Embed Size (px)
Citation preview

Understanding the role of L-type lectin receptor
kinases in Phytophthora resistance
Yan Wang

Thesis committee
Promotor
Prof. Dr Francine P.M. Govers
Personal chair at the Laboratory of Phytopathology
Wageningen University
Co-promotors
Prof. Dr Weixing Shan
Professor of Plant Protection
Northwest A&F University, Yangling, Shaanxi, China
Dr Klaas Bouwmeester
Research fellow, Laboratory of Phytopathology
Wageningen University
Other members
Prof. Dr M. Eric Schranz, Wageningen University
Dr A.F.J.M. (Guido) van den Ackerveken, Utrecht University
Dr Karin I. Posthuma, Enza Zaden R&D BV, Enkhuizen
Dr Vivianne G.A.A. Vleeshouwers, Wageningen University
This research was conducted under the auspices of the Graduate School of
Experimental Plant Sciences

Understanding the role of L-type lectin receptor
kinases in Phytophthora resistance
Yan Wang
Thesis
submitted in fulfilment of the requirements for the degree of doctor
at Wageningen University
by the authority of the Rector Magnificus
Prof. Dr M.J. Kropff,
in the presence of the
Thesis Committee appointed by the Academic Board
to be defended in public
on Monday 24 November 2014
at 4 p.m. in the Aula.

Yan Wang
Understanding the role of L-type lectin receptor kinases in Phytophthora resistance,
216 pages.
PhD thesis, Wageningen University, Wageningen, NL (2014)
With references, with summaries in English and Dutch
ISBN: 978-94-6257-132-7

Contents
Chapter I General introduction 7
Chapter II Characterization of the Phytophthora capsici-Arabidopsis pathosystem
IIa A novel Arabidopsis-oomycete pathosystem; differential interactions with Phytophthora capsici reveal a role for camalexin, indole glucosinolates and
salicylic acid in defence 28
IIb Induced expression of defence-related genes in Arabidopsis upon infection
with Phytophthora capsici 56
Chapter III Phenotypic analyses of Arabidopsis T-DNA insertion lines and expression profiling reveal that multiple L-type lectin receptor kinases are involved in plant immunity 63
Chapter IV Phytophthora resistance conferred by Arabidopsis L-type lectin receptor
kinases LecRK-IX.1 and LecRK-IX.2 is not associated with activation of
plant cell death 97
Chapter V Ectopic expression of Arabidopsis L-type lectin receptor kinase genes
LecRK-I.9 and LecRK-IX.1 confers Phytophthora resistance in
Nicotiana benthamiana 139
Chapter VI L-type lectin receptor kinases in Nicotiana benthamiana and tomato
and their role in Phytophthora resistance 155
Chapter VII General discussion 183
Summary 203
Samenvatting 206
Acknowledgements 209
Curriculum vitae 211
Publications 212
Education statement of the Graduate School 213


Chapter I
General introduction

Chapter I
8
INTRODUCTION
Plants are constantly attacked by a wide variety of microbes. Natural selection
shapes plants resistant to the majority of microbes, and thus plant diseases are
usually an exception rather than a rule. Plant diseases are often a major constraint to
crop production and those that destroy important staple crops are of great threat to
food security (Strange and Scott, 2005; Fisher et al., 2012). Several catastrophic
plant diseases have exerted profound socioeconomic impact on human history. For
example, the Great Irish Famine in the 1840s was due to the emergence of potato
late blight, and led to over a million people dying of starvation. The Great Bengal
Famine in 1943 is another disaster which caused starvation of over two million
people because of the failure of rice production after attack by the fungal pathogen
Cochliobolus miyabeanus (Strange and Scott, 2005; Vurro et al., 2010). Integrated
disease management is of vital importance to reduce losses caused by plant
pathogens, but yet a continuous challenge. Improvement of plant resistance by
breeding is hitherto considered as one of the most effective strategies to protect
crops from diseases. A major challenge for breeders is to obtain “durable resistance”,
and therefore a continuous introgression of novel resistance genes in breeding
programs is required. Exploring versatile disease resistance components in plants as
well as integrated deployment of these resistance components could be the key
leading to durable and broad-spectrum plant resistance.
Arms race between plants and pathogens
Disease resistance in plants is mediated by both preformed and induced defence.
Preformed defence, such as those conferred by cell walls and toxic secondary
metabolites, provides physical and chemical barriers to impede microbial penetration
(Huckelhoven, 2007; Underwood, 2012). However, pathogens developed strategies
to overcome these barriers, for example by secreting hydrolytic enzymes to
decompose plant cell walls, thereby facilitating pathogen invasion (Nuhse, 2012).
Once the layer of preformed plant defence has been breached, pathogens are then
confronted with another barrier raised by various inducible plant defence responses
(Jones and Dangl, 2006).
Unlike animals, plants lack adaptive immunity. Nevertheless, plants have
developed a sophisticated innate immune system to defend themselves against
potential pathogens. Recognition of conserved microbial structures known as
microbe-associated molecular patterns (MAMPs) by cell surface receptors
constitutes the first layer of innate immunity (Jones and Dangl, 2006; Zipfel, 2009).
MAMPs are not only present in pathogenic microbes but also in non-pathogenic or
even beneficial microbes (Mackey and McFall, 2006; Boller and Felix, 2009). MAMPs

General introduction
9
are in general pivotal for the survival of microbes, and therefore they are unlikely to
be modified by microbes to evade plant recognition because of severe fitness
penalties (Naito et al., 2008; Zhang and Zhou, 2010; Thomma et al., 2011). MAMPs
are perceived as “non-self” molecules by plant pattern recognition receptors (PRRs)
(Dubery et al., 2012). Besides, host-derived compounds released during pathogen
infection or wounding, termed damage-associated molecular patterns (DAMPs), can
also be recognized by PRRs, as “modified-self” molecules (Boller and Felix, 2009).
Thus far, most of the identified PRRs are plasma membrane-localized receptor-like
kinases (RLKs) or receptor-like proteins (RLPs) (Schwessinger and Ronald, 2012;
Zipfel, 2014). Recognition of MAMPs/DAMPs leads to activation of PRR-mediated
downstream signal transduction that ultimately results in basal resistance, referred to
as pattern-triggered immunity (PTI). The PTI-associated immune responses include
reactive oxygen species (ROS) bursts, ion fluxes, callose deposition, defence-related
gene expression and secondary metabolite production (Boller and Felix, 2009). This
layer of defence is considered to play a major role in circumventing microbial
invasions in most cases (Boller and Felix, 2009; Dubery et al., 2012).
To achieve successful infection, pathogens acquired the capacity to deliver
effectors into plants to suppress PTI. Effectors interfere with plant defence by
sequestering MAMPs, by blocking or degrading PRRs or by hijacking key
components in PRR-mediated signaling (Boller and He, 2009; de Jonge et al., 2010;
Dou and Zhou, 2012; Giraldo and Valent, 2013; Pel and Pieterse, 2013). In turn,
plants are able to recognize specific pathogen effectors by an additional class of
immune receptors, known as resistance (R) proteins (Jones and Dangl, 2006).
Recognition occurs either directly through the interaction between R proteins and
matching effectors, or indirectly through the perception of modifications in the host
targets of effectors by R proteins (van der Hoorn and Kamoun, 2008; Krasileva et al.,
2010). Defence responses triggered upon recognition of pathogen effectors by R
proteins are similar to those associated with PTI, but differ in timing and magnitude
(Dubery et al., 2012). R protein-mediated recognition triggers stronger host defence
responses, often culminating in a hypersensitive response (HR), a form of
programmed cell death (Chisholm et al., 2006). This type of resistance is termed
effector-triggered immunity (ETI) and plays a dominant role in plant resistance (Jones
and Dangl, 2006). However, ETI only occurs when a matching R/effector pair is
present. Lacking either of these two components will lead to plant susceptibility.
Therefore, ETI is also known as race- or cultivar-specific resistance. Recognition by
R proteins has been a driving force for the evolution of pathogen effectors (Trivedi
and Wang, 2014). To counteract ETI, pathogens developed several strategies, for
example generating effector variants by mutations, silencing effector genes or
secreting additional virulence factors to suppress R-gene function (Chisholm et al.,

Chapter I
10
2006; Jones and Dangl, 2006; van Poppel et al., 2008; Takken and Rep, 2010;
Kasuga and Gijzen, 2013).
Phytophthora species: notorious plant pathogens
The “fungus-like” Phytophthora pathogens belong to the oomycetes, filamentous
organisms which are evolutionally distant from fungi and more closely related to
photosynthetic brown algae and diatoms (Baldauf, 2003). The first Phytophthora
species described in 1876 was Phytophthora infestans. Many other Phytophthora
species have been identified thereafter (Erwin and Ribeiro, 1996). In the last
decades, large scale surveys and advanced strategies for species delimitation have
resulted in a significant increase in the number of described Phytophthora species
(Cooke et al., 2007; Kroon et al., 2012; Martin et al., 2012; Martin et al., 2014). To
date, over 120 species are known which are divided into ten clades based on
combined phylogenetic analysis of mitochondrial and nuclear multilocus traits (Martin
et al., 2014) (Fig. 1).
All identified Phytophthora species are destructive plant pathogens (Kroon et
al., 2012). Most of them are hemibiotrophic; they behave like biotrophic pathogens in
the early stage of infection but act as necrotrophs in a late stage (Bouwmeester et
al., 2009). The most notorious species is P. infestans, the causal agent of potato and
tomato late blight, which causes billions of dollars loss annually through damage in
potato and tomato production (Fry, 2008; Nowicki et al., 2011). Infection on tomato
and potato by P. infestans occurs on leaves, stems and fruits/tubers at any stage of
development, even after harvest (Fig. 2, A and B). Infection causes dark necrotic
lesions that expand quickly and is accompanied by the production of massive
numbers of sporangia that are spread by wind and water (Fry, 2008). Subsequently,
sporangia will germinate directly or indirectly to start new infection cycles (Judelson
and Blanco, 2005). Phytophthora sojae is another economically important species
causing disease mainly on soybean (Tyler, 2007). It causes seed decay, “damping
off” of seedlings and root and stem rot of mature plants (Fig. 2C). While P. infestans
and P. sojae are species with narrow host ranges, others are capable of infecting
many plants across different families. For example, Phytophthora capsici, a highly
dynamic species, causes rot on roots, foliage and fruits of a number of important
vegetables belonging to the Solanaceae and Cucurbitaceae families (Lamour et al.,
2012a; Lamour et al., 2012b) (Fig. 2D). In addition to damaging crops, many
Phytophthora pathogens cause devastating diseases on woody plants (Hansen et al.,
2012). For example, the invasive species, Phytophthora ramorum, is the causal
agent of sudden oak death and ramorum blight on trees and woody ornamentals
(Fig. 2E). P. ramorum is a species with a wide-host-range that can infect over
hundred plant species (Grünwald et al., 2008). The economic impact caused by

General introduction
11
Figure 1. Phylogeny of Phytophthora species.
The tree on the left is reproduced from Martin et al. (2014). Phytophthora species mentioned in this
chapter are listed on the right. Black arrowheads point to the position in the phylogenetic tree. Species
marked by ♦ are reported capable to infect Arabidopsis.

Chapter I
12
P. ramorum was estimated to be in the tens of millions of dollars in the USA
(Grünwald et al., 2008). Due to the substantial economic and ecological impact,
P. ramorum is often highlighted in the same rank as P. infestans in case of destructive
potential (Hansen et al., 2012).
Figure 2. Disease symptoms caused by different Phytophthora species.
A-B, Late blight symptoms caused by P. infestans on tomato (A) and potato (B). C, Stem and root rot
of soybean caused by P. sojae. D, Disease caused by P. capsici on pepper and tomato. E, Sudden
oak death caused by P. ramorum. Pictures are courtesy of Klaas Bouwmeester and Francine Govers
(panels A, B and E) and Anne E. Dorrance (panel C).
During interaction with plants, Phytophthora pathogens secrete effectors to
modulate plant defence and facilitate infection (Kamoun, 2006). Genome sequence
analysis revealed that Phytophthora genomes encode a large repertoire of effectors
that can be divided into two categories, i.e. apoplastic and intracellular effectors
(Tyler et al., 2006; Jiang et al., 2008; Haas et al., 2009). Apoplastic effectors are
instrumental in protecting pathogens against apoplastic defence and in mediating
invasion (Stassen and Van den Ackerveken, 2011). For example, some apoplastic
effectors inhibit host hydrolytic enzymes while others disrupt plant cell wall integrity to
facilitate plant infection (Tian et al., 2007; Kaschani et al., 2010; Bouwmeester et al.,
2011; Stassen and Van den Ackerveken, 2011). Intracellular effectors are currently
classified into two types, i.e. RXLR-effectors and Crinklers (CRN). RXLR-effectors
are named after a conserved four amino acid (R: Arginine, X: any amino acid, L:

General introduction
13
Arginine) motif in the N-terminus that is considered to be important for effector
translocation into hosts, while CRN effectors contain yet another conserved motif,
namely LXLFLAK (L: Leucine, X: any amino acid, F: Phe, A: Ala, K: Lys) in the N-
terminus also with a potential translocation function (Dou et al., 2008a; Govers and
Bouwmeester, 2008). The genome of P. infestans contains 563 genes encoding
RXLR effectors and 196 genes encoding CRN effectors (Haas et al., 2009), whereas
the P. capsici genome contains 357 RXLR effector genes and 84 CRN genes
(Lamour et al., 2012b; Stam et al., 2013). P. sojae and P. ramorum have a similar
number of RXLR effector genes, but less CRN genes (Tyler et al., 2006; Jiang et al.,
2008). There is now ample evidence showing that multiple RXLR effectors as well as
several CRN effectors are capable of suppressing host innate immunity (Bos et al.,
2006; Dou et al., 2008b; Liu et al., 2011; Stassen and Van den Ackerveken, 2011;
Qiao et al., 2013; Zheng et al., 2014). In this respect, the large amount of effectors
may account for the rapid adaptation of Phytophthora pathogens.
Plant resistance against Phytophthora pathogens
Through co-evolution with pathogens, plants developed immune receptors that
monitor the presence of pathogens and mount effective innate immune responses.
So far, most of the identified immune receptors against Phytophthora pathogens
belong to the class of intracellular nucleotide-binding leucine-rich repeat (NLR)
proteins, termed R proteins. These proteins are characterized by the fact that they
recognize RXLR effectors in a “gene-for-gene” manner (Tyler, 2007). Therefore, R
proteins mediate resistance specifically to the Phytophthora races that secrete
matching RXLR effectors. Based on this specialized recognition, the implementation
of effectoromics and comparative genomics approaches accelerated R gene cloning
and specificity profiling (Vleeshouwers et al., 2008). Up to now, at least 23 and 17 R
genes have been cloned from Solanum species and soybean, respectively (Tyler,
2002; Li et al., 2011; Vleeshouwers et al., 2011; Lin et al., 2013; Zhang et al., 2013;
Sun et al., 2014). Unfortunately, race-specific resistance conferred by most of the
identified R genes proved to be not durable due to rapid adaptability of Phytophthora
pathogens (Fry, 2008; Vleeshouwers et al., 2011). Nevertheless, there are several
resistance genes, such as Rpi-blb1 isolated from Solanum bulbocastanum that
confer resistance to a broad spectrum of P. infestans isolates (Song et al., 2003; van
der Vossen et al., 2003). This could be due to the fact that the matching effector of
Rpi-blb1, i.e. IPI-O1 or its functional variants is present in all P. infestans isolates
(Champouret et al., 2009). Thus, R genes which recognize highly conserved effectors
are therefore more promising to confer broad-spectrum and durable resistance in
crops. Since the benefit of exploiting single R gene has been limited due to extremely
diversified Phytophthora field populations (Fry, 2008), stacking of multiple R genes is

Chapter I
14
anticipated to confer durable resistance to a broad set of isolates (Zhu et al., 2012).
Apart from recognizing effectors delivered into plant cells, plants are also
capable to recognize apoplastic effectors or Phytophthora cell wall components to
initiate defence responses (Tyler, 2002). For example, perception of INF1, the major
secreted elicitin of P. infestans, by N. benthamiana induces plant cell death and
compromises P. infestans infection (Kamoun et al., 1998). Plasma membrane-
localized receptors are considered as potential candidates to perceive pathogen-
secreted elicitors and activate plant defence (Dubery et al., 2012). Plant genomes
encode a large number of receptors, including RLPs and RLKs. Both RLPs and RLKs
contain diverse extracellular domains which define ligand specificity (Shiu and
Bleecker, 2003). RLKs contain a cytosolic kinase domain that can transduce signals
via phosphorylation of signaling partners. Given the lack of an obvious signaling
domain in the short cytoplasmic tail, RLPs are more likely to recruit (a) co-receptor(s)
for downstream signal transduction, as has been found for tomato protein Cf4, one of
the RLPs that function in resistance against the leaf mold fungus Cladosporium
fulvum (Liebrand et al., 2014). Another RLP from the wild potato species Solanum
microdontum, termed ELR, has recently been found to recognize INF1 and other
Phytophthora elicitins and seems to play a role in resistance against P. infestans (Du,
2014). The importance of RLKs in Phytophthora resistance has also been
demonstrated. For example, BAK1/SERK3, a RLK with leucine-rich repeats (LRR) in
the extracellular domain, acts as a central modulator of plant defence (Chinchilla et
al., 2009; Liebrand et al., 2014). BAK1/SERK3 plays an important role in transducing
signals upon recognition of elicitors secreted by Phytophthora pathogens. For
example, it is required for INF1-induced cell death (Chaparro-Garcia et al., 2011).
N. benthamiana plants silenced for NbSerk3 showed compromised INF1-induced cell
death and increased susceptibility to P. infestans. BAK1 is also implicated in defence
responses elicited by the cellulose binding elicitor lectin (CBEL), a glycoprotein
secreted by Phytophthora parasitica, and plays a role in Arabidopsis resistance to
P. parasitica (Larroque et al., 2013). Another RLK implicated in Phytophthora
resistance is the L-type lectin receptor kinase (LecRK) LecRK-I.9 in Arabidopsis
(Bouwmeester et al., 2011). Since this thesis focuses on LecRKs, LecRK-I.9 will be
described in more detail later.
Efforts have also focused on partial resistance to Phytophthora pathogens.
Several quantitative trait loci (QTL) have been identified conferring partial resistance,
for example in potato and soybean (Odeny et al., 2010; Johnson et al., 2012; Wang
et al., 2012; Mallard et al., 2013; Lee et al., 2014; Naegele et al., 2014). However, the
key genes governing QTL-mediated resistance and the molecular mechanisms
underlying the activation of partial Phytophthora resistance are largely unknown.
Phytophthora infection often triggers a subset of defence responses. ROS

General introduction
15
production is one of the early defence responses in plants upon pathogen infection
(Boller and Felix, 2009). The role of ROS in plant resistance against Phytophthora
pathogens was supported by the finding that silencing the genes encoding the
plasma membrane-bound NADPH oxidases RbohA and RbohB compromised
P. infestans induced H2O2 accumulation as well as Phytophthora resistance in
N. benthamiana (Yoshioka et al., 2003). Secondary metabolites and related signaling
cascades are also involved in plant resistance against Phytophthora pathogens. For
example, both salicylic acid (SA) and ethylene (ET) pathways are essential for
N. benthamiana resistance to P. infestans (Shibata et al., 2010).
Reducing host susceptibility by mutation of susceptibility genes provides
another avenue to improve plant resistance (Lapin and Van den Ackerveken, 2013).
Screening of Arabidopsis EMS mutants led to the identification of six independent
loci, termed dmr1 to dmr6, that showed increased resistance to the downy mildew
pathogen Hyaloperonospora arabidopsidis, a species that similar to Phytophthora
species belongs to the oomycete (van Damme et al., 2005). Mapped-based cloning
revealed that increased resistance of the dmr1 mutant is determined by mutations in
a single gene that encodes a homoserine kinase which is required for metabolizing
homoserine. Point mutations affected the enzyme activity of DMR1 and led to
increased accumulation of homoserine which is capable to induce Arabidopsis
resistance to H. arabidopsidis (van Damme et al., 2009). Similarly, increased
resistance of dmr6 is determined by mutations in a single gene that encodes 2-
oxoglutarate (2OG)-Fe(II)-dependent oxygenase (van Damme et al., 2008).
Microarray analysis revealed that DMR6 negatively regulates defence-related gene
expression. A role of the substrate of the DMR6 encoded 2OG-Fe(II) oxygenase in
pathogen inhibition was proposed but this needs to be confirmed. A study focused on
plant susceptibility genes to Phytophthora pathogens was also carried out. By mutant
screening, several Arabidopsis mutants have been found that show increased
resistance to Phytophthora parasitica (Shan, W., personal communication). Detailed
functional analysis of the corresponding genes is in progress and will help to uncover
how plants host Phytophthora pathogens.
Arabidopsis, a model to study plant-Phytophthora interaction
Arabidopsis has been used extensively as a model plant to dissect plant-pathogen
interactions. As a favourite model plant, Arabidopsis has many advantages such as
complete genome sequences, feasible molecular modification, large sets of gene
expression data and extensive collections of natural accessions and mutant lines
(Koornneef and Meinke, 2010). Although natural infection of Arabidopsis by
Phytophthora has not been documented, laboratory infection assays have shown that
several Phytophthora species are capable to infect Arabidopsis. In the case of

Chapter I
16
Phytophthora brassicae, Phytophthora cinnamomi and Phytophthora parasitica,
successful infection was observed, as well as interaction specificity among different
Phytophthora isolates and Arabidopsis accessions (Roetschi et al., 2001; Robinson
and Cahill, 2003; Attard et al., 2010; Wang et al., 2011). In contrast, P. infestans and
P. sojae appeared to be non-pathogenic on Arabidopsis (Huitema et al., 2003; Sumit
et al., 2012). Penetration of P. infestans induces non-host defence responses such as
cell death and induced defence-related gene expression (Huitema et al., 2003). To
decipher plant resistance mechanisms against Phytophthora pathogens, Arabidopsis
mutant resources have proven to be instrumental. For example, non-host resistance
to P. infestans was compromised in pen2 (Lipka et al., 2005), a mutant of a
peroxisome-associated myrosinase that hydrolyses the toxic plant secondary
metabolite indole glucosinolate (iGS). In pen2 or other iGS mutants, Arabidopsis
resistance to another Phytophthora species, i.e. P. brassicae, is also slightly
attenuated (Schlaeppi et al., 2010; Schlaeppi and Mauch, 2010). The role of iGS in
Arabidopsis resistance to P. brassicae became evident when introducing a PAD3
mutation into pen2 since the resistance of the pen2pad3 double mutant to
P. brassicae was significantly reduced (Schlaeppi et al., 2010; Schlaeppi and Mauch,
2010). PAD3 encodes a cytochrome P450 monooxygenase that is required for
camalexin biosynthesis (Schuhegger et al., 2006). Interestingly, the pad3 mutant
itself or other mutants defective in camalexin biosynthesis did not show significant
disease susceptibility to P. brassicae (Schlaeppi et al., 2010; Schlaeppi and Mauch,
2010). In this way, the combined effect of camalexin and iGS on Arabidopsis
resistance become evident since blocking biosynthesis of either compound is not
sufficient to render Arabidopsis susceptibility (Schlaeppi and Mauch, 2010). However,
Arabidopsis mutants do not always respond similarly to different Phytophthora
species or even different isolates of a certain species. For example, the ethylene-
insensitive mutant ein2 showed increased susceptibility to P. parasitica but not to
P. brassicae (Roetschi et al., 2001; Attard et al., 2010). This indicates that
Arabidopsis employs sophisticated strategies to orchestrate resistance against
different Phytophthora species. In the Phytophthora genus, the aforementioned
species are scattered over different clades (Fig. 1). Comparative analysis of
interactions of Arabidopsis with these diverse Phytophthora species will likely lead to
new insights into host determinants underpinning interactions with pathogens.
Lectin receptor kinases in plants
Lectin receptor kinases are typical RLKs that are composed of an extracellular lectin
domain, a transmembrane domain and an intracellular kinase domain. Based on the
lectin domain, three types of lectin receptor kinases are found in plants, namely G, C
and L types (Bouwmeester and Govers, 2009). G-type lectin receptor kinases, also

General introduction
17
known as S-locus RLKs contain a so-called Galanthus nivalis agglutinin (GNA)-
related lectin domain (Shiu and Bleecker, 2001). Members of this family have been
shown to be involved in plant self-recognition and plant defence (Sherman-Broyles et
al., 2007; Boggs et al., 2009; Gilardoni et al., 2011; Cheng et al., 2013). C-type lectin
receptor kinases contain a calcium-dependent lectin domain. They are rare in plants
and only one has been found in the Arabidopsis genome with yet unknown function
(Bouwmeester and Govers, 2009). The third type of lectin receptor kinases, defined
as LecRKs, contain a lectin domain that structurally resembles soluble legume lectins
(Barre et al., 2002). LecRK-I.9, one of the Arabidopsis LecRKs, was identified as a
potential host target of the P. infestans RXLR effector IPI-O and is involved in
mediating plasma membrane-cell wall adhesions (Gouget et al., 2006). Mutating
LecRK-I.9 reduces cell wall-plasma membrane adhesion and renders Arabidopsis
susceptible to non-adapted P. brassicae isolates. LecRK-I.9 is also implicated in
mediating plant responses upon PAMP perception. For example, flg22-triggered
callose deposition is reduced in LecRK-I.9 mutants (Bouwmeester et al., 2011).
LecRK-I.9 is not only functional as a resistance component in Arabidopsis, but also
confers resistance to P. infestans when transferred into potato (Bouwmeester et al.,
2014). Recently, LecRK-I.9 was identified as the first extracellular ATP (eATP)
receptor in plants and was found to be required for eATP-mediated responses,
including Ca2+ fluxes, mitogen-activated protein kinase activation and gene
expression (Choi et al., 2014). However, whether LecRK-I.9-mediated Phytophthora
resistance is dependent on eATP recognition remains elusive. LecRK-I.9 belongs to a
multigene family consisting of 45 members in Arabidopsis (Bouwmeester and
Govers, 2009). Several of these LecRKs are induced upon exposure to pathogens or
pathogen-derived elicitors, indicating involvement of LecRKs in plant defence.
OUTLINE OF THIS THESIS
The research described in this thesis focuses on the role of LecRKs in plant
resistance to Phytophthora pathogens and defence signaling cascades downstream
of LecRKs. The function of LecRKs in different plant species was analysed and the
potential of LecRKs to confer Phytophthora resistance when transferred to other
plant species was also addressed.
In Chapter II, we describe a novel Phytophthora-Arabidopsis pathosystem.
We monitored the ability of Phytophthora capsici to infect Arabidopsis and analysed
the interaction specificity in different P. capsici isolate-Arabidopsis accession
combinations. We assessed the role of secondary metabolite-based defence in
Arabidopsis resistance to P. capsici by performing infection assays on mutants

Chapter I
18
(Chapter IIa). Involvement of secondary metabolites and associated pathways in
Arabidopsis-P. capsici interactions was further studied by monitoring expression
patterns of corresponding marker genes (Chapter IIb).
In Chapter III, we present an inventory of the function of LecRKs in
Arabidopsis. All the publicly available LecRK T-DNA insertion mutants were
assembled and challenged with various pathogens and abiotic stresses. The
obtained phenotypic data were combined with in silico LecRK expression data
derived from public databases to investigate correlations between LecRK expression
and biological function. For one particular LecRK that functions in plant defence, we
also tested overexpression lines to further confirm its function.
Chapter IV focuses on two closely-related Arabidopsis LecRKs, namely
LecRK-IX.1 and LecRK-IX.2. We performed an in-depth functional analysis on their
role in Phytophthora resistance. We monitored basal defence responses in
Arabidopsis LecRK mutants and analysed the contribution of the lectin domain and
kinase activity through domain deletion and point mutation. In addition, we transiently
expressed both LecRKs in the Solanaceous plant Nicotiana benthamiana to study
LecRK-mediated defence responses and downstream signaling components. Finally,
a potential LecRK-interacting protein was identified using in planta co-
immunoprecipitation and mass spectrometry, and its role in Phytophthora resistance
was further characterized.
Chapter V describes the generation of transgenic N. benthamiana plants that
constitutively express Arabidopsis LecRK-I.9 and LecRK-IX.1. Transgenic lines were
subjected to molecular characterization and infection assays with P. capsici and
P. infestans.
Chapter VI focuses on Solanaceous LecRKs. By in silico screening of protein
sequences, genome sequences, ESTs and RNA-seq data, we identified LecRKs in
N. benthamiana and Solanum lycopersicum. These obtained LecRKs were subjected
to phylogenetic analysis together with Arabidopsis LecRKs to explore evolutionary
relationships. The role of Solanaceous LecRKs in Phytophthora resistance was
analysed by TRV-mediated gene silencing and infection assays with Phytophthora
pathogens.
In Chapter VII, the results presented in this thesis are discussed in a broader
perspective to formulate the role of LecRKs in plant immunity. The potentials and
obstacles in exploiting LecRKs for disease resistance in crops are also discussed.

General introduction
19
REFERENCES
Attard, A., Gourgues, M., Callemeyn-Torre, N. and Keller, H. (2010). The immediate activation of
defense responses in Arabidopsis roots is not sufficient to prevent Phytophthora parasitica
infection. New Phytologist, 187, 449-460.
Baldauf, S.L. (2003). The deep roots of eukaryotes. Science, 300, 1703-1706.
Barre, A., Hervé, C., Lescure, B. and Rougé, P. (2002). Lectin receptor kinases in plants. Critical
Reviews in Plant Sciences, 21, 379-399.
Boggs, N.A., Nasrallah, J.B. and Nasrallah, M.E. (2009). Independent S-locus mutations caused self-
fertility in Arabidopsis thaliana. PLoS Genetics, 5, e1000426.
Boller, T. and Felix, G. (2009). A renaissance of elicitors: Perception of microbe-associated molecular
patterns and danger signals by pattern-recognition receptors. Annual Review of Plant Biology, 60,
379-406.
Boller, T. and He, S.Y. (2009). Innate immunity in plants: an arms race between pattern recognition
receptors in plants and effectors in microbial pathogens. Science, 324, 742-744.
Bos, J.I.B., Kanneganti, T.D., Young, C., Cakir, C., Huitema, E., Win, J., Armstrong, M.R., Birch, P.R.J.
and Kamoun, S. (2006). The C-terminal half of Phytophthora infestans RXLR effector AVR3a is
sufficient to trigger R3a-mediated hypersensitivity and suppress INF1-induced cell death in
Nicotiana benthamiana. The Plant Journal, 48, 165-176.
Bouwmeester, K. and Govers, F. (2009). Arabidopsis L-type lectin receptor kinases: phylogeny,
classification, and expression profiles. Journal of Experimental Botany, 60, 4383-4396.
Bouwmeester, K., van Poppel, P.M.J.A. and Govers, F. (2009). Genome biology cracks enigmas of
oomycete plant pathogens. In: Molecular Aspects of Plant Disease Resistance, Wiley-Blackwell,
102-133.
Bouwmeester, K., de Sain, M., Weide, R., Gouget, A., Klamer, S., Canut, H. and Govers, F. (2011). The
lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a potential
host target for a RXLR effector. PLoS Pathogens, 7, e1001327.
Bouwmeester, K., Han, M., Blanco-Portales, R., Song, W., Weide, R., Guo, L., van der Vossen, E.A.G.
and Govers, F. (2014). The Arabidopsis lectin receptor kinase LecRK-I.9 enhances resistance to
Phytophthora infestans in Solanaceous plants. Plant Biotechnology Journal, 12, 10-16.
Champouret, N., Bouwmeester, K., Rietman, H., van der Lee, T., Maliepaard, C., Heupink, A., van de
Vondervoort, P.J.I., Jacobsen, E., Visser, R.G.F., van der Vossen, E.A.G., et al. (2009).
Phytophthora infestans isolates lacking class I ipiO variants are virulent on Rpi-blb1 potato.
Molecular Plant-Microbe Interactions, 22, 1535-1545.
Chaparro-Garcia, A., Wilkinson, R.C., Gimenez-Ibanez, S., Findlay, K., Coffey, M.D., Zipfel, C.,
Rathjen, J.P., Kamoun, S. and Schornack, S. (2011). The receptor-like kinase SERK3/BAK1 is
required for basal resistance against the late blight pathogen Phytophthora infestans in Nicotiana
benthamiana. PLoS One, 6, e16608.
Cheng, X., Wu, Y., Guo, J., Du, B., Chen, R., Zhu, L. and He, G. (2013). A rice lectin receptor-like
kinase that is involved in innate immune responses also contributes to seed germination. The Plant
Journal, 76, 687-698.
Chinchilla, D., Shan, L., He, P., de Vries, S. and Kemmerling, B. (2009). One for all: the receptor-
associated kinase BAK1. Trends in Plant Science, 14, 535-541.
Chisholm, S.T., Coaker, G., Day, B. and Staskawicz, B.J. (2006). Host-microbe interactions: Shaping
the evolution of the plant immune response. Cell, 124, 803-814.
Choi, J., Tanaka, K., Cao, Y., Qi, Y., Qiu, J., Liang, Y., Lee, S.Y. and Stacey, G. (2014). Identification of

Chapter I
20
a plant receptor for extracellular ATP. Science, 343, 290-294.
Cooke, D.E.L., Schena, L. and Cacciola, S.O. (2007). Tools to detect, identify and monitor
Phytophthora species in natural ecosystems. Journal of Plant Pathology, 89, 13-28.
de Jonge, R., van Esse, H. P., Kombrink, A., Shinya, T., Desaki, Y., Bours, R., van der Krol, S.,
Shibuya, N., Joosten, M.H.A.J. and Thomma, B.P.H.J. (2010). Conserved fungal LysM effector
Ecp6 prevents chitin-triggered immunity in plants. Science, 329, 953-955.
Dou, D., Kale, S.D., Wang, X., Jiang, R.H.Y., Bruce, N.A., Arredondo, F.D., Zhang, X. and Tyler, B.M.
(2008a). RXLR-mediated entry of Phytophthora sojae effector Avr1b into soybean cells does not
require pathogen-encoded machinery. Plant Cell, 20, 1930-1947.
Dou, D., Kale, S.D., Wang, X., Chen, Y., Wang, Q., Wang, X., Jiang, R.H.Y., Arredondo, F.D.,
Anderson, R.G., Thakur, P.B., McDowell, J.M., Wang, Y. and Tyler, B.M. (2008b). Conserved C-
terminal motifs required for avirulence and suppression of cell death by Phytophthora sojae effector
Avr1b. Plant Cell, 20, 1118-1133.
Dou, D. and Zhou, J.M. (2012). Phytopathogen effectors subverting host immunity: different foes,
similar battleground. Cell Host & Microbe, 12, 484-495.
Dubery, I., Sanabria, N. and Huang, J.C. (2012). Nonself perception in plant innate immunity. Advances
in Experimental Medicine and Biology, 738, 79-107.
Du J. 2014. Elicitin-triggered apoplastic immunity against late blight in potato. PhD thesis. Wageningen
University, 49-79.
Erwin D.C. and Ribeiro O.K. (1996). Phytophthora diseases worldwide. The American
Phytopathological Society, St. Paul, Minnesota USA.
Fisher, M.C., Henk, D.A., Briggs, C.J., Brownstein, J.S., Madoff, L.C., McCraw, S.L. and Gurr, S.J.
(2012). Emerging fungal threats to animal, plant and ecosystem health. Nature, 484, 186-194.
Fry, W. (2008). Phytophthora infestans: the plant (and R gene) destroyer. Molecular Plant Pathology, 9,
385-402.
Gilardoni, P.A., Hettenhausen, C., Baldwin, I.T. and Bonaventure, G. (2011). Nicotiana attenuata
LECTIN RECEPTOR KINASE1 suppresses the insect-mediated inhibition of induced defense
responses during Manduca sexta herbivory. Plant Cell, 23, 3512-3532.
Giraldo, M.C. and Valent, B. (2013). Filamentous plant pathogen effectors in action. Nature Reviews
Microbiology, 11, 800-814.
Gouget, A., Senchou, V., Govers, F., Sanson, A., Barre, A., Rouge, P., Pont-Lezica, R.P. and Canut, H.
(2006). Lectin receptor kinases participate in protein-protein interactions to mediate plasma
membrane-cell wall adhesions in Arabidopsis. Plant Physiology, 140, 81-90.
Govers, F. and Bouwmeester, K. (2008). Effector trafficking: RXLR-dEER as extra gear for delivery into
plant cells. Plant Cell, 20, 1728-1730.
Grünwald, N.J., Goss, E.M. and Press, C.M. (2008). Phytophthora ramorum: A pathogen with a
remarkably wide host range causing sudden oak death on oaks and ramorum blight on woody
ornamentals. Molecular Plant Pathology, 9, 729-740.
Haas, B.J., Kamoun, S., Zody, M.C., Jiang, R.H.Y., Handsaker, R.E., Cano, L.M., Grabherr, M., Kodira,
C.D., Raffaele, S., Torto-Alalibo, T., et al. (2009). Genome sequence and analysis of the Irish potato
famine pathogen Phytophthora infestans. Nature, 461, 393-398.
Hansen, E.M., Reeser, P.W. and Sutton, W. (2012). Phytophthora beyond agriculture. Annual Review
of Phytopathology, 50, 359-378.
Huckelhoven, R. (2007). Cell wall-associated mechanisms of disease resistance and susceptibility.
Annual Review of Phytopathology, 45, 101-127.
Huitema, E., Vleeshouwers, V.G.A.A., Francis, D.M. and Kamoun, S. (2003). Active defence responses

General introduction
21
associated with non-host resistance of Arabidopsis thaliana to the oomycete pathogen
Phytophthora infestans. Molecular Plant Pathology, 4, 487-500.
Jiang, R.H.Y., Tripathy, S., Govers, F. and Tyler, B.M. (2008). RXLR effector reservoir in two
Phytophthora species is dominated by a single rapidly evolving superfamily with more than 700
members. Proceedings of the National Academy of Sciences of the United States of America, 105,
4874-4879.
Johnson, E.B., Haggard, J.E. and St Clair, D.A. (2012). Fractionation, stability, and isolate-specificity of
QTL for resistance to Phytophthora infestans in cultivated tomato (Solanum lycopersicum). G3-
Genes Genomes Genetics, 2, 1145-1159.
Jones, J.D.G., and Dangl, J.L. (2006). The plant immune system. Nature, 444, 323-329.
Judelson, H.S. and Blanco, F.A. (2005). The spores of Phytophthora: weapons of the plant destroyer.
Nature Reviews Microbiology, 3, 47-58.
Kamoun, S., van West, P., Vleeshouwers, V.G.A.A., de Groot, K.E. and Govers, F. (1998). Resistance
of Nicotiana benthamiana to Phytophthora infestans is mediated by the recognition of the elicitor
protein INF1. Plant Cell, 10, 1413-1425.
Kamoun, S. (2006). A catalogue of the effector secretome of plant pathogenic oomycetes. Annual
Review of Phytopathology, 44, 41-60.
Kaschani, F., Shabab, M., Bozkurt, T., Shindo, T., Schornack, S., Gu, C., Ilyas, M., Win, J., Kamoun, S.
and van der Hoorn, R.A.L. (2010). An effector-targeted protease contributes to defense against
Phytophthora infestans and is under diversifying selection in natural hosts. Plant Physiology, 154,
1794-1804.
Kasuga, T. and Gijzen, M. (2013). Epigenetics and the evolution of virulence. Trends in Microbiology,
21, 575-582.
Koornneef, M. and Meinke, D. (2010). The development of Arabidopsis as a model plant. The Plant
Journal, 61, 909-921.
Krasileva, K.V., Dahlbeck, D. and Staskawicz, B.J. (2010). Activation of an Arabidopsis resistance
protein is specified by the in planta association of its leucine-rich repeat domain with the cognate
oomycete effector. Plant Cell, 22, 2444-2458.
Kroon, L.P.N.M., Brouwer, H., de Cock, A.W.A.M. and Govers, F. (2012). The genus Phytophthora anno
2012. Phytopathology, 102, 348-364.
Lamour, K.H., Mudge, J., Gobena, D., Hurtado-Gonzales, O.P., Schmutz, J., Kuo, A., Miller, N.A., Rice,
B.J., Raffaele, S., Cano, L.M., et al. (2012a). Genome sequencing and mapping reveal loss of
heterozygosity as a mechanism for rapid adaptation in the vegetable pathogen Phytophthora
capsici. Molecular Plant-Microbe Interactions, 25, 1350-1360.
Lamour, K.H., Stam, R., Jupe, J. and Huitema, E. (2012b). The oomycete broad-host-range pathogen
Phytophthora capsici. Molecular Plant Pathology, 13, 329-430
Lapin, D. and Van den Ackerveken, G. (2013). Susceptibility to plant disease: more than a failure of
host immunity. Trends in Plant Science, 18, 546-554.
Larroque, M., Belmas, E., Martinez, T., Vergnes, S., Ladouce, N., Lafitte, C., Gaulin, E. and Dumas, B.
(2013). Pathogen-associated molecular pattern-triggered immunity and resistance to the root
pathogen Phytophthora parasitica in Arabidopsis. Journal of Experimental Botany, 64, 3615-3625.
Lee, S., Mian, M.A.R., Sneller, C.H., Wang, H., Dorrance, A.E. and McHale, L.K. (2014). Joint linkage
QTL analyses for partial resistance to Phytophthora sojae in soybean using six nested inbred
populations with heterogeneous conditions. Theoretical and Applied Genetics, 127, 429-444.
Li, G., Huang, S., Guo, X., Li, Y., Yang, Y., Guo, Z., Kuang, H., Rietman, H., Bergervoet, M.,
Vleeshouwers, V.G.A.A., et al. (2011). Cloning and characterization of R3b; members of the R3

Chapter I
22
superfamily of late blight resistance genes show sequence and functional divergence. Molecular
Plant-Microbe Interactions, 24, 1132-1142.
Liebrand, T.W.H., van den Burg, H.A. and Joosten, M.H.A.J. (2014). Two for all: Receptor-associated
kinases SOBIR1 and BAK1. Trends in Plant Science, 19, 123-132.
Lin, F., Zhao, M., Ping, J., Johnson, A., Zhang, B., Abney, T.S., Hughes, T.J., and Ma, J. (2013).
Molecular mapping of two genes conferring resistance to Phytophthora sojae in a soybean
landrace PI 567139B. Theoretical and Applied Genetics, 126, 2177-2185.
Lipka, V., Dittgen, J., Bednarek, P., Bhat, R., Wiermer, M., Stein, M., Landtag, J., Brandt, W., Rosahl,
S., Scheel, D., et al. (2005). Pre- and postinvasion defenses both contribute to nonhost resistance
in Arabidopsis. Science, 310, 1180-1183.
Liu, T., Ye, W., Ru, Y., Yang, X., Gu, B., Tao, K., Lu, S., Dong, S., Zheng, X., Shan, W., et al. (2011).
Two host cytoplasmic effectors are required for pathogenesis of Phytophthora sojae by
suppression of host defenses. Plant Physiology, 155, 490-501.
Mackey, D. and McFall, A.J. (2006). MAMPs and MIMPs: proposed classifications for inducers of
innate immunity. Molecular Microbiology, 61, 1365-1371.
Mallard, S., Cantet, M., Massire, A., Bachellez, A., Ewert, S. and Lefebvre, V. (2013). A key QTL cluster
is conserved among accessions and exhibits broad-spectrum resistance to Phytophthora capsici: a
valuable locus for pepper breeding. Molecular Breeding, 32, 349-364.
Martin, F.N., Abed, Z.G., Baldi, Y. and Ivors, K. (2012). Identification and detection of Phytophthora:
reviewing our progress, identifying our needs. Plant Disease, 96, 1080-1103.
Martin, F.N., Blair, J.E. and Coffey, M.D. (2014). A combined mitochondrial and nuclear multilocus
phylogeny of the genus Phytophthora. Fungal Genetics and Biology, 66, 19-32.
Naegele, R.P., Ashrafi, H., Hill, T.A., Chin-Wo, S.R., Van Deynze, A.E. and Hausbeck, M.K. (2014).
QTL mapping of fruit rot resistance to the plant pathogen Phytophthora capsici in a recombinant
inbred line Capsicum annuum population. Phytopathology, 104, 479-483.
Naito K., Taguchi F., Suzuki T., Inagaki Y., Toyoda K., Shiraishi T., Ichinose Y. (2008). Amino acid
sequence of bacterial microbe-associated molecular pattern flg22 is required for virulence.
Molecular Plant-Microbe Interactions, 21, 1165-1174.
Nowicki, M., Foolad, M.R., Nowakowska, M. and Kozik, E.U. (2011). Potato and tomato late blight
caused by Phytophthora infestans: An overview of pathology and resistance breeding. Plant
Disease, 96, 4-17.
Nuhse, T.S. (2012). Cell wall integrity signaling and innate immunity in plants. Frontiers in Plant
Science, 3, 280.
Odeny, D.A., Stich, B. and Gebhardt, C. (2010). Physical organization of mixed protease inhibitor gene
clusters, coordinated expression and association with resistance to late blight at the StKI locus on
potato chromosome III. Plant, Cell & Environment, 33, 2149-2161.
Pel, M.J.C. and Pieterse, C.M.J. (2013). Microbial recognition and evasion of host immunity. Journal of
Experimental Botany, 64, 1237-1248.
Qiao, Y., Liu, L., Xiong, Q., Flores, C., Wong, J., Shi, J., Wang, X., Liu, X., Xiang, Q., Jiang, S., et al.
(2013). Oomycete pathogens encode RNA silencing suppressors. Nature Genetics, 45, 330-333.
Robinson, L.H. and Cahill, D.M. (2003). Ecotypic variation in the response of Arabidopsis thaliana to
Phytophthora cinnamomi. Australasian Plant Pathology, 32, 53-64.
Roetschi, A., Si-Ammour, A., Belbahri, L., Mauch, F. and Mauch-Mani, B. (2001). Characterization of an
Arabidopsis-Phytophthora pathosystem: Resistance requires a functional PAD2 gene and is
independent of salicylic acid, ethylene and jasmonic acid signalling. The Plant Journal, 28, 293-
305.

General introduction
23
Schlaeppi, K., Abou-Mansour, E., Buchala, A. and Mauch, F. (2010). Disease resistance of Arabidopsis
to Phytophthora brassicae is established by the sequential action of indole glucosinolates and
camalexin. The Plant Journal 62, 840-851.
Schlaeppi, K. and Mauch, F. (2010). Indolic secondary metabolites protect Arabidopsis from the
oomycete pathogen Phytophthora brassicae. Plant Signaling & Behavior, 5, 1099-1101.
Schuhegger, R., Nafisi, M., Mansourova, M., Petersen, B.L., Olsen, C.E., Svatos, A., Halkier, B.A., and
Glawischnig, E. (2006). CYP71B15 (PAD3) catalyzes the final step in camalexin biosynthesis. Plant
Physiology, 141, 1248-1254.
Schwessinger, B. and Ronald, P.C. (2012). Plant Innate Immunity: Perception of conserved microbial
signatures. Annual Review of Plant Biology, 63, 451-482.
Sherman-Broyles, S., Boggs, N., Farkas, A., Liu, P., Vrebalov, J., Nasrallah, M.E. and Nasrallah, J.B.
(2007). S locus genes and the evolution of self-fertility in Arabidopsis thaliana. Plant Cell, 19, 94-
106.
Shibata, Y., Kawakita, K. and Takemoto, D. (2010). Age-related resistance of Nicotiana benthamiana
against hemibiotrophic pathogen Phytophthora infestans requires both ethylene- and salicylic acid-
mediated signaling pathways. Molecular Plant-Microbe Interactions, 23, 1130-1142.
Shiu, S.H. and Bleecker, A.B. (2001). Plant receptor-like kinase gene family: diversity, function, and
signaling. Science Signaling, 113, re22.
Shiu, S.H. and Bleecker, A.B. (2003). Expansion of the receptor-like kinase/Pelle gene family and
receptor-like proteins in Arabidopsis. Plant Physiology, 132, 530-543.
Song, J., Bradeen, J.M., Naess, S.K., Raasch, J.A., Wielgus, S.M., Haberlach, G.T., Liu, J., Kuang, H.,
Austin-Phillips, S., Buell, C.R., Helgeson, J.P., and Jiang, J. (2003). Gene RB cloned from Solanum
bulbocastanum confers broad spectrum resistance to potato late blight. Proceedings of the
National Academy of Sciences of the United States of America, 100, 9128-9133.
Stam, R., Jupe, J., Howden, A.J.M., Morris, J.A., Boevink, P.C., Hedley, P.E. and Huitema, E. (2013).
Identification and characterisation CRN effectors in Phytophthora capsici shows modularity and
functional diversity. PLoS One, 8, e59517.
Stassen, J.H.M. and Van den Ackerveken, G. (2011). How do oomycete effectors interfere with plant
life? Current Opinion in Plant Biology, 14, 407-414.
Strange, R.N. and Scott, P.R. (2005). Plant disease: A threat to global food security. Annual Review of
Phytopathology, 43, 83-116.
Sumit, R., Sahu, B.B., Xu, M., Sandhu, D. and Bhattacharyya, M.K. (2012). Arabidopsis nonhost
resistance gene PSS1 confers immunity against an oomycete and a fungal pathogen but not a
bacterial pathogen that cause diseases in soybean. BMC Plant Biology, 12, 87.
Sun, J., Li, L., Zhao, J., Huang, J., Yan, Q., Xing, H. and Guo, N. (2014). Genetic analysis and fine
mapping of RpsJS, a novel resistance gene to Phytophthora sojae in soybean Glycine max (L.)
Merr. Theoretical and Applied Genetics, 127, 913-919.
Takken, F. and Rep, M. (2010). The arms race between tomato and Fusarium oxysporum. Molecular
Plant Pathology, 11, 309-314.
Thomma, B.P.H.J., Nurnberger, T. and Joosten, M.H.A.J. (2011). Of PAMPs and effectors: The blurred
PTI-ETI dichotomy. Plant Cell, 23, 4-15.
Tian, M., Win, J., Song, J., van der Hoorn, R.A.L., van der Knaap, E. and Kamoun, S. (2007). A
Phytophthora infestans cystatin-like protein targets a novel tomato papain-like apoplastic protease.
Plant Physiology, 143, 364-377.
Trivedi, P. and Wang, N. (2014). Host immune responses accelerate pathogen evolution. The ISME
Journal, 8, 727-731.

Chapter I
24
Tyler, B.M. (2002). Molecular basis of recognition between Phytophthora pathogens and their hosts.
Annual Review of Phytopathology, 40, 137-167.
Tyler, B.M., Tripathy, S., Zhang, X., Dehal, P., Jiang, R.H.Y., Aerts, A., Arredondo, F.D., Baxter, L.,
Bensasson, D., Beynon, J.L., et al. (2006). Phytophthora genome sequences uncover evolutionary
origins and mechanisms of pathogenesis. Science, 313, 1261-1266.
Tyler, B.M. (2007). Phytophthora sojae: root rot pathogen of soybean and model oomycete. Molecular
Plant Pathology, 8, 1-8.
Underwood, W. (2012). The plant cell wall: a dynamic barrier against pathogen invasion. Frontiers in
Plant Science, 3, 85.
van Damme, M., Andel, A., Huibers, R.P., Panstruga, R., Weisbeek, P.J. and Van den Ackerveken, G.
(2005). Identification of Arabidopsis loci required for susceptibility to the downy mildew pathogen
Hyaloperonospora parasitica. Molecular Plant-Microbe Interactions, 18, 583-592.
van Damme, M., Huibers, R.P., Elberse, J. and Van den Ackerveken, G. (2008). Arabidopsis DMR6
encodes a putative 2OG-Fe(II) oxygenase that is defense-associated but required for susceptibility
to downy mildew. The Plant Journal, 54, 785-793.
van Damme, M., Zeilmaker, T., Elberse, J., Andel, A., de Sain-van der Velden, M. and van den
Ackerveken, G. (2009). Downy mildew resistance in Arabidopsis by mutation of homoserine kinase.
Plant Cell, 21, 2179-2189.
van der Hoorn, R.A.L. and Kamoun, S. (2008). From guard to decoy: A new model for perception of
plant pathogen effectors. Plant Cell, 20, 2009-2017.
van der Vossen, E., Sikkema, A., Hekkert, B.T.L., Gros, J., Stevens, P., Muskens, M., Wouters, D.,
Pereira, A., Stiekema, W. and Allefs, S. (2003). An ancient R gene from the wild potato species
Solanum bulbocastanum confers broad-spectrum resistance to Phytophthora infestans in cultivated
potato and tomato. The Plant Journal, 36, 867-882.
van Poppel, P.M.J.A., Guo, J., de Vondervoort, P., Jung, M.W.M., Birch, P.R.J., Whisson, S.C. and
Govers, F. (2008). The Phytophthora infestans avirulence gene Avr4 encodes an RXLR-dEER
effector. Molecular Plant-Microbe Interactions, 21, 1460-1470.
Vleeshouwers, V.G.A.A., Rietman, H., Krenek, P., Champouret, N., Young, C., Oh, S.K., Wang, M.,
Bouwmeester, K., Vosman, B., Visser, R.G.F., et al. (2008). Effector genomics accelerates
discovery and functional profiling of potato disease resistance and Phytophthora infestans
avirulence genes. PLoS One, 3, e2875.
Vleeshouwers, V.G.A.A., Raffaele, S., Vossen, J.H., Champouret, N., Oliva, R., Segretin, M.E.,
Rietman, H., Cano, L.M., Lokossou, A., Kessel, G., Pel, M.A. and Kamoun, S. (2011).
Understanding and exploiting late blight resistance in the age of effectors. Annual Review of
Phytopathology, 49, 507-531.
Vurro, M., Bonciani, B. and Vannacci, G. (2010). Emerging infectious diseases of crop plants in
developing countries: impact on agriculture and socio-economic consequences. Food Security, 2,
113-132.
Wang, Y., Meng, Y., Zhang, M., Tong, X., Wang, Q., Sun, Y., Quan, J., Govers, F. and Shan, W. (2011).
Infection of Arabidopsis thaliana by Phytophthora parasitica and identification of variation in host
specificity. Molecular Plant Pathology, 12, 187-201.
Wang, H., Wijeratne, A., Wijeratne, S., Lee, S., Taylor, C., St Martin, S., McHale, L. and Dorrance, A.
(2012). Dissection of two soybean QTL conferring partial resistance to Phytophthora sojae through
sequence and gene expression analysis. BMC Genomics, 13, 428.
Yoshioka, H., Numata, N., Nakajima, K., Katou, S., Kawakita, K., Rowland, O., Jones, J.D.G. and
Doke, N. (2003). Nicotiana benthamiana gp91(phox) homologs NbrbohA and NbrbohB participate

General introduction
25
in H2O2 accumulation and resistance to Phytophthora infestans. Plant Cell, 15, 706-718.
Zhang, J. and Zhou, J.M. (2010). Plant immunity triggered by microbial molecular signatures.
Molecular Plant, 3, 783-793.
Zhang, C., Liu, L., Zheng, Z., Sun, Y., Zhou, L., Yang, Y., Cheng, F., Zhang, Z., Wang, X., Huang, S., et
al. (2013). Fine mapping of the Ph-3 gene conferring resistance to late blight (Phytophthora
infestans) in tomato. Theoretical and Applied Genetics, 126, 2643-2653.
Zheng, X., McLellan, H., Fraiture, M., Liu, X., Boevink, P.C., Gilroy, E.M., Chen, Y., Kandel, K., Sessa,
G., Birch, P.R.J. and Brunner, F. (2014). Functionally redundant RXLR effectors from Phytophthora
infestans act at different steps to suppress early flg22-triggered immunity. PLoS Pathogens, 10,
e1004057.
Zhu, S., Li, Y., Vossen, J.H., Visser, R.G.F. and Jacobsen, E. (2012). Functional stacking of three
resistance genes against Phytophthora infestans in potato. Transgenic Research, 21, 89-99.
Zipfel, C. (2009). Early molecular events in PAMP-triggered immunity. Current Opinion in Plant Biology,
12, 414-420.
Zipfel, C. (2014). Plant pattern-recognition receptors. Trends in Immunology, 35, 345-351.


Chapter II
Characterization of the
Phytophthora capsici-Arabidopsis pathosystem

IIa
A novel Arabidopsis-oomycete pathosystem;
differential interactions with Phytophthora capsici
reveal a role for camalexin, indole glucosinolates
and salicylic acid in defence
Yan Wang, Klaas Bouwmeester, Judith E. van de Mortel, Weixing Shan♯, Francine
Govers
This chapter has been published in: Plant, Cell & Environment (2013); 36, 1192-1203.
In collaboration with: ♯ College of Plant Protection, Northwest A&F University, Yangling, Shaanxi, China

Arabidopsis-Phytophthora capsici interaction
29
ABSTRACT
Phytophthora capsici causes devastating diseases on a broad range of plant species.
To better understand the interaction with its host plants, knowledge obtained from a
model pathosystem can be instrumental. Here we describe the interaction between
P. capsici and Arabidopsis and the exploitation of this novel pathosystem to assign
metabolic pathways involved in defence against P. capsici. Inoculation assays on
Arabidopsis accessions with different P. capsici isolates revealed interaction
specificity among accession-isolate combinations. In a compatible interaction,
appressorium-mediated penetration was followed by the formation of invasive
hyphae, haustoria and sporangia in leaves and roots. In contrast, in an incompatible
interaction, P. capsici infection elicited callose deposition, accumulation of active
oxygen species and cell death, resulting in early pathogen encasement in leaves.
Moreover, Arabidopsis mutants with defects in salicylic acid signaling, camalexin or
indole glucosinolates biosynthesis pathways displayed severely compromised
resistance to P. capsici. It is anticipated that this model pathosystem will facilitate the
genetic dissection of complex traits responsible for resistance against P. capsici.

