Embed Size (px)
Citation preview

UNCLASSIFIED
'AD 296__097
ARMED SERVICES TECHNICAL INFORMATION AGENCYARLINGTON HALL STATIONARLINGTON 12, VIRGINIA
UNCLASSIFIED

NOTICE: When government or other drawings, speci-fications or other data are used for any purposeother than in connection with a definitely relatedgovernment procurement operation, the U. S.Government thereby incurs no responsibility, nor anyobligation whatsoever; and the fact that the Govern-ment may have formulated, furnished, or in any waysupplied the said drawings, specifications, or otherdata is not to be regarded by implication or other-wise as in any manner licensing the holder or anyother person or corporation, or conveying any rightsor permission to manufacture, use or sell anypatented invention that may in any way be relatedthereto.

,IN3TIcS AND ilOHANI3LS GI PD( AND I mI-
V 4 CATAIMSD AND I zD lTIONB
By D. T. IMORE, E. F. WRAG, and N. J. BAINE
room Queen's University, Belfast, N. Ireland.
Contract Number M2A-91 -591i-EUC-i 686
Final. Technicatl Report. F. No. i.
m I Pc.-'iod: 11st March, 1961 - 30th September, 1962.
I" Te research reported in this document has been
nada possible th-rough the support and sponsorship
if the U.S. Department of Iamy, through its European
Researc' Oif'ice.
1~E 13' 1963
L Ljl 7 ,. .
b
OL"AL Ir; -
,, . copies, .. _

- I
In Part 1, confirmetory evidence has been adduced or the mechanism of lacta-
alzation of methyl eaters of a--toluen6-r-sulphoryl derivatives of aw-diamino-acids
in aqueous solution, which waa described in the Pinal Technioal Report under Contract
DA-91-599-NC-1301. Participation of the sulphonamide-group has been excluded, since
the kinetics of lactasization of methyl y-amino-.--F-methyltoluene-k-sulphonmidobuty-
rate (M!aI), in which ionisation of the sulphonamido group is precluded, are exaotly
parallel to those of the compounds studied earlier.
In Part II, kinetic studies of the action of trypsin on various synthetic sub-
strates have been amplified. The significance of the experimentally observed values
of various kinetic constants is considered theoretically in the light of the probable
three-step mechanism for catalysis by proteolytic enzymes such as trypsin. In the
trypsin-catalysed hydrolysis of esters of a-F-toluene-2-sulphonyl--arginine, aoylation
and deacylation both appear to influence the overall rate of reaction, while in the
case of eaters of a-Lf-benzoyl-k-arginine, deacylation is rate-determining. In
ionizable group, whose apparent pKa and heat of ionization are consistent with those
of an imidazole nucleus, is implicated in the trypsin-catalysed hydrolysis of a-LI-
toluene-g-sulphonyl--arginine mothyl ester. Cholate ion accelerates the trypsin-
catalysed hydrolysis of esters of a-Lj-toluen-.L>-slphoryl-k.arginine and -homoarginine
slightly inhibits the hydrolysis of a-f-benzoyl-L-arginine ethyl ester (B=), Using
substrato concentrations high enough to ensure obedience to zero-order kinetics,
activation constants have been dctermined. Those for the trypsin-catalysed hydrolysis
of the methyl and ethyl esters of a-I-toluene-gI-ulphonyl-,-arginine (TANE and TAn)
converge to practically identical values with increasing cholate-conoentration. This
suggests that in presence of sufficient cholate the acylation step is sufficiently
accelerated so that deacylation of a common intermediate becomes rate-determining.
The same effect is possibly exerted by cholate on the hydrolysis of a-f-toluene--
aulphonyl-k-homoarginine mothyl ester, and supporting evidence is adduced from deter-
mination of apparent % values. a-I1-Toluene-2-sulphonyl-ig-ornithine methyl ester is
slowly hydrolysed by trypsin at pH 7.0 and the kinetics are first-crder with respect
to substrate. There is evidence that this reaction is not completely stereospecifio.
The hydrolysis is appreciably accelerated by cholate with an attendent decrease in the
apparent energy of activation for the reaction.
In Part III, we report a study of the kinetics of thrombin-catalysed reactions.
Using the Ehr(npreis and Scheraga unit (. Bil. Chem., 1957, U 1%.3) as a means of
expressing thrombin activity, it has been ahown that the velocity of a thrombin-
catalysed reaction is proportional to onzyme concentration. In contrast to

-2-
trypsin-oatelysed reactions, the velocity of hydrolysis of TAME by thrombin is decreased
by sodium or potassium chloride. Like trypsin-catalysed reactions, on the other hand,
the rate-determining step for the hydrolysis of esters of L-2-benzoyl--arginine by
thrombin appears to be deacylation. In the case of esters of as--toluene-j>-sulphorWl-
-einine, both acylation and deacylation influence the overall rate of hydrolysis.
The methyl ester of a -toluene-k-sulphocl-k-lysine (TIE) is hydrolysed by thrombin
at a rate similar to that of TAME. Rate oonotants for the thrombin-catalysed hydroly~d
of TAME and BIM do not obey the Arrhenius law. The apparent dissociation constant of
the ionizable group, which is involved in the thrombin-catalysed hydrolysis of TAME, is
rather lower than that associated with the trypsin-catajlysed hydrolysis of this substrete.
Cholate and glycocholate accelerate the thrombin-catalysed hydrolyses of substrates con-
taining the a-A-toluenew-sulphonamide group, but inhibit the hydrolysis of B . The
apparent % values for the TIME- and BAZ-thrombin systems are both decreased by bile
salts. The mechanism by which bile salts modify the kinetics of throubin-catalysed
reactions is probably similar to that postulated for trypsin-catalysed processes.
Non-ionic detergents such as Tween 20, Twceen 40 and Tween 80 accelerate the hydrolysis
of both TAME and BM by thrombin.

