Embed Size (px)
Citation preview

Universidad de Buenos Aires Facultad de Farmacia y Bioquímica
Cátedra de Microbiología
Papel del “quorum sensing” y del hierro en la formación de biofilms y en la producción de otros
potenciales factores de virulencia de
Stenotrophomonas maltophilia
Tesis presentada para optar al título de Doctor de la Universidad
de Buenos Aires en el área Microbiología
Bioq. y Farm. Carlos Adrián García
Directora: Dra. Mirta A. Franco
Directora Adjunta: Dra. Beatriz Passerini de Rossi
2014

i
Agradecimientos
A mi directora la Dra. Mirta Franco, gracias por la confianza ofrecida desde que comencé
a trabajar en este proyecto y por brindarme la oportunidad de realizar mi investigación
libremente.
A mi directora adjunta la Dra Beatriz Passerini de Rossi agradezco la paciencia y
dedicación que le dedicó a este trabajo. Gracias por acompañarme en todas las
dificultades que se plantearon en este periodo.
A la Dra Laura Friedman gracias por brindarme toda su ayuda en ei trabajo de
investigación y en la docencia.
A mis compañeros de la cátedra de Microbiología con quienes compartí tantos agradables
años de docencia
Agradezco al Dr Tomas Santa Coloma por su amabilidad e invaluable contribución a este
trabajo al enseñarme a utilizar el microscopio confocal, a la cátedra química analítica por
el gentil asesoramiento y por permitirme utilizar el equipo de IR.
Un agradecimiento general a todas y cada una de las personas que han compartido
conmigo todos estos años de trabajo, con sus altos y bajos. Desde los más profundo de
mi corazón, agradezco el haberme brindado apoyo, colaboración, ánimo y sobre todo
cariño y amistad.
Agradecimiento a mis padres que siempre alentaron mi formación.
Gracias a la educación pública y gratuita, que me formó como profesional y me brindó la
oportunidad de realizar esta tesis. A la Universidad de Buenos Aires y a la Facultad de
Farmacia y Bioquímica, por ser el lugar donde pude crecer académicamente

ii
Contenidos
Agradecimientos ................................................................................................................. i
Contenidos ......................................................................................................................... ii
Abreviaturas ..................................................................................................................... vii
Introducción ....................................................................................................................... 1
1. Stenotrophomonas maltophilia .................................................................................... 2
1.1. Taxonomia ........................................................................................................... 2
1.2. Características ecológicas y bioquímicas ............................................................ 2
1.3. Aspectos epidemiológicos ................................................................................... 3
1.4. Factores de virulencia .......................................................................................... 4
1.5. Modelos de virulencia .......................................................................................... 5
2. Biofilms bacterianos .................................................................................................... 5
2.1. Implicancia de la formación de biofilms ................................................................ 7
3. Sistemas de “Quorum sensing” ................................................................................... 9
3.1. El sistema rpf/DSF ............................................................................................. 10
3.1.1. El sistema de señalizacion rpf/DSF de S. maltophilia ..................................... 12
4. El hierro en la fisiología bacteriana ........................................................................... 13
4.1. Sistemas de captación de hierro ........................................................................ 14
4.2. Hierro y estrés oxidativo .................................................................................... 15
5. El hierro como señal: El sistema Fur ......................................................................... 16
Hipótesis y Objetivos ....................................................................................................... 18
1. Hipótesis ................................................................................................................... 19
2. Objetivo general........................................................................................................ 19
3. Objetivos parciales ................................................................................................... 19
Materiales y Métodos ....................................................................................................... 20
1. Cepas bacterianas .................................................................................................... 21
2. Tipificación molecular ............................................................................................... 21
2.1. ERIC-PCR ......................................................................................................... 21
2.2. REP-PCR .......................................................................................................... 22
3. Producción de potenciales factores de virulencia...................................................... 22

iii
3.1. Detección de la presencia de los genes narG y stmPr-1 en aislamientos locales de S. maltophilia ........................................................................................................... 22
3.2. Prueba de reducción de nitratos ........................................................................ 23
3.3. Proteasas .......................................................................................................... 24
3.4. Lipasa ................................................................................................................ 24
3.5. DNasa ............................................................................................................... 24
3.6. Resistencia al estrés oxidativo ........................................................................... 24
3.7. Producción de sideróforos ................................................................................. 25
3.8. Determinación de la naturaleza química de sideróforos ..................................... 26
3.8.1. Condiciones de cultivo ................................................................................... 26
3.8.2. Ensayo de Arnow ........................................................................................... 26
3.8.3. Ensayo de Csáky ........................................................................................... 26
4. Curvas de Crecimiento ............................................................................................. 27
5. Bioensayo para determinar la producción DSF ......................................................... 27
5.1. Extracción de DSF de sobrenadantes de cultivo ................................................ 27
5.2. Bioensayo .......................................................................................................... 27
6. Formación de biofilms ............................................................................................... 28
6.1. Estandarización del inoculo ............................................................................... 28
6.2. Ensayo en Tubos ............................................................................................... 28
6.3. Ensayo en microplacas ...................................................................................... 29
6.4. Microscopia óptica ............................................................................................. 29
7. Microscopia confocal laser de barrido. ...................................................................... 29
8. Movilidad asociada a superficies o “twitching” .......................................................... 30
9. Producción de sustancias poliméricas extracelulares (EPS) ..................................... 30
10. Espectroscopia de reflexión total atenuada infrarroja con transformada de Fourier (ATR-FTIR) ...................................................................................................................... 31
11. Aislamiento de mutantes Fur ................................................................................. 31
12. Ensayo de reducción del Azul de Nitrotetrazolio (NBT) ......................................... 32
13. Actividad de la enzima superoxido dismutasa (SOD) ............................................ 32
13.1. Obtención de extractos crudos ....................................................................... 32
13.2. Actividad en cultivos planctónicos .................................................................. 33
13.3. Actividad en el biofilm..................................................................................... 33
13.4. Geles nativos para SOD y determinación del tipo de isoenzima ..................... 33
14. Evaluación de la virulencia con el modelo de infección de Galleria mellonella ...... 34

iv
15. Preparación de fracciones enriquecidas en OMPs ................................................ 35
16. Electroforesis en geles de poliacrilamida (SDS-PAGE) ......................................... 35
17. Digestión en gel e identificación de proteínas mediante espectrometría de masa . 35
18. Cromatografía en placa delgada (TLC) ................................................................. 36
19. Cromatografía de alta performance (HPLC) y detección UV con arreglos de diodos 37
19.1. Preparación de la muestra ............................................................................. 37
19.2. HPLC con arreglo de diodos .......................................................................... 37
20. Estadística ............................................................................................................ 37
Resultados y Discusión .................................................................................................... 38
Sección I .......................................................................................................................... 39
Tipificación de aislamientos locales de S. maltophilia recuperados de infecciones asociadas al uso de productos médicos........................................................................... 39
1. Identificación y fenotipificación .................................................................................. 40
2. Tipificación por métodos genotípicos ........................................................................ 42
2.1. ERIC-PCR ......................................................................................................... 43
2.2. REP-PCR .......................................................................................................... 45
3. Discusión .................................................................................................................. 46
Sección II ......................................................................................................................... 48
Producción de potenciales factores de virulencia en aislamientos locales de S. maltophilia ........................................................................................................................................ 48
1. Formación de biofilms de aislamientos locales ......................................................... 48
2. Presencia del gen codificante de la proteasa StmPr1, actividad de proteasa y lipasa50
3. Producción de sideróforos ........................................................................................ 50
4. Resistencia al estrés oxidativo .................................................................................. 52
5. Presencia del gen narG, codificante de una nitrato reductasa y su relación con la utilización nitrato .............................................................................................................. 52
6. Virulencia en el modelo de infección de Galleria mellonella ...................................... 54
7. Discusión .................................................................................................................. 56
Sección III ........................................................................................................................ 60
Sistema de captación de hierro de alta afinidad de S. maltophilia K279a y su posible regulación por el sistema Fur ....................................................................................... 60
1. Aislamiento de mutantes Fur .................................................................................... 60
2. Caracterización de proteínas de membrana externa reguladas por hierro en S. maltophilia K279a ............................................................................................................ 62

v
2.1. Análisis de los perfiles de OMPs ........................................................................ 62
2.2. Identificación de las OMPs reguladas por hierro mediante espectrometría de masa 63
3. Caracterización de los sideróforos producidos en S. maltophilia ............................... 65
3.1. Determinación de la naturaleza química de los sideróforos ............................... 65
3.2. Análisis mediante cromatografía en capa delgada (TLC) de extractos provenientes de sobrenadantes de cultivos de K279a .................................................. 66
3.3. Análisis mediante cromatografía líquida de alta performance (HPLC) de extractos provenientes de sobrenadantes de cultivos de K279a .................................................. 67
4. Discusión .................................................................................................................. 69
Sección IV ....................................................................................................................... 71
Función del sistema de señalización rpfF/DSF en la formación de biofilms y en la virulencia de S. maltophilia .............................................................................................. 71
1. Crecimiento planctónico ............................................................................................ 71
2. Papel del “quorum sensing” en la formación de biofilms ........................................... 72
2.1. Análisis de biofilms formados sobre superficies hidrofílicas e hidrofóbicas ........ 72
2.2. Análisis de biofilms formados en microplacas .................................................... 74
2.3. Análisis de biofilms por microscopía óptica ........................................................ 76
2.4. Análisis de biofilms por microscopia confocal laser de barrido (CLSM) .............. 77
3. Producción de sustancias poliméricas extracelulares (EPS) ..................................... 79
4. Motilidad tipo “twitching” ........................................................................................... 83
5. Hidrofobicidad de la superficie celular ....................................................................... 85
6. Resistencia al estrés oxidativo .................................................................................. 86
7. Producción de exoenzimas ....................................................................................... 87
8. Evaluación de la virulencia mediante el modelo de Galleria mellonella ..................... 88
9. Discusión .................................................................................................................. 89
Sección V ........................................................................................................................ 94
Función del hierro en la formación de biofilms y en la resistencia al estrés oxidativo de S. maltophilia ....................................................................................................................... 94
1. Efecto de la disponibilidad de hierro en la formación de biofilms y la producción de EPS ................................................................................................................................. 94
1.1. Análisis de biofilms formados en microplacas .................................................... 95
1.2. Análisis de biofilms por CLSM ........................................................................... 96
1.3. Producción de EPS ............................................................................................ 98
2. Efecto de la disponibilidad de hierro en la respuesta al estrés oxidativo ................. 101

vi
2.1. Determinación del tipo de isoenzima de superóxido dismutasa (SOD) ............ 101
2.2. Actividad de SOD en cultivos planctónicos y biofilms ...................................... 102
2.3. Niveles de especies reactivas del oxigeno (ERO) en el biofilm ........................ 104
3. Evaluación del efecto del hierro en la producción de DSF ...................................... 105
4. Evaluación de la virulencia en el modelo de infección de Galleria mellonella .......... 106
5. Discusión ................................................................................................................ 108
Conclusiones ................................................................................................................. 111
Resumen ....................................................................................................................... 117
Publicaciones ................................................................................................................ 121
Anexo I .......................................................................................................................... 125
Bibliografía ..................................................................................................................... 127

vii
Abreviaturas
ADN: Acido desoxirribonucleico ARN: ácido ribonucleico ATCC: American type culture colection CAS: Cromoasurol S CirA: receptor de colicina I NaCN: Cianuro de sidio CV: Cristal violeta °C: Grados centígrados DBHA: Acido dihidroxibenzóico BS: Borosilicato Dip: 2-2´ dipiridilo DMSO: Dimetil sulfóxido DO546 : Densidad óptica a 546 nm DSF: “Diffusible signal factor” EPS: Sustancias poliméricas extracelulares ERIC: Secuencias consenso repetitivas intragénicas de enterobacterias ERO: Especies reactivas del oxigeno Fe2+: Ion ferroso Fe3+: Ion férrico FepA: Receptor de membrana externa de enterobactina Fur: “Ferric uptake regulator” h: horas H2O2: Peróxido de hidrogeno HPLC: Cromatografía líquida de alta performance kDa: kilodalton LB: Luria Bertani LBA: Lavado bronquealveolar LB-glu 0,1%: Luria Bertani suplementado con 0,1% de glucosa MATH: “Microbial adherence to hydrocarbons” min: Minutos mm: Milímetros NBT: Azul de nitrotetrazolio nm: Nanómetros OMP: Proteína de membrana externa P: Nivel justo de significación pb: Pares de bases PBS: Buffer fosfato salino PCR: Reacción en cadena de la polimerasa PM: Peso molecular PP: Polipropileno PS: Poliestireno QS: “Quorum sensing” rpf: “regulation of pathogenicity factors” REP: Secuencias repetitivas palindrómicas extragénicas SAT: Salt aggregation test SDS-PAGE: Gel de poliacrilamida con dodecilsulfato sodico SOD: Superóxido dismutasa SS: Stainer-Scholte medio químicamente definido

viii
SSC: Stainer-Scholte con Casamino ácidos 0,1 % (P/V) SSC-CAS: SSC con el agregado del reactivo CAS y agar SSC-CAS-Dip: SSC-CAS con el agregado de Dip 100 µM TLC: Cromatografía en capa delgada TSB: Caldo tripteina soya TSA: Agar tripteina soya UFC/ml: Unidades formadoras de colonias por mililitro UPGMA: “Unweighted Pair Group Method with Arithmetic Averages” UV: Ultravioleta Xcc: Xanthomonas campestris pv campestris ZnSe: Seleniuro de zinc

Introducción

Introducción
2
1. Stenotrophomonas maltophilia
1.1. Taxonomia
El género Stenotrophomonas pertenece, junto con Xanthomonas, a la subclase γ-β
de las proteobacterias (6). Esta especie fue inicialmente denominada Pseudomonas
maltophilia (84), y posteriormente, en base a estudios de homología del ARN ribosomal
fue clasificada como Xanthomonas maltophilia, sin embargo esta clasificación no fue
universalmente aceptada. Finalmente, debido a estudios de hibridación de ADN con ARN
ribosomal, junto a diferencias fenotípicas entre aislamientos de esta especie y los del
genero Xanthomonas, se propuso en 1993 la creación de un nuevo género denominado
Stenotrophomonas (Stenos, del griego estrecho; trophos, del griego alimento; monas, del
griego única; malt, del inglés maltosa; philia, del griego amistad) (115) siendo S.
maltophilia la única especie (115). En 1997 S. maltophilia dejó de ser la única integrante
del género al describirse la especie S. africana que presentaba características
bioquímicas idénticas a S. maltophilia, excepto por su incapacidad para hidrolizar cis-
aconitato (49). Sin embargo, un estudio posterior demostró que S. africana es una cepa
de S. maltophilia (26). Actualmente existen nuevas especies dentro del género: S.
acidaminiphila (9), S. chelatiphaga (90), S. daejeonensis (95), S. ginsengisoli (91), S.
humi, S. terrae (77), S. koreensis (154), S. nitritireducens (61), S. pavanii (122) y S.
rhizophila (153).
S. maltophilia es la única especie descripta como patógeno humano oportunista
(3).
1.2. Características ecológicas y bioquímicas
S. maltophilia es un bacilo Gram-negativo no fermentador de la glucosa, aerobio
obligado, móvil debido a la presencia de varios flagelos polares. Su temperatura optima
de crecimiento es 35 °C, es oxidasa negativo aunque algunos aislamientos son oxidasa
positivo (22), catalasa positivo, indol negativo y lisina decarboxilasa positivo. Además
presenta la capacidad de hidrolizar esculina, gelatina y ADN (41).

Introducción
3
Es una especie ampliamente distribuida en el ambiente que se aísla de numerosas
fuentes como el agua, el suelo, la rizósfora e incluso fue aislada de suelos antárticos
(147).
1.3. Aspectos epidemiológicos
S. maltophilia es un patógeno nosocomial emergente multiresistente que se aisla
con frecuencia creciente. Esta especie ha llegado a considerarse el tercer bacilo Gram-
negativo más común de los aislamientos clínicos después de P. aeruginosa y
Acinetobacter spp. (68). Aunque es considerada como una bacteria de patogenicidad
limitada, causa infecciones en pacientes inmunocomprometidos o sometidos al uso de
productos médicos. Otro factor que predispone a la infección por S. maltophilia es la
terapia prolongada con antimicrobianos de amplio espectro, principalmente
carbapenemes, debido a su multirresistencia (41, 97, 131). Entre las infecciones que
produce se pueden mencionar neumonía asociada a respiración mecánica, bacteriemia
asociada a catéteres venosos o de diálisis, infecciones urinarias, y con menor frecuencia
puede producir meningitis y endocarditis (41, 45). Esta especie ha sido aislada con
frecuencia creciente en pacientes con fibrosis quística (42).
Una gran variedad de técnicas de tipificación se han utilizado para estudiar la
epidemiologia y las vías de transmisión de esta especie. Entre ellas, la tipificación basada
en la comparación de los antibiogramas (antibiotipificaión) resultó ser poco discriminativa
porque la especie presenta resistencia intrínseca a numerosos antimicrobianos (145). La
biotipificacion también resultó ser de limitada utilidad debido a su metabolismo
relativamente inerte (41). Por el contrario numerosas técnicas de tipificación molecular
demostraron ser útiles para tipificar aislamientos de esta especie, entre ellas la
electroforesis en campo pulsado (PFGE) presenta el mayor poder discriminatorio, pero es
una técnica laboriosa y costosa, lo que la hace poco practica. Por el contrario, las técnicas
basadas en la amplificación mediante la reacción en cadena de la polimerasa son
accesibles, menos laboriosas y han demostrado tener un poder de discriminación
aceptable aunque menor al del PFGE (133). Otros autores desarrollaron un esquema de
tipificación por secuenciación de múltiples locus (MLST) que presenta buen poder
discriminatorio para el estudio de la estructura poblacional de la especie. En la actualidad
existe una base de datos (http://pubmlst.org/smaltophilia/) con genogrupos definidos para
realizar el análisis (89)

Introducción
4
Los estudios epidemiológicos realizados por numerosos grupos de trabajo, sobre
aislamientos clínicos, mostraron una gran diversidad genética en esta especie incluso
entre aislamientos recuperados dentro de un mismo hospital (23, 68, 144).
1.4. Factores de virulencia
A pesar del amplio espectro de síndromes clínicos asociados a S. maltophilia poco
se conoce acerca de sus factores de virulencia. Enzimas extracelulares tales como
DNasa, fibrinolisina, lipasas y proteasas, han sido relacionadas con la patogénesis de S.
maltophilia (18). La producción de proteasa juega un papel significativo en la patogénesis
bacteriana ya que participa en la invasión y el daño tisular. Windhorst et al. (150)
caracterizaron una serina proteasa alcalina denominada StmPr1 que degrada proteínas
del suero y tejido conectivo.
Otro factor de virulencia son los sideróforos, los cuales facilitan la adquisición de
hierro en el huésped. En S. maltophilia existen reportes controversiales ya que mientras
algunos autores detectaron sideróforos tipo hidroxamato (ornibactina), otros detectaron
catecoles como enterobactina (36, 126).
Un importante factor de virulencia es su capacidad de formar biofilms ya que los
mismos facilitan la persistencia en el huésped, la evasión de la respuesta inmune y la
supervivencia frente a altas concentraciones de antimicrobianos (38, 45, 119). Diferentes
estructuras de superficie y la presencia de ciertos genes han sido relacionados con la
formación de biofilms. De Oliveira-Garcia et al. (40) caracterizaron las fimbrias SMF-1,
relacionadas con la adherencia a células Hep-2, y con la formación de biofilms. El genoma
de la cepa K279a recientemente secuenciado (EMBL/GeneBank AM743169) contiene
genes que codifican pili tipo I y tipo IV, relacionados con la adhesión y la motilidad
asociada a superficies o “twitching” (32). McKay et al. (101) describieron el papel de la
fosfoglucomutasa (PGM) de S. maltophilia en la biosíntesis del lipopolisacárido (LPS), la
virulencia, y la resistencia a antibióticos. El gen xanA, también conocido como spgM,
presenta homología con algC responsable en P. aeruginosa de la producción de PGM
asociada con la biosíntesis del LPS y de alginato, exopolisacarido de la matriz del biofilm.
Por otra parte, en S. maltophilia K279a se ha descripto la presencia de una nitrato
reductasa unida a membrana codificada por el gen nar y de una formiato deshidrogenasa,

Introducción
5
codificada por el gen fdn, que soportarían el crecimiento en condiciones de
microoxigenación y así facilitarían la formación de biofilms (32).
1.5. Modelos de virulencia
Diferentes organismos hospedadores fueron utilizados como modelo para evaluar la
virulencia de esta especie y estudiar los mecanismos moleculares relacionados con su
patogenicidad. En este sentido Nicoletti et al (110) utilizaron el modelo de infección de
Galleria mellonella para estudiar la relevancia de las proteasas StmPr1 y StmPr2 y de
estearasas en la virulencia de aislamientos clínicos de S. maltophilia. De este modo
pudieron determinar que la expresión de proteasa StmPr1 es relevante para la virulencia
de este microorganismo. Además, McCarthy et al. utilizaron el modelo de G. mellonella y
determinaron que la proteína Ax21 de S. maltophilia K279a es una señal célula-célula que
regula la virulencia (100).
Fouhy et al (62) evaluaron el efecto de la mutación en el gen rpfF sobre la virulencia
de S. maltophilia con el modelo de Caenorhabditis elegans y encontraron que la cepa
K279a presenta una virulencia similar P. aeruginosa PA14 mientras que la mutante
presenta una disminución en la virulencia.
Por otra parte se utilizaron dos amebas Dictyostelium discoideum Ax2 y
Acanthamoeba castellanii ATTC 30234 que, a través de un ensayo de muerte en placa,
permitieron evaluar la virulencia de 59 aislamientos de S. maltophilia de origen clínico y
ambiental (1).
Por último, Ferrer-Navarro et al (59) utilizaron el modelo de infección del pez cebra
Danio rerio y encontraron que el mismo presenta una elevada capacidad para discriminar
la virulencia de aislamientos clínicos de S. maltophilia, además, se ha reportado que este
modelo correlaciona con el modelo de virulencia en ratones (101).
2. Biofilms bacterianos
Los biofilms son comunidades de microorganismos que crecen adheridos a una
superficie embebidos en una matriz extracelular que ellos mismos producen (38). En la
actualidad se acepta que la mayoría de los microorganismos persisten en la naturaleza
formando parte de un biofilm (66). Las bacterias en el biofilm presentan un fenotipo

Introducción
6
diferente al de sus contrapartes planctónicas caracterizado por una reducción en la
velocidad de crecimiento y una expresión diferencial de genes (46). Incluso dentro de un
mismo biofilm hay zonas con niveles metabólicos diferentes. En las zonas más profundas
hay poca o nula división celular y la tasa metabólica es baja, mientras que en las zonas
más cercanas a la superficie hay mayor disponibilidad de nutrientes lo que conlleva a una
mayor actividad metabólica y tasa de división celular (123).
La formación de biofilms es un proceso complejo, finamente regulado, que se inicia
con la adhesión de las bacterias a una superficie, que puede ser tanto biótica como
abiótica. La movilidad mediada por flagelos es importante para que la bacteria establezca
el contacto inicial con la superficie (111). La adhesión, que primero es reversible y luego
irreversible, depende de estructuras de la superficie bacteriana tales como pili y
adhesinas, y de su hidrofobicidad, y además, de propiedades del sustrato tales como la
hidrofobicidad y la rugosidad, y de las interacciones electrostáticas entre ambas
superficies (46).
La movilidad tipo “twitching”, mediada por el pili de tipo IV, es necesaria para la
migración de las células a lo largo de la superficie con la consecuente formación de
agregados celulares (111). El pili tipo IV estabiliza las interacciones con la superficie
abiótica y las interacciones entre células, necesarias para la formación de microcolonias.
Luego, se produce la expansión clonal de las bacterias y comienza la producción de
exopolisacarido que junto con proteínas y ácidos nucleicos forman la matriz del biofilm
(124). Posteriormente, como resultado de un proceso de maduración, se forman las
estructuras tridimensionales, características del biofilm, compuestas por microcolonias de
gran tamaño, matriz extracelular y canales de flujo de agua por los que se produce el
consecuente acceso a nutrientes para las células del biofilm (138). Finalmente ocurre la
dispersión, en la cual se liberan algunas células plantónicas o agregados de células que
eventualmente podrán adherirse en otros lugares que les resulten favorables (Figura 1).

Introducción
7
Figura 1. Etapas de formación de biofilm. Se distinguen 5 etapas discretas. 1) Adhesión
reversible; 2) Adhesión irreversible y síntesis de matriz, 3) formación de microcolonias; 4)
maduración y 5) dispersión. Imagen modificada de Monroe, D. 2007 (106)
2.1. Implicancia de la formación de biofilms
La formación de biofilms es relevante ya que muchas infecciones bacterianas
crónicas están ligadas a este fenómeno. Los productos médicos invasivos son propensos
a la colonización bacteriana y a la formación de biofilms, lo que genera un nicho de
microorganismos que provocan infecciones recurrentes. Los biofilms son difíciles de
erradicar porque son extremadamente resistentes frente a fuerzas de corte,
antimicrobianos y sustancias biocidas, que pueden eliminar células planctónicas pero no
biofilms, además las células en el biofilm evaden la respuesta inmune del hospedador (28,
29). Los esquemas terapéuticos de control de las infecciones agudas generalmente no
han sido exitosos en las infecciones causadas por biofilms, la susceptibilidad del biofilm
es hasta 1000 veces menor que la de su contraparte planctónica (28).
En nuestro grupo de trabajo hemos determinado la sensibilidad a fluoroquinolonas
de cultivos planctónicos (concentración inhibitoria mínima, CIM y concentración
bactericida mínima, CBM) y de biofilms (MBIC: minimal biofilm inhibition concentration y
MRC: minimal regrowth concentration) de S. maltophilia. De los 32 aislamientos locales
estudiados, 27 fueron sensibles a levofloxacina y 14 a ciprofloxacina. Los aislamientos

Introducción
8
sensibles a fluoroquinolonas de acuerdo a los valores de CIM fueron altamente
resistentes según los valores de MRC. La sensibilidad de los biofilms de S. maltophilia fue
entre 100 y 1000 veces menor que la de sus contrapartes planctónicas (119). Por lo tanto,
las concentraciones terapéuticas de fluoroquinolonas no pueden ser usadas como
monoterapia para la erradicación de biofilms, sin embargo, son candidatos para estudios
de sellado antibiótico (“lock solutions”).
La técnica de sellado antibiótico involucra la instilación de una solución
concentrada de un antimicrobiano en el catéter. Se comparó la actividad in vitro de
diferentes “lock solutions” (levofloxacina 2.5 mg/ml, EDTA 30 mg/ml, levofloxacina-EDTA,
etanol 25% y 40%, etanol 25%-EDTA) sobre biofilms preformados en catéteres de
silicona. Después de 24h de exposición, Levofloxacina logró erradicar el biofilm, mientras
que EDTA redujo su viabilidad a 1.0-5.0%. La combinación EDTA-Levofloxacina no
mostró sinergia. En cambio, etanol 40%, 25% o la asociación etanol-EDTA, erradicaron el
biofilm luego de 1h de tratamiento. Estos resultados, que dieron origen a una publicación
(118), sugieren que etanol en baja concentración, solo o en combinación con EDTA, sería
una alternativa útil en la prevención y el tratamiento, en combinación con ATB sistémicos,
de bacteriemias por S. maltophilia asociadas al uso de catéteres.
Se han propuesto diferentes mecanismos para explicar la resistencia del biofilm
frente a sustancias biocidas, uno de ellos se basa en la existencia de una barrera para la
difusión de compuestos hacia el interior del biofilm debido a la presencia de la matriz
compuesta principalmente por exopolisacáridos. El impedimento en la difusión de
compuestos podría ser generado por adsorción de los mismos o por reacción con los
componentes de la matriz (99). En este sentido, se observó que la difusión de el cloro a
través de la matriz de un biofilm mixto de Klebsiella pneumoniae y P. aeruginosa es del 20
% o menor (39) y que la difusión de ciprofloxacina se encuentra impedida por la adsorción
a la matriz de biofilms de P. aeruginosa (139). Por otra parte, se reportó que el biofilm de
Staphylococcus epidermidis permite la difusión de rifampicina y vancomicina (53). Por lo
tanto, la matriz es una barrera para la difusión de algunas sustancias pero no explica la
mayor resistencia del biofilm frente a todos los biocidas. Otro mecanismo propuesto es la
baja velocidad de crecimiento de las células generada por la limitación de nutrientes. Por
otra parte la generación de una respuesta al estrés oxidativo provocada por la limitación
de nutrientes en el biofilm, genera cambios fisiológicos que protegen a las células. Por

Introducción
9
último, la existencia de un fenotipo especifico del biofilm asociado a la expresión de
mecanismos que contrarrestan los efectos de los biocidas (99).
3. Sistemas de “Quorum sensing”
El “quorum sensing” (QS) es un mecanismo de comunicación célula-célula mediante
el cual las bacterias regulan, en forma concertada, la expresión de ciertos genes en
respuesta a la densidad celular. Las bacterias sintetizan una molécula difusible llamada
autoinductor que les permite sensar la densidad poblacional activando un regulador
específico que modifica la expresión de genes. El sistema de QS regula una amplia
variedad de funciones fisiológicas como la esporulación, los mecanismos de conjugación,
movilidad, la formación de biofilms y la virulencia (104). En bacterias Gram-negativas los
autoinductores son moléculas de bajo peso molecular de diversa naturaleza química pero
los mas difundidos y estudiados son los acilos de homoseril lactona (acil-HSL), mientras
que en bacterias Gram-positivas los autioinductres son péptidos (116).
El sistema de QS de Vibrio fischeri fue el primero que se describió y es el modelo
prototipo en bacterias Gram-negativas (63). Consta de dos proteínas LuxI encargada de
sintetizar el autoinductor N-(3-oxohexanoil)-homoserinolactona y LuxR encargada de la
percepción de la señal. Cuando hay baja densidad celular se sintetizan pequeñas
cantidades del autoinductor, por lo tanto la producción de luz no es evidente. Cuando
aumenta la densidad celular, la concentración del autoinductor aumenta hasta alcanzar un
umbral a partir del cual se une a LuxR y el complejo se une a la región promotora del
operon luxCDABE e inducen la expresión de luciferasa. Además la expresión de luxI
también es controlada positivamente por este sistema, lo que ocasiona un rápido aumento
de la cantidad de autoinductor (Figura 2).

Introducción
10
Figura 2. Sistema de QS de Vibrio fischeri, LuxI sintetiza el autoinductor acil-HSL y LuxR sintetiza
el regulador de respuesta. A baja densidad celular se sintetizan pequeñas cantidades del
autoinductor, por lo tanto no hay producción de luz. Cuando aumenta la densidad celular, el
autoinductor alcanza un umbral, se une a LuxR y el complejo se une a la región promotora del
operon luxCDABE e inducen la expresión de luciferasa. Además la expresión de luxI también es
controlada por este sistema. Disponible en
http://www.cheme.caltech.edu/groups/fha/old_website_2010.6.25/quorum.html
3.1. El sistema rpf/DSF
En X. campestris el sistema de QS no está mediado por un autionductor del tipo
acil-HSL como en la mayoría de las bacterias Gram-negativas. En esta especie el
autoinductor es un derivado de un ácido graso conocido como DSF (diffusible signal
factor) que fue identificado como ácido cis-11-metil-2-dodecenoico (148). Esta molécula o
derivados estructurales han sido encontrados en otras especies bacterianas como
Xanthomonas spp., Xylella fastidiosa y Burkholderia cenocepacia (17, 27, 76).
Los genes involucrados en este sistema de QS pertenecen al operon rpf (regulation
of pathogenicity factors) (48). La síntesis de DSF depende en parte de la proteína RpfB,
una ligasa de ácidos grasos de cadena larga, y completamente de la proteína RpfF, cuya

Introducción
11
secuencia de aminoácidos comparte similitud con una enoil-CoA hidrolasa involucrada en
el metabolismo de los ácidos grasos (13, 31, 135).
La transducción de la señal es llevada a cabo a través de la proteína RpfC que tiene
un dominio histidina quinasa y otro histidina fosfotransferasa (135), y la proteína RpfG que
tiene un dominio c-terminal del tipo HD-GYP que hidroliza di-GMPc. El mecanismo
funciona de la siguiente manera: a baja densidad celular hay poco DSF, en este contexto,
RpfC interacciona con RpfF lo que conduce a la baja producción de DSF. Además la
proteína RpfG se encuentra desfosforilada, en consecuencia el dominio HD-GYP está
inactivo y los niveles de di-GMPc se mantienen elevados. La proteína Clp es un regulador
transcripcional que cuando se encuentra unida a di-GMPc presenta una conformación que
no le permite unirse al ADN para activar la transcripción de genes que codifican por ej.
enzimas extracelulares y EPS. Por el contrario, cuando hay alta densidad celular, RpfC
interactúa con DSF y se autofosforila, lo que produce la liberación de RpfF y el incremento
de la producción de DSF. Además RpfC fosforila a RpfG y activa su dominio HD-GYP lo
que conlleva a la disminución de los niveles de di-GMPc. En estas condiciones Clp activa
la transcripción de genes regulados por QS (Figura 3). Sin embargo, existen controversias
sobre el efecto en la formación o dispersión de biofilms (136).

Introducción
12
Figura 3. El sistema de QS de X. campestris pv. campestris modula los niveles de di-GMPc. La
señal DSF es sintetizada por RpfF y sensada por la histidina quinasa RpfC unida a la membrana
citoplasmática. a) baja densidad celular: La concentración de DSF es baja, RpfC interactúa con
RpfF,y disminuye su actividad catalítica. En esta condición, RpfG se encuentra defosforilada e
inactiva, por lo tanto, el nivel del di-GMPc es alto y une al activador transcripcional Clp impidiendo
su unión al ADN. b) alta densidad celular: La concentración de DSF aumenta, RpfC se une a DSF,
se autofosforila y activa la actividad fosfodiesterasa de RpfG, lo cual conduce a la disminución de
di-GMPc que se transforma en GMP. RpfF ya no está unido a RpfC y se produce más DSF. La
disminución de di-GMPc activa el regulador Clp que a su vez activa la transcripción de distintos
genes. Figura adaptada de Srivastava y waters, 2012 (136).
3.1.1. El sistema de señalizacion rpf/DSF de S. maltophilia
El genoma de S. maltophilia K279a codifica un sistema de QS estrechamente
relacionado al sistema basado en DSF descripto en Xanthomonas campestris pv.
campestris, (Figura 4) y se diferencia de este en que no presenta un gen homologo a rpfH
(62).

