Embed Size (px)
Citation preview

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
ESTUDIO DE LA ACTIVIDAD ANTIMICROBIANA Y ANTIOXIDANTE DEL EXTRACTO
ETANÓLICO DEL ALGA Padina mexicana (PHAEOPHYCEAE: DICTYOTALES).
TESIS
Que, como uno de los requisitos para obtener el título de
BIÓLOGO MARINO
PRESENTA
ELENA STEPHANIE CASTRO SILVA
La Paz, Baja California Sur, Octubre del 2010.

Agradecimientos
A mi familia, principalmente a mi bella madre Clara Silva Geraldo por su apoyo y
comprensión, así también a mis queridas hermanas Paola y Yulissa Cota Silva por aguantar mis
malos ratos de estrés.
A mis preciosas Fernanda y Nubia Carballo Ramírez (mis cuyitos) por su atención, aliento y
apoyo incondicional en todo y siempre, a Sergio Negrete por cuidar de mis niñas y brindar su
apoyo; a mi flaca hermosa Cynthia Arrevillaga por escucharme y aguantar mis rabietas durante el
estrés de la escuela, por ayudarme y estar conmigo. A mi loca bella Yazkara Olachea (la Kazkara)
por retornar a mi vida y brindarme su amistad.
A mi novio precioso Mario Vergara por su ayuda, amor y comprensión en todo momento
así como a su hermosa familia que me ha abierto las puertas de su casa con cariño.
A mis amigos(as) y compañeros(as) de la carrera, Bernardo Sánchez, Joel Soto, José
Manuel Marrón, Ceacatl Arce, Uriel Valdez, Diego Rivera, Sara de las Heras, Merit Mora, Diana
Zaleta, Valerie Schnoler, Pablo Juárez, etc.. que sin sus constantes carrillas, buen humor y apoyo
no hubiese disfrutado tanto estar ahí. Los compas del Cbtis # 230, Sergio Morales Lucero (Mi hijo),
Sarahi Quintero, Juan Manuel Castro, Jorge Pratt, Aaron Chihuahua, Carmen Camacho, Jorge
Ochoa y Christian Higuera, que en su momento me regalaron palabras de aliento y apoyo así como
buenos recuerdos generados en todo este tiempo compartido.
A mis asesores, Dr. Iván Murillo Álvarez, M.C. Mauricio Muñoz Ochoa y Dr. Rafael
Riosmena Rodríguez por toda la ayuda, paciencia y comprensión que me han brindado..¡sobre
todo por aguantarme! A mis compañeros en el Laboratorio de Química de Algas Marinas CICIMAR,
M.C. Elizabeth Rodríguez Montesinos, M.C. Dora Arvizu Higuera, Dr. Gustavo Carmona por toda
su ayuda, y paciencia.
A mis profesores, principalmente a B.M. Marco Antonio Medina López, Dra. Rosalba
Encarnación Dimayuga, Dr. Héctor Reyes, Dr. Volker Kotch, etc. por sus extraordinarias clases que
me dieron el entusiasmo para seguir y terminar la carrera.
Al Centro Interdisciplinario en Ciencias Marinas (CICIMAR-IPN) por prestar sus
instalaciones para llevar a cabo todo el proceso de experimentación, así también al proyecto
CONACYT (ref.: SEP-47942/A-1) e IPN (ref.: CGPI: 20050324, SIP-20060049, SIP-20070020 y
SIP-20082016) por financiar dicho proyecto de donde se deriva el presente trabajo.
A los que han creído en mí, que me han dado palabras de aliento en los momentos que me
sentí flaquear, a los que han compartido conmigo y he olvidado mencionar.. ¡Muchas gracias!

Dedicatoria
A todos aquellos que me han visto crecer, a los que se unieron para verme florecer. A
todos los que en algún momento estuvieron en mi vida y tuvieron que marcharse. A los que me
quieren, a los que me odian. A esos que dijeron que no podría, a los que sinceramente me apoyan.
A todos, les dedico lo siguiente:
Y ella es así, torrente de fuego apasionada. De lustrosos ojos café desesperantes. llorona en naturaleza de la lluvia, seca cuando la arena del desierto le sugiere. Amante de lo bellamente enternecido, imprudente como la ola de tormenta, dócil tal cordero domesticado o fiera leona tras presa sin distintivo. Qué al asno a veces se asemeja, floja –rebuzna entre fantasías de algodón-; se levanta luego y anda-sagaz e inteligente-, ¡águila en el vuelo de plumas rojas!. El corazón le sangra si el amado sufre, su bien antepone antes del propio; vida en guerra daría codo a codo si felicidad para él consiguiere. Pliega sonrisa con la risa de los otros esos que se burlan y le subestiman, no se imaginan que pudiere levantar una montaña entre sus hombros.
Piel trigueña, caderas anchas; me susurra al oído amor, y como bruma se levanta. Soñadora loca, enamorada empedernida, le amo a esa pelirroja desentonada.
....Y así será ella, mientras el aire siga entrando a sus pulmones y el corazón le palpite tan fuerte como las olas en un rompiente... loca..cuerda..floja..inteligente..de vez en cuando distraída e imprudente..empalagozamente dulce y enojona amargosamente ...así es como es.. y no le necesito diferente.

i
Índice general página
Índice general. i Lista de tablas y figuras. ii Lista de gráficos. iii Lista de abreviaciones. iv Glosario. v Resumen. vii Abstract. viii 1. Introducción. 1 2. Antecedentes. 2 2.1. Polifenoles. 2 2.2. Terpenos. 5 2.3. Mecanismo del radical libre estable 2,2-difenil-1-picrilhidracilo (DPPH) y la actividad antioxidante.
7
2.4. Actividad antimicrobiana de las algas pardas. 12 3. Justificación. 17 4. Descripción y distribución de Padina mexicana. 18 4.1. Clasificación taxonómica de Padina mexicana. 20 5. Objetivo general. 21 5.1. Objetivos particulares. 21 6. Materiales y métodos. 22 6.1. Trabajo en campo. 22 6.2. Trabajo de laboratorio. 23 6.2.1. Obtención del extracto crudo. 23 6.2.2. Fraccionamiento del extracto crudo. 23 6.2.3. Actividad antimicrobiana. 26 6.2.3.1. Ensayo de actividad antimicrobiana. 26 6.2.3.2. Microorganismos de prueba 26 6.2.3.3. Medios de cultivo e inóculos. 27 6.2.3.4. Preparación de discos. 27 6.2.4. Pruebas de actividad antioxidante. 28 6.2.4.1. Método espectrofotométrico. 28 6.2.4.2. Método bioautográfico. 28 7. Resultados. 28 7.1. Cromatografía de capa fina (TLC) y fraccionamiento en columna cromatográfica del extracto etanólico de Padina mexicana.
28
7.2. Análisis de actividad antimicrobiana. 31 7.3. Pruebas de actividad antioxidante. 33 7.3.1. Ensayo con el radical libre estable 2,2-difenil-1-picril hidracilo (DPPH). 33 7.3.1.1. Análisis espectrofotométrico. 33 7.3.1.2. Análisis bioautográfico. 34 8. Discusión. 40

ii
9. Conclusiones. 43 10. Literatura citada. 45
Lista de tablas y figuras
página
Figura 1. Ruta biosintética de los phlorotaninos. 3
Figura 2. Derivados de los phlorotaninos identificados en varios géneros y especies de
algas pardas. Donde (1) phloroglucionol, (2) eckol, (3) dieckol, (4) 8,8´-bieckol y (5)
phlorofucofuroeckol A, reproducido de Kang (2004).
4
Figura 3. Unidades de isopreno y estructura reproducido de Dewick, (2002). 5
Figura 4. Ruta biosintética de los terpenoides reproducido de Dewick, (2002). 6
Figura 5. Estructura del radical libre 2,2-difenil-1-picrilhidracilo (DPPH) y mecanismo de
acción reproducido de Pakrash et. al, (2001).
7
Figura 6. Estructura del compuesto diphloretohidroxicarmaloilo extraído de Ishige
okamurae reproducido de Soo-Jin y colaboradores (2008).
11
Figura 7. Estructuras de los compuestos 1) 1-(3´,5´-dihidroxi-peroxi)-7-(2”,4”-
trihidroxiperoxi)-2,4,9-trihidroxidibenzo-1,4-dioxina y 2) 6,6´-bieckol identificados en
Ecklonia cava reproducido de Quang-To y colaboradores (2009).
12
Figura 8. Compuesto derivado del fucosterol activo contra células KB extraído a partir de
Padina pavonica reproducido de Ktari y Guyot (1999).
15
Figura 9. Estructura del compuesto Lobophorido 1 obtenido de Lobophora variegata
reproducido de Kubanek y colaboradores (2002).
16
Figura 10. Apariencia de Padina mexicana. 21
Figura 11. Mapa del área de colecta. 22
Figura 12. Fraccionamiento del extracto etanólico de Padina mexicana. 25
Figura 13. Fraccionamiento de CCC F1 hasta la obtención de un compuesto puro. 26
Figura 14. Cromatografía en capa fina en fase reversa (TLC) realizada con diferentes
solventes para determinar el mejor fraccionamiento del extracto ELS 1F2.
29
Figura 15. Cromatografía del primer fraccionamiento de ELS1F2. Se realizó un punteo en
placa de 10 µL por muestra aproximadamente.
30
Figura 16. Cromatografía del fraccionamiento realizado a CCAF8. Donde 1a representa
las facciones de la columna cromatográfica CCB1, 1b muestra las fracciones de la
columna CCB2 y 1c la columna CCB3.
30
Figura 17. Cromatografía de la columna CCC. La muestra CCB1F17 se fraccionó con un

iii
gradiente de elusión de EtOH: H2O (9:1). 31
Figura 18. Columna cromatográfica DD. Se observa la obtención de un compuesto
parcialmente puro.
31
Figura 19. Placa cromatográfica en fase reversa impregnada con DPPH. 1-2=CCAFU1-2,
3=CCAF3, 4= CCAF4, 5=CCAF5, 6-7=CCAFU6-7,8=CCAF8, 9=CCAF9, 10=CCAF10,
11=CCAF11, 12=CCAF12, 13= CCAF13, 14=CCAF14 y 15=CCAF15
34
Tabla I. Ensayos de actividad antibacteriana de las fracciones CCAFU1-2 –CCAF15 donde
los halos de inhibición están expresados por el promedio ± SD (n=2).
33
Tabla II. Valores de concentración inhibitoria media (IC50 ) requerida para reducir 50% el
radical DPPH.
40
Lista de gráficos
página
Grafica 1. Curva logarítmica donde se muestra la dosis y el efecto del phloroglucinol en el
ensayo espectrofotométrico con DPPH.
35
Grafica 2. Curva logarítmica donde se muestra la dosis y el efecto de la fracción CCAF10
en el ensayo espectrofotométrico con DPPH.
36
Grafica 3. Curva logarítmica donde se muestra la dosis y efecto de la fracción CCAF11 en
el ensayo espectrofotométrico con DPPH.
37
Grafica 4. Curva logarítmica donde se muestra la dosis y efecto de la fracción CCAF12 en
el ensayo espectrofotométrico con DPPH.
38
Grafica 5. Se muestra la dosis y el efecto de la fracción CCAF13 en el ensayo
espectrofotométrico con DPPH.
39

iv
Lista de abreviaciones
nm nanómetros
mg miligramos
mm milímetros
mL mililitros
µm micrómetros
µL microlitros
g gramos
kg kilogramos
IC50 concentración inhibitoria media
H2O agua
DPPH 2,2-difenil-1-picril hidracilo
EtOH etanol
ERO especies reactivas de oxígeno.
HPLC siglas en inglés para cromatografía de líquidos de alta resolución
TLC siglas en inglés para cromatografía en capa fina
CH2Cl2 diclorometano
MeOH metanol
NaCO3 carbonato de sodio
NMR resonancia magnética nuclear.
H2SO4 ácido sulfúrico
LDL lipoproteínas de baja densidad.

v
Glosario
Agar: mezcla heterogénea de dos clases de polisacáridos sin ramificaciones
obtenido a partir de varias especies de algas rojas.
Alga: organismos totipotenciales que llevan a cabo reacciones fotosintéticas,
tienen origen polifilético, principalmente acuáticos.
Anteridios: órgano pluricelular donde se forman los gametos masculinos.
Antibiótico: agente químico producido por un organismo o de manera sintética que es
dañino para otros organismos.
Antimicrobiano: sustancia que puede inhibir el crecimiento de microorganismos o matarlos.
Dioicas: organismos con sexos separados.
Esporangio: estructura que se encuentra en el esporófito y se especializa en llevar a
cabo la meiosis que dará las esporas haploides (n).
Gram-negativo: célula procariota que contiene una membrana externa conformada por
lipopolisácaridos, lipoproteínas y otras macromoléculas complejas.
Gram-positivo: célula procariota que tiene una pared conformada principalmente por
peptidoglucano y carece de membranas externas.
In vitro: se refiere a la técnica o procedimiento realizado en un tubo de vidrio en
condiciones controladas fuera de un organismo.
Macroalga: nombre informal que agrupa organismos dentro de tres divisiones y que
comparten ciertas características: productores primarios, oxigénicos,
principalmente bentónicos con reproducción sexual mediante historias de
vida, acuáticas, etc.
Meristemos: tejido vegetal o zona de células indiferenciadas capaces de dividirse
activamente. A de este se forman los otros tejidos que conforman el
cuerpo vegetal. Metabolitos primarios: compuestos orgánicos esenciales para el organismo que los produce
debido a que forman parte de su metabolismo (crecimiento y
reproducción). Metabolitos secundarios: compuestos orgánicos sintetizados por un organismo los cuales no tienen
un papel esencial en su metabolismo.

vi
Monoico: organismo que puede contener órganos de ambos sexos.
Oogonios: estructuras reproductoras femeninas.
Paráfisis: prolongaciones vegetativas que se forman sobre la superficie del talo.
Patógenos: organismos que causan daño a su hospedero.
Phlorotaninos: compuestos fenólicos solubles en agua, con pesos moleculares entre 500
y 3000 los cuales se encuentran en las algas pardas. Precipitación de carbonato de calcio:
depósito de carbonato de calcio en la pared celular de las macroalgas en
forma de cristales de aragonita y calcita.
Radicales libres: átomo o grupos de átomos con un electrón desapareado, extremadamente
inestable y con gran poder reactivo.
Rotavapor: equipo utilizado para evaporar disolventes a presión reducida.
Shikimatos: compuestos orgánicos derivados de la ruta del acido shikímico.
Soro: Estructuras reproductoras que se encuentran concentradas en una región
del talo. Sujetador secundario:
estructura prensil que se desarrolla en cualquier parte del talo para
adherirse a sustratos diferentes.
Talo: estructura de una macroalga compuesta por tres regiones (sujetador,
estipe y fronda) que contienen células totipotenciales que no forman
tejidos verdaderos.

vii
Resumen.
En experimentos preliminares el extracto etanólico del alga parda Padina mexicana resultó
ser activo por el método de difusión en agar con discos contra las cepas resistentes
Staphylococcus aureus y Streptococcus pyogenes a una concentración de 2.0 mg/disco, por lo que
fue seleccionado para su fraccionamiento en búsqueda de los compuestos responsables de la
actividad mostrada. Adicionalmente las fracciones obtenidas fueron sometidas a un ensayo de
actividad antioxidante por medio de los métodos bioautográfico y espectrofotométrico utilizando la
reducción del radical libre estable 2,2-difenil-1-picrilhidracilo como indicador de actividad. La
extracción sólido-líquido (ESL1) dio como resultado 4 fracciones denominadas ESL1F1 a ESL1F4
de las cuales ESL1F2 (2.42 g) mostró actividad contra S. aureus, S. pyogenes y Escherichia coli,
además de mostrar interesante actividad antioxidante por el método bioautográfico. Del
fraccionamiento de ESL1F2 por medio de columna cromatográfica se obtuvieron 15 fracciones
denominadas CCAFU1-2 ---CCAF15. La mayoría de las fracciones fueron activas contra S. aureus
excepto CCAF9, CCAF11 y CCAF15, en el caso de S. pyogenes solo la fracción CCAFU6-7 no
mostró actividad contra este microorganismo. Las fracciones CCAFU6-7, CCAF10 y CCAF12
mostraron actividad contra Vibrio parahaemolyticus. En el ensayo de actividad antioxidante de
estas fracciones, CCAF10, CCAF11, CCAF12, CCAF13 y CCAF14 resultaron ser las más activas
en el método bioautográfico, la medición espectrofotométrica de la actividad antioxidante de estas
fracciones mostró un IC50 de 0.20 mg/mL para CCAF10 y de 0.22 mg/mL para CCAF11 mientras
que CCAF12 mostró actividad de 0.13 mg/mL y CCAF13 mostró un IC50 de 0.023 mg/mL siendo
menos activas que el stock de phloroglucinol (0.012 mg/mL). El fraccionamiento de CCAF8 por
cromatografía en fase reversa dio como resultado 17 fracciones (CCB1 a CCBF17). La fracción
CCBF17 resultó ser un sólido de color amarillo claro que fue nuevamente fraccionado en columna
cromatográfica de fase reversa dando 17.9 mg de un compuesto aparentemente puro denominado
DDF1.
Palabras clave: Actividad antimicrobiana, actividad antioxidante, Padina mexicana, metabolitos
secundarios, Baja California Sur.

viii
Abstract.
In previous experiments the ethanolic extract of brown algae Padina mexicana showed to
be active using the agar diffusion method with discs against resistant strains of Staphylococcus
aureus and Streptococcus pyogenes at the concentration of 2.0 mg/disk, because it’s activity the
ethanolic extract was selestec for fractionation in searching for compounds responsible for the
activity showed. Also the fractions obtained were tested for antioxidant activity for the
spectrophotometric and bioautographyc methods by the reduction of the stable free radical 2,2-
diphenyl-1-picrilhidracilo as an indicator of activity. The solid-liquid extraction (ESL1) resulted in four
fractions, named ESL1F1 to ESL1F4 of which ESL1F2 (2.42 g) showed activity against S. aureus,
S. pyogenes and Escherichia coli, with interesting antioxidant activity by the bioautographyc
method. From the ESL1F2 fractionation using chromatographic column were obtained 15 sections,
named CCAFU1-2--- CCAF15. Mostly the fractions were active against S. aureus except CCAF9,
CCAF11 and CCAF15, in the case of S. pyogenes only the fraction CCAFU6-7 was not active
against this organism. Fractions CCAFU6-7, CCAF10 and CCAF12 showed activity against Vibrio
parahaemolyticus. In the experiment of antioxidant activity with those fractions, the CCAF10,
CCAF11, CCAF12, CCAF14 and CCAF13 were the most active with the bioautographyc method,
also spectrophotometric measurement of antioxidant activity showed an IC50 of 0.20 mg/mL for
CCAF10 and 0.22 mg/mL for CCAF11 while the highest activity were obteined with the fractions
CCAF12 (0.13 mg/mL) and CCAF13 (IC50 of 0.023 mg/mL) have less activity that stock of
phloroglucinol (0.012 mg/mL). The fractionation of CCAF8 by reversed phase chromatography
resulted in 17 fractions (CCB1 to CCBF17). The CCBF17 fraction was a light yellow solid that was
fractionated once again by reverse phase chromatographic column giving 17.9 mg of a compound
named DDF1 apparently pure.
Keywords: Antimicrobial activity, antioxidant activity, Padina mexicana, secondary metabolites,
Baja California Sur.

