Embed Size (px)
Citation preview

0
Universidad Autónoma del Estado de Morelos
Facultad de Ciencias Biológicas
“Efectos de la perturbación del hábitat y la estacionalidad
climática en la dieta del ratón espinoso mexicano Liomys
irroratus (Gray, 1868) en una selva seca del sur del estado de
Morelos”
TESIS
QUE PARA OBTENER EL TÍTULO DE
BIÓLOGO
PRESENTA:
EDGARD DAVID MASON ROMO
DIRECTOR DE TESIS:
DR. DAVID VALENZUELA GALVÁN
CUERNAVACA, MORELOS. DICIEMBRE 2005

I
Índice de contenido Página
I. Dedicatoria………………………………………………………………………………...2 II. Agradecimientos………………………………………………………………………...3 1. Resumen………………………………………………………………………………….5 2. Introducción……………………………………………………………………………...6 3. Antecedentes…………………………………………………………………………...10
3.1. Comprobación empírica del factor de estabilidad ambiental………………...10 3.2. Acercamientos metodológicos al problema de determinar la dieta…………11 3.3. La anatomía vegetal como un apoyo para determinar la
dieta de los herbívoros………………………………………………………………...17
3.4. Diferentes enfoques en los estudios de dieta………………………………….21 4. Justificación……………………………………………………………………………23 5. Objetivos………………………………………………………………………………..28
5.1. Objetivo general…………………………………………………………………..28
5.2. Objetivos particulares…………………………………………………………….28
6. Hipótesis………………………………………………………………………………..29 7. Especie de estudio……………………………………………………………………31
7.1. Taxonomía………………………………………………………………………....31 7.2. Características generales………………………………………………………..33
7.3. Biología…………………………………………………………………………….35
7.4. Ecología……………………………………………………………………………37
7.5. Dieta………………………………………………………………………………..38
8. Sitio de Estudio…………………………………………………………………….….43 9. Metodología…………………………………………………………………………….46
9.1. Delimitación de periodos estacionales………………………………………….46 9.2. Elección de los sitios y determinación del grado de perturbación…………...46
9.3. Metodología de trampeo………………………………………………………….49
9.4. Estimación de la diversidad y disponibilidad de recursos alimentarios……..50
9.5. Obtención y preparación del catálogo de referencia de los caracteres anatómico

II
foliares de las herbáceas disponibles para Liomys irroratus……………………...52
9.6. Análisis del contenido de abazones…………………………………………….53
9.7. Análisis de las excretas…………………………………………………………..54
9.8. Evaluación de la selectividad o preferencia relativas……………………….. 56
9.9. Análisis estadísticos…………………………………………………………….. 59
10. Resultados………………………………………………………………………..…...61 10.1. Resultados del trampeo de ratones……………………………………...……61
10.2. Dinámica de la comunidad de herbáceas………………………………...…..61
10.3. Dinámica del banco de semillas…………………………...………………......66
10.4. Resultados de las muestras del contenido de abazones………………..….70
10.5. Resultados del análisis de las excretas…………………………………..…...80
11. Discusión…………………………………………………………………….………. 90 12. Conclusiones…………………………………………………………………….....101 13. Referencias………………………………………………………………………….103
Tablas y Anexos
Tablas……………………………………………………………………………………..122
Tabla 1. Nomenclatura, condición y coordenadas UTM de la ubicación de los
cuadrantes. (Se tomó el punto de una esquina del cuadrante)…………………122
Tabla 2. Especies de plantas recolectadas durante los muestreos de vegetación
……………………………………………………………………………………….....122
Tabla 3. Especies recolectadas en el banco de semillas………………………..123
Tabla 4. Especies de semillas recolectadas de los abazones…………………..124
Tabla 5. Resultados del análisis de preferencia para las semillas de diferentes
especies encontradas en los contenidos de abazones, por condición por temporada
y total. El número indica el lugar relativo de preferencia en el que se encuentra la
especie por cada tratamiento. P=perturbado Np=no perturbado Ll=lluvias S1=secas
tempranas S2= secas tardías…………………………………………………….....124
Tabla 6. Especies de plantas herbáceas identificadas dentro de las excretas, incluye
cuales tienen datos de diversidad y disponibilidad en el

III
medio………………............................................................................................125
Tabla 7. Resultados del análisis de preferencia para las especies encontradas en
excretas, por condición por temporada y total. El número indica el lugar relativo de
preferencia en el que se encuentra la especie por cada tratamiento. P=perturbado,
Np=no perturbado, Ll=lluvias, S1=Secas iniciales, S2=secas finales, SD= Sin datos
de disponibilidad……………………………………………………………………...125
Tabla 8. Géneros y especies cuyas semillas se reportan como presentes en
abazones o madrigueras de Liomys pictus en las selvas secas de Chamela, Jalisco y
que estan presentes en la Sierra de Huautla (Dorado et al., en prensa) y fueron
encontrados para Liomys irroratus en el presente trabajo. 1. Este trabajo 2.Pérez-
Saldaña, 1978 3. Janzen, 1986, 4. Ceballos, 1989; 5. Mendoza, 1997. 6.
Domínguez, 2000, 7. Hernández, 2000. *son las spp no reportadas para L. pictus y
descritas para L. irroratus en este trabajo. Los datos (3.) de Janzen (1986) de
contenidos de los abazones de Liomys salvini en selvas secas de Costa Rica.126
Anexos……………………………………………………………………………………..129 Anexo 1. Medidas y datos tomados a los ratones capturados………………….129
Anexo 2. Fórmula del formaldehído alcohol; (F. A. A.)………………………….130
Anexo 3. Fórmula de la gelatina glicerinada……………………………………...130
Anexo 4. Técnica para la preparación de laminillas temporales……………….130
Anexo 5. Técnica para la preparación de laminillas permanentes……………..131
Anexo 6. Técnica para la preparación de las laminillas de las excretas……....133

1
or mí se va a la ciudad del llanto; por mí se va al eterno
dolor; por mí se va hacia la raza condenada: La justicia
animó a mi noble arquitecto; que hizo la divina potestad, la suprema
sabiduría y el primer amor. Antes que yo no hubo nada creado, a
excepción de lo eterno, y yo duro eternamente. ¡Oh vosotros los que
entráis, abandonad toda esperanza!
Canto III Infierno.
La Divina Comedia
Dante Alighieri
IN MEMORIAM:
Edgard Mason Villalobos; por el ejemplo paternal.
Manuel Víctor Romo Martínez; por el ejemplo de vida.
J. Félix Frías Sánchez; por el ejemplo académico.
P

2
I. Dedicatoria.
A ti fuerza invisible, intangible, omnipresente y amorosa que llamo con todo orgullo y
admiración: Dios, porque has estado conmigo en cada momento de mi vida cuidándome y
dándome fuerza para sobrevivir lo indeseable y humildad para no perder el camino, a
pesar que muchas veces, tu bien lo sabes, no te hago mucho caso, gracias por TODOS tus
ángeles.
A mi madre, quien me ha marcado con su amor y sin quien no estaría en este mundo e
igualmente importante; no sería quien soy, te amo y espero enorgullecerte.
A mi padre, con quien me faltó tanto por vivir juntos, pero quien está conmigo en todo
momento y siempre lo estará, en mi amor por el y por nuestra familia.
A mi hermano, a quien amo y con quien he compartido más de lo que ambos podemos y
queremos aceptar y con quien quisiera compartir muchos más.
A mi hermana, a quien amo mucho más de lo que he demostrado, pero quien finalmente es
mi hermanita y le tengo que dar lata y cuidarla, siempre.

3
II. Agradecimientos.
¡Muchas gracias!
A mi familia sin cuyo amor y apoyo incondicionales este proyecto nunca
hubiera sido posible.
Para todos los aquellos que, directa o indirectamente, ayudaron a que este
proyecto llegara a buen puerto, especialmente a los que, de manera involuntaria y
torpe pudiera olvidar en este apartado. ¡Ustedes disculparán!
Dr. David Valenzuela por haberme aceptado, asesorado y aguantado a lo
largo de todo el proyecto, así como por introducirme al mundo de la investigación
científica.
A CONACyT (proyecto J34490-V] por el financiamiento del proyecto y la beca
que fue me otorgada como parte del proyecto.
A mi compañero de tesis Martín Cadena por la gran ayuda prestada en el
trabajo de campo sin la cual este trabajo no hubiera sido posible.
Goyo Rendón; por que sin su apoyo, amistad y buena disposición, más allá de
cualquier recompensa u horario, en todo el trabajo de campo y la identificación del
material vegetal, no se hubiera podido llevar a cabo este proyecto.
M. en C. (ya casi Dra.) Estela Sandoval Zapotitla por su amistad, paciencia,
disposición y apoyo para el catálogo de referencia sin el cual esta tesis nunca
hubiera concluido de manera satisfactoria, así como a todos en el Jardín,
especialmente a: Barbarita, Alicia, Conchita, Ito y Miguel, por su siempre util ayuda.
Biol. Alejandro Flores, Biol. Rolando Ramírez, la M. en C. Martha Olvera y

4
Amado García por su invaluable apoyo en la identificación del material vegetal de
este proyecto.
Para los Drs: Jerry Holechek, Guy Cameron y Fernando Clemente, por sus
consejos de larga distancia que fueron básicos para estructurar la parte
metodológica del proyecto.
A mis revisores, por sus valiosas aportaciones durante las diferentes etapas
del proyecto; Biols. Marco Lozano, Rafael Monroy, Salvador Santillán, Drs. Antonio
Guerrero, y Nestor Mariano, quien sacó al proyecto y a mí de incontables problemas
con la estadística.
Al Dr. Valentino Sorani, jefe del Laboratorio de sistemas de información
geográfica del CEAMISH, quien otorgó los mapas de vegetación con los que se
eligieron los sitios de muestreo.
A los Drs: Ella Vázquez, Gerardo Ceballos, Rodolfo Dirzo, Daniel Janzen, J.
Félix Frías, Douglas Johnson, y Álvaro Miranda, por las interesantes sugerencias
que directa o indirectamente fortalecieron al proyecto.
A las (casi) M. en C. Yola y Bety quienes han ayudado, desde el inicio, tanto
con su trabajo como con su amistad.
A toda la banda, ellos bien saben quienes son, que me brindaron su cariño y
amistad incondicionales y siempre estuvieron prestos a escucharme y apoyarme en
lo que estuvo en sus manos, en especial a los que dieron su tiempo, sudor,
cansancio y trabajo sin recompensa alguna y que merecen más que este
agradecimiento y los que ya les di personalmente.

5
1. Resumen
Se estudiaron los efectos de la perturbación del hábitat y la estacionalidad
climática en la dieta de Liomys irroratus (Heteromyidae) en la selva seca de la
Reserva de la Biósfera Sierra de Huautla, Morelos, México. De agosto del 2002 a
mayo del 2003 se realizaron trampeos mensuales en seis sitios de muestreo: tres
perturbados y tres no perturbados. De los ratones se tomaron medidas
morfométricas estándares, muestras de excretas y contenido de abazones. Para
evaluar la estacionalidad, se determinaron tres épocas: lluvias, secas tempranas y
secas tardías. Se realizaron muestreos de herbáceas y banco de semillas por época,
para evaluar la disponibilidad y diversidad de alimento por sitio. La dieta de este
roedor se determinó con el contenido de los abazones y análisis microhistológicos de
excretas. La diversidad y disponibilidad de recursos variaron a lo largo del tiempo, en
lluvias fueron más importantes las herbáceas y en secas las semillas. La dieta de los
ratones reflejó el cambio en la disponibilidad siendo más herbívoros en lluvias y más
granívoros en las secas. La perturbación cambió la cantidad y la composición de los
recursos que utiliza L. irroratus, en los sitios no perturbados siendo más selectivo
mientras en los perturbados es más generalista.

6
2. Introducción
El conocer la dieta de una especie y la forma en la que consume los recursos
alimenticios en relación a su disponibilidad en el medio, es una pregunta relevante
para la ecología, puesto que ayuda a entender las causas de su distribución y
abundancia así como los cambios que se puedan suscitar en estos aspectos (Begon
et al., 1999; Litvaitis, 2000; Krebs, 1985; Kronfeld & Dayan, 1998; Smith & Smith,
2001).
Entre las consecuencias de la perturbación de los hábitats naturales se
encuentran la modificación de la heterogeneidad y el cambio en la disponibilidad de
los recursos alimentarios, que a su vez impactan en la diversidad biológica de un
lugar en particular (Begon et al., 1999; Brown, 2000; Laurance & Bierregaard, 1997;
Litvaitis, 2000; Miller, 1994).
Por otra parte, la estacionalidad ambiental, es decir, los cambios periódicos
en los factores abióticos particularmente en la temperatura y la precipitación (Krebs,
1985), también genera cambios en la disponibilidad temporal de recursos.
La perturbación y la estacionalidad causan cambios en la heterogeneidad,
disponibilidad y la diversidad de los recursos alimenticios (Begon et al., 1999; Brown,
2000; Laurance & Bierregaard, 1997; Litvaitis, 2000; Miller, 1994) y estos cambios a
su vez impactan en la diversidad biológica de un lugar en particular (Begon et al.,
1999; Brown, 2000; Laurance & Bierregaard, 1997; Litvaitis, 2000; Miller, 1994).
Algunos de los efectos sobre la diversidad biológica, pueden manifestarse con
cambios en aspectos de su ecología de forrajeo (sensu Rosenzweig, citado en

7
Brown, 2000) como la amplitud de nicho (Feisinger et al., 1981; Levins, 1961; May &
MacArthur, 1972). Esto es más notorio en sistemas con una alta variabilidad
estacional de recursos (Brown, 2000; Krebs, 1985).
Los comportamientos de forrajeo y la heterogeneidad (tanto espacial como
temporal) están muy relacionados (Brown, 2000). La mayoría de las especies tienen
comportamientos de forrajeo flexibles que permiten respuestas adaptativas a
contingencias (como la estacionalidad y la perturbación) en el ambiente en el que
forrajean. La forma en la que responde el animal a la contingencia incluye
modificaciones a sus tiempos de alimentación, distribución de su actividad y tácticas
de alimentación (Brown, 2000).
La escala a la que sucede la contingencia puede ser espacial o temporal y
ambas pueden variar en su duración o área de afectación, lo cual causa que se
generen diferentes respuestas por parte de los animales (discutidas a detalle por
Brown, 2000). A las contingencias con escalas temporales y espaciales pequeñas
(como podría ser la estacionalidad ambiental de la selva baja caducifolia y
perturbación relativamente reciente) los animales responden con cambios en sus
ecologías de forrajeo variando la selección de su dieta (Brown, 2000).
La distribución y abundancia de los recursos alimenticios disponibles para un
animal pueden influenciar de manera importante su comportamiento de forrajeo y por
lo tanto su dieta (Brown, 2000; Larter & Gates, 1991; Pyke, 1984).
El nicho (trófico) está definido como: la función (alimentaria) de un organismo
(o especie) en una comunidad (Krebs, 1985). La amplitud de nicho está definida
como la cantidad utilizada de un nicho disponible (Colwell & Futuyma, 1971). La
amplitud del nicho trófico de un animal (Levins, 1968) está determinada por la

8
cantidad de recursos (riqueza de especies o categorías tróficas) que consume, pero
también está determinada por la forma en que los consume. La amplitud de nicho
trófico será menor para una especie que muestra preferencia por algunas especies
(especialista) que para una generalista (Colwell & Futuyma, 1971; Swihart et al.,
2003).
La amplitud de nicho trófico es un factor determinante en la tolerancia de una
especie a la pérdida y fragmentación del hábitat. La habilidad de una especie a
depender de una diversidad mayor de alimentos (estrategia generalista) en su rango
geográfico le permitirá una mayor tolerancia a la perturbación del hábitat (Swihart et
al. 2003).
El factor de estabilidad ambiental (Krebs, 1985) establece que las regiones
con climas más estables (biológicamente más diversos) permitirán la evolución de
nichos “más pequeños” y más especializados, y viceversa para los organismos que
viven en ambientes menos estables, que se ven forzados a ser más flexibles (menos
selectivos).
Una mayor heterogeneidad ambiental, que promueve una mayor
biodiversidad, es seguida por comportamientos de selección de hábitat y de dieta
(Brown, 2000; Rozensweig, 1987; Rozensweig, 1991). En ambientes con más
especies para consumir, un forrajeador puede seleccionar algún (o algunos)
recurso(s) en particular (Brown, 2000).
En el caso específico de las selvas secas la estacionalidad causa cambios
abruptos en la fenología de las plantas y en la disponibilidad de agua y alimentos, lo
que presiona a los animales a generar estrategias para poder sobrevivir (Ceballos,
1995). Y la perturbación muchas veces causa disminución en la heterogeneidad

9
ambiental (Begon et al. 1999; Krebs, 1985).
Las selvas tropicales secas son uno de los ecosistemas con mayores
problemas de perturbación en el mundo (Dirzo & Raven, 2003; Janzen, 1986b;
Janzen, 1988; Lerdau et al., 1991; Murphy & Lugo, 1986).
Uno de los principales cambios, asociados a la perturbación de la selva seca
es una disminución de la cobertura arbórea (Cabin et al., 2002: Dirzo & Miranda,
1990; Janzen, 1988; Lerdeau et al., 1991; Maas, 1995; Murphy & Lugo, 1986).
También se ha documentado que la perturbación de la selva seca ocasiona una
menor disponibilidad de frutos carnosos (De León, 2005). Hay evidencias de
cambios estacionales notables en la disponibilidad y composición específica de
semillas en el banco de semillas (Arias, 1986). Ambos trabajos realizados en la
Sierra de Huautla.
En este escenario, se espera que las especies animales sean más selectivas
cuando el ambiente tenga más diversidad de recursos y más generalistas cuando el
ambiente sea menos diverso. De esta forma se decidió estudiar la dieta del ratón
Liomys irroratus, que es el único roedor granívoro presente en la zona y la segunda
o tercera especie más abundante (Cadena, 2003) y los efectos que sobre su dieta
pudieran tener la perturbación antropogénica, la estacionalidad climática y los
cambios en la heterogeneidad ambiental.

10
3. Antecedentes
3.1. Comprobación empírica del factor de estabilidad ambiental
Los factores que modifican la amplitud de nicho son: la riqueza de especies
disponibles y la forma de utilizar la riqueza por parte del consumidor (Colwell &
Futuyma, 1971; Swihart et al., 2003), para conocer en que forma interactúan estos
factores se analizaron los cambios en la amplitud de nicho trófico (Levins, 1968) con
datos de Cortés et al., (2002), Morris & Schwartz (1957), Murray & Dickman (1994),
Reichman (1977), Smith (1952) y Taylor (1954) y se encontró que:
1. La riqueza específica y la selección de elementos en la dieta afectan de
manera directa la amplitud de nicho trófico, como señala la teoría (Colwell &
Futuyma, 1971; Swihart et al., 2003).
2. En situaciones en las que la selección es indistinta (estrategia generalista)
el aumento de la riqueza específica disponible aumenta la amplitud de nicho trófico.
3. Cuando la riqueza de especies disponibles se mantiene estable en
diferentes circunstancias (ej. estacionalidad, perturbación, competencia) y la especie
selecciona los elementos disponibles de manera diferencial (especialista) la amplitud
de nicho trófico disminuye, comparativamente con la de una especie generalista en
la misma circunstancia.
4. Por lo tanto: en ambientes diversos (ej. tropicales, no perturbados o en la
temporada en la que tienen más recursos disponibles) una especie generalista
tendrá una amplitud de nicho trófico mayor que una especie con estrategia
especialista en sitios de baja diversidad (ej. perturbados, temporada baja en

11
recursos), a pesar que la primera consuma mayor riqueza de especies, lo que indica
que la amplitud de nicho trófico estará afectada en mayor medida por la estrategia
de forrajeo de un animal que por la diferencia en la diversidad del medio.
5. Estos datos empíricos corresponden a lo señalado por la teoría del factor
de estabilidad ambiental (Krebs, 1985): en ambientes menos estables los animales
tendrán dietas más generalistas y en ambientes más estables su dieta será más
especializada.
3.2. Acercamientos metodológicos al problema de determinar la dieta.
La dieta de las especies ha sido ampliamente estudiada durante muchas
décadas, con enfoques distintos, para un variado número de grupos y con
metodologías diversas.
Existen diferentes metodologías utilizadas para determinar la dieta de los
animales herbívoros. Holechek et al., (1982a.) las dividen en las siguientes
categorías:
(1) La utilización (utilization technique): es una de las formas más antiguas
para evaluar la dieta de herbívoros que pastan. Se evalúa mediante la comparación
del grado de remoción de vegetación por un herbívoro (Norbury & Sanson, 1992).
Entre sus ventajas se cuenta la rapidez además de que proveé información de
donde y en que grado es usada la vegetación. Esta metodología tiene desventajas
importantes como: pérdidas a larga escala por efectos diferentes a la herbivoría (ej.
climáticos, daño físico) y consumo por otras especies, así como en lugares donde el
crecimiento vegetal es muy activo y no permite medir la pérdida por herbivoría.

12
(2) La técnica de observación directa: en la cual se intenta observar e
identificar lo que el animal está ingiriendo en el momento en que lo consume (ej.
Laylock et al., 1972; Reppert, 1960; Smith & Hubbard, 1954; Weckerly, 1994). Esta
técnica tiene como ventajas el ser rápida, tanto en el campo como en el cómputo de
los datos (Laylock et al., 1972) y requiere un mínimo de equipo (Holechek et al.,
1982b; Norbury & Sanson, 1992). Entre las desventajas que tiene es que requiere un
amplio entrenamiento para identificar las especies vegetales que son consumidas
(Laylock et al., 1972; Holechek et al., 1982b), así como la dificultad para observar a
los animales en topografías agrestes, o cuando hay arbustos u otros obstáculos
bloqueando al observador (Sanders et al., 1980). Es complicado calcular la cantidad
de alimento que consume un animal en cada mordisco, pues ésto varía entre
individuos (Holechek et al., 1982b). Pero posiblemente la mayor limitante que
presenta esta técnica es que el observador necesita estar muy cerca del animal y
por lo tanto necesita de animales domesticados (Holechek et al., 1982b) y a pesar de
esto, la presencia del observador puede alterar la conducta del animal.
(3) Las metodologías que requieren de análisis microscópico (el cual se
discutirá a detalle más adelante) para determinar la dieta, entre las cuales destacan:
(a) Las sondas o fístulas esofágicas y, (b) la evacuación o fistulación de rumen.
Estas metodologías tienen como ventaja el presentar resultados parecidos a los de
la observación directa, relativamente precisos, sin los errores aducidos al observador
(Laylock et al., 1972), por lo tanto, muchas veces son preferidas para el estudio con
ganado (Free et al., 1970; Galt et al., 1969; Peden et al., 1974). Sin embargo, estas
metodologías también tienen problemas importantes, puesto que, en el mejor de los
casos, la cirugía de esófago es difícil y el gran número de pérdidas ha causado

13
escepticismo en su uso (Van Dyne & Torell, 1964). El comportamiento anormal de
los individuos con fistulación del rumen es un problema grave (Holechek et al.,
1982b; Peden et al., 1974), así como el alto costo de mantenimiento y cuidado (de
ambas) problemas que se acentúan cuando se le aplica a ungulados silvestres
(Peden et al., 1974). Estas técnicas también consumen una gran cantidad de tiempo,
tanto en campo como en el laboratorio (Laylock et al., 1972; Van Dyne & Torell,
1964), están limitadas a animales grandes (Holechek et al., 1982b; Olson, 1991) y
requieren un alto número de animales para poder generar datos precisos (Holechek
et al., 1982b). En específico, la recolecta de material del rumen no es posible en
lugares abiertos ni durante el invierno en localidades templadas (Olson, 1991).
En su trabajo, Galt et al., (1969) compararon los resultados de estas
metodologías contra los de la observación directa y demostraron diferencias
considerables entre ambas metodologías.
(c) El análisis de contenidos estomacales e intestinales (ej. Baumgartner &
Martín, 1939; Campos et al., 2001; Johnson, 1961) presenta la ventaja de poder
determinar con precisión la composición de la dieta, tanto cualitativa como
cuantitativamente. Su principal desventaja radica en la necesidad de sacrificar a los
animales, por lo tanto, está restringida a ser utilizada primordialmente con animales
silvestres con poblaciones grandes (Holechek et al., 1982b). La destrucción
diferencial de las especies durante la digestión causa cambios en las proporciones
de los diferentes elementos consumidos (Holechek et al., 1982b), el lugar donde
fueron consumidos no puede ser determinado (Holechek et al., 1982b) así como la
subestimación o simplemente el no detectar grupos completos (Westoby et al.,
1976).

14
(d) La obtención y análisis de excretas (ej. Dusi, 1949; Vázquez, 1997;
Stewart, 1967; Kronfeld & Dayan, 1998). Las ventajas que esta metodología confiere
han sido discutidas por: Anthony & Smith (1974), Fitzgerald & Waddington (1979),
Holechek et al., (1982a), Kronfeld & Dayan (1998), Johnson & Wofford (1983),
Litvaitis (2000), Norburg & Sanson (1992), Peden et al., (1974), Putman (1984),
Rogers & Dahlgren (1982), Sanders et al., (1980), Smith et al., (2002), Todd &
Hansen (1973), Vavra et al., (1978), Williams (1969) y Zyznar & Urness (1969), y
son las siguientes:
1. No interfiere con los hábitos naturales de los animales.
2. No restringe el movimiento de los animales.
3. Permite la determinación de la dieta sin importar las condiciones del terreno.
4. Es igual de útil para animales domésticos y silvestres.
5. No requiere de sacrificar animales.
6. Reduce al mínimo el contacto con los animales.
7. Es relativamente objetiva.
8. Tamaño de muestreo casi ilimitado.
9. Las excretas son relativamente fáciles de recolectar.
10. Se pueden comparar las dietas de dos o más especies a la vez.
11. En la actualidad el muestreo requiere de poco equipo.
12. Tiene un valor particular cuando los animales se desplazan en distintas
comunidades.
13. Es el único método adecuado para trabajar con especies raras o
amenazadas.
14. No requiere ningún método quirúrgico, por lo cual es el método más

15
apropiado para trabajar con ganado (Mohammad et al., 1995).
Pero esta metodología también tiene importantes desventajas y problemáticas, las
cuales fueron discutidas por: Fitzgerald & Waddington (1979), Hansen et al., (1973),
Holechek (1982), Holechek et al., (1982a), Holechek & Gross (1982a), Johnson &
Wofford (1983), Katona & Altbäcker (2002), Litvaitis (2000), Norbury & Sanson
(1992), Owen (1975), Sanders et al., (1980), Smith et al. (2002), Smith & Shandruk
(1979), Stewart, (1967), Vavra & Holechek (1980) y Williams (1969). Y son las
siguientes:
1. La precisión es un problema importante pues la proporción en que las
especies son registradas en las excretas muchas veces no es igual, o parecida,
a las consumidas.
2. Los índices de preferencia no pueden ser precisos porque no se puede
determinar donde fue consumido el alimento.
3. La identificación de las excretas (a que especie pertenecen) recolectadas en
campo puede ser un problema. Howard (1967) describe un método (que
involucra el pH) que puede servir para diferenciar entre animales diferentes con
excretas parecidas.
4. Equipo, trabajo y tiempo considerables son necesarios para el análisis.
5. Se requiere una vasta colección de referencia.
6. El observador requiere un entrenamiento considerable para identificar
correctamente los fragmentos de tejido vegetal (Holechek & Gross, 1982a).
7. En muchas plantas es difícil la identificación a nivel de especie y a veces de
género.
8. La identificación de los fragmentos es laboriosa y tardada.