Introducción
13
Figure 4. Organización del grupo de genes rpf de Xanthomonas campestris pv. campestris y S.
maltophilia K279a, tomado de Fouhy et al. (62).
Se ha detectado actividad de DSF en varias cepas de S. maltophilia. Huang y Wong
(82) encontraron que la actividad de DSF en la cepa WR-C se debe a un grupo de ácidos
grasos relacionados estructuralmente, el acido cis-11-metill-2-dodecenoico (al igual que
en X. campestris) y siete derivados mas.
Fouhy et al. (62) demostraron que la disrupción del gen rpfF en S. maltophilia K279a
produce efectos pleiotrópicos. La mutante rpfF posee una severa disminución en la
motilidad tipo “swimming”, menor actividad de proteasa y un perfil alterado de LPS, y
además forma agregados cuando crece en el medio L. Además, no es virulenta en un
modelo de nematodo.
4. El hierro en la fisiología bacteriana
El hierro es un nutriente esencial para los microorganismos, es cofactor de
proteínas, enzimas, actúa como trasportador de electrones y regula la expresión génica
en muchas especies bacterianas. En su forma reducida, Fe2+, es relativamente soluble
(0,1 M a pH 7,0) pero en presencia de oxigeno es oxidado a Fe3+ y precipita en forma de
hidróxidos insolubles dificultando la incorporación de este nutriente (5, 109). Además, en
el organismo hospedador la biodisponibilidad del hierro es escasa ya que se encuentra
unido a la hemoglobina, almacenado intracelularmente en forma de ferritina o quelado por
la transferrina en el suero.

Introducción
14
4.1. Sistemas de captación de hierro
La baja biodisponibilidad de hierro dificulta el crecimiento de los microorganismos,
por lo tanto éstos han desarrollado diferentes estrategias para captarlo, una de ellas
consiste en disminuir el pH del medio para incrementar la solubilidad del ion férrico, otra
se basa en reducir el Fe3+ a Fe2+ que es mas soluble, y otra se basa en sintetizar y
secretar sideróforos, compuestos que quelan hierro con alta afinidad e interactúan con un
receptor que internaliza el complejo hierro-sideróforo formado (67).
4.1.1. Sistema captación de hierro basado en sideróforos de
bacterias Gram-negativas
Los sideróforos son compuestos de bajo peso molecular (<1000 Da) con grupos
funcionales que quelan hierro con elevada afinidad. La mayoría de los sideróforos
descriptos son compuestos de tipo hidroxamato, catecol o sideróforos mixtos (78).
En las bacterias Gram-negativas, receptores específicos localizados en la
membrana externa, denominados receptores dependientes de TonB (57), unen los
complejos Fe3+-sideróforo, los cuales son internalizados por el sistema TonB. El complejo
transmembrana TonB-ExbB-ExbD (20) conecta el potencial de membrane citoplasmática
con el transporte de los siderofóros a través de la membrana externa (57). Además, en el
transporte participa el sistema de trasporte tipo ABC (ATP Binding Cassette) a través del
cual el sideróforo pasa desde el periplasma al citoplasma (Figura 5). Los sistemas de
captación de hierro y otros genes son reprimidos por Fur en diferentes especies
bacterianas (30).
En E. coli el sistema de transporte de sideróforos de tipo catecol es llevado a cabo a
través de tres receptores de membrana externa FepA, que transporta principalmente
enterobactina, Fiu y CirA que transportan amonabactina y dihidroxibenzoil serina
(monómero de enterobactina), respectivamente (Hantke,1990; Rabsch y Winkelmann,
1991).
En S. maltophilia poco se conoce sobre la producción de sideróforos y sistemas de
captación.

Introducción
15
Figura 5. Representación esquemática del sistema de captación de hierro mediado por sideróforos
en bacterias Gram-negativas. El complejo Fe3+-sideróforo es reconocido por receptores específicos
de la membrana externa. El complejo TonB-ExbB-ExbD conecta el potencial de membrana
citoplasmática con el transporte del complejo Fe3+-sideróforo a través de la membrana externa. El
complejo es transportado hacia el periplasma donde se une a proteínas, y en su pasaje hacia el
citoplasma participa el sistema de trasporte tipo ABC. Adaptado de Andrews et al. (5).
4.2. Hierro y estrés oxidativo
A pesar de la importancia biológica del hierro, las bacterias deben mantener su
concentración libre dentro de la célula en niveles seguros. El hierro libre actúa como
catalizador en las reacciones de Fenton y Haber-Weiss (142) que generan especies
reactivas del oxigeno (ERO) tales como radical hidroxilo (OH•), anión superoxido (O2
-) y
peróxidos (RO2). Estas ERO pueden oxidar lípidos, proteínas y ADN, entre ellas, el radical
hidroxilo generado en la reacción de Fenton es el mas reactivo.

Introducción
16
Reducción del hiero O2-+ Fe3+ Fe2+ + O2
Reacción de Fenton Fe2+ + H2O2 Fe3+ + OH- + OH•
Reacción de Haber Weiss O2-+ H2O
Catalizado por Fe OH- + OH•+ O2
Las bacterias con metabolismo aeróbico poseen enzimas antioxidantes como
catalasa, superóxido dismutasa y peroxidadas que les permiten deshacerse de estas
sustancias nocivas. Estas enzimas cumplen la función de defensa frente al estrés
oxidativo generado por el oxigeno del ambiente y por la respuesta inmune innata.
5. El hierro como señal: El sistema Fur
La proteína Fur (Ferric Uptake Regulator) forma un homodímero al unirse con el ion
ferroso. El complejo Fe2+-Fur reconoce secuencias de ADN específicas ubicadas en la
región promotora de ciertos genes conocidas como cajas Fur. La unión del complejo Fe2+-
Fur a dichas secuencias produce la represión de la transcripción de los genes que se
encuentran rio abajo ya que se impide el acceso de la ARN polimerasa (5). El
funcionamiento del sistema Fur se esquematza en la figura 6.
La principal función del sistema Fur es reprimir la expresión de genes implicados en
la captación de hierro cuando hay abundancia del mismo, de esta forma se puede
mantener su correcta concentración intracelular. No obstante se han descripto otras
funciones fisiológicas, por ej. en E. coli el sistema Fur reprime la producción de toxinas,
enzimas antioxidantes (manganeso superoxido dismutasa) y otros factores de virulencia
(55)

Introducción
17
Figura 6. Regulación de la expresión génica mediante el sistema Fur. La proteína Fur forma un
homodímero al unirse al Fe2+. El complejo Fe2+-Fur se une a secuencias de ADN específicas
conocidas como cajas Fur y reprime la transcripción de los genes que se encuentran rio abajo al
impedir el acceso de la ARN polimerasa.

Hipótesis y Objetivos

Hipótesis y Objetivos
19
1. Hipótesis
En S. maltophilia los genes relacionados con la formación de biofilms y con la
producción de otros potenciales factores de virulencia estarían regulados positivamente
por “quorum sensing”, y negativamente por concentraciones no limitantes de hierro, a
través del sistema Fur (“ferric uptake regulator”).
2. Objetivo general
Estudiar la regulación, por “quorum sensing” (QS) y/o por hierro, de la expresión de
productos génicos relacionados con la formación de biofilms y de la producción de otros
potenciales factores de virulencia.
3. Objetivos parciales
1- Tipificar aislamientos locales de S. maltophilia recuperados de infecciones asociadas
al uso de productos médicos. Evaluar la producción de potenciales factores de
virulencia y determinar la distribución del gen narG, codificante de nitrato reductasa
asociada al crecimiento en microaerofilia, y del gen codificante de la exoproteasa
StmPr en aislamientos locales de S. maltophilia.
2- Obtener mutantes Fur espontáneas de S. maltophilia.
3- Estudiar el sistema de captación de hierro de alta afinidad de S. maltophilia K279a:
caracterizar los sideróforos sintetizados por S. maltophilia K279a e identificar las
OMPs reguladas por la concentración de hierro.
4- Estudiar la función de DSF, señal de QS de S. maltophilia, en: a) la formación y
arquitectura de biofilms y la expresión de caracteres fenotípicos asociados a su
desarrollo (hidrofobicidad, motilidad “twitching”, producción de EPS) y b) en la
virulencia (producción de enzimas extracelulares, sensibilidad al estrés oxidativo),
utilizando un modelo de Galleria mellonella.
5- Evaluar el efecto de la limitación de hierro en la producción de DSF.
6- Estudiar la influencia de la concentración de hierro en la formación y arquitectura de
biofilms y en la virulencia. Estudiar el efecto de la limitación de hierro en la respuesta
al estrés oxidativo de biofilms de S. maltophilia (actividad de SOD y niveles de las
especies reactivas del oxígeno).

Materiales y Métodos

Materiales y Métodos
21
1. Cepas bacterianas
Para este estudio se utilizaron 63 aislamientos locales de S. maltophilia recuperados
de infecciones nosocomiales asociadas al uso de productos médicos en la Sección
Bacteriología del Hospital de Clínicas José de San Martín, Buenos Aires, durante el
periodo 2004-2010.
La cepa de origen clínico, S. maltophilia K279a, cuyo genoma ha sido secuenciado y
la mutante K279a rpfF que no produce DSF (62) se utilizaron para estudios comparativos.
Xanthomonas campestris pv. campestris 8004, y la cepa biosensora Xcc 8523 (mutante
rpfF, DSF- ) se utilizaron en el bioensayo descripto por Barber et al. (13). Estas cepas
fueron gentilmente cedidas por el Dr. Maxwell Dow (BIOMERIT Resarch Centre,
University college Corck, Ireland).
Todos los aislamientos fueron conservados en glicerol 15 % a -20 ºC.
2. Tipificación molecular
2.1. ERIC-PCR
La genotipificación se realizó mediante la técnica ERIC-PCR (secuencias consenso
repetitivas intragénicas de enterobacterias) utilizando el oligonucleótido ERIC-2 (Tabla 1)
(85). El ADN genómico se obtuvo a partir de una colonia de cada aislamiento cultivado en
agar tripteina soya (TSA) que fue resuspendida en 200 µl de agua destilada estéril,
hervida durante 10 min y centrifugada a 12000 rpm 3 min (34). La reacción en cadena de
la polimerasa se realizó en un volumen final de 50 µl que contenía 10 µl de buffer GoTaq
(Promega) 5X, MgCl2 4 mM, dimetilsulfoxido al 10 %, 400 µM de cada nucleótido, 60
pmoles del oligonucleótido cebador ERIC-2, 2 U de GoTaq polimerasa (Promega) y 3 µl
de ADN genómico. La reacción de amplificación se realizó en un termociclador Biometra
TPersonal, el ciclo de amplificación utilizado consistió en un intervalo inicial de 2 min a 94
°C y 30 ciclos de 30 seg a 94 °C, 1 min a 50 °C, 4 min a 72 °C, seguido de una extensión
final de 7 min a 72 °C. En cada experimento se incluyó un control negativo al que se le
agregó agua en lugar de ADN molde. Los productos de amplificación se analizaron por
electroforesis en geles de agarosa al 1,5 % con un marcador de peso molecular de 1 kb.
Para la visualización se utilizó la tinción con bromuro de etidio y las imágenes fueron
capturadas utilizando el transiluminador Molecular Imager® Gel Doc™ XR (Bio-Rad). Los

Materiales y Métodos
22
patrones de bandas se analizaron visualmente y se consideraron idénticos cuando la
posición de las bandas coincide sin importar la intensidad. Se utilizó además el
dendograma basado en el algoritmo UPGMA obtenido mediante el programa TREECON
versión 1.3 b para Windows.
2.2. REP-PCR
Se utilizó la técnica REP-PCR (secuencias repetitivas palindrómicas extragénicas)
con los oligonucleótidos REP-1 y REP-2 (Tabla 1). El ADN genómico se obtuvo de la
misma forma que para ERIC-PCR. La reacción se realizó en un volumen final de 50 µl que
contenía 10 µl de buffer GoTaq (Promega) 5X, MgCl2 2,5 mM, dimetilsulfoxido al 10%, 400
µM de cada nucleótido 60 pmoles de cada oligonucleótido cebador, 2 U de GoTaq
polimerasa (Promega) y 3 µl de ADN genómico. La reacción de amplificación se realizó en
un termociclador Biometra TPersonal, el ciclo de amplificación utilizado consistió en un
intervalo inicial de 5 min a 95 °C y 35 ciclos de 1 min a 94 °C, 1 min a 45 °C, 6 min a 68
°C, seguido de una extensión final de 18 min a 68 °C. En cada experimento se incluyó un
control negativo. Los productos de amplificación se analizaron del mismo modo que ERIC-
PCR.
3. Producción de potenciales factores de virulencia
3.1. Detección de la presencia de los genes narG y stmPr-1 en
aislamientos locales de S. maltophilia
La detección de los genes narG y stmPr-1 se realizó mediante la reacción en
cadena de la polimerasa utilizando oligonucleótidos diseñados en base al genoma
publicado de K279a (Tabla 1). Los mismos fueron contrastados con otros genomas
publicados para que eventuales polimorfismos no influyan negativamente en la
amplificación. El ADN genómico se obtuvo como se describió en el punto 2.
Para el gen narG, la reacción en cadena de la polimerasa se realizó en un volumen
final de 25 µl que contenía 5 µl de buffer GoTaq (Promega) 5X, MgCl2 2,5 mM,
dimetilsulfoxido al 10 %, 400 µM de cada nucleótido 60 pmoles de cada oligonucleótido
cebador, 1 U de GoTaq polimerasa (Promega) y 4 µl de ADN genómico. La reacción de
amplificación se realizó en un termociclador Biometra TPersonal, el ciclo utilizado
consistió en un intervalo inicial de 5 min a 95 °C y 35 ciclos de 1 min a 95 °C, 30 seg a 50
°C, 1 min a 72 °C, seguido de una extensión final de 6 min a 72 °C.

Materiales y Métodos
23
Para el gen stmPr-1, la reacción se realizó en un volumen final de 25 µl y la mezcla
de reacción se preparó como se describió para el gen narG. El ciclo utilizado consistió en
5 min a 95 °C, 30 ciclos de 1 min a 95°C, 30 seg a 48 °C, 1 min a 72 °C, seguido de una
extensión final de 6 min a 72 °C.
En cada experimento se incluyó un control negativo al que se le agregó agua en
lugar de ADN molde. Los productos de amplificación se analizaron por electroforesis en
geles de agarosa al 1,5 % con un marcador de peso molecular de 1 kb. Para la
visualización se utilizó la tinción con bromuro de etidio.
Tabla 1. Oligonucleótidos utilizados en este estudio
Gen Secuencia Origen
ERIC-2 5’-AAGTAAGTGACTGGGGTGAGCG-3’ Hulton et al. (85)
REP-1 5`-IIIGCGCCGICATCAGGC-3` Seral et al.
REP-2 5`-ACGTCTTATCAGGCCTAC-3`. (132)
narG F 5´-GGCTTGAGCACGATGCGGGT-3´
F 5´-GTGGGCAAGGAGCACGAGGC-3´
Este estudio
stmPr -1 F 5´-CGTGCCAGCTTCTCCAACTA-3´
F 5´-AGGACTGTTGATGGTGCAGG-3´
Este estudio
fur F 5´-GGCGGTTGGGGAAATCAAAC-3´
R 5´-CAACGAAAAACCCCGGGCA-3´
Este estudio
3.2. Prueba de reducción de nitratos
Con esta prueba se detecta la respiración anaeróbica que utiliza nitrato como
aceptor final de electrones. Los nitratos se reducen a nitritos y algunas bacterias reducen
los nitritos a nitrógeno gaseoso (N2) por la presencia de las enzimas nitrato y nitrito
reductasa, respectivamente. Con un cultivo fresco del microoganismo a ensayar, se

Materiales y Métodos
24
inoculó el Caldo nitrato (ver anexo I) y se incubó a 35 °C, 24 h. Para determinar si el
nitrato ha sido reducido a nitrito, después de la incubación, se añadieron al medio de
cultivo, 2 gotas de solución A, y luego 2 gotas de la solución B del reactivo de Griess (ver
anexo I). La aparición de una coloración rosada o roja indica que los nitratos han sido
reducidos a nitritos. Si la reacción es negativa, se añade una muy pequeña cantidad de
polvo de Zinc. La aparición de una coloración rosada o roja confirma la negatividad de la
reacción; en caso de que esto no suceda, significa que los nitratos han sido reducidos a
N2 gaseoso.
3.3. Proteasas
Se determinó la actividad proteolítica utilizando placas de agar nutritivo
suplementado con leche descremada 1,5 % (p/v) (83). El inoculo se preparó a partir de
cultivos en caldo TSB de 20 h de incubación. Las células fueron centrifugadas y
resuspendidas en el mismo medio hasta obtener aproximadamente 108 ufc/ml y se
sembraron 5 µl de esta suspensión en la placa con leche. Luego de 24 h de incubación a
30 °C, la actividad proteolítica fue determinada midiendo el halo de proteólisis alrededor
de la colonia.
3.4. Lipasa
Se determinó la actividad lipolítica utilizando placas con agar base tibutirina
suplementado con glicerol tributirato 1 % (Merck).Se realizó una estría de las cepas a
estudiar a partir de cultivos en TSA de 18 h de incubación y se incubaron a 35 °C durante
24 h (83).
3.5. DNasa
Se determinó la actividad de DNasa utilizando placas conteniendo agar nutritivo y
ácido desoxirribonucleico. Se realizó una estría de las cepas a estudiar a partir de cultivos
en TSA de 18 h de incubación. Después de la incubación de 24 h la placa se inunda con
ácido clorhídrico 1 %, que causa la precipitación del ADN polimerizado y hace al medio
opaco. Los organismos que degradan el ADN producen una zona transparente alrededor
de la estría (149).
3.6. Resistencia al estrés oxidativo
Ensayo de difusión en placa

Materiales y Métodos
25
La sensibilidad al peróxido de hidrogeno se determinó por el ensayo de difusión (73)
con algunas modificaciones. Cultivos de 24 h en TSB se estandarizaron a una DO546 de
0,300, una alícuota de 300 µl se utilizó para inocular 30 ml de TSB con 0,7 % de agar
termostatizado a 45 °C, 3 ml de esta suspensión fueron transferidos a una placa de Petri
que contenía 16 ml de TSA. Discos de papel de filtro (Whatman) estériles de 7 mm de
diámetro fueron impregnados con 3 µl de una solución de H2O2 12 % (Merck). La
sensibilidad al peróxido de hidrogeno fue determinada midiendo el halo de inhibición
alrededor del disco luego de la incubación durante 24 h a 37 °C. Los experimentos fueron
realizados por triplicado.
Ensayo de supervivencia frente a H2O2
Cultivos de 18 h incubados con agitación fueron diluidos con TSB hasta obtener una
densidad celular de aproximadamente 5x107 a 1x108 ufc/ml. Previo al tratamiento con
H2O2 de determinó el número inicial de ufc/ml (N0), para esto a una alícuota del cultivo se
le realizaron diluciones seriadas en solución fisiológica y se espatuló 100 µl de las
diluciones apropiadas en placas conteniendo TSA. Luego de la determinación del N0 los
cultivos fueron tratados con H2O2 20 mM (Merck) e incubados inmediatamente con
agitación a 35 °C. A los 15 y 30 min de incubación se tomaron alícuotas y se realizaron
recuentos de viables utilizando solución fisiológica con tiosulfato de sodio 0,2 % para
inactivar el H2O2. Las diluciones apropiadas fueron espatuladas en placas conteniendo
TSA e incubadas a 35 °C. Luego de la incubación se determino el número de ufc/ml y el
efecto del tratamiento con H2O2 se expresó como el porcentaje de sobrevivientes
relacionado al número inicial de microorganismos (54).
3.7. Producción de sideróforos
Como método de tamizaje se empleó el ensayo universal Cromoazurol S (CAS) en
placa descripto por Schwyn y Neilands (128), modificado por Armstrong et al. (8) que
utiliza como medio de cultivo Stainer-Scholte sin suplemento con agar al 1,5 %. A este
medio se le agregó Casamino ácidos 0,1 % (p/v) (SSC) para favorecer el crecimiento de
S. maltophilia. El método se basa en la incorporación del colorante cromoazurol S en el
medio agarizado para formar complejos estables con el hierro, generando un color verde-
azul. Cuando un quelante con mayor afinidad, como el sideróforo, remueve el hierro del
complejo, el medio se vuelve de color al naranja. Para preparar el medio de cultivo, un

Materiales y Métodos
26
volumen de 100 ml del reactivo CAS-HDTMA-Fe (ver anexo I) se agregó asepticamente a
900 ml del medio SSC con 1,5 % de agar-agar, pH 7,00 previamente esterilizado. Cuando
fue necesario generar condiciones de limitación de hierro se agregó el quelante de hierro
2,2’ dipiridilo (Dip) 100 µM. Cada placa contenía 20 ml de medio. Cultivos de 18 h en TSB
se estandarizaron a una DO546 de 0,300 y se utilizaron para inocular las placas de agar
CAS con escarbadientes estériles. Las placas inoculadas se incubaron a 35 °C por 48 h.
La producción de sideróforos se evidenció como un halo naranja alrededor de la colonia
3.8. Determinación de la naturaleza química de sideróforos
3.8.1. Condiciones de cultivo
Los aislamientos de S. maltophilia fueron cultivados en el medio SSC suplementado
con Dip 100 µM o con FeCl3 100 µM durante 48 h con agitación (200 rpm). Luego de la
incubación se llevaron a una DO546 de 1,00 y los respectivos sobrenadantes fueron
obtenidos por centrifugación a 10.000 rpm 5 min.
3.8.2. Ensayo de Arnow
La presencia de compuestos tipo catecol en los sobrenadantes de cultivo se
determinó por el ensayo de Arnow (25). Para este ensayo a 1 ml de sobrenadante se
agregaron los siguientes reactivos en orden: 1 ml de HCl 0,5 N, 1 ml nitrito-molibdato
(cantidades de c/u) y 1 ml NaOH 1,0 N. Los compuestos de tipo catecol producen un
compuesto de color rosa.
3.8.3. Ensayo de Csáky
La presencia de compuestos tipo hidroxamato en los sobrenadantes se determinó
por el ensayo de Csáky (25). En este ensayo 1 ml de sobrenadante es hidrolizado con 1
ml de H2SO4 6 N a 130 °C por 30 min. Luego se agregan 3 ml de acetato de sodio, 1 ml
de ácido sulfanilico y 0,5 ml de iodo. Luego de 3-5 min el exceso de iodo es eliminado con
1 ml de arsenito de sodio. Finalmente se agrega 1 ml de α-naftillamina y luego de 20-30
min aparece un color rosa si hay presentes compuestos tipo hidroxamato. Bordetella
bronchiseptica 7865 y Acinetobacter baumannii ATCC19606 se emplearon como
controles de cepas productoras de sideróforos tipo hidroxamato y catecol
respectivamente.

Materiales y Métodos
27
4. Curvas de Crecimiento
Las curvas de crecimiento en medio líquido se realizaron en TSB y se incubaron a
35 °C en condiciones estáticas y con agitación (150 rpm). Para la preparación del inoculo
se partió de placas de TSA de 18 h de incubación y se realizó una suspensión en TSB de
DO540 de 0,300. Alícuotas de 100 µl se utilizaron para inocular tubos con 5 ml de TSB. El
crecimiento se siguió espectrofotométricamente a 450 nm (Metrolab 1700).
5. Bioensayo para determinar la producción DSF
Se utilizó una modificación del método desarrollado por Barber et al. (13) basado en
la recuperación de la actividad endoglucanasa, enzima regulada por “quorum sensing”, de
una mutante de Xanthomonas campestris pv. campestris rpfF (Xcc 8523).
5.1. Extracción de DSF de sobrenadantes de cultivo
Los compuestos presentes en el sobrenadante de cultivo fueron extraídos con las
columnas Oasis MAX 30 mg de soporte (Waters Oasis® Milford, USA) de extracción en
fase solida que se basan en el principio combinado de intercambio aniónico e interacción
hidrofóbica. Se realizaron cultivos en 20 ml del medio SSC y SSC suplementado con
FeCl3 100 µM y se incubaron a 35 °C por 48 h con agitación. Los cultivos fueron
centrifugados a 10000 rpm por 10 min, los sobrenadantes se acidificaron hasta pH 3,00
con ácido clorhídrico. Luego, se utilizaron 8 ml de cada sobrenadante para la extracción
en fase solida. Las columnas fueron acondicionadas haciendo pasar 2 ml de metanol y
luego 2 ml de agua destilada estéril. Luego se pasaron 8 ml del sobrenadante acidificado
y para el lavado se pasaron 3 ml de solución de acetato de sodio 50 mM:metanol (95:5).
La elución se realizó con 2 ml de metanol y luego con 2 ml de de solución de acido
acético al 2 % en metanol. El extracto obtenido se evaporó a sequedad y se resuspendió
en 500 µl de agua estéril para ser utilizado en el bioensayo.
5.2. Bioensayo
Las células de Xcc 8523 provenientes de cultivos de 18 h de incubación en el medio
NYGB fueron centrifugadas, lavadas y resuspendidas en el medio MMXC hasta una
concentración de aproximadamente 5x107 ufc/ml. El bioensayo se realizó en 1 ml de
MMXC al que se le agregó 100 µl de esta suspensión y 100 µl del extracto de
sobrenadante de cultivo del cual se quiere determinar la concentración de DSF. Las
mezclas fueron incubadas por 22 hs a 28 °C. Previa medición de la DO540, las células

Materiales y Métodos
28
fueron removidas por centrifugación. La actividad relativa de endoglucanasa fue
determinada sembrando alícuotas de 50 µl del sobrenadante en pocillos de 6 mm hechos
en placas con 20 ml de agar carboximetilcelulosa. Después de incubar a 37 °C por 20 h,
las placas fueron teñidas con rojo congo 0,1 % durante 30 min y reveladas con una
solución de NaCl 1M. Se midió el diámetro de los halos de cada condición, se cuantificó la
actividad mediante la utilización de una curva estándar realizada con diluciones de
concentración conocida de Celulasa I (SIGMA). Como control negativo se realizó el
ensayo en las mismas condiciones pero en lugar de agregar 100 µl del extracto se agregó
agua. Como control positivo se utilizó una suspensión estandarizada de la cepa salvaje
Xcc 8004 en lugar de utilizar la mutante Xcc 8523 y se realizó el ensayo en las mismas
condiciones utilizando agua en lugar del extracto.
6. Formación de biofilms
La formación de biofilms se estudió bajo las siguientes condiciones: se realizaron
cultivos en caldo TSB y TSB suplementado con DSF comercial 200 µM (Santa Cruz
Biotechnology Inc.) o con el quelante 2,2’ dipiridilo (Dip) 200 µM según se indique. La
incubación se realizó en condiciones estáticas a 35 ºC durante 48 h.
6.1. Estandarización del inoculo
Los inóculos se estandarizaron en base a la densidad óptica (DO540) equivalente a
1x108 ufc/ml aproximadamente (117). Las suspensiones estandarizadas fueron utilizadas
para inocular el medio de cultivo apropiado realizando una dilución 1/100. Todos los
ensayos se realizaron al menos tres veces.
6.2. Ensayo en Tubos
La formación de biofilms en tubos se realizó como describieron Passerini de Rosi et
al (117). Alícuotas de 2 ml de TSB inoculado como se describió en el punto anterior fueron
transferidas asépticamente a tubos de borosilicato (BS) y polipropileno (PP) e incubadas
con agitación durante 6, 24 y 48 h. Para cada condición, se utilizaron 8 tubos. Luego de la
incubación 2 tubos fueron utilizados para estimar el crecimiento midiendo la DO450nm
previa homogeneización. Los otros seis tubos fueron utilizados para estimar la biomasa
del biofilm formado. A dichos tubos se les aspiro el cultivo y las células adheridas fueron
lavadas con agua destilada y teñidas con 2 ml de CV (0,01 %) por 30 min. El biofilm

Materiales y Métodos
29
formado fue visualizado y posteriormente cuantificado espectrofotométricamente a 456
nm (Metrolab 1700) mediante la elución del CV adherido con 2 ml etanol.
6.3. Ensayo en microplacas
Los ensayos en microplacas de poliestireno de 96 pocillos (Greiner Bio-One) fueron
realizados como describieron O’Toole y Kolter (111) con algunas modificaciones (117).
Los pocillos de las microplacas se llenaron con 180 µl del medio de cultivo inoculado, que
fue preparado como se describió en el punto anterior. Luego de la incubación por 48 h a
35 °C, se determinó la DO546 del crecimiento y se eliminaron las células planctónicas. El
biofilm formado en los pocillos se lavó con PBS y se tiñó con cristal violeta (CV) 0,01 %.
La cuantificación de la biomasa (células adheridas y matriz extracelular) se realizó
mediante la elución del CV con 200 µl de etanol. La lectura de la DO546 se realizó en un
lector de ELISA (Metertech 960). Cada aislamiento se ensayó por octuplicado.
6.4. Microscopia óptica
Para la observación microscópica los biofilms fueron formaron en cubreobjetos de
borosilicato. Cubreobjetos estériles fueron transferidos asépticamente a placas de Petri
conteniendo TSB inoculado como se describió anteriormente y se incubaron durante 48 h
a 35 °C. Luego fueron lavados con agua destilada, teñidos con CV (0,1 %) durante 10 min y
lavados nuevamente. Los cubreobjetos montados sobre portaobjetos fuero visualizados
en el microscopio Olympus BX51 con magnificación de 400X. Las imágenes fueron
capturadas con la cámara digital Q color 3 y procesadas con Image-Pro plus 6.2 sofware
en la Cátedra de Inmunología.
7. Microscopia confocal laser de barrido.
La arquitectura del biofilm de S. maltophilia se estudió mediante microscopia
confocal laser de barrido. Para este propósito los biofilms se formaron en unas cámaras
de cultivo especiales con base de borosilicato de 100 µm de espesor (Lab-Tek Nunc; N°
155411). Las cámaras se llenaron con 400 µl del medio de cultivo inoculado que fue
preparado como se describió en el punto 6.1. Luego de la incubación por 48 h a 35 °C se
eliminaron las células planctónicas y se lavó el biofilm con solución fisiológica. La
visualización se realizó por tinción con colorante fluorescente Syto9® (Molecular probes)
2,5 µM disuelto en solución fisiológica. Luego de la inciubacion por 30 min con el
colorante, las cámaras se examinaron en un microscopio confocal Carl Zeiss LSM510-

Materiales y Métodos
30
Axiovert 100 M con objetivo de 40X de inmersión en agua. Para la excitación de la
muestra se utilizó el laser de argón de 488 nm y la emisión se midió entre 505-530 nm.
Todas las imágenes fueron tomadas escaneando secuencialmente cada 10 µM.
Las imágenes fueron analizadas con el programa ZEN 2009 Light Edition de Carl
Zeiss mientras que las reconstrucciones tridimensionales fueron realizadas con el
programa ImageJ. Las imágenes fueron tomadas en el servicio de micoscopia confocal
del Instituto de Investigaciones Biomédicas (BIOMED) Pontificia Universidad Católica
Argentina (UCA)-CONICET.
8. Movilidad asociada a superficies o “twitching”
La movilidad asociada a superficies se estudió en medio LB solidificado con agar-
agar 1 %. Se realizaron suspensiones de DO546 1,0 de las cepas a estudiar que se
utilizaron para inocular las placas de twitching por punción. Luego de la incubación se
removió el agar y se tiñeron las células adheridas con CV 0,1 % (130). Las zonas de
movilidad fueron medidas y, además, visualizadas microscópicamente a una
magnificación de 400X.
9. Producción de sustancias poliméricas extracelulares (EPS)
La producción de EPS fue determinada como describieron Boon et al (17) con
algunas modificaciones. A partir de cultivos en caldo LB de 24 h se realizaron
suspensiones de DO540 0,200, y alícuotas de 100 µl de éstas se utilizaron para inocular
erlenmeyers con 100 ml de caldo LB suplementado con glucosa 0,1 % (LB-glu 0,1 %). Los
cultivos fueron incubados a 35 °C en agitación (200 rpm) durante 48 h y se obtuvieron los
sobrenadantes por centrifugación a 14000 rpm durante 20 min mientras que la biomasa,
obtenida en el precipitado, fue secada a 56 °C. Luego de filtrar los sobrenadantes (0,45
µM) se precipitaron las EPS mediante el agregado de 2 volúmenes de etanol absoluto y
posterior incubación a -20 °C durante 24 h. Las EPS precipitadas se obtuvieron por
centrifugación a 10000 rpm durante 10 min. Finalmente el precipitado obtenido fue secado
a 56 °C hasta peso constante para luego determinar el peso seco en balanza analítica.
Los resultados fueron expresados como la cantidad de EPS (µg) relacionado al peso seco
de la biomasa (mg). El contenido de proteínas totales fue cuantificado con el reactivo de
Bradford (Sigma Aldrich) y utilizó seroalbumina bovina (Sigma Aldrich) como patrón. El

Materiales y Métodos
31
contenido de carbohidratos total fue cuantificado con el método de fenol y acido sulfúrico
utilizando glucosa como estándar (19, 50).
10. Espectroscopia de reflexión total atenuada infrarroja con
transformada de Fourier (ATR-FTIR)
El precipitado de EPS obtenido como se describió en el punto anterior fue
resuspendido con una relación de 20 µl de agua Milli-Q por cada mg de EPS y se
realizaron las diluciones necesarias hasta obtener una señal que no sature el sistema.
Alícuotas de 150 µl de las suspensiones fueron transferidos a celdas ópticas de ZnSe y
posteriormente secados hasta obtener una delgada película sobre la superficie.
El análisis por ATR-FTIR fue realizado empleando el espectrofotómetro Nicolet 380
FT-IR (Thermo Scientific Electron Corporation). Los espectros de absorción de las
muestras fueron adquiridos en el intervalo de lectura de 4000 a 650 cm-1 con una
velocidad de medición de 1 cm/seg realizándose 32 escaneos por espectro y empleando
una resolución de 4 cm-1. Estos parámetros fueron ajustados por medio del programa
OMNIC (Thermo Scientific Electron Corporation).
11. Aislamiento de mutantes Fur
Para aislar mutantes espontaneas Fur de S. maltophilia se utilizó la técnica de
selección por resistencia al manganeso descripta por Hantke con algunas modificaciones
(69). Cultivos de K279a en caldo LB de 18 h de incubación fueron espatulados en placas
que contenían MnSO4 20 mM y Dip 200 µM. Luego de 72 h de incubación las colonias
resistentes al manganeso que crecieron en el medio fueron cultivadas en placas de agar
CAS suplementadas con FeCl3 20 mM para detectar la mutantes Fur ya que éstas
producen sideróforos constitutivamente. La mutación en el gen fur fue confirmada por
secuenciación del gen completo junto a una región de 100 nucleótidos rio arriba del codón
de inicio. Los oligonucleótidos utilizados para tal fin fueron diseñados en base al genoma
publicado de K279a (Tabla 1). La secuenciación del ADN fue realizada en la Unidad de
Genómica del Instituto de Biotecnología, C.I.C.V.y A. Instituto Nacional de Tecnología
Agropecuaria (INTA), Castelar. Las secuencias fueron analizadas con los programas
BioEdit sequence ailgnmet editor y MEGA versión 5.