PART I. T KINETICS OF LACTAMIZATION OF METHYL ES7M CP SCME m-N-TVWUM-
p-SUWHONL UIVATVE OF ao-DIJDC=IO S IN AQUB SOLUTION.
In Part I of the Final Technical Report under the preoedinp contract
(No. DA-91-591-N JC-1301), it was shown that laotamization of the methyl esters of
G--toluene-2-sulphonyl-J-crnithine (TOE) and y-amino-c-toluene-L-sulphonamido-
butyric acid (AME) proceeded with hydroxyl-ion oatalysis in the pH range 7.8 -
8.5. We proposed the following mechanism for this reaction:
I H3 H
0 rT s.NRd.C Tos.NRI.. Tos.N.I. -C
"-C~e Me
MI (II) (inI)I-
To .NR. - W----.UIWC - 0(iv)
The initial rate of proton release at constant pH, U., was given by the equation:
+ ............. ()
where k is the second-order rate constant for the lactamization process K is the
dissociation constant of the protonated amino-group of the estcr (I), XW is the
ionic product of water, and [S] 0 is the initial concentration of substrate. The
value of K, as evaluated from the application of equation (i) to our experimental
data, was inconsistent with the participation of the sulphonamide group, and such
a mechanism appeared unlikely. In order to obtain definitive evidence on this
point, m have now synthesised methyl y-amino-L-a--methyltoluene-L-sulphonamido-
butyrate (AMTMM) in which ionization of the sulphonanmide group is precluded. This
compound was readily obtained by methylation of y-benzylo3carborlamido-k-a-toluen.-
]-oulphonamidobutyric acid in a manner similar to that described for the synthesis of
sarcosine (Cocker and Lapworth, g. Chem. §2., 1931, 189.; Cocker, . C 8.,
1 937, 1693). Hydrogenation and esterification completed the synthesis. The y-
benzyloxycarbonylamido-group was proved to be unaffected by methylation, since the
acid afforded by hydrogenation yielded nitrogen when treated with nitrous acid in
"-a Van Slyke apparatus. In addition, the ultraviolet spectrum of the J-methyl acid,
like that of fi-, oluen- .ulphs 'lsaeoosine, was identical in acid and alkaline
solution, whereas that of .-amino-k-a-toluene-2-suphonamidobutyrio acid exibited
a pronounced hyerchromic effect at high pH values resulting from ionization of the
aulphonamide grou. The kinetics of laotamization of AM were founC to be very
similar to those of the two substrates previously studied, and our earlier views
concerning the mechanism of lactamization are fully confirmed. Laotamization of
these esters appears not to be subject to general-base catalysis, since neither
pyridine nor imidazole affected the velocity of reaction. More kinetic runs have
been carried out on TOME and ATMU since the last Final Technical Report, and the
complete results are given in Table I and 2.
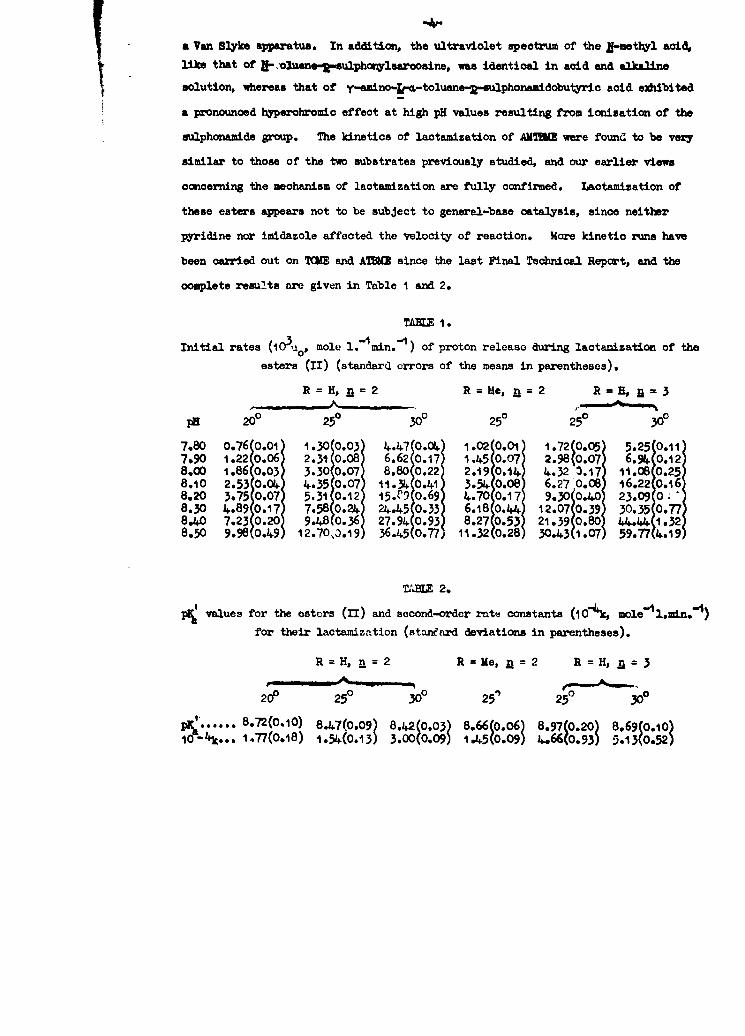
TAXL I.
Initial rates (I0 .Uo, mole l.1xmin.-') of proton release during lactamization of the
esters (II) (standard errors of the means in parentheses).
R = H, 2=2 R=Me, =2 R=H, =3
PH 20° 250 30o 250 250 300
7.80 0.76(0.01 1.30(0.03 4.4 0.04 1.02 0.01 1.72(0.05 5.25(0.117.90 1.220.06 2.31 0.08 6.62 0. 17 1 .45 0.07 2.98(0.07 6.9070.128.00 1.86 0.03 3.30 0.07 8.80 0.22 2.19 0,4 4.32 .17 11.08 0.258.10 2.53 - + 4.35 0.7 11.34 0.41 3.5 0.08 6.27 0.08 16.22 O.168.20 3.75 0.07 5.31 0.12 15.C o.69 4.700.1 7 9.300.40 23.09 08.30 4.89 A0.17 7.58O 02 2 5 0.33 6.18 0.4 12.07(0.39 30.35 0.778.A 7.23 0.20 9.i8 0.36 27.94 0.93 8.27 0.53 21.39(0.80 44.44 1.328.50 9.980.0491 12.70,30.19 36.450.77, 11.32(0.28 30.4311.07 59.77(4.19
TABE 2.
pl values for the esters (II) and second-order rate constants (10-4k, mole Lun.for their lactamiztion (stanarrd deviations in parentheses).
R =H, =2 R =Me, =2 R =H,n= 3
20P 250 300 25' 250 30
1 ...... 8.72(0.10) 8.47(0.09) 8.42(0.03) 8.66(0.06) 8.97(0.20) 8.69(0.10)1O- 4 k... 1.77(0.18) 1.5(0.13) 3.0(0.0 9) 1.450.09) 4-.66(0.93) 5.13(0.52)

-"
PARTI.m ' T1 KINnTIC. AJ3 (7 T' mPIN-&AI
In Parts II and III of the Final Technical Report under contract M-9i-51-C-
1301, we reviewed the available evidence related to the meohanism of aotion of
trypsin and we presented the results of our kinetic studies using various synthetic
substrates. We reported also that the trypsin-catalysed bydrolyses of the methyl
esters of a-L-toluen-sulphorwl-k-arginine and -- homoarginine (TAME and TaW
respectively) were accelerated in the presence of cholate ions. We do not propose
to review the literature again at length here, and we shall confine ourselves to
presenting the results of our continued studies of trypsin mentioning only those
publications which are pertinent to the development of our concept of the mechanism
of action of trypsin.
Shwert and Eisenberg (. Biol. chem., i91 9, jD, 665) found that, at substrate
concentrations high enough to ensure that zero-order kinetics were obeyed, several
esters of a--benzoyl-j-argininc were hydrolysed by trypsin at identical rates.
The inference to be drawn from these experiments is that a commrn intermediate,
(a-fi-benzoyl-L-argiryl) trypsin, is formed and that the deacylation of this is
rate-determining. The hydroityl group in the serine residue at the active centre
is probably the site of acylation. Strong evidence for such intermediates has
been obtained in the case of chymotrypsin-catalysed reactions (P. Desnuefle in
"The Enzymes", edited by P. D. Boyer, H. Lardy, and K. Myrblick, Academic Press,
N.Y. 1960, Vol. 4, p.93). The formation of cinnamoyl-chymotrypsin as an inter-
mediate in the hydrolysis of esters of cinnamic acid by cklnotrypsin has been
particularly firmly established (Bender & Zerner, j. -a 9=0 12200 196i, U
2391). The hydrolysis of J-.Van-cinnamoylimidazole is catalysed by either trypain
or chymotrypsin at pH 5.2, and the formation of an intermediate ciramoyl-derivative
of the enzyme has been demonstrated in each case (Bender, Kaiser and Zerner, 1. AM.
Chem. AM., 1961, 83, 4656). The kinetic mechanism of action of these enzymes can
thus be summarized in the expression:
RICEk +
R.C0.0 + Enye "~ acyl- -!'-- pm +hzaymesubstrate enzymecomplex (3) °(,.
Both the enzyme-auastrate complex and the acyl-ensymo are assumed to contain acidic
groups which must lose a proton for reaction to proceed. The p% values of these