16
9. La destrucción de algunas plantas puede ocurrir al preparar la laminilla
(Holechek, 1982; Vavra & Holechek, 1980).
10. Algunos procedimientos para la separación y preparación de la muestra
pueden causar errores.
11. Algunas especies pueden no ser identificables en las heces.
12. La identificación se complica mientras más tiempo tenga la excreta antes de
la recolecta.
13. La fragmentación puede diferir entre las especies durante la digestión
haciendo parecer diferente la proporción relativa de especies observadas.
1. El índice de digestión (uno de los problemas más notorios y más estudiados
de la técnica) puede variar dependiendo tanto de la especie consumidora como
de las características de la planta que esta consuma (Urness et al., 1977) por lo
cual, en algunos casos las estructuras de alguna(s) especie(s) puede(n) no
conservarse a pesar de ser ingeridas, o ser tan pequeñas que la identificación es
laboriosa y tardada (Williams, 1969). Bartolomé et al., (1995) encuentran
diferencias significativas en la digestión del tejido vegetal que generan la
subestimación de las hierbas y la sobreestimación de los pastos. En contraste
Johnson & Wofford (1983) y Todd & Hansen (1973) sostienen que el grado de
digestión no afecta la identificación de los fragmentos de manera importante.
Existen metodologías complementarias a las ya mencionadas: la comparación
de concentración de n-alcanos en las heces (Smith et al., 2002), la creación de una
válvula a control remoto ubicada en el esófago (Smith et al., 2002), así como otras
metodologías comparadas en ovinos por Laylock et al., (1972).
Existen otras metodologías menos comunes que son utilizadas por algunos

17
autores sin lograr generalizarse. Norbury & Sanson (1992) mencionan una técnica
poco utilizada que se basa en la toma directa del material vegetal recién comido de
la boca del herbívoro; el problema principal de la metodología es que la cantidad de
vegetación recabada por boca es muy baja y la técnica requiere del sacrificio de una
cantidad muy grande de animales, por lo que la refieren como indeseable.
Kronfeld & Dayan (1998) diseñaron una metodología, específicamente para
roedores y otros mamíferos pequeños, que consiste en la regurgitación del alimento
ingerido, por medio del bombeo del contenido estomacal. Ésta es una opción más
para evitar el sacrificio de los animales que es necesario al aplicar el análisis de
contenido estomacal e intestinal y que son las técnicas comúnmente utilizadas para
el trabajo con mamíferos pequeños.
En algunos casos, también se aplican fórmulas estadísticas para corregir las
fallas propias de la estimación por métodos de observación directa y las
metodologías indirectas (Peden et al., 1974; Smith et al., 2002).
3.3. La anatomía vegetal como un apoyo para determinar la dieta de los
herbívoros.
Los trabajos para determinar la composición de la dieta, por medio de
excretas (de mamíferos carnívoros y aves, específicamente rapaces) han sido
utilizados por muchas décadas [ej. Aranda et al., (2002); Clevenger et al., (1992);
Selko (1937); Sperry (1934); Taylor (1954)]; sin embargo; la identificación de las
especies a las que pertenecen los fragmentos en estos estudios está restringida a
grandes partes no digeridas, tales como exoesqueletos quitinosos de insectos, pelo,

18
huesos, semillas y agujas de pinos (Zyznar & Urness, 1969). Habitualmente los
mamíferos herbívoros mastican y degradan los elementos de su dieta tan finamente
que la utilización de la anatomía se vuelve necesaria para la identificación de las
especies (Zyznar & Urness, 1969).
La aplicación de análisis microhistológicos para la determinación de la dieta
de animales herbívoros fue publicada por primera vez por Baumgartner & Martin
(1939), para determinar los hábitos alimentarios de ardillas (Sciurius Níger, por
medio de contenidos estomacales). Posteriormente, Dusi (1949) aplicó esta
metodología para el trabajo con excretas de Sylvilagus floridanus, desde entonces
son muchos los autores quienes han trabajado con diversos grupos de animales, por
ejemplo: Hansen & Dearden (1975) con venado bura (Odocoileus hemionus);
Crawford (1982) con venado cola blanca (Odocoileus virginianus); Galt et al., (1969)
con ganado (Bos taurus); Rumble & Anderson (1993) con guajolotes (Meleagris
gallipavo); Sparks (1968) con liebres (Lepus californicus); Smith & Shandruk (1979)
con berrendos (Antilocapra americana), roedores (Johnson, 1961; Petersen, 1993),
Fitzgerald & Waddington (1979) con zarigüeyas (Trichosurus vulpecula) y, Owen
(1975) con gansos (Branta leucopsis y B. ruficollis). Esta técnica también ha
probado ser de utilidad para la determinación de la dieta de insectívoros, pero con
ciertas reservas en la cuantificación de insectos de cuerpos suaves (Dickman &
Huang, 1988). Esta técnica es útil también para conocer el traslape en la dieta de
especies que coinciden en una zona geográfica, como lo señalan en sus trabajos
Cameron (1983); Hansen et al., (1973); Hansen & Reid (1975); Hansen & Ueckert
(1970); Johnson (1961); Moro & Bradshaw (2002); Petersen (1993); Stancampiano &
Caire (1995); Wondolleck (1978).

19
La precisión de la técnica microhistológica, específicamente aplicada a
excretas, ha sido puesta en duda por varios autores (Anthony & Smith, 1974;
Johnson & Pearson, 1981; Norbury & Sanson, 1992; Sanders et al., 1980). A pesar
de ello existen trabajos teóricos que analizan dietas artificiales, compuestas a mano,
que sustentan la eficiencia de esta metodología para medir cualitativa y
cuantitativamente la dieta de los herbívoros (Holechek, 1982; Holechek et al., 1982;
Sparks & Malechek, 1968; Vavra & Holechek, 1980; Williams, 1969). En el trabajo de
Todd & Hansen (1973) sostienen que la técnica es igualmente eficiente al analizar
rumen y excretas, pues a pesar de que existe mayor grado de digestión en las
excretas ésta no afecta la discernibilidad del mismo.
La técnica descrita por Baumgartner & Martin (1939) genera la posibilidad de
analizar cualitativamente (presencia y ausencia) y cuantitativamente (que especie o
parte es preferida en relación con otras) la dieta de los herbívoros. En el caso de
este segundo enfoque, el cuantitativo, existen varios trabajos que han intentado
definir la preferencia de la dieta de diversos grupos (teniendo problemas en lograr su
objetivo).
Stewart (1967) alimentó con cantidades conocidas de plantas previamente
seleccionadas a siete especies de herbívoros africanos. Smith & Shandruk (1979)
consideran que “el conteo de fragmentos epidérmicos fue considerado inválido por la
diferencia en sus tamaños”.
Zyznar & Urness (1969) mencionan en su trabajo que: “Pocas veces las
muestras de excretas contenían algo más que un porcentaje bajo de fragmentos
identificables” eso puede ser explicable pues el método con el que fueron aclaradas
las excretas (hirviéndolas 15 minutos en hidróxido de sodio) destruye las estructuras

20
identificables (Holechek, 1982). Esto logró generar incertidumbre en la posibilidad de
cuantificar la dieta por medio de esta metodología.
Sparks & Malechek (1968) estimaron los porcentajes de composición por
medio de los pesos en seco de los elementos de una mezcla de vegetales
identificados, en proporciones conocidas, obteniendo que: “La relación entre el
estimado del porcentaje de peso en seco (densidad relativa) y los porcentajes reales
de peso en seco es aproximadamente de 1:1 para las tres categorías,”…“Por lo
tanto, la composición porcentual basada en el peso en seco de las mezclas puede
ser estimado directamente de la densidad relativa”.
Free et al., (1970) verificaron la eficiencia de esta metodología en la
cuantificación de los fragmentos de especies de plantas perennes en excretas de
becerros, desde entonces esta técnica ha sido ampliamente utilizada (Hansen &
Dearden, 1975; Hansen & Reid, 1975; Hansen et al., 1973; Havstad & Donart, 1978;
Holechek et al., 1982b; Holechek, 1982; Peden et al., 1974; Sanders et al., 1980;
Vavra & Holechek, 1980) obteniendo resultados positivos.
La parte matemática de la técnica ha sido tratada en extenso en el artículo
publicado por Johnson (1982) en el que describe detalladamente la fórmula por la
cual se obtiene la relación entre densidad relativa y peso en seco, tema nunca antes
tratado “presumiblemente pasado por alto dado que (el procedimiento) es
relativamente sencillo de entender para personas con una amplia formación en
matemáticas”.
Otra cuestión de suma importancia para la cuantificación de dietas por
métodos microhistológicos es el apropiado adiestramiento de quien vaya a preparar
y observar las muestras, ésto es destacado en los trabajos de Holechek & Gross

21
(1982a), Holechek et al. (1982), Vavra & Holechek (1980) y Westoby et al., (1976).
3.4. Diferentes enfoques en los estudios de dieta.
Además de existir gran cantidad de trabajos que se enfocan al conocimiento
de la dieta, de manera descriptiva de diversos taxa de herbívoros (Galt et al., 1969;
Holechek et al., 1982b; Reppert, 1960; Smith & Hubbard, 1954; Sparks, 1968; Uresk,
1984); existen trabajos dirigidos a mejorar las técnicas aplicadas al conocimiento de
la dieta, generando metodologías nuevas (Baumgartner & Martin, 1939; Peden et al.,
1974; Sparks & Malechek, 1968; Williams, 1969).
Reichman & Van de Graaf (1975) relacionan el consumo de material vegetal
fresco con la reproducción de roedores de zonas áridas.
Se han realizado distintos análisis y comparaciones de las metodologías
existentes para buscar la mejoría de los trabajos posteriores (Holechek, 1982;
Holechek et al., 1982; Mohammad et al., 1997; Sanders et al., 1980; Smith &
Shandruk, 1979; Urness et al., 1977; Vavra et al., 1978; Vavra & Holechek, 1980).
Existen de igual manera publicaciones enfocadas a la revisión del avance que las
diferentes metodologías han tenido como es el caso de Holechek et al., (1982a) para
el caso de la determinación botánica de la dieta en general y Van Dyne & Torell
(1964) para el caso específico de la fístula esofágica.
A pesar que Hansen & Dearden (1975), Reppert (1960), Sparks (1960) y
Ward & Keith (1962) sugieren que las preferencias alimentarias y la calidad del
forraje son influenciadas por el proceso estacional, una cantidad relativamente
pequeña de publicaciones analizan, de manera directa o indirecta, las repercusiones

22
de la estacionalidad ambiental como factores determinantes en la dieta (ej. Aleksiuk,
1970; Altrichter et al., 2000; Bradley & Mauer, 1971; Crawford, 1982; Grajales-Tam
et al., 2003; Hansen & Dearden, 1975; Jameson, 1952; Posse et al., 1996; Sparks,
1968; Taylor, 1954; Vázquez, 1997; Vázquez et al., 2000), sólo los dos últimos
fueron realizados en ambientes tropicales.
Las posibles repercusiones de la perturbación sobre la dieta es un enfoque
más reciente y existen muchos menos trabajos que lo analicen (Vázquez, 1997,
Vázquez et al., 2000).

23
4. Justificación
Los roedores son un eslabón fundamental en las redes tróficas dado que son
presa de muchos animales (Ceballos y Galindo, 1984; Fleming, 1983a; Fleming,
1983b; MacCracken et al., 1985; Sánchez-Cordero & Fleming, 1993) son
importantes como depredadores y dispersores de semillas, así como depredadores
de insectos y plántulas; también participan en mantener la fertilidad y la diversidad
vegetal a nivel local (Ceballos y Galindo, 1984; Ceballos y Miranda, 2000; Dhillion,
1999; Huntly & Reichman, 1994; MacCracken et al., 1985; McDonald, 2001; Villa y
Cervantes, 2003). Por lo tanto, los estudios dirigidos hacia el conocimiento y
conservación de éste grupo de mamíferos son de relevancia para conservar los
demás niveles tróficos que dependen de ellos para su existencia.
Como parte del proyecto (CONACyT J34490-V): “Mastofauna de la Reserva
de la Biósfera Sierra de Huautla, Morelos: diversidad, patrones espacio-temporales y
conservación”, ésta tesis está encaminada a conocer mejor los aspectos biológicos
y ecológicos de las especies de la zona de manera que se pueda generar
conocimiento básico y aplicado a la conservación y el manejo de los mamíferos de la
reserva y sus alrededores.
Liomys irroratus es un buen sujeto de estudio para alcanzar los objetivos
(determinados más adelante) de este trabajo porque:
1. Es el único heterómido en la zona, por lo que es fácil de identificar.
2. Presenta características de especialización a medios semiáridos, como son los
abazones: estructuras especializadas de almacenamiento y transporte de alimentos

24
(Ceballos y Galindo, 1984; Ceballos & Miranda, 2000; Dowler & Genoways, 1978) y
una alta capacidad de regulación de temperatura y agua, típicas de los heterómidos
(Hudson & Rummel, 1966).
3. Tomando en cuenta las características estacionales de la Sierra de Huautla se
puede inferir que su eficiencia biológica será alta lo que conllevará a un éxito
reproductivo alto y una densidad elevada (Begon et al., 1999; Darwin, 2001; Krebs,
1985). Esto está comprobado por estudios anteriores realizados en la zona (Álvarez-
Castañeda, 1996; Cadena (2003); García-Estrada et al., 2002; Morán, 1994 y
Sánchez & Romero, 1995) que reportan abundancias altas, por lo que se pueden
obtener tamaños de muestra suficientemente grandes. Cadena (2003) señala que el
35% de sus capturas, en los mismos sitios de muestreo, pertenecen a esta especie,
con densidades que varían de 0.96 a 40.37 individuos por hectárea (ind/ha) en sitios
no perturbados, y de 0.96 a 30.76 ind/ha en zonas perturbadas, estos cambios
fueron marginalmente significativos. La densidad entre ambas condiciones de
perturbación, si muestra diferencias significativas a lo largo del año (Cadena, 2003).
4. Se alimentan de semillas, hojas, tallos mientras que su consumo de invertebrados
es bastante bajo (Ceballos y Galindo, 1984; Ceballos & Miranda, 2000; Domínguez,
2000; Vázquez, 1997).
Por lo tanto, al generar un catálogo de formas micro histológicas de la cutícula
de estas estructuras vegetales y comparar el contenido de los abazones con lo
encontrado en el banco de semillas se puede suponer que se está tomando en
cuenta la gran mayoría de los componentes de su dieta, así como la frecuencia de
cada uno de ellos con un grado de confianza satisfactorio.
La utilización de la metodología de análisis de excretas por medios micro

25
histológicos, fue elegida para este estudio (por encima de otras metodologías
mencionadas) por que los ratones son muy pequeños para colocarles una fístula en
el esófago, así como por sus hábitos nocturnos, su talla y las condiciones
topográficas y de vegetación de la zona. La observación directa es ineficiente (si no
imposible). Se descartaron las metodologías habituales para mamíferos pequeños,
como el análisis de contenidos estomacales aplicado por Campos et al., (2001);
Dickman & Huang (1988); Kincaid & Cameron (1982); Jameson (1952); Luce et al.,
(1980); Reichman (1975), puesto que este trabajo se realizó en conjunto con una
tesis sobre la demografía de los roedores de la zona (Cadena, 2003) que requirió
marcar y recapturar a los ratones.
Existen una cantidad importante de estudios sobre la dieta de diferentes
grupos de mamíferos (Baumgartner & Martin, 1939; Dickman & Huang, 1988; Dusi,
1949; Katona & Altbäcker, 2002; Kincaid & Cameron, 1982; Preen, 1995; Smith &
Shandruk, 1979; Stewart, 1967; Wilson & Bertram, 1987).
La cantidad de trabajos con roedores es importante (ej. Cortés et al., 2002;
Dusi, 1949; Escobar & González, 1973; Fagerstone et al., 1981; Fleharty & Olson,
1969; Hansen & Ueckert, 1970; Jameson, 1952; Johnson, 1961; Luce et al., 1980;
Moro & Bradshaw, 2002; Petersen, 1993; Reichman, 1975; Torres-Contreras &
Bozinovic, 1997; Uresk, 1984; Voth & Black, 1973; Whitaker, 1966).
Un porcentaje importante de los trabajos sobre dieta de roedores se han
realizado con especies desérticas de la familia Heteromyidae (ej. Bradley & Mauer,
1971; Brown et al. 1979; Brown & Lieberman, 1973; Chapman, 1972; Frank, 1988;
Johnson, 1961; Lockard & Lockard, 1971; Reichman, 1975; Reichman, 1977;
Reichman & Van de Graaf, 1975; Rosenzweig & Sterner, 1970; Shaw, 1934; Smigel

26
& Rosenzweig, 1974; Wondolleck, 1978).
Para el caso de heterómidos tropicales es mucho menos lo que se conoce
(Reichman & Price, 1993; Sánchez-Cordero, 1993), existen muy pocos trabajos
sobre la dieta del género Heteromys, la información sobre la dieta de H. gaumeri, H.
nelsoni y H. oresterus es muy poca (Schmidt et al. 1989; Rogers & Rogers, 1992a y
Rogers & Rogers, 1992b; respectivamente) para Heteromys desmarestianus existen
algunos estudios sobre sus preferencias alimenticias en Costa Rica (Fleming, 1974a;
Fleming, 1977; Fleming & Brown, 1975; Vandermeer, 1979; Martínez-Gallardo &
Sánchez –Cordero, 1993).
Los trabajos sobre la dieta del género Liomys están limitados a dos especies:
L. pictus y L. salvini. La dieta de Liomys salvini ha sido estudiada a nivel de los
contenidos de abazones en varios trabajos, con enfoques diversos, en las selvas
secas de Costa Rica (Brewer, 2001; Fleming, 1974a, Fleming, 1977; Fleming &
Brown, 1975; Janzen, 1971; Janzen, 1981; Janzen, 1982a, Janzen, 1982b, Janzen,
1982c; Janzen, 1982d; Janzen, 1986a).
Los estudios realizados sobre la dieta de los Heterómidos tropicales
mexicanos han sido realizados con Liomys pictus, en Jalisco y solo se analizan
contenidos de abazones y madrigueras (Ceballos, 1989, Domínguez, 2000;
Hernández, 2000; Mendoza, 1997; Pérez-Saldaña, 1978). En un estudio de
laboratorio, Briones y Sánchez-Cordero (1999) compararon el nivel de consumo de
frutos y semillas por Liomys pictus y encontraron que los ratones son capaces de
distinguir las semillas más nutritivas y las potencialmente dañinas, encontraron
también que algunas de las especies que los ratones consumen de manera
importante no tienen altos niveles de nutrientes, por lo que suponen que pueden

27
utilizarlas con base en su disponibilidad o su humedad.
Existen muy pocos estudios que busquen entender el impacto de la
perturbación en la dieta de roedores en el país, como es el caso de los trabajos de
Vázquez (1997) y Vázquez et al., (2000) quienes determinaron la dieta de dos
especies de roedores (Reithrodontomys fulvecens y Peromyscus aztecus) en hábitat
con diferente grado de alteración en la Reserva de la Biosfera Sierra de Manantlán,
en Jalisco, México, en los que encontraron que la dieta varía dependiendo de la
estacionalidad y la perturbación del hábitat, debido a las diferencias en la
disponibilidad y la productividad generadas por la perturbación.
La importancia de esta investigación se debe a que no se han realizado
trabajos sobre dieta, con este enfoque particular, de esta especie en la zona de
estudio, siendo de este modo un proyecto original con un enfoque nunca antes
abordado en el estado de Morelos.

28
5. Objetivos
5.1. Objetivo general.
Conocer el efecto que tiene el grado de perturbación del hábitat y la
estacionalidad ambiental en la dieta de Liomys irroratus en la selva baja caducifolia
de la región de la Sierra de Huautla.
5.2. Objetivos particulares.
1. Determinar la abundancia, diversidad y valor de importancia de los recursos
alimenticios (ej. semillas y herbáceas) disponibles para Liomys irroratus en los sitios
de estudio.
2. Describir la dieta de Liomys irroratus (con base en el análisis de muestras del
contenido de abazones y en el análisis de muestras de excretas) y evaluar cómo
cambia entre sitios y temporadas.
3. Determinar la diversidad de la dieta y la amplitud de nicho trófico en la dieta de
Liomys irroratus en diferentes sitios y temporadas en la zona de estudio.
4. Evaluar si Liomys irroratus es selectivo con algunas de las especies que
consume y si esto cambia entre sitios y temporadas.

29
6. Hipótesis
Existe mayor disponibilidad y diversidad de semillas y de herbáceas en los
sitios no perturbados que en los perturbados. Por lo tanto, en vista de que se trata
de un ambiente más estable, se espera que la dieta de este roedor muestre mayor
selectividad y menor amplitud de nicho trófico en los sitios no perturbados que en los
perturbados.
Existe mayor disponibilidad y diversidad de semillas en la temporada de secas
en general comparativamente con las lluvias y una menor disponibilidad y diversidad
de herbáceas en la temporada seca. Se espera que la amplitud de nicho trófico para
semillas y herbáceas disminuya cuando existan más recursos disponibles (secas
para semillas y lluvias para plantas), por efecto de la selección que se da en
ambientes más estables, o que si L. irroratus forrajea de manera generalista una
mayor diversidad disponible aumente su amplitud de nicho trófico.
Se espera que en la dieta de este roedor la herbivoría sea más importante
durante la temporada de lluvias y disminuya al aumentar la desecación en el
ambiente y que la granivoría siga el patrón opuesto.
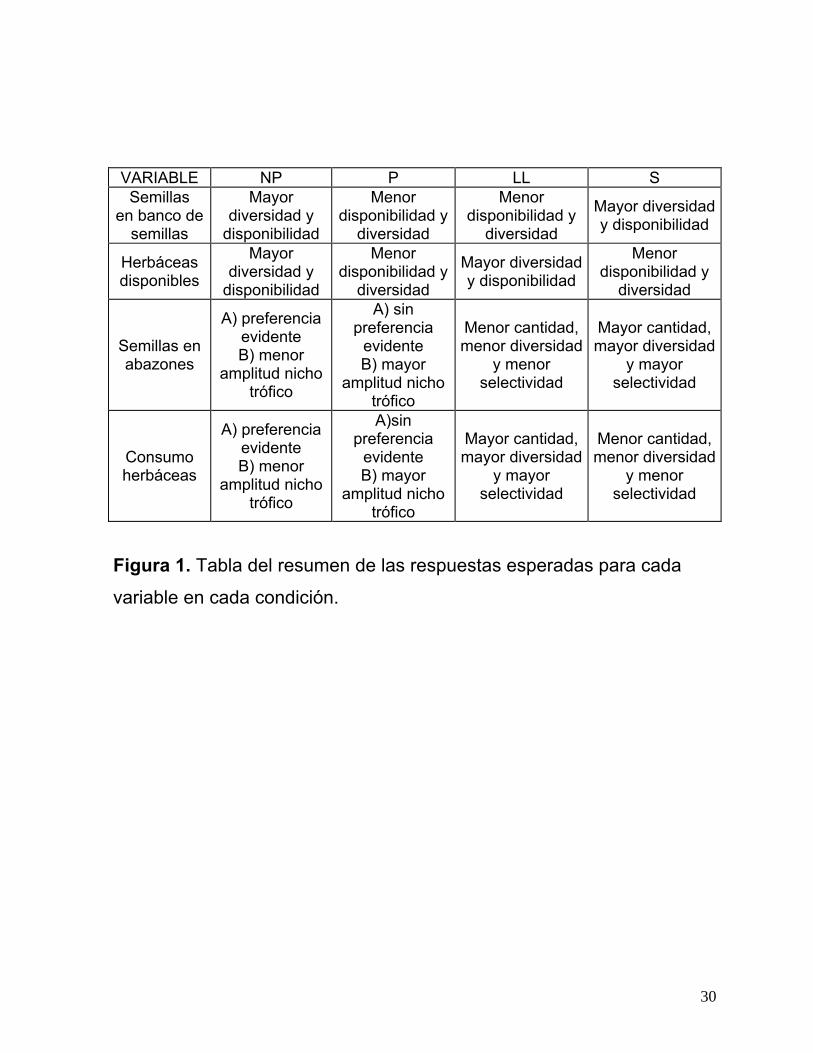
En la figura 1 se muestran, en una tabla, el tipo de respuestas que se
esperan para cada variable en los diferentes sitios y temporadas.
30
VARIABLE NP P LL S Semillas
en banco de semillas
Mayor diversidad y
disponibilidad
Menor disponibilidad y
diversidad
Menor disponibilidad y
diversidad
Mayor diversidad y disponibilidad
Herbáceas disponibles
Mayor diversidad y
disponibilidad
Menor disponibilidad y
diversidad
Mayor diversidad y disponibilidad
Menor disponibilidad y
diversidad
Semillas en abazones
A) preferencia evidente B) menor
amplitud nicho trófico
A) sin preferencia
evidente B) mayor
amplitud nicho trófico
Menor cantidad, menor diversidad
y menor selectividad
Mayor cantidad, mayor diversidad
y mayor selectividad
Consumo herbáceas
A) preferencia evidente B) menor
amplitud nicho trófico
A)sin preferencia
evidente B) mayor
amplitud nicho trófico
Mayor cantidad, mayor diversidad
y mayor selectividad
Menor cantidad, menor diversidad
y menor selectividad
Figura 1. Tabla del resumen de las respuestas esperadas para cada
variable en cada condición.

31
7. Especie de estudio
Liomys irroratus. Gray, 1868. (Proc. Zool. Soc. London, p. 205.).
(Ratón espinoso, Mexican Spiny Pocket Mouse).
7.1. Taxonomía.
Este ratón pertenece a la familia Heteromyidae la cual es endémica del
continente americano (Dowler & Genoways, 1978) y agrupa seis géneros y
aproximadamente 57 especies (Alexander & Riddle, 2005). La distribución de la
familia es principalmente en las zonas áridas y tropicales, siendo abundante en
vegetación xerófita, matorrales, pastizales, selvas secas y algunos bosques de pino-
encino (Ceballos y Galindo, 1984; Ceballos & Miranda, 2000; Dowler & Genoways,
1978; Schmidly et al., 1993). Los heterómidos son constructores de madrigueras y
comedores de semillas con ecologías superficialmente parecidas (Rosenzweig &
Sterner, 1970).
La familia se encuentra dividida en tres subfamilias: Perognathinae,
Dipodomyinae y Heteromyinae, en ésta última se encuentran dos géneros
cercanamente emparentados (Alexander & Riddle, 1993; Best, 1993; Dowler &
Genoways, 1978): Heteromys y Liomys (éste último descrito por Merriam en 1902).
Liomys comprende cinco especies, cuatro de éstas (L. irroratus, L. salvini, L. pictus y
L. spectabilis) habitan en México (Dowler & Genoways, 1978, Schmidly et al. 1993),
Liomys adspersus se encuentra en Panamá (Williams et al, 1993).
La distribución de los roedores pertenecientes a la subfamilia Heteromyinae

32
está fuertemente influenciada por factores como la humedad y la aridez, ninguna
especie se distribuye en zonas con menos de 250mm de precipitación anual y tienen
una marcada propensión ecológica por ambientes tropicales y sub-tropicales
(Hafner, 1993), Liomys es más tolerante a ambientes con mayor aridez (ej. selvas
secas) que Heteromys, quienes se encuentran en zonas menos áridas como selvas
medianas y altas (Schmidly et al., 1993).
Liomys irroratus se distribuye desde una porción al sur de Texas (EUA) hasta
el sur de Veracruz (México) siguiendo la vertiente del Golfo y en su distribución
occidental desde el centro norte de Chihuahua a partir de la parte norte de Jalisco en
todos los estados del centro de la República hasta llegar a Oaxaca (Figura 2)
habitando 20 estados de la República Mexicana (Ceballos y Galindo, 1984; Dowler &
Genoways, 1978; Schmidly et al., 1993; Villa & Cervantes, 2003). Se le encuentra
generalmente en selvas secas o matorrales espinosos (Sánchez-Cordero & Fleming,
1993; Schmidly et al., 1993). En zonas donde es simpátrico con L. pictus se ha
observado que este último prefiere hábitat más húmedos en comparación con los
hábitat xéricos preferidos por L. irroratus.
Junto con Perognathus flavus son las únicas especies de heterómidos
presentes en el estado de Morelos, pero P. flavus sólo esta registrado para la zona
norte del estado (Alvarez-Castañeda, 1996).