Materiales y Métodos
32
12. Ensayo de reducción del Azul de Nitrotetrazolio (NBT)
Este ensayo permite evaluar las especies reactivas del oxigeno (ERO) mediante la
utilización NBT oxidado (de color amarillo) que, en presencia de anión superoxido, es
reducido a azul de formazán, un compuesto de color azul, que pude ser evaluado por
espectrofotometría. Las ERO se determinaron en biofilms que se formaron en microplacas
de PS de 96 pocillos cultivaos en TSB en presencia o ausencia de Dip 200µM. Se
utilizaron 180 µl del medio de cultivo inoculado (preparado como se describió en el ensayo
de formación de biofilms) para llenar 12 pocillos de la microplaca por cada condición.
Luego de la incubación de 48 h a 35 °C, se determinó la DO546 del crecimiento y se
eliminaron las células planctónicas. El biofilm formado en los pocillos se lavó con PBS y 6
de ellos fueron teñidos con cristal violeta (CV) 0,1 % los restantes 6 pocillos se reservaron
para la reacción con NBT. La cuantificación de la biomasa (células adheridas y matriz
extracelular) se realizó mediante la elución del CV con 200 µl de etanol. La formación de
biofilms se evaluó con la relación DO546 del CV/ DO546 del cultivo (biomasa relativa del
biofilm). Por otra parte los biofilms formados en los restantes 6 pocillos se trataron con
200 µl de NBT 0,5 mg/ml, (Sigma Aldrich) y luego de la incubación durante 30 minutos a
37 °C en oscuridad, se frenó la reacción agregando 20 µl de HCl 0,1 N, finalmente se
agregaron 50 µl dimetilsulfóxido para extraer el NBT reducido. El producto de la reacción
se midió espectrofotométricamente a 546 nm. Los niveles de ERO se expresaron en
relación a la biomasa relativa del biofilm (2, 7).
13. Actividad de la enzima superoxido dismutasa (SOD)
La actividad de la enzima SOD se determinó con el sistema Riboflavina/metionina y
NBT (15). En presencia de luz, la riboflavina pierde un electrón y genera anión superoxido
(O2.-). Debido a que la enzima SOD actúa disminuyendo la concentración de O2
.-, a mayor
actividad SOD, menor es la reducción del NBT. Por lo tanto, se definió una unidad de
actividad SOD como la cantidad de enzima requerida para disminuir en un 50 % la
producción de NBT reducido.
13.1. Obtención de extractos crudos
Las células bacterianas provenientes de cultivos planctónicos de 25 ml fueron
obtenidas por centrifugación, lavadas y resuspendidas en 7 ml de PBS con el inhibidor de
porteasas PMSF 0,25 mM (Fluka Biochemika). Las suspensiones fueron sonicadas por 6

Materiales y Métodos
33
min en dos pulsos de 3 min (Vibra cell, Sonics & materials inc.) y centrifugadas a 10000
rpm durante 10 min a 4°C. La concentración de proteínas se determinó con el reactivo de
Bradford (Sigma Aldrich). Se utilizó seroalbumina bovina (Sigma Aldrich) como patrón
(19).
13.2. Actividad en cultivos planctónicos
Alícuotas de 100 µl de cada extracto crudo fueron tratadas con 300 µl de metionina
13 mM (Sigma Aldrich), 100 µl de NBT 1 mg/ml, 300 µl de EDTA 100 nM y 300 µl de
riboflavina (Sigma Aldrich) en presencia de luz. Luego de 15 min se determinó la DO560.
La actividad se expresó como unidades de SOD por mg de proteínas totales.
13.3. Actividad en el biofilm
Para este propósito los biofilms fueron formados en microplacas de 12 pocillos
(Greiner Bio-One) con 3 ml de cultivos estandarizados como se describió en el ítem 6.1.
Luego de la incubación a 35 °C por 48 h, se eliminó el medio de cultivo y los biofilms se
lavaron con PBS. A cada pocillo se le agregó 1 ml de PBS y para despegar las células
adheridas se raspó la superficie y las paredes con un cepillo. Así se obtuvo el material de
10 pocillos por cada tratamiento que luego fueron centrifugados y resuspendidos en 1 ml
de PBS con PMSF 0,25 mM. Las suspensiones fueron sonicadas con un microtip por 6
min en dos pulsos de 3 min (Branson Sonifier 250) y centrifugadas a 10000 rpm durante
10 min a 4 °C. La concentración de proteínas se determinó como se describió en el ítem
13.1
13.4. Geles nativos para SOD y determinación del tipo de isoenzima
Las proteínas presentes en extractos obtenidos como se describió en el ítem 13.1
fueron separadas en geles de poliacrilamida en condiciones no desnaturalizantes en
presencia de Tris 50 mM, glicina 300 mM y EDTA 1,8 mM a 120 voltios. En cada calle se
cargó 20 µg de proteínas totales y se utilizó como control Mn-SOD purificada de E. coli
(Sigma Aldrich). La actividad SOD fue visualizada por la tinción con NBT descripta por
Beauchamp y Fridovich (15). Luego de la electroforesis, los geles fueron lavados con
agua destilada e incubados durante 30 min en oscuridad y agitación con una solución de
PBS 50 mM, pH 7,80 que contenía NBT 500 µM, EDTA 1 mM, TEMED 20 mM (Invitrogen,
Carlsbad) y riboflavina 30 µM. Los geles fueron expuestos a la luz hasta la aparición de
bandas acromáticas en un fondo azul que indican actividad SOD. Para identificar el tipo
de isoenzima SOD, se utilizaron ensayos de inhibición. Los geles se trataron con NaCN

Materiales y Métodos
34
10 mM o H2O2 3,7 mM para inactivar la isoenzima Cu/Zn-SOD o Fe-SOD,
respectivamente (51). La resistencia a ambos tratamientos es característica de Mn-SOD.
Luego de incubar los geles con los inhibidores se tiñeron con NBT como se describió
anteriormente. Se utilizó como control Mn-SOD purificada de E. coli.
14. Evaluación de la virulencia con el modelo de infección de Galleria mellonella
Para evaluar la virulencia se utilizó el modelo de infección del lepidóptero Galleria
mellonella. Las larvas de G. mellonella fueron mantenidas en condiciones de temperatura
y humedad adecuada y alimentadas con una dieta artificial que consistió en salvado de
trigo 250 g, levadura seca 10 g, leche en polvo 25 g, miel 100 g, glicerina 50 g, nipagin 0,1
%, agua 100 ml esterilizada en autoclave. Se infectaron larvas de G. mellonella con
suspensiones bacterianas preparadas a partir de cultivos en TSA de 24 h. Algunas
colonias de cada cepa fueron resuspendidas en solución fisiológica estéril hasta una
DO540 de 0,300 equivalente a 108 ufc/ml. Esta suspensión y diluciones 1/10 y 1/100 se
utilizaron para inocular las dosis de 106, 105 y 104 ufc/larva respectivamente. Se
seleccionaron al azar grupos de 10 a 15 larvas en el estadío larval final de peso entre 0,25
y 0,35 g y se utilizó un grupo por cada condición. Se inyectaron alícuotas de 10 µl de las
suspensiones con una jeringa Hamilton dentro del hemocele de cada larva a través de la
última pata derecha, previa desinfección con alcohol. Todos los inoculos fueron
confirmados mediante recuentos de viables de cada suspensión estandarizada. Después
de la inyección las larvas fueron colocadas en placas de Petri e incubadas a 30°C en
oscuridad. Se contabilizó el número de larvas muertas cada 24 h por un periodo de 96 h.
En cada experimento se incluyó un grupo control de larvas inyectadas con 10 µl de
solución fisiológica para evaluar la muerte que se podría producir debido al trauma físico
generado por la inyección. Los experimentos con más de 1 larva muerta en el grupo
control se descartaron (129). Las curvas de supervivencia fueron graficadas con el
método de Kaplan-Meier y las diferencias significativas fueron analizadas con la prueba
Log-rank mediante el programa Graph Pad Prism versión 3.5”

Materiales y Métodos
35
15. Preparación de fracciones enriquecidas en OMPs
Cultivos bacterianos de 50 ml en TSB en presencia y ausencia de Dip fueron
centrifugaron a 10000 rpm a 4 °C y resuspendidos en HEPES 10 mM (pH 7,4). Los pellets
fueron lavados con HEPES 10 mM y aproximadamente 1,5 g (peso húmedo) de células
fueron resuspendidas en 7 ml del mismo buffer con PMSF (concentración final de 0,1
mM). Las fracciones enriquecidas en OMPs se obtuvieron utilizando la técnica de Leyh y
Griffith (96). Las suspensiones fueron sonicadas durante 10 min con pulsos de 1 min,
mientras se mantenían en un baño de hielo-agua. Las células enteras y los desechos se
separaron por centrifugación a 5.000 rpm durante 20 min y el sobrenadante se procesó
mediante ultracentrifugación a 100.000 rpm durante 60 min a 4 °C. El pellet, que
constituye la fracción de membranas totales, se resuspendió en Sarkosyl al 1,5 % (p/v)
disuelto en HEPES 10 mM con PMSF 0,1 mM y se incubó durante 30 min a temperatura
ambiente. La suspensión se centrifugó a 100000 rpm durante 60 min a 4 °C para obtener
el pellet que contiene el material insoluble en detergente donde se encuentra la fracción
enriquecida en proteínas de membrana externa. Finalmente el pellet se resuspendió en
agua desionizada.
16. Electroforesis en geles de poliacrilamida (SDS-PAGE)
Las fracciones enriquecidas en OMPs se solubilizaron en el buffer muestra de
Laemmli (94) y se calentaron a ebullición durante 5 min. La separación se realizó por
SDS-PAGE utilizando un gel discontinuo en gradiente y el sistema Tris/Tricina SDS-PAGE
(127). El gel se tiñó con azul de Coomassie G-250. Como sustancias de referencia se
emplearon marcadores de masa molecular (Mr) 205, 116, 97, 66, 45 y 29 (SDS-6H,
Sigma).
17. Digestión en gel e identificación de proteínas mediante
espectrometría de masa
Mediante la estrategia denominada huella de masas peptídicas, una proteína es
convertida en péptidos por la acción de una enzima. El conjunto de péptidos derivados de
la proteína es una característica propia de ésta y puede utilizarse para su identificación.
Así, la determinación de la masa de los péptidos de digestión de una proteína permite
hacer comparaciones con las masas teóricas del conjunto de péptidos esperados para las

Materiales y Métodos
36
secuencias almacenadas en una base de datos y sugerir aquellas que se ajustan más a
los datos experimentales.
Luego de la electroforesis se seleccionaron y cortaron las bandas de interés para
ser procesadas por esta técnica. Se realizó la digestión de dichas bandas con 20 µl de
una solución de tripsina de 25 ng/µl y el producto de digestión de cada una de las
muestras se analizó fue separado HPLC en fase reversa con la fase móvil en gradiente de
2-100 % de Solvente B a lo largo de 90 minutos (Solvente A: fluoruro de hidrógeno 0,1 %;
Solvente B: 80 % acetonitrilo en fluoruro de hidrógeno 0,1 %) y con una columna Vydac
RP-C18 (1,0 x 200 mm). Los péptidos separados se analizaron en el Espectrómetro de
Masa LCQ Duo ESI/TRAP (Thermo Fisher) acoplado al cromatografo. Como control
positivo se utilizó seroalbumina bovina (BSA, SIGMA) y como blanco, una zona del gel sin
muestra. Los datos se obtuvieron mediante el programa Triple Play y Data Dependent. Se
aplicó el programa Mascot MSMS (Matrix Science) del servidor público para el estudio de
los digeridos. La base de datos utilizada fue “nr”. Las muestras se analizaron en el
Laboratorio Nacional de Investigación y Servicios en Péptidos y Proteínas (LANAIS–PRO,
CONICET-UBA).
18. Cromatografía en placa delgada (TLC)
Se obtuvieron sobrenadantes de las cepas K279a y Fur60 a partir de 100 ml de
cultivos en SSC y SSC suplementado con FeCl3 100 µM, incubados durante 48 h,
mediante centrifugación a 10000 rpm. Como control se utilizaron sobrenadantes de
Escherichia coli K12 cultivada en los mismos medios con 18 h de incubación. Los
sobrenadantes fueron acidificados con HCl concentrado hasta pH 2,0 y extraídos dos
veces con 70 ml de acetato de etilo. Luego el extracto fue secado con sulfato de sodio
anhidro, evaporado a sequedad y el residuo fue resuspendido en 500 µl de acetato de
etilo. Los extractos se analizaron con placas de Silica gel 60 F254 (Merck) y como fase
móvil se utilizó una mezcla de cloroformo-metanol (2:1). Para revelar las placas
cromatográficas se atomizaron sobre la superficie los reactivos de la reacción de Arnow
para detectar compuestos de tipo catecol.

Materiales y Métodos
37
19. Cromatografía de alta performance (HPLC) y detección UV con arreglos de diodos
Extractos crudos obtenidos con acetato de etilo a partir de sobrenadantes de cultivo
en condiciones limitantes y no limitantes de hierro se analizaron mediante HPLC en fase
reversa.
19.1. Preparación de la muestra
Se obtuvieron sobrenadantes de cultivo a partir de 100 ml de cultivos incubados
durante 48 h en SSC y SSC suplementado con FeCl3 100 µM mediante centrifugación a
10000 rpm. El sobrenadante fue acidificado con HCl concentrado hasta pH 2,0 y extraído
dos veces con 70 ml de acetato de etilo. Luego el extracto se secó con sulfato de sodio
anhidro, el solvente se evaporó a sequedad y el residuo fue resuspendido en una mezcla
de acetonitrilo al 10 % en agua con ácido trifluoroacetico al 0,1 %.
19.2. HPLC con arreglo de diodos
Los compuestos presentes en el extracto crudo fueron separados mediante HPLC
en fase reversa con una columna C-18. Para la separación de los compuestos se aplicó
un gradiente de acetonitrilo (10-50 %) en agua con acido trifluoroacetico al 0,1 %, para
mantener el pH en 3,0, y un flujo de 1 ml min-1. Se inyectaron 20 µl de muestra. La
detección se realizó a 220 nm y a cada pico se le realizó un barrido espectral desde 210
hasta 360 nm (151).
20. Estadística
Todos los experimentos fueron realizados por triplicado y los datos son presentados
como la media con barras de error que representan el desvío estándar. Los datos fueron
analizados estadísticamente utilizando el test ANOVA de una vía y el test de comparación
a posteriori Student-Neuman-Keuls. Las diferencias en las medias fueron consideradas
estadísticamente significativas a partir de un P < 0,05.

Resultados y Discusión

Resultados y Discusión
39
Sección I
Tipificación de aislamientos locales de S. maltophilia recuperados de infecciones asociadas al uso de productos médicos
S. maltophilia es un microorganismo multirresistente, ampliamente distribuido en el
ambiente. Diferentes factores, tales como hospitalización por periodos prolongados,
terapia con antimicrobianos de amplio espectro y utilización de productos médicos han
sido asociados con un mayor riesgo de infección por esta especie.
Debido a que S. maltophilia posee un metabolismo relativamente inerte y a su
resistencia a múltiples antimicrobianos, las técnicas fenotípicas utilizadas en los estudios
epidemiológicos tales como el perfil bioquímico y de sensibilidad a antimicrobianos, han
resultado ser poco efectivas (41). Diferentes técnicas de tipificación molecular como
ribotipificación, electroforesis en campo pulsado (PFGE), amplificación al azar de
secuencias palindrómicas del ADN (RAPD) y amplificación de secuencias consenso
repetitivas intragénicas de enterobacterias (ERIC) entre otras, se utilizaron para estudiar
la epidemiología de esta especie. Numerosos autores encontraron una gran diversidad
genética entre aislamientos de S. maltophilia, esto condujo a pensar que dichos
aislamientos son adquiridos de forma independiente a partir de fuentes ambientales en
lugar de ser transmitidos persona a persona. Sin embargo existen algunos reportes de
pequeños brotes por transmisión cruzada (65, 144).
El objetivo de esta sección fue tipificar aislamientos locales de S. maltophilia
recuperados de infecciones nosocomiales asociadas al uso de productos médicos en la
Sección Bacteriología del Hospital de Clínicas José de San Martin, en el periodo 2004-
2012. Se estudiaron 63 aislamientos locales provenientes de 60 pacientes y se utilizó la
cepa K279a como cepa de referencia.

Resultados y Discusión
40
1. Identificación y fenotipificación
Se estudiaron 63 aislamientos recuperados de diferentes fuentes, la mayoría de
origen respiratorio (73 %) dentro de los cuales el 19 % provenían de muestras de lavado
bronqueo-alveolar y 54 % de aspirados traqueales. Los restantes aislamientos fueron
recuperados de muestras de sangre (21 %), orina (3 %), biopsia renal (1,5 %) y fluido
peritoneal (1,5 %). Las manifestaciones clínicas fueron infección urinaria asociada a
catéter, neumonía asociada a ventilación mecánica, y bacteriemia, asociada a catéter o a
infecciones relacionadas con hemodiálisis (Tabla 2).
Tabla 2. Origen de los aislamientos de S. maltophilia
Aislamiento de S. maltophilia Fuente de aislamiento (1)
Sm9 Sm11 Orina (2)
Sm13 Sm17 Sm18 Sm30 Sm33 Sm34 Sm44 Sm58 Sm61
Sm63 Sm68 Sm74 Sm76 Sangre (3)
Sm14 Biopsia Renal (4)
Sm15 Fluido peritoneal (5)
Sm19 Sm20 Sm26 Sm27 Sm28 Sm36 Sm38 Sm39 Sm40
Sm45 Sm47 Sm60bis LBA (6)
Sm10 Sm29 Sm31 Sm32 Sm35 Sm37
Sm42 Sm41 Sm46 Sm48 Sm43 Sm50
Sm51 Sm49 Sm53 Sm54 Sm52 Sm56
Sm57 Sm55 Sm59bis Sm60 Sm59 Sm64
Sm65 Sm62 Sm67 Sm69 Sm66 Sm71
Sm72 Sm70 Sm75 Sm73
Aspirado traqueal (6)

Resultados y Discusión
41
(1) Infecciones nosocomiales asociadas al uso de productos médicos: (2) catéter urinario, (3) catéter
vascular, (4) hemodiálisis, (5) diálisis peritoneal y (6) ventilación mecánica (LBA: lavado bronqueo-
alveolar).
La identificación de los aislamientos fue realizada en el servicio de Bacteriología del
Hospital de Clínicas mediante pruebas bioquímicas convencionales y fue confirmada en
nuestro laboratorio mediante el sistema API20NE, una galería conformada por 20 pruebas
bioquímicas que combina 8 pruebas convencionales y 12 pruebas de asimilación de
diferentes sustratos. Esta galería permitió agrupar a los aislamientos en 3 biocódigos, en
base a la capacidad de reducir nitratos y utilizar citrato como única fuente de carbono. El
biocódigo I, que se caracteriza por reducir el nitrato y utilizar citrato, fue el más frecuente e
incluyó al 52 % (33/64) de los aislamientos, en este grupo se encuentra la cepa de
referencia K279a. El biocódigo II, caracterizado por no reducir nitrato y utilizar citrato,
incluyó al 42 % (27/64) de los aislamientos. El biocódigo III, caracterizado por reducir
nitrato y no utilizar citrato, fue el grupo minoritario, compuesto por el 6 % (4/64) de los
aislamientos (Tabla 3 y Figura 7).
Figura 7. Porcentajes de distribución de los aislamientos de S. maltophilia según el biocódigo de
API20NE al que pertenecen.
52% 42%
6%
Biocódigo I
Biocódigo II
Biocódigo III

Resultados y Discusión
42
Tabla 3. Distribución de los aislamientos de S. maltophilia según el biocódigo de API20NE
Biocódigo API20NE Aislamiento de S. maltophilia
Biocodigo I
Reducción de nitrato y utilización de citrato
positivas
K279a, Sm9, Sm13, Sm14, Sm15, Sm17,
Sm18, Sm26, Sm27, Sm29, Sm31, Sm32,
Sm33, Sm39, Sm41, Sm44, Sm45, Sm47,
Sm48, Sm49, Sm50, Sm53, Sm55, Sm56,
Sm57, Sm58, Sm62, Sm64, Sm66, Sm67,
Sm69, Sm70, Sm74.
Biocodigo II
Reducción de nitrato negativa y utilización de
citrato positiva
Sm10, Sm11, Sm20, Sm28, Sm30, Sm34,
Sm35, Sm36, Sm37, Sm38, Sm42, Sm43,
Sm46, Sm51, Sm52, Sm59, Sm59bis,
Sm60, Sm60bis, Sm61, Sm63, Sm65,
Sm68, Sm71, Sm72, Sm75, Sm76
Biocodigo III
Reducción de nitrato positiva y utilización de
citrato negativa
Sm19, Sm40, Sm54, Sm73
2. Tipificación por métodos genotípicos
La genotipificación se realizó mediante dos técnicas basadas en la reacción en
cadena de la polimerasa (PCR), conocidas como Rep. Estas técnicas utilizan
oligonucleótidos cebadores que hibridan con secuencias del ADN repetitivas (secuencias
Rep) que se encuentran distribuidas en el cromosoma bacteriano. Por su elevado poder
discriminatorio y su simplicidad, permiten trabajar con un gran número de muestras (114).
Las técnicas Rep-PCR que se utilizaron en este estudio fueron ERIC-PCR (secuencias
consenso repetitivas intragénicas de enterobacterias) y REP-PCR (secuencias repetitivas
palindrómicas extragénicas) (58).

Resultados y Discusión
43
2.1. ERIC-PCR
Con la técnica de tipificación molecular ERIC-PCR se obtuvieron patrones de
bandas heterogéneos que presentaron entre 6 y 12 bandas, en su mayoría, menores de 2
Kb. Para comparar la relación entre los patrones de bandas obtenidos se realizó un
análisis de “clusters” mediante la utilización del algoritmo UPGMA (Unweighted Pair
Group Method with Arithmetic Averages). En el dendograma obtenido se pudo observar
que la mayoría de los aislamientos mostraron patrones de bandas únicos (Figura 8),
acorde con la gran diversidad intraespecie descripta por numerosos grupos de
investigaciçon (23, 68). Sin embargo, se obtuvieron algunos patrones idénticos a partir de
pacientes con infecciones respiratorias, en los cuales la posición de todas las bandas
coincide, sin importar la intensidad. Entre ellos, los de los aislamientos Sm41 y Sm45
procedentes de diferentes pacientes que estuvieron internados en la misma cama del
Hospital de Clínicas José de San Martin, con un mes de diferencia en el año 2008.
También se observaron patrones idénticos entre los aislamientos Sm56 y Sm57
recuperados de una paciente de 2 meses, aislados con 9 días de diferencia en octubre de
2009. Por último, 4 aislamientos recuperados en el periodo de una semana en noviembre
de 2009 presentaron perfiles de bandas idénticos por esta técnica (Figura 9). Dichos
aislamientos fueron: Sm59 y Sm59bis, recuperados de un paciente masculino de 92 años
con 4 días de diferencia, y los aislamientos Sm60 y Sm60bis, provenientes de un paciente
de sexo femenino de 72, recuperados con 2 días de diferencia.

Resultados y Discusión
44
Figura 8. Dendograma de similitud de los perfiles de bandas obtenidos por ERIC-PCR de algunos
aislamientos locales de S. maltophilia
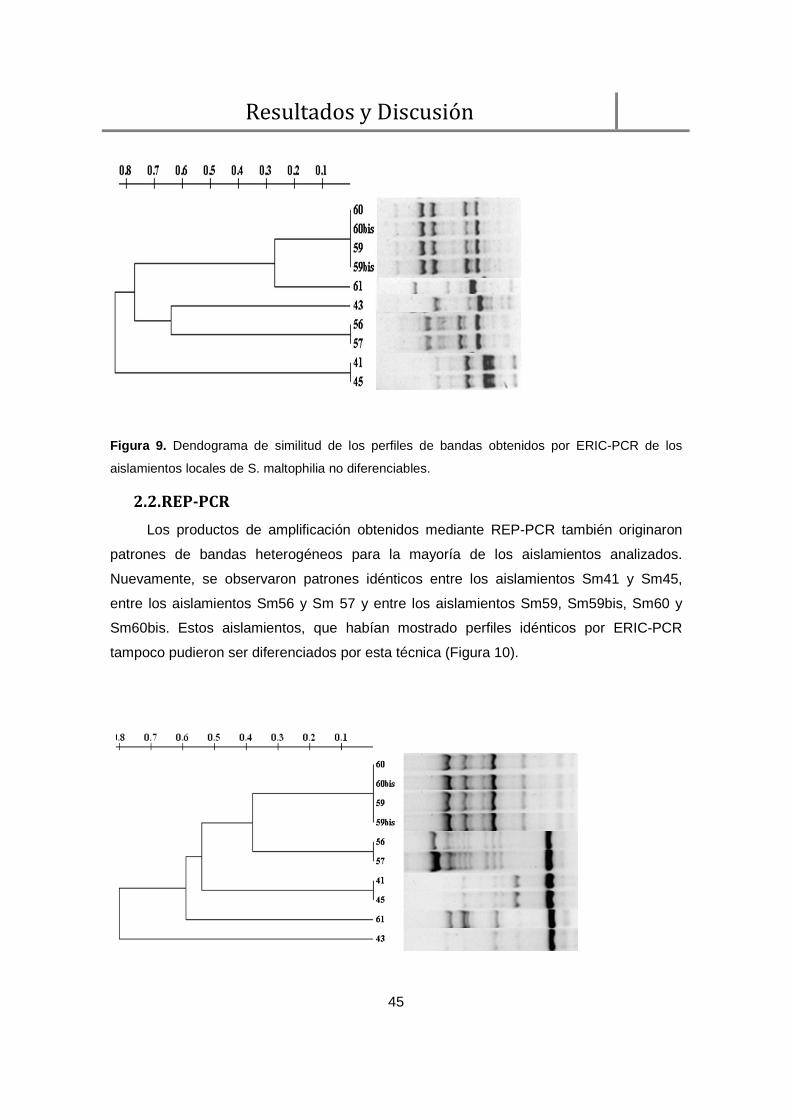
Resultados y Discusión
45
Figura 9. Dendograma de similitud de los perfiles de bandas obtenidos por ERIC-PCR de los
aislamientos locales de S. maltophilia no diferenciables.
2.2. REP-PCR
Los productos de amplificación obtenidos mediante REP-PCR también originaron
patrones de bandas heterogéneos para la mayoría de los aislamientos analizados.
Nuevamente, se observaron patrones idénticos entre los aislamientos Sm41 y Sm45,
entre los aislamientos Sm56 y Sm 57 y entre los aislamientos Sm59, Sm59bis, Sm60 y
Sm60bis. Estos aislamientos, que habían mostrado perfiles idénticos por ERIC-PCR
tampoco pudieron ser diferenciados por esta técnica (Figura 10).

Resultados y Discusión
46
Figura 10. Dendograma de similitud de los perfiles de bandas obtenidos por REP-PCR de los
aislamientos locales de S. maltophilia no diferenciables.
3. Discusión
El sistema API20NE resultó de utilidad para confirmar la identificación de los
aislamientos de S. maltophilia. A pesar de estar conformado por 20 pruebas bioquímicas
fue de escasa utilidad para la tipificación de los mismos ya que solo permitió agruparlos
en 3 biocódigos, esto concuerda con la característica de esta especie de poseer un
metabolismo relativamente inerte (41).
Para la elección de un método de tipificación molecular hay que tener en cuenta el
poder discriminatorio del mismo, la reproducibilidad, la complejidad y el costo. El método
con mayor poder discriminatorio y reproducibilidad es PFGE, sin embargo, su complejidad
y el alto costo del equipamiento necesario, lo transforman en un método poco accesible.
Por el contario, los métodos de PCR son relativamente fáciles de implementar, arrojan los
resultados en el mismo día y presentan alta reproducibilidad (133). Silbert et al (133),
evaluaron comparativamente las técnicas PFGE, ribotipificación y ERIC-PCR para tipificar
20 aislamientos de S. maltophilia. En este estudio, PFGE y ERIC-PCR mostraron un
poder discriminatorio elevado (D = 0,90) mientras que la ribotipificación presentó un poder
discriminatorio menor (D = 0,82).
En este estudio la tipificación molecular se realizó con dos técnicas de PCR: ERIC y
REP. Dichas técnicas revelaron una gran heterogeneidad genética entre los aislamientos
locales de S. maltophilia, sugiriendo que la mayoría de los episodios nosocomiales fueron
independientes. La gran diversidad de perfiles observados concuerda con estudios
previos (23, 68). Se ha sugerido que dicha diversidad se debe a que los aislamientos
provienen de la microbiota habitual del paciente y se seleccionan por una terapia con
antimicrobianos (23).
Los aislamientos Sm41 y Sm45 presentaron perfiles que no pudieron ser
diferenciados con las técnicas utilizadas. Del análisis de los datos epidemiológicos surgió
que los mismos fueron recuperados de diferentes pacientes que habían sido internados
en la misma cama con un mes de diferencia. Este hallazgo estaría en concordancia con el

Resultados y Discusión
47
estudio de Garcia de Viedma et al. (65) donde encontraron aislamientos provenientes de
neonatos con perfiles que presentaban una alta homología. Los resultados obtenidos por
ERIC-PCR fueron confirmados mediante PFGE, y basados en datos epidemiológicos los
autores atribuyeron la generación del brote a una transmisión horizontal. Por otra parte,
Caylan et al (23) detectaron tres pequeños brotes en un hospital universitario. En dicho
estudio los autores detectaron el mismo clon por periodos de 10-12 meses, lo que
atribuyeron a una posible capacidad de S. maltophilia de sobrevivir en el ambiente
hospitalario por periodos prolongados, y sugirieron que los brotes pudieron haber sido
generados por transmisión cruzada.
También se observaron patrones de bandas idénticos entre los aislamientos Sm56 y
Sm57 recuperados con 9 días de diferencia de un paciente de sexo femenino de 2 meses,
lo que significaría una infección no resuelta. Se ha reportado que el hallazgo de patrones
idénticos en esta especie se encuentra principalmente relacionado con aislamientos
sucesivos de un mismo paciente (65).
Los aislamientos Sm59 y Sm59bis de un paciente, y Sm60 y Sm60bis de otro
paciente fueron recuperados en el periodo de una semana y presentaron perfiles
idénticos. Sin embargo, por el carácter retrospectivo del estudio, no contamos con
mayores datos que los relacionen ya que no fueron pacientes de una misma sala ni existe
evidencia de transmisión por parte del personal de salud.
Las técnicas de ERIC-PCR y REP-PCR que son rápidas y sencillas, generaron
perfiles reproducibles y altamente discriminatorios. Estas técnicas, junto con la
información epidemiológica, serían útiles para la tipificación de aislamientos de S.
maltophilia (68). Sin embargo, en casos de aislamientos con perfiles indistinguibles se
debería utilizar la técnica de PFGE, que posee un mayor poder discriminatorio.

Resultados y Discusión
48
Sección II
Producción de potenciales factores de virulencia en aislamientos
locales de S. maltophilia
En S. maltophilia se conoce poco acerca de sus factores de virulencia. La dificultad
de diferenciar entre colonización e infección ha llevado a considerar a esta especie como
poco patogénica (41). Enzimas extracelulares tales como DNasa, fibrinolisina, lipasas y
proteasas, han sido relacionadas con su patogenicidad (18). Windhorst et al. (150)
caracterizaron una serina proteasa alcalina denominada StmPr1 que degrada proteínas
del suero y tejido conectivo.
Un factor de virulencia que contribuye a la patogenicidad bacteriana es la capacidad
de formar biofilms, debido a que las bacterias que crecen formando biofilms son más
resistentes a la acción de sustancias antibacterianas y al ataque por células fagocíticas.
Se ha descripto que S. maltophilia es capaz de formar biofilms en superficies bióticas y
abióticas (97).
El objetivo de esta sección fue detectar la presencia de los potenciales factores de
virulencia de S. maltophilia en aislamientos locales recuperados de infecciones asociadas
al uso de productos médicos.
1. Formación de biofilms de aislamientos locales
Para evaluar la capacidad de formar biofilms de los aislamientos locales, se utilizó el
ensayo cuantitativo en microplacas de poliestireno (PS). Los cultivos en caldo TSB se
incubaron en las microplacas a 35 °C sin agitación durante 48 h. El crecimiento se evaluó
mediante la determinación de la DO546 del cultivo, los aislamientos presentan lecturas de
DO546 entre 0,79 y 1,38 (Figura 11 a). La biomasa del biofilm (células adheridas y matriz)
se cuantificó mediante la tinción con cristal violeta (CV) y elución con etanol. Los biofilms
fueron clasificados en base al criterio de Stepanovic et al (137) que considera un valor de
corte (DOc) correspondiente al promedio de las DO546 de CV del control sin inocular más

Resultados y Discusión
49
3 desvíos estándar, y divide a los aislamientos en grupos: no productores de biofilms
(DO546 < DOc), productores de biofilms débiles (DOc < DO546 ≤ 2 DOc), moderados (2
DOc <DO546≤ 4 DOc) y fuertes (DO546 > 4 DOc).
La mayoría de los aislamientos locales y K279a produjeron biofilms fuertes, con
valores de DO546 de CV entre 0,80 y 1,80. En cambio, el aislamiento Sm61 se consideró
no productor de biofilms, siendo el único de nuestra colección que presentó esta
característica (Figura 11 b, Tabla 4).
K279a
Sm13Sm61
Sm54Sm10
Sm55Sm
58
Sm72Sm9
Sm41
Sm20Sm46
0.0
0.5
1.0
1.5
Cre
cim
ient
o (D
O54
6)
K279a
Sm13Sm61
Sm54
Sm10Sm55
Sm58
Sm72Sm9
Sm41Sm20
Sm460.0
0.5
1.0
1.5
2.0
2.5
Bio
mas
a (D
O54
6de
CV
)
a
b
Figura 11. Crecimiento y formación de biofilms en microplacas de PS de aislamientos de S.
maltophilia cultivados en TSB durante 48 h. a) crecimiento del cultivo b) Biomasa del biofilm
evaluada por tinción con CV. Los resultados corresponden a algunos aislamientos de la colección

Resultados y Discusión
50
con diferentes capacidades de formar biofilms y representan la media ± la desviación estándar
correspondientes a un experimento representativo.
2. Presencia del gen codificante de la proteasa StmPr1, actividad de
proteasa y lipasa
La presencia del gen de la proteasa StmPr1 se estudió mediante la reacción en
cadena de la polimerasa con oligonucleótidos que amplifican un fragmento interno de 570
pb. Los mismos se diseñaron en este estudio en base al genoma de la cepa K279a y de
otros genomas de S. maltophilia publicados (D457, JV3, R551-3) a fin de disminuir la
posibilidad de que eventuales polimorfismos influyan negativamente en la amplificación. El
producto de reacción esperado fue detectado en 52/63 aislamientos locales (82,53 %) y
en la cepa K279a (Tabla 4). En 11 aislamientos, entre los que se encuentran Sm10 y
Sm61, no se detectó producto de amplificación. No obstante, todos los aislamientos
fueron proteolíticos, independientemente de la ausencia del producto de amplificación.
Los valores de los halos de proteólisis producidos fueron de entre 14 y 21 mm. En base al
tamaño del halo los aislamientos se clasificaron como: (+) baja actividad, con halos de 14-
16 mm; (++) actividad intermedia, con halos de > 16 -18 mm, y (+++) alta actividad
proteolítica, con halos de > 18-21 mm. La cepa K279a y 33/63 aislamientos locales (52.38
%) presentaron alta actividad proteolítica (Tabla 4).
Por otra parte, el análisis de la actividad de lipasa reveló que dicha enzima está
presente en todos los aislamientos de S. maltophilia estudiados (Tabla 4).
3. Producción de sideróforos
La producción de sideróforos fue estudiada mediante el ensayo cromo azurol S
(CAS) en placa (8, 128). La técnica de CAS se basa en que el colorante cromo azurol S
incorporado en el medio agarizado forma complejos estables con el hierro, produciendo
un color azul-verde. Cuando un quelante de hierro, como el sideróforo, está presente
remueve el hierro del complejo, el medio cambia de color y se produce un halo naranja.
La utilización de esta técnica requirió su previa optimización. Primero, en lugar de
utilizar el método descripto por Schwyn y Neilands (128) se utilizó el método modificado
por Armstrong et al. (8), que utiliza el medio mínimo Stainer-Scholte (SS) ya que S.

