Embed Size (px)
Citation preview

0
UNIVERSIDADE FEDERAL DE PELOTAS
Programa de Pós-Graduação em Ciência e Tecnologia de Alimentos
Tese
Características probióticas, tecnológicas e aspectos de segurança de
Lactobacillus curvatus P99 e produção, caracterização e aplicação de
suas substâncias antimicrobianas
Graciele Daiana Funck
Pelotas, 2016

1
Graciele Daiana Funck
Características probióticas, tecnológicas e aspectos de segurança de
Lactobacillus curvatus P99 e produção, caracterização e aplicação de
suas substâncias antimicrobianas
Trabalho de Tese apresentado ao
Programa de Pós-Graduação em Ciência
e Tecnologia de Alimentos da
Universidade Federal de Pelotas, como
requisito à obtenção do título de Doutor
em Ciência e Tecnologia de Alimentos.
Comitê de orientação: Profª. Drª. Ângela Maria Fiorentini
Prof. Dr. Wladimir Padilha da Silva
Pelotas, 2016

1

2
Graciele Daiana Funck
Características probióticas, tecnológicas e aspectos de segurança de
Lactobacillus curvatus P99 e produção, caracterização e aplicação de
suas substâncias antimicrobianas
Tese aprovada, como requisito, para obtenção do grau de Doutora em Ciência e Tecnologia de Alimentos, Programa de Pós-Graduação em Ciência e Tecnologia de Alimentos, Faculdade de Agronomia Eliseu Maciel, Departamento de Ciência e Tecnologia Agroindustrial, Universidade Federal de Pelotas.
Data da Defesa: 26/04/2016.
Banca examinadora:
_______________________________________ Profa. Dra. Ângela Maria Fiorentini
Doutora em Ciências dos Alimentos – UFSC
_______________________________________ Prof. Dr. Eliezer Ávila Gandra
Doutor em Ciência e Tecnologia Agroindustrial - UFPel
_______________________________________ Profa. Dra. Leidi Daiana Preichardt
Doutora em Ciência e Tecnologia Agroindustrial – UFPel
_______________________________________ Profa. Dra. Liziane Schittler
Doutora em Ciência e Tecnologia Agroindustrial – UFPel
_______________________________________ Profa. Dra. Simone Pieniz
Doutora em Microbiologia Agrícola e do Ambiente - UFRGS

3
AGRADECIMENTOS
A Deus por sempre abrir novas portas e proteger a minha vida.
A Universidade Federal de Pelotas e ao Departamento de Ciência e Tecnologia
Agroindustrial, pela oportunidade de realizar o curso.
A CAPES, CNPq e FAPERGS pela bolsa de doutorado e apoio financeiro ao
projeto de pesquisa.
A minha orientadora, Ângela Maria Fiorentini, em especial, pela amizade,
confiança, incentivo e orientação.
Ao meu co-orientador, Wladimir Padilha da Silva, pela orientação e
oportunidade.
A todos os professores do Departamento de Ciência e Tecnologia
Agroindustrial que contribuíram para a minha formação.
Aos membros da banca de qualificação e defesa de tese, pela disponibilidade e
enormes contribuições sugeridas para este trabalho.
Aos meus pais, meus irmãos e cunhados, pelo incentivo, compreensão e
paciência que souberam dispensar nesta caminhada. Sem eles nada seria
possível. Em especial, a minha mãe, exemplo de vida.
A minha sobrinha, Laura, mesma tão pequena, com seu lindo sorriso, inspirou-
me nos momentos finais desta etapa.
Aos colegas da Planta de Processamento de Produtos de Origem Animal do
DCTA/UFPel, Ana Rita, Camila, Claudio, Guilherme e Juliana, que além da
parceria no trabalho e ajuda no desenvolvimento da minha pesquisa, tornaram-
se grandes amigos, dividindo aflições e alegrias.
A minha “dupla loira”, Carla Pohl Sehn, pela amizade, cumplicidade, incentivo,
conhecimentos compartilhados, ajuda na pesquisa e muitos momentos loucura
e diversão.

4
Aos colegas do Laboratório de Microbiologia de Alimentos do DCTA/UFPel,
Andréia, Cristiano, Darla, Greici, Helena, Isabela, Louise, Maiara, Marcia,
Mariana, Natalie, Sônia e Tassiana pelo apoio, incentivo e momentos de
descontração.
As amigas e irmãs de coração, Cristiane Stöcker, Gislaine Hermanns e Leidi
Daiana Preichardt, pelo ombro amigo em todas as situações, pelo incentivo,
força e por toda a experiência e ensinamentos compartilhados.
E, a todos que de alguma forma colaboraram para a conclusão desta etapa de
minha jornada.
MUITO OBRIGADA!

5
RESUMO
FUNCK, Graciele Daiana. Características probióticas, tecnológicas e aspectos de segurança de Lactobacillus curvatus P99 e produção, caracterização e aplicação de suas substâncias antimicrobianas. 2016. 146f. Tese (Doutorado) – Programa de Pós-Graduação em Ciência e Tecnologia de Alimentos. Universidade Federal de Pelotas, Pelotas, 2016. O uso de novas culturas iniciadoras, com potencial industrial e funcional, têm sido explorado. Culturas funcionais podem ser utilizadas para melhorar e otimizar processos de produção de alimentos, bem como, características sensoriais, segurança e, ainda, promover benefícios à saúde. A aplicação de substâncias antimicrobianas naturais (como as bacteriocinas), combinada com diferentes tecnologias, promovem novas oportunidades no controle de patógenos, melhorando a qualidade e segurança dos alimentos. Filmes incorporados de bacteriocinas podem aumentar a proteção dos alimentos e evitar a contaminação pós-processamento. Assim, objetivou-se neste estudo, caracterizar o isolado Lactobacillus curvatus P99 quanto as suas características probióticas, tecnológicas e aspectos de segurança, bem como verificar as condições ótimas de multiplicação e produção de substâncias antimicrobianas (bacteriocin-like), caracterizar o sobrenadante livre de células (SLC) quanto a estabilidade ao tratamento térmico, diferentes pH e agentes químicos, quantificar sua ação antimicrobiana e toxicidade, produzir filmes antimicrobianos e aplicar estes filmes em um sistema alimentar. Testes fenotípicos, genotípicos e estatísticos foram utilizados para isto. Quanto as características probióticas, tecnológicas e segurança, o isolado L. curvatus P99 apresentou: tolerância ao trânsito gastrointestinal simulado quando protegido por alimento, autoagregação e coagregação com Listeria monocytogenes, incapacidade em aderir a células intestinais HCT-116, atividade antimicrobiana contra bactérias Gram-positivas e Gram-negativas, ausência da atividade β-galactosidase, multiplicação a 10 e 45°C e em 4,5 e 6% de NaCl, fermentação da glicose sem produção gás, hidrólise da caseína, inaptidão de atividade lipolítica, ausência de genes para resistência a antimicrobianos de uso clínico e para descarboxilação de aminoácidos e ausência da atividade β-hemolítica, DNAse e gelatinase. A partir da otimização das condições de multiplicação e produção de substâncias antimicrobianas (bacteriocin-like), obteve-se como condição ótima de multiplicação pH 6,3, a 29,3°C em 18,6h de incubação e a condição ótima de produção de substâncias antimicrobianas pH 6,22, a 30,6°C

6
no tempo de 17,9h. A produção de substâncias antimicrobianas seguiu uma cinética de metabolismo primário. A concentração inibitória mínima (CIM) do SLC foi de 6.400 UA.mL-1 e a concentração bactericida mínima (CBM) foi 1.600 UA.mL-1. O SLC apresentou ação antimicrobiana contra bactérias Gram-positivas e Gram-negativas, estabilidade ao tratamento térmico, a diferentes pH e agentes químicos, quando armazenado sob congelamento (- 18°C) e refrigeração (4°C) se manteve estável por 48 semanas e não foi tóxico nas concentrações de 10.000, 20.000 e 40.000 UA.mL-1 utilizando Drosophila melanogaster como modelo. Os filmes produzidos com CBM do SLC tiveram atividade antimicrobiana contra bactérias Gram-positivas e Gram-negativas, mantiveram sua atividade durante 30 dias e tiveram ação contra L. monocytogenes em queijo Prato. O isolado L. curvatus P99 apresentou características probióticas, tecnológicas e aspectos de segurança desejáveis para utilização como cultura adjunta de tecnologia. Os dados de otimização da multiplicação celular e produção de substâncias antimicrobianas fornece informações básicas para produção deste micro-organismo e seus metabólitos em escala industrial. O SLC pode ser utilizado como agente antimicrobiano para produção de filmes biodegradáveis de uso alimentar. PALAVRAS-CHAVE: bactérias ácido-láticas; antimicrobianos; sakacinas; superfície de resposta; filme biodegradável.

7
ABSTRACT
FUNCK, Graciele Daiana. Probiotics, technological and safety characteristics of Lactobacillus curvatus P99 and production, characterization and application of its antimicrobial substances. 2016. 146f. Tese (Doutorado) – Programa de Pós-Graduação em Ciência e Tecnologia de Alimentos. Universidade Federal de Pelotas, Pelotas, 2016.
The use of new starter cultures with industrial and functional potential have been explored. Functional cultures can be used to improve and optimize the food production process, as well as sensory characteristics, safety, and promote health benefits. The application of natural antimicrobial substances (such as bacteriocins), combined with differents technologies, promote new opportunities to pathogens control, improving the quality and safety of food. Biodegradable films incorporated with bacteriocins can increase the food protection and avoid post-processing contamination. Food contamination by pathogenic and spoilage bacteria is a problem for the food industry. Biopreservantes cultures and / or their metabolites have been identified and characterized as a alternative for food preservation. This study aimed to evaluate the probiotic, technological and safety characteristics of Lactobacillus curvatus P99 as well as verify the optimal conditions for growth and antimicrobial substances production (bacteriocin-like), characterize the stability at different temperatures, pH values and chemical agents of the cell free supernatant (CFS), quantify its antimicrobial activity, produce biodegradable films with CFS and apply this films in a food system. For this, phenotypic, genotypic and statistics tests were used. The isolate L. curvatus P99 presented probiotic, technological and safety characteristics as: tolerance to simulated gastrointestinal transit when protected for food, autoaggregation and coaggregation with Listeria monocytogenes, inability to adhere to intestinal cells HCT-116, antimicrobial activity against Gram-positive and Gram-negative micro-organisms, the absence of β-galactosidase activity, growth at 10 and 45°C, 4.5 and 6% NaCl, non-production of gas from glucose, casein hydrolysis, non-lipolytic activity, no antimicrobial resistance and amino acids decarboxylase genes nor β-hemolytic, DNAse and gelatinase activity. The optimal conditions for growth were pH 6.3, 29.3°C and 18.6h of incubation and the optimal conditions for antimicrobial substances production were pH 6.22, 30.6°C and 17.9h of incubation. The antimicrobial substances production was produced

8
following primary metabolite kinetics. The minimum inhibitory concentration (MIC) of CFS was 6400 AU.mL-1 while the minimum bactericidal concentration (MBC) was 1.600 AU.mL-1. The CFS showed antimicrobial activity against Gram-positive and Gram-negative microorganisms, stability at different temperatures, pH values and chemical agents, when stored at frozen (-18°C) and refrigerated (4°C) temperature, maintained its stability for 48 weeks and non-toxic at concentrations of 10,000, 20,000 and 40,000 AU.mL-1 in Drosophila melanogaster model. Biodegradable films containing CFS in the MBC showed antimicrobial activity against Gram-positive and Gram-negative microorganisms, maintained their activity for 30 days and inhibited L. monocytogenes in Prato cheese. The isolate L. curvatus P99 showed desirable probiotics, technological and safety characteristics for use as an adjunct culture. The optimization data provides basic information for the growth and antimicrobial substances production of L. curvatus P99 on an industrial scale. The CFS may be used as an antimicrobial agent in biodegradable films for use in food. KEYWORDS: lactic acid bacteria; antimicrobial; sakacins; response surface; biodegradable film.

9
LISTA DE FIGURAS
Figura 1 - Contagem das células viáveis do isolado Lactobacillus curvatus P99
quando submetido à presença de suco gástrico artificial (pepsina 3mg.mL-1 +
pH 2,5) na presença/ausência de alimento. ..................................................... 71
Figura 2 - Contagem das células viáveis do isolado Lactobacillus curvatus P99
quando submetido à presença de suco intestinal artificial (pancreatina 1mg.mL-
1 + pH 8,0) na presença/ausência de alimento e sais bilires (0,3%)................. 74
Figura 3 - Percentual de autoagregação (A) do Lactobacillus curvatus P99 e
Listeria monocytogenes Scott A (LM) nas temperaturas de 20 e 37°C em
função do tempo (2, 20 e 24h) e percentual de coagregação (B) de
Lactobacillus curvatus P99 e Listeria monocytogenes Scott A sob diferentes
temperaturas 20 e 37°C e tempos (2, 4 e 24h). ............................................... 77
Figura 4 - Amplificação do gene blaZ (172 pb) associado a resistência a
antimicrobianos β-lactâmicos. .......................................................................... 84
Figura 5 - Amplificação dos genes (A) sul1 (840 pb), (B) sul2 (293 pb) e (C)
dfrG (422 pb) associados a resistência a antimicrobianos Inibidores da Via
Folato. .............................................................................................................. 85
Figura 6 - Amplificação dos genes (A) ermB (424 pb) e (B) ermC (299 pb)
associados a resistência a antimicrobianos Macrolídeos. ................................ 85
Figura 7 - Amplificação dos genes (A) tetA (953 pb), (B) tetB (1.169 pb), (C)
tetK (360 pb), (D) tetL (1.028 pb), (E) tetM (158 pb), (F) tetO (781 pb) e (G)
Tn916-1545 (1.028 pb) associados a resistência a antimicrobianos
Tetraciclinas. .................................................................................................... 86
Figura 8 - Amplificação dos genes (A) strA (645 pb) e (B) strB (510 pb)
associados a resistência a antimicrobianos Aminoglicosídeos. ....................... 87

10
Figura 9 - Amplificação dos genes tyrdc (1.133 pb) e hdc (440 pb) associados a
produção de aminas biogênicas tirosina e histamina. ...................................... 89
Figura 10 – Perfil plasmidial do isolado Lactobacillus curvatus P99. ............... 90
Figura 11 - Superfície de resposta e gráficos de contorno para contagem de
células viáveis (log UFC.mL-1) de Lactobacillus curvatus P99. ........................ 94
Figura 12 - Superfície de resposta e gráficos de contorno para atividade
antimicrobiana (UA.mL-1) de Lactobacillus curvatus P99. ................................ 96
Figura 13 - Mudanças na contagem de células viáveis (log UFC.mL-1) e
atividade antimicrobiana (UA.mL-1) durante o crescimento do isolado
Lactobacillus curvatus P99 a 30ºC durante 24h. .............................................. 97
Figura 14 - Efeito de diferentes concentrações de sobrenadante livre de células
contendo substância antimicrobiana (bacteriocin-like) produzido por
Lactobacillus curvatus P99 sobre a mortalidade de Drosophila melanogaster
após 7 dias de tratamento. ............................................................................. 105
Figura 15 - Modo de ação dos filmes incorporados com a concentração
inibitória mínima (CIM) e a concentração bactericida mínima (CBM) do
sobrenadante livre de células contendo substâncias antimicrobianas
(bacteriocin-like) produzidas pelo isolado Lactobacillus curvatus P99. .......... 107
Figura 16 - Contagem de Listeria monocytogenes Scott A em queijo Prato
fatiado adicionado de filme biodegradável incorporado com sobrenadante livre
de células contendo substâncias antimicrobianas (bacteriocin-like) produzidas
por Lactobacillus curvatus P99 na concentração inibitória mínima (CIM) e
concentração bactericida mínima (CBM) durante dez dias de armazenamento a
4ºC. ................................................................................................................ 110
Figura 17 - Resistência e susceptibilidade a antimicrobianos de uso clínico por
Lactobacillus curvatus P99 utilizando o teste de disco difusão em ágar. ....... 144
Figura 18 - Quantificação da atividade antimicrobiana do sobrenadante livre de
células contendo substâncias antimicrobianas (bacteriocin- like) produzidas por
Lactobacillus curvatus P99. ............................................................................ 145
Figura 19 - Atividade antimicrobiana de filmes incorporados de sobrenadante
livre de células contendo substância antimicrobiana de origem proteica
(bacteriocin-like) contra Listeria monocytogenes Scott A. .............................. 146

11
LISTA DE TABELAS
Tabela 1 – Genes alvo e primers utilizados na identificação de bacteriocinas
produzidas por Lactobacillus curvatus ............................................................. 37
Tabela 2 - Características das principais sakacinas identificadas, produzidas
por linhagens de Lactobacillus curvatus e Lactobacillus sakei ......................... 41
Tabela 3 - Oligonucleotídeos para detecção dos genes de resistência de
Lactobacillus curvatus P99 a antimicrobianos de uso clínico ........................... 53
Tabela 4 - Contagem de células viáveis (log UFC.mL-1) de Lactobacillus
curvatus P99 quando submetido ao trânsito gástrico simulado ....................... 72
Tabela 5 - Contagem de células viáveis (log UFC.mL-1) de Lactobacillus
curvatus P99 quando submetido ao trânsito intestinal simulado ...................... 74
Tabela 6 - Percentual de autoagregação de Lactobacillus curvatus P99 e
Listeria monocytogenes (LM) em função da temperatura e tempo .................. 75
Tabela 7 - Percentual de coagregação entre Lactobaciilus curvatus P99 e
Listeria monocytogenes Scott A em função da temperatura e do tempo ......... 76
Tabela 8 - Espectro de inibição da atividade antimicrobiana produzida pelo
isolado Lactobacillus curvatus P99 .................................................................. 81
Tabela 9 – Análise de susceptibilidade fenotípica a antimicrobianos de uso
clínico de Lactobacillus curvatus P99 ............................................................... 84
Tabela 10 - Coeficientes de regressão (dados codificados), R2 e R2 adj. dos
modelos de superfície de resposta para contagem de células viáveis e
atividade antimicrobiana de Lactobacillus curvatus P99 .................................. 92
Tabela 11 - Resultados da análise de variância (ANOVA) para equação de
regressão para contagem de células viáveis e atividade antimicrobiana de
Lactobacillus curvatus P99 ............................................................................... 93

12
Tabela 12 - Linhagens indicadoras e sua sensibilidade para o sobrenadante
livre de células contendo substância antimicrobiana (bacteriocin-like) produzido
por Lactobacillus curvatus P99 ......................................................................... 99
Tabela 13 - Estabilidade a refrigeração e ao congelamento do sobrenadante
livre de células contendo substâncias antimicrobianas (bacteriocin-like)
produzido por Lactobacillus curvatus P99 (% de atividade em relação ao
controle sem tratamento) ............................................................................... 102

13
SUMÁRIO
1 Introdução ..................................................................................................... 18
2 Objetivo geral ................................................................................................ 22
2.1 Objetivos específicos ................................................................................. 22
3 Revisão de literatura ..................................................................................... 23
3.1 Bactérias ácido-láticas ............................................................................... 23
3.2 Gênero Lactobacillus spp. .......................................................................... 24
3.2.1 Lactobacillus curvatus ............................................................................. 25
3.3 Culturas probióticas .................................................................................... 26
3.4 Culturas biopreservantes............................................................................ 29
3.4.1 Bacteriocinas ........................................................................................... 30
3.4.1.1 Classificação ........................................................................................ 31
3.4.1.2 Organização genética ........................................................................... 34
3.4.1.3 Biossíntese ........................................................................................... 38
3.4.1.4 Sakacinas ............................................................................................. 40
3.5 Filmes antimicrobianos ............................................................................... 43
4 Material e métodos ........................................................................................ 45
4.1 Linhagem bacteriana .................................................................................. 45
4.2 Caracterização do isolado L. curvatus P99 ................................................ 45
4.2.1 Características probióticas ...................................................................... 45

14
4.2.1.1 Tolerância ao trânsito gastrointestinal de forma simulada (TGS) ......... 45
4.2.1.2 Capacidade de autoagregação, coagregação e hidrofobicidade .......... 46
4.2.1.3 Capacidade de adesão de L. monocytogenes ATCC 7644 e do isolado
L. curvatus P99 a células intestinais HCT-116 ................................................. 47
4.2.1.4 Atividade antimicrobiana ...................................................................... 48
4.2.1.5 Avaliação da atividade β-galactosidase ............................................... 49
4.2.2 Características tecnológicas .................................................................... 49
4.2.2.1 Multiplicação em diferentes temperaturas e concentrações de sal (NaCl)
......................................................................................................................... 49
4.2.2.2 Fermentação da glicose ....................................................................... 50
4.2.2.3 Atividade proteolítica e lipolítica ........................................................... 50
4.2.3 Aspectos de segurança ........................................................................... 51
4.2.3.1 Testes de susceptibilidade aos antimicrobianos de uso clínico ............ 51
4.2.3.2 Determinação dos genes para descarboxilação de aminoácidos ......... 56
4.2.3.3 Perfil plasmidial .................................................................................... 56
4. 2.3.4 Atividade da β-hemolisina, DNAse e gelatinase .................................. 57
4.3 Modelagem do crescimento e produção de substâncias antimicrobianas de
origem proteica (bacteriocin-like) pelo isolado L. curvatus P99 em resposta as
condições de pH, temperatura e tempo de incubação ..................................... 57
4.3.1 Inóculo e condições de crescimento e produção de substâncias
antimicrobianas de origem proteica (bacteriocin-like) ...................................... 57
4.3.2 Ensaios .................................................................................................... 58
4.4 Caracterização do sobrenadante livre de células (SLC) contendo
substâncias antimicrobianas de origem proteica (bacteriocin-like) ................... 58
4.4.1 Atividade antimicrobiana do SLC: concentração inibitória mínima (CIM) e
concentração bactericida mínima (CBM) ......................................................... 58
4.4.2 Atividade antimicrobiana ......................................................................... 59

15
4.4.3 Estabilidade ao tratamento térmico, refrigeração, congelamento, pH e
agentes químicos ............................................................................................. 59
4.4.4 Avaliação da toxicidade em modelo de Drosophila melanogaster .......... 61
4.5 Elaboração de filmes biodegradáveis incorporados de substância
antimicrobiana de origem proteica presente no SLC de L. curvatus P99 ........ 62
4.5.1 Avaliação da atividade antimicrobiana dos filmes biodegradáveis .......... 62
4.5.1.1 Quantificação e manutenção da atividade antimicrobiana ................... 62
4.5.1.2 Modo de ação ....................................................................................... 63
4.5.1.3 Atividade antimicrobiana ...................................................................... 64
4.6 Aplicação dos filmes biodegradáveis incorporados de SLC (substância
antimicrobiana de origem proteica) produzido pelo isolado L. curvatus P99 em
queijo Prato fatiado .......................................................................................... 64
4.7 Desenho experimental e análise estatística ............................................... 65
4.7.1 Otimização simultânea ............................................................................ 68
4.7.2 Correlação de Pearson............................................................................ 69
5 Resultados e discussão................................................................................. 70
5.1 Caracterização do isolado L. curvatus P99 ................................................ 70
5.1.1 Avaliação das características probióticas ................................................ 70
5.1.1.1 Tolerância ao trânsito gastrointestinal de forma simulada (TGS) ......... 70
5.1.1.2 Capacidade de autoagregação, coagregação e hidrofobicidade .......... 74
5.1.1.3 Ensaio de adesão celular ..................................................................... 79
5.1.1.4 Atividade antimicrobiana ...................................................................... 80
5.1.1.5 Avaliação da atividade da enzima β-galactosidase .............................. 81
5.2.2 Avaliação das características tecnológicas ............................................. 82
5.2.2.1 Multiplicação em diferentes temperaturas e concentrações de sal (NaCl)
......................................................................................................................... 82
5.2.2.2 Fermentação da glicose ....................................................................... 82

16
5.2.2.3 Atividade proteolítica e lipolítica ........................................................... 83
5.2.3 Aspectos de segurança ........................................................................... 83
5.2.3.1 Avaliação qualitativa e quantitativa da susceptibilidade aos
antimicrobianos de uso clínico ......................................................................... 83
5.2.3.2 Determinação dos genes para descarboxilação de aminoácidos ......... 88
5.2.3.3 Perfil plasmidial .................................................................................... 89
5.2.3.4 Atividade β-hemolisina, DNAse e gelatinase ........................................ 90
5.3 Modelagem do crescimento e produção de substâncias antimicrobianas de
origem proteica (bacteriocin-like) pelo isolado L. curvatus P99 em resposta ao
pH e temperatura ............................................................................................. 91
5.3.1 Multiplicação microbiana e produção de substâncias antimicrobianas de
origem proteica (bacteriocin-like) ..................................................................... 91
5.4 Caracterização do sobrenadante livre de células contendo substâncias
antimicrobianas de origem proteica (bacteriocin-like) ...................................... 98
5.4.1 Atividade antimicrobiana: Concentração inibitório mínima (CBM) e
concentração bactericida mínima (CBM) ......................................................... 98
5.4.1 Atividade antimicrobiana ......................................................................... 99
5.4.2 Estabilidade ao tratamento térmico, refrigeração, congelamento, pH e
agentes químicos ........................................................................................... 101
5.4.2.1 Estabilidade ao tratamento térmico .................................................... 101
5.4.2.4 Estabilidade aos agentes químicos .................................................... 103
5.4.3 Avaliação da toxicidade da substância antimicrobiana de origem proteica
em modelo de Drosophila melanogaster ........................................................ 104
5.5 Caracterização dos filmes biodegradáveis incorporados de SLC, contendo
substâncias antimicrobianas de origem proteica, produzido pelo isolado L.
curvatus P99 .................................................................................................. 106
5.5.1 Quantificação e manutenção da atividade antimicrobiana .................... 106
5.5.2 Modo de ação ........................................................................................ 106

17
5.5.3 Espectro de ação .................................................................................. 108
5.6 Aplicação dos filmes biodegradáveis incorporados de SLC, contendo
substâncias antimicrobianas de origem proteica, produzido pelo isolado L.
curvatus P99 em queijo Prato ........................................................................ 109
6 Conclusões .................................................................................................. 112
7 Perspectivas futuras .................................................................................... 114
Referências .................................................................................................... 115
Apêndices....................................................................................................... 143

18
1 Introdução
Bactérias ácido-láticas (BAL) têm um longo histórico de uso e até hoje
apresentam papel essencial nos processos de fermentação de alimentos. A
contribuição mais importante destes micro-organismos é extender a vida útil
dos alimentos, porém elas também têm influência sobre as características
nutricionais, sensoriais e padronização dos produtos finais (BAKA et al., 2011;
BARBOSA et al., 2015). Devido a apresentarem propriedades probióticas,
bioprotetoras e tecnológicas, as BAL têm sido utilizadas pela indústria de
alimentos como culturas adjuntas. Culturas adjuntas são aquelas que
promovem propriedades desejáveis aos alimentos, como benefícios à saúde,
segurança e/ou atributos sensoriais (GIRAFFA, 2003).
Os benefícios atribuídos as culturas de BAL probióticas incluem:
colonização do trato gastrointestinal, promovendo o balanço da microbiota
nativa nesse ambiente; redução dos níveis de colesterol; aumento das
imunoglobulinas A (IgA) que são produzidas pelos plasmócitos em resposta a
um imunógeno e funcionam como anticorpos; bloqueio da adesão de bactérias
patogênicas nos tecido epiteliais; tratamento para doenças inflamatórias do
intestino, como síndrome do colo irritado; redução de constipação e
desconforto intestinal; tratamento do câncer intestinal; controle de infecções
orais entre outros benefícios (VASILJEVIC; SHAH, 2008). Adicionalmente,
outros efeitos terapêuticos são atribuídos a bactérias probióticas como a
redução dos sintomas da intolerância a lactose devido à produção da enzima β-
D-galactosidase (GHEYTANCHI et al., 2009).
As BAL também são conhecidas como culturas biopreservantes e assim
são denominadas por apresentarem atividade antagonista contra micro-

19
organismos patogênicos e/ou deteriorantes importantes em alimentos. Esta
atividade antagonista ocorre por diversos mecanismos, principalmente pela
produção de substâncias inibitórias potencialmente letais aos micro-organismos
alvos. As principais substâncias com potencial antimicrobiano são ácidos
orgânicos, dióxido de carbono, álcoois, aldeídos, peróxido de hidrogênio e
bacteriocinas (HUGAS, 1998; OLIVEIRA; OLIVEIRA; GLÓRIA, 2008).
Diversas pesquisas vêm sendo realizadas sobre o potencial
antimicrobiano de metabólitos produzidos, em especial, por BAL, visando a
substituição dos conservadores sintéticos, pois alguns estudos têm
demonstrado riscos toxicológicos e carcinogênicos relacionados a ingestão
frequente e/ou de altas concentrações destes aditivos (GUILLARD et al., 2009;
MPOUNTOUKAS et al., 2008), o que vai ao encontro do crescente interesse da
indústria de alimentos por culturas biopreservantes (ANANOU et al., 2005a;
DAL BELLO et al., 2012; HUANG et al., 2013; CÁLIX-LARA, et al. 2014).
As bacteriocinas são peptídeos ou proteínas antimicrobianas
sintetizadas nos ribossomos das células bacterianas e liberadas no meio
extracelular, que apresentam ação bactericida ou bacteriostática sobre diversos
micro-organismos (JACK; TAGG; RAY, 1995). Além disso, são efetivas em
baixas concentrações e não promovem alteração nas propriedades sensoriais
do produto (RODGERS, 2002). Muitas BAL são capazes de produzir
bacteriocinas com atividade antagonista específica contra determinado grupo
de bactérias ou a diferentes espécies (HOLZAPFEL et al., 2001).
A aplicação de bacteriocinas como biopreservantes nos alimentos pode
seguir três orientações, adição de BAL no alimento e consequente produção
de bacteriocina, adição de um extrato bruto contendo bacteriocinas ou
aplicação da bacteriocina purificada no produto (HUGAS et al., 2003). A técnica
que utiliza culturas biopreservantes e/ou seus metabólitos para estender a vida
útil dos alimentos e aumentar a sua segurança é denominada biopreservação
(ROSS et al., 2002).
Lactobacillus curvatus é uma espécie de BAL comumente associada à
produtos cárneos fermentados e carnes refrigeradas embaladas à vácuo
(HEBERT et al., 2012). Algumas linhagens de L. curvatus apresentam
propriedades probióticas (AHN et al., 2015a; PARK, 2013; TODOROV et al.,

20
2011b) e outras são comercializadas como culturas iniciadoras e
biopreservantes para embutidos cárneos fermentados (MEAT AND
SAUSAGES, 2015). Alguns estudos relatam o isolamento L. curvatus e
avaliação de seu potencial bacteriocinogênico (AHMADOVA et al., 2013a;
BARBOSA et al., 2015), atribuído a diferentes bacteriocinas como as
curvacinas e as sakacinas, com importante ação antimicrobiana (MACWANA;
MURIANA, 2012a; RIVAS et al., 2014).
Em razão de suas propriedades bioquímicas, as curvacinas e sakacinas
estão agrupadas como peptídeos da classe II, as quais tem peso molecular
baixo (< 10 kDa), são termoestáveis e possuem atividade antilisterial
(KLAENHAMMER, 1988; SIP et al., 2012). As bacteriocinas desta classe
formam poros na membrana plasmática da célula alvo, promovendo a
dissipação da força próton motriz, aceleração no consumo do ATP e,
consequentemente, a morte celular do micro-organismo alvo (KAISER;
MONTVILLE, 1996).
Sabe-se que as condições de incubação como pH, temperatura e tempo
podem influenciar a produção de bacteriocinas a qual é induzida por um
mecanismo de quorum sensing, ou seja, a quantidade de bacteriocina formada
esta correlacionada com a biomassa produzida (LEROY; DE VUYST, 2002).
Mataragas et al. (2003), em seu estudo, demonstraram que o pH e a
temperatura, influenciaram a produção de bacteriocinas por L. curvatus L442.
Em alguns casos, bacteriocinas são recuperadas em condições não favoráveis
de nutrientes, pH e temperatura para a multiplicação das linhagens
(HWANHLEM et al., 2014; TODOROV; DICKS, 2006; TODOROV; VAN
REENEN; DICKS, 2004; TODOROV; DICKS, 2005).
A contaminação de alimentos por bactérias patogênicas e deteriorantes
é um problema para a indústria alimentícia. Apesar das diversas tecnologias
para a conservação de alimentos já disponíveis, ainda é um desafio para os
pesquisadores encontrar uma substância conservadora que apresente baixa
toxicidade e mantenha as características sensoriais desejáveis em alimentos
(ACUÑA; MORERO; BELLOMIO, 2011).
Para reduzir o problema da contaminação de alimentos prontos para o
consumo, bacteriocinas têm sido incluídas em polímeros para produção de

21
embalagens ativas (DEEGAN et al., 2006; MARCOS et al., 2010; MASSANI et
al., 2014a). Grande parte da matéria-prima utilizada para a fabricação de
embalagens é constituída por polímeros sintéticos, que provêm de fontes não
renováveis e que não são biodegradáveis. Os polímeros naturais têm sido
utilizados na produção de embalagens como uma alternativa para redução da
poluição ambiental causada pelo descarte de embalagens não biodegradáveis
no meio ambiente. Neste sentido, o amido tem sido estudado como material
constituinte de embalagens devido à sua biodegradabilidade, baixo custo e
disponibilidade em todo o mundo (HALAL et al., 2015; ZAVAREZE et al., 2012).
A utilização de bacteriocinas purificadas na matriz alimentícia pode
acarretar na redução da atividade antimicrobiana, perdendo sua eficiência, à
medida que possa sofrer degradação enzimática ou ainda interagir com outras
proteínas e lipídeos. A utilização do sobrenadante livre de células contendo
bacteriocinas (SLC) é uma alternativa, já que outros metabólitos
biologicamente ativos podem estar presentes e causar efeitos sinérgicos
(HARTMANN; WILKE; ERDMANN, 2011). A incorporação de SLC em filmes
biodegradáveis de amido pode contribuir significativamente para a indústria de
alimentos, já que além de auxiliar na separação das fatias, quando utilizado em
produtos prontos para consumo, contribuem eliminando possíveis
contaminações superficiais por patógenos.
Desta forma, no presente trabalho, objetivou-se avaliar as características
probióticas, tecnológicas e aspectos de segurança de Lactobacillus curvatus
P99 bem como, modelar a produção de suas substâncias antimicrobianas,
caracterizá-las e aplicá-las em filmes biodegradáveis.

