandial plasma bile salts up to 8 h weredetermined by an enzymatic fluorimetric assay [35]. Results were expressed as µmol L -1
plasma. Fecal bile salts were extracted from an aliquot of dried homogenate of a 24-h fecesfraction [36] and fluorimetrically measured [35].
Gas liquid chromatography. Fatty acid methyl esters were separated and quantifiedby gas liquid chromatography on a Hewlett Packard gas chromatograph Model 5880 equippedwith a CP-SIL 88 capillary column (Chrompack; 50 m x 0.32 mm) and an FID detector[37,38]. The gas chromatograph oven was programmed from an initial temperature of 150°Cto 240°C in 2 temperature steps (150°C held 5 min; 150-200°C, ramp 3°C min-1, held 1 min;200-240°C, ramp 20°C min-1, held 10 min). Adequate separation of linoleic acid could be
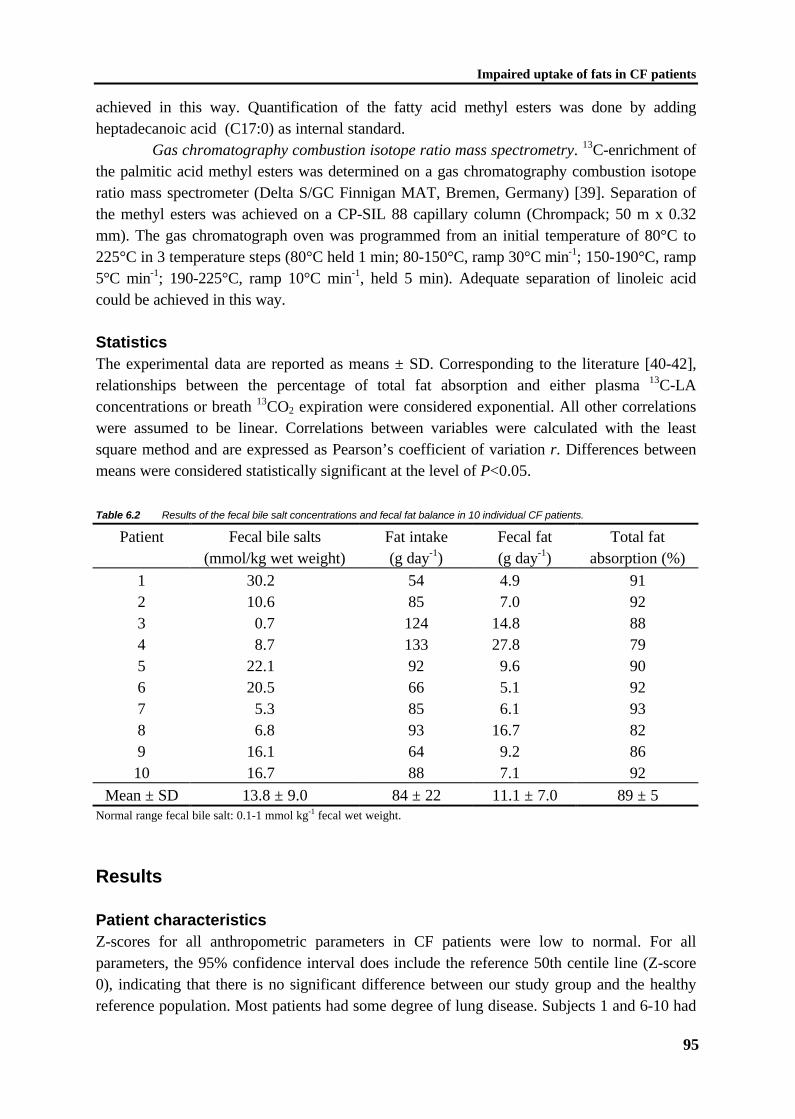
Impaired uptake of fats in CF patients
95
achieved in this way. Quantification of the fatty acid methyl esters was done by addingheptadecanoic acid (C17:0) as internal standard.
Gas chromatography combustion isotope ratio mass spectrometry. 13C-enrichment ofthe palmitic acid methyl esters was determined on a gas chromatography combustion isotoperatio mass spectrometer (Delta S/GC Finnigan MAT, Bremen, Germany) [39]. Separation ofthe methyl esters was achieved on a CP-SIL 88 capillary column (Chrompack; 50 m x 0.32mm). The gas chromatograph oven was programmed from an initial temperature of 80°C to225°C in 3 temperature steps (80°C held 1 min; 80-150°C, ramp 30°C min-1; 150-190°C, ramp5°C min-1; 190-225°C, ramp 10°C min-1, held 5 min). Adequate separation of linoleic acidcould be achieved in this way.
StatisticsThe experimental data are reported as means ± SD. Corresponding to the literature [40-42],relationships between the percentage of total fat absorption and either plasma 13C-LAconcentrations or breath 13CO2 expiration were considered exponential. All other correlationswere assumed to be linear. Correlations between variables were calculated with the leastsquare method and are expressed as Pearson’s coefficient of variation r. Differences betweenmeans were considered statistically significant at the level of P<0.05.
Table 6.2 Results of the fecal bile salt concentrations and fecal fat balance in 10 individual CF patients.
Patient Fecal bile salts(mmol/kg wet weight)
Fat intake(g day-1)
Fecal fat(g day-1)
Total fatabsorption (%)
1 30.2 54 4.9 912 10.6 85 7.0 923 0.7 124 14.8 884 8.7 133 27.8 795 22.1 92 9.6 906 20.5 66 5.1 927 5.3 85 6.1 938 6.8 93 16.7 829 16.1 64 9.2 8610 16.7 88 7.1 92
Mean ± SD 13.8 ± 9.0 84 ± 22 11.1 ± 7.0 89 ± 5Normal range fecal bile salt: 0.1-1 mmol kg-1 fecal wet weight.
Results
Patient characteristicsZ-scores for all anthropometric parameters in CF patients were low to normal. For allparameters, the 95% confidence interval does include the reference 50th centile line (Z-score0), indicating that there is no significant difference between our study group and the healthyreference population. Most patients had some degree of lung disease. Subjects 1 and 6-10 had

Chapter 6
96
normal liver biochemistry. Previously, subject 3 was diagnosed as having liver cirrhosis withportal hypertension. This patient receives ursodeoxycholic acid (750 mg day-1) and thecondition of this patient has been stable for the past few years. The bile salt concentration inplasma of this subject is in the same range as that of the other patients (Table 6.1). Analysis of3-day dietary food records is shown in Table 6.1. Energy intake of 7 patients exceeded therecommended dietary allowance. In all patients approximately 50% of the energy was derivedfrom carbohydrates, 35% from fat, and 15% from protein. Patients took pancreatic enzymesupplements in a dosage of approximately 440 - 1820 IU lipase per gram fat ingested (Table6.1).
Table 6.3 Results of the 13C-LA test, and 13C-MTG test in 10 individual CF patients.
13C-LA test 13C-MTG testBreath Plasma Feces Breath
Patient 6-h cum 13CO2
(% dose)
13C-LA at 8 h(% dose L-1)
13C-LA(% dose)
Breath 6-h cum 13CO2 (% dose)
1 11.0 1.5 0.6 2.42 1.6 1.9 0.2 9.73 2.2 0.9 1.8 15.14 1.7 0.6 0.6 5.85 0.2 1.3 0.0 30.86 3.6 2.0 0.7 11.37 1.1 1.3 0.2 40.28 3.1 0.5 1.3 28.99 2.4 0.5 0.3 11.510 0.5 1.2 0.2 8.7
Mean ± SD 2.7 ± 3.1 1.2 ± 0.5 0.6 ± 0.6 16.4 ± 12.5LA, linoleic acid; MTG, mixed triglyceride. Normal range fecal bile salt: 0.1-1 mmol kg-1 fecal wet weight.
Fat balanceIn the studied CF patients, dietary intake of fat over the 3-day period ranged from 54 to 130 gday-1, and the excretion of fat in feces ranged from 4.9 to 27.8 g day-1 (Table 6.2). Thepercentage of total fat absorption ranged from 79 to 93% (Table 6.2). Under physiologicalconditions, healthy individuals excrete approximately 4-6 g day-1 of fat via the feces [43],which generally means that over 96% of the dietary fats entering the intestinal lumen isabsorbed [43]. These observations were confirmed by experiments performed in our ownlaboratory with dietary records and feces of healthy human adults (n=13, fecal fat excretion:3.0 ± 0.9 g day-1, total fat absorption: 97 ± 2%, data not shown). Despite standard pancreaticenzyme replacement therapy, fecal fat excretion in 8 out of 10 patients was higher than 6 g fatper day, and the percentage of total fat absorption was below 96% in all patients studied.According to the prevailing reference values [43], all patients but 2 have fat malabsorption.
In studies in infants between 0 and 6 months, Fomon et al. [44] found that fecal fatexcretion per kg body weight correlated with fat intake per kg body weight. In our study weobserved a similar curvilinear correlation (r=0.71, P<0.05) despite a considerably lower intake
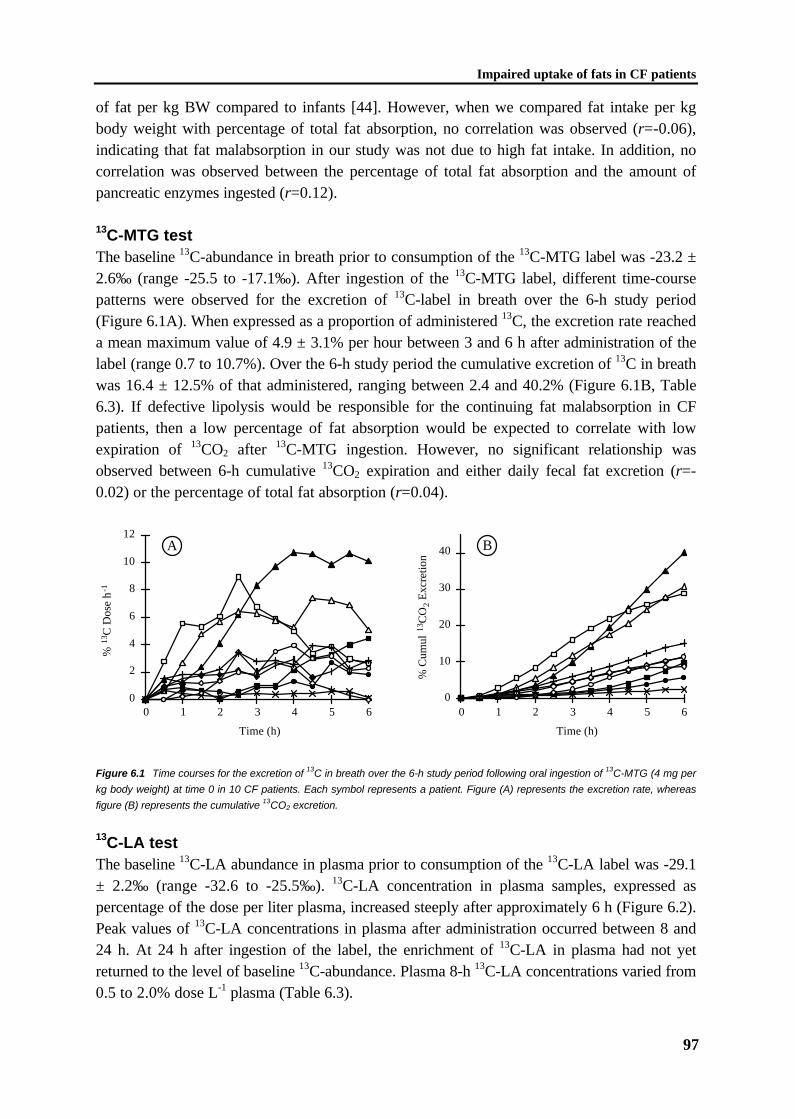
Impaired uptake of fats in CF patients
97
of fat per kg BW compared to infants [44]. However, when we compared fat intake per kgbody weight with percentage of total fat absorption, no correlation was observed (r=-0.06),indicating that fat malabsorption in our study was not due to high fat intake. In addition, nocorrelation was observed between the percentage of total fat absorption and the amount ofpancreatic enzymes ingested (r=0.12).
13C-MTG testThe baseline 13C-abundance in breath prior to consumption of the 13C-MTG label was -23.2 ±2.6‰ (range -25.5 to -17.1‰). After ingestion of the 13C-MTG label, different time-coursepatterns were observed for the excretion of 13C-label in breath over the 6-h study period(Figure 6.1A). When expressed as a proportion of administered 13C, the excretion rate reacheda mean maximum value of 4.9 ± 3.1% per hour between 3 and 6 h after administration of thelabel (range 0.7 to 10.7%). Over the 6-h study period the cumulative excretion of 13C in breathwas 16.4 ± 12.5% of that administered, ranging between 2.4 and 40.2% (Figure 6.1B, Table6.3). If defective lipolysis would be responsible for the continuing fat malabsorption in CFpatients, then a low percentage of fat absorption would be expected to correlate with lowexpiration of 13CO2 after 13C-MTG ingestion. However, no significant relationship wasobserved between 6-h cumulative 13CO2 expiration and either daily fecal fat excretion (r=-0.02) or the percentage of total fat absorption (r=0.04).
0
2
4
6
8
10
12
0 1 2 3 4 5 6
% 13
C D
ose
h-1
A
Time (h)
0
10
20
30
40
0 1 2 3 4 5 6
% C
umul
13C
O2
Exc
retio
n
B
Time (h)
Figure 6.1 Time courses for the excretion of 13C in breath over the 6-h study period following oral ingestion of 13C-MTG (4 mg per
kg body weight) at time 0 in 10 CF patients. Each symbol represents a patient. Figure (A) represents the excretion rate, whereas
figure (B) represents the cumulative 13CO2 excretion.
13C-LA testThe baseline 13C-LA abundance in plasma prior to consumption of the 13C-LA label was -29.1± 2.2‰ (range -32.6 to -25.5‰). 13C-LA concentration in plasma samples, expressed aspercentage of the dose per liter plasma, increased steeply after approximately 6 h (Figure 6.2).Peak values of 13C-LA concentrations in plasma after administration occurred between 8 and24 h. At 24 h after ingestion of the label, the enrichment of 13C-LA in plasma had not yetreturned to the level of baseline 13C-abundance. Plasma 8-h 13C-LA concentrations varied from0.5 to 2.0% dose L-1 plasma (Table 6.3).
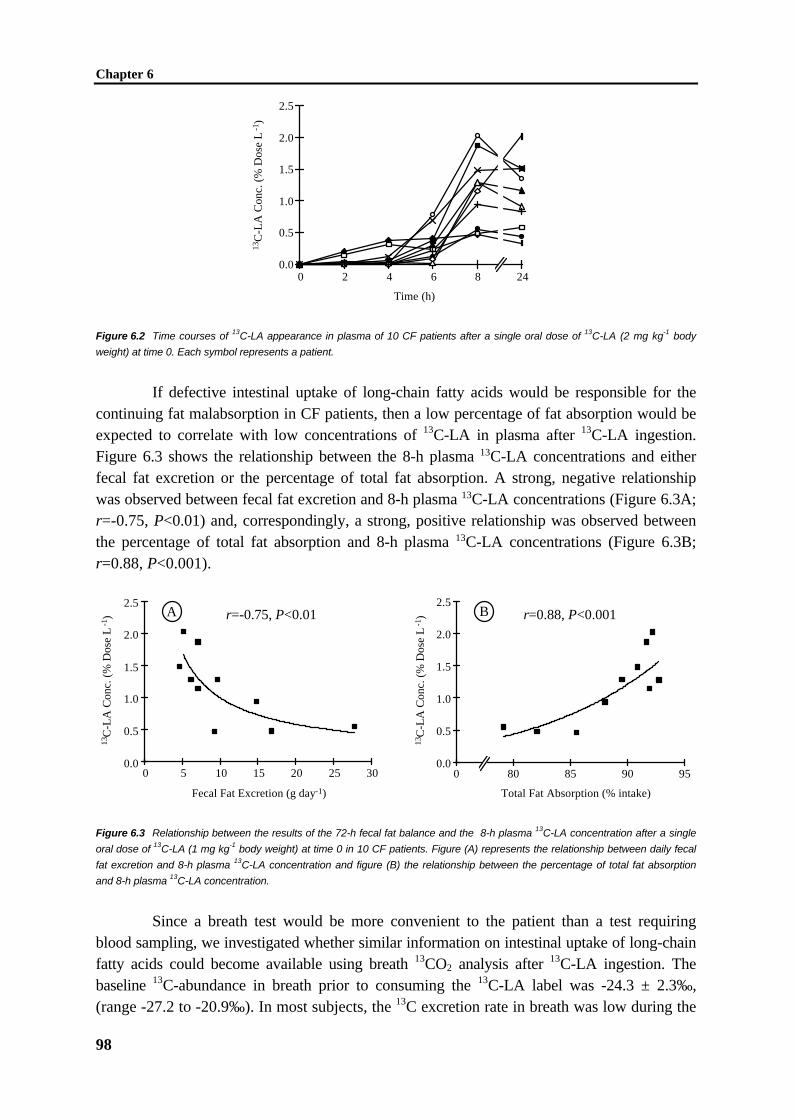
Chapter 6
98
Time (h)
13C
-LA
Con
c. (
% D
ose
L-1
)0.0
0.5
1.0
1.5
2.0
2.5
0 2 4 6 8 24
Figure 6.2 Time courses of 13C-LA appearance in plasma of 10 CF patients after a single oral dose of 13C-LA (2 mg kg-1 body
weight) at time 0. Each symbol represents a patient.
If defective intestinal uptake of long-chain fatty acids would be responsible for thecontinuing fat malabsorption in CF patients, then a low percentage of fat absorption would beexpected to correlate with low concentrations of 13C-LA in plasma after 13C-LA ingestion.Figure 6.3 shows the relationship between the 8-h plasma 13C-LA concentrations and eitherfecal fat excretion or the percentage of total fat absorption. A strong, negative relationshipwas observed between fecal fat excretion and 8-h plasma 13C-LA concentrations (Figure 6.3A;r=-0.75, P<0.01) and, correspondingly, a strong, positive relationship was observed betweenthe percentage of total fat absorption and 8-h plasma 13C-LA concentrations (Figure 6.3B;r=0.88, P<0.001).
13C
-LA
Con
c. (
% D
ose
L-1
)
0 5 10 15 20 25 300.0
0.5
1.0
1.5
2.0
2.5
Fecal Fat Excretion (g day-1)
A r=-0.75, P<0.01
Total Fat Absorption (% intake)
950.0
0.5
1.0
1.5
2.0
2.5
0 80 85 90
B r=0.88, P<0.001
13C
-LA
Con
c. (
% D
ose
L-1
)
Figure 6.3 Relationship between the results of the 72-h fecal fat balance and the 8-h plasma 13C-LA concentration after a single
oral dose of 13C-LA (1 mg kg-1 body weight) at time 0 in 10 CF patients. Figure (A) represents the relationship between daily fecal
fat excretion and 8-h plasma 13C-LA concentration and figure (B) the relationship between the percentage of total fat absorption
and 8-h plasma 13C-LA concentration.
Since a breath test would be more convenient to the patient than a test requiringblood sampling, we investigated whether similar information on intestinal uptake of long-chainfatty acids could become available using breath 13CO2 analysis after 13C-LA ingestion. Thebaseline 13C-abundance in breath prior to consuming the 13C-LA label was -24.3 ± 2.3‰,(range -27.2 to -20.9‰). In most subjects, the 13C excretion rate in breath was low during the