-6-
aups are not necossarily identical. Indeed, if an imidazole group is implicated,
hydrogen-bonding between the hydroxyl group of the serine residue Fnd imidazole could
occur in the enzyme-substrate complex, but not in the acyl-enzyme. On the other hand,
hydrogen-bonding could occur between the imidazolium group and the carborWl-group in the
acyl-enryme. Consequently, the p% of the group in the enzyme-substrate complex would
be expected to be lower than that of the group in the acyl-enzyme. Such a difference
has been found in the chyotrypsin-catalysed hydrolysis of R-nitrophenyl acetate
(utfreund and Sturtevant, Prec. Nat. Acad. §c., 1956, h, 719). These authcs have
also derived the kinetic uquations which govern the three-stage mechanism outlined above.
When the steady state has been established, the velocity of reaction is given by:, I
V k2 Z [S] [3] (3)
where k2. .. .............. (4)2 K , + T H 4
and k k 3 K2 ......................3
Experimentally an apparent rate constant k;(nl ) end an apparent Michaelis constant
(sapp) are obtained, and these are related to the true rate constants as follows:-
-4. A .4. * ........... %......... (6)
(app) k k3 k + k2 (7)
k2"+k; 3 k2 +k3 k1
The determination of the effect of pH on kl(app) leads to an apparent dissociation oon-
stant, K(,p),wich is a function of the dissociation constants X and K2 and the rats
constants k and k.
K (app) W11Y2 k+ k 32(8
When deacylation of the acyl-enyme is rrte-determining, k2. t)k 3 anid, provided that 9
does not gretly exceed K
Conversely, if the acylation of enzyve is rate-determining, k2 ( , and, provided that
K, does not greatly exceed 12
,(app) V
In general, let k2 ook 3 andk =i R 12Y
Hence, K(app) = + + (n J)
If Z> 1, N) K(app)) 12 and conversely, if m( i, Y "(app) )

-7-
Thus, K(app) always lies between the two dissociation oonstants, but under special
oonditions may approximate to one of them.
We have studied the kinetios of the trypsin-catalysed hydrolysis of the methyl,
ethyl, and cyclohexyl esters of a-f-toluene-2-sulphonyl-k-arrinine (TAME, TAEM and
TAM respectively). By using substrate conoentrations high enough to ensure zero-
order kinetics, we were able to measure k3 (ap) over a range of temperatures and thus
to determine the activation constants. The results are presented in Tables 3 and 4.
Tabl 3
Rate constants at rH 8.4 for the tryesin-catalvsed hydrolysis of derivatives of arainim
andhmL1 D&.
Rate constants (k3(app)) are quoted with standard deviations; the number of deter-
minations is given in parcntheses.
t°C TAR TALI TACI t BLAM
-(sec. (sect (secZ;) (sec.)
15 49.22±2.21 (4) 63.4±-2.-9() 7.086-0.174(4) 2.286 -±0.098(7)
20 67.442.80(4) 85.44;t8.86(20 10, O20.66 (4) 3.042 !0.108(8)
23 85.72t.35 (.)
25 93.0 .73(7) 93.45±o.47(3) 108.6 :4..O (6) %."0.62 (4) 4.ai !0.212(8)
28 10.6 t5.5 (6)
30 124.7 !6.6 (10) 123.8 -*.7 (3) 146.7 8.7 (s) 21.1, .37 (6) 5.33- 0.231(8)
37 176.8 ±7.5 (6) 183.8 ±7.7 (8)
Tabl 4Activation constants at rH 8.. for the trvosin-catalvscd hydrolysis of derivtives
of ine and hoaogrinine.
Activation constints, with standard deviations, were calculated by the method of
weighted leat squares.
Ra(as./nole) a (cens./mole at 250) W (cas./mole/oC at 250) AGIG'Ccsawbe at
TM .0 -+ 10 883.0 10 -19.8 -0.4 14749± 2
TAZ310450 320 9860 320 -16.4 - 1.1 1.762± 2
8700 390 8110 390 -22.0 ±1.3 14679 11
BM12510- 350 11920 350 -13.2 ±1.2 15856 10
BM 9810 70 9220 70 -21.8 ±0.2 16621± 2

-It will be seen that there are small but definite differences between the values
obtained using TAME, TAZE, and TACO1. There are two possible explanations. The
hydrolysis of the acylated trypsin may not be rate-determining and k 3 (aP) mayI I
include contributions from both k2 amd k • Alternatively, the mechanism is more
complex than that outlined above. Bender and Glasson (I. AM. g=. IM., 1960, Z
3336) have obtained strong evidence for the presence of a site in ohymotrypsin wioh
binds water or alcohols, and it is possible that a similar site exists in trypsin.
The hydrolysis of the acylated enzyme may be an intramolecular process involving a
molecule of water bound at a site close to the acyl group. If the molecule of
alcohol, which is formed during the acylation of trypsin by the eater substrnte, oom-
pates with water for this site, the acyl-trypsin intermediates formed from the three
substrates can no longer be regarded as identical.
We pointed out in the previous Final Technical Report that the entropy of acti-
vation for the hydrolysis of MAL'E is more negative than for the hydrolysis of VJME,
suggesting a greater degree of structural rAganisation in the activatee complex.
This may reflect a deformation of the enzyme, an increased binding of solvent or a
more complex distribution of charges over one of the activated complexes involved in
the hydrolytic mechanism. Unfortunately, we have no information so far concerning
the nature of the rate-determining step.
"e kinetic constants for the trypain-catalysed hydrolysis of ea-benzoy1-L-
arginine ethyl ester (BAER) are close to those recorded for the related methyl ester
(DIME) (Schwert and Eisenberg, g. "o .g., 194~9, IM 665) after correction of
the value frr S* reported earlier (Final Technical Report, D -91-59it-E-301). This
supports the postulate that a comeon intermediate (W-Mbenzoyl-k-argirYl)trypsin is
formed and that deacylation of this by water is rate-determining. It also implies
that ethanol and methanol are not appreciably bound in the acylated intermediate.
Determination of k (app) at various 1 values and temperatures has been carried
out using the TAM-trypsin system. We were able to calculate the apparent disso-
ciation constants at the different temperatures (Table 5), K(ap)9 and the apparent
heat of ionisation.
Temperature ("C). PNl
TMJ~ !HME
15 7.770 0.o220 7.688 0 O.04425 7.529 O.OB 6.9N ! 0.037
30 7. - 0.038