22
2 Objetivo geral
Avaliar as características probióticas, tecnológicas e aspectos de
segurança do isolado Lactobacillus curvatus P99 bem como, produzir,
caracterizar e aplicar suas substâncias antimicrobianas de origem proteica em
filmes biodegradáveis.
2.1 Objetivos específicos
1. Avaliar as características probióticas, tecnológicas e aspectos de
segurança de L. curvatus P99;
2. Modelar o crescimento e a produção das substâncias
antimicrobianas de origem proteica (bacteriocin-like) por L. curvatus P99 em
resposta ao pH e temperatura;
3. Caracterizar o sobrenadante livre de células (SLC) contendo
substâncias de origem proteica (bacteriocin-like) produzido por L. curvatus P99;
4. Elaborar filmes biodegradáveis incorporados de SLC produzido por
L. curvatus P99;
5. Aplicar os filmes biodegradáveis incorporados de SLC produzido por
L. curvatus P99 em queijo Prato.

23
3 Revisão de literatura
3.1 Bactérias ácido-láticas
As bactérias ácido-láticas (BAL) constituem um grupo heterogêneo de
gêneros que compartilham características fisiológicas. Incluem os gêneros
Lactobacillus, Lactococcus, Carnobacterium, Enterococcus, Lactosphaera,
Leuconostoc, Melissococcus, Oenococcus, Pediococcus, Streptococcus,
Tetragenococcus, Vagococcus, Weissella, Microbacterium, Bifidobacterium e
Propionibacterium classificação baseada na comparação de análise de
sequência 16S rDNA (HOLZAPFEL et al., 2001). As bactérias pertencentes a
este grupo possuem capacidade de fermentar açúcares, principalmente em
ácido lático, através de metabolismo homofermentativo ou heterofermentativo.
Estas se caracterizam por serem Gram-positivas, catalase negativa, não
formadoras de esporos e anaeróbias facultativas (AXELSSON, 1998).
A heterogeneidade deste grupo de bactérias é expressa em suas
características morfológicas, uma vez que elas podem se apresentar na forma
de bacilos ou cocos, em células individuais ou em duplas, tétrades e em
cadeias curtas ou longas. Devido a capacidade limitada de biossíntese e suas
altas exigências em termos de carbono e fontes de nitrogênio, o habitat natural
das BAL são ambientes ricos nutricionalmente. Estão, geralmente, associados
com plantas e matérias-primas animais, produtos alimentícios fermentados,
pele de animais, mucosas e membranas (SETTANNI; MOSCHETTI, 2010).
As BAL estão amplamente distribuídas na natureza e podem ser
encontradas no solo, na água, no esterco, no esgoto e na silagem
(NASCIMENTO et al., 2008; NETO et al. 2005). Elas são também habitantes

24
dos tratos digestório, respiratório superior e urogenital inferior dos animais. A
maioria delas é inativada a temperatura superior a 70°C (NETO et al., 2005).
Muitas BAL apresentam potencial de inibição contra patógenos como
Staphylococcus spp., Listeria spp., Salmonella spp., Bacillus spp. e bactérias
do grupo coliforme e deteriorantes como Pseudomonas spp. (DE MARTINIS;
FREITAS, 2002; HERNÁNDEZ; CARDELL; ZÁRATE, 2005; HERREROS et al.,
2005; LIMA et al., 2009; MACEDO et al., 2004; NESPOLO; BRANDELLI, 2010;
NETO et al., 2005; MORAES et al., 2009; ORTIGOSA et al., 2006; ORTOLANI
et al., 2010; VELJOVIC et al., 2007).
A fermentação por BAL é uma forma de biopreservação utilizada há
muito tempo. A aplicação de um conjunto de técnicas e de processos que
empregam organismos vivos ou substâncias provindas destes é denominada
biopreservação. Esta permite melhorar a qualidade do produto, controlar os
processos e aumentar a vida útil e a segurança alimentar em todas as suas
vertentes (CORBO et al., 2009).
Essas bactérias têm relevância, tanto na indústria de alimentos quanto
em saúde pública, por apresentarem características transformadoras,
deteriorantes, biopreservantes e probióticas. São utilizadas como culturas
iniciadoras (starter), provocando transformações nas matérias-primas,
produzindo ácidos e outras substâncias que conferem sabores e aromas
específicos em produtos fermentados (LEROY; DE VUYST, 2004).
3.2 Gênero Lactobacillus spp.
O gênero Lactobacillus foi isolado e identificado em 1901, por Otto
Kandler e Norbert, sendo que hoje já são identificadas mais de 180 espécies
(http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=1578, 2016).
São bactérias Gram-positivas, de formas longas a delgadas, às vezes tendem
a formar hastes curtas, denominada cocobacilos, normalmente não apresentam
motilidade, são microaerófilos, não esporulam, tendo o produto final do
metabolismo de carbono o lactato, que normalmente não é fermentado e são
microaerofílicos (KANDLER; WEISS, 1986).

25
Estritamente fermentativos e exigentes em termos de uma nutrição
complexa a base de carboidratos, aminoácidos, peptídeos, derivados de ácidos
nucléicos e vitaminas. As condições físico-químicas influenciam e controlam a
população de micro-organismos presentes no intestino. A produção de
substâncias antimicrobianas pelos lactobacilos do intestino, contribuem para o
efeito de proteção contra o ataque de patógenos (HAMMES; HERTEL, 2006).
Este gênero é o mais diversificado do grupo BAL, amplamente
utilizado devido a sua importância para a saúde e aplicação em alimentos
(COUSIN et al., 2015). Lactobacillus spp. apresentam status GRAS (grau-
alimento) e tem longa história de uso como cultura iniciadora em alimentos
fermentados (AYENI et al., 2011). Porém, algumas espécies podem trazer
risco à saúde humana, devido a produção de metabólitos tóxicos, como as
aminas biogênicas (KIM; KIM, 2014; LADERO et al., 2015) ou transferir genes
de resistência a antimicrobianos de uso clínico para patógenos alimentares
(HUMMEL et al., 2007; LIU et al., 2009).
3.2.1 Lactobacillus curvatus
Lactobacillus curvatus são BAL, normalmente associadas a produtos
cárneos (HEBERT et al., 2012). Alguns pesquisadores têm isolado e
identificado L. curvatus a partir de embutidos fermentados tradicionais da Itália
(CASABURI et al., 2016) e salame tipo italiano (BARBOSA, M. DE S. et al.,
2015), no entanto, outros isolaram de produto fermentado à base de peixe
tradicional do Japão (COUSIN et al., 2015) e de queijo tradicional do Azerbaijão
(AHMADOVA et al., 2013b) .
Esta espécie é filogeneticamente relacionadas com Lactobacillus sakei,
Lactobacillus fuchuensis e Lactobacillus graminis, espécies também
associadas com produtos cárneos (BERTHIER; EHRLICH, 1999; ERCOLINI et
al., 2006; SAKALA et al., 2002). Lactobacillus sakei e L. curvatus possuem uma
similaridade fenotípica, diferindo-se apenas na hidrólise da arginina,
fermentação da melibiose, xilose e catalase heme-dependente, porém, muitas
vezes, linhagens atípicas têm sido encontradas (BERTHIER; EHRLICH, 1999).

26
Este micro-organismo é utilizado como cultura iniciadora em embutidos
fermentados (CASABURI et al., 2016) e como cultura biopreservativa, com o
intuito de inibir patógenos e/ou prolongar a vida útil em produtos cárneos
(FADDA; LÓPEZ; VIGNOLO, 2010). Embora a principal atividade
antimicrobiana de L. curvatus seja devido a produção de ácidos orgânicos, a
inibição pode estar associada a bacteriocinas. Linhagens de L. curvatus têm
sido descritas como produtoras de diferentes bacteriocinas relatadas como
ativas contra espécies relacionadas e patógenos (AHMADOVA et al., 2013a).
Isolados de L. curvatus provenientes de diversos alimentos foram
caracterizados como carreadores de genes para produção de curvacinas e
sakacinas (AHMADOVA et al., 2013a; CASABURI et al., 2016; COCOLIN;
RANTSIOU, 2007).
Por ser um micro-organismo bem adaptado a ambientes cárneos, L.
curvatus melhora o flavour e acelera os processos de fermentação de produtos
devido a habilidade de hidrólise de proteínas miofibrilares e sarcoplasmáticas
gerando pequenos peptídeos e aminoácidos (CASABURI et al., 2008; FADDA
et al., 1999).
Além de contribuir com propriedades sensoriais e promover segurança
aos alimentos, algumas linhagens de L. curvatus têm sido caracterizadas como
culturas probióticas. Todorov et al. (2011b) avaliaram L. curvatus isolados de
salmão defumado e obtiveram resultados positivos para as características
probióticas de autoagregação, coagregação, hidrofobicidade e adesão a
células Caco-2. Outros estudos, utilizando uma cepa probiótica de L. curvatus
HY7601, têm demonstrado sua capacidade de reduzir os níveis de
triglicerídeos do plasma em indivíduos diabéticos e não diabéticos (AHN et al.,
2015a, 2015b).
3.3 Culturas probióticas
Os probióticos consistem em micro-organismos vivos, os quais, quando
ingeridos em quantidades suficientes, conferem benefícios à saúde do
hospedeiro (FAO/WHO, 2006). Algumas linhagens de Lactobacillus são boas
candidatas a culturas probióticas por fazerem parte da microbiota intestinal e

27
exibirem propriedades promotoras de saúde in vivo (LEROY; VERLUYTEN; DE
VUYST, 2006).
Os benefícios atribuídos a culturas probióticas incluem: colonização do
trato gastrointestinal, promovendo o balanço da microbiota nativa nesse
ambiente; redução dos níveis de colesterol; aumento das imunoglobulinas A
(IgA) que são produzidas pelos plasmócitos em resposta a um imunógeno e
funcionam como anticorpos; bloqueio da adesão de bactérias patogênicas nos
tecido epiteliais; tratamento para doenças inflamatórias do intestino, como
síndrome do cólon irritado; redução de constipação e desconforto intestinal;
tratamento do câncer intestinal; controle de infecções bucais entre outros
benefícios (VASILJEVIC; SHAH, 2008).
Adicionalmente, outros efeitos terapêuticos são atribuídos a bactérias
probióticas como a redução dos sintomas da intolerância à lactose devido à
produção de β-D-galactosidase. Esta enzima é responsável pela hidrólise da
lactose em glicose e galactose, resultando em baixos níveis de lactose em
leites fermentados, consequentemente baixos níveis de lactose no intestino
humano (GHEYTANCHI et al., 2009).
O modo de ação dos probióticos não foi ainda completamente
esclarecido, embora tenham sido sugeridos vários processos que podem atuar
independentemente ou associados. Diferentes probióticos têm sido associados
a diferentes efeitos, relacionados com suas capacidades específicas de
expressar moléculas de superfície ou secretar proteínas e metabólitos que
interagem diretamente com as células dos hospedeiros (NAGPAL et al., 2012).
A exclusão competitiva tem sido relatada como um dos processos
relacionados ao modo de ação, uma vez que o probiótico competiria com os
patógenos por sítios de fixação e nutrientes, impedindo sua ação
transitoriamente. A exclusão competitiva explicaria a necessidade da
administração continuada e a elevadas doses dos probióticos, para manifestar
seus efeitos. Os probióticos podem também afetar patógenos através da
síntese de bacteriocinas, de ácidos orgânicos voláteis, de peróxido de
hidrogênio e pela liberação enzimas como a lactase. Seus efeitos
anticarcinogênicos podem ser atribuídos à inibição de enzimas pro-

28
carcinogênicas ou a estimulação do sistema imunitário do hospedeiro (KUMAR;
VIJAYENDRA; REDDY, 2015).
Para um micro-organismo ser considerado probiótico ele deve: ser viável
durante todo tempo de estocagem e transporte do produto; sobreviver à
presença de ácido gástrico e a bile; não apresentar genes de resistência a
antibióticos; apresentar genes de adesão à mucosa; manter-se viável na
presença de fármacos comerciais não antibióticos; não apresentar genes que
codificam fatores de virulência; apresentar níveis de autoagregação,
coagregação e hidrofobicidade (COLLADO; MERILUOTO; SALMINEN, 2008;
RANADHEERA; BAINES; ADAMS, 2010).
Também é sugerido que haja uma ingestão diária de 108 a 109 UFC de
micro-organismos probióticos para que seus benefícios à saúde sejam
conferidos. Baseado no consumo diário de 100 g ou mL de alimentos
probióticos, sugere-se que este contenha no mínimo 106 a 107 células por
grama ou mL do alimento (BRASIL, 2002; FAO/WHO, 2006). O conteúdo de
gordura, proteínas, açúcares e pH dos produtos são alguns dos fatores que
afetam a multiplicação e sobrevivência dos probióticos (NAGPAL et al., 2012).
Os alimentos probióticos mais conhecidos são os derivados lácteos,
como, por exemplo, iogurtes, queijos, sorvetes e sobremesas lácteas.
Limitações de produtos lácteos como a presença de compostos alergênicos e a
necessidade de estocagem refrigerada, bem como o aumento da demanda por
novos alimentos têm iniciado a pesquisa e o desenvolvimento de outros
produtos probióticos que não lácteos (KUMAR; VIJAYENDRA; REDDY, 2015).
Bebidas fermentadas e a base de frutas, cereais, alimentos a base de vegetais
e produtos cárneos têm sido testados como veículo de micro-organismos
probióticos (GONZÁLEZ-VÁZQUEZ et al., 2015; KHAN; KANG, 2016;
NAGPAL; KUMAR; KUMAR, 2012; OJHA et al., 2015; KUMAR; VIJAYENDRA;
REDDY, 2015).
A regulamentação sobre o uso de probióticos em alimentos tem sido
estabelecida em nível internacional. As normas estão relacionadas a eficácia,
aspectos de segurança, rotulagem, fraudes e alegações. A Food and Drug
Administration – FDA, dos Estados Unidos, define quatro categorias de
alimentos: alimentos convencionais, dietas especiais, medicinais e

29
suplementos dietéticos. De um ponto de vista da legislação, os alimentos
contendo probióticos poderiam ser incluídos em qualquer uma das quatro
categorias descritas pelo FDA, porém não há um reconhecimento explícito dos
benefícios para a saúde de probióticos nos Estados Unidos (NAGPAL et al.,
2012).
As legislações, em matéria de avaliação de segurança dos probióticos e
como componentes dos alimentos, diferem entre os países e não está
estabelecida em uma base internacional. Na maior parte destas, os probióticos
são regulamentados sob alimentos e suplementos dietéticos. Os fatores que
devem ser abordados na avaliação da segurança dos probióticos incluem
patogenicidade, infecciosidade e virulência que compreendem toxicidade,
atividade metabólica e as propriedades intrínsecas dos micro-organismos
(NAGPAL et al., 2012).
No Brasil, a Agência Nacional de Vigilância Sanitária – ANVISA, através
da RDC nº 2 de 07 de janeiro de 2002, padroniza os procedimentos a serem
adotados para a avaliação de segurança, registro e comercialização de
substâncias bioativas e probióticos isolados com alegação de propriedades
funcional e ou de saúde. São permitidos pela legislação brasileira para uso
como culturas probióticas, as espécies de L. acidophilus, L. casei shirota, L.
casei variedade rhamnosus, L. casei variedade defensis, L. paracasei, L. lactis,
B. bifidum, B. animallis, B. longum e Enterococcus faecium (BRASIL, 2002).
3.4 Culturas biopreservantes
O principal efeito antimicrobiano de BAL, responsáveis pela
biopreservação, é a capacidade de acidificação. Porém, outras substâncias
inibitórias potencialmente letais aos micro-organismos tem papel fundamental,
não só na segurança dos alimentos, como também na prevenção do
crescimento de BAL autóctones que podem produzir flavour indesejável
(LEROY; VERLUYTEN; DE VUYST, 2006). As principais substâncias com
potencial antimicrobiano são os ácidos orgânicos, dióxido de carbono, álcoois,
aldeídos, peróxido de hidrogênio e bacteriocinas (HUGAS, 1998; OLIVEIRA;
OLIVEIRA; GLÓRIA, 2008).

30
Culturas produtoras de bacteriocinas podem ser utilizadas para inibir o
crescimento de micro-organismos deteriorantes e patogênicos durante a vida
útil de alimentos não fermentados pela produção de bacteriocinas in situ. Este
tipo de aplicação ocorre devido à necessidade de uma legislação específica
para utilização de bacteriocinas purificadas, sendo que no Brasil, apenas a
nisina tem autorização de uso (SUSKOVIC et al., 2010). A nisina é produzida
por Lactococcus lactis subsp. lactis e seu uso é permitido como conservante
natural de origem biológica em queijos pasteurizados e superfície de salsichas
no limite máximo de 12,5 mg.kg-1 (BRASIL, 1996; MORI, 2013).
O Joint Expert Committee on Food Additives da FAO/WHO indica os
benefícios do uso da nisina e outras bacteriocinas, como antimicrobiano. Ainda,
recomenda uma ingestão diária de 60 mg de nisina pura para uma pessoa de
70kg. O FDA determina um limite máximo de 500 mg.kg-1 de nisina para
aplicação em produtos cárneos e lácteos nos Estados Unidos (JONES et al.,
2005).
3.4.1 Bacteriocinas
A literatura científica e a comunidade acadêmica tem adaptado a
definição de Klaenhammer (1988) para bacteriocinas: “são um grupo
heterogêneo de peptídeos ou proteínas bioativos sintetizados
ribossomalmente, secretados extracelularmente, com efeito antimicrobiano”.
Historicamente, em 1976, Tagg; Dajani e Wannamaker (1976) definiram
bacteriocinas como compostos proteicos sintetizados por bactérias Gram-
positivas e Gram-negativas que exibiam atividade inibitória contra espécies
relacionadas com a linhagem produtora de bacteriocina. Informações sobre
bacteriocinas foram primeiramente publicadas em 1925 quando pesquisadores
encontraram uma substância biologicamente ativa produzida por Escherichia
coli V, apresentando atividade antagonista contra outra linhagem da mesma
espécie (E. coli F). Mais tarde, substâncias antimicrobianas similares
produzidas por E. coli foram encontradas e denominadas colicinas
(BESHKOVA; FRENGOVA, 2012).

31
Os primeiros relatos de bacteriocinas produzidas por BAL são de 1928
(ROGERS, 1928). Esta substância, mais tarde, foi caracterizada
bioquimicamente, definida como um polipeptídeo e denominada “nisina”
(WHITEHEAD, 1933; MATTICK; HIRSCH, 1947). Depois disso, diversos
pesquisadores vêm estudando diferentes bacteriocinas produzidas por BAL
(JACK; TAGG; RAY, 1995; MASUDA et al., 2011; HARTMANN et al., 2011;
HÉCHARD et al., 1992; KAUR et al., 2013).
As bacteriocinas produzidas por BAL oferecem muitas propriedades
desejáveis que as tornam adequadas para a utilização na conservação de
alimentos. As bacteriocinas são conhecidas como substâncias seguras, não
são ativas ou citotóxicas em células eucarióticas, são inativadas por enzimas
digestivas (proteases), com pouca influência sobre a microbiota intestinal, são
normalmente tolerantes ao pH baixo e ao aquecimento, tem amplo espectro
antimicrobiano contra muitas bactérias patogênicas e deteriorantes,
apresentam modo de ação bactericida e/ou bacteriostático, normalmente seu
alvo é a membrana citoplasmática, não apresentam resistência a antibióticos e
seus determinantes genéticos são geralmente codificados em plasmídeos,
facilitando a manipulação genética (GÁLVEZ et al., 2007).
3.4.1.1 Classificação
As bacteriocinas produzidas por BAL são divididas em três classes
maiores, conforme sua estrutura, tamanho, características físico-químicas e
moleculares. No entanto, esta classificação é continuamente revisada devido
ao acúmulo de conhecimento e a caracterização de novas bacteriocinas.
Klaenhammer (1993) definiu quatro classes de bacteriocinas produzidas
por BAL. A classe I, conhecida como lantibióticos (< 5 kDa); a classe II,
composta por pequenos peptídeos (4 – 6 kDa) termoestáveis e subdividida em
três subclasses, IIa, IIb e IIc; a classe III, composta de grandes proteínas
termolábeis (> 30 kDa); e a classe IV, são proteínas complexas, compostas de
peptídeos e uma ou mais partes químicas (lipídeo, carboidrato) necessárias
para a atividade antimicrobiana.

32
Nes et al. (1996) reagruparam a classe II das bacteriocinas, mantendo
as subclasses I e II e alterando a subclasse IIc para incluir bacteriocinas que
contém um peptídeo sinal típico e que são secretados via translocase geral
(sec) da célula. Estes autores também excluíram a classe IV das bacteriocinas
visto que elas não tinham sido bem caracterizadas. No sistema proposto por
Van Belkum e Stiles (2000), a classe II foi subdividida, baseada no número de
resíduos de cisteína, usando a nomenclatura proposta por Jack; Tagg e Ray
(1995). Na sua classificação, as bacteriocinas que são secretadas via
translocase (sec), tipo IIc proposto por Nes et al. (1996), não são classificadas
em uma subclasse separada porque há uma diversidade de bacteriocinas que
são secretadas por esta via (FRANZ et al., 2007). A classificação mais aceita
atualmente, está descrita abaixo.
3.4.1.1.1 Classe I ou lantibióticos
É constituída por peptídeos de baixo peso molecular (< 5 kDa), entre 19
a 38 resíduos de aminoácidos, termoestáveis, catiônicos, hidrofóbicos,
diferenciados dos demais pela presença de lantionina e derivados. São
divididas em Tipo A (moléculas alongadas e carregadas positivamente) -
subtipo A1 e A2 - e Tipo B (moléculas globulares e não carregadas). Os
exemplos do Tipo A são a nisina A e Z, subtilina, epidermina, mutacina,
lacticina 481 e 3147, lactocina S, entre outras. Os exemplos do Tipo B são
cinamicina, duramicina B e C, ancovenina, entre outras (MCAULIFFE; ROSS;
HILL, 2001; BESHKOVA; FRENGOVA, 2012).
A nisina é a bacteriocina mais estudada e mais bem documentada pela
pesquisa científica. Ela é produzida por linhagens de L. lactis subsp. lactis e é
composta por 34 resíduos de aminoácidos. Duas ocorrências naturalmente
variantes da nisina são a nisina A e a nisina Z, que diferem estruturalmente em
apenas um resíduo de aminoácido, porém, com atividade semelhante
(MULDERS et al., 1991). Apresenta amplo espectro antimicrobiano com ação
sobre L. monocytogenes, S. aureus, B. cereus, além de outros patógenos e
espécies de BAL. É adicionada como aditivo alimentar e foi aceita por Joint

33
Expert Commitee da FAO/WHO em 1969 e usada até hoje em mais de 50
países (JONES et al., 2005).
3.4.1.1.2 Classe II
É composta por pequenos peptídeos (< 10 kDa), são termoestáveis,
catiônicos, hidrofóbicos, geralmente estes apresentam uma estrutura helicoidal
anfifílica, a qual permite sua inserção na membrana citoplasmática da célula
alvo, promovendo a despolarização da membrana e a morte celular. São
propostas 3 subdivisões para esta classe (DRIDER et al., 2006):
- Classe IIa: composta por bacteriocinas que apresentam especificidade
contra L. monocytogenes. Seus representantes possuem 37 a 48 resíduos de
aminoácidos, com porção N-terminal com configuração de folha pregueada e
uma porção C-terminal contendo duas hélices (FIMLAND et al., 2005). As
bacteriocinas desta classe se inserem na membrana celular do micro-
organismo alvo pela porção C-terminal, promovendo a formação de poros e
consequente dissipação da força próton motriz. Na tentativa de manter ou
restaurar a força próton motriz, ocorre uma aceleração no consumo de ATP e,
consequentemente, a morte celular (KAISER; MONTVILLE, 1996). Pediocina
AcH, enterocina A, sakacina A e P, leucocina A, mesentericina Y105, entre
outras, são exemplos de bacteriocinas desta subclasse.
- Classe IIb: esta classe é composta por bacteriocinas heterodiméricas
que requerem a atividade combinada de dois peptídeos. Seu mecanismo de
ação também envolve a dissipação do potencial de membrana e diminuição da
concentração intracelular de ATP (COTTER; HILL; ROSS, 2005). São
exemplos de bacteriocinas desta subclasse a plantaricina EF, JK e S, a
enterocina 1071, a lactococina G e MN, entre outras.
- Classe IIc: as bacteriocinas pertencentes a esta classe apresentam
uma união covalente das terminações C e N, resultando em uma estrutura
cíclica (COTTER; HILL; ROSS, 2005). São representantes desta classe a
enterocina AS-48, a circularina A e a reutericina 6.

34
3.4.1.1.3 Classe III
Esta classe é composta por grandes proteínas (> 10 kDa), termolábeis,
complexas quanto à atividade e à estrutura proteica. São subdivididas em duas
subclasses: IIIa ou bacteriolisinas e IIIb. A subclasse IIIa tem seu mecanismo
de ação diferenciado das demais bacteriocinas, promovendo a lise celular
através da degradação da parede celular do micro-organismo alvo. A melhor
bacteriolisina estudada é a lisostafina, um peptídeo de peso molecular de 27
kDa, responsável pela hidrólise da parede celular de muitas espécies de
Staphylococcus, principalmente S. aureus (BASTOS; COUTINHO, COELHO,
2010). A subclasse IIIb, em contraste, compreende os peptídeos que não
causam lise, mas sim dissipação do potencial de membrana e diminuição da
concentração intracelular de ATP. A helveticina J é exemplo desta subclasse.
As bacteriocinas apresentam uma porção N-terminal homóloga a uma
endopeptidase, responsável pela catálise da parede celular, e uma porção C-
terminal responsável pelo reconhecimento da célula alvo (LAI; TRAN,
SIMMONDS, 2002).
3.4.1.2 Organização genética
Os genes associados com à expressão de bacteriocinas são
organizados em operons encontrados em diferentes elementos do genoma. Os
genes relacionados a bacteriocinas são arranjados em genes clusters (DRIDER
et al., 2006). Uma vez que muitos genes de bacteriocinas estão sob a
regulação de vários promotores, sua estrutura tecnicamente não corresponde à
descrição de um operon, embora o termo seja usado frequentemente. Genes
clusters são complexos, com genes divergentemente transcritos, codificados
em cadeias complementares de DNA e genes auxiliares codificados em
diferentes locus do gene estrutural (KOTELNIKOVA; GELFAND, 2002).
Sabe-se, porém que, a presença do operon da bacteriocina não
necessariamente indica sua expressão. Operons parcialmente ativos ou
inativos têm sido identificados (HÜHNE et al., 1996). Operons funcionais têm
sido identificados no cromossomo, como é o caso da sakacina P (URSO et al.,

35
2006), em plasmídeos, como no caso da pediocina PA-1 (PAPAGIANNI;
ANASTASIADOU, 2009) e em transposons, como a nisina (JACK; TAGG; RAY,
1995).
Os genes clusters são comumente associados a elementos transferíveis,
como plasmídeos ou transposons o que permite sua ampla distribuição na
natureza (MCAULIFFE; ROSS; HILL, 2001). Entretanto, existem exemplos de
bacteriocinas codificadas cromossomalmente, alguns destes resultam da
introdução de elementos transferíveis no cromossomo, como exemplo o
transposon contendo o gene estrutural da nisina A que é encontrado no
cromossomo de L. lactis (ENTIAN; DE VOS, 1996). Em todos os casos, o
operon das bacteriocinas contém um gene estrutural, codificando para suas
bacteriocinas ou um pré-peptídeo para modificação adicional e genes
auxiliares, codificando para peptídeos de imunidade, enzimas modificadoras,
maquinário de secreção e fator indutor. A composição exata do operon é
dependente da estrutura, regulação e expressão da bacteriocina, mas têm sido
comuns entre grupos distintos produtores de bacteriocinas. Porém, há
exceções para este esquema em termos de regulação e organização genética,
como por exemplo, as carnobacteriocinas A, B1 e B2, as quais apresentam três
genes estruturais codificando para elas, localizados em dois plasmídeos
separados em uma linhagem de Carnobacterium (JACK; SAHL, 1995).
O gene que codifica para a produção do precursor da curvacina A
(curA), uma bacteriocina produzida por Lactobacillus curvatus, foi localizado em
um fragmento de 1,2kb AceI-EcoRI de um plasmídeo de 60kb e verificou-se
preceder ainda, um gene adicional, ORF2. Embora a função da proteína ORF2
de 51 aminoácidos não tenha sido definida ainda, esta foi predita como tendo
um papel na imunidade celular. Um promotor foi identificado a upstrem do gene
cura, juntamente com um sítio de ligação do ribossomo. Entretanto, nenhum
sítio de ligação do ribosso foi identificado para ORF2. Nenhum outro gene que
possa estar relacionado ao processamento, maturação e translocação do
peptídeo estava presente na porção sequenciada (TICHACZEK; VOGEL;
HAMMES, 1993).
Em outro estudo, o gene estrutural da sakacina A (sakA) foi identificado
eum um fragmento de 1,4kb EcoRI presente em um plasmídeo de 60kb de

36
Lactobacillus sakei LB 706. A sequência de nucleotídeo da sakA foi
demonstrada como idêntica a curA. O sequenciamento de um fragmento
separado de 1,8kb HindIII-BglI proveniente do mesmo plasmídeo revelou a
presença de outro gene, sakB, com um promotor e um sítio de ligação próprio,
mas de orientação oposta da sakA e separada por 1,6kb. O gene sakB de 430
aminoácidos pode estar relacionado com a imunidade da sakacina A
(AXELSSON; HOLCK, 1995; SCHILLINGER; LÜCKE, 1989).
Na tabela 1 podemos visualizar os genes alvo e primers utilizados para
identificação das principais bacteriocinas produzidas por L. curvatus.