1
1. Introducción.
Los productos naturales también conocidos como metabolitos secundarios son
compuestos (o mezcla de ellos) que poseen una gran diversidad estructural, siendo moléculas
relativamente pequeñas, pueden presentar una amplia gama de efectos biológicos entre los que se
encuentran la actividad inhibitoria del crecimiento bacteriano, actividad inmunoestimulante,
citotóxica, antiviral, anti-incrustante y antioxidante por nombrar algunas (Moreau et al., 1984; Ktari y
Guyot, 1999; De Lara-Isassi et al., 2000; Nagayama et al., 2002; Mi-Jeon et al., 2004; Shibata et al,
2008; Zou et al, 2008). El estudio de los metabolitos secundarios provenientes de organismos
marinos es relativamente reciente, sin embargo en las 3 últimas décadas se han publicado
artículos donde el papel de las algas marinas como fuente de compuestos bioactivos es
preponderante (Blunt et al., 2006). Las algas marinas al ser organismos sésiles están expuestas y
sometidas a diversas condiciones bióticas y abióticas, lo que provoca que sinteticen moléculas que
no tienen equivalencia con las terrestres, lo cual es gran interés debido al gran potencial de
aplicaciones derivadas de la diversidad estructural y bioactividad que presentan. En la actualidad
cerca del 2 % de las especies conocidas de macroalgas han sido estudiadas (Garateix, 2005;
Magallanes et al., 2003). Las investigaciones de manera general han demostrado que muchas
especies de las 3 principales clases de macroalgas (Chlorophyceae, Phaeophyceae y
Rhodophyceae) son fuente potencial de compuestos bioactivos, tan sólo para el 2008 se habían
reportado alrededor de 3000 compuestos y aproximadamente 1,140 de ellos fueron de la clase
Phaeophyceae (Magallanes et al., 2003; Amsler, 2008). Los organismos pertenecientes a dicha
clase tienen un gran potencial bioactivo debido a su capacidad de sintetizar metabolitos mixtos de
origen terpeno-aromático; dentro de esta clase se han encontrado compuestos como
phlorotaninos, polisacáridos, alginatos, manitol, ácidos grasos e inclusive clorofila c2 con actividad
biológica (Mabrouk, 1985; Rosell y Srivastava, 1987; De Lara-Isassi y Álvarez-Hernández, 1999;
Magallanes et al., 2003; Mohsen et al., 2007; Kamei y Aoki, 2007; Zubia et al., 2008; González et
al., 2009).
La familia Dictyotaceae ha sido la más estudiada, cerca del 30 % de los compuestos
reportados han sido obtenidos de especies del género Dictyota, no obstante, dentro del género
Padina también se ha reportado que los extractos muestran actividad antimicrobiana así como
presencia de compuestos de origen fenólico con actividad antioxidante (Mabrouk et al., 1985;
Chkhikvishvili y Ramazanov, 1999; Kang, 2003; Amornlerdpison et al., 2007; Matanjun et al., 2008;
Kumar et al., 2009).

2
Tomando en cuenta lo anterior, el presente estudio se enfocó en la evaluación de la
fracción ESL1F2 obtenida a partir de un extracto crudo de Padina mexicana la cual se fraccionó
nuevamente por medio de columna cromatográfica y aquellas fracciones derivadas fueron
analizadas en pruebas contra las cepas resistentes de Staphylococcus aureus, Streptococcus
pyogenes, Escherichia coli y Vibrio parahaemolyticus así como su potencialidad como agentes
antioxidantes in vitro al ser sometida a pruebas con el radical libre estable 2,2-difenil-1-
picrilhidracilo.
2. Antecedentes.
2.1. Polifenoles.
En las macroalgas podemos encontrar un grupo de moléculas denominadas policétidos,
los cuales son los segundos mayoritarios en el metabolismo secundario de estos organismos;
estas moléculas tienen un origen biosintético muy similar a los ácidos grasos y son polímeros de
acetato y ocasionalmente de propionato. Su clasificación está basada en la policétido sintetasa
responsable de su biosíntesis, principalmente de los policétidos
del tipo I y tipo II. Los policétidos de tipo II condensan sistemas cíclicos aromáticos como
flavonoides que se encuentran en las plantas y los phlorotaninos en las algas (Dewick, 2002;
Amsler, 2008).
Los phlorotaninos o polifenoles son una clase estructural de policétidos que se encuentra
exclusivamente en las algas pardas. Estas moléculas están constituidas por unidades de
phloroglucinol ligadas en enlaces carbono-carbono o carbono-oxígeno cuya estructura base es el
esqueleto dibenzo-1,4-dioxina; se sintetizan por la ruta del malonil-CoA que produce el bloque de
construcción de phloroglucinol y su peso molecular varía entre los 500 y 5000 (Figura 1, Isaza,
2007; Amsler, 2008). Debido a su carácter polar, estos se pueden extraer con mezclas de alcohol
y agua en diversas proporciones, sin embargo, la mezcla mayormente usada es acetona-agua
(70:30) (Isaza, 2007).
Los phlorotaninos se clasifican en seis grupos basados en las variaciones de las unidades
de polimerización del phloroglucinol, entre los cuales tenemos a los fucoles, phloretoles,
fucoforetoles, fuhaloles, isofuhaloles y eckoles (Figura 2) que difieren en el número de grupos
hidroxilo presentes en los vínculos de su enlace y donde todos a excepción de los fucoles están
unidos por enlaces éter. Las unidades son a menudo phloroglucinol esterificado o acilado y puede
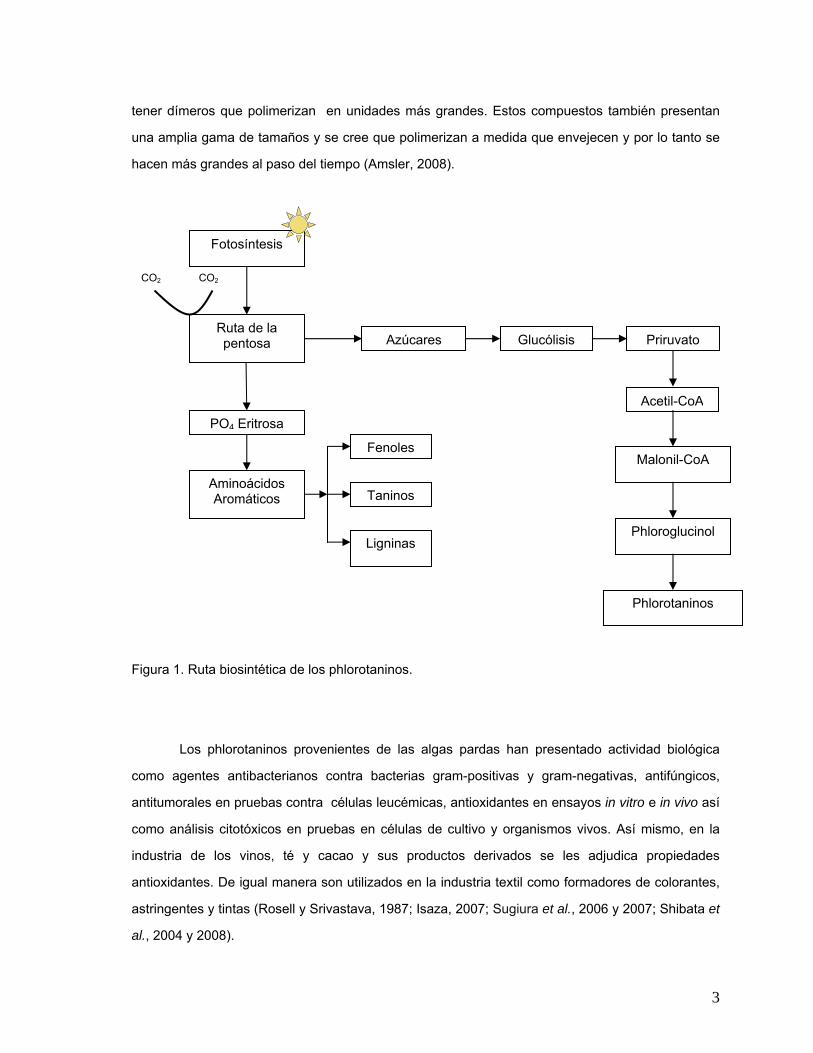
3
tener dímeros que polimerizan en unidades más grandes. Estos compuestos también presentan
una amplia gama de tamaños y se cree que polimerizan a medida que envejecen y por lo tanto se
hacen más grandes al paso del tiempo (Amsler, 2008).
Figura 1. Ruta biosintética de los phlorotaninos.
Los phlorotaninos provenientes de las algas pardas han presentado actividad biológica
como agentes antibacterianos contra bacterias gram-positivas y gram-negativas, antifúngicos,
antitumorales en pruebas contra células leucémicas, antioxidantes en ensayos in vitro e in vivo así
como análisis citotóxicos en pruebas en células de cultivo y organismos vivos. Así mismo, en la
industria de los vinos, té y cacao y sus productos derivados se les adjudica propiedades
antioxidantes. De igual manera son utilizados en la industria textil como formadores de colorantes,
astringentes y tintas (Rosell y Srivastava, 1987; Isaza, 2007; Sugiura et al., 2006 y 2007; Shibata et
al., 2004 y 2008).
Fotosíntesis
Ruta de la pentosa
CO2 CO2
PO4 Eritrosa
Aminoácidos Aromáticos
Fenoles
Taninos
Ligninas
Azúcares Glucólisis Priruvato
Acetil-CoA
Malonil-CoA
Phloroglucinol
Phlorotaninos
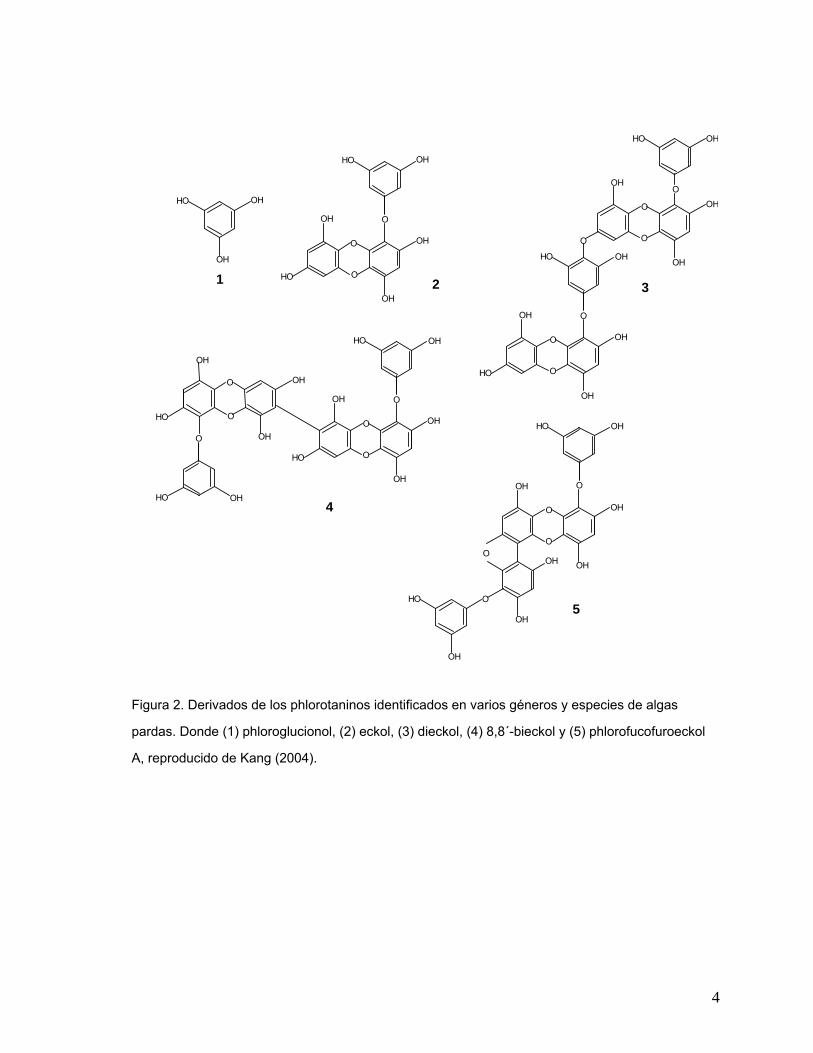
4
Figura 2. Derivados de los phlorotaninos identificados en varios géneros y especies de algas
pardas. Donde (1) phloroglucionol, (2) eckol, (3) dieckol, (4) 8,8´-bieckol y (5) phlorofucofuroeckol
A, reproducido de Kang (2004).
HO OH
OH
O
OHHO
O
O
OH
HO
OH
OH
OH
OH
HO
OH
O
O
HO
O
O O
O OH
HO
OH
OH
OH
O
OH
O
HO
O
O
OH
OH
OH
OH
HO
OH
OH
O
O
O
OH
OH
HO
HO
OH
O
OHO
O
O OH
OH
OH
OH
O
HO
HO
OH
1 2 3
4
5

5
2.2. Terpenos.
Más de la mitad de los metabolitos secundarios obtenidos de las macroalgas son de origen
terpenoidal, los cuales forman una gran y estructuralmente diversa familia de productos naturales
derivados de unidades de isopreno (C5); sus estructuras típicas contienen carbono representado
por esqueletos (C5)n, y pueden clasificarse en función del número de unidades isoprenoides que
contienen incorporados como hemiterpenos (C5), monoterpenos (C10), sesquiterpenos (C15),
diterpenos (C20), sesterterpenos (C25), triterpenos (C30) y tetraterpenos (C40).
Figura 3. Unidades de isopreno y estructura reproducido de Dewick, (2002).
Los terpenos se forman por la vía del mevalonato y la ruta recién descubierta de
mevalonato independiente a través de fosfato deoxixilulosa. Las estructuras son conformadas a
través de un amplio uso de los mecanismos de carbocatión y reordenamientos posteriores Wagner-
Meerwein (Dewick, 2002). Los esteroides, carotenoides, quinonas preniladas e hidroquinonas son
algunos de los compuestos derivados. Muchos otros productos naturales que contienen
terpenoides o elementos de sus moléculas en combinación con esqueletos de carbono son
procedentes del acetato y shikimato que generan alcaloides, compuestos fenólicos, vitaminas,
entre otros.
C5 unidad de isopreno Isopreno
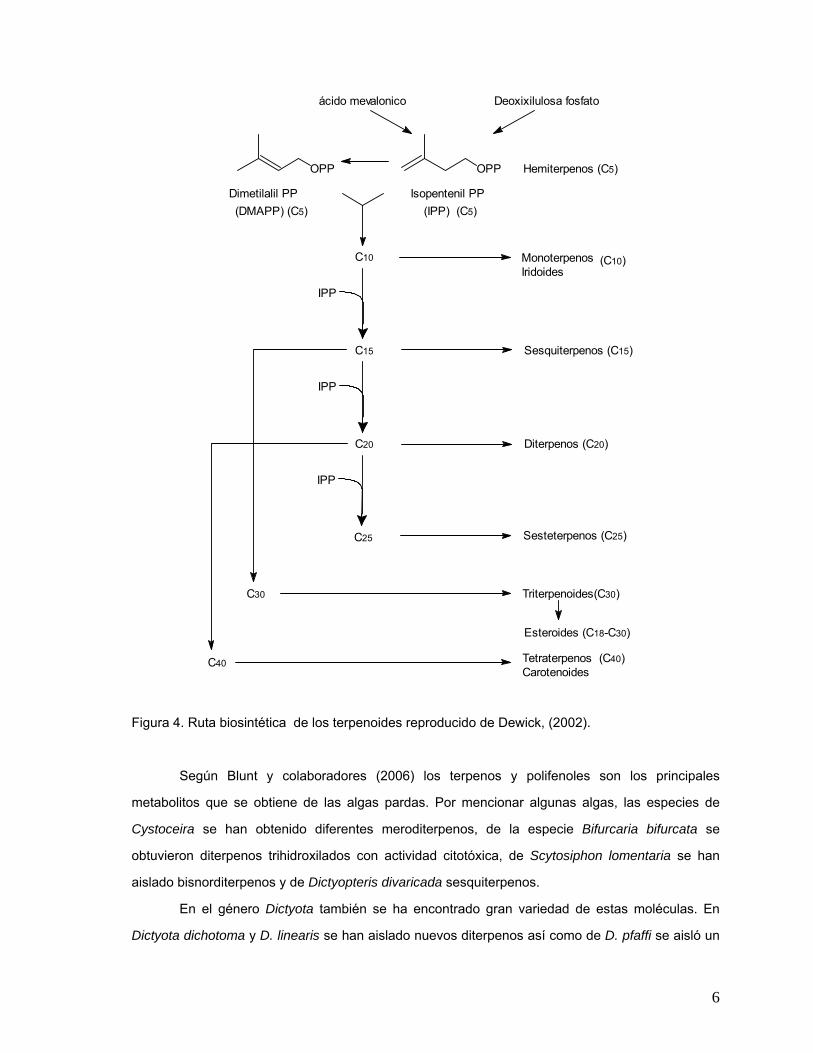
6
Figura 4. Ruta biosintética de los terpenoides reproducido de Dewick, (2002).
Según Blunt y colaboradores (2006) los terpenos y polifenoles son los principales
metabolitos que se obtiene de las algas pardas. Por mencionar algunas algas, las especies de
Cystoceira se han obtenido diferentes meroditerpenos, de la especie Bifurcaria bifurcata se
obtuvieron diterpenos trihidroxilados con actividad citotóxica, de Scytosiphon lomentaria se han
aislado bisnorditerpenos y de Dictyopteris divaricada sesquiterpenos.
En el género Dictyota también se ha encontrado gran variedad de estas moléculas. En
Dictyota dichotoma y D. linearis se han aislado nuevos diterpenos así como de D. pfaffi se aisló un
OPP
Dimetilalil PP Isopentenil PP(DMAPP) (C5) (IPP) (C5)
OPP
ácido mevalonico Deoxixilulosa fosfato
Hemiterpenos (C5)
C10 MonoterpenosIridoides
(C10)
IPP
C15
C20
IPP
IPP
C25
C30
C40
Sesquiterpenos (C15)
Diterpenos (C20)
Sesteterpenos (C25)
Triterpenoides(C30)
Tetraterpenos (C40)Carotenoides
Esteroides (C18-C30)