Chapter IIa
30
INTRODUCTION
In nature, plants are being endangered by a broad range of microbial pathogens.
Development of durable strategies to control plant diseases is therefore a major
challenge when striving for successful plant protection. To this end, increased
knowledge on the mechanisms underlying plant-pathogen interactions would be
helpful and this can greatly benefit from information obtained from model
pathosystems. Arabidopsis thaliana has been put forward as a model plant to study
plant-pathogen interactions due to its unique attributes such as the enriched genetic
and genomic resources (Schlaich 2011). New insights gained from Arabidopsis
research can be exploited to generate pathogen resistance in crop plants (Lacombe
et al., 2010; Zhang et al., 2010).
Successful defence in plants against pathogen attack relies on the initiation of
a range of inducible responses, including cell wall reinforcement, accumulation of
reactive oxygen species, programmed cell death, transcriptional activation of
defence-related genes and synthesis of biologically active secondary metabolites.
The plant hormones salicylic acid (SA), jasmonic acid (JA) and ethylene (ET) play
important roles in resistance to various pathogens as induction of defence responses
in many plant species is coordinated by SA-, JA- and ET-dependent signaling
pathways (Robert-Seilaniantz et a., 2011). In the case of Arabidopsis and other
Brassicaceae species, accumulation of indolic secondary metabolites, such as
camalexin and indole glucosinolates (iGS), is also of central importance to limit
pathogen infection (Thomma et al., 1999; Ferrari et al., 2003; Schlaeppi et al., 2010).
Defence strategies, however, vary between host-pathogen interactions, as was
shown in a number of studies on Arabidopsis, in which mutants impaired in defence-
related pathways and collections of accessions were tested for differential responses
to various pathogens (Thomma et al., 2001; Kover and Schaal 2002; Bohman et al.,
2004; Glazebrook 2005; Spoel et al., 2007). For example, disease resistance in
Arabidopsis to the fungal pathogen Alternaria brassicicola requires camalexin and
JA-dependent pathways (Van Wees et al., 2003), while the bacterial pathogen
Pseudomonas syringae and the oomycete pathogen Hyaloperonospora arabidopsidis
can be inhibited by SA-dependent defence responses (Liu et al., 2010). Moreover,
closely related pathogens not necessarily activate the same defence signaling
pathways. This is, for example, illustrated in studies on interactions of Arabidopsis
with Phytophthora. In the three Phytophthora-Arabidopsis pathosytems that have
been described so far, the role of canonical defence signaling pathways has been
investigated and there seems to be no overlap (Roetschi et al., 2001; Rookes et al.,
2008; Attard et al., 2010; Schlaeppi et al., 2010). In this study, we describe a novel

Arabidopsis-Phytophthora capsici interaction
31
Phytophthora-Arabidopsis pathosystem in which yet another distinct defence network
is activated in order to prevent pathogen invasion.
The novel pathosystem presented here is the interaction between Arabidopsis
and Phytophthora capsici, a renowned plant pathogen that causes severe diseases
on a broad range of economically important crops such as tomato, eggplant,
cucurbits and pepper and is responsible for $1 billion loss in worldwide vegetable
production every year (Lamour et al., 2012a). The P. capsici-Arabidopsis system has
attractive features that make it an ideal model system for dissecting broad-host-range
oomycete-plant interactions, as it not only benefits from the genetic and genomic
resources and tools available for the host, but also those available for the pathogen.
Compared to the other Phytophthora species that interact with Arabidopsis,
P. capsici has several advantages that will greatly facilitate the functional analyses of
candidate effectors and pathogenicity factors. It is one of the few (heterothallic)
species for which genetic crosses and backcrosses can be generated fairly easy,
molecular markers (including numerous single nucleotide variations) and high density
genetic linkage maps are available, it is amenable to DNA transformation, and its
genome has been sequenced (Huitema et al., 2011; Lamour et al., 2012a; Lamour et
al., 2012b).
The aims of this study were to analyse whether or not Arabidopsis can
function as a host for P. capsici and to gain more insight into the defence pathways
that are activated in P. capsici-Arabidopsis interactions. First, we inoculated different
Arabidopsis accessions with a set of P. capsici isolates in order to analyse natural
variation in resistance towards P. capsici. Subsequently, we analysed the cellular
defence responses in Arabidopsis by microscopy during compatible and incompatible
interactions. Finally, we tested mutants with specific defects in defence for enhanced
susceptibility to P. capsici and found that in Arabidopsis SA signaling, as well as
camalexin and iGS are required for resistance to P. capsici.
RESULTS AND DISCUSSION
Arabidopsis is a host for Phytophthora capsici
To determine whether P. capsici is capable to infect Arabidopsis, eight P. capsici
isolates (Table 1) were tested using mycelial plugs or zoospores on mature Col-0
leaves. In most cases, infection was observed, but disease development varied from
complete wilting to no visible symptoms three days post inoculation (dpi) (Fig. 1A).
Based on these observations, a disease severity index (DSI) was adopted to
evaluate disease development at 3 dpi, in which the lowest score (‘0’) represents no
infection and the highest score (‘4’) represents collapse of the inoculated leaf (Fig.

Chapter IIa
32
1B). As shown in Fig. 1C, plug inoculation revealed that five of eight isolates (i.e.
LT263, LT3241, LT3112, LT51 and LT3239) were capable to produce rapid
spreading lesions on Col-0, and more than 90% of the inoculated leaves showed a
DSI ≥ 3 at 3 dpi, resulting in a mean DSI ≥ 3.6. Isolate LT3145 was less virulent on
Col-0 as the proportion of infected leaves with a DSI ≥ 3 was approximately 60%,
with a mean DSI of 3.0. In contrast, isolates LT123 and LT62 were not virulent on
Col-0 as they produced no visible disease symptoms (mean DSI = 0.2 and 0,
respectively). Quantification of the P. capsici biomass in Col-0 leaves at 3 dpi by
quantitative(q) PCR revealed similar levels in leaves inoculated with LT263, LT3241,
LT3112, LT51 and LT3239 whereas P. capsici was hardly detectable in leaves
inoculated with isolates LT123 and LT62 (Supplemental Fig. S1). These results show
that disease severity is highly correlated with pathogen development. Upon zoospore
inoculation, disease symptoms on Col-0 developed at a slower rate when compared
with the plug inoculation, but the outcomes were similar. At 7 dpi, the mean DSI of
LT123 and LT3145 infected leaves was 0 and 3, respectively, whereas the DSI of
leaves infected by the remaining isolates, with the exception of LT62, was ≥ 3. LT62
was not included in this assay because this isolate could not produce enough
zoospores for inoculation.
Table 1. Phytophthora capsici isolates used in this study.
Origin
Isolate Year Country/state Collected from MT
LT51 1997 USA, Michigan cucumber A1a
LT62 1998 USA, Michigan squash A2b
LT123 1998 USA, Michigan cucumber A1a
LT263 2004 USA, Tennessee pumpkin A2a
LT3112 2006 USA, Tennessee pumpkin A1 a
LT3145 2006 USA, Tennessee pumpkin A2b
LT3239 2006 USA, Tennessee pumpkin A2b
LT3241 2006 USA, Tennessee pumpkin A2a a As previously reported (Donahoo and Lamour, 2008). b Determined as described by Harutyunyan et al., (2008).
Since P. capsici is known as a soil-borne pathogen, inoculation assays with
various isolates were also performed on Arabidopsis roots by dipping roots of 16-
day-old seedlings into zoospore suspensions. Consistent with the leaf inoculation
assays, disease phenotypes to individual isolates varied from no symptoms to 100%
mortality of the inoculated seedlings. Inoculation of Col-0 roots with the six isolates
that were found to infect Col-0 leaves, resulted in wilting of the lower leaves within 3

Arabidopsis-Phytophthora capsici interaction
33
dpi, and a complete collapse of the seedlings at 4 dpi (Fig. 1D). In contrast, no
apparent disease symptoms were observed on seedlings inoculated with LT123 (Fig.
1D).
Figure 1. P. capsici isolates vary in virulence on Arabidopsis Col-0.
A, Lesions on plug-inoculated Arabidopsis Col-0 leaves three days post inoculation (dpi) with three
isolates of P. capsici. White arrowheads point to the inoculated leaf. B, Disease severity index (DSI)
presented by numbers from ‘0’ to ‘4’ and increasing levels of gray shading. ‘0’ no infection; ‘1’ lesion
diameter less than 0.5 cm; ‘2’ lesion covering less than 50% of the leaf; ‘3’ lesion covering between 50%
and 75% of the leaf; ‘4’ collapse of the inoculated leaf. C, Disease severity on leaves of Arabidopsis
Col-0 inoculated with eight P. capsici isolates at 3 dpi. Each circle represents the DSI (shown from
light to dark gray as in b) caused by the indicated P. capsici isolate on 40 leaves collected in three
independent experiments. D, Disease symptoms on Col-0 seedlings at 4 dpi. Roots of 16-day-old
seedlings were dipped into water (mock) or P. capsici zoospore suspensions (105 zoospores ml-1)
(LT263; LT123).
To know whether the behavior of P. capsici isolates on Arabidopsis is
comparable to that on host plants of P. capsici, all eight isolates were tested for their
ability to infect tomato. Notably, isolates LT123 and LT62 that were not capable of
infecting Arabidopsis Col-0 could infect tomato leaves (Supplemental Fig. S2). Both

Chapter IIa
34
isolates caused lesions that were visible at 3 dpi but, as on Arabidopsis Col-0, the
lesions caused by the other six isolates were much larger.
In summary, pathogenicity tests on Arabidopsis and tomato revealed striking
differences among P. capsici isolates. Since five out of eight isolates (i.e. LT263,
LT3241, LT3112, LT51 and LT3239) behaved similarly on Arabidopsis and tomato,
isolate LT263 was selected for further analysis, together with isolate LT123 which
caused no disease symptoms on Arabidopsis Col-0.
Natural variation of Arabidopsis accessions in response to P. capsici
Different responses of Arabidopsis Col-0 to P. capsici isolates point to interaction
specificity between Arabidopsis and P. capsici. To get a further understanding of the
interaction specificity, another 34 Arabidopsis accessions originating from different
geographic locations were analysed for their response to P. capsici isolates LT263
and LT123 (Fig. 2).
Figure 2. Disease severity on Arabidopsis accessions inoculated with P. capsici.
Disease severity was assessed at 3 dpi according to the DSI shown in Fig.1B. Each circle represents
the DSI (from light to dark gray as shown at the bottom on the right) caused by the indicated P. capsici
isolate on at least 30 leaves collected from three independent experiments. ***: accessions that show
significant susceptibility to isolate LT123 compared with Col-0 based on a t test (p < 0.001). **:
accessions that show significant resistance to isolate LT263 compared with Col-0 (p < 0.05).

Arabidopsis-Phytophthora capsici interaction
35
All accessions could be infected by isolate LT263, but the disease severity on
the accessions varied significantly (Fig. 2). Of the 35 accessions, 29 including Col-0
were classified as susceptible. On these accessions, lesions extended quickly across
the leaves, resulting in a mean DSI ≥ 2.5 at 3 dpi. The remaining six accessions (i.e.
An-1, Can-0, Est-0, Mt-0, Rsch-4 and Ty-0) were moderately tolerant to LT263; there
was restricted lesion development, with around 70% of the inoculated leaves
showing a DSI ≤ 2. Furthermore, the variations in disease severity were consistent
with the level of colonization by P. capsici and the amount of sporangia present on
the lesions (data not shown).
After inoculation with LT123, only two of the tested accessions, Wei-0 and Ler-
0 developed obvious disease symptoms at 3 dpi (Fig. 2). Lesions spreading from the
inoculation spots as well as formation of sporangia confirmed the susceptibility of
Wei-0 and Ler-0. Nevertheless, infection of Wei-0 and Ler-0 leaves by LT123
resulted in lesions with a mean DSI of 2.2 and 2.1 at 3 dpi, respectively. This was not
comparable to the lesions caused by isolate LT263, with a mean DSI of 4.0 and 3.9,
respectively.
Compatible interaction between Arabidopsis and P. capsici
To further confirm the compatible interaction between Arabidopsis and P. capsici, we
analysed the infection process of Arabidopsis Col-0 with isolate LT263 by microscopy.
Col-0 plants were inoculated by applying LT263 zoospores on the upper side of
leaves. Initially, zoospores attached to the leaf surface and readily germinated, and
approximately 96% of cysts developed appressoria at the tips of germ tubes within 3
h (Fig. 3A). Three hours post inoculation (hpi), penetration pegs emerged beneath
the appressoria. Unlike Phytophthora brassicae (Roetschi et al., 2001) and
Phytophthora parasitica (Attard et al., 2010; Wang et al., 2011), P. capsici did not
only enter the leaf tissue at the junction between epidermal cell walls (Fig. 3B), but
was occasionally observed penetrating epidermal cells directly (Fig. 3C), or via
stomatal cavities (Fig. 3D). Upon penetration, haustoria differentiated from the
penetration hyphae adjacent to the epidermal cell. Subsequently, the infection
hyphae progressed to the adjacent epidermal cells or mesophyll cell layers and more
haustoria were observed. From 1 dpi onwards, intercellular hyphae with haustoria
increased markedly and massive ramifying mycelia were evident in the leaf at 2 dpi
(Fig. 3E). After that, hyphae started to emerge from the leaf through stomatal
openings or via the intercellular space between epidermal cells (Fig. 3, F and G) and
eventually developed sporangia on the leaf surface (Fig. 3H).
To evaluate cellular responses of Arabidopsis upon infection, aniline blue
staining was used to detect the callose deposition, while DAB and NBT were used as
marker dyes for H2O2 and O2- production, respectively. The first day after inoculation,

Chapter IIa
36
neither microscopic detectable callose deposition nor NBT-stained cells were
observed. Only some faint brown-colored cells were found at a few infection sites
after DAB staining (data not shown). Furthermore, infected cells did not show
recognizable trypan blue staining within 2 dpi. This indicates that Arabidopsis cells
remain viable after penetration which is likely a prerequisite for the initial biotrophic
relationship between P. capsici and Arabidopsis.
Figure 3. Cytology of a compatible P. capsici–Arabidopsis interaction.
Arabidopsis Col-0 leaves and roots inoculated with P. capsici LT263 zoospores (105 zoospores ml-1)
were harvested and stained with trypan blue. A, Zoospores attached to the leaf surface, germinated
and formed appressoria within 3 hpi. B-D, Appressorium-mediated penetration of the leaf through the
anticlinal cell wall junction (B), directly through the epidermis (► in C) or via stomata (D). E, Massive
invasive hyphae with haustoria in leaf tissue at 2 dpi. F-G, Infection hyphae emerging from leaves via
stomata (F) or the space between epidermal anticlinal cell walls (G). H, Sporangia development on the
leaf surface. I, Invasive hyphae with haustoria in root cells at 1 dpi. J, Sporangia on the root surface at
2 dpi. ap: appressorium; aw: anticlinal cell wall junction; cy: cyst; gt: germ tube; ha: haustorium; sp:
sporangium; st: stomata. Bars represent 20 µm.

Arabidopsis-Phytophthora capsici interaction
37
P. capsici development in Col-0 roots followed a pattern similar to that
observed in leaves. After penetration into the roots, invading hyphae progressed
rapidly along cortex cells with hardly any branches but formed haustoria within 6 hpi.
Hereafter, infection hyphae started to branch, developed abundant haustoria and
perturbed into the vascular system (Fig. 3I). Sporangia were observed at the root
surface within 2 dpi (Fig. 3J). No recognizable host responses were observed during
the course of infection, i.e. no callose deposition or DAB- and NBT-stained cells (data
not shown).
Incompatible interaction between Arabidopsis and P. capsici
For microscopic analysis of an incompatible interaction between Arabidopsis and
P. capsici, Col-0 leaves were drop-inoculated with LT123 zoospore suspensions.
Approximately 90% of the LT123 zoospores germinated and formed appressoria on
the leaf surface within 6 hpi, but were unable to successfully penetrate plant cells. In
most cases, failure of the first penetration led to the formation of secondary germ
tubes and appressoria (Fig. 4A). However, this did not lead to successful infection as
cell death response, visualized by trypan blue-stained zones, was observed at each
penetration site (Fig. 4B). In addition, infection induced the accumulation of H2O2 and
O2-, which was revealed by the DAB- and NBT-stained cells, respectively, at every
attempted infection site (Fig. 4, C and D). Local accumulation of reactive oxygen
species (ROS), i.e. H2O2 and O2-, is often linked to cell wall-based defence responses,
such as the production of callose, a phenomenon considered to be an active
resistance response to invading pathogens (Hückelhoven, 2007). Consistent with this,
localized callose deposition as shown in Fig. 4E, was detected at nearly every
attempted penetration site during the early phase of infection. In a few instances,
invasive hyphae were detected in the mesophyll cell layer at the inoculation sites in
samples collected after 1 dpi; however, infection hyphae were accompanied by
extensive cell death which was observed not only in mesophyll cells in direct contact
with the infection hyphae but also in the neighboring mesophyll cells (Fig. 4G). No
haustoria or sporangia were detected in inoculated Col-0 leaves. Apparently, during
an incompatible interaction, ROS burst, callose deposition and cell death,
phenomena not commonly observed in the compatible interaction, contributed to
restricting the infection.
In order to examine whether the resistance phenotype observed on Arabidopsis
Col-0 leaves upon dip-inoculation of roots with LT123 zoospores (Fig. 1D, right panel)
is due to failed infection of root tissue, we cytologically examined these inoculated
roots. LT123 zoospores encysted and germinated, and infection of the root tissue
resulted in proliferation of infection hyphae and sporulation (Fig. 4, I and J). This
disease development in Col-0 roots was similar to that observed in roots in the

Chapter IIa
38
Figure 4. Cytology of an incompatible P. capsici–Arabidopsis interaction.
Arabidopsis Col-0 leaves and roots were inoculated with P. capsici LT123 zoospores. A, Abortive
penetration by LT123 led to the formation of secondary germ tubes and appressoria on the leaf
surface as was observed at 6 hpi. B, Cell death of epidermal cells indicated by the dark blue stained
Arabidopsis cells. C-D, DAB-reacting H2O2 accumulation (C) and NBT staining of O2- (D) in
Arabidopsis epidermal cells at 12 hpi. E, Cell wall deposition at the penetration site along the junction
between epidermal cell walls, revealed by aniline blue staining under UV light; F, Bright field image of
(E) showing the germinated cyst at the penetration site; G, Invasive hyphae surrounded by extensive
trypan blue-stained plant cells observed at 1 dpi. H-J, Arabidopsis roots were dipped in water (H) or in
a zoospore suspension of LT123 (I-J) and stained with trypan blue. Invasive hyphae of LT123
colonizing Col-0 roots (I) and sporulation on the root surface (J). cy: cyst; hy: invasive hypha; sa:
secondary appressorium; sg: secondary germ tube; sp: sporangium. Bars represent 20 µm.
compatible interaction between Col-0 and LT263. However, root colonization by
LT123 did not lead to systemic invasion as no classical disease symptoms were
observed on the aerial parts of the seedlings (Fig. 1D). This tolerance response was
also observed on Arabidopsis Col-0 when challenged with Phytophthora cinnamomi
(Rookes et al., 2008) and on cucurbits inoculated with a pathogenicity mutant of the
anthracnose fungus Colletotrichum magna (Freeman and Rodriguez, 1993).
Apparently, in these pathosystems, the ability to establish infection and the ability to

Arabidopsis-Phytophthora capsici interaction
39
further colonize host tissues are separate events.
Phenotypic analysis of JA and ET signaling mutants
To further investigate the relevance of various defence signaling pathways and
secondary metabolites in resistance of Arabidopsis to P. capsici, a set of Col-0
mutants defective in SA, JA, and ET signaling pathways, or in camalexin and iGS
biosynthesis was selected (Fig. 5; Supplemental Table S2) and screened for loss of
resistance to P. capsici LT123, one of the isolates that is unable to infect Col-0 (Fig.
1). Disease severity on the selected mutants was determined according to the
disease severity index (DSI) as described above and is shown by circles in Fig. 5.
Quantitative values are included in Supplemental Table S2.
The analysis included seven mutants that are positioned in JA/ET pathways.
Of these seven, four retained Col-0-like resistance towards LT123, i.e., jin1 and jar1
in which JA signaling is blocked (Berger et al.,1996; Devoto and Turner, 2003), and
ein2 and ein3 which are mutated in ET signaling (Chao et al., 1997; Alonso et al.,
1999). In contrast, resistance was attenuated in the JA-insensitive mutants mpk4 and
eds8-1 (Petersen et al., 2000; Brodersen et al., 2006) and in etr1-1 that has a
mutation in the ET receptor gene ETR1 (Bleecker et al., 1988). These three mutants
all exhibited severe disease symptoms at 3 dpi with 90% of the inoculated leaves
having a DSI ≥ 2. The JA pathway mutants jin1 and jar1 are known to be relatively
weak alleles (Lorenzo et al., 2004) which may explain the difference in phenotype
with mpk4 and eds8-1. Because of the discrepancy in the four JA signaling mutants a
role for JA in P. capsici resistance cannot be ruled out. In the case of ET signaling
mutants differences in disease susceptibility have been reported before, for example
by Geraats et al. (2002), who tested susceptibility of these mutants to various
Pythium species, and by Johansson et al. (2006) and Pantelides et al. (2010), who
analysed the response to Verticillium dahliae. EIN2 and EIN3 act downstream of
ETR1 and the ein2-1 mutant has a stronger ethylene-insensitive phenotype than
etr1-1 (Geraats et al., 2002). Nevertheless, upon P. capsici inoculation etr1-1 is much
more susceptible than ein2-1 and hence there is no positive correlation between the
P. capsici susceptibility phenotype and the ethylene insensitive phenotype. Since
ein3-1 shows a slight gain of susceptibility compared to ein2-1 and Col-0, we cannot
rule out that ET has a role in resistance to P. capsici. However, since etr1-1 is not
only affected in the sensitivity to ET but also in hydrogen peroxide signaling and C6-
aldehyde-induced defence responses (Desikan et al., 2005; Kishimoto et al., 2006),
we speculate that ETR1 is involved in an ethylene independent pathway that affects
disease resistance to P. capsici.

Chapter IIa
40
Figure 5. Disease severity on Arabidopsis mutants impaired in defence related pathways after
inoculation with P. capsici.
A schematic model illustrating the network of defence related signaling pathways and secondary
metabolic biosynthesis routes, and the genes mutated in the selected Arabidopsis mutants
(Supplemental Table S2). Note that NahG is not a mutant but a transgenic line producing an enzyme
that breaks down SA. In the network the undotted arrows represent established relationships between
substrates, the enzymes encoded by the genes and the products; whereas dotted arrows represent
relationships that are deduced from genetic studies and mutant analysis. The disease severity on the
inoculated mutants at 3 dpi with P. capsici isolate LT123 is shown by circles. Each circle represents
the DSI (from light to dark gray as shown on the right) on 30 to 40 leaves collected in three
independent experiments. Mutants that, according to a t test, show significant gain of susceptibility
when compared with Col-0 are marked by *** (P < 0.001) or * (P < 0.05). Quantitative values are
included in Supplemental Table S2. SA: salicylic acid; JA: jasmonic acid; ET: ethylene; iGS: indole
glucosinolates; aGS: aliphatic glucosinolates; Trp: tryptophan; IAOx: indole-3-acetaldoxime; IAN:
indole-3-acetonitrile; DHCA: dihydrocamalexic acid; I3M: indole-3-yl-methyl glucosinolates; 4HO-I3M:
4-hydroxy-indole-3-yl-methyl glucosinolate.

Arabidopsis-Phytophthora capsici interaction
41
SA, camalexin or iGS deficiency compromises resistance of Arabidopsis to
P. capsici
In the SA signaling pathway seven mutants were tested as well as the SA-deficient
line NahG (Delaney et al., 1994). NahG, pad4, eds1-2 and ndr1 (Century et al., 1997;
Zhou et al., 1998; Falk et al., 1999) displayed enhanced susceptibility towards
P. capsici LT123 with fast spreading lesions accompanied by sporulation at 3 dpi,
and resulting in mean DSIs significantly higher than 3. In contrast, inoculation of sid2-
2, npr1-1, npr1-3 and eds5 (Cao et al., 1994; Cao et al., 1997; Glazebrook et al.,
1996; Nawrath and Métraux, 1999; Ton et al., 2002) resulted in visible lesions with
mean DSIs smaller than 2. The difference in susceptibility of, on the one hand the
NahG, eds1-2, pad4 and ndr1 and on the other hand sid2-2, npr1-1, npr1-3 and eds5,
could be due to the functional redundancy of genes involved in the SA pathway; it is
known that SA synthesis is not solely dependent on isochorismate synthase (ICS/SID)
activity, but also on phenylalanine ammonium lyase (PAL) (Wildermuth et al., 2001;
Ferrari et al., 2003). Alternatively, the reduced resistance anticipated in the SA
signaling mutants sid2-2 and eds5 might be recovered by the enhanced
accumulation of camalexin observed in these mutants (Nawrath and Métraux, 1999;
Nawrath et al., 2002). Last but not least, mutations in EDS1, PAD4, and NDR1 not
only suppress SA-mediated defence responses; they can also suppress resistance
controlled by a subset of resistance (R) genes (Coppinger et al., 2004; Zhu et al.,
2011). Thus the enhanced susceptibility of these mutants could also point to a
potential role of R genes in the interaction between Col-0 and LT123. Because of the
pleiotropic effects encountered in the SA signaling mutants, the results on the role of
SA were not conclusive. To address the question whether loss of resistance to P.
capsici in the NahG plants is due to deficiency of SA, we tested the effect of
exogenous application of SA on lesion development. On NahG plants sprayed with
2.5 mM SA one day prior to inoculation with P. capsici LT123, lesions were smaller
than on NahG plants sprayed with water (Supplemental Fig. S3), confirming that SA
plays a role in defence against P. capsici.
In the pathways leading to the production of indolic secondary metabolites,
nine mutants were considered including pad4 mentioned above. The phytoalexin-
deficient mutants pad3 (cyp71b15) and pad4 (Glazebrook and Ausubel, 1994;
Glazebrook et al., 1997) displayed a high level of susceptibility to P. capsici LT123
(Fig. 5). Lesions became clearly visible as early as 2 dpi, and these fast advancing
lesions led to a mean DSI of 3.3 at 3 dpi. CYP71B15 (PAD3) catalyzes the last step
of camalexin biosynthesis (Schuhegger et al., 2006); PAD4 is a lipase-like protein
with a role in camalexin biosynthesis as well as SA/R gene-mediated defence
(Glazebrook et al., 1997; Zhou et al., 1998; Zhu et al., 2011). Unlike pad4, the mutant
pad3 has no other deficiencies apart from camalexin synthesis, and therefore the

Chapter IIa
42
observed susceptibility of pad3 plants to LT123 revealed an important role for
camalexin in disease resistance against P. capsici. Compared to pad3, the cyp71a13
mutant showed much smaller lesion sizes. This difference could be due to different
levels of camalexin. In pad3, camalexin production is completely abolished; even
upon pathogen attack or during abiotic stress there is no camalexin accumulation
(Glazebrook and Ausubel, 1994; Schuhegger et al., 2006). In contrast, in cyp71a13,
which has a mutation in the pathway upstream of CYP71B15, camalexin is still
produced albeit at a reduced level (Nafisi et al., 2007). Moreover, the cytochrome
P450 enzyme CYP71A12, which shows 89% identity with CYP71A13 at the amino
acid level, was reported to function in the camalexin biosynthesis pathway (Nafisi et
al., 2007; Millet et al., 2010). Hence, in the cyp71a13 mutant the absence of
CYP71A13 might be compensated by the activity of CYP71A12. Different
phenotypes for cyp71a13 and pad3 were also reported by Van de Mortel et al. (2012)
who tested the response to non-pathogenic, root-colonizing Pseudomonas
fluorescens bacteria.
PAD2 is a γ-glutamylcysteine synthetase that is required for synthesis of
glutathione (Parisy et al., 2007). The pad2 mutant, which is deficient in both iGS and
camalexin showed increased susceptibility to insect herbivory and P. brassicae
(Schlaeppi et al., 2008; Schlaeppi et al., 2010). Consistently, in our study, pad2 was
found to be very susceptible to P. capsici LT123 with a mean DSI of 3.9 at 3 dpi, thus
indicating that disease resistance to P. capsici is also largely dependent on
camalexin and iGS. Furthermore, the double mutant cyp79b2cyp79b3, which fails to
convert tryptophan into indole-3-acetaldoxime (IAOx), the precursor of camalexin and
iGS (Zhao et al., 2002), showed a similar high DSI (i.e. 4) as pad2. Schlaeppi et al.
(2010) reported that the mutation in CYP79B2 and CYP79B3 does not affect the
level of other defence-associated responses, and hence the susceptibility of the
double mutant cyp79b2cyp79b3 to P. capsici could be attributed to the deficiency of
iGS and camalexin.
To further confirm the potential function of iGS in disease resistance, the iGS-
related mutants pen2, myb51 and cyp81f2 were included in the screening. In the iGS
metabolic pathway, MYB51 is a positive regulator of indole-3-yl-methyl glucosinolate
(I3M) biosynthesis (Gigolashvili et al., 2007). I3M can be converted to 4-hydroxy-
indole-3-yl-methyl glucosinolate (4HO-I3M) by the cytochrome P450 monooxygenase
CYP81F2 (Pfalz et al., 2009). Subsequently, PEN2 myrosinase catalyzes the
hydrolysis of both I3M and 4HO-I3M (Bednarek et al., 2009). Consequently, the
compromised resistance in myb51, cyp81f2 and pen2 indicates that iGS, as well as
iGS hydrolysis products, are important for P. capsici resistance in Arabidopsis.
Arabidopsis produces besides iGS, also aliphatic glucosinolates (aGS), which have
been shown to function in defence towards pest insects (Beekweelder et al., 2008).

Arabidopsis-Phytophthora capsici interaction
43
Biosynthesis of aGS is dependent on the transcription factors MYB28 and MYB29,
and the synthesis of aGS is completely abolished in the double mutant myb28myb29
(Beekweelder et al., 2008). Inoculation of myb28myb29 with P. capsici did not result
in any changes in disease susceptibility when compared with Col-0, indicating that
blocking aGS synthesis by mutation in MYB28 and MYB29 is not sufficient to confer
Arabidopsis susceptibility to P. capsici.
Arabidopsis exploits a different basal defence network for each Phytophthora
species
Collectively, the results described here indicate that basal resistance to P. capsici in
Arabidopsis involves the activation of SA signaling and the biosynthesis of camalexin
and iGS. Blocking the accumulation of SA, camalexin or iGS results in susceptibility
to P. capsici. In comparison, resistance to P. parasitica depends on JA/ET signaling
in addition to SA signaling, as the mutants etr1-3, ein2-1 and jar1-1 were found to be
more susceptible to this pathogen (Attard et al., 2010). In contrast, Arabidopsis
resistance to P. brassicae was shown to be independent of SA and JA/ET signaling
pathways; NahG plants, nor etr1, ein2 and jar1 mutants showed a change in
phenotype demonstrating that blocking SA, or JA/ET signaling had no significant
effect on resistance (Roetschi et al., 2001). Moreover, mutants with defects in the
accumulation of either camalexin or iGS, like pad3 or myb51, showed no significant
change in resistance to P. brassicae (Schlaeppi et al., 2010). However, the combined
deficiency of iGS and camalexin as manifested in either pad2 or cyp79b2cyp79b3,
resulted in susceptibility, demonstrating that the resistance to P. brassicae requires
the sequential action of iGS and camalexin (Schlaeppi et al., 2010). In the case of
P. cinnamomi, resistance in Arabidopsis seems to be independent of any of above
mentioned pathways. All mutants including pad2, pad3, pad4, ein2 and jar1 as well
as NahG plants tested by Rookes et al. (2008) retained wild-type resistance. Taken
together, it is evident that every attacking pathogen elicits the activation of specific
signaling networks and that Phytophthora species vary in their ability to tolerate
different defence components.
CONCLUSIONS
In this study, we demonstrated that P. capsici is capable to infect Arabidopsis and
that there is natural variation among Arabidopsis accessions in response to different
P. capsici isolates. Cell death, callose deposition and ROS accumulation were
induced in Arabidopsis during the early infection stages in an incompatible interaction.
Blocking SA signaling, camalexin or iGS biosynthesis pathways in Arabidopsis,

Chapter IIa
44
conferred susceptibility to P. capsici demonstrating that SA signaling and
biosynthesis of camalexin and iGS are essential for basal resistance to this
Phytophthora species. This newly established pathosystem will facilitate the
identification of novel traits responsible for resistance to the broad-host-range
oomycete pathogen P. capsici.
MATERIALS AND METHODS
P. capsici culture conditions and inoculum preparation
P. capsici isolates (Table 1) were routinely cultured in the dark at 25°C on 20% (v/v)
V8 juice agar plates (Erwin and Ribeiro, 1996). Zoospores for plant infection assays
were obtained by incubating mycelial plugs (Ø 1.0 cm) from the margin of a four-day
old growing colony in 10% (v/v) cleared V8 broth in the dark at 25°C for two days.
Hereafter, the V8 medium was replaced with sterilized mineral solution (Wang et al.,
2011) and refreshed once every hour up to four times. After two days of incubation
under continuous light, numerous sporangia developed. Zoospore release was
induced with a cold shock by flooding the plates with cold water (4°C) for 30 min.
Zoospores were filtered through one layer of Miracloth (Merck) and adjusted to the
desired concentrations.
Plant material and growth conditions
Arabidopsis seeds were sown on soil, or in vitro on 0.5 MS medium (Murashige and
Skoog, Duchefa) supplemented with 1% (w/v) sucrose and 1.2% (w/v) plant agar
(Duchefa), and were stratified by placing them in the dark at 4°C for three days.
Subsequently, Arabidopsis was grown in a conditioned growth chamber at 19-21°C,
with a 16 h photoperiod and a relative humidity (RH) of 75-80%. Tomato plants of
cultivar Moneymaker were grown under standard greenhouse conditions.
Plant inoculation procedures and SA treatment
Four-week-old Arabidopsis or tomato leaves were sprayed with water, and
subsequently inoculated on the abaxial leaf side with fresh mycelial plugs (Ø 0.5 cm).
Mock-inoculation was conducted with blank V8 plugs. Mycelial plugs were removed
from Arabidopsis leaves two days post inoculation (dpi) and from tomato leaves 1 dpi.
Zoospore inoculation of Arabidopsis leaves was performed by inoculating 10 μl
droplets containing 105 zoospores ml-1 on each leaf. For root inoculation, roots of 16-
day-old Arabidopsis seedlings were dipped in water or a zoospore suspension (105
zoospores ml-1) for 5 seconds, and then transplanted to soil. During the first 24 h
after inoculation, plants were kept in the dark at 22°C in trays covered with lids to

Arabidopsis-Phytophthora capsici interaction
45
maintain a high humidity. Subsequently, plants were incubated at a 75-80% RH and
a 10 h photoperiod.
For SA treatment, plants were exposed to SA (Merck Schuchardt) 1 d prior to
inoculation with P. capsici by spraying the leaves with a solution containing 2.5 mM
SA, 0.1 % ethanol (v/v) and 0.02% Silwet L-77 (v/v). Control plants were sprayed
with water supplemented with 0.1 % ethanol (v/v) and 0.02% Silwet L-77 (v/v).
Disease severity index
Disease development on Arabidopsis leaves was evaluated using a disease severity
index (DSI) on a scale of 0 to 4. A score of ‘0’ indicates no visible disease symptoms
or small necrotic flecks; ‘1’, a lesion diameter less than 0.5 cm; ‘2’, a lesion covering
less than 50% of the leaf surface; ‘3’, a lesion covering 50-75% of the leaf surface,
and a score of ‘4’ refers to 75-100% collapse of the leaf. The mean DSI was
calculated according to the equation:
MeanDSI∑ . .
Tomato disease severity was monitored by measuring lesion sizes at 3 dpi. All
experiments were repeated at least three times.
Quantification of P. capsici biomass in infected Arabidopsis leaves
Six leaves inoculated with P. capsici collected at 3 dpi were used for genomic DNA
isolation with a DNeasy Plant Mini Kit (Qiagen). qPCR was performed for quantifying
the P. capsici biomass using an ABI 7300 PCR machine (Applied Biosystems). The
Arabidopsis Rubisco primer pair AtRub-F4/AtRub-R4 was used as endogenous
control, while the primer pair CAP-Fw/CAP-Rv1 was used to target P. capsici internal
transcribed spacer regions (Silvar et al., 2005; Yadeta et al., 2011; Supplemental
Table S1). The ratio of P. capsici genomic DNA to Arabidopsis DNA was calculated
using a 2−∆∆CT method. Two biological replicates were analysed, each with a
technical triplicate.
Histological staining and microscopy
Staining with trypan blue (Sigma) visualizing infection hyphae and plant cell death,
and aniline blue (Acros Organics) visualizing callose deposition, was carried out as
described by Bouwmeester et al. (2011). Detection of H2O2 and O2- was performed
as described by Thordal-Christensen et al. (1997) and Dunand et al. (2007).
Microscopy was performed with a Nikon 90i microscope (Nikon, Amstelveen, The
Netherlands) using differential interference contrast and epifluorescence settings (i.e.
DAPI filter, EX 340-380, DM 400, BA 435-4850).

Chapter IIa
46
ACKNOWLEDGEMENTS
We thank Kurt Lamour for providing Phytophthora capsici isolates, and Bart P.H.J.
Thomma and Joost Keurentjes for Arabidopsis accessions and mutant lines. This
project was financially supported by a Wageningen University sandwich PhD
fellowship (YW) and a Huygens scholarship (YW), by the Centre for BioSystems
Genomics (CBSG) (KB, FG) and by EcoLinc (JEvdM). CBSG and EcoLinc are
centres of the Netherlands Genomics Initiative that is affiliated with the Netherlands
Organisation for Scientific Research.
REFERENCES
Alonso J.M., Hirayama T., Roman G., Nourizadeh S. and Ecker J.R. (1999). EIN2, a bifunctional
transducer of ethylene and stress responses in Arabidopsis. Science, 284, 2148-2152.
Attard A., Gourgues M., Callemeyn-Torre N. and Keller H. (2010). The immediate activation of defense
responses in Arabidopsis roots is not sufficient to prevent Phytophthora parasitica infection. New
Phytologist, 187, 449-460.
Bednarek P., Piślewska-Bednarek M., Svatoš A., Schneider, B., Doubský, J., Mansurova, M., Humphry,
M., Consonni, C., Panstruga, R., Sanchez-Vallet, A. et al. (2009). A glucosinolate metabolism
pathway in living plant cells mediates broad-spectrum antifungal defense. Science, 323, 101-106.
Beekweelder J., van Leeuwen W., van Dam N.M., Bertossi, M., Grandi, V., Mizzi, L., Soloviev, M.,
Szabados, L., Molthoff, J. W., Schipper, B. et al. (2008). The impact of the absence of aliphatic
glucosinolates on insect herbivory in Arabidopsis. PLoS One, 3, e2068.
Berger S., Bell E. and Mullet J.E. (1996). Two methyl jasmonate-insensitive mutants show altered
expression of AtVsp in response to methyl jasmonate and wounding. Plant Physiology, 111, 525-
531.
Bleecker A.B., Estelle M.A., Somerville C. and Kende H. (1988). Insensitivity to ethylene conferred by a
dominant mutation in Arabidopsis thaliana. Science, 241, 1086-1089.
Bohman S., Staal J., Thomma B.P.H.J., Wang M. and Dixelius C. (2004). Characterisation of an
Arabidopsis–Leptosphaeria maculans pathosystem: resistance partially requires camalexin
biosynthesis and is independent of salicylic acid, ethylene and jasmonic acid signalling. The Plant
Journal, 37, 9-20.
Bouwmeester K., de Sain M., Weide R., Gouget A., Klamer S., Canut H. and Govers F. (2011). The
lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a potential
host target for a RXLR effector. PLoS Pathogens, 7, e1001327.
Brodersen P., Petersen M., Nielsen H.B., Zhu S., Newman M.A., Shokat K.M., Rietz S., Parker J. and
Mundy J. (2006). Arabidopsis MAP kinase 4 regulates salicylic acid- and jasmonic acid/ethylene-
dependent responses via EDS1 and PAD4. The Plant Journal, 47, 532-546.
Cao H., Bowling S.A., Gordon A.S. and Dong X. (1994). Characterization of an Arabidopsis mutant that
is nonresponsive to inducers of systemic acquired resistance. Plant Cell, 6, 1583-1592.
Cao H., Glazebrook J., Clarke J.D., Volko S. and Dong X. (1997). The Arabidopsis NPR1 gene that
controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell, 88,
57-63.

Arabidopsis-Phytophthora capsici interaction
47
Century K.S., Shapiro A.D., Repetti P.P., Dahlbeck D., Holub E. and Staskawicz B.J. (1997). NDR1, a
pathogen-induced component required for Arabidopsis disease resistance. Science, 278, 1963-
1965.
Chao Q., Rothenberg M., Solano R., Roman G., Terzaghi W. and Ecker J.R. (1997). Activation of the
ethylene gas response pathway in Arabidopsis by the nuclear protein ETHYLENE-INSENSITIVE3
and related proteins. Cell, 89, 1133-1144.
Coppinger P., Repetti P.P., Day B., Dahlbeck D., Mehlert A. and Staskawicz B.J. (2004).
Overexpression of the plasma membrane-localized NDR1 protein results in enhanced bacterial
disease resistance in Arabidopsis thaliana. The Plant Journal, 40, 225-237.
Friedrich, L., Weymann, K., Negrotto, D., Gaffney, T., Gut-Rella, M., Kessmann, H., Ward, E., Ryals, J.
(1994). A central role of salicylic acid in plant disease resistance. Science, 266, 1247-1250.
Desikan R., Hancock J.T., Bright J., Harrison J., Weir L., Hooley R. and Neill S.J. (2005). A role for
ETR1 in hydrogen peroxide signaling in stomatal guard cells. Plant Physiology, 137, 831-834.
Devoto A. and Turner J.G. (2003). Regulation of jasmonate-mediated plant responses in Arabidopsis.
Annals of Botany, 92, 329-337.
Donahoo R.S. and Lamour K.H. (2008). Interspecific hybridization and apomixis between Phytophthora
capsici and Phytophthora tropicalis. Mycologia, 100, 911-920.
Dong X. (2004). NPR1, all things considered. Current Opinion in Plant Biology, 7, 547-552.
Dunand C., Crevecoeur M. and Penel C. (2007). Distribution of superoxide and hydrogen peroxide in
Arabidopsis root and their influence on root development: possible interaction with peroxidases.
New Phytologist, 174, 332-341.
Erwin D.C. and Ribeiro O.K. (1996). Phytophthora diseases worldwide. The American
Phytopathological Society, St. Paul, Minnesota USA.
Falk A., Feys B.J., Frost L.N., Jones J.D.G., Daniels M.J. and Parker J.E. (1999). EDS1, an essential
component of R gene-mediated disease resistance in Arabidopsis has homology to eukaryotic
lipases. Proceedings of the National Academy of Sciences of the United States of America, 96,
3292-3297.
Ferrari S., Plotnikova J.M., De Lorenzo G. and Ausubel F.M. (2003). Arabidopsis local resistance to
Botrytis cinerea involves salicylic acid and camalexin and requires EDS4 and PAD2, but not SID2,
EDS5 or PAD4. The Plant Journal, 35, 193-205.
Freeman S. and Rodriguez R.J. (1993). Genetic conversion of a fungal plant pathogen to a
nonpathogenic, endophytic mutualist. Science, 259, 75-78.
Geraats B.P.J., Bakker P.A.H.M. and Van Loon L.C. (2002). Ethylene insensitivity impairs resistance to
soilborne pathogens in tobacco and Arabidopsis thaliana. Molecular Plant-Microbe Interactions, 15,
1078-1085.
Gigolashvili T., Berger B., Mock H.P., Müller C., Weisshaar B. and Flügge U.I. (2007). The transcription
factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. The Plant
Journal, 50, 886-901.
Glazebrook J. and Ausubel F.M. (1994). Isolation of phytoalexin-deficient mutants of Arabidopsis
thaliana and characterization of their interactions with bacterial pathogens. Proceedings of the
National Academy of Sciences of the United States of America, 91, 8955-8959.
Glazebrook J., Rogers E.E. and Ausubel F.M. (1996). Isolation of Arabidopsis mutants with enhanced
disease susceptibility by direct screening. Genetics, 143, 973-982.
Glazebrook J., Zook M., Mert F., Kagan I., Rogers E.E., Crute I.R., Holub E.B., Hammerschmidt R. and
Ausubel F.M. (1997). Phytoalexin-deficient mutants of Arabidopsis reveal that PAD4 encodes a

Chapter IIa
48
regulatory factor and that four PAD genes contribute to downy mildew resistance. Genetics, 146,
381-392.
Glazebrook J. (2005). Contrasting mechanisms of defense against biotrophic and necrotrophic
pathogens. Annual Review of Phytopathology, 43, 205-227.
Harutyunyan S.R., Zhao Z., Den Hartog T., Bouwmeester K., Minnaard A.J., Feringa B.L. and Govers
F. (2008). Biologically active Phytophthora mating hormone prepared by catalytic asymmetric total
synthesis. Proceedings of the National Academy of Sciences of the United States of America, 105,
8507-8512.
Hückelhoven R. (2007). Cell wall-associated mechanisms of disease resistance and susceptibility.
Annual Review of Phytopathology, 45, 101-127.
Huitema E., Smoker M. and Kamoun S. (2011). A straightforward protocol for electro-transformation of
Phytophthora capsici zoospores. Methods in Molecular Biology, 712, 129-135.
Johansson A., Staal J. and Dixelius C. (2006). Early responses in the Arabidopsis-Verticillium
longisporum pathosystem are dependent on NDR1, JA- and ET-associated signals via cytosolic
NPR1 and RFO1. Molecular Plant-Microbe Interactions, 19, 958-969.
Kishimoto K., Matsui K., Ozawa R. and Takabayashi J. (2006). ETR1-, JAR1- and PAD2-dependent
signaling pathways are involved in C6-aldehyde-induced defense responses of Arabidopsis. Plant
Science, 171, 415-423.
Kover P.X. and Schaal B.A. (2002). Genetic variation for disease resistance and tolerance among
Arabidopsis thaliana accessions. Proceedings of the National Academy of Sciences of the United
States of America, 99, 11270-11274.
Lacombe S., Rougon-Cardoso A., Sherwood E., Peeters N., Dahlbeck D., van Esse H.P., Smoker M.,
Rallapalli G., Thomma B.P.H.J., Staskawicz B.J., et al. (2010). Interfamily transfer of a plant
pattern-recognition receptor confers broad-spectrum bacterial resistance. Nature Biotechnology, 28,
365-394.
Lamour K.H., Stam R., Jupe J. and Huitema E. (2012a). The oomycete broad-host-range pathogen
Phytophthora capsici. Molecular Plant Pathology, 13, 329-430
Lamour K.H., Mudge J., Gobena D., Hurtado-Gonzales O.P., Schmutz J., Kuo A., Miller N.A., Rice B.J.,
Raffaele S., Cano L., et al. (2012b). Genome sequencing and mapping reveal loss of
heterozygosity as a mechanism for rapid adaptation in the vegetable pathogen Phytophthora
capsici. Molecular Plant-Microbe Interactions, 25, 1350-1360.
Liu P., Yang Y., Pichersky E. and Klessig D.F. (2010). Altering expression of benzoic acid/salicylic acid
carboxyl methyltransferase 1 compromises systemic acquired resistance and PAMP-triggered
immunity in Arabidopsis. Molecular Plant-Microbe Interactions, 23, 82-90.
Leon-Reyes A., Spoel S.H., De Lange E.S., Abe H., Kobayashi M., Tsuda S., Millenaar F.F., Welschen
R.A.M., Ritsema T. and Pieterse C.M.J. (2009). Ethylene modulates the role of NONEXPRESSOR
OF PATHOGENESIS-RELATED GENES1 in cross talk between salicylate and jasmonate signaling.
Plant Physiology, 149, 1797-1809.
Lorenzo O., Chico J.M., Sanchez-Serrano J.J. and Solano R. (2004). Jasmonate-insensitive 1 encodes
a MYC transcription factor essential to discriminate between different jasmonate-regulated defense
responses in Arabidopsis. Plant Cell, 16, 1938-1950.
Millet Y.A., Danna C.H., Clay N.K., Songnuan W., Simon M.D., Werck-Reichhart D. and Ausubel F.M.
(2010). Innate immune responses activated in Arabidopsis roots by microbe-associated molecular
patterns. Plant Cell, 22, 973-990.

Arabidopsis-Phytophthora capsici interaction
49
Nafisi M., Goregaoker S., Botanga C.J., Glawischnig E., Olsen C.E., Halkier B.A. and Glazebrook J.
(2007). Arabidopsis cytochrome P450 monooxygenase 71A13 catalyzes the conversion of indole-
3-acetaldoxime in camalexin synthesis. Plant Cell, 19, 2039-2052.
Nawrath C. and Métraux J.P. (1999). Salicylic acid induction-deficient mutants of Arabidopsis express
PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell, 11,
1393-1404.
Nawrath C., Heck S., Parinthawong N. and Metraux J.P. (2002). EDS5, an essential component of
salicylic acid-dependent signaling for disease resistance in Arabidopsis, is a member of the MATE
transporter family. Plant Cell, 14, 275-286.
Pantelides I.S., Tjamos S.E. and Paplomatas E.J. (2010). Ethylene perception via ETR1 is required in
Arabidopsis infection by Verticillium dahliae. Molecular Plant Pathology, 11, 191-202.
Parisy V., Poinssot B., Owsianowski L., Buchala A., Glazebrook J. and Mauch F. (2007). Identification
of PAD2 as a γ-glutamylcysteine synthetase highlights the importance of glutathione in disease
resistance of Arabidopsis. The Plant Journal, 49, 159-172.
Penninckx I.A.M.A., Thomma B.P.H.J., Buchala A., Métraux J.P. and Broekaert W.F. (1998).
Concomitant activation of jasmonate and ethylene response pathways is required for induction of a
plant defensin gene in Arabidopsis. Plant Cell, 10, 2103-2113.
Petersen M., Brodersen P., Naested H., Andreasson E., Lindhart U., Johansen B., Nielsen H.B., Lacy
M., Austin M.J., Parker J.E., Sharma S.B., Klessig D.F., Martienssen R., Mattsson O., Jensen A.B.
and Mundy J. (2000). Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance.
Cell, 103, 1111-1120.
Pfalz M., Vogel H. and Kroymann J. (2009). The gene controlling the Indole Glucosinolate Modifier1
quantitative trait locus alters indole glucosinolate structures and aphid resistance in Arabidopsis.
Plant Cell, 21, 985-999.
Pieterse C.M.J., Van Wees S.C.M., Van Pelt J.A., Knoester M., Laan R., Gerrits H., Weisbeek P.J. and
Van Loon L.C. (1998). A novel signaling pathway controlling induced systemic resistance in
Arabidopsis. Plant Cell, 10, 1571-1580.
Robert-Seilaniantz A., Grant M. and Jones J.D.G. (2011). Hormone crosstalk in plant disease and
defense: more than just jasmonate-salicylate antagonism. Annual Review of Phytopathology, 49,
317-343.
Roetschi A., Si-Ammour A., Belbahri L., Mauch F. and Mauch-Mani B. (2001). Characterization of an
Arabidopsis-Phytophthora pathosystem: Resistance requires a functional PAD2 gene and is
independent of salicylic acid, ethylene and jasmonic acid signalling. The Plant Journal, 28, 293-305.
Rookes J.E., Wright M.L. and Cahill D.M. (2008). Elucidation of defence responses and signalling
pathways induced in Arabidopsis thaliana following challenge with Phytophthora cinnamomi.
Physiological and Molecular Plant Pathology, 72, 151-161.
Schlaeppi K., Bodenhausen N., Buchala A., Mauch F. and Reymond P. (2008). The glutathione-
deficient mutant pad2-1 accumulates lower amounts of glucosinolates and is more susceptible to
the insect herbivore Spodoptera littoralis. The Plant Journal, 55, 774-786.
Schlaeppi K., Abou-Mansour E., Buchala A. and Mauch F. (2010). Disease resistance of Arabidopsis
to Phytophthora brassicae is established by the sequential action of indole glucosinolates and
camalexin. The Plant Journal, 62, 840-851.
Schlaich N.L. (2011). Arabidopsis thaliana-the model plant to study host-pathogen interactions. Current
Drug Targets, 12, 955-966.

Chapter IIa
50
Schuhegger R., Nafisi M., Mansourova M., Petersen B.L., Olsen C.E., Svatoš A., Halkier B.A. and
Glawischnig E. (2006). CYP71B15 (PAD3) catalyzes the final step in camalexin biosynthesis. Plant
Physiology, 141, 1248-1254.
Silvar C., Duncan J.M., Cooke D.E.L., Williams N.A., Diaz J. and Merino F. (2005). Development of
specific PCR primers for identification and detection of Phytophthora capsici Leon. European
Journal of Plant Pathology, 112, 43-52.
Spoel S.H., Johnson J.S. and Dong X. (2007). Regulation of tradeoffs between plant defenses against
pathogens with different lifestyles. Proceedings of the National Academy of Sciences of the United
States of America, 104, 18842-18847.
Thomma B.P.H.J., Eggermont K., Tierens K. and Broekaert W.F. (1999). Requirement of functional
ethylene-insensitive 2 gene for efficient resistance of Arabidopsis to infection by Botrytis cinerea.
Plant Physiology, 121, 1093-1101.
Thomma B.P.H.J., Nelissen I., Eggermont K. and Broekaert W.F. (1999). Deficiency in phytoalexin
production causes enhanced susceptibility of Arabidopsis thaliana to the fungus Alternaria
brassicicola. The Plant Journal, 19, 163-171.
Thomma B.P.H.J., Tierens K.F.M., Penninckx I.A.M.A., Mauch-Mani B., Broekaert W.F. and Cammue
B.P.A. (2001). Different micro-organisms differentially induce Arabidopsis disease response
pathways. Plant Physiology and Biochemistry, 39, 673-680.
Thordal-Christensen H., Zhang Z., Wei Y. and Collinge D.B. (1997). Subcellular localization of H2O2 in
plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery
mildew interaction. The Plant Journal, 11, 1187-1194.
Ton J., De Vos M., Robben C., Buchala A., Metraux J.P., Van Loon L.C. and Pieterse C.M.J. (2002).
Characterization of Arabidopsis enhanced disease susceptibility mutants that are affected in
systemically induced resistance. The Plant Journal, 29, 11-21.
van de Mortel J.E., De Vos R.C.H., Dekkers E., Pineda A., Guillod L., Bouwmeester K., van Loon
J.J.A., Dicke M. and Raaijmakers J.M. (2012). Metabolic and transcriptomic changes induced in
Arabidopsis by the rhizobacterium Pseudomonas fluorescens SS101. Plant Physiology, 160, 2173-
2188.
van Wees S.C.M., Chang H.S., Zhu T. and Glazebrook J. (2003). Characterization of the early
response of Arabidopsis to Alternaria brassicicola infection using expression profiling. Plant
Physiology, 132, 606-617.
Wang Y., Meng Y., Zhang M., Tong X., Wang Q., Sun Y., Quan J., Govers F. and Shan W. (2011).
Infection of Arabidopsis thaliana by Phytophthora parasitica and identification of variation in host
specificity. Molecular Plant Pathology, 12, 187-201.
Wildermuth M.C., Dewdney J., Wu G. and Ausubel F.M. (2001). Isochorismate synthase is required to
synthesize salicylic acid for plant defence. Nature, 414, 562-565.
Yadeta K.A., Hanemian M., Smit P., Hiemstra J.A., Pereira A., Marco Y. and Thomma B.P.H.J. (2011).
The Arabidopsis thaliana DNA-binding protein AHL19 mediates Verticillium wilt resistance.
Molecular Plant-Microbe Interactions, 24, 1582-1591.
Zhang X., Francis M.I., Dawson W.O., Graham J.H., Orbovic V., Triplett E.W. and Mou Z. (2010). Over-
expression of the Arabidopsis NPR1 gene in citrus increases resistance to citrus canker. European
Journal of Plant Pathology, 128, 91-100.
Zhao Y., Hull A.K., Gupta N.R., Goss K.A., Alonso J., Ecker J.R., Normanly J., Chory J. and Celenza
J.L. (2002). Trp-dependent auxin biosynthesis in Arabidopsis: Involvement of cytochrome P450s
CYP79B2 and CYP79B3. Genes and Development, 16, 3100-3112.

Arabidopsis-Phytophthora capsici interaction
51
Zhou N., Tootle T.L., Tsui F., Klessig D.F. and Glazebrook J. (1998). PAD4 functions upstream from
salicylic acid to control defense responses in Arabidopsis. Plant Cell, 10, 1021-1030.
Zhu S., Jeong R.-D., Venugopal S.C., Lapchyk L., Navarre D., Kachroo A. and Kachroo P. (2011).
SAG101 forms a ternary complex with EDS1 and PAD4 and is required for resistance signaling
against turnip crinkle virus. PLoS Pathogens, 7, e1002318.