37
Tabela 1 – Genes alvo e primers utilizados na identificação de bacteriocinas produzidas por Lactobacillus curvatus
Bacteriocina Genes alvo Sequências de Primers (5’ – 3’) Tamanho Amplicon
(pb) Condições da PCR Referência
Sakacina Tα sakTα
FW: TCGGTGGCTATACTGTCTAAACA 160 95ºC por 15 min; 95ºC por 30 s; 58ºC por 1 min; 72ºC por 1 min
Macwana; Muriana (2012a)
RV: TGTCCTAAAAATCCACCAATGC
Sakacina T sakT
FW: AAGAAATGATAGAAATTTTTGGAGG 151 95ºC por 15 min; 95ºC por 30 s; 56ºC por 1 min; 72ºC por 1 min
Macwana; Muriana (2012a)
RV: TGTGAAATCCAATCTTGTCCTG
Sakacina P sakP
FW: ATG GAA AAG TTT ATT GAA TTA 186 94ºC por 3 min; 94ºC por 30 s; 40ºC por 1 min; 72ºC por 1 min
Remiger; Ehrmann; Vogel (1996)
RV: TTA T TT ATT CCA GCC AGC GTT
Sakacina G1 sakgA1
FW: TTA GAA CTA CAC TGA TCG TG 94ºC por 4 min; 94ºC por 30 s; 38ºC por 30 s; 72ºC por 30 s
Todorov et al. (2011a)
RV: TGG AAG AAT GAG TAC TTG TT
Sakacina G2 sakgA2
FW: CGT TAC AAC AGA ACT TCA AG 94ºC por 4 min; 94ºC por 30 s; 38ºC por 30 s; 72ºC por 30 s
Todorov et al. (2011a)
RV: CGT TAC AAC AGA ACT TCA AG
Curvacina A curA
FW: GTA AAA GAA TTA AGT ATG ACA 171 94ºC por 3 min; 94ºC por 30 s; 40ºC por 1 min; 72ºC por 1 min
Reminger; Ehrmann; Vogel (1996)
RV: TTA CAT TCC AGC TAA ACC ACT
Sakacina A sakA
FW: GAAT⁄ATA⁄GC⁄AA⁄CANCAATTAC⁄TA⁄CGGTGG 150 95ºC por 15 min; 95ºC por, 30 s; 55ºC por 1 min; 72ºC por 1 min
Dortu et al. (2008)
RV: CAGCCGCTAATCATACCACC
FW: Forward RV: Reverse

38
3.4.1.3 Biossíntese
As bacteriocinas são produzidas por um processo dependente da
multiplicação e da atividade biológica da bactéria (LEROY; DE VUYST, 2002).
Em geral, a produção de bacteriocinas por BAL ocorrem durante a fase
exponencial, sendo que as condições favoráveis para a multiplicação da
bactéria e altas densidades celulares, beneficiam a produção de bacteriocinas
(YANG; RAY, 1994). Entretanto, nem sempre esta condição prevalece
(HWANHLEM et al., 2013).
A expressão de bacteriocinas é um mecanismo de defesa de diversas
bactérias, a qual é regulada por um mecanismo de quorum sensing, isto é,
expressão ocorre em resposta às flutuações de densidade celular. As bactérias
produzem e liberam sinais químicos, denominados auto-indutores, que
aumentam à medida que aumenta a densidade celular. A detecção de um
limiar mínimo estimula a liberação dos auto-indutores. À medida que a
densidade celular aumenta, a necessidade de produção de metabólitos de
defesa diminui (MILLER; BASSLER, 2001).
Quanto à biossíntese, observa-se que, enquanto muitos antibióticos
(considerados metabólitos secundários) são sintetizados durante a fase
estacionária de crescimento, quase todas as bacteriocinas produzidas pelas
BAL manifestam cinética de metabólito primário, iniciando a produção na fase
logarítmica de crescimento. Por serem substâncias de natureza protéica, são
sintetizadas pelo mesmo mecanismo ribossomal de síntese de proteínas,
envolvendo transcrição e tradução. A síntese das bacteriocinas está
determinada pela estrutura genética de quatro genes diferentes, os quais
codificam funções básicas requeridas para a produção da atividade
antimicrobiana extracelular. Estes quatro genes são: o gene estrutural que
codifica para a pré-bacteriocina, o gene da imunidade específica, o gene que
codifica para o sistema de transporte ABC e o gene da proteína acessória
necessária para a externalização da bacteriocina (NES et al., 1996).
As bacteriocinas são sintetizadas, primeiramente, na forma de pré-
peptídios ou pré-bacteriocinas biologicamente inativos. Esses pré-peptídios
contêm uma sequência de 18 a 27 aminoácidos, apresentando 2 glicinas na

39
região N-terminal. As funções dessa sequência de aminoácidos são a de evitar
que a bacteriocina seja biologicamente ativada dentro da célula produtora e a
de servir como sinal de reconhecimento para o sistema de transporte que
envolve as proteínas do transporte ABC e uma proteína acessório (NES et al.,
1996). As duas glicinas presentes na sequência de aminoácidos são as
responsáveis pelo reconhecimento da pré-bacteriocina no sistema de
transporte (MOLL et al., 1996). Após o reconhecimento do pré-peptídio, a
sequência de aminoácidos é removida, e a bacteriocina excretada da célula
(ENNAHAR et al., 2000).
As condições ambientais estão intimamente relacionadas com o
crescimento celular e a produção de bacteriocinas. Schiru et al. (2014) afirmam
que cada linhagem tem sua condição ótima de crescimento e produção de
bacteriocina, sendo afetada fortemente pelas condições físico-químicas do
ambiente, como meio de cultura, pH, temperatura e tempo de incubação. Por
exemplo, o pH e a temperatura de crescimento tem mostrado efeito na
produção de pediocina AcH (BISWAS et al., 1991) e pediocina PD-1 (NEL et
al., 2001) produzida por Pediococcus spp., bem como na bacteriocina
produzida por E. faecium RZS C5 (LEROY; DE VUYST, 2002) e nas
bacteriocinas produzidas por E. faecium e E. faecalis isoladas de produtos
lácteos fermentados do Paquistão (JAVED et al., 2011).
Outros estudos demonstraram a influência de diferentes meios de
cultura na produção de bacteriocinas em diversas linhagens de BAL. Todorov;
van Reenen e Dicks (2004) observaram que a produção de bacteriocina
ST341LD produzida por L. plantarum foi estimulada pela presença de triptona.
Já, a adição de 10 e 40 g.L-1 de glicose ou de 5 ou 10 g.L-1 de K2HPO4, reduziu
em 50% a produção de bacteriocina e que a presença de glicerol reprimiu a
produção. De Vuyst e Vandamme (1992) estudaram a influência de fontes de
carbono sobre a produção de nisina produzida por linhagens de L. lactis e
observaram que estas parecem ser um importante mecanismo de controle para
a produção desta bacteriocina. Já para Yi et al. (2013), a bacteriocina Lac-B23
produzida por L. paracasei J23 estudada quanto aos efeitos do tempo de
incubação, temperatura e fatores exógenos para sua produção, observaram
que cisteína e glicina estimularam a produção da mesma. O glicerol e o ácido

40
pirúvico promoveram o aumento da produção da bacteriocina e as
concentrações ótimas foram de 1% e 30 g.L-1, respectivamente.
3.4.1.4 Sakacinas
Sakacinas são bacteriocinas da classe II, produzidas por cepas de L.
sakei, com atividade antimicrobiana contra L. monocytogenes. Porém, além de
L. sakei, cepas de L. curvatus também têm demonstrado a presença de genes
que codificam para a produção de sakacinas. Cocolin e Rantsiou (2007)
descreveram pela primeira vez a presença de genes estruturais que codificam
para produção de sakacinas P e Q em linhagens de L. curvatus. Hequet et al.
(2007) observaram em seu estudo que o isolado L. curvatus 2711 produziu
sakacina X e ainda, apresentou os genes estruturais para sakacina T e P.
Diversas sakacinas têm sido identificadas e caracterizadas como podemos
observar na tabela 2.

41
Tabela 2 - Características das principais sakacinas identificadas, produzidas por linhagens de Lactobacillus curvatus e Lactobacillus sakei
Bacteriocina Classe Tamanho (a.a.) Linhagem produtora Origem Referências
Sakacina P IIa, Grupo I, sub-
grupo I-1
43 L. curvatus LTH1174
L. curvatus L442
L. sakei I151
L. sakei LTH673
L. curvatus CRL 705
Carne fermentada
Embutido fermentado Grego
Embutido
Carne fermentada
Embutido fermentado
Tichaczek et al. (1992)
Cocolin et al. (2007)
Urso et al. (2006)
Tichaczek; Vogel e Hammes (1994)
Castellano et al. (2010)
Sakacina X IIa, Grupo I, sub-
grupo I-1
43 L. sakei 5
L. curvatus CRL 705
Malte
Embutido fermentado
Vaughan et al. (2001)
Castellano et al. (2010)
Sakacina G IIa, Grupo III, Sub-
grupo III-1
37 L. sakei 2512
L. sakei R1333
L. sakei CWBI-B1365
Coleção de alimentos Rhodia
Salmão defumado
Carne de aves
Simon et al. (2002)
Todorov et al. (2011)
Dortu et al. (2008)
Sakacina A IIa, Grupo IV 41 L. sakei Lb706 Carne Holck et al. (1992)
Sakacina K IIa, Grupo IV 40 L. sakei CTC494 Embutido fermentado Aymerich et al. (2000)
Sakacina T IIb 41 L. sakei 5 Malte Vaughan et al. (2001)
Sakacina Q IIc 49 L. sakei Lb674
L. curvatus ACU-1
Carne de cordeiro
Embutidos artesanais
Mathiesen et al. (2005)
Rivas et al. (2014)

42
As bacteriocinas da classe IIa são sintetizadas ribossonalmente como
peptídeos precursores com uma sequência N-terminal líder. A sequência líder
de muitas bacteriocinas contém dois resíduos de glicina, os quais servem como
um sinal de reconhecimento para o processamento e secreção da proteína
(DRIDER et al., 2006). Entretanto, algumas bacteriocinas da classe IIa são
secretadas por um sistema geral sec-dependente. Estas bacteriocinas tem uma
sequência líder N-terminal hidrofóbica sec-dependente as quais dirigem a
proteína secretória para a membrana citoplasmática, sendo processada por
uma peptidase signal através da membrana citoplasmática durante a
translocação (HERRANZ; DRIESSEN, 2005; KALMOKOFF et al., 2001).
Muitas vezes um locus pode incluir a produção de mais de uma
bacteriocina. Esta habilidade pode ser vantajosa para as espécies produtoras,
reforçando sua capacidade de competir com outras bactérias em um mesmo
ambiente (VAUGHAN et al., 2001). L. sakei 5 produz sakacina P codificada por
plasmídeo, bem como duas bacteriocinas codificadas cromossomalmente, a
sakacina T, a qual é uma bacteriocina duplo-peptídeo pertencente a classe IIb
e a sakacina X, pertencente a classe IIa. O locus TX sakacina codifica os
genes estruturais da sakacina T e X, incluindo dois genes clusters adjacentes
mas divergentes. O primeiro gene cluster stxPRKT codifica um peptídeo indutor
e três proteínas envolvendo a regulação e secreção destas bacteriocinas. O
segundo gene cluster inclui sakTα, sakTβ, sakIT, sakX e sakIX os quais
codificam para os genes estruturais e imunidade para as sakacinas T e X
(VAUGHAN.; EIJSINK; VAN SINDEREN, 2003).
As sakacinas T e Tβ são parte de um complexo duplo-peptídeo e a
presença de ambas é requerida para a atividade do peptídeo maduro.
Carregando apenas um dos genes para o duplo-peptídeo, a solicitação primária
é da sakacina Tβ no ciclo de vida das linhagens, porém este peptídeo não é
antimicrobiano ou de defesa, tendo provavelmente um papel diferente no
complexo (MACWANA; MURIANA, 2012a).

43
3.5 Filmes antimicrobianos
Filmes antimicrobianos têm sido introduzido como um novo conceito de
embalagens para uso em alimentos, em resposta a demanda dos
consumidores. Os filmes antimicrobianos promovem, além de uma barreira
entre o produto e o ambiente externo, a redução, a inibição ou retardam a
multiplicação de micro-organismos nos alimentos acondicionados, com
consequente aumento da vida útil e obtenção de produtos seguros (OJAGH et
al., 2010).
Para que uma embalagem antimicrobiana exerça seu efeito, ela deve
estar em contato com o alimento, o que restringe o número de compostos
utilizados em sua produção. Existem dois tipos embalagens antimicrobianas, as
que promovem a migração do agente antimicrobiano para a superfície do
alimento e aquelas que promovem seu efeito sem a necessidade de migração
para o alimento (VERMEIREN; DEVLIEGHERE; DEBEVERE, 2002).
Os polímeros naturais têm sido utilizados na produção de embalagens
como uma alternativa para redução da poluição ambiental causada pelo
descarte de embalagens não biodegradáveis no meio ambiente. Neste sentido,
o amido tem sido estudado como material constituinte de embalagens devido à
sua biodegradabilidade, baixo custo e disponibilidade em várias partes do
mundo (HALAL et al., 2015; ZAVAREZE et al., 2012).
Culturas bacteriocinogênicas ou bacteriocinas têm sido incluídas em
polímeros para produção de embalagens ativas visando reduzir a
contaminação de alimentos prontos para o consumo. Iseppi et al. (2011)
elaboraram filmes a base de polivinil álcool incorporando uma cultura
bacteriocinogênica, E. casseliflavus IM 416K1, ou sua bacteriocina, enterocina
416K1 e observaram a ação destes filmes contra L. monocytogenes.
Marcos et al. (2007) produziram filmes biodegradáveis utilizando
alginato, zeína e polivinil álcool adicionados de enterocinas. Os autores
testaram a ação antimicrobiana destes filmes durante a estocagem de presunto
cozido fatiado e concluíram que as embalagens antimicrobianas podem
melhorar a segurança deste alimento, retardando ou reduzindo a multiplicação
de L. monocytogenes.

44
Os filmes antimicrobianos a base de glúten, elaborados por Massani et
al. (2014b), utilizando bacteriocinas produzidas pelo L. curvatus CRL705
evitaram a multiplicação de L. innocua e L. plantarum. Estes filmes foram
avaliados, também, quanto a manutenção da atividade antimicrobiana durante
a sua estocagem e foi observado que mantiveram a atividade por 50 dias. Para
cumprir o objetivo de proteger e expandir a vida útil dos alimentos, as
embalagens ativas devem manter suas propriedades de atividade ao longo de
um período de tempo definido de acordo com o prazo de validade dos
alimentos que embalado.

45
4 Material e métodos
4.1 Linhagem bacteriana
Foi objeto deste estudo um isolado de BAL bacteriocinogênico,
denominado L. curvatus P99 proveniente de presunto cozido fatiado. A
confirmação do potencial bacteriocinogênico se deu através da sensibilidade da
substância produzida frente a tripsina e a presença e expressão dos genes que
codificam para a produção de, sakacina X, T- e T-β. O isolado P99 pertence
a coleção de culturas bacterianas do Laboratório de Microbiologia de Alimentos
- Departamento de Ciência e Tecnologia Agroindustrial da Universidade
Federal de Pelotas (UFPel) e se encontrava armazenado a -80°C em caldo
MRS adicionado de 20% de glicerol.
4.2 Caracterização do isolado L. curvatus P99
4.2.1 Características probióticas
4.2.1.1 Tolerância ao trânsito gastrointestinal de forma simulada (TGS)
A avaliação da TGS foi realizada conforme Huang e Adams (2004).
Para isto, o meio (caldo de Man, Rogosa and Sharpe – MRS) contendo a
cultura cultivada a 37°C por 24h foi centrifugado a 6.800 x g, por 10 minutos, a
4ºC e o pellet resultante, após descarte do sobrenadante, foi lavado duas
vezes com solução salina a 0,5% e ressuspendida em 1 mL da mesma
solução. Uma alíquota de 0,2mL da suspensão celular foi misturada a 0,3mL de

46
solução salina e 1,0mL de suco gástrico ou intestinal artificial e incubada a
37ºC. A contagem de células viáveis à passagem ao trato gástrico foi realizada
nos tempos inicial e em 15, 30, 60, 120, 180 e 240 minutos enquanto que a
contagem para a passagem ao trato intestinal foi realizada no tempo inicial e
240 minutos em placas contendo ágar MRS após incubação a 37°C por 72h.
O suco gástrico artificial consistiu em pepsina (Sigma-Aldrich®) na
concentração de 3mg.mL-1 e pH 2,5 enquanto o suco intestinal artificial foi
composto por 1mg.mL-1 de pancreatina (Sigma-Aldrich®) e pH 8,0, adicionados
ou não de 0,5% de bile bovina (Sigma-Aldrich®).
A influência da presença de um alimento na sobrevivência durante o
trânsito gastrointestinal foi avaliada da mesma forma, porém, substituindo a
solução salina por 0,3mL de leite em pó integral ou desnatado (Elegê)
reconstituído em água destilada a 10% (m/v) e esterilizado a 121°C.
4.2.1.2 Capacidade de autoagregação, coagregação e hidrofobicidade
As propriedades in vitro de autoagregação e coagregação foram
realizadas conforme descrito por Collado; Meriluoto e Salminen (2008) e a
hidrofobicidade foi realizada como descrito por e Vinderola e Reinhemer
(2003). Suspensões celulares das culturas crescidas em caldo MRS a 37°C por
24h, a uma absorbância de 0,25 ± 0,02 a 600nm, foram preparadas para os
testes.
A autoagregação foi determinada a 20 e 37°C nos tempos de 2, 20 e
24h. A absorbância foi medida a 600nm e os resultados foram expressos em
percentual, conforme [1 – (A600 nm da suspensão inicial/A600 nm da suspensão
final) x 100]. Para determinar a capacidade de coagregação do isolado ao
patógeno L. monocytogenes Scott A, volumes iguais das suspensões celulares
bacterianas foram misturados e incubados a 37ºC. A absorbância foi
monitorada em 2, 4 e 24h. Os resultados foram expressos em percentual,
conforme [(Apat + Aisol)/2 – (Amix )/( Apat + Aisol)/2] x 100, onde Apat e Aisol
representam a absorbância das suspensões bacterianas em separado em
tubos controle e Amix representa a absorbância das suspensões celulares
misturadas nos diferentes tempos (inicial e final).

47
Para avaliar a adesão bacteriana ao hidrocarboneto, reagente octano
(Synth) foi empregado. A suspensão celular (3mL) foi agitada em vórtex por 60
segundos com 400µL de octano. Depois de 2h a 37°C, a fase aquosa foi
cuidadosamente removida e a absorbância a 600nm foi medida. A
hidrofobicidade foi determinada como o percentual de adesão, conforme [(A0-
A)/A] x 100, onde A0 e A são as absorbâncias antes e depois da extração com
octano, respectivamente.
4.2.1.3 Capacidade de adesão de L. monocytogenes ATCC 7644 e do
isolado L. curvatus P99 a células intestinais HCT-116
4.2.1.3.1 Ensaio de adesão a células HCT-116
O ensaio de adesão foi realizado segundo Garriga et al. (2015), com
modificações descritas por Sehn (2015). As células utilizadas para este ensaio
são linhagens de câncer de colorretal HCT-116 obtidas da coleção de células
do Laboratório de Imunologia Aplicada do Centro de Desenvolvimento
Tecnológico (CDTEc), da Universidade Federal de Pelotas.
Primeiramente, as células foram cultivadas em Dulbecco’s Modified
Eagle Medium - DMEM (Sigma-Aldrich®) suplementado com 10 % de soro fetal
bovino - SFM (Sigma-Aldrich®) (DMEM + SFB) e mantidas a 37 ºC em
atmosfera úmida com 5% de CO2. A seguir, as placas para cultivo celular foram
semeadas com 106 células por poço. Após a semeadura, as placas foram
incubadas nas mesmas condições descritas acima, até atingirem confluência
de 90%.
Separadamente, foram preparadas suspensões celulares de cultivos de
18h a 37ºC de L. curvatus P99 e de L. monocytogenes ATCC 7644. Para isto,
os cultivos foram centrifugados a 6.000 x g por 3min, os pellets lavados três
vezes com tampão fosfato-salino (PBS) e ressuspendidos em 1 mL do mesmo
(109 UFC.mL-1). Diluições seriadas dos inóculos foram preparadas e semeadas
em placas de ágar tripticase soy (TSA) para quantificação de L.
monocytogenes ATCC 7644 e de ágar MRS para o isolado P99. As placas
semeadas foram incubadas a 37ºC por 24h a 48h.

48
Concomitantemente, alíquotas de 100µL de cada uma das suspensões
foram adicionadas a 900µL de DMEM (concentração final de 107 UFC.mL-1). A
seguir, 100µL da suspensão celular em DMEM foram adicionados aos poços,
em triplicata. A placa de cultivo foi incubada por 1h a 37ºC. Transcorrido este
período, o meio de cultura foi removido e as cavidades foram lavadas três
vezes com meio DMEM para remoção das células não aderidas. A seguir,
100µL de PBS contendo 0,1% (v/v) de Triton X-100 (Synth) foram adicionados
aos poços e mantidos por 5 minutos a 37ºC. Então, os 100µL foram removidos
e a quantificação das células foi realizada, utilizando-se os meios e condições
de incubação descritos acima.
Os resultados foram expressos em porcentagem de adesão às células
intestinais HCT-116, conforme [(nº células do micro-organismo aderido x 100)/
nº total de células do micro-organismo adicionado por poço].
4.2.1.4 Atividade antimicrobiana
A atividade antimicrobiana produzida pelo isolado foi verificada a partir
da técnica spot-on-the-lawn, segundo Fleming et al. (1975) com adaptações
descritas por Marques (2014), contra as seguintes culturas de referência:
Bacillus cereus ATCC 11778, L. innocua CLIP 12612, L. monocytogenes Scott
A, L. monocytogenes ATCC 7644, L. monocytogenes Siliken, S. aureus ATCC
25923, S. hycus ATCC 11249, S. intermedius ATCC 29663, E. coli O157:H7
ATCC 43895, E. coli ATCC 8739, Pseudomonas aeruginosa ATCC 15442,
Salmonella Tiphymurium ATCC 14028 e Shigella dysenteriae ATCC 13313.
Inicialmente, foram retirados 2 μL da cultura do isolado P99 crescido
em caldo MRS a 37°C por 24h e inoculados sobre ágar MRS em placas,
seguido de incubação em anaerobiose a 37ºC por 24h. A seguir, o ágar das
placas foi recoberto com uma sobrecamada de ágar Brin Heart Infusion - BHI
(Himedia) semi-sólido contendo aproximadamente 105 UFC.mL-1 das culturas
indicadoras (concentração determinada através da escala de McFarland),
incubadas a 37ºC por 24h. A formação de halos inibitórios maiores ou iguais a
2mm ao redor da colônia do isolado indicou atividade antagonista. O diâmetro
dos halos foi medido com o auxílio de paquímetro (King.tools®).

49
4.2.1.5 Avaliação da atividade β-galactosidase
A atividade da enzima β-galactosidase foi determinada de acordo com
Vinderola e Reinheimer (2003), com modificações descritas por Sehn (2015). O
isolado foi cultivado em caldo MRS suplementado de 1% de lactose (Synth) a
37°C por 24h. Uma suspensão celular a absorbância de 1,0 ± 0,02 a 560nm foi
preparada a partir do cultivo. Em seguida, 1mL da suspensão celular foi
permeabilizada com 50µL de solução de tolueno:acetona (Synth) (1:9 v/v),
através de agitação por 7min. Após, 100µL da suspensão permeabilizada foi
adicionado à 900µL de tampão fosfato e 200µL de o-nitrophenyl-β-D-
galactopyranoside – ONPG na concentração de 4 mg.mL-1 (Sigma-Aldrich®). A
mistura foi mantida em banho-maria a 37ºC por 15 minutos e a reação foi
paralisada pela adição de 500µL de Na2CO3 (Vetec) a 1mol.L-1. Os valores de
absorbância foram medidos a 420 e 560 nm e os resultados expressos como
unidades Miller, conforme 1000 x [(A420 – 1,75x A2 560)/(15min x 1mL x A1
560)], onde A1 representa a absorbância imediatamente antes e A2 a
densidade das células da reação.
4.2.2 Características tecnológicas
4.2.2.1 Multiplicação em diferentes temperaturas e concentrações de sal
(NaCl)
Uma alíquota de 100µL do cultivo do isolado P99 em caldo MRS a
37°C por 24h foi utilizada para testar a multiplicação a diferentes temperaturas
(10 e 45°C por 7 dias). Para testar a multiplicação na presença de sal, utilizou-
se caldo MRS modificado com 4,5 e 6% de NaCl (Synth) e incubação a 37ºC
por 7 dias. A habilidade de se multiplicar nestas condições foi verificada através
da turvação do meio de cultura e presença de massa celular depositada no
fundo do tubo (DROSINOS et al., 2005).

50
4.2.2.2 Fermentação da glicose
A capacidade de fermentar glicose com produção de gás foi
determinada em tubos de ensaio contendo tubos de Durhan e caldo MRS
suplementado com 3% de glicose (Synth). A incubação foi realizada a 37ºC por
48h (LIMA et al., 2009). As culturas de referência, L. fermentum ATCC 9338 e
L. plantarum ATCC 8014 foram usadas como controle positivo e negativo,
respectivamente.
4.2.2.3 Atividade proteolítica e lipolítica
A produção de proteases foi avaliada pelo teste da hidrólise da caseína
(COLLINS et al., 2004). Primeiramente, um meio base consistindo de 0,75g de
extrato de carne (Oxoid), 1,25g de triptona (Difco), 0,25g de glicose (Synth) e
4,25g de ágar-ágar (Acumedia) em 190mL de água destilada foi preparado. A
seguir, o pH foi ajustado para 6,5 e procedeu-se a autoclavagem por 15
minutos a 121ºC. Separadamente, 6g de leite em pó desnatado (Elegê) foi
adicionado a 60mL de água destilada, o pH da mistura foi ajustado para 6,5 e
procedeu-se a autoclavagem por 15 minutos a 121°C. Por fim, a suspensão de
leite foi misturada com o meio base, seguida de plaqueamento em placas de
petri. Separadamente foi preparada uma suspensão celular, a partir da cultura
do isolado P99 crescida em caldo MRS a 37°C por 48h, por centrifugação a
13.000 g por 5 minutos, descarte do sobrenadante, sendo as células
posteriormente ressuspendidas em 1 mL de tampão PBS. Por fim, 10 μL da
suspensão celular foi adicionada sobre o ágar e procedeu-se a incubação a
30°C por 24h. O teste foi realizado em triplicata. A visualização de zona
transparente ao redor de uma colônia indicou a produção de proteases.
A atividade proteolítica foi determinada utilizando albumina de soro
bovino (BSA) de acordo com Mauriello et al. (2004), com adaptações descritas
por Cruxen (2016). Primeiramente, foi preparado um meio base contendo 0,5g
de triptona de soja (Himedia), 0,25g de extrato de levedura (Himedia), 0,1g de
glicose e 1,5g de ágar-ágar (Vetec, Brasil) em 100mL de água destilada. A

51
seguir, procedeu-se a autoclavagem a 121°C por 15 minutos. Após o meio
base ser resfriado a 60°C, adicionou-se 0,57 mg.mL-1 de proteína BSA (Sigma-
Aldrich®) seguido de plaqueamento em placas de petri. Então, 10μL da
suspensão celular, preparada como descrito anteriormente para o teste da
hidrólise da caseína, foi adicionada as placas e estas foram incubadas a 37°C
por 48h. Após este período, o ágar foi removido e corado por 5 minutos com
solução contendo 0,05% de Brilhante Blue R (Sigma-Aldrich®) em
metanol:ácido acético:água na proporção de 50:40:10 e descorado em
metanol:ácido acético:água na proporção de 25:5:70. Os reagentes metanol e
ácido acético são provenientes da Vetec. O teste foi realizado em triplicata.
Zonas claras ao redor da colônia indicaram atividade proteolítica.
Atividade lipolítica foi testada conforme Kouker e Jaeger (1987).
Inicialmente, foi preparado um meio base contendo 0,8g de caldo nutriente
(Oxoid®), 0,4g de NaCl e 1g de ágar-ágar em 100mL de água destilada. Este
meio foi autoclavado e resfriado a 60ºC. Em seguida, adicionou-se 2,5mL de
azeite de oliva extra virgem e homogeneizou-se em ultraturrax a 5.000 rpm por
3 minutos. Após, adicionou-se 1mL de Rodamina R (1 mg.mL-1) (Synth) e
procedeu-se o plaqueamento. Por fim, foram adicionados 10μL da suspensão
celular, preparada como descrito acima para o teste da hidrólise da caseína. As
placas foram incubadas a 37°C por 48h. O teste foi realizado em triplicata. O
aparecimento de halos fluorescentes sob luz UV a 350 nm indicou atividade
lipolítica. Para as análises atividade proteolítica e lipolítica, utilizou-se como
controle positivo, S. xylosus ATCC 29971.
4.2.3 Aspectos de segurança
4.2.3.1 Testes de susceptibilidade aos antimicrobianos de uso clínico
A avaliação fenotípica da susceptibilidade aos antimicrobianos foi
realizada pelo método de difusão em ágar, utilizando-se o ágar Müller-Hinton –
MH (Merck), conforme descrito por European Food Safety Authority (EFSA,
2003). Os antibióticos (Laborclin, Brasil) testados foram amicacina (30μg),
ampicilina (10μg), ciprofloxacina (5μg), clindamicina (2μg), cloranfenicol (30μg),

52
eritromicina (15μg), estreptomicina (10μg), gentamicina (10μg), penicilina G
(10μg), sulfametoxazol-trimetoprim (25μg), sulfonamida (300μg), vancomicina
(30μg) e tetraciclina (30μg). Os diâmetros das zonas de inibição foram medidos
e os resultados foram expressos como isolados resistentes (≤ 15mm), com
sensibilidade intermediária (16 – 20mm) ou sensíveis (≥ 21mm), conforme
descrito por Liase et al. (2009). A avaliação genotípica foi realizada para os
genes de resistência aos antimicrobianos de uso clínico através da técnica da
Reação em Cadeia da Polimerase (PCR) de acordo com os autores dos
primers e condições descritas na tabela 3. A reação foi preparada contendo
12,5 μL de GoTaq® Green MasterMix 2X (Promega Corp.), 1 μL de cada
primer, 2 μL de DNA (5ng/µL) e 8,5 μL de água ultrapura para completar o
volume final de 25 μL.
Os produtos da PCR foram submetidos a eletroforese a 80V por 70
minutos em gel de agarose 1,5% em tampão TBE (solução de Tris, ácido
bórico e EDTA) 0,5X, utilizando-se o marcador de peso molecular de 1Kb
(Invitrogen®). Os produtos amplificados foram corados com GelRedTM e
visualizados sob luz UV em transiluminador (Loccus®, L-Pix Touch).
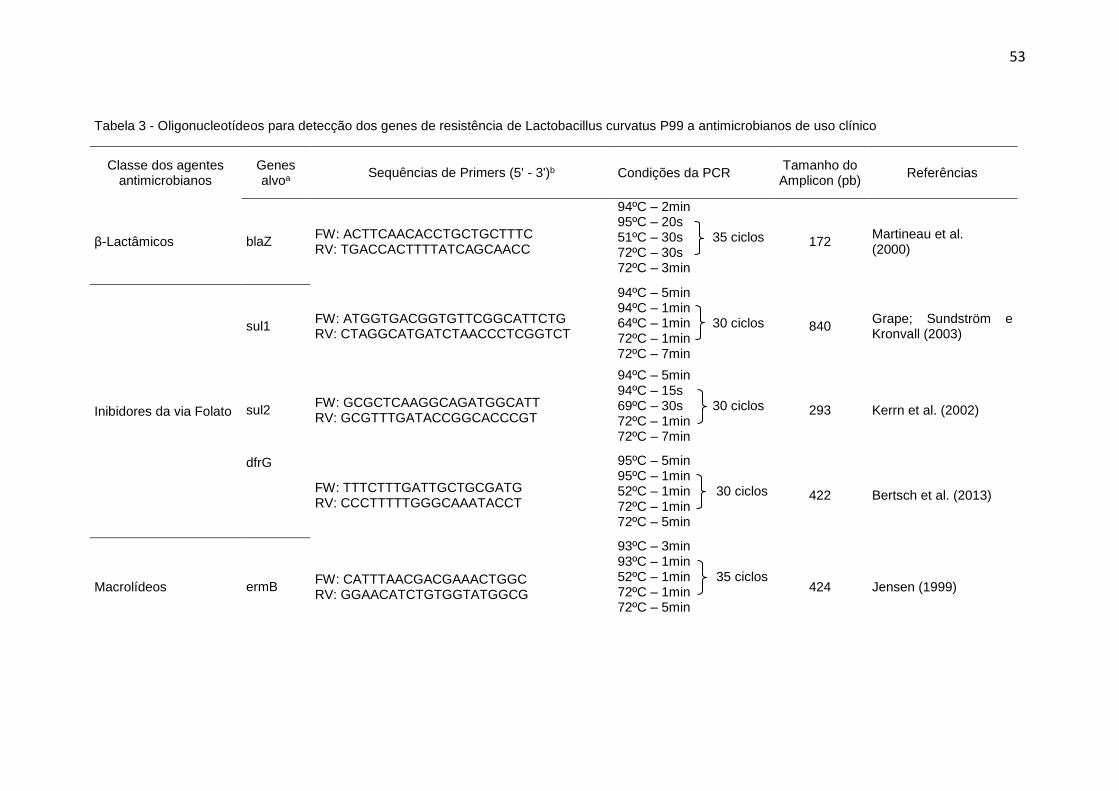
53
Tabela 3 - Oligonucleotídeos para detecção dos genes de resistência de Lactobacillus curvatus P99 a antimicrobianos de uso clínico
Classe dos agentes antimicrobianos
Genes alvoa
Sequências de Primers (5' - 3')b Condições da PCR Tamanho do
Amplicon (pb) Referências
β-Lactâmicos blaZ FW: ACTTCAACACCTGCTGCTTTC RV: TGACCACTTTTATCAGCAACC
94ºC – 2min 95ºC – 20s 51ºC – 30s 35 ciclos 72ºC – 30s 72ºC – 3min
172 Martineau et al. (2000)
Inibidores da via Folato
sul1 FW: ATGGTGACGGTGTTCGGCATTCTG RV: CTAGGCATGATCTAACCCTCGGTCT
94ºC – 5min 94ºC – 1min 64ºC – 1min 30 ciclos 72ºC – 1min 72ºC – 7min
840 Grape; Sundström e Kronvall (2003)
sul2 FW: GCGCTCAAGGCAGATGGCATT RV: GCGTTTGATACCGGCACCCGT
94ºC – 5min 94ºC – 15s 69ºC – 30s 30 ciclos 72ºC – 1min 72ºC – 7min
293 Kerrn et al. (2002)
dfrG
FW: TTTCTTTGATTGCTGCGATG RV: CCCTTTTTGGGCAAATACCT
95ºC – 5min 95ºC – 1min 52ºC – 1min 30 ciclos 72ºC – 1min 72ºC – 5min
422 Bertsch et al. (2013)
Macrolídeos ermB FW: CATTTAACGACGAAACTGGC RV: GGAACATCTGTGGTATGGCG
93ºC – 3min 93ºC – 1min 52ºC – 1min 35 ciclos 72ºC – 1min 72ºC – 5min
424 Jensen (1999)1999)

54
ermC FW: AATCGTCAATTCCTGCATGT RV: TAATCGTGGAATACGGGTTTG
94ºC – 3min 94ºC – 30s 55ºC – 30s 30 ciclos 72ºC – 30s 72ºC – 4min
299 Strommenger et al. (2003)
Tetraciclinas
tetA FW: GTAATTCTGAGCACTGT RV: CCTGGACAACATTGCTT
94ºC – 4min 94ºC – 1min 43ºC – 2min 34 ciclos 72ºC – 3min 72ºC – 7min
953 Frech e Schwarz (2000)
tetB FW: ACGTTACTCGATGCCAT RV: AGCACTTGTCTCCTGTT
94ºC – 4min 94ºC – 1min 50ºC – 2min 34 ciclos 72ºC – 3min 72ºC – 7min
1169 Frech e Schwarz (2000)
tetK FW: GTAGCGACAATAGGTAATAGT RV: GTAGTGACAATAAACCTCCTA
94ºC – 3min 94ºC – 30s 55ºC – 30s 30 ciclos 72ºC – 30s 72ºC – 4min
360 Strommenger et al. (2003)
tetL FW: CAAACTGGGTGAACACTG RV: CCTGTTCCCTCTGATAAA
95ºC – 3min 95ºC – 20s 45ºC – 30s 35 ciclos 72ºC – 30s 72ºC – 2min
1028 Pang; Bosch e Roberts (1994)
tetM FW: AGTGGAGCGATTACAGAA RV: CATATGTCCTGGCGTGTCTA
94ºC – 3min 94ºC – 30s 55ºC – 30s 30 ciclos 72ºC – 30s 72ºC – 4min
158 Strommenger et al. (2003)