7
derivado de dolabeladieno que presentó actividad anti-HSV-1 y contra la reverso transcriptasa del
VIH in vritro. Así mismo, de la especie Dictyota mertensii se han aislado diterpenos que previenen
la herbivoría.
2.3. Mecanismo del radical libre estable 2,2-difenil-1-picrilhidracilo (DPPH) y la
actividad antioxidante.
El radical libre estable 2,2-difenil-1-picrilhidracilo (DPPH) es un polvo de color púrpura
compuesto por moléculas estables de radicales libres. Su estructura está conformada por tres
anillos aromáticos unidos por enlaces nitrógeno-nitrógeno y en uno de estos anillos contiene tres
moléculas de NO2 como grupos funcionales (Pakrash et al., 2001).
El DPPH tiene dos aplicaciones principales, por un lado es utilizado como un monitor de
reacciones químicas que involucra a dichos radicales y por otro es un estándar en las resonancias
paramagnéticas de electrones; este método se ha utilizado para cuantificar muestras sólidas o
líquidas y no es específico a un componente antioxidante particular (Pakrash et al., 2001).
Al someter en contacto al DPPH con un compuesto antioxidante, su electrón desapareado
genera una fuerte absorción máxima a 517 nm en un espectrofotómetro y se visualiza el cambio de
coloración de púrpura a amarillo como resultado de la oxidación. La oxidación es una reacción
simple donde interviene un agente reductor o antioxidante por ejemplo phloroglucinol el cual
suministra electrones al agente oxidante o DPPH que capta dichos electrones quedando con un
estado de reducción inferior al que tenía, es decir, se reduce (Figura 5; Pakrash et al., 2001).
Figura 5. Estructura del radical libre 2,2-difenil-1-picrilhidracilo (DPPH) y mecanismo de acción
reproducido de Pakrash et al. (2001).
N
NNO2 O2N
NO2
+
OH
OH HO
NO2
O2N NO2
NH
N
OH
OH
+O
DPPH Phloroglucinol

8
Esto mismo sucede en los sistemas biológicos, donde la acción de componentes
enzimáticos o no enzimáticos intervienen en la regulación de los radicales libres. Los componentes
enzimáticos son básicamente las enzimas superóxido dismutasa (SOD), glutation peroxidasa
(GSHPx) y la catalasa (CAT). La enzima SOD dismuta el oxígeno para formar peróxido de
hidrógeno y su principal función es la protección contra el anión superóxido, la enzima GSHPx se
relaciona principalmente con las membranas celulares pues destruye a los peróxidos orgánicos
formados a partir de ácidos grasos poli-insaturados derivados de la acción de radicales libres
oxigenados en las membranas. También reacciona activamente con el peróxido de hidrógeno,
destruyéndolo. Finalmente, la enzima CAT destruye al peróxido de hidrógeno formado en los
peroxisomas como producto de la actividad metabólica. Esta enzima es particularmente importante
en los glóbulos rojos, ya que estas células que no poseen peroxisomas en su estado maduro y la
enzima permanece libre en el citoplasma cumpliendo una actividad citoprotectora de gran
relevancia (Gutiérrez, 2002; Avello y Suwalsky, 2006).
Los componentes no enzimáticos del sistema de defensa antioxidante son muy
numerosos. Sin embargo, los más importantes son el glutation reducido (GSH), la ceruloplasmina
(proteína transportadora de cobre en el plasma), la ferritina (proteína transportadora de hierro), el
ácido úrico, la vitamina E, la vitamina C, el β-caroteno y los aminoácidos derivados de la taurina e
hipotaurina. La actividad antioxidante de estas moléculas no sólo depende del metabolismo celular,
sino también de la nutrición, ya que algunas de estas moléculas no son sintetizadas por el
organismo y deben ser aportados por la dieta (Gutiérrez, 2002; Avello y Suwalsky, 2006).
Al perderse la efectividad de los componentes enzimáticos y no enzimáticos se genera el
estrés oxidativo debido a que el contenido intracelular de EROs sobrepasa las defensas
antioxidantes de la célula induciendo daño a moléculas biológicas como lípidos, proteínas, ácidos
nucleicos, entre otros. Las ERO actúan sobre lípidos poli-insaturados en las membranas celulares
generando pérdida de fluidez y lisis como consecuencia de la peroxidación lipídica. También
alteran funciones de los glúcidos asociados a la actividad de las interleuquinas y la formación de
prostaglandinas, hormonas y neurotransmisores; en cuanto a las proteínas, llevan a cabo una
desnaturalización e inactivación de las mismas y en ácidos nucleicos modifican las bases
produciendo mutaciones y cáncer. Las ERO se han asociado con la aparición de desordenes como
el Alzheimer, cáncer, inflamación, artritis reumatoide y ateroesclerosis, entre otras enfermedades
(Kang et al., 2004; Mosquera et al., 2005; Zou et al., 2008).

9
Por lo anterior, en los últimos 30 años se ha desarrollado cada día más el interés por los
problemas relacionados con el estrés oxidativo, los radicales libres, las especies reactivas del
oxígeno y los antioxidantes, por lo tanto se han buscado nuevas fuentes de moléculas efectivas
contra esta problemática (Gutiérrez, 2002). Las macroalgas tropicales contienen moléculas
antioxidantes denominados phlorotaninos capaces de secuestrar radicales libres. Debido a su alto
potencial redox esas moléculas son especialmente importantes. La efectividad de estos
antioxidantes varía dependiendo de los factores abióticos (Truus et al., 2004; Zubia et al., 2008),
siendo capaces de actuar contra especies reactivas de oxígeno (ERO) tales como el anión
peróxido (O2-), hidroxilo (OH), alcóxilo (RO), peroxilo (ROO) y peróxido de hidrógeno (H2O2) los
cuales tienen efecto dañino en lípidos, proteínas, carbohidratos, enzimas y ácidos nucleicos. Por
otro lado los phlorotaninos poseen mecanismos de fotoprotección contra la radiación solar,
específicamente la radiación ultravioleta y se asume que éstos protegen la fotodestrucción del talo
algal (Kang et al., 2004; Mosquera et al., 2005; Zou et al., 2008).
En los análisis de Le Tutour y colaboradores (1998) mencionan en el texto descubrimientos
en los años 70 sobre fosfolípidos en Porphyra tenera y Eisenia bicyclis así como análisis en los
años 80 enfocados en substitutos de fenoles y o-difenoles en Polysiphonia urceolata. Sin embargo,
Le Tutour y colaboradores (1998) realizaron pruebas en extractos de Laminaria digitata,
Himanthalia elongata, Fucus vesiculosus, F. serratus y Ascophyllum nodosum referente a
captación de radicales libres, con ello determinaron actividad antioxidante por un periodo inducido,
no obstante, mostraron sinergia con la vitamina E adquiriendo gran capacidad oxidativa.
Para el año 2000, Chkhikvishvili y Ramazanov determinan el porcentaje en peso seco de
compuestos fenólicos en varias algas pardas, encontrando mayor porcentaje en Cystoceira
compressa con 4.83% mientras que Padina pavonica fue una de las más bajas con 0.69%;
identificaron phlorotaninos acetilados en Cystoceira compressa los cuales se sometieron a la
prueba de actividad antioxidante con DPPH al igual que los antioxidantes comerciales picnogenol y
phloroglucina siendo estos últimos los que presentaron mayor actividad.
Posteriormente Kang y colaboradores (2003) analizaron extractos fenólicos de Eisenia
bicyclis, Ecklonia estolonifera, Ecklonia cava, Ecklonia kurome y Hizikia fusiformis encontrando en
ellos capacidad de protección vascular, específicamente evaluando la protección de las
lipoproteínas de baja densidad (LDL) y la disfunción eréctil en 31 pacientes; determinaron la
utilidad de los compuestos polifenólicos, como fundamentales agentes preventivos contra los
factores de riesgo vascular procedentes del estrés oxidativo.

10
En el 2004 Truus y colaboradores estimaron que F. vesiculosus contiene 1 µg/mL de
phlorotaninos y discernieron elementos inorgánicos bioactivos como metales pesados y halógenos
en ella. En este mismo año Kang (2004) y colaboradores analizaron 17 extractos y realizaron
pruebas contra las ERO utilizando el marcador 2’,7’-diclorodihidrofluoresceina diacetato.
Determinaron que a una concentración de 25 mg/mL de Ecklonia stolonifera inhibe un 44.30 ±
7.33% la generación total de ERO. Se siguió el estudio con E. stolonifera debido a su uso como
alimento en Korea y se aislaron de ella phloroglucinol, eckstolonol, eckol, phlorofucofuroeckol A y
dieckol (Figura 2).
Chang-Seon (2005) y colaboradores analizaron dieckol y phlorofucofuroeckol (Figura 2)
proveniente de Ecklonia cava verificando su acción en los neurotransmisores ya que la aplicación
de estos en ratas (estudiando tres áreas del cerebro: corte frontal, hipocampo y zona estriada)
incrementó el nivel de acetilcolina inhibiendo la acetilcolinoesterasa. También Gin-Nae y
colaboradores (2007) determinaron phloroglucinol, eckol y dieckol (Figura 2) en el alga Ecklonia
cava con actividad antioxidante; establecieron que dichos compuestos mostraron una alta
inhibición de radicales libres y detienen el daño en el ADN proponiéndolos como compuestos
efectivos y aplicables como antioxidantes.
Amornlerdison y colaboradores (2007) realizaron un estudio de actividad antioxidante en
Padina minor donde evaluaron la reducción del radical DPPH y estimaron los compuestos
fenólicos. En su estudio determinaron que esta alga contenía 217.40 ± 11.28 de fenoles
equivalentes de ácido gálico y con tal cantidad inhibió un 77% el radical DPPH. Por igual, Zubia y
colaboradores (2007) determinaron en Padina gymnospora una cantidad de 5.58 ± 0.30 (peso
seco) de polifenoles y la reducción del radical DPPH con tal cantidad fue de 3.45 mg/mL, sin
embargo, en su estudio encuentran que el mayor porcentaje de reducción está en Avrainvillea
longicaulis, Chondria baileyana y Lobophora variegata.
Matanjun y colaboradores (2008) realizaron análisis de actividad antioxidante con los
métodos, TEAC (capacidad antioxidante equivalente a trolox) y FRAP (poder antioxidante de
reducción férrica); determinaron la capacidad de las tres algas rojas Eucheuma cottoni, E.
spinosum y Halymenia durvillaei, dos algas verdes Caulerpa lentillifera y C. racemosa y las tres
algas pardas Dictyota dichotoma, Sargassum polysystum y Padina sp. Encontrando que C.
lentillifera presentó mayor actividad en el ensayo TEAC (2.16 ± 0.04 mM/mg) y S. polysystum en el
ensayo FRAP (366.69 ± 11.85 mM/mg) así mismo estas dos algas presentaron un alto contenido
de compuestos fenólicos.

11
Shibata y colaboradores (2008) evaluaron el potencial de los phlorotaninos como
inhibidores de la peroxidación fosfolipídica en el sistema liposomal y determinaron la capacidad
oxidativa con DPPH; los oligómeros eckol, phlorofucofuroeckol A, dieckol y 8,8”-bieckol (Figura 2)
aislados de Ecklonia cava, E. kurome y Eisenia bicyclis mostraron potente inhibición en la
peroxidación fosfolipídica (1 µM en el sistema liposomal) y una reducción de DPPH (50%) en
concentración de 12-26 µM.
Soo-Jin y colaboradores (2008) elucidaron el compuesto diphloretohidroxicarmaloilo
(Figura 6) vía NMR proveniente del alga parda Ishige okamurae, midieron la capacidad de
captación en radicales siendo comparada la actividad contra la del ácido ascórbico; se determinó
que el compuesto es más efectivo contra radicales hidroxilo que el mismo ácido. Así también, este
mismo autor determinó junto a You-Jin (2008) los efectos antioxidantes de enzimas extraídas de la
misma alga, encontrando que los extractos generados por proteasas tienen alta captación
reductiva frente a radicales de peróxido de hidrógeno.
Figura 6. Estructura del compuesto diphloretohidroxicarmaloilo extraído de Ishige
okamurae reproducido de Soo-Jin y colaboradores (2008).
Por igual, Zou y colaboradores (2008) analizaron el alga Ishige okamurae y aislaron
phloroglucinol (Figura 2), diphlorethohidroxicarmaloilo (Figura 6) y 6,6´-bieckol (Figura 7) siendo
estas dos últimas moléculas las que presentaron la mayor actividad reductiva frente al DPPH,
radical alquilo, hidroxilo y superóxido, así como también redujeron significativamente el nivel de
especies reactivas de oxígeno en fibroblastos fetales, macrófagos celulares de ratón y en células
leucémicas.
O O
OH
OH
HO
HO
5a
9a8
7
6
1''
2''3''
4''
5''6''
O
O
OH
OH
OH
OH
HO
1'
2'3'
4'
5'6'
1
2
34
4a
10a

12
Quang-To y colaboradores (2009) identificaron en Ecklonia cava phloroglucinol, dieckol,
6,6´-bieckol y 1-(3´,5´-dihidroxi-peroxi)-7-(2”,4”-trihidroxiperoxi)-2,4,9-trihidroxidibenzo-1,4-dioxina
que posteriormente fueron probados en un ensayo de inhibición sobre la liberación de histamina en
células de cultivo (basófilas de leucemia); encontraron un potencial antialérgico a base de un buen
mecanismo de supresión en la liberación de histamina (Figura 7).
Es por esto que en el presente estudio se dio importancia en analizar el extracto etanólico
del alga Padina mexicana con el fin de identificar la actividad antioxidante y antimicrobiana de las
respectivas fracciones, esto basado en la bibliografía antes mencionada donde claramente se han
encontrado que diferentes especies del género presentan dichas actividades.
Figura 7. Estructuras de los compuestos 1) 1-(3´,5´-dihidroxi-peroxi)-7-(2”,4”-trihidroxiperoxi)-2,4,9-
trihidroxidibenzo-1,4-dioxina y 2) 6,6´-bieckol identificados en Ecklonia cava reproducido de
Quang-To y colaboradores (2009).
2.4. Actividad antimicrobiana de las algas pardas.
A lo largo de la historia, ha habido una continua batalla entre los seres humanos y la
multitud de microorganismos que causan enfermedades infecciosas. Esto mejoró con la aparición
de la penicilina en 1940, no obstante la euforia por la conquista potencial de enfermedades
HO
O
OH
OH
OH
OHO
O
OH
OH
OHO
O
OHO
OH
HO
1
24a5a
10a9a
9
7
1'
3'5'
1''
3''5''
O
1'
3'5'HO
O
OH
HO 10a 9a
34a 5a 6
8
HO7
5a 4a3
19a 10a
OH
OH
OH
OHHO
6'2'
O
O
1 2

13
infecciosas fue de corta duración pues casi tan pronto como los fármacos antibacterianos se
desplegaron, las bacterias respondieron manifestando diversas formas de resistencia (Tenover,
2006).
Se conoce que las principales formas de acción que contienen los agentes antibacterianos
son la interferencia en la síntesis de la pared celular, interferencia en la formación de ácidos
nucleicos y proteínas así como inhibición de rutas metabólicas pero esto fue contrarrestado por la
manifestación de la resistencia antibacteriana generada al adquirir genes que codifican enzimas,
tales como β-lactamasas, que destruyen el agente antibacteriano antes de que pueda surtir efecto.
Otro mecanismo, es la adquisición de bombas de flujo que expulsan el agente antibacteriano de la
célula antes de que pueda llegar a su sitio de destino. También pueden sintetizar varios genes por
vía metabólica tomando la unión del agente antimicrobiano de bacterias muertas que inhiban sus
efectos o pueden adquirir mutaciones que limitan accesos a dichos agentes. Se reporta que las
penicilinas, cefalosporinas, tetraciclinas, macrólidos, fluroquinolonas, entre otros ya no resultan
efectivos por lo que se ha enfocado en la búsqueda de compuestos provenientes de nuevas
fuentes como las macroalgas que tengan potencialidad (Tenover, 2006; Džidič et al., 2008).
Shirahama en 1942 publicó el primer reporte de las propiedades antibióticas en algas
donde consiguió aislar una sustancia a partir de Cystophyllum hakodatense que tubo efectos sobre
Lactobacillus vulgaricus y L. helveticus. Pratt y colaboradores en 1951 obtuvieron extractos de
algas marinas solubles en éter capaces de inhibir el crecimiento de Staphylococcus aureus,
Escherichia coli y Pseudomonas aeruginosa (De Lara- Isassi et al., 1989; Durand, 2001).
En contraste con estos años, Colin y Augier (1939) estudiaron el alga Polysiphonia
fastigiata y la presencia de componentes fenólicos en ella, también Leman (1944) aisló una
sustancia cristalina de carácter fenólico. Augier (1949) reaisló el ácido dibromohidroxibenzoico
(Scheuer, 1973).
Así mismo, en los sesenta se produjeron avances aun más significativos en la
investigación de los productos naturales algales consiguiendo con ello el aislamiento de un gran
número de compuestos con actividad antimicrobiana hasta la actualidad (Jing-wen y Wei-ci, 1984).
Durante marzo y junio de 1982, Jing-wen y Wei-ci colectaron 60 algas las cuales fueron extraídas
con tolueno-metanol (1:3) para determinar su actividad antimicrobiana contra Bacillus subtilis,
Escherichia coli, Saccharomyces cerevisiae y Vibrio sp. En ese estudio se encontró que solo
algunos miembros de las Rhodophyceae y Phaeophyceae presentaron dicha actividad.