33
Figura 2. Mapa de la distribución de Liomys irroratus. (Villa & Cervantes, 2003).
7.2. Características generales.
Las características generales de este ratón son (Figura 3): pelaje hirsuto, formado
de pelos duros rodeados de pelos más suaves; las partes superiores son de color
gris oscuro con tonos cafés, los pelos rígidos son de color claro, cuentan con una
línea lateral, usualmente presente, de color rosa claro a beige, las partes inferiores
son blancas (Ceballos y Galindo, 1984; Dowler & Genoways, 1978). El pelaje del
juvenil es más gris y consiste de pelos más suaves (Dowler & Genoways, 1978). La
cola es bicolor con punta pincelada, las patas traseras tienen cinco tubérculos
plantares; la garra del segundo dedo de las patas traseras tiene forma de espolón,
diseñada para la huida, los últimos molares son más estrechos que los premolares.
Los huesos pterigoideos tienen un ala ancha, en forma de “U”, las bulas timpánicas

34
son aplanadas, sus incisivos superiores no tienen surcos y el báculo es oval y ancho
en la base terminando en una punta afilada, ligeramente dirigida hacia arriba y un
poco comprimida dorso-ventralmente. La fórmula dentaria de esta especie es la
misma de todos los heterómidos: i1/1, c0/0, p 1/1, m 3/3 para un total de 20. El
número de cromosomas diploides (2n) de este ratón es de 60, con un número
fundamental (FN) de 62 (Dowler & Genoways), algunos trabajos sobre aspectos
genéticos de esta especies son: Cervantes et al., (1999) Patton & Rogers (1993a),
Patton & Rogers (1993b) y Rogers (1990).
Figura 3. Fotografía de Liomys irroratus.
El dimorfismo sexual de esta especie es notorio, el macho es de mayor talla
que la hembra (Ceballos y Galindo, 1984; Dowler & Genoways, 1978; Sánchez &
Romero, 1995). Las medidas (en milímetros, primero se da el promedio y después el
rango, las medidas de machos preceden a las hembras) para esta especie (largo
total (LT) largo de cola (LC) Longitud de pata trasera (LPT)) son: LT: 238 ± 5.18mm

35
(216-262mm); 226 ± 3.14mm (207-251mm). LC: 120.1 ± 3.31mm (106-138mm);
112.5 ± 1.98mm (102-131mm). LPT: 29.2 ± 0.49mm (26-31mm); 28.2 ± 0.27mm (27-
30mm). Éstas y más medidas se pueden consultar en la revisión de la especie
(Dowler & Genoways, 1978). El peso de este ratón varía entre los 34-50 gramos
(Ceballos y Galindo, 1984). Best (1993) señala que la mayor diferencia se da en
medidas anatómicas no craneales, sólo encontrando cuatro medidas que fueron
significativamente diferentes, no siendo tan notorio el dimorfismo como antes se
señalaba, sólo en cuatro medidas, no en siete como señalan Dowler & Genoways
(1978).
7.3. Biología.
Tiene actividad reproductiva durante casi todo el año, aunque el pico
reproductivo parece ocurrir de agosto a noviembre (Dowler & Genoways, 1978).
Álvarez-Castañeda (1996) reporta para Morelos la presencia de hembras preñadas
en mayo y junio, así como lactantes de marzo a junio. Sánchez y Romero (1995)
reportan la mayor actividad reproductiva de machos de esta especie (en El Limón,
dentro de la Reserva de la Biosfera Sierra de Huautla) en julio y agosto, mientras
García-Estrada et al., (2002) reportan, para El Limón también, que la mayoría de las
crias y juveniles los capturaron en noviembre. Aunque en general se reporta que los
heterómidos tropicales tienen períodos reproductivos más prolongados que los de
los heterómidos de zonas desérticas, Sánchez-Cordero y Fleming (1983) reportan
que, varias especies de Liomys (entre las que no se tomó en cuenta a L. irroratus)
tiene un periodo reproductivo restringido estacionalmente y sobre todo a la

36
temporada seca y el periodo inicial de las lluvias.
En los mismos sitios de estudio de este proyecto, la actividad reproductiva de
L. irroratus se encuentra restringida por el régimen de lluvias, Cadena (2003)
encontró individuos reproductivos principalmente en la época de lluvias (de agosto a
noviembre), aunque reporta algunos individuos reproductivos en diciembre, enero,
mayo y junio.
Su ciclo reproductivo es generalmente monoéstrico y dura entre 21 y 28 días,
en cada parto nacen de dos a ocho crías y el promedio es de cuatro (Ceballos y
Galindo, 1984; Ceballos & Miranda, 2000; Dowler & Genoways, 1978).
Los heterómidos en general tienen un metabolismo del agua y una regulación
de su temperatura altamente eficiente, basada en un metabolismo basal bajo
(French, 1993), una actividad renal muy eficiente, la producción de excretas secas,
un bajo índice de pérdida de agua por evaporación y lactancia así como actividad
nocturna. Esto le permite a este grupo sobrevivir en medios secos y cálidos. En el
caso de Liomys estas adaptaciones están menos desarrolladas que en el resto del
grupo (Brylski, 1993; French, 1993; Hafner, 1993; Hudson & Rummel, 1966) ya que
no presenta, por ejemplo, ciclos de torpor comunes en otros miembros de su familia
(French, 1993).
El plano corporal de la subfamilia Heteromyinae es considerado como una
aproximación a la condición ancestral del resto de la familia Heteromyidae y
Geomyidae (Hafner, 1993).

37
7.4. Ecología.
Al igual que la mayoría de los roedores, tiene un lugar fundamental en la base
de diversas redes tróficas siendo un consumidor primario, especialmente de
semillas, por sus hábitos granívoros es un importante dispersor y depredador de
semillas (Ceballos y Galindo, 1984; Ceballos y Miranda, 2000; Dhillion, 1999; Huntly
& Reichman, 1994; MacCracken et al., 1985; McDonald, 2001; Villa y Cervantes,
2003). Es depredado por diversas especies de mamíferos, reptiles y aves (Ceballos
y Galindo, 1984; MacCracken et al., 1985; Sánchez-Cordero & Fleming, 1993; Villa y
Cervantes, 2003).
Su estructura social es compleja, es un ratón, igual que los demás miembros
de su familia, muy territorial que habitualmente tiene zonas de influencia bien
definidas por individuo (Ceballos & Miranda, 2000; Jones, 1993; Sánchez & Romero,
1995; Vázquez, 1997) y que construye madrigueras comunales clasificadas en muy
diversas formas y funciones (Domínguez, 2000). Aparentemente la distribución de
algunas especies está restringida por la presencia de otra especie de Liomys
(Schmidly et al., 1993).
Se ha argumentado que este ratón puede llegar a convertirse en una plaga
para la agricultura debido principalmente a la ausencia de depredadores y a sus
hábitos granívoros (Ceballos y Galindo, 1984). Sin embargo, la evidencia recopilada
por Cadena (2003) en la misma zona de muestreo en la que se realizó el presente
estudio, sugiere que es una especie que se ve afectada por la perturbación natural,
en este estudio se encontraron densidades bajas en hábitat previamente señalados
como muy utilizados por la especie (e. g. bardas de piedra y cercanías de campos

38
de cultivo) según Davis & Russell (1954), sembradíos y bordes (Mellink, 1995).
García-Estrada et al., (2002), éste último trabajo se realizó también en la Sierra de
Huautla.
7.5. Dieta.
Como fue señalado anteriormente (ver sección 4) la mayoría de los trabajos
de dieta de heterómidos han sido centrados en especies de zonas desérticas por lo
que existe muy poca información sobre la dieta de las especies tropicales
(Heteromyinae), aunque se ha observado que son principalmente granívoras y que
consumen, en general, semillas de mayor tamaño que las especies de zonas
desérticas (Reichman & Price, 1993). Se reporta que los heterómidos tropicales
pueden remover una gran proporción de las semillas de las sombras de semillas,
sobre todo en las selvas secas y que son importantes depredadores de semillas
post-dispersión (Howe & Smallwood, 1982; Janzen, 1986a; Sánchez-Cordero &
Fleming, 1993).
Los heterómidos tropicales consumen frutos y semillas encontrados en el
suelo pero su dieta también incluye otras partes de material vegetal y artrópodos
(Sánchez-Cordero & Fleming, 1993). Sin duda, su comportamiento de remoción de
semillas y almacenamiento de las mismas, tiene y ha tenido un papel preponderante
en la demografía de las plantas de bosques tropicales (Fleming, 1983a, Fleming,
1983b, Sánchez-Cordero & Fleming, 1993).
La evidencia sugiere (aunque es muy variable) que la dieta de los
heterómidos refleja la disponibilidad de recursos en un momento determinado y que

39
sus dietas varían substancialmente entre estaciones y años en la proporción total de
semillas, material vegetal e insectos, esencialmente de acuerdo a los patrones
temporales de abundancia de estas categorías (Reichman, 1975; Reichman y Price,
1993).
El consumo de una especie particular de semilla puede cambiar abruptamente
dependiendo de su abundancia y la de especies con diferente palatabilidad
(Reichman, 1975). En general los heterómidos pueden modificar la selección de
semillas en respuesta a cambios ambientales y a sus requerimientos nutricionales
(Reichman, 1975; Reichman y Price, 1993) o reproductivos (Reichman & Van de
Graaff, 1975). Por medio del almacenamiento de semillas, tanto en madrigueras
como en almacenes externos (Shaw, 1934), puede mitigar la escasez de estas
durante períodos estacionales (Sánchez-Cordero & Fleming, 1993). Reichman & Van
de Graaff (1975) reportan (para heterómidos de zonas desérticas) que al parecer
consumen plantas frescas en la temporada reproductiva.
Dado que no se ha hecho un estudio completo de la ecología de esta especie
es poca la información disponible sobre la dieta de Liomys irroratus. Se ha reportado
que se alimenta de una gran variedad de semillas y de hojas de plantas silvestres
(Villa y Cervantes, 2003). Sin embargo, se tiene mucha información sobre su especie
hermana (Cervantes et al., 1999) L. pictus que también habita en selvas secas y
dado que los heterómidos tienen ecologías parecidas (Rosenzweig & Sterner, 1970)
se puede suponer que L. irroratus se comportará de forma muy parecida. Villa y
Cervantes (2003) señalan que la cantidad de plantas y artrópodos en la dieta de
Liomys irroratus parece variar de acuerdo a las poblaciones, lo que sugiere su
carácter oportunista.

40
Hernández (2000) encontró en sus madrigueras semillas de 155 especies,
equivalentes al 23% de las plantas reportadas para la zona (Lott, 1985).
Predominaban las semillas de tres familias: Fabaceae, Convolvulaceae y
Euphorbiaceae, de especies arbóreas y lianas. Cuatro especies representaron el
80% de la biomasa total de semillas encontrada en las madrigueras: Spondias
purpurea, Jatropha sp., Cordia sp. y Serjania brachicarpa. Del total de géneros y
especies registradas en las madrigueras, 30 géneros (entre ellos 12 especies)
también están presentes en la Sierra de Huautla. En el mismo estudio se
encontraron 177 especies de semillas en muestras de suelo y la diversidad de estas
no difirió entre estaciones; un dato importante es que algunas de las semillas más
abundantes en las madrigueras no eran las semillas más abundantes encontradas
en el suelo.
Mandujano (2002) reporta a L. pictus como consumidor de frutos de Spondias
purpurea. Bullock (2002) reporta a este roedor como el depredador más importante
para las semillas de la especie Cordia eleagnoides, una de las especies arbóreas
más conspícuas y de gran importancia en las selvas secas del occidente del país.
Mendoza (1997) reportó que para este mismo roedor, de un total de 10,300
capturas, el 9.4% de los ratones tenían contenido en sus abazones y este consistió
en un 98% de semillas y apenas el 1.2% de restos de insectos y moluscos.
En dicho estudio se encontraron semillas de 144 diferentes especies en los
abazones, de las que sólo se pudo identificar 77 y de estas, el 60% pertenecían a
las siguientes familias: Fabaceae, Euphorbiaceae, Sapindaceae, Boraginaceae y
Convolvulaceae. Sin embargo, la especie que más semillas acumuló fue el pasto
Panicum sp. (17% del total de semillas), seguida de Lonchocarpus lanceolatus (7%)

41
e Ipomea sp. (5%). De las 77 especies identificadas, sólo se pudo obtener
información sobre la forma de vida para 44 especies, que sin embargo representaron
el 75% de las semillas encontradas en los abazones. La mayor parte fueron
especies que son árboles o lianas, es decir, estas formas de vida son las que
podrían verse más afectadas por la actividad de este ratón (Mendoza, 1997).
El número total de especies y el número total de semillas en los abazones
acumuladas por año varió entre años (Mendoza, 1997). El número de semillas por
muestra de abazón fluctuó entre una y 209, con un promedio de cuatro semillas por
muestra, con los valores mayores para la temporadas de secas temprana y los
valores menores para la temporada de lluvias (Mendoza, 1997).
Ceballos (1989) logró recolectar 46 especies de semillas en abazones en
selva seca y bosque de arroyo de Jalisco, encontró que las familias más frecuentes
fueron: Fabaceae, Euphorbiaceae y Convolvulaceae, siendo Phaseolus microcarpus,
Cyclanthera multifoliolata, Ipomoea spp., y Recchia mexicana las especies más
abundantes.
En la Sierra de Huautla se encuentran presentes varios de los géneros y
especies que se han reportado en abazones de otras especies del género Liomys,
(Carter & Genoways, 1978; Ceballos, 1989; Janzen, 1986a; Pérez-Saldaña, 1978;
Sánchez-Cordero & Fleming, 1993) como: Acacia farnesiana, Ceiba aesculifolia,
Cissus sp., Cochlospermum vitifolium, Crescentia alata, Enterolobium ciclocarpum,
Ficus sp., Ipomoea arborescens, Ipomoea sp., Lasiacis ruscifolia, Lonchocarpus sp.,
Lysiloma divaricata, Mimosa spp., Phaseolus lunatus, Phaseolus sp., Pithecellobium
sp., Serjania brachycarpa, Sida sp. y Tephrosia sp.
Un listado más completo de se encuentra en la Tabla 8 del presente trabajo.

42
Los ratones de la familia Heteromyidae son, junto con las hormigas, los
granívoros más importantes de los desiertos y hábitats áridos de norteamérica
(Brown et al., 1979) y han sido ampliamente estudiados en cuanto a su selectividad
alimentaria y su importancia como depredadores y dispersores de semillas (Briones
& Sánchez-Cordero, 1999; Brown et al., 1979; Fleming, 1983a; Fleming, 1983b;
Janzen; 1971; Janzen, 1986a; Reichman & Van de Graaf, 1975; Rosenzweig &
Sterner; 1970; Shaw, 1934; Smigel & Rosenzweig, 1974).
Los heterómidos tienen una función clave en las interacciones de las redes
tróficas, la dispersión, la depredación de semillas y en la estructura demográfica de
las plantas de los habitats donde se encuentran, sean selvas tropicales (Fleming,
1983a; Fleming, 1983b; Janzen, 1971; Janzen; 1986; Janzen & Martin, 1982;
Sánchez-Cordero & Fleming, 1993) o ambientes semi desérticos y desérticos (Brown
& Harney, 1993; Reichman & Price, 1993; Mares, 1993).
Janzen (1986a) indica que Liomys salvini consume casi la totalidad de las
semillas de Enterolobium ciclocarpum dispersadas por medio de las excretas de
mamíferos grandes.
Fleming (1983a y 1983b) señala que Heteromys desmarestianus y Liomys
salvini son elementos clave para las selvas medianas y secas, respectivamente, de
Costa Rica.

43
8. Sitio de Estudio
El estudio se realizó en la región sur del estado de Morelos, en sitios de selva
seca dentro de la Reserva de la Biosfera Sierra de Huautla y en su área de
influencia (figura 4).
Figura 4. Mapa de la ubicación de la sierra de Huautla.
Esta reserva limita al sur con los estados de Puebla y Guerrero y tiene una
superficie total de 59,310.65 hectáreas ubicadas entre altitudes de los 700 a los
2,400msnm (Dorado et al. En prensa). La mayor parte de dicha extensión protege
selva seca (Bosque Tropical Deciduo según la clasificación de Rzedowski, 1978 o
Dorado et al. En prensa.

44
Selva Baja Caducifolia según la de Miranda y Hernández, 1963). Dentro de la
reserva coexisten 29 comunidades de cinco municipios de Morelos (Amacuzac,
Puente de Ixtla, Jojutla, Tlaquiltenango y Tepalcingo). Con cerca de 20,000
habitantes, dedicados en su mayoría a actividades agropecuarias, como la cosecha
de maíz, frijol, calabaza, chile y cacahuate y la cría de ganado vacuno, caprino,
porcino y equino. Los principales tipos de suelo de la zona son: feozem háplico,
regosol éutrico y en los cuerpos montañosos se encuentran litosoles (Dorado et al.,
en prensa).
El clima en general es Awo"(w) (i’) g, correspondiente a un clima cálido, el
más seco de los subhúmedos, con régimen de lluvias de verano (junio a octubre, con
picos entre julio y septiembre) la precipitación media anual es de aproximadamente
900 mm y presenta canícula (Dorado et al., en prensa).
Para el periodo de 1980 a 2001, la precipitación media anual fue de 851 mm y
la temperatura media mensual oscilo entre 22.5o en diciembre y 26.6oC para mayo
(datos de la estación meteorológica de Huautla, Comisión Nacional del Agua).
A la fecha se han registrado en esta región un total de 937 especies de
plantas vasculares, incluidas en 449 géneros y 130 familias. Las familias más
abundantes en cuanto a número de especies son: Fabaceae, Poaceae, Asteraceae
y Burseraceae (Dorado et al., en prensa).
Se han registrado más de 350 especies de vertebrados (Dorado et al., en
prensa): ocho de peces, 11 de anfibios, 52 de reptiles, 220 de aves y en el caso de
los mamíferos se han reportado 66 especies (de 18 familias) que representan el 47%
de las especies registradas en el estado de Morelos; de éstas, 12 son roedores: dos
de las cuales son ardillas (Sciuridae) y 10 ratones, nueve de ellas pertenecen a la

45
familia Muridae: Baiomys musculus, Hodomys alleni, Reithrodontomys fulvescens
(recolectado por primera vez en la zona por Cadena (2003) y el presente estudio),
Reithrodontomys megalotis, Oligoryzomys fulvescens, Oryzomys couesi (probable,
no recolectado), Peromyscus levipes, Peromyscus melanophrys y Sigmodon
hispidus., y sólo un representante de la familia Heteromyidae: Liomys irroratus
(Sánchez y Romero, 1995).
A pesar de su protección, la selva seca de la Sierra de Huautla sufre de
amenazas tales como la tala de árboles, para venta o uso de la madera o leña; la
cacería de subsistencia y la deforestación de muchas áreas para la creación de
potreros y campos de cultivos (Dorado et al., en prensa). Esto ha ocasionado que
en el total de la superficie de la reserva que aún presenta selva seca (60%),
aproximadamente una tercera parte este cubierta por selva seca perturbada y que
del restante 40% de la reserva, la vegetación natural haya sido transformada en un
36% por las actividades humanas, en particular las agropecuarias (Dorado et al., en
prensa).
En un estudio realizado en la Sierra de Huautla se encontró que las familias
más abundantes que en el banco de semillas eran Fabaceae y Burseraceae,
(considerando sólo las especies arbóreas) tanto por el número de especies como por
el número de individuos (Arias, 1986). La cantidad (tanto de especies como de
semillas) y composición de especies del banco de semillas varía mucho a lo largo
del año, pudiendo prolongarse más allá de la temporada de fructificación de las
especies (Arias, 1986).

46
9. Metodología
9.1. Delimitación de períodos estacionales.
Para propósitos del análisis de los resultados y considerando que el factor
ambiental más notable en la zona es la estacionalidad climática, se determinaron
tres temporadas de análisis, con base en la distribución de la precipitación y su
efecto en el ambiente: a) temporada de lluvias de junio-octubre del 2002, meses en
los que hay agua disponible todo el tiempo y todas las plantas tienen follaje; b) la
temporada de transición entre la época lluviosa y los meses más secos del año, seca
temprana de noviembre del 2002-febrero del 2003, en la que ya no hay agua
disponible y la humedad en el suelo y el follaje de los árboles va disminuyendo
paulatinamente y; c) la temporada de mayor sequía o seca tardía de marzo-mayo del
2003, en la que la humedad del suelo se ha perdido, todas las plantas perdieron su
follaje y prácticamente no hay agua disponible.
9.2. Elección de los sitios y determinación del grado de perturbación.
La elección y determinación del grado de perturbación de los sitios de trabajo
se realizó con base en el mapa de vegetación de la Reserva de la Biosfera Sierra de
Huautla elaborado por el Laboratorio de Sistemas de Información Geográfica del
CEAMISH (Figura 5).
En dicho mapa se seleccionaron cuadros de 1 km2 (Figura 5) en áreas de la
reserva con la mayor cobertura de selva continua posible y áreas en donde fuera
evidente que la vegetación fue afectada o transformada. En total se seleccionaron

47
seis sitios pareados (un sitio poco perturbado y otro bastante perturbado) de forma
tal que se minimizarán los problemas logísticos para su estudio y que tuvieran el
mayor contraste posible en términos de su cobertura (Figura 6). Se seleccionaron
por pares para tratar de controlar el efecto que otras variables no evaluadas (e.g.
microcuenca, tipo de suelo) pudieran tener sobre el contraste entre un sitio no
perturbado y uno perturbado.
Se consideraron como sitios no perturbados aquellos con cobertura
continua y conservada de selva seca y una mínima presencia de campos de cultivo o
de tipos de vegetación secundaria en las cercanías, los sitios perturbados fueron
seleccionados en zonas donde la cobertura estuviera alterada y que tuvieran una
mayor cercanía con campos de cultivo, vegetación secundaria y mayor presencia de
ganado.
Una vez seleccionados los sitios en el mapa, se procedió a corroborar en
campo la condición de cada sitio con la obtención de datos sobre las características
de: a) la cubierta arbórea (e. g. diversidad, tamaño) y si había o no un estrato
arbustivo; b) la presencia de especies características de sitios poco perturbados (ej.
Conzattia multiflora; Crescentia alata; Pachycereus sp.; Cissus sp.; Rzedowski,
1978; Miranda & Hernandez, 1963) y, c) que hubiera baja intensidad de pastoreo y
baja evidencia de tala y agricultura.
Los tres pares de sitios fueron ubicados como indican la Figura 5 y la Figura 6.
La nomenclatura, condición y coordenadas geográficas donde se ubicaron los
cuadrantes se encuentran en la Tabla 1.

48
Figura 5. Ubicación de los cuadrantes de ratones (100Ha c/u)
46,9
96,7
25,9
88,6
15,13
38,75
0%
20%
40%
60%
80%
100%
XP XNP
AX
VIRG
EN QP
QNP
Sitios
Por
cent
aje de
tipo
de ve
getacióo
n
Erosion
Vegetación de Galeria
Cuerpos de Agua
Infraestructura
Pastizal
Veg. Sec.
Agricultura
sbc pertSelva Baja
Figura 6. Porcentajes de cobertura en los sitios de estudio (100 ha).
Quilamula
Qnp Qp
Vgn
Ajn Xnp
Xp
Laboratorio de Sistemas de Información Geográfica, CEAMISH.
Laboratorio de Sistemas de Información Geográfica, CEAMISH.

49
9.3. Metodología de trampeo. Dentro de cada sitio de muestreo se ubicó una gradilla 70x70 m cada una,
con estaciones de trampeo separadas entre sí cada 10 m. En cada gradilla se
colocaron un total de 88 trampas Sherman, 64 de ellas en el suelo y 24 en los
árboles o lianas aproximadamente a 1.5 m de altura, en las estaciones pares (Figura
7), estas últimas trampas se colocaron para capturar especies arborícolas en
conjunto con el trabajo de Cadena (2003).
Figura 7. Diseño de los sitios de trampeo. Cada punto es la ubicación de una trampa, los puntos rellenos señalan donde se pusieron las trampas en árboles y lianas.
En total cada gradilla estuvo compuesta por 88 trampas Sherman y un área
de muestreo de 4,900m2. Cada gradilla se operó tres noches consecutivas al mes,
durante 11 muestreos, de agosto de 2002 a mayo de 2003. Las trampas fueron
7
0
1
2
3
4
5
6
A B C D E F G H

50
cebadas con una mezcla de avena, mantequilla de cacahuate y extracto de vainilla,
siguiendo a Romero-Almaraz et al., (2000). Las medidas y datos que se les tomaron
a los ratones capturados están detallados en el Anexo 1.
Específicamente, por haber realizado el marcaje de los animales por medio de
ectomización de falanges (por si misma estresante) se trato de disminuir (en lo
posible) el estrés en la manipulación y el mismo marcaje de los roedores, siguiendo
las recomendaciones pertinentes hechas por diversos autores (Animal Care and Use
comittee, 1998; Jones et al., 1996; The Association for the Study of Animal
Behaviour, 2000; Rudran & Kunz, 1996).
9.4. Estimación de la diversidad y disponibilidad de recursos
alimentarios.
La preferencia de un consumidor para un componente particular (ej. alimento)
es el reflejo de la posibilidad de que dicho componente sea escogido si se oferta en
la misma proporción que todos los elementos (Chesson, 1983; Johnson, 1980). Para
poder determinar la preferencia sobre algún elemento de su dieta es necesario saber
la disponibilidad que tiene en el ambiente (Norbury & Sanson, 1992). Para calcular la
preferencia en campo, donde la disponibilidad no es la misma para los recursos, ésta
se calcula en valores de preferencia relativos. (Norbury & Sanson, 1992).
Muestreo de herbáceas.
Para obtener la información sobre la disponibilidad y la diversidad de los
recursos alimentarios de Liomys irroratus se diseñó un muestreo de cuadrantes para
el que se ubicaron de manera azarosa 10 cuadrantes de 1 m2 en cada uno de los

51
sitios de trampeo por cada temporada (Figura 8). En cada cuadrante se cosechó
toda la biomasa vegetal viva de tallos no leñosos y que midiese menos de 1.5 m de
alto que naciera dentro del cuadrante, el material se separó en campo en
morfoespecies (con base en diferencias en su morfología), para cada morfoespecie
se herborizaron ejemplares para su identificación.
0.25m2
1m2
0.25m2
1m2
Figura 8. Cuadrante de vegetación, el cuadrante de 0.25m2 fue utilizado
para muestraear el banco de semillas.
Del material recolectado se obtuvieron; su abundancia, frecuencia de
ocurrencia, biomasa en fresco y en seco (secando las muestras entre 60-70° C
hasta alcanzar un peso constante), todo para cada sitio en las tres temporadas ya
mencionadas.
Muestreo del banco de semillas.
En una sección de 0.25 m2 de cada cuadrante de 1 m2 (Figura 8), se

52
recolectó todo el suelo y hojarasca disponible (aproximadamente los primeros 5-7
cm) en bolsas de plástico que se etiquetaron con los datos respectivos. Estas
muestras fueron cribadas en el laboratorio, con mallas edafológicas de 1mm,
separando las semillas encontradas para determinar la disponibilidad y diversidad de
especies en el banco de semillas.
Las semillas encontradas fueron determinadas a nivel de especie, cuando fue
posible, consultando a especialistas y pobladores de la zona (sus nombres están
incluidos en los agradecimientos). También se comparó el material con colecciones
privadas de semillas y directamente con las plantas, que en ese momento tuvieran
semillas, en la zona de estudio. De cada muestra se contabilizó el número de
semillas, la biomasa (gr) por especie, el promedio de especies, de semillas
(individuos) y biomasa. Las especies que no pudieron ser identificadas se
agregaron al análisis como morfoespecies
9.5. Obtención y preparación del catálogo de referencia de los
caracteres anatómico foliares de las herbáceas disponibles para Liomys
irroratus.
Para la identificación de los fragmentos microscópicos de material vegetal en
las excretas, fue necesario generar un catálogo de referencia de estructuras
anatómico foliares de las herbáceas encontradas en los muestreos de vegetación y
que potencialmente podrían ser consumidas por los herbívoros (Baumgartner &
Martin, 1939; Dusi, 1949; Stewart, 1967). Para ello se hicieron preparaciones
microscópicas de las plantas, siguiendo las recomendaciones de Sandoval-Zapotitla