55
TetO FW: AATGAAGATTCCGACAATTT RV: CTCATGCGTTGTAGTATTCCA
95ºC – 5min 94ºC – 1min 55ºC – 1min 30 ciclos 72ºC – 2min 72ºC – 10min
781 Li; Sherwood e Logue (2007)
Tn916-1545
FW: GCGTGATTGTATCTCACT RV: GACGCTCCTGTTGCTTCT
95ºC – 5min 94ºC – 1min 50ºC – 1min 30 ciclos 72ºC – 2min 72ºC – 10min
1028 Li; Sherwood e Logue (2007)
Aminoglicosídeos
strA FW: TGACTGGTTGCCTGTCAGAGG RV: CCAGTTGTCTTCGGCGTTAGCA
94ºC – 2min 94ºC – 1min 64ºC – 1min 30 ciclos 72ºC – 1min 72ºC – 7min
645 Kehrenberg e Schwarz (2001)
strB FW: ATCGTCAAGGGATTGAAACC RV: GGATCGTAGAACATATTGGC
94ºC – 2min 94ºC – 1min 56ºC – 1min 30 ciclos 72ºC – 1min 72ºC – 7min
510 Kikuvi et al. (2007)
a Controle positivo: Campylobacter jejuni 119 (tetO); Staphylococcus aureus S2 (tetK, tetL); Staphylococcus aureus S10 (blaZ, dfrG, ermC, tetM, Tn916-1545); Staphylococcus aureus 218 (ermB); Salmonella Typhimurium 22C (sul1, tetA, strA); Salmonella Typhimurium 140B1 (strB); Salmonella Typhimurium 171B2 (sul2, tetB). b FW: Forward; RV: Reverse

56
4.2.3.2 Determinação dos genes para descarboxilação de aminoácidos
A determinação dos genes histidina descarboxilase (hdc) e tirosina
descarboxilase (tyrdc) foi realizado por PCR multiplex, conforme Coton e Coton
(2005). Para identificação do gene hdc foram utilizados os primers forward (5’-
ATGTCAGAGTTTGATAAAAAG-3’) e reverse (5’-
TTAATAATTGATGTTTCCACC-3’). Para identificação do gene tyrdc foram
utilizados os primers forward (5’-ACATAGTCAACCATRTTGAA-3’) e reverse
(5’-CAAATGGAAGAAGAAGTAGG-3’). Utilizou-se como controle interno da
reação os primers universais 16S r-RNA como referência. A concentração do
DNA utilizado foi de 20 ng/μL, concentração de 2 pmol para o primer 16S rDNA
(controle interno da reação) e 20 pmol para os primer Hdc e Tyrdc. A reação foi
preparada contendo 12,5 μL de GoTaq® Green Master Mix 2X (Promega
Corp.), 1,0 μL de cada primer (forward e reverse), 2 μL do DNA e 8,5 μL de
água para completar o volume final de 25 μL. As condições do programa de
amplificação foram: 95°C por 5 minutos, 32 ciclos de 95°C por 45 segundos,
52°C por 45 segundos, 72°C por 1 minuto e 15 segundos e extensão final de
72 °C por 5 minutos. Os produtos da PCR foram submetidos a eletroforese a
100V por 50 minutos em gel de agarose 1,5% em tampão TBE (solução de
Tris, ácido bórico e EDTA) 0,5X, utilizando-se o marcador de peso molecular de
1Kb (Invitrogen®). Os produtos amplificados foram corados com GelRedTM e
visualizados sob luz UV em transiluminador (Loccus®, L-Pix Touch).
4.2.3.3 Perfil plasmidial
A extração do DNA plasmidial foi realizada utilizando NucleoSpin®
Tissue Kits (Macherey-Nagel, Duren, Germany). Cinco microlitros do extrato
plasmidial adicionado de 3µL de GelRedTM e 3 µL de tampão de carga foi
submetido a eletroforese (500mA, 150W, 150V, 30 min) em gel de agarose
0,8% em tampão TBE (solução de Tris, ácido bórico e EDTA) 0,5X. Os
produtos amplificados foram visualizados sob luz UV em transiluminador
(Loccus®, L-Pix Touch). E. coli V517 contendo os plasmídeos de 54,38kb,

57
7,30kb, 5,56kb, 5,14kb, 3,98kb, 3,08kb, 2,71kb e 2,06kb (MACRINA et al.,
1978) foi utilizada como padrão para comparação.
4. 2.3.4 Atividade da β-hemolisina, DNAse e gelatinase
A atividade da β-hemolisina foi avaliada em ágar Tripticase Soy Agar -
TSA (Himedia) suplementado com 7% de sangue de cavalo, incubado a 37°C
durante 24h. Zonas claras ao redor da colônia indicam produção de β-
hemolisina (EATON; GASSON, 2001).
A atividade da DNAse foi determinada em ágar DNA azul de toluidina,
incubado a 37°C por 24h (APHA, 2002). Zonas claras ao redor da colônia
indicam atividade DNAse positiva. A cepa S. aureus ATCC 25923 foi utilizada
como controle positivo.
A atividade da gelatinase foi avaliada em um meio de cultura preparado
com 1% de extrato de levedura,1,5% de triptona e 12% de gelatina. O isolado
foi cultivado em caldo TSA e uma alíquota de 100µL deste cultivo foi transferido
para 4mL do meio contendo gelatina. Os tubos foram incubados a 30°C por
sete dias. A atividade da gelatinase foi verificada quando o meio se tornou
líquido quando colocado em refrigerador (PEREIRA et al., 2009).
4.3 Modelagem do crescimento e produção de substâncias
antimicrobianas de origem proteica (bacteriocin-like) pelo isolado L.
curvatus P99 em resposta as condições de pH, temperatura e tempo de
incubação
4.3.1 Inóculo e condições de crescimento e produção de substâncias
antimicrobianas de origem proteica (bacteriocin-like)
O inóculo foi obtido a partir da semeadura do isolado de L. curvatus
P99 em 10mL de caldo MRS e incubação a 37°C por 18h sob agitação
constante a 100 oscilações/minuto. Para realização do experimento, foi
adicionado 1mL (1%) do inóculo em 100mL de caldo MRS com diferentes pH
(4, 6,5 e 9) e a incubação procedeu-se em diferentes temperaturas (25, 30 e

58
35ºC) durante um período de 24h. Alíquotas de 2mL foram retiradas, de hora
em hora, para determinação do crescimento microbiano e quantificação da
atividade antimicrobiana.
4.3.2 Ensaios
O crescimento microbiano foi monitorado através da contagem de
células viáveis em placas contendo ágar MRS e incubação a 37°C por 48 a
72h. Para a determinação da atividade antimicrobiana do sobrenadante livre de
células contendo substâncias antimicrobianas de origem proteica (bacteriocin-
like) – SLC, foi utilizada a técnica de difusão em ágar conforme descrito por
Lewus e Montville (1991), com as modificações descritas a seguir. Para isto, 10
µl do sobrenadante bruto e diluído foram adicionados sobre ágar BHI,
previamente inoculado com a bactéria indicadora L. monocytogenes Scott A na
concentração de 105 UFC.mL-1 determinada através da escala McFarland (0,5)
e diluições sucessivas até a concentração desejada. A seguir, as placas foram
incubadas a 35°C por 18h e os halos de inibição foram medidos (BISCOLA et
al., 2013). O SLC produzido por L. lactis subsp. lactis Dy13, contendo nisina, foi
utilizado como controle positivo e como controle negativo, o tampão PBS. O
experimento foi realizado em duplicata com três repetições. A atividade
antimicrobiana foi expressa em unidade arbitrária por mililitro (UA.mL-1).
4.4 Caracterização do sobrenadante livre de células (SLC) contendo
substâncias antimicrobianas de origem proteica (bacteriocin-like)
4.4.1 Atividade antimicrobiana do SLC: concentração inibitória mínima
(CIM) e concentração bactericida mínima (CBM)
A determinação da CIM foi realizada pela técnica de microdiluição em
caldo (CLSI, 2012). O SLC obtido do cultivo do isolado P99 foi diluído em caldo
BHI obtendo concentrações de 50 a 0,1% (v/v). Em seguida, L. monocytogenes
Scott A foi adicionada ao caldo, obtendo a concentração final de 104 UFC.mL-1
em cada poço. As placas de microtitulação foram incubadas a 37°C por 24h, e

59
a leitura realizada em espectrofotômetro de placas (Robonik® Readwel plate) a
625nm, considerando a maior diluição onde não houve multiplicação celular
(OJEDA-SANA et al., 2013).
A CBM foi determinada a partir da transferência de alíquotas de 10µL de
cada poço onde houve inibição no teste de CIM para ágar Oxford (Himedia). A
CBM foi considerada como a menor concentração onde não houve
multiplicação microbiana neste novo meio de cultura. Todos os testes foram
realizados em triplicata.
4.4.2 Atividade antimicrobiana
A determinação foi realizada utilizando a técnica de difusão em ágar
conforme descrito no item 4.3.2. Os micro-organismos alvos foram: B. cereus
ATCC 11778, L. innocua CLIP 12612, L. monocytogenes Scott A, L.
monocytogenes ATCC 7644, L. monocytogenes Siliken, S. aureus ATCC
25923, S. hycus ATCC 11249, S. intermedius ATCC 29663, E. coli O157:H7
ATCC 43895, E. coli ATCC 8739, P. aeruginosa ATCC 15442, S. Tiphymurium
ATCC 14028 e S. dysenteriae ATCC 13313. Os halos de inibição formados
foram medidos em milímetros. O experimento foi realizado em duplicata. O
título de UA.mL-1 foi definido como a recíproca da maior diluição que
apresentou um halo de inibição maior que 2mm.
4.4.3 Estabilidade ao tratamento térmico, refrigeração, congelamento, pH
e agentes químicos
4.4.3.1 Estabilidade ao tratamento térmico
Alíquotas de 200µl do SLC bruto foram submetidas ao tratamento
térmico de 40, 50, 60, 70, 80, 90 e 100°C em banho-maria por 30 minutos e
121°C por 10 e 20 minutos em autoclave. Após os tratamentos, a atividade
antimicrobiana foi avaliada de acordo com a técnica de difusão em ágar como
descrito no item 4.3.2, utilizando L. monocytogenes Scott A como cepa de
referência. Um controle sem tratamento térmico foi utilizado para posterior

60
cálculo da atividade antimicrobiana final. As placas foram incubadas por 18h a
37°C. Os halos foram medidos em milímetros. O experimento foi realizado em
duplicata. A atividade final foi expressa em porcentagem conforme [(HT X 100) /
HC], sendo HT = Halo do sobrenadante após tratamento (mm) e HC = Halo do
controle (mm)
4.4.3.2 Estabilidade a refrigeração e ao congelamento
Alíquotas dos SLC bruto foram incubadas por doze (12) meses sob
refrigeração (4°C) e congelamento (-20°C). A cada 07 dias, durante os dois
primeiros meses, e após 6, 9 e 12 meses, foi determinada a atividade
antimicrobiana do SLC utilizando a técnica de difusão em ágar descrita no item
4.3.2 com a bactéria L. monocytogenes Scott A como linhagem de referência.
As placas foram incubadas por 18h a 37°C. Os halos foram medidos em
milímetros. O experimento foi realizado em duplicata.
4.4.3.3 Estabilidade frente a variações de pH
Alíquotas de 200µl do SLC bruto foram incubadas com 200µl de solução
tampão de pH entre 2 a 10 a 37°C por 2h em banho-maria. Os tampões
utilizados foram: tampão glicina para pH 2 e 3; tampão citrato para pH 4 e 5;
tampão fosfato para pH entre 6 e 8 e tampão TRIS para pH 9 e 10, todos numa
concentração de 0,25M. Após incubação foi realizada a determinação da
atividade antimicrobiana, de acordo com a técnica de difusão em ágar descrita
no item 4.3.2, utilizando a bactéria L. monocytogenes Scott A como linhagem
de referência. Um controle sem adição de tampão foi utilizado para posterior
cálculo da atividade antimicrobiana final. As placas foram incubadas por 18h a
37°C. Os halos foram medidos em milímetros. O experimento foi realizado em
duplicata.

61
4.4.3.4 Estabilidade aos agentes químicos
A ação dos agentes químicos sob a estabilidade da substância
antimicrobiana foi analisada pela adição de 10% (v/v ou m/v) de Tween 20
(Synth), Tween 80 (Synth), Triton X-100 (Vetec), Triton X-114 (Vetec), Dodecil
sulfato de sódio - SDS (Vetec), ureia (Synth), NaCl (Synth), álcool etílico
(Dinâmica), glicerol (Synth) e acetona (Synth) a alíquotas de 1mL de SLC
bruto. Os mesmos foram incubados a 37ºC por 1h (TODOROV et al., 2010).
Decorridos os períodos de incubação de cada uma das análises, o pH
das amostras foi corrigido para 6,0 pela adição de solução de HCl (Synth) 1M
ou NaOH (Synth) 1M e a atividade antimicrobiana dos SLC avaliada conforme
descrito no item 4.3.2. Todos os ensaios foram realizados em duplicata e, como
controle positivo, foi utilizado SLC sem nenhum tratamento.
4.4.4 Avaliação da toxicidade em modelo de Drosophila melanogaster
A avaliação da toxicidade da substância antimicrobiana de origem
proteica presente no SLC do cultivo de L. curvatus P99 foi realizada de acordo
com método descrito por Sudati et al. (2013), utilizando como modelo a mosca
Drosophila melanogaster (linhagem Harwich), proveniente do Laboratório de
Avaliações Farmacológicas e Toxicológicas Aplicadas às Moléculas Bioativas –
Laftambio - UNIPAMPA. As moscas foram mantidas em incubadora BOD, ciclo
claro/escuro 12h (25 ± 1°C; 60% de umidade), alimentadas em meio padrão do
laboratório, composto por 1% de farinha de milho, 1% de levedura, 1% de
germe de trigo, 2% de sacarose, 1% de leite em pó e 0,08% do conservante
nipagin (Sigma-Aldrich®).
Moscas fêmeas, com 3 a 5 dias de idade, foram divididas em 5 grupos,
contendo 25 moscas cada: (1) controle, (2) SLC 10.000 UA.mL-1, (3) SLC
20.000 UA.mL-1, (4) SLC 40.000 UA.mL-1 e (5) SLC 80.000 UA.mL-1. Cada
grupo foi exposto ao tratamento durante 7 dias. A dieta, durante este período,
consistiu de 2% de sacarose, 1% de leite em pó, 1% de ágar, 0,08% nipagin e
nos grupos 2, 3, 4 e 5 o SLC na concentração descrita acima. A taxa de
sobrevivência foi avaliada por contagem diária do número de moscas vivas até

62
ao final do período experimental (7 dias). O ensaio foi realizado em triplicada.
Todas as etapas foram realizadas no Biotério Central da Universidade Federal
do Pampa (UNIPAMPA), Campus Itaqui.
4.5 Elaboração de filmes biodegradáveis incorporados de substância
antimicrobiana de origem proteica presente no SLC de L. curvatus P99
Foi testada a incorporação do SLC de L. curvatus P99 em filmes
biodegradáveis elaborados com amido de batata inglesa adquirida no comércio
local de Pelotas. Os filmes foram produzidos utilizando a técnica de casting
(MÜLLER; LAURINDO; YAMASHITA, 2009). As soluções filmogênicas (SF)
foram compostas por 3g de amido, 0,9g de glicerol, 100µL de Tween 20 e duas
concentrações do SLC (CIM e CBM) e 100mL de água destilada. Um filme sem
a adição de SLC foi utilizado como controle negativo.
O amido, o glicerol e o Tween 20 foram suspensos em água e aquecidos
em banho-maria a 85ºC por 10 minutos. A seguir, a SF foi resfriada a 60ºC e o
SLC incorporado, seguida de homogeneização manual. Em seguida, 20g de
cada SF foi espalhada em placas de Petri acrílicas de 9cm de diâmetro e
mantidas em estufa com circulação de ar a 35ºC por 24h. Após a secagem, os
filmes foram acondicionados em ambiente com umidade relativa de 75 ± 5%
para posterior análise. Foram preparadas duas repetições de cada tratamento.
4.5.1 Avaliação da atividade antimicrobiana dos filmes biodegradáveis
4.5.1.1 Quantificação e manutenção da atividade antimicrobiana
A atividade antimicrobiana dos filmes incorporados com SLC de L.
curvatus P99 foi determinada utilizando-se o método de difusão em ágar. Os
filmes foram assepticamente cortados em discos, de cerca de 8mm de
diâmetro e expostos à luz ultravioleta (UV) em cabine de segurança biológica
classe II por 15 minutos em cada lado, para esterilização. Em seguida, os
filmes foram adicionados à superfície de placas de Petri contendo ágar BHI
(Acumedia), previamente inoculado com L. monocytogenes Scott A (micro-

63
organismo indicador), na concentração de 105 UFC.mL-1 (determinada através
da escala McFarland) (CAGRI; USTUNOL; RYSER, 2001). Como controle
positivo foi utilizado o SLC puro, e como controle negativo foram utilizados
filmes sem a adição de SLC. As placas foram incubadas a 37°C por 18h e
analisadas quanto à formação de halos de inibição, expressos em milímetros,
como a média de duas medições.
A avaliação do tempo de manutenção da atividade antimicrobiana dos
filmes foi realizada nos tempos 0, 2, 4, 6, 8, 10, 14, 21 e 30 dias de
armazenamento, conforme descrito anteriormente, utilizando o micro-
organismo de referência L. monocytogenes Scott A.
4.5.1.2 Modo de ação
A ação in vitro do filme incorporado com SLC contendo substância
antimicrobiana, ao longo do tempo, sobre o patógeno de interesse foi verificado
pelo teste de modo de ação conforme descrito por Diao et al. (2014), com
adaptações.
Uma cultura de 18h a 37°C de L. monocytogenes foi suspensa em
água peptonada - AP 0,1% (Acumedia) até concentração de 108 UFC.g-1 (0,5
McFarland) e diluída em caldo BHI (Acumedia) para obter a concentração final
de 105 UFC.g-1. O caldo BHI inoculado com o patógeno foi dividido em três
grupos: o grupo I (controle), contendo o caldo BHI inoculado com o patógeno e
o filme sem incorporação de SLC; o grupo II, contendo o caldo BHI inoculado
com o patógeno e o filme incorporado da CIM (SLC); e o grupo III, contendo o
caldo BHI inoculado com o patógeno e o filme incorporado com a CBM (SLC).
Foram utilizados recortes de 4cm2 dos filmes para realização deste teste. Os
tubos foram incubados a 37°C, retirando-se amostras de cada grupo nos
tempos 0, 3, 6, 9, 12, 24 e 48h para a quantificação de células viáveis.
Alíquotas dos tubos foram diluídas em AP (0,1%) e inoculadas em Ágar
Oxford (Himedia), seguido de incubação a 30ºC por 24h para posterior
contagem de colônias. Cada grupo continha uma repetição biológica e as
análises foram realizadas em duplicata.

64
4.5.1.3 Atividade antimicrobiana
Os filmes biodegradáveis foram cortados e esterilizados conforme
descrito no item 4.5.1.1. A seguir, foram colocados sobre a superfície de ágar
BHI previamente inoculado com as cepas de referência inibidas no teste
utilizando o SLC. As placas foram incubadas por 24h a 37ºC e, após,
analisadas quanto à formação de halos de inibição, expressos em milímetros,
como a média de duas medições.
4.6 Aplicação dos filmes biodegradáveis incorporados de SLC
(substância antimicrobiana de origem proteica) produzido pelo isolado L.
curvatus P99 em queijo Prato fatiado
Foi pesado e esterilizado 1g de queijo Prato, conforme descrito no item
4.4.5.1. A seguir, a superfície das fatias de queijo foram contaminadas com
10µL de uma suspensão celular de L. monocytogenes Scott A (5 x 104 UFC.g-1)
nas superfícies das fatias. A suspensão celular foi preparada a partir de
diluições seriadas do cultivo ajustado na escala 0,5 de McFarland. Então, os
filmes antimicrobianos foram sobrepostos nas fatias contaminadas. Os
tratamentos dos filmes consistiram em: (1) filme contendo SLC com a CIM; (2)
filme contendo SLC com a CBM; (3) filme sem adição de SLC (controle). As
amostras foram colocadas em placas de petri estéreis e armazenadas a
temperatura de 4ºC durante 10 dias.
A contagem de L. monocytogenes Scott A foi avaliada imediatamente
após a inoculação no queijo e nos tempos 2, 4, 6, 8 e 10 dias de
armazenamento refrigerado. Foram realizadas diluições decimais seriadas a
partir de 1g da amostra, e estas foram inoculadas em ágar Oxford em
duplicata. As placas foram incubadas a 30ºC por 48h e os resultados
expressos em log UFC.g-1.

65
4.7 Desenho experimental e análise estatística
Para as variáveis de tolerância ao trânsito gástrico e intestinal de forma
simulada, capacidade de autoagregação e coagregação, toxicidade do SLC e
modo de ação e aplicação dos filmes contendo SLC o delineamento
experimental utilizado foi completamente casualizado com três repetições. Para
o ensaio da tolerância ao trânsito gástrico de forma simulada, coagregação,
modo de ação e aplicação dos filmes contendo SLC, os tratamentos foram
arranjados em esquema bifatorial. Para a tolerância ao trânsito gástrico de
forma simulada o fator de tratamento A testou, o suco gástrico artificial na
ausência/presença de alimento (SS, LI e LD) e o fator B, o tempo (0, 15, 30, 60,
120, 180 e 240 minutos). Para o ensaio da coagregação, o fator de tratamento
A testou, as temperaturas (20 e 37°C) e o fator B, o tempo (2, 4 e 24h). Para o
ensaio de toxicidade do SLC, o fator de tratamento A testou, os tratamentos
(Controle, 10.000, 20.000, 40.000 e 80.000 UA.mL-1) e o fator B, o tempo (0, 1,
2, 3, 4, 5, 6 e 7 dias). Para o ensaio da modo de ação dos filmes contendo
SLC, o fator de tratamento A testou, os filmes (CT, CIM e CBM) e o fator B, o
tempo (0, 3, 6, 9, 12, 24 e 48h). Para o ensaio da aplicação dos filmes
contendo SLC, o fator de tratamento A testou, os filmes (CT, CIM e CBM) e o
fator B, o tempo (0, 2, 4, 6, 8 e 10 dias).
Para o ensaio da tolerância ao trânsito intestinal de forma simulada e a
capacidade de autoagregação, os tratamentos foram arranjados em esquema
trifatorial. Para o ensaio da tolerância ao trânsito intestinal, o fator de
tratamento A testou o suco intestinal artificial na ausência/presença de alimento
(SS, LI e LD), o fator de tratamento B, os sais biliares (sem e com) e o fator de
tratamento C, o tempo (0, 60, 120, 180 e 240 minutos). Para a capacidade de
autoagregação, o fator de tratamento A testou as temperaturas (20 e 37°C), o
fator B, os micro-organismos (L. curvatus P99 e L. monocytogenes Scott A -
LM) e o fator C, os tempos (2, 20 e 24h).
Os dados obtidos foram analisados quanto à normalidade pelo teste de
Shapiro Wilk; à homocedasticidade pelo teste de Hartley; e, a independência
dos resíduos, por análise gráfica. Posteriormente, os dados foram submetidos
à análise de variância através do teste F (p ≤ 0,05). Constatando-se

66
significância estatística, os efeitos dos sucos gástrico/intestinal simulados e
diferentes filmes foram comparados pelo teste de Tukey (p ≤ 0,05); sal biliar,
temperatura e micro-organismo, foram comparados pelo teste t (p ≤ 0,05); e, os
efeitos dos tempos, foram avaliados por modelos de regressão (p 0,05).
Regressão linear foi utilizada para as variáveis de tolerância ao trânsito intestinal,
autoagregação e coagregação: y = yo + ax, onde: y = variável resposta; yo =
variável resposta correspondente ao ponto mínimo da curva; a = inclinação da
reta ou curva; x = tempo; e regressão não linear (p 0,05) para a variável
tolerância ao trânsito gástrico: y = y0 + a (1 - e-bx), onde: y = variável resposta; y0
= mínima contagem de células; a = valor máximo estimado para a variável
resposta; b = inclinação da curva; x = tempo; e = constante. A seleção dos
modelos foi baseada em: (a) baixo resíduo; (b) baixo p-valor; e (c) alto R2 e R2
adj..
Os dados obtidos do teste de adesão a células HCT-116 foram
submetidos à análise de variância (ANOVA) com intervalo de confiança de 99%
e ao teste de Dunnett (p ≤ 0,001). Os dados obtidos nos testes de toxicidade,
modo de ação e aplicação dos filmes incorporados com SLC foram submetidos
à análise de variância com intervalo de confiança de 95% e ao teste de Tukey
(p ≤ 0,005).
A metodologia de superfície de resposta (MSR) com desenho Box–
Behnken - BB (BOX; BEHNKEN, 1960) foi utilizada para a análise do efeito da
combinação do pH (4, 6,5 e 9), temperatura (25, 30 e 35ºC) e tempo de
incubação (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21,
22, 23 e 24h) na multiplicação celular e produção da substância antimicrobiana
de origem proteica (bacteriocin- like). BB é um planejamento esférico que
envolve um ponto central e pontos médios das arestas.
Três fatores explicativos, nomeados como pH (x1), temperatura (x2) e
tempo de incubação (x3) podem afetar a multiplicação celular e atividade
antimicrobiana de BAL potencialmente bacteriocinogênica. Para cada um dos
fatores, um desenho experimental foi determinado baseado em resultados de
experimentos preliminares. O desenho experimental BB de 3 fatores com três
repetições no ponto central foi adotado nesse estudo, totalizando 120 unidades
experimentais, nas quais cada variável foi testada em três diferentes níveis

67
codificados: mínimo (−1), médio (0) e máximo (+1). A equação de regressão
está apresentada abaixo:
Onde Xi é o código do valor da variável independente ith, xi é o valor
natural da variável independente ith, x0i é o valor natural da variável
independente ith no ponto central e Δxi é a mudança de valor abrupto. Todos os
experimentos foram realizados aleatoriamente para minimizar os efeitos dos
erros estranhos nas respostas observadas e foram repetidos três vezes
independentemente. O desenho experimental conteve os níveis codificados (x)
e reais (X) das variáveis e as respostas medidas foram contagem de células
viáveis (log UFC.mL-1) e atividade antimicrobiana (log UFC.mL-1). Os valores
foram ajustados para as variáveis codificadas (xi, i = 1, 2 e 3) por polinômio de
segunda ordem. A forma geral do polinômio de segunda ordem usado para
ajustar as respostas medidas, segue representada abaixo:
y = b0 + b1x1 + b2x2 + b3x3 + b12x1x2 + b13x1x3 + b23x2x3 + b11x12 + b22x22 + b33x32
Onde y é a variável dependente; b0 é o intercepto; b1, b2 e b3 são os
coeficientes lineares; b11, b22 e b33 são os coeficientes quadráticos; b12, b13 e
b23 são os coeficientes para a interação dos efeitos e Xi é um valor
adimensional codificado de xi (variável independente).
A qualidade do ajuste do modelo foi avaliada baseada em: (a) baixos
resíduos; (b) baixo p-value; (c) baixo desvio padrão; (d) alto coeficiente de
determinação (R2) e R2 adj.; e (e) e pelo teste para verificar a falta de ajuste do
modelo através da análise de variância (ANOVA), utilizando o modelo de
Regressão de Superfície de Resposta (RSREG). O teste para verificar a falta
de ajuste do modelo é realizado para determinar se o modelo escolhido é
adequado para descrever os dados observados, ou se um modelo mais
complexo deverá ser usado. Os testes do modelo foram realizados pelo teste
estatístico de Fischer. A robustez do modelo foi avaliada pela determinação do
coeficiente (R2), ou teste F.

68
4.7.1 Otimização simultânea
Durante a otimização, diversas variáveis respostas que descrevem as
características de qualidade, são otimizadas. Algumas destas variáveis são
maximizadas enquanto algumas são minimizadas. Em muitos casos, estas
respostas competem, como por exemplo, melhorando uma resposta, pode-se
ter um efeito oposto em outra, o que complica ainda mais a situação. Neste
trabalho, as respostas preditivas do modelo foram otimizadas por meio da
metodologia de otimização superfície de resposta (DERRINGER, 1994). A
abordagem geral é a primeira a converter cada resposta (yn) em uma função de
superfície de resposta individual (dn). A escala de superfície de resposta varia
de 0 a 1, onde, se a reposta é sua meta ou alvo, então dn = 1, e se sua
resposta está fora da região aceitável, então dn = 0. Cada resposta é
padronizada em funções desejáveis do tipo dn.
dn = hn(yn)
Onde n é o número total de respostas da medição. Derringer e Suich
(1980) usaram a seguinte função modificada:
Onde minn
y é o mínimo valor aceitável de yn, maxn
y é o máximo valor que
é considerado desejável e r é uma constante positiva. Se r = 1, o dn aumenta
linearmente quando yn aumenta; se r > 1, o dn muda mais rapidamente em
direção a maxn
y e se r < 1, o dn muda menos rapidamente em direção a maxn
y .
As funções de superfície de resposta individuais das respostas ponderadas são
então combinadas para obter D desejável global, definida como a média
geométrica da superfície de resposta individual.
D = (d1, d2, . . ., dn)1/n

69
Onde 0 ≤ D ≤ 1, um valor alto de D indica a maior e melhor função do
sistema, sendo considerado como a solução ótima do sistema. Os valores
ótimos dos fatores são determinados a partir de valores da função de superfície
de resposta individual que maximiza D.
4.7.2 Correlação de Pearson
A presença de correlações entre as variáveis dependentes neste estudo
foram analisadas utilizando o coeficiente de correlação de Pearson (r) geral
para o experimento e também considerando separadamente os fatores de
tratamento (pH, temperatura e tempo de incubação).

70
5 Resultados e discussão
5.1 Caracterização do isolado L. curvatus P99
5.1.1 Avaliação das características probióticas
5.1.1.1 Tolerância ao trânsito gastrointestinal de forma simulada (TGS)
Para a tolerância ao trânsito gástrico ocorreu interação entre os fatores
de tratamentos testados (suco gástrico artificial na ausência/presença de
alimento e tempo) (F = 4031,28; p < 0,0001) (Figura 1 e Tabela 4). Pode-se
visualizar que para os três diferentes sucos gástricos artificiais houve diferença
significativa na contagem de células a partir do tempo 15 minutos, sendo que o
tratamento que continha leite desnatado manteve as contagens em torno de 9
log UFC.mL-1 até 240 minutos (Tabela 4).
Os dados de tolerância ao trânsito gástrico demonstraram ajuste de
modelos não lineares exponenciais para os diferentes sucos gástricos
artificiais, SS (F = 6080995; p < 0,0001), LI (F = 342,1452; p < 0,0001) e LD (F
= 92,8544; p < 0,0001). O isolado P99 não manteve sua viabilidade quando na
presença somente de suco gástrico artificial. Porém, este se manteve viável
quando protegido pelo alimento (Figura 1). O isolado P99 apresentou uma taxa
de sobrevivência de 90,58% quando submetido a presença de leite desnatado
e de 72,91% quando submetido a presença de leite integral. Outros trabalhos
demonstram o efeito protetor do alimento sobre linhagens probióticas durante
estocagem e testes in vitro de tolerância ao trato gastrointestinal (HERMANNS
et al., 2014; SENTER et al., 2015; WANG et al., 2009).

71
Produtos probióticos devem manter a viabilidade das culturas durante a
vida útil do alimento (SUMERI et al., 2008). Além disso, é essencial para a
escolha de um micro-organismo probiótico é a capacidade de sobreviver no
ambiente no qual irá agir. Desta forma, as linhagens precisam sobreviver ao
baixo pH e as enzimas digestivas para que, ao chegarem no intestino, possam
aderir às células epiteliais e competir pela exclusão de patógenos
(MARAGKOUDAKIS et al., 2006).
O aumento do pH, quando ocorre a adição do alimento, é um dos
fatores que contribuem para o aumento da viabilidade das linhagens testadas
quanto ao potencial probiótico. A baixa tolerância de algumas linhagens
quando submetidas ao suco gástrico simulado, não deveria ser suficiente para
excluí-las de aplicações como probióticas, isto porque, segundo Huang e
Adams (2004), as linhagens podem chegar ao intestino em alta concentração
quando tamponados por alimentos ou encapsuladas e promover efeitos
benéficos.
Tempo (minutos)
000 151515 303030 606060 120120120 180180180 240240240
Co
nta
ge
m d
e c
élu
las
(log
UF
C m
L-1 )
0
2
4
6
8
10
y (SS) = 9,62 - 9,62 (1 - e-1,27x
) R2 = 0,99
y (LI) = 9,94 - 3,31 (1 - e-0,007x
) R2 = 0,97
y (LD) = 9,80 - 1,21 (1- e-0,006x
) R2 = 0,91
Figura 1 - Contagem das células viáveis do isolado Lactobacillus curvatus P99 quando submetido à presença de suco gástrico artificial (pepsina 3mg.mL-1 + pH 2,5) na presença/ausência de alimento. Suco gástrico artificial (SS); suco gástrico artificial + leite integral (LI); suco gástrico artificial + leite desnatado (LD).