14
Por su parte, Moreau y colaboradores (1984) reportaron varios géneros de Dictyotales
sometidos a estudios antifúngicos y antibacterianos; encontrando que los extractos hexánicos de
especies del género Dictyota fueron los más activos contra los hongos Microsporum gypseum, M.
fulvum, M. canis y contra bacterias Gram-negativas como Sarcina lutea. Así mismo, en 1988
realizó un muestreo estacional (12 meses) determinando con ello que la variación de la actividad
es dependiente de la madurez sexual, la ecología del sitio y el ciclo de vida del alga.
Bakus (1985) catalogó más de 220 compuestos fenólicos, provenientes de organismos
marinos como bacterias, algas, esponjas y anélidos, entre otros. En ese mismo año Mabrouk
estudió la inhibición del crecimiento micelial y acumulación de aflatoxinas en Aspergillus flavus,
donde encontró que los ácidos grasos y aceites volátiles de Padina pavonica podían inhibirles en
un 35% y 50% a concentraciones de 100, 150, 200 g/L.
Rosell y Srivastava (1987) establecieron que los ácidos grasos de algunas especies de
algas pardas poseen actividad antimicrobiana contra Bacillus cereus, Bacillus subtilis, Branhamella
catarrhalis, Microccocus luteus y Staphylococcus aureus. Un año después Paul (1988) reportó la
presencia de polifenoles en las algas pardas, a los cuales les atribuyó la defensa química contra
depredadores como invertebrados y peces.
En 1989 De Lara-Isassi y colaboradores reportan actividad antimicrobiana en el extracto
etanólico de Padina crispata (halo de 1-2 mm) así como en los extractos acuoso y etanólico de
Padina durvillaei (halos de 1-2 mm respectivamente). Este análisis fue realizado por el método de
difusión en agar por disco en el cual se añadió 0.1 mL de muestra pero no establece la cantidad de
extracto. Por igual, De Lara-Isassi y colaboradores en 1993 colectaron 57 algas en las costas
mexicanas, 13 de las cuales se encontraban dentro de las Phaeophyceae y se analizaron en
búsqueda de actividad antimicrobiana; para el análisis colocaron 0.1 mL de las muestras en cada
disco encontrando que el extracto acetónico del alga Padina boergesenii presentó un halo de
inhibición de 7 mm contra una cepa de Staphylococcus aureus y un halo de 13 mm en la cepa de
Streptococcus pyogenes, así mismo en un extracto etanólico de la misma alga se obtuvo un halo
de inhibición de 11 mm en la cepa de S. pyogenes, desafortunadamente, es imposible determinar
la concentración a la cual el extracto fue probado.
Ktari y Guyot (1999) aislaron el compuesto derivado del fucosterol (Figura 8) con actividad
citotóxica proveniente de Padina pavonica que inhibió el crecimiento de células KB (carcinoma
epidérmico-bucal humano); el extracto obtenido con diclorometano presentó una inhibición del

15
100% a 10 µg/mL mientras que el extracto obtenido con una combinación de diclorometano-
metanol fue moderadamente activo.
Figura 8. Compuesto derivado del fucosterol activo contra células KB extraído a partir de Padina
pavonica reproducido de Ktari y Guyot (1999).
En el 2000 De Lara-Isassi y colaboradores estudiaron la toxicidad de 73 especies de algas.
La medición de la toxicidad fue clasificada en tres rangos: tóxica (T), moderadamente tóxica (MT) y
no tóxica (NT) esto de acuerdo a las reacciones que los peces Carassius aratus presentaron; se
consideró alta toxicidad si el organismo sometido a prueba moría en un periodo de 2 h después de
aplicar el extracto, la moderada toxicidad implicó la observación del comportamiento como
movimiento rápido, equilibrio, cambio de coloración, entre otros, durante un periodo de 20 h; al no
presentar ningún efecto dentro de los intervalos anteriores se consideró no tóxico. Con lo anterior
se determinó que los extractos acetónicos y etanólicos de Padina boergesenii eran altamente
tóxicos al igual que el extracto etanólico de Padina durvillaei, mientras que los extractos acetónicos
y etanólicos de Padina gymnospora fueron moderadamente tóxicos. Gonzáles del Val y
colaboradores (2001) reporta a Padina pavonica con actividad antimicrobiana contra Bacillus
subtilis así como otras algas pardas con potencial bioactivo.
Nagayama y colaboradores (2002) dicen haber realizado el primer estudio que demuestra
actividad bactericida de cinco phlorotaninos purificados del alga parda Ecklonia kurome (Figura 4);
se probó phloroglucinol, eckol, phlorofucofuroeckol A, dieckol y 8,8′-bieckol contra patógenos como
S. aureus, S pyogenes y V. parahaemolyticus entre otros encontrando en el análisis que no había
una diferente susceptibilidad entre las Gram-negativas y Gram-positivas.
1
2
34
HO
10
56
7
89
19 11 13
12
1415
1617
21
20
22
23HOO
27
25
2624
2829

16
Después, Kubanek y colaboradores (2002) aislaron el compuesto denominado Lobophorido
1 del alga parda Lobophora variegata (Figura 9); se determinó que dicho compuesto es utilizado
por la misma como defensa química contra los hogos Lindra thalassiae y Dendryphiella salina, así
también mencionan que podría ser producto de un simbionte microbiano del alga al tener similitud
estructural con metabolitos bacterianos.
Figura 9. Estructura del compuesto Lobophorido 1 obtenido de Lobophora variegata reproducido de
Kubanek y colaboradores (2002).
Mi-Jeon y colaboradores (2004) aislaron de Ecklonia cava cuatro derivados de
phloroglucinol denominados eckol, 8,8’-bieckol, 8,4”-dieckol y phlorofucofuroeckol A (Figura 2),
posteriormente los probaron como inhibidor de la transcriptasa reversa y proteasa del virus VIH-1;
con ello encontraron que 8,8’-bieckol y 8,4”-dieckol (Figura 2) tienen potente actividad antiviral,
comparable con otros supresores de dichas enzimas. Por otro lado, Shibata y colaboradores (2004)
determinaron la localización de los phlorotaninos en las células vegetativas de la parte externa en
el talo de E. kurome, E. cava y E. bicyclis, así mismo elucidaron el contenido de compuestos como
eckol, dieckol, 8,8” bieckol, phlorofucofuroeckol A (Figura 2), tetrámero de phloroglucinol y
phloroglucinol en varias concentraciones utilizando la técnica de HPLC y microscopia con vainillina-
HCI.
Tuney (2006) analizó cuatro extractos de Padina pavonica obtenidos con acetona, metanol,
dietil éter y etanol respectivamente, siendo éste último el único que mostró actividad antimicrobiana
sobre Candida albicans, S. faecalis, P. aeruginosa y E. coli. Por su parte, Kamei y Aoki (2007)
analizaron 342 especies de algas en la búsqueda de un compuesto capaz de inhibir el virus
infeccioso de necrosis hematopoyética en peces, encontrando 15 especies de algas pardas con
actividad antiviral. Padina crassa, Padina arborescens y Eisenia bicyclis mostraron una inhibición
O
OMe
42
2829
3031
27 26
25
40
24 23
O OH
22
39
2120
OMe37
1918
O OMe OMeOH
OMe
36
17
O
16
35
15
34
14
1312
11
10
9
8 7
65
43
21
32 38
O

17
del 90% del virus, siendo esta última seleccionada para la determinación de su acción. El estudio
de E. bicyclis condujo al aislamiento e identificación del pigmento fotosintético clorofila c2 como el
responsable de la actividad en E. bicyclis.
Kandhasamy y Arunachalam (2008) reportan que el extracto metanólico de Padina
tetrastomatica inhibe el crecimiento de S. aureus (entre otros) al igual que un extracto etanólico de
Kumar y colaboradores (2009). Chiheb y colaboradores (2009) determinaron la acción del extracto
metanólico de Padina pavonica encontrando que presentó alta actividad contra S. aureus.
3. Justificación.
La Secretaría de Salud reportó para el 2006 que las enfermedades del sistema urinario
ocupaba el séptimo lugar en egresos hospitalarios mientras que enfermedades en tejido conjuntivo
el treceavo, enfermedades infecciosas intestinales el dieciseisavo y enfermedades respiratorias
como la neumonía el decimoctavo. Así mismo, el Sistema Nacional de Información en Salud
reportó para el 2007 que la causa de muerte por enfermedades infecciosas respiratorias bajas son
de un 27.6 % y las enfermedades de vías urinarias como nefritis o nefrosis 22.2%.
Se ha establecido, que los microorganismos causantes de tales enfermedades han
adquirido resistencia a los fármacos disminuyendo la susceptibilidad a estos, tal es el caso de E.
coli que comúnmente es causante de infecciones en el tracto urinario y bacteremia pero que ahora
es resistente a los productos que eran utilizados para su erradicación como amoxicilina o
ampicilina y cefalosporinas de alto espectro (Rosales y Rodríguez, 2006; Tenover, 2006). Por su
parte S. aureus es causante de enfermedades respiratorias que regularmente se trataban con
vacomicina, tienen inmunidad a su acción y a otros antibióticos como la meticilina (Sá-Leáo et al.,
1999; Tenover, 2006).
En cuanto a Streptococcus pyogenes se refiere, este microorganismo es causante de
faringitis, infecciones cutáneas y de tejidos blandos, sepsis puerperal, neumonía, endocarditis,
meningitis y artritis las cuales eran tratadas con penicilina, eritromicina u otros macrólidos pero este
microorganismo ha adquirido residencia a ellos (Bassetti et al, 2000; Rodríguez et al, 2000)
Los organismos del género Vibrio son causantes de enfermedades como la gastroenteritis
y al no ser tratada podría ser mortal; se reporta que este microorganismo es susceptible a
tetraciclina, doxiciclina, furazolidona, cotrimoxazol, cloranfenicol, cefalosporinas de tercera
generación, aminoglucósidos y quinolonas. Sin embargo, pueden presentar resistencia a las

18
sulfonamidas, penicilinas, colistín, ampicilina y cefalosporinas de primera y segunda generación
(cefalotina y cefuroximaes) (Dabanch et al, 2009).
Los cambios en las epidemiologías infecciosas causadas por estos microorganismos y las
secuelas producidas tienen una estrecha relación con la disponibilidad de antibióticos, la mejoría
en el acceso a la atención médica para precisar con oportunidad el diagnóstico de las infecciones
así como la eficacia de los antibióticos y el adecuado huso de los mismos (Rodríguez et al, 2000).
Es por esto que la búsqueda de compuestos que presenten actividad antimicrobiana contra cepas
resistentes es de gran importancia ya que es necesario obtener moléculas con estructuras
novedosas que ayuden a contrarrestar las enfermedades de manera efectiva y que al mismo
tiempo puedan ser obtenidas de manera rápida y con bajos costos.
En cuanto a enfermedades degenerativas crónicas, el Sistema Nacional de Información en
Salud establece para el 2007 que el 60.5% de la población masculina y el 45 % de la población
femenina para ese año murió de enfermedades isquémicas del corazón, el 26.7% y 28.6% de
enfermedades cerebro-vasculares y el 64% y 69.2 % de diabetes mellitus respectivamente. Las
enfermedades cancerígenas causantes de muerte para ese año en la población masculina fueron
de próstata en un 15.7%, tráquea, bronquios y pulmón en un 14% y estómago en 9 % mientras que
para la población femenina el cáncer de mama fue mayoritario con 13.8 %, en el cuello del útero
con 12.1% e hígado con 7.6%.
Por lo tanto, la búsqueda de moléculas con potencial para contrarrestar enfermedades
degenerativas causadas por el estrés oxidativo ha adquirido auge en los últimos años (Rosales y
Rodríguez, 2006). Las macroalgas tropicales del género Padina contienen moléculas antioxidantes
capaces de secuestrar radicales libres. La efectividad de estos antioxidantes varía dependiendo de
los factores abióticos, siendo capaces de actuar contra especies reactivas de oxígeno (ERO) las
cuales tienen efecto dañino en lípidos, proteínas, carbohidratos, enzimas y ácidos nucleicos lo que
les asocia con las enfermedades degenerativas (Kang et al., 2004; Mosquera et al., 2005; Truus et
al., 2004; Zubia et al., 2008; Zou et al., 2008).
4. Descripción y distribución de Padina mexicana.
Las algas de la clase Phaeophycea son organismos multicelulares fotosintéticos que se
reproducen sexualmente y contienen clorofila a, c, c1 y c2 así como también pigmentos accesorios

19
como fucoxantinas y ß-caroteno; esta clase tiene alrededor de 14 órdenes dentro de los cuales se
encuentra el orden Dictyotales (Van den Hoek et al., 1995).
Dentro de dicho Orden se encuentra la familia Dictyotaceae, la cual presenta 26 géneros y
222 especies. Las Dictyotales contienen talos aplanados con diferentes tipos de meristemos,
mechones de pelos superficiales y crecimiento apical; usualmente presentan frondas erectas,
dicotómicas subdivididas o irregulares dependiendo si el crecimiento es inducido por una célula
apical o un grupo pequeño de células apicales, flaveadas o irregulares (Ceseña, 2003).
Uno de los géneros que se encuentra dentro de este orden es Padina, el cual se
caracteriza por tener un talo aplanado y en forma de abanico con una coloración que va desde
amarillo a café claro (Chávez, 2000; Ceseña, 2003). Las estructuras reproductivas denominadas
oogonios, anteriodios y esporangios se encuentran regularmente entre bandas concéntricas de
paráfisis; muchas especies se consideran dioicas, no obstante, en algunas poblaciones esto se ve
influenciado por la profundidad y estación del año. Los soros concéntricos alternantes y la línea de
pelos son consistentes en el medio apical de frondas bien desarrolladas, siendo rasgo distintivo
para determinar las especies (Ceseña, 2003).
Por otra parte, la especie Padina mexicana tiene un talo de aproximadamente 5 a 15 cm
de longitud y de 5 a 20 cm de amplitud, el cual no siempre se presentará erecto pues en ocasiones
no cuenta con sujetadores secundarios sobre la superficie del talo. Estas son capaces de precipitar
carbonato de calcio que puede ser depositado como aragonita en las zonas extracelulares
concéntricas comenzando en el espacio intercelular formando el margen apical del talo (Lüning,
1990; Chávez, 2000). Se pueden encontrar dos fenotipos de ella, en el primero los sujetadores
secundarios no precipitan carbonato de calcio por uno de los lados, midiendo de largo entre 10 y
15 cm con un número de células basales de 5 a 6 (dos células corticales) y el segundo, el
crecimiento se origina en forma de roseta y desarrolla sujetadores secundarios en uno de los lados
(sin células corticales), teniendo una altura máxima de 10 cm (Chávez, 2000).
En cuanto a su reproducción se refiere, posee talos monoicos con reproductores
arreglados en soros encontrados en la superficie del abanico; los oogonios miden 50 a 55 µm de
largo y 40 a 45 µm de ancho localizados en la región subapical entre las bandas de paráfisis y
están dentro de una cápsula de 220 µm. Los anteridios miden de 80 a 85 µm de largo y 10 a 15 µm
de ancho encontrándose sobre las bandas de paráfisis en la región apical como filamentos cortos
pluriloculares; los esporangios se localizan a lo largo de todo el talo entre las bandas de paráfisis y

20
carbonato de calcio, pueden medir entre 55 y 60 µm de largo y 40 a 45 µm de ancho (Chávez,
2000).
Padina mexicana se puede definir como una especie esporádica ya que puede estar
presente en primavera y verano y ausente durante el otoño e invierno; se ha registrado en todo el
Golfo de California sin embargo, en la región media y norte no es constante (Riosmena-Rodríguez
et al, 2009). Su principal distribución es en la región sur del golfo con una extensión al Pacífico
tropical mexicano; el crecimiento se ha reportado en la zona intermareal asociada a otras algas del
mismo género, así como también en sustratos arenoso-rocoso y en canto rodado (Chávez, 2000;
Riosmena-Rodríguez et al, 2009).
4.1. Clasificación taxonómica de Padina mexicana.
Dominio: Eucariota (Chatton)
Reino: Cromista (T. Cavalier-Smith)
Subreino: Cromobionta (T. Cavalier-Smith)
Infrareino: Heteroconta (T. Cavalier-Smith)
Phylum: Heterocontofita (Moestrup)
Clase: Phaeophyceae (Kjellman)
Orden: Dictyotales (Bory de Saynt)
Familia: Dictyotaceae (Lamouroux ex Dumortier)
Tribu: Zonarieae
Género: Padina (Adanson)
Especie: Padina mexicana (Thivy, 1945)

21
Figura 10. Apariencia de Padina mexicana.
5. Objetivo general.
Evaluar las fracciones obtenidas de Padina mexicana por su actividad antibacteriana y
antioxidante.
Identificar las fracciones del alga Padina mexicana que contengan actividad inhibitoria
contra cepas bacterianas resistentes a los antibióticos así como determinar la actividad
antioxidante en las mismas.
5.1. Objetivos particulares.
a) Fraccionar por cromatografía en columna la fracción ESL1F2 obtenida del extracto
etanólico de Padina mexicana.
b) Evaluar la actividad antibacteriana de las fracciones obtenidas, por medio del método de
difusión en agar contra tres cepas patógenas de humanos S. aureus (ATCC BAA-42), S.
pyogenes (ATCC BAA-946) y E. coli (ATCC BAA-196).