Impaired uptake of fats in CF patients
99
first hours, then increased rapidly and reached a possible maximum value at 6 h afteradministration of the label (Figure 6.4A). In most subjects no decay was observed during thetime course of the study. This time course pattern was very similar to the pattern obtained for13C-LA concentrations in plasma, except for subject 1, whose 13C excretion rate in breathalready increased after 90 min. The 6-h cumulative 13CO2 expiration (Table 6.3) amounted to2.7 ± 3.1% dose. In Figure 6.4B the 6-h cumulative 13CO2 expiration for all patients is plotted.In contrast to plasma values, no significant relationship between 6-h cumulative 13CO2
expiration and either fecal fat excretion (r=0.00) or the percentage of total fat absorption (r=-0.13) was observed. In addition, there was no correlation between plasma 13C-LAconcentrations and cumulative breath 13CO2 expiration (r=0.32), indicating that the multitudeof metabolic processes limits the utility of breath samples to measure uptake of long-chainfatty acids [45]. The results indicate that for the measuring intestinal uptake of long-chain fattyacids, plasma sampling cannot be easily replaced by breath sampling.
Finally, we investigated the excretion of 13C-LA in feces. The apparent absorption of13C-label was determined from the difference between the amount of 13C-LA administered andthat excreted in feces. 13C-LA excretion in feces over the 3-day period was very low andvaried between 0.0 and 1.8% of the dose administered (Table 6.3). No metabolites of 13C-LAwere observed in the feces. There was no significant correlation observed between theexcretion of 13C-LA and of total fat in feces (r=0.22, P=0.54).
% 13
C D
ose
h-1
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
0 1 2 3 4 5 6
A
Time (h)
% C
umul
13C
O2
Exc
retio
n
0
2
4
6
8
10
12
0 1 2 3 4 5 6
B
Time (h)
Figure 6.4 Time courses for the excretion of 13C in breath over the 6-h study period following oral ingestion of 13C-LA (1 mg kg-1
body weight) at time 0 in 10 CF patients. Figure (A) represents the excretion rate, whereas figure (B) represents the cumulative13CO2 excretion.
Total bile salt concentrations in plasma and fecesTotal bile salt concentrations were determined in plasma and feces. Fasting plasma total bilesalt concentrations in CF patients were high when compared with normal healthy controlvalues and ranged from 11.6 to 30.3 µmol L -1 (mean 17.2 µmol L -1) (Table 6.1). Following ameal there was no significant change in total plasma bile salts (data not shown). Fecal total bilesalt concentrations in most CF patients were elevated (range 0.7-30.2; mean 13.8 mmol per kgfecal wet weight) when compared with healthy control values, indicating that they had bile saltmalabsorption (Table 6.2). Bile salt malabsorption could result in a decreased amount of bilesalts available for the formation of mixed micelles, leading to fat malabsorption. However, no

Chapter 6
100
significant correlation was found between percentage of dietary fat absorption and fecal bilesalt concentrations (r=0.26).
Discussion
In CF patients, pancreatic enzyme replacement therapy frequently does not correct disorderedfat absorption to values obtained in controls. Our results of the 3-day fat balance confirm thepresence of mild to moderate fat malabsorption (percentage of total fat absorption: 79-93%) ina group of pediatric CF patients on enzyme replacement therapy despite good clinicalconditions. The aim of the present study was to elucidate whether fat malabsorption in CFpatients receiving habitual pancreatic enzyme replacement therapy is due to deficient lipolysisof triacylglycerols or due to impaired intestinal uptake of fatty acids.
We applied two fat substrates with different physical and chemical properties, i.e.13C-MTG and 13C-LA. The principle of the 13C-MTG breath test is based on lipolysis-dependent 13CO2 excretion via the breath. Efficient absorption of the 13C label from the mixedtriglyceride is limited primarily by lipolysis [23], and the 13C-MTG test therefore distinguishespancreatic insufficiency from deficient intestinal uptake of long-chain fatty acids. After 13C-MTG ingestion, no relationship was observed between recovery of 13CO2 in breath andpercentage of total fat absorption, indicating that fat malabsorption in CF patients on theirhabitual enzyme replacement therapy is probably not related to defective lipolysis. Therecovery of expired 13CO2 obtained in the present study was similar to those obtained in otherstudies, indicating sufficient supplementation of pancreatic enzymes to the CF patients in thisstudy. In healthy adults the 6-h cumulative percentage of 13C expired via the breath afteringestion of 13C-MTG varied between 23 and 52% of the dose in one study [23] and between3 and 48% in another study [24]. The recovery of expired 13CO2 in CF patients receivingregular amounts of pancreatic enzymes varied between 0 and 45% [23,25]. In neither of thesestudies total fat absorption was related to the percentage of 13C recovered in the breath.
Efficient absorption of 13C-LA, a long-chain unesterified fatty acid, differspredominantly from 13C-MTG in its dependence on adequate intestinal uptake [27]. Minich etal. [28] showed in a rat model for fat malabsorption (permanently interrupted enterohepaticcirculation) that measuring plasma 13C-LA concentrations is a valuable method to assess theintestinal uptake of long-chain fatty acids and correlates with fat absorption. The 13C-LA testtherefore distinguishes deficient intestinal uptake of long-chain fatty acids from pancreaticinsufficiency [28]. After ingestion of 13C-LA, a strong relationship was observed between 8-hplasma 13C-LA concentrations and total fat absorption, indicating that the observed fatmalabsorption in CF patients on their habitual enzyme replacement therapy is due to defectiveintestinal uptake of long-chain fatty acids.
Impaired intestinal uptake of long-chain fatty acids may result from several processes.In the absence of adequate bicarbonate secretion, gastric acid entering the duodenum maylower intestinal pH until well into the jejunum [11]. Bile salts are readily precipitated in an acidmilieu [17], and duodenal bile salt concentration may fall below the critical micellarconcentration, thereby exacerbating fat malabsorption. Precipitated bile salts also appear to be

Impaired uptake of fats in CF patients
101
lost from the enterohepatic circulation in greater quantities, thus reducing the total bile saltpool and decreasing the fraction of bile salts conjugated with glycine [20]. Intracellullar eventsmay also contribute to impaired uptake of long-chain fatty acids in CF patients, e.g. due toabsent fatty acid binding proteins or impaired chylomicron assembly and secretion [46].Viscous, thick intestinal mucus, with altered physical properties, may have a deleterious effecton the thickness of the intestinal unstirred water layer, limiting translocation of long-chainfatty acids over the intestinal epithelium [5,21]. Our data on increased fecal bile salt losses arein agreement with several other studies [47-49] and could be in agreement with a diminishedbile salt pool in CF patients. Watkins et al. [18] showed that bile acid pool size was nearlydoubled upon treatment with pancreatic enzymes in a group of CF patients with normal fecalbile salt losses. Although the present data suggest that the problem is related to insufficientlong-chain fatty acid uptake, they do not allow a clear identification of the individual processresponsible for impaired uptake.
The 13C-LA bolus was administered in an acid-resistant coated capsule, preventingthe capsule from being opened at a low pH environment (gastric or intestinal). In patients witha low intestinal pH, e.g. due to inadequate bicarbonate secretion [10,11], the bioavailability of13C-LA was hypothesized to be impaired, resulting in a decreased amount of 13C-LAincorporated into plasma linoleic acid. Since low intestinal pH affects uptake of long-chainfatty acids, we reasoned that the acid-resistant capsule probably enhances the effect of the 13C-LA test in correctly diagnosing solubilization disorders. The release of the substrate may bedelayed in some patients, which can explain the differences in timing for the onset of theindividual 13C-LA curves. In addition, delayed time courses for the onset of 13CO2 in breathhave been observed before and may be explained by, e.g., delayed gastric emptying [24,50,51].
The study was designed such that the patients served as their own controls. Thus, ineach individual patient we calculated the percentage of total fat absorption and related theseresults to the measurements of the 13C-MTG breath test and the 13C-LA test. We reasoned thatthese controls would be the most appropriate, given the fact that neither the optimal positivecontrol group (pancreatic sufficient CF patients with known impaired intestinal uptake) nor theoptimal negative control group (pancreatic sufficient CF patients without intestinal uptakedisorder) exists or is available. The present approach allowed us to relate the results of totalfat absorption to the results of lipolysis and intestinal uptake in the individual patient.
In conclusion, fat balance data indicate that, despite enzyme replacement therapy,pediatric CF patients have increased fecal fat excretion and, correspondingly, decreasedpercentage of fat absorption. The results of the 13C-MTG test and 13C-LA test indicate thatcontinuing fat malabsorption is not likely due to insufficient enzyme replacement therapy, butrather due to either incomplete intraluminal solubilization and/or reduced mucosal uptake oflong-chain fatty acids. Indirect indications exist that an increased bile salt loss leading to adiminished bile salt pool may contribute to this problem. Therapeutic attempts to normalize fatabsorption in pediatric CF patients need to include a strategy to improve intestinal uptake oflong-chain fatty acids.

Chapter 6
102
References
1. Carey MC, Hernell O. Digestion and absorption of fat. Sem Gastrointest Dis 1992;3:189-208.
2. Tso P. Intestinal lipid absorption. In: Johnson LR, ed. Physiology of the gastrointestinaltract. 3rd Ed. New York: Raven Press, 1994:1867-1907.
3. Staggers JE, Hernell O, Stafford RJ, Carey MC. Physical-chemical behavior of dietary andbiliary lipids during intestinal digestion and absorption. 1. Phase behavior and aggregationstates of model lipid systems patterned after aqueous duodenal contents of healthy adulthuman beings. Biochemistry 1990;29:2028-2040.
4. Hernell O, Staggers JE, Carey MC. Physical-chemical behavior of dietary and biliary lipidsduring intestinal digestion and absorption. 2. Phase analysis and aggregation states ofluminal lipids during duodenal fat digestion in healthy adult human beings. Biochemistry1990;29:2041-2056.
5. Pencharz PB, Durie PR. Nutritional management of cystic fibrosis. Annu Rev Nutr1993;13:111-136.
6. Shalon LB, Adelson JW. Cystic fibrosis. Gastrointestinal complications and gene therapy.In: Lebenthal E, ed. The pediatric clinics of North America. Pediatric gastroenterology I.Philadelphia: W.B. Saunders Company, 1996:157-196.
7. Zentler-Munro PL, Fine DR, Batten JC, Northfield TC. Effect of cimetidine on enzymeinactivation, bile acid precipitation, and lipid solubilisation in pancreatic steatorrhoea dueto cystic fibrosis. Gut 1985;26:892-901.
8. Regan PT, Malagelada J-R, DiMagno EP, Go VLW. Reduced intraluminal bile acidconcentrations and fat maldigestion in pancreatic insufficiency: correction by treatment.Gastroenterology 1979;77:285-289.
9. Carroccio A, Pardo F, Montalto G, Iapichino L, Soreso M, Averna MR, Iacono G,Notarbartolo A. Use of famotidine in severe exocrine pancreatic insufficiency withpersistent maldigestion on enzymatic replacement therapy. A long-term study in cysticfibrosis. Dig Dis Sci 1992;37:1441-1446.
10. Abrams CK, Hamosh M, Hubbard VS, Dutta SK, Hamosh P. Lingual lipase in cysticfibrosis. Quantitation of enzyme activity in the upper small intestine of patients withexocrine pancreatic insufficiency. J Clin Invest 1984;73:374-382.
11. Weber AM, Roy CC. Intraduodenal events in cystic fibrosis. J Pediatr Gastroenterol Nutr1984;3(suppl. 1):S113-S119.
12. Robinson PJ, Smith AL, Sly PD. Duodenal pH in cystic fibrosis and its relationship to fatmalabsorption. Dig Dis Sci 1990;35:1299-1304.
13. Robinson P, Sly P. High dose pancreatic enzymes in cystic fibrosis. Arch Dis Child1990;65:311-312.
14. Smyth R, Van Velzen D, Smyth A, Lloyd D, Heaf D. Strictures of ascending colon incystic fibrosis and high strength pancreatic enzymes. Lancet 1994;343:85-86.
15. MacSweeney E, Oades PJ, Buchdahl RM, Phelan M, Bush A. Relationship of thickeningof colon wall to pancreatic enzyme treatment in cystic fibrosis. Lancet 1995;345:752-756.