72
Tabela 4 - Contagem de células viáveis (log UFC.mL-1) de Lactobacillus curvatus P99 quando submetido ao trânsito gástrico simulado
Tempo (minutos)
Suco gástrico artificial SS LI LD
0 9,62±0,00 b1/ 9,71±0,00 a 9,69±0,00 a 15 0,00±0,00 b 9,85±0,00 a 9,81±0,03 a 30 0,00±0,00 c 9,41±0,02 b 9,59±0,01 a 60 0,00±0,00 c 8,87±0,00 b 9,48±0,02 a 120 0,00±0,00 c 7,90±0,00 b 9,03±0,11 a 180 0,00±0,00 c 7,68±0,02 b 9,00±0,00 a 240 0,00±0,00 c 7,31±0,06 b 8,87±0,04 a 1/ Médias (± erro padrão) acompanhadas por mesma letra minúscula na linha não diferem entre si pelo teste de Tukey (p≤0,05) comparando os sucos gástricos artificiais dentro de cada tempo. SS: solução salina; LI: leite integral; LD: leite desnatado.
Para a contagem de células do trânsito intestinal ocorreu interação tripla
entre os fatores de tratamentos testados (suco intestinal artificial na
ausência/presença de alimento, sais biliares e tempo) (F = 12,46; p < 0,0001)
(Figura 2 e Tabela 5). O isolado foi capaz de se manter viável em todas as
condições testadas. Pode-se observar que nos tempos 180 e 240 minutos, o
tratamento com adição de leite desnatado sem a adição de sais biliares, diferiu
significativamente do tratamento que continha apenas suco intestinal artificial,
mas não diferiu do tratamento contendo leite integral. Os tratamentos contendo
sais biliares e alimento (LI e LD), diferiram significativamente nos tempos 120,
180 e 240 minutos. O tratamento contendo apenas suco intestinal artificial e
sais biliares diferiu do tratamento contendo leite integral no tempo 120 minutos
e do tratamento contendo leite desnatado no tempo 240 minutos. A presença
de sais biliares interferiu significativemente nos três tratamentos testados nos
tempos 180 e 240 minutos.
Os dados de contagem de células do trânsito intestinal demonstraram
ajuste de modelos lineares, tanto para ausência de sais biliares e presença do
alimento SS (F = 14,1102; p = 0,0024), LI (F = 26,1947; p = 0,0003) e LD (F =
16,6424; p = 0,0266) quanto na amostra contendo sais biliares nos tratamentos
SS (F = 18,8596; p = 0,0225), LI (F = 12,7145; p = 0,0377) e LD (F = 78,4163;
p < 0,0001) (Figura 2). O isolado P99 manteve um percentual de sobrevivência
acima de 80% em todas as condições intestinais simuladas, sendo as maiores

73
reduções nas contagens de células relacionadas aos tratamentos com a
presença de sais biliares.
Para colonizar o intestino e promover seus efeitos benéficos, a cultura
probiótica deve chegar ao intestino com uma população de no mínimo 6 log
UFC.mL-1 (CHARTERIS et al., 1998). Neste estudo, pode-se observar que, o
isolado P99 manteve sua viabilidade acima de 7,7 log UFC.mL-1. Diversos
autores ao avaliar diferentes espécies de Lactobacillus, quanto a passagem ao
trânsito intestinal simulado demonstraram tolerância dos micro-organismos
quando expostos as condições intestinais (JERONYMO-CENEVIVA et al.,
2014; RUIZ-MOYANO et al., 2008; SENTER et al., 2015).

74
Tempo (minutos)
000 606060 120120120 180180180 240240240
Co
nta
ge
m d
e c
élu
las
(log
UF
C m
L-1)
7,5
8,0
8,5
9,0
9,5
10,0
y (SS SSB) = 9,35 - 0,003x R2 = 0,52
y (LI SSB) = 9,38 - 0,004x R2 = 0,69
y (LD SSB) = 8,98 - 0,002x R2 = 0,85
y (SS CSB) = 9,41 - 0,006x R
2 = 0,86
y (LI CSB) = 9,39 - 0,006x
R2 = 0,81
y (LD CSB) = 9,42 - 0,007x
R2
= 0,87
Figura 2 - Contagem das células viáveis do isolado Lactobacillus curvatus P99 quando submetido à presença de suco intestinal artificial (pancreatina 1mg.mL-1 + pH 8,0) na presença/ausência de alimento e sais bilires (0,3%). SS SSB: suco intestinal artificial sem sais biliares; LI SSB: suco intestinal artificial sem sais biliares + leite integral; LD SSB: suco intestinal artificial sem sais biliares + leite desnatado; SS CSB: suco intestinal artificial com sais biliares; LI CSB: suco intestinal artificial com sais biliares + leite integral; LD CSB: suco intestinal artificial com sais biliares + leite desnatado. Tabela 5 - Contagem de células viáveis (log UFC.mL-1) de Lactobacillus curvatus P99 quando submetido ao trânsito intestinal simulado
Tempo (minutos)
Sal biliar Sem Com
Suco intestinal simulado SS LI LD SS LI LD
0 9,68±0,00 a1/ ns 9,64±0,00 a ns 8,93±0,07 b * 9,68±0,00 a 9,64±0,00 b 9,65±0,00 b 60 8,80±0,05 a ns 8,78±0,02 a ns 8,83±0,07 a ns 8,76±0,06 a 8,67±0,07 a 8,72±0,04 a 120 8,86±0,07 a * 8,81±0,04 a ns 8,86±0,11 a * 8,47±0,04 b 8,70±0,06 a 8,39±0,05 b 180 8,87±0,05 a * 8,81±0,02 ab * 8,65±0,05 b * 8,44±0,02 ab 8,61±0,04 a 8,39±0,05 b 240 8,74±0,02 a * 8,40±0,00 b * 8,40±0,00 b * 8,00±0,02 a 8,00±0,00 a 7,70±0,00 b 1/ Médias (± erro padrão) acompanhadas por mesma letra minúscula na linha não diferem entre si pelo teste de Tukey (p≤0,05) comparando os sucos intestinais artificiais dentro de sal biliar e tempo. *, ns
Significativo e não significativo, respectivamente, pelo teste t (p≤0,05) comparando os sais biliares dentro de cada suco gástrico artificial e tempo. SS: solução salina; LI: leite integral; LD: leite desnatado.
5.1.1.2 Capacidade de autoagregação, coagregação e hidrofobicidade
Para o percentual de autoagregação ocorreu interação tripla entre os
fatores de tratamento (F = 6,22; p = 0,0072) (Figura 3A e Tabela 6). O isolado
P99 apresentou maior percentual de autoagregação na temperatura de 37°C
após 24h de incubação, diferindo significativamente no mesmo período,

75
quando incubado a 20°C. Esse mesmo comportamento foi verificado para o
patógeno L. monocytogenes. Na comparação dos micro-organismos, L.
curvatus P99 diferiu de L. monocytogenes, na temperatura de 20°C no tempo
de 24h, enquanto que na temperatura de 37°C as diferenças ocorreram nos
tempos de 2 e 20h (Tabela 6).
Tabela 6 - Percentual de autoagregação de Lactobacillus curvatus P99 e Listeria monocytogenes (LM) em função da temperatura e tempo
Tempo (horas)
Micro-organismo P99 LM
Temperatura (°C) 20 37 20 37
Autoagregação (%) 2 15,53±1,66 a1/ ns 15,96±0,20 a * 20,28±0,45 a 21,40±0,52 a 20 43,41±1,59 b ns 50,88±1,02 a * 45,50±1,79 b 58,78±1,07 a 24 47,71±1,05 b * 53,33±1,17 a ns 36,73±3,27 b 55,25±0,15 a 1/ Médias (± erro padrão) acompanhadas por mesma letra minúscula na linha não diferem entre si pelo teste t (p≤0,05) comparando as temperaturas dentro de cada micro-organsimo e tempo. *, ns Significativo e não significativo, respectivamente, pelo teste t (p≤0,05) comparando os micro-organismos dentro de cada temperatura e tempo.
Os dados de autoagregação ajustaram-se adequadamente ao modelo
de regressão linear para o isolado P99 nas temperaturas de 20°C (F =
335,7032; p < 0,0001) e 37°C (F = 376,8048; p < 0,0001). Os dados de L.
monocytogenes caracterizaram ajuste de modelo polinomial quadrático, tanto
na temperatura de 20°C (F = 58,5147; p = 0,0003) quanto em 37°C (F =
883,7961; p < 0,0001) (Figura 3A).
No isolado P99, na temperatura de 20°C, ocorreu acréscimo de 208% no
percentual de autoagregação ao comparar o tempo 2 e 24h (Figura 3A). Já, na
temperatura de 37°C, o acréscimo foi superior, 237% para a mesma
comparação. O patógeno L. monocytogenes teve acréscimo inferior ao P99, ao
comparar 2 com 24h, foram verificados acréscimos de 90% para temperatura
de 20°C e 170% em 37°C.
Para o percentual de coagregação ocorreu interação entre os fatores de
tratamentos testados (temperatura e tempo) (F = 6,74; p = 0,0109) (Figura 3B e
Tabela 7). O maior percentual de coagregação entre os dois micro-organismos
foi verificado na temperatura de 37°C após 24h de incubação, diferindo

76
significativamente no mesmo período quando a incubação se deu a 20°C
(Tabela 7).
Tabela 7 - Percentual de coagregação entre Lactobaciilus curvatus P99 e Listeria monocytogenes Scott A em função da temperatura e do tempo
Tempo (horas) Temperatura (°C)
20 37 Coagregação (%)
2 9,48±0,42 a1/ 12,12±1,98 a 4 13,61±0,08 a 17,61±2,62 a 24 42,56±5,49 b 66,08±4,44 a 1/ Médias (± erro padrão) acompanhadas por mesma letra minúscula na linha não diferem entre si pelo teste t (p≤0,05) comparando as temperaturas dentro de cada tempo.
Os dados de coagregação ajustaram-se adequadamente ao modelo de
regressão linear estabelecido nas temperaturas de 20°C (F = 74,2141; p <
0,0001) e 37°C (F = 201,8492; p < 0,0001) (Figura 3B). Na temperatura de
20°C, ocorreu acréscimo de 325% no percentual de coagregação ao comparar
o tempo 2 e 24h (Figura 3B). Já, na temperatura de 37°C, o acréscimo foi
superior, 433% para a mesma comparação.
A habilidade para aderir às células epiteliais e a superfície da mucosa
tem sido sugerida como uma importante propriedade de muitas linhagens
usadas como probióticas. A adesão ao epitélio intestinal é importante para a
colonização do trato gastrointestinal, prevenindo a eliminação por peristaltismo
e promovendo vantagem competitiva no ecossistema (DEL RE et al., 2000). Há
indícios de que este critério funcional esteja relacionado ao aumento da
habilidade de estimulação do sistema imune (SCHILLINGER; GUIGAS;
HOLZAPFEL, 2005).

77
Tempo (horas)
222 202020 242424
Au
toa
gre
ga
çã
o (
%)
15
30
45
60
75
y (P99 - 20°C) = 12,69 + 1,48x R2 = 0,98
y (P99 - 37°C) = 12,88 + 1,77x R2 = 0,98
y (LM - 20°C) = 10,94 + 4,99x - 0,16x2 R2 = 0,96
y (LM - 37°C) = 11,87 + 5,04x - 0,13x2 R2 = 0,99
A
Tempo (horas)
222 444 242424
Coagre
gação (
%)
15
30
45
60
75
y (20°C) = 7,07 + 1,48x R2 = 0,91
y (37°C) = 7,53 + 2,44x R
2 = 0,97
B
Figura 3 - Percentual de autoagregação (A) do Lactobacillus curvatus P99 e Listeria monocytogenes Scott A (LM) nas temperaturas de 20 e 37°C em função do tempo (2, 20 e 24h) e percentual de coagregação (B) de Lactobacillus curvatus P99 e Listeria monocytogenes Scott A sob diferentes temperaturas 20 e 37°C e tempos (2, 4 e 24h).
Muitas vezes, a habilidade de agregação é relatada como uma
propriedade de aderência celular. A agregação bacteriana entre linhagens da
mesma espécie (autoagregação) e entre linhagens de diferentes espécies
(coagregação) é considerada importante em muitos nichos, especialmente no
intestino humano, onde os probióticos são bioativos. A relação entre
autoagregação e habilidade de adesão tem sido reportada para algumas
espécies de bifidobactérias (DEL RE et al., 2000; PÉREZ et al., 1998) e
também, uma relação entre habilidade de adesão e hidrofobicidade para alguns
lactobacilos tem sido observada (DEL RE et al., 2000).
Autoagregação é considerada necessária para a adesão de linhagens
probióticas às células epiteliais e a habilidade de coagregação pode formar
uma barreira para prevenir a colonização por patógenos. Geralmente, as

78
linhagens probióticas mostram maior habilidade de autoagregação do que os
patógenos (COLLADO; MERILUOTO; SALMINEN, 2008).
Todorov et al. (2011) observaram que o percentual de autoagregação de
isolados de L. curvatus a 37°C foi menor que 10%. Os autores visualizaram
ainda que as linhagens apresentaram baixos níveis de coagregação com L.
monocytogenes 603 (< 20%). Collado; Meriluoto e Salminen (2008)
observaram que a melhor temperatura para autoagregação das linhagens
probióticas foi 37°C e dos patógenos foi a de 20°C. O resultado difere do
encontrado no presente estudo, pois tanto o isolado P99 quanto o patógeno
testado, tiveram maior habilidade de autoagregação a 37°C. Este resultado é
positivo visto que 37°C é a temperatura corporal humana, podendo-se inferir
que este micro-organismo terá a capacidade de se aderir ao epitélio intestinal.
Baixos níveis de coagregação com patógenos pode prevenir a formação
de biofilmes, eliminando, desta maneira, os patógenos do trato gastrointestinal.
Por outro lado, altos níveis de coagregação com patógenos pode facilitar a
ação antimicrobiana e, consequentemente, a exclusão dos patógenos do trato
gastrointestinal (SANTOS et al., 2015).
Apenas 13,17% das células do isolado P99 aderiu ao solvente apolar.
Hidrofobicidade da superfície celular é uma interação não específica entre as
células microbianas e hospedeiro. A interação inicial pode ser fraca, muitas
vezes reversível e precede processos de adesão subsequentes mediadas por
mecanismos mais específicos que envolvem proteínas da superfície celular e
ácidos lipoteicóicos (SADRANI et al., 2014; TODOROV et al., 2011).
A determinação da adesão de micro-organismos a hidrocarbonetos,
como uma forma de estimar a capacidade de uma estirpe de aderir às células
epiteliais é uma abordagem qualitativa. Porém, Todorov et al. (2011)
demonstraram em seu estudo que as linhagens que tiveram alta capacidade de
hidrofobicidade não aderiram em altos níveis às células HT-29. Portanto, a
hidrofobicidade pode auxiliar na adesão, mas não é pré-requisito para que esta
ocorra.

79
5.1.1.3 Ensaio de adesão celular
Lactobacillus curvatus P99 demonstrou capacidade de adesão de 0,5%
às células intestinais HCT-116. Listeria monocytogenes ATCC 7644 apresentou
um percentual de adesão de 3,41%, o qual foi significativamente maior que a
do isolado P99. Normalmente, ensaios in vitro da capacidade de adesão a
células intestinais são realizados utilizando linhagens celulares Caco-2 ou HT-
29 derivadas de adenocarcinoma. Porém, em estudo realizado por Ayeni et al.
(2011), pode-se perceber que a linhagem celular utilizada influenciou a adesão
de isolados de L. paracasei. Quando utilizadas as células Caco-2 e HT-29, os
autores observaram que a adesão de L. paracasei foi menor que a adesão de
L. rahmnosus utilizado como controle. Enquanto que, quando utilizaram-se
células HT-29 MTX e HeLa, a adesão dos isolados foi similar ou maior. Pode-
se inferir que a linhagem de células intestinais pode ter influenciado na
capacidade de adesão do mesmo.
A adesão de BAL ao epitélio intestinal é influenciada por múltiplos
fatores, incluindo interações eletrostáticas, interações hidrófobicas e estruturas
bacterianas específicas, tais como cápsula, proteínas de superfície e muco
epitelial (YU et al., 2013). Contudo, a aderência in vitro de uma linhagem não
pode ser relacionada com a aderência in vivo ao trato intestinal humano, pois
diferentes fatores influenciam a adesão influência, como os mecanismos de
defesa do hospedeiro, a concorrência com microbiota residente, a presença de
muco e fluido peristáltico (GARCÍA-RUIZ et al., 2014).
Xu et al. (2009) avaliaram linhagens de micro-organismos probióticos de
diferentes espécies quanto a habilidade de aderir a células Caco-2 e
observaram que L. acidophilus ADH, L. brevis KACC 10553 e L. casei KACC
12413 tiveram baixa capacidade de adesão (< 6%). Todorov et al. (2011)
observaram em seu estudo que três isolados de L. curvatus apresentaram altos
percentuais de adesão a células Caco-2 (ET06 – 12,6%; ET30 – 13,9%; ET31
– 12,3%).
A adesão in vitro a linhagens de células intestinais é um teste
frequentemente utilizado para avaliar o potencial de isolados candidatos a

80
probióticos, uma vez que simula a capacidade da bactéria de realizar a
colonização transitória do epitélio intestinal, a qual permitiria o probiótico
exercer seu efeito benéfico (FAO/WHO, 2006).
A fixação a superfície das mucosas pode prolongar a persistência dos
probióticos o que permite conferir benefícios ao hospedeiro (JERONYMO-
CENEVIVA et al., 2014). Entretanto, outros mecanismos, como a agregação e
altas taxas de multiplicação para evitar a lavagem, ou no caso de probióticos, o
consumo contínuo, podem estar envolvidos na persistência bacteriana no
intestino (VAUGHAN; MOLLET; DEVOS, 1999).
5.1.1.4 Atividade antimicrobiana
Lactobacillus curvatus P99 apresentou atividade antimicrobiana contra
todas as linhagens Gram-positivas e Gram-negativas testadas. O tamanho dos
halos de inibição formados estão apresentados na tabela 8. As linhagens
produtoras de bacteriocinas da classe II tem sido reportadas, principalmente
pela sua forte ação antilisterial. Hartmann et al. (2011) observaram em seu
estudo que o isolado L. curvatus IDE0444 inibiu L. monocytogenes, mas não
inibiu B. cereus, S. aureus e Salmonella enterica.
O principal efeito antimicrobiano de BAL é a taxa de acidificação
(LEROY; VERLUYTEN; DE VUYST, 2006). Porém, outros metabólitos, como
as bacteriocinas, têm sido relatados como os responsáveis pela ação
antagonista contra bactérias patogênicas e deteriorantes (AHMADOVA et al.,
2013a; BARBOSA et al., 2015; ÜNLÜ; NIELSEN; IONITA, 2015). A ação
antagonista de BAL já foi descrita por diversos autores (BELLO et al., 2010;
NERO et al., 2008; SAWITZKI et al., 2008), principalmente, em relação a
patógenos Gram-positivos.
A presença da dupla camada lipídica (membrana externa) de bactérias
Gram-negativas impede a interação das substâncias antagonistas específicas,
como as bacteriocinas, com a parede e a membrana plasmática. Porém, outros
trabalhos demonstram o isolamento de BAL com potencial antagônico também
contra patógenos Gram-negativos (MEIRA et al., 2010; NERO et al., 2008).

81
Tabela 8 - Espectro de inibição da atividade antimicrobiana produzida pelo isolado Lactobacillus curvatus P99
Micro-organismos Origem Zona de inibição (mm) Gram-positiva Bacillus cereus ATCC 11778 ATCC1 16.5 Listeria inoccua CLIP 12612 CLIP2 16.5 Listeria monocytogenes Scott A ATCC 20.0 Listeria monocytogenes ATCC 7644 ATCC 16.5 Listeria monocytogenes Siliken TIAR3 16.5 Staphylococcus aureus ATCC 25923 ATCC 15.0 Staphylococcus hycus ATCC 11249 ATCC 17.5 Staphylococcus intermedius ATCC 29663 ATCC 17.0 Gram-negativa Escherichia coli O157:H7 ATCC 43895 ATCC 13.0 Escherichia coli ATCC 8739 ATCC 23.0 Pseudomonas aeruginosa ATCC 15442 ATCC 26.5 Salmonella Typhimurium ATCC 14028 ATCC 18.5 Shigella dysenteriae ATCC 13313 ATCC 19.0 1 ATCC: American Type Culture Collection. 2 CLIP: Listeria Collection, Instituto Pasteur 3 TIAR: Tasmanian Institute of Agricultural Research, Universidade da Tasmania, Austrália
5.1.1.5 Avaliação da atividade da enzima β-galactosidase
O isolado P99 não apresentou atividade da enzima β-galactosidase.
Dentre os lactobacilos provenientes de ambientes cárneos, muitos são
incapazes de fermentar lactose. Obst et al. (1992), que observaram que todas
as linhagens de L. curvatus provenientes de produtos cárneos investigadas
foram β-galactosidase negativa. Já, Gheytanchi et al. (2009) observaram que,
L. curvatus isolado de leite, analisado para a atividade da β-galactosidase,
apresentou resultado positivo para a enzima.
Lactose é o principal carboidrato do leite, sendo essencial para a
multiplicação das culturas iniciadoras utilizadas na produção de produtos
lácteos fermentados (OBST et al., 1992). Além de, também garantir o
desenvolvimento de culturas iniciadoras, na indústria de produtos cárneos
fermentados, a lactose é utilizada como agente emulsificante e ligante, uma
vez que ele se liga a água livre diminuindo assim o tempo de fermentação
(MARIANSKI; MARIANSKI, 2009).

82
5.2.2 Avaliação das características tecnológicas
5.2.2.1 Multiplicação em diferentes temperaturas e concentrações de sal
(NaCl)
O isolado P99 foi capaz de se multiplicar nas temperaturas de 10 e
45°C, bem como nas concentrações de 4,5 e 6% de NaCl. A capacidade de se
desenvolver em ampla faixa de temperatura e tolerar grandes concentrações
de NaCl sugere que este micro-organismo pode persistir durante processos de
maturação de alimentos fermentados como queijos e salames (BARBOSA et
al., 2015; GIRAFFA, 2003).
Barbosa et al. (2015) verificaram o efeito da adição de diferentes
concentrações NaCl no desenvolvimento de dois isolados de L. curvatus
provenientes de salame tipo italiano e observaram que estes foram capazes de
se multiplicar em meio suplementado com até 6% de NaCl. Rai; Tamang e
Palni (2010), em seu estudo, observaram que dois isolados de L. curvatus
provenientes de produtos cárneos avaliados foram capazes de se multiplicar na
presença de 6,5% de NaCl e na temperatura de 8°C, porém, apenas um deles
foi capaz de se multiplicar a temperatura de 45°C.
5.2.2.2 Fermentação da glicose
O isolado não produziu gás a partir da fermentação da glicose. Este
resultado se assemelha aos resultados encontrados por Cousin et al. (2015) e
Rai et al. (2010) ao avaliarem isolados de L. curvatus. Espécies que não
produzem gás a partir da glicose são desejáveis como culturas iniciadoras em
embutidos fermentados, uma vez que, não irão propiciar a formação de
cavidades indesejáveis e, por serem homofermentativas produzirão ácido lático
como principal metabólito acidificando favoravelmente o meio
(BUCKENHÜSKES, 1993).

83
5.2.2.3 Atividade proteolítica e lipolítica
O isolado P99 foi capaz de hidrolisar a caseína, porém não foi capaz de
hidrolisar a albumina de soro bovino. Proteólise e lipólise são eventos
bioquímicos que ocorrem durante a maturação de diferentes alimentos
fermentados e contribuem para a formação do flavour. A principal contribuição
das BAL para o flavour, atribui-se ao ácido lático e aos compostos voláteis
produzidos a partir da fermentação de carboidratos (TULINI et al., 2015) e,
geralmente, não possuem intensa atividade proteolítica e lipolítica (LEROY;
VERLUYTEN; DE VUYST, 2006).
Papamanoli et al. (2003) ao avaliarem a capacidade de hidrolisar a
caseína de 24 isolados de L. curvatus observaram que todos apresentaram
atividade proteolítica. O isolado não foi capaz de hidrolisar lipídios, sendo este
resultado esperado, uma vez que Lactobacillus apresentam fraca atividade
lipolítica, como já citado por Leroy; Verluyten; De Vuyst (2006). Casaburi et al.
(2016) também não observaram atividade lipolítica pelo isolado de L. curvatus
54M16.
5.2.3 Aspectos de segurança
5.2.3.1 Avaliação qualitativa e quantitativa da susceptibilidade aos
antimicrobianos de uso clínico
O isolado P99 apresentou resistência fenotípica a ciprofloxacina,
sulfametoxazol-trimetoprim, sulfonamida e vancomicina. Resistência
intermediária foi observada para ampicilina. L. curvatus P99 foi sensível a
amicacina, cefalotina, clindamicina, cloranfenicol, eritromicina, estreptomicina,
gentamicina penicilina e tetraciclina (Tabela 9 e Apêndice A). Porém, não foi
detectada a presença de nenhum gene de resistência aos antimicrobianos de
uso clínico pesquisados (Figuras 4, 5, 6, 7 e 8).

84
Tabela 9 – Análise de susceptibilidade fenotípica a antimicrobianos de uso clínico de Lactobacillus curvatus P99
Antimicrobianos Zona de inibição (mm) S – I – R*
Amicacina 40 S
Ampicilina 16 I
Cefalotina 40 S
Ciprofloxacina 10 R
Clindamicina 45 S
Cloranfenicol 45 S
Eritromicina 55 S
Estreptomicina 35 S
Gentamicina 40 S
Penicilina 35 S
Sulfametoxazol-trimetoprim 0 R
Sulfonamida 0 R
Tetraciclina 35 S
Vancomicina 0 R
*S = sensível; I = sensibilidade intermediária; R = resistente
3
Figura 4 - Amplificação do gene blaZ (172 pb) associado a resistência a antimicrobianos β-lactâmicos. Número 1: Ausência do gene blaZ em Lactobacillus curvatus P99. Número 2: Controle positivo - Staphylococcus aureus S10. Número 3: Controle negativo - Escherichia coli NCTC 12900; Número 4: Controle da reação – água ultrapura. Letra M: marcador de tamanho molecular 1 kb.
1 2 3 4 M

85
(A) (B) (C)
Figura 5 - Amplificação dos genes (A) sul1 (840 pb), (B) sul2 (293 pb) e (C) dfrG (422 pb) associados a resistência a antimicrobianos Inibidores da Via Folato. Número 1: Ausência do gene sul1 em Lactobacillus curvatus P99. Número 2: Controle positivo gene sul1 - Salmonella Typhymurium 22C. Número 3: Controle negativo gene sul1 - Escherichia coli 25922. Número 4: Controle da reação gene sul1 – água ultrapura. Número 5: Ausência do gene sul2 em Lactobacillus curvatus P99. Número 6: Controle positivo gene sul2 - Salmonella Typhymurium 171B2. Número 7: Controle negativo gene sul2 - Escherichia coli 25922. Número 8: Controle da reação gene sul2 – água ultrapura.Número 9: Ausência do gene dfrG em Lactobacillus curvatus P99. Número 10: Controle positivo gene dfrG - Staphylococcus aureus S10. Número 11: Controle negativo gene dfrG - Escherichia coli NCTC 12900. Letra M: marcador de tamanho molecular 1Kb.
(A) (B)
Figura 6 - Amplificação dos genes (A) ermB (424 pb) e (B) ermC (299 pb) associados a resistência a antimicrobianos Macrolídeos. Número 1: Ausência do gene ermB em Lactobacillus curvatus P99. Número 2: Controle positivo gene ermB - Staphylococcus aureus 218. Número 3: Controle negativo gene ermB - Salmonella Typhimurium ATCC 14028. Número 4: Controle da reação gene ermB – água ultrapura. Número 5: Ausência do gene ermC em Lactobacillus curvatus P99. Número 6: Controle positivo gene ermC - Staphylococcus aureus S10. Número 7: Controle negativo gene ermC - Salmonella Typhimurium ATCC 14028. Número 8: Controle da reação gene ermC – água ultrapura. Letra M: marcador de tamanho molecular 1Kb.
1 2 3 4 M 5 6 7 8 M M 9 10 11 12
1 2 3 4 M 5 6 7 8 M

86
(A) (B) (C)
(D) (E)
(F) (G)
Figura 7 - Amplificação dos genes (A) tetA (953 pb), (B) tetB (1.169 pb), (C) tetK (360 pb), (D) tetL (1.028 pb), (E) tetM (158 pb), (F) tetO (781 pb) e (G) Tn916-1545 (1.028 pb) associados a resistência a antimicrobianos Tetraciclinas. Número 1: Ausência do gene tetA em Lactobacillus curvatus P99. Número 2: Controle positivo gene tetA - Salmonella Tiphymurium 22C. Número 3: Controle negativo gene tetA - Staphylococcus aureus 25923. Número 4: Controle da reação gene tetA – água ultrapura. Número 5: Ausência do gene tetB em Lactobacillus curvatus P99. Número 6: Controle positivo gene tetB - Salmonella Typhymurium 171B2. Número 7: Controle negativo gene tetB –
M 1 2 3 4 5 6 7 8 M 9 10 11 12 M
13 14 15 16 M 17 18 19 20 M
21 22 23 24 M 25 26 27 28 M

87
Staphylococcus aureus ATCC 25923. Número 8: Controle da reação gene tetB – água ultrapura. Número 9: Ausência do gene tetK em Lactobacillus curvatus P99. Número 10: Controle positivo gene tetK - Staphylococcus aureus S2. Número 11: Controle negativo gene tetK – Salmonella Enteritidis ATCC 13076. Número 12: Controle da reação gene tetK – água ultrapura. Número 13: Ausência do gene tetL em Lactobacillus curvatus P99. Número 14: Controle positivo gene tetL - Staphylococcus aureus S2. Número 15: Controle negativo gene tetL – Staphylococcus aureus ATCC 25923. Número 16: Controle da reação gene tetL – água ultrapura. Número 17: Ausência do gene tetM em Lactobacillus curvatus P99. Número 18: Controle positivo gene tetM - Staphylococcus aureus S10. Número 19: Controle negativo gene tetM – Staphylococcus aureus ATCC 25923. Número 20: Controle da reação gene tetM – água ultrapura. Número 21: Ausência do gene tetO em Lactobacillus curvatus P99. Número 22: Controle positivo gene tetO - Campylobacter termófilo 119. Número 23: Controle negativo gene tetO – Listeria monocytogenes ATCC 7644. Número 24: Controle da reação gene tetO – água ultrapura. Número 25: Ausência do gene Tn916-1545 em Lactobacillus curvatus P99. Número 26: Controle positivo gene Tn916-1545 - Staphylococcus aureus S10. Número 27: Controle negativo gene Tn916-1545 – Listeria monocytogenes ATCC 7644. Número 28: Controle da reação gene Tn916-1545 – água ultrapura. Letra M: marcador de tamanho molecular 1Kb.
(A) (B)
Figura 8 - Amplificação dos genes (A) strA (645 pb) e (B) strB (510 pb) associados a resistência a antimicrobianos Aminoglicosídeos. Número 1: Ausência do gene strA em Lactobacillus curvatus P99. Número 2: Controle positivo gene strA - Salmonella Typhimurium 140B1. Número 3: Controle negativo gene strA - Escherichia coli ATCC 25922. Número 5: Ausência do gene strB em Lactobacillus curvatus P99. Número 6: Controle positivo gene strB - Staphylococcus aureus S10. Número 7: Controle negativo gene strB - Listeria monocytogenes ATCC 7644. Letra M: marcador de tamanho molecular 1Kb.
A resistência aos antimicrobianos por micro-organismos utilizados em
alimentos é um elemento de extrema relevância para demonstrar a segurança
para utilização de novas linhagens como culturas coadjuvante de tecnologia,
iniciadoras e/ou probióticas (FAO/WHOO, 2006). A resistência aos
antimicrobianos de uso clínico pode ser intrínseca ou adquirida. A resistência
intrínseca não é associada a elementos genéticos móveis e representam
baixos riscos de disseminação (SAARELA et al., 2007). Genes de resistência a
1 2 3 M 5 6 7 M

88
antibióticos adquiridos, especialmente os associados com elementos genéticos
móveis, como plasmídeos e transposons, podem transferir estes genes para
patógenos desde o produto alimentício até a sua consumação (NAWAZ et al.,
2011).
Métodos genotípicos podem confirmar a presença de genes que
conferem a resistência a antimicrobianos, entretanto, o perfil de resistência
fenotípico deve ser levado em consideração. Muñoz et al. (2014)
demonstraram que isolados multirresistentes fenotipicamente não apresentam
genes de resistência para os mesmos antimicrobianos. Este fato sugere que
outros mecanismos podem contribuir para a resistência fenotípica ou que
outros genes podem estar envolvidos no mecanismo de aquisição da
resistência.
Além da presença de determinados genes de resistência, as bactérias
podem ser capazes de evitar antimicrobianos por inativação direta da molécula
ativa, perda da susceptibilidade bacteriana ao antimicrobiano através da
modificação do alvo da ação e redução da concentração de droga que atinge a
molécula alvo sem qualquer modificação do próprio composto (bomba de
efluxo) (KUMAR; SCHWEIZER, 2005). A resistência à eritromicina, tetraciclina
e vancomicina é uma característica intrínseca comum para linhagens de
Lactobacillus (ASHRAF; SHAH, 2011). Mecanismos bioquímicos similares aos
da resistência adquirida estão relacionado para a resistência intrínseca. Porém,
em alguns casos, nem mecanismos bioquímicos nem genes responsáveis pela
resistência instrínseca foram identificados (EFSA, 2003)
5.2.3.2 Determinação dos genes para descarboxilação de aminoácidos
No presente estudo, não foi detectada a presença dos genes que
codificam para produção de histamina e tiramina (Figura 9). Este resultado é
significativo, pois estas AB tem sido as mais estudadas, sendo consideradas as
aminas de origem bacteriana mais importantes em alimentos e por apresentam
efeitos toxicológicos derivados das suas propriedades psicoativas e vasoativas
(BOVER-CID; HOLZAPFEL, 1999). Muitas linhagens de L. curvatus são
associadas com a produção de AB (KIM ; KIM, 2014; LADERO et al., 2015;