22
c) Determinar el potencial antioxidante de todas las fracciones obtenidas utilizando el
ensayo del radical libre estable 2,2-difenil-1-picril hidracilo (DPPH) por los métodos
bioautográfico y espectrofotométrico.
6. Materiales y Métodos.
6.1. Trabajo en campo.
Padina mexicana fue recolectada el 5 de junio de 2004 por Dr. Jesús Iván Murillo Álvarez y
M. en C. Mauricio Muñoz Ochoa en de San Juan de la Costa, Baja California Sur a los 24° 24’
47.08” latitud Norte, 110° 41’ 28.66” longitud Oeste. El alga se tomó del sustrato rocoso en la zona
intermareal, fue limpiada de material extraño y colocada en bolsas de plástico previamente
etiquetada como 04-002. Posteriormente, fue identificada en Escuela Nacional de Ciencias
Biológicas por la Dr. Luz Elena Mateo Cid y la Bióloga Catalina Mendoza y confirmada por el Dr.
Rafael Riosmena Rodríguez de la Universidad Autónoma de Baja California Sur.
Figura 11. Mapa del área de colecta.

23
6.2 Trabajo de laboratorio.
6.2.1. Obtención del extracto crudo.
El alga fue secada al sol y almacenada a -20° C hasta el momento de su extracción. 839 g
del alga Padina mexicana (etiquetada como 04-002) seca y molida fueron extraídos por
maceración con etanol destilado en proporción de 1:2 (masa:volumen). Cada 3 días se cambió el
disolvente. El extracto resultante fue concentrado a sequedad a presión reducida en un rotavapor
Yamato RE500. El sólido resultante (extracto etanólico crudo) fue transferido a un vial tarado y
etiquetado como 04-002-41.
6.2.2. Fraccionamiento del extracto crudo.
Inicialmente, el extracto crudo sé sometió a un proceso de extracción en fase sólida para
separar sus componentes en fracciones gruesas; esto se realizó disolviendo el extracto en
etanol:agua (9:1) seguidamente se mezcló con 250 g de sílica gel de fase normal (40 μm, 60 Å); se
mantuvo en movimiento constante hasta evaporar el disolvente y se dejó en la campana de
extracción. La mezcla completamente seca, fue extraída por percolación utilizando CH2Cl2 (1.5 L),
EtOAc (1.5 L), EtOH (0.9 L) y H2O (0.9 L), obteniéndose 4 fracciones denominadas
ESL1F1…ESL1F4 respectivamente. Todas las fracciones fueron sometidas a los ensayos de
actividad antimicrobiana y antioxidante. La fracción ESL1F2 (eluída con EtOAc) mostró actividad
contra S. aureus y S. pyogenes en el ensayo de difusión en agar con discos y actividad reductora
del radical libre estable 2,2’-difenil-1-picrilhidracilo. Considerando esto, la fracción ESL1F2 fue
seleccionada para continuar su fraccionamiento, por lo que 2.41 g de esta fracción fueron
sometidos a un fraccionamiento en columna cromatográfica sobre sílica gel de fase normal (40 μm,
60 Å). La muestra fue adsorbida en sílica gel a una proporción muestra: adsorbente igual a 1:2.5.
La columna fue empacada por vía húmeda con 120 g de sílica gel suspendida en CH2Cl2. La
muestra fue depositada en la parte alta de la columna. La elusión fue realizada utilizando CH2Cl2 (3
volúmenes, equivalentes a 690 mL), CH2Cl2:EtOH en gradiente 98:2, 96:4, 92:8, 90:10, 85:15,
80:20, 50:50 y finalmente EtOH y H2O. Después de analizar el patrón de separación por medio de
cromatografía en capa fina, y las fracciones fueron unidas por similitud en 15 fracciones nombradas

24
como CCAF1…CCAF15. Todas las fracciones fueron sometidas nuevamente a los ensayos de
actividad antimicrobiana y antioxidante.
Entre otras, CCAF8 mostró actividad inhibitoria contra S. aureus, S. pyogenes, Por lo que
fue fraccionada adicionalmente por cromatografía en columna de fase reversa. Para ello, 330.6 mg
de CCAF8 fueron disueltos en 2.5 mL de EtOH:H2O (9:1) e inyectados a una columna Lobar 240
LiChroprep RP18 (Merck) acoplada a una bomba peristáltica Masterflex (Cole-Parmer Instrument).
La elusión se realizó con EtOH:H2O (9:1) a 1 mL/min. El total de la fracción CCAF8 fue procesado
en 3 inyecciones sucesivas de donde, después de unir las fracciones semejantes se obtuvieron 18
nuevas fracciones las cuales fueron denominadas CCBF1…CCBF17. De estas fracciones,
CCBF17 (52 mg) resultó ser un sólido amorfo ligeramente amarillo, el cual fue adicionalmente
procesado en las mismas condiciones que la anterior habiéndose obtenido el compuesto puro
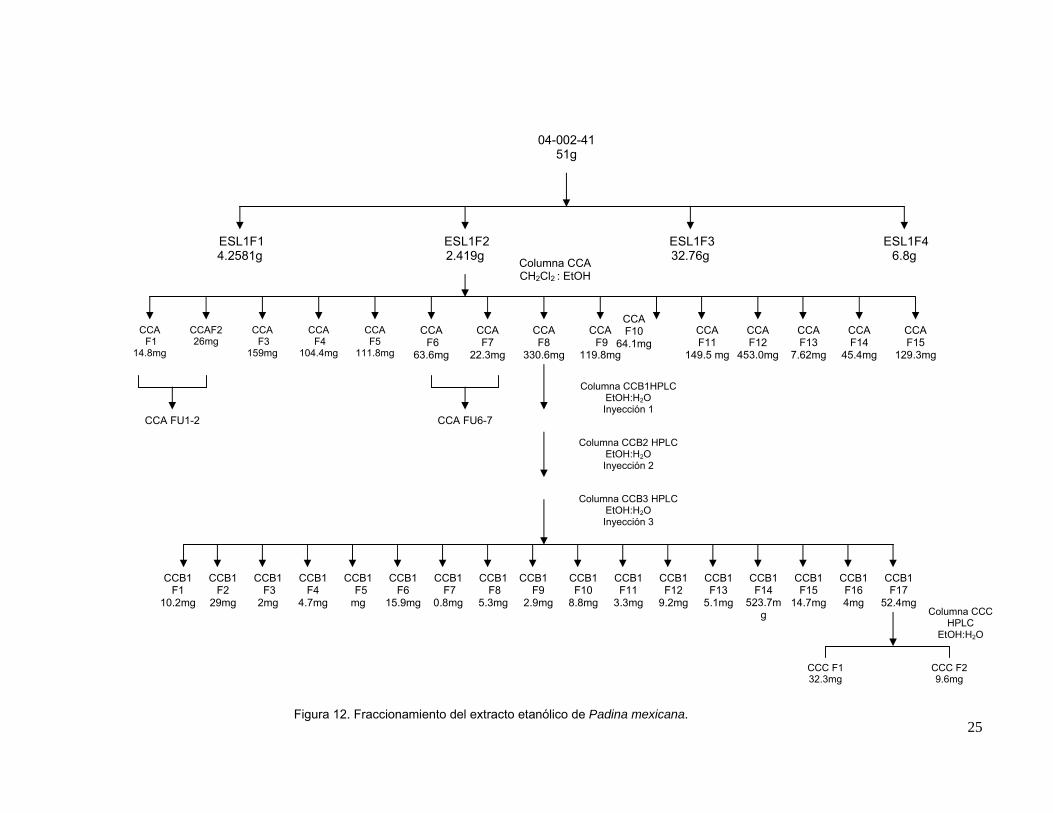
DDF1 (Figuras 12 y 13).
25
CCA F10
64.1mg
Columna CCC HPLC
EtOH:H2O
Columna CCB3 HPLC EtOH:H2O Inyección 3
Columna CCB2 HPLC EtOH:H2O Inyección 2
Columna CCB1HPLC EtOH:H2O Inyección 1
04-002-41 51g
ESL1F4 6.8g
ESL1F3 32.76g
ESL1F2 2.419g
ESL1F1 4.2581g
CCA F1
14.8mg
CCAF2 26mg
CCA F3
159mg
CCA F4
104.4mg
CCA F5
111.8mg
CCA F6
63.6mg
CCA F7
22.3mg
CCA F8
330.6mg
CCA F9
119.8mg
CCA F11
149.5 mg
CCA F12
453.0mg
CCA F13
7.62mg
CCA F14
45.4mg
CCA F15
129.3mg
CCA FU1-2 CCA FU6-7
CCB1 F1
10.2mg
CCB1 F2
29mg
CCB1 F3 2mg
CCB1 F4
4.7mg
CCB1 F5 mg
CCB1 F6
15.9mg
CCB1 F7
0.8mg
CCB1 F8
5.3mg
CCB1 F9
2.9mg
CCB1 F10
8.8mg
CCB1 F11
3.3mg
CCB1 F12
9.2mg
CCB1 F13
5.1mg
CCB1 F14
523.7mg
CCB1 F15
14.7mg
CCB1 F16 4mg
CCB1 F17
52.4mg
CCC F1 32.3mg
CCC F2 9.6mg
Columna CCA CH2Cl2 : EtOH
Figura 12. Fraccionamiento del extracto etanólico de Padina mexicana.

26
Figura 13. Fraccionamiento de CCC F1 hasta la obtención de un compuesto puro.
6.2.3. Actividad antimicrobiana.
6.2.3.1. Ensayo de actividad antimicrobiana.
Las fracciones de Padina mexicana fueron sometidas a un ensayo de actividad
antimicrobiana utilizando el método de difusión en agar con discos. El ensayo consiste en
colocar discos de papel filtro (6.4 mm de diámetro) impregnados individualmente con una
solución stock de cada fracción, sobre placas de agar inoculadas con la suspensión del
microorganismo de prueba. Las placas fueron incubadas a 37º C por 24 h, después de este
lapso el diámetro de los halos de inhibición fue medido. Cada ensayo se realizó por duplicado
y los resultados expresados como el promedio ± la desviación estándar en milímetros.
6.2.3.2. Microorganismos de prueba.
Los organismos de prueba utilizados en este estudio fueron obtenidos de la American
Type Culture Collection (ATCC), proveedor de cepas bacterianas previamente aisladas de
sitios específicos; se etiquetaron con el número de lote proporcionado por dicha institución.
El microorganismo Staphylococcus aureus (ATCC BAA-42) fue aislado de pacientes en
el hospital pediátrico Ward, Lisboa, Portugal en 1996; ésta bacteria es clasificada como grado
de seguridad 2 por su resistencia contra meticilina, penicilina, oxacilina, ampicilina y cefalotina.
La temperatura óptima de crecimiento es de 37º C.
CCC F1 32.4mg
DDF2 0.6mg
DDF3 --
DDF4 0.7mg
DDF5 2.1mg
DDF6 8.7mg
DDF7 6mg
DDF8 1.6mg
DDF9 5.2mg
DD F1 17.9mg
Compuesto puro
Columna cromatográfica
DD HPLC
EtOH:H2O

27
Por su parte Streptococcus pyogenes (ATCC BAA-946) se aisló durante la epidemia de
faringitis originada en el 2001 dentro de una escuela elemental en Pittsburgh, E.U.A. La
bacteria presenta resistencia a la eritromicina, grado de seguridad 2 y crecimiento óptimo a los
37º C.
Escherichia coli (ATCC BAA-196) se obtuvo de un aislamiento clínico de Klebsiella
pneumoniae; con características similares de bioseguridad que las otras 2 cepas anteriores. E.
coli mostró resistencia a todos los antibióticos probados y es capaz de producir β-lactamasas.
Vibrio parahaemolyticus (ATCC 17802) se aisló de un stock de Shirasu contaminado,
presentando susceptibilidad moderada a oxitetraciclina, ciproxina, tetraciclina y una alta
susceptibilidad al ácido oxalínico y sulfonamida.
6.2.3.3. Medios de cultivo e inóculos.
Se preparó agar Muller-Hinton en condiciones asépticas, fue esterilizado en autoclave a
120º C durante 15 minutos para verterse en cajas petri (100 x 15 mm) asignadas a cultivo de
Escherichia coli (ATCC BAA-196), Staphylococcus aureus (ATCC BAA-42) S. pyogenes
(ATCC BAA-946). De igual manera se preparó medio de cultivo Vibrio parahaemolyticus el agar
fue preparado con una solución salina al 2%.
A partir de un cultivo masivo de 24 h se prepararon suspensiones de cada
microorganismo de prueba en solución salina (al 2.5 % para V. parahaemolyticus y 0.85% para
los demás microorganismos), la suspensión fue estandarizada a una densidad óptica igual a 1
UA a 585 nm en el espectrofotómetro Merck SQ 118.
6.2.3.4. Preparación de discos.
En condiciones asépticas dentro de la campana de flujo laminar se impregnaron discos
de papel filtro de 6.4 mm de diámetro con 100 μL de una solución stock (20 mg/mL) de cada
una de las fracciones de prueba una concentración de 2 mg/disco. Así mismo, se impregnaron
discos con el disolvente utilizado en la disolución de las fracciones como control negativo.

28
6.2.4. Pruebas de actividad antioxidante.
6.2.4.1. Método espectrofotométrico.
Se preparo una solución stocks con 20 mg /mL de phloroglucionol en EtOH, se
realizaron diluciones para obtener concentraciones de 10, 5, 2.5, 1.25 mg/mL respectivamente.
Así mismo, se realizó una solución de 0.008g de DPPH (2,2-difenil-1-picrilhidracilo) en 200 mL
MeOH. Dentro de celdas espectrofotométricas de vidrio de 10 mL, se añadieron 5 mL de la
solución stock de DPPH y 50 μL phloroglucinol y un blanco (DPPH); se procedió a leer en el
espectrofotómetro Spectronic 20D (Milton Roy Company) a una longitud de onda de 517 nm.
Se leyó la absorbancia inicial y a los 30 minutos para luego determinar el porcentaje de
reducción mediante la formula: AbsBLANCO-AbsMUESTRA/AbsBLANCO*100 tomada de Lee et al.
(2003).
Posteriormente, de las fracciones CCAF10—CCAF13 se tomaron 20 mg y fueron
disueltos con 1 mL de EtOH realizando después diluciones seriadas en diferentes
concentraciones: 10, 5, 2.5, 1.25, 0.0625 y 0.3125 mg respectivamente. De cada fracción, se
tomó 250 μL y se vertieron en la celda espectrofotométrica con 4.75 mL de DPPH. Se leyó a
una longitud de onda de 517 nm la absorbancia inicial y después de los 30 minutos; esto se
realizó por duplicado y de igual manera, el porcentaje de reducción se estimó con la ecuación
anteriormente mencionada.
6.2.4.2. Método bioautográfico.
Se tomaron 150 µg de las fracciones CCAFU1-2—CCAF15 y se diluyeron en 360 µL
de EtOH. Posteriormente se impregnó una placa de cromatografía en fase reversa con 10 µL
de cada muestra. La placa fue sometida a una fase móvil con el sistema EtOH:H2O (9:1) y una
vez que estuvo seca se impregnó con DPPH diluido al 0.02%.
7. Resultados.
7.1. Cromatografía de capa fina (TLC) y fraccionamiento en columna
cromatográfica del extracto etanólico de Padina mexicana.
Como se observa en la figura 14 de los diferentes disolventes con distintos gradientes
de polaridad probados, la combinación CH2Cl2:EtOH resultó satisfactoria para el