Cha
pter
IIa
52
SU
PP
LE
ME
NT
AL
INF
OR
MA
TIO
N
Tab
le S
2. A
rab
ido
psi
s d
efen
ce-r
elat
ed m
uta
nts
an
d t
ran
sgen
ic l
ines
use
d i
n t
his
stu
dy
and
th
e d
isea
se s
ever
ity
at t
hre
e d
ays
aft
er i
no
cula
tio
n
wit
h P
. ca
psi
ci i
sola
te L
T12
3. T
he in
form
atio
n on
the
pat
hway
s or
met
abol
ites
that
are
affe
cted
and
the
pat
hoge
ns t
hat
wer
e st
udie
d in
the
inte
ract
ion,
was
col
lect
ed fr
om th
e re
fere
nces
list
ed.
Mu
tan
t D
isea
se s
ever
itya
A
ffec
ted
pat
hw
ays
o
r m
etab
olit
esb
Pat
ho
gen
s R
efer
ence
s
L
T12
3 (↓
: d
ecre
ased
; ↑:
incr
ease
d)
eds1
-2
3.4
± 0.
3***
S
A↓;
R
-gen
e-m
edia
ted
resi
stan
ce↓
Pse
udom
onas
syr
inga
e
Gla
zebr
ook
et a
l., 1
996;
Fal
k et
al.,
199
9;
F
eys
et a
l., 2
001;
Ven
ugop
al e
t al.,
200
9
eds5
1.
4 ±
0.4*
S
A↓;
cam
alex
in↑
P
seud
omon
as s
yrin
gae
G
laze
broo
k et
al.,
199
6; N
awra
th e
t al.,
199
9;
N
awra
th e
t al.,
200
2; T
on e
t al.,
200
2;
Hec
k et
al.,
200
3
ndr1
3.
4 ±
0.2*
**
SA↓;
R
-gen
e-m
edia
ted
resi
stan
ce↓
Pse
udom
onas
syr
inga
e
Cen
tury
et a
l., 1
997;
Sha
piro
and
Zha
ng 2
001
Nah
G
3.8
± 0.
3***
S
A↓;
cam
alex
in↓
Pse
udom
onas
syr
inga
e
Naw
rath
and
Mét
raux
199
9; H
eck
et a
l., 2
003
sid2
-2
1.6
± 0.
4*
SA↓;
cam
alex
in↑
Pse
udom
onas
syr
inga
e
Naw
rath
and
Mét
raux
199
9; H
eck
et a
l., 2
003
npr1
-3
1.5
± 0.
2*
SA↓
Pse
udom
onas
syr
inga
e
Cao
et a
l., 1
997;
Don
g, 2
004
npr1
-1
1.8
± 0.
3*
SA↓;
JA↓;
ET↓
Pse
udom
onas
syr
inga
e
A
ltern
aria
bra
ssic
icol
a C
ao e
t al.,
199
4; P
iete
rse
et a
l., 1
998;
Leon
-Rey
es e
t al.,
200
9
jar1
0.
6 ±
0.3
Inse
nsiti
ve to
JA
Dev
oto
and
Tur
ner,
200
3
jin1
0.7
± 0.
4 S
A↑;
inse
nsiti
ve to
JA
P
seud
omon
as s
yrin
gae
B
erge
r et
al.,
199
6; N
icks
tadt
et a
l., 2
004
mpk
4 2.
1 ±
0.2*
**
SA↑;
JA↓;
ET↓
Alte
rnar
ia b
rass
icic
ola
Pet
erse
n et
al.,
200
0; B
rode
rsen
et a
l., 2
006
eds8
-1
3.2
± 0.
4***
In
sens
itive
to J
A
Pse
udom
onas
syr
inga
e
Ton
et a
l., 2
002
Chapter IIa
52

Ara
bido
psis
-Phy
toph
thor
a ca
psic
i int
erac
tion
53
ein2
-1
0.1
± 0.
1 JA↑;
inse
nsiti
ve to
ET
A
ltern
aria
bra
ssic
icol
a
B
otry
tis c
iner
ea
Alo
nso
et a
l., 1
999;
Tho
mm
a et
al.,
199
9;
P
enni
nckx
et a
l., 1
998
ein3
-1
0.9
± 0.
4 In
sens
itive
to E
T
C
hao
et a
l., 1
997
etr1
-1
2.5
± 0.
2***
In
sens
itive
to E
T; H
2O2↓
; C
6-al
dehy
de-in
duce
d re
sist
anc
e↓
B
leec
ker
et a
l., 1
988;
Kis
him
oto
et a
l., 2
006
pad2
3.
9 ±
0.2*
**
SA↓;
cam
alex
in↓;
iGS↓
P
hyto
phth
ora
bras
sica
e
Gla
zebr
ook
and
Aus
ubel
199
4; R
oets
chi e
t al.,
20
01; S
chla
eppi
et a
l., 2
010
pad3
(cy
p71b
15)
3.3
± 0.
4***
C
amal
exin↓
Pse
udom
onas
syr
inga
e P
hyto
phth
ora
bras
sica
e
Gla
zebr
ook
and
Aus
ubel
199
4;
S
chuh
egge
r et
al.,
200
6
pad4
3.
3 ±
0.3*
**
SA↓;
cam
alex
in↓;
R
-gen
e-m
edia
ted
resi
stan
ce↓
H
yalo
pero
nosp
ora
para
sitic
a
Pse
udom
onas
syr
inga
e
Zho
u et
al.,
1998
; Fey
s et
al.,
200
1;
M
ert-
Tur
k et
al.,
200
3; B
arts
ch e
t al.,
200
6;
Ven
ugop
al e
t al.,
200
9
cyp7
1a13
1.
4 ±
0.2*
C
amal
exin↓
Alte
rnar
ia b
rass
icic
ola
Pse
udom
onas
syr
inga
e N
afis
i et a
l., 2
007
cyp7
9b2c
yp79
b3
4.0
± 0.
1***
C
amal
exin↓;
iGS↓
Alte
rnar
ia b
rass
icic
ola
P
seud
omon
as s
yrin
gae
B
lum
eria
gra
min
is
Naf
isi e
t al.,
200
7; B
edna
rek
et a
l., 2
009
cyp8
1f2-
2 2.
5 ±
0.2*
**
iGS↓
Blu
mer
ia g
ram
inis
P
falz
et a
l., 2
009;
Bed
nare
k et
al.,
200
9
myb
51
2.6
± 0.
4***
iG
S↓
G
igol
ashv
ili e
t al.,
200
7
pen2
3.
6 ±
0.3*
**
iGS↓
Blu
mer
ia g
ram
inis
B
edna
rek
et a
l., 2
009
myb
28m
yb29
0.
1 ±
0.1
aGS↓
Mam
estr
a br
assi
cae
Bee
kwee
lder
et a
l., 2
008
a Val
ues
repr
esen
t m
ean
DS
I (±
SD
) of
at
leas
t th
ree
inde
pend
ent
expe
rimen
ts e
ach
with
mor
e th
an 1
0 in
ocul
ated
leav
es.
Val
ues
that
ref
lect
a s
igni
fican
t
gain
of
susc
eptib
ility
com
pare
d w
ith C
ol-0
acc
ordi
ng t
o a
t te
st a
re m
arke
d w
ith *
(P
<0.
05)
or ***
(P
<0.
001)
. b ↓
: de
crea
sed;
↑:
incr
ease
d; S
A:
salic
ylic
acid
; JA
: jas
mon
ic a
cid;
ET
: eth
ylen
e; iG
S: i
ndol
e gl
ucos
inol
ates
; aG
S: a
lipha
tic g
luco
sino
late
s.
53
Arabidopsis-Phytophthora capsici interaction

Chapter IIa Arabidopsis-Phytophthora capsici interaction
54
Table S1. Primers used in this study.
Primersa 5'-3'
AtRub-F4 GCAAGTGTTGGGTTCAAAGCTGGTG
AtRub-R4 CCAGGTTGAGGAGTTACTCGGAATGCTG
CAP-Fw TTTAGTTGGGGGTCTTGTACC
CAP-Rv1 CCTCCACAACCAGCAACA a Previously reported (Silvar et al., 2005; Yadeta et al., 2011).
Figure S1. Relative quantification (RQ) of P. capsici biomass in Arabidopsis Col-0 leaves at 3
dpi by qPCR. Bars represent mean values (± SD) relative to that of LT123-inoculated leaves.
Experiments were repeated twice with similar results.
0
40
80
120
RQ

Arabidopsis-Phytophthora capsici interaction
55
Figure S2. Disease severity on tomato leaves inoculated with different P. capsici isolates.
Four-week-old tomato leaves were plug-inoculated with P. capsici. Lesion diameters were measured
at 3 dpi (two per lesion perpendicular to each other, assigned as diameter a and b) and lesion sizes
were calculated with the formula: lesionsize . Values represent mean lesion size (± SD) from
three independent experiments each with 16 inoculated leaves.
Figure S3. SA treatment of Arabidopsis NahG plants decreases susceptibility to P. capsici
isolate LT123.
NahG plants were sprayed with (A) water supplemented with 0.1% ethanol and 0.02% Silwet L-77 or
(B) a solution containing 2.5mM SA, 0.1 % ethanol and 0.02% Silwet L-77 one day before inoculation.
Photographs were taken at 3 dpi. Disease severity index of leaves treated with water and SA was 3.4
and 2.1, respectively and was determined based on 20 leaves collected in two independent
experiments.
0
2
4
6
8
10
Lesi
on s
ize
(cm
2)

IIb
Induced expression of defence-related genes in
Arabidopsis upon infection with
Phytophthora capsici
Yan Wang, Klaas Bouwmeester, Judith E. van de Mortel, Weixing Shan♯, Francine
Govers
This chapter has been published in: Plant Signaling & Behavior (2013); 8, e24618.
In collaboration with: ♯ College of Plant Protection, Northwest A&F University, Yangling, Shaanxi, China

Defence-related gene expression in Arabidopsis upon P. capsici infection
57
ABSTRACT
Recognition of pathogens by plants initiates defence responses including activation
of defence-related genes and production of antimicrobial compounds. Recently, we
reported that Phytophthora capsici can successfully infect Arabidopsis and revealed
interaction specificity among various accession-isolate combinations. We used this
novel pathosystem to demonstrate that camalexin, indole glucosinolates (iGS) and
salicylic acid (SA) have a role in defence against P. capsici. To further investigate the
role of camalexin-, iGS- and SA-related pathways in the differential interaction
between Arabidopsis and P. capsici, we monitored expression of marker genes over
time during infection. In both compatible and incompatible interactions, induction of
expression was detected, but in compatible interactions transcript levels of camalexin
and iGS marker genes were higher.

Chapter IIb
58
INTRODUCTION Phytophthora capsici is a pathogen with a broad host range and is particularly
devastating in vegetable crops worldwide. Recently, we showed that P. capsici can
also infect Arabidopsis and we anticipate that such a model pathosystem will
facilitate the genetic dissection of complex traits responsible for P. capsici resistance
in crops (Wang et al., 2013). Microscopic investigation of the infection process
revealed that inoculation of Arabidopsis Col-0 with zoospores of the highly virulent P.
capsici isolate LT263 resulted in colonization of leaf and root tissue and formation of
haustoria followed by sporulation. In contrast, inoculation of Col-0 with zoospores of
the intermediate virulent isolate LT123 resulted in an incompatible interaction
characterized by callose deposition, accumulation of active oxygen species and cell
death (Wang et al., 2013). We also observed natural variation among Arabidopsis
accessions in response to the two isolates; LT263 caused severe disease symptoms
on 29 out of 35 Arabidopsis accessions tested, while LT123 could infect only 2 out of
the 35, i.e. Ler-0 and Wei-0. Moreover, Arabidopsis mutants deficient in SA signaling
and the biosynthesis of camalexin and iGS showed compromised resistance against
P. capsici, indicating a role for these components in defence. The aim of this study
was to find additional support for the involvement of these secondary metabolites in
defence against P. capsici, and therefore we monitored the expression of several
defence-related genes in Arabidopsis upon inoculation with P. capsici over time using
real-time PCR. For inoculation we used mycelial plugs instead of zoospores; the
outcome is similar but disease development is faster thus shortening the time course.
Based on previous work, we selected two P. capsici isolates and two Arabidopsis
accessions. This allowed us to compare an incompatible and compatible interaction
on either the same accession (Col-0) inoculated with different isolates (LT123 and
LT263, respectively), or different accessions (Col-0 and Ler-0, respectively)
inoculated with the same isolate (LT123). In these interactions the disease severity
index (DSI) as defined previously, ranged from 0.2 with hardly any visible symptoms,
to 3.9 showing complete collapse of the inoculated leaves (Fig. 1).
RESULTS AND DISCUSSION The cytochrome P450 monooxygenase (CYP) encoding genes CYP71B15 (PAD3)
and CYP71A13 were selected as marker genes for the biosynthesis of camalexin
(Nafisi et al., 2007; Schuhegger et al., 2006). Another CYP gene, CYP81F2, and the
gene encoding the transcription factor MYB51 were used as markers for the iGS
biosynthesis pathway (Gigolashvili et al., 2007; Pfalz et al., 2009). In Col-0,

Defence-related gene expression in Arabidopsis upon P. capsici infection
59
Figure 1. Symptoms on Arabidopsis accessions Col-0 and Ler-0 three days after plug-
inoculation with P. capsici isolate LT263 or LT123.
The disease severity index (DSI) determined according to Wang et al. (2013) is indicated. White
arrowheads point to inoculated leaves.
inoculation with either P. capsici LT123 or LT263 leads to an increase in transcript
levels of these four genes over time with a peak at 36 hours post inoculation (hpi),
regardless of the compatible and incompatible nature of the interaction. In particular
for CYP71B15 and CYP71A13, drastic increases in transcript levels were observed
(Fig. 2) whereas this increase did not occur in mock-inoculated leaves. As shown
previously by Wang et al. (2013), loss-of-function mutants of these four genes
showed gain of susceptibility upon infection with P. capsici LT123, suggesting a role
in resistance and thus in incompatible interactions. Remarkably, at all time points the
transcript levels were significantly higher in the compatible interaction than in the
incompatible interaction (Fig. 2). This might be due to the fact that in LT263-infected
Col-0 leaves, many more cells are colonized thus resulting in a stronger response to
infection, especially for the camalexin-related marker genes. In several pathosystems,
camalexin accumulation as well as transcripts of relevant biosynthesis genes were
found to be strictly limited to the infection sites and correlated with the spatial
expansion of the pathogen (Glawischnig et al., 2004; Schuhegger et al., 2007). To
investigate if this holds for the P. capsici-Arabidopis pathosystem, we checked
expression of these marker genes during infection of Arabidopsis Ler-0. This
accession is susceptible to LT123 (DSI 2.1), but even more susceptible to LT263
(DSI 3.9). In these compatible interactions transcript levels of CYP71A13 were higher
than in the incompatible interaction of Col-0 with LT123 (DSI 0.2) but overall lower
than in the compatible interaction of Col-0 with LT263 (DSI 3.7). Remarkably, in Ler-0
basal expression of CYP71B15 is extremely low when compared to Col-0.
Nevertheless, also in infected Ler-0 leaves the CYP71B15 transcript levels increased
dramatically (more than 100-fold after 24 hpi), as shown in the inset in Fig.2. During
time, the peak in expression in LT263-infected Ler-0 leaves (with a higher DSI) was
at 24 hpi, so slightly earlier than in LT123-infected Ler-0 leaves (with a lower DSI)

Chapter IIb
60
and in Col-0, but overall the transcript changes followed a similar pattern in all
interactions. Taken together we found no evidence for a strict correlation between
transcript levels and DSI. We do however, observe a consistent pattern for the two
camalexin marker genes, namely induction of expression upon inoculation and
overall higher transcript levels in compatible compared to incompatible interactions. A
similar strong increase in the compatible interaction was found for one of the iGS
biosynthesis marker genes, i.e. CYP81F2. For the other one, MYB51, the differences
between transcript levels in the Col-0 compatible and incompatible interaction were
less pronounced and in Ler-0, MYB51 transcript levels in leaves with the lower DSI
(2.1) were overall higher than in those with a higher DSI (3.9).
Figure 2. Transcript levels of CYP71B15, CYP71A13, CYP82F2, MYB51, PR1 and PDF1.2 genes
during compatible and incompatible Arabidopsis-P. capsici interactions.
Four to five week-old Arabidopsis leaves were plug-inoculated with P. capsici LT123 (gray bars) and
LT263 (black bars). Six circular leaf discs (Ø 1 cm), with the inoculation spot in the center, were
collected from three plants at the time points indicated on the X-axes (hours post inoculation).
Subsequently, total RNA was isolated and used as template for cDNA synthesis. Transcript levels
were quantified by real-time PCR using gene-specific primers of which the sequences are shown in
the figure and were normalized with Arabidopsis Actin2 (F: 5’- TAACTCTCCCGCTATGTATGTC GC-
3’; R: 5’- GAGAGAAACCCTCGTAGATTGGC -3’). Values on the Y-axes are expressed as mean fold
changes (± SD) relative to the transcript level in mock-inoculated Col-0 leaves (white bars) at time
point 0 which was arbitrarily set as 1. Experiments were repeated twice with similar results.

Defence-related gene expression in Arabidopsis upon P. capsici infection
61
PR1, the gene that encodes a pathogenesis-related protein, was selected as
marker gene for the SA pathway (Leon-Reyes et al., 2010). In Col-0, PR1 expression
was found to be induced during the incompatible interaction with P. capsici isolate
LT123 with a maximum at 36 hpi followed by a rapid decline to the basal level. In the
compatible interaction with isolate LT263, PR1 transcript accumulation was triggered
faster, but to a lesser extent. Also in the compatible interactions of Ler-0 with LT123
and LT263, PR1 expression was triggered rather fast and the fold increase
approached the levels observed in the incompatible interaction in Col-0. As a marker
gene for the JA/ET-pathway we used PDF1.2 (Leon-Reyes et al., 2010). In Col-0, the
PDF1.2 transcript levels started to increase from 12 hpi, reaching a peak at 24 hpi
and then decreasing steadily in both the compatible and incompatible interaction.
Transcript levels in the compatible interaction, however, are significantly higher than
in the incompatible interaction. Also in Ler-0 PDF1.2 transcript levels increase upon
inoculation but to a much lesser extent than in Col-0.
CONCLUSIONS
In summary and in line with our recent report in which we screened the response of
Arabidopsis mutants to P. capsici and revealed requirement of SA, camalexin and
iGS for resistance to this pathogen, we found induced expression of relevant marker
genes in Arabidopsis challenged with P. capsici. Variations in expression levels were
detected in different accession-isolate combinations, and for nearly all marker genes
expression was higher in compatible interactions as compared to incompatible
interactions. The expression profiles, however, do not reveal any obvious correlation
between expression levels and disease phenotype. These results raise the question
why in the absence of proteins encoded by the camalexin and iGS marker genes, so
in the mutants, the plants are susceptible and can no longer stop the pathogen while
in wild-type plants these genes are highly induced upon successful infection. When
assuming that expression indeed leads to protein production, one would expect that
in the compatible interaction camalexin and iGS are produced but apparently not in
time or at sufficient levels to block disease development.
ACKNOWLEDGEMENTS
This project was financially supported by a Wageningen University sandwich PhD
fellowship (YW) and a Huygens scholarship (YW) and by the Centre for BioSystems

Chapter IIb
62
Genomics (CBSG) (KB, FG), which is part of the Netherlands Genomics Initiative/
Netherlands Organisation for Scientific Research.
REFERENCES
Gigolashvili T., Berger B., Mock H.P., Müller C., Weisshaar B. and Flügge U.I. (2007). The
transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis
thaliana. The Plant Journal, 50, 886-901.
Glawischnig E, Hansen B.G., Olsen C.E. and Halkier B.A. (2004). Camalexin is synthesized from
indole-3-acetaidoxime, a key branching point between primary and secondary metabolism in
Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America,
101, 8245-8250.
Leon-Reyes, A., Van der Does, D., De Lange, E.S., Delker, C., Wasternack, C.,Van Wees, S.C.M.,
Ritsema, T. and Pieterse, C.M.J. (2010). Salicylate-mediated suppression of jasmonate-
responsive gene expression in Arabidopsis is targeted downstream of the jasmonate biosynthesis
pathway. Planta, 232, 1423-1432.
Nafisi M., Goregaoker S., Botanga C.J., Glawischnig E., Olsen C.E., Halkier B.A. and Glazebrook J.
(2007). Arabidopsis cytochrome P450 monooxygenase 71A13 catalyzes the conversion of indole-
3-acetaldoxime in camalexin synthesis. Plant Cell, 19, 2039-2052.
Pfalz M., Vogel H. and Kroymann J. (2009). The gene controlling the Indole Glucosinolate Modifier1
quantitative trait locus alters indole glucosinolate structures and aphid resistance in Arabidopsis.
Plant Cell, 21, 985-999.
Schuhegger R., Nafisi M., Mansourova M., Petersen B.L., Olsen C.E., Svatoš A., Halkier B.A. and
Glawischnig E. (2006). CYP71B15 (PAD3) catalyzes the final step in camalexin biosynthesis.
Plant Physiology, 141, 1248-1254.
Schuhegger R., Rauhut T. and Glawischnig E. (2007). Regulatory variability of camalexin biosynthesis.
Plant Physiology, 164, 636-644.
Wang Y., Bouwmeester K., van de Mortel J.E., Shan W. and Govers F. (2013). A novel Arabidopsis-
oomycete pathosystem; differential interactions with Phytophthora capsici reveal a role for
camalexin, indole glucosinolates and salicylic acid in defense. Plant, Cell & Environment, 36,
1192-1203.

Chapter III
Phenotypic analyses of Arabidopsis T-DNA insertion
lines and expression profiling reveal that multiple
L-type lectin receptor kinases are involved in
plant immunity
Yan Wang, Klaas Bouwmeester, Patrick Beseh, Weixing Shan♯, Francine Govers
This chapter has been published in: Molecular Plant-Microbe Interactions (2014); 27,
1390-1402.
In collaboration with: ♯ College of Plant Protection, Northwest A&F University, Yangling, Shaanxi, China

Chapter III
64
ABSTRACT
L-type lectin receptor kinases (LecRKs) are membrane-spanning receptor-like
kinases with putative roles in biotic and abiotic stress responses and in plant
development. In Arabidopsis, 45 LecRKs were identified but their functions are
largely unknown. Here, a systematic functional analysis was carried out by evaluating
phenotypic changes of Arabidopsis LecRK T-DNA insertion lines in plant
development and upon exposure to various external stimuli. None of the LecRK T-
DNA insertion lines showed clear developmental changes, neither under normal
conditions nor upon abiotic stress treatment. However, many of the T-DNA insertion
lines showed altered resistance to Phytophthora brassicae, Phytophthora capsici,
Pseudomonas syringae or Alternaria brassicicola. One mutant defective in LecRK-
V.5 expression was compromised in resistance to two Phytophthora spp. but showed
enhanced resistance to P. syringae. LecRK-V.5 overexpression confirmed its dual
role in resistance and susceptibility depending on the pathogen. Combined analysis
of these phenotypic data and LecRK expression profiles retrieved from public
datasets revealed that LecRKs which are hardly induced upon infection or even
suppressed are also involved in pathogen resistance. Computed co-expression
analysis revealed that LecRKs with similar function displayed diverse expression
patterns. Since LecRKs are widespread in plants, the results presented here provide
invaluable information for exploring the potential of LecRKs as novel sources of
resistance in crops.

Function of Arabidopsis LecRKs in resistance
65
INTRODUCTION
Plasma membrane-localized receptor-like kinases (RLKs) are critical for plants to
perceive and process environmental stimuli. In Arabidopsis, there are over 400
plasma membrane-localized RLKs with a conserved cytosolic kinase domain and
divergent extracellular domains (Shiu and Bleecker, 2003). Several of these RLKs
have been analysed in detail and demonstrated to play essential roles in plant
development and/or plant immunity. For example, the RLK CLAVATA1 controls
meristem cell proliferation and differentiation (Dievart et al., 2003), and RLKs such as
FLS2, EFR and CERK1 recognize microbe-associated molecular patterns (MAMPs)
to initiate MAMP-triggered immunity (Boller and Felix, 2009). Other RLKs, such as
BAK1/SERK3 and SOBIR1, function as central components in regulation of various
receptor-mediated signaling processes (Chinchilla et al., 2009; Liebrand et al., 2013).
More insight into the function of a wider and more diverse range of RLKs would help
to better understand how plants interact with their environment.
Based on the divergent extracellular domains, RLKs can be divided into
different classes. One of the largest RLK classes in plants comprises the L-type lectin
receptor kinases, abbreviated as LecRKs. LecRKs are characterized by an
extracellular lectin domain structurally resembling soluble lectins of leguminous
plants (Hervé et al., 1996). In Arabidopsis, 45 LecRKs have been identified that were
divided into nine clades (i.e. Clade I to IX) and seven singletons (Bouwmeester and
Govers, 2009). Given this large number, LecRKs are expected to play diverse roles
in plant development and defence. Thus far, functional characterization has been
limited to a few Arabidopsis LecRKs. For example, LecRK-IV.2 (alias SGC lectin RLK)
was suggested to be essential for proper pollen development since a T-DNA
insertion mutant showed deformed and collapsed pollen grains (Wan et al., 2008).
LecRK-V.2 (alias LecRK-b2) and three LecRKs from clade VI (previously named
LecRK4.1, LecRK4.2 and LecRK4.3) were shown to be involved in regulating
abscisic acid (ABA) response during seed germination (Deng et al., 2009; Xin et al.,
2009). Later studies also demonstrated a role for LecRKs in plant immunity. LecRK-
I.9, a mediator of plant cell wall-plasma membrane integrity (Gouget et al., 2006),
was shown to be crucial for Phytophthora resistance in Arabidopsis (Bouwmeester et
al., 2011). More recently the same LecRK was identified as the first ATP receptor in
plants and shown to be required for ATP-mediated responses (Choi et al., 2014). As
yet it is not clear if the function of LecRK-I.9 as Phytophthora resistance component
is linked to its function as ATP receptor. Two other LecRKs, LecRK-V.5 and LecRK-
VI.2, were shown to be involved in bacterial resistance by regulating stomatal
immunity (Desclos-Theveniau et al., 2012; Singh et al., 2012). The latter was found
to associate with the bacterial flagellin-receptor FLS2 and to be a positive regulator of

Chapter III
66
the flagellin-mediated defence responses (Huang et al., 2014). LecRK-IV.3 (alias
AtLPK1) might have a role in resistance to fungal pathogens because LecRK-IV.3
overexpression in Arabidopsis resulted in increased resistance to Botrytis cinerea
(Huang et al., 2013). For several other LecRKs, a function in plant growth and/or
stress response was anticipated based on in silico expression analysis
(Bouwmeester and Govers, 2009), but as yet experimental evidence for this is
lacking.
The objective of the current work was to elucidate the biological function of
individual LecRKs in Arabidopsis. LecRK T-DNA insertion lines were obtained and
analysed for phenotypic changes (i) in plant development under normal conditions or
abiotic stress and (ii) upon exposure to pathogens. The phenotypic data were then
integrated with in silico expression data with the aim to uncover potential
associations between LecRK gene expression and biological function. In addition,
LecRK-V.5, of which a mutant showed altered disease phenotypes, was selected for
more detailed study to validate its function.
RESULTS
Assembling the collection of Arabidopsis LecRK T-DNA insertion lines
The T-DNA Express database of the SALK Institute Genome Analysis Laboratory
(SIGnAL; http://signal.salk.edu) was used to search for T-DNA insertion lines for all
45 LecRKs (Table 1). Preferably, homozygous lines bearing T-DNA insertions within
the coding regions were selected to enhance the possibility of complete disruption of
gene function. If not available, homozygous lines with insertions in the promoter (5
lines), 5’UTR (14) or 3’UTR (2) or segregating lines were obtained. In total we
obtained 62 T-DNA insertion lines covering 41 LecRKs, all in a Col-0 background.
The seeds of the only T-DNA insertion line available for LecRK-IV.3 (i.e.
SAIL_529_D07) did not geminate, thereby reducing the number of lines to 61
covering 40 LecRKs. Of these 61, 52 were indicated as homozygous lines and 9 as
segregating lines. To confirm this, we performed PCR analysis and found that 42 out
of the 52 lines were indeed homozygous with correct T-DNA insertion sites. Of the
remaining ten, eight were found to be segregating at the site of the T-DNA insertion,
and two (i.e. SALK_053703C and SALK_019496C) did not show a T-DNA insertion
in the anticipated target gene (i.e. LecRK-IV.1) indicating that these lines are either
untransformed Col-0 or contain a T-DNA insertion elsewhere in the genome. So in
total we ended up with 17 segregating lines. These were propagated and the
offspring was genotyped to select the homozygous ones. For five lines however, we
were not able to detect any homozygous offspring among over 60 plants tested for

Function of Arabidopsis LecRKs in resistance
67
each individual line. This was the case for the single T-DNA insertion line that was
available for LecRK-II.2 as well as for the two lines mutated in LecRK-V.6 and the
two in LecRK-S.5. The consistency among the two independent LecRK-V.6 lines and
the two independent LecRK-S.5 lines suggests that a complete knockout of these
genes is lethal raising the possibility that these LecRKs have a crucial role in
development. In total 54 lines representing 36 individual LecRKs were confirmed as
homozygous T-DNA insertion lines and used for further analysis (Table 1).
Table 1. Overview of the clades and members of LecRK gene family in Arabidopsis, the T-DNA
insertion lines used in this study and their phenotypes in response to pathogen infection and
abiotic stress. The response to P. brassicae and P. capsici is depicted by bars that show the DSI
obtained from 20 to 40 inoculated leaves in three independent experiments. The DSI scale ranges
from 0 (light grey) to 4 (dark grey) as shown below the table. The response to P. capsici, P. parasitica,
A. brassicicola, Pst DC3000 and to abiotic stress is depicted by a ● when there is no difference
compared with Col-0, and by S (Susceptible), R (Resistant), EN (Enhanced necrotic symptoms but not
increased bacterial proliferation) or ES (Enhanced necrotic symptoms and increased bacterial
proliferation) to indicate the disease phenotype. Infection assays with P. capsici LT62 were performed
once while infection assays with all other pathogens were repeated at least three times.
Footnotes to Table 1 (see next page) a For alternative gene names and alternative mutant names see Table S1. b No T-DNA insertion lines
available. c The obtained SAIL_529_D07 line did not germinate. d SALK_053703C and
SALK_019496C were indicated as lines with T-DNA insertions in At2g37710 but in our analysis this
could not be confirmed. e >< : homozygous; >: segregating; /: no insertion in the gene under study. f
With 10-fold higher inoculation densities lecrk-I.9-1 and lecrk-I.9-2 show enhanced susceptibility to Pst
DC3000 as reported by Bouwmeester et al. (2011). g Similar results were obtained as described by
Desclos-Theveniau et al. (2012) and Singh et al. (2012). * T-DNA insertion lines showing significantly
higher or lower DSI when compared with Col-0 based on a t test, p < 0.05.

Chapter III
68

Function of Arabidopsis LecRKs in resistance
69
Arabidopsis LecRK T-DNA insertion lines showed no phenotypic changes in
growth and development
To determine whether LecRKs are involved in plant growth and development, LecRK
T-DNA insertion lines were grown in soil and analysed for phenotypic changes in
rosette size, inflorescence emergence, and in flower and silique morphology for up to
8 weeks. In addition, seeds were grown in vitro on vertical 0.5 MS plates to monitor
shoot and root growth. Under these conditions, no consistent growth alterations were
observed in comparison to Col-0 (Table 1). Previously, Wan et al. (2008) described
that a homozygous mutant with two insertions in the region encoding the lectin
domain of LecRK-IV.2 developed abnormal siliques and they concluded that LecRK-
IV.2 is required for proper pollen development. The T-DNA insertion line lecrk-IV.2
that we analysed harbours a T-DNA insertion in the 5’UTR, at ~70 bp before the start
codon. Since this line showed no defects in silique development we analysed the
LecRK-IV.2 expression by qRT-PCR and found that the transcript level is only slightly
reduced when compared with Col-0 (approximately 40%) (Supplemental Fig. S1, A
and B). This reduction is apparently not sufficient to disturb normal silique
development (Supplemental Fig. S1C). Unfortunately the mutant analysed by Wan et
al. (2008) is no longer available in public resources thus hampering further functional
analysis of LecRK-IV.2.
Response of LecRK T-DNA insertion lines to abiotic stress
To evaluate the role of LecRKs in resistance to abiotic stresses, the LecRK T-DNA
insertion lines were grown on 0.5 MS medium supplemented with 100 mM NaCl or
200 mM mannitol. Plants were checked for changes in rosette size and growth of
shoots and roots. No consistent differences were observed between any of the
LecRK T-DNA insertion lines and Col-0. Previously, Deng et al. (2009) observed that
a line with a T-DNA insertion in LecRK-V.2 showed enhanced tolerance to NaCl and
mannitol and developed longer primary roots than Col-0 and concluded that LecRK-
V.2 is involved in salt and osmotic stress. In our study, lecrk-V.2, a T-DNA insertion
line with an insertion in the 3’UTR, ~60 bp after the stop codon showed similar
growth as Col-0 upon treatment with NaCl or mannitol, also when treated using
exactly the same conditions as described by Deng et al. (2009) (Table 1;
Supplemental Fig. S2, A and B). To find a reason for this discrepancy, we
determined the LecRK-V.2 expression in lecrk-V.2 and Col-0 by qRT-PCR.
Remarkably, the transcript level in lecrk-V.2 was higher than in Col-0, instead of
lower (Supplemental Fig. S2C). Possibly the T-DNA insertion in the 3’UTR in one
way or another stabilizes the mRNA and this may explain why we did not find defects
in salt and osmotic stress responses. T-DNA insertion line lecrk-V.2 was excluded
from further analysis.

Chapter III
70
Response of LecRK T-DNA insertion lines to Phytophthora pathogens
Previously, LecRK-I.9 was reported as a novel resistance component against
Phytophthora brassicae in Arabidopsis (Bouwmeester et al., 2011). To determine the
involvement of other LecRKs in Phytophthora resistance, we performed infection
assays on the LecRK T-DNA insertion lines with P. brassicae, Phytophthora capsici
and Phytophthora parasitica. Recently it was reported that latter two are able to infect
Arabidopsis (Wang et al., 2011; Wang et al., 2013).
In order to identify LecRKs contributing to Phytophthora resistance, infection
assays were performed with isolates that are not able to infect Arabidopsis accession
Col-0 i.e. P. brassicae HH and P. capsici LT123. As shown by Bouwmeester et al.
(2011), isolate HH can successful infect LecRK-I.9 mutants. Inoculation of lecrk-I.9-1
and lecrk-I.9-2 with P. brassicae HH resulted in water-soaked lesions at four days
post inoculation (dpi) with a mean DSI of 2.2 and 2.4, respectively (Supplemental Fig.
S3A). P. capsici LT123 caused similar disease symptoms on these mutants and at 4
dpi the mean DSI was 2.0 and 1.9, respectively (Supplemental Fig. S3B). This shows
that LecRK-I.9 also contributes to resistance against P. capsici.
Inoculation of all available LecRK T-DNA insertion lines with P. brassicae HH
and P. capsici LT123 resulted in evident disease symptoms on several lines. For
example, lecrk-S.1, which contains a T-DNA insertion in the region encoding the
extracellular lectin domain and has a 90% reduction in LecRK-S.1 expression level
(Fig. 1, A and B), showed a mean DSI of 2.5 and 2.1 with HH and LT123,
respectively (Fig. 1C). In total 19 T-DNA insertion lines representing 13 LecRKs from
different clades showed a significantly higher DSI at 4 dpi than Col-0 (Table 1). From
these 13 LecRKs, three are from clade V, two from clade IX and four are singletons.
The four remaining ones are from clade I, III, IV and VIII, respectively. For four out of
13, two T-DNA insertion lines are available which both showed enhanced disease
severity when challenged with Phytophthora, with at least one of the two having an
insertion in the coding region (Table 1). For two other LecRKs for which two or more
T-DNA insertion lines are available, T-DNA insertion lines with different insertion sites
showed discrepancy in susceptibility. In the case of LecRK-VIII.2 with three lines
having insertions in the coding region, only lecrk-VIII.2-3 developed rapid spreading
lesions, resulting in a DSI ≥ 2 on approximately 80% of the inoculated leaves at 4 dpi.
The other two lines, lecrk-VIII.2-1 and lecrk-VIII.2-2 also showed disease symptoms
but the disease severity was less pronounced (Table 1). In the case of LecRK-S.7,
the two T-DNA insertion lines showed opposite responses. lecrk-S.7-1 is fully
resistant to both P. brassicae HH and P. capsici LT123, while lecrk-S.7-2 showed
clear disease symptoms with a mean DSI of 2.0 or 1.9, respectively (Table 1). Both
lines have the T-DNA insertion in the 5’UTR but at different positions. Expression
analysis revealed that the difference in phenotype between lecrk-S.7-1 and lecrk-S.7-

Function of Arabidopsis LecRKs in resistance
71
Figure 1. Response of Arabidopsis LecRK T-DNA insertion lines to infection by Phytophthora
spp.
A, Schematic model representing LecRK-S.1. The black arrowhead points to the T-DNA insertion site
in mutant lecrk-S.1 while the head-to-head arrow pairs indicate the position of the primers (S.1-RT)
used for qRT-PCR. B, Relative quantification of LecRK-S.1 transcript levels in Col-0 and lecrk-S.1 by
qRT-PCR. C, Disease symptoms on Col-0 and lecrk-S.1 at four days after plug-inoculation (dpi) with
P. brassicae HH and P. capsici LT123. White arrowheads point to inoculated leaves. Each bar
represents the DSI obtained from at least 30 leaves in three independent experiments with the number
showing the average DSI and a * a significant difference (p < 0.05) according to a t test. D, Disease
symptoms on T-DNA insertion line lecrk-I.7 and Col-0 inoculated with P. brassicae CBS686.95 and P.
capsici LT263 at 3 dpi. * indicates significant difference in DSI (p < 0.05) according to a t test.

Chapter III
72
2 correlates with LecRK-S.7 expression levels. In lecrk-S.7-2 expression is reduced
compared with Col-0 whereas in lecrk-S.7-1 it is increased (Supplemental Fig. S4B).
For the seven remaining LecRKs of which T-DNA insertion lines showed a change in
phenotype in the Phytophthora disease assays, only a single T-DNA insertion line
was available with six (i.e. lecrk-III.2, lecrk-V.4, lecrk-V.7, lecrk-IX.2, lecrk-S.1 and
lecrk-S.6) having a T-DNA insertion in the coding region and one (i.e. lecrk-V.5-3) in
the 5’UTR (Table 1).
When challenging the LecRK T-DNA insertion lines with Phytophthora isolates
that are less virulent, in this case P. capsici LT62 and P. parasitica Pp009, none of
the LecRK T-DNA insertion lines showed lesions (Table 1). They remained resistant
similar to Col-0. This suggests that these LecRKs are not essential to stop less
virulent isolates but might function as extra barrier for more virulent isolates.
To investigate the potential role of LecRKs in suppressing Phytophthora
resistance, LecRK T-DNA insertion lines were challenged with Phytophthora isolates
that are capable of infecting Col-0, i.e. P. brassicae CBS686.95 and P. capsici LT263.
Out of all tested LecRK T-DNA insertion lines, only lecrk-I.7, with an insertion in the
region encoding the kinase, showed slightly smaller lesions when compared with Col-
0 (Fig. 1D). All the other T-DNA insertion lines showed a comparable or slightly
higher disease severity than Col-0 (Table 1).
Response of LecRK T-DNA insertion lines to Pseudomonas syringae and
Alternaria brassicicola
Previously, LecRK-VI.2 was reported to play a role in resistance to bacterial
pathogens (Singh et al., 2012). To determine the involvement of other LecRKs in
bacterial resistance, we performed infection assays with Pseudomonas syringae
(Pst) DC3000. Upon spray inoculation, T-DNA insertion lines of eight LecRKs
showed enhanced yellowing and necrosis at 4 dpi when compared with Col-0 (Table
1; Fig. 2A). For three of these, two T-DNA insertion lines are available which both
showed enhanced symptoms when challenged with Pst DC3000, with at least one of
the two having an insertion in the coding region (Table 1). To determine to what
extent the visible symptoms are correlated with enhanced Arabidopsis susceptibility,
bacterial colonization was quantified in rosette leaves. In line with Singh et al., (2012),
we detected significantly higher bacterial titers in lecrk-VI.2-2 in comparison to Col-0
(Fig. 2B). Also in lesions on the two T-DNA insertion lines from LecRK-IV.4, the one
from LecRK-S.1 and two from LecRK-S.4, significantly higher bacterial titers were
detected and this was reproducible (Fig. 2B). Thus, these T-DNA insertion lines were
scored as ES (enhanced susceptibility). The phenotype of the remaining four T-DNA
insertion lines (i.e. lecrk-IV.2, lecrk-V.4, lecrk-V.7 and lecrk-S.6) was scored as EN
(enhanced yellowing and necrosis) rather than ES (Table 1). This is because the

Function of Arabidopsis LecRKs in resistance
73
bacterial titers were not consistently higher in all assays despite the fact that the
enhanced susceptibility phenotypes were reproducible.
Figure 2. Response of Arabidopsis LecRK T-DNA insertion lines to infection by Pseudomonas
syringae and Alternaria brassicicola.
A, Disease symptoms on Col-0, lecrk-S.1 and lecrk-VI.2-2 four days after inoculation with Pst DC3000
(108 cfu ml-1). B, Bacterial growth in Col-0 and various LecRK T-DNA insertion lines at 3 dpi. Bars
depict mean bacterial titers of four plants (± SD). * indicates p < 0.05 according to a t test for
comparisons between Col-0 and LecRK T-DNA insertion lines. Experiments were repeated three
times with similar results. C, Disease symptoms on Col-0, pad3 and various LecRK T-DNA insertion
lines five days after inoculation with A. brassicicola MUCL20297.
To determine the potential involvement of LecRKs in non-host resistance, we
tested the response of the LecRK T-DNA insertion lines to Alternaria brassicicola, a
fungal pathogen that is not pathogenic on Col-0 but is capable to cause disease
symptoms on the camalexin deficient mutant pad3 (van Wees et al., 2003). The

Chapter III
74
majority of the LecRK T-DNA insertion lines showed no visible symptoms up to 5 dpi.
Five T-DNA insertion lines however, representing four clade I LecRKs, showed a
slight gain of susceptibility (Table 1; Fig. 2C). Two of three LecRK-I.3 T-DNA
insertion lines showed the phenotype, one with an insertion in the exon (lecrk-I.3-3)
and the other in the 5’UTR (lecrk-I.3-1). The third one (i.e. lecrk-I.3-2) also with a
5’UTR insertion was not infected. Also in the case of LecRK-I.8 the exon mutant
(lecrk-I.8-2) showed gain of susceptibility but not the T-DNA insertion line with a
5’UTR insertion (lecrk-I.8-1).
LecRK-V.5 plays a dual role in resistance to Pseudomonas syringae and
Phytophthora pathogens
Our infection assays revealed that the mutant lecRK-V.5-3 showed gain of
susceptibility to P. brassicae HH and P. capsici LT123 (Table 1; Fig. 3, A-C). In
contrast, Desclos-Theveniau et al. (2012) reported that mutants of LecRK-V.5
showed enhanced resistance towards Pst DC3000. Because of these opposite
phenotypes with respect to different pathogens we decided to investigate the function
of LecRK-V.5 in more detail.
Gain of Phytophthora susceptibility in lecrk-V.5-3 points to a role of LecRK-V.5
in Phytophthora resistance. To confirm this, we generated transgenic Arabidopsis
lines overexpressing LecRK-V.5 in a Col-0 background. Two lines (i.e. 35S-LecRK-
V.5-1/2) with constitutive and strongly enhanced LecRK-V.5 expression were
selected (Fig. 3B). Neither of the 35S-LecRK-V.5 lines showed any visible
developmental defects. Inoculation of Col-0 with P. brassicae CBS686.95 and
P. capsici LT263 resulted in almost fully colonized leaves at 3 dpi (Fig. 3D). In
contrast, the two 35S-LecRK-V.5 lines showed reduced lesion sizes with a DSI of
approximately 2.0 (Fig. 3D). To determine if the reduced disease severity on the
LecRK-V.5 overexpression lines is indeed correlated with enhanced resistance,
Phytophthora biomass was quantified by qRT-PCR in leaves inoculated with
P. capsici LT263 at 3 dpi. This showed that the P. capsici biomass in the 35S-LecRK-
V.5 lines was reduced by at least 50% compared with Col-0 (Fig. 3E).
To confirm the role of LecRK-V.5 in resistance to Pst DC3000 as postulated
by Desclos-Theveniau et al. (2012), we compared the phenotype of lecrk-V.5-3 with
the phenotypes of the previously analysed mutants lecrk-V.5-1 and lecrk-V.5-2.
Similar to lecrk-V.5-1 and lecrk-V.5-2, lecrk-V.5-3 showed less necrotic symptoms
and reduced bacterial proliferation compared with Col-0 (Fig. 3, F and G). Moreover,
both 35S-LecRK-V.5 lines showed enhanced susceptibility to Pst DC3000 with
increased necrosis and higher bacterial titers than Col-0 (Fig. 3, F and G). Taken
together, these results demonstrate that LecRK-V.5 has a dual role in regulating
defence to different pathogens.

Function of Arabidopsis LecRKs in resistance
75

Chapter III
76
Figure 3. LecRK-V.5 plays opposite roles in resistance to Phytophthora and Pseudomonas
syringae.
A, Schematic model representing LecRK-V.5. The black arrowhead points to the T-DNA insertion site
in lecrk-V.5-3 while the head-to-head arrow pairs indicate the position of the primers (V.5-RT) used for
qRT-PCR. B, qRT-PCR analysis of LecRK-V.5 transcript levels in Col-0, lecrk-V.5-3 and two
overexpression lines. * a significant difference (p < 0.05) according to a t test. C, Disease symptoms
on Col-0 and lecrk-V.5-3 four days after inoculation with P. brassicae HH and P. capsici LT123. White
arrowheads point to inoculated leaves. Each bar represents the DSI obtained from at least 30 leaves
in three independent experiments with the number showing the average DSI and a * a significant
difference (p < 0.05) according to a t test. D, Disease symptoms on Col-0 and two LecRK-V.5
overexpression lines three days after inoculation with P. brassicae CBS686.95 and P. capsici LT263.
E, Relative quantification of P. capsici biomass (RQ) in Col-0 and overexpression lines at 3 dpi by
qPCR. Bars represent mean values (± SD) relative to that of infected Col-0 leaves. * indicates
significantly less biomass (p < 0.05) according to a t test. Experiments were repeated twice with
similar results. F, Disease symptoms on Col-0, lecrk-V.5-3 and two overexpression lines four days
after inoculation with Pst DC3000. G, Bacterial growth in Col-0, lecrk-V.5-3 and two overexpression
lines at 3 dpi. Bars shown are mean values (± SD). * indicates significantly lower or higher bacterial
titers (p < 0.05) according to a t test; Experiments were repeated twice with similar results.
Are there associations between LecRK mutant phenotypes and LecRK
expression profiles?
Expression profiling analysis has been widely used to predict potential biological
function of genes (Li et al., 2006; Fukushima et al., 2012). Here, we explored the
transcriptome-to-phenotype relations by comparing LecRK expression profiles
retrieved from publicly available microarray data, with the mutant phenotypes

Function of Arabidopsis LecRKs in resistance
77
obtained in this study (Fig. 4). Since there is no genome-wide expression data
available from Arabidopsis inoculated with P. brassicae or P. capsici, we used the
expression data derived from Arabidopsis leaves inoculated with
Phytophthora infestans, a Phytophthora species that is not capable of infecting
Arabidopsis (Huitema et al., 2003). The 13 LecRKs, of which T-DNA insertion lines
showed compromised resistance to P. brassicae HH and P. capsici LT123, were
found to be differentially expressed during P. infestans infection (Fig. 4A). LecRK-
IX.2 was highly induced and the expression level was increased up to 50-fold (log2
value of 5.7) at 6 hpi. LecRK-I.9, LecRK-V.5 and LecRK-V.7 showed induced
expression with a maximum of four-fold-increase, whereas the other nine LecRKs are
either hardly expressed or even down-regulated during infection process. The same
holds for LecRK-I.7, the only LecRK of which the mutant showed slightly enhanced
Phytophthora resistance.
The nine LecRKs of which T-DNA insertion lines showed altered phenotypes
in response to Pst DC3000 (EN, ES or R) were all down-regulated at one or more
time points after inoculation with Pst DC3000 (Fig. 4A). The five LecRKs with T-DNA
insertion lines showing significantly altered bacterial colonization (ES or R) were
down-regulated at 12 hpi with log2 values ranging from -0.7 to -1.4.
Despite the fact that several LecRKs were differently expressed in various
tissues and in response to NaCl or mannitol treatment, none of the tested T-DNA
insertion lines showed consistent phenotypic changes during growth and
development or when exposed to abiotic stress. Hence, it is difficult to reconcile the
biological function of these LecRKs with expression profiles. However, lack of
phenotypic changes in mutants does not exclude that certain LecRKs have a function
in these processes. It could well be that functional redundancy masks the
phenotypes caused by a single gene disruption.
Genes that exhibit similar expression patterns are often expected to be
involved in the same biological processes (Eisen et al., 1998; Fukushima et al., 2012).
We computed co-expression profiles of all LecRKs in leaves inoculated with
P. infestans and Pst DC3000 over time. In order to easily visualize LecRK expression
patterns, expression levels (log2 values) were normalized by Z-score transformation
(Cheadle et al., 2003). LecRKs with similar expression patterns were grouped
together using hierarchical clustering with Pearson’s correlation coefficient (r) and
average linkage. Pearson’s r is a metric to evaluate the strength of the correlation
between variables, ranging from -1 to 1 (Usadel et al., 2009) and therefore we used
the cut-off value of r ≥ 0.6 for clustering which, in general, indicates strong positive
correlation (Aoki et al., 2007; Fukushima et al., 2012). With this criterion, LecRKs
could be grouped into five clusters based on their expression patterns upon infection

Chapter III
78
Figure 4. Compilation of expression profiles of LecRKs and phenotypes of LecRK T-DNA
insertion lines.

Function of Arabidopsis LecRKs in resistance
79
A, The heat map in the left panel shows LecRK expression levels in different plant tissues and in
response to various biotic and abiotic stresses. Signal intensity log2 ratios are color-coded in the range
of -3 (purple) to 6 (green) as indicated in the scale bar, and represent the relative expression levels
compared with corresponding controls. The table on the right shows the phenotypes of the LecRK T-
DNA insertion lines compiled from the results presented in Table 1 with ‘R’ standing for ‘resistant’, ‘S’
for ‘susceptible’, ‘EN’ for ‘enhanced necrotic symptoms but no increased bacterial proliferation’ and
‘ES’ for ‘enhanced necrotic symptoms and increased bacterial proliferation. ● depicts ‘no consistent
phenotypic changes’ and ─ ‘not tested’. B-D, Hierarchical clustering of LecRKs based on expression
patterns at 6, 12 and 24 hours after inoculation with P. infestans (C), 2, 6 and 24 hours after
inoculation with Pst DC3000 (D) or combined (B) using Pearson’s correlation coefficient (r) of per
gene Z-score transformed log2 values (sample relative to control). Clusters were created using a cut-
off value of r ≥ 0.6. In (C) and (D) clusters are indicated by vertical colored bars connected to panels
showing LecRK expression patterns in each cluster. The lengths of the branches (da) are the scaled
(black lines) and unscaled (grey lines) average Pearson correlation distances (dp = 2da = 1 - r).
Normalized expression data are color-coded to indicate higher (yellow, saturated at 1.2) and lower
(blue, saturated at -1.2) expression relative to the respective controls. œ On the microarray the same
probes represent LecRK-V.7 and LecRK-V.8 and thus the expression values are identical. In (A-D)
LecRKs involved in Phytophthora resistance (S or R) are depicted in green and those involved in
Pst DC3000 resistance (ES or R) in dark red. LecRKs involved in both are depicted in combined
green-red.
with P. infestans (Fig. 4C) and another five clusters upon infection with Pst DC3000
(Fig. 4D). Of the 14 LecRKs of which T-DNA insertion lines showed altered
susceptibility to Phytophthora spp., five were grouped in cluster II and showed a
slight reduction in expression between 6 and 24 hpi. The four LecRKs that grouped in
cluster V show an opposite expression pattern compared with LecRKs in cluster II. Of
the remaining ones, two are in cluster I, i.e. LecRK-I.9 and LecRK-S.1, two in cluster
III, i.e. LecRK-S.4 and LecRK-S.7 and one in cluster IV, i.e. LecRK-III.2. Similar
transcriptome-to-phenotype patterns were observed upon challenging with
Pst DC3000 (Fig. 4D). Of the five LecRKs of which T-DNA insertion lines showed
altered susceptibility to Pst DC3000, LecRK-V.5 and LecRK-VI.2 are combined in
cluster I, LecRK-S.1 and LecRK-S.4 in cluster II while LecRK-IV.4 was allocated to
cluster III. When combining the expression patterns of the two time courses obtained
from leaves inoculated with the two different pathogens, 27 of the 45 LecRKs
grouped in clusters, 11 in total, while the remaining 18 did not share their expression
pattern with any other LecRK (Fig. 4B). Although the four LecRKs that are implicated
in resistance to both Phytophthora pathogens and Pst DC3000, do not group in one
cluster and are far apart in the dendrogram, the clustering of two, respectively three
relevant LecRKs is maintained, i.e. LecRK-V.4 and LecRK-VIII.2 and the combination
of LecRK-V.5, LecRK-VI.2 and LecRK-IX.2.