Resultados y Discusión
51
maltophilia no desarrolla en el medio originalmente propuesto. Aun así los aislamientos
presentaron un crecimiento pobre que fue promovido por el agregado de Casamino ácidos
0,1 % (P/V) (SSC).
Se estudiaron un total de 63 aislamientos locales y la cepa de referencia K279a en
placas conteniendo el medio de cultivo SSC con el agregado del reactivo CAS y agar
(SSC-CAS). En estas condiciones solo 10 aislamientos locales fueron débilmente
positivos, con halos naranja alrededor de la colonia menor a 4,0 mm, mientras que la cepa
control Acinetobacter baumannii ATCC19606, produjo un halo de 11.0 mm (Figura 12 a,
Tabla 4). Como el componente Casamino ácidos agregado es una fuente de hierro, esos
resultados negativos podrían reflejar la presencia de niveles de hierro suficientes para
inhibir la síntesis de sideróforos. Por este motivo se agregó 2,2'-dipiridilo (Dip) al medio
para alcanzar la condición de limitación de hierro y estimular la producción de sideróforos.
En el medio SSC-CAS con el agregado de Dip 100 µM (SSC-CAS-Dip) los 63
aislamientos clínicos y la cepa de referencia K279a produjeron sideróforos (Figura 12 b).
Los aislamientos fueron divididos en tres grupos en base al diámetro del halo, lo que
indica diferencias en la producción de sideróforos (Tabla 4). La cepa K279a y 10/63
aislamientos locales (15,87 %) fueron fuertes productores de sideróforos. Por otra parte,
ninguno de los aislamientos produjo sideróforos cuando se agregó FeCl3 100 µM al medio
SSC-CAS (Figura. 12 c)
Figura 12. Producción de sideróforos mediante el ensayo agar CAS en diferentes medios de
cultivo a) SSC-CAS, b) SSC-CAS suplementado con Dip 100 µM, y c) SSC-CAS suplementado con

Resultados y Discusión
52
FeCl3 100 µM. 1: Acinetobacter baumannii ATCC19606; 2: Sm13; 3: Sm11; 4: Sm64; 5: Sm30; 6:
Sm29; 7: Sm40; 8: K279a; 9: Sm9.
4. Resistencia al estrés oxidativo
La sensibilidad al peróxido de hidrogeno de los aislamientos locales de S.
maltophilia y de K279a se determinó mediante el ensayo de difusión en placa. Todos los
aislamientos estudiados presentaron halos de inhibición de 14 a 22 mm. En base al
tamaño del halo de inhibición los aislamientos se clasificaron como: (+++) de alta
resistencia, con halos de 14-16 mm; (++) de resistencia intermedia, con halos > 16-18
mm, y (+) de baja resistencia, con halos > 18 mm (Tabla 4). La cepa K279a y 33/63
aislamientos locales (52,38 %) presentaron alta resistencia al estrés oxidativo (Tabla 4).
5. Presencia del gen narG, codificante de una nitrato reductasa y su relación con la utilización nitrato
La presencia del gen narG codificante de una nitrato reductasa asociada al
crecimiento en microaerofilia, se estudió mediante PCR con oligonucleótidos que
amplifican un fragmento interno de 623 pb, diseñados en este estudio siguiendo la misma
estrategia que se describió en el ítem 2. El producto de la PCR esperado fue detectado en
38/63 de los aislamientos locales (60,31 %) y en la cepa K279a.
Por otra parte, se estudió la capacidad de reducir el nitrato a nitrito y se observó
correlación con la presencia del gen narG en la mayoría de los aislamientos. Sin embargo,
en los aislamientos Sm10 y Sm11 se obtuvo el producto de amplificación del gen narG
pero los mismos no presentaron la capacidad de reducir el nitrato a nitrito.
Resulta interesante remarcar que Sm61 que no posee la capacidad de reducir
nitrato y no presentó producto de amplificación en la reacción en cadena de la polimerasa,
tampoco forma biofilms

Resultados y Discusión
53
Tabla 4. Producción de potenciales factores de virulencia de S. maltophilia
Amplificación por PCR 3
Aislamiento Actividad Lipolítica 1
Actividad Proteolítica 2 gen stmPr1 gen
narG Reducción de Nitrato 4
Formación de
Biofilm 5
Resistencia al estrés
oxidativo 6
Producción de
Sideróforos 7
K279a + +++ + + + +++ +++ +++ Sm9 + ++ + + + +++ ++ ++ Sm10 + + - + - +++ + + Sm11 + + + + - +++ + + Sm13 + +++ + + + +++ +++ +++ Sm14 + +++ + + + +++ ++ ++ Sm15 + + + + + +++ ++ + Sm17 + ++ - + + +++ ++ ++ Sm18 + ++ - + + +++ ++ + Sm19 + +++ + + + +++ ++ + Sm20 + ++ - - - +++ +++ ++ Sm26 + +++ + + + +++ ++ ++ Sm27 + + + + + +++ +++ +++ Sm28 + + + - - +++ + ++ Sm29 + +++ + + + +++ + ++ Sm30 + +++ + - - +++ +++ +++ Sm31 + +++ + + + +++ + ++ Sm32 + +++ + + + +++ + ++ Sm33 + +++ + + + +++ ++ ++ Sm34 + +++ + - - +++ ++ + Sm35 + ++ + - - +++ ++ + Sm36 + ++ + - - +++ ++ ++ Sm37 + + + - - +++ +++ ++ Sm38 + +++ + - - +++ +++ ++ Sm39 + +++ + + + +++ +++ ++ Sm40 + ++ + + + +++ +++ +++ Sm41 + ++ + + + +++ ++ +++ Sm42 + ++ + - - +++ ++ ++ Sm43 + ++ - - - +++ ++ ++ Sm44 + +++ + + + +++ ++ + Sm45 + ++ + + + +++ ++ ++ Sm46 + +++ + - - +++ ++ + Sm47 + + - + + +++ +++ + Sm48 + ++ + + + +++ +++ + Sm49 + ++ - + + +++ +++ ++ Sm50 + +++ + + + +++ + ++ Sm51 + +++ + - - +++ ++ ++ Sm52 + ++ + - - +++ ++ + Sm53 + +++ + + + +++ ++ ++ Sm54 + +++ + + + +++ + ++ Sm55 + ++ + + + +++ ++ + Sm56 + +++ + + + +++ +++ ++ Sm57 + +++ + + + +++ ++ ++ Sm58 + +++ + + + +++ + ++ Sm59 + +++ + - - +++ ++ ++
Sm59bis + ++ + - - +++ ++ ++ Sm60 + ++ + - - +++ ++ ++
Sm60bis + ++ + - - +++ ++ ++ Sm61 + +++ - - - - +++ +++ Sm62 + +++ + + + +++ ++ +++ Sm63 + ++ + - - +++ ++ +++ Sm64 + ++ + + + +++ ++ +++ Sm65 + ++ + - - +++ ++ ++ Sm66 + +++ + + + +++ + ++ Sm67 + +++ + + + +++ ++ + Sm68 + +++ + - - +++ ++ ++ Sm69 + +++ + + + +++ +++ ++ Sm70 + +++ + + + +++ +++ + Sm71 + +++ - - - +++ ++ + Sm72 + +++ - - - +++ ++ + Sm73 + +++ + + + +++ ++ + Sm74 + ++ - + + +++ + ++ Sm75 + ++ + - - +++ ++ ++ Sm76 + ++ + - - +++ +++ +

Resultados y Discusión
54
1 Evaluada en placas con agar base tibutirina suplementado con glicerol tributirato 1 %.
2 Evaluada en placas de agar nutritivo suplementado con leche descremada: (+) baja actividad, con
halos de 14-16 mm; (++) actividad intermedia, con halos de > 16 -18 mm, y (+++) alta actividad
proteolítica, con halos de > 18-21 mm.
3 (+) amplificación por PCR del fragmento esperado del gen correspondiente (-) ausencia de
amplificación.
4 Evaluado en caldo nitrato y revelado con el reactivo de Griess.
5 Ensayo en microplacas de PS. La biomasa del biofilm se cuantificó mediante la tinción con CV.
Los biofilms fueron clasificados en base al criterio de Stepanovic et al (137) que divide a los
aislamientos en grupos: (-) no productores de biofilms, (+) productores de biofilms débiles, (++)
moderados, y (+++) fuertes.
6 Evaluada mediante la determinación de los halos de inhibición generados por peróxido de
hidrogeno en el ensayo de difusión en placa (73). Los aislamientos se clasificaron como: (+++) de
alta resistencia, con halos de 14-16 mm; (++) de resistencia intermedia, con halos > 16-18 mm, y
(+) de baja resistencia, con halos > 18 mm.
7 Evaluada en placas de agar SSC-CAS suplementadas con Dip 100 µM. Los aislamientos se
clasificaron como: (+) productores débiles, con halos de 4,0-5,9 mm; (++) moderados, con halos de
6,0-8,9 mm, y (+++) productores fuertes de sideróforos, con halos > 9 mm.
6. Virulencia en el modelo de infección de Galleria mellonella
Con el objetivo de evaluar la virulencia de algunos aislamientos de nuestra colección
se utilizó el modelo de infección del lepidóptero Galleria mellonella, cuyo sistema inmune
comparte homología estructural y funcional con el de los mamíferos, mediada por
péptidos con actividad antimicrobiana y células fagocíticas (80). Este modelo presenta la
ventaja de poder utilizar un gran número de individuos por experimento que además son
fáciles de manipular.
Como primer paso se determinó la dosis infectiva que permitiera comparar la
virulencia de diferentes aislamientos de S. maltophilia. Para este propósito se utilizó la
cepa K279a y se infectaron grupos de 11 individuos con 104, 105 y 106 ufc/larva. Al utilizar
la dosis de 104 ufc/larva no se observó efecto letal en ninguna de las larvas a las 96 h,
mientras que la dosis de 106 ufc/larva resultó altamente letal y produjo la muerte del 90 %
de las larvas dentro de las primeras 24 h post infección (Figura 13). En cambio, con dosis

Resultados y Discusión
55
de 105 ufc/larva, K279a produjo la muerte del 20 % de las larvas a las 48 h, y del 40 % a
las 96h. En consecuencia, se eligió esta dosis para realizar los ensayos de virulencia.
0 20 40 60 80 1000
20
40
60
80
100K279a 104ufc/larvaK279a 105ufc/larvaK279a 106ufc/larva
Tiempo (h)
Sup
ervi
venc
ia (
%)
Figura 13. Ensayo de infección en larvas de G. mellonella. Luego de la inyección de 11 larvas por
grupo con una dosis cada uno se registró diariamente el número de individuos sobrevivientes y se
graficaron las curvas de Kaplan-Mayer. El grupo control de larvas inyectadas con solución
fisiológica no presentó individuos muertos (datos no graficados). Los datos corresponden a un
experimento representativo.
Los estudios realizados con los aislamientos locales Sm10, Sm13 y Sm61, con
dosis de 105 ufc/larva, evidenciaron tres niveles de virulencia significativamente diferentes
(P < 0,01). Sm10 no mostró efecto letal ya que a las 96 h todas las larvas inoculadas
continuaron con vida. Sm13 resultó letal para un 10 % de la población a las 24 h,
porcentaje que aumentó al 50 % a las 96 h. En cambio, Sm61 fue significativamente (P <
0,01) más virulenta ya que produjo un 30 % de muerte a las 24 h y alcanzó el 70% de
letalidad a las 96 h (figura 14).

Resultados y Discusión
56
0 20 40 60 80 10020
40
60
80
100Sm10Sm13K279aSm61
Tiempo (h)
Sup
ervi
venc
ia (
%)
Figura 14. Ensayo de infección en larvas de G mellonella. Luego de la inyección de 10 a 13 larvas
por cepa con la dosis de 105 ufc/larva se registró diariamente el número de individuos
sobrevivientes y se graficaron las curvas de Kaplan-Mayer. El grupo control de larvas inyectadas
con solución fisiológica no presentó individuos muertos (datos no graficados). Los datos
corresponden a un experimento representativo. El análisis estadístico se realizó con el método de
Log-rank.
7. Discusión
El uso cada vez más frecuente de productos médicos ha llevado a un significativo
aumento de las infecciones asociadas a los mismos. En esta sección se investigó la
diversidad de los aislamientos locales de S. maltophilia recuperados de infecciones
asociadas a asistencia respiratoria, catéteres venosos, para diálisis y urinarios, con
respecto a la producción de potenciales factores de virulencia.
La capacidad de formar biofilms sobre superficies abióticas es un factor que
favorece la colonización de productos médicos (Denton & Kerr, 1998). Dicha capacidad es
además un importante factor de virulencia bacteriano ya que facilita la persistencia en el
huésped, la evasión de la respuesta inmune y la sobrevivencia bacteriana frente a altas
concentraciones de antimicrobianos. Todos de los aislamientos locales de S. maltophilia,
salvo Sm61 que no forma biofilms, produjeron biofilms fuertes, al igual que la cepa K279a,
en el ensayo en microplacas de PS. En cambio, Pompilio et al. detectaron biofilms fuertes
y débiles en 47 aislamientos clínicos de S. maltophilia no provenientes de enfermos
fibroquísticos, mientras que los aislamientos de pacientes fibroquísticos produjeron en

Resultados y Discusión
57
general biofilms moderados (121). Cabe destacar que en un trabajo anterior de nuestro
grupo se determinó que el aislamiento Sm13 posee la capacidad de formar biofilms en
catéteres uretrales de goma, silicona y PVC (117).
La producción de proteasa juega un papel significativo en la patogénesis bacteriana
ya que participa en la invasión y el daño tisular. Nicoletti et al. (110) determinaron la
presencia de los genes stmPr1 y stmPr2, que codifican la principal proteasa de S.
maltophilia y otra proteasa menor, respectivamente, en 43 aislamientos clínicos
provenientes de pacientes con fibrosis quística. Además, diseñaron cebadores para
detectar 2 alelos de stmPr1, unos en base al genoma de K279a y otros, en base al gen
descripto por Windhorst et al. (150) . La mayoría de los aislamientos presentó el alelo
correspondiente a K279a, mientras que en 6 aislamientos no detectaron ninguno de los
alelos. Por otra parte, detectaron la presencia de stmPr2 en todos los aislamientos (110).
Mediante la utilización del modelo de virulencia de G. mellonella estos autores
encontraron que la proteasa StmPr1 es un factor de virulencia relevante en los
aislamientos clínicos analizados.
Los resultados presentados muestran que utilizando los cebadores diseñados en
este estudio se detectó la presencia del gen stmPr1 en el 82,53 % de los aislamientos
locales, sin embargo, todos los aislamientos presentaron actividad proteolítica. Dicha
actividad, podría deberse a la presencia de 1 alelo de stmPr1 no detectado o a la
presencia de la proteasa StmPr2. Estos resultados están en concordancia con los de
Nicoletti et al. y difieren con lo observado en 11 aislamientos provenientes de pacientes
fibroquísticos por Di Bonaventura et al. quienes solo detectaron el gen stmPr1 en 2
aislamientos y sugirieron que el mismo no está ampliamente distribuido en esta especie
(44).
Por otra parte, la producción de lipasa podría contribuir a la virulencia de S.
maltophilia mediante la degradación de tejidos ricos en lípidos como los presentes en
pulmón, o desencadenando una intensa respuesta inflamatoria (143). Todos los
aislamientos locales y K279a presentaron actividad lipolítica.
Otro factor de virulencia son los sideróforos, los cuales facilitan la adquisición de
hierro en el huésped. El ensayo universal en placa CAS agar es un método ampliamente
utilizado para detectar la producción de sideróforos. Sin embargo, en S. maltophilia
algunos autores no detectaron sideróforos por este ensayo mientras que otros

Resultados y Discusión
58
describieron halos muy pequeños (3-5 mm) (52, 105). Con el objetivo de estudiar la
producción de sideróforos en los aislamientos locales se optimizó su detección mediante
modificaciones en la composición del medio. Los resultados muestran que la producción
de sideróforos de S. maltophilia requiere el agregado de casamino ácidos y Dip. La
necesidad del agregado de quelantes fue también descripta por Chatterjee y Sonti
quienes reportaron que Xanthomonas oryzae pv. oryzae no produce sideróforos en el
medio peptona-sucrosa agar-CAS pero el agregado de Dip al medio permite su detección
(35).
La resistencia al estrés oxidativo es una característica importante para la
patogénesis bacteriana ya que durante el proceso de infección las bacterias se enfrentan
a especies reactivas del oxigeno producidas por las células fagocíticas. Nuestros
aislamientos mostraron diferentes niveles de resistencia al estrés oxidativo in vitro. La
cepa K279a y el 52,38 % de los aislamientos locales, entre los que figuran Sm13 y Sm61,
presentaron alta resistencia al estrés oxidativo, mientras, que Sm10 fue uno de los menos
resistente (Tabla 4).
La presencia del gen narG codificante de una nitrato reductasa asociada al
crecimiento en microaerofilia, fue detectada en el 60,31 % de los aislamientos estudiados
y mostró correlación con la capacidad de reducir el nitrato a nitrito. Sin embargo, en los
aislamientos Sm10 y Sm11 se obtuvo el producto de amplificación del gen narG pero los
mismos no presentaron la capacidad de reducir el nitrato, lo que sugiere la presencia de
una variante no funcional del gen. La presencia del gen narG y la capacidad de reducir
nitrato, relacionadas con el crecimiento en microaerofilia, detectadas en la mayoría de los
aislamientos de S. maltophilia facilitaría el crecimiento en biofilms, y de esta forma
incrementaría la patogenicidad. Por ejemplo, Sm61 no presentó estas características y no
formó biofilms en el ensayo en microplacas. Sin embargo, 24 aislamientos, en los que no
se obtuvo el producto de amplificación del gen narG ni presentaron la capacidad de
reducir el nitrato, formaron biofilms fuertes en microplacas. Esto podría deberse a la
multiplicidad de factores involucrados en dicha formación.
Por último, se puso a punto el modelo de infección de G. mellonella que permitió
comparar la virulencia de algunos aislamientos locales y K279a. Se seleccionaron los
aislamientos Sm13, Sm10 y Sm61 en función de sus potenciales factores de virulencia.
Sm13, al igual que Sm39, Sm56, Sm69 y Sm70, presentó junto con K279a la capacidad

Resultados y Discusión
59
de formar biofilms, alta actividad proteolítica, capacidad de reducir nitratos y alta
resistencia al estrés oxidativo, además de haberse detectado la presencia de los genes
stmPr1 y narG. El aislamiento Sm10 formó biofilms, pero presentó baja actividad
proteolítica, baja resistencia al estrés oxidativo, no presentó la capacidad de reducir el
nitrato, y no se detectó la presencia de los genes stmPr1 y narG. Por último, Sm61 es el
único aislamiento que no formó biofilms, además, no presentó la capacidad de reducir el
nitrato, y no se detectó la presencia de los genes stmPr1 y narG, sin embargo, presentó
alta actividad proteolítica y alta resistencia al estrés oxidativo.
Sm13 y la cepa K279a fueron virulentas en el modelo de infección de G. mellonella,
en concordancia con la detección de los potenciales factores de virulencia mencionados.
En cambio, Sm10 no mostró efecto letal, lo cual podría estar relacionado a su baja
actividad proteolítica y baja resistencia al estrés oxidativo. Por último, Sm61 presentó el
mayor efecto letal a pesar de no formar biofilms y no haberse detectado el gen stmPr1,
gen relacionado con la virulencia en este modelo (110). Esto podría deberse su alta
resistencia al estrés oxidativo, lo cual podría ser importante en este modelo debido a que
G. mellonella tiene un sistema inmune, mediado por células fagocíticas.

Resultados y Discusión
60
Sección III
Sistema de captación de hierro de alta afinidad de S. maltophilia
K279a y su posible regulación por el sistema Fur
El hierro es un nutriente esencial para los microorganismos pero, a pesar de ser un
compuesto abundante en la naturaleza, su biodisponibilidad es limitada. En presencia de
oxigeno es oxidado a Fe3+ y precipita en forma de hidróxidos insolubles (142). La baja
disponibilidad de hierro libre dificulta el crecimiento de los microorganismos, por lo tanto
éstos han desarrollado diferentes estrategias para captarlo del medio. Una de ellas es el
sistema que se basa en sintetizar y secretar sideróforos, compuestos de bajo peso
molecular, que quelan hierro con alta afinidad. Una vez formado el complejo hierro-
sideróforo es reconocido por un receptor específico y es transportado al interior celular
(5).
El hierro libre actúa como catalizador en las reacciones de Fenton y Haber-Weiss
(142) que generan especies reactivas del oxigeno (ERO) tales como radicales hidroxilo
(OH•), aniones superoxido (O2
-) y peróxidos (RO2). Dichos compuestos causan daños a
los componentes celulares, por eso la homeostasis del hierro debe ser finamente
controlada. La producción de sideróforos y la expresión de los componentes involucrados
en la captación del complejo hierro-sideróforo están reguladas negativamente a través del
sistema Fur (ferric uptake regulator) en muchas especies bacterianas (5). En S.
maltophilia no hay estudios sobre el sistema Fur ni sobre los genes que regula.
El objetivo de esta sección fue obtener mutantes Fur de S. maltophilia, caracterizar
las OMPs reguladas por la concentración de hierro, y determinar la naturaleza química de
los sideróforos sintetizados por esta especie bacteriana.
1. Aislamiento de mutantes Fur
Como paso inicial se analizó el genoma de S. maltophilia K279a (32) y se encontró
un gen fur putativo (Smlt1986), que codifica una proteína de 135 aminoácidos. El
alineamiento múltiple de la secuencia de nucleótidos del gen fur de K279a reveló que

Resultados y Discusión
61
presenta un 86 % de identidad con el de Xanthomonas campestris pv campestris ATCC
33913 (XCC1470) (37) y un 76% de identidad con el de Pseudomonas aeruginosa PAO1
(PA4764) (152).
Las mutantes espontáneas Fur fueron asiladas mediante la técnica de selección con
manganeso (69). Se seleccionaron un total de 51 clones independientes de K279a luego
de 72 h de incubación en el medio agar LB suplementado con manganeso. Dichos clones
resistentes al manganeso fueron cultivados en el medio agar CAS suplementado con
hierro para identificar mutantes Fur, debido a que dichas mutantes producen sideróforos
constitutivamente. En estas condiciones 2 clones, denominados Fur42 y Fur60, mostraron
el halo naranja alrededor de la colonia indicativo de la producción de sideróforos. Por el
contrario la cepa salvaje K279a no presentó dicho halo en presencia de hierro (Figura 15).
Para confirmar la presencia de mutaciones, el gen fur completo y un fragmento de
100 pb de la región rio arriba del codón de inicio de la transcripción fueron amplificados
por PCR utilizando los cebadores diseñados en base al genoma de K279a (Tabla 1) y
posteriormente secuenciados. Tanto el clon Fur42 como el Fur60 presentaron una
mutación puntual. Fur42 presentó una mutación que causa la sustitución de un único
aminoácido (Asp�Gly en la posición 5) y Fur60 presentó una mutación en la secuencia -
10 de la región promotora putativa (T�C).
Figura 15. El ensayo universal agar CAS a) SSC-CAS suplementado con FeCl3 20 µM y b) SSC-
CAS suplementado con Dip 100 µM. El halo naranja es indicativo de la producción de sideróforos.

Resultados y Discusión
62
La ausencia de un gen fur funcional causa la producción constitutiva de sideróforos, mientras que
la cepa salvaje solo produce sideróforos bajo condiciones de limitación de hierro.
2. Caracterización de proteínas de membrana externa reguladas por
hierro en S. maltophilia K279a
2.1. Análisis de los perfiles de OMPs
Se estudiaron los perfiles de fracciones enriquecidas en proteínas de membrana
externa de la cepa K279a y de su mutante Fur60 en presencia y ausencia del quelante de
hierro 2-2´ dipiridilo (Dip) mediante SDS-PAGE. Los perfiles de OMPs de K279a
presentaron una variación en el patrón de bandas en la región de 70-90 kDa cuando las
bacterias crecieron en condiciones limitantes de hierro. Se observó un aumento de la
expresión de la denominada Banda 1 (de mayor peso molecular) y la aparición de la
Banda 2. Este perfil fue similar al de la mutante Fur60 cultivada tanto en TSB como en
TSB-Dip (Figura 16).
Figura 16. Perfiles de fracciones enriquecidas en OMPs. 1: K279a cultivada en TSB; 2: K279a
cultivada en TSB-Dip; 3: Fur60 cultivada en TSB; 4: Fur60 cultivada en TSB-Dip y 5: patrón de
peso molecular. Las flechas negras indican la posición de la Banda 1 (de mayor PM) y de la Banda
2, reguladas por hierro.

Resultados y Discusión
63
2.2. Identificación de las OMPs reguladas por hierro mediante
espectrometría de masa
Para la identificación de las OMPs reguladas por hierro se utilizó la estrategia
denominada huella de masas peptídicas. Mediante esta técnica una proteína es
convertida en péptidos por la acción de una enzima. El conjunto de péptidos derivados de
la proteína es una característica propia de ésta y puede utilizarse para su identificación.
Así, la determinación de la masa de los péptidos de digestión de la proteína permite hacer
comparaciones con las masas teóricas del conjunto de péptidos esperados para las
secuencias almacenadas en una base de datos y sugerir aquellas que se ajustan más a
los datos experimentales.
El análisis por espectrometría de masa reveló que la Banda 1 presenta homología
con FepA (gi|190573428), un potencial receptor de membrana externa de enterobactina
en S. maltophilia K279a de 80,7 kDa, con un score de 283 y un 57 % de cobertura de la
secuencia de la proteína (Figura 17 a). La Banda 2 presenta homología con CirA
(gi|190575965), un potencial receptor de colicina I de S. maltophilia K279a de 77,3 kDa,
con un score de 419 y un 60 % de cobertura de la secuencia (Figura 17 b). Dichos
potenciales receptores de K279a fueron propuestos por comparación de la secuencia del
genoma de K279a depositada por Crossman et al. (32) con las secuencias codificantes de
proteínas ya caracterizadas experimentalmente en otras especies bacterianas.

Resultados y Discusión
64
Figura 17. Secuencia de aminoácidos de las proteínas de K279a identificadas por espectrometría
de masa: a) receptor de membrana externa de enterobactina FepA, (gi|190573428) y b) receptor
de colicina I CirA, (gi|190575965). En rojo y negrita se encuentran resaltados los péptidos
detectados experimentalmente.
Con el objetivo de profundizar el estudio de FepA y CirA se realizó un análisis in
silico con el algoritmo BPROM (Softberry, Inc.) de las regiones que comprenden 200
nucleótidos río arriba de los respectivos codones de inicio de la transcripción. Este
análisis permitió localizar las regiones promotoras de fepA y cirA (Figura 18 a).
Para detectar potenciales cajas Fur en el genoma de K279a (número de acceso
AM743169), se utilizó el programa MAST (11) y 112 secuencias de cajas Fur
caracterizadas en otras especies bacterianas (155) Se obtuvieron 41 secuencias de
K279a con potenciales cajas Fur. La correspondiente a fepA, 5`-
GCATTTGAGAATCACTCGC-3`, entre las posiciones -144 y -125 respecto al codón de
inicio, se encuentra solapada con la región -35 del presunto promotor. La potencial caja
Fur de cirA, 5`-CGCAACGGTTATCATTTCA-3`, localizada entre las posiciones -82 y -63,
se encuentra solapada con la región -10 del presunto promotor (Figura 18 a). Finalmente,
las secuencias de S. maltophilia obtenidas se analizaron con el programa MEME, una
herramienta que permite encontrar motivos repetidos en un grupo de secuencias (10). De
esta forma se obtuvo una secuencia consenso de la caja Fur putativa de K279a (Figura 18
b).

Resultados y Discusión
65
Figura 18. a) Análisis in silico de fepA y cirA: las secuencias conservadas -10 y -35 de los
respectivos promotores se muestran recuadradas y las cajas Fur putativas se muestran resaltadas
en amarillo. b) Logo de la secuencia consenso que representaría la caja Fur putativa de K279a.
3. Caracterización de los sideróforos producidos en S. maltophilia
3.1. Determinación de la naturaleza química de los sideróforos
La mayoría de los sideróforos producidos por los microorganismos son compuestos
de tipo catecol o hidroxamatos. Para determinar la naturaleza química de los sideróforos
producidos por S. maltophilia se emplearon las técnicas de Csáky y Arnow que detectan
compuestos tipo hidroxamato y catecol, respectivamente. Para ello se utilizaron
sobrenadantes de cultivo en medio SSC con el agregado de Dip 100 µM o de FeCl3 100
µM. Bordetella bronchiseptica 7865 y Acinetobacter baumannii ATCC19606 se emplearon
como controles de cepas productoras de sideróforos tipo hidroxamato y catecol,
respectivamente. El ensayo de Arnow reveló que todos los aislamientos locales y K279a

Resultados y Discusión
66
producen sideróforos de tipo catecol en presencia de Dip y no los producen en presencia
de hierro. Ninguno de los aislamientos estudiados produjo sideróforos de tipo
hidroxamato.
3.2. Análisis mediante cromatografía en capa delgada (TLC) de
extractos provenientes de sobrenadantes de cultivos de K279a
Los sideróforos presentes en el sobrenadante de cultivo se extrajeron, previa
acidificación, con acetato de etilo y los compuestos presentes en el extracto se separaron
por cromatografía en capa delgada en placas de silica gel 60 F254. Las placas fueron
reveladas mediante la atomización de los reactivos de la reacción de Arnow, lo que
permitió visualizar las manchas correspondientes a compuestos de tipo catecol.
Los extractos de K279a obtenidos en condiciones de limitación de hierro y de la
mutante Fur60, tanto en condiciones limitantes como no limitantes de hierro, mostraron
tres manchas correspondientes a compuestos de tipo catecol con Rf de 0,72, 0,36 y 0,2, y
una mancha que quedó en el punto de siembra. Dichos compuestos no se encontraron en
los extractos de K279a obtenidos en condiciones no limitantes de hierro (Figura 19). Se
utilizaron como control extractos de E. coli K12, que solo produce el sideróforo
enterobactina, obtenidos a partir de cultivos con concentraciones limitantes de hierro (5).
El perfil de compuestos de tipo catecol del control presentó los mismos Rf que los
observados en extractos de las cepas de S. maltophilia en estudio.