89
LATORRE-MORATALLA et al., 2014; O’SULLIVAN et al., 2015), porém, para
usar uma BAL como cultura iniciadora, esta não deve apresentar atividade
aminoácido-descarboxilase (AMMOR; MAYO, 2007).
Figura 9 - Amplificação dos genes tyrdc (1.133 pb) e hdc (440 pb) associados a produção de aminas biogênicas tirosina e histamina. Número 1: Controle positivo - Lactobacillus curvatus LC254. Número 2: Lactobacillus curvatus P99. Número 3: Controle da reação Letra M: marcador de tamanho molecular 1Kb. Controle interno da reação: primers universal 16S rRNA.
As aminas biogênicas (AB) são compostos derivados da
descarboxilação de aminoácidos através do metabolismo microbiano
(CAPOZZI et al., 2011). Diferentes alimentos podem conter altos níveis de AB
como peixes, queijos, produtos cárneos e vegetais fermentados, bebidas e
vinhos (LINARES et al., 2011). O acúmulo de AB em alimentos requer a
presença de precursores (aminoácidos provenientes da atividade proteolítica),
micro-organismos com atividade aminoácido-descarboxilase e condições
favoráveis para multiplicação e descarboxilação (AMMOR; MAYO, 2007).
5.2.3.3 Perfil plasmidial
Lactobacillus curvatus P99 não apresentou plasmídeos (Figura 10).
Plasmídeos podem carrear diversos genes como para produção de
antimicrobianos, para resistência a antimicrobianos de uso clínico e genes de
toxicidade (ASHRAF; SHAH, 2011; ENTIAN; DE VOS, 1996; LADERO et al.,
2015). A ausência de plasmídeo no isolado P99 é um resultado positivo e

90
corrobora com os resultados obtidos para a ausência de genes de resistência a
antimicrobianos de uso clínico e de descarboxilação de aminoácidos. Pode-se
inferir também que as substâncias antimicrobianos de origem proteica
(bacteriocin-like) são codificadas por genes cromossomais uma vez que
elementos móveis do tipo plasmídeo não estão presentes.
1 2 3 M
Figura 10 – Perfil plasmidial do isolado Lactobacillus curvatus P99. Número 1: DNA genômico de Lactobacillus curvatus P99; Número 2: DNA plasmideal de Lactobacillus curvatus P99; Número 3: Escherichia coli V517; Letra M: marcador de tamanho molecular (Ladder 1kb).
Aymerich et al. (2006) analisaram o perfil plasmidial de 88 isolados de
BAL e observaram que 14 não apresentaram plasmídeos. O genoma de L.
plantarum ZJ95, isolado de fezes de recém-nascido, foi sequenciado por Li; Gu
(2016) e estes observaram a presença de dois plasmídeos, identificados como
ZJ95p1 (48.556 pb) e ZJ95p2 (12484 pb). Hebert et al (2012) sequenciaram o
genoma de L. curvatus CRL705, produtor das bacteriocinas lactocina 705 e
“AL705” e observaram que este apresentou dois plasmídeos circulares, pRC12
(12.342 pb) e pRC18 (18.664 pb).
5.2.3.4 Atividade β-hemolisina, DNAse e gelatinase
O isolado P99 não apresentou atividade hemolítica, da DNAse e nem da
gelatinase. A β-hemolisina é uma enzima capaz de lisar eritrócitos humanos e
de animais (EATON; GASSON, 2001). A DNAse é uma enzima capaz de

91
degradar o ácido nucleico (DNA). A gelatinase é uma protease que hidrolisa
gelatina, colágeno, caseína, hemoglobina e outros peptídeos bioativos, por isso
sugerem um papel importante em processos inflamatórios (BARBOSA; GIBBS;
TEIXEIRA, 2010; SEMEDO et al., 2003). Em muitos casos, Enterococcus e
Streptococcus são mais importantes quando há envolvimento de BAL em
infecções hospitalares do que o gênero Lactobacillus. A presença de fatores de
virulência nestas linhagens pode aumentar a habilidade em causar doenças,
contriubuindo para os riscos de infecções (JERONYMO-CENEVIVA et al.,
2014).
Muñoz-Atienza et al. (2013) investigaram a produção da hemolisina para
99 isolados de BAL, dentre os quais, avaliaram um isolado de L. curvatus e não
observaram atividade hemolítica para este. Barbosa; Gibbs e Teixeira (2010)
avaliaram isolados de Enterococcus provenientes de fermentados cárneos para
e observaram que 26% (47 isolados) foram gelatinase positiva e que nenhum
isolado apresentou atividade para DNAse. Sieladie et al., (2011) não
observaram atividade hemolítica e nem da gelatinase para isolados de
Lactobacillus potencialmente probióticos. Mahasneh; Hamdan e Mahasneh,
(2015) analisaram 17 isolados de Lactobacillus com potencial probiótico e
observaram que estes não apresentaram atividade hemolítica e foram DNAse e
gelatinase negativa.
5.3 Modelagem do crescimento e produção de substâncias
antimicrobianas de origem proteica (bacteriocin-like) pelo isolado L.
curvatus P99 em resposta ao pH e temperatura
5.3.1 Multiplicação microbiana e produção de substâncias
antimicrobianas de origem proteica (bacteriocin-like)
Os dados experimentais foram utilizados para calcular a equação
polinomial de segunda ordem com os coeficientes dos modelos. Os
coeficientes de regressão para as equações polinomiais de segunda ordem e
resultados para os efeitos linear, quadrático e interação, bem como os
coeficientes de determinação, estão apresentados na tabela 10.

92
Tabela 10 - Coeficientes de regressão (dados codificados), R2 e R2 adj. dos modelos de superfície de resposta para contagem de células viáveis e atividade antimicrobiana de Lactobacillus curvatus P99
Coeficientes Respostas
Contagem de células viáveis (log UFC.mL-1)
Atividade antimicrobiana (UA.mL-1)
b0 (intercepto) 8,681* 69102* b1 (pH) 0,370* 10429* b2 (T) -0,140* 4370,83** b3 (TI) 0,796** 32201** b12 (pH x T) 0,257* 4187,50** b13 (pH x TI) 0,554* 20734* b23 (T x TI) -0,128** 7270,50** b11 (pH x pH) -1,026* -57604* b22 (T x T) 0,001** 0,001** b33 (TI x TI) -0,479* 1511,85** R2 0,91 0,94 R2 adj. 0,90 0,93
T, temperatura. TI, tempo de incubação. * Significativo a p < 0,001. ** Significativo a p < 0,01.
A significância de cada coeficiente foi determinada pelo teste t de
Student e p-valor. Um resumo das análises de variância para os modelos
quadráticos, estão apresentados na tabela 11. A ANOVA dos modelos de
regressão indica que os modelos resultantes foram altamente significativos (p <
0,05), demonstrando falta de ajuste não significativa. Estes modelos podem ser
usados para descrever os efeitos das variáveis independentes selecionadas
(pH, temperatura e tempo de incubação) na multiplicação celular e produção de
substâncias antimicrobianas.
A multiplicação celular foi significativamente afetada pelo pH,
temperatura e tempo de incubação (Tabela 10). Foi observado efeito linear e
quadrático e suas interações com pH, temperatura e tempo de incubação, para
multiplicação celular. A equação de superfície de resposta resultante descreve
a multiplicação celular perfeitamente (R2 = 0,91 e R2 adj. = 0,90). A falta de
ajuste não foi significativa (p = 0,3497), demonstrando que a equação de
superfície de resposta descreve adequadamente os dados (Tabelas 10 e 11).
O p-valor para o modelo (< 0,0001) e a falta de ajuste não significativa
(0,1034) indicam que os dados experimentais obtidos tem um bom ajuste com
o modelo, para atividade antimicrobiana. A equação de regressão resultante

93
apresenta coeficiente de determinação (R2) de 0,94 e R2 adj. de 0,93, indicando
que 94% da variação total foi explicada pelo modelo. Foi observado efeito
linear e quadrático e suas interações com pH, temperatura e tempo de
incubação na atividade antimicrobiana (Tabelas 4 e 5).
Tabela 11 - Resultados da análise de variância (ANOVA) para equação de regressão para contagem de células viáveis e atividade antimicrobiana de Lactobacillus curvatus P99
Resposta Fonte SQ QM Valor F Pr > F
Contagem de células viáveis
Linear 90,646 247,10 <0,0001 Quadrática 61,473 251,36 <0,0001 Interação 35,175 95,89 <0,0001 Total 187,294 191,46 <0,0001 Falta de ajuste 13,143 0,1991 1,07 0,3497 Erro puro 4,234 0,0295
Atividade antimicrobiana
Linear 123730726695 91,93 <0,0001 Quadrático 173818154512 193,72 <0,0001 Interação 36948537304 27,45 <0,0001 Total 334497418512 93,20 <0,0001 Falta de ajuste 6143936 13,924 0,1034 Erro puro 6933333 48148
SQ, soma dos quadrados; QM, quadrado médio.
A análise de regressão dos dados mostrou que a multiplicação celular foi
significativamente afetada pelo pH, temperatura e tempo de incubação. A
relação entre a multiplicação celular e as variáveis é retratada na figura 11 (A,
B e C). A análise rotacional canônica indicou que o ponto estacionário foi um
ponto de sela, sugerindo que um movimento longe destes pontos causaria um
aumento ou diminuição da resposta, dependendo do sentido do movimento. De
acordo com a otimização matemática, as melhores condições foram: pH 6,3, a
temperatura de 29,3°C e um tempo de incubação de 18,6 h. Nesta condições, a
contagem de células viáveis foi de 8,86 log UFC.mL-1.

94
6,5
7,0
7,5
8,0
8,5
9,0
252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525
353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535
303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030
252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525
353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535
4,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,0
6,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,5
9,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,0
Conta
gem
de c
élu
las v
iáveis
(Log
UF
C m
L-1)
Tem
pera
tura
(ºC
)
pH
6,5 7,0 7,5 8,0 8,5 9,0
A
6,0
6,5
7,0
7,5
8,0
8,5
9,0
9,5
36
912
1518
2124
4,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,0
6,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,5
9,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,0
Conta
gem
de c
élu
las v
iáveis
(Log
UF
C m
L-1)
Tem
po d
e in
cuba
ção
(hor
as)
pH
6,0 6,5 7,0 7,5 8,0 8,5 9,0 9,5
B
7,0
7,5
8,0
8,5
9,0
9,5
10,0
10,5
36
912
1518
2124
252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525
353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535
303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030
252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525
353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535
Conta
gem
de c
élu
las v
iáveis
(Log
UF
C m
L-1)
Tem
po d
e in
cuba
ção
(hor
as)
Temperatura (ºC)
7,0 7,5 8,0 8,5 9,0 9,5 10,0 10,5
C
Figura 11 - Superfície de resposta e gráficos de contorno para contagem de células viáveis (log UFC.mL-1) de Lactobacillus curvatus P99. A: efeito do pH e temperatura para uma incubação de 12h; B: efeito do pH e tempo de incubação a uma temperatura de 30°C; C: efeito da temperatura e tempo de incubação para o pH 6,5.

95
Os efeitos das variáveis independentes na atividade antimicrobiana
estão apresentados na figura 12 (A, B e C). A atividade antimicrobiana foi
significativamente afetada pelo pH, temperatura e tempo de incubação. A
análise rotacional canônica indicou que o ponto estacionário foi um ponto de
sela. De acordo com a otimização matemática, as melhores condições foram:
pH 6,22, uma temperatura de 30,6°C e tempo de incubação de 17,9h. Nestas
condições, a atividade antimicrobiana foi de 79.686 UA.mL-1.
As variáveis de multiplicação celular e atividade antimicrobiana (r = 0,79;
p < 0,0001) evidenciaram elevado coeficiente de correlação positivo.
Representando que quando ocorre aumento na contagem de células viáveis,
igualmente é verificado acréscimo na atividade antimicrobiana.
Com os resultados obtidos podemos perceber que a produção de
substâncias antimicrobianas (bacteriocin-like) pelo isolado L. curvatus P99
seguiu uma cinética de metabolismo primário, ou seja, a produção de
metabólito ocorreu durante a fase logarítmica de crescimento microbiano (SIP
et al., 2012), chegando ao nível máximo no final desta fase (100.000 UA.mL-1).
Podemos observar ainda que a máxima multiplicação celular coincidiu com a
máxima produção de substâncias antimicrobianas quando o isolado P99 foi
submetido à temperatura de 30ºC e pH 6,5 (Figura 13). Os resultados obtidos
na otimização pela MSR diferem da máxima eficiência encontrada durante os
experimentos realizados, uma vez que a máxima multiplicação e produção de
substâncias antimicrobianas já ocorreu a partir de 8h de incubação.

96
0
20000
40000
60000
80000
100000
252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525
353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535
303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030
252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525
353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535
4,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,0
6,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,5
9,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,0Ativ
idad
e an
timic
robi
ana
(UA
mL-1
)
Tem
pera
tura
(ºC
)
pH
0 20000400006000080000
A
-20000
0
20000
40000
60000
80000
100000
120000
36
912
1518
2124
4,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,04,0
6,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,56,5
9,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,09,0Ativ
idad
e an
timic
robi
ana
(UA
mL-1
)
Tem
po d
e in
cuba
ção
(hor
as)
pH
-20000 0 20000 40000 60000 80000 100000 120000
B
-20000
0
20000
40000
60000
80000
100000
36
912
1518
2124
252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525
353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535
303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030303030
252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525252525
353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535353535Ativ
idad
e an
timic
robi
ana
(UA
mL-1
)
Tem
po d
e in
cuba
ção
(hor
as)
Temperatura (ºC)
-200000 20000400006000080000100000
C
Figura 12 - Superfície de resposta e gráficos de contorno para atividade antimicrobiana (UA.mL-
1) de Lactobacillus curvatus P99. A: efeito do pH e temperatura para uma incubação de 12h; B: efeito do pH e tempo de incubação a uma temperatura de 30°C; C: efeito da temperatura e tempo de incubação para o pH 6,5.

97
Figura 13 - Mudanças na contagem de células viáveis (log UFC.mL-1) e atividade antimicrobiana (UA.mL-1) durante o crescimento do isolado Lactobacillus curvatus P99 a 30ºC durante 24h. Resultados da análise de variância com índice de confiança de 95%. Cada ponto representa a média de 3 repetições testados em duplicata.
Uma alta correlação entre a produção de biomassa e de substâncias
antimicrobianas (bacteriocin-like) foram também descritas para espécies de L.
sakei (MALHEIROS et al., 2015), Brevibacterium linens (MOTTA; BRANDELLI,
2003) e L. casei LA-1 (KUMAR et al., 2012). Em estudo realizado por
Mataragas et al. (2003), foi observado que a linhagem de L. curvatus L442
sofreu influência do pH e temperatura na produção de bacteriocinas, porém
estes parâmetros não influenciaram, consideravelmente, a multiplicação
celular. Os resultados mostraram que a diminuição do pH e temperatura abaixo
do ótimo para a multiplicação celular aumentaram a produção de bacteriocina,
devido a melhor utilização de energia e metabólitos essenciais. Os autores
observaram ainda que, o pH ótimo para multiplicação celular foi pH entre 6,0 –

98
6,5 e temperatura entre 25 – 30°C e a máxima produção de bacteriocina (1.280
UA.mL-1) foi em pH 5,5, a temperatura de 25°C em 16h.
Messens et al. (2003) também observaram influência das diferentes
condições de pH e temperatura na multiplicação celular e atividade
antimicrobiana de L. curvatus LTH 1174. O valor ótimo de temperatura para
multiplicação celular foi 34,5°C e não correspondeu ao ótimo de produção de
curvacina A que foi a 25°C (3.010 UA.mL-1). Os autores observaram ainda que,
tanto a produção de bacteriocina, quanto a multiplicação celular, ocorreram a
uma ampla faixa de pH.
Malheiros et al. (2015) observaram que um isolado de L. sakei também
apresentou cinética de metabolismo primário na produção de bacteriocina. A
curvacina A, produzida por L. curvatus LTH 1174, segue uma cinética de
metabolismo primário de acordo com o estudo realizado por Messens et al.
(2003). Comparando os trabalhos citados anteriormente ao presente estudo,
percebe-se que o isolado de L. curvatus P99 teve uma atividade antimicrobiana
superior em relação as outras bacteriocinas, estudadas por aqueles
pesquisadores.
Portanto, estes parâmetros devem ser avaliados para cada isolado, uma
vez que, as condições ótimas de multiplicação celular e produção de
metabólitos são cepa dependentes (MALHEIROS et al., 2015).
5.4 Caracterização do sobrenadante livre de células contendo substâncias
antimicrobianas de origem proteica (bacteriocin-like)
5.4.1 Atividade antimicrobiana: Concentração inibitório mínima (CBM) e
concentração bactericida mínima (CBM)
A menor concentração do SLC do isolado P99 necessária para promover
efeito bacteriostático (CIM) sobre L. monocytogenes Scott A foi de 6.400
UA.mL-1 (15,6 µL.mL-1), enquanto a ação bactericida (CBM) contra o mesmo
micro-organismo, foi de 1.600 UA.mL-1 (62,5 µL.mL-1). No apêndice F, pode-se
observar uma imagem da quantificação da CIM do SLC.

99
Anacarso et al. (2011) avaliaram a CIM do SLC produzido por E.
casseliflavus IM 416K1, contendo enterocina 416K1 e, observaram que, esta foi
de 2.560 UA.mL-1 (39 µL.mL-1). Pode-se perceber que o SLC produzido pela
linhagem E. casseliflavis IM 416K1, teve uma atividade antimicrobiana inferior a
atividade antimicrobiana no SLC, produzido pelo isolado P99, em estudo.
5.4.1 Atividade antimicrobiana
Pode-se observar que o SLC, produzido por L. curvatus P99, apresentou
atividade antimicrobiana contra diferentes espécies de L. monocytogenes e E.
coli O157:H7 (Tabela 12).
Tabela 12 - Linhagens indicadoras e sua sensibilidade para o sobrenadante livre de células contendo substância antimicrobiana (bacteriocin-like) produzido por Lactobacillus curvatus P99
Micro-organismo Origem Atividade antimicrobiana (UA.mL-1) Gram-positivas Listeria monocytogenes ATCC 7644 ATCC1 6.400 Listeria monocytogenes Scott A ATCC 6.400 Listeria monocytogenes Siliken TIAR2 400 Listeria ivanovii ATCC 19119 ATCC 12.800 Listeria innocua CLIP 12612 CLIP3 800 Staphylococcus aureus ATCC 25923 ATCC -4
Bacillus cereus ATCC 11778 ATCC -
Gram-negativas Escherichia coli O157:H7 ATCC 43895 ATCC 200 Escherichia coli ATCC 43895 ATCC - Pseudomonas aeruginosa ATCC 15442 ATCC - Salmonella Enteritidis ATCC 13076 ATCC - Salmonella Typhimurium ATCC 14028 ATCC - 1 ATCC: American Type Culture Collection 2 CLIP: Listeria Collection of Institute Pasteur 3 TIAR: Tasmanian Institute of Agricultural Research, University of Tasmania, Australia 4 -: ausência de atividade antimicrobiana.
Listeria monocytogenes é considerado um dos mais importantes
patógenos alimentares, sendo responsável por 1.600 casos de infecção em
humanos e aproximadamente 260 mortes/ano nos Estados Unidos (CDC,
2014). No Brasil, a listeriose é subdiagnosticada e subnotificada, portanto
poucos dados existem sobre este micro-organismo (FOOD SAFETY BRAZIL,
2013). Uma forma de controle desta bactéria é a utilização de bacteriocinas

100
provenientes da classe II, como por exemplo, as sakacinas, devido ao seu
poder de inibição, principalmente contra Listeria spp., como demonstrado no
presente estudo.
Ahmadova et al. (2013a) testaram o SLC produzido por um isolado de L.
curvatus, produtor de curvacina A, e não observaram atividade contra
Staphylococcus aureus nem contra as bactérias Gram-negativas testadas (E.
coli e Salmonella spp.). Porém, os autores obtiveram resultados positivos para
espécies de B. cereus, L. inoccua e L. monocytogenes. Barbosa et al. (2015)
avalariam o SLC de dois isolados de L. curvatus, MBSa2 e MBSa3, produtores
da sakacina P, quanto ao seu espectro de ação e observaram que estes foram
efetivos contra L. monocytogenes, porém não inibiram B. cereus, S. aureus, E.
coli e Salmonella spp.
Casaburi et al. (2016) quantificaram a atividade antimicrobiana do SLC
de um isolado de L. curvatus 54M16, produtor das sakacinas X, T e P contra
micro-organismos Gram-positivos e Gram-negativos. Os autores observaram
atividade contra L. monocytogenes 7644 (3.200 UA.mL-1), L. monocytogenes
Scott A (12.800 UA.mL-1), L. inoccua (800 UA.mL-1) e B. cereus (400 - 800
UA.mL-1), porém linhagens de S. aureus e E. coli O157:H7 não foram inibidas
pelo SLC de L. curvatus 54M16.
No presente estudo, o isolado P99 não foi capaz de inibir S. aureus.
Zygmunt; Browder e Tavormina (1967) sugerem que a presença de pontes
cruzadas de peptidioglicano com pentaglicina, na parede celular de S. aureus,
podem torná-los resistentes a enzimas. Ainda, algumas cepas de S. aureus são
produtoras de bacteriocinas da classe II. Linhagens produtoras são protegidas
contra os efeitos dos suas bacteriocinas por sistemas de imunidade cognatos,
cuja expressão está invariavelmente associada a produção de bacteriocina
(COELHO et al., 2014). Portanto, supõe-se que a inatividade do SLC do isolado
P99 contra S. aureus pode ser relacionada a estes fatores.
A atividade antimicrobiana das bacteriocinas é basicamente limitada a
bactérias Gram-positivas. Não é usual a inibição de bactérias Gram-negativas
por bacteriocinas produzidas por BAL. Isto pode ser explicado pela diferença
na composição das paredes celulares entre bactérias Gram-positivas e Gram-
negativas (ABEE; KROCKEL; HILL, 1995). Um dos principais problemas para a

101
aplicação de bacteriocinas contra as bactérias Gram-negativas é o efeito
protetor atribuído à membrana externa bacteriana.
Alguns estudos demonstram que, o uso combinado de bacteriocinas
com agentes quelantes ou ácidos, provocam a permeabilização da membrana
externa, fazendo com que a bacteriocina seja ativa contra bactérias Gram-
negativas (ANANOU et al., 2005b; MOLINOS et al., 2009). Porém, alguns
estudos já demonstraram a eficácia de bacteriocinas contra bactérias Gram-
negativas (KWON et al., 2002; LAUKOVÁ et al., 1998; LINE et al., 2008).
A habilidade do isolado P99 em inibir L. monocytogenes e outros
patógenos de interesse alimentar, como E. coli O157:H7, sugere o uso deste
SLC como uma ferramenta para o controle da contaminação de alimentos.
5.4.2 Estabilidade ao tratamento térmico, refrigeração, congelamento, pH
e agentes químicos
5.4.2.1 Estabilidade ao tratamento térmico
O SLC do cultivo de L. curvatus P99 manteve sua atividade
antimicrobiana estável até 90ºC (100%). A atividade residual aos 100ºC, 121ºC
por 10 minutos, 121ºC por 20 minutos foi de 96,43%, 89,29% e 82,14%,
respectivamente. A estabilidade ao tratamento térmico é uma característica das
bacteriocinas provenientes da classe II (COTTER; HILL; ROSS, 2005).
Outros autores também demonstraram termoestabilidade para o SLC de
isolados de L. curvatus. Lactobacillus curvatus A61 (AHMADOVA et al., 2013)
manteve a sua atividade antimicrobiana em temperaturas de 100 e 121ºC. Já L.
curvatus, identificado como isolado 5, manteve sua atividade total quando
submetido as temperaturas de 60°C (60 minutos) e 100 °C (30 minutos),
enquanto L. curvatus, identificado como isolado 14, teve sua atividade reduzida
a 50%, após 15 minutos a 60 e 100°C (MARTINIS; SANTAROSA; FREITAS,
2003).

102
5.4.2.2 Estabilidade a refrigeração e ao congelamento
Os resultados para a estabilidade da atividade antimicrobiana em
refrigeração e ao congelamento estão apresentados na tabela 13.
Tabela 13 - Estabilidade a refrigeração e ao congelamento do sobrenadante livre de células contendo substâncias antimicrobianas (bacteriocin-like) produzido por Lactobacillus curvatus P99 (% de atividade em relação ao controle sem tratamento)
Tempo/Temperatura Refrigeração (4 °C)
% Congelamento (- 18°C)
% 1 semana 100 100 2 semanas 100 100 3 semanas 92,86 96,43 4 semanas 92,86 92,86 5 semanas 92,86 92,86 6 semanas 89,29 89,29 7 semanas 89,29 89,29 8 semanas 85,71 89,29 24 semanas 82,14 89,29 36 semanas 82,14 82,14 48 semanas 42,86 53,57
O SLC do isolado L. curvatus L442 produtor da curvaticina L442
manteve a atividade final em 60% a 4 e – 20°C, após 4 semanas de
armazenamento (XIRAPHI et al., 2006). Xiraphi et al. (2008) observaram que o
SLC proveniente de um isolado de L. mesenteroides teve uma redução de 60%
na sua atividade após seis semanas de armazenamento a 4ºC e, quando este
SLC foi submetido ao congelamento a – 20ºC pelo mesmo período, a redução
da atividade antimicrobiana foi apenas de 20%.
Podemos observar que o SLC do isolado P99 manteve uma atividade
antimicrobiana final acima 80% após 36 semanas de armazenamento
congelado e refrigerado. Estes resultados foram superiores aos encontrados
pelos pesquisadores descritos acima, uma vez que os SLC tiveram uma
redução maior na atividade antimicrobiana quando armazenados sob
refrigeração e congelamento e ainda, mantiveram sua estabilidade por um
menor período de tempo. A estabilidade ao armazenamento faz com que, o
SLC produzido pelo isolado P99, seja uma das características que o tornam
bom candidato a biopreservante.

103
5.4.2.3 Estabilidade frente a variações de pH
A atividade antimicrobiana final do SLC do isolado P99 foi de 92,8%
submetido aos pH 7 e 8. Nos pH 2, 3, 4, 5, 6 e 9, foi mantida uma atividade de
89,3% e em pH 10 de 85,7%. A estabilidade a baixos pH é muito importante
para que as bacteriocinas possam ser aplicadas em alimentos, principalmente
aqueles fermentados, nos quais prevalecem as condições ácidas e isto foi
observado neste estudo. Ahmadova et al. (2013) e Martinis; Santarosa e
Freitas (2003) também observaram estabilidade a baixos pH quando estes
testaram a atividade do SLC provenientes de linhagens de L. curvatus.
5.4.2.4 Estabilidade aos agentes químicos
Observando os resultados de estabilidade da atividade antimicrobiana
do SLC produzido pelo isolado P99, na presença de agentes químicos, foi
possível verificar que a atividade antimicrobiana se manteve estável frente a
Tween 80, Triton X-100 e Triton X-114. Percebe-se que a atividade
antimicrobiana final, após tratamento com SDS, NaCl e ureia reduziu 10% e
quando tratado com Tween 20, reduziu 5% em relação ao controle. Resultados
similares foram relatados por Ahmadova et al. (2013a) para a atividade do SLC
produzido por L. curvatus A61 (curvacina A), por Todorov e Dicks (2004) para
SLC contendo a bacteriocina ST99, produzida por L. mesenteroides subsp.
dextranicum ST99 e por Favaro et al. (2014) para atividade dos SLC
produzidos por quatro isolados de E. faecium, ST209GB (enterocina A, P e B),
ST278GB (enterocina A e B), ST315GB (enterocina A, P e B) e ST711GB (A, P
e L50B).
A estabilidade a diferentes agentes químicos é de grande importância
sob o ponto de vista tecnológico e laboratorial, pois estes compostos podem
ser utilizados nas etapas de purificação da bacteriocina, em técnicas de
biologia molecular, em práticas bioquímicas, ou mesmo aplicados
concomitantemente a substância antimicrobiana, como por exemplo, o NaCl,
que é utilizado no processamento de alimentos (JERONYMO-CENEVIVA et al.
2014).

104
A estabilidade ao tratamento térmico e a ampla faixa de pH,
especialmente ao pH ácido e a temperatura de 121ºC, demonstram a potencial
aplicação de substâncias antimicrobianas com essas características, como
biopreservantes em alimentos fermentados (JERONYMO-CENEVIVA et al.
2014) ou em alimentos submetidos a tratamento térmico (BARBOSA et al.,
2015).
5.4.3 Avaliação da toxicidade da substância antimicrobiana de origem
proteica em modelo de Drosophila melanogaster
As substâncias antimicrobianas de origem proteica no SLC do isolado
P99, quando administrado até uma concentração de 40.000 UA.mL-1 não
apresentam efeito tóxico sobre células eucarióticas. As moscas adultas
expostas a concentração de 80.000 UA.mL-1 de SLC apresentaram mortalidade
significativa apenas no sétimo dia de exposição ao SLC do isolado P99 (Figura
14). Nos demais dias do período de administração, não houve diferença
significativa do controle.
Uma vez que não são permitidos testes toxicológicos em humanos,
devido a razões éticas, outros organismos vêm sendo utilizados para isto. D.
melanogaster, vulgarmente conhecida como ''mosca da fruta'' é um exemplo de
organismo eucariótico utilizado como modelo para o estudo de genética e
biologia celular, pois os processos fisiológicos importantes são bem
conservados da mosca para humanos. Além disso, um ciclo curto de vida,
diferentes estágios de desenvolvimento, o cultivo fácil, prole numerosa e a
possibilidade de induzir mutações fazem da Drosophila um organismo modelo
para estudar muitos processos biológicos, incluindo os testes de toxicidade
(PANACEK et al., 2011).

105
Tratamento (UA.mL-1)
1
So
bre
viv
ên
cia
(%
)
0
20
40
60
80
Controle 10.000 20.000 40.000 40.000: 60 80.000
Figura 14 - Efeito de diferentes concentrações de sobrenadante livre de células contendo
substância antimicrobiana (bacteriocin-like) produzido por Lactobacillus curvatus P99 sobre a
mortalidade de Drosophila melanogaster após 7 dias de tratamento.
Gupta; Sarkar e Srivastava (2014) avaliaram a toxicidade de uma
mistura de quatro peptídeos antimicrobianos produzidos por L. plantarum LR/14
utilizando a D. melanogaster como modelo e verificaram que concentrações
abaixo de 10 mg.mL-1 não apresentaram toxicidade. Porém, valores superiores
a 10 mg.mL-1 exerceram efeito tóxico, os quais foram dependentes da dose
aplicada, promovendo atraso no desenvolvimento das larvas e pupas e
afetando também a capacidade reprodutiva das moscas adultas.
Sehn (2015) avaliou o efeito de diferentes concentrações do SLC (10,
20, 500, 1.000, 2.000 e 10.000 UA.mL-1) produzido pelo isolado L. curvatus
LC254, contendo sakacina P, em Drosophilas adultas e observou que não
houve mortalidade significativa destas (p < 0,05) após 7 dias de tratamento.
Sugeriu ainda, que o resultado obtido indica que a sakacina P não apresentou
efeito tóxico sobre células eucarióticas. No presente estudo, foram avaliadas
concentrações maiores que as avaliadas por Sehn (2015), inferindo-se que, na

106
aplicação de concentrações menores do SLC, o efeito tóxico não seria
significativo.
Cabe ressaltar que, apesar dessas substâncias antimicrobianas serem
produzidas por BAL, as quais apresentam status GRAS, ensaios clínicos
adequados são necessários para confirmar a segurança destas substâncias
para utilização em alimentos.
5.5 Caracterização dos filmes biodegradáveis incorporados de SLC,
contendo substâncias antimicrobianas de origem proteica, produzido
pelo isolado L. curvatus P99
5.5.1 Quantificação e manutenção da atividade antimicrobiana
Tanto os filmes biodegradáveis de amido elaborados com a CIM do SLC
quanto os elaborados com a CBM apresentaram ação contra L.
monocytogenes Scott A e mantiveram a atividade durante os 30 dias
analisados (Apêndice G). Para cumprir o objetivo de proteger e expandir a vida
útil dos alimentos, as embalagens antimicrobianas devem manter sua atividade
ao longo de um período de tempo definido, de acordo com o prazo de validade
dos alimentos embalados. Massani et al. (2014b) também observaram a
manutenção da atividade antimicrobiana de filme a base de gluten de trigo
incorporado de bacteriocinas produzidas por L. curvatus CRL705 durante 50
dias de estocagem.b
5.5.2 Modo de ação
Ao analisar o efeito in vitro dos filmes incorporados com as CIM e CBM
do SLC de L. curvatus P99 sobre L. monocytogenes Scott A ao longo de 48h
de incubação, verificou-se diferença significativa (p < 0,05) entre os todos os
tratamentos, nos tempos de 3 a 48h de incubação, como apresentado na figura
15.
No tratamento controle, o patógeno praticamente duplicou a população
inicial ao final do experimento (97,4% de multiplicação). No filme com a CIM, o

107
desenvolvimento de L. monocytogenes Scott A foi expressivamente reduzido, o
que representou uma contagem 18,6% inferior que a contagem final do
tratamento controle. O filme contendo a CBM promoveu uma desaceleração
ainda mais acentuada na multiplicação do patógeno em relação ao tratamento
contendo a CIM. Neste tratamento, a contagem celular foi 28,7% inferior ao
controle (2,83 log). Quando comparado a CIM houve uma redução de 1 log,
representando 12,5% na contagem final. Provavelmente, a proporção filme:
suspensão de Listeria utilizada na análise, não foi adequada, pois a partir de 3h
houve multiplicação em escala logarítmica do patógeno. Novos testes, em
diferentes proporções devem ser realizados para verificar o efeito
bacteriostático e bactericida dos filmes antimicrobianos.
Tempo (Horas)
0 10 20 30 40 50
Con
tage
m d
e c
élu
las v
iáve
is (
log U
FC
.mL
-1)
0
2
4
6
8
10
Controle CIM CBM
Figura 15 - Modo de ação dos filmes incorporados com a concentração inibitória mínima (CIM) e a concentração bactericida mínima (CBM) do sobrenadante livre de células contendo substâncias antimicrobianas (bacteriocin-like) produzidas pelo isolado Lactobacillus curvatus P99.
Listeria monocytogenes é um patógeno capaz de crescer em
temperatura de refrigeração (COLE; JONES; HOLYOAK, 1990). O uso de

108
agentes antimicrobianos (SLC e bacteriocinas purificadas) adicionados
diretamente ou através de embalagens ativas provaram ser eficazes contra a
contaminação superficial de queijos (DAL BELLO et al., 2012; ISEPPI et al.,
2008; KALLINTERI; KOSTOULA; SAVVAIDIS, 2013).
Considerando que a embalagem isola o alimento de agentes externos,
possíveis contaminações microbiológicas do produto acabado são inerentes de
etapas anteriores. Sendo assim, é de interesse que as embalagens
antimicrobianas tenham rápida ação, impedindo e/ou retardando a
multiplicação celular bacteriana durante o armazenamento. Neste contexto, o
mecanismo de liberação, determinado pelas interações entre o material
polimérico que compõem a matriz e o agente antimicrobiano incorporado a ela
(HYLDGAARD et al., 2012), deve possibilitar uma rápida liberação.
No presente estudo, o ensaio de modo de ação do filme biodegradável
demonstrou que o amido de batata, utilizado na elaboração do filme, permitiu a
rápida liberação da substância antimicrobiana do SLC de L. curvatus P99,
promovendo a redução nas contagens de células viáveis até o tempo 3h nas
duas concentrações avaliadas.
5.5.3 Espectro de ação
Os filmes biodegradáveis elaborados com a CBM do SLC apresentaram
atividade antimicrobiana contra todas as cepas de referência testadas. Os
filmes elaborados com a CIM do SLC não foram eficientes contra as culturas de
referência L. monocytogenes Siliken, L. inoccua CLIP 12612 e E. coli O157:H7
ATCC 43895, assim como os isolados L. monocytogenes L6P4, L101P5,
L137P2, L142Q2, L148Q5, L149Q2 e L167Q5.
Massani et al. (2014a) testaram a atividade de filme sintético e
biodegrável incorporados de bacteriocinas e comprovaram seu efeito
antimicrobiano contra L. inoccua 7. Iseppi et al. (2008) e Marcos et al. (2007)
demonstraram efeito antimicrobiano contra L. monocytogenes de filme
contendo enterocina, para uso como embalagem biodegradável em alimentos.
Outros estudos demonstraram que a combinação de bacteriocinas com
outros agentes antimicrobianos em filmes promoveram a ação antimicrobiana

109
contra outros patógenos que não do gênero Listeria, como por exemplo, S.
aureus, S. Typhimurium e S. Enteretidis (JOFRÉ; AYMERICH; GARRIGA,
2008; PATTANAYAIYING; H-KITTIKUN; CUTTER, 2015).
5.6 Aplicação dos filmes biodegradáveis incorporados de SLC, contendo
substâncias antimicrobianas de origem proteica, produzido pelo isolado
L. curvatus P99 em queijo Prato
Foi observado que a multiplicação celular de L. monocytogenes Scott A
variou em função da concentração do SLC utilizado. Os resultados
demostraram que os filmes contendo a CBM do SLC foram capazes de reduzir
a contagem do patógeno em fatias de queijo Prato. Durante os dez dias de
armazenamento refrigerado, a enumeração do micro-organismo permaneceu
abaixo do limite de detecção da técnica (2,7 log UFC.g-1). Nos filmes controle
(sem SLC), pode-se perceber um comportamento típico da multiplicação de L.
monocytogenes uma vez que este patógeno é capaz de se desenvolver a
temperatura de refrigeração (4°C), atingindo contagens superiores a 4,3 log
UFC.g-1 em 10 dias de armazenamento. A CIM do SLC não demonstrou ser
eficiente no controle do patógeno, já que não apresentou diferença estatística
(p ≤ 0,05) quando comparado ao controle (Figura 16).