The apparent heat of iciasation for the TAN-trypsin system was 7070 - 530 oals./
mole, a value consistent with that far an imidazole nucleus. It is notabie that the
pg. for the HAIM-trypsin is considerably lower than that for the TAME-trypsin system
at the same temperature. Since it is likely that K. the lower' K2 or p "PX2
apparent pKa for the THAM-trypsin system may be an indication that the ratio k 2 /k 3
is much lower for this system than for the TAMI-trypsin system. In other words, the
acylation step may be the main rate-limiting stage in the mechanism. Our results
differ somewhat from those of Ronwin 4oZhin. Bio i Ada 1959, N, 326), but the
conditions were rather different.
We next compared the effect of cholate on the kinetics of the trypsin-catalysed
hydrolyses of TAIM and TA at high substrate oonoentrations. Unfortunately a
similar investigation using TAMH was precluded, since addition of cholate to an
aqueous solution of this substrate gave a precipitate. The hydrolysis of both sub-
strates was accelerated by cholate and the results presented in Tables 6 and 7, show
that the rates of hydrolysis, k' p and activation constants for the two substrates
become much closer for the two substrates in presence of cholate. Thus k 2 and/orI
k 3mst be increased by cholate. It is, of course, possible that one 6f these rate
constants is decreased by cholate provided that the other is sufficient3ly increased
to give rise to an overall increase in kapp, and an increase in the ratio k 2 3
might explain the convergenoe of the activation constants for TAM and TAU in presence
of cholate. This phenomenon could be also explained if cholate helps to remove the
bound alcohol from the specific site on the acylated trypuin, and permit its replace-
ment by water. Such a process would increase the rate of hydrolysis of the acylated
trypsin (i.e. increase k 3 ) and would explain, at least in pert, the observed accele-
ration of the hydrolyses of TAME, TA1U, and IA by oholate. Cholate may intervene
in these reactions in other ways, however, and the relative importance of the various
effects may depend on the particular substrate. Thus it will be seen (Table 7) that
when TAIM or T are used as substrates, cholate tends to decrease o and to make
more negative. Conversely, in the hydrolysis of MAE, although this also is
accelerated by cholate, addition of cholate increases WA and makes M less negative.
If it is assumed for the moment that deacylation of the acylated enzyme is the rate-
determining step, cholate apparently affects the structure of the aotivated state as
wenl as possibly influencing the binding of water or alcohol in the ground state.
sach perturbations of the activated state might include (1) alteration of the helical
content of the polypeptide chain, (2) alteration of the degree of solvent binding
(3) alteration of the number and distribution of charges. Alternatively, if the
foregoing postulate that the TMJ-trypsin and THMh-trypsin systems differ with regard

_iito the k 2 3 ratio is correct, the opposite effects of oholate on the apparent acti-
vation constants may reflect the differing weights of k 2 and k 3 in the composite
rate oonstnnt k 3(,Pp)for the two systems. It is interesting to note that increasing
cholate oonentrrtion above 0.002 M has little further effect on the kinetics of the
hydrolysis of UU0, unlike the onse of TAM, Yhere cholite oontinues to modify the
kinetics up to a concentration of 0.006 M. The effects of cholate on the hydrolysis
of the above substrates by trypsin is not due to general-bae catalysis, since acetate
ion w.s inactive.
Taue6.
Rate constonts at pH 8.4 for the trypsin-catalysed hydrolysis of derivatives
of r dnine and homoarinine in presence of cholate.
Rate constants (k'3 (11)) are quoted with standard devictions;
the number of determinations is given in parentheses.
J& holate (0() TAM V=TAM
15 0 57.98t 2.08(101,9.29 2.21(4) 2.286t 0.098(7 7.0,6±.174(4.)
15 2 69.20t 3.71 (4) 71.27± 2.39(4) 2.45*3 0.105(8 7.072±0.068(3)
15 4 81.661 2.97 (4 89.17-+ 7.11(4) 2.589± 0.062(w 6.406±o.169(4)is 6 92,5 33t 44t] 976&26( .&S2,tf.8 0.07L.(& 6.127-;,.,291[(jj
20 0 79.09± 2.21 (81 67.41+ 2.80(4) 3.00± 0.108(8 10.48 ±0.66 (4
20 2 90.21t 2.89 (4 98. 11 4.69(4) 3.10 0.065(7 9.767-0.310(4)
20 4 j04.5 - 8.1 (4 17.4 - 5.9 (4) 3.69t 0.228(4 9.52810.369(4)20 6 12J.5 7A (4 1.4 .3( 3.672+ 0.107(41 9.06co.i26(4)
23 0 85.72± 1.35 0+23 2 114.7 ± 3.3 (23 1+ 1330± 3.6 ()+
25 0 93.-0 ±8.73'%*17: 93.45 ±0.47 (3 4.04-1 ±0.212(8 14.38 10.62 (4)25 2 110.8 4. (15 124.6 - 0.8 (4. 4.411 !0.194(8 14.16 !1.27 (4)
25 . 129.1 + 7.7 05 152.3 ± 9.8 (4 4.766 !0.067(4 13 38 -0.-7 (4)25 6 I 6.9 65 66., 1 j. LAX 10 , J.75 .L6 (L)28 0 110.6 5.5 (6'
28 2 141.41.5 (5
28 4 166.4 5.4 (6'28 6 _8_.8 t _ (6+ ,
30 0 124.7 - 6.6 (10 123.8 ± 1.7 (3 5.331 10.231(8 .ei49 ± .37 (6)30 2 159.2 9.1 (4 162.4 ± 4.1 (4' 6.083 ±0.229(8 20.27 ±+-.60 (4)
30 4 189.5 3.7 (411%.7 ±-1,5 (). 6.437 !0-065( 19.86 ±1.26 (40 6 209,0 -12.6 (207,0 ! L.2 . 6,42 ..37 + ,
37 a 176.8 ± 7.5 (6)37 2 221,7 3.2 (5.
37 4 250.4 5.C (ii.
37 6 279.4±5.7 (04_________________
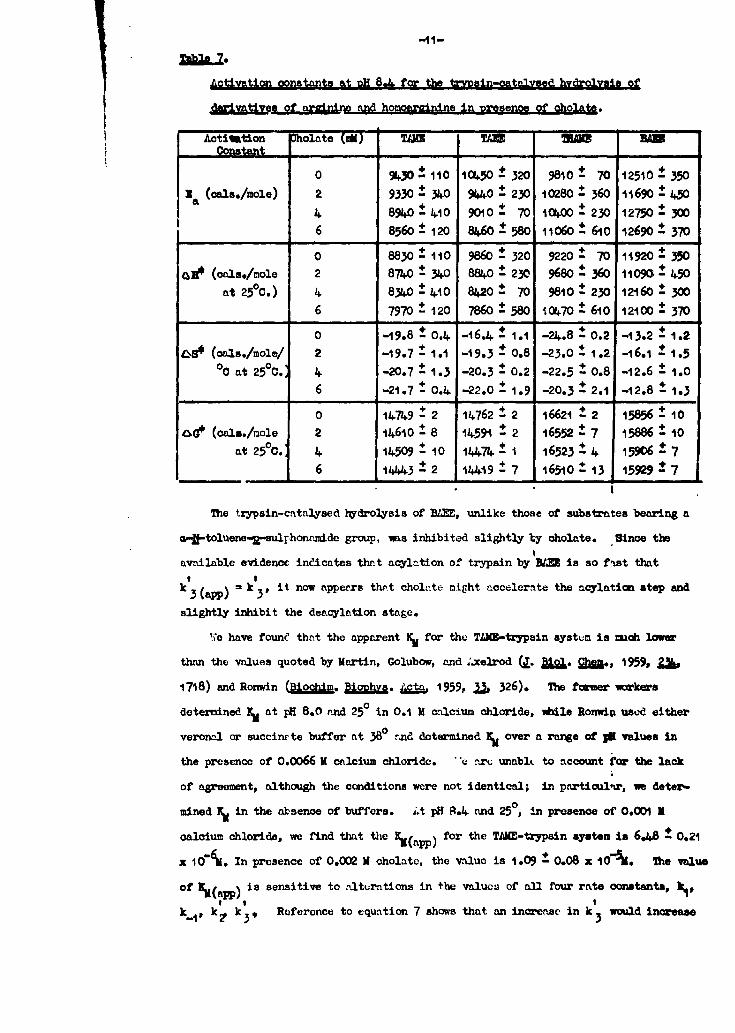
Activation oonstants at uH 8&L for the trypsin-oatalvsed hydrolysis of
derivativL of snirn and homoarainine in Dresenoe of oholate.
Aotivation Cholate () TAW Tku lu . BMConstant , ,.
0 9430-110 1,50-320 9810- 70 12510-350
3 (oais./mole) 2 9330 310 90 230 10280 360 11690 4504 8940 4.10 9010± 70 t0oo00 230 12750 ±306 8560 120 860 580 11060 610 12690 370
0 8830 110 9860 320 9220± 70 11920 350OR (oals./mole 2 8740 310 884.0 ±230 9680 360 11090 450
at 250C.) 4 8340±-i0 8420± 70 980 230 12160 3006 7970 ± 120 7860 580 10..70 610 12100 370
0 -i9.8 0.4 -16.4- 1.1 -24.8 ±0.2 -13.2 ±1.21-S (oals./mole/ 2 -19.7 ± 1.1 -19.3 ± 0.8 -23.0 ± 1.2 -16.o ± 1.5
°0 at 2500. 4 -20.7 ± 1.3 -20.3 ± 0.2 -22.5 0 0.8 -12.6 ± 1.0
6 -21.7 0.4 -22.0 1.9 -20.3± 2 -12.8 1.3
0 14749- 2 14762 2 16621 ±2 15856 10
oe (oals./aole 2 14610 ± 8 14591 - 2 16552 - 7 15886 ± 10at 250.: 4 I1509 -10 14474 - 16523 -4 15906- 7
6 14443 2 1419 7 16510 13 15929 7
The trypsin-catalysed t~drolysis of BAEE, unlike those of substrates bearing a
am*-toluene-2-sulhonjude group, m.s inhibited slightly by oholate. Since the
avnilable evideno. ineicates that acylation of trypsin by Rt- is so fPst thatI I
k 3 (app) = k 3 , it now appears that cholv-te might accelerate the acylation step and
slightly inhibit the deacylation stage.
We have foune thnt the apparent K. for the TIA-tzypuin system is ach lower
than the values quoted by Martin, Golubow, and axelrod (A. Al. n., 1959, 116
1718) and Ronwin (Bioch1m . y. igct 1959, j 326). The tcrer workers
determined % at ;H 8.0 end 250 in 0.1 X cnlcium chloride, 1hile Ronwip used either
veronal or succinrte buffer at 380 -nd determined % over a range of V1 values in
the presence of 0.0066 U calcium chloride. -c -rc unable to account Cor the lack
of agreement, although the conditions were not identical; in particular, we deter,-
mined % in the absence of buffers. Lt pH R.4 and 250, in prosence of 0.00 N
oalcium chloride, we find that the N (app) for the TAE-tzypsin system is 6.48 - 0.21
x id"%. In presence of 0.002 M cholate, the value is t.09 ± 0.08 x i0 5 . 7he value
of %(p)is sensitive to alterations in the valucs of all four rate constants, kipI k ik l, k2 , k,, Reference to equation 7 shows that an increase in k 3 would inorease