Impaired uptake of fats in CF patients
103
16. FitzSimmons SC, Burkhart GA, Borowitz D, Grand RJ, Hammerstrom T, Durie PR,Lloydstill JD, Lowenfels AB. High-dose pancreatic-enzyme supplements and fibrosingcolonopathy in children with cystic fibrosis. N Engl J Med 1997;336:1283-1289.
17. Zentler-Munro PL, FitzPatrick WJF, Batten JC, Northfield TC. Effect of intrajejunalacidity on aqueous phase bile acid and lipid concentrations in pancreatic steatorrhoea dueto cystic fibrosis. Gut 1984;25:500-507.
18. Watkins JB, Tercyak AM, Szczepanik P, Klein PD. Bile salt kinetics in cystic fibrosis:influence of pancreatic enzyme replacement. Gastroenterology 1977;73:1023-1028.
19. Roy CC, Weber EA, Morin CL, Combes J-C, Nusslé D, Mégevand A, Lasalle R.Abnormal biliary lipid composition in cystic fibrosis. Effect of pancreatic enzymes. N EngJ Med 1977;297:1301-1305.
20. Belli DC, Levy E, Darling P, Leroy C, Lepage G. Taurine improves the absorption of afat meal in patients with cystic fibrosis. Pediatrics 1987;80:517-523.
21. Eggermont E, De Boeck K. Small intestinal abnormalities in patients with cystic fibrosis.Eur J Pediatr 1991;150:824-828.
22. Corey M, McLaughlin FJ, Williams M, Levison H. A comparison of survival, growth, andpulmonary function in patients with cystic fibrosis in Boston and Toronto. J ClinEpidemiol 1988;41:583-591.
23. Vantrappen GR, Rutgeerts PJ, Ghoos YF, Hiele MI. Mixed triglyceride breath test: Anoninvasive test of pancreatic lipase activity in the duodenum. Gastroenterology1989;96:1126-1134.
24. Kalivianakis M, Verkade HJ, Stellaard F, Van der Werf M, Elzinga H, Vonk RJ. The 13C-mixed triglyceride breath test in healthy adults: determinants of the 13CO2 response. Eur JClin Invest 1997;27:434-442.
25. Amarri S, Harding M, Coward WA, Evans TJ, Weaver LT. 13Carbon mixed triglyceridebreath test and pancreatic enzyme supplementation in cystic fibrosis. Arch Dis Child1997;76:349-351.
26. Swart GR, Baartman EA, Wattimena JL, Rietveld T, Overbeek SE, Van den Berg JW.Evaluation studies of the 13C-mixed triglyceride breath test in healthy controls and adultcystic fibrosis patients with exocrine pancreatic insufficiency. Digestion 1997;58:415-420.
27. Watkins JB, Klein PD, Schoeller DA, Kirschner BS, Park R, Perman JA. Diagnosis anddifferentiation of fat malabsorption in children using 13C-labeled lipids: trioctanoin, trioleinan palmitic acid breath tests. Gastroenterology 1982;82:911-917.
28. Minich DM, Kalivianakis M, Havinga R, Kuipers F, Stellaard F, Vonk RJ, Verkade HJ. Anovel 13C-linoleic acid absorption test detects lipid malabsorption due to impairedsolubilization in rats. Gastroenterology 1997;112 (suppl.):A894(Abstract).
29. Kerem B-S, Rommens JM, Buchanan JA, Markiewicz D, Cox TK, Chakravarti A,Buchwald M, Tsui L-C. Identification of the cystic fibrosis gene: genetic analysis. Science1989;245:1073-1080.
30. Gerver WJM, De Bruin R. Paediatric morphometrics: a reference manual. Utrecht:Bunge, 1996:1-262.
31. Zapletal A, Samánek M, Paul T. Lung function in children and adolescents. Methods,reference values. Basel: Karger, 1987:1-218.

Chapter 6
104
32. Lepage G, Roy CC. Direct transesterification of all classes of lipids in a one-step reaction.J Lipid Res 1986;27:114-120.
33. Van de Kamer JH, Ten Bokkel Huinink H, Weyers HA. Rapid method for thedetermination of fat in feces. J Biol Chem 1949;177:347-355.
34. Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can JBiochem Physiol 1959;37:911-917.
35. Murphy GM, Billing BH, Baron DN. A fluorimetric and enzymatic method for theestimation of serum total bile acids. J Clin Pathol 1970;23:594-598.
36. Kalek H-D, Stellaard F, Kruis W, Paumgartner G. Detection of increased bile acidexcretion by determination of bile acid content in single stool samples. Clin Chim Acta1984;140:85-90.
37. Eder K. Gas chromatography analysis of fatty acid methyl esters. J Chromatogr B1995;671:113-131.
38. Gutnikov G. Fatty acid profiles of lipid samples. J Chromatogr B 1995;671:71-89.39. Guo Z, Nielsen S, Burguera B, Jensen MD. Free fatty acid turnover measured using
ultralow doses of [U-13C]palmitate. J Lipid Res 1997;38:1888-1895.40. Newcomer AD, Hofmann AF, DiMagno EP, Thomas PJ, Carlson GL. Triolein breath
test; A sensitive and specific test for fat malabsorption. Gastroenterology 1979;76:6-13.41. Einarsson K, Björkhem I, Eklöf R, Blomstrand R. 14C-triolein breath test as a rapid and
convenient screening test for fat malabsorption. Scand J Gastroenterol 1983;18:9-12.42. Mills PR, Horton PW, Watkinson G. The value of the 14C breath test in the assessment of
fat absorption. Scand J Gastroenterol 1979;14:913-921.43. Carey MC, Small DM, Bliss CM. Lipid digestion and absorption. Ann Rev Physiol
1983;45:651-677.44. Fomon SJ, Ziegler EE, Thomas LN, Jensen RL, Filer LJ. Excretion of fat by normal full-
term infants fed various milks and formulas. Am J Clin Nutr 1970;23:1299-1313.45. Pedersen NT, Jorgensen BB, Rannem T. The [14C]-triolein breath test is not valid as a test
of fat absorption. Scand J Clin Lab Invest 1991;51:699-703.46. Minich DM, Vonk RJ, Verkade HJ. Intestinal absorption of essential fatty acids under
physiological and essential fatty acid-deficient conditions. J Lipid Res 1997;38:1709-1721.
47. O'Brien S, Mulcahy H, Fenlon H, O'Broin A, Casey M, Burke A, FitzGerald MX,Hegarty JE. Intestinal bile acid malabsorption in cystic fibrosis. Gut 1993;34:1137-1141.
48. Weber AM, Roy CC, Morin CL, Lasalle R. Malabsorption of bile acid in children withcystic fibrosis. N Eng J Med 1973;289:1001-1005.
49. Weber AM, Roy CC, Chartrand L. Relationship between bile acid malabsorption andpancreatic insufficiency in cystic fibrosis. Gut 1976;17:295-299.
50. Duncan A, Cameron A, Stewart MJ, Russell RI. Limitations of the triolein breath test.Clin Chim Acta 1992;205:51-64.
51. Jones PJH, Pencharz PB, Clandinin MT. Absorption of 13C-labeled stearic, oleic, andlinoleic acids in humans: Application to breath tests. J Lab Clin Med 1985;105:647-652.

CHAPTER 7
Increased fecal bile salt excretion is independent ofthe presence of dietary fat malabsorption in two
mouse models for cystic fibrosis
M. Kalivianakis, I. Bronsveld, H.R. de Jonge, M. Sinaasappel,R. Havinga, F. Kuipers, B.J. Scholte, H.J. Verkade

Chapter 7
106
CHAPTER 7
Increased fecal bile salt excretion is independent ofthe presence of dietary fat malabsorption in two
mouse models for cystic fibrosis
Abstract
Background & Aim: Recent studies from our laboratory suggested that fat malabsorption incystic fibrosis (CF) patients on pancreatic enzyme replacement therapy is partially due toimpaired intestinal uptake of long-chain fatty acids [1], which may be due to bile-relatedprocesses. To obtain more insight into the effects of CF on fat absorption and bile formation,we studied two mouse models for CF: mice homozygous for the ∆F508 mutation in the cftrgene (∆F508/∆F508), and mice in which the cftr gene is disrupted (cftr -/-). Methods: Fatabsorption was studied by means of a 3-day fat balance, after feeding a standard (14 en% fat)or a high-fat (35 en% fat) diet for 2 weeks. Biliary bile salt secretion was determined during80 min after cannulation of the gallbladder. Fecal bile salts were determined for 3 days.Results: In ∆F508/∆F508 mice, dietary fat absorption was not significantly different fromcontrols, and above 94% in all groups. However, dietary fat absorption in cftr -/- mice wassignificantly decreased compared to controls: standard diet: 82.8 ± 3.0% (mean ± SEM) and93.9 ± 1.3% (P<0.01); high-fat diet: 88.8 ± 1.6% and 95.0 ± 1.4% (P<0.01), respectively.Biliary bile salt secretion rates were similar for the CF mouse models and their respectivecontrols on either diet. The contribution of cholic acid to the biliary bile salt pool was slightlyincreased in both CF mice at the expense of deoxycholic acid. Primary bile salts were slightlyincreased, whereas secondary bile salts were slightly decreased in CF mice. Fecal bile saltexcretion was increased in ∆F508/∆F508 and in cftr -/- mice when compared with theirrespective controls (10 versus 5 µmol g -1 feces, respectively, P<0.01). No significantcorrelation was observed between fecal excretion of bile salts and of fats. Conclusion: Cftr -/-mice, but not ∆F508/∆F508 mice, have an impaired dietary fat absorption, which does notresult from alterations in bile production. In both CF mouse models, fecal bile salt excretionwas increased, but this was not related to increased fecal fat excretion. Bile composition dataindicate that the increased fecal loss of bile salts is compensated for by an increased bile saltneosynthesis.

Fat (mal)absorption in CF mice
107
Introduction
Cystic fibrosis (CF), the most common recessive disorder in Caucasian populations, is causedby mutations in the gene for the cystic fibrosis transmembrane conductance regulator (CFTR).The CFTR gene encodes for a phosphorylation-regulated Cl- channel and is expressed in theapical membrane of various epithelial cells [2-4]. Malfunction of this chloride channel in CFpatients is associated with obstruction and inflammation of airways, pancreatic ducts, intestineand bile ducts, frequently resulting in intestinal fat malabsorption [5,6]. The most commonmutation in the CFTR gene in Caucasian populations is a deletion of a phenylalanine residue atamino acid position 508 of the protein (∆F508), which is found in 90% of the CF patients inNorthern Europe [3,4]. The ∆F508 mutation disrupts the biosynthetic processing of CFTR toits mature glycosylated form [7], so that the protein is retained in the endoplasmic reticulumand subsequently degraded [8]. The protein does not reach the apical plasma membrane, andas a result, affected epithelia lack CFTR in the apical membrane and are deficient in cAMP-stimulated Cl- permeability [8].
In an attempt to further elucidate the pathophysiology of CF, mouse models weredeveloped [9-11]. The initial excitement generated by the emergence of these mouse modelswas somewhat tempered by the finding that none of the models showed spontaneous airwaydisease, which is primarily responsible for most of the morbidity and mortality in the humanCF population. However, the various CF mouse models are remarkably similar to their humancounterparts with respect to intestinal pathophysiology [11-14]. Most importantly, theintestinal tract of the CF mouse models demonstrates the absence or decrease of cAMP-mediated chloride transport which often results in intestinal obstructions, a hallmark of CF.
Most CF patients display a considerable malabsorption of dietary fats due topancreatic insufficiency resulting in impaired lipolysis [5,15]. Pancreatic insufficiency can bealleviated by oral supplementation of pancreatic enzymes, however, many patients continue toexperience a degree of steatorrhoea, with fat absorption ranging from 80 to 90% of theirdietary fat intake [16-18]. Recently, we reported strong indications that fat malabsorption inCF patients on pancreatic enzyme replacement therapy is partially due to impaired intestinaluptake of long-chain fatty acids [1]. Impaired uptake may involve bile-related processes suchas altered bile composition, decreased bile salt secretion, or bile salt inactivation at lowintestinal pH [18-22]. Furthermore, small bowel mucosal dysfunction or alterations in themucus layer have been suggested to contribute to inefficient intestinal uptake of long-chainfatty acids in CF patients [5,23]. So far it has been difficult to differentiate between theseprocesses, partially due to the relative inaccessibility of the processes for mechanisticinvestigations in humans. Yet, a more detailed insight into the processes causing impaireduptake of long-chain fatty acids would allow the development of improved nutritionaltherapies. This will likely benefit CF patients, because the positive correlation between a goodnutritional status and long-term survival or well-being of CF patients is well documented [24].
To obtain more insight into the effects of CF on fat absorption and bile formation westudied two recently generated CF mouse models: (i) Mice homozygous for the ∆F508mutation in the cftr gene [11]. This mouse model was chosen because the ∆F508 mutation isthe most frequently observed mutation in cystic fibrosis patients in Caucasian population; (ii)

Chapter 7
108
Mice with complete inactivation of the cftr gene, cftr -/- “null” mice [14], expected to result incomplete inactivation of the cftr gene, the most severe phenotype.
Materials and Methods
AnimalsThe male and female mice with the ∆F508 mutation (∆F508/∆F508) used in this study weregenerated by Van Doorninck et al. and are described in reference [11]. Mice with a targeteddisruption in the cftr gene (cftrm1cam knockout mice, cftr -/-), resulting in complete loss ofCFTR function, and their controls (cftr +/+) were described by Ratcliff et al. [14].Experiments involving ∆F508/∆F508 mice and their controls (N/N) were performed with thestrain in 129/FVB genetic background [11], whereas experiments involving cftr -/- and cftr+/+ mice were performed with the strain in 129/C57/Bl6 genetic background [14]. All micewere obtained from the breeding colony at the Erasmus University Rotterdam, TheNetherlands. The animals used for the experiments reported here were approximately 2-3months old, had no obvious signs of disease or discomfort, and an average weight of 29 ± 1 g.The genotype of each individual animal was tested by Southern blotting of tail DNA [4]. Fortwo weeks prior to the experiments, animals were kept on a semi-synthetic diet with standardamounts of fat (14 en% fat; 4.538 kcal kg-1 food; fatty acid composition: C8-C12, 2.6%;C16:0, 13.5%; C18:0, 4.2%; C18:1n-9, 21.3%; C18:2n-6, 48.4%; C18:3n-3, 1.1%) or anisocaloric high fat diet (35 en% fat; 4.538 kcal kg-1 food; fatty acid composition: C8-C12,4.4%; C16:0, 28.5%; C18:0, 3.9%; C18:1n-9, 33.2%; C18:2n-6, 29.3%; C18:3n-3, 0.2%)(Hope Farms BV, Woerden, The Netherlands). The high-fat diet was applied to challenge theabsorptive system for fats in the mouse intestine. Mice were housed in an environmentallycontrolled facility with diurnal light cycling and had free access to chow and tap water.Experimental protocols were approved by the Ethical Committee for Animal Experiments,Erasmus University Rotterdam.
Study protocolFor fat balance measurements, feces was collected and food intake was recorded for 3 days.The gallbladder of mice was cannulated under Hypnorm (fentanyl/fluanisone, 1 mL kg-1 bodyweight) and Diazepam anesthesia (10 mg kg-1 body weight) and bile was collected in 20-minutes fractions for 80 minutes. Bile production was assessed by weight assuming that 1 mLof bile corresponds to 1 g of bile. After bile collection, a large blood sample (0.5-1 mL) wascollected by cardiac puncture.
Analytical techniquesLipids. Rat chow and feces were freeze-dried and mechanically homogenized. Aliquots ofchow and feces were extracted, hydrolyzed and methylated [25]. Resulting fatty acid methylesters were analyzed by gas chromatography to calculate fat intake and fecal fat excretion, asdetailed below. Percentage of total fat absorption was calculated from the daily fat intake andthe daily fecal fat excretion and expressed as a percentage of the daily fat intake.