Chapter III
80
DISCUSSION
Most of the plant immune receptors that have been identified to date are either
plasma membrane-localized receptors including RLKs and receptor-like proteins
(RLPs) or intracellular nucleotide binding-leucine-rich repeat proteins (NLRs) (Spoel
and Dong, 2012). The majority was discovered based on their ability to activate
defence responses upon recognition of MAMPs or pathogen effectors. For example,
the RLK gene encoding the MAMP receptor FLS2 was isolated by a forward genetics
approach based on screening random mutants for flagellin-insensitive mutants,
followed by map-based cloning of the locus harbouring the mutation (Gómez-Gómez
and Boller, 2000). Also the isolation of several NLR genes that function as major R-
gene in ‘gene-for-gene’ interactions was the result of combining phenotype (i.e.
pathogen recognition) and map based cloning, in this case facilitated by linkage
mapping using segregating progenies of resistant and susceptible parents (Marone
et al., 2013; Rodewald and Trognitz, 2013). Genome sequencing has revealed that
defence-related RLKs and NLRs often have many homologs of which the function is
unknown. They are grouped into families and sub-families based on sequence
similarity and shared domain architecture.
In this study we focused on one such RLK sub-family in Arabidopsis, namely
the LecRK family of which three out of 45 members were previously identified as
critical players in Arabidopsis innate immunity. Two of the three were selected for
functional studies based on their increased expression in plants treated with priming
agent BABA (LecRK-VI.2; Singh et al., 2012) or oligogalacturonic acid (LecRK-V.5;
Desclos-Theveniau et al., 2012). The third one was selected via a phage display
aimed at identifying RGD-binding proteins in plants (Gouget et al., 2006) and
appeared to be a potential host target of a P. infestans RXLR effector (LecRK-I.9;
Bouwmeester et al., 2011). We anticipated that functional screening of other
members of the LecRK family would reveal several novel candidates with a role in
disease resistance. In an unbiased strategy we monitored all available LecRK T-DNA
insertion lines and indeed, besides LecRK-I.9, LecRK-V.5 and LecRK-VI.2 mentioned
above, we found 16 additional LecRKs of which mutants showed phenotypic changes
in response to pathogens. T-DNA insertion lines are effective tools for determining
gene function and for linking genotype to phenotype (O’Malley and Ecker, 2010;
Lloyd and Meinke, 2012). In other studies similar unbiased screenings of T-DNA
insertion line collections resulted in novel insight in gene function. An example is the
study of ten Hove et al. (2011) who performed large scale screening of T-DNA
insertion lines of root-expressed leucine-rich repeat (LRR) RLKs and identified LRR-
RLKs implicated in hormone and abiotic stress responses. In a similar way Wang et
al. (2008) performed a screening of a broad range of T-DNA insertion lines in 56

Function of Arabidopsis LecRKs in resistance
81
genes encoding RLPs and assigned functions to these RLPs in plant development
and resistance. It should be noted that the success rate in these two studies was
relatively low; only few T-DNA insertion lines were found that could be linked to the
phenotype of interest. This could well be due to redundancy in function among
members of these sub-families. When one gene is inactivated because of the T-DNA
insertion another family member might take over its function resulting in the wild-type
phenotype. For example, SOMATIC EMBRYOGENESIS RECEPTOR KINASE1
(SERK1) and the closest homolog SERK2 were concluded to function redundantly in
controlling anther development and male gametophyte maturation based on the
observation that the single mutants serk1 and serk2 did not show any developmental
defects whereas serk1serk2 double mutants were defective in pollen development
(Colcombet et al., 2005). This implies that such screenings, including ours, are by
definition not conclusive for all members of the family; the ones of which mutants
don’t show a phenotype could still have an important role. Another issue to keep in
mind is the variation in knock-out efficiency among mutants with a T-DNA insertion in
the same gene. According to a survey on 1084 published Arabidopsis insertion lines,
an insertion in the coding region leads to inactivation of the target gene in over 98%
of the cases; in mutants with an insertion before the start codon or after the stop
codon, this percentage decreases but is still high (86% and 75%, respectively)
(Wang, 2008). Variation in knock-out efficiency might explain why in some cases we
found differences in phenotype among T-DNA insertion lines of the same gene like
for example, the three LecRK-I.3 lines. In one case where we analysed expression of
the target gene, i.e. LecRK-IV.2, we indeed detected residual expression in a mutant
that contains a T-DNA insertion in the 5’UTR, and no change in phenotype, whereas
a line with insertions in the exon of that same gene showed a phenotype
(Supplemental Fig. S1; Wan et al., 2008). In the case of LecRK-V.2 we even found
that a T-DNA insertion in the 3’UTR led to increased transcript levels of the target
gene (Supplemental Fig. S2), a phenomenon that was also noted in the survey of
Wang (2008). It is thus not surprising that we could not confirm the phenotype of a
LecRK-V.2 mutant observed by Deng et al. (2009). Since we have not systematically
checked all T-DNA insertion lines for expression there is a chance that some of them
are not true mutants in the sense that they lost gene function. This implies that
LecRKs with T-DNA insertion lines showing no phenotype might still be relevant for
pathogen resistance.
In the unbiased strategy that we used here successful identification of mutants
is highly dependent on the design and the methodology of the phenotypic screening.
In our screening none of the LecRK T-DNA insertion lines showed phenotypic
changes upon exposure to mannitol or NaCl despite the fact that multiple LecRKs are
induced during abiotic stress. It could well be that the conditions that we used for

Chapter III
82
screening were not optimal. We only tested one concentration of mannitol or one of
NaCl instead of a whole range of concentrations. When screening for disease
resistance the choice of pathogen isolates is extremely important. In the large scale
screening performed by Wang et al. (2008), none of the four RLP30 T-DNA insertion
lines showed altered susceptibility towards the necrotrophic fungal pathogens
Sclerotinia sclerotiorum or Botrytis cinerea. Recently, however, it was found that two
of these RLP30 T-DNA insertion lines showed increased susceptibility to S.
sclerotiorum and B. cinerea which is likely due to the fact that Zhang et al. (2013)
used other isolates that are more virulent. In our screening we choose to use
different Phytophthora species and different isolates next to each other. The P.
capsici and P. brassicae isolates were selected based on their differential behavior
on Arabidopsis (Roetschi et al., 2001; Wang et al., 2013). We confirmed the role of
LecRK-I.9 in resistance to P. brassicae and showed that this LecRK also inhibits P.
capsici. In addition, we found several T-DNA insertion lines representing 12 LecRKs
with the same phenotype as lecrk-I.9-1 and lecrk-I.9-2, i.e. compromised resistance
to both Phytophthora spp. For four of these LecRKs (i.e. LecRK-IV.4, LecRK-VIII.2,
LecRK-IX.1 and LecRK-S.4) two or more independent T-DNA insertion lines showed
consistent gain of susceptibility demonstrating that they function as Phytophthora
resistance component. For another one, i.e. LecRK-V.5, we confirmed the mutant
phenotype by showing that overexpression of LecRK-V.5 enhances Phytophthora
resistance. For the remaining ones we have to be more cautious; in the phenotypic
analysis only a single T-DNA insertion line was included, the only one available or, in
the case of LecRK-S.7, only one of the two lines was a reliable mutant. Therefore, for
these LecRKs only a potential role in resistance against Phytophthora pathogens can
be suggested. With respect to P. brassicae and P. capsici, LecRK T-DNA insertion
lines showed no specificity suggesting that LecRKs do not discriminate between
Phytophthora species. Overall the T-DNA insertion lines are more susceptible to
P. brassicae HH than to P. capsici LT123 and less susceptible to P. brassicae
CBS686.95 than to P. capsici LT263. None of the LecRK T-DNA insertion lines was
infected by the less virulent isolate P. capsici LT62 and P. parasitica Pp009. These
two isolates are probably too weak to reach the stage where LecRKs come into play
as barrier for infection.
Four of the LecRKs that have a function in Phytophthora resistance were also
found to have a role in the interaction of Arabidopsis with the bacterial pathogen
Pst DC3000. Three of these promote resistance suggesting that they can monitor
different types of pathogens and potentially confer a broad-spectrum-resistance. The
fourth one however, LecRK-V.5, was found to have opposite roles in these two
pathosystems, i.e. a resistance component against Phytophthora pathogens but a
susceptibility factor for Pst DC3000. One explanation for this dual role is that

Function of Arabidopsis LecRKs in resistance
83
Phytophthora and bacteria have different modes to enter host tissues. Bacteria
usually penetrate via natural opening such as stomata, and stomatal closure can
essentially restrict bacterial invasion (Melotto et al., 2006). Phytophthora pathogens
can also penetrate via stomata but instead, they prefer to penetrate at the junctions
between epidermal cells (Hardham, 2007; Wang et al., 2011). Therefore, the function
of LecRK-V.5 in suppressing stomatal immunity is dispensable for preventing
Phytophthora penetration. In addition, LecRK-V.5 was found negatively regulating the
ABA response in Arabidopsis (Desclos-Theveniau et al., 2012). ABA plays a
multifaceted role in regulating plant resistance (Ton et al., 2009). On the one hand,
ABA enhances resistance against bacterial pathogens by regulating stomatal closure;
on the other hand, ABA compromises defence to various pathogens via suppression
of salicylic acid-signaling (Cao et al., 2011; Mohr and Cahill, 2007; Sanchez-Vallet et
al., 2012), a process that in Arabidopsis is essential for resistance to P. capsici
(Wang et al., 2013). Hence, the gain of susceptibility found in lecrk-V.5-3 upon
Phytophthora infection might be linked to ABA signaling.
Previously LecRK-VI.2 was found to be required for resistance to various
bacterial pathogens but not to the fungal pathogen Botrytis cinerea (Singh et al.,
2013; Huang et al., 2014). In our study, we confirmed the role of LecRK-VI.2 in
bacterial resistance and found that it is neither required for resistance to
Phytophthora pathogens nor the fungal pathogen A. brassicicola. This specificity
could be explained by the recent finding that LecRK-VI.2 associates with FLS2 and
participates in flg22-mediated defence (Huang et al., 2014), which is essential for
bacterial pathogen resistance but might be dispensable for resistance to
Phytophthora or fungal pathogens.
Considering the fact that the primary sequence of a protein is a prime
determinant of biological function, LecRKs with high sequence similarity might share
similar biological function. This might be the case for the four LecRKs in clade I of the
T-DNA insertion lines showed compromised resistance to A. brassicicola. This
clustering was not observed for LecRKs involved in Phytophthora or Pst DC3000
resistance. They are dispersed over different clades and thus sequence similarity as
such does not account for a similar function. Moreover, the computed expression
profiling revealed that LecRKs with similar function displayed rather different
expression patterns upon pathogen infection and therefore, expression is also not a
determinant of the functional similarity among LecRKs.
In summary, phenotypic analysis of a large set of T-DNA insertion lines
uncovered multiple Arabidopsis LecRKs with so far unknown function that contribute
to defence against pathogens, thus pointing to LecRKs as potential immune
receptors. The incentive for this screening was the finding that LecRK-I.9 is a
Phytophthora resistance component and, as importantly, that transgenic potato lines

Chapter III
84
expressing Arabidopsis LecRK-I.9 are more tolerant to P. infestans, the causal agent
of potato late blight (Bouwmeester et al., 2014). This suggests that LecRKs can be
exploited to confer resistance in crops. Since LecRKs are widespread in the plant
kingdom, understanding the function of LecRKs in Arabidopsis will facilitate the
functional analysis of LecRKs in crop species and this will be helpful to explore novel
resistance sources against damaging plant pathogens. Further studies should reveal
how the different LecRKs are activated, what their ligands are, and how they feed
into downstream signaling and activate defence. The overall importance of LecRKs in
plants is demonstrated by the recent finding that LecRK-I.9 (alias DORN1) is capable
to bind extracellular ATP (Choi et al., 2014). It is the first eATP receptor identified in
plants and has a completely different structure than eATP receptors in animals. The
collection of LecRK T-DNA insertion lines described in this study will be instrumental
to further explore the role of LecRKs as eATP receptors.
MATERIALS AND METHODS Analysis of T-DNA insertion lines
LecRK T-DNA insertion lines were obtained from the European Arabidopsis Stock
Center (http://arabidopsis.info; Alonso et al., 2003). T-DNA insertion sites and
homozygosity were checked by PCR using T-DNA primers LBb1.3 (SALK lines), LB1
(SAIL lines) or p745 (WiscDsLox lines) in combination with gene-specific primers
(Supplemental Table S2).
Plant growth conditions
Arabidopsis seeds were sown on soil or in vitro on 0.5 MS medium, stratified in the
dark at 4°C for three days and subsequently placed in a growth chamber at 19-21°C
with 75-78% relative humidity and a 12 h photoperiod. Alternatively, seeds were
grown on vertically orientated 0.5 MS plates or 0.5 MS plates supplemented with 100
mM NaCl or 200 mM mannitol or under the same conditions as described previously
(Deng et al., 2009). Seedling growth was evaluated from the 7th-to-16th day after
seed-sowing. Each treatment included at least three replicates and the experiment
was repeated four times.
Pathogen growth and infection assays
P. brassciae, P. capsici and P. parasitica were grown in the dark on 20% (v/v) V8
agar plates at appropriate temperatures. Infection assays on Arabidopsis with fresh
mycelial plugs were carried out as described previously (Bouwmeester et al., 2011).
Disease symptoms scoring using a disease severity index (DSI) and the

Function of Arabidopsis LecRKs in resistance
85
Phytophthora biomass quantification were conducted as described by (Wang et al.,
2013).
Pst DC3000 was grown on King's B medium supplemented with 50 µg ml-1
rifampicin. Arabidopsis plants were spray-inoculated with bacterial suspensions with
a concentration of 108 colony forming units (cfu) ml-1 in 10 mM MgSO4, 0.02% (v/v)
Silwet-L77, or mock-treated with 10 mM MgSO4, 0.02% (v/v) Silwet-L77. Bacterial
growth was determined at 3 dpi. Whole rosettes were collected and measured for
fresh weight and subsequently homogenized in 10 mM MgSO4. Homogenates were
serially diluted in 10 mM MgSO4 and were plated on Pseudomonas agar (BD)
containing 50 µg ml-1 rifampicin. Bacterial growth was evaluated by counting bacterial
cfu two days after incubation at 28°C and normalized as cfu/g using the total weight
of the inoculated rosette leaves. This experiment was repeated three times, each
with at least three replicates.
A. brassicicola MUCL20297 was cultured in dark at 22°C on potato dextrose
agar. Four to five-week-old Arabidopsis rosette leaves were drop-inoculated with 5 µl
of spore suspensions (106 spores ml-1) or mock-inoculated with 5 µl water. Plants
were placed in trays covered with lids and kept in the dark during the first 24 h.
Thereafter, they were moved to a climate chamber with aforementioned settings.
Disease symptoms were checked from 3 to 5 dpi.
Plasmid construction and plant transformation
The coding sequence of LecRK-V.5 was amplified by PCR using Pfu DNA
polymerase (Promega) with primers listed in Supplemental Table S2. Purified PCR
products were cloned into the vector pENTR™/D-TOPO® (Invitrogen). Plasmids with
correct sequence were recombined by LR reaction into the binary vector pGWB2.
The resulting vector was transformed into the Agrobacterium tumefaciens GV3101.
Arabidopsis Col-0 plants were transformed using the floral dip method as previously
described (Zhang et al., 2006).
RNA isolation and qRT-PCR
Total RNA was isolated from four-week-old Arabidopsis leaves with a NucleoSpin
RNA plant Kit (Macherey-Nagel) and thereafter used as the template for cDNA
synthesis with an oligo-dT primer using a M-MLV reverse transcriptase kit (Promega)
(Supplemental Table S2). qRT-PCR was performed using an ABI7300 Real-Time
PCR Systems (Applied Biosystems). Data were analysed using a 2−∆∆CT method
(Livak and Schmittgen, 2001). Transcript levels of LecRKs were normalized with
Arabidopsis Actin2 and expressed as mean fold changes (± SD) relative to that in
Col-0 leaves which was arbitrarily set as 1.

Chapter III
86
Expression data analyses
LecRK expression data during plant growth or upon treatment with different stimuli
were obtained from the public microarray datasets using eFP-Browser at the Bio-
Array Resource (http://www.bar.utoronto.ca) (Winter et al., 2007). Signal intensity
log2 ratios of the samples relative to respective controls were used as a measure for
LecRK expression. Of the datasets sampled from Arabidopsis in different growth
stages and exposed to abiotic stresses, the maximal (highest or lowest) signal
intensity relative to the mean throughout all arrays in the database was retrieved.
Expression data of Arabidopsis treated with biotic stresses were derived from five-
week-old Arabidopsis Col-0 rosette leaves inoculated with Phytophthora infestans or
infiltrated with Pst DC3000. LecRK-V.7 (At3g59740) and LecRK-V.8 (At3g59750)
share the same probe-set and obtained the same values. For expression pattern
analysis, per gene log2 values were normalized by the Z-score method according to
Cheadle et al. (2003). Co-expression analysis of LecRKs was conducted using
hierarchical clustering with Pearson’s correlation coefficient (r) and average linkage
(da) as implemented in MultiExperiment Viewer (MeV4.0).
ACKNOWLEDGEMENTS We thank Laurent Zimmerli (National Taiwan University) for providing LecRK-VI.2
mutants, Michael F. Seidl (Wageningen University) for discussion and Kolawole
Oluwaseun (University of Ibadan) for technical assistance. This project was
financially supported by a Wageningen University sandwich-PhD fellowship (YW), a
Huygens scholarship (YW), a VENI grant (KB) from the Netherlands Organization for
Scientific Research (Technology Foundation NWO-STW) and the Food-for-Thought
campaign of the Wageningen University Fund.
REFERENCES
Alonso, J.M., Stepanova, A.N., Leisse, T.J., Kim, C.J., Chen, H., Shinn, P., Stevenson, D.K.,
Zimmerman, J., Barajas, P., Cheuk, R., et al. (2003). Genome-wide insertional mutagenesis of
Arabidopsis thaliana. Science, 301, 653-657.
Aoki, K., Ogata, Y. and Shibata, D. (2007). Approaches for extracting practical information from gene
co-expression networks in plant biology. Plant & Cell Physiology, 48,381-390.
Boller, T. and Felix, G. (2009). A renaissance of elicitors: Perception of microbe-associated molecular
patterns and danger signals by pattern-recognition receptors. Annual Review of Plant Biology, 60,
379-406.
Bouwmeester, K. and Govers, F. (2009). Arabidopsis L-type lectin receptor kinases: phylogeny,
classification, and expression profiles. Journal of Experimental Botany, 60, 4383-4396.

Function of Arabidopsis LecRKs in resistance
87
Bouwmeester, K., de Sain, M., Weide, R., Gouget, A., Klamer, S., Canut, H. and Govers, F. (2011).
The lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a potential
host target for a RXLR effector. PLoS Pathogens, 7, e1001327.
Bouwmeester, K., Han, M., Blanco-Portales, R., Song, W., Weide, R., Guo, L., van der Vossen, E.A.G.
and Govers, F. (2014). The Arabidopsis lectin receptor kinase LecRK-I.9 enhances resistance to
Phytophthora infestans in Solanaceous plants. Plant Biotechnology Journal, 12, 10-16.
Cao, F.Y., Yoshioka, K. and Desveaux, D. (2011). The roles of ABA in plant-pathogen interactions.
Journal of Plant Research, 124, 489-499.
Cheadle, C., Vawter, M.P., Freed, W.J. and Becker, K.G. (2003). Analysis of microarray data using Z
score transformation. Journal of Molecular Diagnostics, 5, 73-81.
Chinchilla, D., Shan, L., He, P., de Vries, S. and Kemmerling, B. (2009). One for all: the receptor-
associated kinase BAK1. Trends in Plant Science, 14, 535-541.
Choi, J., Tanaka, K., Cao, Y., Qi, Y., Qiu, J., Liang, Y., Lee, S.Y. and Stacey, G. (2014). Identification
of a plant receptor for extracellular ATP. Science, 343, 290-294.
Colcombet, J., Boisson-Dernier, A., Ros-Palau, R., Vera, C.E. and Schroeder, J.I. (2005). Arabidopsis
SOMATIC EMBRYOGENESIS RECEPTOR KINASES1 and 2 are essential for tapetum
development and microspore maturation. Plant Cell, 17, 3350-3361.
Deng, K., Wang, Q., Zeng, J., Guo, X., Zhao, X., Tang, D. and Liu, X. (2009). A lectin receptor kinase
positively regulates ABA response during seed germination and is involved in salt and osmotic
stress response. Journal of Plant Biology, 52, 493-500.
Desclos-Theveniau, M., Arnaud, D., Huang, T.Y., Lin, G.J.C., Chen, W.Y., Lin, Y.C. and Zimmerli, L.
(2012). The Arabidopsis lectin receptor kinase LecRK-V.5 represses stomatal immunity induced by
Pseudomonas syringae pv. tomato DC3000. PLoS Pathogens, 8, e1002513.
Dievart, A., Dalal, M., Tax, F.E., Lacey, A.D., Huttly, A., Li, J.M. and Clark, S.E. (2003). CLAVATA1
dominant-negative alleles reveal functional overlap between multiple receptor kinases that regulate
meristem and organ development. Plant Cell, 15, 1198-1211.
Eisen, M.B., Spellman, P.T., Brown, P.O. and Botstein, D. (1998). Cluster analysis and display of
genome-wide expression patterns. Proceedings of the National Academy of Sciences of the United
States of America, 95, 14863-14868.
Fukushima, A., Nishizawa, T., Hayakumo, M., Hikosaka, S., Saito, K., Goto, E. and Kusano, M. (2012).
Exploring tomato gene functions based on coexpression modules using graph clustering and
differential coexpression approaches. Plant Physiology, 158, 1487-1502.
Gómez-Gómez, L. and Boller, T. (2000). FLS2: An LRR receptor–like kinase involved in the perception
of the bacterial elicitor flagellin in Arabidopsis. Molecular Cell, 5, 1003-1011.
Gouget, A., Senchou, V., Govers, F., Sanson, A., Barre, A., Rouge, P., Pont-Lezica, R.P. and Canut, H.
(2006). Lectin receptor kinases participate in protein-protein interactions to mediate plasma
membrane-cell wall adhesions in Arabidopsis. Plant Physiology, 140, 81-90.
Hardham, A.R. (2007). Cell biology of plant-oomycete interactions. Cellular Microbiology, 9, 31-39.
Hervé, C., Dabos, P., Galaud, J.P., Rougé, P. and Lescure, B. (1996). Characterization of an
Arabidopsis thaliana gene that defines a new class of putative plant receptor kinases with an
extracellular lectin-like domain. Journal of Molecular Biology, 258, 778-788.
Huang, P., Ju, H.W., Min, J.H., Zhang, X., Kim, S.H., Yang, K.Y. and Kim, C.S. (2013). Overexpression
of L-type lectin-like protein kinase 1 confers pathogen resistance and regulates salinity response in
Arabidopsis thaliana. Plant Science, 203, 98-106.

Chapter III
88
Huang, P.Y., Yeh, Y.H., Liu, A.C., Chien, C.P., Zimmerli, L. (2014). The Arabidopsis LecRK-VI.2
associates with the pattern-recognition receptor FLS2 and primes Nicotiana benthamiana pattern-
triggered immunity. The Plant Journal, 79, 243–255.
Huitema, E., Vleeshouwers, V.G.A.A., Francis, D.M. and Kamoun, S. (2003). Active defence
responses associated with non-host resistance of Arabidopsis thaliana to the oomycete pathogen
Phytophthora infestans. Molecular Plant Pathology, 4, 487-500.
Li, X.L., Tan, Y.C. and Ng, S.K. (2006). Systematic gene function prediction from gene expression data
by using a fuzzy nearest-cluster method. BMC Bioinformatics, 7, S23.
Liebrand, T.W.H., van den Burg, H.A. and Joosten, M.H.A.J. (2013). Two for all: receptor-associated
kinases SOBIR1 and BAK1. Trends in Plant Science, 19, 123-132.
Livak, K.J. and Schmittgen, T.D. (2001). Analysis of relative gene expression data using real-time
quantitative PCR and the 2−∆∆CT method. Methods, 25, 402-408.
Lloyd J. and Meinke D. (2012). A comprehensive dataset of genes with a loss-of-function mutant
phenotype in Arabidopsis. Plant Physiology, 158, 1115-1129.
Marone, D., Russo, M.A., Laido, G., De Leonardis, A.M. and Mastrangelo, A.M. (2013). Plant
nucleotide binding site-leucine-rich repeat (NBS-LRR) genes: active guardians in host defense
responses. International Journal of Molecular Sciences, 14, 7302-7326.
Melotto, M., Underwood, W., Koczan, J., Nomura, K. and He, S.Y. (2006). Plant stomata function in
innate immunity against bacterial invasion. Cell, 126, 969-980.
Mohr, P. and Cahill, D. (2007). Suppression by ABA of salicylic acid and lignin accumulation and the
expression of multiple genes, in Arabidopsis infected with Pseudomonas syringae pv. tomato.
Functional & Integrative Genomics, 7, 181-191.
O’Malley R.C. and Ecker J.R. (2010). Linking genotype to phenotype using the Arabidopsis unimutant
collection. The Plant Journal, 61, 928-940.
Rodewald, J. and Trognitz, B. (2013). Solanum resistance genes against Phytophthora infestans and
their corresponding avirulence genes. Molecular Plant Pathology, 14, 740-757.
Roetschi, A., Si-Ammour, A., Belbahri, L., Mauch, F. and Mauch-Mani, B. (2001). Characterization of
an Arabidopsis-Phytophthora pathosystem: Resistance requires a functional PAD2 gene and is
independent of salicylic acid, ethylene and jasmonic acid signalling. The Plant Journal, 28, 293-305.
Sanchez-Vallet, A., Lopez, G., Ramos, B., Delgado-Cerezo, M., Riviere, M.P., Llorente, F., Fernandez,
P.V., Miedes, E., Estevez, J.M., Grant, M. and Molina, A. (2012). Disruption of abscisic acid
signaling constitutively activates Arabidopsis resistance to the necrotrophic fungus
Plectosphaerella cucumerina. Plant Physiology, 160, 2109-2124.
Shiu, S.H. and Bleecker, A.B. (2003). Expansion of the receptor-like kinase/Pelle gene family and
receptor-like proteins in Arabidopsis. Plant Physiology, 132, 530-543.
Singh, P., Kuo, Y.C., Mishra, S., Tsai, C.H., Chien, C.C., Chen, C.W., Desclos-Theveniau, M., Chu,
P.W., Schulze, B., Chinchilla, D., Boller, T. and Zimmerli, L. (2012). The lectin receptor kinase-VI.2
is required for priming and positively regulates Arabidopsis pattern-triggered immunity. Plant Cell,
24, 1256-1270.
Singh, P., Chien, C.C., Mishra, S., Tsai, C.H., Zimmerli, L. (2013). The Arabidopsis LECTIN
RECEPTOR KINASE-VI.2 is a functional protein kinase and is dispensable for basal resistance to
Botrytis cinerea. Plant Signaling & Behavior, 8,1-3.
Spoel, S.H. and Dong, X. (2012). How do plants achieve immunity? Defence without specialized
immune cells. Nature Reviews Immunology, 12, 89-100.

Function of Arabidopsis LecRKs in resistance
89
ten Hove, C.A., Bochdanovits, Z., Jansweijer, V.M.A., Koning, F.G., Berke, L., Sanchez-Perez, G.F.,
Scheres, B. and Heidstra, R. (2011). Probing the roles of LRR-RLK genes in Arabidopsis thaliana
roots using a custom T-DNA insertion set. Plant Molecular Biology, 76, 69-83.
Ton, J., Flors, V. and Mauch-Mani, B. (2009). The multifaceted role of ABA in disease resistance.
Trends in Plant Science, 14, 310-317.
Usadel, B., Obayashi, T., Mutwil, M., Giorgi, F.M., Bassel, G.W., Tanimoto, M., Chow, A., Steinhauser,
D., Persson, S. and Provart, N.J. (2009). Co-expression tools for plant biology: opportunities for
hypothesis generation and caveats. Plant, Cell & Environment, 32, 1633-1651.
van Wees, S.C.M., Chang, H.S., Zhu, T. and Glazebrook, J. (2003). Characterization of the early
response of Arabidopsis to Alternaria brassicicola infection using expression profiling. Plant
Physiology, 132, 606-617.
Wan, J., Patel, A., Mathieu, M., Kim, S.Y., Xu, D. and Stacey, G. (2008). A lectin receptor-like kinase is
required for pollen development in Arabidopsis. Plant Molecular Biology, 67, 469-482.
Wang, G., Ellendorff, U., Kemp, B., Mansfield, J.W., Forsyth, A., Mitchell, K., Bastas, K., Liu, C.M.,
Woods-Toer, A., Zipfel, C., et al. (2008). A genome-wide functional investigation into the roles of
receptor-like proteins in Arabidopsis. Plant Physiology, 147, 503-517.
Wang, Y. (2008). How effective is T-DNA insertional mutagenesis in Arabidopsis? Journal of
Biochemical Technology, 1, 11-20.
Wang, Y., Meng, Y., Zhang, M., Tong, X., Wang, Q., Sun, Y., Quan, J., Govers, F. and Shan, W.
(2011). Infection of Arabidopsis thaliana by Phytophthora parasitica and identification of variation in
host specificity. Molecular Plant Pathology, 12, 187-201.
Wang, Y., Bouwmeester, K., Van de Mortel, J.E., Shan, W. and Govers, F. (2013). A novel
Arabidopsis-oomycete pathosystem: differential interactions with Phytophthora capsici reveal a role
for camalexin, indole glucosinolates and salicylic acid in defence. Plant, Cell & Environment, 36,
1192-1203.
Winter, D., Vinegar, B., Nahal, H., Ammar, R., Wilson, G.V. and Provart, N.J. (2007). An "Electronic
Fluorescent Pictograph" browser for exploring and analyzing large-scale biological data sets. PLoS
One, 2, e718.
Xin, Z., Wang, A., Yang, G., Gao, P. and Zheng, Z.L. (2009). The Arabidopsis A4 subfamily of lectin
receptor kinases negatively regulates abscisic acid response in seed germination. Plant Physiology,
149, 434-444.
Zhang, W., Fraiture, M., Kolb, D., Löffelhardt, B., Desaki, Y., Boutrot, F.F.G., Tör, M., Zipfel, C., Gust,
A.A. and Brunner, F. (2013). Arabidopsis RECEPTOR-LIKE PROTEIN30 and receptor-like kinase
SUPPRESSOR OF BIR1-1/EVERSHED mediate innate immunity to necrotrophic fungi. Plant Cell,
25, 4227-4241.
Zhang, X., Henriques, R., Lin, S.S., Niu, Q.W. and Chua, N.H. (2006). Agrobacterium-mediated
transformation of Arabidopsis thaliana using the floral dip method. Nature Protocols, 1, 641-646.

Chapter III
90
SUPPLEMENTAL INFORMATION Table S1. Summary of the available LecRK T-DNA insertion lines.
a No T-DNA insertion lines available. b SALK_053703C, SALK_019496C are listed in the NASC database as mutants with insertions in At2g37710, but in our analysis this
could not be confirmed. c The obtained SAIL_529_D07 could not germinate. d Mutant in Ler-0 background. e T-DNA insertion line show increased LecRK mRNA levels.

Function of Arabidopsis LecRKs in resistance
91
Table S2. Primers used in this study.
HM: homozygosity test. RT: primers used for q-RT-PCR. FL: primers for full length LecRK-V.5 cloning.

Chapter III
92
Figure S1. SALK_119402C with a T-DNA insertion in the 5’UTR of LecRK-IV.2 does not show
defects in growth and development.
A, Schematic model representing LecRK-IV.2. The black arrowhead points to the T-DNA insertion site
in lecrk-IV.2 (i.e. SALK_119402C) and the head-to-head arrow pairs indicate the position of the
LecRK-IV.2 specific primers (IV.2-RT) used for qRT-PCR analysis. B, Relative quantification of
LecRK-IV.2 transcript levels in Col-0 and lecrk-IV.2. Transcript levels were normalized with
Arabidopsis Actin2 and expressed as mean fold changes (± SD) relative to the transcript level in Col-0
leaves which was arbitrarily set as 1. C, Silique morphology of seven-week-old lecrk-IV.2 and Col-0
plants.

Function of Arabidopsis LecRKs in resistance
93
Figure S2. SALK_014678C with a T-DNA insertion in the 3’UTR of LecRK-V.2 does not show an
altered response to NaCl and mannitol.
A, Schematic model representing LecRK-V.2. The black arrowhead points to the T-DNA insertion site
in lecrk-V.2 (i.e. SALK_014678C) and the head-to-head arrow pairs indicate the position of the LecRK-
V.2 specific primers (V.2-RT) used for qRT-PCR analysis. B, 16-day-old seedlings of Col-0 and lecrk-
V.2 grown on 0.5 MS plate or on 0.5 MS supplemented with either 100 mM NaCl or 200 mM mannitol.
C, Relative quantification of LecRK-V.2 transcript levels in Col-0 and lecrk-V.2. Transcript levels were
normalized with Arabidopsis Actin2 and expressed as mean fold changes (± SD) relative to the
transcript level in Col-0 leaves which was arbitrarily set as 1.

Chapter III
94
Figure S3. Disease symptoms on LecRK-I.9 mutants upon inoculation with Phytophthora
pathogens.
A-B, Disease symptoms on lecrk-I.9-1 and lecrk-I.9-2 four days after plug-inoculation with
P. brassicae HH (A) and P. capsici LT123 (B). White arrowheads point to inoculated leaves.

Function of Arabidopsis LecRKs in resistance
95
Figure S4. Expression analysis of LecRK-S.7 in the T-DNA insertion lines lecrk-S.7-1 and lecrk-
S.7-2.
A, Schematic model representing LecRK-S.7. Black arrowheads point to the T-DNA insertion sites in
lecrk-S.7-1 (i.e. SALK_140480C) and lecrk-S.7-2 (SALK_008479C) and the head-to-head arrow pairs
indicate the position of the LecRK-S.7 specific primers (V.7-RT) used for qRT-PCR analysis. B,
Relative quantification of LecRK-S.7 transcript levels in Col-0, lecrk-S.7-1 and lecrk-S.7-2. Transcript
levels were normalized with Arabidopsis Actin2 and expressed as mean fold changes (± SD) relative
to the transcript level in Col-0 leaves which was arbitrarily set as 1.


Chapter IV
Phytophthora resistance conferred by Arabidopsis
L-type lectin receptor kinases LecRK-IX.1 and
LecRK-IX.2 is not associated with activation of
plant cell death
Yan Wang, Klaas Bouwmeester, Jan H. G. Cordewenerⱡ, Antoine H.P. Americaⱡ,
Weixing Shan♯, Francine Govers
In collaboration with: ⱡ Plant Research International, Wageningen UR, Wageningen, The Netherlands. ♯ College of Plant Protection, Northwest A&F University, Yangling, Shaanxi, China

Chapter IV
98
ABSTRACT
Plasma membrane-localized L-type lectin receptor kinases (LecRKs) are considered
as potential immune receptors. In this study, we characterized the two closely-related
Arabidopsis LecRKs, i.e. LecRK-IX.1 and LecRK-IX.2. For both LecRKs, T-DNA
insertion mutants showed compromised Phytophthora resistance as well as reduced
defence-related gene expression. In comparison, the double mutants showed additive
susceptibility. Overexpression of each of the two LecRKs in Arabidopsis increased
resistance to Phytophthora pathogens but also induced cell death. For their function
as Phytophthora resistance component, both the lectin domain and kinase activity are
required, while the ability to induce cell death is determined by the kinase activity but
not the lectin domain. Transient expression of both Arabidopsis LecRKs in
N. benthamiana induces cell death, with LecRK-IX.2 showing a stronger capacity to
induce cell death than LecRK-IX.1. Silencing of NbSIPK/NTF4 in N. benthamiana
completely abolished LecRK-IX.1-induced cell death but not Phytophthora resistance.
In planta co-immunoprecipitation with LecRK-IX.1 and LecRK-IX.2 as baits followed
by LC-MS analysis resulted in the identification of the N. benthamiana ABC
transporter NbPDR1 that interacts with both LecRK-IX.1 and LecRK-IX.2. Also its
closest Arabidopsis homolog ABCG40 interacts with LecRK-IX.1 and LecRK-IX.2 in
planta. In both mutants lecrk-IX.1 and lecrk-IX.2, expression of ABCG40 is reduced
upon Phytophthora infection. Similar to the LecRK mutants, ABCG40 mutants showed
compromised Phytophthora disease resistance. This study demonstrates that
LecRK-IX.1 and LecRK-IX.2 function as Phytophthora resistance components. Both
as well induce cell death when overexpressed, but the Phytophthora resistance
function is independent of cell death. Since both LecRK-IX.1 and LecRK-IX.2 interact
with the same ABC transporters, this suggests that they share similar signaling
components. Nevertheless, they were found to act independently to confer resistance
to Phytophthora pathogens.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
99
INTRODUCTION
Plants are constantly threatened by microbes and have evolved strategies to detect
microbial infection via various cell surface and intracellular receptors (Spoel and Dong,
2012). Defence responses initiated by pathogen perception through cell surface
pattern recognition receptors (PRRs) provide the first line of innate immunity and
collectively suppress the majority of invading microbes (Boller and Felix, 2009). PRRs
rapidly form complexes via association with other components to initiate signal
transduction cascades upon recognition of conserved microbial molecules, the so
called microbe-associated molecular patterns (MAMPs), or endogenous elicitors
released from plants, known as damage-associated molecular patterns (DAMPs)
(Sun et al., 2013; Liebrand et al., 2014; Macho and Zipfel, 2014). As a result, various
cellular responses are initiated, including production of reactive oxygen species
(ROS), activation of mitogen-activated protein kinase (MAPK) cascades, secretion of
antimicrobial compounds and induction of defence-related gene expression (Boller
and Felix, 2009). In some cases, activation of cell surface receptors also leads to
plant cell death (Gao et al., 2009).
Plant genomes encode a large number of cell surface receptors with an
intracellular kinase domain. These so-called receptor-like kinases (RLKs) not only act
as PRRs to perceive pathogens, but also can function as signaling partners
participating in PRR-mediated downstream signaling (Chinchilla et al., 2009; Liebrand
et al., 2014). RLKs are classified based on their versatile extracellular domains, which
are considered to determine ligand perception (Shiu and Bleecker, 2001). L-type lectin
receptor kinases (LecRKs) are a group of RLKs with an extracellular legume-like
lectin domain (Herve et al., 1999; Bouwmeester and Govers, 2009). Previously, we
found that one of the LecRKs in Arabidopsis, i.e. LecRK-I.9 plays a crucial role in
resistance to Phytophthora brassicae and acts as a mediator of plant cell wall-plasma
membrane adhesion (Bouwmeester et al., 2011). More recently, the same LecRK was
identified as the first plant receptor for extracellular ATP (eATP) and found to be
required for eATP-mediated downstream signaling (Choi et al., 2014). LecRK-I.9 is a
member of a family consisting of 45 LecRKs in Arabidopsis. Based on sequence
similarity and phylogeny, these 45 LecRKs were divided into nine clades (clade I to
clade IX) and seven singletons (Bouwmeester and Govers, 2009). In recent years,
several Arabidopsis LecRKs other than LecRK-I.9 have been found to be implicated in
plant immunity. For example, LecRK-VI.2, which is required for priming BABA-induced
resistance, associates with the flagellin receptor FLS2 and positively regulates
MAMP-mediated plant defence (Singh et al., 2012; Huang et al., 2014). LecRK-V.5
was found to play a dual role in plant resistance against different pathogens. It
negatively regulates plant stomatal immunity against bacterial pathogens, but

Chapter IV
100
positively contributes to Phytophthora resistance (Desclos-Theveniau et al., 2012;
Wang et al., 2014/Chapter III). A systematic screen of Arabidopsis LecRK T-DNA
insertion lines identified several additional LecRKs that are involved in resistance
against either Alternaria brassicicola, Phytophthora or Pseudomonas pathogens
(Wang et al., 2014/Chapter III). Among these are LecRK-IX.1 and LecRK-IX.2, the
only two members in clade IX (Bouwmeester and Govers, 2009).
In this study, we performed more detailed analyses on the role of Arabidopsis
LecRK-IX.1 and LecRK-IX.2 in Phytophthora resistance. We compared the
susceptibility of the null mutants lecrk-IX.1 and lecrk-IX.2 with double mutants and
generated overexpression lines to confirm the role of LecRK-IX.1 and LecRK-IX.2 in
Phytophthora resistance. Subsequently, we assayed the response of transgenic
Arabidopsis plants expressing mutated versions of these two LecRKs and pinpointed
the domains required for resistance and cell death responses. In addition, we
transiently expressed the two LecRKs in the Solanaceous plant Nicotiana
benthamiana to monitor induced plant responses. Finally, by means of in planta
co-immunoprecipitation and LC-MS, we identified an interacting protein that seems to
be required for Phytophthora resistance.
RESULTS
Arabidopsis mutants lecrk-IX.1 and lecrk-IX.2 show compromised Phytophthora
resistance
Previously, LecRK-IX.1 and LecRK-IX.2 were identified as potential resistance
components based on the finding that the Arabidopsis mutants lecrk-IX.1 and
lecrk-IX.2 showed gain of susceptibility to the non-adapted isolates
Phytophthora brassicae HH and Phytophthora capsici LT123. In contrast, the
response to the fungal pathogen Alternaria brassicicola MUCL20297 or the bacterial
pathogen Pseudomonas syringae was not changed in these mutants (Wang et al.,
2014/Chapter III; Supplemental Fig. S1). In Arabidopsis, secondary metabolites such
as salicylic acid (SA), jasmonic acid (JA), ethylene (ET), camalexin and indole
glucosinolates (iGS) have been found to be required for resistance to Phytophthora
pathogens, and expression of marker genes indicative for the related pathways was
found to be induced upon inoculation with P. brassicae or P. capsici (Roetschi et al.,
2001; Schlaeppi et al., 2010; Wang et al., 2013a; Wang et al., 2013b). To determine
whether these pathways are implicated in resistance mediated by LecRK-IX.1 and
LecRK-IX.2, we compared the expression levels of these marker genes in lecrk-IX.1,
lecrk-IX.2 and Col-0 during infection by P. capsici LT123. As shown in Fig. 1A,
induction of all selected marker genes is less pronounced in lecrk-IX.1 and lecrk-IX.2

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
101
Figure 1. Mutants lecrk-IX.1 and lecrk-IX.2 show defects in defence against Phytophthora
pathogens.
A, Expression of defence-related genes in lecrk-IX.1, lecrk-IX.2 and Col-0 upon inoculation with
P. capsici LT123. Arabidopsis leaves plug-inoculated with P. capsici isolate LT123 were harvested at 12
and 24 hours post inoculation (hpi). Relative transcript levels were normalized to Arabidopsis Actin2.
Values are expressed as mean fold changes (± SD) relative to the transcript level in mock-inoculated
leaves. B, Schematic presentation of LecRK-IX.1 and LecRK-IX.2. Domains were predicted by SMART.
Black arrowheads point to the T-DNA insertion sites and primers used for semi-qRT-PCR are indicated.
TM, transmembrane domain; STK, kinase domain. C, Transcript levels of LecRK-IX.1 and LecRK-IX.2
in Col-0, lecrk-IX.1, lecrk-IX.2 and double mutants (i.e. lecrk-IX.1/2 and lecrkK-IX.2/1) determined by
semi-qRT-PCR. Actin2 was used as control. D, Disease symptoms observed on Col-0, lecrk-IX.1,
lecrk-IX.2 and double mutants four days post inoculation (dpi) with P. capsici LT123. White arrowheads
point to the inoculated leaves.

Chapter IV
102
when compared to Col-0, in particular for PR1, a SA marker gene and CYP71B15,
which is required for camalexin biosynthesis. Compared to lecrk-IX.1, overall lower
expression levels of PR1 and CYP71B15 were found in lecrk-IX.2. In addition, PDF1.2,
a JA/ET marker gene also showed a lower expression in lecrk-IX.2, whereas the
expression of CYP81F2, a marker gene for iGS was comparable in the two mutants.
To determine whether LecRK-IX.1 and LecRK-IX.2 are functionally redundant, two
homozygous double mutants, termed lecrk-IX.1/2 and lecrk-IX.2/1 were generated
with T-DNA insertions in both LecRK-IX.1 and LecRK-IX.2. Absence of full length
mRNA of both LecRK-IX.1 and LecRK-IX.2 was confirmed in the double mutants
(Fig.1C). Compared to Col-0 and the parental null mutants, the double mutants
showed no visible growth alterations or developmental defects. Upon inoculation with
P. capsici LT123, the double mutants showed increased susceptibility compared to the
single mutants (Fig. 1D). These results indicate that LecRK-IX.1 and LecRK-IX.2
function as independent Phytophthora resistance components.
Overexpression of LecRK-IX.1 or LecRK-IX.2 in Arabidopsis induces cell death
and Phytophthora resistance
To further confirm the role of LecRK-IX.1 and LecRK-IX.2 in Phytophthora resistance,
we generated Arabidopsis transgenic lines expressing LecRK-IX.1 or LecRK-IX.2
under control of the CaMV 35S promoter, named 35S-IX.1 and 35S-IX.2, respectively
(Fig. 2). During the first two weeks of growth, the 35S-IX.1 and 35S-IX.2 lines were
morphologically indistinguishable from the recipient Col-0 plants or Col-0 transformed
with a GFP construct (35S-GFP). Thereafter, part of the 35S-IX.1 and 35S-IX.2 lines
displayed spontaneous necrosis and retarded growth in comparison to Col-0 or
35S-GFP (Fig. 2C). When examined under the microscope, these plants showed
intensive staining with trypan blue indicative of cell death (Fig. 2C). When transgene
expression levels were monitored by qRT-PCR, it appeared that transgenic lines with
a cell death phenotype showed higher transgene expression levels than those without
(Fig. 2B). Apparently, the induced cell death correlates with transgene expression
levels.
To determine whether overexpression of LecRK-IX.1 or LecRK-IX.2 increases
Arabidopsis resistance to Phytophthora pathogens, transgenic lines with a similar
growth phenotype as Col-0 plants, i.e. showing neither macroscopically nor
microscopically visible cell death, were selected for infection assays with P. capsici
LT263. Col-0 and 35S-GFP lines developed severe disease symptoms three days
post inoculation (dpi). In comparison, the 35S-IX.1 and 35S-IX.2 lines showed less
severe disease symptoms and less P. capsici biomass (Fig. 3, A and D; Supplemental
Fig. S2). These lines also showed enhanced resistance to P. brassicae CBS686.95

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
103
Figure 2. Phenotypic changes of Arabidopsis transgenic lines expressing LecRK-IX.1,
LecRK-IX.2 or derivatives lacking the lectin domain or mutated in the kinase domain.
A, Schematic representation of LecRK-IX.1, LecRK-IX.2 and their derivatives used in this study.
B, Correlation of plant cell death phenotype and expression levels of LecRK-IX.1, LecRK-IX.2 or
derivatives in different transgenic lines. Relative transcript levels were normalized to Arabidopsis Actin2
and values are expressed as mean fold changes (± SD) relative to the transcript level in Col-0 plants
which was arbitrarily set as 1. ‘+’ / ‘-’ indicates transgenic lines with or without a cell death phenotype.
C, Growth of transgenic lines constitutively expressing GFP, LecRK-IX.1, LecRK-IX.2 or their
derivatives. Inset figures are microscopic images showing leaf pieces upon trypan blue staining. Scale
bars represent 200 µm.

Chapter IV
104
(Supplemental Fig. S3), demonstrating that expression of LecRK-IX.1 and LecRK-IX.2
can inhibit infection by different Phytophthora species. To determine whether
overexpression of LecRK-IX.1 and LecRK-IX.2 causes increased plant cell death
during P. capsici infection, inoculated leaves were stained with trypan blue and
investigated by microscopy two days after drop-inoculation with LT263 zoospores.
Preliminary data indicate that overexpression of LecRK-IX.1 or LecRK-IX.2 blocks the
early steps of infection given the fact that only a few penetration events were
observed on 35S-IX.1 and 35S-IX.2 lines. In the mesophyll cell layers of Col-0 and
35S-GFP-1, expanding hyphae with haustoria were observed at 2 dpi. In contrast,
only penetration pegs or short primary infection hyphae were observed in the leaves
of 35S-IX.1-1 and 35S-IX.2-1 (Fig. 3B). Nevertheless, there was no increase in cell
death upon P. capsici penetration in the 35S-IX.1-1 or 35S-IX.2-1 leaves when
compared to Col-0 (Fig. 3B).
Collectively, these results indicate that both LecRK-IX.1 and LecRK-IX.2 function
in plant cell death induction and Phytophthora resistance. The induced cell death,
however, is not a prerequisite for Phytophthora resistance.
Contribution of the lectin domain and kinase activity to LecRK-mediated cell
death and Phytophthora resistance in Arabidopsis
The extracellular domains and a functional kinase domain have been shown to be
essential for the biological function of several RLKs (Gomez-Gomez et al., 2001;
Schwessinger and Ronald, 2012). Hence, we investigated whether both domains are
required for LecRK-IX.1 and LecRK-IX.2 to mediate plant cell death and Phytophthora
resistance. Truncated constructs lacking the lectin domain (i.e. LecRK-IX.1-∆lectin
and LecRK-IX.2-∆lectin) were generated and transformed to Col-0 (Fig. 2A). Similar to
35S-IX.1 and 35S-IX.2 lines, plants expressing either LecRK-IX.1-∆lectin or
LecRK-IX.2-∆lectin (35S-IX.1-∆lectin and 35S-IX.2-∆lectin lines, respectively) varied
in cell death phenotype (Fig. 2C). Transgenic lines with relatively high transgene
expression levels showed cell death (Fig. 2, B and C). To assess the importance of
the kinase domain, kinase-dead mutants were generated by substitution of the
essential RD residues in the catalytic loop to RN or AA (Fig. 2A). In the transgenic
lines, − i.e. 35S-IX.1-RN/AA and 35S-IX.2-RN/AA, respectively − different transgene
expression levels were detected (Fig. 2B), but none of these lines displayed cell death
or any other apparent phenotypic change compared to Col-0 (Fig. 2, B and C). Overall,
these results indicate that the cell death phenotype triggered by overexpression of
LecRK-IX.1 or LecRK-IX.2 is dependent on the kinase activity, but not on the lectin
domain.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
105
Figure 3. Infection of P. capsici on Col-0 and transgenic lines expressing LecRK-IX.1,
LecRK-IX.2 or derivatives lacking the lectin domain or mutated in the kinase domain.
A, Disease symptoms on Col-0 and transgenic lines 35S-GFP, 35S-IX.1-1 and 35S-IX.2-1 three days
after plug-inoculation with P. capsici LT263. White arrowheads point to the inoculated leaves. B,
Microscopic investigation of P. capsici infection on Col-0 and transgenic lines 35S-GFP, 35S-IX.1-1 and

Chapter IV
106
35S-IX.2-1. Arabidopsis leaves inoculated with P. capsici LT263 zoospores (105 zoospores ml-1) were
harvested at 2 dpi and stained with trypan blue. White arrowheads point to the penetration sites. ha,
haustorium; hy, invasive hyphae. Scale bars represent 20 µm. C, Disease symptoms on Col-0 and
different transgenic lines at 3 dpi. White arrowheads point to the inoculated leaves. D, Relative
quantification of P. capsici biomass in Col-0 and different transgenic lines using qPCR. Bars represent
mean values (± SD) relative to that of infected Col-0 leaves. * indicates significant difference in
biomass (p < 0.05) according to a t test. Experiments were repeated twice with similar results.
Infection assays were performed on transgenic lines that are morphologically
undistinguishable from Col-0. These transgenic lines 35S-IX.1-∆lectin and
35S-IX.2-∆lectin showed similar disease symptoms as Col-0 or 35S-GFP upon
inoculation with P. capsici LT263 (Fig. 3C; Supplemental Fig. S2A). Similar
susceptibility was also observed on the transgenic lines expressing the kinase-dead
mutants (Fig. 3C; Supplemental Fig. S2). In line with these phenotypes, P. capsici
biomass was only reduced in 35S-IX.1 and 35S-IX.2 lines when compared to that in
Col-0, but not in the transgenic lines expressing lectin-deletion or kinase-dead
mutants (Fig. 3D; Supplemental Fig. S2B). Therefore, we conclude that both the lectin
domain and the kinase activity of LecRK-IX.1 and LecRK-IX.2 are essential to
mediate Phytophthora resistance.
Cell death induced by overexpression of LecRK-IX.1 and LecRK-IX.2 is not
correlated with leaf senescence
In silico gene expression analysis using publicly available microarray data indicated
that LecRK-IX.2 but not LecRK-IX.1 is highly induced in senescent Arabidopsis leaves
(Fig. 4A). This was verified by monitoring the transcript levels of LecRK-IX.1 and
LecRK-IX.2 by qRT-PCR in Col-0 leaves undergoing senescence (Fig. 4B).
LecRK-IX.1 showed no significant change in expression but LecRK-IX.2 expression
levels increased in leaves with more severe symptoms of senescence (Fig. 4B). This
expression pattern was comparable to that of SAG12, a gene encoding a cysteine
protease that has been widely used as a reliable senescence marker gene. SAG12 is
only expressed in senescing tissues but not in stress- or hypersensitive
response-related cell death (Noh and Amasino, 1999; Pontier et al., 1999; Zhou et al.,
2009). To determine whether cell death induced by overexpression of LecRK-IX.2 is
correlated with plant senescence, we compared the expression levels of SAG12 in
different LecRK-IX.2 transgenic lines with or without cell death phenotype (Fig. 4C).
No correlation was found between SAG12 expression and LecRK-IX.2 expression
levels or plant phenotypic changes (Fig. 2B and Fig. 4C). This also holds for the
LecRK-IX.1 transgenic lines (Fig. 2B and Fig. 4C). Apparently, the cell death
phenotype induced by overexpression of LecRK-IX.1 or LecRK-IX.2 is not linked to

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
107
plant senescence.
Figure 4. Cell death induced by overexpression of LecRK-IX.1 or LecRK-IX.2 is not correlated
with leaf senescence.
A, Expression profiles of LecRK-IX.1 and LecRK-IX.2 in different Arabidopsis tissues. Signal intensity
log2 ratios are color-coded in a range of -2 (blue) to 6 (yellow) as indicated in the scale bar. B,
Expression levels of LecRK-IX.1, LecRK-IX.2 and SAG12 in Col-0 leaves in different extent of
senescence quantified by qRT-PCR. Relative transcript levels were normalized to Arabidopsis Actin2
and values are expressed as mean fold changes (± SD) relative to that in Col-0 leaves at the
senescence stage 1 (S1) which was arbitrarily set as 1. C, Expression levels of SAG12 in Col-0 and the

Chapter IV
108
presented transgenic lines. Total RNA was isolated from the whole rosette and used for qRT-PCR.
Relative transcript levels were normalized to Arabidopsis Actin2 and values are expressed as mean
fold changes (± SD) relative to the transcript levels in Col-0 plants which was arbitrarily set as 1.
Transient expression of LecRK-IX.1 or LecRK-IX.2 induces H2O2 production and
cell death in N. benthamiana
To further confirm the role of LecRK-IX.1 and LecRK-IX.2 in Phytophthora resistance
and cell death induction, C-terminal GFP-tagged LecRK-IX.1 and LecRK-IX.2 were
expressed in N. benthamiana using Agrobacterium-mediated transient gene
expression and induced defence responses were analysed.
Accumulation of LecRK-IX.1-eGFP and LecRK-IX.2-eGFP was determined in the
agro-infiltrated N. benthamiana leaves. Total protein was isolated, immunopurified
using GFP-trap_A® beads and detected by western blot with anti-GFP. Both fusion
proteins could be detected, but the sizes were slightly larger than that of the predicted
monomers (Fig. 5A). Since both LecRKs contain putative N-glycosylation sites
(Supplemental Fig. S4), the discrepancy in size could be partially due to glycosylation.
This type of post-translational modification has been shown to be important for some
plant receptors in mediating immune responses (Haweker et al., 2010; Sun et al.,
2012). To test this hypothesis, we investigated whether the two LecRKs are
glycosylated using anti-horseradish peroxidase (HRP). Purified LecRK-IX.1-eGFP
and LecRK-IX.2-eGFP detected with anti-HRP had the similar size as that detected
with anti-GFP (Fig. 5A), indicating that both LecRK-IX.1 and LecRK-IX.2 are
N-glycosylated. Also, the kinase-dead mutants could be detected with both anti-GFP
and anti-HRP and showed similar sizes as the corresponding full length LecRKs (Fig.
5A). It should be noted that in all cases the detected LecRK-IX.2 protein level was less
than LecRK-IX.1. Also, accumulation of LecRK-IX.1 and LecRK-IX.2 protein was
lower when compared to the kinase-dead mutants, indicating that the latter are more
stable or show lower turnover rate.
A hallmark of defence following activation of receptors is the rapid production of
ROS (Boller and Felix, 2009). Hence, we monitored H2O2 production using DAB ((3,
3’-diaminobenzidine)) staining in N. benthamiana upon transient expression of
Arabidopsis LecRK-IX.1 and LecRK-IX.2. Agro-infiltrated leaves expressing GFP did
not show any distinguishable DAB staining whereas the other half of the leaves
expressing LecRK-IX.1 or LecRK-IX.2 clearly showed DAB staining visualized as a

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
109
Figure 5. H2O2 accumulation and cell death induced by transient expression of LecRK-IX.1,
LecRK-IX.2 and derivatives in N. benthamiana.
A, LecRK-IX.1-eGFP, LecRK-IX.2-eGFP and kinase-dead mutants are expressed and glycosylated in
N. benthamiana. eGPF-tagged LecRKs and kinase-dead mutants were co-expressed with P19 in
N. benthamiana by Agrobacterium-mediated transformation. Total protein was isolated and subjected
to immunopurification (IP) using GFP-trap_A® beads. The immunopurified proteins were detected by
western blot (IB) with anti-GFP or anti-HRP. Coomassie staining shows the 50 KDa Rubisco band
present in the total protein extract to indicate equal loading of protein in each lane. B, H2O2
accumulation in N. benthamiana leaves expressing GFP, LecRK-IX.1-eGFP, LecRK-IX.2-eGFP and
LecRK-S.4-eGFP with (+P19) or without P19 (-P19). Leaves were collected two days after

Chapter IV
110
agro-infiltration (dpa), stained with DAB solution and destained with ethanol. C, Cell death on
N. benthamiana leaves expressing GFP, LecRK-IX.1-eGFP and LecRK-IX.2-eGFP with (+P19) or
without P19 (-P19) at 3 dpa. D, Quantification of cell death induced by expression of LecRK-IX.1-eGFP
and LecRK-IX.2-eGFP in N. benthamiana by ion leakage measurement. Six leaf discs (Ø 1cm)
collected from infiltration zones were incubated in deionized water for 2 h with 200 rpm shaking.
Supernatants were collected for measuring ion leakage. Bars represent mean values (± SE) of six
replicates from each experiment. * indicates significant difference (p < 0.05) according to a t test. This
experiment was repeated three times with similar results. E, H2O2 accumulation in N. benthamiana
leaves expressing LecRK-IX.1-eGFP, LecRK-IX.2-eGFP and derivatives with P19. Leaves collected at
1.5 dpa were stained with DAB and destained with ethanol. F, Cell death development in
N. benthamiana leaves expressing LecRK-IX.1-eGFP, LecRK-IX.2-eGFP and derivatives with P19 at 3
dpa.
brown precipitate (Fig. 5B). Co-expression with the silencing suppressor P19 resulted
in more intense DAB staining demonstrating significantly enhanced H2O2 production
(Fig. 5B). Irrespective of P19, DAB staining was always stronger in leaves expressing
LecRK-IX.2 than those expressing LecRK-IX.1, indicating that the presence of
LecRK-IX.2 causes a higher H2O2 production (Fig. 5B). In contrast, DAB staining was
not observed in leaves expressing another LecRK, i.e. LecRK-S.4, suggesting that
induction of H2O2 is typical for the two clade IX LecRKs. Also the spontaneous cell
death phenotype that was observed in leaves expressing LecRK-IX.2 was much
stronger than in leaves expressing LecRK-IX.1 (Fig. 5C). The extent of cell death was
quantified by measuring ion leakage and this showed that the conductivity was higher
in N. benthamiana leaves expressing LecRK-IX.2 than in those expressing
LecRK-IX.1 (Fig. 5D). Upon co-expression with P19, LecRK-IX.1 produced a slightly
delayed but eventually similar extent of cell death as LecRK-IX.2 (Fig. 5, C and D).
None of the leaves expressing GFP, with or without P19, showed any visible cell death
(Fig. 5, C and D).
Expression of lectin-deletion mutant of LecRK-IX.1 or LecRK-IX.2 in
N. benthamiana induced a similar extent of H2O2 production and cell death as
expression of the full length proteins (Fig. 5, E and F). In contrast, leaves expressing
kinase-dead mutants showed neither H2O2 production nor cell death. Collectively,
these results show that the kinase activity but not the lectin domain plays a role in
LecRK-IX.1- and LecRK-IX.2-mediated H2O2 production and cell death induction.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
111
Silencing of NbSIPK/NTF4 abolishes LecRK-IX.1-induced cell death in
N. benthamiana
To identify downstream signaling components required for LecRK-IX.1 and
LecRK-IX.2-induced cell death in N. benthamiana, we analysed the role of several
genes known to be involved in cell death induction (Rusterucci et al., 2001; Yoshioka
et al., 2003; Gabriëls et al., 2007; Takahashi et al., 2007). NbNPR1, NbEDS1,
NbMEK2, NbNRC1, NbRbohA/B and NbSIPK/NTF4 were individually silenced in
N. benthamiana using TRV-mediated virus induced gene silencing (VIGS).
TRV:GUS-treated plants were used as control. Three to four weeks after
TRV-inoculation, LecRK-IX.1-eGFP, LecRK-IX.2-eGFP and GFP were expressed in
leaves using agro-infiltration. In all cases, expression of LecRK-IX.1-eGFP or
LecRK-IX.2-eGFP resulted in cell death on TRV:GUS plants (Fig. 6A). Similarly, cell
death was observed in leaves silenced for NbEDS1, NbNRC1 and NbRbohA/B,
whereas it was attenuated in leaves silenced for NbMEK2 or NbNPR1 (Supplemental
Fig. S5). These results indicate that LecRK-mediated cell death is not dependent on
NbEDS1, NbNRC1 or NbRbohA/B, but partially dependent on NbMEK2 and NbNPR1.
In contrast, cell death induced by LecRK-IX.1-eGFP was completely abolished in
leaves silenced for NbSIPK and its closest homolog NbNTF4 (Fig. 6A). The efficiency
of silencing was checked and shown to be close to 100% (Fig. 6B). Loss of
LecRK-IX.1-induced cell death in NbSIPK/NTF4-silenced leaves was confirmed by
staining with trypan blue (Fig. 6C). In addition, cell death induced by
LecRK-IX.2-eGFP was also remarkably compromised in the NbSIPK/NTF4-silenced
leaves (Fig. 6 A and C). But under the same conditions, cell death elicited by the
Phytophthora infestans elicitor NPP1 was not altered. Compared to TRV:GUS-treated
leaves, leaves silenced for NbSIPK/NTF4 showed a slight delay in cell death
development, but eventually the extent of cell death induced by NPP1 was
comparable (Fig. 6A). In line with this, NbSIPK/NTF4-silenced leaves expressing
LecRK-IX.1-eGFP or LecRK-IX.2-eGFP showed a significant reduction in ion leakage
compared to TRV:GUS-treated leaves; but for leaves expressing NPP1, this was not
the case (Fig. 6D). To determine whether the loss of cell death observed in
NbSIPK/NTF4-silenced plants is due to reduced protein accumulation, GFP and
LecRK-IX.1-eGFP protein levels were determined but no differences were found when
compared to those detected in the TRV:GUS-treated plants (Fig. 6E), indicating that
inhibition of LecRK-IX.1-induced cell death in NbSIPK/NTF4-silenced leaves is not
due to reduced protein accumulation. In conclusion, NbSIPK and NbNTF4 are
indispensable downstream signaling components for LecRK-IX.1- and LecRK-IX.2-
mediated cell death.