Resultados y Discusión
67
Figura 19. TLC en placas de silica gel 60 F254 fase móvil cloroformo-metanol (2:1). revelada
mediante la atomización con los reactivos de Arnow. Las flechas señalan las manchas
correspondientes a compuestos tipo catecol
3.3. Análisis mediante cromatografía líquida de alta performance
(HPLC) de extractos provenientes de sobrenadantes de cultivos de
K279a
Los compuestos presentes en los extractos de los sobrenadantes de cultivo se
separaron mediante HPLC en fase reversa y se detectaron mediante espectroscopia UV
con arreglos de diodos. En el cromatograma obtenido de los extractos de los
sobrenadantes de cultivo en condiciones de limitación de hierro se observó la aparición de
cuatro picos (Figura 20 b) que no estuvieron presentes en los extractos obtenidos en
presencia de concentraciones no limitantes del mismo (Figura 20 a). El primer pico tuvo
un tiempo de retención de 12,754 min y presentó un espectro UV con tres bandas de
absorción a 210 nm, 247 nm y 307 nm, respectivamente, similares a las del espectro de
enterobactina (60). Además, se observaron otros dos picos, uno con tiempo de retención
de 18,043 min, que presentó un espectro UV con tres bandas similares a las antes
mencionadas, y otro con tiempo de retención de retención de 15,453 min cuyo espectro
UV comparte las primeras 2 bandas de absorción. Por último, se observó la presencia de
un pico con tiempo de retención de 14,754 min que presentó un espectro UV cuyas
bandas difirieron de las de los picos anteriores (217,5 y 277,9 nm) (Figura 20 b y c)

Resultados y Discusión
68

Resultados y Discusión
69
Figura 20. Cromatograma de HPLC de los extractos de a) cultivos en condiciones no limitantes de
hierro y b) cultivos en condiciones de limitación de hiero. c) Espectros UV-DAD de los picos
presentes en condiciones de limitación de hierro: en rojo (tiempo de retención 12,7min), verde
(15,4 min), azul (14,7 min) y violeta (18,04 min).
4. Discusión
Durante el desarrollo de esta tesis se obtuvieron, por primera vez, mutantes Fur de
S. maltophilia. Para ello se utilizó la técnica de selección por manganeso que permite
obtener mutantes puntuales en el gen fur. Esta técnica fue ampliamente utilizada para
aislar mutantes Fur en numerosas especies tales como P. aeruginosa (74) y
Xanthomonas campestris pv campestris (86) donde no se han podido aislar mutantes
completamente “knock out” probablemente debido a la gran variedad de funciones
esenciales que controla esta proteína (14). Las mutantes Fur de S. maltophilia produjeron
sideróforos en forma constitutiva, lo que sugiere la falta de regulación por el sistema Fur-
Fe2+. La secuenciación del gen fur permitió poner en evidencia que Fur42 presenta una
mutación que causa la sustitución de un único aminoácido (Asp�Gly en la posición 5) y
Fur60, una mutación en la secuencia -10 de la región promotora putativa (T�C). La
mutante Fur60 se utilizó en los estudios presentados en la Sección V sobre la función del
hierro en la formación de biofilms y en la resistencia al estrés oxidativo de S. maltophilia.
El análisis de las fracciones enrriquecidas en OMPs mediante SDS-PAGE mostró
que en condiciones limitantes de hierro los perfiles de K279a presentan un aumento de la
expresión de la denominada Banda 1 y la aparición de la Banda 2, similar a lo observado
en los perfiles de OMPs de Fur60 cultivada tanto en condiciones limitantes como no
limitantes de hierro. Este comportamiento, sugiere que la expresión de ambas proteínas
está regulada por el complejo Fur-Fe2+. El análisis por espectrometría de masa reveló que
la Banda 1 presenta homología con FepA, un potencial receptor de enterobactina, y la
Banda 2, con CirA, un potencial receptor de colicina I de K279a. En E. coli K-12 FepA es
el receptor del complejo hierro-enterobactina mientras que CirA es el receptor de los
productos de degradación lineales de dicho complejo y actúa como una vía alternativa
para incorporar hierro (103). Mediante análisis in silico se determinaron las potenciales
cajas Fur de fepA y cirA. La correspondiente a fepA, 5`-GCATTTGAGAATCACTCGC-3`,
entre las posiciones -144 y -125 respecto al codón de inicio, se encuentra solapada con la

Resultados y Discusión
70
región -35 del presunto promotor. La potencial caja Fur de cirA, 5`-
CGCAACGGTTATCATTTCA-3`, localizada entre las posiciones -82 y -63, se encuentra
solapada con la región -10 del presunto promotor (Figura 18 a). Finalmente, se obtuvo
una secuencia consenso de la caja Fur putativa de K279a (Figura 18 b).
Por otra parte, se determinó la naturaleza química de los sideróforos de S.
maltophilia. Todos los aislamientos locales y la cepa K279a produjeron sideróforos de tipo
catecol en condiciones limitantes de hierro. Estos resultados no concuerdan con los
obtenidos por Chhibber et al quienes reportaron la producción de ornibactina por S.
maltophilia (36). Sin embargo, Ryan et al. reportaron que S. maltophilia K279a y R551-3
producen el sideróforo de tipo catecol enterobactina, basados en la informacion de los
genomas recientemente secuenciados (126). El genoma de S. maltophilia K279a
(Smlt2817, Smlt2818 y Smlt2822) codifica los potenciales componentes A, F y C de
enterobactina sintasa, lo que sugiere la producción de enterobactina en esta especie
(http://www.sanger.ac.uk/Projects/S_maltophilia/).
El análisis por TLC de los extractos obtenidos a partir de los sobrenadantes de
cultivos de K279a, en condiciones limitantes de hierro y de F60, en presencia o ausencia
de Dip, permitió revelar la presencia de varios compuestos de tipo catecol. Dichos
compuestos presentaron los mismos valores de Rf que los presentes en el extracto de E.
coli K12, que solo produce enterobactina. Como lo describieron Furrer et al. (64) dichos
compuestos podrían corresponder al monómero, dímero y trímero de enterobactina, y el
compuesto con mayor Rf a enterobactina. La separación mediante HPLC se realizó
siguiendo las condiciones utilizadas por Winkelmann el al. (151) para separar
enterobactina y sus derivados lineales quienes identificaron los picos separados mediante
espectrometría de masa y determinaron que un pico con tiempo de retención de 18,3 min
correspondía a enterobactina, otros con 16,2 min y 14,5 correspondian al trímero y
dimero, respectivamente. La similitud de los picos detectados en K279a, junto con el
espectro UV obtenido, sugiere que dichos compuestos podrían ser enterobactina y sus
derivados.

Resultados y Discusión
71
Sección IV
Función del sistema de señalización rpfF/DSF en la formación de biofilms y en la virulencia de S. maltophilia
En Xanthomonas campestris pv. campestris (Xcc) la producción de enzimas
extracelulares está regulada de forma positiva por el operon rpf (“regulation of
pathogenicity factors”). Los genes rpfB y rpfF son los responsables de la síntesis del
autoinductor DSF (“diffusible signal factor”). La disrupción del gen rpfF conlleva a la
ausencia de producción del autoinductor, lo que ocasiona importantes cambios
fenotípicos. El sistema de QS está implicado en la regulación de factores de virulencia y
en la formación de biofilms en Xcc (13, 47, 135).
En S. maltophilia la señalización por QS también está mediada a través de la
molécula DSF. La mutante K279arpfF presenta una severa disminución en la motilidad
tipo “swimming”, en la producción de proteasa, en la tolerancia frente a anitibioticos y
metales pesados, presenta una alteración en el perfil de lipopolisacáridos y además,
forma agregados en el medio L. Por otra parte, en un medio artificial de esputo, que
simula el ambiente en el pulmón de un paciente fibroquístico, la cepa K279a forma
microcolonias mientras que su mutante rpfF no las forma. (62).
El objetivo de esta sección fue estudiar la función de DSF en el crecimiento
planctónico, la formación y la arquitectura de biofilms, la expresión de caracteres
fenotípicos asociados a su desarrollo, y la virulencia de S. maltophilia. Para este propósito
se realizaron estudios comparativos entre la cepa salvaje K279a y su mutante K279arpfF.
1. Crecimiento planctónico
Se evaluó el posible efecto de la señalización por QS en el crecimiento planctónico
de S. maltophilia en condiciones estáticas y con agitación (150 rpm). El crecimiento de las
cepas K279a y su mutante K279arpfF cultivadas en TSB a 35 °C fue determinado
espectrofotométricamente a 540nm a diferentes tiempos de incubación.
Los resultados presentados en la figura 21 muestran que en ambas condiciones la
cosecha máxima alcanzada por la mutante rpfF fue inferior a la correspondiente a la cepa
salvaje. Cabe resaltar que en condiciones estáticas esta diferencia fue marcadamente

Resultados y Discusión
72
mayor (DO540 K279a 1,120 y DO540 K279arpfF 0,391) que con agitación (DO540 K279a
1,758 y DO540 K279arpfF 1,239). Asimismo, en ambas condiciones la mutante rpfF
presentó menor velocidad de crecimiento que la cepa salvaje, especialmente en
condiciones estáticas.
b
0 20 40 600.0
0.5
1.0
1.5
2.0K279a
K279arpfF
Tiempo (h)
DO
540
a
0 20 40 600.0
0.5
1.0
1.5
2.0
Tiempo (h)
DO
540
Figura 21. Curvas de crecimiento de S. maltophilia K279a y de su mutante rpfF en: a) condiciones
estáticas y b) con agitación. Las bacterias fueron cultivadas en TSB a 35 °C, el crecimiento fue
determinado espectrofotométricamente a 540nm. Los resultados se expresan como la media ± la
desviación estándar de un experimento representativo.
2. Papel del “quorum sensing” en la formación de biofilms
Para evaluar la capacidad de formar biofilms de la cepa salvaje y de su mutante rpfF
sobre diferentes superficies abióticas se utilizaron el ensayo en tubos de borosilicato (BS)
y de polipropileno (PP), y el ensayo en microplacas de poliestireno (PS). Además, se
analizaron los biofilms formados sobre BS mediante microscopia óptica y microscopia
confocal laser de barrido (CLSM).
2.1. Análisis de biofilms formados sobre superficies hidrofílicas e
hidrofóbicas
Se evaluó la formación de biofilms sobre superficies hidrofílicas (BS) e hidrofóbicas
(PP) utilizando el ensayo en tubos. Los cultivos en TSB se incubaron con agitación (150

Resultados y Discusión
73
rpm) durante 6, 24 y 48 h. Luego, se cuantificó espectrofotométricamente la biomasa de
los biofilms teñidos con CV.
S. maltophilia K279a presentó una marcada adherencia en BS a las 6 h de
incubación, la biomasa alcanzó su máximo nivel a las 24 h y disminuyó ligeramente a las
48 h (Figura 22 a). En cambio, no se observó adherencia en PP a las 6 h de incubación, y
los valores de DO546 del CV obtenidos a las 24 h fueron menores que los
correspondientes a las 6 h en BS (P < 0,01). Finalmente, a las 48 h de incubación K279a
presentó una adherencia en PP semejante a la alcanzada a las 6 h en BS (Figura 22 b).
La mutante mostró una adherencia significativamente menor (P < 0,001) que la de la cepa
salvaje en ambas superficies durante todo el periodo estudiado (Figura 22 a y b).
En la figura 22 c y d se presenta las imágenes de biofilms formados sobre tubos de
BS y PP, respectivamente, teñidos con CV. La inspección visual de los biofilms formados
en tubos ha sido ampliamente utilizada como tamizaje para determinar la capacidad de
formar biofilms de aislamientos bacterianos (112). En las imágenes se puede observar
que K279a se adhiere a la interfase aire-medio de cultivo y tapiza el fondo y las paredes
de BS a partir de las 6 h mientras que solo se visualizan imágenes similares en PP a las
48 h de incubación. Por su parte, la mutante formó anillos incompletos en la interfase aire-
medio de cultivo en ambas superficies.

Resultados y Discusión
74
Figura 22. Formación de biofilms de S. maltophilia K279a y su mutante rpfF en superficies
hidrofílicas e hidrofóbicas. Los cultivos en TSB se incubaron a 35 ºC durante 6, 24 y 48 h. a) y b)
biomasa del biofilm formado sobre tubos de BS y PP, respectivamente, cuantificada por tinción con
CV. Los resultados se expresan como la media ± desviación estándar de un experimento
representativo.*representa diferencias estadísticamente significativas. c) y d) Imagen de biofilms
formados sobre tubos de BS y PP, respectivamente, teñidos con CV.
2.2. Análisis de biofilms formados en microplacas
En el ensayo cuantitativo en microplacas de PS el crecimiento fue evaluado
mediante la determinación de la DO546 del cultivo, y la biomasa (células adheridas y matriz
extracelular) de los biofilms teñidos con CV fue evaluada espectrofotométricamente. Los
resultados obtenidos en condiciones estáticas, a diferentes tiempos de incubación, se
presentan en la figura 23 a y b, respectivamente.
La cepa salvaje presentó una marcada adherencia a las 6 h de cultivo, a pesar de
su escaso crecimiento y alcanzó el máximo nivel de adherencia a las 24 h (Figura 23 b),
comportamiento similar al observado en BS. En concordancia con los resultados del
crecimiento planctónico en condiciones estáticas descripto en el punto 1 (Figura 23a), el
crecimiento de la mutante en la microplaca se vio severamente reducido, respecto al de la
cepa salvaje. La mutante mostró una adherencia significativamente menor (P < 0,001) que
la de la cepa salvaje en todo el periodo estudiado (Figura 23 b).

Resultados y Discusión
75
a
6 24
48
0.0
0.4
0.8
1.2
1.6
K279aK279arpfF*
* *
Tiempo (h)
Cre
cim
ient
o (D
O54
6)
b
6 24
48
0.0
0.4
0.8
1.2
1.6
2.0
2.4
2.8
*
** *
Tiempo (h)
Bio
mas
a (D
O54
6de
CV
)
Figura 23. Crecimiento y formación de biofilms de S. maltophilia K279a y su mutante rpfF en
microplacas de PS cultivadas en TSB a 35ºC. a) crecimiento, y b) biomasa del biofilm cuantificada
por tinción con CV. Los resultados se expresan como la media ± desviación estándar de un
experimento representativo. *representa diferencias estadísticamente significativas.
Además la adición de DSF comercial 200 µM al medio de cultivo produjo un
incremento significativo (P < 0,05) de la biomasa del biofilm de la mutante evaluada a las
48 h de incubación (figura 24).

Resultados y Discusión
76
K279a rpfF rpfF+ DSF200µM 0.0
0.5
1.0
1.5
2.0
2.5*
*B
iom
asa
(DO
546
de C
V)
Figura 24. Formación de biofilms de S. maltophilia K279a y su mutante rpfF en microplacas de PS
cultivadas a 35 ºC, durante 48 h en TSB y TSB suplementado con DSF comercial 200 µM. Biomasa
del biofilm cuantificada por tinción con CV. Los resultados se expresan como la media ± desviación
estándar de un experimento representativo. *representa diferencias estadísticamente significativas.
2.3. Análisis de biofilms por microscopía óptica
Las características de los biofilms de K279a y de la mutante rpfF desarrollados
sobre cubreobjetos de BS, a partir de cultivos en TSB incubados 48 h, fueron analizadas
por microscopia óptica previa tinción con CV (Figura 25). El biofilm de K279a presentó
una monocapa de células adheridas con extensas áreas de la superficie cubiertas por
microcolonias. Por el contrario, el patrón de adherencia de la mutante rpfF se caracterizó
por presentar bacterias aisladas y agregados celulares.

Resultados y Discusión
77
Figura 25 . Biofilms de S. maltophilia desarrollados sobre cubreobjetos de BS. Los biofilms
desarrollados por K279a y su mutante rpfF a partir de cultivos en TSB incubados 48 h fueron
teñidos con CV y examinados por microscopia de campo claro con una magnificación de 400x.
2.4. Análisis de biofilms por microscopia confocal laser de barrido
(CLSM)
Se estudió la arquitectura de los biofilms formados por ambas cepas mediante
CLSM que permite la visualización de muestras totalmente hidratadas y, de esta forma,
revela la elaborada estructura tridimensional del biofilm. Los biofilms se formaron en las
cámaras especiales de cultivo Lab.Tek, Nunc® con base de borosilicato (BS). Los cultivos
en TSB y TSB suplementado con DSF comercial se incubaron durante 48 h en
condiciones estáticas. El biofilm formado en la base de la cámara se visualizó con el
colorante fluorescente Syto9®, un agente que atraviesa fácilmente las membranas, se
une a los ácidos nucleicos y tiñe de color verde las bacterias. Como se observa en la
imagen de un plano x-y (Figura 26) el biofilm de K279a en TSB presentó una monocapa
con microcolonias distribuidas por toda la superficie. La reconstrucción tridimensional
realizada a partir de la serie de imágenes tomadas en los planos x-y a través del eje z
permitió evidenciar la arquitectura del biofilm, compuesta por picos de hasta 30 µm
intercalados por canales acuosos (Figura 27). La mutante rpfF no mostró un crecimiento
confluente sino que formó un gran número de agregados celulares dispersos y su
estructura tridimensional mostró solo algunos picos de apenas 10 µm (Figura 26 y Figura
27). Por otra parte, los biofilms de la mutante formados en TSB suplementado con DSF

Resultados y Discusión
78
presentaron un crecimiento casi confluente con más cantidad de microcolonias y picos de
20 µm (Figura 26 y Figura 27).
Figura 26. Imágenes de biofilms de S. maltophilia obtenidas mediante CLSM. Se realizaron
cultivos en las cámaras Lab.Tek de K279a y su mutante rpfF en TSB y TSB suplementado con
DSF. Luego de 48 h de incubación los biofilms fueron teñidos con Syto9®. Cada imagen central
corresponde a un plano x-y, mientras que las imágenes de la parte superior y de la derecha de
cada plano x-y representan la proyección en el eje z de la serie de imágenes obtenidas a partir de
los cortes en profundidad. La barra representa 20 µm.

Resultados y Discusión
79
Figura 27. Reconstrucción tridimensional de la serie de imágenes obtenidas mediante CLSM en
los planos x-y a través del eje z de biofilms de S. maltophilia. Las reconstrucciones fueron
realizadas con el programa ImageJ.
3. Producción de sustancias poliméricas extracelulares (EPS)
Se evaluó la producción de EPS mediante la técnica de precipitación con etanol y
determinación del peso seco. Para ello, se utilizaron sobrenadantes de cultivos

Resultados y Discusión
80
planctónicos en LB-glu 0,1 % de S. maltophilia incubados con agitación durante 48 h a 35
ºC. La figura 28 muestra que la cepa K279a produce cantidades significativamente
menores de EPS que la mutante rpfF. El análisis de los componentes químicos de los
precipitados realizado mediante la cuantificación de hidratos de carbono con glucosa
como estándar (50) y de proteínas (19) demostró que la cepa K279a presenta mayor
cantidad de exopolisacáridos por mg de EPS (0,27 ± 0,02) que la mutante (0,12 ± 0,05).
Por el contrario, la mutante produjo cantidades mucho mayores de proteínas por mg de
EPS (0,85 ± 0,04) que la cepa salvaje (0,56 ± 0,07) (Tabla 5). Por lo tanto, relación
hidratos de carbono/proteínas en la EPS es mucho mayor para la cepa salvaje (0,48) que
para la mutante (0,14).
K279a K279arpfF0
100
200
300
400 *
EP
S( µµ µµ
g)/B
iom
asa(
mg)
Figura 28. Producción de EPS de S. maltophilia K279a y su mutante K279arpfF cultivadas durante
48 h, a 35 ºC con agitación. Los resultados se expresan como la media ± desviación estándar de la
relación entre el peso seco de las EPS (µg) y el peso seco de la biomasa del cultivo (mg), de un
experimento representativo.*representa diferencias estadísticamente significativas.

Resultados y Discusión
81
Tabla 5. Composición de las EPS de cepas de S. maltophilia
Cepas de S.
maltophilia
EPS
(µg de EPS/mg de
biomasa)
Análisis químico de EPS
Hidratos de
carbono
(mg de glucosa/mg de
peso seco de EPS)
Proteínas
(mg de proteínas/mg
de peso seco de EPS)
Relación Hidratos de
carbono/Proteínas
(mg de glucosa/mg de
proteínas)
K279a 77,4 ±12,2 0,27 ± 0,02 0,56 ± 0,07 0,48
K279arpfF 301,4 ± 20,3 0,12 ± 0,05 0,85± 0,04 0,14
Para profundizar el estudio de la composición molecular de las EPS se utilizó
espectroscopia de reflexión total atenuada infrarroja con transformada de Fourier (ATR-
FTIR). Como primer paso, se determinó la cantidad de EPS adecuada a ser empleada
para la adquisición de espectros IR. Con este propósito, el precipitado de EPS obtenido
con etanol a partir del sobrenadante de K279a fue resuspendido en una relación de 20 µl
de agua Milli-Q por cada mg de EPS (peso seco) y diluido 1/4, 1/10 y 1/20. Alícuotas de
150 µl de las suspensiones fueron transferidas al cristal del modulo ATR y se les evaporó
el agua para formar una película sobre el mismo. Luego se obtuvieron los
correspondientes espectros de cada dilución. Los valores de absorbancia de los picos
amida I y amida II se utilizaron para evaluar la saturación de la señal por ser los más
intensos. La dilución 1/20 demostró ser adecuada para la adquisición de espectros (datos
no mostrados).
El espectro de ATR-FTIR obtenido con la suspensión de los precipitados es el
resultado de la superposición de las huellas digitales de cinco biomoléculas. El agua está
representada por el estiramiento simétrico y asimétrico del grupo OH en el rango de 3800-
3000 cm-1 y el balanceo a 1640 cm-1. Las proteínas están representadas principalmente
por estiramiento y vibraciones del grupo N-H de la unión peptídica entre 1700-1500 cm-1.
La banda amida I corresponde al estiramiento del grupo C=O de las amidas y la banda
amida II corresponde a la combinación fuera de fase del balanceo en un plano del grupo

Resultados y Discusión
82
N-H y el estiramiento C-N. Los lípidos presentan bandas entre 3100 a 2800 cm-1
correspondientes a vibraciones debidas a estiramientos y balanceos de C-H en los grupos
funcionales CH3 y CH2. Los picos de la región 1200-900 cm-1 son debidos principalmente
a un fuerte acoplamiento de los estiramientos C-C y C-O y de las deformaciones C-O-C y
C-O-H correspondientes a oligo y polisacáridos. Asimismo en esta ventana se encuentra
el estiramiento simétrico del grupo PO2- perteneciente a ácidos nucléicos (1080 cm-1). Por
último la zona entre 1300 y 1500 cm-1 es la denominada zona mixta, ya que involucra
absorciones de grupos funcionales presentes en lípidos, proteínas, carbohidratos y ácidos
nucléicos (21).
El espectro de ATR-FTIR de las EPS de la mutante mostró mayor intensidad de
absorción en los picos de amida I (1643,08 cm-1) y amida II (1550,31 cm-1) que el de la
cepa salvaje. Por el contrario el pico de 1053,40 cm-1 de polisacáridos presentó una
intensidad ligeramente mayor en la cepa K279a que en su mutante (Figura 29). Ambos
resultados están en concordancia con la relación hidratos de carbono/proteínas de las
EPS determinada por métodos químicos, la cual es mucho mayor para la cepa salvaje
que para la mutante (Tabla 5). En la zona de los grupos funcionales CH3 y CH2 de los
lípidos (3100 a 2800 cm-1) no se observó diferencia entre ambas cepas.
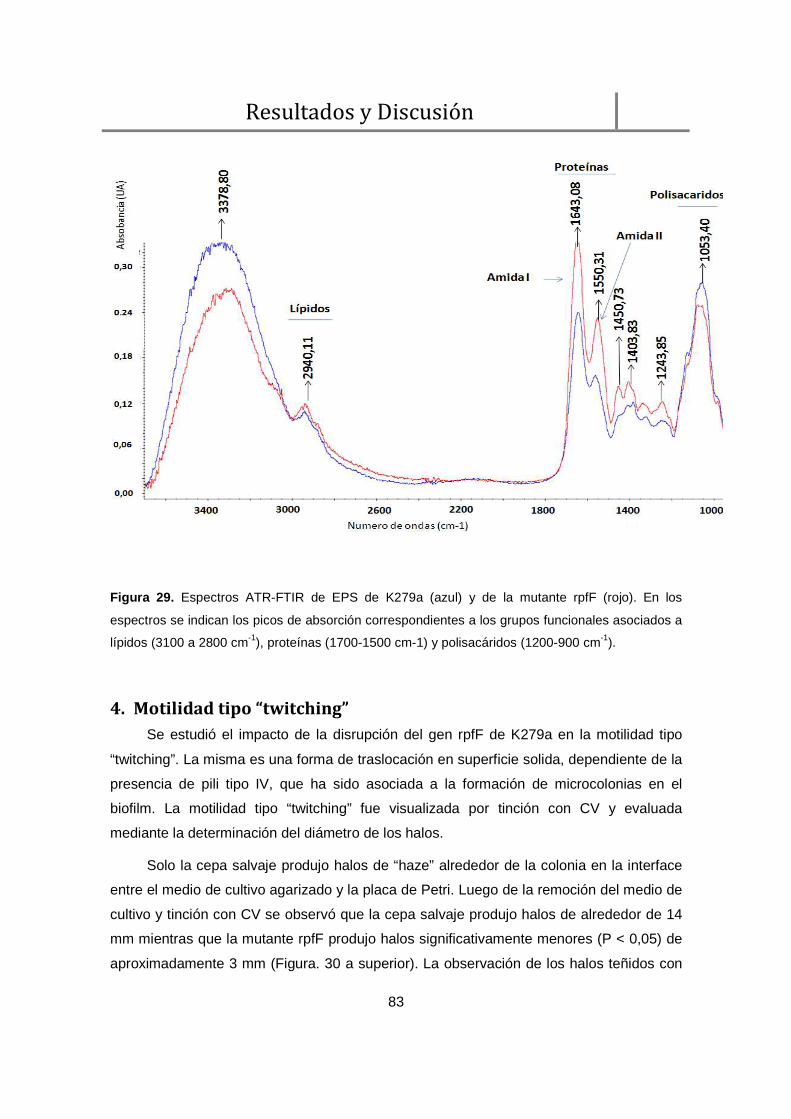
Resultados y Discusión
83
Figura 29. Espectros ATR-FTIR de EPS de K279a (azul) y de la mutante rpfF (rojo). En los
espectros se indican los picos de absorción correspondientes a los grupos funcionales asociados a
lípidos (3100 a 2800 cm-1), proteínas (1700-1500 cm-1) y polisacáridos (1200-900 cm-1).
4. Motilidad tipo “twitching”
Se estudió el impacto de la disrupción del gen rpfF de K279a en la motilidad tipo
“twitching”. La misma es una forma de traslocación en superficie solida, dependiente de la
presencia de pili tipo IV, que ha sido asociada a la formación de microcolonias en el
biofilm. La motilidad tipo “twitching” fue visualizada por tinción con CV y evaluada
mediante la determinación del diámetro de los halos.
Solo la cepa salvaje produjo halos de “haze” alrededor de la colonia en la interface
entre el medio de cultivo agarizado y la placa de Petri. Luego de la remoción del medio de
cultivo y tinción con CV se observó que la cepa salvaje produjo halos de alrededor de 14
mm mientras que la mutante rpfF produjo halos significativamente menores (P < 0,05) de
aproximadamente 3 mm (Figura. 30 a superior). La observación de los halos teñidos con

Resultados y Discusión
84
CV en la placa de Petri de poliestireno mostró importantes diferencias en el patrón de
adherencia. La cepa K279a presentó una zona de adherencia expansiva mientras que la
mutante permaneció adherida cerca del sitio de inoculación (Figura. 30 a inferior). El
análisis microscópico de los halos de K279a reveló un borde externo compuesto por
grupos bacterias que se alejan de una monocapa de células, y más cerca del punto de
inoculación, una monocapa con microcolonias típica de los biofilms. La mutante, por el
contrario, no presentó un patrón definido de adherencia (Figura 30 b).
Figura 30. Motilidad “twitching” de S. maltophilia K279a y su mutante rpfF. Las cepas fueron
inoculadas por punción en LB agar 1% e incubadas a 35 ºC por 48 h. a) Diámetros de los halos de
“twitching” teñidos con CV e imagen de los mismos. Los resultados muestran la media ± la
desviación estándar de un experimento representativo. Las diferencias son estadísticamente
significativas a un valor de P < 0,05 (*). b) Imagen de microscopía de campo claro de los halos
teñidos con CV (magnificación de 400X).

Resultados y Discusión
85
5. Hidrofobicidad de la superficie celular
Se evaluó la hidrofobicidad de la superficie celular de S. maltophilia mediante: 1- el
ensayo de agregación bacteriana en presencia de sulfato de amonio (SAT, salt
aggregation test), 2- la afinidad al n-hexadecano (MATH, microbial adherence to
hydrocarbons) y 3- la afinidad por superficies de poliestireno (PS) a través del ensayo de
réplica (125) (Tabla 6). La cepa K279a fue clasificada como moderadamente hidrofóbica
en base a los valores de SAT (1,00) y MATH (21 %). Por el contrario K279a rpfF se
comportó como no hidrofóbica según el valor de SAT (4,00) y no presentó adherencia al
n-hexadecano.
En concordancia con estos resultados, el método de réplica, en el cual la
hidrofobicidad de la superficie celular correlaciona con la adherencia bacteriana al PS,
mostró que solo la cepa salvaje se adhiere a esa superficie (Figura 31).
Tabla 6. Hidrofobicidad de la superficie celular de cepas de S.maltophilia
Cepas de S. maltophilia MATH SAT Método de replica
K279a 21 % 1,00 ++
K279arpfF - 4,00 -
MATH (microbial adherence to hydrocarbons): < 20 % no hidrofóbica, ≥ 20 % < 50 %,
moderadamente hidrofóbica, y ≥ 50 %, fuertemente hidrofóbica (93). SAT (salt aggregation test): <
1 M, fuerte; 1 M, moderada; o 2 M, débil hidrofobicidad; y ≥ 4 M, no hidrofóbica (93). Método de
replica: adherencia al PS, - negativa, (+) reducida, + positiva, o ++ positiva fuerte (125)

Resultados y Discusión
86
Figura 31. Hidrofobicidad celular de S. maltophilia evaluada mediante el método de replica: a)
colonias de K279a (mitad superior) y de su mutante rpfF (mitad inferior) en el medio TSA y b)
replica de dichas colonias sobre PS, teñida con CV.
6. Resistencia al estrés oxidativo
Se evaluó la resistencia al estrés oxidativo en S. maltophilia K279a y de su mutante
rpfF. Cultivos en fase exponencial de ambas cepas fueron expuestos a una concentración
de H2O2 20 mM por diferentes períodos de tiempo. Se calculó el porcentaje de células
sobrevivientes como el número de ufc/ml luego del tratamiento relativo al número de
ufc/ml inicial.
Luego de 15 y 30 min de exposición la cepa salvaje presentó un porcentaje de
sobrevida de 2,96 ± 0,89 y 1,71 ± 0,78, respectivamente, mientras que la supervivencia de
la mutante rpfF fue significativamente menor (P < 0,01) que la de K279a (0,10 ± 0,04 y
0,03 ± 0,01 % de sobrevida a los 15 o 30 min de exposición) (Figura 32).
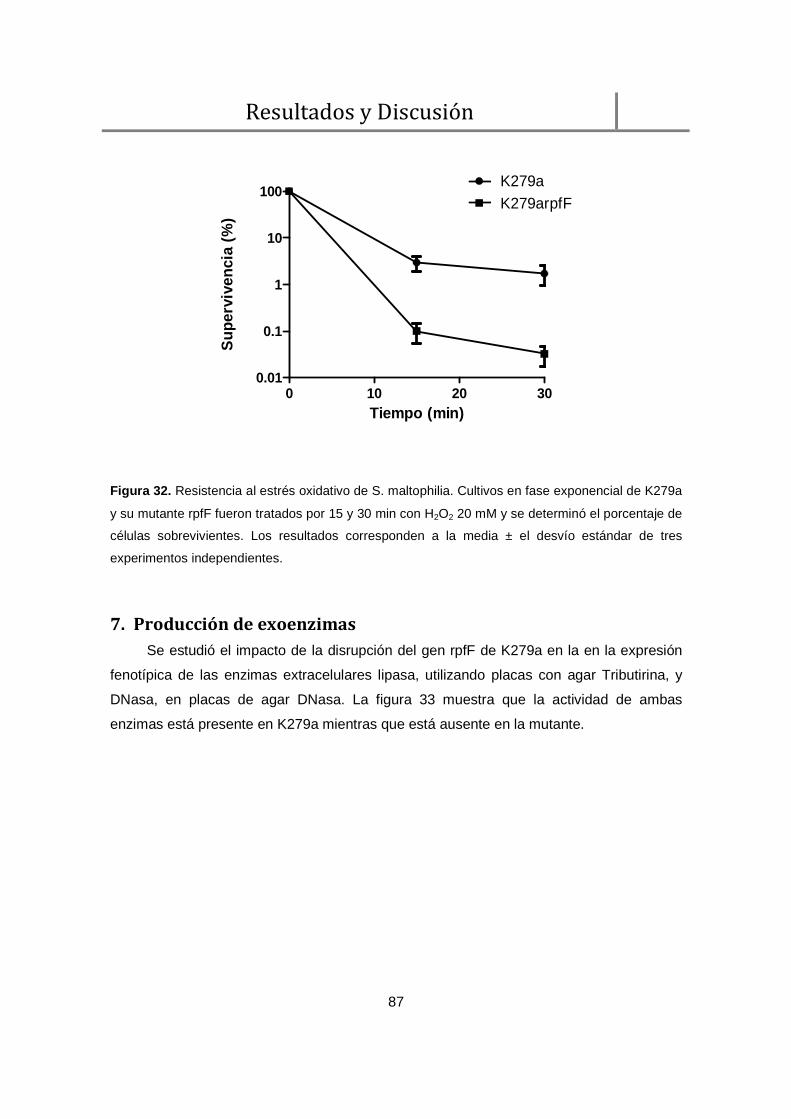
Resultados y Discusión
87
0 10 20 300.01
0.1
1
10
100K279aK279arpfF
Tiempo (min)
Sup
ervi
venc
ia (
%)
Figura 32. Resistencia al estrés oxidativo de S. maltophilia. Cultivos en fase exponencial de K279a
y su mutante rpfF fueron tratados por 15 y 30 min con H2O2 20 mM y se determinó el porcentaje de
células sobrevivientes. Los resultados corresponden a la media ± el desvío estándar de tres
experimentos independientes.
7. Producción de exoenzimas
Se estudió el impacto de la disrupción del gen rpfF de K279a en la en la expresión
fenotípica de las enzimas extracelulares lipasa, utilizando placas con agar Tributirina, y
DNasa, en placas de agar DNasa. La figura 33 muestra que la actividad de ambas
enzimas está presente en K279a mientras que está ausente en la mutante.

Resultados y Discusión
88
Figura 33. Actividad de las exoenzimas lipasa (agar Tributirina) y DNasa (agar DNasa, revelado
con HCl) en S. maltophilia. Las placas sembradas en estría con cultivos de K279a y su mutante
rpfF se incubaron durante 48 h a 35°C.
8. Evaluación de la virulencia mediante el modelo de Galleria
mellonella
Con el objetivo de evaluar el efecto de la mutación en el gen rpfF sobre la virulencia
de S. maltophilia se utilizó el modelo de infección del lepidóptero G. mellonella con la
dosis infectiva de 105 ufc/larva que demostró ser adecuada para estudios comparativos
(sección II).
Se observó una diferencia significativa (P < 0,0174) de la virulencia entre ambas
cepas, K279a resultó letal para un 20 % y 40 % de la población a las 48 h y 96 h,
respectivamente, mientras que la mutante rpfF no mostró efecto letal y el porcentaje de
supervivencia fue similar al del grupo control inyectado con solución fisiológica (Figura 34,
datos del grupo control no presentados).