I1 -12 -
Twp) whl nices nk rl! ouddces h feto le
rations to k 2 depend on the relative mgnitude of k_1 and k 3" If k_, Ik y anincrase inwhilandincrease decreaseTheetioal, if k ( , the converse
would be true, but this woule" bo most unusual for an enzyme-catalysed reaction. It
in not possible to determine fron the present data which rate constants are affected
by oholate in the T1.M-trypain system.
Values of %(,Pp) for the TWJM-trypsin system under various conditions are
recorded in Table 8.
Tale8.
K( ) values for the TUH-trypsin system at 259.
(Values of K?(app) a quoted with stn&'C1C devir.tions; the
number of complete deturminations involving 6 - 8 runs is
given in parentheses).
PH Cholate oonc'n.(cU) i& K(ip)(moles/litre)
7.0 0 2.391 0.070 (3)
8.4 0 3.320 0.017 ()
8.4 2 2.51+3 ± 0.288 (1)
84 6 2.243 - 0.058 (2)
It will be seen that %(app) increases with pH over the range 7.0 - 84. Since
K, the dissociation constant of the group in the Michaelis complex is hiher than K0I I
it in likely that k 3 incrcnass considerably over this range, while k 2 is much le
sensitie. The incre.se in ) with increasing pH is probably attributable,
therefore, to the increase in k * The decrease in K,(,p) in presence of cholate,I I
on the other hand, is probably due to an increase in k2 The value of k3 probably
does not alter appreciably, as suggestee by the foregoing work, but a small chonge in
either direction is possible.
We were unable to study the trypsin-c'tri7ysed hydrolysis of Gj-toluene-2-
sulphonl-L-ornithine methyl ester (TOM) at ,H 8.4, in view of the latAnisation of
the substate, which is c:..t]ysed by hydroxyl ions (P-rt I). Trypain ws found to
cattlyse the hydrolysis of TE .t AH 7.0, however, with little oncomit-nt lact.-
mization. The %(,Pp) -ast be very high, since th,m was no dvyprture frcm first-
order kinetics with respect to substr te even when the substr-.te conoentrntion was
's "'gh is 5 x id . Pseudo-first-oreer r.t constatnts nre recorded in T.ble 9,

P..aaoa-ffrt-,orde- rate constant for the 1lstrvDin system at xE 7.0.
(lat constants are quoted with standard dbviati Ls; the number of
determinations is given in parentheses).
t 0 k (seo.-'/mole trypsin/i'tre).
No dcholate 2 di cholate
15 66.69 1 1.71 (,) 89.o6 - 2.18 (.)
20 97.83 3.06 (4) 123.08 - 2.57 (6)
25 133.02 ± 348 (4) 169.58 ± 2.78 (4)
30 193.39 ± 2.61 (6) 241.06 !11.90 (6)
In the presence of 0.002 M cholp-te, the apparent energy of aotivation is decrersed
from 12280 ± 290 oals./mole. to 11180 ± 260 cals./mole. It is not possible to decide
from the present &ta which step of the ensymic mechanism is affected by cholate. The
hydrolysis of N by trypsin does not appear to be completely stereospecifio. Using
the 5, ester, we found that the rr.te of alkali uptake in presence of trypsin was
peater than when the &-ester of half the molar concentration was used as substrate.
Moreover, when the B-ester w'.s used as substrate, alkali uptake continued beyond the
value representing 5CW hydrolysis. This was shomn not to be due to laotamization,
since a sample wns hydrolysed with typsin until 7).8% of the theoretical amount of
alknli required for complete hydrolysis had been used. 4t that point a S~remsen
titration indicated thAt only 8.6% of the total amount of ester had lctamised.
Finally, although good first-orCer plots coule be obtained from the hydrolysis of the
L-estwr in presence of trypsin, similr analysis of kinetic runs with the &[-ester
gave ourves, indicating that at least two reactions with different rate constants
were proceeding simultaneously. It is possible thrt the loss of complete stereo-
specificity is due to the shortness of the sido-chain of ornithine compared with
arginine rnd lysine. Attachment of TO to a specific site on the eneyme moleoule
by the protonated amino group my be weakened. Further shortening of the side-oh-An
might preclude 1jW attachment to the specific site and this presumably explains the
failure of trypsin to catalyse the hydrolysis of A IE.