110
Tempo (Dias)
Inóculo 0 2 4 6 8 10
Co
nta
ge
m d
e c
élu
las v
iáve
is (
log
UF
C.m
L-1
)
0
1
2
3
4
5
ControleCIMCBM
Figura 16 - Contagem de Listeria monocytogenes Scott A em queijo Prato fatiado adicionado de filme biodegradável incorporado com sobrenadante livre de células contendo substâncias antimicrobianas (bacteriocin-like) produzidas por Lactobacillus curvatus P99 na concentração inibitória mínima (CIM) e concentração bactericida mínima (CBM) durante dez dias de armazenamento a 4ºC.
No ensaio in situ, as contagens de L. monocytogenes Scott A, nas fatias
de queijo, foram reduzidas abaixo do limite de detecção da técnica na presença
dos filmes com a CBM do SLC de L. curvatus P99 e, o filme com a CIM do SLC
não diferiu significativamente (p ≤ 0,05) do controle. Este resultado corrobora
com o encontrado por Marcos et al. (2007) ao avaliarem a utilização de filmes
biodegradáveis de alginato, zeína e álcool polivinil contendo enterocinas em
diferentes concentrações no controle de L. monocytogenes em fatias de
presunto cozido. A maior concentração de enterocina utilizada pelos autores
apresentou resultado mais eficiente.
Produtos prontos para o consumo são um desafio para a seguridade
alimentar, uma vez que, devido a manipulação pós-processamento e o contato
com superfícies, utensílios e equipamentos contaminados, é possível a
incidência de patógenos alimentares. Queijos fatiados tem vida útil de

111
aproximadamente 7 dias, após este período ele pode vir a se tornar inaceitável
para os consumidores. A utilização de filmes antimicrobianos pode ajudar, não
só no controle de micro-organismos patogênicos, como também a aumentar a
vida útil destes alimentos pelo controle de micro-organismos deteriorantes,
como já foi demonstrado no estudo realizado por Massani et al. (2014b).
Os resultados apresentados neste estudo confirmam o potencial
antimicrobiano do filme adicionado da CBM de SLC contendo substâncias
(bacteriocin-like) produzidas por L. curvatus P99 no controle de L.
monocytogenes, aumentando assim a qualidade e segurança em alimentos
prontos para o consumo.

112
6 Conclusões
No trabalho realizado, pode-se verificar que:
O isolado L. curvatus P99 foi tolerante ao trânsito gastrointestinal
simulado quando protegido por alimento, foi capaz de se autoagregar e
de se coagregar com L. monocytogenes, não apresentou capacidade de
adesão a células intestinais HCT-116, apresentou atividade
antimicrobiana contra bactérias Gram-positivas e Gram-negativas, não
apresentou atividade da enzima β-galactosidase, foi capaz de se
multiplicar a 10 e 45°C e em 4,5 e 6% de NaCl, não produziu gás a partir
da glicose, foi capaz de hidrolisar caseína, não apresentou atividade
lipolítica, genes para resistência a antimicrobianos de uso clínico e
descarboxilação de aminoácidos e nem atividade da enzima β-
hemolisina;
A partir da otimização das condições de multiplicação e produção de
substâncias antimicrobianas (bacteriocin- like), obteve-se como condição
ótima de multiplicação pH 6,3, temperatura de 29,3°C em 18,6h de
incubação e a condição ótima de produção de substâncias
antimicrobianas foi pH 6,22, temperatura de 30,6°C no tempo de 17,9h.
A produção de substâncias antimicrobianas seguiu uma cinética de
metabolismo primário;
A CIM do SLC foi de 6.400 UA.mL-1 e a CBM foi 1.600 UA.mL-1. O SLC
teve ação antimicrobiana contra bactérias Gram-positivas e Gram-
negativas, manteve-se estável ao tratamento térmico, diferentes pH e
agentes químicos, ao congelamento e a refrigeração por 48 semanas e

113
não foi tóxico nas concentrações de 10.000, 20.000 e 40.000 UA.mL-1
utilizando Drosophila melanogaster como modelo;
Os filmes produzidos com a CBM do SLC teve atividade antimicrobiana
contra todas as linhagens de referência testadas, mantiveram sua
atividade durante 30 dias e tiveram ação in situ contra L.
monocytogenes.
O isolado L. curvatus P99 apresentou características probióticas,
tecnológicas e aspectos de segurança compatíveis para utilização como cultura
coadjuvante de tecnologia em alimentos. Os dados de otimização da
multiplicação celular e produção de substâncias antimicrobianas forneceram
informações básicas para produção deste micro-organismo e seus metabólitos
em escala industrial. Filmes biodegradáveis, incorporados de SLC contendo
substâncias antimicrobianas, podem ser aplicados como agentes
antimicrobianos na biopreservação de queijo Prato, quando armazenado em
refrigeração.

114
7 Perspectivas futuras
Mais estudos são necessários para determinar o potencial probiótico de L.
curvatus P99, utilizando critérios de seleção in vivo.
Em relação às substâncias antimicrobianas, produzidas pelo isolado L.
curvatus P99, a purificação e caracterização das diferentes bacteriocinas
produzidas pelo isolado L. curvatus P99, são necessárias, bem como aplicação
das mesmas in situ.

115
Referências
ABEE, T.; KROCKEL, L.; HILL, C. Bacteriocins: modes of action and potentials in food preservation and control of food poisoning. International Journal of Food Microbiology, v. 28, p. 169–185, 1995. ACUÑA, L.; MORERO, R. D.; BELLOMIO, A. Development of Wide-Spectrum Hybrid Bacteriocins for Food Biopreservation. Food and Bioprocess Technology, v. 4, p. 1029–1049, 2011. AHMADOVA, A. et al. Antimicrobial and antifungal activities of Lactobacillus curvatus strain isolated from homemade Azerbaijani cheese. Anaerobe, v. 20, p. 42-49 2013. AHMADOVA, A. et al. Evaluation of antimicrobial activity, probiotic properties and safety of wild strain Enterococcus faecium AQ71 isolated from Azerbaijani Motal cheese. Food Control, v. 30, p. 631–641, 2013. AHN, H. Y. et al. Supplementation with two probiotic strains, Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032, reduces fasting triglycerides and enhances apolipoprotein A-V levels in non-diabetic subjects with hypertriglyceridemia. Atherosclerosis, v. 241, n. 2, p. 649–656, 2015. AHN, H. Y. et al. The triglyceride-lowering effect of supplementation with dual probiotic strains, Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032: Reduction of fasting plasma lysophosphatidylcholines in nondiabetic and hypertriglyceridemic subjects. Nutrition, Metabolism and Cardiovascular Diseases, v. 25, n. 8, p. 724–733, 2015. AMERICAN PUBLIC HEALTH ASSOCIATION - APHA. Compendium of methods for the microbiological examination of foods. 4. ed. Washington: [s.n.], 2002.

116
AMMOR, M. S.; MAYO, B. Selection criteria for lactic acid bacteria to be used as functional starter cultures in dry sausage production: An update. Meat Science, v. 76, p. 138–146, 2007. ANACARSO, I. et al. Anti-listerial activity of chitosan and Enterocin 416K1 in artificially contaminated RTE products. Food Control, v. 22, n. 12, p. 2076–2080, 2011. ANANOU, S. et al. Control of Listeria monocytogenes in model sausages by enterocin AS-48. International Journal of Food Microbiology, v. 103, p. 179–190, 2005. ANANOU, S. et al. Synergistic effect of enterocin AS-48 in combination with outer membrane permeabilizing treatments against Escherichia coli O157:H7. Journal of Applied Microbiology, v. 99, p. 1364–1372, 2005. ASHRAF, R.; SHAH, N. P. Antibiotic resistance of probiotic organisms and safety of probiotic dairy products. International Food Research Journal, v. 18, n. 3, p. 837–853, 2011. AXELSSON, L. Lactic acid bacteria: classification and physiology. Lactic acid bacteria: microbiology and functional aspects. 2. ed. New York: Marcel Dekker, 1998. p. 1–72. AXELSSON, L.; HOLCK, A. The genes involved in production of and immunity to sakacin A, a bacteriocin from Lactobacillus sake Lb706. Journal of Bacteriology, v. 177, n. 8, p. 2125–2137, 1995. AYENI, F. A. et al. Evaluation of the functional potential of Weissella and Lactobacillus isolates obtained from Nigerian traditional fermented foods and cow’s intestine. International Journal of Food Microbiology, v. 147, n. 2, p. 97–104, 2011. AYMERICH, M. et al. Bacteriocin-producing lactobacilli in Spanish-style fermented sausages: characterization of bacteriocins. Food Microbiology, v. 17, n. 1, p. 33–45, 2000. AYMERICH, T. et al. Safety properties and molecular strain typing of lactic acid bacteria from slightly fermented sausages. Journal of Applied Microbiology, v. 100, n. 1, p. 40–49, 2006.

117
BAKA, A. M. et al. Effect of selected autochthonous starter cultures on processing and quality characteristics of Greek fermented sausages. LWT - Food Science and Technology, v. 44, n. 1, p. 54–61, 2011. BARBOSA, J.; GIBBS, P. A.; TEIXEIRA, P. Virulence factors among enterococci isolated from traditional fermented meat products produced in the North of Portugal. Food Control, v. 21, n. 5, p. 651–656, 2010. BARBOSA, M. S. et al. Improving safety of salami by application of bacteriocins produced by an autochthonous Lactobacillus curvatus isolate. Food Microbiology, v. 46, p. 254–62, 2015. BASTOS, M. D. C. D. F.; COUTINHO, B. G.; COELHO, M. L. V. Lysostaphin: A staphylococcal bacteriolysin with potential clinical applications. Pharmaceuticals, v. 3, p. 1139–1161, 2010. BELLO, B. D. et al. Microbial ecology of artisanal products from North West of Italy and antimicrobial activity of the autochthonous populations. LWT - Food Science and Technology, v. 43, n. 7, p. 1151–1159, 2010. BERTHIER, F.; EHRLICH, S. D. Genetic diversity within Lactobacillus sakei and Lactobacillus curvatus and design of PCR primers for its detection using randomly amplified polymorphic DNA. International Journal of Systematic Bacteriology, v. 49 Pt 3, n. 1 999, p. 997–1007, 1999. BERTSCH, D. et al. pDB2011, a 7.6kb multidrug resistance plasmid from Listeria innocua replicating in Gram-positive and Gram-negative hosts. Plasmid, v. 70, n. 2, p. 284–287, 2013. BESHKOVA, D.; FRENGOVA, G. Bacteriocins from lactic acid bacteria: Microorganisms of potential biotechnological importance for the dairy industry. Engineering in Life Sciences, v. 12, n. 4, p. 419–432, 2012. BISCOLA, V. et al. Isolation and characterization of a nisin-like bacteriocin produced by a Lactococcus lactis strain isolated from charqui, a Brazilian fermented, salted and dried meat product. Meat Science, v. 93, n. 3, p. 607–613, 2013. BISWAS, S. R. et al. Influence of growth conditions on the production of a bacteriocin, pediocin AcH, by Pediococcus acidilactici H. Applied and

118
Environmental Microbiology, v. 57, n. 4, p. 1265–1267, 1991. BOVER-CID, S.; HOLZAPFEL, W. H. Improved screening procedure for biogenic amine production by lactic acid bacteria. International Journal of Food Microbiology, v. 53, p. 33–41, 1999. BOX, G. E. P.; BEHNKEN, D. W. Some New Three Level Designs for the Study of Quantitative Variables. Technometrics, v. 2, p. 455-475, 1960. BRASIL. Ministério da Saúde. Portaria no 29 de 22 de janeiro de 1996. Aprova a extensão de uso da NISINA com a função de conservador para queijos pasteurizados no limite máximo de 12,5mg/kg. Brasília, DF, janeiro, 1996. BRASIL. Ministério da Saúde. Resolução RDC n°2 de 07 de janeiro, 2002. Regulamento Técnico de Substâncias Bioativas e Probióticos Isolados com Alegação de Propriedades Funcional e ou de Saúde. Brasil, DF, janeiro , 2002 BUCKENHÜSKES, H. J. Selection criteria for lactic acid bacteria to be used as cultures for various food commodities. FEMS Microbiology Reviews, v. 12, p. 253–272, 1993. CAGRI, A.; USTUNOL, Z.; RYSER, E. T. Antimicrobial, mechanical, and moisture barrier properties of low pH whey protein based edible films containing paminobenzoic or sorbic acids. Journal of Food Science, v. 66, n. 6, p. 865–870, 2001. CÁLIX-LARA, T. F. et al. Inhibition of Escherichia coli O157: H7 and Salmonella enterica on spinach and identification of antimicrobial substances produced by a commercial Lactic Acid Bacteria food safety intervention. Food Microbiology, v. 38, p. 192–200, 2014. CAPOZZI, V. et al. Isolation and characterization of tyramine-producing Enterococcus faecium strains from red wine. Food Microbiology, v. 28, n. 3, p. 434–439, 2011. CASABURI, A. et al. Proteolytic and lipolytic starter cultures and their effect on traditional fermented sausages ripening and sensory traits. Food Microbiology, v. 25, n. 2, p. 335–347, 2008.

119
CASABURI, A. et al. Technological properties and bacteriocins production by Lactobacillus curvatus 54M16 and its use as starter culture for fermented sausage manufacture. Food Control, v. 59, p. 31–45, 2016. CASTELLANO, P. et al. Protective action of Lactobacillus curvatus CRL705 on vacuum-packaged raw beef. Effect on sensory and structural characteristics. Meat Science, v. 85, n. 3, p. 394–401, 2010. CENTER FOR DISEASE CONTROL AND PREVENTION - CDC. Listeria (Listeriosis). 2014 Disponível em: <http://www.cdc.gov/listeria/statistics.html>. Acesso em: 25 de maior de 2015. CHARTERIS, W. P. et al. Ingredient selection criteria for probiotic microorganisms in functional dairy foods. International Journal of Dairy Technology, v. 51, n. 4, p. 123–136, 1998. CLINICAL AND LABORATORY STANDARDS INSTITUTE - CLSI, 2012. Performance Standards for Antimicrobial Disk Susceptibility Tests; Approved standard - Eleventh Edition. CLSI document M02-A11. Wayne, PA: Clinical and Laboratory Standards Institute. COCOLIN, L. et al. Description of the bacteriocins produced by two strains of Enterococcus faecium isolated from Italian goat milk. Food Microbiology, v. 24, p. 752–758, 2007. COCOLIN, L.; RANTSIOU, K. Sequencing and expression analysis of sakacin genes in Lactobacillus curvatus strains. Applied Microbiology and Biotechnology, v. 76, n. 6, p. 1403–1411, 2007. COELHO, M. L. V. et al. Immunity to the Staphylococcus aureus leaderless four-peptide bacteriocin aureocin A70 is conferred by AurI, an integral membrane protein. Research in Microbiology, v. 165, p. 50–59, 2014. COLE, M. B.; JONES, M. V; HOLYOAK, C. The effect of pH, salt concentration and temperature on the survival and growth of Listeria monocytogenes. The Journal of Applied Bacteriology, v. 69, p. 63–72, 1990. COLLADO, M. C.; MERILUOTO, J.; SALMINEN, S. Adhesion and aggregation properties of probiotic and pathogen strains. European Food Research and

120
Technology, v. 226, p. 1065–1073, 2008. COLLINS, C. H. et al. Microbiological Methods. 8 ed. New York: Arnold, 2004. CORBO, M. R. et al. Prolonging microbial shelf life of foods through the use of natural compounds and non-thermal approaches - A review. International Journal of Food Science and Technology, v. 44, p. 223–241, 2009. COTON, E.; COTON, M. Multiplex PCR for colony direct detection of Gram-positive histamine- and tyramine-producing bacteria. Journal of Microbiological Methods, v. 63, n. 3, p. 296–304, 2005. COTTER, P. D.; HILL, C.; ROSS, R. P. Bacteriocins: Developing innate immunity for food. Nature reviews, v. 3, p. 777–788, 2005. COUSIN, F. J. et al. Detection and Genomic Characterization of Motility in Lactobacillus curvatus: Confirmation of Motility in a Species outside the Lactobacillus salivarius Clade. Applied and Environmental Microbiology, v. 81, n. 4, p. 1297–1308, 2015. CRUXEN, C. E. S. Caracterização fenotípica, molecular e avaliação do potencial tecnológico de Estafilococos coagulase negativa isolados de queijos da região sudoeste do Rio Grande do Sul/Brasil. 2016. 76 f. Dissertação (Mestrado em Ciência e Tecnologia de Alimentos) - Faculdade de Agronomia Eliseu Maciel, Universidade Federal de Pelotas, Pelotas, 2016. DAL BELLO, B. et al. Technological characterization of bacteriocin producing Lactococcus lactis strains employed to control Listeria monocytogenes in Cottage cheese. International Journal of Food Microbiology, v. 153, n. 1-2, p. 58–65, 2012. DE MARTINIS, E. C. P.; FREITAS, F. Z. Screening of lactic acid bacteria from Brazilian meats for bacteriocin formation. Food Control, v. 14, p. 197–200, 2002. DE VUYST, L.; VANDAMME, E. J. Influence of the carbon source on nisin production in Lactococcus lactis subsp. lactis batch fermentations. Journal of General Microbiology, v. 138, p. 571–578, 1992.

121
DEEGAN, L. H. et al. Bacteriocins: Biological tools for bio-preservation and shelf-life extension. International Dairy Journal, v. 16, p. 1058–1071, 2006. DEL RE, B. et al. Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Letters in Applied Microbiology, v. 31, p. 438–442, 2000. DERRINGER, G. C. A Balancing Act : Optimizing a Product’s Properties. Quality Progress, v. 27, n. 6, p. 51–58, 1994. DERRINGER, G.; SUICH, R. Simultaneous optimization of several response variables. Journal of Quality Technology, v. 12, n. 4, p. 214–219, 1980. DIAO, W. R. et al. Chemical composition, antibacterial activity and mechanism of action of essential oil from seeds of fennel (Foeniculum vulgare Mill.). Food Control, v. 35, n. 1, p. 109–116, 2014. DORTU, C. et al. Anti-listerial activity of bacteriocin-producing Lactobacillus curvatus CWBI-B28 and Lactobacillus sakei CWBI-B1365 on raw beef and poultry meat. Letters in Applied Microbiology, v. 47, n. 6, p. 581–586, 2008. DRIDER, D. et al. The continuing story of class IIa bacteriocins. Microbiology and Molecular Biology Reviews, v. 70, n. 2, p. 564–582, 2006. DROSINOS, E. H. et al. Characterization of the microbial flora from a traditional Greek fermented sausage. Meat Science, v. 69, n. 2, p. 307–317, 2005. EATON, T. J.; GASSON, M. J. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Applied and Environmental Microbiology, v. 67, p. 1628–1635, 2001. European Food Safety Authority - EFSA. Opnion of the Scientific Committee on Animal Nutrition on the criteria for assessing the safety of micro-organisms resistant to antibiotics of human clinical and veterinary importance. The European Food Safety Authority Journal, v. 226, p. 1–16, 2003. ENNAHAR, S. et al. Class IIa bacteriocins: Biosynthesis, structure and activity. FEMS Microbiology Reviews, v. 24, p. 85–106, 2000.

122
ENTIAN, K. D.; DE VOS, W. M. Genetics of subtilin and nisin biosyntheses. Antonie van Leeuwenhoek, v. 69, n. 2, p. 109–117, 1996. ERCOLINI, D. et al. Changes in the spoilage-related microbiota of beef during refrigerated storage under different packaging conditions. Applied and Environmental Microbiology, v. 72, n. 7, p. 4663–4671, 2006. FADDA, S. et al. Hydrolysis of pork muscle sarcoplasmic proteins by Lactobacillus curvatus and Lactobacillus sake. Applied and Environmental Microbiology, v. 65, n. 2, p. 578–584, 1999. FADDA, S.; LÓPEZ, C.; VIGNOLO, G. Role of lactic acid bacteria during meat conditioning and fermentation: Peptides generated as sensorial and hygienic biomarkers. Meat Science, v. 86, n. 1, p. 66–79, 2010. Food and Agriculture Organization of the United Nations/World Health Organization - FAO/WHO, 2006. Probiotics in food. Health and nutritional properties and guidelines for evaluation. FAO Food and Nutrition Paper 85. FAO, Rome, Italy. FAVARO, L. et al. Bacteriocinogenic potential and safety evaluation of non-starter Enterococcus faecium strains isolated from home made white brine cheese. Food Microbiology, v. 38, p. 228–239, 2014. FIMLAND, G. et al. Pediocin-like antimicrobial peptides (class IIa bacteriocins) and their immunity proteins: Biosynthesis, structure, and mode of action. Journal of Peptide Science, v. 11, p. 688–696, 2005. FLEMING, H. P.; ETCHELLS, J. L.; COSTILOW, R. N. Microbial inhibition by an isolate of Pediococcus from cucumber brines. Applied microbiology, v. 30, n. 6, p. 1040–1042, 1975. FOOD SAFETY BRAZIL, 2013. Listeria e E. coli não atormentam os brasileiros? Disponível em: <http://foodsafetybrazil.com/listeria-e-e-coli-nao-atormentam-aos-brasileiros/>. Acesso em: 10 fev. 2015. FRANZ, C. M. A P. et al. Diversity of enterococcal bacteriocins and their grouping in a new classification scheme. FEMS Microbiology Reviews, v. 31, p. 293–310, 2007.

123
FRECH, G.; SCHWARZ, S. Molecular analysis of tetracycline resistance in Salmonella enterica subsp. enterica serovar Typhimurium, Enteritidis, Dublin, Choleraesuis, Hadar and Saintpaul: Construction and application of specific gene probes. Journal of Applied Microbiology, v. 89, n. 4, p. 633–641, 2000. GÁLVEZ, A. et al. Bacteriocin-based strategies for food biopreservation. International Journal of Food Microbiology, v. 120, p. 51–70, 2007. GARCÍA-RUIZ, A. et al. Assessment of probiotic properties in lactic acid bacteria isolated from wine. Food Microbiology, v. 44, p. 220–225, 2014. GARRIGA, M. et al. Potentially probiotic and bioprotective lactic acid bacteria starter cultures antagonise the Listeria monocytogenes adhesion to HT29 colonocyte-like cells. Beneficial Microbes, v. 6, n. 3, p. 337–343, 2015. GHEYTANCHI, E. et al. Study on b-galactosidase enzyme produced by isolated lactobacilli from milk and cheese. African Journal of Microbiology, v. 4(6), n. 6, p. 454–458, 2009. GIRAFFA, G. Functionality of enterococci in dairy products. International Journal of Food Microbiology, v. 88, n. 2-3, p. 215–222, 2003. GONZÁLEZ-VÁZQUEZ, R. et al. Isolation, Identification and Partial Characterization of a Lactobacillus casei Strain with Bile Salt Hydrolase Activity from Pulque. Probiotics and Antimicrobial Proteins, v. 7, n. 4, p. 242–248, 2015. GRAPE, M.; SUNDSTRÖM, L.; KRONVALL, G. Sulphonamide resistance gene sul3 found in Escherichia coli isolates from human sources. Journal of Antimicrobial Chemotherapy, v. 52, n. 6, p. 1022–1024, 2003. GUILLARD, V. et al. Food preservative content reduction by controlling sorbic acid release from a superficial coating. Innovative Food Science and Emerging Technologies, v. 10, n. 1, p. 108–115, 2009. HALAL, S. L. M. et al. Films based on oxidized starch and cellulose from barley. Carbohydrate Polymers, v. 133, p. 644-653, 2015. HAMMES, W. P.; HERTEL, C. The genera Lactobacillus and Carnobacterium.

124
The Prokariotes. 3. ed. New York: Springer-Verlag, 2006. p. 320–406. HARTMANN, H. A.; WILKE, T.; ERDMANN, R. Efficacy of bacteriocin-containing cell-free culture supernatants from lactic acid bacteria to control Listeria monocytogenes in food. International Journal of Food Microbiology, v. 146, n. 2, p. 192–199, 2011. HEBERT, E. M. et al. Genome Sequence of the bacteriocin-producing Lactobacillus curvatus strain CRL705. Journal of Bacteriology, v. 194, n. 2, p. 538–539, 2012. HÉCHARD, Y. et al. Characterization and purification of mesentericin Y105, an anti-Listeria bacteriocin from Leuconostoc mesenteroides. Journal of General Microbiology, v. 138, p. 2725–2731, 1992. HÉQUET, A. et al. Characterization of new bacteriocinogenic lactic acid bacteria isolated using a medium designed to simulate inhibition of Listeria by Lactobacillus sakei 2512 on meat. International Journal of Food Microbiology, v. 113, n. 1, p. 67–74, 2007. HERMANNS, G. et al. Evaluation of probiotic characteristics of lactic acid bacteria isolated from artisan cheese. Journal of Food Safety, v. 34, n. 4, p. 1–8, 2014. HERNÁNDEZ, D.; CARDELL, E.; ZÁRATE, V. Antimicrobial activity of lactic acid bacteria isolated from Tenerife cheese: Initial characterization of plantaricin TF711, a bacteriocin-like substance produced by Lactobacillus plantarum TF711. Journal of Applied Microbiology, v. 99, n. class I, p. 77–84, 2005. HERRANZ, C.; DRIESSEN, A. J. M. Sec-mediated secretion of bacteriocin enterocin P by Lactococcus lactis. Applied and Environmental Microbiology, v. 71, n. 4, p. 1959–1963, 2005. HERREROS, M. A. et al. Antimicrobial activity and antibiotic resistance of lactic acid bacteria isolated from Armada cheese (a Spanish goats’ milk cheese). Food Microbiology, v. 22, p. 455–459, 2005. HOLCK, A. et al. Purification and amino acid sequence of sakacin A, a bacteriocin from Lactobacillus sake Lb706. Journal of General Microbiology, v. 138, n. 12, p. 2715–2720, 1992.

125
HOLZAPFEL, W. H. et al. Taxonomy and importante features of probiotic microorganisms in food and nutrition. The American Journal of Clinical Nutrition, v. 73, p. 365S–73S, 2001. HUANG, E. et al. Characterization and application of enterocin RM6, a bacteriocin from Enterococcus faecalis. BioMed Research International, v. 2013, p. 1–7, 2013. HUANG, Y.; ADAMS, M. C. In vitro assessment of the upper gastrointestinal tolerance of potential probiotic dairy propionibacteria. International Journal of Food Microbiology, v. 91, p. 253–260, 2004. HUGAS, M. Bacteriocinogenic lactic acid bacteria for the biopreservation of meat and meat products. Meat Science, v. 49, n. 98, p. S139–S150, 1998. HUGAS, M.; GARRIGA, M.; AYMERICH, M. T. Functionalty of enterococci in meat products. International Journal of Food Microbiology, v. 88, p. 223–233, 2003. HÜHNE, K. et al. Analysis of the sakacin P gene cluster from Lactobacillus sake Lb674 and its expression in sakacin-negative Lb. sake strains. Microbiology, v. 142, n. 6, p. 1437–1448, 1996. HUMMEL, A. S. et al. Antibiotic resistances of starter and probiotic strains of lactic acid bacteria. Applied and Environmental Microbiology, v. 73, p. 730–739, 2007. HWANHLEM, N. et al. Bacteriocin-Producing Lactic Acid Bacteria Isolated from Mangrove Forests in Southern Thailand as Potential Bio-Control Agents: Purification and Characterization of Bacteriocin Produced by Lactococcus lactis subsp. lactis KT2W2L. Probiotics and Antimicrobial Proteins, v. 5, p. 264–278, 2013. HWANHLEM, N.; CHOBERT, J. M.; H-KITTIKUN, A. Bacteriocin-producing lactic acid bacteria isolated from mangrove forests in southern Thailand as potential bio-control agents in food: Isolation, screening and optimization. Food Control, v. 41, p. 202–211, 2014. HYLDGAARD, M. et al. Essential oils in food preservation : mode of action , synergies , and interactions with food matrix components. Frontiers in

126
Microbiology, v. 3, n. January, p. 1–24, 2012. ISEPPI, R. et al. Anti-listerial activity of a polymeric film coated with hybrid coatings doped with Enterocin 416K1 for use as bioactive food packaging. International Journal of Food Microbiology, v. 123, p. 281–287, 2008. ISEPPI, R. et al. Anti-listerial activity of coatings entrapping living bacteria. Soft Matter, v. 7, p. 8542-8548, 2011. JACK, R. W.; SAHL, H. G. Unique peptide modifications involved in the biosynthesis of lantibiotics. Trends in Biotechnology, v. 13, n. 7, p. 269–278, 1995. JACK, R. W.; TAGG, J. R.; RAY, B. Bacteriocins of gram-positive bacteria. Microbiological Reviews, v. 59, n. 2, p. 171–200, 1995. JAVED, A. et al. Enterocins of Enterococcus faecium, emerging natural food preservatives. Annals of Microbiology, v. 61, p. 699–708, 2011. JENSEN, L. B. Presence of erm gene classes in Gram-positive bacteria of animal and human origin in Denmark. FEMS Microbiology Letters, v. 31, n. 1, p. 60–158, 1999. JERONYMO-CENEVIVA, B. A. et al. Probiotic properties of lactic acid bacteria isolated from Water-Buffalo Mozzarella cheese. Probiotics and Antimicrobial Proteins, v. 6, n. 3, p. 141–156, 2014. JOFRÉ, A.; AYMERICH, T.; GARRIGA, M. Assessment of the effectiveness of antimicrobial packaging combined with high pressure to control Salmonella sp. in cooked ham. Food Control, v. 19, p. 634–638, 2008. JONES, E.; SALIN, V.; WILLIAMS, G. Nisin and the Market for Commercial Bacteriocins. Reports, n. July, 2005. KAISER, A. L.; MONTVILLE, T. J. Purification of the bacteriocin bavaricin MN and characterization of its mode of action against Listeria monocytogenes Scott A cells and lipid vesicles. Applied and Environmental Microbiology, v. 62, n. 12, p. 4529–4535, 1996.