53
(en prensa). Para hacer las preparaciones es necesario tener material vegetal en
fresco o fijado utilizando formol-acético-alcohol, FAA, (Anexo 2) posteriormente se
prepararon laminillas siguiendo dos técnicas distintas: montadas con gelatina
glicerinada (fórmula en el Anexo 3 y técnica en Anexo 4) para un montaje temporal
con una duración de dos a tres años o por medio de un montaje permanente, con
tinción de Safranina (Anexo 5).
A partir de las laminillas se generó un catálogo de fotografías de estructuras
anatómicas de las especies recolectadas durante los muestreos de vegetación para
permitir la identificación (en las preparaciones microscópicas de las excretas) de
dichas estructuras y por tanto de la composición de la dieta de este roedor.
La nomenclatura e identificación de las estructuras anatómicas de las plantas
se logró con el apoyo de botánicos (incluidos en los agradecimientos) y con el uso
de bibliografía especializada para monocotiledóneas (Ellis, 1979; Johnson et al.,
1983; Sandoval-Zapotitla, en prensa) y dicotiledóneas (Johnson et al., 1983;
Sandoval-Zapotitla, en prensa).
9.6. Análisis del contenido de abazones.
Todos los abazones de los ratones capturados (capturas y recapturas) fueron
revisados visual y tactilmente en campo y todos los contenidos encontrados dentro
de ellos fueron recolectados, etiquetados y guardados para su análisis en el
laboratorio.
Éstos contenidos fueron identificados por medio de la comparación directa
con semillas encontradas en los muestreos de banco de semillas para tener un

54
catálogo de semillas unificado, en los casos en los que fue necesario para su
determinación se consultó a especialistas, pobladores de la zona de muestreo,
colecciones tanto privadas como públicas de semillas (mencionadas en los
agradecimientos) y se recolectaron posteriormente las plantas que las producen.
Para poder estimar la frecuencia relativa de cada especie de semilla en la
dieta, la diversidad de especies, la equidad, el valor de importancia y el análisis de
similitud del número relativo de individuos, selectividad y preferencia (todos por
condición por temporada) Se tomaron datos del material identificado (todos ellos por
condición y temporada) como: número de especies por muestra, número de
muestras, biomasa (gr) por semilla por muestra, promedio de especies, individuos,
biomasa y medida (mm) promedio por muestra .
9.7. Análisis de las excretas.
Las muestras de excretas se procesaron siguiendo la técnica (Anexo 6)
sugerida por Holechek (1982). Esta técnica es similar a la usada para la preparación
del catálogo de referencia, es más sencilla y rápida que otras (ej. Sparks &
Malechek, 1968; Williams, 1969) y ha sido ampliamente utilizada (Holechek & Gross,
1982a; Holechek et al., 1982a; Holechek et al., 1982b; Vavra & Holechek, 1980).
Se montaron y analizaron tres laminillas (cada laminilla compuesta por tres
pellets de una sola muestra, osea un individuo) por sitio y por temporada (para un
total de 54 laminillas analizadas). Cada laminilla se dividió en 20 campos, elegidos al
azar, siguiendo a Holechek & Vavra (1981) que reportan que éste es el número
mínimo de observaciones por laminilla para asegurar la representatividad de cada

55
muestra y permitir la comparación entre laminillas. La observación y conteo se hizo a
20X, aumentando a 40X o 100X cuando se requirío para facilitar la identificación de
un fragmento particular. Se siguió la recomendación de Holechek & Gross (1982b)
moviendo las laminillas al revisarlas para facilitar la identificación de fragmentos.
En cada laminilla analizada se determinó la frecuencia relativa de aparición de
cada fragmento diferente encontrado (Holechek, 1982; Sparks & Malechek, 1968). El
material se identificó a nivel específico hasta donde fue posible, si no a nivel de
familia tomando en cuenta por separado el material no determinable, para saber la
proporción de fragmentos que pudieron ser definidos.
Para conocer diferentes variables de la dieta se calcularon, por condición por
temporada, la diversidad, la equidad, la frecuencia relativa, la amplitud del nicho
trófico y un índice de similitud del número relativo de individuos.
La frecuencia de ocurrencia de cada especie en la dieta se estimó como el
número de campos microscópicos en los que se encontró esa especie, dividido entre
la suma total de ocurrencias en todos los campos analizados y multiplicado por 100.
Estoa valores se calcularon para cada condición y temporada.
Los fragmentos encontrados en las laminillas se determinaron por medio de la
comparación directa con las fotografías obtenidas del catálogo de referencia, las
principales características comparadas fueron:
1. La forma, alineación y tamaño de las células epidérmicas.
2. La forma y el grosor de las paredes anticlinales (perpendiculares al
mesodermo) y paradermales (paralelas al mesodermo) de las células epidérmicas.
3. La presencia o ausencia de tricomas y bases de tricomas, así como su
forma, grosor y características de las paredes, la cantidad y forma de sus células,

56
tanto de la proyección como de la base, en particular la célula Terminal.
4. La presencia de contenidos celulares (cristales de oxalato de calcio en
forma de drusas, rafidios, prismáticos, solitarios o agregados; sílice, especialmente
para monocotiledoneas; contenidos en idioblastos; contenidos de apariencias
resinosas, aceitosas, acuosas) tanto en tricomas como en células epidérmicas.
5. Refringencia a la luz polarizada, tanto de las paredes celulares como de los
contenidos.
9.8. Evaluación de la selectividad y la preferencia relativas.
Tanto la evaluación de la selectividad como de la preferencia de las especies de
semillas en abazones como las de herbáceas en excretas se analizaron de manera
relativa y no absoluta, siguiendo las sugerencias de Johnson (1980).
La selección de un componente de la dieta es el proceso por el cual el
consumidor elige dicho componente (Johnson, 1980) por lo que para este trabajo la
selectividad relativa (en adelante referida como selectividad) se determinó como
las especies que son seleccionadas dependiendo de su disponibilidad en el medio.
Una especie seleccionada es la que se consume por encima de su disponibilidad,
mientras que una especie utilizada puede ser consumida de manera accidental o por
debajo de su disponibilidad en el ambiente.
Para evaluar la selectividad de los diferentes recursos alimentarios en la dieta de
L. irroratus, se compararon los valores de las frecuencias de ocurrencia de las
diferentes categorías tróficas (especies) encontradas tanto en los contenidos de
abazones como en las excretas con los valores de disponibilidad de semillas y

57
herbáceas en los sitios de estudio utilizando el índice estandarizado de selección
de Manly-Gresson (Bi).
Éste índice es una estandarización del índice de selección (Krebs, 1999) de
forma en que la suma del Bi todos los recursos sea 1.0, utilizando la siguiente
fórmula:
Bi=ŵi / nΣi=1ŵi
Donde Bi= al índice estandarizado de selección para la especie i
ŵi= índice de selección para la especie i calculado como: ŵi=oi / pi.
Donde oi es la frecuencia relativa en que la especie i es consumida y pi es la
frecuencia de ocurrencia de la especie i en el medio, los valores superiores a 1.0
indican preferencia y valores menores a 1.0 indican aversión (Fagerstone et al.,
1981; Krebs, 1999; Reichman, 1975).
La estandarización del índice se llevó al cabo siguiendo las sugerencias de
Chesson (1978) y Manly et al., (1993, Citado de Krebs, 1999) para utilizar el índice
de modo que los radios estandarizados (los valores por medio de los cuales se
determina si una especie es o no preferida) se encuentren entre 0.0 y 1.0. Cuando
los radios estandarizados son iguales a 1/número de recursos indican que no existe
preferencia. Valores menores a éste indican que el recurso es relativamente evitado
y valores superiores indican que es seleccionado (Krebs, 1999).
La preferencia de un consumidor por una especie en particular es un reflejo
de la probabilidad de que este componente sea elegido si la disponibilidad en el
ambiente es igual en todas las especies (Johnson, 1980); por lo que la preferencia
relativa ( en adelante referida como preferencia) en este trabajo se define como la

58
posición de un componente de la dieta conforme a los demás, utilizando un método
por el cual la disponibilidad no varía dependiendo de los componentes incluidos en
el análisis.
La preferencia se determinó de acuerdo al índice de Johnson (tij) quien
señala que es importante que los índices de preferencia no sean dependientes de
las especies incluidas en ellos (pues índices como el de electibidad de Ivlev o el de
selectividad de Manly-Gresson tienen problemas al definir el orden de las especies
preferidas dependiendo de la inclusión o exclusión de especies al análisis) así como
de eliminar la probabilidad de que los resultados sean interpretados de forma
absoluta (Johnson, 1980).
El índice de Johnson (1980) se calcula con la siguiente fórmula:
tij =rij-sij
Donde tij es el índice de preferencia de la especie i para el consumidor j.
rij: el uso (consumo) de un componente i por un consumidor j.
sij: la disponibilidad de un componente i para un consumidor j.
A partir de los resultados de este índice se ordenan las especies
(componentes) de mayor a menor valor de tij para obtener una medida relativa del
orden en que son consumidos, siendo los componentes de valores más altos más
preferidos que los de valores menores.
Para ambos índices, (selectividad y preferencia) por requisito de las
metodologías, solo se incluyeron las especies que son consumidas y que tienen
valores de disponibilidad, las especies que no cumplen esto fueron excluidas del
análisis, dado que no es posible determinar ni la selectividad ni la preferencia de

59
especies para las cuales no se conoce la disponibilidad en el medio.
9.9. Análisis estadísticos.
Para calcular la eficiencia del trabajo de muestreo en todos los resultados
(herbáceas y semillas en el medio, así como en semillas en abazones y contenido de
excretas) se calcularon curvas de acumulación de especies (Spp observadas,
Chao1, Chao2, Jacknife1 y Jacknife2) que permiten observar que porcentaje del
total de especies se obtuvo en el muestreo, dependiendo de la tasa de
descubrimiento de nuevas especies. Éstas se calcularon con el programa
Biodiversity Pro (Mac Aleece et al., 1997).
Los análisis que se realizaron de: diversidad, Shannon (H´ log base 10),
Simpson (1/D), equidad (J´) y de similitud (distancias euclidianas, con anclaje
promedio mínimo). Todos fueron calculados con el programa Biodiversity Pro (Mac
Aleece et al., 1997).
El análisis de las diferencias en los valores de número de muestras, número
de especies (o morfoespecies), biomasa, frecuencia de ocurrencia, diversidad,
equidad y amplitud de nicho tanto para los datos de muestreo de ratones en campo,
herbáceas, semillas en suelo, contenido de abazones y dieta, fue realizado para la
condición de perturbación y la estacionalidad, por medio de un análisis de varianza
multivariado (MANOVA) de medidas repetidas con efecto de bloque (Von Ende,
1993). El diseño con bloques se realizó para disminuir el error experimental debido a
la heterogeneidad de los sitios. En los casos en los que la varianza no fuera
homogénea o que los datos tuvieran distribuciones no normales fueron normalizados

60
por medio de una conversión con Log base 10, para después analizarlos de nuevo.
Todos estos análisis fueron realizados con el programa JMP 3.2.2. (SAS institute
inc., 1997).
Para determinar la amplitud de nicho trófico de Liomys irroratus se utilizó la
fórmula de Levins (1968):
B= 1/ Σpi²
Donde pi es la sumatoria de los cuadrados de las frecuencias relativas de
cada categoría trófica (ej. especie de semilla o herbácea). Los valores de B están
entre 1 y el número de categorías tróficas (especies) consideradas. Estos valores se
calcularon por condición y temporada.
Para complementar los resultados de los índices de diversidad, equidad y
amplitud de nicho trófico y para tomar en cuenta la variación en la composición de
especies (la cual no es calculada por los índices anteriores) se calcularon:
1. El valor de importancia (Krebs, 1985): que es la sumatoria de la
abundancia, la biomasa y la densidad relativas de la especie i para conocer su
importancia ecológica comparativamente con las demás especies disponibles.
2. Y análisis de χ2 para determinar si la condición y la estacionalidad causan un
cambio en la composición de especies que L. irroratus transporta o consume (para lo
cual también se usó JMP (SAS institute inc., 1997).

61
10. Resultados
10.1. Resultados del trampeo de ratones.
El esfuerzo de trampeo (sensu Jones et al., 1996) fue de 9,124
noches/trampa reales. En este trabajo el esfuerzo de trampeo fue considerado como
el número de trampas colocadas por el número de noches operando, menos el
número de trampas encontradas inactivas (ej. cerradas sin ratón, abiertas sin cebo,
en las que se capturó otra especie) restando también el número de trampas en árbol
(no disponibles para Liomys irroratus).
Se capturaron 720 ratones (incluidas 400 recapturas), con un éxito de captura
del 7.9%. Las capturas fueron marginalmente más frecuentes (F(2,1)=135.214 p=0.06)
en los sitios no perturbados; (198 capturas y 266 recapturas contra 122 capturas y
134 recapturas en los perturbados).
10.2. Dinámica de la comunidad de herbáceas.
En los muestreos de vegetación se recolectaron 38 morfoespecies de
herbáceas de las cuales 29 fueron identificadas a nivel especie, siete hasta género,
una a familia y una que no pudo ser identificada (tabla 2). Se recolectaron, de
manera no significativa, más especies en lluvias que en cualquiera de las
temporadas secas. Los datos muestran un ligero aumento del número de especies
en sitios no perturbados sobre sitios perturbados a lo largo de la temporada seca,
ésta tendencia tampoco fue significativa.

62
Las curvas de acumulación de especies indican que la representatividad del
muestreo fue de 68-97% de las especies calculadas para la zona de muestreo.
Se encontraron diferencias marginalmente significativas (F(1,2)=10.965 p=0.08)
en la biomasa a lo largo del año, existe una mayor biomasa en la época de lluvias
que en cualquiera de las temporadas secas. Los datos muestran una tendencia no
significativa a que en los sitios no perturbados exista más biomasa que los
perturbados.
El número promedio de plantas y de biomasa por m2 no fueron
significativamente diferentes por la estacionalidad o condición.
A pesar que la diversidad no difirió significativamente por condición o
temporada se pueden observar dos tendencias (Figura 9 y Figura 10): para el caso
de los sitios no perturbados se observa la disminución de la diversidad en el inicio de
la temporada seca y un importante aumento al final de la temporada seca. Por el
contrario, los sitios perturbados muestran un aumento en el inicio de la temporada
seca y una disminución al finalizar la temporada seca.
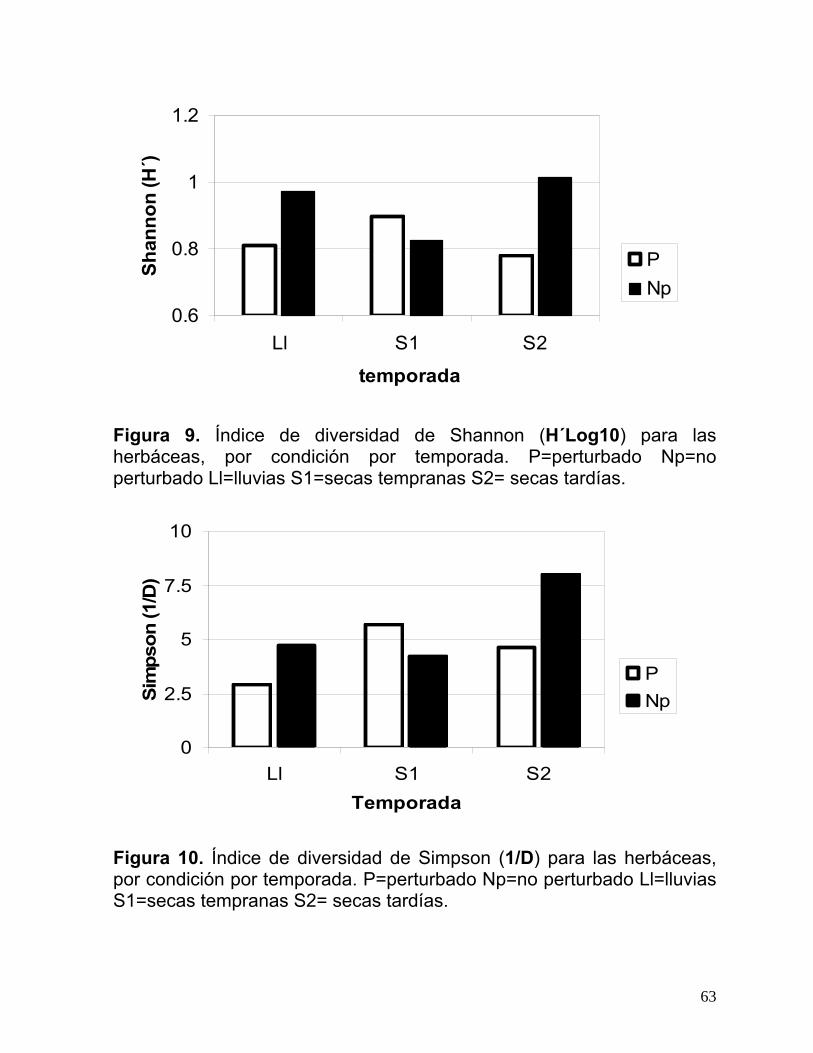
63
0.6
0.8
1
1.2
Ll S1 S2
temporada
Shan
non
(H´)
PNp
Figura 9. Índice de diversidad de Shannon (H´Log10) para las herbáceas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
0
2.5
5
7.5
10
Ll S1 S2Temporada
Sim
pson
(1/D
)
PNp
Figura 10. Índice de diversidad de Simpson (1/D) para las herbáceas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
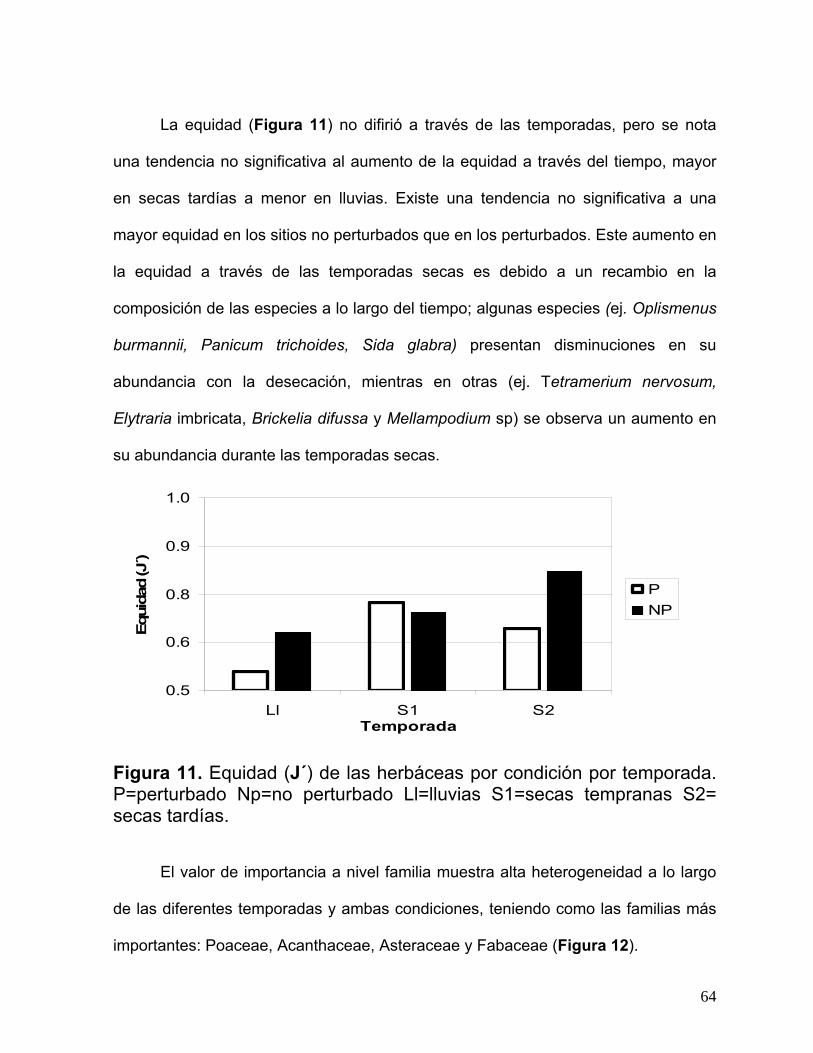
64
La equidad (Figura 11) no difirió a través de las temporadas, pero se nota
una tendencia no significativa al aumento de la equidad a través del tiempo, mayor
en secas tardías a menor en lluvias. Existe una tendencia no significativa a una
mayor equidad en los sitios no perturbados que en los perturbados. Este aumento en
la equidad a través de las temporadas secas es debido a un recambio en la
composición de las especies a lo largo del tiempo; algunas especies (ej. Oplismenus
burmannii, Panicum trichoides, Sida glabra) presentan disminuciones en su
abundancia con la desecación, mientras en otras (ej. Tetramerium nervosum,
Elytraria imbricata, Brickelia difussa y Mellampodium sp) se observa un aumento en
su abundancia durante las temporadas secas.
0.5
0.6
0.8
0.9
1.0
Ll S1 S2Temporada
Equi
dad
(J́)
PNP
Figura 11. Equidad (J´) de las herbáceas por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
El valor de importancia a nivel familia muestra alta heterogeneidad a lo largo
de las diferentes temporadas y ambas condiciones, teniendo como las familias más
importantes: Poaceae, Acanthaceae, Asteraceae y Fabaceae (Figura 12).

65
0
50
100
150
Aca
ntha
ceae
Am
aran
thac
ea
Ast
erac
eae
Con
volv
ulac
ea
Eup
horb
iace
ae
Faba
ceae
Lam
iace
ae
Mal
vace
ae
Mar
anta
ceae ND
Poa
ceae
Por
tula
cace
ae
Rub
iace
ae
Ste
rcul
iace
ae
Tilia
ceae
Familias
VI
PLlPS1PS2NpLlNpS1NpS2
Figura 11. Valores de importancia (VI) de las familias de herbáceas por condición por temporada. Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
0
40
80
120
Ely
traria
imbr
icat
a
Des
mod
ium
proc
umbe
ns
Sid
a gl
abra
Hab
raca
nthu
ssi
lvat
icus
Tetra
mer
ium
nerv
osum
Pan
icum
trich
oide
s/O
plis
men
usbu
rman
ii
Laga
scea
mol
lis
especies
VI
PLlPS1PS2NpLlNpS1NpS2
Figura 12. Valores de importancia (VI) de las especies ecológicamente más importantes por condición por temporada. Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.

66
Al igual que sucede a nivel familia existe un cambio en los valores de
importancia (Figura 12) de las especies a través del ciclo estacional y en las
diferentes condiciones. Las especies con valores de importancia más altos durante
las tres temporadas son, por orden de importancia de mayor a menor: Oplismenus
burmanii, Panicum trichoides, Tetramerium nervosum, Elytraria imbricata, Lagascea
mollis, Desmodium procumbens var. transversum y Sida glabra. Estas especies
constituyen el 82% de los ejemplares recolectados y el 69% de la biomasa total.
10.3. Dinámica del banco de semillas.
A partir del material obtenido de los muestreos de banco semillas se
determinaron 84 morfo-especies de semillas, de las cuales se lograron identificar 33
a nivel especie (pertenecientes a 25 familias), 20 a nivel género, 7 a nivel familia y
24 no pudieron ser identificas (Tabla 3). Las curvas de acumulación de especies
señalan que se recolectó entre el 88 y 95% de las especies esperadas para la zona
de muestreo.
El número de especies y la cantidad de semillas no variaron
significativamente a través del tiempo, pero muestran una tendencia a ser mayores
al final de la temporada seca que en el resto del año. Se encontraron tendencias no
significativas a mayor riqueza de especies de semillas en los sitios no perturbados y
a un mayor número de semillas en los sitios perturbados.
Se encontraron tendencias, no significativas, a menor diversidad y equidad
conforme avanza la época seca en los sitios perturbados y un aumento en los sitios
no perturbados (Figura 14 y Figura 15 para diversidad y Figura 16 para equidad).

67
0
0.5
1
1.5
Ll S1 S2
Temporada
Shan
non
(H´)
PNp
Figura 14. Índice de diversidad de Shannon (H´Log10) para el banco de semillas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
0
5
10
15
20
25
Ll S1 S2Temporada
Sim
pson
(1/D
)
PNp
Figura 15. Índice de diversidad de Simpson (1/D) para el banco de semillas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.

68
0.5
0.6
0.7
0.8
0.9
Ll S1 S2 temporada
Equi
dad
(J́)
PNp
Figura 16. Equidad (J´) del banco de semilas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
Existen cambios notorios en los valores de importancia de las diferentes
familias encontradas en las muestras de semillas del suelo. Acanthaceae,
Convolvulaceae, Fabaceae, Malpighiaceae y Sapindaceae representan el 49% del
valor de importancia total. Los valores de importancia de las familias varían mucho a
través de las temporadas, posiblemente por las marcadas épocas de fructificación de
algunas especies (Figura 17).
Existen cambios notorios en los valores de importancia de las diferentes
especies; 14 de ellas (Aeschynomene americana, Amphipterygium adstringens,
Combretum farinosum, Ficus sp., Justicia candicans, Ipomoea pauciflora, Ipomoea
sp.1, Ipomoea sp.2, Mascagnia aff. Polybotrya, morfo 19., Nissolia fructicosa,
Ruprechtia fusca, Serjania triquetra y Vitex hemsleyi) representan el 70% de los
valores de importancia de todas las especies recolectadas (Figura 18).

69
0
40
80
120
160
Acanth
acea
e
Anaca
rdiac
eae
Convo
lvulac
eae
Faba
ceae
Malphig
iacea
e
Morac
eae
ND
Familias
VI
PLlPS1PS2NpLlNpS1NpS2
Figura 17. Valores de importancia (VI) de las familias del banco de semillas por condición por temporada. Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
0
30
60
90
120
150
Ipomoe
a sp.
1
I. pau
ciflor
a
S. triqu
etra
Morfo
19
J. ca
ndica
ns
R. fusc
a
Ficus s
p.
A. ameri
cana
A. ads
tringe
ns
C. fruc
ticos
um
N. fruti
cosa
M. poly
botry
a
V. hem
sleyi
Ipomoe
a sp.
2
Especies
VI
PLlPS1PS2NpLlNpS1NpS2
Figura 18. Valores de importancia (VI) de las especies más importantes del banco de semillas por condición por temporada (nombres completos en Tabla 3). Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.