PAR m. sTMMS CP M r x u
Thrombin in a proteolytic enzyme which is responsible for Ahe oonversion of
fibrinogen into fibrin during blood-clotting; it also displays esaterase activity
tow rds substrates like TAS. Inhibition of thrombin by di-isprofl phosphovo-
fluoridte, followed by partial hydrolysis, has revealed that this ensyme, le
bypsin, chymotrypsin, elastase, and cholinesterase, has the amino-acid sequence
- gly.asp.ser.V - at the active centre. Unfortunately, t1nombin has not been
purified as completely as trypsin or ckmotrypsin, and it is necessary to define a
suitable unit of activity. The literature abounds with different units for expres-
sing thrombin activity, and we have selected that due to Shrempreis and 8cheraga
(&. .20., 1957, 22, I ) as the most suitable for our purpose. One
thrombin unit is defined as that amount of enzyme required to liberate 0.1 poole of
acid per manute from I al. of 0.1M TAU at pH 8 in 0.15 M I at 2!P. We were
able to confirm the observation of 1hrenpreis and Scheraga that thrombin solutions
at pH 7 and 0eC retain their esterase activity for at least nine days. On the
other hand Ronwin (CanaL J. Bio ham. Physiol., 1957, Zo 743) reports that thrombin,
dissolved in glyoerol-mater (i : i) or isotonic saline, becomes more active as an
eaterase during storage at 0-4o. Using BAEZ (5 x 10)M) as substrate, we found
that zero-order kinetics were obeyed and the velocity was proportional to the
enzyme concentration.
The throbin-oatalysed hydrolysis of TAU is markedly dependent en ionic
strength. Ronwin (Canad. J. Biochem. Ptwasol., 1957, 3, 743) has reported that
increase of ionic strength inhibited this reaction, but since he used buffers as
well, the results are difficult to interpret. Our results (Table 10) were obtained
in the gbeence of buffers, and it can be clearly seen that k3 (app) decreases
asymptotically with increasing strength to a limiting value which depends on the
cation. Our results do not afford a linear relationship between log k and
where p is the ionic strength, as might have been expected. Scheraga and
2brenpreis . tM. 2=. Bichmir, Venna ergamon Press,
Vol. X, p.212) have briefly reported that sodium chloride inhibits the hydrolysis
of TiME by thrombin as the ionic strength is increased. Their results qualita-
tively resemble ours and afford no linear relationship between log k and , .
A possible explanation for the complex connexion between log k and p may be the
presence of more than one esterase activity in the thrombin preparations.

Tbe0.
Effect of ionic strength on k' for the thrombin-oata~lsed
! hvg Iya ofTAKE at yH 8.1& and 2500.
k 3 (aPP) values are quoted with standard deviations; the number
of -',.-trminations is given in parentheses.
Cne'.. of salt (M) 10A31 (app)(mo3eslitelmn.lsheraga uit/i~tre).
0.00 18.071 - 0.076 (5) 18.071 ± 0.076 (5)
0.05 9.082 0.663 (3) i3.1i. + 0.381 (3)
0.10 6.7,54 0.1*31 (4) 12.361 ± 0.238 (2)
0.20 5.114 ± 0.808 (2) 10.802 ± 0.751 (2)
0.30 4.289 4. 0.10 (2) 8.832 ± 0.125 (2)
0.40 3.880 - 0.287 (2) 8.809 ±. 0166 (2)
Surprisingly, the rate of hyerolysis of BAM by thrombin was insensitive to
the ionic strength when sodium or potassium chloride was added to the system. A
possible explanation of the inhibition of hydrolysis of TAME by thrombin with in-
creasing ionic strength is the existence of a mechanism which involves the approach
of ions of unlike charges. Since TAME has a positive charge on the guanidine group,
it sight be argued that a negative charge exists at the active centre of thrombia.
Baoh a charge does exist on the A- 002- group of the aspartyl residue, adjacent to
the serine residue which is phosphorylated by di-isopropyl phosphorofluoridate.
This explanation is weakened, however, by the insensitivity of the BA3-thrombin
and TAME-trypsin systems to change of ionic strength. On the other hand, it is
possible, as outlined below, that the acylation step controlled by ks contributes
to ki (app) in the TAME-thrombin system, but not in the BABB-system. In this
event, the acylation step only could be sensitive to change of ionic strength, in
which case, it could be an indication that a negatively charged group is the
nuoleophile which attacks the oarboiyl-carbon atom in the substrate. Further
evidence must be obtained before this point can be established. It is puzzling
that the TAM-trypsin system is insensitive to change of ionic strength when other
evidence indicates that the acylation step is partially responsible for the observed
value of k' It is of interest to recall that the velocities of certain3 (app)-hydrolyses catalysed by chymotrypsin and ficin are increased as the ionic strength is
inreased (Shine and Niemann, J. Amer. Chem. Soc., 1955, Z 4275,; Martin and
Niemarz, J. Imer. Chem. Soo., 1958, _ 1481; Hamond and Gutfreund, lodhe. J.,
059, 2L X9)-

-16.-
We have determined k (app) for the hydrolysis of a range of esters in presence
of thrombin at pH 8.4 and 25 0 C. These include TAME, TAN!, TAM, RA, DAME, --
toluene-2-sulphorWlj--arginine -propyl esttcr (TAPE), and -J-touene-2sulphory:L..-
lysine methyl ester (TMUE). The results are presented in Table 11.
Table 1i.
k/ ) for the hydrolysis of various substrates by thrombin at pH 8.4 and.
2500 in 0.1 M sodium chloride solution.
Valuestctkl 'app) are quoted with standard deviations; the number of deter-
minations is given in parentheses.
Substate iO5k'Substrte I A app)(moleslitre/mn./scheraaa unit/litre).
TAME 6.754 0.431 (4)
TAU 2.988 - 0.051(10)
TM 2.611 ± o.o56 (6)
T6.538 0.088 (6)
U 10.430 o.6o (6)
BAE 3.233 ± 0.071 (5)
BARE3.246 ±0.013(05)
it will be noticed that T.AQE is hydrolysed considerably morm rapidly than TAM Or
TAPE, and it is intereting tbat this order is the opposite of that expected in the
ncn-ermymic alkaline hydrolysis of esters. Ls wrs found in the case of trypaim,
esters ontaining a a-1-toluene-R-sulphonamide group do not hydrolyse at the saem
rate. Henoe, we may conclude that for thesc substrrtes, deacylation of an aq:L-
thrombin is not uniquely rnte-determining; the acylation step governed by 14 makes
a significant contribution to k' ., On the other hand BAME and BM are3 'app)bydrolysed by thrombin at indistinguishable rates, and it may be concluded that
deaoylation is rate-determininp, i~e k I Ic, I'* The foregoing observatiora3 ap) 3are thus similar to but rather more striking than those found using tr'ypsin. On
further point may be noted; T and TAM are hydrolysed at about the same rate by
thrombin. The distanct between the charged group in the side-chain and the ester-
carboryl group is almost identical for the two substrates ihen the .oleaules eve in
their energetically most favoured conformations. '5e have indicated in the studies
on trypain that the length of the side-chain of the substrate is important in cleter-
mining specificity, and the same seems to be true in the case of thrcmino In
preliminary experiments, m have found that TOM is slowly hydrolysed by throdbis