Chapter IV
112
Figure 6. Silencing of NbSIPK/NTF4 suppresses LecRK-IX.1- and LecRK-IX.2-induced cell death
in N. benthamiana leaves.
A, Cell death induced by LecRK-IX.1-eGFP, LecRK-IX.2-eGFP and NPP1 on control (TRV:GUS) and
NbSIPK/NTF4-silenced N. benthamiana leaves. GFP, NPP1, LecRK-IX.1-eGFP and LecRK-IX.2-
eGFP were expressed by Agrobacterium-mediated transient expression. Pictures were taken at 3 dpa.
B, Relative quantification of NbSIPK and NbNTF4 transcript levels in N. benthamiana leaves three
weeks after inoculation with TRV:GUS or TRV:NbSIPK/NTF4 constructs. Transcript levels were
normalized using NbActin and expressed as mean fold changes (± SD) relative to the transcript level in
TRV:GUS-treated leaves which was arbitrarily set as 1. C, Microscopic investigation of cell death
induced by LecRK-IX.1-eGFP and LecRK-IX.2-eGFP in TRV:GUS- and TRV:NbSIPK/NTF4-treated
N. benthamiana leaves. Leaves were collected at 3 dpa and stained with trypan blue. Scale bars
represent 200 µm. D, Quantification of ion leakage in TRV:GUS- or TRV:NbSIPK/NTF4-treated
N. benthamiana leaves expressing GFP, NPP1, LecRK-IX.1-eGFP or LecRK-IX.2-eGFP. Quantified ion
leakage was relative to that in GFP expressing leaves which was arbitrarily set as 1. * indicates

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
113
significant difference (p < 0.05) according to a t test. Similar results were obtained in three independent
experiments. E, Accumulation of GFP and LecRK-IX.1-eGFP in TRV:GUS- and
TRV:NbSIPK/NTF4-treated N. benthamiana leaves. Total protein was immunopurified by GFP-trap_A®
beads and immunodetected with anti-GFP. Coomassie staining indicates the amount of loading in each
lane.
Silencing of NbSIPK/NTF4 compromises LecRK-IX.1-mediated Phytophthora
resistance in N. benthamiana
To determine whether LecRK-IX.1 and LecRK-IX.2 are capable to enhance
Phytophthora resistance in N. benthamiana, we inoculated P. capsici on
N. benthamiana leaves with one half of the leaf expressing GFP and the other half
expressing LecRK-IX.1-eGFP or LecRK-IX.2-eGFP. Disease symptoms were
evaluated at 3 dpi. Leaves expressing LecRK-IX.1-eGFP showed significantly
reduced lesion sizes compared to those expressing GFP (Fig. 7A). Leaves expressing
LecRK-IX.2 showed severe cell death and were not included in the infection assays
(Fig. 5C).
Host cell death often associates with disease resistance, especially against
(hemi)biotrophic pathogens (Mur et al., 2008). Absence of LecRK-IX.1-induced cell
death in NbSIPK/NTF4-silenced plants raised the possibility that also the resistance
function of LecRK-IX.1 might be affected. Hence, we transiently expressed
LecRK-IX.1-eGFP and GFP in TRV:GUS- or TRV:NbSIPK/NTF4-treated plants and
inoculated these plants with P. capsici. Lesion sizes on TRV:GUS-treated leaves
expressing LecRK-IX.1-eGFP were significantly reduced compared to those
expressing GFP (Fig. 7, B and C). Similarly, TRV:NbSIPK/NTF4-treated leaves
expressing LecRK-IX.1-eGFP also showed reduced lesion sizes compared to those
expressing GFP. However, the reduction in lesion size was not as strong as on
TRV:GUS-treated leaves (Fig. 7, B and C), indicating that silencing of NbSIPK/NTF4
did not completely abolish the function of LecRK-IX.1 in Phytophthora disease
resistance. Since cell death was completely abolished in NbSIPK/NTF4-silenced
leaves, it can be concluded that LecRK-IX.1-mediated Phytophthora resistance is not
entirely a consequence of induced cell death. It is likely that additional components or
responses other than cell death play a role in inhibiting Phytophthora colonization.

Chapter IV
114
Figure 7. Phytophthora resistance conferred by transient expression of LecRK-IX.1 in
N. benthamiana.
A, Disease symptoms and lesion sizes on N. benthamiana leaves expressing LecRK-IX.1 or GFP three
days after plug-inoculation with P. capsici LT263. P. capsici was inoculated on N. benthamiana leaves
24 hours after agro-infiltration. Each experiment included at least 18 infiltrated N. benthamiana leaves.
Bars represent mean lesion sizes (± SE). * indicates significant difference in lesion size (p < 0.05)
according to a t test. This experiment was repeated three times with similar results. B-C, Disease
symptoms and quantified lesion sizes on TRV:GUS- and TRV:NbSIPK/NTF4-treated N. benthamiana
leaves expressing LecRK-IX.1-eGFP or GFP three days after inoculation with P. capsici LT263. The
infection assay included at least 12 infiltrated N. benthamiana leaves per treatment. Bars represent
mean lesion sizes (± SE). * indicates significant difference in lesion size (p < 0.05) according to a t test.
This experiment was repeated three times with similar results.
LecRK-IX.1 and LecRK-IX.2 interact with N. benthamiana NbPDR1 and
Arabidopsis ABCG40 in planta
To identify potential LecRK-interacting proteins, our idea was to co-immunoprecipitate
proteins that form a complex with the eGFP-tagged LecRK-IX.1 and eGFP-tagged
LecRK-IX.2 in the stable transgenic Arabidopsis lines and subsequently analyse these
co-immunoprecipitated proteins by liquid chromatography mass spectrometry
(LC-MS). However, we were unable to purify sufficient amounts of protein for LC-MS
analysis either because transgene expression was too low or because lines with a

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
115
higher transgene expression showed severe cell death (Fig. 2). Since Arabidopsis
LecRK-IX.1 and LecRK-IX.2 retained their function when transiently expressed in
N. benthamiana, we anticipated also that the interaction partners might be conserved
in the two plant species. Hence, we transiently expressed LecRK-IX.1-eGFP,
LecRK-IX.2-eGFP and the control GFP in N. benthamiana and analysed the
(co-)immunoprecipitated proteins with LC-MS after trypsin digestion. The resulting
masses were used as queries to search in the protein database containing sequences
from N. benthamiana and the input LecRKs and GFP. A large number of peptides
matching LecRK-IX.1 and LecRK-IX.2 were detected, demonstrating successful
purification of the input LecRKs (Fig. 8A). Further analysis revealed several potential
candidates that were detected only in the LecRK samples but not in the control GFP
sample. One of these is an ATP-binding cassette (ABC) transporter. Multiple peptides
in both the LecRK-IX.1 and LecRK-IX.2 samples matched uniquely to NbPDR1
(NbS00038999g0004.1), a pleiotropic drug resistance-type ABC transporter (Fig. 8, A
and B; Supplemental Table S2). Apparently, both LecRK-IX.1 and LecRK-IX.2 interact
with NbPDR1 in planta.
We subsequently determined the interaction of both LecRKs with Arabidopsis
ABCG40 (alias PDR12), the closest homolog of NbPDR1 in Arabidopsis.
LecRK-IX.1-eGFP, LecRK-IX.2-eGFP or GFP were co-expressed with ABCG40-Myc
in N. benthamiana and immunoprecipitated using GFP-trap_A® beads. As shown by
western blots probed with Myc antibody, ABCG40-Myc co-immunoprecipitated with
both LecRK-IX.1-eGFP and LecRK-IX.2-eGFP, but not with GFP (Fig. 8C). Note that
two bands were detected, with the lower band closer to the predicted protein size of
ABCG40-Myc. Interaction was further confirmed by swapping the epitope tags.
Consistently, both LecRK-IX.1-Myc and LecRK-IX.2-Myc, but not GUS-Myc
co-immunoprecipitated with ABCG40-eGFP and again two bands were detected for
ABCG40-eGFP (Fig. 8C).
Since both LecRK-IX.1 and LecRK-IX.2 interact with NbPDR1 and ABCG40, we
checked whether LecRK-IX.1 associates with LecRK-IX.2 in planta, but no interaction
was detected (Fig. 8D). Also in preliminary experiments, no interaction was detected
between kinase-dead mutants of LecRK-IX.1 and LecRK-IX2. However, LecRK-IX.1
and LecRK-IX.2 were found to interact with themselves, indicating homodimer
formation. Similarly, the kinase-dead mutants of both LecRKs were also found to form
homodimers (Fig. 8D).
Collectively, these results indicate independent interaction of LecRK-IX.1 and
LecRK-IX.2 with ABCG40 in Arabidopsis and the ability to form homodimers.

Chapter IV
116
Figure 8. NbPDR1 and ABCG40 interact with LecRK-IX.1 and LecRK-IX.2 in planta.
A, The total number of peptides identified by LC-MS matching LecRK-IX.1, LecRK-IX.2 and the
potential interactor NbPDR1. B, Protein sequence of NbPDR1 with peptides detected by LC-MS

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
117
highlighted. Peptides detected only in LecRK-IX.1 sample are depicted as yellow, peptides detected in
both LecRK-IX.1 and LecRK-IX.2 samples are depicted as green while peptides that match uniquely to
NbPDR1 are underlined. C, ABCG40 interacts with LecRK-IX.1 and LecRK-IX.2 in planta. ABCG40
was co-expressed with LecRK-IX.1, LecRK-IX.2, GFP or GUS in N. benthamiana. Immunoprecipitation
was performed using anti-GFP Immunoblotting was performed using anti-GFP or anti-Myc. Coomassie
staining indicates equal amount of loading in each lane. D, Dimerization of LecRK-IX.1, LecRK-IX.2
and kinase-dead mutants in N. benthamiana. GPF- and Myc-tagged LecRKs were co-expressed with
P19 in N. benthamiana. Total protein was isolated, immunopurified by GFP-trap_A® beads and
detected with anti-GFP or anti-Myc. Coomassie staining indicates equal amount of loading in each
lane.
Arabidopsis ABCG40 is a potential Phytophthora resistance component
To determine the biological relevance of the interaction between ABCG40 and the two
LecRKs, we monitored the expression of ABCG40 in Col-0 and LecRK mutants during
infection with P. capsici LT123. In Col-0, expression of ABCG40 was increased
significantly from 12 to 24 hpi. In comparison, the expression levels were much lower
in both lecrk-IX.1 and lecrk-IX.2 (Fig. 9A), indicating that ABCG40 functions
downstream of the two LecRKs. To determine the role of ABCG40 in Phytophthora
resistance, two homozygous Arabidopsis mutants, i.e. abcg40-1 and abcg40-2 with
T-DNA insertions in the coding regions (Fig. 9B), were analysed in Phytophthora
infection assays. Absence of full-length ABCG40 mRNA in the mutants was confirmed
by semi-qRT-PCR (Fig. 9C). Upon inoculation with P. brassicae HH and P. capsici
LT123, both lines developed clear disease symptoms at 4 dpi, while Col-0 plants
remained fully resistant (Fig. 9D). This points to a function for ABCG40 in
Phytophthora resistance.

Chapter IV
118
Figure 9. Arabidopsis ABCG40 is a potential Phytophthora resistance component.
A, ABCG40 expression in Col-0, lecrk-IX.1 and lecrk-IX.2 upon inoculation with P. capsici LT123.
Relative transcript levels were normalized to Arabidopsis Actin2. Values are expressed as mean fold
changes (± SD) relative to the transcript levels in mock-inoculated leaves. B, Schematic representation
of ABCG40. Domains were predicted by SMART. Black arrowheads point to the T-DNA insertion sites in
mutants abcg40-1 and abcg40-2 while ABCG40-P1 indicates the primer pair used for semi-qRT-PCR.
C, Transcript levels of ABCG40 in Col-0, abcg40-1 and abcg40-2 detected by semi-qRT-PCR. Actin2
was used as control. D, Disease symptoms on Col-0, abcg40-1 and abcg40-2 four days after
inoculation with P. brassicae HH and P. capsici LT123. White arrowheads point to the inoculated
leaves.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
119
DISCUSSION
As one of the largest RLK subfamilies in Arabidopsis, LecRKs are considered to play
diverse roles in plant adaptation. In recent years, detailed studies on three members
of this subfamily, i.e. Arabidopsis LecRK-I.9, LecRK-V.5 and LecRK-IV.2, revealed
their importance in plant immunity (Bouwmeester et al., 2011; Desclos-Theveniau et
al., 2012; Singh et al., 2012). In this study, we focus on two Arabidopsis LecRKs
belonging to clade IX, namely LecRK-IX.1 and LecRK-IX.2 that were previously
identified as potential Phytophthora resistance components based on phenotypic
analyses of T-DNA insertion lines (Wang et al., 2014/Chapter III). We confirmed the
role of LecRK-IX.1 and LecRK-IX.2 in Phytophthora resistance and found that they act
as functional analogs. Knockout of either LecRK-IX.1 or LecRK-IX.2 in Arabidopsis
leads to a gain of susceptibility to Phytophthora pathogens and reduced expression of
defence-related genes upon infection. Overexpression of each of the two in
Arabidopsis increases Phytophthora resistance, but also induces spontaneous plant
cell death, in particular in transgenic lines with a relatively high transgene expression
level. In addition, LecRK-IX.1 retained its function as a Phytophthora resistance
component when transiently expressed in the Solanaceous plant N. benthamiana.
Both LecRKs as well induced cell death when expressed in N. benthamiana,
suggesting conservation of downstream components for LecRK-mediated signaling in
different plant species. Both LecRKs were found to associate with ABC transporters
independent of each other suggesting that they participate in the same signaling
network. In Arabidopsis, the ABC transporter ABCG40 turned out to be a potential
Phytophthora resistance component.
Programmed cell death often occurs in response to pathogen invasion (Coll et al.,
2011; Dickman and de Figueiredo, 2013; Dickman and Fluhr, 2013). The effect of cell
death on plant resistance depends largely on the lifestyle of the invading pathogen. In
general, activation of cell death at early infection stages often limits proliferation of
biotrophic and hemibiotrophic pathogens but promotes infection of necrotrophic
pathogens. However, several reports showed that plant cell death is not always
essential for resistance (Yu et al., 1998; Genger et al., 2008; Takahashi et al., 2012).
The cell death phenotype induced by overexpression of LecRK-IX.1 and LecRK-IX.2
raised the question whether cell death is involved in LecRK-mediated Phytophthora
resistance. Here, we provide three lines of evidence to show that LecRK-mediated
Phytophthora resistance is not due to induction of cell death. First of all, for both
LecRKs, increased Phytophthora resistance was detected in transgenic Arabidopsis
lines that did not show the cell death phenotype. The intensity of cell death was found
to be correlated with transgene expression levels and cell death was only observed in
the transgenic lines with relative high LecRK-IX.1 or LecRK-IX.2 expression levels;

Chapter IV
120
whereas LecRK-mediated Phytophthora resistance was also found in the lines with
relative low transgene expression. Microscopic analysis showed that the early stage
of Phytophthora infection was compromised on Arabidopsis LecRK overexpression
lines, but this was not correlated with increased cell death. Secondly, the lectin
domain was found to be required for LecRK-mediated Phytophthora resistance but
dispensable for cell death execution. Apparently, the extracellular lectin domain is
required for monitoring pathogen attack by recognizing either effectors/PAMPs of
Phytophthora or DAMPs released upon infection. This recognition could in turn lead to
activation of plant defence mediated by the kinase domain. The spontaneous cell
death phenotype may be a consequence of constitutive activation of pathogen-
independent responses, such as production of ROS that has been shown to play a
prominent role in the execution of plant cell death (van Breusegem and Dat, 2006).
Along with induced cell death, elevated H2O2 production was detected in
N. benthamiana expressing LecRK-IX.1 or LecRK-IX.2, either full length or truncated
versions lacking the lectin domain. This indicates that the H2O2 production induced by
expression of LecRK-IX.1 or LecRK-IX.2 is tightly correlated with cell death induction.
Nevertheless, there is still a possibility that both LecRKs are involved in regulating
plant cell death, similar to SOBIR1, an Arabidopsis RLK which also activates plant cell
death upon overexpression (Gao et al., 2009). The third line of evidence is that
LecRK-mediated cell death and Phytophthora resistance in N. benthamiana require
different downstream components. One of the important events leading to plant cell
death is the sequential activation of MAPK cascades (Meng and Zhang, 2013). By
virus-induced gene silencing in N. benthamiana, we identified NbSIPK/NbNTF4 as
being essential for LecRK-IX.1 and LecRK-IX.2-mediated cell death. Both NbSIPK
and NbNTF4 have been shown to be involved in defence responses downstream of
receptors and capable to induce cell death when expressed in N. benthamiana
(Zhang and Liu, 2001; Ren et al., 2006; Segonzac et al., 2011; Huang et al., 2014).
Silencing of NbSIPK/NbNTF4 completely abolished LecRK-IX.1-mediated cell death,
whereas Phytophthora resistance was decreased, but not completely lost. This
indicates that NbSIPK and NbNTF4 participate in, but not determine
LecRK-IX.1-mediated Phytophthora resistance in N. benthamiana.
For RLKs, the kinase domain was found to be crucial in initiating signal
transduction via phosphorylation of downstream substrates as point mutations often
abolish RLK-mediated functions (Gomez-Gomez et al., 2001; Morillo and Tax, 2006;
Schwessinger et al., 2011; Liebrand et al., 2013). Although it is very likely that the
kinase domain of the various LecRKs has a similar role in signal transduction, there is
as yet no proof that their kinase activity is essential for biological function. LecRK-VI.2
has been shown that is capable to auto-phosphorylate in vitro (Singh et al., 2013), but
whether this kinase activity is essential for its role in priming plant defence is unknown.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
121
In this study, we analysed the relevance of the kinase activity of LecRK-IX.1 and
LecRK-IX.2 for mediating resistance and cell death induction. LecRKs belong to
RD-kinases with an aspartate (D) in the catalytic loop preceded by a conserved
arginine (R) (Nolen et al., 2004; Kornev et al., 2006). It has been demonstrated that
mutation of the conserved RD motif into RN or AA blocks kinase activity
(Schwessinger et al., 2011; van Damme et al., 2012). Upon mutation of the RD motif,
both LecRK-mediated Phytophthora resistance and cell death were completely
abolished. These results demonstrated that both LecRKs harbor an active kinase and
that kinase activity is indispensable for their function.
Plant receptors do not work alone but rather function as part of multi-protein
complexes (Liebrand et al., 2014; Macho and Zipfel, 2014). Recently, one of the
Arabidopsis LecRKs, namely LecRK-VI.2, was found to associate with the flagellin
receptor FLS2 and to prime flg22-mediated defence in Arabidopsis and
N. benthamiana (Huang et al., 2014). In our hands, the identification of potential
LecRK-IX.1- and LecRK-IX.2-interacting proteins in Arabidopsis was hampered by the
severe cell death phenotype. Due to the conserved function of the two LecRKs in
N. benthamiana, we exploited N. benthamiana as a convenient alternative. This
resulted in the identification of NbPDR1 and its closest homolog in Arabidopsis, i.e.
ABCG40, as potential LecRK-interacting proteins. NbPDR1 and ABCG40 belong to a
pleiotropic drug resistance transporter subfamily (van den Brule and Smart, 2002;
Rea, 2007). ABCG40 in Arabidopsis was previously reported to be involved in lead
detoxification and ABA transport and has also been implicated in transport of the
diterpenoid sclareol to restrict pathogen growth (Lee et al., 2005; Kang et al., 2010).
For example, inhibition of Ralstonia solanacearum in tobacco by ABCG40 and its
homolog NtPDR1 is associated with sclareol-related compounds (Seo et al., 2012).
Similarly, its close homolog in Nicotiana plumbaginifolia (i.e. NpPDR1) contributes to
transport of sclareol and resistance to Botrytis cinerea, Fusarium oxysporum,
Rhizoctonia solani and Phytophthora nicotianae (Stukkens et al., 2005; Bultreys et al.,
2009). In Arabidopsis, however, ABCG40 does not confer resistance to
Fusarium oxysporum, A. brassicicola and P. syringae, indicating that it is not a
universal resistance component (Campbell et al., 2003). In this study, the
compromised resistance of the two independent Arabidopsis ABCG40 mutants to
Phytophthora pathogens indicates that, similar to LecRK-IX.1 and LecRK-IX.2,
ABCG40 functions as a Phytophthora resistance component. It is not surprising that
both LecRK-IX.1 and LecRK-IX.2 interact with NbPDR1 and ABCG40 in planta since
they were also found to activate similar downstream defence responses. It is unlikely
though that LecRK-IX.1, LecRK-IX.2 and ABCG40 form a ternary complex because
LecRK-IX.1 and LecRK-IX.2 do not form a heterodimer. The fact that both clade IX
LecRKs interact with the same ABC transporter but yet function independently raises

Chapter IV
122
the possibility that their interaction with the ABC transporter is temporally and spatially
different. Since ABC transporters contribute to transport of a wide variety of
compounds, including ions, nucleic acids and anti-microbial compounds (Rea, 2007),
binding between ABCG40 and the clade IX LecRKs might cause rearrangement of
plant molecules and the subsequent activation of signal transduction culminating in
effective defence.
MATERIALS AND METHODS
Plant material and growth conditions
Arabidopsis T-DNA insertion lines SALK_042414 (henceforth named as lecrk-IX.1),
SALK_111817 (lecrk-IX.2), SALK_148565.27.50 (abcg40-1) and SALK_005635.51.50
(abcg40-2) were obtained from the European Arabidopsis stock center NASC
(http://arabidopsis.info) and homozygosity of the T-DNA insertions was determined as
previously described (Wang et al., 2014/Chapter III). Arabidopsis Col-0 and derived
mutants or transgenic lines were grown on soil and maintained in a conditioned
growth chamber at 19-21°C with a 12 h photoperiod and a 75-80% relative humidity.
N. benthamiana plants were grown in soil in a greenhouse at 19-21°C with a
75-78% relative humidity and a 16/8 h photoperiod. Supplementary light (100 W m2-)
was applied when the light intensity dropped below 150 W m2-.
Pathogen maintenance and infection assays
Culturing of Phytophthora pathogens, Pst DC3000 and A. brassicicola, infection
assays on Arabidopsis with these pathogens and biomass measurements were
performed as previously described (Wang et al., 2013/Chapter IIa; Wang et al.,
2014/Chapter III).
Infection assays on N. benthamiana with P. capsici were performed by inoculating
fresh mycelial plugs (Ø 0.5 cm) on leaves of six-week-old plants one day after
agro-infiltration. Inoculated plants were kept in a climate chamber with
aforementioned conditions. Disease symptoms were evaluated by measuring lesion
sizes at 3 dpi as described by Vleeshouwers et al. (1999).
Gene expression analysis
Arabidopsis RNA was isolated from eight circular leaf discs (Ø 0.8 cm), each with the
inoculation spots in the center from four inoculated Arabidopsis plants or from rosette
leaves collected from transgenic plants. For N. benthamiana, six leaves from three
individual plants were pooled together and ground in liquid nitrogen. Total RNA
isolation and qRT-PCR analysis were carried out following description in Chapter III.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
123
First strand cDNA was synthesized using 1.5 µg RNA in a total volume of 25 µl and
the resultant cDNA was diluted 20-fold. Semi-quantative RT-PCR was performed
using 1 µl of diluted cDNA with gene-specific primers (Supplemental Table S1) in 45
cycles at 94°C for 30s, 55°C for 30s, and 72°C for 1 min. Actin2 was used as control
and amplified using the same PCR conditions in 30 cycles.
LecRK expression data were obtained from publicly available microarray datasets
using the eFP-Browser at the Bio-Array Resource website (http://www.bar.utoronto.ca;
Winter et al., 2007). Signal intensity log2 ratios of the samples relative to the mean
were used as a measure for LecRK expression. Of the datasets sampled from
Arabidopsis seeds, roots and flowers, the highest value throughout all the arrays was
retrieved.
Plasmid construction
The coding sequences of the full-length or truncated Arabidopsis LecRK-IX.1,
LecRK-IX.2, LecRK-S.4 and ABCG40 were amplified by PCR using Pfu DNA
polymerase (Promega) with primers listed in Supplemental Table S1. Point mutations
in the kinase domain were generated using overlap extension PCR with primers
shown in Supplemental Table S1. The purified PCR fragments were ligated into
pENTR/D-TOPO® vector (Invitrogen) for sequencing. Thereafter, the correct
sequences were introduced into binary vectors pSol2095 or pGWB20 using Gateway®
LR Clonase II® (Invitrogen). The resulting vectors were transformed into
Agrobacterium tumefaciens strain GV3101.
The fragment used for gene silencing of NbRbohA/B (Yoshioka et al., 2003) was
synthesized (Eurofins MWG Operon) while the fragment used for NbSIPK/NTF4
silencing was amplified using primers listed in Supplemental Table S1. Synthesized or
PCR-amplified fragments were cloned into the vector pTRV-RNA2 and transformed
into A. tumefaciens GV3101.
Generation of Arabidopsis transgenic lines and double mutants
Arabidopsis plants were transformed with A. tumefaciens GV3101 carrying the
aforementioned constructs using the floral dip method (Zhang et al., 2006).
Transformed seeds were selected on 0.5 MS (Murashige and Skoog, Duchefa) plates
containing 50 µg ml-1 kanamycin or 20 µg ml-1 hygromycin B. Transgene expression in
the different transgenic lines was detected by qRT-PCR using primers described in
Supplemental Table S1.
Double mutants were generated by reciprocally crossing mutant lecrk-IX.1 with
lecrk-IX.2. Homozygosity of the T-DNA insertion in both LecRK-IX.1 and LecRK-IX.2
in F2 progeny was confirmed by PCR-based genotyping. Expression levels of
LecRK-IX.1 and LecRK-IX.2 in the double mutants were analysed by semi-qRT-PCR

Chapter IV
124
with Actin2 as control.
A. tumefaciens-mediated transient expression in N. benthamiana leaves
A. tumefaciens strains carrying binary vectors were grown overnight at 28°C in Yeast
Extract Broth with appropriate antibiotics. A. tumefaciens cells were collected by
centrifugation, resuspended and incubated in MMA induction medium (10 mM MES,
10 mM MgCl2, 50 μM acetosyringone, pH 5.6) for 3 h. Thereafter, A. tumefaciens cells
were collected, resuspended in an infiltration medium (10 mM MES, 10 mM MgCl2,
150 μM acetosyringone, pH 5.6) and incubated for another 1 h.
For gene silencing, A. tumefaciens suspensions carrying pTRV-RNA2 constructs
and A. tumefaciens carrying pTRV1 were mixed at a ratio of 1:1 to a final OD600 of
1.0 before infiltration into cotyledons of two-week-old N. benthamiana plants. For
transient gene expression, A. tumefaciens suspensions with a final OD600 of 0.6 were
syringe-infiltrated into five-week-old N. benthamiana leaves or co-infiltrated with the
silencing suppressor P19 at a ratio of 1:1 to a final OD600 of 0.6.
Protein extraction, immunoprecipitation and western blotting
Agro-infiltrated N. benthamiana leaves were collected and gound in liquid nitrogen.
Total protein was extracted by incubating ground leaf samples for 30 min in an
extraction buffer containing 150 mM NaCl, 50 mM Tris-HCl pH 8.0, 1.0% IGEPAL®
CA-630 (Sigma) and one protease inhibitor cocktail tablet (Roche) per 50 ml. The
homogenate was centrifuged at 18,000 rpm for 20 min and the supernatant was
subsequently incubated with GFP-trap_A® beads (Chromotek) at 4°C for 1-2 h. After
incubation, GFP-beads were pelleted and washed with extraction buffer for 6 times.
Proteins were eluted from GFP-beads by boiling for 5 min, separated on a 8%
SDS-PAGE gel and electroblotted onto a PVDF membrane (Bio-Rad). Accumulation
of GFP-tagged protein was detected by immunoblotting with anti-GFP-HRP (Miltenyi
Biotec). Detection of Myc-tagged protein was performed by incubating the blots in
1:2,000 diluted anti-c-Myc (Santa Cruz Biotechnology) and subsequently 1:2,000
diluted anti-Mouse Ig-HRP (Amersham). Protein N-glycosylation was detected using
anti-HRP as previously described (Liebrand et al., 2012). Supersignal West Femto
Chemiluminescent Substrate (Thermo Scientific) was used for signal development.
Equal loading was demonstrated by staining the membrane with coomassie brilliant
blue.
Tryptic digestion of immunopurified proteins and mass spectrometry analysis
For mass spectrometry analysis of LecRK-interacting proteins, 5 g agro-infiltrated
N. benthamiana leaves were used for protein isolation and the resultant protein
extract was incubated with 60 μl GFP-trap_A® beads (Chromotek) at 4°C for 2 h. The

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
125
purified proteins on GFP-trap_A® beads were subjected to on-bead digestion followed
by LC-MS using both Data-Independent Acquisition (DIA/MSE) and Data-Dependent
Acquisition (DDA) as previously described (Liebrand et al., 2012).
Measurement of electrolyte leakage
Six N. benthamiana leaf discs (Ø 1.0 cm) collected three days after agro-infiltration
were floated on 4 ml of deionized water for 2 h under continuous shaking (200 rpm) at
room temperature. Conductivity was measured using a Mettler Toledo InLab®741 ISM
conductivity meter (Mettler Toledo, Tiel). Three biological repeats were performed
each with six replicates.
Trypan blue and DAB staining
Trypan blue staining was performed as previously described (Wang et al., 2011). For
DAB staining, agro-infiltrated N. benthamiana leaves were immersed into DAB
solution (1 mg ml-1 DAB-HCl, pH 3.7) for 8 h. Leaves were subsequently washed with
dH2O to remove the staining solution on the surface and destained with 96% ethanol.
Experiments were repeated three times each with at least six leaves from three
independent plants.
ACKNOWLEDGEMENTS
We thank Patrick Smit and Daniela Sueldo for providing the pSol2095 vector and the
GFP-HA construct, Henk Smid and Bert Essenstam from Unifarm for taking care of
the plants and Hagos M. Juhar for assistance. This project was financially supported
by a Wageningen University sandwich-PhD fellowship (YW), a Huygens scholarship
(YW), a VENI grant (KB) from the Netherlands Organization for Scientific Research
(Technology Foundation NWO-STW), CBSG2012/NPC2 proteomics hotel and the
Food-for-Thought campaign of the Wageningen University Fund.
REFERENCES
Boller, T. and Felix, G. (2009). A renaissance of elicitors: Perception of microbe-associated molecular
patterns and danger signals by pattern-recognition receptors. Annual Review of Plant Biology, 60,
379-406.
Bouwmeester, K. and Govers, F. (2009). Arabidopsis L-type lectin receptor kinases: phylogeny,
classification, and expression profiles. Journal of Experimental Botany, 60, 4383-4396.
Bouwmeester, K., de Sain, M., Weide, R., Gouget, A., Klamer, S., Canut, H. and Govers, F. (2011). The
lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a potential host
target for a RXLR effector. PLoS Pathogens, 7, e1001327.

Chapter IV
126
Bultreys, A., Trombik, T., Drozak, A. and Boutry, M. (2009). Nicotiana plumbaginifolia plants silenced for
the ATP-binding cassette transporter gene NpPDR1 show increased susceptibility to a group of
fungal and oomycete pathogens. Molecular Plant Pathology, 10, 651-663.
Campbell, E.J., Schenk, P.M., Kazan, K., Penninckx, I.A.M.A., Anderson, J.P., Maclean, D.J., Cammue,
B.P.A., Ebert, P.R. and Manners, J.M. (2003). Pathogen-responsive expression of a putative
ATP-Binding cassette transporter gene conferring resistance to the diterpenoid sclareol is regulated
by multiple defense signaling pathways in Arabidopsis. Plant Physiology, 133, 1272-1284.
Chinchilla, D., Shan, L., He, P., de Vries, S. and Kemmerling, B. (2009). One for all: the
receptor-associated kinase BAK1. Trends in Plant Science, 14, 535-541.
Choi, J., Tanaka, K., Cao, Y., Qi, Y., Qiu, J., Liang, Y., Lee, S.Y. and Stacey, G. (2014). Identification of a
plant receptor for extracellular ATP. Science, 343, 290-294.
Coll, N.S., Epple, P. and Dangl, J.L. (2011). Programmed cell death in the plant immune system. Cell
Death and Differentiation, 18, 1247-1256.
Dickman, M.B. and de Figueiredo, P. (2013). Death be not proud-cell death control in plant fungal
interactions. PLoS Pathogens, 9, e1003542.
Dickman, M.B. and Fluhr, R. (2013). Centrality of host cell death in plant-microbe interactions. Annual
Review of Phytopathology, 51, 543-570.
Gabriëls, S.H.E.J., Takken, F.L.W., Vossen, J.H., De Jong, C.F., Liu, Q., Turk, S.C.H.J., Wachowski,
L.K., Peters, J., Witsenboer, H.M.A., de Wit, P.J.G.M. and Joosten, M.H.A.J. (2006). cDNA-AFLP
combined with functional analysis reveals novel genes involved in the hypersensitive response.
Molecular Plant-Microbe Interactions, 19, 567-576.
Gabriëls, S.H.E.J., Vossen, J.H., Ekengren, S.K., Ooijen, G.V., Abd-El-Haliem, A.M., Berg, G.C.M.V.D.,
Rainey, D.Y., Martin, G.B., Takken, F.L.W., de Wit, P.J.G.M. and Joosten, M.H.A.J. (2007). An
NB-LRR protein required for HR signalling mediated by both extra- and intracellular resistance
proteins. The Plant Journal, 50, 14-28.
Gao, M., Wang, X., Wang, D., Xu, F., Ding, X., Zhang, Z., Bi, D., Cheng, Y., Chen, S., Li, X. and Zhang,
Y. (2009). Regulation of cell death and innate immunity by two receptor-like kinases in Arabidopsis.
Cell Host & Microbe 6, 34-44.
Genger, R.K., Jurkowski, G.I., McDowell, J.M., Lu, H., Jung, H.W., Greenberg, J.T. and Bent, A.F.
(2008). Signaling pathways that regulate the enhanced disease resistance of Arabidopsis “Defense,
No Death” mutants. Molecular Plant-Microbe Interactions, 21, 1285-1296.
Gomez-Gomez, L., Bauer, Z. and Boller, T. (2001). Both the extracellular leucine-rich repeat domain
and the kinase activity of FLS2 are required for flagellin binding and signaling in Arabidopsis. Plant
Cell, 13, 1155-1163.
Haweker, H., Rips, S., Koiwa, H., Salomon, S., Saijo, Y., Chinchilla, D., Robatzek, S. and von
Schaewen, A. (2010). Pattern recognition receptors require N-glycosylation to mediate plant
immunity. Journal of Biological Chemistry, 285, 4629-4636.
Herve, C., Serres, J., Dabos, P., Canut, H., Barre, A., Rouge, P. and Lescure, B. (1999).
Characterization of the Arabidopsis lecRK-a genes: members of a superfamily encoding putative
receptors with an extracellular domain homologous to legume lectins. Plant Molecular Biology, 39,
671-682.
Huang, P.Y., Yeh, Y.H., Liu, A.C., Chien, C.P. and Zimmerli, L. (2014). The Arabidopsis LecRK-VI.2
associates with the pattern-recognition receptor FLS2 and primes Nicotiana benthamiana
pattern-triggered immunity. The Plant Journal, 79, 243–255..
Kang, J., Hwang, J.U., Lee, M., Kim, Y.Y., Assmann, S.M., Martinoia, E. and Lee, Y. (2010). PDR-type
ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proceedings of the

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
127
National Academy of Sciences of the United States of America, 107, 2355-2360.
Kornev, A.P., Haste, N.M., Taylor, S.S. and Ten Eyck, L.F. (2006). Surface comparison of active and
inactive protein kinases identifies a conserved activation mechanism. Proceedings of the National
Academy of Sciences of the United States of America, 103, 17783-17788.
Lee, M., Lee, K., Lee, J., Noh, E.W. and Lee, Y. (2005). AtPDR12 contributes to lead resistance in
Arabidopsis. Plant Physiology, 138, 827-836.
Liebrand, T.W.H., Smit, P., Abd-El-Haliem, A., de Jonge, R., Cordewener, J.H.G., America, A.H.P.,
Sklenar, J., Jones, A.M.E., Robatzek, S., Thomma, B.P.H.J., Tameling, W.I.L. and Joosten, M.H.A.J.
(2012). Endoplasmic reticulum-quality control chaperones facilitate the biogenesis of Cf
receptor-like proteins involved in pathogen resistance of tomato. Plant Physiology, 159, 1819-1833.
Liebrand, T.W.H., van den Berg, G.C.M., Zhang, Z., Smit, P., Cordewener, J.H.G., America, A.H.P.,
Sklenar, J., Jones, A.M.E., Tameling, W.I.L., Robatzek, S., Thomma, B.P.H.J. and Joosten, M.H.A.J.
(2013). Receptor-like kinase SOBIR1/EVR interacts with receptor-like proteins in plant immunity
against fungal infection. Proceedings of the National Academy of Sciences of the United States of
America, 110, 13228-13228.
Liebrand, T.W.H., van den Burg, H.A. and Joosten, M.H.A.J. (2014). Two for all: Receptor-associated
kinases SOBIR1 and BAK1. Trends in Plant Science, 19, 123-132.
Macho, A.P. and Zipfel, C. (2014). Plant PRRs and the activation of innate immune signaling. Molecular
Cell, 54, 263-272.
Meng, X.Z. and Zhang, S.Q. (2013). MAPK cascades in plant disease resistance signaling. Annual
Review of Phytopathology, 51, 245-266.
Morillo, S.A. and Tax, F.E. (2006). Functional analysis of receptor-like kinases in monocots and dicots.
Current Opinion in Plant Biology, 9, 460-469.
Mur, L.A., Kenton, P., Lloyd, A.J., Ougham, H. and Prats, E. (2008). The hypersensitive response; the
centenary is upon us but how much do we know? Journal of Experimental Botany, 59, 501–520.
Noh, Y.S. and Amasino, R.M. (1999). Identification of a promoter region responsible for the
senescence-specific expression of SAG12. Plant Molecular Biology, 41, 181-194.
Nolen, B., Taylor, S. and Ghosh, G. (2004). Regulation of protein kinases: Controlling activity through
activation segment conformation. Molecular Cell, 15, 661-675.
Pontier, D., Gan, S.S., Amasino, R.M., Roby, D. and Lam, E. (1999). Markers for hypersensitive
response and senescence show distinct patterns of expression. Plant Molecular Biology, 39,
1243-1255.
Rea, P.A. (2007). Plant ATP-binding cassette transporters. Annual Review of Plant Biology, 58,
347-375.
Ren, D., Yang, K.Y., Li, G.J., Liu, Y. and Zhang, S. (2006). Activation of Ntf4, a tobacco
mitogen-activated protein kinase, during plant defense response and its involvement in
hypersensitive response-like cell death. Plant Physiology, 141, 1482-1493.
Roetschi, A., Si-Ammour, A., Belbahri, L., Mauch, F. and Mauch-Mani, B. (2001). Characterization of an
Arabidopsis-Phytophthora pathosystem: Resistance requires a functional PAD2 gene and is
independent of salicylic acid, ethylene and jasmonic acid signalling. The Plant Journal, 28, 293-305.
Rusterucci, C., Aviv, D.H., Holt, B.F., Dangl, J.L. and Parker, J.E. (2001). The disease resistance
signaling components EDS1 and PAD4 are essential regulators of the cell death pathway controlled
by LSD1 in Arabidopsis. Plant Cell, 13, 2211-2224.
Schlaeppi, K., Abou-Mansour, E., Buchala, A. and Mauch, F. (2010). Disease resistance of Arabidopsis
to Phytophthora brassicae is established by the sequential action of indole glucosinolates and
camalexin. The Plant Journal, 62, 840-851.

Chapter IV
128
Schwessinger, B., Roux, M., Kadota, Y., Ntoukakis, V., Sklenar, J., Jones, A. and Zipfel, C. (2011).
Phosphorylation-dependent differential regulation of plant growth, cell death, and innate immunity
by the regulatory receptor-like kinase BAK1. PLoS Genetics, 7, e1002046.
Schwessinger, B. and Ronald, P.C. (2012). Plant innate immunity: Perception of conserved microbial
signatures. Annual Review of Plant Biology, 63, 451-482.
Segonzac, C., Feike, D., Gimenez-Ibanez, S., Hann, D.R., Zipfel, C. and Rathjen, J.P. (2011).
Hierarchy and roles of pathogen-associated molecular pattern-induced responses in Nicotiana
benthamiana. Plant Physiology, 156, 687-699.
Seo, S., Gomi, K., Kaku, H., Abe, H., Seto, H., Nakatsu, S., Neya, M., Kobayashi, M., Nakaho, K.,
Ichinose, Y., Mitsuhara, I. and Ohashi, Y. (2012). Identification of natural diterpenes that inhibit
bacterial wilt disease in tobacco, tomato and Arabidopsis. Plant & Cell Physiology, 53, 1432-1444.
Shiu, S.H. and Bleecker, A.B. (2001). Plant receptor-like kinase gene family: diversity, function, and
signaling. Science Signaling, 2001, re22.
Singh, P., Kuo, Y.-C., Mishra, S., Tsai, C.H., Chien, C.C., Chen, C.W., Desclos-Theveniau, M., Chu,
P.W., Schulze, B., Chinchilla, D., Boller, T. and Zimmerli, L. (2012). The lectin receptor kinase-VI.2 is
required for priming and positively regulates Arabidopsis pattern-triggered immunity. Plant Cell, 24,
1256-1270.
Singh, P., Chien, C.C., Mishra, S., Tsai, C.H. and Zimmerli, L. (2013). The Arabidopsis LECTIN
RECEPTOR KINASE-VI.2 is a functional protein kinase and is dispensable for basal resistance to
Botrytis cinerea. Plant Signaling & Behavior, 8, 1-3.
Spoel, S.H. and Dong, X. (2012). How do plants achieve immunity? Defence without specialized
immune cells. Nature Reviews Immunology, 12, 89-100.
Stukkens, Y., Bultreys, A., Grec, S., Trombik, T., Vanham, D. and Boutry, M. (2005). NpPDR1, a
pleiotropic drug resistance-type ATP-binding cassette transporter from Nicotiana plumbaginifolia,
plays a major role in plant pathogen defense. Plant Physiology, 139, 341-352.
Sun, W.X., Cao, Y.R., Labby, K.J., Bittel, P., Boller, T. and Bent, A.F. (2012). Probing the Arabidopsis
flagellin receptor: FLS2-FLS2 association and the contributions of specific domains to signaling
function. Plant Cell, 24, 1096-1113.
Sun, Y., Li, L., Macho, A.P., Han, Z., Hu, Z., Zipfel, C., Zhou, J.-M. and Chai, J. (2013). Structural basis
for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex. Science, 342,
624-628.
Takahashi, Y., Bin Nasir, K.H., Ito, A., Kanzaki, H., Matsumura, H., Saitoh, H., Fujisawa, S., Kamoun, S.
and Terauchi, R. (2007). A high-throughput screen of cell-death-inducing factors in Nicotiana
benthamiana identifies a novel MAPKK that mediates INF1-induced cell death signaling and
non-host resistance to Pseudomonas cichorii. The Plant Journal, 49, 1030-1040.
Takahashi, H., Kai, A., Yamashita, M., Ando, S., Sekine, K.T., Kanayama, Y. and Tomita, H. (2012).
Cyclic nucleotide-gated ion channel-mediated cell death may not be critical for R gene-conferred
resistance to Cucumber mosaic virus in Arabidopsis thaliana. Physiological and Molecular Plant
Pathology, 79, 40-48.
van Breusegem, F. and Dat, J.F. (2006). Reactive oxygen species in plant cell death. Plant Physiology,
141, 384-390.
van Damme, M., Bozkurt, T.O., Cakir, C., Schornack, S., Sklenar, J., Jones, A.M.E. and Kamoun, S.
(2012). The Irish Potato Famine pathogen Phytophthora infestans translocates the CRN8 kinase
into host plant cells. PLoS Pathogens, 8, e1002875.
van den Brule, S. and Smart, C.C. (2002). The plant PDR family of ABC transporters. Planta, 216,
95-106.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
129
Vleeshouwers, V.G.A.A., van Dooijeweert, W., Keizer, L.C.P., Sijpkes, L., Govers, F. and Colon, L.T.
(1999). A laboratory assay for Phytophthora infestans resistance in various Solanum species
reflects the field situation. European Journal of Plant Pathology, 105, 241-250.
Wang, Y., Meng, Y., Zhang, M., Tong, X., Wang, Q., Sun, Y., Quan, J., Govers, F. and Shan, W. (2011).
Infection of Arabidopsis thaliana by Phytophthora parasitica and identification of variation in host
specificity. Molecular Plant Pathology, 12, 187-201.
Wang, Y., Bouwmeester, K., van de Mortel, J.E., Shan, W. and Govers, F. (2013a). A novel
Arabidopsis-oomycete pathosystem: Differential interactions with Phytophthora capsici reveal a role
for camalexin, indole glucosinolates and salicylic acid in defence. Plant, Cell & Environment, 36,
1192-1203.
Wang, Y., Bouwmeester, K., van de Mortel, J.E., Shan, W. and Govers, F. (2013b). Induced expression
of defense-related genes in Arabidopsis upon infection with Phytophthora capsici. Plant Signaling &
Behavior, 8, e24618.
Wang, Y., Bouwmeester, K., Beseh, P., Shan, W. and Govers, F. (2014). Phenotypic analyses of
Arabidopsis T-DNA insertion lines and expression profiling reveal that multiple L-type lectin receptor
kinases are involved in plant immunity. Molecular Plant-Microbe Interactions, 27, 1390-1402.
Winter, D., Vinegar, B., Nahal, H., Ammar, R., Wilson, G.V. and Provart, N.J. (2007). An "Electronic
Fluorescent Pictograph" browser for exploring and analyzing large-scale biological data sets. PLoS
One, 2, e718.
Yoshioka, H., Numata, N., Nakajima, K., Katou, S., Kawakita, K., Rowland, O., Jones, J.D.G. and Doke,
N. (2003). Nicotiana benthamiana gp91(phox) homologs NbrbohA and NbrbohB participate in H2O2
accumulation and resistance to Phytophthora infestans. Plant Cell, 15, 706-718.
Yu, I.C., Parker, J. and Bent, A.F. (1998). Gene-for-gene disease resistance without the hypersensitive
response in Arabidopsis dnd1 mutant. Proceedings of the National Academy of Sciences of the
United States of America, 95, 7819-7824.
Zhang, S. and Liu, Y. (2001). Activation of salicylic acid-induced protein kinase, a mitogen-activated
protein kinase, induces multiple defense responses in tobacco. Plant Cell, 13, 1877-1889.
Zhang, X., Henriques, R., Lin, S.S., Niu, Q.W. and Chua, N.H. (2006). Agrobacterium-mediated
transformation of Arabidopsis thaliana using the floral dip method. Nature Protocols, 1, 641-646.
Zhou, C., Cai, Z., Guo, Y. and Gan, S. (2009). An Arabidopsis mitogen-activated protein kinase
cascade, MKK9-MPK6, plays a role in leaf senescence. Plant Physiology, 150, 167-177.

Chapter IV
130
SUPPLEMENTAL INFORMATION Table S1. Primers used in this study.
Primer name 5'-3' Used to / for
IX.1-P1-F CACCATGGCCAACTCAATTCTG Detect T-DNA insertion in lecrk-IX.1
IX.1-P1-R TAACCAAATGTTCCTGCTAACC Detect T-DNA insertion in lecrk-IX.1
IX.1-P2-F CAAGGCGAGTAATGTGATGCT qRT-PCR
IX.1-P2-R TAACCAAATGTTCCTGCTAACC qRT-PCR
LecRK-IX.1-FL-F CACCATGGCCAACTCAATTCTG Clone Arabidopsis LecRK-IX.1
LecRK-IX.1-R ACGACCATGTTGAGCACTTG Clone Arabidopsis LecRK-IX.1
LecRK-IX.1(RD/RN)-R CGCCTTGATGTTTCTGTGTACAAC Generate Arabidopsis LecRK-IX.1 RD/RN mutant
LecRK-IX.1(RD/RN)-F GTACACAGAAACATCAAGGCG Generate Arabidopsis LecRK-IX.1 RD/RN mutant
LecRK-IX.1(RD/AA)-R CCTTGATTGCTGCGTGTACAAC Generate Arabidopsis LecRK-IX.1 RD/AA mutant
LecRK-IX.1(RD/AA)-F TGTACACGCAGCAATCAAGGC Generate Arabidopsis LecRK-IX.1 RD/AA mutant
LecRK-IX.1-SP(+TM)-R TCCAGGCTTGAGACAAAAG Generate LecRK-IX.1-∆lectin
LecRK-IX.1-(SP+)TM-F CTTTTGTCTCAAGCCTGGAGC Generate LecRK-IX.1-∆lectin
IX.2-P1-F TTGGAATGCAAGCTCTCACAG Detect T-DNA insertion in lecrk-IX.2
IX.2-P1-R GGCAGCTATAAACCCGAACAT Detect T-DNA insertion in lecrk-IX.2; qRT-PCR
IX.2-P2-F GGCTCGGCTAATGAACCATG qRT-PCR
IX.2-P2-R CGTTCCAGCGATTTTCTCCC qRT-PCR
LecRK-IX.2-FL-F CACCATGCTTTATTTCATTTTCTG Clone Arabidopsis LecRK-IX.2
LecRK-IX.2-R ACGACCATACTCGATGCCTGAG Clone Arabidopsis LecRK-IX.2
LecRK-IX.2(RD/RN)-R GCCTTGATGTTTCTGTGCAGT Generate Arabidopsis LecRK-IX.2 RD/RN mutant
LecRK-IX.2(RD/RN)-F ACTGCACAGAAACATCAAGGC Generate Arabidopsis LecRK-IX.2 RD/RN mutant
LecRK-IX.2(RD/AA)-R CTTGATTGCTGCGTGCAGTAC Generate Arabidopsis LecRK-IX.2 RD/AA mutant
LecRK-IX.2(RD/AA)-F ACTGCACGCAGCAATCAAGGC Generate Arabidopsis LecRK-IX.2 RD/AA mutant
LecRK-IX.2-SP(+TM)-R ACTTGAAACAACAAAAGGGAGG Generate LecRK-IX.2-∆lectin
LecRK-IX.2-(SP+)TM-F TTGTTGTTTCAAGTTTGGACAGCG Generate LecRK-IX.2-∆lectin
PDR12-FL-F CACCATGGAGGGAACTAGTTTTC Clone Arabidopsis ABCG40
PDR12-R TCGTTTTTGGAAATTGAAACTC Clone Arabidopsis ABCG40; semi-qRT-PCR
PDR12-RT-F GAAGCGGCTTTAGGAGTCGATTTCGC Arabidopsis ABCG40 qRT-PCR; semi-qRT-PCR
PDR12-RT-R CGTCCACTCGAATCCTATCATAGCG Arabidopsis ABCG40 qRT-PCR
Actin 2-RT-F TAACTCTCCCGCTATGTATGTCGC Arabidopsis Actin2 qRT-PCR; semi-qRT-PCR
Actin 2-RT-R GAGAGAAACCCTCGTAGATTGGC Arabidopsis Actin2 qRT-PCR
Actin 2-SemiRT-R GAGGGCTGGAACAAGACTTCTG Arabidopsis Actin2 semi-qRT-PCR
AtRub-F4 GCAAGTGTTGGGTTCAAAGCTGGTG Arabidopsis Rubisco qRT-PCR
AtRub-R4 CCAGGTTGAGGAGTTACTCGGAATGCTG Arabidopsis Rubisco qRT-PCR
Nbactin-RT-F TATGGAAACATTGTGCTCAGTGG N. benthamiana Actin qRT-PCR
Nbactin-RT-R CCAGATTCGTCATACTCTGCC N. benthamiana Actin qRT-PCR
NbSIPK1-VIGS-F GAATTCCAAGATTGATGCCAAGAGG Clone fragment for NbSIPK/NTF4 silencing

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
131
NbSIPK1-VIGS-R GAGCTCGCAATTATTCACATGTGCTGGT Clone fragment for NbSIPK/NTF4 silencing
NbSIPK1/NTF4-RT-F GGTGGCAGGTTCATTCAATACAAT NbSIPK/NTF4 qRT-PCR
NbSIPK1/NTF4-RT-R CTTCTCTCTGTGGTGGTGGTAT NbSIPK/NTF4 qRT-PCR
NbNTF4-RT-F CACAGCAACCAGCTCCTCCTC NbNTF4 qRT-PCR
NbNTF4-RT-R TGCCATAAGCTCCTTTACCG NbNTF4 qRT-PCR
NbSIPK1-RT-F GGAGCAGCCACCTCCGGCGG NbSIPK qRT-PCR
NbSIPK1-RT-R GTAAGCACCTCTACCAATAG NbSIPK qRT-PCR
CAP-Fw TTTAGTTGGGGGTCTTGTACC Detect P. capsici biomass
CAP-Rv1 CCTCCACAACCAGCAACA Detect P. capsici ITS amplification
Oligo-dT GACTCGAGTCGACATCGATTTTTTTTTTTTTTT cDNA synthesis

Chapter IV
132
Table S2. Peptides matching NbPDR1 that were identified using LC-MS.
Input Output
Annotation DDA MSE
peptidea No. peptidea No.
LecR
K-I
X.1
-GF
P
NbPDR1
TVVCTIHQPSIDIFEAFDELFLMK 1 AANIKPDADIDMFMK 1
TTLMDVLAGR 1 ALFMDEISTGLDSSTTYSIVNSLK 1
TTLLLALAGK 1 EIYVGPLGR 1
MMFVEEVMDLVELTPLR 1 EMTVR 1
AANIKPDADIDMFMK 1 FEHLNIDADAYVGSR 1
FIFTTFIALIFGTMFWDIGTK 1
GAADFLQEVTSK 1
GGQEIYVGPLGR 1
IGEMTVR 1
LLLALAGK 1
LMGVSGAGK 1
MPRDTAEDGGIYSGALFFVVIMIMFNGLSELPMTLYK 1
NGLSTEQR 1
SALVGLPGVNGLSTEQR 1
SGAFRPGVLTALMGVSGAGK 1
SVSGAFRPGVLTALMGVSGAGK 1
TATYISQHDLHIGEMTVR 2
TTLLLALAGK 1
TVVCTIHQPSIDIFEAFDELFLMK 1
VCTERELLLMQR 2
LecR
K-I
X.2
-GF
P
NbPDR1
TTLLLALAGK 1
TTLMDVLAGR 1
a Peptides that are not unique to NbPDR1 are in grey.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
133
Figure S1. Disease symptoms on Arabidopsis LecRK mutants upon infection by various
pathogens.
A, Disease symptoms on Col-0, lecrk-IX.1 and lecrk-IX.2 upon inoculation with Phytophthora brassicae
HH and Phytophthora capsici LT123 four days post inoculation (dpi), Alternaria brassicicola
MUCL20297 at 5 dpi and Pseudomonas syringae (Pst) DC3000 at 4 dpi. White arrowheads point to
inoculated leaves. B, Bacterial growth in Col-0, lecrk-IX.1 and lecrk-IX.2 at 3 dpi. Bars depict mean
bacterial titers of four plants (± SE). Experiments were repeated three times with similar results.

Chapter IV
134
Figure S2. Infection assays with P. capsici LT263 on transgenic lines expressing LecRK-IX.1,
LecRK-IX.2 or derivatives.
A, Disease symptoms on Col-0 and different transgenic lines at 3 dpi. White arrowheads point to
inoculated leaves. B, Relative quantification of P. capsici biomass in Col-0 and different transgenic lines
using qPCR. Bars represent mean values (± SD) relative to that detected in infected Col-0 leaves. *
indicates significant difference in biomass (p < 0.05) compared to Col-0 according to a t test.
Experiments were repeated twice with similar results.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
135
Figure S3. Disease symptoms on Col-0 and transgenic lines expressing GFP, LecRK-IX.1,
LecRK-IX.2 or derivatives upon inoculation with P. brassicae CBS686.95 at 3 dpi.

Chapter IV
136
Figure S4. Alignment of protein sequences of LecRK-IX.1 and LecRK-IX.2 using ClustalW.
Identical amino acids are shaded in black. The N-terminal lectin domain is depicted by a blue bar, the
predicated transmembrane domain by a yellow bar and the kinase domain by a green bar. The RD
motif in the catalytic loop is marked with asterisks while predicted N-glycosylation sites according to
NetNGlyc 1.0 Server are highlighted in red.

LecRK-IX.1 and LecRK-IX.2 function in Phytophthora resistance and cell death induction
137
Figure S5. Cell death induced by LecRK-IX.1, LecRK-IX.2 and NPP1 on TRV-treated
N. benthamiana leaves.
N. benthamiana leaves treated with TRV:GUS TRV:NbEDS1, TRV:NbMEK2, TRV:NbNPR1,
TRV:NbNRC1, TRV:NbRbohA/B and TRV:NbSIPK/NTF4 were agro-infiltrated to express GFP, NPP1,
LecRK-IX.1 or LecRK-IX.2. Pictures were taken three days after agro-infiltration.


Chapter V
Ectopic expression of Arabidopsis L-type lectin
receptor kinase genes LecRK-I.9 and LecRK-IX.1
confers Phytophthora resistance in
Nicotiana benthamiana
Yan Wang, Klaas Bouwmeester, David L. Nsibo, Hagos M. Juhar, Francine Govers

Arabidopsis LecRK-I.9 and LecRK-IX.1 confer Phytophthora resistance in N. benthamiana
140
ABSTRACT
L-type lectin receptor kinases LecRK-I.9 and LecRK-IX.1 were previously found to
function as Phytophthora resistance components in Arabidopsis. In this study, we
checked the role of these two LecRKs in Phytophthora resistance when transferred
into the Solanaceous plant Nicotiana benthamiana. Multiple transgenic lines
expressing AtLecRK-I.9 or AtLecRK-IX.1 were generated. Molecular analyses
revealed that the obtained transgenic lines vary in transgene copy number, transgene
expression level and protein accumulation. Infection assays showed that transgenic
N. benthamiana plants expressing either AtLecRK-I.9 or AtLecRK-IX.1 are more
resistant to Phytophthora capsici and Phytophthora infestans. These results
demonstrate that Arabidopsis LecRK-I.9 and LecRK-IX.1 retained their Phytophthora
resistance function after transfer into N. benthamiana.