29
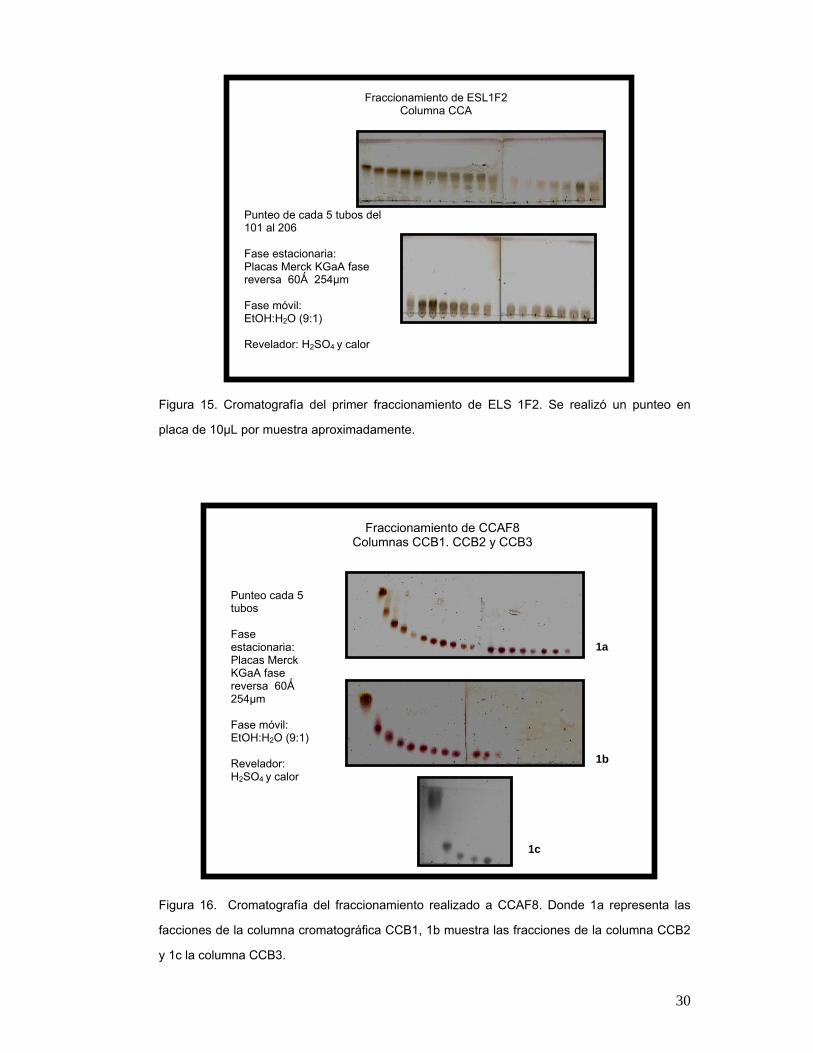
fraccionamiento de ESL1F2. La fase móvil con CH2Cl2:EtOH permitió una separación clara
entre una mancha con respecto a otra dentro de la fase estacionaria. Con lo anterior, se
obtuvieron 15 fracciones (Figura 15) etiquetadas como CCAFU1-2–CCAF15 respectivamente,
en las cuales la fase móvil fue EtOH:H2O (9:1) permitió discernir en la fase estacionaria las
diferentes manchas afines para la posterior unión de las fracciones en base a su semejanza.
Seguidamente, se eligió la fracción CCAF8 aplicándose en su fraccionamiento una
proporción 9:1 de EtOH:H2O que permitió una lejanía de la fracción con respecto al punto de
aplicación en la placa cromatográfica. Así mismo se eligió la fracción CCAF8 por presentar
actividad antimicrobiana en pruebas posteriores del la obtención (Sección 7.2). Por lo tanto, al
desarrollarse las columnas cromatográficas denominadas CCB1, CCB2 y CCB3 (Figura 16,
1a, 1b y 1c respectivamente) se obtuvieron 17 fracciones etiquetadas como CCB1F1–
CCB1F17 donde CCB1F17 presentó una coloración blanca y pequeños cristales signo de
menor impureza. Como se puede ver en la figura 17, del fraccionamiento de CCB1F17 se
obtuvieron dos fracciones denominadas CCCF1 y CCCF2 donde la fracción CCCF1 presentó
una coloración blanca indicativa de menor impurezas así como también se obtuvo una cantidad
factible (32.4mg). Al someter a fraccionamiento la fracción CCCF1 se obtuvieron nueve
fracciones etiquetadas como DDF1 a DDF9 de las cuales se determinó que DDF1 se
encontraba visiblemente libre de impurezas lo que llevó a asumirle como un compuesto
parcialmente puro (Figura 18).
Figura 14. Cromatografía en capa fina en fase reversa realizada con diferentes disolventes
para determinar el mejor fraccionamiento del extracto ELS1F2.
1 2 3 4 5
Determinación del fraccionamiento de ELS 1F2 con distintos gradientes
Fase móvil: 1) 100% CH2Cl2 2) 98:2 CH2Cl2:EtOH 3) 94:6 CH2Cl2:EtOH 4) 92:8 CH2Cl2:EtOH 5) 9:1 CH2Cl2:EtOH Revelador: H2SO4 y calor Fase estacionaria: Placas Merck KGaA fase reversa 60Ǻ 254µm
30
Figura 15. Cromatografía del primer fraccionamiento de ELS 1F2. Se realizó un punteo en
placa de 10µL por muestra aproximadamente.
Figura 16. Cromatografía del fraccionamiento realizado a CCAF8. Donde 1a representa las
facciones de la columna cromatográfica CCB1, 1b muestra las fracciones de la columna CCB2
y 1c la columna CCB3.
Fraccionamiento de ESL1F2 Columna CCA
Punteo de cada 5 tubos del 101 al 206 Fase estacionaria: Placas Merck KGaA fase reversa 60Ǻ 254µm Fase móvil: EtOH:H2O (9:1) Revelador: H2SO4 y calor
Fraccionamiento de CCAF8 Columnas CCB1. CCB2 y CCB3
Punteo cada 5 tubos Fase estacionaria: Placas Merck KGaA fase reversa 60Ǻ 254µm Fase móvil: EtOH:H2O (9:1) Revelador: H2SO4 y calor
1a
1b
1c

31
Figura 17. Cromatografía de la columna CCC. La muestra CCB1F17 se fraccionó con un
gradiente de elusión de EtOH: H2O (9:1).
Figura 18. Columna cromatográfica DD. Se observa la obtención de un compuesto
parcialmente puro.
7.2. Análisis de actividad antimicrobiana.
Una vez realizada la primera columna cromatográfica del extracto ESL1F2 se
obtuvieron 15 fracciones que se sometieron a pruebas de actividad antimicrobiana. Como se
muestra en la tabla I, ninguna fracción presentó actividad contra E. coli (ATCC BAA-196).
Por otro lado, la fracción CCAFU1-2, presentó inhibición contra S. aureus (ATCC
BAA-42) de 9 ± 0 y contra S. pyogenes (ATCC BAA-946) de 10 ± 0 mientras que CCAF3,
Punteo cada 5 tubos Fase estacionaria: Placas Merck KGaA fase reversa 60Ǻ 254µm Fase móvil: EtOH: H2O 9:1 Revelador: H2SO4 y calor
Fraccionamiento de CCB1F17 Columna CCC
Fase estacionaria: Placas Merck KGaA fase reversa 60Ǻ 254µm Fase móvil: EtOH: H2O 9:1 Revelador: H2SO4 y calor
Fraccionamiento de CCC F1 Columna DD

32
CCAF4, CCAF5 mostraron actividad similar contra ambas bacterias (Tabla I). A diferencia de
las anteriores, la fracción CCAFU6-7 sólo mostró actividad contra S. aureus (ATCC BAA-42)
con halos de 9.5 ± 0.7 y actividad contra V. parahaemolyticus (ATCC 17802) con 8 ± 0.
La fracción CCAF8 probada contra S. pyogenes (ATCC BAA-946) mostró halos de
inhibición de 15 ± 2.8 y en las pruebas realizadas contra S. aureus (ATCC BAA-42) una
inhibición de 8 ± 0. Esta fracción fue elegida para seguir su fraccionamiento en base a lo
anterior así como también por la fácil dilución en EtOH en comparación con las otras y por
tener una cantidad considerable de 330.6 mg.
Por su parte, CCAF10 presentó un comportamiento semejante a la fracción CCAF8 en
las pruebas contra S. aureus (ATCC BAA-42), sin embargo en las pruebas contra S. pyogenes
(ATCC BAA-946) y V. parahaemolyticus (ATCC 17802) los halos de inhibición fueron de 17.5 ±
3.5 y 8 ± 0 respectivamente. Las fracciones CCAF11 y CCAF15 presentaron actividad
solamente contra la cepa de S. pyogenes (ATCC BAA-946) teniendo halos de inhibición muy
similares (13.5 ± 2.1 y 14 ± 0, respectivamente).
Así mismo, CCAF12 mostró actividad contra las cepas de S. aureus (ATCC BAA-42),
S. pyogenes (ATCC BAA-946) y V. parahaemolyticus (ATCC 17802) al igual que CCAFU1-2 y
CCAF10 difiriendo en los valores ya que CCAF12 inhibió 12 ± 0, 18 ± 1.4 y 8.5 ± 0.7 en cada
caso siendo mayoritario.
Finalmente, como se muestra en la tabla I, las fracciones CCAF13 y CCAF14
presentaron un comportamiento semejante en las pruebas contra S. aureus (ATCC BAA-42) y
S. pyogenes (ATCC BAA-946).

33
Tabla I. Ensayos de actividad antibacteriana de las fracciones CCAFU1-2–CCAF15 donde los
halos de inhibición están expresados por el promedio ± SD (n=2).
+ halo marginal
-- actividad nula
7.3. Pruebas de actividad antioxidante.
7.3.1. Ensayo con el radical libre estable 2,2-difenil-1-picril hidracilo (DPPH).
7.3.1.1. Análisis espectrofotométrico.
Dentro de las gráficas 1 a 5 se muestra la acción de las fracciones sometidas al análisis
espectrofotométrico. Las fracciones CCAF10 y CCAF11 tuvieron una acción similar entre si
presentando un IC50 de 0.20 mg/mL y 0.22 mg/mL respectivamente (gráficas 2 y 3, tabla II). Por
su parte, la fracción CCAF12 presentó un IC50 de 0.13 mg/mL y CCAF13 mostró un valor de
0.023 mg/mL (graficas 4 y 5, tabla II) para inhibir el radical libre estable 2,2-difenil-1-
picrilhidracilo siendo estas las más activas. El análisis para el stock de phloroglucinol mostró un
Fracción E. coli
(ATCC BAA-196)
S. aureus
(ATCC BAA-42)
S. pyogenes
(ATCC BAA-946)
V. parahaemolyticus
(ATCC 17802)
CCAFU1-2 -- 9 ± 0 10 ± 0 --
CCAF3 -- 14.5 ± 0.7 20 ± 0 +
CCAF4 -- 14.5 ± 0.7 22 ± 0 +
CCAF5 -- 14.5 ± 2.1 15 ± 0 +
CCAFU6-7 -- 9.5 ± 0.7 -- 8 ± 0
CCAF8 -- 8 ± 0 15 ± 2.8 --
CCAF9 -- -- 14 ± 1.4 --
CCAF10 -- 8 ± 0 17.5 ± 3.5 8 ± 0
CCAF11 -- -- 13.5 ± 2.1 --
CCAF12 -- 12 ± 0 18 ± 1.4 8.5 ± 0.7
CCAF13 -- 10.5 ± 0.7 13.5 ± 2.1 --
CCAF14 -- 10.5 ± 0.7 14 ± 2.8 +
CCAF15 -- -- 14 ± 0 --
Blanco -- -- -- --

34
valor de IC50 de 0.012 mg/mL (grafica 1, tabla II) por lo que las cuatro fracciones anteriormente
mencionadas mostraron menor actividad reductora en comparación con este.
7.3.1.2. Análisis bioautográfico.
En la figura 19 se muestra la cromatografía en capa fina de fase reversa impregnada
con DPPH (0.02%) después de haber colocado las fracciones; es notable la coloración
amarilla indicativa de la oxidación del radical. La aplicación homogénea de las fracciones (10
µL/muestra) permitió observar claramente la potente actividad de las fracciones CCAF10,
CCAF11, CCAF12, CCAF13 y CCAF14 donde la coloración amarilla apareció inmediatamente
al impregnar la placa.
Figura 19. Placa cromatográfica en fase reversa revelada con DPPH al 0.2%. 1-2=CCAFU1-2,
3=CCAF3, 4= CCAF4, 5=CCAF5, 6-7=CCAFU6-7,8=CCAF8, 9=CCAF9, 10=CCAF10,
11=CCAF11, 12=CCAF12, 13= CCAF13, 14=CCAF14 y 15=CCAF15.
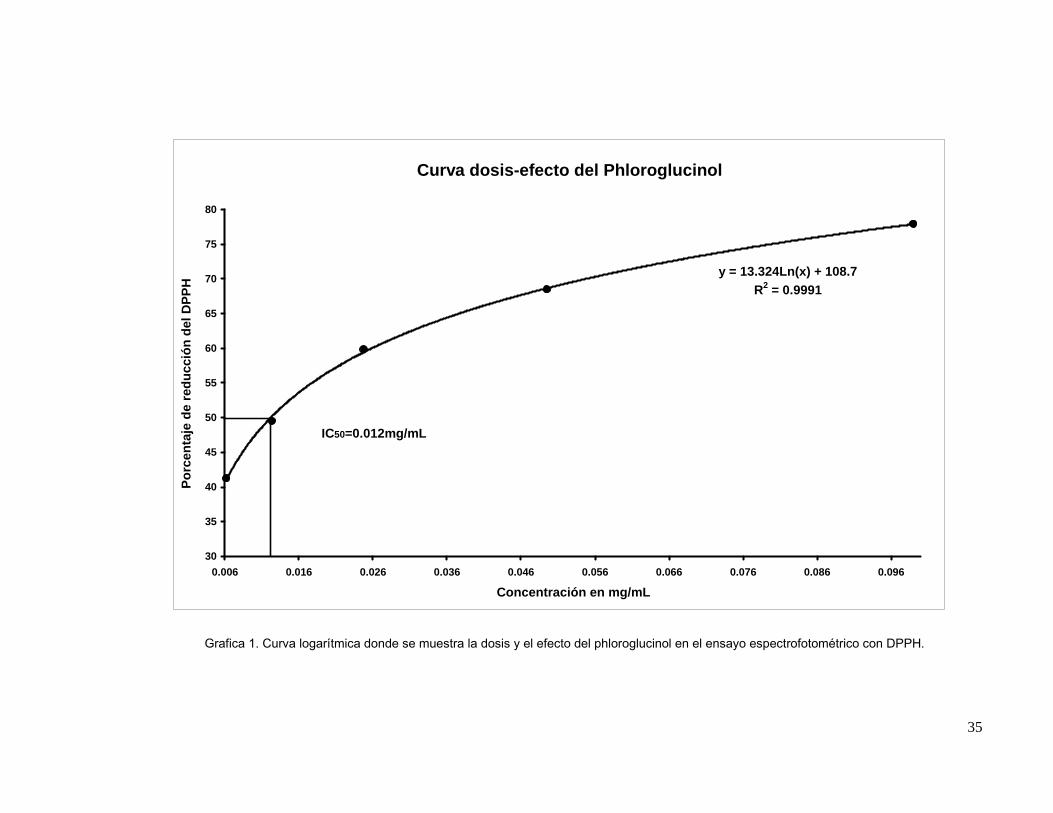
35
Curva dosis-efecto del Phloroglucinol
y = 13.324Ln(x) + 108.7R2 = 0.9991
30
35
40
45
50
55
60
65
70
75
80
0.006 0.016 0.026 0.036 0.046 0.056 0.066 0.076 0.086 0.096
Concentración en mg/mL
Porc
enta
je d
e re
ducc
ión
del D
PPH
IC50=0.012mg/mL
Grafica 1. Curva logarítmica donde se muestra la dosis y el efecto del phloroglucinol en el ensayo espectrofotométrico con DPPH.