-17-
and the apparent % is high, as in the oae of trypsin. TRAM, on the other hand,
is not appreoably bydro3.ysed by thrombin, and in this respect contrasts sharply with
trpsil.
In the previous Final Teohnioal Report we recorded the results of a limited
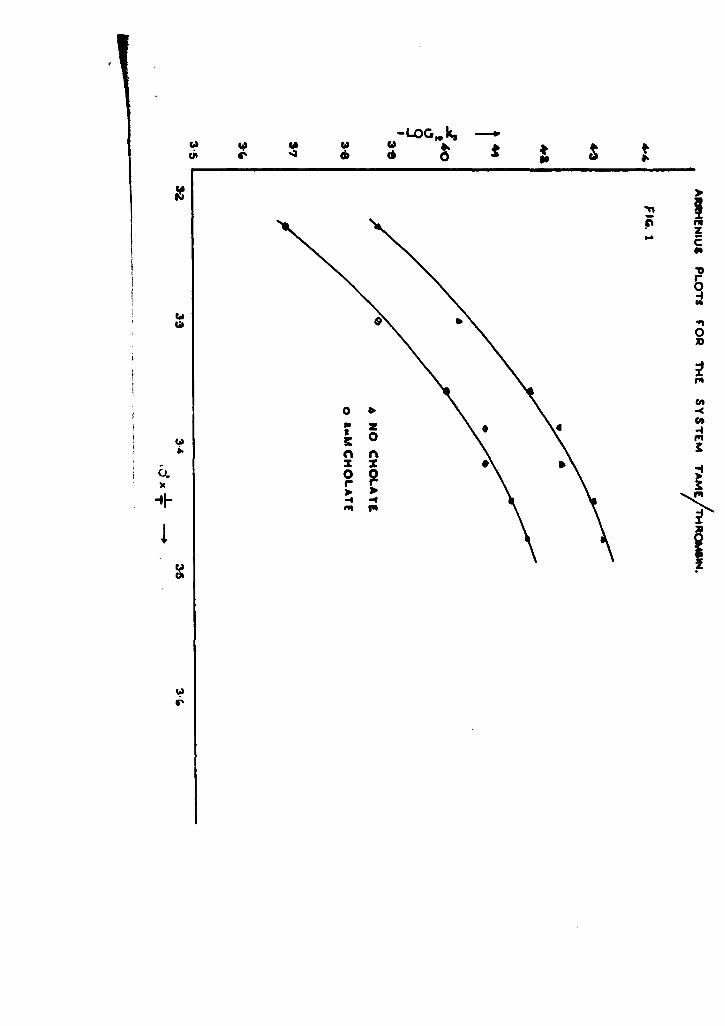
stu43 of the effect of temperature variation on k' (aPP) for the TAM-thrombin my&-
tern. The results available at that time indicated that an Arrhenius type of plot
was linear. More extensive measurements using both TAME and BABE as substrates
have since shown that plots of log k', (app) against I/T have a marked curvature
(Tables 12 and 13, Figs. I and 2). We are thus unable to calculate activation con-
el -1 for tbrombin-eatalysed reactions. It is likely that this abnormal behaviot
results from the presence of more than one type of esterase activity in the thrombin
preparation. This is substantiated by the very recent iselation by Maroiniak and
Seegers (91 . & Bio .-zwol., 1962, J& 597) of two fractions possessing
esterase activity from highly purified thrombin. Ronwin (goa. g. Roah. Physiol.,
1957, 2, 743) found that zero-order kinetics in the TAME-thrombin system were not
obeyed at 380 when small concentrations of enzyme were employed, but normal behaviour
was found in experiments when relatively large amounts of thrombin were used.
Neither we nor Sherry and Troll (J. Biol. Chem., 1954, 2, 95) have observed this
pheInon.
Effeot of tewerature n kj (app) for the TafL-t bin avstem at
m1 8Ja. alone and in the uresenoe of cholate.
Values of k3 (app) are quoted with standard deviations; the rum-
ber of determinations is given in parentheses.
No cholate P Idi cholate
15 4.826 097 '3 6.786 1 0.252 317.5 5.058 -0.8 4 7.289 0.095 420 5.869 ± 0.031 '4 8.231 0.35322.5 5.965 - 0.124 4 8.207 ; 0.15725 6.754 - 0.431 4 9.875 ; 0.337 '530 9.486 0.517 41 13.533 ; 0.(00 437 13.499 - 0.015 4 20.512 0.056 1
-LOG~, -
x z
00
300
0 4 4

-LOG,,ks .
* CA
100
C3.-r"

Effetof teme on k _ (app) for the EAMI-thrombin asvtem
at r1 8.Ji. alone and in the ioresence of oholate.
Values of k Iap are quoted with standard deviations; the
number of determinations is given in parentheses.
tera 1o 5km (app) (moleqitre/min. cherage unit/litre)
Vj No cholate 2m( cholate &aM cholate
1o 1.283 0 0.023 (4) 1-043 ±- o.04 (4)
15 1.627 0.037 (3) 1.197 0.051 () 1.116 0.055 (3) 1.047 ±0.053 (3)
17.5 1.695 ± o.017 (4) 1.279 0 o.07 ()
20 2.10 o.061 (4) 1.703± .011 ()
22.5 2.75+ 0.013 (4) 1.773 -.065 (3)
25 3.24.6 ± o.M3(15) 2.370 o.o88 (3) 1.798 ± 0.059 (3) 1.603 ± 0.069 (3)27.5 3.-7 - 0.025 (4) 2.388 0.096 (3) 1.826 ± 0.067 (3) 1.690 ± 0.1O (3)
30 .212 ± 0.051 (3) 2.926 ± 0.091 (5) 2.457 ± 0.093 (5) 2.217 - 0.065 (5)
37 8,8937 ±0.021 0) 5.236 ± 0.011 (3) 1.1 05 o.o2o () 1. 519 o.016 (4)
Ronwin (M.him " j os. . A21 1959, j 326) has studied the effect of pH-
variation on k for the TAME-thrombin system and has reported that there are
two dissociating groups in the ES complex, and three in the free enzyme. In con-
treat, e detected only one dissociable group, P%app) = 6.46 t 0.06, in the TAME-
trypsin system by measuring k(ap) at various pH values at high substrate con-
centration. This could bu an imidasole group, but it is likely that the observed
pK, lies between those of the dissociable group an the HS complex and that on the
acylated enzyme, since it appears from the results described above tlmt mylation
is partly rate-determining.
The thrombin-catalysed hydrolyses of substrates such as TAME, TmL, 7194 and
MME, Which contain a L--toluene-2-sulphonamide group are accelerated by cholate
and even more by glycocholate. The thrombin-catalysec hydrolysis of BARM, on the
other hand, is markedly inhibited by both cholate and glyoocholate. The inhibition
of this system is more pronounced than in the case of the tt- in-oatalyaed
hydrolysis of BAR. The kinetic modification by bile salts of both trypsin- and
thrombin-catalysed reactions appears to operate by a similar mechanism; acylaticn
is accelerated while deacylation is retarded. The results are recorded in Tables
12, 13, and 14. There is a quantitative difference between the effects of cholate
and glycocholate. The former, from its effect on the byholyais of BAIE, appears
to be the more powerful inhibitor of the deacylation step. Glyoocholate, on the
other hand, is the more effectiv nccelerator of the hydrolysis of TAME. This ic
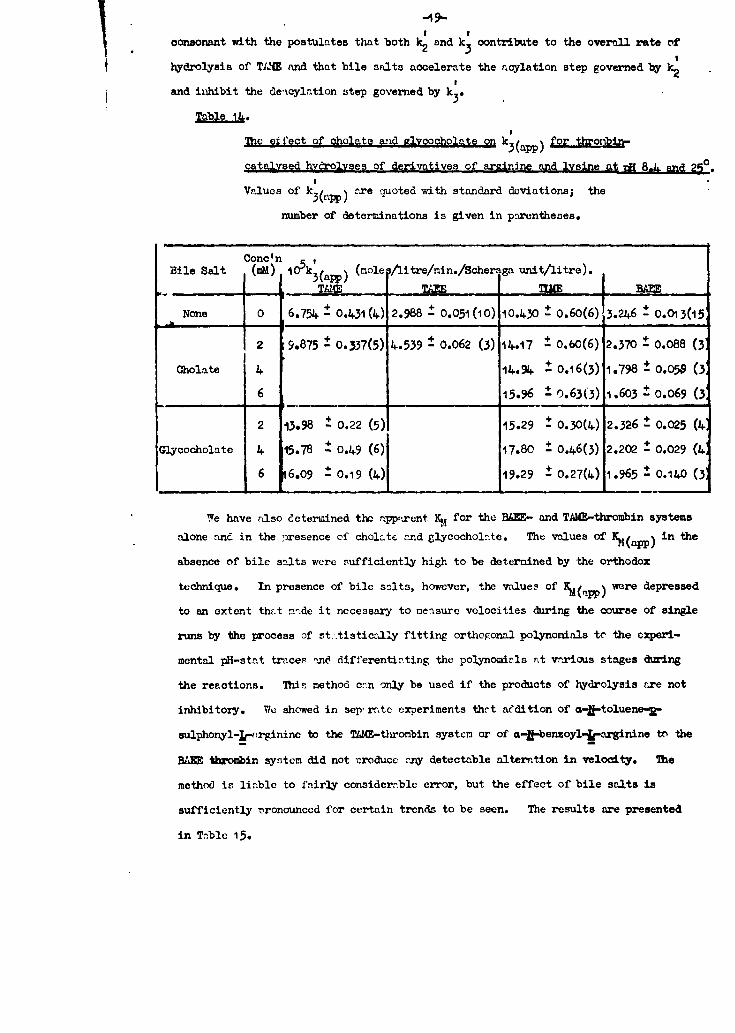
-49-consonant with the postulates that both k2 and k 3 contribute to the overall rate or
hydrolysis of TIJ and that bile salts aoelerate the acylation step governed by%
osand ihibit the de-tcylation step gover n e d by k 3 t
The effect of oholate and a1vcoocholate on k3(app) for throni
catalvsed hycrolyses of derivatives of arinine and lvsine at M 8.1 an, 250.
Values of k36rPP) ae quoted with standard deviations; the
number of determinations is given in parentheses.
Conc'nBile Salt (MM) 10 5 k 3 (,p) (mole /litre/min./Scherga unit/litre).
_______TAC ________ _______ BAM
None 0 6.754 - 0.431(4) 2.988 + 0.051(10) 10.430 0 0.60(6)13.246 - 0.013(15
2 9.875 4 0.337(5) 4.539 ± 0.062 (3) 14.17 0.0(6) 2.370 - 0.088 (3,
Cholate 4 14.94 ± 0.16(3) 1.798 4 0.058 (3:
6 15.96 0 o.63(3) 1.603 ± 0.069 (0
2 13.98 ± 0.22 (5) 15.29 0 0.30(4) 2.326 0 0.025 (4'
Glycocholate 4 15.78 ± 0.+9 (6) 17.80 0.46(3) 2.202 - 0.029 (4',
6 6.09 .0.19 (4) 19.29 ± 0.27(4) 1.965 , 0.140 (3
We have also determined the npptrent K4 for the BAZE- and TAME-throubin systems
alone and in the presence of cholate and glycochol: te. The values of N(app) in the
absence of bile salts were sufficiently high to be determined by the orthodox
technique. In presence of bile salts, however, the values of K(,p ) were depressed
to an extent that -nde it necessary to measure velocities during the course of single
runs by the process of stctistically fitting orthoFonal polynomials to the experi-
mental pH-stat traces -nd differentiating the polynomials at various stages during
the reactions. This method c-n only be used if the products of hydrolysis are not
inhibitory. We showed in sep rate experiments thrt adition of a-n-toluene- R -
sulphonyl-L-erginine to the T1M-thrombin systcm or of a-l-benzoyl-k-orginine to the
BAE thrombin system did not oroduce any detectable alteration in velocity. The
method is liable to fairly considernble error, but the effect of bile salts is
sufficiently Dronounced for certain trends to be seen. The results are presented
in Table 15.