127
KALLINTERI, L. D.; KOSTOULA, O. K.; SAVVAIDIS, I. N. Efficacy of nisin and/or natamycin to improve the shelf-life of Galotyri cheese. Food Microbiology, v. 36, n. 2, p. 176–181, 2013. KALMOKOFF, M. L. et al. Identification of a new plasmid-encoded sec-dependent bacteriocin produced by Listeria innocua 743. v. 67, n. 9, p. 4041–4047, 2001. KANDLER, O.; WEISS, N. Regular, nonsporing gram-positive rods. In: J. G. HOLT & P. H. A. SNEATH (Org.). . Bergey’s Manual of Systematic Bacteriology. 2. ed. Baltimore: Williams & Wilkins, 1986. p. 1208–1234. KAUR, G. et al. Antibacterial efficacy of nisin, pediocin 34 and enterocin FH99 against L. monocytogenes, E. faecium and E. faecalis and bacteriocin cross resistance and antibiotic susceptibility of their bacteriocin resistant variants. Journal of Food Science and Technology, v. 51, p. 233–244, 2014. KEHRENBERG, C.; SCHWARZ, S. Occurrence and linkage of genes coding for resistance to sulfonamides, streptomycin and chloramphenicol in bacteria of the genera Pasteurella and Mannheimia. FEMS Microbiology Letters, v. 205, n. 2, p. 283–290, 2001. KERRN, M. B. et al. Susceptibility of Danish Escherichia coli strains isolated from urinary tract infections and bacteraemia, and distribution of sul genes conferring sulphonamide resistance. The Journal of Antimicrobial Chemotherapy, v. 50, n. 4, p. 513–516, 2002. KHAN, I.; KANG, S. C. Probiotic potential of nutritionally improved Lactobacillus plantarum DGK-17 isolated from Kimchi – A traditional Korean fermented food. Food Control, v. 60, p. 88–94, 2016. KIKUVI, G. M. et al. Streptomycin and chloramphenicol resistance genes in Escherichia coli isolates from cattle, pigs, and chicken in Kenya. Microbial Drug Resistance, v. 13, n. 1, p. 62–68, 2007. KIM, M. J.; KIM, K. S. Tyramine production among lactic acid bacteria and other species isolated from kimchi. LWT - Food Science and Technology, v. 56, n. 2, p. 406–413, 2014. KLAENHAMMER, T. R. Bacteriocins of lactic acid bacteria. Biochimie, v. 70, n.

128
11171, p. 337–349, 1988. KLAENHAMMER, T. R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiology Reviews, v. 12, p. 39–85, 1993. KOTELNIKOVA, E. A; GELFAND, M. S. Bacteriocin production of bacteriocins by gram-positive bacteria and the mechanisms of transcriptional regulation. Genetika, v. 38, n. 6, p. 758–772, 2002. KOUKER, G.; JAEGER, K. E. Specific and sensitive plate assay for bacterial lipase. Applied and Environmental Microbiology, v. 53, n. 1, p. 211–213, 1987. KUMAR, A.; SCHWEIZER, H. Bacterial resistance to antibiotics: Active efflux and reduced uptake. Advanced Drug Delivery Reviews, v. 57, n. 10, p. 1486–1513, 2005. KUMAR, M. et al. Statistical optimization of physical parameters for enhanced bacteriocin production by L. casei. Biotechnology and Bioprocess Engineering, v. 17, n. 3, p. 606–616, 2012. KUMAR, B. V.; VIJAYENDRA, S. V. N.; REDDY, O. V. S. Trends in dairy and non-dairy probiotic products - a review. Journal of Food Science and Technology, v. 52, n. 10, p. 6112–6124, 2015. KWON, A. et al. Bacteriocin produced by Pediococcus sp. in kimchi and its characteristics. Journal of Microbiology and Biotechnology, v. 12, p. 96–105, 2002. LADERO, V. et al. Genetic and functional analysis of biogenic amine production capacity among starter and non-starter lactic acid bacteria isolated from artisanal cheeses. European Food Research and Technology, v. 241, n. 3, p. 377–383, 2015. LAI, A. Y.; TRAN, S.; SIMMONDS, R. S. Functional characterization of domains found within a lytic enzyme produced by Streptococcus equi subsp. zooepidemicus. FEMS Microbiology Letters, v. 215, p. 133–138, 2002. LATORRE-MORATALLA, M. L. et al. Amino acid availability as an influential

129
factor on the biogenic amine formation in dry fermented sausages. Food Control, v. 36, n. 1, p. 76–81, 2014. LAUKOVÁ, A. et al. Occurrence of bacteriocin production among environmental enterococci. Letters in Applied Microbiology, v. 27, n. 1992, p. 178–182, 1998. LEROY, F.; DE VUYST, L. Bacteriocin production by Enterococcus faecium RZS C5 is cell density limited and occurs in the very early growth phase. International Journal of Food Microbiology, v. 72, p. 155–164, 2002. LEROY, F.; DE VUYST, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends in Food Science and Technology, v. 15, p. 67–78, 2004. LEROY, F.; VERLUYTEN, J.; DE VUYST, L. Functional meat starter cultures for improved sausage fermentation. International Journal of Food Microbiology, v. 106, p. 270–285, 2006. LEWUS, C. B.; MONTVILLE, T. J. Detection of bacteriocins produced by lactic acid bacteria. Journal of Microbiological Methods, v. 13, p. 145–150, 1991. LI, Q.; SHERWOOD, J. S.; LOGUE, C. M. Antimicrobial resistance of Listeria spp. recovered from processed bison. Letters in Applied Microbiology, v. 44, n. 1, p. 86–91, 2007. LIASI, S. A. et al. Antimicrobial activity and antibiotic sensitivity of three isolates of lactic acid bacteria from fermented fish product, Budu. Malaysian Journal of Microbiology, v. 5, n. 1, p. 33–37, 2009. LIMA, C. D. L. C. et al. Bactérias do acido láctico e leveduras associadas com o queijo-de-minas artesanal produzido na região da Serra do Salitre, Minas Gerais. Arquivo Brasileiro de Medicina Veterinaria e Zootecnia, v. 61, p. 266–272, 2009. LINARES, D. M. et al. Biogenic amines in dairy products. Critical Reviews in Food Science and Nutrition, v. 51, n. July, p. 691–703, 2011. LINE, J. E. et al. Isolation and purification of enterocin E-760 with broad

130
antimicrobial activity against Gram-positive and Gram-negative bacteria. Antimicrobial Agents and Chemotherapy, v. 52, n. 3, p. 1094–1100, 2008. LIU, C. et al. Antibiotic resistance of probiotic strains of lactic acid bacteria isolated from marketed foods and drugs. Biomedical and environmental sciences : BES, v. 22, n. 5, p. 401–12, 2009. MACEDO, A. C.; TAVARES, T. G.; MALCATA, F. X. Influence of native lactic acid bacteria on the microbiological, biochemical and sensory profiles of Serra da Estrela cheese. Food Microbiology, v. 21, p. 233–240, 2004. MACRINA, F. L. et al. A multiple plasmid-containing Escherichia coli strain: convenient source of size reference plasmid molecules. Plasmid, v. 1, p. 417–420, 1978. MACWANA, S. J.; MURIANA, P. M. A “bacteriocin PCR array” for identification of bacteriocin-related structural genes in lactic acid bacteria. Journal of Microbiological Methods, v. 88, n. 2, p. 197–204, 2012. MACWANA, S.; MURIANA, P. M. Spontaneous bacteriocin resistance in Listeria monocytogenes as a susceptibility screen for identifying different mechanisms of resistance and modes of action by bacteriocins of lactic acid bacteria. Journal of Microbiological Methods, v. 88, n. 1, p. 7–13, 2012. MAHASNEH, A. M.; HAMDAN, S.; MAHASNEH, S. A. Probiotic Properties of Lactobacillus Species Isolated from Local Traditional Fermented Products. Jordan Journal of Biological Sciences, v. 8, n. 2, p. 81–87, 2015. MALHEIROS, P. S. et al. Optimization of growth and bacteriocin production by Lactobacillus sakei subsp. sakei 2a. Brazilian Journal of Microbiology, v. 46, n. 3, p. 825–834, 2015. MARAGKOUDAKIS, P. A. et al. Probiotic potential of Lactobacillus strains isolated from dairy products. International Dairy Journal, v. 16, p. 189–199, 2006. MARCOS, B. et al. Physical performance of biodegradable films intended for antimicrobial food packaging. Journal of Food Science, v. 75, n. 8, p. 502–507, 2010.

131
MARCOS, B. et al. Use of antimicrobial biodegradable packaging to control Listeria monocytogenes during storage of cooked ham. International Journal of Food Microbiology, v. 120, p. 152–158, 2007. MARIANSKI, S.; MARIANSKI, A. The Art of Making Fermented Sausage. 2 ed. Florida: Bookmagic, 2009. MARQUES, J. L. Caracterização fenotípica e molecular de bactérias ácido láticas isoladas de presunto cozido e verificação do potencial tecnológico e bacteriocinogênico contra Listeria monocytogenes e Staphylococcus aureus. 2014. 69 f. Departamento de Ciência e Tecnologia Agroinustrial. Universidade Federal de Pelotas, Pelotas, 2014. MARTINEAU, F. et al. Multiplex PCR assays for the detection of clinically relevant infected after cardiac surgery. Journal of Antimicrobial Chemotherapy, v. 46, p. 527–533, 2000. MARTINIS, E. C. P. DE; SANTAROSA, P. R.; FREITAS, F. Z. Caracterização preliminar de bacteriocinas produzidas por seis cepas de bactérias láticas isoladas de produtos cárneos embalados a vácuo. Ciência e Tecnologia de Alimentos, v. 23, n. 2, p. 195–199, 2003. MASSANI, M. B. et al. Active polymers containing Lactobacillus curvatus CRL705 bacteriocins: effectiveness assessment in Wieners. International Journal of Food Microbiology, v. 178, p. 7–12, 2014. MASSANI, M. B. et al. Development of an active wheat gluten film with Lactobacillus curvatus CRL705 bacteriocins and a study of its antimicrobial performance during ageing. Food Additives and Contaminants, v. 31, n. 1, p. 164–171, 2014. MASUDA, Y. et al. Identification and characterization of Leucocyclicin Q, a novel cyclic: Bacteriocin produced by Leuconostoc mesenteroides TK41401. Applied and Environmental Microbiology, v. 77, n. 22, p. 8164–8170, 2011. MATARAGAS, M. et al. Influence of pH and temperature on growth and bacteriocin production by Leuconostoc mesenteroides L124 and Lactobacillus curvatus L442. Meat Science, v. 64, p. 265–271, 2003. MATHIESEN, G. et al. Characterization of a new bacteriocin operon in sakacin

132
P-producing Lactobacillus sakei, showing strong translational coupling between the bacteriocin and immunity genes. Applied and Environmental Microbiology, v. 71, n. 7, p. 3565–3574, 2005. MATTICK, A T. R.; HIRSCH, A. Further observations on an inhibitory substance (nisin) from lactic streptococci. Lancet, v. 2, p. 5–8, 1947. MAURIELLO, G. et al. Isolation and technological properties of coagulase negative staphylococci from fermented sausages of Southern Italy. Meat science, v. 67, n. 1, p. 149–58, 2004. MCAULIFFE, O.; ROSS, R. P.; HILL, C. Lantibiotics structure, biosynthesis and mode of action. FEMS Microbiology Reviews, v. 25, p. 285–308, 2001. MEAT AND SAUSAGES, 2015. Starter cultures for making fermented sausages. Disponível em: <http://www.meatsandsausages.com/sausage-types/fermented-sausage/cultures>. Acesso em: 19 mar. 2016. MEIRA, S. M. M. et al. Identificação e resistência a barreiras biológicas de bactérias lácticas isoladas de leite e queijo de ovelha. Brazilian Journal of Food Technology, v. 13, n. EE01, p. 75–80, 2010. MESSENS, W. et al. Modelling growth and bacteriocin production by Lactobacillus curvatus LTH 1174 in response to temperature and pH values used for European sausage fermentation processes. International Journal of Food Microbiology, v. 81, p. 41–52, 2003. MILLER, M. B.; BASSLER, B. L. Quorum sensing in bacteria. Annual Review of Microbiology, v. 55, p. 165–199, 2001. MOLINOS, A. C. et al. Inhibition of Salmonella enterica Cells in Deli-Type Salad by Enterocin AS-48 in Combination with Other Antimicrobials. Probiotics and Antimicrobial Proteins, v. 1, p. 85–90, 2009. MOLL, G. N. et al. Mechanism of lantibiotic-induced pore-formation. Antonie van Leeuwenhoek, v. 69, p. 185–191, 1996. MORAES, P. M. et al. Foodborne pathogens and microbiological characteristics of raw milk soft cheese produced and on retail sale in Brazil. Foodborne

133
Pathogens and Disease, v. 6, n. 2, p. 245–249, 2009. MORI, M. T. S. S. Aplicação de nisina em envoltório celulósico para mortadela. 2013. 83 f. Escola de Engenharia Mauá do Centro Universitário do Instituto Mauá de Tecnologia, São Caetano do Sul, 2013. MOTTA, A. S.; BRANDELLI, A. Influence of growth conditions on bacteriocin production by Brevibacterium linens. Applied Microbiology and Biotechnology, v. 62, n. 2-3, p. 163–167, 2003. MPOUNTOUKAS, P. et al. Cytogenetic study in cultured human lymphocytes treated with three commonly used preservatives. Food and Chemical Toxicology, v. 46, n. 7, p. 2390–2393, 2008. MULDERS, J. W. M. et al. Identification and characterization of the lantibiotic nisin Z, a natural nisin variant. European Journal of Biochemistry, v. 201, p. 581–584, 1991. MÜLLER, C. M. O.; LAURINDO, J. B.; YAMASHITA, F. Effect of cellulose fibers addition on the mechanical properties and water vapor barrier of starch-based films. Food Hydrocolloids, v. 23, n. 5, p. 1328–1333, 2009. MUÑOZ, M. D. C. C. et al. Antibiotic resistance of Lactobacillus pentosus and Leuconostoc pseudomesenteroides isolated from naturally-fermented Aloreña table olives throughout fermentation process. International Journal of Food Microbiology, v. 172, p. 110–118, 2014. MUÑOZ-ATIENZA, E. et al. Antimicrobial activity, antibiotic susceptibility and virulence factors of Lactic Acid Bacteria of aquatic origin intended for use as probiotics in aquaculture. BMC Microbiology, v. 13, n. 1, p. 15, 2013. NAGPAL, R. et al. Probiotics, their health benefits and applications for developing healthier foods: A review. FEMS Microbiology Letters, v. 334, n. 1, p. 1–15, 2012. NAGPAL, R.; KUMAR, A.; KUMAR, M. Fortification and fermentation of fruit juices with probiotic lactobacilli. Annals of Microbiology, v. 62, n. 4, p. 1573–1578, 2012.

134
NASCIMENTO, MARISTELA DA SILVA DO, MORENO, IZILDINHA, KUAYE, A. Y. Bacteriocinas em alimentos : uma revisão Bacteriocins in food : a review. Brazilian Journal of Food Technology, v. 11, n. 2, p. 120–127, 2008. NAWAZ, M. et al. Characterization and transfer of antibiotic resistance in lactic acid bacteria from fermented food products. Current Microbiology, v. 62, p. 1081–1089, 2011. NEL, H. A. et al. Growth optimization of Pediococcus damnosus NCFB 1832 and the influence of pH and nutrients on the production of pediocin PD-1. Journal of Applied Microbiology, v. 91, p. 1131–1138, 2001. NERO, L. A. et al. Listeria monocytogenes and Salmonella spp. in raw milk produced in Brazil: Occurrence and interference of indigenous microbiota in their isolation and development. Zoonoses and Public Health, v. 55, p. 299–305, 2008. NES, I. F. et al. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie van Leeuwenhoek, v. 70, p. 113–128, 1996. NESPOLO, C. R.; BRANDELLI, A. Production of bacteriocin-like substances by lactic acid bacteria isolated from regional ovine cheese. Brazilian Journal of Microbiology, v. 41, p. 1009–1018, 2010. NETO, L. G. G. et al. Atividade antimicrobiana de bactérias ácido-lácticas isoladas de queijos de coalho artesanal e industrial frente a microrganismos indicadores. Arquivo Brasileiro de Medicina Veterinaria e Zootecnia, v. 57, p. 245–250, 2005. O’SULLIVAN, D. J. et al. High-throughput DNA sequencing to survey bacterial histidine and tyrosine decarboxylases in raw milk cheeses. BMC Microbiology, v. 15, n. 1, p. 266, 2015. OBST, M. et al. Lactose metabolism in Lactobacillus curvatus and Lactobacillus sake. FEMS Microbiology Letters, v. 97, n. 3, p. 209–214, 1992. OJAGH, S. M. et al. Development and evaluation of a novel biodegradable film made from chitosan and cinnamon essential oil with low affinity toward water. Food Chemistry, v. 122, n. 1, p. 161–166, 2010.

135
OJEDA-SANA, A. M. et al. New insights into antibacterial and antioxidant activities of rosemary essential oils and their main components. Food Control, v. 31, n. 1, p. 189–195, 2013. OJHA, K. S. et al. Technological advances for enhancing quality and safety of fermented meat products. Trends in Food Science & Technology, v. 44, n. 1, p. 105–116, 2015. OLIVEIRA, R. B. P.; OLIVEIRA, A. D. L.; GLÓRIA, M. B. A. Screening of lactic acid bacteria from vacuum packaged beef for antimicrobial activity. Brazilian Journal of Microbiology, v. 39, p. 368–374, 2008. ORTIGOSA, M. et al. Effect of lactobacillus adjunct cultures on the microbiological and physicochemical characteristics of Roncal-type ewes’-milk cheese. Food Microbiology, v. 23, p. 591–598, 2006. ORTOLANI, M. B. T. et al. Microbiological quality and safety of raw milk and soft cheese and detection of autochthonous lactic acid bacteria with antagonistic activity against Listeria monocytogenes, Salmonella spp., and Staphylococcus aureus. Foodborne Pathogens and Disease, v. 7, n. 2, p. 175–180, 2010. PANACEK, A. et al. Acute and chronic toxicity effects of silver nanoparticles (NPs) on Drosophila melanogaster. Environmental Science & Technology, v. 45, n. 11, p. 4974–4979, 2011. PANG, Y.; BOSCH, T.; ROBERTS, M. C. Single polymerase chain reaction for the detection of tetracycline-resistant determinants Tet K and Tet L. Molecular and Cellular Probes, v. 8, n. 5, p. 417–422, out. 1994. PAPAGIANNI, M.; ANASTASIADOU, S. Pediocins: The bacteriocins of Pediococci. Sources, production, properties and applications. Microbial cell factories, v. 8, n. 3, p. 1-16, 2009. PAPAMANOLI, E. et al. Characterization of lactic acid bacteria isolated from a Greek dry-fermented sausage in respect of their technological and probiotic properties. Meat Science, v. 65, n. 2, p. 859–67, 2003. PARK, D.-Y. Dual probiotic strains suppress high fructose-induced metabolic syndrome. World Journal of Gastroenterology, v. 19, n. 2, p. 274, 2013.

136
PATTANAYAIYING, R.; H-KITTIKUN, A.; CUTTER, C. N. Incorporation of nisin Z and lauric arginate into pullulan films to inhibit foodborne pathogens associated with fresh and ready-to-eat muscle foods. International Journal of Food Microbiology, v. 207, p. 77–82, 2015. PEREIRA, V. et al. Characterization for enterotoxin production , virulence factors , and antibiotic susceptibility of Staphylococcus aureus isolates from various foods in Portugal. Food Microbiology, v. 26, n. 3, p. 278–282, 2009. PÉREZ, P. F. et al. Surface properties of bifidobacterial strains of human origin. Applied and Environmental Microbiology, v. 64, n. 1, p. 21–26, 1998. RAI, A. K.; TAMANG, J. P.; PALNI, U. Microbiological studies of ethnic meat products of the Eastern Himalayas. Meat Science, v. 85, n. 3, p. 560–7, 2010. RANADHEERA, R. D. C. S.; BAINES, S. K.; ADAMS, M. C. Importance of food in probiotic efficacy. Food Research International, v. 43, n. 1, p. 1–7, 2010. REMIGER, A.; EHRMANN, M. A.; VOGEL, R. F. Identification of Bacteriocin-Encoding Genes in Lactobacilli by Polymerase Chain Reaction (PCR). Systematic and Applied Microbiology, v. 19, n. 1, p. 28–34, 1996. RIVAS, F. P. et al. Sakacin Q produced by Lactobacillus curvatus ACU-1: Functionality characterization and antilisterial activity on cooked meat surface. Meat Science, v. 97, n. 4, p. 475–479, 2014. RODGERS, S. Preserving non-fermented refrigerated foods with microbial cultures — a review. Trends in Food Science & Technology, v. 12, n. 2001, p. 276–284, 2002. ROGERS, L. A. The inhibiting effect of Streptococcus lactis on Lactobacillus bulgaricus. Journal Bacteriological, v. 16, p. 321–325, 1928. ROSS, R. P.; MORGAN, S.; HILL, C. Preservation and fermentation: Past, present and future. International Journal of Food Microbiology, v. 79, p. 3–16, 2002. RUIZ-MOYANO, S. et al. Screening of lactic acid bacteria and bifidobacteria for potential probiotic use in Iberian dry fermented sausages. Meat Science, v. 80,

137
n. 3, p. 715–21, 2008. SAARELA, M. et al. Tetracycline susceptibility of the ingested Lactobacillus acidophilus LaCH-5 and Bifidobacterium animalis subsp. lactis Bb-12 strains during antibiotic/probiotic intervention. International Journal of Antimicrobial Agents, v. 29, n. 3, p. 271–280, 2007. SADRANI, H. et al. Screening of potential probiotic Lactobacillus strains isolated from fermented foods, fruits and of human origin. Asian Journal of Pharmaceutical and Clinical Research, v. 7, n. 2, p. 216–225, 2014. SAKALA, R. M. et al. Lactobacillus fuchuensis sp. nov., isolated from vacuum-packaged refrigerated beef. International Journal of Systematic and Evolutionary Microbiology, v. 52, n. 4, p. 1151–1154, 2002. SANTOS, K. M. O. et al. Safety, beneficial and technological properties of Enterococcus faecium isolated from Brazilian cheeses. Brazilian Journal of Microbiology, v. 46, n. 1, p. 237–249, 2015. SAWITZKI, M. C. et al. Lactobacillus plantarum AJ2 isolated from naturally fermented sausage and its effects on the technological properties of Milano-type salami. Ciência e Tecnologia de Alimentos, v. 28, n. 3, p. 709–717, 2008. SCHILLINGER, U.; GUIGAS, C.; HOLZAPFEL, W. H. In vitro adherence and other properties of lactobacilli used in probiotic yoghurt-like products. International Dairy Journal, v. 15, p. 1289–1297, 2005. SCHILLINGER, U.; LUCKE, F. K. Antibacterial activity of Lactobacillus sake isolated from meat. Applied and Environmental Microbiology, v. 55, n. 8, p. 1901–1906, 1989. SCHIRRU, S. et al. Comparison of bacteriocins production from Enterococcus faecium strains in cheese whey and optimised commercial MRS medium. Annals of Microbiology, v. 64, p. 321–331, 2014. SEHN, C. P. Avaliação da atividade bacteriocinogênica e características probióticas do isolado Lactobacillus curvatus LC254 e da utilização da sua bacteriocina em filmes biodegradáveis. 2015. 116 f. Departamento de Ciência e Tecnologia Agroindustrial. Universidade Federal de Pelotas, Pelotas,

138
2015. SEMEDO, T. et al. Virulence factors in food, clinical and reference Enterococci: A common trait in the genus? Systematic and Applied Microbiology, v. 26, p. 13–22, 2003. SENTER, L. et al. Isolation and characterization of Lactobacillus plantarum BLS29 as a potential probiotic starter culture for pork sausage production. African Journal of Microbiology Research, v. 9, n. 32, p. 1887–1895, 2015. SETTANNI, L.; MOSCHETTI, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiology, v. 27, n. 6, p. 691–697, 2010. SIELADIE, D. V. et al. Probiotic Properties of Lactobacilli Strains Isolated From Raw Cow Milk in the Western Highlands of Cameroon. Innovative Romanian Food Biotechnology, v. 9, p. 12–28, 2011. SIMON, L. et al. Sakacin G, a new type of antilisterial bacteriocin. Applied and Environmental Microbiology, v. 68, n. 12, p. 6416–6420, 2002. SIP, A. et al. Anti-Listeria activity of lactic acid bacteria isolated from golka, a regional cheese produced in Poland. Food Control, v. 26, p. 117–124, 2012. STROMMENGER, B. et al. Multiplex PCR assay for simultaneous detection of nine clinically relevant antibiotic resistance genes in Staphylococcus aureus. Journal of Clinical Microbiology, v. 41, n. 9, p. 4089–94, 2003. SUDATI, J. H. et al. Valeriana officinalis attenuates the rotenone-induced toxicity in Drosophila melanogaster. NeuroToxicology, v. 37, p. 118-126, 2013. SUMERI, I. et al. Single bioreactor gastrointestinal tract simulator for study of survival of probiotic bacteria. Applied Microbiology and Biotechnology, v. 80, p. 317–324, 2008. SUSKOVIC, J. et al. Antimicrobial activity - The most important property of probiotic and starter lactic acid bacteria. Food Technology and Biotechnology, v. 48, n. 3, p. 296–307, 2010.

139
TAGG, JOHN R., DAJANI, ADNAN S., WANNAMAKER, L. W. Bacteriocins of Gram-Positive Bacteria. Bacteriological Reviews, v. 40, n. 3, p. 722–756, 1976. TICHACZEK, P. S. et al. Characterization of the bacteriocins curvacin A from Lactobacillus curvatus LTH1174 and sakacin P from L. sake LTH673. Systematic and Applied Microbiology, v. 15, n. 3, p. 460–468, 1992. TICHACZEK, P. S.; VOGEL, R. F.; HAMMES, W. P. Cloning and sequencing of cur A encoding curvacin A, the bacteriocin produced by Lactobacillus curvatus LTH 1174. Archives of Microbiology, v. 160, p. 279–283, 1993. TICHACZEK, P. S.; VOGEL, R. F.; HAMMES, W. P. Cloning and sequencing of sakP encoding sakacin P, the bacteriocin produced by Lactobacillus sake LTH 673. Microbiology, v. 140, p. 361–367, 1994. TODOROV, S. D. et al. Characterization of a bacteriocin produced by Lactobacillus sakei R1333 isolated from smoked salmon. Anaerobe, v. 17, n. 1, p. 23–31, 2011. TODOROV, S. D. et al. Potential beneficial properties of bacteriocin-producing lactic acid bacteria isolated from smoked salmon. Journal of Applied Microbiology, v. 110, p. 971–986, 2011. TODOROV, S. D.; DICKS, L. M. T. Characterization of mesentericin ST99, a bacteriocin produced by Leuconostoc mesenteroides subsp. dextranicum ST99 isolated from boza. Journal of Industrial Microbiology and Biotechnology, v. 31, n. 7, p. 323–329, 2004. TODOROV, S. D.; DICKS, L. M. T. Effect of medium components on bacteriocin production by Lactobacillus plantarum strains ST23LD and ST341LD, isolated from spoiled olive brine. Microbiological Research, v. 161, p. 102–108, 2006. TODOROV, S. D.; DICKS, L. M. T. Optimization of bacteriocin ST311LD production by Enterococcus faecium ST311LD, isolated from spoiled black olives. Journal of Microbiology, v. 43, n. 4, p. 370–374, 2005. TODOROV, S. D.; VAN REENEN, C. A.; DICKS, L. M. T. Optimization of bacteriocin production by Lactobacillus plantarum ST13BR, a strain isolated from barley beer. The Journal of General and Applied Microbiology, v. 50, p.

140
149–157, 2004. TULINI, F. L. et al. Evaluation of the proteolytic activity of Enterococcus faecalis FT132 and Lactobacillus paracasei FT700, isolated from dairy products in Brazil, using milk proteins as substrates. European Food Research and Technology, v. 241, n. 3, p. 385–392, 2015. ÜNLÜ, G.; NIELSEN, B.; IONITA, C. Production of antilisterial bacteriocins from lactic acid bacteria in dairy-based media: a comparative study. Probiotics and Antimicrobial Proteins, v. 7, n. 4, p. 259–274, 2015. URSO, R. et al. Sequencing and expression analysis of the sakacin P bacteriocin produced by a Lactobacillus sakei strain isolated from naturally fermented sausages. Applied Microbiology and Biotechnology, v. 71, n. 4, p. 480–485, 2006. VAN BELKUM, M. J.; STILES, M. E. Nonlantibiotic antibacterial peptides from lactic acid bacteria. Natural Product Reports, v. 17, p. 323–335, 2000. VASILJEVIC, T.; SHAH, N. P. Probiotics from metchnikoff to bioactives. International Dairy Journal, v. 18, p. 714–728, 2008. VAUGHAN, A. et al. An analysis of bacteriocins produced by lactic acid bacteria isolated from malted barley. Journal of Applied Microbiology, v. 91, n. 1, p. 131–138, 2001. VAUGHAN, A.; EIJSINK, V. G. H.; VAN SINDEREN, D. Functional characterization of a composite bacteriocin locus from malt isolate Lactobacillus sakei 5. Applied and Environmental Microbiology, v. 69, n. 12, p. 7194–7203, 2003. VAUGHAN, E. E.; MOLLET, B.; DEVOS, W. M. Functionality of probiotics and intestinal lactobacilli: Light in the intestinal tract tunnel. Current Opinion in Biotechnology, v. 10, n. 5, p. 505–510, 1999. VELJOVIC, K. et al. Preliminary characterization of lactic acid bacteria isolated from Zlatar cheese. Journal of Applied Microbiology, v. 103, p. 2142–2152, 2007.

141
VERMEIREN, L.; DEVLIEGHERE, F.; DEBEVERE, J. Effectiveness of some recent antimicrobial packaging concepts. Food Additives and Contaminants, v. 19, p. 163–171, 2002. VINDEROLA, C. G.; REINHEIMER, J. A. Lactic acid starter and probiotic bacteria: A comparative “in vitro” study of probiotic characteristics and biological barrier resistance. Food Research International, v. 36, p. 895–904, 2003. WANG, J. et al. Fermentation characteristics and transit tolerance of probiotic Lactobacillus casei Zhang in soymilk and bovine milk during storage. Journal of Dairy Science, v. 92, n. 6, p. 2468–2476, 2009. WHITEHEAD, H. R. CCXLII. A substance inhibiting bacterial. Biochemistry Journal, v. 27, p. 1793–1800, 1933. XIRAPHI, N. et al. Purification and characterization of a bacteriocin produced by Leuconostoc mesenteroides E131. Meat Science, v. 80, p. 194–203, 2008. XIRAPHI, N. et al. Purification and characterization of curvaticin L442, a bacteriocin produced by Lactobacillus curvatus L442. Antonie van Leeuwenhoek, v. 89, n. 1, p. 19–26, 2006. XU, H. et al. Assessment of cell surface properties and adhesion potential of selected probiotic strains. Letters in Applied Microbiology, v. 49, n. 4, p. 434–442, 2009. YANG, R.; RAY, B. Factors influencing production of bacteriocins by lactic acid bacteria. Food Microbiology, v. 11, p. 281–291, 1994. YI, H. et al. Effect of exogenous factors on bacteriocin production from Lactobacillus paracasei J23 by using a resting cell system. International Journal of Molecular Sciences, v. 14, p. 24355–24365, 2013. YU, Z. et al. Evaluation of probiotic properties of Lactobacillus plantarum strains isolated from Chinese sauerkraut. World Journal of Microbiology and Biotechnology, v. 29, p. 489–498, 2013. ZAVAREZE, E. D. R. et al. Development of oxidised and heat-moisture treated potato starch film. Food Chemistry, v. 132, n. 1, p. 344–350, 2012.

142
ZYGMUNT, W. A.; BROWDER, H. P.; TAVORMINA, P. A. Lytic action of lysostaphin on susceptible and resistant strains of Staphylococcus aureus. Canadian Journal of Microbiology, v. 13, n. 16, p. 845–853, 1967.

143
Apêndices

144
Apêndice A – Resistência e susceptibilidade a antimicrobianos de uso clínico por Lactobacillus curvatus P99 utilizando o teste de disco difusão em ágar MRS.
Figura 17 - Resistência e susceptibilidade a antimicrobianos de uso clínico por Lactobacillus curvatus P99 utilizando o teste de disco difusão em ágar. A: Cloranfenicol (halo de inibição – 45mm); B: Ampicilina (halo de inibição – 16mm); C: Vancomicina (halo de inibição – não detectado).
Fonte: Autor.
B
A
C

145
Apêndice B – Quantificação da atividade antimicrobiana do sobrenadante livre de células contendo substâncias antimicrobianas (bacteriocin- like) produzidas por Lactobacillus curvatus P99
Figura 18 - Quantificação da atividade antimicrobiana do sobrenadante livre de células contendo substâncias antimicrobianas (bacteriocin- like) produzidas por Lactobacillus curvatus P99. Fonte: Autor.

146
Apêndice C – Atividade antimicrobiana de filmes incorporados de sobrenadante livre de células contendo substância antimicrobiana de origem proteica (bacteriocin-like) contra Listeria monocytogenes Scott A. Figura 19 - Atividade antimicrobiana de filmes incorporados de sobrenadante livre de células contendo substância antimicrobiana de origem proteica (bacteriocin-like) contra Listeria monocytogenes Scott A. CT: filme controle, CIM: filme elaborado com concentração inibitória minima do SLC; CBM: filme elaborado com concentração bactericida minima do SLC. Fonte: Autor.





