70
10.4. Resultados de las muestras del contenido de abazones.
Se recolectaron 102 muestras de contenido de abazones (14.16% del total de
ratones). El 64.6% de las muestras se recolectaron en sitios no perturbados y el
82.3% fueron recolectadas durante las dos temporadas secas.
El número de muestras difirió significativamente entre condiciones durante el
inicio de la temporada de lluvias y la parte final de las secas (F(2,1)=1663.48 p=0.017)
siendo el número de muestras más de 4 veces mayor, en promedio, en los sitios no
perturbados que en perturbados, ésto puede deberse a que en los sitios no
perturbados hay más ratones.
35.29% de los morfos de semillas se recolectaron exclusivamente en las dos
temporadas secas del año (ej. Pictecoctenium crucigerum, Ipomoea sp y Lysiloma
divaricata). El 30% de las morfoespecies se encontraron exclusivamente en sitios no
perturbados y el 21% es exclusivo de los sitios perturbados.
Se determinaron 34 morfos diferentes de semillas (26 de ellos se encontraron
en las muestras de suelo también), se pudieron identificar 13 de ellas a nivel
específico, ocho a nivel de género, 2 a nivel familia y 11 no pudieron ser
determinados (Tabla 4). Todas las identificadas fueron agrupadas en 14 familias.
Las curvas de acumulación de especies señalan que se encontraron entre el 43-68%
de las especies esperadas.
La riqueza de especies no difirió significativamente entre estaciones ni
condiciones, pero si existe una tendencia a un aumento en la riqueza en los sitios no
perturbados hacia el final de la temporada seca, mientras que los sitios perturbados
sufren una disminución, la diferencia es de 32% más especies en sitios no

71
perturbados que en perturbados.
En cuanto al número total de semillas recolectadas en las muestras de
abazones (341; promedio total de semillas por muestra de 1.71 semillas por
muestra) a pesar que no se encontraron diferencias significativas, sí existe una
tendencia que muestra una mayor cantidad de semillas en ambas épocas seca (300
semillas) en comparación con la época de lluvias (41 semillas). Y también una mayor
cantidad total de semillas recolectadas en las muestras de abazones (en promedio)
en los sitios no perturbados durante el final de las secas que en los sitios
perturbados (1.90 contra 0.90 de los perturbados).
El tamaño promedio (largo máximo de la semilla) por sitio de las semillas varió
de forma marginalmente significativa (F(1,2)=9.54 p=0.09) entre las dos condiciones
de perturbación, siendo ligeramente más grandes en los sitios no perturbados que
en los perturbados.
En cuanto a la diversidad y la equidad en semillas de las muestras de
abazones, los sitios no perturbados, muestran una tendencia (no significativa) a ser
más diversos y equitativos que los sitios perturbados. A través de las diferentes
temporadas se muestran dos tendencias (no significativas) dependientes de la
condición de los sitios, la diversidad y la equidad disminuyen de forma importante en
los sitios perturbados y hacia la temporada seca tardía, mientras que los sitios no
perturbados mantienen su diversidad y su equidad a lo largo del año (Figura 19,
Figura 20 y Figura 21 respectivamente).

72
0
0.4
0.8
1.2
Ll S1 S2
Temporada
Shan
non
(H´)
PNP
Figura 19. Índice de Shannon (H´) del contenido de abazones, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
0
2.5
5
7.5
10
Ll S1 S2Temporadas
Sim
pson
(1/D
)
PNp
Figura 20. Índice de Simpson (1/D) del contenido de abazones, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.

73
0.7
0.8
0.9
1.0
Ll S1 S2Temporadas
Equi
dad
(J´)
PNp
Figura 21. Equidad (J´) del contenido de abazones, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
Las familias más importantes, encontradas en abazones, son: Burseraceae,
Cochlospermataceae, Convolvulaceae, Fabaceae y Sapindaceae, las cuales
representan el 78.8% del valor de importancia total. Los valores de importancia de
estas familias cambian de manera notable a lo largo de las tres temporadas y en
ambas condiciones (Figura 22). Familias como Burseraceae, Caricaceae y
Cochlospermataceae son más importantes en los sitios no perturbados (Caricaceae
y Cochlospermataceae sólo se encuentran en sitios no perturbados) mientras
Sapindaceae y Apocynaceae tienen valores de importancia más altos en sitios
perturbados. Existen tres patrones estacionales delimitados de manera general: 1.
Familias con valores de importancia mayores en lluvias (Caricaceae, Sapindaceae y
ND). 2. Con valores de importancia estables durante el año (Bignoniaceae,
Convolvulaceae) y 3. Con incremento en sus valores de importancia durante las

74
temporadas secas (Apocynaceae, Burseraceae, Cochlospermataceae, Fabaceae y
Rutaceae).
Los valores de importancia a nivel de especies muestran el mismo patrón que
se observa a nivel familia (Figura 23). Existen especies que son transportadas de
manera más importante en sitios perturbados que en no perturbados (ej. Bursera
aloexylon, Lysiloma divaricata, Aeschynomene americana, Mimosa, sp., Ruprechtia
fusca, Senna sp., Stemmadenia bella). Otras con valores más altos en sitios no
perturbados que en los perturbados (ej. Cochlospermum vitifolium, Bursera
copallifera, Ipomoea pauciflora, Jacaratia mexicana, Justicia candicans).
Los patrones temporales para especies son parecidos a los encontrados para
familias: 1. Especies con valores de importancia mayores en la temporada de lluvias
(ej. Bursera copallifera, Jacaratia mexicana, Serjania triquetra.) 2. Especies que
mantienen sus valores de importancia a lo largo del año (ej. Desmanthus virgatus,
Zanthoxylum culantrillo). En el tercer caso vale la pena separar la tercera categoría
en dos subcategorías: 3.1. Especies que tienen un aumento notable en el valor de
importancia en el inicio de la temporada de secas (ej. Aeschynomene americana,
Guazuma ulmifolia, Ipomoea sp., Lysiloma divaricata, Stemmadenia bella, Triumfetta
semitriloba) y 3.2. Especies que tienen un aumento en sus valores de importancia al
final de la temporada seca (ej. Cochlospermum vitifolium, Ipomoea pauciflora,
Justicia candicans, Mimosa sp., Pithecoctenium crucigerum, Ruprechtia fusca).

75
0
50
100
150
200
Acanth
acea
e
Apocy
nace
ae
Bursera
ceae
Cochlo
sperm
acea
e
Convo
lvulac
eae
Faba
ceae
Rutace
ae
Sapind
acea
e
Sin de
termina
r
Familias
VI
PLlPS1PS2NpLlNpS1NpS2
Figura 22. Valores de importancia (VI) de las familias en el contenido de abazones, por condición por temporada. Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
Figura 23. Valores de importancia (VI) de las especies más importantes en el contenido de abazones, por condición por temporada (nombres completos en la Tabla 4). Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
Cabe también hacer dos excepciones a los patrones antes mencionados,
Bursera grandifolia y Conzattia multiflora mantienen algunos de los valores de
importancia más altos a lo largo del estudio y parece ser que su transporte en
abazones tiene un patrón diferente a las demás especies, pues en ambos casos la
importancia es muy alto en la temporada de lluvias, disminuye de manera importante
en la temporada inicial de secas y aumenta, de nuevo, a niveles altos al final de la
temporada seca.

76
Para las semillas en abazones existe, una amplitud de nicho trófico, a nivel de
familia, significativamente menor (F(1,2)=19.637 p=0.047) en los sitios no perturbados
que en los perturbados, al inicio de la temporada seca y mayor al final temporada
seca, en estos últimos la amplitud es similar a lo largo del año, mientras en los sitios
perturbados disminuye en la época seca tardía (Figura 24).
0
4
8
12
Ll S1 S2Temporadas
Am
plitu
d ni
cho
trófic
o (B
)
PNp
Figura 24. Amplitud de nicho trófico (B) del contenido de abazones, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
Los cambios en la selectividad de semillas de diversas especies por Liomys
irroratus a lo largo de las temporadas son muy notorios, (Figura 25), existen
especies que son seleccionadas durante una sola temporada (ej. Bursera aloexylon
en lluvias; Ipomoea sp al inicio de las lluvias e Ipomoea pauciflora al final de las
secas) y otras que son seleccionadas durante todo el año de forma importante (ej.
Bursera grandifolia, Conzattia multiflora y Serjania triquetra).

77
Especie PLl PS1 PS2 NpLl NpS1 NpS2 Morfo 3 0.533 0.103 0.090 0.028Morfo 5 0.182 0.042Morfo 6 0.182 0.135 0.042Morfo 9 0.061 0.180 Morfo 19 0.010 B. aloexylon 0.267 B. copallifera 0.141 B. grandifolia 0.250 0.061 0.207 0.030 0.617C. multiflora 0.125 0.091 0.621 0.424 0.131Ipomoea sp. 1 0.364 0.113 0.014I. pauciflora 0.090 0.025J. candicans 0.011Mimosa sp. 1 0.061 Mimosa sp 2 0.028P. crucigerum 0.056R. fusca 0.069 S. triquetra 0.125 0.424 0.271 0.006T. semitriloba 0.090
Figura 25. Tabla del índice estandarizado de selección de Manly-Gresson (Bi) para los contenidos de abazones, por condición y temporada (nombres completos en Tabla 4). P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías. Los valores sombreados indican lo seleccionado por encima de su disponibilidad, los no sombreados indican lo seleccionado por debajo de su disponibilidad.
Al analizar la preferencia se observó que, entre sitios perturbados y no
perturbados existen diferencias importantes relacionadas con la composición y la
preferencia de las especies encontradas en los abazones. Existen especies sólo
preferidas en sitios no perturbados (ej. Bursera copallifera, Pithecoctenium
crucigerum, Triumfetta semitriloba) y otras sólo en sitios perturbados (ej. Bursera
aloexylon, Mimosa sp). Aun en especies ampliamente preferidas (ej. Bursera
grandifolia y Conzattia multiflora) existen cambios dependientes de la temporada del

78
año que hacen que su grado de preferencia sea distinto y disminuya en algunas
temporadas. (Tabla 5).
El dendrograma basado en el índice de similitud (Figura 26) muestra que los
sitios no perturbados durante la parte final de la temporada seca se comportan de
forma diferente a todos los demás. También indica que no existe diferencia entre
condiciones durante el principio de las secas y que durante las lluvias los sitios de
ambas condiciones son similares. Lo más notorio es que los sitios perturbados
durante la temporada seca tardía son muy cercanos al comportamiento de estos
mismos sitios durante la época de lluvias, en que la granivoría es mínima.
Figura 26. Dendrograma del índice de similitud para número relativo de semillas en el contenido de abazones, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2=

79
secas tardías.
Los análisis de X2 para ambos tipos de condiciones, existe una composición
de especies significativamente diferente (p=0.0021) entre ambas condiciones, esto
es especialmente notorio en algunas especies como Bursera grandifolia, los morfos
3, 5, 6 y Serjania triquetra. En contraste Conzattia multiflora está representada de
manera muy similar en ambos sitios, haciendo notorio que: algunas especies son
recolectadas de manera diferente entre las condiciones hay algunas que son
buscadas independientemente de la perturbación.
Para el caso de las diferentes temporadas la X2 sólo se analizó para tres
especies (Bursera grandifolia, Conzattia multiflora y Serjania triquetra) dado que son
las únicas que especies que se encontraron en todas las temporadas y que
cumplieron los requerimientos de la prueba (menos del 20% de las especies con #
ind<5). La prueba determinó que existen diferencias muy significativas (<0.001) en la
abundancia de estas especies entre las temporadas, particularmente en los casos
de Bursera grandifolia y Serjania triquetra.
10.5. Resultados del análisis de las excretas.
Del muestreo de ratones se obtuvo un total de 477 (66% de los ratones
capturados) muestras de excretas, (160 en la temporada de lluvias, 204 en secas
tempranas y 113 en secas tardías), estos resultados no fueron significativamente
diferentes. En los sitios no perturbados se obtuvieron, de manera no significativa,
más muestras que en los perturbados (339 contra 138 de los perturbados).
Se observaron entre el 75 y 85% de las especies esperadas por los

80
estimadores en la dieta.
Se encontraron 31 (17.75%) fragmentos de material que no pudieron ser
determinados y 144 que si pudieron identificarse. Estos últimos se dividieron en 28
morfo especies, que fueron comparadas con el catálogo de referencia (Figura 27),
por esta comparación se pudieron determinar 10 especies pertenecientes a siete
familias, la más diversa fue Acanthaceae (4), mientras las demás estan
representadas por una especie. Para dos especies (Ludwigia octovalvis y Plumbago
scandens) no se tienen datos de diversidad y disponibilidad, por lo que se
excluyeron de los análisis (Tabla 6).

81
Figura 27. Ejemplos de comparaciones entre el contenido de las
excretas (40X izquierda) y el catálogo de referencia (100X derecha). Las
especies que se observan son, de arriba abajo: Tetramerium nervosum
(Acanthaceae), Desmodium procumbens (Fabaceae) y Oplismenus
burmannii (Poaceae).

82
Resulta importante destacar que en el caso de los resultados de excretas, al
igual ocurrió con la disponibilidad de recursos y contenido de abazones, existe un
importante recambio de especies entre sitios de ambas condiciones. En el caso de
excretas el 27% de las morfoespecies son exclusivamente de sitios perturbados y el
23% son de sitios no perturbados solamente. Este recambio también es notorio para
las diferentes temporadas pues el 30% de las morfoespecies sólo se encontró
durante las temporadas secas, siendo también el 23% exclusivo de la temporada de
lluvias.
Los índices de diversidad y equidad (Figura 28, Figura 29 para diversidad y
Figura 30 para equidad) muestran que la dieta de los ratones de los sitios
perturbados es ligeramente más diversa y equitativa que en los sitios no
perturbados, estas diferencias no fueron significativas. También se muestran dos
tendencias, no significativas, a través de las temporadas; en los sitios perturbados
existe un leve decremento durante la temporada seca inicial y un aumento muy
notorio hacia el final de la temporada seca (el índice de Simpson marca un aumento
para seca inicial y un decremento para seca tardía, pero el aumento es en general).
En el caso de los sitios no perturbados se da el caso opuesto, un leve aumento al
inicio de la temporada de estiaje y un declive al final de la temporada seca.
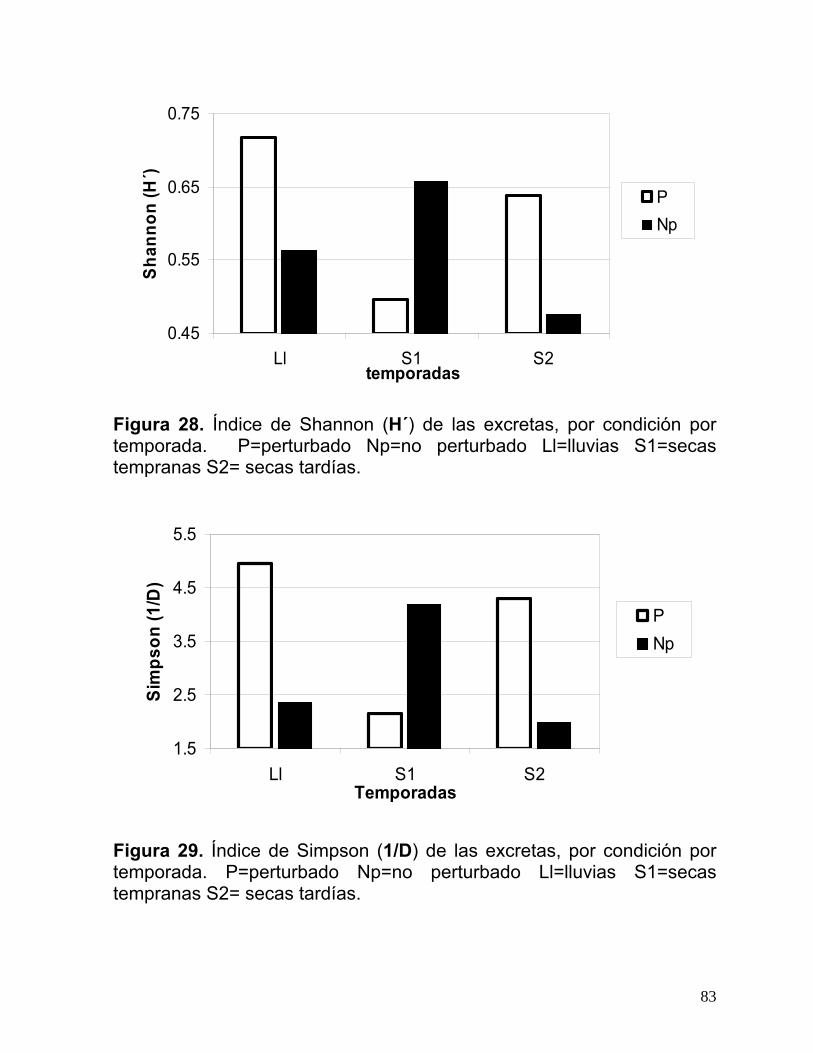
83
0.45
0.55
0.65
0.75
Ll S1 S2temporadas
Shan
non
(H´)
PNp
Figura 28. Índice de Shannon (H´) de las excretas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
1.5
2.5
3.5
4.5
5.5
Ll S1 S2Temporadas
Sim
pson
(1/D
)
PNp
Figura 29. Índice de Simpson (1/D) de las excretas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
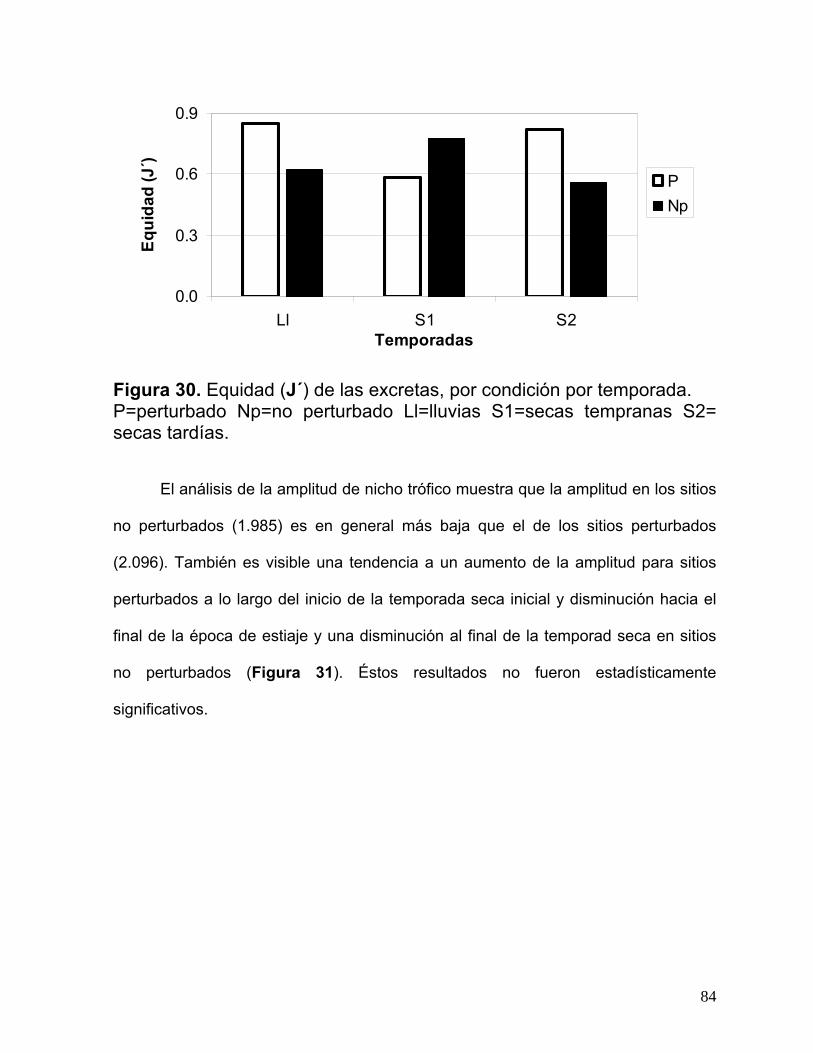
84
0.0
0.3
0.6
0.9
Ll S1 S2Temporadas
Equi
dad
(J´)
PNp
Figura 30. Equidad (J´) de las excretas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
El análisis de la amplitud de nicho trófico muestra que la amplitud en los sitios
no perturbados (1.985) es en general más baja que el de los sitios perturbados
(2.096). También es visible una tendencia a un aumento de la amplitud para sitios
perturbados a lo largo del inicio de la temporada seca inicial y disminución hacia el
final de la época de estiaje y una disminución al final de la temporad seca en sitios
no perturbados (Figura 31). Éstos resultados no fueron estadísticamente
significativos.

85
0
1
2
3
4
5
Ll S1 S2Temporadas
Am
plitu
d de
nic
ho tr
ófic
o (B
)
PNp
Figura 31. Amplitud de nicho trófico (B) de las excretas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
Al analizar la selectividad se observa (Figura 32) que el consumo de
herbáceas es selectivo para el pasto más común en el estudio, Oplismenus
burmannii, cuyo consumo es muy alto en ambas condiciones y todas las
temporadas. Aunadas a este patrón existen varias especies con índices de
selectividad menores que van cambiando su importancia en la alimentación de L.
irroratus a lo largo de las temporadas (Desmodium procumbens y Elytraria imbricata
durante las lluvias, Habracanthus silvaticus y Maranta arundinacea para el inicio de
la temporada seca), particularmente en la temporada de lluvias y el inicio de la seca.
Los sitios perturbados presentan una mayor cantidad de especies de manera
simultánea que los no perturbados. Al analizar la selección por condición se observa
de nuevo que Oplismenus burmannii es ampliamente buscado en ambas
condiciones y que Maranta arundinacea es seleccionada en sitios perturbados. Las
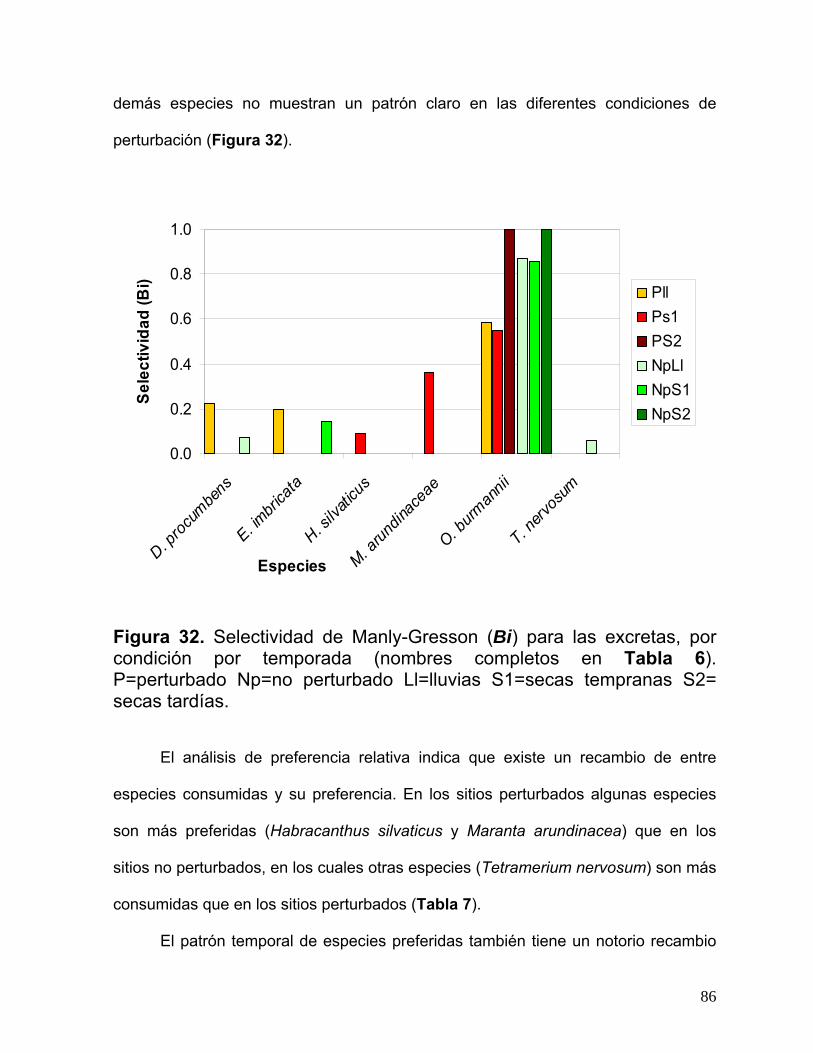
86
demás especies no muestran un patrón claro en las diferentes condiciones de
perturbación (Figura 32).
0.0
0.2
0.4
0.6
0.8
1.0
D. proc
umbe
ns
E. imbri
cata
H. silv
aticu
s
M. arun
dinac
eae
O. burm
annii
T. nerv
osum
Especies
Sele
ctiv
idad
(Bi) Pll
Ps1PS2NpLlNpS1NpS2
Figura 32. Selectividad de Manly-Gresson (Bi) para las excretas, por condición por temporada (nombres completos en Tabla 6). P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
El análisis de preferencia relativa indica que existe un recambio de entre
especies consumidas y su preferencia. En los sitios perturbados algunas especies
son más preferidas (Habracanthus silvaticus y Maranta arundinacea) que en los
sitios no perturbados, en los cuales otras especies (Tetramerium nervosum) son más
consumidas que en los sitios perturbados (Tabla 7).
El patrón temporal de especies preferidas también tiene un notorio recambio

87
en las (Tabla 7). Durante las lluvias y el inicio de la temporada seca se observa una
mayor riqueza de especies (más especies preferidas) en la dieta que al final de la
temporada seca en la cual solo se consume Oplismenus burmannii; entre la
temporada de lluvias y la de inicio de secas se observan también diferencias en la
composición de la dieta. Desmodium procumbens y Tetramerium nervosum que son
consumidas durante las lluvias son cambiadas por Habracanthus silvaticus y
Maranta arundinacea, mientras Elytraria imbricata es consumida a lo largo de ambas
temporadas. Durante todo el año Oplismenus burmannii es preferida más que todas
las demás especies consumidas. (Tabla 7).
El dendrograma del análisis de similitud de especies (Figura 33) genera dos
grupos, el primero constituido por sitios no perturbados durante las lluvias y el final
de las secas y sitios perturbados durante la parte inicial de las secas. El segundo
grupo se compone de los sitios perturbados durante lluvias y el final de las secas y
sitios no perturbados durante el inicio de las secas. Es importante señalar que la
menor similitud se dio entre los sitios de ambas temporadas secas. Se observa
también que los sitios no perturbados durante lluvias y los perturbados al inicio de
las secas son los más parecidos entre si. Los sitios peturbados durante secas
tardías son diferentes a todos los demás sitios en las otras temporadas. De estos
últimos resultados se puede inferir que durante este período, en los sitios
perturbados los ratones están consumiendo especies y proporciones de las mismas,
diferentes con respecto a las que consumen durante los demás muestreos (Figura
33).

88
Figura 33. Dendrograma del índice de similitud para número relativo de especies de herbáceas en las excretas, por condición por temporada. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías.
Las pruebas de X2 señalan que existen cambios significativos (p=0.013) entre
ambas condiciones de perturbación. Estos cambios son especialmente notorios en
algunas especies (Gomphrena decumbens y Oplismenus burmannii) indicando que
el consumo realizado por L. irroratus sobre estas especies está influenciado por la
perturbación.
Oplismenus burmannii es consumida en mayor proporción en los sitios no
perturbados que en los perturbados, en estos últimos el consumo de Gomphrena
decumbens (y algunas especies no determinadas) aumenta, posiblemente por que el
ratón no es más generalista en su dieta en los sitios no perturbados o por que estas

89
especies sean comunes de sitios perturbados.
Para el caso de las diferentes temporadas la X2 no muestra ninguna
diferencia significativa. Posiblemente las mismas especies son consumidas a lo largo
del año y sus proporciones cambian de manera ligera.

90
11. DISCUSIÓN.
Varios de los resultados estadísticos de este estudio no fueron significativos
pero, en ellos, se observan tendencias como lo muestra la figura 33.
VARIABLE NP P Resultado LL S Resultado
Herbáceas en suelo
Mayor div y disp
Menor disp y div
tendencia no
significativa
Mayor div y disp
Menor disp y div Si
Banco de semillas
Mayor div y disp
Menor disp y div
tendencia no
significativa
Menor disp y div
Mayor div y disk Si
pref evidente
sin pref evidente
ambas muestran
pref Semillas en abazones menor
amplitud nicho trófico
mayor amplitud nicho trófico
Si
Menor cantidad y menor div
Mayor cantidad y mayor div
SI
pref evidente
sin pref evidente Si
Consumo herbáceas
menor amplitud nicho trófico
mayor amplitud nicho trófico
tendencia no
significativa
Mayor cantidad y mayor div
Menor cantidad y menor div
tendencia solo en algunas
temporadas
Figura 33. Tabla comparativa de las respuestas esperadas y los resultados obtenidos para cada variable en cada condición y por temporada. Disp= disponibilidad. Div=diversidad. Pref=preferencia. P=perturbado, Np= no perturbado, Ll=lluvias, S=secas (incluye ambas temporadas secas).