Fat (mal)absorption in CF mice
109
Percentage of total fat absorptionFat intake (g day ) Fecal fat output (g day )
Fat intake (g day )
-1 -1
-1=−
×100%
Bile salts. Total bile salt concentrations in bile and plasma were determined by anenzymatic fluorimetric assay [26]. Individual bile salts in bile were analyzed by gaschromatography after extraction with commercially available Sep-Pak-C18 cartridges (WatersAssociates, Milford, MA, USA) [27]. Total fecal bile salt concentrations were extracted froman aliquot of freeze-dried homogenate [28] and fluorimetrically measured [26].
Gas liquid chromatography. Fatty acid methyl esters were separated and quantifiedby gas liquid chromatography on a Hewlett Packard gas chromatograph Model 6890 equippedwith a capillary column (Hewlett Packard - Ultra 1, crosslinked methyl silicone gum; 50 m x0.2 mm) and an FID detector. The gas chromatograph oven was programmed from an initialtemperature of 160°C to 290°C in 2 temperature steps (160°C held 2 min; 160-240°C, ramp2°C min-1, held 1 min; 240-290°C, ramp 10°C min-1, held 10 min). Quantification of the fattyacid methyl esters was performed by adding heptadecanoic acid (C17:0) as internal standard.
Bile salts were separated and quantified with a CP-SIL 19CB capillary column (25 mx 0.25 mm; Chrompack, Middelburg, The Netherlands). The gas chromatograph oven wasprogrammed from an initial temperature of 240°C to 280°C in 1 temperature step (240°C held4 min; ramp 10°C min-1; held 26 min). Quantification of bile salts was performed by adding5β-cholestane-3βol (coprostanol) as internal standard.
Calculations and statisticsThe experimental data are reported as means ± SEM. Differences between sample means ofCF mice and their controls were analyzed by the two-tailed Student’s t-test for unpaired dataor one-way ANOVA followed by post-hoc analysis (Student-Newman-Keuls). Differencesbetween means were considered statistically significant at the level of P<0.05. Analysis wasperformed with SPSS for Windows software (SPSS, Chicago, IL, USA).
Results
Fecal fat balanceNutritional data of ∆F508/∆F508 mice, cftr -/- mice and their respective controls on standardchow and high fat chow are shown in Table 7.1. No differences were observed between∆F508/∆F508 and their controls with respect to fat intake, fecal fat excretion, net fat uptakeor percentage of dietary fat absorption on either standard or high-fat diet. In cftr -/- mice onstandard diet, fecal fat excretion was significantly increased when compared with controls (82± 12 vs. 31 ± 4 µmol day-1, respectively, P<0.01). Accordingly, percentage of dietary fatabsorption was significantly decreased from 93.9 ± 1.3% in cftr +/+ mice to 82.8 ± 3.0% incftr -/- mice (P<0.01). Similar results were obtained when cftr mice were fed the high-fat diet,although net fat uptake was approximately three-fold higher when compared with the standarddiet: fecal fat excretion was significantly increased and percentage of dietary fat absorption
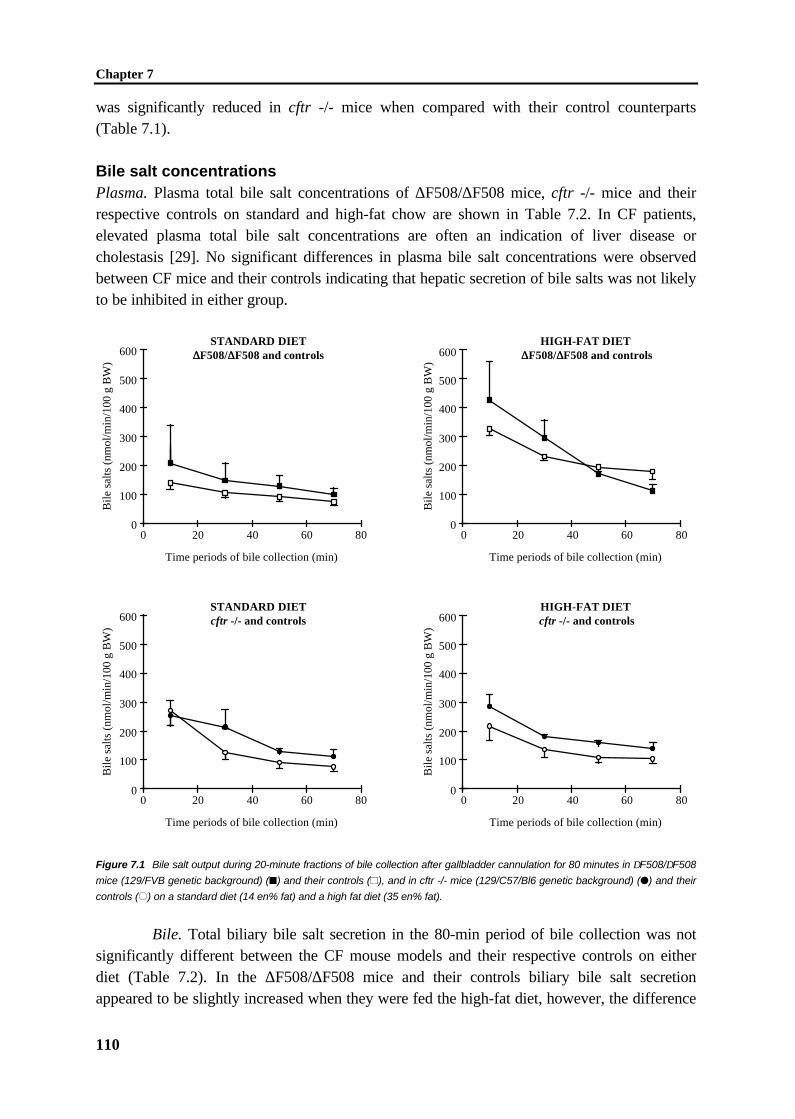
Chapter 7
110
was significantly reduced in cftr -/- mice when compared with their control counterparts(Table 7.1).
Bile salt concentrationsPlasma. Plasma total bile salt concentrations of ∆F508/∆F508 mice, cftr -/- mice and theirrespective controls on standard and high-fat chow are shown in Table 7.2. In CF patients,elevated plasma total bile salt concentrations are often an indication of liver disease orcholestasis [29]. No significant differences in plasma bile salt concentrations were observedbetween CF mice and their controls indicating that hepatic secretion of bile salts was not likelyto be inhibited in either group.
STANDARD DIET∆∆F508/∆∆F508 and controls
Bile
sal
ts (
nmol
/min
/100
g B
W)
0
100
200
300
400
500
600
Time periods of bile collection (min)
20 40 60 800
STANDARD DIETcftr -/- and controls
Bile
sal
ts (
nmol
/min
/100
g B
W)
0
100
200
300
400
500
600
Time periods of bile collection (min)
20 40 60 800
HIGH-FAT DIET ∆∆F508/∆∆F508 and controls
0
100
200
300
400
500
600
Time periods of bile collection (min)
20 40 60 800
HIGH-FAT DIET cftr -/- and controls
0
100
200
300
400
500
600
Time periods of bile collection (min)
20 40 60 800
Bile
sal
ts (
nmol
/min
/100
g B
W)
Bile
sal
ts (
nmol
/min
/100
g B
W)
Figure 7.1 Bile salt output during 20-minute fractions of bile collection after gallbladder cannulation for 80 minutes in ∆F508/∆F508
mice (129/FVB genetic background) (�) and their controls (~), and in cftr -/- mice (129/C57/Bl6 genetic background) (�) and their
controls (±) on a standard diet (14 en% fat) and a high fat diet (35 en% fat).
Bile. Total biliary bile salt secretion in the 80-min period of bile collection was notsignificantly different between the CF mouse models and their respective controls on eitherdiet (Table 7.2). In the ∆F508/∆F508 mice and their controls biliary bile salt secretionappeared to be slightly increased when they were fed the high-fat diet, however, the difference
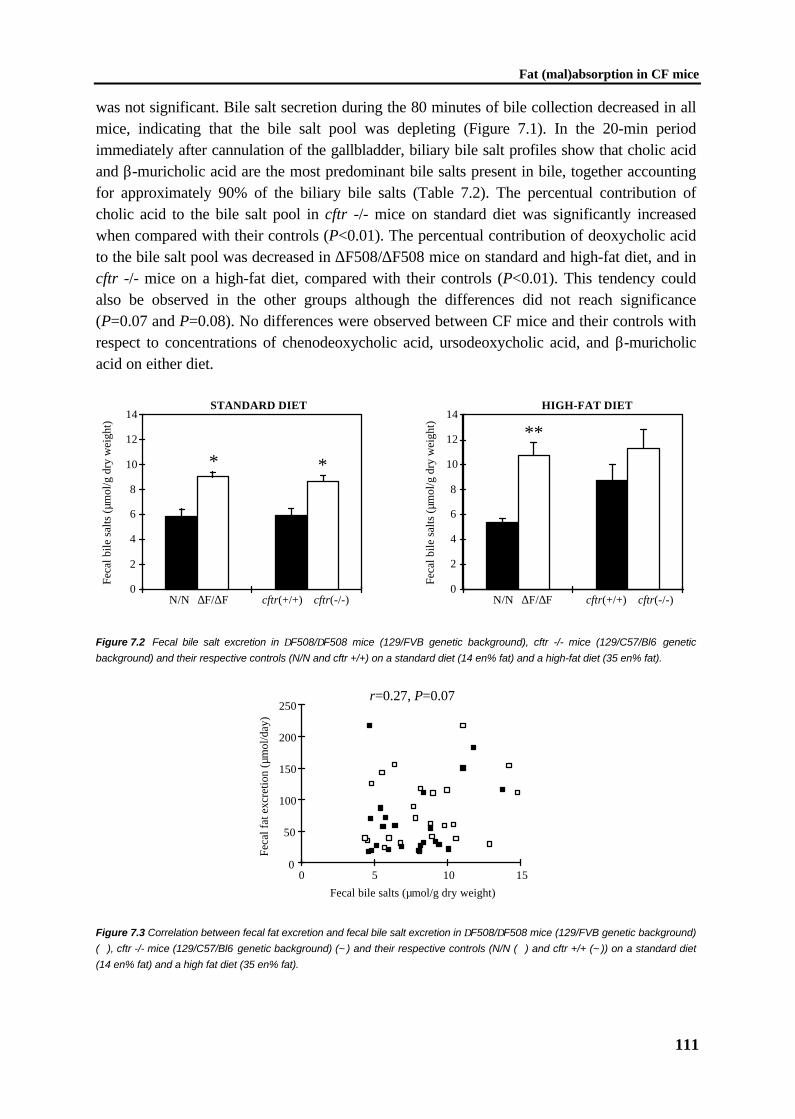
Fat (mal)absorption in CF mice
111
was not significant. Bile salt secretion during the 80 minutes of bile collection decreased in allmice, indicating that the bile salt pool was depleting (Figure 7.1). In the 20-min periodimmediately after cannulation of the gallbladder, biliary bile salt profiles show that cholic acidand β-muricholic acid are the most predominant bile salts present in bile, together accountingfor approximately 90% of the biliary bile salts (Table 7.2). The percentual contribution ofcholic acid to the bile salt pool in cftr -/- mice on standard diet was significantly increasedwhen compared with their controls (P<0.01). The percentual contribution of deoxycholic acidto the bile salt pool was decreased in ∆F508/∆F508 mice on standard and high-fat diet, and incftr -/- mice on a high-fat diet, compared with their controls (P<0.01). This tendency couldalso be observed in the other groups although the differences did not reach significance(P=0.07 and P=0.08). No differences were observed between CF mice and their controls withrespect to concentrations of chenodeoxycholic acid, ursodeoxycholic acid, and β-muricholicacid on either diet.
HIGH-FAT DIET
0
2
4
6
8
10
12
14
∆F/∆F cftr(-/-)cftr(+/+)N/N
**
STANDARD DIET
Feca
l bile
sal
ts (
µmol
/g d
ry w
eigh
t)
∆F/∆F cftr(-/-)cftr(+/+)N/N0
2
4
6
8
10
12
14
* *
Feca
l bile
sal
ts (
µmol
/g d
ry w
eigh
t)
h
Figure 7.2 Fecal bile salt excretion in ∆F508/∆F508 mice (129/FVB genetic background), cftr -/- mice (129/C57/Bl6 genetic
background) and their respective controls (N/N and cftr +/+) on a standard diet (14 en% fat) and a high-fat diet (35 en% fat).
0
50
100
150
200
250
0 5 10 15
Feca
l fat
exc
retio
n (µ
mol
/day
)
Fecal bile salts (µmol/g dry weight)
r=0.27, P=0.07
Figure 7.3 Correlation between fecal fat excretion and fecal bile salt excretion in ∆F508/∆F508 mice (129/FVB genetic background)
(Ä), cftr -/- mice (129/C57/Bl6 genetic background) (~) and their respective controls (N/N (Ä) and cftr +/+ (~)) on a standard diet
(14 en% fat) and a high fat diet (35 en% fat).
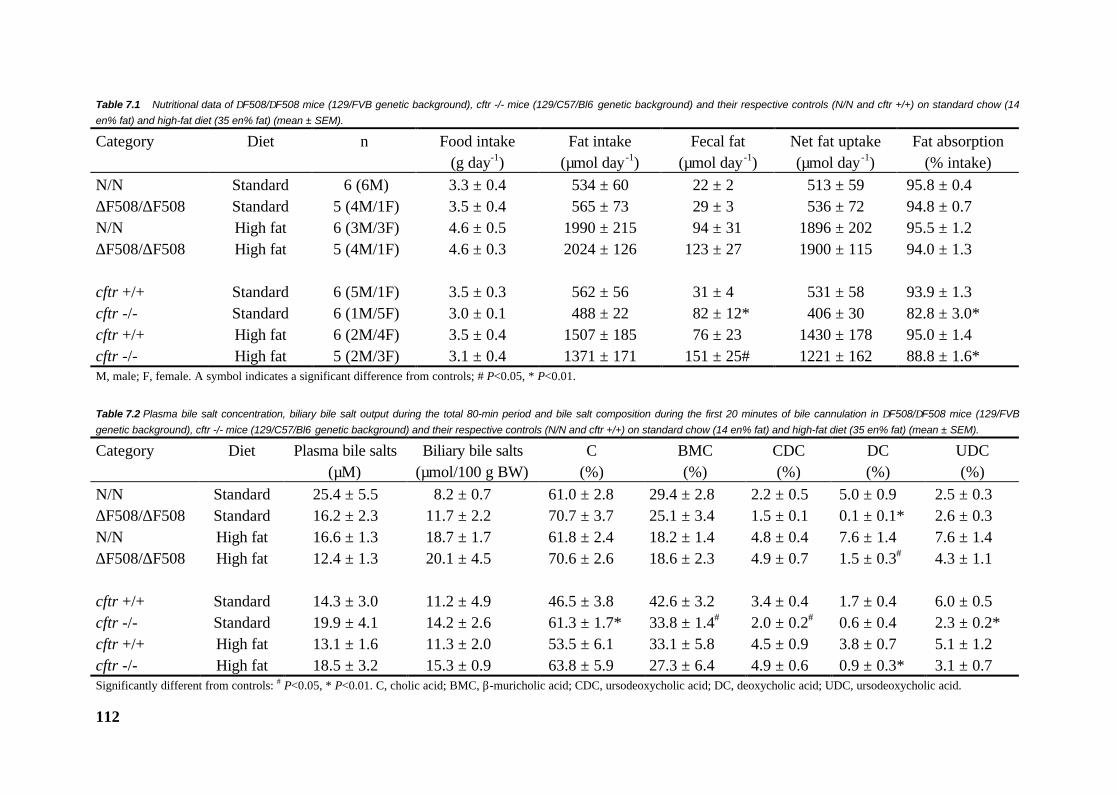
112
Table 7.1 Nutritional data of ∆F508/∆F508 mice (129/FVB genetic background), cftr -/- mice (129/C57/Bl6 genetic background) and their respective controls (N/N and cftr +/+) on standard chow (14
en% fat) and high-fat diet (35 en% fat) (mean ± SEM).
Category Diet n Food intake(g day-1)
Fat intake(µmol day -1)
Fecal fat(µmol day -1)
Net fat uptake(µmol day -1)
Fat absorption(% intake)
N/N Standard 6 (6M) 3.3 ± 0.4 534 ± 60 22 ± 2 513 ± 59 95.8 ± 0.4∆F508/∆F508 Standard 5 (4M/1F) 3.5 ± 0.4 565 ± 73 29 ± 3 536 ± 72 94.8 ± 0.7N/N High fat 6 (3M/3F) 4.6 ± 0.5 1990 ± 215 94 ± 31 1896 ± 202 95.5 ± 1.2∆F508/∆F508 High fat 5 (4M/1F) 4.6 ± 0.3 2024 ± 126 123 ± 27 1900 ± 115 94.0 ± 1.3
cftr +/+ Standard 6 (5M/1F) 3.5 ± 0.3 562 ± 56 31 ± 4 531 ± 58 93.9 ± 1.3cftr -/- Standard 6 (1M/5F) 3.0 ± 0.1 488 ± 22 82 ± 12* 406 ± 30 82.8 ± 3.0*cftr +/+ High fat 6 (2M/4F) 3.5 ± 0.4 1507 ± 185 76 ± 23 1430 ± 178 95.0 ± 1.4cftr -/- High fat 5 (2M/3F) 3.1 ± 0.4 1371 ± 171 151 ± 25# 1221 ± 162 88.8 ± 1.6*M, male; F, female. A symbol indicates a significant difference from controls; # P<0.05, * P<0.01.
Table 7.2 Plasma bile salt concentration, biliary bile salt output during the total 80-min period and bile salt composition during the first 20 minutes of bile cannulation in ∆F508/∆F508 mice (129/FVB
genetic background), cftr -/- mice (129/C57/Bl6 genetic background) and their respective controls (N/N and cftr +/+) on standard chow (14 en% fat) and high-fat diet (35 en% fat) (mean ± SEM).
Category Diet Plasma bile salts(µM)
Biliary bile salts(µmol/100 g BW)
C(%)
BMC(%)
CDC(%)
DC(%)
UDC(%)
N/N Standard 25.4 ± 5.5 8.2 ± 0.7 61.0 ± 2.8 29.4 ± 2.8 2.2 ± 0.5 5.0 ± 0.9 2.5 ± 0.3∆F508/∆F508 Standard 16.2 ± 2.3 11.7 ± 2.2 70.7 ± 3.7 25.1 ± 3.4 1.5 ± 0.1 0.1 ± 0.1* 2.6 ± 0.3N/N High fat 16.6 ± 1.3 18.7 ± 1.7 61.8 ± 2.4 18.2 ± 1.4 4.8 ± 0.4 7.6 ± 1.4 7.6 ± 1.4∆F508/∆F508 High fat 12.4 ± 1.3 20.1 ± 4.5 70.6 ± 2.6 18.6 ± 2.3 4.9 ± 0.7 1.5 ± 0.3# 4.3 ± 1.1
cftr +/+ Standard 14.3 ± 3.0 11.2 ± 4.9 46.5 ± 3.8 42.6 ± 3.2 3.4 ± 0.4 1.7 ± 0.4 6.0 ± 0.5cftr -/- Standard 19.9 ± 4.1 14.2 ± 2.6 61.3 ± 1.7* 33.8 ± 1.4# 2.0 ± 0.2# 0.6 ± 0.4 2.3 ± 0.2*cftr +/+ High fat 13.1 ± 1.6 11.3 ± 2.0 53.5 ± 6.1 33.1 ± 5.8 4.5 ± 0.9 3.8 ± 0.7 5.1 ± 1.2cftr -/- High fat 18.5 ± 3.2 15.3 ± 0.9 63.8 ± 5.9 27.3 ± 6.4 4.9 ± 0.6 0.9 ± 0.3* 3.1 ± 0.7Significantly different from controls: # P<0.05, * P<0.01. C, cholic acid; BMC, β-muricholic acid; CDC, ursodeoxycholic acid; DC, deoxycholic acid; UDC, ursodeoxycholic acid.