Chapter V
141
INTRODUCTION
Plant diseases caused by Phytophthora species are widespread and cause enormous
economic losses on a large variety of crops (Tyler, 2007; Fry, 2008; Lamour et al.,
2012). Under favorable conditions, Phytophthora pathogens reproduce rapidly and
become epidemic within a short period of time. Phytophthora disease control is costly
and often depends on excessive application of preventive fungicides. Hence,
development of plant cultivars with durable resistance against different Phytophthora
species is under high demand. Currently, breeding programs are mainly focused on
the exploitation of cytoplasmic resistance (R) genes that encode intracellular
nucleotide-binding leucine-rich repeat (NLR) proteins to restrict Phytophthora
pathogens (Vleeshouwers et al., 2011). However, these attempts are often hampered
by the quick adaptation of Phytophthora pathogens that leads to evasion of the
R-gene mediated recognition (Fry, 2008; Vleeshouwers et al., 2011; Rodewald and
Trognitz, 2013).
Plants also respond to pathogens by activation of plasma membrane-localized
receptors (Zipfel, 2014). Recently, L-type lectin receptor kinases (LecRKs) have been
found to play a role in plant resistance to different Phytophthora pathogens. One of
the Arabidopsis LecRKs, i.e. LecRK-I.9, was identified as a potential host target of the
Phytophthora infestans RXLR effector IPI-O and function as a Phytophthora
resistance component (Bouwmeester et al., 2011). LecRK-I.9 belongs to a family of 45
members in Arabidopsis (Bouwmeester and Govers, 2009). To unravel the function of
other LecRKs, we previously carried out infection assays on Arabidopsis T-DNA
insertion mutants with different Phytophthora pathogens (Wang et al., 2014/Chapter
III). Mutants of 14 LecRKs showed compromised resistance, indicating that these
LecRKs are also implicated in Phytophthtora resistance. One of these LecRKs,
namely LecRK-IX.1, was analysed in more detail and was confirmed to be a
Phytophthora resistance component in Arabidopsis (Chapter IV).
Engineering plants via interfamily transfer of resistance components has the
potential to improve disease resistance in crop plants. A successful example is the
transfer of Arabidopsis EFR into Solanaceous plants (Lacombe et al., 2010). EFR is a
receptor of bacterial elongation factor EF-TU and is restricted to the Brassicaceae
family (Kunze et al., 2004; Zipfel et al., 2006). Nicotiana benthamiana and tomato
transgenic plants expressing EFR gained the capacity to respond to EF-Tu and
showed enhanced resistance to various bacterial pathogens (Lacombe et al., 2010).
In a similar way, Arabidopsis LecRK-I.9 was shown to confer Phytophthora resistance
in potato (Bouwmeester et al., 2014). Consistently, transient expression of LecRK-I.9
in N. benthamiana also increased resistance to P. infestans. Likewise, Arabidopsis
LecRK-VI.2 maintained its function in bacterial resistance when expressed in

Arabidopsis LecRK-I.9 and LecRK-IX.1 confer Phytophthora resistance in N. benthamiana
142
N. benthamiana (Huang et al., 2014).
The objective of this work was to generate stable transgenic N. benthamiana lines
expressing Arabidopsis LecRK-I.9 or LecRK-IX.1 and to analyse their response to
Phytophthora infection. To this end, transgenic N. benthamiana plants were generated
using Agrobacterium-mediated transformation. The obtained transgenic lines were
subjected to molecular analyses to determine transgene copy number, transgene
expression level and protein accumulation. Thereafter, we monitored the phenotypic
changes of these transgenic N. benthamiana lines in growth and response to different
Phytophthora pathogens.
Figure 1. Flowchart of the generation, selection and characterization of transgenic
N. benthamiana lines harboring Arabidopsis LecRK-I.9 or LecRK-IX.1.
RESULTS AND DISCUSSION
Generation of transgenic N. benthamiana ectopically expressing Arabidopsis
LecRK-I.9 or LecRK-IX.1
Agrobacterium-mediated transformation of N. benthamiana with the binary vectors
pBIN-KS-35S::AtLecRK-I.9-eGFP, pBIN-KS-35S::AtLecRK-IX.1-eGFP and pBIN61-
35S::GFP (Fig. 2A) resulted in 17, 12 and 4 putative T0 kanamycin-resistant lines,
respectively. These were named as I.9-OE-1 to 17, IX.1-OE-1 to 12 and EV-1 to 4
(Table 1). The flowchart in Fig. 1 gives an overview of the various steps in the process
of selection and analysis of the T0 and T1 transgenic lines. For the molecular

Chapter V
143
characterization, we first determined whether AtLecRK-I.9 and AtLecRK-IX.1 were
successfully transferred into N. benthamiana by PCR using gene specific primers (Fig.
2A). Of the selected kanamycin-resistant plants, the four empty vector transgenic
lines (EV-1 to EV-4) and one LecRK-I.9 line, i.e. I.9-OE-7, gave no PCR product (Fig.
2B).
To determine transgene copy number, we performed qPCR analysis. For both
AtLecRK-I.9 and AtLecRK-IX.1, the copy number in transgenic lines ranges from 0 to
5 and this number was not always consistent with that of the calculated NPTII copy
number (Table 1). In the transgenic lines I.9-OE-7, I.9-OE-10, I.9-OE-11, IX.1-OE-2,
IX.1-OE-3, IX.1-OE-5 and IX.1-OE-6, more copies were detected for NPTII than for
LecRKs, indicating the presence of truncated T-DNA fragments. This likely explains
why the T0 line I.9-OE-7 lacks AtLecRK-I.9 but is still kanamycin-resistant.
Expression of AtLecRK-I.9 and AtLecRK-IX.1 in the T0 plants was determined by
quantifying mRNA levels using qRT-PCR. For both AtLecRK-I.9 and AtLecRK-IX.1,
the expression level varies among individual transgenic lines (Fig. 2C). Variations in
transgene expression level in stable transgenic lines have often been attributed to the
site(s) of transgene insertion and transgene copy number (Kole et al., 2010). In T0
plants, however, no correlation was found between copy number and transgene
expression level. Transgene expression level varies among individual transgenic lines
with same transgene copy number. Some of the transgenic lines with a single copy of
the transgene showed even higher expression levels than those with two or more
copies. For example, line I.9-OE-16 contains a single AtLecRK-I.9 copy but showed
the highest transgene expression level of all I.9-OE lines. Even transgenic lines with
the same transgene number, e.g. line IX.1-OE-3 and IX.1-OE-11, showed quite
different expression levels (Fig. 2C). In the two EV lines, different amounts of GFP
with the expected size around 28 kDa were detected using GFP antibody. To
determine whether the transgenic lines produce the LecRK proteins, a subset of
transgenic lines was selected for protein isolation. Among the seven tested I.9-OE
lines, variable amount of LecRK-I.9-eGFP was detected (Fig. 2D). Comparison with
the expression levels suggests that the accumulation of the LecRK-I.9 is correlated
with the transgene expression level in these lines. LecRK-IX.1-eGFP was only
detected in line IX.1-OE-10, the one with the highest transgene expression levels of
the three selected IX.1-OE lines (Fig. 2D).

Arabidopsis LecRK-I.9 and LecRK-IX.1 confer Phytophthora resistance in N. benthamiana
144
Figure 2. Molecular characterization of transgenic N. benthamiana lines.
A, Schematic representation of T-DNA regions of the vectors used for N. benthamiana transformation.
PCR amplified fragments and position of the primers are indicated by the head-to-head arrow pairs.
The fragments F-I.9-1, F-I.9-2, F-IX.1-1, F-IX.1-2 and NPTII-RT were amplified to determine transgene
presence in transgenic lines, while fragments I.9-RT and IX.1-RT were amplified to monitor transgene
mRNA levels. B, Presence of AtLecRK-I.9 or AtLecRK-IX.1 in transgenic N. benthamiana lines.
Genomic DNA from each line was used as template for amplification with primers indicated in (A). C,
Relative quantification of transgene expression levels in transgenic N. benthamiana lines. Transcript
levels were normalized to NbActin and values are expressed as mean fold changes (± SD) relative to

Chapter V
145
the transcript level of AtLecRK-I.9 in I.9-OE-4 or the transcript level of AtLecRK-IX.1 in IX.1-OE-8 which
was arbitrarily set as 1. D, GFP, LecRK-I.9-eGFP and LecRK-IX.1-eGFP accumulation in transgenic
N. benthamiana lines. Proteins were immunodetected using anti-GFP-HRP. Coomassie staining was
used to indicate the amount of loading in each lane.
Table 1. Transgenic N. benthamiana lines vary in transgene copy number and growth.
Line No. Transgene copy number Morphology (transformants vs wild type)a Germination rateb (%)
AtLecRK-I.9/IX.1 NPTII T0/T1 plants T1 progeny
EV
-
1 0 1 − 100
2 0 2 − 100
3 0 5 − 100
4 0 2 − 100
I.9-O
E-
1 1 1 Slightly smaller 32
2 2 2 Smaller plants with curly leaves 60
3 3 3 Smaller plants with compact rosettes 40
4 3 3 − 60
5 1 1 − 100
6 4 4 − 55
7 0 1 − /a
8 1 1 − 60
9 1 1 Smaller 85
10 5 6 Smaller 95
11 4 6 Smaller 65
12 1 1 Smaller plants with compact rosettes, thick leaves 55
13 2 2 − 50
14 2 2 − 70
15 1 1 Smaller 60
16 1 1 Smaller plants with compact rosettes 35
17 3 3 Slightly smaller and curly leaves 85
IX.1
-OE
-
1 2 2 − 100
2 1 2 Smaller plants with curly round leaves 100
3 1 2 Smaller 100
4 2 2 − 100
5 2 3 − 100
6 1 2 − 100
7 1 1 Smaller 100
8 2 2 − 100
9 1 1 − 100
10 1 1 Smaller plants with old leaves showing necrosis 100
11 1 1 Smaller plants with old leaves showing necrosis 100
12 1 1 Smaller plants with old leaves showing necrosis /a a ‘─’ means no difference compared with the wild-type N. benthamiana, ‘/’ not tested. b Percentage of germinated seeds (n=18-24) three days after grown on MS. This experiment was only
performed once.

Arabidopsis LecRK-I.9 and LecRK-IX.1 confer Phytophthora resistance in N. benthamiana
146
Morphology and growth alterations of I.9-OE and IX.1-OE lines
In a previous study, it was reported that overexpression of AtLecRK-I.9 in Arabidopsis
led to more compact rosettes with smaller and slightly wrinkled leaves (Bouwmeester
et al., 2011). In line with this, ectopic expression of AtLecRK-I.9 in potato also led to
developmental defects, such as aberrant leaf morphology (Bouwmeester et al., 2014).
In this study, the T0 transgenic N. benthamiana plants were monitored for growth and
developmental alterations starting from one-week after transfer into soil until seed set
(Table 1; Fig. 1). All the four EV lines showed normal development; i.e. leaf
morphology, branching and plant height were similar to untransformed
N. benthamiana. In contrast, several I.9-OE lines were smaller in size and displayed
more compact rosettes or curly leaves. These phenotypic changes, however, were not
correlated with AtLecRK-I.9 expression levels, but this could be due to a combined
effect of varying transgene copy numbers, transgene insertion sites and transgene
expression levels in the different transgenic lines. Three out of the twelve IX.1-OE
lines, namely IX.1-OE-10, IX.1-OE-11 and IX.1-OE-12, showed spontaneous cell
death in leaves of over five-week-old plants (Fig. 3A). Considering that these three
IX.1-OE lines showed much higher AtLecRK-IX.1 expression levels than the rest, the
cell death phenotype might be attributed to the elevated AtLecRK-IX.1 expression.
The cell death phenotype was also found when AtLecRK-IX.1 was overexpressed in
Arabidopsis and is correlated with AtLecRK-IX.1 expression levels (Chapter IV). To
determine whether the capacity of LecRK-IX.1 in inducing cell death can be
maintained in the offspring, line IX.1-OE-10 was propagated and the progeny plants
(T1) were assayed for both the presence of AtLecRK-IX.1 and development of
spontaneous cell death. Cell death was only found in the T1 plants harboring
AtLecRK-IX.1 (Fig. 3B). These plants also showed retarded growth when compared to
those without AtLecRK-IX.1 (Fig. 3C). Taken together, these results indicate that
AtLecRK-IX.1 plays a role in mediating plant cell death which could hamper plant
growth.
For all the EV and IX.1-OE lines, seeds harvested from T0 plants have a similar
germination efficiency as untransformed N. benthamiana when grown in soil or on MS
(Murashige and Skoog) medium (Table 1; Fig. 3D). However, ten out of the sixteen
I.9-OE lines showed severe defects in seed germination, with a germination rate of 60%
or lower (Table 1). Also here, the severity of the phenotype does not correlate with
AtLecRK-I.9 expression levels in the various transgenic lines.

Chapter V
147
Figure 3. Morphology of transgenic N. benthamiana lines.
A, T0 N. benthamiana IX.1-OE-10 plant, but not the T0 EV-2 plant displayed cell death. Ten-week-old
plants were photographed. B, T1 N. benthamiana IX.1-OE-10 plant, but not EV-2 plant developed cell
death. Six-week-old plants were photographed. C, T1 progeny of the IX.1-OE-10 line harboring
AtLecRK-IX.1 (+) is smaller in size than the T1 progeny without AtLecRK-IX.1 (‒). Six-week-old plants
were photographed. D, Germination of seeds harvested from untransformed and transgenic
N. benthamiana plants. Six seeds were sown in each pot. Two-week-old seedlings were photographed.
I.9-OE and IX.1-OE lines show enhanced resistance to Phytophthora pathogens
Both LecRK-I.9 and LecRK-IX.1 were previously shown to function in Phytophthora
resistance in Arabidopsis (Bouwmeester et al., 2011; Chapter IV). In order to
determine whether both LecRKs retain their function in Phytophthora resistance, we
checked the response of N. benthamiana I.9-OE and IX.1-OE lines upon inoculation
with Phytophthora capsici and Phytophthora infestans. Lines I.9-OE-1, I.9-OE-8,
IX.1-OE-3 and IX.1-OE-10 were selected and leaves from T1 progenies harboring the
transgenes were used for infection assays. It has to be noted that for IX.1-OE-10, only
leaves without any visible cell death phenotype were used for infection assays.

Arabidopsis LecRK-I.9 and LecRK-IX.1 confer Phytophthora resistance in N. benthamiana
148
Figure 4. Infection assays on transgenic N. benthamiana lines with different Phytophthora
pathogens.
A-C, Disease symptoms on transgenic N. benthamiana EV, I.9-OE and IX.1-OE lines three days after
plug-inoculation with P. capsici LT263 (A) or LT3239 (B), or six days after zoospore-inoculation with

Chapter V
149
P. infestans 14-3-GFP (C). Lesions are indicated by black circles. D, Average lesion sizes on
N. benthamiana plants upon inoculation with Phytophthora pathogens. Each experiment included
12-20 inoculation sites. Bars represent the mean lesion sizes (± SE). * indicates significantly difference
in lesion sizes (p < 0.05) compared to the EV lines according to a t test. Infection assays were repeated
three times with both P. capsici isolates and twice with P. infestans with similar results.
Upon plug-inoculation with P. capsici LT263 or LT3239, smaller lesions were found on
the I.9-OE and IX.1-OE lines when compared to EV-1 and EV-2 (Fig. 4, A and B),
indicating that constitutive expression of AtLecRK-I.9 or AtLecRK-IX.1 in
N. benthamiana enhances resistance to different isolates of P. capsici. This increased
resistance was also found in I.9-OE and IX.1-OE lines when inoculated with
P. infestans (Fig. 4C). Irrespective of inoculation with either P. capsici or P. infestans,
lesions on IX.1-OE-10 plants with higher AtLecRK-IX.1 expression levels were
smaller than those on IX.1-OE-3 (Fig. 2C and Fig. 4D). However, this is not the case
for I.9-OE plants. Despite a higher LecRK-I.9 expression level, I.9-OE-8 plants
showed slightly larger lesions when compared to I.9-OE-1 upon inoculation with
P. infestans (Fig. 2C and Fig. 4D).
CONCLUSIONS
In this study, multiple transgenic N. benthamiana lines with constitutive expression of
Arabidopsis LecRK-I.9 or LecRK-IX.1 were obtained. Transgenic lines vary in
transgene copy number, transgene expression level and protein accumulation.
Ectopic expression of either AtLecRK-I.9 or AtLecRK-IX.1 in N. benthamiana
increased the resistance to different Phytophthora species. Our results suggest that
Arabidopsis LecRK-I.9 and LecRK-IX.1 maintained their function in Phytophthora
resistance when transferred into N. benthamiana, which is in line with the previous
study on the transfer of LecRK-I.9 into potato. These results also suggest that LecRKs
could be used as a complementary resistance resource next to R genes for
engineering broad-spectrum disease resistance to Phytophthora pathogens. Since
ectopic expression of both LecRKs also caused several adverse effects on plant
fitness, such as curly leaves, leaf necrosis or reduced plant sizes, further analysis is
needed to reduce these pleiotropic effects. These transgenic N. benthamiana lines
can be used as a valuable experimental tool for further analysis of the components
required for LecRK-mediated resistance, for example via the virus-induced gene
silencing or protein complex pull-down assays.

Arabidopsis LecRK-I.9 and LecRK-IX.1 confer Phytophthora resistance in N. benthamiana
150
MATERIALS AND METHODS
Plant growth conditions and infection assays
N. benthamiana seeds were surface-sterilized by 70% ethanol and 1% NaClO, and
grown on MS medium (4.4 g MS salt, 20 g sucrose and 8 g agar) or MS medium
supplemented with antibiotics in a conditioned growth chamber at 19-21°C, with a 16
h photoperiod and a relative humidity of 75-80%. Plants grown in soil were kept in a
greenhouse with similar settings. Supplementary light (100 W m2-) was applied when
the light intensity dropped below 150 W m2-.
P. capsici isolates LT263 and LT3239 were maintained in the dark on V8 plates at
25°C (Wang et al., 2013/Chapter IIa), and Phytophthora infestans isolate 14-3-GFP
on rye sucrose agar at 18°C (Bouwmeester et al., 2014). P. infestans zoospores were
obtained by treating sporulating mycelia with cold water for 3-4 h. For detached-leaf
assays, leaves from five-week-old N. benthamiana plants were collected and the
abaxial sides were inoculated with fresh P. capsici mycelial plugs (Ø 0.5 cm) or 10 µl
droplets of a P. infestans zoospore suspension with a concentration of 5*105
zoospores ml-1. Inoculated leaves were kept in transparent plastic boxes with high
humidity in the dark overnight and thereafter exposed to a condition with a 12 h
photoperiod and appropriate temperature settings. Disease severity was monitored by
measuring lesion sizes as previously described by Vleeshouwers et al. (1999) three
and six days after inoculation with P. capsici and P. infestans, respectively.
Agrobacterium-mediated transformation of N. benthamiana
A. tumefaciens GV3101 carrying the binary vectors pBIN-KS-35S::AtLecRK-I.9-
eGFP, pBIN-KS-35S::AtLecRK-IX.1-eGFP (Chapter IV) and pBIN61-35S::GFP (Fig.
2A) were grown overnight at 28°C in Yeast Extract Broth with appropriate antibiotics.
A. tumefaciens cells were pelleted, resuspended and incubated in MMA induction
medium (10 mM MES, 10 mM MgCl2, 50 μM acetosyringone, pH 5.6) for 3 h.
A. tumefaciens cells were collected by centrifugation and resuspended in MS broth
supplemented with 150 μM acetosyringone. Leaf pieces (2-3 cm2) were cut from
five-week-old N. benthamiana leaves and incubated with A. tumefaciens cells for 30
min. Thereafter, leaf discs were dried on filter paper to remove excess A. tumefaciens
and incubated on regeneration medium consisting of MS salt, 1 mg/L cytokinins
6-BAP (6-benzyl amino purine), 0.1 mg/L auxin NAA (1-naphthaleneacetic acid) and
0.8% agar for two days at 19-21°C. Leaf pieces were transferred every week to fresh
regeneration medium supplemented with 400 mg/L cefotaxime, 200 mg/L vancomycin
and 200 mg/L kanamycin until the appearance of plantlets. The generated plantlets
were transferred to MS medium containing 200 mg/L kanamycin to allow root
development. Upon root generation, plantlets were transferred into soil and kept in

Chapter V
151
greenhouse for seed harvesting.
Transgene detection in transgenic N. benthamiana
Genomic DNA was isolated using CTAB buffer (0.02% CTAB, 100 mM Tris-HCl pH 8.0,
20 mM EDTA pH 8.0, 1.4 M NaCl and 1% PVP) followed by precipitation with
isopropanol. RNA was removed by RNaseA. Transgene presence was checked by
PCR using specific primers for AtLecRK-I.9 and AtLecRK-IX.1 and a primer matching
sequence in the binary vector (Fig. 2A; Supplemental Table S1).
Transgene copy number in T0 transgenic lines was determined according to
Honda et al. (2002). Briefly, qPCR was performed using genomic DNA as template
with specific primers for AtLecRK-I.9, AtLecRK-IX.1, neomycin phosphotransferase II
gene (NPTII) or Nbactin (NbS00011031g0002.1) (Table S1). The copy number was
calculated by normalizing the amplification of AtLecRK-I.9, AtLecRK-IX.1 or NPTII to
Nbactin.
RNA isolation and qRT-PCR
Total RNA was isolated from leaves collected from six-week-old T0 transgenic
N. benthamiana plants with a NucleoSpin RNA plant Kit (Macherey-Nagel) and
thereafter used as template for cDNA synthesis with an oligo-dT primer and a M-MLV
reverse transcriptase kit (Promega). qRT-PCR was performed as previously
described (Wang et al., 2014/Chapter III). Transcript levels of AtLecRK-I.9 and
AtLecRK-IX.1 were normalized to Nbactin.
Protein extraction, immunoprecipitation and western blotting
Leaves collected from six-week-old T0 transgenic N. benthamiana plants were ground
in liquid nitrogen and subsequently incubated for 30 min in an extraction buffer
containing 150 mM NaCl, 50 mM Tris-HCl pH 8.0, 1.0% IGEPAL® CA-630 (Sigma)
and one protease inhibitor cocktail tablet per 50 ml (Roche). The homogenate was
centrifuged at 18,000 rpm for 20 min and the supernatant was incubated with
GFP-trap_A® beads (Chromotek) at 4°C for 1-2 h. After incubation, GFP-beads were
spinned down and washed six times with extraction buffer. Proteins were eluted from
GFP-trap_A® beads by boiling for 5 min, separated on a 8% SDS-PAGE gel and
electroblotted onto PVDF membrane (Bio-Rad). Accumulation of eGFP-tagged
LecRK-I.9 and LecRK-IX.1 was detected by immunoblotting using anti-GFP-HRP
(Miltenyi Biotec). Supersignal West Femto Chemiluminescent Substrate (Thermo
Scientific) was used for signal development. Coomassie brilliant blue staining was
used to indicate the amount of loading.

Arabidopsis LecRK-I.9 and LecRK-IX.1 confer Phytophthora resistance in N. benthamiana
152
ACKNOWLEDGEMENTS
We thank Bert Essenstam from Unifarm for taking care of the transgenic plants. This
project was financially supported by a Wageningen University sandwich-PhD
fellowship (YW), a Huygens scholarship (YW), a VENI grant (KB) from the
Netherlands Organization for Scientific Research (Technology Foundation NWO-STW)
and the Food-for-Thought campaign of the Wageningen University fund.
REFERENCES
Bouwmeester, K. and Govers, F. (2009). Arabidopsis L-type lectin receptor kinases: phylogeny,
classification, and expression profiles. Journal of Experimental Botany, 60, 4383-4396.
Bouwmeester, K., de Sain, M., Weide, R., Gouget, A., Klamer, S., Canut, H. and Govers, F. (2011). The
lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a potential host
target for a RXLR effector. PLoS Pathogens, 7, e1001327.
Bouwmeester, K., Han, M., Blanco-Portales, R., Song, W., Weide, R., Guo, L., van der Vossen, E.A.G.
and Govers, F. (2014). The Arabidopsis lectin receptor kinase LecRK-I.9 enhances resistance to
Phytophthora infestans in Solanaceous plants. Plant Biotechnology Journal, 12, 10-16.
Fry, W. (2008). Phytophthora infestans: the plant (and R gene) destroyer. Molecular Plant Pathology, 9,
385-402.
Honda, M., Muramoto, Y., Kuzuguchi, T., Sawano, S., Machida, M. and Koyama, H. (2002).
Determination of gene copy number and genotype of transgenic Arabidopsis thaliana by competitive
PCR. Journal of Experimental Botany, 53, 1515-1520.
Huang, P.Y., Yeh, Y.H., Liu, A.C., Chien, C.P. and Zimmerli, L. (2014). The Arabidopsis LecRK-VI.2
associates with the pattern-recognition receptor FLS2 and primes Nicotiana benthamiana
pattern-triggered immunity. The Plant Journal, 79, 243–255.
Kole, C., Michler, C., Abbott, A., Hall, T., Kohli, A., Miro, B. and Twyman, R. (2010). Transgene
integration, expression and stability in plants: strategies for improvements. In: Transgenic Crop
Plants, Springer Berlin Heidelberg, 201-237.
Kunze, G., Zipfel, C., Robatzek, S., Niehaus, K., Boller, T. and Felix, G. (2004). The N terminus of
bacterial elongation factor Tu elicits innate immunity in Arabidopsis plants. Plant Cell, 16,
3496-3507.
Lacombe, S., Rougon-Cardoso, A., Sherwood, E., Peeters, N., Dahlbeck, D., van Esse, H.P., Smoker,
M., Rallapalli, G., Thomma, B.P.H.J., Staskawicz, B.J. et al. (2010). Interfamily transfer of a plant
pattern-recognition receptor confers broad-spectrum bacterial resistance. Nature Biotechnology, 28,
365-369.
Lamour, K.H., Stam, R., Jupe, J. and Huitema, E. (2012). The oomycete broad-host-range pathogen
Phytophthora capsici. Molecular Plant Pathology, 13, 329-430.
Rodewald, J. and Trognitz, B. (2013). Solanum resistance genes against Phytophthora infestans and
their corresponding avirulence genes. Molecular Plant Pathology, 14, 740-757.
Tyler, B. M. (2007). Phytophthora sojae: root rot pathogen of soybean and model oomycete. Molecular
Plant Pathology, 8, 1-8.
Vleeshouwers, V.G.A.A., van Dooijeweert, W., Keizer, L.C.P., Sijpkes, L., Govers, F., and Colon, L.T.
(1999). A laboratory assay for Phytophthora infestans resistance in various Solanum species

Chapter V
153
reflects the field situation. European Journal of Plant Pathology, 105, 241-250.
Vleeshouwers, V.G.A.A., Raffaele, S., Vossen, J.H., Champouret, N., Oliva, R., Segretin, M.E.,
Rietman, H., Cano, L.M., Lokossou, A., Kessel, G., Pel, M.A. and Kamoun, S. (2011).
Understanding and exploiting late blight resistance in the age of effectors. Annual Review of
Phytopathology, 49, 507-531.
Wang, Y., Bouwmeester, K., van de Mortel, J. E., Shan, W. and Govers, F. (2013). A novel
Arabidopsis-oomycete pathosystem: differential interactions with Phytophthora capsici reveal a role
for camalexin, indole glucosinolates and salicylic acid in defence. Plant, Cell & Environment, 36,
1192-1203.
Wang, Y., Bouwmeester, K., Beseh, P., Shan, W. and Govers, F. (2014). Phenotypic analyses of
Arabidopsis T-DNA insertion lines and expression profiling reveal that multiple L-type lectin receptor
kinases are involved in plant immunity. Molecular Plant-Microbe Interactions, 27, 1390-1402. Zipfel, C., Kunze, G., Chinchilla, D., Caniard, A., Jones, J.D.G., Boller, T. and Felix, G. (2006).
Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated
transformation. Cell, 125, 749-760.
Zipfel, C. (2014). Plant pattern-recognition receptors. Trends in Immunology, 35, 345-351.

Arabidopsis LecRK-I.9 and LecRK-IX.1 confer Phytophthora resistance in N. benthamiana
154
SUPPLEMENTAL INFORMATION
Table S1. Primers used in this study. Primer Sequence 5'-3' Used to / for
NPTII-RT-F GGAGAGGCTATTCGGCTATG Check the presence of NPTII
NPTII-RT-R TCGTCCTGCAGTTCATTCAG Check the presence of NPTII
Nbactin-F TATGGAAACATTGTGCTCAGTGG Endogenous control
Nbactin-R CCAGATTCGTCATACTCTGCC Endogenous control
Oligo-dT GACTCGAGTCGACATCGATTTTTTTTTTTTTTT cDNA synthesis
pGRAB-F1 CCCACTATCCTTCGCAAGACCCTTCC Check the presence of T-DNA sequence
LecRK-IX.1-RT-F CAAGGCGAGTAATGTGATGCT Check presence of AtLecRK-IX.1; qRT-PCR
LecRK-IX.1-RT-R TAACCAAATGTTCCTGCTAACC qRT-PCR
LecRK-IX.1-F TCAAGCCTGGAGCTAATAG Check the presence of AtLecRK-IX.1
LecRK-IX.1-R ACGACCATGTTGAGCACTTG Check the presence of AtLecRK-IX.1
LecRK-I.9-RT-F TTTGCCAGATTTCTCACCATACAC qRT-PCR
LecRK-I.9-RT-R TCTGTTGACTGCCAAGCGTAAG qRT-PCR
LecRK-I.9-F ATGGCTCGTTGGTTGCTTCAG Check the presence of AtLecRK-I.9
LecRK-I.9-R GCTTTGACATCTCGGTGCAGAAC Check the presence of AtLecRK-I.9

Chapter VI
L-type lectin receptor kinases in
Nicotiana benthamiana and tomato and their role in
Phytophthora resistance
Yan Wang, Klaas Bouwmeester, Rob Weide, Francine Govers

Chapter VI
156
ABSTRACT
Membrane-bound receptors play crucial roles as sentinels of plant immunity against
a large variety of invading microbes. One class of receptors known to be involved in
self/non-self-surveillance and plant resistance comprises the L-type lectin receptor
kinases (LecRKs). Previously, we reported that several Arabidopsis LecRKs play a
role in Phytophthora resistance. In this study we determined whether homologs of
these LecRKs from the Solanaceous plants Nicotiana benthamiana and tomato
(Solanum lycopersicum) play similar roles in defence against Phytophthora. In
genome-wide screenings a total of 38 (Nb)LecRKs was identified in N. benthamiana
and 22 (Sl)LecRKs in tomato, each consisting of both a lectin and a kinase domain.
Phylogenetic analysis revealed that in contrast to Arabidopsis, which has a LecRK
family comprised of nine clades, Solanaceous species have just five of these nine
clades (i.e. IV, VI, VII, VIII and IX), plus one additional clade that lacks Arabidopsis
homologs. Subsequently, several Solanaceous LecRKs were selected for functional
analysis using virus-induced gene silencing. Infection assays with Phytophthora
capsici and Phytophthora infestans on LecRK-silenced plants revealed that
N. benthamiana and tomato homologs in clade IX play a role in Phytophthora
resistance similar to the two Arabidopsis LecRKs in this clade, hence suggesting
conserved functions of clade IX LecRKs across different plant families. This study
provides a first insight into the diversity of Solanaceous LecRKs and their role in plant
immunity, and shows the potential of LecRKs for Phytophthora resistance breeding.

Solanaceous LecRKs and their role in Phytophthora resistance
157
INTRODUCTION
Plant diseases caused by Phytophthora pathogens are a major constraint to the
production of a large variety of Solanaceous crops (Kroon et al., 2012). Renowned
are Phytophthora infestans, the causal agent of late blight disease on potato and
tomato, and Phytophthora capsici which is highly destructive to multiple Solanaceous
crops, including tomato, eggplant and pepper (Fry, 2008; Bouwmeester et al., 2009;
Lamour et al., 2012). Breeding for Phytophthora resistance has been focused largely
on the introgression of resistance (R) genes encoding the intracellular nucleotide-
binding leucine-rich repeat receptors (NLRs), that mediate effector-triggered
immunity (ETI) upon recognition of cognate effectors (Vleeshouwers et al., 2011).
Since Phytophthora pathogens can quickly adapt, NLR-mediated resistance is often
not durable. In the pathogen population new races emerge that have inactivated or
modified effector genes and can thus circumvent R gene recognition (Vleeshouwers
et al., 2011; Kasuga and Gijzen, 2013).
Besides ETI, plants rely on defence mediated by plasma membrane-localized
receptor-like kinases (RLKs), which play pivotal roles in the surveillance of “non-self”
(e.g. microbe-associated molecular patterns; MAMPs) and “modified-self” (e.g.
damage-associated molecular patterns; DAMPs) molecules as exogenous stress
signals to initiate non-race-specific immunity. These so-called pattern recognition
receptors (PRRs) have been suggested to mediate basal defence, which is thought
to have a high potential to confer broad spectrum disease resistance in plants (Boller
and Felix, 2009). So far, PRRs have received limited attention in resistance breeding.
One class of RLKs that has been suggested to function as PRRs in recognition of
stress signals and subsequent initiation of plant defence, is comprised of the L-type
lectin receptor kinases (LecRKs). Arabidopsis has 45 LecRK genes that are
distributed over nine clades (i.e. clade I to IX) and seven singletons, with several
showing induced expression upon pathogen attack and in response to pathogen-
associated elicitors and MAMPs (Bouwmeester and Govers, 2009). One of these is
LecRK-I.9 that functions in maintaining cell wall integrity and plays a crucial role in
Phytophthora resistance in Arabidopsis (Gouget et al., 2006; Bouwmeester et al.,
2011). Interfamily gene transfer of Arabidopsis LecRK-I.9 to the Solanaceous plants
Nicotiana benthamiana and potato conferred enhanced resistance to P. infestans
(Bouwmeester et al., 2014; Chapter V), suggesting a conserved functionality in
stress signal recognition and immunity among plant species. Recently, it was shown
that LecRK-I.9 (alias DORN1) also functions as a receptor of extracellular ATP
(eATP) (Choi et al., 2014). Possibly, eATP is released upon pathogen attack or
wounding and as such may function as a DAMP (Choi et al., 2014). Two other
Arabidopsis LecRKs that have been studied for their roles in plant defence are

Chapter VI
158
LecRK-V.5 and LecRK-VI.2. Both were found to play a role in bacterial resistance by
mediating stomatal immunity (Desclos-Theveniau et al., 2012; Singh et al., 2012).
Ectopic expression of LecRK-VI.2 in N. benthamiana appears to prime MAMP-
mediated defence, and to increase resistance against various hemibiotrophic and
necrotrophic bacterial pathogens (Huang et al., 2014). Another example is
Arabidopsis LecRK-I.8, which is required for proper PR-1 induction upon treatment
with egg-derived elicitors of the cabbage butterfly Pieris brassicae (Gouhier-Darimont
et al, 2013). Recently, we published a study that was aimed at investigating the role
of all Arabidopsis LecRKs in defence against a variety of plant pathogens. This
revealed that next to LecRK-I.9, there are at least 14 other LecRKs that have a
putative role in resistance against Phytophthora pathogens in Arabidopsis (Wang et
al., 2014/Chapter III). Arabidopsis lines with T-DNA insertions in these 14 LecRK
genes showed altered susceptibility when challenged with P. capsici and
Phytophthora brassicae, suggesting that LecRK family members collectively build up
a basal level of Phytophthora resistance.
Analysis of LecRKs for a role in defence in plant species other than Arabidopsis
has so far been limited to one gene, i.e. NbLRK1 from Nicotiana benthamiana
(Kanzaki et al., 2008). NbLRK1 was reported to interact with INF1, an elicitin
secreted by P. infestans, and suggested to play a role in intensifying INF1-induced
cell death (Kanzaki et al., 2008). In addition, several LecRKs were indicated to be
involved in plant defence since they are induced upon treatment with pathogens or
pathogen-derived elicitors. Multiple cucumber (Cucumis sativus) LecRKs were found
to be induced upon infection by P. capsici and Phytophthora melonis (Wu et al.,
2014); and a cotton (Gossypium hirsutum) LecRK, i.e. GhLecRK-2 was shown to be
up-regulated upon treatment with a cell wall-derived fraction of the vascular wilt
fungus Verticillium dahliae (Phillips et al., 2013).
Characterization of defence mechanisms in model plants, such as Arabidopsis,
paves the way to study similar processes in crops (Koornneef and Meinke, 2010).
Disrupting gene homologs encoding proteins with a conserved physiological function
often results in similar phenotypes in different plant species. For example, the role of
the PRR FLS2 in the perception of bacterial flagellin was first discovered in
Arabidopsis, and thereafter found to be largely conserved in various plant lineages
(Gomez-Gomez and Boller, 2000; Hann and Rathjen, 2007; Robatzek et al., 2007;
Takai et al., 2008).
To determine whether LecRKs are functionally conserved in Solanaceous
species, we set out to investigate the function of Solanaceous LecRKs in
Phytophthora resistance. In this study, we performed a genome-wide identification of
LecRKs in N. benthamiana and tomato (Solanum lycopersicum) and analysed the
phylogenetic relationship of these LecRKs with Arabidopsis LecRKs. Subsequently,

Solanaceous LecRKs and their role in Phytophthora resistance
159
several Solanaceous LecRKs were selected for functional analysis using virus-
induced gene silencing and infection assays to pinpoint their role in Phytophthora
resistance.
RESULTS AND DISCUSSION
Identification of LecRKs in N. benthamiana and tomato
LecRKs in N. benthamiana and tomato were identified following the pipeline depicted
in Fig.1. Protein sequences of Arabidopsis LecRKs (AtLecRKs) were used as queries
for BLAST searches against the predicted protein databases of N. benthamiana and
tomato in the Sol Genomic Network (SGN). Presence of both a lectin domain and a
kinase domain was used as the major criterion in the selection of putative LecRKs.
Using reciprocal BLAST searches, we identified 37 (Nb)LecRKs and 22 (Sl)LecRKs
in N. benthamiana and tomato, respectively (Table 1). Corresponding coding DNA
(cDNA) sequences were obtained and in combination with EST data, RNA-seq data
and genomic DNA sequences used for gene model validation. Strikingly, over half of
the NbLecRK gene models are incomplete due to erroneous open reading frame
(ORF) prediction including erroneous prediction of start codon, intron or stop codon.
The coding sequence for NbLRK1, a N. benthamiana LecRK homologous to
AtLecRK-VIII.2 (deposited in GenBank as AB247455.1; Kanzaki et al., 2008), was
found to contain multiple SNPs when compared with RNA-seq data. Hence, we PCR-
amplified and sequenced the cDNA and it was found to match to
NbS00026192g0010.1. cDNA and protein sequences were therefore corrected
accordingly and used for further analysis (Table 1). Making use of the RNA-seq data,
we found another NbLecRK (i.e. NbS00021029g0001.1) of which the encoded
protein was annotated as missing the kinase domain in SGN (Table 1). For the
SlLecRKs, three were found to be incomplete and were corrected according to the
RNA-seq data (Table 1). The obtained LecRK protein sequences including the
revised ones are available upon request and will be included in a supplemental file
when this chapter is submitted for publication.
In Arabidopsis, only six out of the 45 LecRKs contain introns. Five of these
have one intron and the other one contains two introns. To determine whether
Solanaceous LecRKs have introns, cDNA sequences were compared to genomic
DNA sequences. In this way, 12 NbLecRKs were found to contain introns ranging
from one to four per gene (Table 1). By using RNA-seq data as extra check six
NbLecRKs were confirmed to contain one intron, while for the other six, introns could
not be validated due to lack of RNA-seq data. Three out of the 22 SlLecRKs were
confirmed to contain one intron by the RNA-seq data, whereas another three

Chapter VI
160
SlLecRKs were predicted to contain one, four and five introns, respectively (Table 1).
Figure 1. Pipeline depicting the procedures used for identification and analysis of LecRKs in
Nicotiana benthamiana and tomato.
Chromosomal location of tomato LecRKs
To investigate tandem duplication events, we examined the chromosomal distribution
of the obtained SlLecRKs in tomato. For N. benthamiana no such information is
available, so we were only able to summarize information obtained on SlLecRKs.
SlLecRKs are distributed over eight of the 12 tomato chromosomes, i.e. chromosome
1, 2, 3, 4, 5, 7, 9, 10, with the number of LecRKs ranging from 1 to 6 (Table 1). Most
of the SlLecRKs are located on chromosomes 9 and 10, with each chromosome
contains six SlLecRKs. In addition, two tandem duplicated SlLecRK gene clusters
were found on chromosome 9 and one on chromosome 10 based on the criterion
that tandem duplicated genes are located within 10 adjacent gene models (Shiu and
Bleecker, 2003). In comparison, LecRKs in Arabidopsis show a much higher tandem
duplication rate according to the cluster analysis by Bouwmeester and Govers
(2009). Here nine distinct clusters were found, each with two to six AtLecRKs.

Solanaceous LecRKs and their role in Phytophthora resistance
161
Table 1. Inventory of LecRKs of Arabidopsis, Nicotiana benthamiana and tomato.
a,b,c Tandem duplicated LecRKs. d Gray blocks represent clades of Solanaceous LecRKs. e The
number of introns was predicted by comparing cDNA and genomic DNA. f Signal peptide prediction
based on SignalP 3.0. g +: with RD motif, ─: without RD motif due to lack of the VIb subdomain.

Chapter VI
162
Domain composition of NbLecRKs and SlLecRKs
Domain analysis performed using multiple bioinformatics webtools revealed that of
the 38 identified NbLecRKs, 32 contain the typical composition of a LecRK, i.e. a
putative signal peptide (SP), an extracellular L-type lectin domain, a single
transmembrane (TM) domain and a cytosolic serine/threonine kinase domain. Of the
remaining six, three have no clear TM domain, two have no SP and another one
lacks both the SP and the TM domain. Similarly, 21 of the 22 SlLecRKs contain all
representative LecRK features, whereas the other one was predicted to lack both the
SP and the TM domain (Table 1). In addition, the SlLecRK Solyc09g011060.2.1
contains an additional domain, namely a NTP-binding domain following the kinase
domain in the C-terminus. Although NbS00043874g0006.1, Solyc10g047680.1.1 and
Solyc10g047700.1.1 were predicted to contain a lectin signature, the domain
composition is incomplete (Table 1). Kinase (STK) domains of RLKs are in general
highly conserved and contain 12 subdomains which are essential for kinase activity
(Shiu and Bleecker, 2001; Hanks, 2003; Afzal et al., 2008). Alignment of the
predicted kinase domains revealed that six NbLecRKs and three SlLecRKs lack one
or more subdomains, which could impair kinase activity (Table 1). Protein kinases
that are activated by phosphorylation in the activation loop typically carry a
conserved arginine (R) that precedes the catalytic aspartate (D) in subdomain VIb
(Nolen et al., 2004; Kornev et al., 2006). They are therefore known as RD kinases
whereas kinases that lack the RD motif are collectively termed non-RD kinases. It
has been found that activation of non-RD kinases does not require phosphorylation
of the activation loop (Dardick and Ronald, 2006). Nearly all identified Solanaceous
LecRKs contain a RD motif with the exception of three NbLecRKs and two SlLecRKs.
NbC25369236g0004.1 and Solyc10g047700.1.1 do not contain a RD motif due to
absence of the kinase subdomain VIb, whereas NbS00029224g0003.1,
NbS00001395g0006.1 and Solyc03g080060.1.1 have an RD motif substituted by KN
(Table 1). This is also the case for some of the Arabidopsis LecRKs, i.e. AtLecRK-I.2
lacks the kinase subdomain VIb and AtLecRK-III.1 and AtLecRK-III.2 contain GN
residues instead of RD.
Phylogeny of LecRKs in Arabidopsis, N. benthamiana and tomato
In Arabidopsis, LecRKs were divided into nine clades based on the definition that a
clade contains at least two homologs with a minimum of 50% similarity in both the
DNA sequence and the amino acid sequence (Bouwmeester and Govers, 2009).
However, due to high sequence divergence this criterion is not applicable for the
Solanaceous LecRKs (data not shown). Hence, Solanaceous LecRKs were grouped
together if they share over 50% similarity at the amino acid level. In this way, twelve
clades were found by means of pairwise alignment (Table 1), ranging in size from

Solanaceous LecRKs and their role in Phytophthora resistance
163
three to 10 members.
To evaluate the evolutionary relationship among LecRKs from different plant
species, an un-rooted phylogenetic tree containing 43 full length AtLecRKs, 38
NbLecRKs and 22 SlLecRKs was generated using the Neighbor-Joining algorithm
(Fig. 2). The reliability of the phylogram was determined by bootstrap analysis with
10,000 replicates. A comparable consensus tree was obtained using the Maximum-
Likelihood algorithm with the same bootstrap replicates (data not shown). In addition,
a similar phylogenetic analysis was performed using solely the extracellular lectin
domains or the intracellular kinase domains. The resulting tree topologies are overall
in agreement with the one shown in Fig. 2, which was constructed based on the full
length LecRKs (Supplemental Fig. S1).
In the phylogenetic tree, 38 of the 43 AtLecRKs fall into nine distinct clades (I-
IX) and the other five are singletons, which is in agreement with the previous study
by Bouwmeester and Govers (2009). In addition, the degree of similarity in LecRKs
was found to be indicative of the phylogenetic relationship among these three
species (Table 1; Fig. 2). In the tree, most of the NbLecRKs and SlLecRKs group
together with AtLecRKs in the clades IV, VI, VII, VIII.2 and IX and with the five
AtLecRK singletons. In those branches often the AtLecRK clade members or the
AtLecRK singletons group with one SlLecRK and two NbLecRKs in one sub-branch.
In such a sub-branch, the NbLecRKs and SlLecRK are often closer to each other
than to their Arabidopsis counterpart, as expected, and the duplicated number of
NbLecRKs is most probably due to the allo-tetraploid nature of N. benthamiana. In
addition, closely related sub-branches comprising only Solanaceous LecRKs are
observed in the tree, with up to eight SlLecRKs and NbLecRKs, often in a 1:2 ratio.
These sub-branches are most prominent next to those containing the singletons
AtLecRK-S.4 and AtLecRK-S.5 and the two clade IX AtLecRKs. In contrast to these
expansions in numbers on certain branches of the tree, none of the Solanaceous
LecRKs fall into clades I, II, III and V. Similarly, no LecRKs belonging to these four
clades were identified in cucumber (Wu et al., 2014). It should be noted that the
AtLecRKs in clade I, II, III and V exhibit high frequency of tandem duplication
(Bouwmeester and Govers, 2009). In addition, these four LecRK clades appear to be
conserved among plants of the Brassicales order (Nsibo et al., submitted).
Collectively, these indicate a lineage-specific expansion of LecRKs after the split of
the Brassicales. Several LecRKs belonging to these four Arabidopsis-specific clades
were found to be involved in resistance to Phytophthora, Alternaria brassicicola and
bacterial pathogens (Bouwmeester et al., 2011; Wang et al., 2014/Chapter III),
suggesting that this lineage-specific expansion of LecRKs might be important for
Arabidopsis adaptation. Next to the Arabidopsis-specific clades, the phylogenetic
analysis revealed one Solanaceous-specific (Sol-specific) clade and one NbLecRK

Chapter VI
164
that ends up as a singleton (Fig. 2). The three Solanaceous LecRKs carrying a
substitution in the RD motif form a well-supported clade separated from other
LecRKs. The singleton NbS00037263g0008.1 is on a branch in the vicinity of the
cluster comprising AtLecRK clade IX.
Figure 2. Phylogenetic tree of LecRKs from Arabidopsis, N. benthamiana and tomato.
Neighbor-Joining tree based on predicted full length amino acid sequences, including 43 AtLecRKs,
38 NbLecRKs and 22 SlLecRKs. Branches are colored according to the clades delineated by
Bouwmeester and Govers (2009). Numbers above the branches represent the level of clade support
inferred by 10,000 bootstrap replicates.

Solanaceous LecRKs and their role in Phytophthora resistance
165
Arabidopsis clade IX LecRK homologs in N. benthamiana are functional in
Phytophthora resistance
To determine the role of NbLecRKs in Phytophthora resistance we selected
homologs of AtLecRKs that, in a previous study, were identified as putative
resistance components in Arabidopsis (Wang et al., 2014/Chapter III). That study
revealed 14 AtLecRKs implicated in Phytophthora resistance of which seven have
close homologs in N. benthamiana. TRV-based constructs were made to silence
NbLecRKs that are homologous to six of these AtLecRKs, i.e. AtLecRK-VIII.2,
AtLecRK-IX.1/LecRK-IX.2, AtLecRK-S.1, AtLecRK-S.6 and AtLecRK-S.7. Loss of
function of a single gene might fail to cause any phenotypic change as its function
can be either completely or partially complemented by the presence of homologous
genes. To avoid this problem, we made TRV-based constructs to simultaneously
silence several LecRKs with high sequence similarity. As shown in Fig. 3A, seven
TRV constructs were made, of these three were made to target different AtLecRK-
IX.1/LecRK-IX.2 homologs. In all cases, at least 60% reduction in transcript levels of
the target NbLecRKs was observed in the NbLecRK-silenced N. benthamiana plants
when compared to TRV:GUS-treated plants (Fig. 3B). NbLecRK silencing did not
affect plant growth and development as none of the NbLecRK-silenced plants
showed consistent phenotypic changes in terms of plant size, leaf color, leaf
morphology and shoot growth (Supplemental Fig. S2). Upon inoculation with
P. capsici LT263 or P. infestans 14-3-GFP, TRV:NbIX.1b and TRV:NbIX.1c-treated
N. benthamiana leaves showed significantly larger lesions compared to those
observed on TRV:GUS-treated plants (Fig. 3, C and D). The increased susceptibility
of TRV:NbIX.1b- and TRV:NbIX.1c-treated N. benthamiana plants indicates that the
silenced LecRKs are involved in Phytophthora resistance. In contrast, silencing of the
other two NbLecRKs (i.e. NbS00048421g0010.1 and NbS00051756g0005.1) that
group with clade IX but are relatively distant from AtLecRK-IX.1 and AtLecRK-IX.2
(Fig. 2), did not result in increased susceptibility to Phytophthora (Fig. 3C). In
addition, none of the plants treated by TRV:NbVIII.2, TRV:NbS.1 or TRV:NbS.7
showed significant differences in susceptibility to either P. capsici or P. infestans
when compared to TRV:GUS-treated plants (Fig. 3C). TRV:NbS.6-treated plants
though, showed increased susceptibility upon infection by P. infestans 14-3-GFP, but
not when inoculated with P. capsici LT263 (Fig. 3C). It is possible that the activity of
some of these NbLecRKs is simply too weak to prevent Phytophthora infection. This
is exemplified by the fact that TRV:NbS.6-treated plants allow P. infestans 14-3-GFP
to expand but not P. capsici LT263, an isolate with relatively higher virulence (Fig.
3D). Also the sequence divergence between the AtLecRKs and their N. benthamiana
homologs may contribute to functional divergence that we observed.

Chapter VI
166

Solanaceous LecRKs and their role in Phytophthora resistance
167
Figure 3. Response of NbLecRK-silenced N. benthamiana to Phytophthora infection.
A, NbLecRKs targeted by different TRV constructs. The phylogenetic tree was made as described in
Figure 2. B, Relative NbLecRK transcript levels in TRV:GUS- and TRV:NbLecRK-treated
N. benthamiana leaves. Transcript levels were normalized with NbActin and expressed as mean fold
changes (± SD) relative to the transcript level in TRV:GUS-treated leaves which was arbitrarily set as
1. This experiment was repeated four times with similar results. C, Lesion sizes on TRV:GUS- and
TRV:NbLecRK-treated N. benthamiana leaves upon inoculation with Phytophthora capsici and
Phytophthora infestans three days post inoculation (dpi) and 6 dpi, respectively. Bars represent mean
lesion sizes (± SE) of over 20 inoculation sites. * indicates significant difference in lesion sizes (p <
0.05, t test) between TRV:GUS and TRV:NbLecRK-treated plants. This experiment was repeated four
times with similar results. D, Disease symptoms on TRV:GUS-, TRV:NbIX.1- and TRV:NbS.6-treated
N. benthamiana leaves after inoculation with P. capsici at 3 dpi and P. infestans at 6 dpi. Pictures
were taken under daylight and UV light. E-F, Disease symptoms (E) and quantified lesion sizes (F) on
N. benthamiana leaves transiently expressing GFP or NbLecRK-IX.1/2 after inoculation with P. capsici
and P. infestans. Bars represent the mean lesion sizes (± SE) of at least twenty infiltrated
N. benthamiana leaves. * indicates significant difference in lesion sizes (p < 0.05) according to a t test.
This experiment was repeated three times with similar results.
To further investigate the role of NbLecRK-IX.1 homologs in Phytophthora
resistance, we selected NbS00000562g0002.1 for further study and renamed it
NbLecRK-IX.1/2 to emphasize its relation with the two clade IX AtLecRKs. The full
length coding sequence of NbLecRK-IX.1/2 was PCR-amplified and cloned into a
binary vector enabling transient expression in N. benthamiana leaves. Upon
inoculation with P. capsici or P. infestans, N. benthamiana leaves transiently
expressing NbLecRK-IX.1/2 showed much smaller lesions than that growing on

Chapter VI
168
leaves expressing GFP (Fig. 3, E and F). This confirms that NbLecRK-IX.1/2 indeed
functions in resistance to Phytophthora. It should be noted that transient expression
of NbLecRK-IX.1/2 in N. benthamiana did not cause any visible cell death, a
phenomenon that was observed when transiently expressing AtLecRK-IX.1 or
AtLecRK-IX.2 in N. benthamiana (Chapter IV).
NbLecRK silencing does not alter elicitor-induced cell death in N. benthamiana
NbLRK1 was previously found to interact with the P. infestans elicitor INF1 via its
kinase domain (Kanzaki et al., 2008). Silencing of NbLRK1 in N. benthamiana was
reported to compromise INF1-induced cell death and it was proposed to be an
important component of a host receptor complex that recognizes INF1 (Kanzaki et
al., 2008). We found four LecRKs in N. benthamiana that are homologous to NbLRK1
(Table 1). Sequence alignment of the predicted cDNA sequences revealed that three
of these four are likely silenced by the fragment used by Kanzaki et al. (2008)
(termed NbVIII.2-2 in Supplemental Fig. S3) raising the question which of the three
genes is responsible for the compromised cell death that they observed. In this study,
we used another fragment named TRV:NbVIII.2 to simultaneously silence these three
NbLecRKs (Fig. 3A; Supplemental Fig. S3) and confirmed by qRT-PCR that
expression of the three genes is reduced (Fig. 3B). In contrast to Kanzaki et al.
(2008) we found no difference in the appearance of cell death upon transient
expression of inf1 between N. benthamiana plants treated with TRV:NbVIII.2 and
TRV:GUS (Fig. 4). This discrepancy could be due to the fact that Kanzaki et al.
(2008) infiltrated the leaves with INF1 protein whereas we expressed the inf1 gene in
planta by Agrobacterium-mediated transient expression. In the natural situation, INF1
is secreted by P. infestans into the plant apoplast and is assumed to remain in the
apoplastic space (Kamoun, 2006). Hence, it is puzzling how INF1 can interact with
the cytoplasmic kinase domain of NbLRK1 to mediate cell death induction. Kanzaki
et al. (2008) hypothesized that INF1 could be translocated into plant cells via
receptor-mediated endocytosis but they were not able to show interaction between
INF1 and NbLRK1 in planta.
We also investigated whether other NbLecRKs affect cell death triggered by
INF1 and whether NbLecRKs play a role in cell death triggered by two other
Phytophthora elicitors i.e. CRN2 (Crinkling and necrosis-inducing protein) and NPP1
(necrosis-inducing Phytophthora protein 1). Transient expression of inf1, crn2 and
npp1 in NbLecRK-silenced plants resulted in cell death in all cases and no visible
differences in the appearance of cell death were observed when compared with
TRV:GUS-treated leaves (Fig. 4). Apparently, none of the NbLecRKs tested here play
a role in cell death induced by these three elicitors.

Solanaceous LecRKs and their role in Phytophthora resistance
169
Figure 4. Silencing of NbLecRKs does not affect cell death induced by Phytophthora elicitors.
Cell death in TRV:GUS- and TRV:NbLecRK-treated N. benthamiana leaves expressing inf1, crn2 and
npp1. Each experiment included at least six leaves per construct. This experiment was repeated three
times with similar results.
Silencing of the tomato LecRK homolog of AtLecRK-IX.1/LecRK-IX.2 reduces
Phytophthora resistance
To assess the role of tomato LecRKs in Phytophthora resistance, we used the same
strategy as for the NbLecRKs and silenced SlLecRKs that group with the six
AtLecRKs mentioned above. The TRV constructs were designed to silence a single
SlLecRK gene, with the exception of TRV:SlVIII.2 that targets two SlLecRKs

Chapter VI
170
simultaneously, i.e. Solyc10g084250.1.1 and Solyc09g007510.1.1 (Fig. 5A).
Quantitative analysis of transcript levels in TRV-treated plants revealed that
Solyc03g043710.1.1 and Solyc07g065610.1, targeted by TRV:SlIX.1 and TRV:SlS.7,
respectively, are reduced up to 70% in expression in comparison to TRV:GUS-treated
plants. Silencing of the other SlLecRKs was found to be less efficient (Fig. 5B).
Inoculation of TRV:SlIX.1-treated plants with P. capsici LT263 or P. infestans 14-3-
GFP, resulted in significantly larger lesions than those on inoculation of TRV:GUS-
treated plants (Fig. 5, C and D; Supplemental Fig. S4), indicating that the most
closely related tomato homolog of AtLecRK-IX.1 and AtLecRK-IX.2 plays a role in
Phytophthora resistance similar to NbLecRK-IX.1/2, the homolog in N. benthamiana.
In plants treated with silencing constructs targeting the other SlLecRKs, no increased
susceptibility was found, neither to P. infestans nor to P. capsici (Supplemental Fig.
S4).
CONCLUSIONS In total, 38 and 22 LecRKs were identified in the genomes of N. benthamiana and
tomato, respectively. Multiple deposited Solanaceous LecRK gene models were
found to be erroneous due to mis-prediction of ORF and these sequences were
corrected based on the transcriptome data. Domain composition analysis indicated
that most of the identified LecRKs have the typical LecRK structure, but there are
several LecRKs, especially the ones from N. benthamiana, that lack a signal peptide,
a TM domain or both. Phylogenetic analysis revealed that most of the Solanaceous
LecRKs group together with Arabidopsis LecRKs, whereas one clade containing two
NbLecRKs and one SlLecRK seems to be a Solanaceous-specific clade. Functional
analysis of Solanaceous LecRKs using TRV-mediated gene silencing and
Agrobacterium-mediated transient gene expression demonstrated that homologs of
AtLecRK-IX.1/LecRK-IX.2 in both N. benthamiana and tomato function in resistance
to different Phytophthora pathogens. These data demonstrated that the Phytophthora
resistance function of clade IX LecRKs is conserved across different plant species.

Solanaceous LecRKs and their role in Phytophthora resistance
171
Figure 5. Silencing of SlLecRK-IX.1/LecRK-IX.2 compromises Phytophthora resistance in
tomato.
A, SlLecRKs targeted by different TRV constructs. The phylogenetic tree was made as described in
Figure 2. B, Relative SlLecRK transcript levels in TRV:GUS- and TRV:SlLecRK-treated tomato leaves.
Transcript levels were normalized with SlActin and expressed as mean fold changes (± SD) relative to
the transcript level in TRV:GUS-treated leaves which was arbitrarily set as 1. C-D, Disease symptoms
(C) and quantified lesion sizes (D) on TRV:GUS- and TRV:SlIX.1-treated tomato leaves inoculated
with P. capsici LT263 (1*105 zoospores ml-1) and P. infestans 14-3-GFP (5*105 zoospores ml-1). Each
experiment included at least 20 leaves treated for each construct. Bars represent the mean lesion
sizes (± SE). * indicates significant difference (p < 0.05) in lesion sizes between TRV:GUS and
TRV:SlIX.1-treated plants according to a t test. The infection assay with P. capsici was performed once
whereas infection assays with P. infestans were repeated three times with similar results.