91
Esto podría sugerir que los efectos de la perturbación ambiental y la
estacionalidad no son notorios sobre la dieta de Liomys irroratus, aunque por los
resultados obtenidos se sabe que los ratones consumen diferentes especies, tanto
de herbáceas como de semillas, entre las diferentes condiciones y temporadas de
estudio.
Un factor a considerar en los resultados estadísticos encontrados, es que el
diseño experimental usado es muy conservador (Von Ende, 1993) y tiende a no dar
como significativos resultados que con análisis más potentes podrían considerarse
significativos. En conjunto con lo anterior es importante destacar que el diseño con
efecto de bloque substrae grados de libertad al modelo lo cual resta aún más la
potencia del MANOVA, dado que este estudio tuvo pocas replicas.
Una serie de análisis exploratorios, sin el efecto del bloque, mostraron que:
en muy pocos casos el bloque tuvo un efecto significativo, y que si se realizan los
análisis sin él, varios resultados son significativamente diferentes a los encontrados
con el efecto de bloque.
Von Ende (1993) sugiere varias posibilidades para resolver este tipo de
problemas con el MANOVA de medidas repetidas, entre las cuales sugiere modificar
la α para que resultados de p≤0.1 sean significativos; o cambiar el modelo a un
ANOVA de medidas repetidas. El diseño de este proyecto se mantuvo como estaba
establecido para no romper con las formas ni caer en el error de ajustar el diseño
experimental de forma que favorezca los resultados que se esperan obtener,
intentando que sea lo más imparcial posible.
Desde luego, los datos generados presentan una gran variabilidad natural y

92
posiblemente eso lo reflejan los resultados no significativos de los análisis, a pesar
de la robustez del MANOVA para éste tipo de problemas. Este factor pudo haberse
minimizado aumentando el número de replicas, aunque logísticamente eso
complicaba mucho el desarrollo del trabajo.
Un factor adicional a considerar es que un par de sitios de estudio (un bloque)
aún cuando tenía contraste entre los dos sitios (perturbado vs no perturbado) en el
grado de perturbación, éste no fue tan intenso como el que indicaron los otros cuatro
sitios. Esta falta de contraste pudo haber resultado en respuestas más parecidas
entre esos sitios y por tanto afectar los análisis generales.
Otras variables, no consideradas en este estudio, que podrían estar
afectando la ecología de forrajeo de este ratón son: la interacción con otras especies
animales (competidoras o depredadoras), la calidad nutricional de semillas y
herbáceas y el efecto de la ganadería sobre la vegetación, sobre todo en el
sotobosque, lo que pudo causar que la perturbación y la estacionalidad no
mostrasen un efecto más contundente sobre la dieta de L. irroratus.
Es importante resaltar, de nuevo, que se detectaron cambios en la
composición de especies disponibles y utilizadas (tanto en abazones como excretas)
por este ratón, tanto estacionalmente como entre condiciones de perturbación. Esto
puede ser omitido por índices como el de diversidad, de forma tal que si el número
de especies y sus abundancias relativas son similares, no se detectan diferencias en
la diversidad o amplitud de nicho trófico en los análisis, pese a que las especies
sean distintas. El cambio en la composición del banco de semillas también es
señalado por Arias (1986), en la misma zona.

93
Los resultados de este trabajo muestran una tendencia (no significativa) hacia
una mayor disponibilidad y diversidad de herbáceas durante la temporada de lluvias
y en los sitios no perturbados y el patrón estacional inverso para el banco de
semillas, más diversidad y disponibilidad mientras más avanza la temporada de
estiaje y también mayor diversidad en los sitios no perturbados sobre los
perturbados. Ésto concuerda con lo esperado en las hipótesis de éste trabajo.
Se observó una tendencia a que la perturbación y la estacionalidad causen
una disminución en la heterogeneidad ambiental de la Sierra de Huautla, ésto
también es señalado en la teoría (Begon et al., 1999; Brown, 2000; Krebs, 1985;
Ceballos, 1995; Laurance & Bierregaard, 1997; Litvaitis, 2000; Miller, 1994).
Considerando que los lugares menos heterogéneos son menos estables (Krebs,
1985) es posible señalar que la estacionalidad climática de la zona, aunada con la
perturbación genera que el ambiente sea menos estable que en los sitios sin
perturbación.
Se encontró, tanto en la dieta como en el contenido de abazones, que existe
una flexibilidad en la selección alimentaria de Liomys irroratus, esto concuerda con lo
señalado por el factor de estabilidad ambiental (Krebs, 1985) que indica que el
alimentarse de manera más generalista (mayor amplitud de nicho trófico) es una de
las estrategias de las especies para enfrentar ambientes menos estables. Y que la
especialización se da en sitios más estables (no perturbados y con estacionalidad
menos marcada) en los que se alimentan de manera más selectiva (menor amplitud
de nicho trófico).
El patrón observado, para el contenido de abazones, a la disminución en la
amplitud de nicho trófico (generada por la selectividad) encontrada en este trabajo

94
concuerda con lo descrito en otros estudios (Cortés et al., 2002; Morris & Schwartz,
1957; Murray & Dickman, 1994; Reichman, 1977; Smith, 1952; Taylor, 1954), en los
que se observa como influye de manera notoria la especialización en la dieta en
disminuir la amplitud de nicho trófico, a pesar que los animales consuman una
riqueza de especies mayor a otras condiciones en las que son más selectivas.
Estudios compilatorios y de inventario realizados anteriormente (Ceballos &
Galindo, 1984, Davis & Russell, 1952, Villa & Cervantes, 2003), así como el de
García-Estrada et al. (2002) realizado en la Sierra de Huautla sugieren que por sus
características de adaptación a los medios semiáridos Liomys irroratus es resistente
a cambios ambientales como la aridez y la perturbación. Cadena (2003) en un
trabajo diseñado para comprobar los efectos de la perturbación y la estacionalidad
climática en la comunidad de ratones en la misma zona y tiempo en los que se
realizó este estudio sugiere que la perturbación y la estacionalidad si causan
cambios en la estructura poblacional de L. irroratus.
El presente trabajo sugiere de qué forma podrían darse estos cambios. Si
bien es cierto que las características morfofisiológicas de L. irroratus pueden
disminuir los efectos de los cambios en la cobertura vegetal o del aumento en la
aridez, asociados a la perturbación y la estacionalidad. También es cierto que los
cambios en la diversidad y disponibilidad de alimentos generan una presión de
índole diferente para los ratones. Al menos, los resultados de este trabajo evidencian
que si la selección de las especies que consumen, si cambia en función de la época
del año y del grado de perturbación de la selva y esto a su vez, podría reflejarse a
nivel de la dinámica poblacional, ocasionando diferencias como las reportadas por
Cadena (2003).

95
Poblaciones en sitios perturbados, en donde los recursos estan más limitados,
podrían tener mayor emigración y mayor competencia intrapoblacional e
interespecífica. Esto se ha encontrado para otras especies de roedores (Brown,
1973; Brown & Lieberman, 1973; Rosenzweig & Sterner, Reichman, 1975 con
comunidades de heterómidos desérticos; Cameron, 1977 con Sigmodon hispidus y
Reithrodontomys fulvescens; Cameron, 1982 con Sigmodon hispidus,
Reithrodontomys fulvescens y Oryzomys palustris; Cortés et al., 2002 con Lagidium
viscacia y Abrocoma cinerea; Kotler & Brown, 1990 con Gerbillus allenby y G.
pyramidum; Stancampiano & Caire con tres especies de Peromyscus (P. attwateri,
P. leucopus y P. maniculatus) y Reithrodontomys fulvescens). En estas
cirunstancias, una disminución en la selectividad alimentaria, tiende a disminuir la
competencia con congeneres o especies que consumen dichos recursos limitados,
pero podría aumentar la competencia con otras especies generalistas.
En este trabajo se considera que el contenido de abazones estima de manera
adecuada la dieta granívora de Liomys irroratus, sobre esto se tienen opiniones
diferentes, Jameson (1954) y Reichman (1975) sugieren que el contenido de
abazones no es un buen indicador de lo que los ratones consumen, pero que da
una idea cercana de los recursos que pueden utilizar y, posteriormente, varios
trabajos (Bradley & Mauer, 1971; Chapman; 1972; Johnson, 1961) complementan
ésta información encontrando que la cantidad relativa de insectos en la dieta es lo
que puede generar que el contenido de abazones sea diferente a lo encontrado en la
dieta, sugiriendo que en especies con un menor consumo de insectos el contenido
de abazones representa de manera objetiva lo encontrado en la dieta. Al no haber
encontrado rastros de insectos en la dieta o en los abazones de Liomys irroratus es

96
posible que el contenido de los abazones posiblemente indique lo que consume de
manera confiable, teniendo siempre en mente que bien puede ser que los consuman
en el lugar donde los encuentren o casualmente no hayan estado representados en
las muestras de excretas. También es importante tomar en cuenta que Liomys
salvini guarda semillas que nunca ha visto en los abazones, posiblemente para
llevarlas a su madriguera donde las prueba evitando ulteriormente las especies no
palatables, peligrosas o que simplemente no son de su gusto (Janzen, com. pers).
Para poder verificar la funcionalidad del contenido de los abazones como
indicador de la dieta de Liomys irroratus es necesario realizar el análisis de las
excretas comparando las estructuras con un catálogo de referencia de las semillas
disponibles, ésto no fue posible en este estudio por que, a pesar de existir una
metodología a seguir (Dacar & Giannoni, 2001) ésta fue localizada cuando el
proyecto estaba casi terminado y por cuestiones de tiempo y logística no fue posible
utilizarla).
El consumo de diferentes especies de semillas y herbáceas por Liomys
irroratus parece estar influenciado por su disponibilidad, como está descrito para
otros miembros de la familia Heteromyidae (Fleming, 1983; Janzen, 1986a;
Reichman, 1975; Reichman & Price, 1993, Villa & Cervantes, 2003) a los que se
considera consumidores más bien generalistas.
De forma contrastante se encontró que Liomys irroratus también selecciona
algunas especies (ej. Bursera grandifolia y Conzattia multiflora) a las cuales prefiere
independientemente de su disponibilidad (o de la de otras especies) el que las esté
transportando aun cuando no estén disponibles en el banco de semillas y no se
encuentren en la temporada de fructificación (tomadas de Arias, 1986) puede

97
significar que estos ratones tienen comportamiento parecido a L. pictus que
almacena una mayor cantidad de semillas en madrigueras con una gran cantidad de
túneles y cámaras, que no contienen nidos (Domínguez, 2000; Hernández, 2000); o
en almacenes superficiales externos como lo hacen otros heterómidos (Lockard &
Lockard, 1971; Shaw, 1934), por lo que posiblemente las vuelven a transportar
cuando las llevan a sus nidos para consumirlas, lo cual puede explicar por qué se
encontraron en los abazones durante todo el año. O que ésta especie esté
consumiendo dichas especies de manera preferente por encima de las demás, como
sugieren los resultados del índice de preferencia de Johnson (1980) obtenidos en
éste estudio.
En algunos casos Liomys irroratus selecciona familias (ej. Convolvulaceae,
Fabaceae, Sapindaceae) y especies (ej. Cochlospermum vitifolium, Ficus sp.,
Ipomoea sp., Jacaratia mexicana) que son reportadas como muy consumidas por L.
pictus (Ceballos, 1989, Mendoza, 1997) y L. salvini salvini (Fleming, 1974a; Fleming,
1974b; Janzen, 1986a) lo cual sugiere que los efectos de la perturbación en las
dietas de las tres especies pueden ser parecidos, puesto a que algunas de estas
especies se encuentran ausentes en sitios perturbados analizados en este trabajo.
Al comparar las especies encontradas en abazones durante este estudio
(Tabla 8) con las reportadas para Liomys pictus (Ceballos, 1989, Domínguez, 2000;
Hernández, 2000; Mendoza, 1997; Pérez, 1978) en Jalisco y para Liomys salvini en
Costa Rica (Fleming, 1974a; Fleming, 1974b; Janzen, 1971 y Janzen, 1986a), se
observa que algunas de las especies (ej. Bursera alloexylon, B. grandifolia, B.
copallifera) no estaban registradas anteriormente en abazones del género Liomys.
A pesar que muchas especies encontradas en abazones de L. pictus y L.

98
salvini se encuentran presentes en la Sierra de Huautla no fueron encontradas en
los abazones de L. irroratus, a pesar de que algunas son sugeridas por los autores
como preferidas por los roedores (ej. Enterolobium cyclocarpum por Liomys salvini
(Janzen, 1986a) y Serjania brachycarpa para Liomys pictus (Ceballos, 1989;
Mendoza, 1997)). Esto es posiblemente porque no están presentes en la zona de
muestreo o porque casualmente no fueron encontradas en los abazones.
Las semillas que L. irroratus transporta son de mayor tamaño que las
descritas para heterómidos desérticos (≤3mm) como lo señalan Reichman & Price
(1993), pero también parece haber diferencias con respecto a la perturbación. Los
ratones transportan semillas de especies más grandes en sitios no perturbados, y a
pesar de esto el número de semillas no varía significativamente entre condiciones, y
existe una tendencia a un mayor número de semillas por muestra de abazones en
los sitios no perturbados. Esto se relaciona a que las semillas preferidas por Liomys
irroratus son, generalmente, semillas más grandes, que posiblemente sean de
árboles poco comunes en sitios perturbados, las semillas más grandes fueron las de
Conzattia multiflora (5 a 12mm) y Bursera grandifolia (7 a 11mm) o también es
posible que sean más nutritivas y éste sea una forma de Liomys irroratus para
forrajear optimizamente (Cassini, 1994; Reichman, 1977; Pyke, 1984).
Para el caso de las herbáceas existe poca información con la cual comparar
este trabajo y la existente es básicamente sobre heterómidos desérticos (Reichman,
1975; Reichman, 1977; Reichman & Van de Graaf, 1975) por lo que se discutirán
estos resultados de manera menos extensa que los de contenido de abazones.
Liomys irroratus consume de manera muy preferente Oplismenus burmannii por
encima de todas las demás especies y mantiene su consumo a pesar de la condición

99
de perturbación y la temporada en la que se encuentre, esto es posiblemente
generado porque almacenan fragmentos de esta especie en las madrigueras y la
consumen durante todo el año. Éste ratón consume otras especies de forma menos
generalizada, por una tempor ada o una condición, pero durante la parte final de las
secas sólo consumen Oplismenus burmannii, lo cual indica que la están
almacenando para esa temporada desde las lluvias lo cual es más notorio en los
sitios no perturbados, situación que concuerda con la primera hipótesis del proyecto,
ésta supone que la disponibilidad y diversidad son mayores en sitios no perturbados
que en perturbados y por lo tanto la selectividad será mayor que en los perturbados.
Estos resultados permiten inferir que, como lo reportado por Reichman & Van de
Graaf (1975) para Dipodomys merriami y Perognathus amplus, en el caso de Liomys
irroratus es posible que el consumo de plantas esté ligado a sus ciclos reproductivos.
Se puede señalar, basándose en la información de éste trabajo y la literatura
disponible de la familia Heteromyidae (ej. Fleming, 1983a; Fleming, 1983b; Janzen,
1971; Janzen, 1986a; Sánchez-Cordero & Fleming, 1993) que L. irroratus tiene una
importante labor como depredador de semillas (Ceballos y Galindo, 1984; Ceballos y
Miranda, 2000; Dhillion, 1999; Huntly & Reichman, 1994; MacCracken et al., 1985;
McDonald, 2001; Villa y Cervantes, 2003), pues utiliza diversas especies y las utiliza
de manera diferencial, por que puede estar afectando su reproducción y causando
cambios en la fisonomía de la selva.
La importancia de este ratón como dispersor tiene que ser analizada por
medio de estudios de la estructura de las madrigueras y de germinación de semillas
encontradas en los abazones, a pesar que las mayoría de las especies encontradas
en los abazones no tenían daños que aparentemente pudiesen hacerlas inviables,

100
es probable sean almacenadas en madrigueras, a mayor profundidad que la que
requiere para germinar y no se dispersen de manera adecuada o que el ratón las
consuma cuando empiezan a germinar y pierden dureza como es señalado por
Janzen (1986a) para Liomys salvini en Costa Rica.

101
12. CONCLUSIONES.
1. Liomys irroratus tiene una dieta herbívora basada en 10 especies distintas
de las que consume la herbácea Oplismenus burmannii (la más común en las
muestras de vegetación) de forma preferente y en magnitud independiente a su
disponibilidad, a la perturbación y la estacionalidad. Mientras que la selectividad y la
composición específica de las demás categorías tróficas que consume cambian
conforme su disponibilidad, la estacionalidad y la condición.
2. La dieta granívora de Liomys irroratus es más diversa que la herbívora,
utiliza 34 especies diferentes; de las cuales la mayoría las usa en la misma
proporción que en la que se encuentran en el medio. Mientras que Bursera
grandifolia y Conzattia multiflora (especies muy abundantes en la zona) las recolecta
de manera preferente en ambas condiciones de perturbación y en todas las
estaciones.
3. Liomys irroratus consume especies indicadoras de un buen estado de
conservación (ej. Cochlospermum vitifolium y Jacaratia mexicana) lo cual puede
tener efectos en la estructura vegetal de la selva, sugiriendo la importancia en los
cambios en la composición de especies en el banco de semillas de L. irroratus como
depredador o dispersor de semillas en la Sierra de Huautla.
4. Aún cuando se notan tendencias que sugieren un efecto claro de la
perturbación y la estacionalidad en la dieta de Liomys irroratus en la zona, pocos
resultados fueron significativos (solo el promedio de medida de las semillas en
abazones, la cantidad de muestras de abazones, la amplitud de nicho trófico de

102
semillas en abazones).
5. Por otra parte fue notorio que, asociado a la perturbación y la
estacionalidad, hubo cambios en la composición especies que son los recursos
alimentarios que selecciona este ratón.
6. De manera general se observa que los sitios no perturbados son más
estables que los perturbados en la disponibilidad y diversidad de recursos
alimenticios para este ratón, lo que fue particularmente notorio para la parte final de
la época seca.
7. Liomys irroratus utiliza sus recursos alimentarios (semillas) de forma más
selectiva (menor amplitud de nicho trófico) en sitios no perturbados y temporadas
secas y más generalista (mayor amplitud de nicho trófico) en sitios perturbados y
temporada de lluvias.
8. Es importante complementar los resultados de este trabajo con estudios
sobre las madrigueras, análisis de nutrientes de los recursos que consume ésta
especie contra los disponibles y comparación del contenido de abazones con lo
encontrado en las excretas.

103
13. Referencias.
American Psychological Association. (1983). Manual of the American Psychological
Association. [Criterios para la elaboración de documentos psicológicos] tercera
edición. México. 110pp.
Animal Care and Use committee. (1998). Guidelines for the capture, handling and care of
mammals. As approved by the American Society of Mamalogists.
(http://www.mammalsociety.org/pubsociety/index.html)
Aleksiuk, M. (1970). The seasonal food regime of artic beavers. Ecology. 51(2): 264-270.
Alexander, L. F. & Riddle, B. R. (2005). Phylogenetics of the new world rodent family
Heteromyidae. Journal of Mammalogy 86(2): 366-379.
Altrichter, M., Saenz, J. C., Carrillo, E. & Fuller, T. K. (2000). Dieta estacional del
Tayassu pecari (Artiodactyla: Tayassuidae) en el parque nacional Corcovado, Costa
Rica. Biología tropical. 48(2-3):42-56.
Alldredge, J. R. & Ratti, J. T., (1986). Comparison of some statistical techniques for
analysis of resource selection. Journal of Wildlife Management 50:157-165.
Álvarez-Castañeda, S. T. (1996). Los Mamíferos del Estado de Morelos. Centro de
Investigaciones Biológicas del Noroeste S. C. 211pp.
Ángeles, G., Ortega, F & Zendejas, P. (2002). Anatomía vegetal sin bisturí. Ciencia y
Desarrollo. 164(28):19-23.
Animal Behavior Society. (2000). Guidelines for the treatment of animals in behavioral
research and teaching. Animal Behavior. 59:253-257.
Anthony, R. G. & Smith, N. S. (1974). Comparison of rumen and fecal analysis to describe
deer diets. Journal of Wildlife Management. 38(3):535-540. Aranda, M., Rosas, O., Rios, J. de J. y García, N. (2002). Análisis de la alimentación del
gato montés (Lynx rufus) en dos diferentes ambientes en México. Acta Zoológica
Mexicana (n.s.) 87:99-109.
Arias, D. M. (1986). Análisis de la comunidad de semillas del estrato arbóreo en suelos
superficiales de selva baja caducifolia. Tesis de Licenciatura, UAEM, Escuela de
Ciencias Biológicas. 92pp.

104
Bartolomé, J., Franch, J., Gutman, M. & Seligman, N. G. (1995). Technical note: Physical
factors that influence fecal análisis estimates of herbivore diets. Journal of Range
Management. 48(3): 267-270.
Baumgartner, L. L., & Martin, A. C. (1939). Plant histology as an aid in squirrel food-
habitat studies. Journal of Wildlife Management. 3(3):266-268.
Begon, M., Harper, J. L. & Townsend, C. R. (1999). Ecología: individuos, poblaciones y
comunidades. Tercera edición. Ed. Omega. 1068pp.
Best. T. L. (1993). Patterns of morphologic and morphometric variation in heteromyid
rodents. (Capítulo 3:197-235). En: Genoways, H. H. & Brown, J. H. (eds) Biology of
the Heteromyidae. American Society of Mammalogists. Special publication #10. pp.
719.
Bradley, G. W. & Mauer, R. A. (1971). Reproduction and food habits of Merriam´s
kangaroo rat, Dipodomys merriami. Journal of Mammalogy 52(3): 497-507.
Brewer, S. W. (2001). Predation and dispersal of large and small seeds of a tropical palm.
Oikos 92(2):245-255
Briones, M. & Sánchez-Cordero, V. (1999). Dietary value of fruits and seeds to spiny
pocket mice (Liomys pictus) in a tropical deciduos forest in México. Studies on
neotropical fauna and environment. 34: 65-71.
Brown, J. H. (1973). Species diversity of seed-eating desert rodents in sand dune habitats.
Ecology 54(4):775-787.
Brown, J. H. & Harney, B. A. (1993). Population and community ecology of heteromyid
rodents in temperate habitats. (Capítulo 17:618-651) En: Genoways, H. H. & Brown,
J. H. (eds) Biology of the Heteromyidae. American Society of Mammalogists. Special
publication #10. pp. 719.
Brown, J. H. & Lieberman, G. A. (1973). Resource utilization and coexistence of seed-
eating desert rodents in sand dune habitats. Ecology 54(4):788-797
Brown, J. H., Reichman, O. J., Davidson, D. W. (1979). Granivory in desert ecosystems.
Annual Review of Ecology and Systematics 10:201-227.
Brown, J. S. (2000). Foraging ecology of animals in response to heterogeneous
environments. (Capítulo 10: 181-214) En Hutchings, M. J., John, E. A. & Stewart, A.
J. A. The ecological consequences of environmental heterogeneity. Blackwell

105
Science Ltd. Oxford, Inglaterra.
Brylski, P. (1993). The evolutionary morphology of heteromyids. (capítulo 9:357-385). En
Genoways, H. H. & Brown, J. H. (eds) Biology of the Heteromyidae. American
Society of Mammalogists. Special publication #10. 719pp.
Bullock, S. (2002). Cordia eleagnoides DC. (Boraginaceae) Bárcino. (151-153). En:
Noguera, F. A., Vega-Rivera, J. H., García-Aldrete A. N. y Quesada-Avendaño, M.
(Eds). Historia Natural de Chamela. Instituto de Biología, Universidad Nacional
Autónoma de México. México. 568pp.
Byers, C. R. & Steinhorst, R. K. (1984). Clarification of a technique for analysis of
utilization-availability data. Journal of Wildlife Management 48:1050-1053.
Cabin, J. R., Weller, S. G., Lorence, D. H., Cordell, S. & Hadway, L. J. (2002). Effects of
microsite, water, weeding, and direct seeding on the regeneration of native and alien
species within a Hawaiian dry forest preserve. Biological Conservation 104 (2002)
181–190.
Cadena, M. (2003). Efectos de la perturbación y estacionalidad en la
comunidad de roedores en una selva baja caducifolia en Morelos, México. Tesis de
licenciatura. Universidad de las Américas, Puebla. Escuela de Ciencias. 92pp.
Cameron, G. N. (1977). Experimental species removal: demographic responses by
Sigmodon hispidus and Reithrodontomys fulvescens. Journal of Mammalogy 58:488-
506. Campos, C., Ojeda, R., Monge, S. & Dacar, M. (2001). Utilization of food resources by
small and medium-sized mammals in the Monte Desert biome, Argentina. Austral
Ecology 26:142-149.
Carter, C. H. & Genoways, H. H. (1978). Liomys salvini Mammalian Species 84:1-5.
Cassini, M. H. (1994). Behavioral mechanisms of selection of diet components and their
ecological implications in herbivorous mammals. Journal of Mammalogy 75(3):733-
740.
Ceballos, G. (1989). Population and community ecology of small mammals from tropical
deciduos and arroyo forest in Western Mexico. Unpublished Ph. D. Thesis. University
of Arizona, Tucson, Arizona, 158pp.
Ceballos, G. (1995). Vertebrate diversity, ecology, and conservation en Neotropical dry

106
forests. (195-220) En Bullock, S. H., Mooney, H. A. & Medina, E. (eds.) Seasonally
Dry Tropical Forests. . Cambridge University Press. EUA. 568pp.
Ceballos, G. & Galindo, C. (1984). Mamíferos silvestres de la cuenca de México (pp. 179,
180). Limusa. México. 299pp.
Ceballos. G. & Miranda A. (2000). Guía de campo de los mamíferos de la costa de
Jalisco, México. Fundación Ecológica de Cuixmala. UNAM, México. 502pp.
Cervantes, F. A., Lorenzo, C. & Ward, O. G. (1999). Chromosomal relationships among
spiny pocket mice, Liomys (Heteromyidae), from Mexico. Journal of Mammalogy
80(3):823-832.
Chapman, B. R. (1972). Food habits of Loring´s kangaroo rat, Dipodomys elator. Journal of
Mammalogy 53(4): 877-880.
Chesson, J. (1978). Measuring preference in selective predation. Ecology 59(2): 211-215.
Chesson, J. (1983). The estimation and analysis of preference and its relationship to
foraging models Ecology 64(5):1297-1304.
Colwell, R.K. & Futuyma, D. J. (1971). On the measurement of niche breadth and overlap.
Ecology, 52 (3):567–576.
Cooperrider, A. Y., Boyd, R. J. & Stuart, H. R. (1986). Inventory and monitoring of wildlife
habitat. U.S. Dept. Inter., Bur. Land Manage. Service Center. Col. USA. 858pp.
Cortés, A., Rau. J, Miranda. E, y Jimenez E. J. (2002). Hábitos alimenticios de Lagidium
viscacia y Abracoma cinerea; roedores sintopticos en ambientes altoandinos del
norte de Chile, Revista Chilena de Historia Natural 75: 583-594.
Clevenger, A. P., Purroy, F. J. & Pelton, M. R. (1992). Food habits of brown bears (Ursus
arctos) in the cantabrian mountains, Spain. Journal of Mammalogy 73(2): 415-421.
Crawford, H. S., (1982). Seasonal food selection and digestibility by tame white-tailed deer
in central Maine. Journal of Wildlife Management. 46(6): 974-982.
Dacar, M. A. & Giannoni, S. M. (2001). A simple method for preparing reference slides of
seed (Technical note) Journal of Range Management 54(2): 191-193.
Darwin, C. R. (2001). El origen de las especies Segunda edición. Editores Mexicanos
Unidos S. A. México. 506pp.
De León, M. A. (2005). Fenología de especies de plantas con frutos carnosos y
disponibilidad espacial y temporal de este recurso en la Reserva de la Biosfera

107
Sierra de Huautla: implicaciones para los vertebrados. Tesis de licenciatura.
Universidad Autónoma del Estado de Morelos, Facultad de Ciencias Biológicas.
66pp.
Dhillion, S. S. (1999). Environmental heterogeneity, animal disturbances, microsite
characteristics, and seeding establishment in a Quercus havardii community.
Restoration Ecology. 7(4): 399-406.
Dickman, C. R. & Huang, C. (1988). The reliability of fecal analysis as a method for
determining the diet of insectivorous mammals. Journal of Mammalogy. 69(1):108-
113.
Dirzo, R. & Miranda. A. (1990). Contemporary neotropical defaunation and forest structure,
function and diversity: a sequel to John Terborgh. Conservation Biology 4:444-447.
Dirzo, R. & Raven, P. (2003). Global state of biodiversity and loss. Annual Review of
Environment and Natural Resources 28:137-167.
Domínguez, Y. C. (2000). Estructura del contenido de las madrigueras de Liomys pictus
en selva mediana subperennifolia, de la estación de biología Chamela, Jalisco. Tesis
de licenciatura UNAM, Campus Iztacala. 86pp.
Dorado, O., Valenzuela, D. y Ramírez, R. (En prensa). Ficha técnica 29 Sierra de
Huautla. En: Ceballos, G., Maass, M., Bezaury, J. & Espinoza, E. (eds). Diversidad,
amenazas y regiones prioritarias para la conservación de las selvas secas del
pacífico de México. CONABIO / CONANP / UNAM / WWF.
Dowler, R. C. & Genoways, H. H., (1978). Liomys irroratus Mammalian Species 82:1-6.
Dusi, J. L. (1949). Methods for the determination of food habits by plants microtechniques
and histology and their application to cottontail rabbit. Journal of Wildlife
Management. 13(3):295-298. Ellis, R. P. (1979). A procedure for standardizing comparative leaf anatomy in the Poaceae.
II. The epidermis as seen in surface view. Bothalia 12(4):641-671.
Escobar, A. & Gonzalez, E. (1973). Estudio de la competencia alimenticia de los
herbívoros mayores del llano inundable con especial al chigüire (Hydrochoerus
hydrochaeris) Agronomía Tropical 26(3):229-236.
Fagerstone, K. A., Tietjen, H. P. & Williams, O. (1981). Seasonal variation in the diet of
black-tailed prairie dogs. Journal of Mammalogy 62(4): 820-824.