Fat (mal)absorption in CF mice
113
Feces. Fecal total bile salts were significantly increased in ∆F508/∆F508 mice andcftr -/- mice when compared with their respective controls on a standard diet (∆F508/∆F508mice: 9.1 ± 0.4 versus 5.9 ± 0.6 µmol per g dry weight of feces, respectively, P<0.01; cftr -/-mice: 10.8 ± 1.0 versus 5.4 ± 0.3 µmol per g dry weight of feces, respectively, P<0.01, Figure7.2). A significant increase was also obtained for ∆F508/∆F508 mice on a high-fat diet (10.8 ±1.0 versus controls 5.4 ± 0.3 µmol per g dry weight P<0.001). There was no significantcorrelation between fecal fat excretion and fecal bile salt excretion in the group as a whole(r=0.27, P=0.07; Figure 7.3), nor when the mice were stratified according to their genotype.
Discussion
Recently, we reported strong indications suggesting that fat malabsorption in CF patients onpancreatic enzyme replacement therapy is partially due to impaired intestinal uptake of long-chain fatty acids [1], which may involve bile-related processes. The aim of the present studywas to investigate in more detail fat absorption and bile formation in two recently generatedCF mouse models: ∆F508/∆F508 mice (129/FVB genetic background) and cftr -/- mice(129/C57/Bl6 genetic background) [11,14].
The current data show that cftr -/- mice exhibit fat malabsorption, in contrast to∆F508/∆F508 mice. No differences were observed with respect to biliary bile salt output.Hence, this is not likely the cause of the fat malabsorption. The observed fat malabsorption isnot likely due to differences in the composition of the bile salt pool either, since thesedifferences were observed in both ∆F508/∆F508 mice and cftr -/- mice, whereas only cftr -/-mice exhibit fat malabsorption. It has been speculated that liver disease develops in patientswith CF as a consequence of the plugging of intrahepatic bile ducts [30-32]. The lack ofCFTR in the apical membrane of bile duct cells may lead to abnormalities in biliary drainagewith chronic cholestasis [30]. Cholestasis would result in increased amounts of bile salts inplasma and decreased amounts of bile in the intestine, and absorption of dietary fats would beimpaired [29,33]. Our results show that intestinal bile salts were similar in all mice, indicatingthat cholestasis is probably not a cause of fat malabsorption in cftr -/- mice. Plasma bile saltswere similar in all mice.
Apparently, the fat malabsorption in the cftr -/- mice can not be explained byprocesses regarding bile formation in the liver. Other mechanisms that may contribute toinefficient fat absorption are intestinal mucosal dysfunction or alterations in the mucus layer.This would also be in concordance with the intestinal histologic abnormalities observed in∆F508/∆F508 mice and in cftr -/- mice [11,14]. Finally, the difference of fat malabsorptionmay be due to different pancreatic functioning. However, CF mice do not show majorhistological abnormalities in the pancreas or pancreatic duct and the secretion of amylase is notimpaired [34]. This lack of pancreatic disease in CF mice is most plausibly due to a lower levelof CFTR expression and a relatively higher contribution of alternative Ca-activated chloridechannels as compared with human pancreas [35,36]. CF-related abnormalities in the intestinerather than hepatobiliary or pancreatic disturbances are therefore the most probable cause offat malabsorption in cftr -/- mice.

Chapter 7
114
The observation that fat malabsorption is present in cftr -/- mice but absent in∆F508/∆F508 mice, may be due to the fact that the “Rotterdam” ∆F508/∆F508 mice exhibitresidual cftr activity [11,37], whereas in cftr -/- mice, cftr function is completely abolished[14]. Low apical cftr activity in ∆F508/∆F508 mice has been observed at physiologicaltemperatures in the gallbladder and in the ileum [11,37]. The level of residual cftr activitycould differ in various tissues and small variations in apical activity levels could have profoundeffects on pathology. A careful analysis of cftr processing kinetics between distinct tissues isnecessary to confirm this hypothesis. Moreover, it can not be excluded that the differentgenetic backgrounds (129/FVB and 129/C57/Bl6 for ∆F508/∆F508 and cftr -/- mice,respectively) are in part responsible for the different phenotypes of the two CF mouse models.Additionally, studies were performed with both male and female mice (Table 7.1), which mayalso influence the results. However, it has not been reported before in the literature that fatabsorption, bile salt pool size or bile composition differ for males and females.
In the present study, bile salt secretion during an 80-min period decreased byapproximately 50%, showing that most of the bile salt pool was collected. Total biliary bilesalt output during this period was similar for CF mice and their respective controls, suggestingthat bile salt pools of CF and control mice were similar. Previous data on the bile salt pool sizein CF patients are conflicting; both a normal and a decreased bile salt pool size have beenreported in CF patients exhibiting fat malabsorption [19,38]. Our results indicate that bile saltsecretion rates and bile salt pool size in the two mouse models for CF are not affected.
The observation of a relative increase in the proportion of cholic acid at the expenseof deoxycholic acid is a well-known phenomenon in CF [38,39]. This result is consistent withthe finding of increased fecal bile salts in CF mice. Interruption of the enterohepatic circulationby fecal bile salt loss is normally accompanied by an increase in bile salt synthesis in order tomaintain bile salt output [40]. The capacity to increase synthesis is estimated to equal three tofour times the pool size [41]. Apparently, the increased fecal loss of bile salts can becompensated for by increased hepatic bile salt neosynthesis.
Both CF mouse models exhibited increased excretion of bile salts in the feces whencompared with their respective controls. Fecal bile salt loss is well recognized in patients withCF and has been attributed to various intraluminal factors: 1. unhydrolyzed triacylglycerolsand phospholipids, 2. precipitation of bile salts in acidic duodenal content, 3. adsorption of bilesalts to non-absorbed dietary residues, 4. modification of bile salts by intestinal microflora and5. defects in the ileal uptake of bile salts [42-44]. In the CF mouse models, we did not find asignificant correlation between fecal bile salt secretion and fecal fat excretion, indicating thatunhydrolyzed triacylglycerols and phospholipids do not contribute to the increased fecal bilesalt loss. The other factors can not be excluded. Since intestinal histologic abnormalities havebeen observed in both ∆F508/∆F508 and in cftr -/- mice [11,14], we speculate that the mostlogical explanation for bile salt malabsorption in these two CF mouse models would be defectsin the ileal uptake of bile salts due to intestinal mucosal dysfunction or alterations in the mucuslayer [45].
In conclusion, in this study we have shown that cftr -/- mice, but not ∆F508/∆F508mice, have an impaired dietary fat absorption, which is not likely due to either decreased bilesalt pool size or altered bile composition. In both CF mouse models, fecal bile salt excretion

Fat (mal)absorption in CF mice
115
was increased, which was not secondary to increased fecal fat excretion. Bile composition dataindicate that the increased fecal loss of bile salts is compensated for by an increased bile saltneosynthesis.
References
1. Kalivianakis M, Minich DM, Bijleveld CMA, Aalderen WMC, Stellaard F, Laseur M,Vonk RJ, Verkade HJ. Fat malabsorption in pediatric cystic fibrosis patients on enzymereplacement therapy is due to impaired intestinal uptake of long chain fatty acids. Am JClin Nutr 1998 (In Press).
2. Rommens JM, Iannuzzi MC, Kerem B-S, Drumm ML, Melmer G, Dean M, Rozmahel R,Cole JL, Kennedy D, Hidaka N, Zsiga M, Buchwald M, Riordan JR, Tsui L-C, CollinsFS. Identification of the cystic fibrosis gene: chromosome walking and jumping. Science1989;245:1059-1065.
3. Riordan JR, Rommens JM, Kerem B-S, Alon N, Rozmahel R, Grzelczak Z, Zielenski J,Lok S, Plavsic N, Chou J-L, Drumm ML, Iannuzzi MC, Collins FS, Tsui L-C.Identification of the cystic fibrosis gene: cloning and characterization of complementaryDNA. Science 1989;245:1066-1073.
4. Kerem B-S, Rommens JM, Buchanan JA, Markiewicz D, Cox TK, Chakravarti A,Buchwald M, Tsui L-C. Identification of the cystic fibrosis gene: genetic analysis. Science1989;245:1073-1080.
5. Pencharz PB, Durie PR. Nutritional management of cystic fibrosis. Annu Rev Nutr1993;13:111-136.
6. Zemel BS, Kawchak DA, Cnaan A, Zhao H, Scanlin TF, Stallings VA. Prospectiveevaluation of resting energy expenditure, nutritional status, pulmonary function, andgenotype in children with cystic fibrosis. Pediatr Res 1996;40:578-586.
7. Cheng SH, Gregory RJ, Marshall J, Paul S, Souza DW, White GA, Riordan JR, SmithAE. Defective intracellular transport and processing of CFTR is the basis of most cysticfibrosis. Cell 1990;63:827-834.
8. Ward CL, Omura S, Kopito RR. Degradation of CFTR by the ubiquitin-proteasomepathway. Cell 1995;83:121-127.
9. Dorin JR. Development of mouse models for cystic fibrosis. J Inher Metab Dis1995;18:495-500.
10. Dickinson P, Dorin JR, Porteous DJ. Modelling cystic fibrosis in the mouse. Mol MedToday 1995;1:140-148.
11. Van Doorninck JH, French PJ, Verbeek E, Peters RHPC, Morreau H, Bijman J, ScholteBJ. A mouse model for the cystic fibrosis delta-F508 mutation. EMBO J 1995;14:4403-4411.
12. Grubb BR, Gabriel SE. Intestinal physiology and pathology in gene-targeted mousemodels of cystic fibrosis. Am J Physiol 1997;36:G258-G266.
13. Ip WF, Bronsveld I, Kent G, Corey M, Durie PR. Exocrine pancreatic alterations in long-lived surviving cystic fibrosis mice. Pediatr Res 1996;40:242-249.