-20-
Tale 15.
AA (app) in the -and !h-throtio_ systems at DH 8.. -d 250
Mean values of %(app) are quoted with standard deviations; the
number of determinations is given in parentheses.
Bile Salt i o0% (app) (oles/itre)
None 1,61 - 0.39 (5) 3.21 ± 0.78 (7)
Cholate (2rr1) 0.6o ± o.1o (6) 0.50 ± 0.23 (6)
Glycholate (2ii) 0.74 - 0.14 (5) 0.54 - 0.26 (5)
I S
The effect on %(ap) of alterations to k2 nd k; is analysed in Part 17. An! aj2
increase in 2 or a decrease in ky both of which appear to result from addition of
bile salts, lead to a decrease in (ap). The observation that the % (app) for the
TANE-thrombin system is more sensitive than that for the BAZ-thrombin system in
presence of cholate is consistent with the view that k 2 )> k 3 in the latter case.
A few experiments have been carried out to determine the effect of the non-ionio
detergents Tween 20, Tween 40, and Tween 80 on thrombin-catalysed reactions. The
hydrolysis of BAEE is slightly accelerated, hile that of TAME is considerably
accelerated. In the light of the foregoing discussion, it appears that the non-
ionic detergents slightly increase k3 and effect a considerable increase in k 2. The
results are given in Table 16.
Table -16,ffect of non-ionic detergents on k3(app) for the th'ombin-catalysed
bwdrolvsis of TM. and AM at nH 8.4 and 250.
Values of k;(aPP) are quoted w ith standard deviations; the rAniber
of determinations is given in parentheses.
Detergent ConW,,t4ron ik3 aP (mole s/litre/isin./cheraga uit/*itre).
TAM BAE
None 0 6.754 ± 0.431 (4) 3.26 0 0.013 (15)
Tween 20 0.3 9.422 0.140 (3) 3.667 - 0.096 (4)
1.2 11.026 0.486 (3)
Tween 40 0.3 9.219 ± 0.331 (4)
1.2 9.59 0.097 (4) 3.- 0.062 (3)
Tween 80 0.3 11.603 - 0.301 (5) 3.920 ± 0.116 (4)

-21-
Prom the foregoing work, it is evident that trypsin and thrombin closely resemble
one another. Thrombin, however, does appear to be somersat more specific (see
Scheraga and Laskowski, AAX. Protein Chem., 1957, Z i), since it has virtually no
action on TRW. The conversion of fibrinogen into fibrin is a further example of
the greater specificity of thrombin compared with trypsin. Although fibrin is not a
substrate for thrombin, except perhaps in presence of high enzyme concentrations
(Quest and Ware, Science 1950, 112, 21), trypsin is capable of effecting considerable
degradation involving the cleavage of peptide bonds. Both thrombin and trypsin appear
to octalyse the hydrolysis of synthetic substrates by a three-step mechanism, and, for
a given substrate, the relative importance of each step in determining the overall
kinetics is very similar. The behaviour of kinetic modifiers such as cholate appears
to be qualitatively similar with both enzymes; the step governed by 4 is accelerated
and that controlled by k' is slightly retarded. If this theory can be confirmed by.3
further experiments using other substrntes, it is possible that such kinetic modifiers
will be useful in elucidating the mechanism of enzyme-catalysed processem. It is not
possible yet to assess with certainty the importance of anionic surface-active Wients
such as bile salts or ohospholipids on biological processes such as protein digestion
and blood-clotting, which depend on trypsin nnd thrombin. The kinetic mechanism of
those reactions described in this report appears to be more complex than at first
seemed to be the case. "se have not had time to study the effect of cholate and glyoo-
cholate on more complex reactions such as the conversion of fibrinogen into fibrin,
have we been able to study the kinetics of renctions catalysed by pla.a. Clearly, it
would be of considerable interest to do this. We would have liked to have studied
the effect of bile salts on the pre-steady state using substrates such as p-nitrphenyl
acetate, but delays in obtaining commercial equipment precluded this. It is hoped,
however, that the authors will be in a position to tackle some of these problems in
the near future.
P'RSOHW TnI 'r AN E
Mr. N. J. Baines worked on this contract from its commencement until the end of
the second quarter. Mr. E. F. Curragh has worked on this contract throughout the
whole period. Both men have devoted their complete attention to the problem during
the time that they have been paid under the contract.
The cost of chomicals and of preparing reports has exceeded the original estimate
of Z79.





