108
Fitzgerald, A. E. & Waddington, D. C. (1979). Comparison of two methods of fecal
analysis of herbivore diet. Journal of Wildlife Management 43 (2):468-473.
Fleharty, E. C. & Olson, L. E. (1969). Summer foods of Microtus ochrogaster and
Sigmodon hispidus. Journal of Mammalogy 50 (3): 475-486.
Fleming, T. H. (1974a). The population ecology of two species of Costa Rican heteromyid
rodents. Ecology 55:493-510.
Fleming, T. H. (1974b). Social organization in two species of Costa Rican heteromyid
rodents. Ecology 55:541-561.
Fleming, T. H. (1977). Response of two species of tropical heteromyid rodents to reduced
food and water availability. Journal of Mammalogy 58(1):102-106.
Fleming, T. H. (1983a). Heteromys desmarestianus (474-475) En Janzen, D. H. (editor)
Costa Rican natural history. The Chicago University Press. 816pp.
Fleming, T. H. (1983b). Liomys salvini (475-477) En Janzen, D. H. (editor) Costa Rican
natural history. The Chicago University Press. 816pp.
Fleming, T. H. & Brown, G. J. (1975). An experimental analysis of seed hoarding and
burrowing behaviour in two species of Costa Rican heteromyid rodents. Journal of
Mammalogy 56(2)301-316.
Frank, C.L. (1988). Diet selection by a heteromyid rodent: role if net metabolic water
selection. Ecology 69(6):1943-1951.
Free, J. C., Hansen, R. M. & Sims, P. L. (1970). Estimating dryweights of foodplants in
feces of herbivores. Journal of Range Management. 23(2):300-302.
French, A. R. (1993). Physiological ecology of the Heteromyidae: economycs of energy and
water utilization. (capítulo 13:509-538). En: Genoways, H. H. & Brown, J. H. (eds)
Biology of the Heteromyidae. American Society of Mammalogists. Special Publication
#10. 719pp.
Galt, H. D., Theurer, B., Ehrenreich, J. H., Hale, W. H. & Martin, S. C. (1969). Botanical
composition of diet of steers grazing a desert grassland range. Journal of Range
Management. 22(1):14-18.
García, E. (1981). Modificaciones al sistema de Köppen para adaptarlo a las condiciones
de la república mexicana. Editorial Larios, México. 150pp.
García-Estrada, C., Romero-Almaraz, Ma. De L. y Sánchez-Hernández, C. (2002).

109
Comparison of rodent communities in sites with different degrees of disturbance in
deciduous forest of southeastern Morelos, Mexico. Acta Zoológica Mexicana (n.s.)
85: 153-168.
Garshelis, D. L. (2000). Delusions in habitat evaluation: measuring use, selection, and
importance (Capítulo 4). En: Boitani, L., Fuller, T. K. (Editores). Research
Techniques in Animal Ecology controversies and consequences. Columbia University
Press. EUA.
Gaviño, G., Juárez-López, C. & Figueroa, H. H. (1999). Técnicas biológicas selectas de
laboratorio y de campo. Segunda edición. Limusa. México.133pp.
Grajales-Tam, K. M., Rodriguez-Estrella, R. y Cancino, J. (2003). Dieta estacional del
coyote Canis latrans durante el periodo 1996-1997 en el desierto del Vizcaíno, Baja
California Sur, México. Acta Zoológica Mexicana (n.s.) 89: 17-28.
Hafner, J. C. (1993). Macroevolutionary diversification in heteromyid rodents: heterochrony
and adaptations in phylogeny. (capítulo 7:291-318). En Genoways, H. H. & Brown, J.
H. (eds). Biology of the Heteromyidae. American Society of Mammalogists Special
Publication #10. 719pp.
Hansen, R. M. (1976). Foods of free roaming horses in southern New Mexico. Journal of
Range Management. 29(4):347.
Hansen, R. M. & Dearden, B. L. (1975). Winter foods of mule deer in Piceance Basin,
Colorado. Journal of Range Management. 28(4):298-300.
Hansen, R. M. & Reid, L. D. (1975). Diet overlap of deer, elk, and cattle in southern
Colorado. Journal or Range Management. 28(1):43-47.
Hansen, R. M., Peden, D. G. & Rice, R. W. (1973). Discerned fragments in faeces
indicates diet overlap. Journal of Range Management. 26(2):103-105.
Hansen, R. M. & Ueckert, D. N. (1970). Dietary similarity of some primary consumers.
Ecology. 51(4):640-648.
Hernández, B. Del C. (2000). Caracterización espacial y contenido de madrigueras de
liomys pictus en una selva baja de jalisco. Tesis de licenciatura, UNAM, Campus
Iztacala. 40pp.
Hinnant, R. T. & Kothmann, M. M. (1988). Collecting, drying and preserving feces for
chemical and microbiological analysis. Journal of Range Management. 41(2):168-

110
171.
Holechek, Jerry. L. (1982). Sample preparation techniques for microhistological analysis.
Journal of Range Management. 35(2):267-268.
Holechek, J. L. & Gross, B. (1982a). Training needed for quantifying simulated diets from
fragmented range plants. Journal of Range Management. 35(5):644-647.
Holechek, J. L. & Gross, B. (1982b). Evaluation of different calculation procedures for
microhistological analysis. Journal of Range Management. 35(6): 721-723.
Holechek, J. L., Gross, B., Dabo, S. M. & Stephenson, T. (1982). Effects of sample
preparation, growth stage, and observer on microhistological analysis of herbivore
diets. Journal of Wildlife Management 46(2):502-505.
Holechek, J. L. & Vavra, M. (1981). The effect of slide and frequency observation
numbers on the precision of microhistological analysis. Journal of Range
Management. 34(3):337-338.
Holechek, J. L., Vavra, M. & Pieper, R. D. (1982a). Botanical composition determination of
range herbivore diets: A review. Journal of Range Management. 35(3):309-315.
Holechek, J. L., Vavra, M., Skovlin, J. & Krueger, W. C. (1982b). Cattle diets in the blue
mountains of Oregon II. Forests. Journal of Range Management. 35(2):239-242.
Howard, V. W. Jr. (1967). Identifying fecal groups by pH analysis. Journal of Wildlife
Management. 34(1):190-191.
Howe, H.F., Smallwood, J., (1982). Ecology of seed dispersal. Annual Review of Ecology
and Systematics 13: 201–228.
Hudson, J. W. & Rummel, J. A. (1966). Water metabolism and temperature regulation of
the primitive heteromyids, Liomys salvini and Liomys irroratus. Ecology. 47(3): 345-
354.
Huntly, N. & Reichman, O. J. (1994). Effects of subterranean mammalian herbivores on
vegetation. Journal of Mammalogy. 75(4):852-859.
Jameson, E. W. Jr. (1952). Foods of deer mice, Peromyscus maniculatus and P. boylei, in
the northern Sierra Nevada, California. Journal of Mammalogy 33(1): 50-60.
Janzen, D. H. (1971). Seed predation by animals. Annual Review of Ecology and
Systematics 2:465-492.
Janzen, D. H. (1981). Lectins and plant-herbivore interactions. Recent Advances in

111
Phytochemistry 15:241-258.
Janzen, D. H. (1982a). Seed removal from fallen guanacaste fruits (Enterolobium
ciclocarpum) by spiny pocket mice (Liomys salvini). Brenesia 19/20:425-429.
Janzen, D. H. (1982b). Removal of seeds from horse dung by tropical rodents: influence of
habitat and amount of dung. Ecology 63:1887-1900.
Janzen, D. H. (1982c). Attraction of Liomys mice to horse dung and the extinction of this
response. Animal Behaviour 30:483-489.
Janzen, D. H. (1982d). Fruit traits, and seed consumption by rodents, of Crescentia alata
(Bignoniaceae) in Santa Rosa National Park, Costa Rica. American Journal of
Botany 69:1258-1268.
Janzen, D. H. (1986a). Mice, big mammals and seeds: it matters who defecates what
where (Capítulo 20:251-271) En: Estrada, A. and Fleming, T. H. (eds.), Frugivores
and seed dispersal. Dr W. Junk Publishers, Dordrecht, The Netherlands. 392 pp.
Janzen, D.H., (1986b). The future of tropical ecology. Annual Review of Ecology and
Systematics 17, 305–324.
Janzen, D.H., (1988). Tropical dry forests: the most endangered major tropical ecosystem.
In: Wilson, E.O. (Ed.), Biodiversity. National Academy Press, Washington, pp. 130–
144.
Janzen, D. H. & Martin, P. S. (1982). Neotropical anachronisms: the fruits the gomphoteres
ate. Science 215 (1): 19-27.
Johnson, D. H. (1980). The comparison of usage and availability measurements for
evaluating resource preference. Ecology 61(1): 65-71.
Johnson, D. R. (1961). The food habits of rodents on rangelands of southern Idaho.
Ecology 42(2): 407-410.
Johnson, M. K. (1982). Frequency sampling for microscopic analysis of botanical
compositions. Journal of Range Management. 35(4):541-542.
Johnson, M. K. & Pearson, H. A. (1981). Esophageal, fecal and exclosure estimates of
cattle diets on a longleaf pine-bluestem range. Journal of Range Management.
34(3):232-234.
Johnson, M. K. & Wofford, H. (1983). Digestion and fragmentation: influence on herbivore
diet analysis. Journal of Wildlife Management. 47(3): 877-879.

112
Johnson, M. K., Wofford, H. & Pearson, H. A. (1983). Microhistological techniques for
food habits analyses. U.S. Department of Agriculture, Research Paper SO-199. New
Orleans, Lo. U. S. A. 40pp.
Jones, C., McShea, W. J., Conroy, M. J. & Kunz, T. H. (1996). Capturing mammals
(capítulo 8 115:156) En: Wilson, D. E., Cole, F. R., Nichols, J. D., Rudran, R. &
Foster, M. S. (Editores). Measuring and monitoring biological diversity. Standard
methods for mammals.. Smithsonian Institute. 408pp.
Jones, W. T. (1993). Social systems of heteromyid rodents (Capítulo 15:575-595). En
Genoways, H. H. & Brown, J. H. (eds). Biology of the Heteromyidae. American
Society of Mammalogists special publication #10. 719pp.
Katona, K. & Altbäcker, V. (2002). Diet estimation by faeces analysis: sampling
optimization for the European hare. Folia Zoológica. 51(1):11-15. Kincaid, W. B. & Cameron, G. N. (1982). Dietary variation in three sympatric rodents on
the Texas coastal prairie. Journal of Mammalogy. 63(4):668-672.
Kincaid, W. B. & Cameron, G. N. (1985). Interactions of cotton rats with patchy
environment: dietary responses and habitat selection. Ecology 66(6):1769-1783.
Krebs, C. J. (1985). Estudio de la distribución y la abundancia. Oxford University Press
segunda edición. México. 753pp.
Krebs, C.J. (1999). Ecological methodology. Benjamin/Cummings. Second edition. E.U.A.
620pp.
Kronfeld, N. & Dayan, T. (1998). A new method of determining diets of rodents. Journal of
Mammalogy. 79(4):1198-1202.
Lacher. T. E. Jr., Willig, M. R. & Mares, M. A. (1982). Food preference as a function of
resource abundance with multiple prey types: An experimental analysis of optimal
foraging theory. The American Naturalist 120(3):297-316.
Larter, N. C. & Gates, C. C. (1991). Diet and Habitat selection of wood bison in relation to
seasonal changes in forage quantity and quality. Canadian Journal of Zoology
69:2677-2685.
Laurance, W. F. & Bierregaard, R. O. jr. (1997). Tropical forest remnants: Ecology,
conservation and management of fragmented communities University of Chicago
Press. (502-514). 632 pp.

113
Laylock, W. A., Buchanan, H. & Krueger, W. C. (1972). Three methods of determinating
diet, utilization and trampling damage on sheep ranges. Journal of Range
Management. 25(2):352-356.
Lerdau, M.J., Whitbeck, J., Holbrook, N.M., (1991). Tropical deciduous forests: death of a
biome. Trends in Ecology and Evolution 6, 201–202.
Levins, R. (1968). Evolution in changing environments: some theoretical explorations.
Princetown University Press, Princetown, USA. 132 pp.
Lockard, R. B. & Lockard, J. S. (1971). Seed preferences and buried retrieval of
Dipodomys deserti. Journal of Mammalogy 52(1):219-221.
Lott, E. (1985). Listados florísticos de México III. La Estación de Biología Chamela. Instituto
de Biología, UNAM. México. 47pp.
Luce, D. G., Case, R. M. & Stubbendieck, J. (1980). Food habits of the plains pocket
gopher on western Nebraska rangeland. Journal of Range Management. 33(2):229-
231.
Litvaitis, J. A. (2000). Investigating food habits of terrestrial vertebrates (Capítulo
5:165:190) En: Boitani, L., Fuller, T. K. (Editores). Research Techniques in Animal
Ecology controversies and consequences. Columbia University Press. EUA.
Maass, J.M., (1995). Conversion of tropical dry forest to pasture and agriculture. In: Bullock,
S.H., Mooney, H.A., Medina, E. (Eds.) Seasonally Dry Tropical Forests. Cambridge
University Press, Cambridge, USA. pp. 399–422.
MacCracken, J. G., Uresk, D. W. & Hansen, R. M. (1985). Burrowing owl foods in Conata
Basin, South Dakota. Great Basin Naturalist.45(2):287-290.
Mandujano, S. (2002). Spondias purpurea L. (Anacardiaceae) Ciruelo. (pags: 45-150).
En: Noguera, F. A., Vega-Rivera, J. H., García-Aldrete A. N. y Quesada-Avendaño,
M. (Eds). Historia Natural de Chamela. Instituto de Biología, Universidad Nacional
Autónoma de México. México. 568pp.
Mares, M. A. (1993). Heteromyids and their ecological counterparts: a pandesertic view of
rodent ecology and evolution. (Capítulo 18:652-714) En: Genoways, H. H. & Brown,
J. H. (eds) Biology of the Heteromyidae. American Society of Mammalogists. Special
publication #10. pp. 719.
May, R. M. & MacArthur, R. H. (1972). Niche overlap as a function of environmental

114
variability. Procedures of the National Academy of Science (USA) 69:1109-1113.
McAleece, N., Lambshead, J., Paterson, G., Gage, J. (1997). Biodiversity Pro Beta 1. The
Natural History Museum / Scottish Association for Marine Science.
McDonald, D.W. (ed.). (2001). The New enciclopedia of mammals. Oxford University
Press, Oxford, UK.
Mellink, E. (1995). Uso del hábitat, dinámica poblacional y estacionalidad reproductiva de
roedores en el altiplano potosino, México. Revista Mexicana de Mastozoología 1:1-8.
Mendoza, M de los A. (1997). Efectos de la adición de alimento en la dinámica de
poblaciones y estructura de comunidades de pequeños mamíferos en un bosque
tropical caducifolio. Tesis de Maestría. UNAM, Facultad de Ciencias. México. 99 pp.
Mendoza, M de los A. (2002). Liomys pictus (Thomas 1893). Ratón espinoso de
abazones. (pp. 423-425). En: Noguera, F. A., Vega-Rivera, J. H., García-Aldrete A.
N. y Quesada-Avendaño, M. (Eds). Historia Natural de Chamela. Instituto de
Biología, Universidad Nacional Autónoma de México. México. 568pp.
Miller, G. T. jr. (1994). Ecología y medio ambiente. Séptima Edición. Grupo Editorial
Iberoamericano. México. 867 pp.
Miranda, F. & Hernández, E. (1963). Los tipos de vegetación de México y su clasificación.
Boletín de la Sociedad Botánica Mexicana. 28:45-55.
Morán, J. (1994). Estudio poblacional de Liomys irroratus torridus (rodentia: Heteromyidae)
en dos áreas con diferente grado de alteración en el sur de Morelos. Tesis de
licenciatura UAEM, Facultad de Ciencias Biológicas. México.
Mohammad, A. G., Pieper, R. P., Wallace, J. D., Holechek, J. L. & Murray, L, W. M. (1995). Comparison of fecal analysis and rumen evacuation techniques for sampling
diet botanical composition of grazing cattle. Journal of Range Management.
48(3):202-205.
Moro, D. & Bradshaw, S. D. (2002). Diets and predictions of feeding rate of house mice
and lakeland downs short-tailed mice inhabiting an arid island in western Australia.
Australian Journal of Zoology. 50:249-265.
Murphy, P.G., Lugo, A.E., (1986). Ecology of tropical dry forest. Annual Review of
Ecological Systems 17, 67–88.
Neu, C. W., Byers, C. R. & Peek, J. M. (1974). A technique for analysis of utilization-

115
availability data. Journal of Wildlife Management 38:541--545.
Norbury, G. L. & Sanson, G. D. (1992). Problems with measuring diet selection of
terrestrial mammalian herbivores (Review). Australian Journal of Ecology 17: 1-7
Olson, K. C. (1991). Diet sample collection by esophageal fistula and rumen evacuation
techniques. Journal of Range Management. 44(5):515-518.
Owen, M. (1975). An assessment of fecal analysis technique in waterfowl feeding studies.
Journal of Wildlife Management. 39(2):271-279.
Patton, J. L., & Rogers, D, S. (1993a). Cytogenetics (Capítulo 4: 236-258). En Genoways,
H. H. & Brown, J. H. (eds). Biology of the Heteromyidae. American Society of
Mammalogists special publication #10. 719pp.
Patton, J. L., & Rogers, D, S. (1993b). Biochemical genetics (Capítulo 5: 259-269). En
Genoways, H. H. & Brown, J. H. (eds). Biology of the Heteromyidae. American
Society of Mammalogists special publication #10. 719pp.
Peden, D. G., Hansen, R. M., Rice, R. W. & Van Dyne, G. M. (1974). A double sampling
technique for estimating dietary composition. Journal of Range Management.
27(4):323-325.
Petersen, M. K. (1993). Dietary overlap and livestock forage relationships in two species of
Sigmodon from Durango, México. (Pags: 289-300) En; Medellín, R. A. y Ceballos, G.
(eds). 1993. Avances en el estudio de los mamíferos de México. Publicaciones
Especiales, vol. 1, Asociación Mexicana de Mastozoología, A.C., México, D.F.
Pérez-Saldaña, G. A. (1978). Observaciones sobre la variación morfológica, alimentación y
reproducción de Liomys pictus pictus, Rodentia, Heteromyidae. Tesis de licenciatura.
UNAM, Facultad de Ciencias. México. 61 pp.
Feisinger, P., Spears, E. E. & Poole, R. W. (1981). A simple measure of niche breadth.
Ecology 62(1):27-32. Posse, G., Anchorena, J. & Collantes, M. B. (1996). Seasonal diets of sheep in the
steppe region of Tierra del Fuego, Argentina. Journal of Range Management.49(1):
24-30.
Preen, A. (1995). Diet of dugongs: are they omnivores? Journal of Mammalogy. 76(1):163-
171.
Putman, R. J. (1984). Facts from faeces. Mammal Reviews. 14(2): 79-97.

116
Pyke, G. H. (1984). Optimal foraging theory: a critical review. Annual Review of Ecology and
Systematics 15:523-575.
Reichman, O. J. (1975). Relation of desert rodent diets to available resources. Journal of
Mammalogy 56 (4): 731-751.
Reichman, O. J. (1977). Optimization of diets through food preferences by heteromyid
rodents. Ecology 58: 454-457
Reichman. O. J. & Price, M.V. (1993). Ecological aspects of heteromyid foraging. (Capítulo
14) En Genoways, H. H. & Brown, J. H. (eds). Biology of the Heteromyidae.
American Society of Mammalogists special publication #10.
Reichman, O. J. & Van de Graaf, K. M. (1975). Association between ingestión of green
vegetation and desert rodent reproduction. Journal of Mammalogy 56(2): 503-506.
Reppert, J. N. (1960). Forage preference and grazing habits of cattle at the eastern
Colorado range station. Journal of Range Management. 13(1):58-65.
Rogers, D. S. (1990). Genetic evolution, historical biogeography, and systematic
relationships among spiny pocket mice (subfamily Heteromyinae). Journal of
Mammalogy 71:668-685.
Rogers, P. & Dahlgren R. B. (1982). A comparison of prairie dog stomach contents and
feces using a microhistological technique. Journal of Wildlife Management 46(4):
1104-1108.
Romero-Almaraz, M. L., Sánchez-Hernández, C., García-Estrada C. & Owen, R. D. (2000). Mamíferos pequeños: Manual de técnicas de captura, preparación,
preservación y estudio. Primera Edición. Facultad de Ciencias UNAM, Instituto de
Biología UNAM. Centro de Investigaciones Biológicas, UAEM. México.
Rosenzweig, M. L. (1987). Habitat selection as a source of biological diversity. Evolutionary
Ecology 1:315-330.
Rosenzweig, M. L. (1991). Habitat selection and population interactions. American
Naturalist 137 (supplement):s5-s28
Rosenzweig, M. L. & Sterner, P. W. (1970). Populational ecology of desert rodent
communities: body size and seed-husking as bases for heteromyid coexistence.
Ecology 51:217-224.
Rudran, R. & Kunz, T. H. (1996). Ethics in research (apéndice 1 251:255) En: Wilson, D.

117
E., Cole, F. R., Nichols, J. D., Rudran, R. & Foster, M. S. (Editores). Measuring and
monitoring biological diversity: standard methods for mammals. Smithsonian Institute.
408pp.
Rumble, M. A. & Anderson, S. H. (1993). Evaluating the microscopic fecal technique for
estimating hard mast in turkey diets. USDA Forest Service Research paper RM-310.
Rzedowski, J. (1978). Vegetación de México. (pp. 189-205). Limusa. México.
Sánchez, H. C. & Romero, M de L. (1995). Mastofauna silvestre del área de reserva sierra
de Huautla. (con énfasis en la región noreste). Universidad Autónoma del Estado de
Morelos, Centro de Investigaciones Biológicas. México.
Sánchez-Cordero, V. & Fleming, T. H. (1993). Ecology of tropical heteromyids (capítulo
8:596-617). En Genoways, H. H. & Brown, J. H. (eds). Biology of the Heteromyidae.
American Society of Mammalogists special publication #10. 719pp.
Sanders, K. D., Dahl, B. E., Scott, G. (1980). Bite-count vs. fecal analysis for range animal
diets. Journal of Range Management. 33(1):146-149.
Sandoval-Zapotitla, E. (En prensa). técnicas aplicadas al estudio de la anatomía vegetal.
UNAM, Jardín Botánico, Instituto de Biología. México.
SAS Institute inc. (1997). JMP. 3.2.2. (programa estadístico1989-1997).
Schmidly, D. J., Wilkins, K. T. & Derr, J. N. (1993). Biogeography. (capítulo 8:319-385).
En Genoways, H. H. & Brown, J. H. (eds). Biology of the Heteromyidae. American
Society of Mammalogists special publication #10. 719pp.
Selko, L. F. (1937). Food habits of Iowa skuns in the fall of 1936. Journal of Wildlife
Management 1:70-76.
Setoguchi, M. (1990). Food habits of red-bellied tree squirrels on a small island in Japan.
Journal of Mammalogy 71(4): 570-578.
Shaw, W. T. (1934). The ability of the giant kangaroo rat as a harvester and storer of seeds.
Journal of mammalogy 15 (4): 275:286.
Smigel, B. W. & Rosenzweig, M. L. (1974). Seed selection in Dipodomys merriami and
Perognathus penicillatus. Ecology 55(2): 329-339.
Smith D. G., Gordon, I. J., Mpendu B. F., Lamani, M. B., Dziba, L. & Kelly, E.(2002). Evaluation of some recently developed techniques for estimating diet composition in
browsing herbivores. (Página de Internet)

118
(www.fao.org/DOCREP/ARTICLE/AGRIPPÄ/522_EN.HTM).
Smith, A. D. & Hubbard, R. L. (1954). Preference ratings for winter deer forages from
northern Utah ranges, based on browsing time and forage consumed. Journal of
Range Management. 7(2):262-264.
Smith, R. L. & Smith, T. M. (2001). Ecología. Cuarta edición. Pearson.
Sparks, D. R. (1968). Diet of black-tailed jackrabbits on sandhill rangeland in Colorado.
Journal or Range Management. 21(2):203-208.
Sparks, D. R. & Malechek, J. C. (1968). Estimating percentage dry weight in diets using a
microscopic technique. Journal of Range Management. 21(2):264-265.
Sperry, C. C. (1934). Winter food habits of coyote: a report of progress 1933. Journal of
Mammalogy 15(4): 286-296.
Stancampiano, A. J. & Caire, W. (1995). Food habits of Peromyscus and Reithrodontomys
in the Wichita Mountains Wildlife Refuge, Oklahoma. Proceedings of the Oklahoma
Academy of Science 75:45-49.
Stewart, D. R. M. (1967). Analysis of plant epidermis in faeces: a technique for studying
food preferences of grazing herbivores. Journal of Applied Ecology. 4(1):83-111.
Swihart, R. K., Gehring, T. M., Kolozsvary, M. B. & Nupp, T. E. (2003). Responses or
“resistant” vertebrates to habitat loss and fragmentation: the importance of niche
breadth and range boundaries. Diversity and Distributions 9:1-18.
Taylor, W. P. (1954). Food habits and notes on life history of the ring-tailed cat in Texas.
Journal of Mammalogy 35(1): 55-63.
The Association for the Study of Animal Behaviour. (2000). Guidelines for the treatment
of animals in behavioural research and teaching. Animal Behaviour 59: 253-257.
Todd, J. W. & Hansen, R. M., (1973). Plant fragments in the feces of bighorns as indicators
of food habits. Journal of wildlife Management 37(3): 363-366.
Torres-Contreras, H. & Bozinovic, F. (1997). Food selection in an herbivorous rodent:
balancing nutrition with thermoregulation. Ecology 78(7): 2230-2237.
Uresk, D. W. (1984). Black-tailed prairie dog food habits and forage relationships in western
South Dakota. Journal of Range Management. 37(4):325-329.
Urness, P. J., Smith, A. D. & Watkins, R. K. (1977). Comparison of in vivo and in vitro dry
matter digestibility of mule deer forages. Journal of Range Management.30(1):119-

119
121.
Vandermeer, J. H. (1979). Hoarding behaviour of captive Heteromys desmarestianus,
(Rodentia) on the fruits of Welfia georgii, a rainforest dominant palm in Costa Rica.
Brenesia 16:107-116.
Van Dyne, G. M. & Torell, D. T. (1964). Development and use of the esophageal fistula: a
review. Journal of Range Management. 17(1):7-18.
Vavra, M. & Holechek, J. L. (1980). Factors influencing microhistological analysis of
herbivore diets. Journal of Range Management. 33(5):371-374.
Vavra, M., Rice, R. W. & Hansen, R. M. (1978). A comparison of esophageal fistula and
fecal material to determinate steer diets. Journal of Range Management. 31(1):11-13.
Vázquez, L. B. (1997). Dieta y demografía de una comunidad de pequeños roedores en
dos hábitat contrastantes. Tesis de licenciatura. Universidad de Guadalajara.
División de Ciencias Ambientales. Jalisco, México.
Vázquez, L. B., Cameron, G. N. y Medellín, R. A. (2000). Hábitos alimentarios y biología
poblacional de dos especies de roedores en el occidente de México. Revista
Mexicana de Mastozoología 4:5-21.
Villa, B. & Cervantes, F. (2003). Los mamíferos de México. Grupo Editorial
Iberoamericana, Instituto de Biología, UNAM. México.
Von Ende, C. N. (1993). Repeated-measures analysis: Growth and other time-dependent
measures (capítulo 6 pp 113-137). En: Scheiner, S. M. & Gurevitch, J. Design and
analysis of ecological experiments. Chapman & Hall. USA. pp. 445.
Voth, E. H. & Black, H. C. (1973). A histologyc technique for determining feeding habits of
small herbivores. Journal of Wildlife Management. 37(2): 223-231.
Ward, A. L. & Keith, J. O. (1962). Feeding habits of pocket gophers on mountain
grasslands, Black Mesa, Colorado. Ecology. 43 (4) 744-749.
Weckerly, F. W. (1994). Selective feeding by black-tailed deer: forage quality or
abundance? Journal of Mammalogy. 75:905-913.
Westoby, M., Rost, G. R. & Wels, J. A. (1976). Problems with estimating herbivore diets
by microscopically identifying plant fragments from stomachs. Journal of
Mammalogy. 57(1):167-172.
Whitaker, J. O. (1966). Foods of Mus musculus, Peromyscus maniculatus bairdi and

120
Peromyscus leucopus in Vigo county, Indiana. Journal of Mammalogy. 47(3): 473-
488.
Williams, O. B. (1969). An Improved technique for identification of plant fragments in
herbivore feces. Journal of Range Management 22(1):51-52.
Wilson, D.L. & Bertram, J. D. (1987). Examination of faeces for the identification of plant
species in the diet of cattle. Technical note No. 046 May 1987 Agdex No. 420/59
ISSN No. 0158/2755.
Wondolleck, J. T. (1978). Forage-area separation and overlap in heteromyid rodents.
Journal of Mammalogy 59(3):510-518.
Zyznar, E. & Urness, P, J. (1969). Qualitative identification of forage remnants in deer
feces. Journal of Wildlife Management. 33(3):506-510.