36
Curva dosis-efecto de la fracción CCAF10
y = 16.036Ln(x) + 75.203R2 = 0.9616
0
10
20
30
40
50
60
70
80
0 0.2 0.4 0.6 0.8 1 1.2
Concentración en mg/mL
Porc
enta
je d
e re
ducc
ión
del D
PPH
IC50 = 0.20mg/mL
Grafica 2. Curva logarítmica donde se muestra la dosis y el efecto de la fracción CCAF10 en el ensayo espectrofotométrico con DPPH.
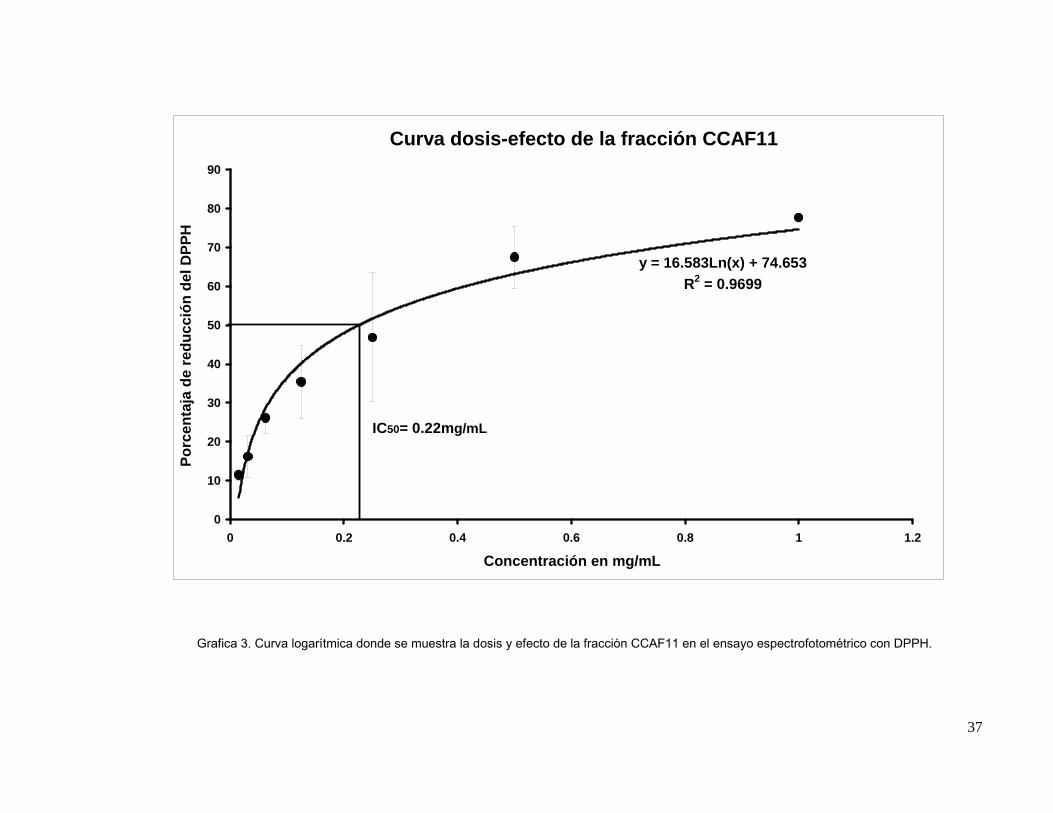
37
Curva dosis-efecto de la fracción CCAF11
y = 16.583Ln(x) + 74.653R2 = 0.9699
0
10
20
30
40
50
60
70
80
90
0 0.2 0.4 0.6 0.8 1 1.2
Concentración en mg/mL
Porc
enta
ja d
e re
ducc
ión
del D
PPH
IC50= 0.22mg/mL
Grafica 3. Curva logarítmica donde se muestra la dosis y efecto de la fracción CCAF11 en el ensayo espectrofotométrico con DPPH.
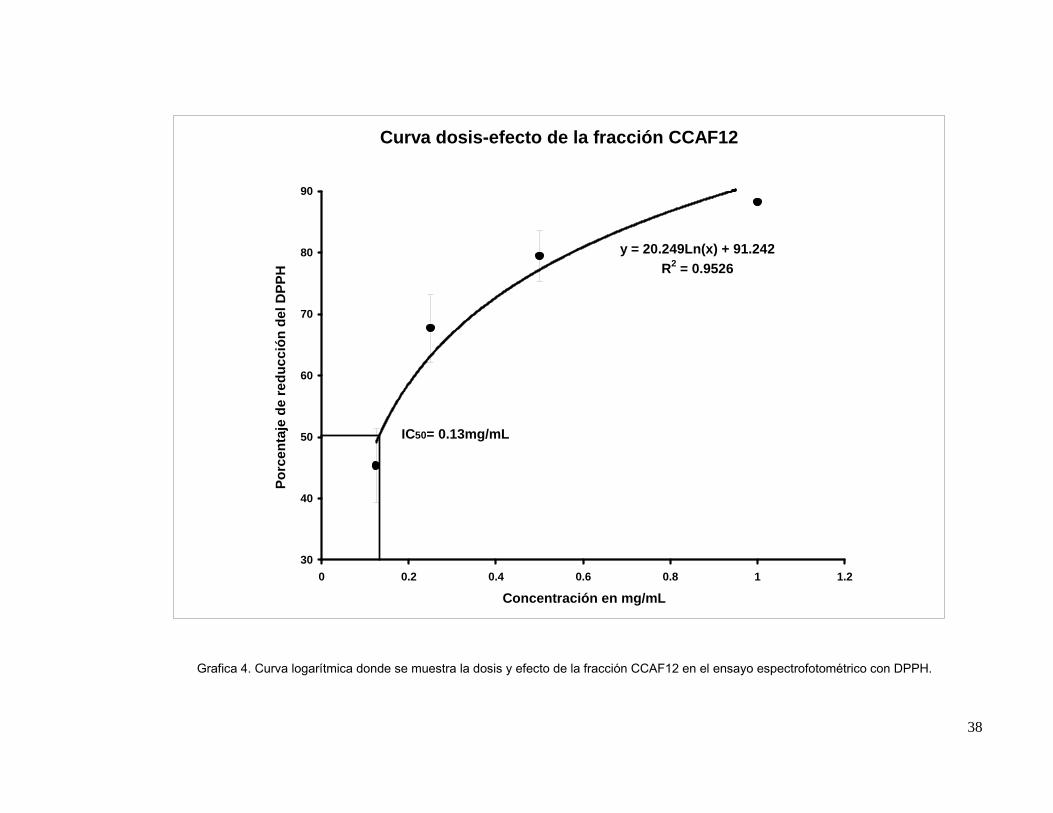
38
Curva dosis-efecto de la fracción CCAF12
y = 20.249Ln(x) + 91.242R2 = 0.9526
30
40
50
60
70
80
90
0 0.2 0.4 0.6 0.8 1 1.2
Concentración en mg/mL
Porc
enta
je d
e re
ducc
ión
del D
PPH
IC50= 0.13mg/mL
Grafica 4. Curva logarítmica donde se muestra la dosis y efecto de la fracción CCAF12 en el ensayo espectrofotométrico con DPPH.
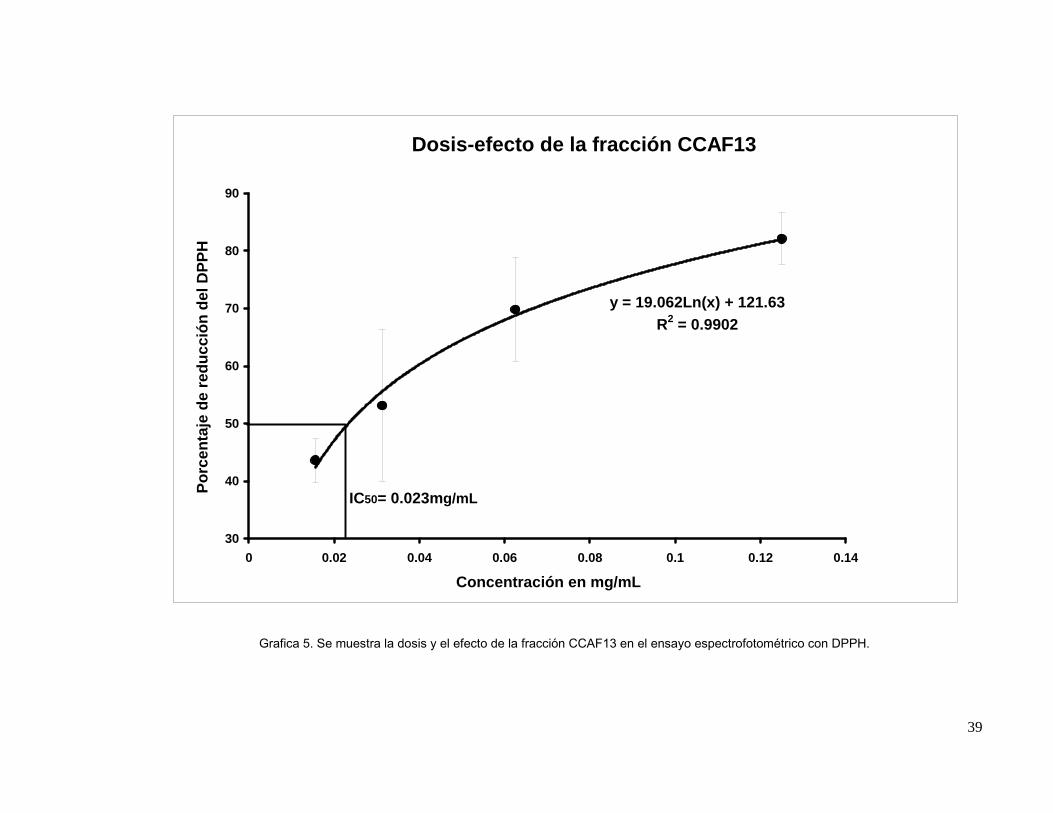
39
Dosis-efecto de la fracción CCAF13
y = 19.062Ln(x) + 121.63R2 = 0.9902
30
40
50
60
70
80
90
0 0.02 0.04 0.06 0.08 0.1 0.12 0.14
Concentración en mg/mL
Porc
enta
je d
e re
ducc
ión
del D
PPH
IC50= 0.023mg/mL
Grafica 5. Se muestra la dosis y el efecto de la fracción CCAF13 en el ensayo espectrofotométrico con DPPH.

40
Tabla II. Valores de concentración inhibitoria media (IC50 ) requerida para reducir 50% el radical
DPPH.
Fracción IC50 (mg/mL)
Phloroglucinol 0.012
CCA10 0.20
CCA11 0.22
CCA12 0.13
CCA13 0.023
8. Discusión
Al igual que el presente estudio, en algunos otros análisis del género Padina no se han
encontrado resultados de actividad antimicrobiana contra cepas Gram-negativas, tal es el caso
de los estudios realizados por Jin-Wen y colaboradores (1984) donde Padina crassa no generó
ningún efecto positivo contra E. coli y Vibrio sp. u otros microorganismos; por igual, Ballesteros
y colaboradores (1992) tampoco encontraron en Padina pavonica actividad antimicrobiana
solamente antimitótica con una inhibición del 25% en pruebas de crecimiento de células
leucémicas y actividad citotóxica contra células de riñón de mono teniendo una zona de
inhibición 1 mm.
Sin embargo, Kumar y colaboradores (2009) reportan actividad de Padina
tetrastomatica contra el organismo Gram-negativo Proteus vulgaris con halos de inhibición de
11 mm. Tuney y colaboradores (2006) así como Chiheb y colaboradores (2009) reportan mayor
susceptibilidad de los extractos algales contra bacterias Gam-positivas principalmente S.
aureus pero muestran resultados contra E. coli (halo de inhibición entre 7-10 mm y 10-16 mm
respectivamente) proveniente de extracto etanólico y metanólico de Padina pavonica; esto
sugiere que el género Padina también puede presentar actividad antimicrobiana contra
microorganismos gram-negativos.
De Lara-Isassi y colaboradores (1989, 1999) publican la acción antimicrobiana de
Padina crispata, Padina divoillaci contra S. aureus y Padina boerguesenii contra S. pyogenes
pero no es posible establecer el rendimiento ya que estos autores no revelan el contenido total
de extracto obtenido, no obstante, sus análisis indican al menos la presencia de componentes
con actividad antimicrobiana en estas especies.

41
Asimismo, González del Val y colaboradores (2001) probaron un extracto metanólico
de Padina pavonica teniendo actividad contra Bacillus subtilis (halo mayor a 15 mm) y
Kandhasamy y Arunachalam (2008) reportan a Padina tetrastomatica contra Gram-positivas.
De las 15 fracciones de Padina mexicana analizadas en el presente estudio, se puede
determinar que existe una alta potencialidad contra bacterias Gram-positivas como S. aureus
(ATCC BAA-42) y S. pyogenes (ATCC BAA-946), esto constata con estudios mencionados
anteriormente y los realizados por Durand (2001), Magallanes y colaboradores (2003) quienes
establecen que los extractos etanólicos presentan más capacidad inhibitoria en Gram positivas
debido a su carácter lipofílico; inclusive Magallanes y colaboradores (2003) dicen que S. aureus
es el microorganismo con mayor susceptibilidad a extractos y exudados algales.
La actividad mayoritaria se observó en las fracciones CCAFU1-2, CCAF3, CCAF4,
CCAF5 contra las bacterias Gram-positivas S. aureus (ATCC BAA-42) y S. pyogenes (ATCC
BAA-946) y las fracciones CCAFU6-7, CCAF9 y CCAF15 mostraron actividad contra una sola
cepa en cada caso (Tabla I). Las fracciones CCAF8 y CCAF10 mostraron actividad
antimicrobiana similar a pesar que visiblemente su naturaleza difiere. La distinta actividad
antimicrobiana observada sobre Gram-positivas y Gram-negativas puede deberse a las
diferencias estructurales de la pared celular puesto que estas últimas son más complejas y los
extractos no logran penetrarlas fácilmente o es posible que las concentraciones de los
principios activos no sean altos u estén siendo opacados por compuestos antagonistas (De
Lara-Isassi, 1999; Durand, 2001; Zubia et al., 2008; Kumar et al., 2009).
Así mismo, los resultados encontrados en las fracciones de Padina mexicana no sólo
pueden diferir por tales razones, si no que también los cambios estaciónales y geográficos
(ecología del sitio), la variabilidad morfológica de cada especie en análisis (madurez sexual y
ciclo de vida) y el método utilizado para obtener el extracto algal (polaridad de disolventes
utilizados) son factores que influyen en la potencialidad de los compuestos (Jin-Wen et al.,
1984; Moreau et al., 1984 y 1988; Ballesteros et al., 1992; De Lara-Isassi et al., 1999; Tuney et
al., 2006; Kandhasamy y Arunachalam, 2008; Chiheb et al., 2009).
Cabe señalar que la posible presencia de phlorotaninos en las fracciones CCA11,
CCA12 y CCA13 pudiere ser causa de tales efectos positivos en las pruebas antimicrobianas,
ya que según Nagayama y colaboradores (2002) establecen que la actividad bactericida de los
phlorotaninos tiende a incrementar con la polimerización del phloroglucinol y la actividad
antimicrobiana de los polifenoles puede ser resultado de la interacción de estos con las
enzimas y proteínas de las bacterias.
Como se puede ver en la gráfica 1 el stock de phloroglucinol mostró IC50 = 0.012
mg/mL lo cual es comparable con el valor de la fracción CCAF13 la cual mostró un IC50 = 0.023

42
mg/mL (gráfica 5, tabla II). Por su parte, las fracciones CCAF10 y CCAF11 mostraron un IC50 =
0.20 mg/mL y 0.22 mg/mL respectivamente mostrando semejanza entre si, pero al ser mayores
al stock de phloroglucinol resultan menos activas (gráficas 2 y 3, Tabla II); en la gráfica 4 se
observa como la fracción CCAF12 mostró un valor IC50 = 0.13 mg/mL.
En cuanto a los resultados mencionados anteriormente y los reportados por
Amornlerdpison y colaboradores (2007) con el extracto de Padina minor, existe discordancia
puesto que dichos autores presentan en los ensayos contra el radical libre DPPH un IC50 = 6.92
± 0.16 mg/mL (expresado en mg equivalentes de ácido gálico) y los valores del presente
estudio son mucho menores. De igual manera, Zubia y colaboradores (2007) determinan en
Padina gymnospora la captación del radical DPPH a un IC50 = 3.45 mg/mL lo cual también
difiere con los ensayos espectrofotométricos. En la tabla II se evidencia que las fracciones del
extracto de Padina mexicana muestran mayor actividad que las especies anteriores pues se
requiere de una menor cantidad de estas para inhibir el radical libre estable 2,2-difenil-1-
picrilhidracilo.
No obstante, Zubia y colaboradores (2007) también reportan contra el radical DPPH un
IC50 = 0.32 ± 0.01 mg/mL y un contenido de fenoles de 29.18 ± 0.32 (en peso seco) en el alga
parda Lobophora variegata perteneciente a la misma familia de Padina mexicana; en sus
análisis, el valor de IC50 de L. variegata es comparado con los antioxidantes comerciales
sometidos al ensayo con DPPH, tales como α-tocopherol (0.31 ± 0.03 mg/mL), BHT (0.16± 0.01
mg/mL), ácido ascórbico (0.09 ± 0.02 mg/mL) y BHA (0.06 ± 0.02 mg/mL). Se puede inferir con
lo anterior, que los valores de las fracciones CCAF10, CCAF11 y CCAF12 (tabla II) son
significativamente concordantes con los del alga L. variegata y los antioxidantes comerciales,
por tanto son igualmente activas contra el radical DPPH y es posible que contengan
compuestos de origen fenólico.
La diferenciación entre los resultados en los análisis de otros autores y el presente
estudio tal vez se deba a que la polaridad de los disolventes con que se obtuvieron los
extractos en cada caso hace que incremente o descienda su efectividad y también el hecho de
que el stock utilizado en cada análisis es diferente (De Lara-Isassi et al., 1999; Matanjun et al.,
2008).
La prueba bioautográfica (figura 19) muestra el comportamiento de las fracciones en la
placa cromatográfica y es notable la coloración amarilla intensa de CCAF10, CCAF11, CCAF12
y CCAF13 las cuales reaccionaron inmediatamente después del rocío con DPPH; sin embargo
las fracciones CCAFU1-2, CCAF3, CCAF4, CCAF5, CCAFU6-7 y CCAF9 denotan ligera
coloración amarilla en este ensayo pero no mostraron actividad en los análisis
espectrofotométricos. Por su parte, la facción CCAF8 no mostró actividad en ninguno de los

43
dos análisis. El hecho de que no hayan mostrado actividad puede deberse a la propia
pigmentación de las fracciones (en cuanto a la absorbancia) o a la presencia de un número alto
de compuestos fenólicos en la fracción inicial (ESL1F2) con estructuras químicas muy
parecidas lo que provoca que en varias fracciones aparezcan los mismos metabolitos en
diferente cantidad (Novoa et al., 2001; Chandrasekar et al., 2005).
Otro aspecto que es importante mencionar, es el sinergismo que puede existir entre los
diferentes compuestos presentes en las fracciones; según Le Tutour y colaboradores (1998) los
compuestos identificados con efecto sinérgico en sus pruebas antioxidantes y pro-oxidantes
más potentes son los pigmentos no polares, especialmente la clorofila a y compuestos
liposolubles relacionados. De igual manera Zhang y colaboradores (2007) menciona que no
solamente compuestos de origen fenólico pueden estar presentes en este tipo de análisis
puesto que es posible la presencia de otros componentes como ácido ascórbico, carotenoides
y tocoferol. Por lo tanto, las fracciones obtenidas del extracto etanólico de Padina mexicana
sometidas al presente análisis no se encontraban puras por lo que se pudiere pensar que
existen otras moléculas en sinergismo con los phlorotaninos siendo causantes de los efectos
positivos.
9. Conclusiones.
• El compuesto parcialmente puro obtenido de CCAF8 derivó del fraccionamiento por
cromatografía en columna. La elección de esta fracción para su procesamiento se basó
en la actividad antimicrobiana observada, la cantidad de la fracción de 330.6 mg y su
fácil solubilidad. Es necesario dar seguimiento a su análisis.
• La fracción CCAF13 presentó potencialidad para reducir radicales libres semejante al
phloroglucinol en los análisis espectrofotométricos.
• Las fracciones CCAF10, CCAF11 y CCAF12 mostraron actividad antioxidante en
menor potencialidad en comparación con el phloroglucinol.
• La presencia de compuestos de origen fenólico en las fracciones de Padina mexicana
probadas contra las cepas pudieron causar efecto bactericida y la actividad
antioxidante contra el radical DPPH. Se puede asumir que la fracción CCAF13 pudiere
contener metabolitos de origen fenólico ya que al observar a la par los análisis de
actividad antimicrobiana (tabla I) bioautográficos (figura 19) y espectrofotométricos
(gráfica 5, tabla II) la fracción CCAF13 muestra un comportamiento semejante al
phloroglucinol e inhibe a S. aureus (ATCC BAA-42) y S. pyogenes (ATCC BAA-946)
(tabla I).