Resultados y Discusión
89
0 20 40 60 80 10040
60
80
100K279a
K279arpfF
Tiempo (h)
Sup
ervi
venc
ia (
%)
Figura 34. Ensayo de infección de larvas de Galleria mellonella con cepas de S. maltophilia. Luego
de la inyección de 10 a 13 larvas por cepa con la dosis de 105 ufc/larva se registró diariamente el
número de individuos sobrevivientes y se graficaron las curvas de Kaplan-Mayer. El grupo control
de larvas inyectadas con solución fisiológica no presentó individuos muertos (datos no graficados).
El análisis estadístico se realizó con el método de Log-rank. Los datos corresponden a un
experimento representativo.
9. Discusión
La mutación en el gen rpfF produjo efectos pleiotrópicos, entre ellos, una marcada
disminución de la cosecha máxima y de la velocidad de crecimiento de K279arpfF,
fundamentalmente en condiciones estáticas con baja tensión de oxígeno (Figura 21 a). Se
ha propuesto que la capacidad de crecer en microaerofilia de K279a está relacionada con
la formación de biofilms (32).
La disrupción del gen rpfF produjo además una disminución de la movilidad tipo
“twitching” y una alteración de la hidrofobicidad superficial. Previamente Fouhy et al (62)
observaron que la mutante K279arpfF presenta alterada la movilidad mediada por flagelos
(“swimming”). Todas estas características influyen en la formación de biofilms. Dicha
formación requiere de la movilidad mediada por flagelos que permite a la bacteria
establecer el contacto inicial con la superficie (111). Por otra parte, la adhesión depende
de estructuras de la superficie bacteriana tales como pili y adhesinas, y de su
hidrofobicidad, y además, de propiedades de la superficie biótica o abiótica tales como la

Resultados y Discusión
90
hidrofobicidad y la rugosidad, y de las interacciones electrostáticas entre ambas
superficies (46).
Nuestros resultados muestran que la cepa K279a es moderadamente hidrofóbica en
base a los valores de SAT y MATH mientras que la mutante se comportó como no
hidrofóbica (Tabla 6). Además, en el método de réplica, solo la cepa salvaje se adhiere al
PS (Figura 31). En consecuencia, la disrupción del gen rpfF en S. maltophilia conduce a
cambios en la hidrofobicidad superficial. En un trabajo anterior de nuestro grupo de
investigación hemos descripto una pobre correlación entre la hidrofobicidad de la
superficie bacteriana y la formación de biofilms, sin embargo, en algunos aislamientos
clínicos de S. maltophilia se observó correlación entre una moderada hidrofobicidad y la
formación de biofilms fuertes (117). Por su parte Pompilio et al. concluyeron que la
hidrofobicidad es un factor importante en la adhesión y formación de biofilms, sin
embargo, observaron que algunos aislamientos con diferente hidrofobicidad poseían
similares capacidades de formar biofilms (120). Esto podría deberse a la multiplicidad de
factores que influyen en el proceso y la redundancia en la función de algunos de ellos.
Con respecto a la hidrofobicidad de las superficies abióticas, los resultados
presentados muestran que la cepa K279a se adhiere rápidamente (6 h) al BS, una
superficie hidrofílica. Esta capacidad estaría relacionada con las cargas positivas de la
superficie de S. maltophilia, proporcionadas por proteínas localizadas en la membrana
externa (88). En cambio, K279a presentó una pobre adhesión a superficies hidróbicas
como los tubos de PP (Figura 22 b). Por su parte, la mutante mostró una adherencia
significativamente menor que la de la cepa salvaje en ambas superficies (Figura 22 a y b).
Estos resultados, sumados a los discutidos respecto a la hidrofobicidad bacteriana,
sugieren que los efectos pleiotrópicos de la mutación del gen rpfF incluyen la modificación
de la superficie bacteriana que conlleva a una menor adhesión.
La mutación en el gen rpfF produjo profundos cambios en la capacidad de formar
biofilms de S. maltophilia evaluada en microplacas. El comportamiento de K279a fue
similar al observado en BS (Figura 22), se observó una marcada adherencia a las 6 h de
incubación y un máximo nivel de adherencia a las 24 h (Figura 23 b). La mutante mostró
una adherencia significativamente menor en todo el periodo estudiado (Figura 23 b).
Además, la adición de DSF comercial produjo un incremento significativo de la biomasa
del biofilm de K279arpfF.

Resultados y Discusión
91
En las etapas de formación del biofilm, una vez que las bacterias se han adherido
comienzan a dividirse y la migración de las células a lo largo de la superficie lleva a la
formación de agregados conocidos como microcolonias. La movilidad tipo “twitching”,
mediada por el pili de tipo IV, participa en este proceso (111). La mutante rpfF produjo
halos de “twitching” significativamente menores que los de la cepa salvaje (Figura. 30 a
superior) y a diferencia de ésta, no presentó un patrón definido de adherencia (Figura 30
b). En P. aeruginosa, ha sido demostrado que la formación de microcolonias depende de
la movilidad tipo “twitching” regulada por QS (43). Nuestros resultados sugieren que en S.
maltophilia el sistema de QS, mediado por DSF, sería responsable de este tipo de
movilidad asociada a superficies. Además, el análisis por microscopía óptica de biofilms
de K279a demostró la presencia de abundantes microcolonias mientras que en los de la
mutante se observaron agregados celulares sin un patrón definido.
Pompilio et al. estudiaron 40 aislamientos clinicos de S. maltophilia y no encontraron
correlación entre la movilidad tipo “twitching” y la formación de biofilms (120). Sin
embargo, en otro estudio, la movilidad tipo “twitching ha sido correlacionada positivamente
con la formación de biofilms en S. maltophilia (81). En este sentido, en un trabajo anterior
de nuestro grupo hemos reportado que la motilidad asociada a superficies presentó buena
correlación con la adhesión a BS, PP y PS de aislamientos locales de S. maltophilia (117).
Estos resultados presentados demuestran que la ausencia de DSF generada por la
mutación del gen rpfF impacta negativamente en la formación de biofilms sobre diferentes
superficies desde la adhesión inicial.
Para profundizar el análisis sobre la arquitectura de biofilms maduros se utilizó la
microscopía confocal. El análisis por CLSM reveló que la cepa salvaje, luego de 48 h de
incubación, forma estructuras tridimensionales características de un biofilm maduro, con
canales de agua entre ellas. Los canales de agua son parte integral de la estructura del
biofilm ya que proveen de una vía de circulación de nutrientes y de intercambio de
productos metabólicos con el medio (28). La mutante rpfF no mostró un crecimiento
confluente sino que formó un gran número de agregados celulares dispersos y no
desarrolló una estructura tridimensional. Sin embargo, en presencia de DSF comercial la
mutante contrarrestó parcialmente dicha deficiencia.
La función del QS en la formación de biofilms de Xcc ha sido analizada por
diferentes autores. Dow et al (47) reportaron que una mutante rpfF de Xcc crece formando

Resultados y Discusión
92
agregados cuando es cultivada en el medio L y la adición de DSF produce la dispersión
de los mismos. Además, como la cepa salvaje no crece formando agregados llegaron a la
conclusión que la función del DSF es la dispersión del biofilm en esta especie. Sin
embargo, Torres et al (141) encontraron que la formación de biofilms de Xcc en medio
mínimo requiere un preciso control de la cantidad de DSF ya que la ausencia o la
sobreproducción del mismo afectan negativamente la formación de biofilms estructurados.
Además observaron que la virulencia de Xcc también requiere un preciso control de la
producción de DSF.
En concordancia con los resultados de Torres et al en Xcc, Alavi et al encontraron
que una mutación en el gen rpfF impacta negativamente en la formación de biofilms de S
maltophilia R551-3, una cepa asociada a plantas (4). Estos resultados concuerdan con los
presentados en esta tesis, los cuales demuestran la importancia de DSF en la formación
de biofilms de la cepa clínica K279a.
Una parte fundamental del biofilm es su matriz de EPS que le otorga resistencia
frente a distintos biocidas. En Xcc la mutación en rpfF genera una disminución de la
producción de EPS (13), sin embargo, los resultados presentados muestran que la cepa
K279a produce cantidades significativamente menores de EPS que su mutante rpfF. Esto
nos llevó a estudiar en profundidad la composición química de las EPS de ambas cepas
mediante la cuantificación de hidratos de carbono y de proteínas, y el análisis por
espectroscopia ATR-FTIR. La cepa K279a produjo mayor cantidad de exopolisacáridos
por mg de EPS que la mutante (Tabla 5). Por el contrario, la mutante produjo cantidades
mayores de proteínas por mg de EPS que la cepa salvaje (Tabla 5). Por lo tanto, la
relación hidratos de carbono/proteínas en la EPS es mucho mayor para la cepa salvaje
(0,48) que para la mutante (0,14). En concordancia, el espectro de ATR-FTIR de las EPS
de la mutante mostró mayor intensidad de absorción en los picos de amida I y amida II
que el de la cepa salvaje. Además, el pico de polisacáridos presentó una intensidad
ligeramente mayor en la cepa K279a que en su mutante. En la zona de los grupos
funcionales CH3 y CH2 de los lípidos no se observó diferencia entre ambas cepas (Figura
29). Por lo tanto, la mutación en el gen rpfF produce un cambio en la composición relativa
de los compuestos presentes en las EPS. El exopolisacarido de S. maltophilia presenta
una estructura novedosa respecto al de otras bacterias, consiste en una unidad repetitiva
de cuatro azucares compuesta por tres residuos de ácido urónico y un residuo D-lactato
que aporta otra carga negativa. Además presenta dos grupos o-acetilo por subunidad. La

Resultados y Discusión
93
abundancia de cargas negativas que posee es una característica compartida con el
alginato de P. aeruginosa y con el exopolisacarido de Inquilinus limosus, y se ha sugerido
que podría resultar en una ventaja para la persistencia en el pulmón de estos dos
organismos de relevancia en infecciones de pacientes fibroquisticos (24).
Por último, se investigó el rol del QS en la virulencia de S. maltophilia. La disrupción
del gen rpfF impactó negativamente en la actividad de DNasa y lipasa. Previamente,
Fouhy et al (62) demostraron que la mutación en el gen rpfF producía una disminución de
la actividad de proteasa. Estas tres enzimas han sido asociadas a la virulencia de S.
maltophilia (18).
La resistencia al estrés oxidativo es otra característica importante para la
patogénesis bacteriana ya que durante el proceso de infección las bacterias se enfrentan
a especies reactivas del oxigeno producidas por las células fagocíticas. La mutante rpfF
mostró una marcada disminución en la resistencia al peróxido de hidrogeno, lo que
sugiere la participación del sistema de QS en la regulación de enzimas antioxidantes. En
P. aeruginosa el QS participa en la resistencia al peróxido de hidrogeno y al anión
superóxido a través de la regulación de la expresión de las enzimas antioxidantes SOD y
catalasa (72).
Para profundizar los estudios sobre el rol biológico del QS en la virulencia de S.
maltophilia se utilizó el modelo de G. mellonella, cuyo sistema inmune comparte
homología estructural y funcional con el sistema inmune innato de mamíferos (80). Este
modelo fue utilizado anteriormente para evaluar la virulencia en esta especie (100, 110).
Solo la cepa salvaje presentó efecto letal en este modelo, sugiriendo el rol del QS en la
regulación de factores de virulencia. La ausencia de efecto letal en la mutante
correlaciona con su menor capacidad de formar biofilms, con la ausencia de actividad de
las exoenzimas lipasa y DNasa, y con su menor resistencia al estrés oxidativo.
En conclusión, el sistema rpfF/DSF juega un papel importante en la formación de biofilms
y en la virulencia de S. maltophilia.

Resultados y Discusión
94
Sección V
Función del hierro en la formación de biofilms y en la resistencia al estrés oxidativo de S. maltophilia
La formación de biofilms y la resistencia al estrés oxidativo son factores de
virulencia de gran relevancia en la patogenia bacteriana. El crecimiento en biofilms facilita
la persistencia en el huésped, la evasión de la respuesta inmune y la supervivencia frente
a altas concentraciones de antimicrobianos (38, 45, 119). Diferentes señales ambientales
regulan la formación de biofilms y la resistencia al estrés oxidativo, entre ellas, el hierro ha
sido ampliamente estudiado en numerosas especies bacterianas, sin embargo, su papel
es complejo. En P aeruginosa se demostró que el transporte activo de hierro y la
presencia de niveles intracelulares adecuados del mismo son necesarios para la
formación de biofilms (12), además, la limitación de hierro generada por la lactoferrina
lleva a la producción de biofilms con poco espesor (134). Por el contrario, en
Acinetobacter baumannii la formación de biofilms se incrementa en condiciones limitantes
de hierro (140). Hasta el momento no hay publicaciones que describan el efecto del hierro
en la formación de biofilms de S. maltophilia.
Por otra parte, el metabolismo en organismos aeróbicos genera especies reactivas
del oxigeno (ERO), también las células fagocíticas del sistema inmune innato generan
grandes cantidades de ERO, sin embargo, los microorganismos han desarrollado
estrategias para contrarrestar el efecto tóxico que éstas producen. Una estrategia consiste
en sintetizar enzimas antioxidantes cuya regulación está sujeta a señales ambientales.
Con el propósito de estudiar el papel del hierro y la participación del sistema Fur en
la formación de biofilms y la resistencia al estrés oxidativo se realizaron estudios
comparativos entre la cepa K279a y su mutante Fur obtenida por primera vez en S.
maltophilia durante el desarrollo de esta tesis.
1. Efecto de la disponibilidad de hierro en la formación de biofilms y la producción de EPS
El efecto del hierro en la formación de biofilms de S. maltophilia fue evaluado
utilizando el ensayo cuantitativo en microplacas de PS y por microscopía confocal. Para
esto se realizaron cultivos de K279a y su mutante Fur60 en TSB (condición no limitante

Resultados y Discusión
95
de hierro) y en TSB suplementado con dipiridilo (Dip 200 µM) para lograr la condición de
limitación de hierro.
1.1. Análisis de biofilms formados en microplacas
Para investigar la participación del sistema Fur en la regulación por hierro de la
formación de biofilms, se utilizó la mutante Fur60 derivada de K279a. Si el sistema Fur
estuviese implicado en la regulación de la formación de biofilms se esperaría que la
mutante Fur, independientemente de la presencia del quelante de hierro en el medio,
forme biofilms con características similares al de la cepa salvaje en condiciones limitantes
de hierro.
La figura 35 muestra la producción de biofilms de 48 h evaluada como la relación
entre la biomasa (tinción con cristal violeta) y la DO546 del crecimiento (DO546 de CV/DO546
del cultivo), que permite independizarse de posibles efectos en el crecimiento generados
por la adición del quelante. La producción de biofilms de K279a en condiciones limitantes
de hierro fue significativamente mayor que la producida en condiciones no limitantes del
mismo (P < 0,05). En medio TSB la producción de biofilm fue mayor en la mutante Fur
que en la cepa salvaje en la misma condición (P < 0,01), además, la producción de biofilm
de Fur60 no fue afectada por el agregado del quelante.
TSB TSB-Dip 200µµµµM0
1
2
3K279aFur60
**
DO
546
de C
V/D
O54
6de
l cul
tivo
Figura 35. Formación de biofilms de S. maltophilia K279a y su mutante Fur60 en microplacas. Los
cultivos en TSB y TSB suplementado con Dip 200 µM se incubaron a 35 ºC por 48 h. La biomasa

Resultados y Discusión
96
del biofilm fue cuantificada por tinción con CV. Los resultados se expresan como la media ±
desviación estándar de la relación entre la DO del CV y del crecimiento (DO546 de CV/DO546 del
cultivo) de un experimento representativo.*representa diferencias estadísticamente significativas.
1.2. Análisis de biofilms por CLSM
Para profundizar el análisis del efecto del hierro en la arquitectura de los biofilms
formados sobre superficies abióticas se utilizó CLSM. La misma permite visualizar
muestras totalmente hidratadas y determinar el espesor del biofilm que proporciona una
medida de la extensión espacial del mismo. Para estos estudios se realizaron cultivos en
condiciones limitantes y no limitantes de hierro en las cámaras Lab Tek, Nunc® y los
biofilms formados en la base de dichas cámaras se visualizaron mediante tinción con
Syto9 (Molecular Probes®).
Como se observa en la imagen de un plano x-y (Figura 36) la cepa salvaje K279a
cultivada en condiciones no limitantes de hierro desarrolló una monocapa de células con
microcolonias distribuidas por toda la superficie, mientras que en condiciones limitantes
de hierro formó un biofilm más denso y estructurado. Por otra parte, la mutante Fur, tanto
en presencia como en ausencia del quelante, formó un biofilm más denso y con
microcolonias de mayor tamaño que la cepa salvaje en ausencia de Dip. La proyección en
el eje z de la serie de imágenes obtenidas a partir de los cortes en profundidad permite
analizar el espesor del biofilm. El espesor promedio del biofilm de la cepa salvaje en
condiciones no limitantes de hierro fue de 30 µm mientras que en condiciones limitantes
del mismo fue de 50 µm. Por otra parte, el biofilm de la mutante Fur60 tanto en
condiciones limitantes como no limitantes de hierro presentó mayor espesor (70 µm) que
la cepa salvaje (Figura 36). La reconstrucción tridimensional realizada con el programa
ImageJ permitió revelar la arquitectura del biofilm. En K279a estaba compuesta por
estructuras con forma de picos separados entre sí por canales por los que podría circular
medio de cultivo con el consiguiente intercambio de nutrientes. Se observó que K279a en
presencia de Dip y la mutante Fur en presencia y ausencia del mismo presentaron no solo
picos más altos sino también mayor densidad de los mismos (Figura 37).
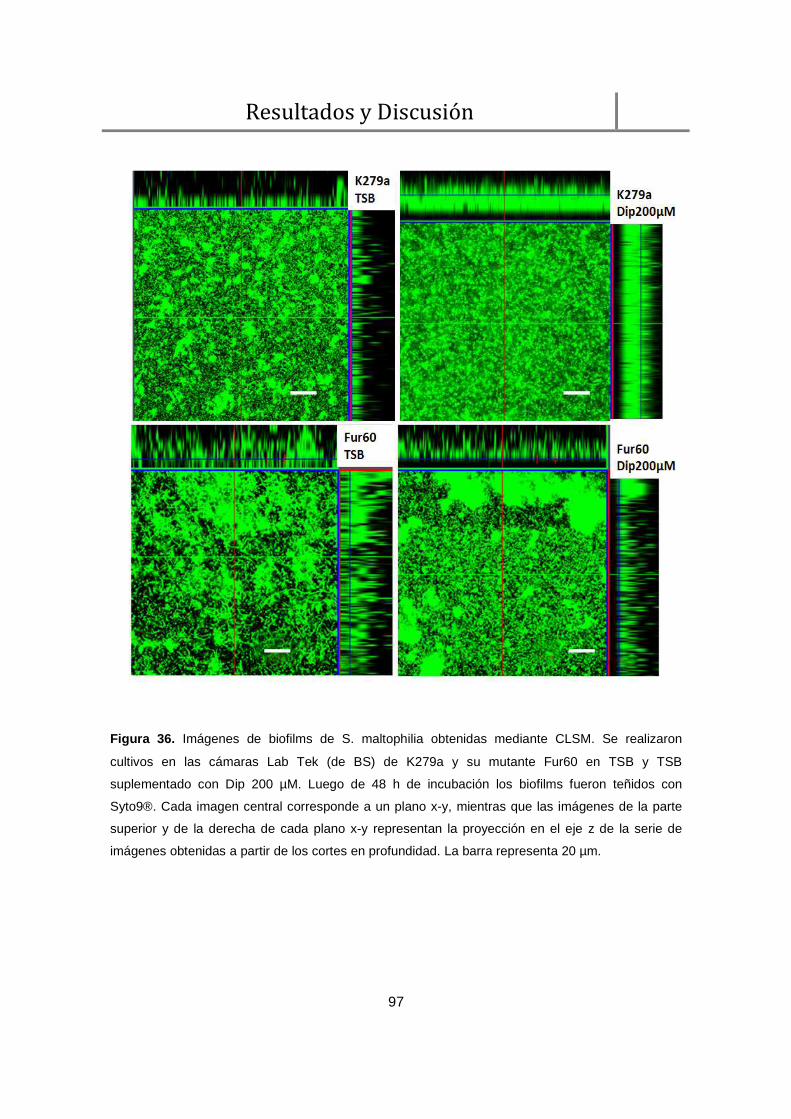
Resultados y Discusión
97
Figura 36. Imágenes de biofilms de S. maltophilia obtenidas mediante CLSM. Se realizaron
cultivos en las cámaras Lab Tek (de BS) de K279a y su mutante Fur60 en TSB y TSB
suplementado con Dip 200 µM. Luego de 48 h de incubación los biofilms fueron teñidos con
Syto9®. Cada imagen central corresponde a un plano x-y, mientras que las imágenes de la parte
superior y de la derecha de cada plano x-y representan la proyección en el eje z de la serie de
imágenes obtenidas a partir de los cortes en profundidad. La barra representa 20 µm.

Resultados y Discusión
98
Figura 37. Reconstrucción tridimensional de la serie de imágenes obtenidas mediante CLSM en
los planos x-y a través del eje z de biofilms de S. maltophilia. Las reconstrucciones fueron
realizadas con el programa ImageJ.
1.3. Producción de EPS
Se evaluó la producción de EPS mediante la técnica de precipitación con etanol y
determinación del peso seco. Para ello, se utilizaron sobrenadantes de cultivos
planctónicos en LB-glu 0,1 % y LB-glu 0,1 % suplementado con Dip 200 µM incubados
con agitación durante 48 h a 35 ºC. La figura 38 muestra que la cepa K279a en
condiciones limitantes de hierro produjo mayor cantidad de EPS (246,3 ± 20,5) que en
condiciones no limitantes del mismo (64,9 ± 3,3). La mutante Fur60 tanto en condiciones
limitantes como no limitantes de hierro produjo mayor cantidad de EPS (225,9 ± 11,5 y
201,1 ± 38,0, respectivamente) que la cepa salvaje en ausencia de Dip.

Resultados y Discusión
99
LB-glu 0,1% LB-glu 0,1% Dip 200µµµµM0
100
200
300
K279aFur60
**
EP
S (
µµ µµg)/
biom
asa
(mg)
Figura 38. Producción de EPS de S. maltophilia K279a y su mutante Fur60 cultivadas durante 48
h, a 35 ºC con agitación. Los resultados se expresan como la media ± desviación estándar de la
relación entre el peso seco de las EPS (µg) y el peso seco de la biomasa del cultivo (mg), de un
experimento representativo.*representa diferencias estadísticamente significativas.
El análisis de los componentes químicos de los precipitados obtenidos a partir de
cultivos de K279a y Fur60 en LB-glu 0,1 % realizado mediante la cuantificación de
hidratos de carbono con glucosa como estándar (50) y de proteínas (19) demostró que la
cepa K279a produjo cantidades similares de exopolisacaridos (0,25 ± 0,02) y de proteínas
(0,49 ± 0,03) por mg de peso seco de EPS que la mutante que la mutante (0,26 ± 0,05 y
0,52 ± 0,05, respectivamente) (Tabla 7). Por lo tanto, la relación hidratos de
carbono/proteínas en la EPS es similar para ambas cepas (0,51 y 0,50, respectivamente)

Resultados y Discusión
100
Tabla 7. Composición de las EPS de cepas de S. maltophilia
Cepas de
S.
maltophilia
EPS
(µg de EPS/mg
de biomasa)
Análisis químico de EPS
Hidratos de carbono
(mg de glucosa/mg de
peso seco de EPS)
Proteínas
(mg de proteínas/mg
de peso seco de EPS)
Relación Hidratos de
carbono/ Proteínas
(mg de glucosa/mg de
proteínas)
K279a 64,9 ±3,3 0,25 ± 0,02 0,49 ± 0,03 0,51
Fur60 201,1 ± 38,0 0,26 ± 0,05 0,52 ± 0,05 0,50
Para profundizar el estudio de la composición molecular de las EPS se utilizó
espectroscopia ATR-FTIR en las condiciones descriptas en la Sección IV, ítem 3.
En los espectros de ATR-FTIR de las EPS de K279a y su mutante Fur60 se puede
observar que la intensidad de absorción de todos los picos es similar para ambas cepas,
en concordancia con los resultados del análisis químico de los componentes de los
precipitados (Figura 39). Por lo tanto, la mutante produce mayor cantidad de EPS pero de
la misma composición química que la cepa salvaje.

Resultados y Discusión
101
Figura 39. Espectros ATR-FTIR de EPS de K279a (azul) y la mutante Fur60 (rojo). En los
espectros se indican los picos de absorción correspondientes a los grupos funcionales asociados a
lípidos (3100 a 2800 cm-1), proteínas (1700-1500 cm-1) y polisacáridos (1200-900 cm-1).
2. Efecto de la disponibilidad de hierro en la respuesta al estrés oxidativo
2.1. Determinación del tipo de isoenzima de superóxido dismutasa
(SOD)
En muchas especies bacterianas las isoenzimas de SOD están reguladas por la
concentración de hierro en el medio y la tensión de oxigeno (56, 75). Teniendo en cuenta
estos factores, se estudió la presencia de esta enzima mediante electroforesis en geles de
poliacrilamida en condiciones no desnaturalizantes y tinción con NBT. Se utilizaron
extractos de la cepa K279a y su mutante Fur60 cultivadas en TSB suplementado con
diferentes concentraciones de Dip (0, 100 y 200 µM Figura 40 a), Además, en la cepa
salvaje, se evaluó el efecto de la agitación y del tiempo de incubación de los cultivos (12,
24, 48 h Figura 40 b).
En condiciones limitantes y no limitantes de hierro se observó una única banda con
actividad de SOD tanto en la cepa salvaje como en la mutante Fur (figura 40). Los

Resultados y Discusión
102
experimentos de inhibición mostraron que la única banda detectada corresponde a la
isoenzima Mn-SOD ya que la actividad enzimática no fue inhibida por H2O2 ni por NaCN,
al igual que el control de Mn-SOD purificada de E. coli (datos no mostrados). Sin
embargo, la movilidad electroforética de la Mn-SOD de K279a fue diferente a la que
presentó la Mn-SOD de E. coli, lo que podría representar diferencias estructurales entre
ambas enzimas.
En la figura 40 b se observa una banda con actividad de SOD en los extractos de
K279a obtenidos a partir de cultivos de 12 h con agitación pero no en los provenientes de
cultivos estáticos. En estos últimos, se observan bandas con actividad SOD a las 24 y 48
h de incubación y las mismas son más tenues que las correspondientes a los cultivos con
agitación. Nuevamente, los experimentos de inhibición mostraron que la única banda
detectada corresponde a la isoenzima Mn-SOD.
Figura 40. Actividad de SOD en geles nativos revelados con NBT. a) Extractos de K279a y Fur60
obtenidos de cultivos en TSB y TSB suplementado con Dip100 y 200 µM. b) Extractos de K279a
obtenidos de cultivos estáticos (Es) y con agitación (Ag) en TSB, de 12, 24 y 48 h de incubación. P:
Mn-SOD purificada de E. coli.
2.2. Actividad de SOD en cultivos planctónicos y biofilms
Se realizaron estudios comparativos de la actividad de SOD en extractos
provenientes de células de cultivos planctónicos y biofilms de K279a y de Fur60 en
condiciones limitantes y no limitantes de hierro. Los biofilms se formaron en placas de
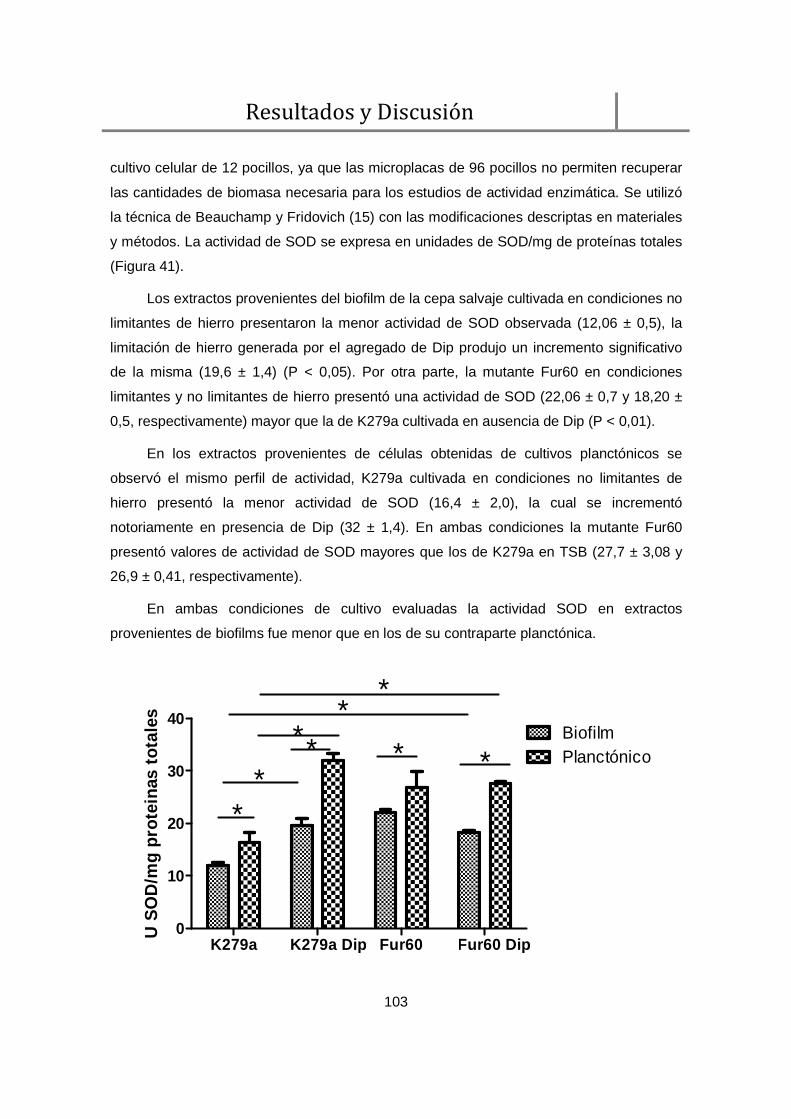
Resultados y Discusión
103
cultivo celular de 12 pocillos, ya que las microplacas de 96 pocillos no permiten recuperar
las cantidades de biomasa necesaria para los estudios de actividad enzimática. Se utilizó
la técnica de Beauchamp y Fridovich (15) con las modificaciones descriptas en materiales
y métodos. La actividad de SOD se expresa en unidades de SOD/mg de proteínas totales
(Figura 41).
Los extractos provenientes del biofilm de la cepa salvaje cultivada en condiciones no
limitantes de hierro presentaron la menor actividad de SOD observada (12,06 ± 0,5), la
limitación de hierro generada por el agregado de Dip produjo un incremento significativo
de la misma (19,6 ± 1,4) (P < 0,05). Por otra parte, la mutante Fur60 en condiciones
limitantes y no limitantes de hierro presentó una actividad de SOD (22,06 ± 0,7 y 18,20 ±
0,5, respectivamente) mayor que la de K279a cultivada en ausencia de Dip (P < 0,01).
En los extractos provenientes de células obtenidas de cultivos planctónicos se
observó el mismo perfil de actividad, K279a cultivada en condiciones no limitantes de
hierro presentó la menor actividad de SOD (16,4 ± 2,0), la cual se incrementó
notoriamente en presencia de Dip (32 ± 1,4). En ambas condiciones la mutante Fur60
presentó valores de actividad de SOD mayores que los de K279a en TSB (27,7 ± 3,08 y
26,9 ± 0,41, respectivamente).
En ambas condiciones de cultivo evaluadas la actividad SOD en extractos
provenientes de biofilms fue menor que en los de su contraparte planctónica.
K279a K279a Dip Fur60 Fur60 Dip0
10
20
30
40BiofilmPlanctónico
*
**
** **
*
U S
OD
/mg
prot
eina
s to
tale
s

Resultados y Discusión
104
Figura 41. Actividad de SOD de extractos crudos obtenidos a partir de células provenientes de
cultivos planctónicos y biofilms de S. maltophilia. Los cultivos fueron realizados en TSB y TSB
suplementado con Dip 200 µM e incubados durante 48 h a 35 °C. Los resultados se expresan
como la media ± desviación estándar de la relación entre las U de SOD y las proteínas totales, de
un experimento representativo.*representa diferencias estadísticamente significativas.
2.3. Niveles de especies reactivas del oxigeno (ERO) en el biofilm
Los niveles de ERO en biofilms de S. maltophilia se evaluaron mediante el ensayo de
NBT en biofilms formados en microplacas de PS (2, 7). En presencia de anión superóxido
la reducción del NBT a azul de formazán es proporcional a los niveles del mismo en el
medio (15).
El biofilm de K279a formado en condiciones no limitantes de hierro presentó niveles
de ERO 2,5 veces mayores que en presencia de Dip, estos últimos fueron similares a los
de los biofilm de Fur60 formado en condiciones limitantes y no limitantes de hierro (P <
0,01) (Figura 42). Estos resultados concuerdan con la actividad de SOD evaluada
anteriormente.
TSB TSB-Dip 200µµµµM0.0
0.1
0.2
0.3
0.4
0.5K279a
Fur60*
*
DO
546
de N
BT/
Bio
mas
a(C
V/D
O)
Figura 42. Determinación de las ERO de biofilms de S. maltophilia. Los cultivos en TSB y TSB
suplementado con Dip 200 µM fueron incubados durante 48 h a 35 °C en condiciones estáticas.
Los resultados se expresan como la media ± desviación estándar de la relación entre la DO546 de

Resultados y Discusión
105
NBT y la biomasa del biofilm evaluada como la relación entre la DO546 de CV y la DO546 del
crecimiento, de un experimento representativo.*representa diferencias estadísticamente
significativas.
3. Evaluación del efecto del hierro en la producción de DSF
Los resultados presentados indican que en S. maltophilia K279a el QS y el hierro, a
través del sistema Fur, están implicados en la formación de biofilms y en la resistencia al
estrés oxidativo. La expresión de dichas propiedades podría ser la resultante de la
interacción de ambas señales regulatorias. Para investigar la eventual relación entre
ambas señales, se evaluó el efecto del hierro en la producción de DSF. Para este
propósito se utilizó el bioensayo descripto por Barber et al (13) que detecta la molécula de
DSF presente en extractos de sobrenadantes de cultivo por la restauración de la
producción de endoglucanasa, una enzima regulada por QS, en una mutante rpfF de Xcc.
Los resultados se expresan como la relación entre las U de endoglucanasa y el
crecimiento bacteriano.
Se realizaron cultivos en el medio SSC y SSC suplementado con FeCl3 100 µM que
representan la condición limitante y no limitante de hierro, respecivamente. En estos
experimentos no se puede utilizar Dip porque en el proceso de extracción también se
extrae el quelante, que actúa como inhibidor del crecimiento de la cepa reportera.
En condiciones de limitación de hierro se produjo un aumento estadísticamente
significativo (P < 0,05) de la producción de la molécula señal en la cepa K279a (18,7 ±
0,6) respecto a cuando crece en condiciones no limitantes de hierro (16,2 ± 0,4). Por su
parte, la mutante Fur60, en ambas condiciones, produjo cantidades de DSF
significativamente mayores (17,9 ± 0,5 y 18,1 ± 0,5, respectivamente. Figura 43.) que la
cepa salvaje en condiciones no limitantes de hierro (P < 0,05).

Resultados y Discusión
106
K279aSSC+Fe3+ K279a SSC F60SSC+Fe3+ F60 SSC14
16
18
20 **
U d
e en
dogl
ucan
asa/
DO
540
Figura 43. Evaluación de la producción de DSF por S. maltophilia mediante el ensayo de Barber et
al (13). Se obtuvieron extractos a partir de cultivos estáticos incubados a 35 °C por 48 h de K279a
y Fur60 en condiciones limitantes (SSC) y no limitantes de hierro (SSC FeCl3 100 µM). Los
resultados se expresan como la media ± desviación estándar de la relación entre las U de
endoglucanasa y el crecimiento bacteriano de un experimento representativo.*representa
diferencias estadísticamente significativas.
4. Evaluación de la virulencia en el modelo de infección de Galleria
mellonella
Con el objetivo de evaluar el efecto que produce la mutación del gen fur sobre la
virulencia de S. maltophilia se utilizó el modelo de infección del lepidóptero G. mellonella.
Se utilizó la dosis de 105 ufc/larva para estudios comparativos entre la cepa K279a y su
mutante Fur60. La cepa K279a resultó letal para un 20 % de la población a las 48 h y para
el 40 % a las 96 h. La mutante Fur60 fue significativamente (P < 0,006) más virulenta ya
que produjo la muerte del 60 % de de la población a las 48 h y alcanzó el 80 % de
letalidad a las 96 h (Figura 44 y 45).