Chapter 7
116
14. Ratcliff R, Evans MJ, Cuthbert AW, MacVinish LJ, Foster D, Anderson JR, ColledgeWH. Production of a severe cystic fibrosis mutation in mice by gene targeting. NatureGenet 1993;4:35-41.
15. Shalon LB, Adelson JW. Cystic fibrosis. Gastrointestinal complications and gene therapy.In: Lebenthal E, ed. The pediatric clinics of North America. Pediatric gastroenterology I.Philadelphia: W.B. Saunders Company, 1996:157-196.
16. Zentler-Munro PL, Fine DR, Batten JC, Northfield TC. Effect of cimetidine on enzymeinactivation, bile acid precipitation, and lipid solubilisation in pancreatic steatorrhoea dueto cystic fibrosis. Gut 1985;26:892-901.
17. Regan PT, Malagelada J-R, DiMagno EP, Go VLW. Reduced intraluminal bile acidconcentrations and fat maldigestion in pancreatic insufficiency: correction by treatment.Gastroenterology 1979;77:285-289.
18. Carroccio A, Pardo F, Montalto G, Iapichino L, Soreso M, Averna MR, Iacono G,Notarbartolo A. Use of famotidine in severe exocrine pancreatic insufficiency withpersistent maldigestion on enzymatic replacement therapy. A long-term study in cysticfibrosis. Dig Dis Sci 1992;37:1441-1446.
19. Watkins JB, Tercyak AM, Szczepanik P, Klein PD. Bile salt kinetics in cystic fibrosis:influence of pancreatic enzyme replacement. Gastroenterology 1977;73:1023-1028.
20. Roy CC, Weber EA, Morin CL, Combes J-C, Nusslé D, Mégevand A, Lasalle R.Abnormal biliary lipid composition in cystic fibrosis. Effect of pancreatic enzymes. N EngJ Med 1977;297:1301-1305.
21. Zentler-Munro PL, FitzPatrick WJF, Batten JC, Northfield TC. Effect of intrajejunalacidity on aqueous phase bile acid and lipid concentrations in pancreatic steatorrhoea dueto cystic fibrosis. Gut 1984;25:500-507.
22. Belli DC, Levy E, Darling P, Leroy C, Lepage G. Taurine improves the absorption of afat meal in patients with cystic fibrosis. Pediatrics 1987;80:517-523.
23. Eggermont E, De Boeck K. Small intestinal abnormalities in patients with cystic fibrosis.Eur J Pediatr 1991;150:824-828.
24. Corey M, McLaughlin FJ, Williams M, Levison H. A comparison of survival, growth, andpulmonary function in patients with cystic fibrosis in Boston and Toronto. J ClinEpidemiol 1988;41:583-591.
25. Lepage G, Roy CC. Direct transesterification of all classes of lipids in a one-step reaction.J Lipid Res 1986;27:114-120.
26. Murphy GM, Billing BH, Baron DN. A fluorimetric and enzymatic method for theestimation of serum total bile acids. J Clin Pathol 1970;23:594-598.
27. Setchell KDR, Worthington J. A rapid method for the quantitative extraction of bile acidsand their conjugates from serum using commercially available reverse-phaseoctadecylsilane bonded silica cartridges. Clin Chim Acta 1982;125:135-144.
28. Kalek H-D, Stellaard F, Kruis W, Paumgartner G. Detection of increased bile acidexcretion by determination of bile acid content in single stool samples. Clin Chim Acta1984;140:85-90.
29. O'Brien SM, Campbell GR, Burke AF, Maguire OC, Rowlands BJ, FitzGerald MX,Hegarty JE. Serum bile acids and ursodeoxycholic acid treatment in cystic fibrosis-relatedliver disease. Eur J Gastroenterol Hepatol 1996;8:477-483.

Fat (mal)absorption in CF mice
117
30. Colombo C, Battezzati PM, Podda M, Bettinardi N, Giunta A, the Italian Group for theStudy of Ursodeoxycholic Acid in Cystic Fibrosis. Ursodeoxycholic acid for liver diseaseassociated with cystic fibrosis: a double-blind multicenter trial. Hepatology 1996;23:1484-1490.
31. Roy CC, Weber AM, Morin CL, Lepage G, Brisson G, Yousef I, Lasalle R. Hepatobiliarydisease in cystic fibrosis: a survey of current issues and concepts. J Pediatr GastroenterolNutr 1982;1:469-478.
32. Colombo C, Battezzati PM, Podda M. Hepatobiliary disease in cystic fibrosis. SeminLiver Dis 1994;14:259-269.
33. Robb TA, Davidson GP, Kirubakaran C. Conjugated bile acids in serum and secretion inresponse to cholecystokinin/secretin stimulation in children with cystic fibrosis. Gut1985;26:1246-1256.
34. Mills CL, Dorin JR, Davidson DJ, Porteus DJ, Alton EWFW, Dormer RL, McPhersonMA. Decreased beta-adrenergic stimulation of glycoprotein secretion in CF micesubmandibular glands: reversal by the methylxanthine, IBMX. Biochem Biophys ResComm 1995;215:674-681.
35. Snouwaert JN, Brigman KK, Latour AM, Iraj E, Schwab U, Gilmour MI, Koller BH. Amurine model of cystic fibrosis. Am J Respir Crit Care Med 1995;151:S59-S64.
36. Clarke LL, Grubb BR, Gabriel SE, Smithies O, Koller BH, Boucher RC. Defectiveepithelial chloride transport in a gene-targeted mouse model of cystic fibrosis. Science1992;257:1125-1128.
37. French PJ, Van Doorninck JH, Peters RHPC, Verbeek E, Ameen NA, Marino CR, DeJonge HR, Bijman J, Scholte BJ. A delta F508 mutation in mouse cystic fibrosistransmembrane conductance regulator results in a temperature-sensitive processing defectin vivo. J Clin Invest 1996;98:1304-1312.
38. Strandvik B, Einarsson K, Lindblad A, Angelin B. Bile acid kinetics and biliary lipidcomposition in cystic fibrosis. J Hepatol 1996;25:43-48.
39. Lefebvre D, Ratelle S, Chartrand L, Roy CC. Reduced microbial transformation of bileacids in cystic fibrosis. Experientia 1977;33:616-618.
40. Dowling RH, Mack E, Small DM. Biliary lipid secretion and bile composition after acuteand chronic interruption of the enterohepatic circulation in the Rhesus monkey: IV.primate biliary physiology. J Clin Invest 1971;50:1917-1926.
41. Small DM, Dowling RH, Redinger RN. The enterohepatic circulation of bile salts. ArchIntern Med 1972;130:552-573.
42. Weber AM, Roy CC. Bile acid metabolism in children with cystic fibrosis. J Clin Invest1985;99:1880-1887.
43. Weber AM, Roy CC, Morin CL, Lasalle R. Malabsorption of bile acid in children withcystic fibrosis. N Eng J Med 1973;289:1001-1005.
44. Weber AM, Roy CC, Chartrand L. Relationship between bile acid malabsorption andpancreatic insufficiency in cystic fibrosis. Gut 1976;17:295-299.
45. O'Brien S, Mulcahy H, Fenlon H, O'Broin A, Casey M, Burke A, FitzGerald MX,Hegarty JE. Intestinal bile acid malabsorption in cystic fibrosis. Gut 1993;34:1137-1141.


CHAPTER 8
General discussion

Chapter 8
120
CHAPTER 8
General discussion
The two main processes involved in fat absorption are lipolysis and solubilization. The firstpart of this thesis deals with the process of lipolysis. Lipolysis is an important step in theoverall process of fat absorption and a shortage of lipase due to pancreatic insufficiency maylead to severely reduced levels of fat absorption. A rat model in which fat malabsorption iscaused by impaired lipolysis of dietary triacylglycerols has been developed and characterized(chapter 2). Impaired lipolysis was induced by feeding rats different doses of orlistat, aninhibitor of gastric and pancreatic lipase. It has been demonstrated that orlistat inactivateslipase enzymes by reacting covalently with serine (Ser-152) in the active site of the catalyticsubunit. Orlistat has been shown to reproducibly induce fat malabsorption in a dose-dependentfashion [1,2]. The percentage of total dietary fat absorption was examined upon feeding therats 4 different doses of orlistat, i.e. 0, 50, 200, and 800 mg orlistat per kg rat chow. Fatabsorption decreased in a dose-dependent way from 80.2 ± 2.2% in control rats (mean ±SEM) to 32.8 ± 3.7% when 800 mg orlistat per kg rat chow was added (P<0.001).

General discussion
121
A potential test for the diagnosis of impaired lipolysis would be the 13C-MTG breathtest, which was previously described by Vantrappen et al. [3]. The advantage of the 13C-MTGis its sensitivity to pancreatic lipase activity, the most important enzyme with respect tohydrolysis of triacylglycerols. The principle of the 13C-MTG breath test is based on lipolysis-dependent 13CO2 excretion via the breath. The relationship between the extent of fatmalabsorption and the recovery of 13CO2 in breath after oral ingestion of 13C-MTG wasinvestigated in control and orlistat-fed rats. Percentage of 13CO2 in breath was examined uponfeeding the rats 0, 50, 200, and 800 mg orlistat per kg rat chow. A significant correlation(r=0.88, P<0.001) was observed between percentage of total fat absorption and 6-h recoveryof 13CO2 in breath. The correlation was especially strong in rats having major fatmalabsorption, indicating that the 13C-MTG breath test can be used as a tool to detectimpaired lipolysis when fat malabsorption is severe. However, in rats with fat absorptionhigher than 75% the coefficient of variation of cumulative breath 13CO2 excretion was large(15%) compared to that of fat absorption (5%). Thus, even under controlled circumstances ina homogeneous group of rats with similar genetic background and standard diet, aconsiderable variation in 13CO2 expiration was observed. Potential causes of this largevariation may be differences in gastric emptying, hepatic clearance and metabolism, β-oxidation, endogenous CO2 production and pulmonary excretion, since all these factors mayinfluence the recovery of 13CO2 in the breath. In summary, when fat malabsorption is severe,impaired lipolysis is evidently the rate-limiting step, which can be identified by the 13C-MTGbreath test. However, upon rather mild fat malabsorption, the rate-limiting step of 13CO2
expiration is shifted from impaired lipolysis to one of the other mentioned factors, resulting inlarge variations upon application of the 13C-MTG breath test. Although no direct extrapolationfrom rats to humans can be performed, these results indicate the diagnostic limitations of the13C-MTG breath test in subjects with mild to moderate fat malabsorption.
The mechanistic studies were extended by characterization of the 13C-MTG breathtest in healthy adults (chapter 3). The effects of various test conditions on the 13CO2 responsehave only partially been elucidated. Therefore, it was determined which factors, apart frompancreatic insufficiency, may influence the quantitative recovery of 13CO2 in breath. Thephysiological variation of the 13CO2 response in healthy human adults was examined byperforming the test twice under exactly the same test circumstances within 4 weeks. Therepeatability was calculated according to Bland and Altman [4] and it was found that when the13C-MTG breath test is repeated in the same individual, the results of both tests areconsiderably different. In order to explore whether the application of the 13C-MTG breath testcould be simplified or the sensitivity improved, the following factors were studied: the effectof two different test meals on the 13CO2 response, the effect of an additional meal during thetest, and the effect of physical exercise during the test. A variety of test meals has beendescribed for breath tests with a diversity of substrates [3,5-7]. So far, no standardized testmeal for clinical purpose of these breath tests has been proposed. A disadvantage of a testmeal containing bread and butter could be the extended time it would take for children toconsume it, and the risk of not consuming it quantitatively. Also, such a test meal is notapplicable to small infants. It was examined whether the 13CO2 response of a liquid test meal(75 mL cream) is similar to the 13CO2 response of a solid test meal (2 slices of bread and 25 g

Chapter 8
122
butter). The results suggest that these two distinct test meals give a similar cumulative 13CO2
response. Since it may be cumbersome to keep patients, in particular infants, fasted for severalhours, the effect of consuming an extra meal during the test on the 13CO2 response wasexamined. The extra meal was ingested 3 h after the start of the experiment and consisted of 2slices of bread and 30 g of strawberry jam. It appeared that stringency on continuous fastingduring the test is unnecessary, which is in favor of the applicability of the test in pediatricpatients. It is known that physical activity considerably affects the production rate of CO2 andnutrient oxidation [8-10], however, it is not established to what extent it influences the resultsof the 13C-MTG test. Subjects were asked to perform moderate exercise (50 Watt) during thefirst 5 h of the test on a bicycle ergometer. Most subjects showed a large increase in their13CO2 recovery in the breath, and thus standardization of resting conditions still seemspreferable.
A second important step in the overall process of fat absorption is solubilization ofthe lipolytic products by bile components. A rat model has been developed and characterizedin which fat malabsorption is induced by long-term bile diversion (chapter 4). Bile diversionwas achieved by providing rats with a permanent catheter in the bile duct in order to interruptthe enterohepatic circulation. This experimental model allows for physiological studies inunanesthetized rats with long-term bile diversion without the interference of stress or restraint.After 6 days of bile diversion, no bile is available in the intestine for the formation of mixedmicelles, and theoretically, lipid absorption is expected to be decreased. When rats were fed astandard diet (14 en% fat), however, percentage of dietary fat absorption still appeared to beefficient and decreased only from 96.7 ± 0.2% in control rats to 87.2 ± 0.9% (P<0.001) inbile-diverted rats. In order to challenge the absorptive system of the rat, fat absorption wasalso studied when rats were fed a high-fat diet (35 en% fat). Percentage of total dietary fatabsorption again was highly efficient in control rats (93.2 ± 0.4%), but was considerablydecreased in bile-diverted rats (53.9 ± 3.9%, P<0.001). The presently characterized rat modelallows the systematic evaluation of the quantitative role of the various bile components onintestinal lipid absorption, in particular by reconstitution experiments involving the continuousinfusion of model bile solutions. In addition, this rat model will allow to evaluate the potencyof novel therapeutic approaches/drugs to improve lipid absorption under conditions ofimpaired bile formation.
A potential substrate for the diagnosis of impaired solubilization would be a 13C-labeled long-chain fatty acid. A 13C-labeled long-chain fatty acid test measures the wholeprocess of uptake of long-chain fatty acids: solubilization of lipolytic products by bilecomponents and translocation of the fatty acids over the intestinal mucosa. In this thesis, thesubstrate [1-13C]palmitic acid was selected, because palmitic acid is the most predominantsaturated fatty acid in the Western diet. The potency of the [1-13C]palmitic acid test toquantify fat malabsorption due to impaired intestinal uptake of long-chain fatty acids wasinvestigated in chronically bile-diverted rats (chapter 4). So far, the use of 13C-labeled long-chain fatty acids for quantitative studies on defective fat absorption has been limited to breathand feces analysis [5,11,12]. The excretion rate of 13C in the form of exhaled 13CO2, however,does not necessarily reflect quantitative differences in the absorption of the 13C-labeled parentcompound, e.g., due to variations in the post-absorptive metabolism [7,13]. The determination

General discussion
123
of plasma concentrations of absorbed 13C-labeled fats as a measure of their absorption offers atheoretical advantage over breath 13CO2 analysis, since numerous steps are involved in thepost-absorptive metabolism of the tracer prior to exhalation of 13CO2 [14]. After intraduodenaladministration of [1-13C]palmitic acid, plasma 13C-palmitic acid concentrations clearlydifferentiated between control rats and chronically bile-diverted rats. When 10% dose L-1
plasma was used as the lower limit of normal plasma values and 91% as the lower limit ofnormal dietary fat absorption, the test had a sensitivity and specificity of 100% for detectingfat malabsorption under the test conditions used. The results were essentially similar on thestandard and high-fat diet, emphasizing the potency of the [1-13C]palmitic acid absorption testto detect impaired intestinal uptake of long-chain fatty acids.
In order to investigate the sensitivity of the [1-13C]palmitic acid test in humans, thetest was applied to a group of healthy volunteers in which fat absorption was slightly reducedby dietary supplementation of calcium (chapter 5). It has been shown that calcium can forminsoluble precipitates, consisting of calcium, phosphate, bile acids and long-chain fatty acids[15,16]. Oral calcium supplementation to healthy subjects has been reported to increase fatexcretion via the feces in a dose-dependent fashion [15,17-20]. Upon calcium administrationfat absorption significantly decreased from 96.6 ± 0.6% to 94.9 ± 0.9% (P<0.05). Thus, oralcalcium supplementation in humans seems to be a method to reduce fat absorption to a minorextent. It was examined whether this effect could be quantified by means of orallyadministered [1-13C]palmitic acid. Upon calcium administration, plasma 13C-palmitic acidconcentrations after 8 h were significantly increased when compared to control values, yet,cumulative expiration of 13CO2 was significantly decreased. This discrepancy between theresults of the [1-13C]palmitic acid test in plasma and breath indicates that post-absorptivemetabolism is changed upon calcium supplementation. Percentage of dietary fat absorption didnot correlate to either breath 13CO2 recovery or plasma 13C-palmitic acid concentrations.Although calcium supplementation clearly affects the outcomes of the [1-13C]palmitic acidtest, present data do not indicate that the test is sensitive enough to reliably quantify this smalldegree of fat malabsorption in human adults.
A frequently encountered disorder in Caucasian populations associated with fatmalabsorption is cystic fibrosis. The pathophysiology of fat malabsorption in human cysticfibrosis patients may involve both pancreatic insufficiency and bile acid deficiency. However,so far it has not been possible to determine in the individual patient which of the processes israte-limiting, especially not when patients are supplemented with pancreatic enzymes. In orderto obtain more insight into the impaired processes of fat malabsorption in cystic fibrosis weperformed a study in pediatric cystic fibrosis patients treated with their usual pancreaticenzyme replacement therapy (chapter 6). The substrates 13C-MTG and uniformly labeled 13C-linoleic acid were both applied to determine whether the rate-limiting step behind their fatmalabsorption was either impaired lipolysis or impaired intestinal uptake of long-chain fattyacids, respectively. 13C-linoleic acid was selected as the substrate because, theoretically, itcould provide information on the essential fatty acid status and metabolism of the patients. The13C-linoleic acid test and the 13C-MTG breath test were both applied to 10 pediatric cysticfibrosis patients receiving their habitual pancreatic enzymes. During the test days, a fat balancewas performed for 3 days to determine dietary fat absorption. Fecal fat excretion ranged from