121
Tablas y Anexos
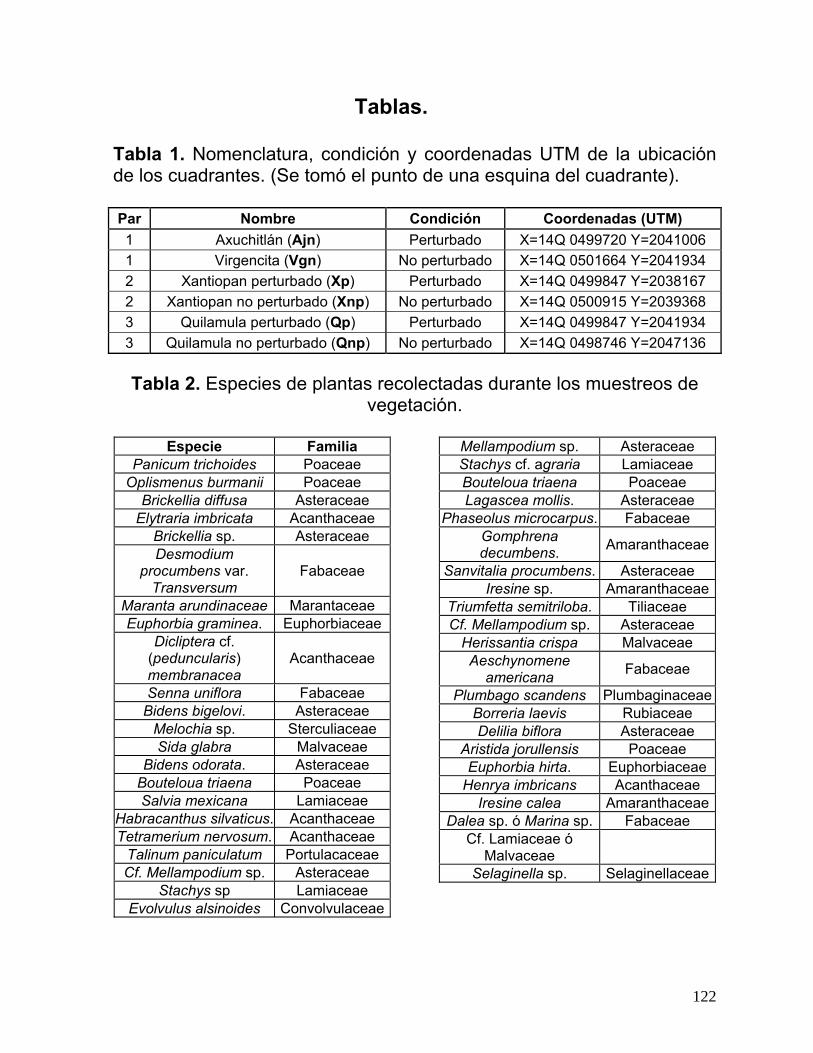
122
Tablas.
Tabla 1. Nomenclatura, condición y coordenadas UTM de la ubicación de los cuadrantes. (Se tomó el punto de una esquina del cuadrante). Par Nombre Condición Coordenadas (UTM) 1 Axuchitlán (Ajn) Perturbado X=14Q 0499720 Y=2041006 1 Virgencita (Vgn) No perturbado X=14Q 0501664 Y=2041934 2 Xantiopan perturbado (Xp) Perturbado X=14Q 0499847 Y=2038167 2 Xantiopan no perturbado (Xnp) No perturbado X=14Q 0500915 Y=2039368 3 Quilamula perturbado (Qp) Perturbado X=14Q 0499847 Y=2041934 3 Quilamula no perturbado (Qnp) No perturbado X=14Q 0498746 Y=2047136
Tabla 2. Especies de plantas recolectadas durante los muestreos de
vegetación.
Especie Familia Panicum trichoides Poaceae
Oplismenus burmanii Poaceae Brickellia diffusa Asteraceae
Elytraria imbricata Acanthaceae Brickellia sp. Asteraceae Desmodium
procumbens var. Transversum
Fabaceae
Maranta arundinaceae Marantaceae Euphorbia graminea. Euphorbiaceae
Dicliptera cf. (peduncularis) membranacea
Acanthaceae
Senna uniflora Fabaceae Bidens bigelovi. Asteraceae
Melochia sp. Sterculiaceae Sida glabra Malvaceae
Bidens odorata. Asteraceae Bouteloua triaena Poaceae Salvia mexicana Lamiaceae
Habracanthus silvaticus. Acanthaceae Tetramerium nervosum. Acanthaceae
Talinum paniculatum Portulacaceae Cf. Mellampodium sp. Asteraceae
Stachys sp Lamiaceae Evolvulus alsinoides Convolvulaceae
Mellampodium sp. Asteraceae Stachys cf. agraria Lamiaceae Bouteloua triaena Poaceae Lagascea mollis. Asteraceae
Phaseolus microcarpus. Fabaceae Gomphrena decumbens. Amaranthaceae
Sanvitalia procumbens. Asteraceae Iresine sp. Amaranthaceae
Triumfetta semitriloba. Tiliaceae Cf. Mellampodium sp. Asteraceae
Herissantia crispa Malvaceae Aeschynomene
americana Fabaceae
Plumbago scandens PlumbaginaceaeBorreria laevis Rubiaceae Delilia biflora Asteraceae
Aristida jorullensis Poaceae Euphorbia hirta. Euphorbiaceae
Henrya imbricans Acanthaceae Iresine calea Amaranthaceae
Dalea sp. ó Marina sp. Fabaceae Cf. Lamiaceae ó
Malvaceae
Selaginella sp. Selaginellaceae

123
Tabla 3. Especies recolectadas en el banco de semillas
Morfo 3 Morfo 6 Morfo 9
Morfo 12 Morfo 14 Morfo 19 Morfo 36 Morfo 39 Morfo 41 Morfo 43 Morfo 44 Morfo 46 Morfo 47 Morfo 51 Morfo 56 Morfo 57 Morfo 58 Morfo 73 Morfo 81 Morfo 83 Morfo 85 Morfo 87 Morfo 91
Justicia sp. Acanthaceae Ruellia sp. Acanthaceae
Amphipterygium adstringens Anacardiaceae
Spondias purpurea Anacardiaceae Stemmadenia bella Apocynaceae
Marsdenia sp. Asclepiadaceae cf. Bidens sp. Asteraceae Delilia biflora Asteraceae Montanoa. cf.
grandifolia Asteraceae
Morfo 59 Asteraceae Pithecoctenium
crucigerum Bignoniaceae
Ceiba aesculifolia Bombacaceae Bursera aloexylon Burseraceae Bursera copallifera Burseraceae Bursera grandifolia Burseraceae
Opuntia sp. Cactaceae Pachycereus grandis Cactaceae
Morfo 74 cf. Fabaceae Morfo 52 cf. Rubiaceae. Morfo 37 Cf. Solanaceae.
Combretum fructicosum Combretaceae
Ipomoea conzattii Convolvulaceae Ipomoea pauciflora Convolvulaceae
Ipomoea sp 1 Convolvulaceae Ipomoea sp 2 Convolvulaceae Ipomoea sp. 3 Convolvulaceae
Morfo 86 Euphorbiaceae. Acacia cochliacantha Fabaceae
Acacia farnesiana Fabaceae Acacia sp. Fabaceae
Aeschynomene americana Fabaceae
cf. Aeschynomene americana Fabaceae
Chamaecrista sp. Fabaceae Conzattia multiflora. Fabaceae
Desmanthus virgatus. Fabaceae Gliricidia sepium Fabaceae
Lysiloma divaricata Fabaceae Mimosa benthamii Fabaceae Mimosa poliantha Fabaceae
Mimosa sp. 1 Fabaceae Mimosa sp. 2 Fabaceae
Morfo 5 Fabaceae Morfo 50 Fabaceae
Nissolia fruticosa. Fabaceae Senna sp 3. Fabaceae Senna sp. 1 Fabaceae Senna sp. 2 Fabaceae Senna sp. 4 Fabaceae Hippocratea acapulcensis Hippocrateaceae
Mascagnia aff. polybotrya Malpighiaceae
Ficus padifolia Moraceae Ficus sp. Moraceae
Polygonum sp. Polygonaceae Ruprechtia fusca Polygonaceae
Zanthoxylum culantrillo Rutaceae Serjania triquetra Sapindaceae
Guazuma ulmifolia. Sterculiaceae Heliocarpus pallidus Tiliaceae
Triumfetta semitriloba. Tiliaceae Vitex hemsleyi. Verbenaceae
Vitex mollis Verbenaceae

124
Tabla 4. Especies de semillas recolectadas de los abazones.
Morfo 3 Morfo 6 Morfo 8 Morfo 9
Morfo 12 Morfo 14
Morfo 15B Morfo 19
Morfo 20B Morfo 31 Morfo 33
Justicia sp. Acanthaceae Stemmadenia bella Apocynaceae
Pithecoctenium crucigerum Bignoniaceae
Bursera aloexylon Burseraceae Bursera copallifera Burseraceae Bursera grandifolia Burseraceae Jacaratia mexicana Caricaceae
Ipomoea pauciflora Convolvulaceae Ipomoea sp. 1 Convolvulaceae
Cochlospermum vitifolium Cochlospermaceae
cf. Aeschynomene sp. Fabaceae Conzattia multiflora Fabaceae
Desmanthus sp. Fabaceae Lysiloma divaricata Fabaceae
Mimosa sp. Fabaceae Morfo 5 Fabaceae
Senna sp. Fabaceae Polygonum sp. Polygonaceae
Ruprechtia fusca Polygonaceae Zanthoxylum cf.
culantrillo Rutaceae
Serjania triquetra. Sapindaceae Guazuma ulmifolia. Sterculiaceae
Triumfetta semitriloba Tiliaceae
Tabla 5. Resultados del análisis de preferencia para las semillas de diferentes especies encontradas en los contenidos de abazones, por condición por temporada y total. El número indica el lugar relativo de preferencia en el que se encuentra la especie por cada tratamiento. P=perturbado Np=no perturbado Ll=lluvias S1=secas tempranas S2= secas tardías. Especie PLl NpLl PS1 NpS1 PS2 NpS2 Total Conzattia multiflora 4 2 2 1 8 1 Bursera grandifolia 2 5 8 2 6 2 5 3 2 3 Ipomoea sp 1 1 1 4 3 1 4 3 9 5 6 4 3 3 6 Serjania triquetra 5 1 2 5 7 Desmanthus virgatus 8 Lysiloma divaricata 9 Stemmadenia bella 10 9 6 5 4 11 Bursera aloexylon 3 1 12 12 13 Mimosa sp 1 7 10 14 Zanthoxylum culantrillo 15

125
cf.Aeschynomene americana 16 14 17 Senna sp. 18 Triumfetta semitriloba 7 19 Pithecoctenium crucigerum 7 20 Mimosa sp. 2 21 Bursera copallifera 3 22 Ipomoea pauciflora 6 23 Ruprechtia fusca 4 24 Justicia candicans 25 19 4 26 Tabla 6. Especies de plantas herbáceas identificadas dentro de las excretas, incluye cuales tienen datos de diversidad y disponibilidad en el medio.
Familia Especie Datos disponibilidad Acanthaceae Elytraria imbricata Si Acanthaceae Habracanthus silvaticus Si Acanthaceae Justicia candicans Si Acanthaceae Tetramerium nervosum Si Amaranthaceae Gomphrena decumbens Si Fabaceae Desmodium procumbens Si Marantaceae Maranta arundinaceae Si Onagraceae Ludwigia octovalvis No Plumbaginaceae Plumbago scandens No Poaceae Oplismenus burmannii Si Tabla 7. Resultados del análisis de preferencia para las especies encontradas en excretas, por condición por temporada y total. El número indica el lugar relativo de preferencia en el que se encuentra la especie por cada tratamiento. P=perturbado, Np=no perturbado, Ll=lluvias, S1=Secas iniciales, S2=secas finales, SD= Sin datos de disponibilidad.
Especie Familia PLl NpLl PS1 NpS1 PS2 NpS2 TotalOplismenus burmannii Poaceae 1 1 1 1 1 1 1 Gomphrena decumbens Amaranthaceae 2 Justicia candicans Acanthaceae 3 Maranta arundinaceae Marantaceae 2 4 Habracanthus silvaticus Acanthaceae 3 5 Desmodium procumbens Fabaceae 2 2 6 Tetramerium nervosum Acanthaceae 3 7 Elytraria imbricata Acanthaceae 3 2 8 Ludwigia octovalvis Onagraceae SD Plumbago scandens Plumbaginaceae SD

126
Tabla 8. Géneros y especies cuyas semillas se reportan como presentes en abazones o madrigueras de Liomys pictus en las selvas secas de Chamela, Jalisco y que estan presentes en la Sierra de Huautla (Dorado et al., en prensa) y fueron encontrados para Liomys irroratus en el presente trabajo. 1.- Este trabajo 2.-Pérez-Saldaña, 1978 3.- Janzen, 1986a, 4.-Ceballos, 1989; 5.-Mendoza, 1997. 6.- Domínguez, 2000. 7.- Hernández, 2000. *son las spp no reportadas para L. pictus y descritas para L. irroratus en este trabajo. Los datos de Janzen (1986a) son de contenidos de abazones de Liomys salvini en Costa Rica.
Familia especie 1 2 3 4 5 6 7 Acanthaceae Dicliptera sp. X Acanthaceae Justicia candicans* X Acanthaceae Justicia sp. X Acanthaceae Ruellia sp. X X X Anacardiaceae Comocladia engleriana X X X X Anacardiaceae Spondias purpurea X X X X Apocynaceae Stemmadenia bella* X Bignoniaceae Crescentia alata X X Bignoniaceae Pithecoctenium crucigerum* X Bignoniaceae Tabebuia sp. X Bombacaceae Ceiba aesculifolia X X Boraginaceae Cordia elaeagnoides X X Boraginaceae Cordia sp. X Burseraceae Bursera aloexylon* X Burseraceae Bursera copallifera* X Burseraceae Bursera grandifolia* X Cactaceae Opuntia sp. X Capparaceae Cleome sp. X Capparaceae Crataeva tapia X X Caricaceae Jacaratia mexicana X X X Cochlospermaceae Cochlospermum vitifolium X X X X Combretaceae Combretum fruticosum X Convolvulaceae Evolvulus sp. X X Convolvulaceae Ipomoea arborescens X Convolvulaceae Ipomoea pauciflora* X Convolvulaceae Ipomoea sp. X X X X X X Convolvulaceae Ipomoea wolcottiana X X X Cucurbitaceae Echinopepon racemosus X X Cucurbitaceae Cucurbita sp. X

127
Euphorbiaceae Cnidoscolus sp. X X Euphorbiaceae Cnidoscolus tubulosus Euphorbiaceae Croton sp. X X X X Euphorbiaceae Euphorbia sp. X X Euphorbiaceae Jatropha sp. X X Euphorbiaceae Manihot sp. X Fabaceae Acacia farnesiana X X Fabaceae Bauhinia sp. X Fabaceae Caesalpinia pulcherrima X X X Fabaceae cf. Aeschynomene americana* X Fabaceae Conzattia multiflora* X fabaceae Crotalaria sp. X X X Fabaceae Desmanthus virgatus* X Fabaceae Entadopsis polystachya X X X Fabaceae Enterolobium cyclocarpum X Fabaceae Hymenaea courbaril X Fabaceae Leucaena lanceolata X Fabaceae Leucaena sp. X Fabaceae Lonchocarpus sp. X X Fabaceae Lysiloma divaricata X X Fabaceae Mimosa sp. X X X Fabaceae Nissolia fruticosa X X X X X Fabaceae Phaseolus lunatus X X X X X Fabaceae Phaseolus microcarpus X X X X Fabaceae Phaseolus sp. X X X Fabaceae Phaseolus vulgaris X Fabaceae Tephrosia sp. X X Fabaceae Senna sp.* X Malvaceae Abutilion sp. X Malvaceae Sida sp. X X Meliaceae Swetenia humilis X Moraceae Ficus cotinifolia X Moraceae Ficus sp. X X X X X X Phytolaccaceae Rivinia humilis X Plumbaginaceae Plumbago scandens X Poaceae Lasiacis ruscifolia X Poaceae Panicum sp. X X X Poaceae Setaria sp. X Polygonaceae Ruprechtia fusca X X X Rubiaceae Randia sp. X X Rutaceae Zanthoxylum culantrillo* X

128
Sapindaceae Cardiospermum halicacabum X X Sapindaceae Serjania brachycarpa X X X Sapindaceae Serjania sp. X Sapindaceae Serjania triquetra* X Sapindaceae Thouinidium decandrum X X Solanaceae Physalis leptophylla X X Solanaceae Solanum sp. X X X Sterculiaceae Guazuma ulmifolia X X Theophrastaceae Jacquinia pungens X X Tiliaceae Heliocarpus pallidus X X X X Tiliaceae Triumfetta semitriloba X
27 81 23 17 3 34 29 26 29

129
Anexos
Anexo 1. Medidas y datos tomados a los ratones capturados.
De todos los animales capturados se registró el número de trampa, la fecha
y sitio de captura y los siguientes datos (Romero-Almaraz et al., 2000):
1. Identificación a nivel especie utilizando las guías de Álvarez-Castañeda
(1996); Ceballos & Miranda (2000); Sánchez & Romero (1995).
2. Datos morfométricos: Longitud total (LT), Longitud de cola (LC), Longitud de la
pata izquierda trasera (LPI), Longitud de la oreja izquierda (LOI), en milímetros y
peso en gramos.
3. Sexo, clase de edad (ej. cría, juvenil o adulto) y condición reproductiva (ej.
inactivo, con testículos escrotados, preñada y en lactancia).
4. Contenido de los abazones y muestras de excretas (de cada individuo
incluyendo recapturas) para la estimación de la dieta de Liomys irroratus
De manera complementaria se recolectaron muestras de ectoparásitos, pelo y
tejido, para la realización de estudios posteriores.
Además, los animales se marcaron por medio de ectomización de falanges
(Martoff, 1963 citado en Romero-Almaraz et al., 2000) para permitir su identificación
posterior y la obtención de información demográfica y para obtener muestras de
tejido, necesario para los objetivos del proyecto general dentro del cual se inserta
este trabajo. Los animales fueron devueltos el día siguiente al lugar exacto de la
trampa donde fueron capturados.

130
Anexo 2. Fórmula del formaldehído alcohol; (F. A. A.).
(Gaviño et al., 1999).
Alcohol de 96°.
Ácido acético glacial.
Formol comercial.
Agua destilada
50ml.
5ml.
10ml.
35ml.
Anexo 3. Fórmula de la gelatina glicerinada. (Gaviño et al., 1999; Sandoval-
Zapotitla, en prensa).
Gelatina (grenetina) 3.5gr.
Glicerina 25 ml.
Ácido fénico 1 gr.
Agua destilada. 21 ml.
Disolver la gelatina en agua caliente, cuando esté tibia, agregar las otras
sustancias. Filtrar utilizando un lienzo fino. Esta solución se debe conservar en
frascos pequeños y en refrigeración. Antes de utilizar se debe fundir en baño María
(o en microondas por 10 segundos) evitando la formación de burbujas.
Anexo 4. Técnica para la preparación de laminillas temporales. (Sandoval-Zapotitla, en prensa).
1. Cortar secciones de aproximadamente 1cm2 del material fijado con F. A. A.
2. Colocar las secciones en frascos con NaOH al 20 o 30%. Hasta que la sección
tenga una apariencia translúcida (normalmente toma entre 20 y 40 minutos).
3. Lavar con agua destilada, al menos dos veces.
4. Colocar las secciones el cloro comercial en concentración similar a la anterior
solución y permanecerán ahí por un mínimo de 30 minutos, hasta que las secciones

131
se tomen blanquecinas (transparentes). Esto es un buen indicio de que la
separación de la cutícula/epidermis será más fácil.
5. Lavar las secciones en repetidas ocasiones con agua destilada para
eliminar el exceso de cloro.
6. Colocar la muestra en un portaobjetos, agregando de tres a cinco gotas de
gelatina glicerinada.
7. Cubrir la laminilla con un cubre objetos y colocarle un peso sobre el para
mantener la muestra en un solo plano, dejándola secar hasta que la gelatina
glicerinada esté totalmente sólida.
Anexo 5. Técnica para la preparación de laminillas permanentes. (Sandoval-Zapotitla, en prensa).
Esta técnica permite observar la forma y tamaño de las células epidérmicas,
orientación de las mismas, ornamentaciones epidérmicas o cuticulares, tricomas,
papilas u otros apéndices epidérmicos, estomas y cristales.
1. Cortar secciones de aproximadamente 1cm2 del material fijado con F. A. A.
2. Colocar las secciones en frascos con NaOH al 20 o 30%. Hasta que la sección
tenga una apariencia translúcida (normalmente toma entre 20 y 40 minutos).
3. Lavar con agua destilada, al menos dos veces.
4. Colocar las secciones en cloro comercial en concentración similar a la anterior
solución y permanecerán ahí por un mínimo de 30 minutos, hasta que las secciones
se tornen blanquecinas (transparentes). Esto es un buen indicio de que la
separación de la cutícula/epidermis será más fácil.
5. Lavar las secciones en repetidas ocasiones con agua destilada para
eliminar el exceso de cloro.
6. Colocar el material en cajas Petri y bajo el microscopio estereoscópico tratar de
separar la cutícula con ayuda de pinzas de punta fina y agujas de disección. Eliminar

132
el tejido adherido a la cutícula con ayuda de un pincel de cerdas finas, cuidando que
esta no se rompa o dañe.
7. Cuando la cutícula está totalmente desprovista de tejido parenquimático se
procede a la tinción. Para evitar que la cutícula se enrolle y sea más fácil su manejo,
se recomienda colocar en un portaobjetos una gota de gelatina glicerinada
diluida con agua. Extenderla en una fina capa y colocar sobre esta la cutícula;
esperar a que la gelatina seque.
8. Asegurarse que la superficie expuesta hacia arriba sea la externa de la
epidermis. Agregar unas gotas de Safranina "O" por un mínimo de 15 minutos.
9. Lavar con agua destilada el excedente de colorante. Deshidratar gradualmente
con etanol al 30, 50, 70, 96% y absoluto, dos cambios en cada alcohol de 1 a 3
minutos en cada uno.
10. Agregar unas gotas de aceite de clavo durante 10 minutos para lograr su
aclaramiento. Lavar el aceite con tres cambios de xilol, un minuto o dos en cada
cambio y montar en resina sintética (o Permount).
Nota: ésta metodología fue modificada para optimizar su funcionamiento con
el material recolectado (herbáceas), el cual al ser delgado requirió de menores
tiempos que el material más grueso (hojas de árboles) por lo que se requiere de un
mayor tiempo en los diferentes reactivos, tiempos que el material del presente
trabajo no puede estar inmerso en dichos químicos pues presenta daño celular.
En múltiples ocasiones se omitieron los pasos 6 y 7, por no haber sido posible
la separación de la epidermis, por lo mismo se tiñó la sección (±1cm2) completa,
dando resultados muy similares al material del cual la epidermis fue separada.

133
Anexo 6. Técnica para la preparación de las laminillas de las excretas.
(Holechek, 1982).
Esta es la técnica utilizada para la preparación de las laminillas de material fecal:
1. Secado del material en horno, entre 60ºC y 70ºC, hasta estabilizar el peso (Hinnart &
Kothman, 1988).
2. El material es molido a mano y cribado a través de una malla de 1mm, antes de
iniciar el tratamiento.
3. El material se coloca en blanqueador casero (hipoclorito de sodio) durante 30
minutos.
4. El material se hierve en agua destilada durante 2 minutos.
5. Se monta en la laminilla con Permount.
Ésta metodología se modificó de Holechek (1982) pues dentro del equipo
necesario se requiere un molino de cuchillas (Wiley mill) para moler las excretas a
1mm, equipo que no se encuentra disponible para este trabajo por lo tanto el
material se maceró a mano a un tamaño que los fragmentos coincidieran con las
dimensiones sugeridas por el trabajo de Holechek (1982), pasándolas a través de
malla de 1mm, esperando poder acceder a ese equipo posteriormente y comparar
los resultados de ambos métodos.
La inmersión en NaOH es recomendada por Holechek (1982) y Vavra &
Holechek (1980, recomendando también el uso del molino de cuchillas en conjunto
con el aclarador. La aplicación de Hipoclorito de sodio (blanqueador casero) se
recomienda en Holechek (1982) y en Williams (1969), a pesar de que ambas son
factibles de generar destrucción en las células, por lo tanto se recomienda probar los
resultados de ambas y compararlos (Holechek, 1982).
La temperatura óptima de secado del material fecal se obtuvo del trabajo de
Hinnant & Kothmann (1988), donde se señala que el secado a esas temperaturas se
puede utilizar para proveer un método rápido y eficaz para analizar el material fecal,
dado a que Holechek (1982) no detalla la temperatura a la que se deben secar las
excretas.





