Resultados y Discusión
107
0 20 40 60 80 1000
20
40
60
80
100K279aFur60
Tiempo (h)
Sup
ervi
venc
ia (
%)
Figura 44. Ensayo de infección en larvas de Galleria mellonella. Luego de la inyección de 10 a 13
larvas por cepa con la dosis de 105 ufc/larva se registró diariamente el número de individuos
sobrevivientes y se graficaron las curvas de Kaplan-Mayer. El grupo control de larvas inyectadas
con solución fisiológica no presentó individuos muertos (datos no graficados). Los datos
corresponden a un experimento representativo. El análisis estadístico se realizó con el método de
Log-rank. Los datos corresponden a un experimento representativo.

Resultados y Discusión
108
Figura 45 . Larvas de G. mellonella a) inmediatamente después de ser inyectadas y b) luego de 96
h de haber sido inyectadas con la dosis 105 ufc/larva de una suspensión de Fur60. Durante la
infección las larvas desarrollan una progresiva melanización hasta morir.
5. Discusión
El hierro es una señal que regula la formación de biofilms en muchas especies
bacterianas, sin embargo, su papel es controversial. Mientras que en Legionella
pneumophila y en Staphylococcus aureus la limitación de hierro actúa induciendo la
formación de biofilms (79, 87) en otras especies, tales como Vibrio cholerae actúa
inhibiéndola (102). Más aún, la regulación por hierro es compleja incluso dentro de la
misma especie. En P. aeruginosa, la limitación de hierro bloquea las etapas tempranas de
la formación de microcolonias, y la actividad quelante de la lactoferrina humana disminuye
la formación de biofilms (12). Sin embargo, altas concentraciones de hierro también
inhiben la formación de biofilms y causan la dispersión de los ya formados (107). En
consecuencia, la formación de biofilms en P. aeruginosa ocurre dentro de un rango
limitado de concentraciones de hierro (1–100 µM) (108).
Los resultados presentados muestran que la limitación de hierro incrementa la
formación de biofilms de S. maltophilia. Dicha limitación produjo además, profundos
cambios en la estructura tridimensional del biofilm de K279a analizada por CLSM, la cual
se caracterizó por una mayor densidad celular y mayor organización espacial con picos de
mayor altura (Figura 36 y 37). Por su parte, la mutante Fur60 en condiciones limitantes y
no limitantes de hierro produjo mayor biomasa de biofilms, con mayor cantidad de picos
de mayor altura que los de la cepa salvaje en condiciones no limitantes de hierro. Por
tanto, el sistema Fur participa, al menos en parte, en la regulación de la formación de
biofilms de S. maltophilia.
En P. aeruginosa el sistema Fur regula numerosos genes involucrados en la
formación de biofilms y la virulencia (16, 71). La limitación de hierro produce un aumento
de la transcripción de genes que codifican alginato (EPS), autoinductores y factores de
virulencia tales como proteasas, sideróforos y superóxido dismutasa (Mn-SOD) (16, 146).
En S. maltophilia, la producción de EPS también se vio afectada por la
disponibilidad de hierro. En presencia de Dip la cepa K279a, al igual que la mutante Fur
en ambas condiciones de cultivo, produjo mayor cantidad de EPS que en condiciones no

Resultados y Discusión
109
limitantes de hierro (Figura 38). Además, a diferencia de lo observado en la mutante
K279arpfF (seción IV ítem 3), la composición relativa de proteínas y polisacáridos de las
EPS de la mutante Fur fue similar a la de las EPS de la cepa salvaje (Tabla 7) y los
espectros de ATR-FTIR corroboraron dicha similitud (Figura 39). Estos resultados
sugieren que el hierro a través del sistema Fur participa en la producción de EPS, un
importante componente estructural de la matriz de los biofilms. Esto concuerda con lo
reportado en P. aeruginosa donde se demostró que mutantes Fur producen niveles de
alginato mayores que la cepa salvaje (71).
Para comenzar a estudiar la influencia del hierro en la resistencia al estrés
oxidativo, se investigó la presencia de isoenzimas de SOD en S. maltophilia. Los
experimentos de inhibición, en diferentes condiciones de cultivo, mostraron que la cepa
K279a posee solo la isoenzima Mn-SOD. Al analizar el genoma de K279a se encontraron
varios genes que codificarían para SOD, 2 genes sodA (Smlt2828 y Smlt3238)
codificantes de Mn-SOD, el gen sodB (Smlt1616b) que codificaría para la isoenzima Fe-
SOD, el gen sodC (Smlt0161) que codificaría para una Cu/Zn-SOD. Sin embargo, los
productos de los genes sodB y sodC no fueron detectados en las condiciones
experimentales utilizadas. Los resultados presentados sugieren que solo el o los genes
sodA se expresan en K279a, en concordancia con un estudio previo que demostró que la
cepa S. maltophilia WZ2 produce solo Mn-SOD (98). De esta forma, al ser la isoenzima
Mn-SOD la única presente en K279a, se puede evaluar la regulación por hierro de SOD
mediante la actividad de la enzima en extractos crudos.
Los estudios comparativos realizados entre la cepa K279a y Fur60 en presencia y
ausencia de Dip mostraron que la limitación de hierro produjo un incremento
estadísticamente significativo en la actividad SOD de la cepa salvaje tanto en cultivos
planctónicos como en biofilms (Figura 41). En consecuencia, el sistema Fur en presencia
de hierro regularía negativamente la transcripción del gen sodA de S. maltophilia. Estos
resultados concuerdan con los de Hassett et al., quienes demostraron que mutantes Fur
de P. aeruginosa presentan un incremento en la actividad de Mn-SOD y catalasa pero una
reducción en la actividad de SOD total (74). También en E. coli la limitación de hierro
incrementa la expresión de Mn-SOD (70).
En todas las condiciones de cultivo evaluadas la actividad SOD en células
provenientes del biofilm fue menor que en las de su contraparte planctónica (Figura 41).

Resultados y Discusión
110
La menor expresión de la enzima en el biofilm podría ser la resultante de una menor
actividad metabólica. La limitación de hierro condujo a una mayor actividad de SOD y
consecuentemente, los niveles de ERO del biofilm fueron menores (Figura 42). En
conclusión, la limitación de hierro protege al biofilm del estrés oxidativo, probablemente al
favorecer la expresión de la isoenzima Mn-SOD.
Para investigar la eventual regulación por hierro de la señal de QS se evaluó la
producción de DSF en cultivos planctónicos de K279a y de su mutante Fur en condiciones
limitantes y no limitantes de hierro. Los resultados demuestran que el hierro, a través del
sistema Fur, es una señal que regula negativamente la síntesis de DSF.
Se han descripto diferentes interacciones entre el hierro, a través del sistema Fur, y el
QS en varias especies bacterianas. En Pseudomonas syringae una mutación en la
proteína Fur redujo la producción del autoinductor, una acil homoserino lactona (33). En
Vibrio vulnificus se demostró que el complejo hierro-Fur regula negativamente la
expresión del gen smcR que codifica un regulador de respuesta de QS similar a LuxR de
V. harveyi (92) y de esta forma coordinan la expresión de factores de virulencia. En P.
aeruginosa la limitación de hierro produce un aumento de la producción del autoinducor
de uno de sus sistemas de QS denominado pseudomonas quinolona (PQS), el cual regula
positivamente la expresión de enzimas extracelulares, metabolitos secundarios y factores
de virulencia. (113).
La mayor virulencia de la mutante Fur en el modelo de G. mellonella correlaciona con
su mayor capacidad de formar biofilms y con su mayor resistencia al estrés oxidativo.
Los resultados presentados en la Sección IV y V indican que en S. maltophilia K279a
los genes relacionados con la formación de biofilms y con la producción de otros
potenciales factores de virulencia están regulados positivamente por QS, y negativamente
por concentraciones no limitantes de hierro, a través del sistema Fur. Además, en esta
tesis se comprobó por primera vez la relación entre el sistema Fur-hierro y el sistema de
QS mediado por DSF en S. maltophilia.

Conclusiones

Conclusiones
112
El análisis de los resultados obtenidos durante el desarrollo de esta Tesis permitió
elaborar las siguientes conclusiones:
Tipificación de aislamientos locales de S. maltophilia recuperados de infecciones
asociadas al uso de productos médicos
� El sistema API20NE resultó de utilidad para confirmar la identificación de los
aislamientos de S. maltophilia pero fue de escasa utilidad para la tipificación de los
mismos ya que solo permitió agruparlos en 3 biocódigos
� Las técnicas de tipificación molecular ERIC-PCR y REP-PCR generaron perfiles
reproducibles y altamente discriminatorios. Las mismas revelaron una gran
heterogeneidad genética entre los aislamientos locales de S. maltophilia,
sugiriendo que la mayoría de los episodios nosocomiales fueron independientes.
Estas técnicas, junto con la información epidemiológica, son útiles para la
tipificación de aislamientos de esta especie, aunque aquellos con perfiles
indistinguibles deben ser analizados con la técnica de PFGE, que posee un mayor
poder discriminatorio.
Producción de potenciales factores de virulencia en aislamientos locales de S.
maltophilia
� La capacidad de formar biofilms es un importante factor de virulencia bacteriano.
Todos los aislamientos locales, salvo Sm61 que no forma biofilms, produjeron
biofilms fuertes.
� Los 63 aislamientos estudiados fueron lipolíticos y proteolíticos, aunque solo en el
82,53 % se detectó el gen stmPr1 que codifica la principal proteasa.
� La producción de sideróforos se detectó en todos los aislamientos clínicos gracias
a las modificaciones realizadas al medio empleado en la técnica en placa CAS
agar. Las modificaciones consistieron en reemplazar el medio M9 por SS con el
agregado de casamino ácidos y Dip (SSC-CAS-Dip).

Conclusiones
113
� El gen narG, codificante de una nitrato reductasa, se detectó en el 60,31 % de los
aislamientos. La presencia del gen narG y la capacidad de reducir nitrato (57,1 %),
relacionadas con el crecimiento en microaerofilia, detectadas en los aislamientos
de S. maltophilia facilitaría el crecimiento en biofilms, y de esta forma
incrementaría la patogenicidad.
� Los aislamientos mostraron diferentes niveles de resistencia al estrés oxidativo, el
52,38 % presentó alta resistencia al estrés oxidativo.
� El modelo de infección de Galleria mellonella permitió comparar la virulencia de
algunos aislamientos locales de S. maltophilia, seleccionados en función de sus
potenciales factores de virulencia, y de K279a. En este modelo se evidenciaron
tres niveles de virulencia y el efecto letal mostró correlación con la resistencia al
estrés oxidativo.
Sistema de captación de hierro de alta afinidad de S. maltophilia K279a y su
regulación por el sistema Fur
� Se obtuvieron por primera vez en S. maltophilia mutantes espontaneas Fur de
K279a. Los clones Fur42 y Fur60 presentaron una mutación puntual. Fur42
presentó una mutación que causa la sustitución de un único aminoácido (Asp�Gly
en la posición 5) y Fur60 presentó una mutación en la secuencia -10 de la región
promotora putativa (T�C). Las mutantes Fur produjeron sideróforos en forma
constitutiva, lo que sugiere la falta de regulación por el sistema Fur-Fe2+.
� Se detectaron dos OMPs de K279a cuya expresión está regulada por el sistema
Fur-Fe2+. Estas OMPs fueron identificadas por espectrometría de masa como
FepA, un potencial receptor de enterobactina, y CirA, un potencial receptor de
colicina I de K279a. Estos receptores permitirian la captación del complejo hierro-
enterobactina y de los productos de degradación lineales de dicho complejo
respectivamente. CirA actuaría como una vía alternativa para incorporar hierro.
Mediante análisis in silico se determinaron las potenciales cajas Fur de fepA y cirA.
Finalmente, se obtuvo una secuencia consenso de la caja Fur putativa de K279a.
� Todos los aislamientos locales de S. maltophilia y la cepa K279a produjeron
sideróforos de tipo catecol en condiciones limitantes de hierro y no produjeron
sideroforos de tipo hidroxamato.

Conclusiones
114
� El análisis por TLC y la separación mediante HPLC sugieren que el sideróforo
producido por S. maltophilia es enterobactina. Se profundizará el estudio mediante
espectrometría de masa y resonancia magnética nuclear para confirmar la
identificación del sideróforo.
Función del sistema de señalización rpfF /DSF en la formación de biofilms y en la
virulencia de S. maltophilia
� La mutación en el gen rpfF de la cepa K279a produjo efectos pleiotrópicos que
afectaron características que participan en diferentes etapas de la formación de
biofilms. Entre estos efectos, la mutación produjo una marcada disminución de la
cosecha máxima y de la velocidad de crecimiento, fundamentalmente en
condiciones estáticas con baja tensión de oxígeno, y produjo la disminución de la
movilidad tipo “twitching” y de la hidrofobicidad superficial.
� La mutación en el gen rpfF impacta negativamente en la capacidad de formar
biofilms sobre diferentes superficies desde la adhesión inicial. El análisis por
CLSM reveló que la cepa salvaje, luego de 48 h de incubación, forma estructuras
tridimensionales características de un biofilm maduro, con canales de agua entre
ellas. La mutante K279arpfF no mostró un crecimiento confluente sino que formó
un gran número de agregados celulares dispersos y no desarrolló una estructura
tridimensional. Sin embargo, la presencia de DSF comercial restauró parcialmente
la capacidad de formar biofilms de la mutante.
� Una parte fundamental del biofilm es su matriz de EPS que le otorga resistencia
frente a distintos biocidas. La cuantificación de hidratos de carbono y de proteínas,
y el análisis por espectroscopia ATR-FTIR revelaron que la mutación en el gen
rpfF produce un cambio en la composición relativa de los compuestos presentes
en las EPS. La cepa K279a produjo mayor cantidad de exopolisacáridos por mg de
EPS que la mutante, mientras que la mutante produjo cantidades mayores de
proteínas.
� La disrupción del gen rpfF también afectó la producción de otros potenciales
factores de virulencia. La misma produjo la pérdida de la actividad de DNasa y
lipasa y una marcada disminución en la resistencia al peróxido de hidrogeno.
Previamente, Fouhy et al (62) demostraron que la mutación en el gen rpfF produce
una disminución de la actividad de proteasa.

Conclusiones
115
� La mutante rpfF, a diferencia de la cepa salvaje, no mostró efecto letal en el
modelo de virulencia de G. mellonella, lo cual correlaciona con su menor
capacidad de formar biofilms, con la ausencia de actividad de las exoenzimas
proteasa, lipasa y DNasa, y con su menor resistencia al estrés oxidativo.
Función del hierro en la formación de biofilms y en la resistencia al estrés oxidativo de S. maltophilia
� El hierro es una señal que regula negativamente la producción de EPS y la
formación de biofilms de K279a. La limitación de hierro incrementó no solo la
biomasa del biofilm sino también su estructura tridimensional determinada por
CLSM. La utilización de la mutante Fur60 reveló que dicho efecto depende del
sistema Fur.
� En la cepa K279a solo se detectó la isoenzima Mn-SOD aunque su genoma posee
también los genes sodB y sodC. La limitación de hierro condujo a una mayor
actividad de la isoenzima Mn-SOD en cultivos planctónicos y biofilms. Además, en
dicha condición, los niveles de ERO del biofilm también fueron menores. El
sistema Fur en presencia de hierro regularía negativamente la transcripción del
gen sodA de S. maltophilia. En conclusión, la limitación de hierro protegería al
biofilm del estrés oxidativo, al favorecer la expresión de la isoenzima Mn-SOD.
� El hierro, a través del sistema Fur, es una señal que regula negativamente la
síntesis de DSF.
� La mutación en el gen fur aumentó la virulencia de K279a en el modelo de G.
mellonella. La mayor virulencia de la mutante Fur60 correlaciona con su mayor
capacidad de formar biofilms y con su mayor resistencia al estrés oxidativo.
Conclusión final
En S. maltophilia K279a la formación de biofilms y la producción de otros
potenciales factores de virulencia están reguladas positivamente por el sistema de
señalización rpfF/DSF, y negativamente por concentraciones no limitantes de hierro, a
través del sistema Fur. Además, en esta tesis se comprobó por primera vez la relación
entre el sistema Fur-Fe2+ y el sistema de QS mediado por DSF en S. maltophilia.
La importancia del hierro como señal nos condujo a estudiar el sistema de
captación del mismo en S. maltophilia. El mismo está compuesto por los receptores de

Conclusiones
116
membrana externa FepA y CirA, regulados por el sistema Fur-hierro, responsables de
internalizar el sideróforo de S. maltophilia.

Resumen

Resumen
118
Stenotrophomonas maltophilia es un patógeno nosocomial emergente
multiresistente. El gen rpfF es responsable de la síntesis de DSF, su señal de quorum
sensing (QS). Para estudiar el papel del QS y del hierro en la formación de biofilms y la
producción de otros potenciales factores de virulencia, se utilizaron las cepas K279a y su
mutante K279arpfF y mutantes Fur (“ferric uptake regulator”) espontáneas obtenidas por
primera vez en S. maltophilia por nuestro grupo de trabajo. Además, se estudiaron 63
aislamientos locales provenientes de infecciones asociadas al uso de productos médicos.
Las técnicas de tipificación molecular ERIC-PCR y REP-PCR generaron perfiles
reproducibles y altamente discriminatorios. Las mismas revelaron una gran
heterogeneidad genética entre los aislamientos locales de S. maltophilia.
Se estudió la producción de potenciales factores de virulencia en los aislamientos
locales de S. maltophilia. La capacidad de formar biofilms es un importante factor de
virulencia bacteriano. La producción de sideróforos se detectó en todos los aislamientos
clínicos gracias a las modificaciones realizadas en la técnica en placa CAS agar. Se
detectó la presencia del gen narG codificante de una nitrato reductasa en el 60,31% de
los aislamientos, y del gen stmPr1, que codifica la principal proteasa, en el 82,53%,
mientras que el 52,38% presentó alta resistencia al estrés oxidativo. En el modelo de
virulencia de Galleria mellonella el efecto letal mostró correlación con la resistencia al
estrés oxidativo.
Se profundizó el estudio sobre los sideróforos y el sistema de captación de hierro de
alta afinidad de S. maltophilia, y su regulación por el sistema Fur. Todos los aislamientos
locales y K279a produjeron sideróforos de tipo catecol. El análisis por TLC y la separación
mediante HPLC sugieren que el sideróforo producido por S. maltophilia es enterobactina.
Para estudiar la posible regulación por el sistema Fur-Fe2+ se obtuvieron por primera
vez en S. maltophilia mutantes espontaneas Fur de K279a que produjeron sideróforos en
forma constitutiva, lo que sugiere la falta de regulación por el sistema Fur-Fe2+. Se
detectaron dos OMPs de K279a cuya expresión está regulada por el sistema Fur-Fe2+.
Estas OMPs fueron identificadas por espectrometría de masa como FepA, un potencial
receptor de enterobactina, y CirA, un potencial receptor de colicina I de K279a. Estos
receptores permitirían la captación del complejo hierro-enterobactina y de los productos
de degradación lineales de dicho complejo respectivamente. CirA actuaría como una vía

Resumen
119
alternativa para incorporar hierroOtros de los objetivos fue estudiar la función del sistema
de señalización rpfF/DSF en la formación de biofilms y en la virulencia de S. maltophilia.
La mutación en el gen rpfF de K279a produjo efectos pleiotrópicos que afectaron
características que participan en diferentes etapas de la formación de biofilms. Entre estos
efectos, produjo la disminución de la movilidad tipo “twitching” y de la hidrofobicidad
superficial.
La mutación en el gen rpfF impactó negativamente en la capacidad de formar
biofilms sobre diferentes superficies desde la adhesión inicial. El análisis por CLSM reveló
que la cepa salvaje forma estructuras tridimensionales características de un biofilm
maduro, con canales de agua entre ellas. La mutante K279arpfF no mostró un crecimiento
confluente ni desarrolló una estructura tridimensional. Sin embargo, la presencia de DSF
comercial restauró parcialmente la capacidad de formar biofilms de la mutante.
Una parte fundamental del biofilm es su matriz de EPS que le otorga resistencia
frente a distintos biocidas. El análisis por espectroscopia ATR-FTIR revelaron que la
mutación en el gen rpfF produce un cambio en la composición relativa de los compuestos
presentes en las EPS. La cepa K279a produjo mayor cantidad de exopolisacáridos por mg
de EPS que la mutante, mientras que la mutante produjo cantidades mayores de
proteínas.
La disrupción del gen rpfF también produjo la pérdida de la actividad de DNasa y
lipasa y una marcada disminución en la resistencia al peróxido de hidrogeno. En
consecuencia, K279arpfF, a diferencia de la cepa salvaje, no mostró efecto letal en el
modelo de virulencia de G. mellonella.
Por último se estudió la función del hierro en la formación de biofilms y en la
resistencia al estrés oxidativo de S. maltophilia. El hierro es una señal que regula
negativamente la producción de EPS y la formación de biofilms de K279a. La limitación de
hierro incrementó no solo la biomasa del biofilm sino también su estructura tridimensional
determinada por CLSM. La utilización de la mutante Fur60 reveló que dicho efecto
depende del sistema Fur.
En K279a solo se detectó la isoenzima Mn-SOD y la limitación de hierro condujo a
una mayor actividad de Mn-SOD en cultivos planctónicos y biofilms. Dicha limitación
protegería al biofilm del estrés oxidativo, al favorecer la expresión de la isoenzima Mn-
SOD.

Resumen
120
La mutación en el gen fur aumentó la virulencia de K279a en el modelo de G.
mellonella. La mayor virulencia de la mutante Fur60 correlaciona con su mayor capacidad
de formar biofilms y con su mayor resistencia al estrés oxidativo.
Los resultados presentados muestran que el hierro, a través del sistema Fur, es una
señal que regula negativamente la síntesis de DSF.
En conclusión, en S. maltophilia la formación de biofilms y la producción de otros
potenciales factores de virulencia están reguladas positivamente por el sistema de
señalización rpfF/DSF, y negativamente por concentraciones no limitantes de hierro, a
través del sistema Fur. Además, en esta tesis se comprobó por primera vez la relación
entre el sistema Fur-hierro y el sistema de QS mediado por DSF en S. maltophilia.

Publicaciones

Publicaciones
122
Publicaciones con referato
Los resultados obtenidos durante el desarrollo de esta Tesis dieron lugar a la siguiente publicación:
García C, Passerini de Rossi B, Alcaraz E, Vay C, Franco M. 2012. “Siderophores of Stenotrophomonas maltophilia : detection and determination of their chemical nature”. Rev Argent Microbiol 44: 150-154. ISSN 0325-7541.
Recientemente hemos recibido una invitación del editor Dr. José Martinez para participar del tópico de investigación "A multidisciplinary look at Stenotrophomonas maltophilia: an emerging multi-drug-resistant global opportunistic pathogen.”,a publicarse en Frontiers in Microbiology, El resumen “Iron is a signal for Stenotrophomonas maltophilia biofilm formation, oxidative stress response, OMPs expression and virulence” ha sido aceptado y próximamente enviaremos el manuscrito para su evaluación.
Parte de los resultados de esta tesis permitieron desarrollar otras líneas de trabajo en las que se investigaron la actividad de antimicrobianos sobre biofilms y el papel de DSF como señal inter especie. Estos estudios dieron lugar a las siguientes publicaciones:
Passerini de Rossi B., García C., Calenda M., Vay C., Franco M. 2009. “Activity of levofloxacin and ciprofloxacin on biofilms and plan ktonic cells of Stenotrophomonas maltophilia isolates from patients with device-associated infections” . Int. J. Antimicrob. Agents 34: 260-264. ISSN 0924-8579.
Passerini de Rossi B., García C., Alcaraz E., Franco M. 2014. “ Stenotrophomonas maltophilia interferes via the DSF-mediated quorum sensing syst em with Candida albicans filamentation and with its planktonic and biofilm modes of growth”. Rev Argent Microbiol. Aceptado para su publicación el 29/10/2014.
Presentaciones en Congresos
� XI Congreso Argentino de Microbiología. Organizador: Asociación Argentina de Microbiología. Córdoba, Argentina, 10-12 de Octubre de 2007. “Tipificacion molecular de aislamientos de Stenotrophomonas maltophilia proveniente de un hospital universitario por ERIC-PCR” . Passerini de Rossi B, Garcia C, Vay C, Franco M. Revista Argentina de Microbiología, Vol. 39, Suplem ento Nº1 Año 2007.

Publicaciones
123
� XIII Jornadas Argentinas de Microbiología. Organizador: Asociación Argentina de Microbiología. Rosario, Argentina, 9-11 de Octubre de 2008. “Efectos pleiotrópicos de una mutación en el gen que codific a la señal de quorum sensing en Stenotrophomonas maltophilia ”. Passerini de Rossi B; Garcia C; Calenda M; Durán L; Feldman L; Franco M.
� XII Congreso Argentino de Microbiología. Organizador: Asociación Argentina de Microbiología. Buenos Aires, Argentina, 17-20 de Octubre de 2010. “ Stenotrophomonas maltophilia interfiere en la formación del tubo germinal y el crecimiento de Candida albicans a través de su señal de quorum sensing” Passerini de Rossi B, Feldman L, Saliba Pineda M, Garcia C, Franco M. Revista Argentina de Microbiología, Vol. 42, Suplemento Nº1 Año 2010.
� XII Congreso Argentino de Microbiología. Organizador: Asociación Argentina de Microbiología. Buenos Aires, Argentina, 17-20 de Octubre de 2010. “Estudio de la producción de sideróforos en aislamientos clínicos de Stenotrophomonas maltophilia ” Garcia C, Passerini de Rossi B, Feldman L, Saliba Pineda M, Vay C, Franco M. Revista Argentina de Microbiología, Vol. 42, Suplem ento Nº1 Año 2010.
� VII Congreso Argentino de Microbiología General. SAMIGE del Bicentenario. San Miguel de Tucumán, Tucumán, Argentina, 18-20 de Mayo de 2011. “Role of iron on biofilm formation of wild type and fur mutants of Stenotrophomonas maltophilia” García C, Passerini de Rossi B, Alcaraz E, Pradelli C, Vay C, Franco M.
� XIV Jornadas Argentinas de Microbiología, III Jornada de Microbiología e Infectología del NEA. Organizador: Asociación Argentina de Microbiología, Filial NEA, Resistencia, Chaco, Argentina 29 de septiembre al 1 de octubre de 2011. “Influencia del hierro en la respuesta al estrés ox idativo de cultivos planctónicos y biofilms de Stenotrophomonas maltophilia ” . C. García, B. Passerini de Rossi, E. Alcaraz, C. Pradelli, M. Franco.
� XIV Jornadas Argentinas de Microbiología, III Jornada de Microbiología e Infectología del NEA. Organizador: Asociación Argentina de Microbiología, Filial NEA, Resistencia, Chaco, Argentina 29 de septiembre al 1 de octubre de 2011. “La señal de quorum sensing de Stenotrophomonas maltophilia interfiere con la transición morfológica de Candida albicans y con la viabilidad de la forma miceliar.” B. Passerini de Rossi, L. Feldman, M. Saliba Pineda, C. García, M. Franco.
� VIII Congreso de Microbiología General. SAMIGE. Mar del Plata, Pcia. de Buenos Aires, Argentina, 4-6 de Julio de 2012. “Influence of oxygen availability and growth phase on the activity of superoxide dismutas e in Stenotrophomonas maltophilia.” C. García, B. Ferraris Rostock, B. Passerini de Rossi, M. Franco
� VIII Congreso de Microbiología General. SAMIGE. Mar del Plata, Pcia. de Buenos Aires, Argentina, 4-6 de Julio de 2012. “Quorum sensing cross-kingdom interactions: Stenotrophomonas maltophilia and Candida albicans biofilms”. B. Passerini de Rossi, C. Pradelli, E. Alcaraz, C. Garcia, M. Franco. Comunicación oral.
� XIII Congreso Argentino de Microbiología. Organizador: Asociación Argentina de Microbiología. Buenos Aires, Argentina, 23-26 de Septiembre de 2013. “Caracterización de proteínas de membrana externa r eguladas por hierro en

Publicaciones
124
Stenotrophomonas maltophilia ” Garcia C, Passerini de Rossi B, Dassoy L, Franco M. Comunicación oral. Revista Argentina de Microbiología, Vol. 45, Suplemento Nº1 Año 2013.
� X Congreso de Microbiología General. SAMIGE. Mar del Plata, Pcia. de Buenos Aires, Argentina, 2-4 de Julio de 2014. “Impact of quorum sensing and iron signals on biofilm architecture and virulence of Stenotrophomonas maltophilia”. C. García, E. Alcaraz, Y. Figueroa, R. Romero, B. Passerini de Rossi, M. Franco. Comunicación oral.

125
Anexo I
Medios de cultivo
Stainer-Scholte suplementado con casamino ácidos 0, 1% (SSC)
� L-glutamato 10 g/l
� L prolina 0,24 g/l
� NaCl 2,5 g/l
� PO4H2K 0,5 g/l
� KCl 0,2 g/l
� MgCl2.6 H2O 0,1 g/l
� Tris 12,6 mM
� CaCl2 0,03 g/l
� Casamino Acidos (Difco) 0,1%
CAS-HDTMA-Fe que se agrega al medio SSC
� 605 mg Cromo azurol S (Sigma Aldrich) disuelto en 500 ml de agua
� 100 ml 1mM FeCl3.6 H2O (Sigma Aldrich) en 10 mM HCl
� 729 mg Hexadeciltrimetilamonio bromuro (HDTMA) (Fluka) en 400 ml de agua
MMXC:
� K2HPO4 10,5g/l
� KH2PO4 4,5g/l
� (NH4)2SO4 1g/l
� Hidrolizado de caseína 1,5g/l
� MgSO4 1mM
� Citrato de sodio 0,5g/l
CMC agar
� CMC 0,125%
� Buffer KH2PO4/K2HPO4, 50 mM pH 6,0
� Agar-agar 1,5%
NYGB: Medio liquido rico para Xanthomonas campestris
� Difco peptona 5g/l
� Extracto de levadura 3g/l
� Glicerol 20g/l
�

126
NYGA
� Difco peptona 5g/l
� Extracto de levadura 3g/l
� Glicerol 20g/l
� Agar-agar 1,5%
LB
• Triptona 10 g/l
• Extracto de levadura 5 g/l
• NaCl 5 g/l
LB Glucosa
� Triptona 10 g/l
� Extracto de levadura 5 g/l
� NaCl 5 g/l
� Glucosa 1 g/l
Caldo nitrato
� Extracto de carne 3 g/l
� Peptona 5 g/l
� Nitrato de potasio 1 g/l
� pH:7,00
Reactivo de Griess:
� Solucion A: Acido sulfanilico 8 g/l
� Solucion B: α naftilamina 5 g en 1 l de ácido acético 5N

Bibliografía

Bibliografía
128
1. Adamek, M., J. Overhage, S. Bathe, J. Winter, R. Fi scher, and T. Schwartz. 2011. Genotyping of environmental and clinical Stenotrophomonas maltophilia isolates and their pathogenic potential. PLoS One 6:e27615.
2. Aiassa, V., A. I. Barnes, and I. Albesa. 2010. Resistance to ciprofloxacin by enhancement of antioxidant defenses in biofilm and planktonic Proteus mirabilis. Biochem Biophys Res Commun 393:84-8.
3. Al-Anazi, K. A., and A. M. Al-Jasser. Infections Caused by Stenotrophomonas maltophilia in Recipients of Hematopoietic Stem Cell Transplantation. Front Oncol 4:232.
4. Alavi, P., H. Muller, M. Cardinale, C. Zachow, M. B . Sanchez, J. L. Martinez, and G. Berg. 2013. The DSF quorum sensing system controls the positive influence of Stenotrophomonas maltophilia on plants. PLoS One 8:e67103.
5. Andrews, S. C., A. K. Robinson, and F. Rodriguez-Qu inones. 2003. Bacterial iron homeostasis. FEMS Microbiol Rev 27:215-37.
6. Anzai, Y., H. Kim, J. Y. Park, H. Wakabayashi, and H. Oyaizu. 2000. Phylogenetic affiliation of the pseudomonads based on 16S rRNA sequence. Int J Syst Evol Microbiol 50 Pt 4:1563-89.
7. Arce Miranda, J. E., C. E. Sotomayor, I. Albesa, an d M. G. Paraje. 2011. Oxidative and nitrosative stress in Staphylococcus aureus biofilm. FEMS Microbiol Lett 315:23-9.
8. Armstrong, S. K., and M. O. Clements. 1993. Isolation and characterization of Bordetella bronchiseptica mutants deficient in siderophore activity. J Bacteriol 175:1144-52.
9. Assih, E. A., A. S. Ouattara, S. Thierry, J. L. Cay ol, M. Labat, and H. Macarie. 2002. Stenotrophomonas acidaminiphila sp. nov., a strictly aerobic bacterium isolated from an upflow anaerobic sludge blanket (UASB) reactor. Int J Syst Evol Microbiol 52:559-68.
10. Bailey, T. L., and C. Elkan. 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc Int Conf Intell Syst Mol Biol 2:28-36.
11. Bailey, T. L., and M. Gribskov. 1998. Combining evidence using p-values: application to sequence homology searches. Bioinformatics 14:48-54.
12. Banin, E., M. L. Vasil, and E. P. Greenberg. 2005. Iron and Pseudomonas aeruginosa biofilm formation. Proc Natl Acad Sci U S A 102:11076-81.
13. Barber, C. E., J. L. Tang, J. X. Feng, M. Q. Pan, T . J. Wilson, H. Slater, J. M. Dow, P. Williams, and M. J. Daniels. 1997. A novel regulatory system required for pathogenicity of Xanthomonas campestris is mediated by a small diffusible signal molecule. Mol Microbiol 24:555-66.
14. Barton, H. A., Z. Johnson, C. D. Cox, A. I. Vasil, and M. L. Vasil. 1996. Ferric uptake regulator mutants of Pseudomonas aeruginosa with distinct alterations in the iron-dependent repression of exotoxin A and siderophores in aerobic and microaerobic environments. Mol Microbiol 21:1001-17.
15. Beauchamp, C., and I. Fridovich. 1971. Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Anal Biochem 44:276-87.
16. Bollinger, N., D. J. Hassett, B. H. Iglewski, J. W. Costerton, and T. R. McDermott. 2001. Gene expression in : evidence of iron override effects on quorum sensing and biofilm-specific gene regulation. J Bacteriol 183:1990-6.