Chapter 8
124
5.1 to 27.8 g day-1 and fat absorption ranged from 79 to 93%. After ingestion of 13C-MTG norelationship was observed between breath 13CO2 recovery and dietary fat absorption (r=0.04).In contrast, a strong relationship was observed between 8-h plasma 13C-linoleic acidconcentrations and dietary fat absorption after ingestion of 13C-linoleic acid (r=0.88,P<0.001). Our results suggest that fat malabsorption in cystic fibrosis patients on enzymereplacement therapy is not likely due to insufficient lipolytic enzyme activity, but rather due todefective intestinal uptake of long-chain fatty acids. Therefore, therapeutic attempts tonormalize fat absorption in cystic fibrosis patients need to include a strategy to improveintestinal uptake of long-chain fatty acids.
Impaired intestinal uptake of long-chain fatty acids may be caused by (combinationsof) processes such as altered bile composition, bile salt precipitation, decreased intestinal bilesalt concentration, small bowel mucosal dysfunction and/or alterations in the mucus layer [21-25]. In order to obtain more mechanistic insight into the involved processes, we studied twocystic fibrosis mouse models (chapter 7). The two cystic fibrosis mouse models are: 1. micewith the ∆F508 mutation in the cftr gene, ∆F508/∆F508 (129/FVB genetic background), and2. mice with complete inactivation of the cftr gene, cftr -/- (129/C57/Bl6 genetic background)[26,27]. The basic defect in cystic fibrosis lies in the cystic fibrosis transmembrane regulator(CFTR), a protein responsible for chloride ion transport. In ∆F508/∆F508 mice, thebiosynthetic processing of the cftr gene product to its mature glycosylated form is disrupted[28], so that the protein is retained in the endoplasmic reticulum and is then degraded [29].However, it has been observed that ∆F508/∆F508 mice exhibit residual cftr activity in thegallbladder and ileum [26]. In cftr -/- mice, cftr function is completely abolished, and epithelialack cftr in the apical plasma membrane and, therefore, lack cAMP-stimulated Cl- permeability[30]. Fat absorption was studied after feeding the mice a standard (14 en% fat) or a high-fat(35 en% fat) diet for 2 weeks. In ∆F508/∆F508 mice, dietary fat absorption was similarcompared with controls on both diets (standard diet: 94.8 ± 0.7% compared to 95.5 ± 0.6%,respectively; high-fat diet: 93.8 ± 2.2% compared to 95.3 ± 2.4 %, respectively). Absorptionof dietary fats by cftr -/- mice, however, was significantly less efficient when compared withtheir control counterparts (standard diet: 82.8 ± 3.0% compared to 93.9 ± 1.3%, respectively,P<0.01; high-fat diet: 88.8 ± 1.6% compared to 95.0 ± 1.4%, respectively, P<0.01). Thesedata indicate that the complete disruption of the cftr gene leads to moderate fat malabsorption,in contrast to introduction of the ∆F508 mutation. However, it can not be completely excludedthat the observed difference in fat malabsorption is also influenced by the different geneticbackground of the mice. In order to study the processes behind defective uptake of long-chainfatty acids in more detail, biliary secretion of bile salts was determined after cannulation of thegallbladder for 80 min and fecal bile salt excretion was determined. Biliary bile salt pool sizesand biliary bile salt secretion rates were similar for the CF mouse models and their respectivecontrols on either diet, indicating that this is not the reason for fat malabsorption in cftr -/-mice. Fecal bile salt excretion was increased in ∆F508/∆F508 and in cftr -/- mice whencompared with their respective controls (10 versus 5 µmol g -1 feces, respectively, P<0.01). Nosignificant correlation was observed between fecal bile salt excretion and fecal fat, indicatingthat the increased excretion of fecal bile salts is not secondary to fat malabsorption. Biliary bilesalts in ∆F508/∆F508, cftr -/- and control mice were predominantly composed of cholate and

General discussion
125
ß-muricholate, with minor contributions of deoxycholate, ursodeoxycholate andhyodeoxycholate. Significantly increased cholate and decreased deoxycholate concentrationswere observed in bile of ∆F508/∆F508 and cftr -/- mice when compared with their controls.This result is consistent with the finding of increased fecal bile salt loss, which upregulatesneosynthesis of bile salts thereby resulting in higher amounts of primary bile salts anddecreased amounts of secondary bile salts. In conclusion, in this study we have shown that cftr-/- mice, but not ∆F508/∆F508 mice, have an impaired dietary fat absorption, which is notlikely due to either decreased bile salt pool size or altered bile composition. In both CF mousemodels, fecal bile salt excretion was increased, which was not secondary to increased fecal fatexcretion. Bile composition data indicate that the increased fecal loss of bile salts iscompensated for by an increased bile salt neosynthesis. Further investigations are needed toestablish whether impaired pancreatic function, intestinal mucosal dysfunction or alterations inthe intestinal mucosa are responsible for the observed fat malabsorption in cftr -/- mice.
In summary, in this thesis more information is obtained with respect to the variouspathophysiological processes involved in fat malabsorption. Various animal models for lipidmalabsorption were described and characterized: orlistat-fed rats to study impaired lipolysis,bile-diverted rats to study bile deficiency, and transgenic and knock-out mice for the study ofcystic fibrosis. These animal models are expected to be of significant importance to investigatethe potency of novel diagnostic and therapeutic strategies in the near future. In addition, thepotency of diagnostic tests such as the 13C-MTG breath test and the 13C-palmitic acid test wasinvestigated to characterize the etiology behind fat malabsorption in animal models and inhumans. The present data in cystic fibrosis patients and mice do not only open the possibilityto determine pathophysiological mechanisms of lipid malabsorption in individual patients, butthey also give way for titration of therapy to individual patients.
References
1. Isler D, Moeglen C, Gains N, Meier MK. Effect of the lipase inhibitor orlistat and ofdietary lipid on the absorption of radiolabelled triolein, tri-gamma-linolenin and tripalmitinin mice. Br J Nutr 1995;73:851-862.
2. Zhi J, Melia AT, Guerciolini R, Chung J, Kinberg J, Hauptman JB, Patel IH.Retrospective population-based analysis of the dose-response (fecal fat excretion)relationship of orlistat in normal and obese volunteers. Clin Pharmacol Ther 1994;56:82-85.
3. Vantrappen GR, Rutgeerts PJ, Ghoos YF, Hiele MI. Mixed triglyceride breath test: Anoninvasive test of pancreatic lipase activity in the duodenum. Gastroenterology1989;96:1126-1134.
4. Bland JM, Altman DG. Statistical methods for assessing agreement between two methodsof clinical measurement. Lancet 1986;307-310.
5. Watkins JB, Klein PD, Schoeller DA, Kirschner BS, Park R, Perman JA. Diagnosis anddifferentiation of fat malabsorption in children using 13C-labeled lipids: trioctanoin, trioleinan palmitic acid breath tests. Gastroenterology 1982;82:911-917.

Chapter 8
126
6. Murphy MS, Eastham EJ, Nelson R, Aynsley-Green A. Non-invasive assessment ofintraluminal lipolysis using a 13CO2 breath test. Arch Dis Child 1990;65:574-578.
7. Murphy JL, Jones A, Brookes S, Wootton SA. The gastrointestinal handling andmetabolism of [1-13C]palmitic acid in healthy women. Lipids 1995;30:291-298.
8. Brooks GA, Mercier J. Balance of carbohydrate and lipid utilization during exercise: the"crossover" concept. J Appl Physiol 1994;76:2253-2261.
9. Coyle EF. Substrate utilization during exercise in active people. Am J Clin Nutr1995;61:968S-979S.
10. Romijn JA, Coyle EF, Sidossis LS, Gastaldelli A, Horowitz JF, Endert E, Wolfe RR.Regulation of endogenous fat and carbohydrate metabolism in relation to exerciseintensity and duration. Am J Physiol 1993;265:E380-E391.
11. Murphy JL, Jones AE, Stolinski M, Wootton SA. Gastrointestinal handling of [1-13C]palmitic acid in healthy controls and patients with cystic fibrosis. Arch Dis Child1997;76:425-427.
12. Odeka EB, Miller V. Evaluation of fat utilization in paediatric Crohn's disease using 13C-labelled fat. J Pediatr Gastroenterol Nutr 1995;21:430-434.
13. Kalivianakis M, Verkade HJ, Stellaard F, Van der Werf M, Elzinga H, Vonk RJ. The 13C-mixed triglyceride breath test in healthy adults: determinants of the 13CO2 response. Eur JClin Invest 1997;27:434-442.
14. Pedersen NT, Jorgensen BB, Rannem T. The [14C]-triolein breath test is not valid as a testof fat absorption. Scand J Clin Lab Invest 1991;51:699-703.
15. Saunders D, Sillery J, Chapman R. Effect of calcium carbonate and aluminium hydroxideon human intestinal function. Dig Dis Sci 1988;33:409-413.
16. Meer Rv, Welberg JWM, Kuipers F, Kleibeuker JH, Mulder NH, Termont DSML, VonkRJ, De Vries HT, De Vries EGE. Effects of supplemental dietary calcium on the intestinalassociation of calcium, phosphate, and bile acids. Gastroenterology 1990;99:1653-1659.
17. Govers MJAP, Termont DSML, Lapré JA, Kleibeuker JH, Vonk RJ, Meer Rv. Calcium inmilk products precipitates intestinal fatty acids and secondary bile acids and thus inhibitscolonic cytotoxicity in humans. Cancer Res 1996;56:3270-3275.
18. Welberg JWM, Monkelbaan JF, De Vries EGE, Muskiet FAJ, Cats A, Oremus ETHGJ,Boersma-van Ek W, Van Rijsbergen H, Meer Rv, Mulder NH, Kleibeuker JH. Effects ofsupplemental dietary calcium on quantitative and qualitative fecal fat excretion in man.Ann Nutr Metab 1994;38:185-191.
19. Denke MA, Fox MM, Schulte MC. Short-term dietary calcium fortification increases fecalsaturated fat content and reduces serum lipids in men. J Nutr 1996;123:1047-1053.
20. Potter SM, Kies CV, Rojhani A. Protein and fat utilization by humans as affected bycalcium phosphate, calcium carbonate, and manganese gluconate supplements. Nutrition1990;6:309-312.
21. Watkins JB, Tercyak AM, Szczepanik P, Klein PD. Bile salt kinetics in cystic fibrosis:influence of pancreatic enzyme replacement. Gastroenterology 1977;73:1023-1028.
22. Roy CC, Weber EA, Morin CL, Combes J-C, Nusslé D, Mégevand A, Lasalle R.Abnormal biliary lipid composition in cystic fibrosis. Effect of pancreatic enzymes. N EngJ Med 1977;297:1301-1305.

General discussion
127
23. Zentler-Munro PL, FitzPatrick WJF, Batten JC, Northfield TC. Effect of intrajejunalacidity on aqueous phase bile acid and lipid concentrations in pancreatic steatorrhoea dueto cystic fibrosis. Gut 1984;25:500-507.
24. Belli DC, Levy E, Darling P, Leroy C, Lepage G. Taurine improves the absorption of afat meal in patients with cystic fibrosis. Pediatrics 1987;80:517-523.
25. Carroccio A, Pardo F, Montalto G, Iapichino L, Soreso M, Averna MR, Iacono G,Notarbartolo A. Use of famotidine in severe exocrine pancreatic insufficiency withpersistent maldigestion on enzymatic replacement therapy. A long-term study in cysticfibrosis. Dig Dis Sci 1992;37:1441-1446.
26. Van Doorninck JH, French PJ, Verbeek E, Peters RHPC, Morreau H, Bijman J, ScholteBJ. A mouse model for the cystic fibrosis delta-F508 mutation. EMBO J 1995;14:4403-4411.
27. Trezise AEO, Ratcliff R, Hawkins TE, Evans MJ, Freeman TC, Romano PR, Higgins CF,Colledge WH. Co-ordinate regulation of the cystic fibrosis and multidrug resistance genesin cystic fibrosis knockout mice. Hum Mol Genet 1997;6:527-537.
28. Cheng SH, Gregory RJ, Marshall J, Paul S, Souza DW, White GA, Riordan JR, SmithAE. Defective intracellular transport and processing of CFTR is the basis of most cysticfibrosis. Cell 1990;63:827-834.
29. Ward CL, Omura S, Kopito RR. Degradation of CFTR by the ubiquitin-proteasomepathway. Cell 1995;83:121-127.
30. Grubb BR, Gabriel SE. Intestinal physiology and pathology in gene-targeted mousemodels of cystic fibrosis. Am J Physiol 1997;36:G258-G266.


SAMENVATTING

Samenvatting
130
SAMENVATTING
Vetten in de voeding zijn belangrijk voor het menselijk lichaam als bronnen van energie en alsbouwstenen, vooral in periodes van groei en ontwikkeling. Gewoonlijk is het menselijklichaam in staat om voedingsvetten zeer efficiënt op te nemen; meer dan 96% van devoedingsvetten wordt opgenomen in het lichaam. Er zijn echter verschillende aandoeningenwaarbij de absorptie van voedingsvetten verstoord is. Voor een goede behandeling vanpatiënten met deze zogenaamde vetmalabsorptiesyndromen is inzicht in het vetabsorptieprocesessentieel. Het proces dat verantwoordelijk is voor de opname van vetten uit de voeding speeltzich af in het maagdarmkanaal. Allereerst worden in de maag en in het duodenum devoedingsvetten (voornamelijk triglyceriden) gesplitst in vetzuren en monoacylglyceriden. Ditsplitsingsproces, dat bekend is onder de naam lipolyse, wordt gekatalyseerd door lipase-enzymen afkomstig uit de maag en de pancreas. Vervolgens worden deze gesplitste vetdeeltjesopgelost in de waterige omgeving van de dunne darm middels een proces dat bekend staat alssolubilisatie. Hiertoe worden micellen gevormd die bestaan uit galcomponenten, zoalsgalzouten en fosfolipiden, en de vetdeeltjes. Vooral lange-keten vetzuren zijn sterk afhankelijkvan een efficiënte solubilisatie, in tegenstelling tot middellange-keten vetzuren, omdat lange-keten vetzuren slecht oplosbaar zijn in water. Tenslotte worden de vetdeeltjes getransporteerdover de celwand van de dunne darm, waarna ze terecht komen in de bloedbaan. De vetten dieniet worden opgenomen in het lichaam, worden uitgescheiden via de ontlasting. Tot nu toewerd vooral aandacht besteed aan de efficiëntie van het totale proces van vetabsorptie,waardoor het inzicht in de bijdrage van de onderliggende mechanismes aan de vetmalabsorptiebeperkt is. Een gedetailleerder inzicht in deze onderliggende mechanismes zou niet alleenkunnen leiden tot verbeteringen in diagnostische methodes, maar ook tot verbeteringen inbehandelingsmethoden voor patiënten met vetmalabsorptie. Het doel van het huidigeonderzoek was dan ook om meer informatie te verkrijgen over de verschillendepathofysiologische processen die betrokken zijn bij vetmalabsorptie en om de diagnostiek voorvetmalabsorptie te verbeteren.