Chapter VI
172
MATERIAL AND METHODS
Sequence identification and gene analysis
Protein sequences of Arabidopsis LecRKs analysed by Bouwmeester and Govers
(2009) were retrieved from the TAIR website (http://www.arabidopsis.org). Protein
sequences were used as queries for reciprocal BLAST searches via the Sol Genomic
Network (SGN) website (http://solgenomics.net) against the genomic databases of
N. benthamiana and tomato. Obtained LecRK sequences were further analysed by
comparative analysis using publicly available ESTs, RNA-seq data derived from the
University of Sydney Nicotiana benthamiana Genome Page (http://sydney.edu.au
/science/molecularbioscience/sites/benthamiana/) (Nakasugi et al., 2012; Naim et al.,
2012) and RNA-seq data derived from tomato leaves (Faino, L., personal
communication). NbS00026192g0010.1 was verified by sequencing after
amplification of the entire coding sequence using Pfu DNA polymerase (Promega)
and gene specific primers (Supplemental Table S1). In addition, the retrieved cDNA
sequences were compared with the genomic DNA sequences, followed by manual
validation of the presence of introns and open reading frames. Amino acid sequences
were subjected to the protein domain and motif annotation webtools SMART
(http://smart.embl-heidelberg.de), SignalP 3.0 (http://www.cbs.dtu.dk/services/
SignalP-3.0) and TMHMM 2.0 Server (http://www.cbs.dtu.dk/services/TMHMM).
Predicted kinase domain sequences were aligned by ClustalW and manually
checked for sub-domains according to those defined based on LecRK-VI.2 and
NtCPK5 (Wang et al., 2005; Singh et al., 2013).
Multiple sequence alignment and phylogenetic analysis
Full length protein sequences and sequences comprising the lectin domain or the
kinase domain were aligned with ClustalW using the protein weight matrix GONNET
with a penalty gap opening of 10 and a gap extension of 0.1. The obtained sequence
alignment was subsequently used as input to construct a Neighbor-Joining tree and a
Maximum Likelihood tree with 10,000 bootstrap replicates using the Jones-Taylor-
Thornton substitution model (Hall, 2013) in MEGA 5.1. Branches corresponding to
partitions reproduced in <60% of bootstrap replicates were collapsed in the
phylogenetic trees.
Plasmid construction
The coding sequence of NbS00000562g0002.1 was amplified by PCR using Phusion
DNA polymerase (Thermo) and primers listed in Supplemental Table S1. Purified
PCR fragments were cloned into the pENTR/D-TOPO® vector (Invitrogen) and
verified by sequencing. The correct PCR fragments were subcloned into the binary

Solanaceous LecRKs and their role in Phytophthora resistance
173
vector pSol2095 via LR reaction using Gateway® LR ClonaseTM II (Invitrogen).
Resulting plasmids were transformed into Agrobacterium tumefaciens strain GV3101.
Gene fragments used to generate silencing constructs were synthesized by Eurofins
Genomics (Supplemental file S1), and were subsequently cloned into the gene-
silencing vector pTRV-RNA2. pTRV-RNA2 derivatives and pTRV-RNA1 vectors were
transformed into A. tumefaciens strain GV3101.
Plant growth conditions
N. benthamiana and tomato (cultivar Moneymaker) were grown in soil in a
conditioned greenhouse at 19-21°C with a 16 h photoperiod and at a relative
humidity of 75-78%. Supplementary light (100 W m-2) was applied when the light
intensity dropped below 150 W m-2.
Agro-infiltration and TRV-mediated silencing assays
A. tumefaciens strains carrying binary vectors were grown overnight at 28°C in Yeast
Extract Broth with appropriate antibiotics. A. tumefaciens cells were pelleted,
resuspended and incubated in an induction medium (10 mM MES, 10 mM MgCl2,
50 μM acetosyringone, pH 5.6) for 3-4 h, and thereafter for 1 h in infiltration medium
(10 mM MES, 10 mM MgCl2, 200 μM acetosyringone, pH 5.6). For gene silencing,
A. tumefaciens cultures carrying pTRV-RNA2 constructs and A. tumefaciens carrying
pTRV1 were mixed at a ratio of 1:1 to a final OD600 of 1.0 and 2.0 before infiltration
into cotyledons of two-week-old N. benthamiana and ten-day-old tomato seedlings,
respectively. TRV:PDS and TRV:GUS were used as controls. For transient gene
expression, A. tumefaciens suspensions with appropriate concentrations were
syringe-infiltrated into N. benthamiana leaves. Co-infiltration with the silencing
suppressor P19 was performed at a ratio of 1:1.
RNA isolation and qRT-PCR
For each silencing construct, six leaves (the fifth and sixth true leaves) from three
individual plants were harvested and ground in liquid nitrogen. Total RNA was
isolated from 100 mg ground sample with the NucleoSpin RNA Plant kit (Macherey-
Nagel), and subsequently used as template for first strand cDNA synthesis using a
M-MLV reverse transcriptase kit (Promega) with an oligo-dT primer (Supplemental
Table S1). qRT-PCR was performed as described by Wang et al. (2014) using Actin
as endogenous control (Supplemental Table S1). Relative expression levels reflect
fold-changes compared to control.
Phytophthora cultivation and infection assays
Culturing of P. capsici isolate LT263 and P. infestans isolate 14-3-GFP and

Chapter VI
174
production of zoospores were performed as previously described (Champouret et al.,
2009; Wang et al., 2013; Bouwmeester et al., 2014).
N. benthamiana and tomato leaves were collected three-to-four weeks after TRV-
treatment or two days after agro-infiltration, and placed in water-saturated floral foam
in trays as described by Vleeshouwers et al. (1999). Leaves were inoculated on the
abaxial sides with fresh mycelial plugs (Ø 0.5 cm), or 10 µl droplets containing 1*105
ml-1 P. capsici zoospores or 5*105 ml-1 P. infestans zoospores. Inoculated leaves
were kept in transparent plastic boxes to maintain high humidity and placed in a
climate chamber with a 12 h photoperiod and appropriate temperature settings.
Boxes were kept the first 24 h in the dark. Lesion diameters were measured for
P. capsici at 3 dpi and for P. infestans at 4 or 6 dpi. Lesion sizes were calculated as
previously described (Vleeshouwers et al., 1999).
ACKNOWLEDGEMENTS
We thank Harold Meijer and Luigi Faino for discussion, Michael Seidl, Huchen Li and
David Nsibo for assistance, Patrick Smit and Daniela Sueldo for providing the
pSol2095 vector and GFP-HA construct, and Henk Smid and Bert Essenstam at
Unifarm for excellent plant care. This research was supported by a Wageningen
University sandwich-PhD fellowship (YW), a Huygens scholarship (YW), a VENI
grant (KB) from The Netherlands Organization for Scientific Research (Technology
Foundation NWO-STW) and the Food-for-Thought campaign of the Wageningen
University Fund.
REFERENCES
Afzal, A.J., Wood, A.J. and Lightfoot, D.A. (2008). Plant receptor-like serine threonine kinases: roles in
signaling and plant defense. Molecular Plant-Microbe Interactions, 21, 507-517.
Boller, T. and Felix, G. (2009). A renaissance of elicitors: Perception of microbe-associated molecular
patterns and danger signals by pattern-recognition receptors. Annual Review of Plant Biology, 60,
379-406.
Bouwmeester, K. and Govers, F. (2009). Arabidopsis L-type lectin receptor kinases: phylogeny,
classification, and expression profiles. Journal of Experimental Botany, 60, 4383-4396.
Bouwmeester, K., van Poppel, P.M.J.A. and Govers, F. (2009). Genome biology cracks enigmas of
oomycete plant pathogens. In: Molecular Aspects of Plant Disease Resistance, Wiley-Blackwell,
102-133.
Bouwmeester, K., de Sain, M., Weide, R., Gouget, A., Klamer, S., Canut, H. and Govers, F. (2011). The
lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a potential
host target for a RXLR effector. PLoS Pathogens, 7, e1001327.

Solanaceous LecRKs and their role in Phytophthora resistance
175
Bouwmeester, K., Han, M., Blanco-Portales, R., Song, W., Weide, R., Guo, L., van der Vossen, E.A.G.
and Govers, F. (2014). The Arabidopsis lectin receptor kinase LecRK-I.9 enhances resistance to
Phytophthora infestans in Solanaceous plants. Plant Biotechnology Journal, 12, 10-16.
Champouret, N., Bouwmeester, K., Rietman, H., van der Lee, T., Maliepaard, C., Heupink, A., van de
Vondervoort, P.J.I., Jacobsen, E., Visser, R.G.F., van der Vossen, E.A.G., et al. (2009).
Phytophthora infestans isolates lacking class I ipiO variants are virulent on Rpi-blb1 potato.
Molecular Plant-Microbe Interactions, 22, 1535-1545.
Dardick, C. and Ronald, P. (2006). Plant and animal pathogen recognition receptors signal through
non-RD kinases. PLoS Pathogens, 2, 14-28.
Desclos-Theveniau, M., Arnaud, D., Huang, T.Y., Lin, G.J.C., Chen, W.Y., Lin, Y.C. and Zimmerli, L.
(2012). The Arabidopsis lectin receptor kinase LecRK-V.5 represses stomatal immunity induced by
Pseudomonas syringae pv. tomato DC3000. PLoS Pathogens, 8, e1002513.
Fry, W. (2008). Phytophthora infestans: the plant (and R gene) destroyer. Molecular Plant Pathology,
9, 385-402.
Gomez-Gomez, L. and Boller, T. (2000). FLS2: An LRR receptor-like kinase involved in the perception
of the bacterial elicitor flagellin in Arabidopsis. Molecular Cell, 5, 1003-1011.
Gouget, A., Senchou, V., Govers, F., Sanson, A., Barre, A., Rouge, P., Pont-Lezica, R.P. and Canut, H.
(2006). Lectin receptor kinases participate in protein-protein interactions to mediate plasma
membrane-cell wall adhesions in Arabidopsis. Plant Physiology, 140, 81-90.
Hall, B.G. (2013). Building phylogenetic trees from molecular data with MEGA. Molecular Biology and
Evolution, 30, 1229-1235.
Hanks, S.K. (2003). Genomic analysis of the eukaryotic protein kinase superfamily: A perspective.
Genome Biology, 4, 111.
Hann, D.R. and Rathjen, J.P. (2007). Early events in the pathogenicity of Pseudomonas syringae on
Nicotiana benthamiana. Plant Journal, 49, 607-618.
Kamoun, S. (2006). A catalogue of the effector secretome of plant pathogenic oomycetes. Annual
Review of Phytopathology, 44, 41-60.
Kanzaki, H., Saitoh, H., Takahashi, Y., Berberich, T., Ito, A., Kamoun, S. and Terauchi, R. (2008).
NbLRK1, a lectin-like receptor kinase protein of Nicotiana benthamiana, interacts with Phytophthora
infestans INF1 elicitin and mediates INF1-induced cell death. Planta, 228, 977-987.
Kasuga, T. and Gijzen, M. (2013). Epigenetics and the evolution of virulence. Trends in Microbiology,
21, 575-582.
Koornneef, M. and Meinke, D. (2010). The development of Arabidopsis as a model plant. Plant
Journal, 61, 909-921.
Kornev, A.P., Haste, N.M., Taylor, S.S. and Ten Eyck, L.F. (2006). Surface comparison of active and
inactive protein kinases identifies a conserved activation mechanism. Proceedings of the National
Academy of Sciences of the United States of America, 103, 17783-17788.
Kroon, L.P.N.M., Brouwer, H., de Cock, A.W.A.M. and Govers, F. (2012). The genus Phytophthora
anno 2012. Phytopathology, 102, 348-364.
Lamour, K.H., Stam, R., Jupe, J. and Huitema, E. (2012). The oomycete broad-host-range pathogen
Phytophthora capsici. Molecular Plant Pathology, 13, 329-430.
Naim F., Nakasugi K., Crowhurst R.N., Hilario E., Zwart A.B., Hellens R.P., Taylor J.M., Waterhouse
P.M. and Wood C.C. (2012). Advanced engineering of lipid metabolism in Nicotiana benthamiana
using a draft genome and the V2 viral silencing-suppressor protein. PLoS One, 7, e52717.
Nakasugi K., Crowhurst R.N., Bally J., Wood C.C., Hellens R.P. and Waterhouse P.M. (2013). De
Novo transcriptome sequence assembly and analysis of RNA silencing genes of Nicotiana

Chapter VI
176
benthamiana. PLoS One, 8, e59534.
Nolen, B., Taylor, S. and Ghosh, G. (2004). Regulation of protein kinases: Controlling activity through
activation segment conformation. Molecular Cell, 15, 661-675.
Phillips, S.M., Dubery, I.A. and van Heerden, H. (2013). Identification and molecular characterisation
of a lectin receptor-like kinase (GhLecRK-2) from Cotton. Plant Molecular Biology Reporter, 31, 9-
20.
Robatzek, S., Bittel, P., Chinchilla, D., Köchner, P., Felix, G., Shiu, S.H. and Boller, T. (2007).
Molecular identification and characterization of the tomato flagellin receptor LeFLS2, an orthologue
of Arabidopsis FLS2 exhibiting characteristically different perception specificities. Plant Molecular
Biology, 64, 539-547.
Shiu, S.H. and Bleecker, A.B. (2001). Receptor-like kinases from Arabidopsis form a monophyletic
gene family related to animal receptor kinases. Proceedings of the National Academy of Sciences of
the United States of America, 98, 10763-10768.
Shiu, S.H. and Bleecker, A.B. (2003). Expansion of the receptor-like kinase/Pelle gene family and
receptor-like proteins in Arabidopsis. Plant Physiology, 132, 530-543.
Singh, P., Kuo, Y.C., Mishra, S., Tsai, C.H., Chien, C.C., Chen, C.W., Desclos-Theveniau, M., Chu, P.
W., Schulze, B., Chinchilla, D., Boller, T. and Zimmerli, L. (2012). The lectin receptor kinase-VI.2 is
required for priming and positively regulates Arabidopsis pattern-triggered immunity. Plant Cell, 24,
1256-1270.
Singh, P., Chien, C.C., Mishra, S., Tsai, C.H. and Zimmerli, L. (2013). The Arabidopsis LECTIN
RECEPTOR KINASE-VI.2 is a functional protein kinase and is dispensable for basal resistance to
Botrytis cinerea. Plant Signaling & Behavior, 8, 1-3.
Takai, R., Isogai, A., Takayama, S. and Che, F.S. (2008). Analysis of flagellin perception mediated by
flg22 receptor OsFLS2 in rice. Molecular Plant-Microbe Interactions, 21, 1635-1642.
Vleeshouwers, V.G.A.A., van Dooijeweert, W., Keizer, L.C.P., Sijpkes, L., Govers, F. and Colon, L.T.
(1999). A laboratory assay for Phytophthora infestans resistance in various Solanum species
reflects the field situation. European Journal of Plant Pathology, 105, 241-250.
Vleeshouwers, V.G.A.A., Raffaele, S., Vossen, J.H., Champouret, N., Oliva, R., Segretin, M.E.,
Rietman, H., Cano, L.M., Lokossou, A., Kessel, G., Pel, M.A. and Kamoun, S. (2011).
Understanding and exploiting late blight resistance in the age of effectors. Annual Review of
Phytopathology, 49, 507-531.
Wang, Y., Zhang, M., Ke, K. and Lu, Y. (2005). Cellular localization and biochemical characterization of
a novel calcium-dependent protein kinase from tobacco. Cell Research, 15, 604-612.
Wang, Y., Bouwmeester, K., van de Mortel, J.E., Shan, W. and Govers, F. (2013). A novel Arabidopsis-
oomycete pathosystem: differential interactions with Phytophthora capsici reveal a role for
camalexin, indole glucosinolates and salicylic acid in defence. Plant, Cell & Environment, 36, 1192-
1203.
Wang, Y., Bouwmeester, K., Beseh, P., Shan, W. and Govers, F. (2014). Phenotypic analyses of
Arabidopsis T-DNA insertion lines and expression profiling reveal that multiple L-type lectin receptor
kinases are involved in plant immunity. Molecular Plant-Microbe Interactions, 27, 1390-1402.
Wu, T., Wang, R., Xu, X., He, X., Sun, B., Zhong, Y., Liang, Z., Luo, S. and Lin, Y. (2014). Cucumis
sativus L-type lectin receptor kinase (CsLecRK) gene family respond to Phytophthora melonis,
Phytophthora capsici and water immersion in disease resistant and susceptible cucumber cultivars.
Gene, 549, 214-222..
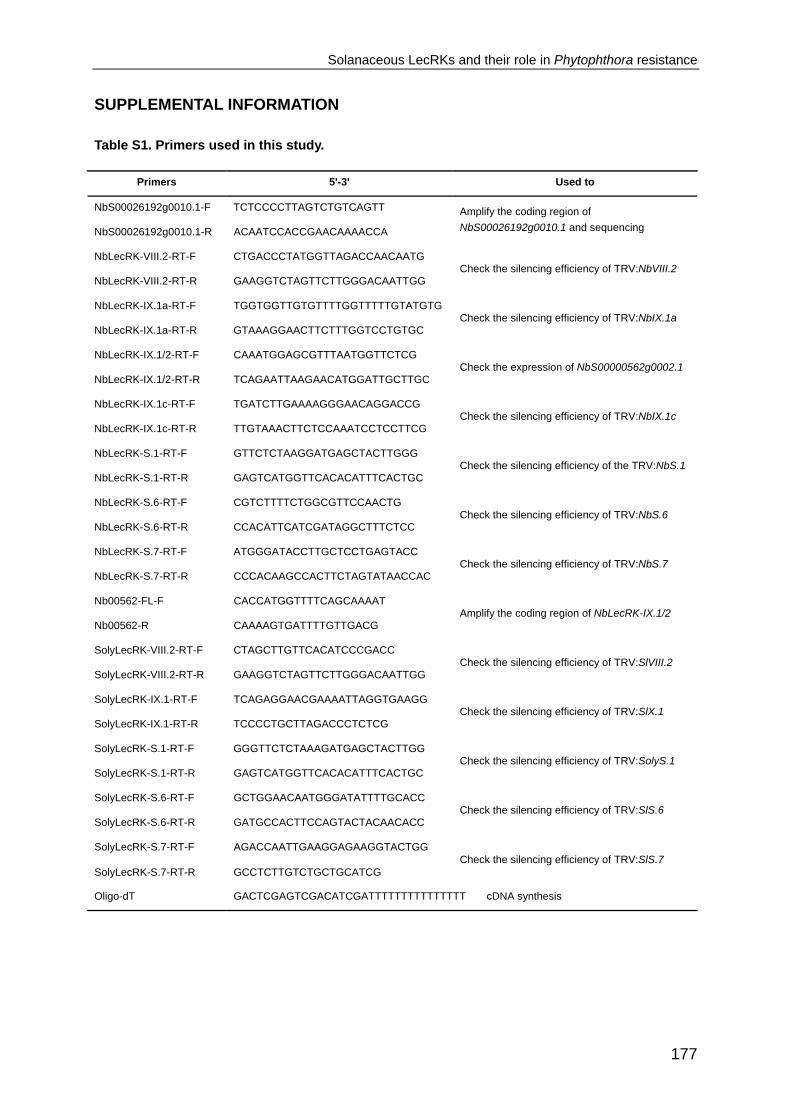
Solanaceous LecRKs and their role in Phytophthora resistance
177
SUPPLEMENTAL INFORMATION
Table S1. Primers used in this study.
Primers 5'-3' Used to
NbS00026192g0010.1-F TCTCCCCTTAGTCTGTCAGTT Amplify the coding region of
NbS00026192g0010.1 and sequencing NbS00026192g0010.1-R ACAATCCACCGAACAAAACCA
NbLecRK-VIII.2-RT-F CTGACCCTATGGTTAGACCAACAATG Check the silencing efficiency of TRV:NbVIII.2
NbLecRK-VIII.2-RT-R GAAGGTCTAGTTCTTGGGACAATTGG
NbLecRK-IX.1a-RT-F TGGTGGTTGTGTTTTGGTTTTTGTATGTG Check the silencing efficiency of TRV:NbIX.1a
NbLecRK-IX.1a-RT-R GTAAAGGAACTTCTTTGGTCCTGTGC
NbLecRK-IX.1/2-RT-F CAAATGGAGCGTTTAATGGTTCTCG Check the expression of NbS00000562g0002.1
NbLecRK-IX.1/2-RT-R TCAGAATTAAGAACATGGATTGCTTGC
NbLecRK-IX.1c-RT-F TGATCTTGAAAAGGGAACAGGACCG Check the silencing efficiency of TRV:NbIX.1c
NbLecRK-IX.1c-RT-R TTGTAAACTTCTCCAAATCCTCCTTCG
NbLecRK-S.1-RT-F GTTCTCTAAGGATGAGCTACTTGGG Check the silencing efficiency of the TRV:NbS.1
NbLecRK-S.1-RT-R GAGTCATGGTTCACACATTTCACTGC
NbLecRK-S.6-RT-F CGTCTTTTCTGGCGTTCCAACTG Check the silencing efficiency of TRV:NbS.6
NbLecRK-S.6-RT-R CCACATTCATCGATAGGCTTTCTCC
NbLecRK-S.7-RT-F ATGGGATACCTTGCTCCTGAGTACC Check the silencing efficiency of TRV:NbS.7
NbLecRK-S.7-RT-R CCCACAAGCCACTTCTAGTATAACCAC
Nb00562-FL-F CACCATGGTTTTCAGCAAAAT Amplify the coding region of NbLecRK-IX.1/2
Nb00562-R CAAAAGTGATTTTGTTGACG
SolyLecRK-VIII.2-RT-F CTAGCTTGTTCACATCCCGACC Check the silencing efficiency of TRV:SlVIII.2
SolyLecRK-VIII.2-RT-R GAAGGTCTAGTTCTTGGGACAATTGG
SolyLecRK-IX.1-RT-F TCAGAGGAACGAAAATTAGGTGAAGG Check the silencing efficiency of TRV:SlX.1
SolyLecRK-IX.1-RT-R TCCCCTGCTTAGACCCTCTCG
SolyLecRK-S.1-RT-F GGGTTCTCTAAAGATGAGCTACTTGG Check the silencing efficiency of TRV:SolyS.1
SolyLecRK-S.1-RT-R GAGTCATGGTTCACACATTTCACTGC
SolyLecRK-S.6-RT-F GCTGGAACAATGGGATATTTTGCACC Check the silencing efficiency of TRV:SlS.6
SolyLecRK-S.6-RT-R GATGCCACTTCCAGTACTACAACACC
SolyLecRK-S.7-RT-F AGACCAATTGAAGGAGAAGGTACTGG Check the silencing efficiency of TRV:SlS.7
SolyLecRK-S.7-RT-R GCCTCTTGTCTGCTGCATCG
Oligo-dT GACTCGAGTCGACATCGATTTTTTTTTTTTTTT cDNA synthesis

Chapter VI
178
Figure S1. Neighbor-Joining trees constructed based on lectin domain (A) and kinase domain
(B) of 43 AtLecRKs, 38 NbLecRKs and 22 SlLecRKs.
Nbc25369236g0004.1 was excluded in (A). Branches of the phylogenetic trees were colored
according to the clades delineated by Bouwmeester and Govers (2009). Numbers above branches
represent the level of clade support inferred by 10,000 bootstrap replicates.

Solanaceous LecRKs and their role in Phytophthora resistance
179
Figure S2. Morphology of N. benthamiana plants treated by TRV:NbLecRKs, TRV:PDS and
TRV:GUS.
Plants were photographed three weeks after inoculation with the silencing constructs.

Chapter VI
180
NbS00026192g0010.1 1315 CTTATGCCTAATGGGAGTCTTGATAAGGCATTATTTGAATCAAGAATGATTCTACCTTGG NbS00026087g0010.1 1315 CTAATGCCTAATGGGAGTCTTGATAAGGCATTATTTGAATCAAGAATGATTCTACCTTGG NbVIII.2 1 --------------------GAATTCGGCATTATTTGAATCAAGAATGATTCTACCTTGG NbS00016101g0013.1 1318 TTGATGCCAAATGGTAGTCTTGATAAGGCATTATTTGAGTCAAGAATGGTACTCCCATGG
NbS00026192g0010.1 1375 GTACATAGGAGGAAAATTTTGCTAGGTGTTGCTTCAGCCTTGGCATATTTACATCAAGAA NbS00026087g0010.1 1375 CTACATAGGAGGAAAATTTTGCTAGGTGTTGCTTCAGCCTTGGCATATTTACATCAAGAA NbVIII.2 41 CTACATAGGAGGAAAATTTTGCTAGGTGTTGCTTCAGCCTTGGCATATTTACATCAAGAA NbS00016101g0013.1 1378 CTACATAGGAGGAAAATTTTGTTAGGTGTTGCTTCAGCCTTAGCATATTTACATCAAGAA NbS00026192g0010.1 1435 TGTGAAAACCAGGTGATTCACAGGGATATAAAGAGTAGTAACATTATGTTGGATGAAGGG NbS00026087g0010.1 1435 TGTGAAAATCAGGTGATTCATAGGGATATAAAGAGTAGTAACATTATGTTAGATGAAGGG NbVIII.2 101 TGTGAAAATCAGGTGATTCATAGGGATATAAAGAGTAGTAACATTATGTTAGATGAAGGG NbS00016101g0013.1 1438 TGTGAAAATCAAGTTATGTTAAGGGATATTAAGACTAGTAATATTATGTTGGATGAAGGG
NbS00026192g0010.1 1495 TTCAATGCAAGATTAGGTGATTTTGGATTAGCAAGACAAGTTGAACATGACAAGTCCCCC NbS00026087g0010.1 1495 TTCAATGCAAGATTAGGTGATTTTGGATTAGCAAGACAAGTTGAACATGACAAGTCCCCC NbVIII.2 161 TTCAATGCAAGATTAGGTGATTTTGGATTAGCAAGACAAGTTGAACATGACAAGTCCCCC NbS00016101g0013.1 1498 TTTAATGCAAGATTAGGTGATTTTGGGTTAGCAAAACAAATTGAACATGACAAATCTCCT
NbS00026192g0010.1 1555 GATGTAACAGTAGCAGCTGGGACAATGGGCTACTTGGCTCCTGAATACTTGTTAACCGGA NbS00026087g0010.1 1555 GATGCAACGGTAGCAGCCGGGACAATGGGCTACTTGGCTCCTGAATACTTGTTAACCGGA NbVIII.2 221 GATGCAACGGTAGCAGCCGGGACAATGGGCTACTTGGCTCCTGAATACTTGTTAACCGGA NbS00016101g0013.1 1558 GATGCGACAGTAGCAGCAGGAACAATGGGATACTTAGCACCTGAATATTTATTAACCGGT
NbS00026192g0010.1 1615 AGAGCAACCGAAAAAACTGATGTTTTTAGCTATGGAGCTGTGGTTCTTGAAGTGGCAAGT NbS00026087g0010.1 1615 AGAGCAACCGAAAAAACTGATGTTTTTAGCTATGGAGCAGTTGTTCTTGAAGTGGCAAGT NbVIII.2 281 AGAGCAACCGAAAAAACTGATGTTTTTAGCTATGGAGCAGTTGTTCTTGAAGTGGCAAGT NbS00016101g0013.1 1618 CGAGCAAGTGAAAAAACTGATGCTTTTAGCTATGGCGCAGTGGTTCTTGAAGTGGCAAGT
NbS00026192g0010.1 1675 GGAAGGAGGCCAATTGAGAGGGAAACAACAAGAGTTGAGAAAGTTGGAGTGAAGAGTAAC NbS00026087g0010.1 1675 GGAAGGAGGCCAATTGAGAGGGAAACAACAAGAGTTGAGAAAGTTGGAGTGAATAGTAAC NbVIII.2 341 GGAAGGAGGCCAATTGAGAGGGAAACAACAAGAGTTGAGAAAGTTGGAGTGAATAGTAAC NbS00016101g0013.1 1678 GGGAGAAGGCCAATTGAGAAAGAAACAAATGGGGTTGGGAAAGTTGGATTAAATAGCAAT
NbS00026192g0010.1 1735 TTAGTTGAATGGGTTTGGGGATTGCATAGAGAAGGGAATTTGCTAATGGCAGCAGATTCA NbS00026087g0010.1 1735 TTAGTTGAATGGGTTTGGGGATTGCATAGAGAAGGGAATTTGCTAATGGCAGCTGATTCA NbVIII.2 401 TTAGTTCCCGGG------------------------------------------------ NbS00016101g0013.1 1738 TTGGTGGAATGGGTTTGGGGATTACATAAAGAAGGGAGGTTATTAGCAGCAGCTGATTCA
NbS00026192g0010.1 1855 TGTTCACAACCTGACCCTATGGTTAGACCAACAATGAGAAGTGTGGTCCAAATGCTAGTA
NbS00026087g0010.1 1855 TGTTCACAACCTGACCCTATGGTTAGACCAACAATGAGAAGTGTGGTCCAAATGCTAGTA NbVIII.2-2 1 ------------GACCCTATGGTTAGACCAACAATGAGAAGTGTGGTCCAAATGCTAGTA NbS00016101g0013.1 1858 TGTTCACACCCTGACCCTATGGCTAGACCAACAATGAGAGGTGTGGTTCAAATGTTAGTT NbS00026192g0010.1 1915 GGTGAAGCTGAAGTTCCTATTGTCCCAAGAACTAAGCCTTGTATGAGTTTCAGCACATCA NbS00026087g0010.1 1915 GGTGAAGCTGAAGTTCCTATTGTCCCAAGAACTAAGCCTTCTATGAGTTTCAGCACATCA NbVIII.2-2 49 GGTGAAGCTGAAGTACCTATTGTCCCAAGAACTAAGCCTTCTATGAGTTTCAGCACATCA NbS00016101g0013.1 1918 GGTGAGGCTGAAGTTCCAATTGTCCCTAGAGCTAAGCCAACTATGAGTTTTAGCACATCT NbS00026192g0010.1 1975 CATCTTCTAATGACTTTGCAAGATAGTGTTTCTGATTTGAATGGTATGATCACACTTTCC NbS00026087g0010.1 1975 CATCTTCTAATGACTTTGCAAGATAGTGTTTCTGATTTGAATGGTATGATCACACTTTCA NbVIII.2-2 109 CATCTTCTAATGACTTTGCAAGATAGTGTTTCTGATTTGAATGGTATGATCACACTTTCC NbS00016101g0013.1 1978 CATCTGCTAATGACTTTGCAAGATAGTGTTTCTGACTTGAATGGTATGATCACACTTTCC NbS00026192g0010.1 2035 TCTTCATCATCTGAAAACAGTTTCACTGGTGTTGCCAATGGGATGGACTTGGTCTAA NbS00026087g0010.1 2035 ACTTCATCATCTGAAAACAGCTTCAATAGTGCTGGCAATGGCATGGACTTGGTCTAA NbVIII.2-2 169 ACTTCATCATCTGAAAACAGTTTCACTGGTGTTGCCCATGGGATGA---TGGA---- NbS00016101g0013.1 2038 ACGTCTTCATCTGAAAATAGCTTCAATGGTGGTGGTGTGGGTATGGACCTAGTCTAG
Figure S3. Sequence alignment of NbS00016101g0013.1, NbS00026192g0010.1, NbS00026087
g0010.1 and the fragments used for silencing.
NbVIII.2 represents the fragment used in this study while NbVIII.2-2 represents the fragment retrieved
according to Kanzaki et al. (2008).

Solanaceous LecRKs and their role in Phytophthora resistance
181
Figure S4. Quantified lesion sizes on SlLecRK-silenced tomato leaves after inoculation with P. capsici
(mycelial plugs Ø 0.5 cm) or P. infestans (5*105 zoospores ml-1). Bars represent the mean lesion size
(± SE). Each experiment included at least twenty leaves silenced for each construct. * indicates
significant difference in lesion sizes (p < 0.05) between TRV:GUS and TRV:SlLecRK-IX.1-treated
plants according to a t test. This experiment was repeated twice, with similar outcome for P. capsici but
not for P. infestans.
Supplemental file 1. DNA sequences of the fragments used for TRV-mediated silencing in
N. benthamiana and tomato.
>TRV:NbVIII.2/TRV:SlVIII.2
gaattcggcattatttgaatcaagaatgattctaccttggctacataggaggaaaattttgctaggtgttgcttcagccttggcatatttacatcaaga
atgtgaaaatcaggtgattcatagggatataaagagtagtaacattatgttagatgaagggttcaatgcaagattaggtgattttggattagcaag
acaagttgaacatgacaagtcccccgatgcaacggtagcagccgggacaatgggctacttggctcctgaatacttgttaaccggaagagcaa
ccgaaaaaactgatgtttttagctatggagcagttgttcttgaagtggcaagtggaaggaggccaattgagagggaaacaacaagagttgaga
aagttggagtgaatagtaacttagttcccggg
>TRV:NbIX.1a
gaattctaattccttttgcgacttcgctgtccttcaatttcgatagttttagacccagcgatcaaaatgtaacatacgagagagatgcttatccagcaa
atggtgcaattcaacttaccacagaccttatcaatcgtgatataaacgcaactataggtcgagccacatattccaagcttctgcatctttgggaca
aggcctcaggaaatgtcacagatttcagtactcacttctcctttagcatcaattcacaaggcagaacaagatatggtgatggtcttgccttcctgag
aacacaaccagaggtggcagtcttggccatacaagtaatactcaacgactgaatacatcggccaatcattttgtagctgtggagtttgatacgta
ccagaacgtccagtatgatcccggg
>TRV:NbIX.1b
gaattcctcagcatcaactggacaattgtttcagaaaaacaatgtcaagtcttgggatttcaattcaagtatgagttttgatccaaacaaagttcctg
aacatgctgaaccgccgagtgcaaatcctcctattactcaagttactcctccttctaaccaagaaatcagcaccagaaaaaaaggaaataagg
gactagtcgttggatcgagcataggtttgcctattctggttatcgtattgattactggtagctgctttttatggaagaaaaagagtaaaaggaatgata

Chapter VI
182
aagaccatgtttttattgatctcagtatggacaatgaatttcaaaagggtactggccctaagaagttttcctatggtgaattagctcgtgcaacgaac
aattttgctgaggtacagcccggg
>TRV:NbIX.1c
gaattccgattcaagttttaatgtcaagctcggtgattttggcttagctagactaatggaccatgaattaggtcctcagactacagggttggctggaa
ctttaggttatttggctcctgaatacataaaaacaggccgagcaagtaaagagtcagacgtatacagcttcggaatagttgcacaagaaattgc
aaccggaagaaaatcagttgatccggggacggggaaatctgatgcagtgttagtggagtatgtttgggagctttataaaggacaacttctttctg
ctgttgatgagaaattaaacctggattttgacgcgaaacaagtagagcgattgatggctaccaggttatggtgtgctcatccggagagcaatctg
aggccatctataagacaagcaattcccggg
>TRV:SlIX.1
ggatccgacatggattacttacaatgctacaacaaataatcttagtgtcatttggaactatggaacaggtccgaattcgagcatgttttacataata
aacctcagagatgttctgcctccgtgggtcacaattggattctctgctgcaacaggactaaatgtcgagagacatacacttgaatcgtgggaattc
agctcgagtcttgttataacagagttaggtggaaatgatcgcgaaaagattggacttattgcagggctaacaacattaggtgggattttgttagtga
gcgcgattttggctttgatagttttgaggaaacgtagacgaaaggtgaagggaaatccagagacaataagcttaacatctttcaatgatgatcttg
aaaagggggcaggaccaagacccggg
>TRV:NbS.1/TRV:SlS.1
gaattcatggaagtctgaataaatggatatttgataagccagagaaggttatgaattggctagacaggaggaaggtcctaactgatgttgctgag
ggtttaaactatttacatcatggttgggaacaagtggttgtacatagggatattaaatctagcaatgtcttgttagattgtgaaatgagagggagact
gggagatttcgggctagcaaagttatacactcacggtggtgtaccaaatacaactagggtagtaggtacattagggtacttggcacctgaagttg
tgacaagggctacaccaactgcagctagtgacccggg
>TRV:NbS.6/TRV:SlS.6
gaattcttgtatatggtggcttagctgcagttgttacacttgttgttggtattctggctattgtgtctgtttttgtgttaaggagaaaaaaaagggatggta
gtagaggggaaaatgaaggtcaaatgtgtagattccaaggaaatagagtgcctcaaagattgtcattatctgaaataaaatcagccacagaa
ggatttaataatgaaaggataattggtgaaggaggatctgctgttgtatatgaaggagatattccttctaagggaactgttgctattaagagatttgtt
caagggactagattaggtccttcacatattccatttaacactgaatttgcttctatggttggctgcttaagacacaagaatttgattcagttccaagga
tggtgttgtgagagcccggg
>TRV:NbS.7
gaattcatttggagatggtttggcttttttcctttcacctgataatcagactttaggtagtccaggtggtttcttgggtctggttaattcatcccaattgacta
agaacaagtttgttgctgttgagttcgacacttggcaagatttgcactttaatgatcctgatgataaccatgtaggtcttgatatagatagccttatctc
aatcaagactgctaatgtaaagttggctggtgttgatctcaaaagtaggaatttgattacttcttggattgattacaagagtgaggaaaagcagttgt
ttgtgttcttgagttactcaagttccaagcctaaaaaaccactcttgactgtggatattgacctgtctgattatctaaaggagtttatgtacgtggggttt
gctcccggg
>TRV:SlS.7
gaattcaagacttcaaagaggaagagatgaaaaagttgctacttgttggactgagctgtgcaaatcctgatagcacagaaaggccttgtatga
ggagagtatttcagatactcaacaatgaggcggaacctatctttgttccgaaagtgaaaccaactctaactttctccactagcatcccgttcaacat
tgatgacattttctcagacagtgaagggagtgaggcaccagaacatgagctcgaaatcagaatacccggg

Chapter VII
General discussion

Chapter VII
184
INTRODUCTION Receptor-like kinases (RLKs) form one of the largest families in plants, with over 600
and 1100 members in Arabidopsis and rice, respectively (Shiu and Bleecker, 2001a;
Shiu et al., 2004). The RLK family is dramatically expanded in plants when compared
to other organisms, such as animal, fungi and protists (Shiu and Bleecker, 2001b;
Shiu and Bleecker, 2003). Within the Arabidopsis RLK family, over 400 members
contain a domain configuration resembling a transmembrane receptor composed of
an extracellular putative ligand binding domain and a cytosolic kinase domain which
transduces signals through phosphorylation of downstream signaling components
(Antolin-Llovera et al., 2012; Kadota et al., 2014; Li et al., 2014). Plant RLKs have
been implicated in many different processes, including development, growth,
hormone perception and response to abiotic and biotic stresses (De Smet et al.,
2009). One of the most important determinants for plant adaptation is the capacity to
perceive environmental stimuli and initiate defence responses accordingly. Several
RLKs are capable to recognize conserved microbial signatures, named microbe-
associated molecular patterns (MAMPs), or molecules derived from plants
themselves upon pathogen attack, the so-called damage-associated molecular
patterns (DAMPs). This type of RLKs is also known as pattern recognition receptors
(PRRs).
One example that has been well-characterized with respect to ligand
perception and defence activation is FLS2, a RLK with an extracellular domain
composed of 28 tandem leucine-rich repeats (LRRs). FLS2 plays an important role in
perception of bacterial pathogens by detecting a conserved epitope of flagellin,
referred to as flg22. Perception occurs via the direct binding of flg22 to the LRR
domain of FLS2 and this leads to the rapid recruitment of BAK1, another RLK which
acts as a co-receptor for FLS2 to bind flg22 (Sun et al., 2013). This intermolecular
interaction leads to the rapid phosphorylation of the cytoplasmic kinase BIK1, which
constitutively interacts with FLS2 and BAK1 (Lu et al., 2010). In turn, BIK1 is
released from the FLS2 complexes and directly binds and phosphorylates the
NADPH oxidase RbohD to activate rapid production of ROS (Kadota et al., 2014; Li
et al., 2014).
RLKs vary greatly in their extracellular domain. Based on the diverse
extracellular domains, RLKs were classified into different subfamilies (Shiu and

General discussion
185
Bleecker, 2001a). Both FLS2 and BAK1 belong to LRR-RLKs, the largest subfamily
which consists of 235 members in Arabidopsis with varying number of LRRs (Shiu
and Bleecker, 2001a). Thus far, four LRR-RLKs in Arabidopsis have been found to
function as PRRs, i.e. FLS2, EFR, PEPR1 and PEPR2 (Huffaker et al., 2006; Zipfel,
2006; Yamaguchi et al., 2010). Another two members, i.e. BAK1 and SOBIR1, act as
co-receptors of multiple RLKs or receptor-like proteins (RLPs) (Chinchilla et al., 2009;
Liebrand et al., 2014). Several other LRR-RLKs, such as ERECTA, are also involved
in plant immunity, but the exact activation mechanisms are yet unknown (Sanchez-
Rodriguez et al., 2009). Next to the LRR-RLK subfamily, a few other large subfamilies
have been implicated in plant defence. One such family comprises the L-type lectin
receptor kinases (LecRKs) which are characterized by an legume lectin-like
extracellular domain (Shiu and Bleecker, 2001a; Barre et al., 2002; Bouwmeester
and Govers, 2009). This chapter focuses on the role of LecRKs in plant defence and
their potential to be exploited for disease resistance management in crops.
LecRKs exhibit species-specific expansions in plants
LecRKs are present in many different plant species, but have so far not been found in
the genome sequences of yeast (Saccharomyces cerevisiae), human or green algae
(Shiu and Bleecker, 2001b; Navarro-Gochicoa et al., 2003; Lehti-Shiu et al., 2009).
The available plant genome sequences enable the identification of LecRKs on a
genome-wide scale. The Arabidopsis genome encodes 45 LecRKs with 41 containing
the typical LecRK composition and four either lacking the signal peptide, the lectin
domain or the transmembrane domain (Bouwmeester and Govers, 2009). A
comparable number of LecRKs, i.e. 38 LecRKs were identified in
Nicotiana benthamiana (Chapter VI) while only about half of that number was found
in the genome of cucumber (Cucumis sativus) and tomato (Solanum lycopersicum),
i.e. 25 and 22, respectively (Wu et al., 2014; Chapter VI). In the monocot rice (Oryza
sativa), in total 69 full length LecRKs were identified (Vaid et al., 2012). Also for a few
other plant species, LecRKs have been reported but not on a genome-wide scale,
such as Gossypium hirsutum (GhLecRK-2), Medicago truncatula (i.e. MtLecRK1;1,
MtLecRK7;1, MtLecRK7;2 and MtLecRK7;3), Populus nigra (PnLPK), Pisum sativum
(PsLecRLK), (Nishiguchi et al., 2002; Navarro-Gochicoa et al., 2003; Joshi et al.,
2010; Phillips et al., 2013).
In Arabidopsis, the 45 LecRKs were classified into nine clades (I to IX) and

Chapter VII
186
seven singletons based on sequence similarity and phylogenetic analyses
(Bouwmeester and Govers, 2009). Similar analyses were performed on LecRKs of
Arabidopsis and two Solanaceous plants, i.e. N. benthamiana and tomato, which
revealed that Solanaceous plants have no LecRKs belonging to clades I, II, III and V
but contain another clade that is absent in Arabidopsis (Chapter VI). To gain a better
insight into the divergence of LecRKs in plants, we compared LecRKs of the dicot
plants Arabidopsis (AtLecRKs), N. benthamiana (NbLecRKs) and tomato (SlLecRKs)
with those of the monocot rice (OsLecRKs) and constructed a phylogenetic tree (Fig.
1). Of the 69 OsLecRKs, the majority does not group into the clades defined based
on the Arabidopsis sequences. None of the OsLecRKs fall into the four Arabidopsis-
specific clades or the Solanaceous-specific clade. However, an extensively expanded
clade was identified with 36 OsLecRKs which is closest to Arabidopsis clade IV.
Similar to what has been found for Arabidopsis-specific clades (Bouwmeester and
Govers, 2009; Chapter VI), OsLecRKs in this clade exhibit high frequency of tandem
duplications based on the definition that tandem duplicated genes are those within 10
genes apart in genome (Lehti-Shiu et al., 2009). Hence, the expansion of this rice-
specific LecRK clade seems to depend largely on tandem duplication events. Another
large cluster of 14 OsLecRKs groups with clade IX indicating that clade IX is an
ancient clade that arose before the separation of monocots and dicots. Yet, the
increased number of clade IX LecRKs found in rice (14), N. benthamiana (6), and
tomato (5) compared to Arabidopsis (2) indicates independent gene loss or gain
events after the monocot-dicot split. This is also the case for the clade comprising the
Arabidopsis singleton LecRK-S.5 homologs, which includes seven OsLecRKs, six
NbLecRKs and three SlLecRKs. OsLecRKs are also present in the clades comprising
clade VI LecRKs, and the Arabidopsis singletons LecRK-S.6 and LecRK-S.7, as are
NbLecRKs and SlLecRKs. Hence these clades are also conserved in both monocot
and dicot plants, but as yet there are no indications for species-specific gene
expansions. Collectively, these data indicate that the LecRK family is widespread in
different plant species, and underwent species-specific expansions.

General discussion
187
Figure 1. Phylogenetic analysis of LecRKs belonging to four plant species.
Full-length protein sequences were aligned in ClustalW using the weight matrix GONNET with a
penalty gap opening of 10 and a gap extension of 0.1. The alignment was subsequently used to
construct a Neighbor-Joining tree with 1,000 bootstrap replicates using the Jones-Taylor-Thornton
substitution model (Hall, 2013) in MEGA 5.1. Branches are colored according to the clades delineated
by Bouwmeester and Govers (2009), and branches with less than 60% bootstrap support are
collapsed. Arabidopsis LecRKs (AtLecRKs) were downloaded via TAIR10 and N. benthamiana and
tomato LecRKs (i.e. NbLecRKs and SlLecRKs) from the Sol Genomics website (Chapter VI). The rice
LecRKs (OsLecRKs) are the ones defined by Vaid et al. (2012) and were downloaded from Rice
Genome Annotation Project website (http://rice.plantbiology.msu.edu/). The tree includes 43
AtLecRKs, 38 NbLecRKs, 22 SlLecRKs and 69 OsLecRKs.

Chapter VII
188
LecRKs function as PRRs
Emerging evidence indicates that the extracellular domains of RLKs determine ligand
binding specificity (Liu et al., 2012; Sun et al., 2013; Macho and Zipfel, 2014). The
extracellular lectin domains of LecRKs resemble the soluble carbohydrate-binding
legume lectins (Shiu and Bleecker, 2001a; Barre et al., 2002). Legume lectins are
metalloproteins with both a Ca2+ and a transition metal ion (usually Mn2+)
incorporated in their structure which are essential for the carbohydrate-binding (van
Damme et al., 2008; Etzler, 2009). Hence, the corresponding ion-binding residues
that form the carbohydrate-binding sites are extremely conserved among the legume
lectins (Etzler, 2009). However, structural analysis and molecular modelling revealed
that the lectin domains of Arabidopsis LecRKs are poorly conserved in the ion-
binding residues but contain a conserved hydrophobic cavity, which was proposed to
bind hydrophobic ligands, such as tryptophan and auxin (Barre et al., 2002). So far,
for only one LecRK, i.e. LecRK-I.9 from Arabidopsis, potential ligands that interact
with the extracellular lectin domain have been reported. Arabidopsis LecRK-I.9 was
first identified as a candidate host target of the Phytophthora infestans RXLR effector
IPI-O, and more recently as the first plant extracellular ATP (eATP) receptor (Gouget
et al., 2006; Bouwmeester et al., 2011a; Choi et al., 2014).
IPI-O belongs to a large family of oomycete effectors that shares the host cell
targeting motif RXLR (Champouret et al., 2009). It also contains an RGD (R:
arginine; G: glycine; D: aspartic acid) motif ‒ that partially overlaps with the RXLR
(RXLRGD) motif ‒which is well-known to interfere with cell adhesion in animals and
to disrupt cell wall (CW)-plasma membrane (PM) adhesions in plants (Ruoslahti,
1996; Mellersh and Heath, 2001; Senchou et al., 2004; Bouwmeester et al., 2011b).
Screening of a phage library for peptides binding to the RGD motif of IPI-O identified
two heptamers, i.e. IHQASYY and AAQPHPR which can inhibit the RGD-binding
activity of plasma membrane proteins (Gouget et al., 2006). Both heptamers were
then used to screen the Arabidopsis proteome for plasma membrane proteins with
potential RGD-binding activity. In this way, Arabidopsis LecRK-I.9 was identified as
one of the candidates (Gouget et al., 2006). The capacity of the LecRK-I.9 lectin
domain to bind IPI-O was further confirmed by binding and photoaffinity labelling
assays. The RGD-binding activity of LecRK-I.9 raised the question to what extent
LecRK-I.9 plays a role in mediating CW-PM adhesion. Bouwmeester et al. (2011a)
found that Arabidopsis LecRK-I.9 mutants show disrupted CW-PM integrity, a

General discussion
189
phenomenon that was also found in transgenic Arabidopsis plants with constitutive
expression of ipiO. Taken together, these results indicate that LecRK-I.9 protects the
CW-PM continuity via binding to the RGD-proteins.
Extracellular ATP is a well-known signaling agent that has been implicated in
diverse physiological processes in animals (Bours et al., 2006). Also in plants, eATP
was found to be an important signaling molecule that regulates various cellular
responses (Cao et al., 2014). For example, ATP treatment rapidly triggers elevation
of cytosolic Ca2+ concentration and production of reactive oxygen species in plants
(Demidchik et al., 2009). In animals, eATP signals through the plasma membrane-
localized purinoceptors, including ligand-gated nonselective cation channels (P2X
receptors) or G-protein-coupled receptors (P2Y receptors) (Ralevic and Burnstock,
1998). Identification of plant eATP receptors based on sequence homology to animal
purinoceptors failed to obtain any suitable candidates (Tang et al., 2003; Choi et al.,
2014). It is conceivable that eATP signals through different types of receptors in
plants. Using a forward genetic screening of Arabidopsis mutants that are defective in
response to ATP, Choi et al. (2014) identified LecRK-I.9 and demonstrated its
function as an eATP receptor. Arabidopsis LecRK-I.9 mutants show defects in Ca2+
response upon treatment with ATP or ADP, but not upon treatment with the pyrimidine
nucleotide CTP, biotic elicitors (i.e. flg22, chitin, elf26 and pep1) or abiotic stresses
(i.e. NaCl, mannitol and cold water) (Choi et al., 2014). In vitro binding assays
indicated that the extracellular lectin domain of LecRK-I.9 has a relative high binding
affinity for ATP (Kd ∼ 46 nM) (Choi et al., 2014). ATP can be released from plants
upon pathogen attack or exposure to other stress stimuli, in particular upon wounding
(Tanaka et al., 2010; Clark and Roux, 2011). It has been documented that the
released eATP in Arabidopsis upon wounding can reach a concentration up to 45 µM
(Song et al., 2006). Transcriptome analysis revealed that 60% of the genes induced
upon treatment with eATP were also induced by wounding (Choi et al., 2014). Based
on these findings, Choi et al. (2014) proposed a model in which eATP acts as a
damage-associated molecular pattern (DAMP) that is recognized by LecRK-I.9 to
initiate downstream responses, including increased cytosolic Ca2+ concentration
(Song et al., 2006; Choi et al., 2014). Given the fact that LecRK-I.9 is absent in
multiple plant species, its function in eATP perception may be taken over by other
LecRKs.
The fact that LecRK-I.9 binds, on the one hand, IPI-O and on the other hand

Chapter VII
190
eATP, raises many questions on how these two processes are regulated. For
example, what is the exact ATP-binding site(s) in the lectin domain? Is IPI-O
competing with eATP for LecRK-I.9 binding and in turn interfering with eATP-
mediated downstream signaling? In animals, the eATP binding sites on the P2X and
P2Y receptors have been well investigated (Chataigneau et al., 2013). However, this
information seems not helpful to understand the eATP binding in plants since LecRK-
I.9 is a different type of eATP receptor. It is expected that a combination of
crystallography and site-directed mutagenesis will lead to the identification of the
eATP binding site(s) in LecRK-I.9. This will unravel the mechanisms underpinning
eATP binding and activation of downstream signaling. The binding of IPI-O to LecRK-
I.9 is highly dependent on the RGD motif and it was found that mutation of RGD to
RGA abolished the binding (Gouget et al., 2006). Hence, eATP binding assays with
Arabidopsis protoplasts derived from transgenic Arabidopsis plants expressing ipiO
or ipiORGA may give some clues on whether IPI-O competes with eATP for LecRK-I.9
binding. In addition, by monitoring the response of ipiO/ipiORGA expressing
Arabidopsis lines upon ATP treatment may answer the question whether IPI-O affects
eATP-mediated signal transduction.
LecRKs function in defence against various biotic agents
Maintenance of CW-PM continuity has been found to be essential for plant defence
responses triggered by pathogens (Kiba et al., 1998; Mellersh and Heath, 2001). The
role of LecRK-I.9 in mediating CW-PM adhesions was an indication that LecRK-I.9 is
involved in plant defence. Indeed, Arabidopsis LecRK-I.9 mutants were found to be
compromised in cell-wall-associated defence response, i.e. reduced callose
deposition upon treatment with the Pseudomonas syringae (Pst) hrcC mutant or flg22
(Bouwmeester et al., 2011a). Moreover, LecRK-I.9 expression is induced upon
infection by Phytophthora brassicae. LecRK-I.9 mutants showed gain of susceptibility
to non-adapted P. brassicae isolates and accordingly overexpression lines showed
enhanced resistance to a virulent P. brassicae isolate CBS686.95 (Bouwmeester et
al., 2011a). Besides LecRK-I.9, several Arabidopsis LecRKs are differentially
expressed upon treatment with pathogens or elicitors according to in silico gene
expression analyses (Bouwmeester and Govers, 2009). Experimental evidence for a
function of other Arabidopsis LecRKs in defence was provided when Arabidopsis
LecRK T-DNA insertion lines showed compromised resistance to different pathogens

General discussion
191
(Wang et al., 2014/Chapter III). In this way, in total 14 LecRKs were found to have a
potential role in Phytophthora resistance. These 14 LecRKs fall into different clades
and display different expression patterns upon P. infestans infection. It should be
noted that in the screening of the T-DNA insertion lines, two phylogenetically distant
Phytophthora species were used for infection. None of the LecRKs mutants,
however, showed specificity to either species, suggesting that the 14 LecRKs confer
broad-spectrum Phytophthora resistance. Recognition of pathogens by receptors is
often correlated with diverse biochemical and cell biological changes (Boller and
Felix, 2009). It is difficult to figure out whether all these changes are involved in
limitation of pathogen colonization. For instance, overexpression of Arabidopsis
LecRK-IX.1 and LecRK-IX.2 induces plant cell death but also increases plant
resistance to Phytophthora pathogens (Chapter IV). Plant cell death could play an
essential role in arresting pathogen growth in early infection stages (Dickman and
Fluhr, 2013). However, Phytophthora resistance and cell death induction mediated by
both LecRK-IX.1 and LecRK-IX.2 appeared to be independent (Chapter IV). Both
LecRK-IX.1 and LecRK-IX.2 were found to interact with the ABC transporters
NbPDR1 in N. benthamiana and ABCG40 in Arabidopsis and the latter was found to
be a Phytophthora resistance component (Chapter IV). ABCG40 and its closest
homologs in Nicotiana plumbaginifolia and Nicotiana tabacum were found to be
required for transport of antimicrobial compounds such as ABA and diterpenes (Kang
et al., 2010; Seo et al., 2012). Hence, we speculate that LecRK-IX.1 and LecRK-IX.2
may interact with ABCG40 or NbPDR1 to activate the transport of antimicrobial
compounds that subsequently leads to Phytophthora resistance. The observation that
LecRK-IX.1 and LecRK-IX.2 are not interacting with each other reduces the chance
that LecRK-IX.1, LecRK-IX.2 and ABCG40 are present in one complex. Although
LecRK-IX.1 and LecRK-IX.2 function in a similar manner, knockout of both LecRKs in
Arabidopsis leads to increased susceptibility compared to the null mutants,
suggesting that these two LecRKs have an additive effect on Phytophthora
resistance (Chapter IV). Since clade IX is evolutionarily ancient, LecRKs belonging
to this clade seem to be important in plant adaptation. In line with this, the homologs
of LecRK-IX.1 and LecRK-IX.2 in N. benthamiana and tomato are also involved in
Phytophthora resistance, demonstrated a conserved biological function of clade IX
LecRKs across different plant species (Chapter VI).
Stomatal immunity plays an important role in limiting entry of bacterial

Chapter VII
192
pathogens into plants (Melotto et al., 2006). One of the Arabidopsis LecRKs, i.e.
LecRK-V.5, which is induced upon wounding and oligogalacturonic acid treatment,
was shown to negatively regulate plant stomatal immunity (Desclos-Theveniau et al.,
2012). LecRK-V.5 mutants are resistant to the hemibiotrophic bacterial pathogen Pst
DC3000, but only upon surface-inoculation, not by infiltration-inoculation (Desclos-
Theveniau et al., 2012). Mutation of LecRK-V.5 compromised stomatal reopening,
and accordingly overexpression of LecRK-V.5 resulted in increased stomatal
reopening when treated with Pst DC3000 or MAMPs (i.e. flg22, lipopolysaccharides
and the elongation factor epitope elf26). Upon MAMP treatment, LecRK-V.5 mutants
demonstrated normal apoplastic PTI responses, such as flg22-triggered ROS
production, bacteria-induced callose deposition and PTI-responsive marker gene
expression. However, LecRK-V.5 mutants showed constitutive increased ROS
production in the guard cells and this was found to be indispensable for the stomatal
closure (Desclos-Theveniau et al., 2012). Additionally, LecRK-V.5 was found to
negatively regulate stomatal closure mediated by abscisic acid (ABA) but not by
other hormones, such as salicylic acid or jasmonic acid (Desclos-Theveniau et al.,
2012). Collectively, these data support the negative role of LecRK-V.5 in mediating
stomatal immunity. In contrast, LecRK-V.5 was found to positively regulate
Arabidopsis resistance to Phytophthora pathogens (Wang et al., 2014/Chapter III).
The determinants for the multifaceted role of LecRK-V.5 in mediating resistance
against different pathogens is not yet known, but apparently LecRK-V.5-regulated
stomatal closure is not affecting Arabidopsis resistance to Phytophthora pathogens
since the penetration by Phytophthora pathogens into plants is not preferentially
occurring via the stomata (Roetschi et al., 2001; Wang et al., 2013). LecRK-VI.2,
which is required for BABA-primed defence, is another Arabidopsis LecRK found to
be important for bacterial resistance (Singh et al., 2012). In contrast to LecRK-V.5,
LecRK-VI.2 positively regulates stomatal closure upon MAMP treatment or bacterial
infection and this regulation is not related to ABA signalling. Upon bacterial infection
or treatment with MAMPs (i.e. flg22 or elf26), the LecRK-VI.2 mutant showed
compromised PTI-related responses, such as reduced callose deposition, defence-
related gene expression and reduced MPK3 and MPK6 activity (Singh et al., 2012).
Recently, LecRK-VI.2 was found to associate with FLS2 in a ligand independent
manner (Huang et al., 2014). However, mutation of LecRK-VI.2 in Arabidopsis did not
affect the flg22-induced early responses in plants, such as FLS2 and BAK1

General discussion
193
association, ROS production or BIK1 phosphorylation (Singh et al., 2012). Since
BIK1 was shown to be immediately activated upon recognition of flg22 (Lu et al.,
2010), these results suggest that either LecRK-VI.2 functions downstream of BIK1 or
that LecRK-VI.2-mediated defence responses signal through a BIK1-independent
pathway. In addition, LecRK-VI.2 is involved in mediating plant response to multiple
MAMPs (Singh et al., 2012), suggesting that LecRK-VI.2 is also involved in other
receptor complexes. Transcriptome profiling revealed that many defence-related
genes were up-regulated in Arabidopsis lines overexpressing LecRK-VI.2 (Singh et
al., 2012), however, LecRK-VI.2 likely functions specifically in resistance against
bacterial pathogens but not to others such as Phytophthora or fungal pathogens
(Singh et al., 2013; Huang et al., 2014; Wang et al., 2014/Chapter III). In addition to
the two aforementioned LecRKs, another three Arabidopsis LecRKs (i.e. LecRK-IV.4,
LecRK-S.1 and LecRK-S.4) were indicated to be involved in bacterial resistance
since the corresponding mutants showed increased susceptibility (Wang et al.,
2014/Chapter III).
LecRKs are also indicated to be involved in fungal pathogen resistance and
perception of insects. For example, overexpression of LecRK-IV.3 (alias AtLPK1)
increased Arabidopsis resistance to Botrytis cinerea (Huang et al., 2013). Infection
assays on Arabidopsis LecRK mutants with Alternaria brassicicola indicated that four
of the clade I LecRKs contribute to A. brassicicola resistance (Wang et al.,
2014/Chapter III). One of the four, i.e. LecRK-I.8, is also implicated in recognition of
eggs deposited by the cabbage butterfly Pieris brassicae (Gouhier-Darimont et al.,
2012). Expression of LecRK-I.8 is induced upon treatment with P. brassicae egg
extract, and the LecRK-I.8 mutant was shown to be defective in expression of the
defence-related gene PR1 upon treatment with egg extract.
Systematic analyses of LecRK function by monitoring response of Arabidopsis
LecRK mutants to different pathogens revealed function specificity of LecRKs, i.e.,
several LecRKs function in resistance against particular pathogens, while others are
implicated in resistance to multiple pathogens. Function specificity of LecRKs could
not be reconciled with the differences in gene expression profiles (Wang et al.,
2014/Chapter III), but may be determined by the recognition specificity of the
extracellular lectin domain. This idea was supported by the finding that the lectin
domain is indispensable for Phytophthora resistance mediated by the two clade IX
LecRKs (Chapter IV).