44
• Todas las fracciones derivadas de Padina mexicana presentaron actividad
antimicrobiana al menos contra un microorganismo Gram-positivo principalmente
contra S. pyogenes (ATCC BAA-946), siendo mayor la actividad en CCAFU1-2,
CCAF3, CCAF4 y CCAF5 en comparación con CCAFU6-7, CCAF8, CCAF9, CCAF10,
CCAF11, CCAF12, CCAF13, CCAF14 y CCAF15.
• Es necesario realizar el análisis cuantitativo del contenido fenólico en las fracciones.

45
10. Literatura citada.
Avello M. y Suwalsky M. 2006. Radicales libres, estrés oxidativo y defensa antioxidante celular.
Ciencia ahora. Universidad de Concepción. Chile. No 17. Año 9. 6 pp.
Amsler C.D. 2008. Algal chemical ecology. Springer-Verlag Berlin Heildelberg. 313pp.
Amornlerdpison D., Peerapornpisal, Y., Taesotiikul, T., Jamjai, U., Nualchareo, M.,
Kanjanapothi, D. 2007. Antioxidan activity of Padina minor Yamada. KMITL Science and
Technology Journal. Vol. 7(S1): 1-7 pp.
Bakus G. J. 1985. Chemical ecology of marine organisms: An Overview. Journal of Chemical
Ecology. Vol. 12(5): 951-987.
Ballesteros E., Martin D. y Uriz M. J. 1992. Biological activity of extracts from Some
Mediterranean Macrophytes. Botánica Marina. Vol. 35: 481 -485 pp.
Blunt W. J., Copp R. B., Munro H. G. M., Northcotec T. P y Prinsepd R. M. 2006. Marine natural
products. Natural Products Report. Vol. 23: 26–78 pp.
Ceseña A. M. 2003. Fenología de Padina caulescens (Thivy) Taylor 1945 (Dictyotales:
Phaeophyta) en la playa “concha”, B.C.S., México. Universidad Autónoma de Baja
California Sur. México. Tesis. 43 pp.
Chandrasekar D., Madhhusudhana k., Ramakrishna S., Diwan V.P. 2005. Determination of
DPPH free radical scavenging activity by reversed –phase HPLC : A sensitive screening
method for polyhereval formulations. Pharmaceutical and Biomedical Analysis. Vol. 40:
460-464 pp.
Chang-Seon M., Hyeon-Cheol S. Hai Y.B., Soo J.Y., Bong H.L., Jong S.K.2005. Improvement
of memory by dieckol and phlorofucofuroeckol in ethanol-treated mice: possible
involvement of the inhibition of acetylcholinesterase. Archives parmacal Research. Vol.
28(6): 691- 698 pp.
Chávez P. L. 2000. Evaluación taxonómica de las especies del género Padina Adarson 1763
(Dictyotales: Phaeophyta) para el golfo de California. Instituto Politécnico Nacional. Tesis.
México. 93 pp.
Chiheb I., Riadi H., Martinez L. J., Dominguez S.J.F., Gomez V.J.A., Bouziane H. y Kadiri M.
2009.Screening of antibacterial activity in marine green and brown macroalgae from the
coast of Morocco. African Journal of Biotechnology. Vol. 8 ( 7) :1258-1262 pp.
Chkhikvishvili I.D. y Ramazanov Z.M. 2000. Phenolic sbstances of brown algae and their
antioxidant activity. Applied biochemistry and microbiology. Vol. 36 (3): 289-291 pp.

46
Dabanch P.J., Herero C. D., Pavéz A.C., Veas P. N., Braun J. S. y Porte T.L. 2009. Bacteriemia
por Vibrio parahaemolyticus: reporte de caso y revisión de la literatura. Revista Chilena
Infectología. Vol 26:(4) 360-362 pp.
Dewick M. P. 2002. Medicinal natural product: A biosynthetic approach. John Wiley & Sons Ltd.
UK. 507 pp.
Džidič S. Šuškovič y Blaženka K. 2008. Antibiotic resistance mechanisms in bacteria:
biochemical and genetics aspects. Food technology biotechnology. Vol. 46 (1): 11-21 pp.
Garateix A. 2005. El mar: fuente de nuevos fármacos. Ciencia y Cultura. Vol. 12 (58): 39-47
pp.
Gin-Nae A., Kil-Nam K., Seon-Heui C., Choon-Bok S., Jehee L., Moon-Soo H., In-Kyu Y., Nam-
Ho L., Young-Heun J., Jin-Soo K., Min-Soo H.y You-Jin J. 2007. Antioxidants activites of
phlorotannins purified from Ecklonia cava on free radical scavenging using ESR and H202-
meditated DNA damage. European food research. Vol. 226:71–79 pp.
Gutiérrez V. J. R. 2002. Daño oxidativo, radicales libres y antioxidantes. Revista Cubana
Medicina Militar. Vol. 31(2):126-133 pp.
De Lara-Isassi, G. Sobrino-Figueroa, A. Lozano-Ramírez, C., Ponce-Marques, M.E. y
Dreckman E.Y. 1989. Evaluación de la actividad antibiótica de las microalgas de las
costas de Michoacán, México. Boletine institute oceanography. Vol. 28(1-2): 99-104 pp.
De Lara-Isassi, G. y Álvarez-Hernández, S. 1999. Nuevas adiciones al conocimiento de la
actividad antibiótica de macroalgas Mexicanas. Hidrobiológica . Vol. 9 (12): 159-169 pp.
De Lara- Issasi, Álvarez-Hernández S. y Collado-Vides L. 2000. Ichtyotoxic activity of extracts
from Mexican marine macroalgae. Journal of Applied Phycology. Vol. 12: 45–52 pp.
Durand E. L. 2001. Propiedades antibacterianas de algunas macroalgas de la costa de
Veracruz. Universidad Autónoma Metropolitana. México, D.F. Tesis. 30 pp.
González B. E. A., Charles B. M., Filho M.J., Novoa V. 2009. Las algas marinas como fuentes
de fitofarmacos. Revista cubana de plantas medicinales. Vol. 14 (2): 64-81pp.
Gonzáles del Val A., Platas G., Basilio A., Cabello A., Gorrochategui J., Suay I., Vicente F.,
Portillo E., Jiménez del Río M., García R.G., Peláez F. 2001. Screening of antimicrobial
activities in red, green and brown macroalgae from Gran Canaria (Canary Islands, Spain).
International microbiology. Vol. 4: 35-40 pp.
Isaza M. J. H. 2007. Taninos o polifenoles vegetales. Scientia et Technica. Vol. 8(2): 13-18 pp.
Jing-wen M. y Wei-ci T. 1984. Screening for antimicrobial activities in marine algae from the
Qingdao coast, China. Hydrobiología. Vol. 22: 517–520 pp.

47
Kandhasamy M. y Arunachalam K. D. 2008. Evaluation of in vitro antibacterial property of
seaweeds of southeast coast of India. African journal of biotechnology. Vol. 7 (12): 1958-
1961 pp.
Kang K., Park Y., Hwang J.H., Kim H.S., Lee G.J. Shin C.H. 2003 .Antioxidative properties of
brown algae polyphenolics and their perspectives as chemopreventive agents against
vascular risk factors. Archives of pharmacal research. Vol. 26 (4): 286-293 pp.
Kang H.S., Chung H. Y., Kim J. Y., Son B. W., Jung H. A. y Sue J. 2004. Inhibitory
phlorotannins from the edible brown alga Ecklonia stolonifera on total reactive oxygen
species (ROS) Generation. Archive pharmaceutical research. Vol. 27 (2): 194-198 pp.
Kubanek J., Jensen Paul R., Keifer P. A., Cameron S. M., Collins D. O. y Fenical W. 2002.
Seaweed resistance to microbial attack: A targeted chemical defense against marine
fungi. USA. Proceedings of the National Academy of Sciences. Vol. 100 (12): 6916–6921
pp.
Kumar R. S., Ramanathan G., Subhakaran M. y Inbaneson J. S.2009. Antimicrobial compounds
from marine halophytes for silkworm disease treatment. International journal of medicine
and medical sciences. Vol. 1(5): 184-191 pp.
Ktari L. y Guyot M. 1999. A cytotoxic oxysterol from the marine alga Padina pavonica (L.) Thivy.
Journal of applied phycology. Vol. 11: 511-513 pp.
Mabrouk S. S., EI-Shayeb N. M. A., EI-Refai A. H., Sallam L. A. R, Hamdy A. A. 1985. Inhibitory
activities of some marine algae on aflatoxin accumulation. Applied microbiology and
biotechnology. Vol. 22:152-155 pp.
Magallanes C., Córdova C. y Orozco R. 2003. Actividad antibacteriana de extractos etanólicos
de microalgas marinas de la costa central del Perú. Revista peruana de biología. Vol. 10
(2): 125-132 pp.
Matanjun P., Mohamed S., Mustapha M.N., Muhammad K., Ming H.C. 2008. Antioxidant
activities and phenolics content of eight species of seaweeds from north Borneo. Journal
applied phycology. Vol. 20:367-373 pp.
Moreau J., Pesando D y Caram B. 1984. Antifungal and antibacterial screening of Dictyotales
from the French Mediterranean coast. Hidrobiología. Vol. (116/117): 521-524 pp.
Moreau J., Pesandol D., Bernard P., Caram B. y Pionnat J. C. 1988. Seasonal variations in the
production of antifungal substances by some dictyotales (brown algae) from the french
mediterranean coast. Hydrobiology. Vol. 162: 157-162 pp.
Mohsen A M.S., Mohamed S. F., Ali F.M. y El-Sayed O. H.. 2007. Chemical structure and
antiviral activity of water-soluble sulfated polysaccharides from Surgassum latifolium.
Journal of applied sciences. Vol 3 (10): 1178-1185 pp.

48
Mosquera O. M., Niño O. J., Correa Y. M. y Bejarano B. 2005. Estandarización del método de
captura de radicales libres para la evaluación de la actividad antioxidante de extractos
vegetales. Scientia et técnica. Año XI, No. 27: 231-234 pp.
Nagayama K., Iwamura Y., Shibata T., Hirayama I. y Nakamura T. 2002. Bactericidal activity of
phlorotannins from the brown alga Ecklonia kurome. Journal of antimicrobial
chemotherapy. Vol. 50: 889–893 pp.
Novoa V. A., Motidome M., Filho M.J., Linares F.A., Tanae M.M., Torres B.M.L., Lapa J.A.
2001. Actividad antioxidante y ácidos fenolicos del alga marina Bryothamnion triquetrum
(S.G. Gmelim) Howe. Brazilian journal of pharmaceutical sciences. Vol. 37 (3): 373- 382
pp.
Lee S.M., Na M.K., An R.B., Min B.S., Lee H.K. 2003. Antioxidant activity of two phloroglucinol
derivatives from Dryopteris crassirhizoma. Biological pharmaceutical bulletin of Korea.
Vol. 26(9): 1354-1356 pp.
Le Tutour B., Benlismane F., Gouleau M.P., Gouygou J.P., Saadan B., Quemeneur F. 1998.
Antioxidant and pro-oxidant activities of the brown algae, Laminaria digitata, Himantharia
elongata, Fucus vesiculosus, Fucus serratos and Ascophyllum nodosum. Journal applied
phycology. Vol. 10: 121-129pp.
Lüning K. 1990. Seaweeds: their environment, biogeography and ecophysiology. Wiley-
Intercience publication. New York, USA. 527 pp.
Paul V. J. 1988. Marine chemical ecology and natural products research. Memories of the
California Academy of Sciences. Ed. Daphne, G. F. San Francisco, California. 23-27 pp.
Pakrash A. Rigelhof F. y Miller E. 2001. Antioxidant activity. Medallion Laboratories. EUA.
Analitical progress. Vol. 19( 2). 6 pp.
Quang-To L., Young L.,Zhong-Ji Q., Moon-Moo K., Se-Kwon K. 2009. Inhibitory effects of
polyphenols isolated from marine alga Ecklonia cava on histamine releasa. Process
biochemistry. Vol. 44: 168-176 pp.
Riosmena-Rodríguez R., Paúl-Chávez L., Hernández- Carmona G., Lopez-Vivas J.M, y Casas-
Valdez M. 2009. Taxonomic Reassessment of the Genus Padina (Dictyotales,
Phaeophyta) from the Gulf of California. Algae. Vol. 24(4): 213-229 pp.
Rosales R. A. y Rodríguez, B. J. R. 2006. Evaluación de la actividad antioxidante de polifenoles
de algas marinas. Manual de Prácticas de Laboratorio de Ficología. Facultad de Ciencias
del Mar. España. 10 pp.
Rosell G. K., Srivastava, M. L. 1987. Fatty acids as antimicrobial substances in brown algae.
Hydrobiologia .151-152 (1): 471-475 pp.

49
Sá-Leáo R., Sanches S. I., Dias D., Peres I., Barros R.M., De Lencastre H. 1999. Detection of
an archaic clone of Staphylococcus aureus with low-level resistance to methicilin in a
pedriatric hospital in Portugal and international samples : relics of a formerly widely
disseminated train?. Journal of clinical microbiology. Vol. 37(6):1913-1920 pp.
Scheuer J. P. 1973. Chemistry of Marine Natural Products. Departament of chemistry university
of Hawaii. Honolulu, Hawaii. Academic press. 201 pp.
Shibata T. , Kawaguchi S., Hama Y., Inagaki M. , Yamaguchi K. y Nakamura T. 2004. Local and
chemical distribution of phlorotannins in brown algae. Journal of applied phycology .
Vol.16: 291–296 pp.
Shibata T., Ishimaru K., Kawaguchi S., Yoshikawa H. y Hama Y. 2008. Antioxidant activities of
phlorotannins isolated from Japanese Laminariaceae. Journal of applied phycology. Vol.
20:705–711 pp.
Soo-Jin H., Jong-Pyung K.,Won-Kyo J., Nam-Ho L.,Hahk-Soo K., Eun-Mi J., Soon-Hye P.,
Sung-Myung K., Young-Jae L., Pyo-Jam P. y You-Jin Y. 2008. Identification of chemical
structure and free radical scavenging activity of diphlorethohydroxycarmamol isolated
from a brown alga, Ishige okamurae. Journal of microbiology and biotechnology. Vol.
18(4):676-681 pp.
Soo-Jin H. y You-Jin Y. 2008. Radical scavenging capacity and cytoprotective effect on
enzimatic digests of Ishige okamurae. Journal Applied Phycology. Vol. 20:1087-1095pp.
Sugiura Y., Takeuchi Y., Kakinuma M. y Amano H. 2006. Inhibitory effects of seaweeds on
histamine release from rat basophile leukemia cells (RBL-2H3). Fisheries science. Vol.
72: 1286-1291 pp.
Sugiura Y., Matsuda K., Yamada Y. Nishikawa M, Shioya K., Katsuzaki H., Imai K. y Amano H.
2007. Anti-Allergic phlorotannins from the edible brown alga, Eisenia Arborea. Food
science and technology research. Vol. 3(1): 54-60 pp.
Tenover F.C. 2006. Mechanisms of antimicrobial resitance in bacteria. The american journal of
medicine. Vol. 199(6A): 4-10 pp.
Truus K., Vaher M., Koel M., Mähar A., Taure I. 2004. Analysis of bioactive ingredients in the
brown alga Fucus vesiculosus by capillary electrophoresis and neutron activation
analysis. Analytical and bioanalytical chemistry. Vol. 379: 849–852 pp.
Tuney Ü., Adirci B. H., Nal D., Sukatar A. 2006.Antimicrobial activities of the extracts of marine
algae from the coast of Urla (Üzmir, Turkey). Turkish journal of biology. Vol. 30, 171–175
pp.
Van Den Hoek C., Mann D.G. y Jahns H. 1995. Algae: An introduction to phycology.Cambridge,
University press. USA. 623 pp.

50
Zhang W.W., Duan X.J., Huang H.L., Zhang Y. y Wang B.G. 2007. Evaluation of 28 marine
algae from the Qingdao coast for antioxidative capacity and determination of antioxidant
efficiency and total phenolic content of fractions and subfractions derived from
Symphyocladia latiuscula (Rhodomelaceae). Journal applied phycology. Vol. 19:97–108
pp.
Zubia M., Robledo D., Pelegrin F. Y. 2007. Antioxidant activities in tropical marine maacroalgae
from the Yucatán Peninsula, Mexico. Journal applied phycology. Vol. 19: 449-458 pp.
Zubia M., Payri C. y Deslandes E. 2008. Alginate, mannitol, phenolic compounds and
biological activities of two range-extending brown algae, Sargassum mangarevense and
Turbinaria ornata (Phaeophyta: Fucales), from Tahiti (French Polynesia). Journal applied
phycology . Vol. 20:1033–1043 pp.
Zou Y., Ji Qian Z., Li Y., Kim M., Lee S. y Kim S. 2008. Antioxidant effects of phlorotannins
isolated from Ishige okamurae in Free Radical Mediated Oxidative Systems. Journal of
Agricultural and food chemistry. Vol. 56:(16):7001-7009 pp.
























![Fisiología Vegetal. Seminario Terpenos.[1] (1)](https://img.dokumen.tips/doc/110x75/557211c5497959fc0b8f7873/fisiologia-vegetal-seminario-terpenos1-1.jpg)