Samenvatting
131
Om deze doelstelling te verwezenlijken zijn studies uitgevoerd met proefdieren,gezonde proefpersonen en patiënten. De studies werden zo opgezet dat, naast de totalevetabsorptie, ook de processen lipolyse en solubilisatie werden onderzocht. De methode die inde kliniek gebruikt wordt om de totale hoeveelheid van geabsorbeerde vetten te bepalen is devetbalans. Dit houdt in dat patiënten gedurende drie dagen in voedseldagboeken alles noterenwat ze eten, waardoor de totale vetinname kan worden bepaald. In deze periode verzamelende patiënten ook al hun ontlasting, waarin de niet-geabsorbeerde hoeveelheid vetten kanworden bepaald. Op deze manier kan dan de hoeveelheid geabsorbeerde vetten uitgedruktworden als percentage van de vetinname. Bijvoorbeeld, als de vetinname gedurende drie dagen100 g per dag is en de hoeveelheid vet in de ontlasting gedurende deze periode 5 g per dag is,dan betekent dit dat de vetabsorptie 95% is. Met behulp van deze methode kan echter geeninformatie over de achterliggende mechanismes (lipolyse en solubilisatie) worden verkregen.Deze informatie kan wel verkregen worden door gebruik te maken van testen waarbij vettenop een onschadelijke manier gelabeld worden met stabiele isotopen. In dit onderzoek zijn dediagnostische capaciteiten van twee verschillende soorten testen beschreven, namelijk een testvoor het diagnosticeren van verstoorde lipolyse en een test voor het diagnosticeren vanproblemen bij solubilisatie.
Verstoorde lipolyse kan worden aangetoond met behulp van de 13C-MTG ademtest.13C-MTG is de afkorting van het gelabelde vet waarvan de volledige chemische naam luidt:1,3-distearoyl, 2[carboxyl-13C]octanoyl glycerol. Na inname van dit substraat splitsen lipase-enzymen, afkomstig van de pancreas, de twee vetzuurketens van het molekuul af (=lipolyse),waarna het 13C-gelabelde restmolekuul opgenomen kan worden in het lichaam. Omdatrestmolekuul een middellange-keten vetzuur is, is de opname van het molekuul niet afhankelijkvan solubilisatie. Na opname wordt dit 13C-gelabelde restmolekuul grotendeels verbrand in hetlichaam en vervolgens uitgeademd als 13CO2. Dus, het principe van de 13C-MTG ademtest isgebaseerd op het feit dat, na inname van het substraat, de hoeveelheid 13CO2 die uitgeademdwordt, afhankelijk is van de activiteit van het pancreas lipase-enzym. Verstoorde solubilisatiekan worden aangetoond met behulp van lange-keten vetzuren (bijv. 13C-palmitinezuur of 13C-linolzuur). Dit zijn molekulen die niet meer gesplitst hoeven te worden door lipase-enzymen ende opname van deze molekulen in het lichaam is dus alleen afhankelijk van een goedesolubilisatie. Na inname van bijvoorbeeld het substraat 13C-palmitinezuur kan de door hetlichaam opgenomen hoeveelheid in het plasma gemeten worden als 13C-gelabeld palmitinezuuren na verbranding in de adem als 13CO2. De hoeveelheid die niet wordt geabsorbeerd, wordtuitgescheiden in de ontlasting en kan eveneens gemeten worden. Als het solubilisatieprocesgoed werkt, zal er veel 13C-gelabeld palmitinezuur aanwezig zijn in plasma en veel 13CO2 in deadem, maar weinig 13C-palmitinezuur in de ontlasting.
In het eerste deel van dit proefschrift werd het proces lipolyse onderzocht. Om ditproces in detail te kunnen bestuderen werd allereerst een proefdiermodel ontwikkeld engekarakteriseerd voor vetmalabsorptie ten gevolge van verstoorde lipolyse (hoofdstuk 2).Verstoorde lipolyse werd bewerkstelligd door ratten via hun gewone voer een stofje toe tedienen, orlistat, dat de activiteit van lipase-enzymen vermindert. De mate van vetmalabsorptiebleek afhankelijk te zijn van de toegediende hoeveelheid orlistat. Bij deze ratten werdonderzocht of vetmalabsorptie kon worden aangetoond met behulp van de 13C-MTG

Samenvatting
132
ademtest. Er bleek een sterk verband te bestaan tussen het percentage vetabsorptie en detotale hoeveelheid 13CO2 in de adem. Echter, als er sprake was van slechts een lichtevetmalabsorptie was, dan bleek er toch een grote variatie in de hoeveelheid 13CO2 in de ademte zijn. Dit zou kunnen komen doordat bij een milde vorm van verstoorde lipolyse de 13CO2
vorming niet alleen afhankelijk is van de lipolysesnelheid maar ook van factoren alsmaagontlediging en verbrandingssnelheid. Dit betekent dat de 13C-MTG ademtestwaarschijnlijk minder geschikt is als diagnostisch middel indien vetmalabsorptie slechts ingeringe mate verstoord is. Om de toepassingsmogelijkheden van de 13C-MTG ademtest verderte onderzoeken werd deze test vervolgens uitgevoerd bij gezonde volwassenen (hoofdstuk 3).De volgende factoren werden bestudeerd: het effect van twee verschillende testmaaltijden opde 13CO2 respons in de adem, het effect van een extra maaltijd tijdens de test, en het effect vanlichamelijke oefening tijdens de test. Uit de experimenten met twee verschillendetestmaaltijden bleek dat geen significante verschillen werden gevonden met betrekking tot de13CO2 respons in de adem na een vloeibare testmaaltijd (75 ml slagroom) of een vastetestmaaltijd (2 sneeën tarwebrood en 25 g roomboter). Dit is vooral een voordeel voorzuigelingen, die nog geen vast voedsel kunnen innemen. Een extra maaltijd drie uur naaanvang van de test (2 sneeën tarwebrood en 25 g aardbeienjam), bleek de resultaten niet teveranderen. Dit is een voordeel voor kinderen omdat die vaak moeite hebben met vastengedurende een langere periode. Ten derde bleek dat lichamelijk activiteit tijdens de test weldegelijk de resultaten beïnvloedde en dat het dus belangrijk is om tijdens de test in rust teblijven.
De tweede stap in het proces van vetabsorptie die onderzocht werd in dit proefschriftis de solubilisatie van vetten. Om dit proces in detail te kunnen bestuderen werd ook hiervooreen proefdiermodel ontwikkeld en gekarakteriseerd (hoofdstuk 4). Verstoorde solubilisatiewerd gerealiseerd door langdurige galonderbreking. Dit houdt in dat ratten werden voorzienvan een catheter in de galgang, waardoor de gal niet meer de dunne darm instroomt, maarbuiten het lichaam van de rat wordt opgevangen. De gal is dan niet meer beschikbaar voorsolubilisatie van vetten in de dunne darm met als gevolg dat de vetten niet goed meergeabsorbeerd kunnen worden. In dit proefdiermodel werd onderzocht of verstoordesolubilisatie kon worden aangetoond met behulp van de 13C-palmitinezuur test. Deconcentraties 13C-palmitinezuur in plasma van gal-onderbroken ratten bleken duidelijk lager tezijn dan de concentraties 13C-palmitinezuur in plasma van controle ratten. De test bleek zeergeschikt te zijn voor het aantonen van een sterk solubilisatieprobleem bij ratten. Om degevoeligheid van de test verder te bepalen werden testen gedaan bij gezonde volwassenenwaarbij een zeer licht vetmalabsorptie was bewerkstelligd ten gevolge van verstoordesolubilisatie (hoofdstuk 5). Dit werd gerealiseerd door de proefpersonen dagelijks 2 g calciumtoe te dienen in de vorm van calcium carbonaat. Calcium bindt aan galzouten, waardoor dezeniet meer beschikbaar zijn voor solubilisatie. Vervolgens werd onderzocht of deze mildevetmalabsorptie kon worden aangetoond met behulp van de [1-13C]palmitinezuur test. Bij deproefpersonen werden zowel adem- als plasmamonsters verzameld gedurende een periode van8 uur. Er bleek echter een discrepantie te zijn tussen de resultaten in het plasma en in de ademen de 13C-palmitinezuur test is dus blijkbaar niet gevoelig genoeg om een zeer lichte vorm vanvetmalabsorptie te detecteren.

Samenvatting
133
Vervolgens werden experimenten verricht om meer inzicht te verkrijgen in devetmalabsorptie die geassocieerd is met de genetische afwijking cystische fibrose. Cystischefibrose patiënten lijden vaak aan een pancreas insufficiëntie wat betekent dat onvoldoendepancreas lipase-enzymen in de dunne darm aanwezig zijn. Deze patiënten worden dan ookbehandeld door bij elke maaltijd lipase-enzymen in te nemen. Het is echter meerdere malenaangetoond dat cystische fibrose patiënten, ondanks behandeling met pancreasenzymen, nogsteeds een bepaalde mate van vetmalabsorptie overhouden. Een hogere dosispancreasenzymen is echter niet gewenst omdat dit kan leiden tot aantasting van de dikke darm.Het mechanisme van vetmalabsorptie in cystische fibrose patiënten behandeld met lipase-enzymen kan veroorzaakt worden door een inefficiënte enzymtherapie en/of een verstoordesolubilisatie. Om meer inzicht te krijgen in deze processen werd een studie uitgevoerd bijcystische fibrose patiënten die behandeld werden met pancreasenzymen (hoofdstuk 6). Desubstraten 13C-MTG en 13C-linolzuur werden gebruikt om te bepalen wat desnelheidsbepalende stap achter deze vetmalabsorptie was: respectievelijk een lipolysedefect ofverstoorde solubilisatie. Na inname van het 13C-MTG substraat werd geen verband gevondentussen de mate van vetabsorptie en de hoeveelheid 13CO2 in de adem. Dit betekent dat deverminderde vetabsorptie in deze patiënten waarschijnlijk niet veroorzaakt wordt door eenverstoorde lipolyse. Er bleek echter wel een sterk verband te bestaan tussen de mate vanvetabsorptie en de concentratie 13C-linolzuur in plasma na inname van 13C-linolzuur. Dus,patiënten met een relatief lage vetabsorptie hadden ook een lage hoeveelheid 13C-linolzuur inhun plasma. Deze resultaten suggereren dat de vetmalabsorptie in cystische fibrose patiëntendie behandeld worden met pancreasenzymen waarschijnlijk veroorzaakt wordt door eensolubilisatiedefect. Om vetabsorptie in cystische fibrose patiënten te bevorderen is het duszinvol om de therapie aan te passen met betrekking tot het solubilisatiedefect. Een verstoringin het solubilisatieproces kan echter verschillende oorzaken hebben, zoals verlaagdegalzoutconcentratie, andere galzoutsamenstelling, inactivatie van de galzouten door een lagepH in de dunne darm. De volgende stap in dit onderzoek was dan ook om meer inzicht teverkrijgen in dit proces bij cystische fibrose. Hiertoe werden studies uitgevoerd met muizendie genetisch gemanipuleerd waren aan het gen voor cystische fibrose (hoofdstuk 7). Bij dezemuizen werd onderzocht of vetmalabsorptie werd veroorzaakt door een verminderdegalzoutpool, een verlaagde galzoutconcentratie, en/of een andere galzoutsamenstelling. Ditbleek niet het geval te zijn. De muizen met cystische fibrose bleken wel verhoogdeconcentraties galzouten in de ontlasting te vertonen, een verschijnsel dat ook bij patiënten metcystische fibrose veel voorkomt. De oorzaak hiervan moet nog verder onderzocht worden.
Samenvattend kan worden gezegd dat diagnostische testen met stabiele isotopen, integenstelling tot de klassieke vetbalans, gedetailleerde informatie geven over de processenlipolyse en solubilisatie. Derhalve kan met behulp van deze testen gerichte farmacotherapie endieettherapie opgesteld worden voor patiënten met vetmalabsorptie. De 13C-MTG test kan bijsterke verstoring van de lipolyse goed gebruikt worden; bij marginale verstoring spelen ookandere experimentele condities een rol, die beter gekarakteriseerd dienen te worden. Met delange-keten vetzuurtest kunnen verstoringen in solubilisatie opgespoord worden. Dezebevinding kan met name van belang zijn voor de verdere behandeling van patiënten metcystische fibrose.


135
NAWOORD
Na ruim 4 jaar noeste arbeid sluit ik met het schrijven van deze allerlaatste pagina de periodeGroningen af. Alhoewel ik steeds heb geroepen dat ik Groningen een gezellige, knusse stadvind (al is het dan een beetje ver verwijderd van de rest van Nederland) en ik een erg leuke tijdin Groningen heb gehad, ben ik toch blij dat het nu dan eindelijk achter de rug is. In deafgelopen periode heb ik veel mensen weten te waarderen voor hun steun en gezelligheid. Eenaantal mensen wil ik hier daarom persoonlijk noemen. Allereerst wil ik graag Henkjan Verkadebedanken voor alle kennis die hij me heeft bijgebracht en voor zijn enthousiaste begeleiding.Mijn promotor Roel Vonk wil ik bedanken voor zijn zinnige suggesties met betrekking tot delijn van mijn onderzoek. Frans Stellaard heeft me vooral in het eerste jaar van mijn onderzoekgeholpen met alles wat er over stabiele isotopen te leren valt. Daarnaast zijn er natuurlijkontzettend veel mensen die me wegwijs hebben gemaakt op het laboratorium, achter decomputer, in de proefdierruimte, en in de Groningse kroegen en eetcafés. Heel hartelijkbedankt voor jullie professionele en emotionele bijdrage aan mijn promotieonderzoek. Wemoeten nog maar eens een avondje wat gaan drinken, lachen en kletsen zodat ik julliepersoonlijk kan bedanken.

Stellingen behorend bij het proefschriftMechanisms involved in malabsorption of dietary lipids
Mini KalivianakisGroningen, 23 september 1998
Severe fat malabsorption due to impaired lipolysis can be identified by the 13C-MTG breath test. However, in situations of mild fat malabsorption, considerableinterindividual variation in the results of the 13C-MTG breath test occurs, whichmay be explained by a shift in the rate-limiting step in the overall process of 13CO2
production.(DIT PROEFSCHRIFT)
The continuing fat malabsorption in cystic fibrosis patients on enzyme replacementtherapy is not likely due to insufficient lipolytic enzyme activity, but rather due toeither incomplete intraluminal solubilization and/or reduced mucosal uptake oflong-chain fatty acids.(DIT PROEFSCHRIFT)
Vrouwelijke wetenschappers moeten ongeveer 2½ maal beter presteren dan hunmannelijke collega’s voor een vergelijkbaar resultaat.(NATURE 1997;387:341)
Een promotieonderzoek is niet alleen een investering is in jezelf maar ook in deuniversiteit. Dat dit besef ook begint door te dringen tot de universitaire wereldblijkt uit het feit dat een aantal universiteiten bereid is de promovendi meer tebetalen.
Mini staat voor meer dan klein.
Ook voor stellingen geldt dat een hogere kwantiteit vaak niet ten goede komt aande kwaliteit.





