Chapter VII
194
Engineering LecRKs for broad-spectrum resistance: potentials and obstacles
LecRKs have several features that make them attractive as potential resistance
components. They are widespread in the plant kingdom, confer broad-spectrum
disease resistance and show conserved biological functions among different plant
species. A disadvantage, however, is that resistance conferred by LecRKs is
relatively mild when compared to the resistance mediated by intracellular nucleotide-
binding leucine-rich repeat proteins (NLRs), which is overall dominant and often
yields clear resistance phenotypes (van der Vossen et al., 2003; van der Vossen et
al., 2005). LecRK-mediated resistance often fails to arrest the infection by the
adapted pathogens. Hence, exploiting LecRKs for improving crop resistance is still a
major challenge. Since overexpression of LecRKs proved to be more effective
(Bouwmeester et al., 2011a; Singh et al., 2012; Chapter IV; Chapter VI), increased
resistance could be achieved by boosting the expression level of LecRKs.
Due to species-specific expansion of LecRKs, not all LecRKs which are
functional in plant defence have homologs present in different plant species (Chapter
VI). For example, LecRK-I.9 from the cruciferous plant Arabidopsis is absent in
N. benthamiana, tomato and rice. Therefore, transfer of LecRKs across plant families
provides a new avenue to increase disease resistance. Thus far, three Arabidopsis
LecRKs have been transferred into the Solanaceous plants and all of them proved to
maintain their function in plant defence. Ectopic expression of LecRK-I.9 in potato
(Solanum tuberosum) increases constitutive expression of protease inhibitor genes
and enhances resistance to Phytophthora infestans (Bouwmeester et al., 2014).
Similarly, ectopic expression of Arabidopsis LecRK-I.9 and LecRK-IX.1 in
N. benthamiana increases resistance to P. infestans (Chapter V). Although LecRK-I.9
and LecRK-IX.1 were identified for resistance against P. brassicae and P. capsici in
Arabidopsis, both confer resistance to P. infestans when transferred into
Solanaceous plants. P. infestans is phylogenetically distant from P. brassicae and
P. capsici and this again supports the idea that LecRKs do not discriminate between
Phytophthora species. N. benthamiana plants ectopically expressing Arabidopsis
LecRK-VI.2 showed increased response to flg22, − e.g. enhanced ROS production,
callose deposition, MAP kinase activity and defence-related gene expression − which
might account for the increased bacterial resistance detected in the transgenic lines
(Huang et al., 2014). The retained function of these LecRKs indicates that the
downstream signaling pathways are conserved between different plant families. As

General discussion
195
such, LecRKs have potential to be transferred into distantly related species to
increase disease resistance.
A disadvantage of ectopic/overexpression of defence-related genes is that it
often results in pleiotropic side effects that affect plant fitness (Gurr and Rushton,
2005). This is also the case for LecRKs. Overexpression of LecRK-I.9 in Arabidopsis
or in other plant species such as potato and N. benthamiana results in aberrant plant
development (Bouwmeester et al., 2011a; Bouwmeester et al., 2014; Chapter V).
The severity of morphological changes in transgenic potatoes was found to be
correlated with transgene expression levels. Similarly, expression of LecRK-IX.1 in
Arabidopsis or in N. benthamiana leads to plant cell death and this phenotype was
also found to be correlated with LecRK-IX.1 expression levels (Chapter IV; Chapter
V). To minimize side effects, LecRKs could be modified by using plant inducible
promoters that allow the fine-tuning of transgene expression (Gurr and Rushton,
2005). In the case of LecRK-IX.1 and LecRK-IX.2, their function in Phytophthora
disease resistance is independent of that in plant cell death induction (Chapter IV),
and therefore it should be possible to manipulate the signaling pathways to suppress
cell death induction without losing the LecRK-mediated disease resistance. Another
alternative is to identify the key residues in LecRKs controlling disease resistance
and cell death induction through random mutagenesis. Mutation of these key
residues is expected to improve the resistance and reduce the fitness costs
associated with expression of LecRKs.
Model pathosystems facilitate functional studies of LecRKs
Undoubtedly, functional analysis of LecRKs benefits from established model
pathosystems. Previously several pathosystems have been described in which the
interactions between Arabidopsis and different Phytophthora species were
characterized. In those studies, different Phytophthora isolates were tested on
Arabidopsis accessions to identify interaction specificity among accession-isolate
combinations. For Phytophthora parasitica and Phytophthora cinnamomi, the tested
isolates turned out to be either infectious on all the selected Arabidopsis accessions
or unable to infect any (Robinson and Cahill, 2003; Wang et al., 2011). In the case of
P. brassicae on Arabidopsis, various compatible and incompatible interactions with
Arabidopsis were detected among accession-isolate combinations (Roetschi et al.,
2001). Isolates with moderate virulence were identified that are virulent only on part

Chapter VII
196
of the selected accessions. For example, isolate HH is not capable to infect Col-0 but
causes disease on Ler-0. Moreover, it causes disease on Col-0 mutants defective in
biosynthesis of the anti-microbial secondary metabolites camalexin and indole
glucosinolates or on the LecRK-I.9 mutants (Schlaeppi et al., 2010; Bouwmeester et
al., 2011a). In Chapter II, we described a pathosystem between Arabidopsis and
P. capsici. We found an isolate, i.e. LT123, that behaves similarly as P. brassicae HH.
Although isolate LT123 is not capable to infect Col-0, it causes disease on a variety
of Col-0 mutants defective in defence-related signaling. Hence, P. capsici isolate
LT123 proved to be useful to distinguish essential components for plant resistance.
The prominent advantage of these model pathosystems is that we can make use of
the public Arabidopsis mutant resources. Screening of Arabidopsis T-DNA insertion
lines has been considered as an effective tool to link phenotype to gene function.
Disease phenotyping of a large set of LecRK T-DNA insertion lines using various
pathogens showing different levels of virulence identified several LecRKs that play a
role in resistance. In this way, we identified LecRKs with function specificity or varying
resistance capacity. Nevertheless, we still may have overlooked some LecRKs that
function redundantly with other genes or play a subtle role in plant defence.
Conclusions and prospective
This thesis describes our efforts to study LecRKs in different plant species to
understand their role in plant immunity. Although LecRKs are not as effective as the
classical NLR proteins that mediate effector-triggered immunity to stop pathogen
infection, the resistance provided by LecRKs was proved to be broad-spectrum
instead of race-specific. Therefore, LecRKs can provide an additional layer of
defence. Pyramiding LecRKs with classical R genes is expected to give a
quantitative synergistic effect that could lead to broad-spectrum disease resistance.
The first research article that made notice of a LecRK being a receptor for
monitoring the plant pathogen effector IPI-O dates from 2006 (Gouget et al., 2006).
Despite the increasing interest in LecRKs in recent years, our current understanding
on the molecular basis of LecRKs in plant defence is still in its infancy. To obtain a
comprehensive overview, many questions remain to be answered. For example, how
do these LecRKs fit into the defence signaling cascades? How are these LecRKs
activated to implement resistance function? Do other LecRKs besides LecRK-I.9
perceive eATP and are LecRKs receptors of DAMPs or MAMPs? Which components

General discussion
197
are required for LecRK-mediated signaling? Since interactions between plants and
pathogens involve bilateral communication, it is also necessary to understand how
pathogens respond to LecRK-mediated resistance and how pathogens with high
virulence suppress LecRK-mediated defence. It is anticipated that integration of
forward and reverse genetics, biochemistry approaches, bioinformatics and
pathogenomics will allow us to answer some the above-mentioned questions in the
near future.
REFERENCES Antolin-Llovera, M., Ried, M.K., Binder, A. and Parniske, M. (2012). Receptor kinase signaling
pathways in plant-microbe interactions. Annual Review of Phytopathology, 50, 451-473.
Barre, A., Hervé, C., Lescure, B. and Rougé, P. (2002). Lectin receptor kinases in plants. Critical
Reviews in Plant Sciences, 21, 379-399.
Boller, T. and Felix, G. (2009). A renaissance of elicitors: Perception of microbe-associated molecular
patterns and danger signals by pattern-recognition receptors. Annual Review of Plant Biology, 60,
379-406.
Bours, M.J.L., Swennen, E.L.R., Di Virgilio, F., Cronstein, B.N. and Dagnelie, P.C. (2006). Adenosine
5'-triphosphate and adenosine as endogenous signaling molecules in immunity and inflammation.
Pharmacology & Therapeutics, 112, 358-404.
Bouwmeester, K. and Govers, F. (2009). Arabidopsis L-type lectin receptor kinases: phylogeny,
classification, and expression profiles. Journal of Experimental Botany, 60, 4383-4396.
Bouwmeester, K., de Sain, M., Weide, R., Gouget, A., Klamer, S., Canut, H. and Govers, F. (2011a).
The lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a
potential host target for a RXLR effector. PLoS Pathogens, 7, e1001327.
Bouwmeester, K., Meijer, H.J.G. and Govers, F. (2011b). At the frontier; RXLR effectors crossing the
Phytophthora-host interface. Frontiers in Plant Science, 2, 75.
Bouwmeester, K., Han, M., Blanco-Portales, R., Song, W., Weide, R., Guo, L., van der Vossen, E.A.G.
and Govers, F. (2014). The Arabidopsis lectin receptor kinase LecRK-I.9 enhances resistance to
Phytophthora infestans in Solanaceous plants. Plant Biotechnology Journal, 12, 10-16.
Cao, Y., Tanaka, K., Nguyen, C.T. and Stacey, G. (2014). Extracellular ATP is a central signaling
molecule in plant stress responses. Current Opinion in Plant Biology, 20, 82-87.
Champouret, N., Bouwmeester, K., Rietman, H., van der Lee, T., Maliepaard, C., Heupink, A., van de
Vondervoort, P.J.I., Jacobsen, E., Visser, R.G.F., van der Vossen, E.A.G., et al. (2009).
Phytophthora infestans isolates lacking class I ipiO variants are virulent on Rpi-blb1 potato.
Molecular Plant-Microbe Interactions, 22, 1535-1545.
Chataigneau, T., Lemoine, D. and Grutter, T. (2013). Exploring the ATP-binding site of P2X receptors.
Frontiers in Cellular Neuroscience, 7, 273.

Chapter VII
198
Chinchilla, D., Shan, L., He, P., de Vries, S. and Kemmerling, B. (2009). One for all: the receptor-
associated kinase BAK1. Trends in Plant Science, 14, 535-541.
Choi, J., Tanaka, K., Cao, Y., Qi, Y., Qiu, J., Liang, Y., Lee, S.Y. and Stacey, G. (2014). Identification of
a plant receptor for extracellular ATP. Science, 343, 290-294.
Clark, G. and Roux, S.J. (2011). Apyrases, extracellular ATP and the regulation of growth. Current
Opinion in Plant Biology, 14, 700-706.
De Smet, I., Vosz, U., Jurgens, G. and Beeckman, T. (2009). Receptor-like kinases shape the plant.
Nature Cell Biology, 11, 1166-1173.
Demidchik, V., Shang, Z.L., Shin, R., Thompson, E., Rubio, L., Laohavisit, A., Mortimer, J.C., Chivasa,
S., Slabas, A.R., Glover, B.J., et al. (2009). Plant extracellular ATP signalling by plasma membrane
NADPH oxidase and Ca2+ channels. The Plant Journal, 58, 903-913.
Desclos-Theveniau, M., Arnaud, D., Huang, T.Y., Lin, G.J.C., Chen, W.Y., Lin, Y.C. and Zimmerli, L.
(2012) The Arabidopsis lectin receptor kinase LecRK-V.5 represses stomatal immunity induced by
Pseudomonas syringae pv. tomato DC3000. PLoS Pathogens, 8, e1002513.
Dickman, M.B. and Fluhr, R. (2013). Centrality of host cell death in plant-microbe interactions. Annual
Review of Phytopathology, 51, 543-570.
Etzler, M.E., Surolia, A. and Cummings, R. D. (2009). L-type Lectins. In: Essentials of glycobiology,
Cold Spring Harbor, 415–424.
Gouget, A., Senchou, V., Govers, F., Sanson, A., Barre, A., Rouge, P., Pont-Lezica, R.P. and Canut, H.
(2006). Lectin receptor kinases participate in protein-protein interactions to mediate plasma
membrane-cell wall adhesions in Arabidopsis. Plant Physiology, 140, 81-90.
Gurr, S.J. and Rushton, P.J. (2005). Engineering plants with increased disease resistance: how are we
going to express it? Trends in Biotechnology, 23, 283-290.
Huang, P., Ju, H.W., Min, J.H., Zhang, X., Kim, S.H., Yang, K.Y. and Kim, C.S. (2013) Overexpression
of L-type lectin-like protein kinase 1 confers pathogen resistance and regulates salinity response in
Arabidopsis thaliana. Plant Science, 203, 98-106.
Huang, P.Y., Yeh, Y.H., Liu, A.C., Chien, C.P. and Zimmerli, L. (2014). The Arabidopsis LecRK-VI.2
associates with the pattern-recognition receptor FLS2 and primes Nicotiana benthamiana pattern-
triggered immunity. The Plant Journal, 79, 243–255.
Huffaker, A., Pearce, G. and Ryan, C.A. (2006). An endogenous peptide signal in Arabidopsis
activates components of the innate immune response. Proceedings of the National Academy of
Sciences of the United States of America, 103, 10098-10103.
Joshi, A., Dang, H., Vaid, N. and Tuteja, N. (2010). Pea lectin receptor-like kinase promotes high
salinity stress tolerance in bacteria and expresses in response to stress in planta. Glycoconjugate
Journal, 27, 133-150.
Kadota, Y., Sklenar, J., Derbyshire, P., Stransfeld, L., Asai, S., Ntoukakis, V., Jones, J.D.G., Shirasu,
K., Menke, F., Jones, A. and Zipfel, C. (2014). Direct regulation of the NADPH oxidase RBOHD by
the PRR-associated kinase BIK1 during plant immunity. Molecular Cell, 54, 43-55.
Kang, J., Hwang, J.U., Lee, M., Kim, Y.Y., Assmann, S.M., Martinoia, E. and Lee, Y. (2010). PDR-type
ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proceedings of the

General discussion
199
National Academy of Sciences of the United States of America, 107, 2355-2360.
Kiba, A., Sugimoto, M., Toyoda, K., Ichinose, Y., Yamada, T. and Shiraishi, T. (1998). Interaction
between cell wall and plasma membrane via RGD motif is implicated in plant defense responses.
Plant & Cell Physiology, 39, 1245-1249.
Lehti-Shiu, M.D., Zou, C., Hanada, K. and Shiu, S.H. (2009). Evolutionary history and stress regulation
of plant receptor-like kinase/Pelle genes. Plant Physiology, 150, 12-26.
Li, L., Li, M., Yu, L, Zhou, Z., Liang, X., Liu, Z., Cai, G., Gao, L., Zhang, X., Wang, Y., Chen, S. and
Zhou, J.M. (2014). The FLS2-associated kinase BIK1 directly phosphorylates the NADPH oxidase
RbohD to control plant immunity. Cell Host & Microbe, 15, 329-338.
Liebrand, T.W.H., van den Burg, H.A. and Joosten, M.H.A.J. (2014). Two for all: Receptor-associated
kinases SOBIR1 and BAK1. Trends in Plant Science, 19, 123-132.
Liu, T., Liu, Z., Song, C., Hu, Y., Han, Z., She, J., Fan, F., Wang, J., Jin, C., Chang, J., Zhou, J.M. and
Chai, J. (2012). Chitin-induced dimerization activates a plant immune receptor. Science, 336, 1160-
1164.
Lu, D., Wu, S., Gao, X., Zhang, Y., Shan, L. and He, P. (2010). A receptor-like cytoplasmic kinase,
BIK1, associates with a flagellin receptor complex to initiate plant innate immunity. Proceedings of
the National Academy of Sciences of the United States of America, 107, 496-501.
Macho, A.P. and Zipfel, C. (2014). Plant PRRs and the activation of innate immune signaling.
Molecular Cell, 54, 263-272.
Mellersh, D.G. and Heath, M.C. (2001). Plasma membrane-cell wall adhesion is required for
expression of plant defense responses during fungal penetration. Plant Cell, 13, 413-424.
Melotto, M., Underwood, W., Koczan, J., Nomura, K. and He, S. (2006). The innate immune function of
plant stomata against bacterial invasion. Cell, 126, 969-980.
Navarro-Gochicoa, M.T., Camut, S., Timmers, A.C.J., Niebel, A., Herve, C., Boutet, E., Bono, J.J.,
Imberty, A. and Cullimore, J.V. (2003). Characterization of four lectin-like receptor kinases
expressed in roots of Medicago truncatula. Structure, location, regulation of expression, and
potential role in the symbiosis with Sinorhizobium meliloti. Plant Physiology, 133, 1893-1910.
Nishiguchi, M., Yoshida, K., Sumizono, T. and Tazaki, K. (2002). A receptor-like protein kinase with a
lectin-like domain from lombardy poplar: gene expression in response to wounding and
characterization of phosphorylation activity. Molecular Genetics and Genomics, 267, 506-514.
Phillips, S.M., Dubery, I.A. and van Heerden, H. (2013). Identification and molecular characterisation
of a lectin receptor-like kinase (GhLecRK-2) from Cotton. Plant Molecular Biology Reporter, 31, 9-
20.
Ralevic, V. and Burnstock, G. (1998). Receptors for purines and pyrimidines. Pharmacological
Reviews, 50, 413-492.
Robinson, L.H. and Cahill, D.M. (2003). Ecotypic variation in the response of Arabidopsis thaliana to
Phytophthora cinnamomi. Australasian Plant Pathology, 32, 53-64.
Roetschi, A., Si-Ammour, A., Belbahri, L., Mauch, F. and Mauch-Mani, B. (2001). Characterization of
an Arabidopsis-Phytophthora pathosystem: Resistance requires a functional PAD2 gene and is
independent of salicylic acid, ethylene and jasmonic acid signalling. The Plant Journal, 28, 293-

Chapter VII
200
305.
Ruoslahti, E. (1996). RGD and other recognition sequences for integrins. Annual Review of Cell and
Developmental Biology, 12, 697-715.
Sanchez-Rodriguez, C., Estevez, J.M., Llorente, F., Hernandez-Blanco, C., Jorda, L., Pagan, I.,
Berrocal, M., Marco, Y., Somerville, S. and Molina, A. (2009). The ERECTA receptor-like kinase
regulates cell wall-mediated resistance to pathogens in Arabidopsis thaliana. Molecular Plant-
Microbe Interactions, 22, 953-963.
Schlaeppi, K., Abou-Mansour, E., Buchala, A. and Mauch, F. (2010). Disease resistance of Arabidopsis
to Phytophthora brassicae is established by the sequential action of indole glucosinolates and
camalexin. The Plant Journal, 62, 840-851.
Senchou, V., Weide, R., Carrasco, A., Bouyssou, H., Pont-Lezica, R., Govers, F. and Canut, H. (2004).
High affinity recognition of a Phytophthora protein by Arabidopsis via an RGD motif. Cellular and
Molecular Life Sciences, 61, 502-509.
Seo, S., Gomi, K., Kaku, H., Abe, H., Seto, H., Nakatsu, S., Neya, M., Kobayashi, M., Nakaho, K.,
Ichinose, Y., et al. (2012). Identification of natural diterpenes that inhibit bacterial wilt disease in
tobacco, tomato and Arabidopsis. Plant & Cell Physiology, 53, 1432-1444.
Shiu, S.H., and Bleecker, A.B. (2001a). Plant receptor-like kinase gene family: diversity, function, and
signaling. Science Signaling, 113, re22.
Shiu, S.H. and Bleecker, A.B. (2001b). Receptor-like kinases from Arabidopsis form a monophyletic
gene family related to animal receptor kinases. Proceedings of the National Academy of Sciences
of the United States of America, 98, 10763-10768.
Shiu, S.H. and Bleecker, A.B. (2003). Expansion of the receptor-like kinase/Pelle gene family and
receptor-like proteins in Arabidopsis. Plant Physiology, 132, 530-543.
Shiu, S.H., Karlowski, W.M., Pan, R., Tzeng, Y.H., Mayer, K.F.X. and Li, W.H. (2004). Comparative
analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell, 16, 1220-1234.
Singh, P., Kuo, Y.C., Mishra, S., Tsai, C.H., Chien, C.C., Chen, C.W., Desclos-Theveniau, M., Chu,
P.W., Schulze, B., Chinchilla, D., Boller, T. and Zimmerli, L. (2012). The lectin receptor kinase-VI.2
is required for priming and positively regulates Arabidopsis pattern-triggered immunity. Plant Cell,
24, 1256-1270.
Singh, P., Chien, C.C., Mishra, S., Tsai, C.H. and Zimmerli, L. (2013). The Arabidopsis LECTIN
RECEPTOR KINASE-VI.2 is a functional protein kinase and is dispensable for basal resistance to
Botrytis cinerea. Plant Signaling & Behavior, 8, 1-3.
Song, C.J., Steinebrunner, I., Wang, X.Z., Stout, S.C. and Roux, S.J. (2006). Extracellular ATP induces
the accumulation of superoxide via NADPH oxidases in Arabidopsis. Plant Physiology, 140, 1222-
1232.
Sun, Y., Li, L., Macho, A.P., Han, Z., Hu, Z., Zipfel, C., Zhou, J.M. and Chai, J. (2013). Structural basis
for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex. Science, 342, 624-
628.
Tanaka, K., Gilroy, S., Jones, A.M. and Stacey, G. (2010). Extracellular ATP signaling in plants. Trends
in Cell Biology, 20, 601-608.

General discussion
201
Tang, W., Brady, S.R., Sun, Y., Muday, G.K. and Roux, S.J. (2003). Extracellular ATP inhibits root
gravitropism at concentrations that inhibit polar auxin transport. Plant Physiology, 131, 147-154.
Vaid, N., Pandey, P.K. and Tuteja, N. (2012). Genome-wide analysis of lectin receptor-like kinase
family from Arabidopsis and rice. Plant Molecular Biology, 80, 365-388.
van Damme, E.J.M., Lannoo, N. and Peumans, W.J. (2008). Plant lectins. Advances in Botanical
Research, 48, 107-209.
van der Vossen, E., Sikkema, A., Hekkert, B.T.L., Gros, J., Stevens, P., Muskens, M., Wouters, D.,
Pereira, A., Stiekema, W. and Allefs, S. (2003). An ancient R gene from the wild potato species
Solanum bulbocastanum confers broad-spectrum resistance to Phytophthora infestans in
cultivated potato and tomato. The Plant Journal, 36, 867-882.
van der Vossen, E.A.G., Gros, J., Sikkema, A., Muskens, M., Wouters, D., Wolters, P., Pereira, A. and
Allefs, S. (2005). The Rpi-blb2 gene from Solanum bulbocastanum is a Mi-1 gene homolog
conferring broad-spectrum late blight resistance in potato. The Plant Journal, 44, 208-222.
Wang, Y., Meng, Y., Zhang, M., Tong, X., Wang, Q., Sun, Y., Quan, J., Govers, F. and Shan, W. (2011).
Infection of Arabidopsis thaliana by Phytophthora parasitica and identification of variation in host
specificity. Molecular Plant Pathology, 12, 187-201.
Wang, Y., Bouwmeester, K., van de Mortel, J.E., Shan, W. and Govers, F. (2013). A novel Arabidopsis-
oomycete pathosystem: differential interactions with Phytophthora capsici reveal a role for
camalexin, indole glucosinolates and salicylic acid in defence. Plant, Cell & Environment, 36, 1192-
1203.
Wang, Y., Bouwmeester, K., Beseh, P., Shan, W. and Govers, F. (2014). Phenotypic analyses of
Arabidopsis T-DNA insertion lines and expression profiling reveal that multiple L-type lectin
receptor kinases are involved in plant immunity. Molecular Plant-Microbe Interactions, 27, 1390-
1402.
Wu, T., Wang, R., Xu, X., He, X., Sun, B., Zhong, Y., Liang, Z., Luo, S. and Lin, Y. (2014). Cucumis
sativus L-type lectin receptor kinase (CsLecRK) gene family respond to Phytophthora melonis,
Phytophthora capsici and water immersion in disease resistant and susceptible cucumber cultivars.
Gene, 549, 214-222..
Yamaguchi, Y., Huffaker, A., Bryan, A., Tax, F. and Ryan, C. (2010). PEPR2 is a second receptor for
the Pep1 and Pep2 peptides and contributes to defense responses in Arabidopsis. Plant Cell, 22,
508-522.
Zipfel, C., Kunze, G., Chinchilla, D., Caniard, A., Jones, J. D. G., Boller, T. and Felix, G. (2006).
Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated
transformation. Cell, 125, 749-760.


203
Summary
Phytophthora pathogens are notorious for causing severe damage to many
agriculturally and ornamentally important plants. Effective plant resistance depends
largely on the capacity to perceive pathogens and to activate rapid defence.
Cytoplasmic resistance (R) proteins are well-known for activation of plant immunity
upon recognition of matching effectors secreted by Phytophthora. However,
Phytophthora pathogens are notoriously difficult to control due to their rapid
adaptation to evade R protein-mediated recognition. Hence, exploring novel
resistance components is instrumental for developing durable resistance.
Receptor-like kinases (RLKs) function as important sentinels in sensing exogenous
and endogenous stimuli to initiate plant defence. One RLK that was previously
identified as a novel Phytophthora resistance component is the Arabidopsis L-type
lectin receptor kinase LecRK-I.9. This RLK belongs to a multigene family consisting of
45 members in Arabidopsis but whether or not the other members function in
Phytophthora resistance was thus far unknown. The research described in this thesis
was aimed at unravelling the role of LecRKs in plant immunity, in particular to
Phytophthora pathogens.
Chapter I describes various Phytophthora diseases and the current
understanding of the mechanisms underlying plant innate immunity with emphasis on
disease resistance to Phytophthora pathogens.
In Chapter II, we describe the development of a new Arabidopsis-Phytophthora
pathosystem. We demonstrated that Phytophthora capsici is capable to infect
Arabidopsis. Inoculation assays and cytological analysis revealed variations among
Arabidopsis accessions in response to different P. capsici isolates. Moreover, infection
assays on Arabidopsis mutants with specific defects in defence showed that salicylic
acid signaling, camalexin and indole glucosinolates biosynthesis pathways are
required for P. capsici resistance (Chapter IIa). The importance of these pathways in
Arabidopsis resistance was supported by the finding that the corresponding marker
genes are induced upon infection by P. capsici (Chapter IIb). This model pathosystem
can be used as an additional tool to pinpoint essential components of Phytophthora
resistance.
We then exploited Arabidopsis-Phytophthora pathosystems to uncover the role of
LecRKs in Phytophthora resistance. In Chapter III we describe a systematic
phenotypic characterization of a large set of Arabidopsis LecRK T-DNA insertion lines.
The T-DNA insertion lines were assembled and assayed for their response towards
different Phytophthora pathogens. This revealed that next to LecRK-I.9, several other
LecRKs function in Phytophthora resistance. We have also analysed whether the

Summary
204
LecRKs are involved in response to other biotic and abiotic stimuli. Several T-DNA
insertion lines showed altered responses to bacterial or fungal pathogens, but none of
the lines showed visible developmental changes under normal conditions or upon
abiotic stress treatment. Combining these phenotypic data with LecRK expression
profiles obtained from publicly available datasets revealed that LecRKs that are hardly
induced or even suppressed upon infection, might still have a function in pathogen
resistance. Computed co-expression analysis revealed that LecRKs with similar
function display diverse expression patterns.
Arabidopsis LecRK clade IX comprises two members. T-DNA insertion mutants of
both LecRK-IX.1 and LecRK-IX.2 showed gain of susceptibility to non-adapted
Phytophthora isolates and therefore the role of these two LecRKs in Phytophthora
resistance was further investigated. In Chapter IV we describe that overexpression of
either LecRK-IX.1 or LecRK-IX.2 in Arabidopsis resulted in increased resistance to
Phytophthora, but also induced plant cell death. A mutation in the kinase domain
abolished the ability of LecRK-IX.1 and LecRK-IX.2 to induce Phytophthora resistance
as did deletion of the lectin domain. Cell death induction however, only required the
kinase, not the lectin domain. Since transient expression of both LecRKs in
Nicotiana benthamiana also resulted in increased Phytophthora resistance and
induction of cell death, we used N. benthamiana to explore downstream components
required for LecRK-IX.1- and LecRK-IX.2-mediated Phytophthora resistance and cell
death. Virus-induced gene silencing of candidate signaling genes revealed that
NbSIPK1/NPT4 is essential for LecRK-IX.1-mediated cell death but not for
Phytophthora resistance. Collectively, these results illustrate that the Phytophthora
resistance mediated by LecRK-IX.1 and LecRK-IX.2 is independent of the cell death
phenotype. By co-immunoprecipitation we identified putative interacting proteins, one
of which was an ATP-binding cassette (ABC) transporter. A homolog in Arabidopsis,
the ABC transporter ABCG40, was found to interact in planta with both LecRK-IX.1
and LecRK-IX.2. Similar to the LecRK mutants, Arabidopsis ABCG40 mutants
showed compromised Phytophthora resistance, indicating that ABCG40 has a
function in Phytophthora resistance.
In Chapter V, we describe the generation of stable transgenic N. benthamiana
plants expressing Arabidopsis LecRK-I.9 or LecRK-IX.1. Multiple transgenic lines
were obtained varying in transgene copy number and transgene expression level.
Ectopic expression of LecRK-I.9 resulted in reduced plant sizes and aberrant leaf
morphology. In addition, expression of LecRK-IX.1 induced plant cell death.
Transgenic N. benthamiana lines expressing either LecRK-I.9 or LecRK-IX.1 showed
increased resistance towards P. capsici or Phytophthora infestans. This demonstrated
that Arabidopsis LecRK-I.9 and LecRK-IX.1 retained their role in Phytophthora
resistance upon interfamily transfer.

Summary
205
Based on the results obtained on Arabidopsis LecRKs, we speculated that
LecRKs in other plant species could play a similar role in Phytophthora resistance. In
Chapter VI, we focus on LecRKs in two Solanaceous plants, i.e. N. benthamiana and
tomato. By exploring genome databases, we identified 38 and 22 LecRKs in
N. benthamiana and tomato, respectively. Phylogenetic analysis revealed that both
N. benthamiana and tomato lack LecRKs homologous to Arabidopsis LecRKs of
clades I, II, III and V, but contain a Solanaceous-specific clade of LecRKs. Functional
analysis of various Solanaceous LecRKs using virus-induced gene silencing followed
by infection assays revealed that homologs of Arabidopsis LecRK-IX.1 and
LecRK-IX.2 in N. benthamiana and tomato are implicated in Phytophthora resistance.
These results indicate that the role of clade IX LecRKs in Phytophthora resistance is
conserved across plant species.
In Chapter VII, the experimental data presented in this thesis are summarized
and discussed in a broader context. We present an overview of the current
understanding of LecRKs in plant immunity and discuss how LecRKs can be exploited
to improve plant resistance.

206
Samenvatting
Plantenziekten veroorzaakt door Phytophthora pathogenen richten ernstige schade
aan in belangrijke land- en tuinbouwgewassen en in de sierteelt. Effectieve
ziekteresistentie in planten is grotendeels gebaseerd op een vroegtijdige herkenning
van pathogenen gevolgd door initiatie van afweer. Immuniteit wordt bewerkstelligd
door intracellulaire resistentie-eiwitten (R) die geactiveerd worden door herkenning
van Phytophthora effectoren. Phytophthora pathogenen zijn echter moeilijk te
bestrijden. Ze kunnen zich gemakkelijk aanpassen waardoor R-gen-afhankelijke
resistentie vaak snel wordt doorbroken. Voor het ontwikkelen van duurzame
resistentie is het daarom van belang om alternatieve resistentie-componenten te
identificeren. Receptor kinases spelen een cruciale rol in de herkenning van exogene
en endogene stimuli en in het activeren van afweer in de plant. Eerder werd
aangetoond dat LecRK-I.9, een Arabidopsis L-type lectine receptor kinase, fungeert
als een Phytophthora resistentie-component. LecRK-I.9 is een van de 45 leden van
de LecRK familie in Arabidopsis. Of, en hoe, andere LecRKs functioneren in
Phytophthora resistentie is tot dusver niet onderzocht. Het in dit proefschrift
beschreven onderzoek was gericht op het ontrafelen van een potentiële rol van
LecRKs in resistentie, in het bijzonder tegen Phytophthora pathogenen.
Hoofdstuk I beschrijft verschillende plantenziekten veroorzaakt door
Phytophthora en geeft een overzicht van de huidige kennis van immuniteit tegen
plantenpathogenen, met nadruk op resistentie tegen Phytophthora pathogenen.
In hoofdstuk II beschrijven we de ontwikkeling van een nieuw
Arabidopsis-Phytophthora pathosysteem. We toonden aan dat Phytophthora capsici
in staat is Arabidopsis te infecteren. Infectietoetsen en cytologische analyses
onthulden verschillen in afweer tussen Arabidopsis accessies in reactie op infectie
met verschillende P. capsici isolaten. Uit infectietoetsen op Arabidopsis mutanten met
verstoorde signaleringsroutes bleek dat salicylzuur-afhankelijke signalering, en
biosynthese van camalexin en indoolglucosinolaten nodig zijn voor een effectieve
resistentie tegen P. capsici (hoofdstuk IIa). Het belang van deze signaleringsroutes
voor afweer werd ondersteund door waarnemingen dat expressie van bijbehorende
markergenen wordt geïnduceerd na infectie met P. capsici (hoofdstuk IIb). Dit
pathosysteem kan dus gebruikt worden als model om essentiële componenten van
resistentie tegen Phytophthora te identificeren.
Vervolgens gebruikten we verschillende Arabidopsis-Phytophthora
pathosystemen om de potentiële rol van LecRKs in Phytophthora resistentie te
onderzoeken. Hoofdstuk III beschrijft een systematische fenotypische karakterisering
van Arabidopsis LecRK mutanten. In een collectie van T-DNA insertielijnen werd de

Samenvatting
207
respons op infectie met verschillende Phytophthora pathogenen onderzocht. Hieruit
bleek dat, naast LecRK-I.9, ook andere LecRKs een rol spelen in Phytophthora
resistentie. Daarnaast onderzochten we of LecRKs ook belangrijk zijn voor de
respons op andere pathogenen en op abiotische stressfactoren. Verscheidene T-DNA
insertielijnen vertoonden fenotypische verschillen in vatbaarheid voor pathogene
bacteriën en schimmels, maar geen van deze Arabidopsis lijnen vertoonden
afwijkingen in groei en ontwikkeling onder normale omstandigheden of na
blootstelling aan abiotische stress. Deze fenotypische gegevens werden gekoppeld
aan LecRK expressieprofielen beschikbaar in publiekelijk toegankelijke
databestanden. Hieruit bleek dat LecRKs waarvan de expressie na infectie niet of
nauwelijks wordt geïnduceerd, of zelfs wordt onderdrukt, mogelijk toch een functie
hebben in resistentie tegen pathogenen. Verder bleek uit deze co-expressie analyses
dat LecRKs die een vergelijkbare rol hebben kunnen verschillen in expressiepatroon.
LecRK clade IX in Arabidopsis bestaat uit slechts twee leden. T-DNA insertielijnen
van zowel LecRK-IX.1 als LecRK-IX.2 vertoonden een verhoogde vatbaarheid voor
Phytophthora, en daarom werden deze twee LecRKs in meer detail onderzocht. In
hoofdstuk IV beschrijven we dat verhoogde constitutieve expressie van LecRK-IX.1
of LecRK-IX.2 in Arabidopsis resulteerde in een verhoogde resistentie tegen
Phytophthora, maar tevens in spontane inductie van celdood. Door een mutatie in het
kinasedomein van LecRK-IX.1 of LecRK-IX.2, verdween niet alleen de verhoogde
Phytophthora resistentie, ook de spontane celdood trad niet meer op. Echter, deletie
van het lectine-domein van beide LecRKs had alleen een effect op de Phytophthora
resistentie; het spontane celdood fenotype bleef behouden en is dus alleen
afhankelijk van het kinase-domein, niet het lectine-domein. Transiënte expressie van
beide LecRK-IX genen in Nicotiana benthamiana resulteerde ook in een verhoogde
resistentie tegen Phytophthora en in spontane celdood. Dit bood de mogelijkheid om
N. benthamiana te gebruiken voor het in kaart te brengen van de benodigde
signaleringscomponenten voor LecRK-IX.1- en LecRK-IX.2-afhankelijke
Phytophthora resistentie en celdood. Met behulp van virus-geïnduceerde silencing
van genen betrokken bij verscheidende signaleringsroutes konden we aantonen dat
NbSIPK1/NPT4 essentieel is voor LecRK-IX.1-afhankelijke celdood, maar niet voor
Phytophthora resistentie. Uit deze resultaten blijkt dat Phytophthora resistentie
gebaseerd op LecRK-IX.1 of LecRK-IX.2 onafhankelijk is van het celdood fenotype.
Door middel van co-immunoprecipitatie werden eiwitten geïdentificeerd die
interacteren met de clade IX LecRKs. Dit resulteerde in de selectie van een ABC
transporter (ATP-binding cassette transporter), die sterke homologie vertoond met de
ABC transporter ABCG40 in Arabidopsis. ABCG40 bleek in planta te interacteren met
zowel LecRK-IX.1 als LecRK-IX.2 en, net als de LecRK-IX mutanten, vertoonden
Arabidopsis ABCG40 mutanten een verhoogde vatbaarheid voor Phytophthora. Deze

Samenvatting
208
resultaten suggereren dat ABCG40 een rol speelt in Phytophthora resistentie.
In hoofdstuk V beschrijven we het genereren van stabiele transgene N.
benthamiana planten die Arabidopsis LecRK-I.9 of LecRK-IX.1 constitutief tot
expressie brengen. Meerdere transgene lijnen werden verkregen met variaties in het
aantal transgen kopieën en in transgen expressieniveaus. Constitutieve expressie
van LecRK-I.9 resulteerde in kleinere planten met een afwijkende bladmorfologie
terwijl hoge constitutieve expressie van LecRK-IX.1 celdood veroorzaakte. De
transgene N. benthamiana lijnen met LecRK-I.9 of LecRK-IX.1 als transgen
vertoonden een verhoogde weerstand tegen P. capsici en Phytophthora infestans. Dit
wijst erop dat de Arabidopsis LecRK-I.9 en LecRK-IX.1 genen hun rol in Phytophthora
resistentie in een N. benthamiana achtergrond kunnen handhaven.
Op basis van de resultaten verkregen met de Arabidopsis LecRKs, speculeerden
we dat LecRKs in andere plantensoorten ook een rol hebben in Phytophthora
resistentie. Hoofdstuk VI is gericht op LecRKs in N. benthamiana en tomaat, twee
soorten in de nachtschadefamilie. Door het exploreren van genoomdatabanken
konden we 38 LecRKs in N. benthamiana en 22 LecRKs in tomaat identificeren. Uit
fylogenetische analyses bleek dat N. benthamiana en tomaat een
nachtschade-specifieke LecRK clade bevatten, maar geen homologen van
Arabidopsis LecRKs in clades I, II, III en V. Uit functionele analyse van LecRKs met
behulp van virus-geïnduceerde silencing van de betreffende genen gevolgd door
infectietoetsen, bleek dat homologen van Arabidopsis LecRK-IX.1 en LecRK-IX.2 in
zowel N. benthamiana als tomaat betrokken zijn bij Phytophthora resistentie. Deze
resultaten geven aan dat de rol van clade IX LecRKs in Phytophthora resistentie
geconserveerd is in verschillende plantensoorten.
In hoofdstuk VII, zijn de experimentele gegevens uit dit proefschrift samengevat
en besproken in een bredere context. Daarnaast geeft het een overzicht van de
huidige kennis van LecRKs in immuniteit, en bespreken we hoe LecRKs kunnen
worden benut om resistentie in planten te verbeteren.

209
Acknowledgements
Time flies. Four years of study in Wageningen almost comes to an end. I am very excited with the completion of my PhD thesis. However, this achievement would not have been possible without the help and support from my supervisors, friends and family. Here I would like to take this opportunity to thank all of them.
First of all, I would like to thank my supervisor Francine Govers for giving me the opportunity to perform my PhD in your group. I appreciate both your critical suggestions and words of encouragement on the research; I appreciate your supervision in making presentations, slides and writing the articles; I appreciate your patience in correcting spelling errors and grammar mistakes in my manuscripts ; and I also appreciate your encouragements to improve my social skills. Your serious attitude and passion towards science impressed me a lot and definitely you set a good example of being a scientist.
Special thanks to my supervisor Weixing Shan. I was so lucky to study under your supervision during my MSc training. I got the chance to learn many experimental techniques in your lab that were very helpful during my PhD study. I am impressed by your commitment to the research and your charming personality, which were important in shaping my view on science. Thank you for encouraging me to learn English, and stimulating me to go abroad for the PhD study. I appreciate your support and inspiration during my PhD study.
My sincere thanks go to my supervisor Klaas Bouwmeester. Thank you for picking me up at Schiphol in 2010 and your daily supervision. Instead of being a ‘real’ supervisor, you are more like a big funny brother. You are very positive, patient and helpful. You were always available when I asked you for a work discussion. I really appreciate your contributions to my PhD project.
My thanks extend to colleagues and students. I am so lucky to work together with friendly and helpful colleagues in Phytopathology. Thank you guys and girls for giving me a hand when I needed help. I want to thank Rob Weide, Harold Meijer, Chara Schoina, Johan van den Hoogen, Elysa Overdijk and Natalie Verbeek for your help in the lab and the great fun in Pinf-activities. Thanks to the MSc students Patrick Beseh, David Nsibo and Hagos Juhar for your hard work and contributions to my PhD project. Thanks to Judith van de Mortel for generating Arabidopsis iGS mutants and your suggestions on writing the manuscripts present in Chapter II. Thanks to Michael Seidl and Luigi Faino for advice related to bioinformatics and data mining. Thanks to Peter van Esse, Koste Yadeta, Emilie Fradin, Mireille van Damme, Patrick Smit, Daniela Sueldo and Thomas Liebrand for suggestions and encouragement. Thanks to Ali Ormel and Marion Rodenburg for administrative issues. Thanks to Jan Cordewener and Twan America for mass spectrometry analysis. Thanks to Henk Smid, Bert

Acknowledgements
210
Essenstam, Gerrit Stunnenberg and Taede Stoker from Unifarm for growing and taking care of the plants.
Living abroad is not an easy thing for me and I am so lucky to have supportive and kind-hearted friends around. Chenlei Hua, Lisha Zhang, Xu Cheng, Wei Qin, Chunxu Song and Freddy Yeo Kuok San, thank you for ‘taking care of me’ when I arrived in the Netherlands. Thank you for introducing me the nice places to visit and delicious Chinese food and learning me how to cook. Yu Du, we studied in the same university in China, and here we are working in the same group. We are like twins and I always feel very close to you. I am very grateful for your company. Thanks to Zhao Zhang, Miao Han, Guozhi Bi, Yin Song, Jinling Li, Xiaoqian Shi, Furong Liu, Xiaolin Wu, Huayi Li and Jinbin Wu for the funny jokes and nice dinners we had together. Thanks to Feng Zhu, Lisong Ma, Fang Xu, Ke Peng, Hieng Ming Ting, Yafen Lin, Tingting Xiao, Hunchen Li, Hanzi He, Yanxia Zhang, Hui Li, Qing Liu, Ting Yang, Bo Wang, Dong Xiao, Xuan Xu, Juan Du, Tao Li, Dongli Gao, Yunmeng Zhang, Chunzhao Zhao, Ying Zhang, Junyou Wang, Junli Guo, Suxian Zhu and Wei Song for your help in life and the pleasant time.
My sincere appreciations go to my family. I want to express my great gratitude to my parents and parents-in-law for always supporting my decisions and your endless love. Great thanks to my dear brothers(-in-law) Bo Wang, Guoshuai Qiu and sister-in-law Jingjing Sun. Thank you for taking care of our parents and your support to Yinyin and me. I also would like to thank my cute nephews Daofeng Wang, Hongen Qiu, Daoheng Wang and my niece Tian Qiu for the great fun when we talk on the telephone. To my dear husband Yinyin Sun (孙银银), thank you for your constant encouragement, support and endless love.
Yan 20th October 2014
Wageningen

Curr
PhD re
Nether
Dr. Klaa
role of
thesis.
riculum
esearch at
lands unde
as Bouwm
L-type lect
m vita
Yan W
In 200
Northw
June 2
Northw
projec
genes
the su
t the Labo
er the supe
meester. Th
tin recepto
ae
Wang (王燕)
03, she sta
west A&F
2007, she c
west A&F
t on the id
required
pervision o
oratory of
ervision of
e results o
or kinases
) was born
arted her
University,
continued
University
dentificatio
for Phytop
of Prof. W
Phytopath
f Prof. Fran
of the resea
in Phytoph
n on May 6
Bachelor
, Yangling
her Maste
y. She pe
n and cha
phthora pa
Weixing Sha
hology, Wa
ncine Gove
arch that fo
hthora resi
th, 1985 in
study in P
, China. A
r study in P
rformed h
racterizatio
arasitica su
an. In 2010
ageningen
ers, Prof. W
ocused at u
stance are
Shandong
Plant Scie
After gradu
Plant Patho
er MSc re
on of Arab
usceptibility
0, she star
University
Weixing Sh
understand
e described
211
g, China.
nces at
ation in
ology at
esearch
bidopsis
y under
rted her
y in the
han and
ding the
d in this

212
Publications Wang, Y., Bouwmeester, K., Cordewener, J.H.G., America, A.H.P., Shan, W. and
Govers, F. Arabidopsis lectin receptor kinases LecRK-IX.1 and LecRK-IX.2 are
functional analogs in regulating Phytophthora resistance and plant cell death.
Submitted.
Wang, Y., Weide, R., Govers, F. and Bouwmeester, K. L-type lectin receptor kinases
in Nicotiana benthamiana and tomato and their role in Phytophthora resistance.
Submitted.
Deng, F., Wang, Y., Jia, J., Li, J., Govers, F. and Shan, W. The Raf-like MAPKKK
gene Raf36 regulates Arabidopsis thaliana susceptibility to Phytophthora
parasitica and programmed cell death. In preparation.
Wang, Y., Bouwmeester, K., Beseh, P., Shan, W. and Govers, F. (2014). Phenotypic
analyses of Arabidopsis T-DNA insertion lines and expression profiling reveal that
multiple L-type lectin receptor kinases are involved in plant immunity. Molecular
Plant-Microbe Interactions, 27, 1390-1402.
Wang, Y., Bouwmeester, K., van de Mortel, J.E., Shan, W. and Govers, F. (2013). A
novel Arabidopsis-oomycete pathosystem: differential interactions with
Phytophthora capsici reveal a role for camalexin, indole glucosinolates and
salicylic acid in defence. Plant, Cell & Environment, 36, 1192-1203.
Wang, Y., Bouwmeester, K., van de Mortel, J.E., Shan, W. and Govers, F. (2013).
Induced expression of defence-related genes in Arabidopsis upon infection with
Phytophthora capsici. Plant Signaling & Behavior, 8, e24618.
Wang, Y., Meng, Y., Zhang, M., Tong, X., Wang, Q., Sun, Y., Quan, J., Govers, F. and
Shan, W. (2011). Infection of Arabidopsis thaliana by Phytophthora parasitica and
identification of variation in host specificity. Molecular Plant Pathology, 12, 187-
201.

213
Education Statement of the Graduate School
Experimental Plant Sciences
Issued to: Yan Wang Date: 24 November 2014 Group: Phytopathology University: Wageningen University & Research Centre
1) Start-up phase date
► First presentation of your project The role of lectin recptor kinases in resistance to Phytophthora Nov 10, 2010 ► Writing or rewriting a project proposal Huygens Scholarship Programme proposal: The role of lectin receptor kinases in disease resistance to Phytophthora pathogens' Dec 2010 ► Writing a review or book chapter ► MSc courses Plant-Microbe Interactions (PHP-30306) Mar-Apr, 2010 ► Laboratory use of isotopes
Subtotal Start-up Phase 5.5 credits*
2) Scientific Exposure date ► EPS PhD student days EPS PhD student day, Utrecht University Jun 01, 2010 2nd European Retreat for PhD Students in Plant Sciences, Cologne, Germany Apr 15-17, 2010 2nd PhD retreat in Northwest A&F University, Yangling Dec 04-05, 2010 4th Eurpean Retreat for PhD Students in Plant Sciences, Norwich, UK Aug 15-17, 2012
EPS PhD student day, University of Amsterdam Nov 30, 2012 ► EPS theme symposia
EPS theme 2 symposium 'Interactions between Plants and Biotic Agents, and Willie Commelin Scholten Day, Wageningen UR Feb 10, 2012
EPS theme 2 symposium 'Interactions between Plants and Biotic Agents, and Willie Commelin Scholten Day, Utrecht University Jan 24, 2013
EPS theme 2 symposium 'Interactions between Plants and Biotic Agents, and Willie Commelin Scholten Day, University of Amsterdam Feb 25, 2014
► NWO Lunteren days and other National Platforms NWO-ALW meeting 'Experimental Plant Sciences', Lunteren Apr 19-20, 2010 The Chinese Society for Plant Pathology, annual meeting Aug 19-21, 2011 NWO-ALW Platform Molecular Genetics Annual Meeting Oct 07, 2011 NWO-ALW meeting 'Experimental Plant Sciences', Lunteren Apr 03-04, 2012
NWO-ALW meeting 'Experimental Plant Sciences', Lunteren Apr 22-23, 2013
NWO-ALW meeting 'Experimental Plant Sciences', Lunteren Apr 14-15, 2014► Seminars (series), workshops and symposia CBSG2012 Summit 2010 Mar 16, 2010 Invited seminar Roeland Boer: 'Structural studies on proteins that prepare DNA for replication and conjugation' Apr 22, 2010
Invited seminar Matteo Brilli: 'Development of an integrated gene regulation-metabolism mathematical model of carbon utilization in E.coli’
Apr 29, 2010
Invited seminar Brigitte Mauch-Mani: 'Grapevine and downy mildew - Wine is not the only difference between grapevine and Arabidopsis'
May 31, 2010
Invited seminar Felix Mauch: 'Old fashioned secondary metabolites save Arabidopsis from Phytophthora brassicae' May 31, 2010
CBSG Seminar by H. Yang, S. Huang and R. Luo (from BGI and CAAS, China) Apr 01, 2010
CBSG workshop: Genome mining: taste the tomato genome Oct 27, 2011
Invited seminar David C. Baulcombe ''Plant versus virus: defense, counter defense, and counter counter defense'' Oct10, 2012
Invited seminar Ralph Panstruga: ‘Comparative pathogenomics of powdery mildew fungi: chasing the molecular secrets of obligate biotrophy and fungal pathogenesis’
Dec 04, 2012
Symposium: Intraspecific pathogen variation-implications and opportunities Jan 22, 2013
Invited seminar Detlef Weigel: “Arabidopsis thaliana as a model system for the study of evolutionary questions” Feb 27, 2013
Invited seminar Marc Boutry: ‘Plant drug smugglers’ about transport of secondary metabolites in plants Mar 13, 2013
Symposium: Food for all sustainable nutrition security, 95th Dies Natalis Mar 15, 2013
Invited seminar Howard S. Judelson 'Biology,pathology and evolution of Phytophthora infestans' May 07, 2013
Invited seminar Rays H.Y. Jiang 'integrative genomics of destructive pathogens from ommycete to malaria parasites' May 07, 2013
Invited seminar Brian Staskawicz:“Effector-Targeted Breeding for Durable Disease Control of Xanthomonas diseases in Tomato and Cassava”
May 21, 2013
Invited seminar Kuang Hanhui 'Using the Nicotiana-TMV system to study resistance gene evolution and plant genome stability' Sep 11, 2013
Invited seminar Jane Parker : 'Reprogramming cells for defence in plant innate immunity' Apr 09, 2014
Invited seminar Frank van Breusegem : 'Plant Metacaspases' Apr 09, 2014
► Seminar plus ► International symposia and congresses Annual Oomycete molecular genetics network meeting, Toulouse, France Jun 06-08, 2010
Annual Oomycete molecular genetics network meeting, Nanjing, China May 25-28, 2012
The 3rd International Conference on Biotic Plant Interactions, Yangling, Shaanxi, China Aug 18-21, 2013
10th International Congress of Plant Pathology, Beijing, China Aug 25-30, 2013
Annual Oomycete molecular genetics network meeting, Norwich, UK Jul 03-05, 2014
► Presentations Poster: Infection of Arabidopsis thaliana by Phytophthora parasitica Jun 06-08, 2010
Poster: Phytophthora capsici-Arabidopsis, a model pathosystem for studying the role of lectin receptor kinases in Phytophthora disease resistance
May 25-28, 2012
Poster: L-type lectin receptor kinases: novel Phytophthora resistance components Aug 18-21, 2013 Poster: L-type lectin receptor kinases: novel Phytophthora resistance components Apr 14-15, 2014 Oral: EPS Summer School Rhizosphere Signaling Aug 23-25, 2010 Oral: ALW meeting 'Experimental Plant Sciences', Lunteren Apr 03-05, 2012 Oral: CBSG Meeting Proteomics Hotel Projects Jan 18, 2013 Oral: EPS Theme 2 Symposium & WCS day Feb 25, 2014 ► IAB interview Meeting with a member of the International Advisory Board of the Graduate School EPS Nov 15, 2012
► Excursions Subtotal Scientific Exposure
23.9 credits*

214
3) In-Depth Studies date ► EPS courses or other PhD courses EPS PhD Summer School Rhizosphere Signaling Aug 23-25, 2010 EPS PhD Autumn School Host-Microbe Interactomics Nov 01-03, 2011 ► Journal club Literature discussion group at Pytopathology, Wageningen UR and at NWSUAF 2010-2014 ► Individual research training
Subtotal In-Depth Studies 4.8 credits*
4) Personal development date ► Skill training courses Information Literacy for PhD including EndNote Introduction Jun 15-16, 2010 Interactive workshop on presentation skills Nov 06, 2012 CBSG matchmaking event Oct 18, 2012 Techniques for Writing and Presenting a Scientific Paper Oct 16-19, 2013 Mobolizing your scientific network Mar 19 and 27, 2014 ► Organisation of PhD students day, course or conference ► Membership of Board, Committee or PhD council
Subtotal Personal Development 3.2 credits*
TOTAL NUMBER OF CREDIT POINTS* 37.4 Herewith the Graduate School declares that the PhD candidate has complied with the educational requirements set by the Educational Committee of EPS which comprises of a minimum total of 30 ECTS credits * A credit represents a normative study load of 28 hours of study.


This research was performed at the Laboratory of Phytopathology, Wageningen
University with financial support of a Wageningen University Sandwich-PhD fellowship,
a Huygens scholarship and the Food-for-Thought campaign of the Wageningen
University Fund.
Cover design and layout: Yan Wang Printed at Wöhrmann Print Service, Zutphen, The Netherlands.





