Bibliografía
129
17. Boon, C., Y. Deng, L. H. Wang, Y. He, J. L. Xu, Y. Fan, S. Q. Pan, and L. H. Zhang. 2008. A novel DSF-like signal from Burkholderia cenocepacia interferes with Candida albicans morphological transition. ISME J 2:27-36.
18. Bottone, E. J., M. Reitano, J. M. Janda, K. Troy, a nd J. Cuttner. 1986. Pseudomonas maltophilia exoenzyme activity as correlate in pathogenesis of ecthyma gangrenosum. J Clin Microbiol 24:995-7.
19. Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248-54.
20. Braun, V. 1995. Energy-coupled transport and signal transduction through the gram-negative outer membrane via TonB-ExbB-ExbD-dependent receptor proteins. FEMS Microbiol Rev 16:295-307.
21. Cao, B., L. Shi, R. N. Brown, Y. Xiong, J. K. Fredr ickson, M. F. Romine, M. J. Marshall, M. S. Lipton, and H. Beyenal. 2011. Extracellular polymeric substances from Shewanella sp. HRCR-1 biofilms: characterization by infrared spectroscopy and proteomics. Environ Microbiol 13:1018-31.
22. Carmody, L. A., T. Spilker, and J. J. LiPuma. 2011. Reassessment of Stenotrophomonas maltophilia phenotype. J Clin Microbiol 49:1101-3.
23. Caylan, R., N. Kaklikkaya, K. Aydin, F. Aydin, G. Y ilmaz, B. Ozgumus, and I. Koksal. 2004. An epidemiological analysis of Stenotrophomonas maltophilia strains in a university hospital. Jpn J Infect Dis 57:37-40.
24. Cescutti, P., B. Cuzzi, G. Liut, C. Segonds, G. Di Bonaventura, and R. Rizzo. 2011. A novel highly charged exopolysaccharide produced by two strains of Stenotrophomonas maltophilia recovered from patients with cystic fibrosis. Carbohydr Res 346:1916-23.
25. Clark, V. L., and P. M. Bavoil. 1997. Bacterial pathogenesis, Methods in Enzymology, vol. 235. Academic Press, San Diego.
26. Coenye, T., E. Vanlaere, E. Falsen, and P. Vandamme . 2004. Stenotrophomonas africana Drancourt et al. 1997 is a later synonym of Stenotrophomonas maltophilia (Hugh 1981) Palleroni and Bradbury 1993. Int J Syst Evol Microbiol 54:1235-7.
27. Colnaghi Simionato, A. V., D. S. da Silva, M. R. La mbais, and E. Carrilho. 2007. Characterization of a putative Xylella fastidiosa diffusible signal factor by HRGC-EI-MS. J Mass Spectrom 42:490-6.
28. Costerton, J. W., Z. Lewandowski, D. E. Caldwell, D . R. Korber, and H. M. Lappin-Scott. 1995. Microbial biofilms. Annu Rev Microbiol 49:711-45.
29. Costerton, J. W., P. S. Stewart, and E. P. Greenber g. 1999. Bacterial biofilms: a common cause of persistent infections. Science 284:1318-22.
30. Crosa, J. H. 1997. Signal transduction and transcriptional and posttranscriptional control of iron-regulated genes in bacteria. Microbiol Mol Biol Rev 61:319-36.
31. Crossman, L., and J. M. Dow. 2004. Biofilm formation and dispersal in Xanthomonas campestris. Microbes Infect 6:623-9.
32. Crossman, L. C., V. C. Gould, J. M. Dow, G. S. Vern ikos, A. Okazaki, M. Sebaihia, D. Saunders, C. Arrowsmith, T. Carver, N. Peters, E. Adlem, A. Kerhornou, A. Lord, L. Murphy, K. Seeger, R. Square s, S. Rutter, M. A. Quail, M. A. Rajandream, D. Harris, C. Churcher, S. D. Ben tley, J. Parkhill, N. R. Thomson, and M. B. Avison. 2008. The complete genome, comparative and functional analysis of Stenotrophomonas maltophilia reveals an organism heavily shielded by drug resistance determinants. Genome Biol 9:R74.

Bibliografía
130
33. Cha, J. Y., J. S. Lee, J. I. Oh, J. W. Choi, and H. S. Baik. 2008. Functional analysis of the role of Fur in the virulence of Pseudomonas syringae pv. tabaci 11528: Fur controls expression of genes involved in quorum-sensing. Biochem Biophys Res Commun 366:281-7.
34. Chatelut, M., J. L. Dournes, G. Chabanon, and N. Ma rty. 1995. Epidemiological typing of Stenotrophomonas (Xanthomonas) maltophilia by PCR. J Clin Microbiol 33:912-4.
35. Chatterjee, S., and R. V. Sonti. 2002. rpfF mutants of Xanthomonas oryzae pv. oryzae are deficient for virulence and growth under low iron conditions. Mol Plant Microbe Interact 15:463-71.
36. Chhibber, S., A. Gupta, R. Sharan, V. Gautam, and P . Ray. 2008. Putative virulence characteristics of Stenotrophomonas maltophilia: a study on clinical isolates. World J Microbiol Biotechnol Volume 24: 2819-2825.
37. da Silva, A. C., J. A. Ferro, F. C. Reinach, C. S. Farah, L. R. Furlan, R. B. Quaggio, C. B. Monteiro-Vitorello, M. A. Van Sluys, N. F. Almeida, L. M. Alves, A. M. do Amaral, M. C. Bertolini, L. E. Camargo, G. Camarotte, F. Cannavan, J. Cardozo, F. Chambergo, L. P. Ciapina, R. M. Cicarel li, L. L. Coutinho, J. R. Cursino-Santos, H. El-Dorry, J. B. Faria, A. J. Fer reira, R. C. Ferreira, M. I. Ferro, E. F. Formighieri, M. C. Franco, C. C. Gregg io, A. Gruber, A. M. Katsuyama, L. T. Kishi, R. P. Leite, E. G. Lemos, M . V. Lemos, E. C. Locali, M. A. Machado, A. M. Madeira, N. M. Martinez-Rossi, E. C. Martins, J. Meidanis, C. F. Menck, C. Y. Miyaki, D. H. Moon, L. M. Moreir a, M. T. Novo, V. K. Okura, M. C. Oliveira, V. R. Oliveira, H. A. Pereira, A. R ossi, J. A. Sena, C. Silva, R. F. de Souza, L. A. Spinola, M. A. Takita, R. E. Tamura , E. C. Teixeira, R. I. Tezza, M. Trindade dos Santos, D. Truffi, S. M. Tsai, F. F . White, J. C. Setubal, and J. P. Kitajima. 2002. Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature 417:459-63.
38. Davey, M. E., and A. O'Toole G. 2000. Microbial biofilms: from ecology to molecular genetics. Microbiol Mol Biol Rev 64:847-67.
39. De Beer, D., R. Srinivasan, and P. S. Stewart. 1994. Direct measurement of chlorine penetration into biofilms during disinfection. Appl Environ Microbiol 60:4339-44.
40. de Oliveira-Garcia, D., M. Dall'Agnol, M. Rosales, A. C. Azzuz, N. Alcantara, M. B. Martinez, and J. A. Giron. 2003. Fimbriae and adherence of Stenotrophomonas maltophilia to epithelial cells and to abiotic surfaces. Cell Microbiol 5:625-36.
41. Denton, M., and K. G. Kerr. 1998. Microbiological and clinical aspects of infection associated with Stenotrophomonas maltophilia. Clin Microbiol Rev 11:57-80.
42. Denton, M., N. J. Todd, K. G. Kerr, P. M. Hawkey, a nd J. M. Littlewood. 1998. Molecular epidemiology of Stenotrophomonas maltophilia isolated from clinical specimens from patients with cystic fibrosis and associated environmental samples. J Clin Microbiol 36:1953-8.
43. Deziel, E., Y. Comeau, and R. Villemur. 2001. Initiation of biofilm formation by Pseudomonas aeruginosa 57RP correlates with emergence of hyperpiliated and highly adherent phenotypic variants deficient in swimming, swarming, and twitching motilities. J Bacteriol 183:1195-204.
44. Di Bonaventura, G., G. Prosseda, F. Del Chierico, S . Cannavacciuolo, P. Cipriani, A. Petrucca, F. Superti, M. G. Ammendolia , C. Concato, E. Fiscarelli, M. Casalino, R. Piccolomini, M. Nicoletti, and B. C olonna. 2007. Molecular characterization of virulence determinants of Stenotrophomonas maltophilia strains

Bibliografía
131
isolated from patients affected by cystic fibrosis. Int J Immunopathol Pharmacol 20:529-37.
45. Donlan, R. M. 2001. Biofilms and device-associated infections. Emerg Infect Dis 7:277-81.
46. Donlan, R. M. 2002. Biofilms: microbial life on surfaces. Emerg Infect Dis 8:881-90.
47. Dow, J. M., L. Crossman, K. Findlay, Y. Q. He, J. X . Feng, and J. L. Tang. 2003. Biofilm dispersal in Xanthomonas campestris is controlled by cell-cell signaling and is required for full virulence to plants. Proc Natl Acad Sci U S A 100:10995-1000.
48. Dow, J. M., and M. J. Daniels. 1994. Pathogenicity determinants and global regulation of pathogenicity of Xanthomonas campestris pv. campestris. Curr Top Microbiol Immunol 192:29-41.
49. Drancourt, M., C. Bollet, and D. Raoult. 1997. Stenotrophomonas africana sp. nov., an opportunistic human pathogen in Africa. Int J Syst Bacteriol 47:160-3.
50. DuBois, M., K. A. Gilles, J. K. Hamilton, P. A. Reb ers, and F. Smith. 1956. Colorimetric Method for Determination of Sugars and Related Substances. Analytical Chemistry 28:350-356.
51. Dunlap, P. V., and H. M. Steinman. 1986. Strain variation in bacteriocuprein superoxide dismutase from symbiotic Photobacterium leiognathi. J Bacteriol 165:393-8.
52. Dunne, C., J. J. Crowley, Y. Moenne-Loccoz, D. N. D owling, F. J. de Bruijn, and F. O'Gara. 1997. Biological control of Pythium ultimum by Stenotrophomonas maltophilia W81 is mediated by an extracellular proteolytic activity. Microbiology 143:3921-3931.
53. Dunne, W. M., Jr., E. O. Mason, Jr., and S. L. Kapl an. 1993. Diffusion of rifampin and vancomycin through a Staphylococcus epidermidis biofilm. Antimicrob Agents Chemother 37:2522-6.
54. Elkins, J. G., D. J. Hassett, P. S. Stewart, H. P. Schweizer, and T. R. McDermott. 1999. Protective role of catalase in Pseudomonas aeruginosa biofilm resistance to hydrogen peroxide. Appl Environ Microbiol 65:4594-600.
55. Escolar, L., J. Perez-Martin, and V. de Lorenzo. 1999. Opening the iron box: transcriptional metalloregulation by the Fur protein. J Bacteriol 181:6223-9.
56. Fee, J. A. 1991. Regulation of sod genes in Escherichia coli: relevance to superoxide dismutase function. Mol Microbiol 5:2599-610.
57. Ferguson, A. D., and J. Deisenhofer. 2002. TonB-dependent receptors-structural perspectives. Biochim Biophys Acta 1565:318-32.
58. Fernandez-Cuenca, F. 2004. [Applications of PCR techniques for molecular epidemiology of infectious diseases]. Enferm Infecc Microbiol Clin 22:355-60.
59. Ferrer-Navarro, M., R. Planell, D. Yero, E. Mongiar dini, G. Torrent, P. Huedo, P. Martinez, N. Roher, S. Mackenzie, I. Gibert, and X. Daura. 2013. Abundance of the Quorum-Sensing Factor Ax21 in Four Strains of Correlates with Mortality Rate in a New Zebrafish Model of Infection. PLoS One 8:e67207.
60. Fiedler, H. P., P. Krastel, J. Muller, K. Gebhardt, and A. Zeeck. 2001. Enterobactin: the characteristic catecholate siderophore of Enterobacteriaceae is produced by Streptomyces species.(1). FEMS Microbiol Lett 196:147-51.
61. Finkmann, W., K. Altendorf, E. Stackebrandt, and A. Lipski. 2000. Characterization of N2O-producing Xanthomonas-like isolates from biofilters as Stenotrophomonas nitritireducens sp. nov., Luteimonas mephitis gen. nov., sp. nov. and Pseudoxanthomonas broegbernensis gen. nov., sp. nov. Int J Syst Evol Microbiol 50 Pt 1:273-82.

Bibliografía
132
62. Fouhy, Y., K. Scanlon, K. Schouest, C. Spillane, L. Crossman, M. B. Avison, R. P. Ryan, and J. M. Dow. 2007. Diffusible signal factor-dependent cell-cell signaling and virulence in the nosocomial pathogen Stenotrophomonas maltophilia. J Bacteriol 189:4964-8.
63. Fuqua, C., S. C. Winans, and E. P. Greenberg. 1996. Census and consensus in bacterial ecosystems: the LuxR-LuxI family of quorum-sensing transcriptional regulators. Annu Rev Microbiol 50:727-51.
64. Furrer, J. L., D. N. Sanders, I. G. Hook-Barnard, a nd M. A. McIntosh. 2002. Export of the siderophore enterobactin in Escherichia coli: involvement of a 43 kDa membrane exporter. Mol Microbiol 44:1225-34.
65. Garcia de Viedma, D., M. Marin, E. Cercenado, R. Al onso, M. Rodriguez-Creixems, and E. Bouza. 1999. Evidence of nosocomial Stenotrophomonas maltophilia cross-infection in a neonatology unit analyzed by three molecular typing methods. Infect Control Hosp Epidemiol 20:816-20.
66. Garnett, J. A., and S. Matthews. 2012. Interactions in bacterial biofilm development: a structural perspective. Curr Protein Pept Sci 13:739-55.
67. Guerinot, M. L. 1994. Microbial iron transport. Annu Rev Microbiol 48:743-72. 68. Gulmez, D., and G. Hascelik. 2005. Stenotrophomonas maltophilia: antimicrobial
resistance and molecular typing of an emerging pathogen in a Turkish university hospital. Clin Microbiol Infect 11:880-6.
69. Hantke, K. 1987. Selection procedure for deregulated iron transport mutants (fur) in Escherichia coli K 12: fur not only affects iron metabolism. Mol Gen Genet 210:135-9.
70. Hassan, H. M., and H. C. Sun. 1992. Regulatory roles of Fnr, Fur, and Arc in expression of manganese-containing superoxide dismutase in Escherichia coli. Proc Natl Acad Sci U S A 89:3217-21.
71. Hassett, D. J., M. L. Howell, U. A. Ochsner, M. L. Vasil, Z. Johnson, and G. E. Dean. 1997. An operon containing fumC and sodA encoding fumarase C and manganese superoxide dismutase is controlled by the ferric uptake regulator in Pseudomonas aeruginosa: fur mutants produce elevated alginate levels. J Bacteriol 179:1452-9.
72. Hassett, D. J., J. F. Ma, J. G. Elkins, T. R. McDer mott, U. A. Ochsner, S. E. West, C. T. Huang, J. Fredericks, S. Burnett, P. S. Stewart, G. McFeters, L. Passador, and B. H. Iglewski. 1999. Quorum sensing in Pseudomonas aeruginosa controls expression of catalase and superoxide dismutase genes and mediates biofilm susceptibility to hydrogen peroxide. Mol Microbiol 34:1082-93.
73. Hassett, D. J., H. P. Schweizer, and D. E. Ohman. 1995. Pseudomonas aeruginosa sodA and sodB mutants defective in manganese- and iron-cofactored superoxide dismutase activity demonstrate the importance of the iron-cofactored form in aerobic metabolism. J Bacteriol 177:6330-7.
74. Hassett, D. J., P. A. Sokol, M. L. Howell, J. F. Ma , H. T. Schweizer, U. Ochsner, and M. L. Vasil. 1996. Ferric uptake regulator (Fur) mutants of Pseudomonas aeruginosa demonstrate defective siderophore-mediated iron uptake, altered aerobic growth, and decreased superoxide dismutase and catalase activities. J Bacteriol 178:3996-4003.
75. Hassett, D. J., W. A. Woodruff, D. J. Wozniak, M. L . Vasil, M. S. Cohen, and D. E. Ohman. 1993. Cloning and characterization of the Pseudomonas aeruginosa sodA and sodB genes encoding manganese- and iron-cofactored superoxide dismutase: demonstration of increased manganese superoxide dismutase activity in alginate-producing bacteria. J Bacteriol 175:7658-65.

Bibliografía
133
76. He, Y. W., J. Wu, J. S. Cha, and L. H. Zhang. 2010. Rice bacterial blight pathogen Xanthomonas oryzae pv. oryzae produces multiple DSF-family signals in regulation of virulence factor production. BMC Microbiol 10:187.
77. Heylen, K., B. Vanparys, F. Peirsegaele, L. Lebbe, and P. De Vos. 2007. Stenotrophomonas terrae sp. nov. and Stenotrophomonas humi sp. nov., two nitrate-reducing bacteria isolated from soil. Int J Syst Evol Microbiol 57:2056-61.
78. Hider, R. C., and X. Kong. 2010. Chemistry and biology of siderophores. Nat Prod Rep 27:637-57.
79. Hindre, T., H. Bruggemann, C. Buchrieser, and Y. He chard. 2008. Transcriptional profiling of Legionella pneumophila biofilm cells and the influence of iron on biofilm formation. Microbiology 154:30-41.
80. Hoffmann, J. A. 1995. Innate immunity of insects. Curr Opin Immunol 7:4-10. 81. Huang, T. P., E. B. Somers, and A. C. Wong. 2006. Differential biofilm formation
and motility associated with lipopolysaccharide/exopolysaccharide-coupled biosynthetic genes in Stenotrophomonas maltophilia. J Bacteriol 188:3116-20.
82. Huang, T. P., and A. C. Wong. 2007. A cyclic AMP receptor protein-regulated cell-cell communication system mediates expression of a FecA homologue in Stenotrophomonas maltophilia. Appl Environ Microbiol 73:5034-40.
83. Huber, B., K. Riedel, M. Hentzer, A. Heydorn, A. Go tschlich, M. Givskov, S. Molin, and L. Eberl. 2001. The cep quorum-sensing system of Burkholderia cepacia H111 controls biofilm formation and swarming motility. Microbiology 147:2517-28.
84. Hugh, R., and E. Ryschenkow. 1961. Pseudomonas maltophilia, an alcaligenes-like species. J Gen Microbiol 26:123-32.
85. Hulton, C. S., C. F. Higgins, and P. M. Sharp. 1991. ERIC sequences: a novel family of repetitive elements in the genomes of Escherichia coli, Salmonella typhimurium and other enterobacteria. Mol Microbiol 5:825-34.
86. Jittawuttipoka, T., R. Sallabhan, P. Vattanaviboon, M. Fuangthong, and S. Mongkolsuk. 2010. Mutations of ferric uptake regulator (fur) impair iron homeostasis, growth, oxidative stress survival, and virulence of Xanthomonas campestris pv. campestris. Arch Microbiol 192:331-9.
87. Johnson, M., A. Cockayne, P. H. Williams, and J. A. Morrissey. 2005. Iron-responsive regulation of biofilm formation in Staphylococcus aureus involves fur-dependent and fur-independent mechanisms. J Bacteriol 187:8211-5.
88. Jucker, B. A., H. Harms, and A. J. Zehnder. 1996. Adhesion of the positively charged bacterium Stenotrophomonas (Xanthomonas) maltophilia 70401 to glass and Teflon. J Bacteriol 178:5472-9.
89. Kaiser, S., K. Biehler, and D. Jonas. 2009. A Stenotrophomonas maltophilia multilocus sequence typing scheme for inferring population structure. J Bacteriol 191:2934-43.
90. Kaparullina, E., N. Doronina, T. Chistyakova, and Y . Trotsenko. 2009. Stenotrophomonas chelatiphaga sp. nov., a new aerobic EDTA-degrading bacterium. Syst Appl Microbiol 32:157-62.
91. Kim, H. B., S. Srinivasan, G. Sathiyaraj, L. H. Qua n, S. H. Kim, T. P. Bui, Z. Q. Liang, Y. J. Kim, and D. C. Yang. 2010. Stenotrophomonas ginsengisoli sp. nov., isolated from a ginseng field. Int J Syst Evol Microbiol 60:1522-6.
92. Kim, I. H., Y. Wen, J. S. Son, K. H. Lee, and K. S. Kim. 2013. The fur-iron complex modulates expression of the quorum-sensing master regulator, SmcR, to control expression of virulence factors in Vibrio vulnificus. Infect Immun 81:2888-98.

Bibliografía
134
93. Krepsky, N., R. B. Rocha Ferreira, A. P. Ferreira N unes, U. G. Casado Lins, F. Costa e Silva Filho, A. L. de Mattos-Guaraldi, and K. R. Netto-dosSantos. 2003. Cell surface hydrophobicity and slime production of Staphylococcus epidermidis Brazilian isolates. Curr Microbiol 46:280-6.
94. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-5.
95. Lee, M., S. G. Woo, M. Chae, M. C. Shin, H. M. Jung , and L. N. Ten. 2011. Stenotrophomonas daejeonensis sp. nov., isolated from sewage. Int J Syst Evol Microbiol 61:598-604.
96. Leyh, R., and R. W. Griffith. 1992. Characterization of the outer membrane proteins of Bordetella avium. Infect Immun 60:958-64.
97. Looney, W. J. 2005. Role of Stenotrophomonas maltophilia in hospital-acquired infection. Br J Biomed Sci 62:145-54; quiz 1 p following 154.
98. Lu, Z., L. Sang, Z. Li, and H. Min. 2009. Catalase and superoxide dismutase activities in a Stenotrophomonas maltophilia WZ2 resistant to herbicide pollution. Ecotoxicol Environ Saf 72:136-43.
99. Mah, T. F., and G. A. O'Toole. 2001. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol 9:34-9.
100. McCarthy, Y., J. M. Dow, and R. P. Ryan. 2011. The Ax21 protein is a cell-cell signal that regulates virulence in the nosocomial pathogen Stenotrophomonas maltophilia. J Bacteriol 193:6375-8.
101. McKay, G. A., D. E. Woods, K. L. MacDonald, and K. Poole. 2003. Role of phosphoglucomutase of Stenotrophomonas maltophilia in lipopolysaccharide biosynthesis, virulence, and antibiotic resistance. Infect Immun 71:3068-75.
102. Mey, A. R., S. A. Craig, and S. M. Payne. 2005. Characterization of Vibrio cholerae RyhB: the RyhB regulon and role of ryhB in biofilm formation. Infect Immun 73:5706-19.
103. Miethke, M., and M. A. Marahiel. 2007. Siderophore-based iron acquisition and pathogen control. Microbiol Mol Biol Rev 71:413-51.
104. Miller, M. B., and B. L. Bassler. 2001. Quorum sensing in bacteria. Annu Rev Microbiol 55:165-99.
105. Minkwitz, A., and G. Berg. 2001. Comparison of antifungal activities and 16S ribosomal DNA sequences of clinical and environmental isolates of Stenotrophomonas maltophilia. J Clin Microbiol 39:139-45.
106. Monroe, D. 2007. Looking for chinks in the armor of bacterial biofilms. PLoS Biol 5:e307.
107. Musk, D. J., D. A. Banko, and P. J. Hergenrother. 2005. Iron salts perturb biofilm formation and disrupt existing biofilms of Pseudomonas aeruginosa. Chem Biol 12:789-96.
108. Musk, D. J., Jr., and P. J. Hergenrother. 2008. Chelated iron sources are inhibitors of Pseudomonas aeruginosa biofilms and distribute efficiently in an in vitro model of drug delivery to the human lung. J Appl Microbiol 105:380-8.
109. Neilands, J. B. 1995. Siderophores: structure and function of microbial iron transport compounds. J Biol Chem 270:26723-6.
110. Nicoletti, M., A. Iacobino, G. Prosseda, E. Fiscare lli, R. Zarrilli, E. De Carolis, A. Petrucca, L. Nencioni, B. Colonna, and M. Casali no. 2011. Stenotrophomonas maltophilia strains from cystic fibrosis patients: genomic variability and molecular characterization of some virulence determinants. Int J Med Microbiol 301:34-43.

Bibliografía
135
111. O'Toole, G. A., and R. Kolter. 1998. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol Microbiol 30:295-304.
112. O'Toole, G. A., L. A. Pratt, P. I. Watnick, D. K. N ewman, V. B. Weaver, and R. Kolter. 1999. Genetic approaches to study of biofilms. Methods Enzymol 310:91-109.
113. Oglesby, A. G., J. M. Farrow, 3rd, J. H. Lee, A. P. Tomaras, E. P. Greenberg, E. C. Pesci, and M. L. Vasil. 2008. The influence of iron on Pseudomonas aeruginosa physiology: a regulatory link between iron and quorum sensing. J Biol Chem 283:15558-67.
114. Olive, D. M., and P. Bean. 1999. Principles and applications of methods for DNA-based typing of microbial organisms. J Clin Microbiol 37:1661-9.
115. Palleroni, N. J., and J. F. Bradbury. 1993. Stenotrophomonas, a new bacterial genus for Xanthomonas maltophilia (Hugh 1980) Swings et al. 1983. Int J Syst Bacteriol 43:606-9.
116. Parsek, M. R., and E. P. Greenberg. 2000. Acyl-homoserine lactone quorum sensing in gram-negative bacteria: a signaling mechanism involved in associations with higher organisms. Proc Natl Acad Sci U S A 97:8789-93.
117. Passerini de Rossi, B., M. Calenda, C. Vay, and M. Franco. 2007. Biofilm formation by Stenotrophomonas maltophilia isolates from device-associated nosocomial infections. Rev Argent Microbiol 39:204-12.
118. Passerini de Rossi, B., L. Feldman, M. S. Pineda, C . Vay, and M. Franco. 2012. Comparative in vitro efficacies of ethanol-, EDTA- and levofloxacin-based catheter lock solutions on eradication of Stenotrophomonas maltophilia biofilms. J Med Microbiol 61:1248-53.
119. Passerini de Rossi, B., C. Garcia, M. Calenda, C. V ay, and M. Franco. 2009. Activity of levofloxacin and ciprofloxacin on biofilms and planktonic cells of Stenotrophomonas maltophilia isolates from patients with device-associated infections. Int J Antimicrob Agents 34:260-4.
120. Pompilio, A., R. Piccolomini, C. Picciani, D. D'Ant onio, V. Savini, and G. Di Bonaventura. 2008. Factors associated with adherence to and biofilm formation on polystyrene by Stenotrophomonas maltophilia: the role of cell surface hydrophobicity and motility. FEMS Microbiol Lett 287:41-7.
121. Pompilio, A., S. Pomponio, V. Crocetta, G. Gherardi , F. Verginelli, E. Fiscarelli, G. Dicuonzo, V. Savini, D. D'Antonio, a nd G. Di Bonaventura. 2011. Phenotypic and genotypic characterization of Stenotrophomonas maltophilia isolates from patients with cystic fibrosis: genome diversity, biofilm formation, and virulence. BMC Microbiol 11:159.
122. Ramos, P. L., S. Van Trappen, F. L. Thompson, R. C. Rocha, H. R. Barbosa, P. De Vos, and C. A. Moreira-Filho. 2011. Screening for endophytic nitrogen-fixing bacteria in Brazilian sugar cane varieties used in organic farming and description of Stenotrophomonas pavanii sp. nov. Int J Syst Evol Microbiol 61:926-31.
123. Rani, S. A., B. Pitts, H. Beyenal, R. A. Veluchamy, Z. Lewandowski, W. M. Davison, K. Buckingham-Meyer, and P. S. Stewart. 2007. Spatial patterns of DNA replication, protein synthesis, and oxygen concentration within bacterial biofilms reveal diverse physiological states. J Bacteriol 189:4223-33.
124. Reisner, A., J. A. Haagensen, M. A. Schembri, E. L. Zechner, and S. Molin. 2003. Development and maturation of Escherichia coli K-12 biofilms. Mol Microbiol 48:933-46.
125. Rosenberg, M. 1981. Bacterial adherence to polystyrene: a replica method of screening for bacterial hydrophobicity. Appl Environ Microbiol 42:375-7.

Bibliografía
136
126. Ryan, R. P., S. Monchy, M. Cardinale, S. Taghavi, L . Crossman, M. B. Avison, G. Berg, D. van der Lelie, and J. M. Dow. 2009. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat Rev Microbiol 7:514-25.
127. Schagger, H., and G. von Jagow. 1987. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal Biochem 166:368-79.
128. Schwyn, B., and J. B. Neilands. 1987. Universal chemical assay for the detection and determination of siderophores. Anal Biochem 160:47-56.
129. Seed, K. D., and J. J. Dennis. 2008. Development of Galleria mellonella as an alternative infection model for the Burkholderia cepacia complex. Infect Immun 76:1267-75.
130. Semmler, A. B., C. B. Whitchurch, and J. S. Mattick . 1999. A re-examination of twitching motility in Pseudomonas aeruginosa. Microbiology 145 ( Pt 10):2863-73.
131. Senol, E. 2004. Stenotrophomonas maltophilia: the significance and role as a nosocomial pathogen. J Hosp Infect 57:1-7.
132. Seral, C., B. Rojo-Bezares, A. Garrido, M. J. Gude, Y. Saenz, and F. J. Castillo. 2012. [Characterisation of a CTX-M-15-producing Shigella sonnei in a Spanish patient who had not travelled abroad]. Enferm Infecc Microbiol Clin 30:469-71.
133. Silbert, S., M. A. Pfaller, R. J. Hollis, A. L. Bar th, and H. S. Sader. 2004. Evaluation of three molecular typing techniques for nonfermentative Gram-negative bacilli. Infect Control Hosp Epidemiol 25:847-51.
134. Singh, P. K., M. R. Parsek, E. P. Greenberg, and M. J. Welsh. 2002. A component of innate immunity prevents bacterial biofilm development. Nature 417:552-5.
135. Slater, H., A. Alvarez-Morales, C. E. Barber, M. J. Daniels, and J. M. Dow. 2000. A two-component system involving an HD-GYP domain protein links cell-cell signalling to pathogenicity gene expression in Xanthomonas campestris. Mol Microbiol 38:986-1003.
136. Srivastava, D., and C. M. Waters. 2012. A tangled web: regulatory connections between quorum sensing and cyclic Di-GMP. J Bacteriol 194:4485-93.
137. Stepanovic, S., I. Cirkovic, L. Ranin, and M. Svabi c-Vlahovic. 2004. Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett Appl Microbiol 38:428-32.
138. Stoodley, P., D. Debeer, and Z. Lewandowski. 1994. Liquid flow in biofilm systems. Appl Environ Microbiol 60:2711-6.
139. Suci, P. A., M. W. Mittelman, F. P. Yu, and G. G. G eesey. 1994. Investigation of ciprofloxacin penetration into Pseudomonas aeruginosa biofilms. Antimicrob Agents Chemother 38:2125-33.
140. Tomaras, A. P., C. W. Dorsey, R. E. Edelmann, and L . A. Actis. 2003. Attachment to and biofilm formation on abiotic surfaces by Acinetobacter baumannii: involvement of a novel chaperone-usher pili assembly system. Microbiology 149:3473-84.
141. Torres, P. S., F. Malamud, L. A. Rigano, D. M. Russ o, M. R. Marano, A. P. Castagnaro, A. Zorreguieta, K. Bouarab, J. M. Dow, and A. A. Vojnov. 2007. Controlled synthesis of the DSF cell-cell signal is required for biofilm formation and virulence in Xanthomonas campestris. Environ Microbiol 9:2101-9.
142. Touati, D. 2000. Iron and oxidative stress in bacteria. Arch Biochem Biophys 373:1-6.
143. Travassos, L. H., M. N. Pinheiro, F. S. Coelho, J. L. Sampaio, V. L. Merquior, and E. A. Marques. 2004. Phenotypic properties, drug susceptibility and genetic

Bibliografía
137
relatedness of Stenotrophomonas maltophilia clinical strains from seven hospitals in Rio de Janeiro, Brazil. J Appl Microbiol 96:1143-50.
144. Valdezate, S., A. Vindel, P. Martin-Davila, B. S. D el Saz, F. Baquero, and R. Canton. 2004. High genetic diversity among Stenotrophomonas maltophilia strains despite their originating at a single hospital. J Clin Microbiol 42:693-9.
145. VanCouwenbergh, C., and S. Cohen. 1994. Analysis of epidemic and endemic isolates of Xanthomonas maltophilia by contour-clamped homogeneous electric field gel electrophoresis. Infect Control Hosp Epidemiol 15:691-6.
146. Vasil, M. L., and U. A. Ochsner. 1999. The response of Pseudomonas aeruginosa to iron: genetics, biochemistry and virulence. Mol Microbiol 34:399-413.
147. Vazquez, S. C., L. N. Rios Merino, W. P. MacCormack , and E. R. Fraile. 1995. Protease-producing psychrotrophic bacteria isolated from Antarctica. Polar Biology 15:131-135.
148. Wang, L. H., Y. He, Y. Gao, J. E. Wu, Y. H. Dong, C . He, S. X. Wang, L. X. Weng, J. L. Xu, L. Tay, R. X. Fang, and L. H. Zhang . 2004. A bacterial cell-cell communication signal with cross-kingdom structural analogues. Mol Microbiol 51:903-12.
149. Weckman, B. G., and B. W. Catlin. 1957. Deoxyribonuclease activity of micrococci from clinical sources. J Bacteriol 73:747-53.
150. Windhorst, S., E. Frank, D. N. Georgieva, N. Genov, F. Buck, P. Borowski, and W. Weber. 2002. The major extracellular protease of the nosocomial pathogen Stenotrophomonas maltophilia: characterization of the protein and molecular cloning of the gene. J Biol Chem 277:11042-9.
151. Winkelmann, G., A. Cansier, W. Beck, and G. Jung. 1994. HPLC separation of enterobactin and linear 2,3-dihydroxybenzoylserine derivatives: a study on mutants of Escherichia coli defective in regulation (fur), esterase (fes) and transport (fepA). Biometals 7:149-54.
152. Winsor, G. L., T. Van Rossum, R. Lo, B. Khaira, M. D. Whiteside, R. E. Hancock, and F. S. Brinkman. 2009. Pseudomonas Genome Database: facilitating user-friendly, comprehensive comparisons of microbial genomes. Nucleic Acids Res 37:D483-8.
153. Wolf, A., A. Fritze, M. Hagemann, and G. Berg. 2002. Stenotrophomonas rhizophila sp. nov., a novel plant-associated bacterium with antifungal properties. Int J Syst Evol Microbiol 52:1937-44.
154. Yang, H. C., W. T. Im, M. S. Kang, D. Y. Shin, and S. T. Lee. 2006. Stenotrophomonas koreensis sp. nov., isolated from compost in South Korea. Int J Syst Evol Microbiol 56:81-4.
155. Zaini, P. A., A. C. Fogaca, F. G. Lupo, H. I. Nakay a, R. Z. Vencio, and A. M. da Silva. 2008. The iron stimulon of Xylella fastidiosa includes genes for type IV pilus and colicin V-like bacteriocins. J Bacteriol 190:2368-78.





























