Embed Size (px)
Citation preview

Ultrastructure of Posterior Sternal Glands ofMacrotermes annandalei (Silvestri): New Members ofthe Sexual Glandular Set Found in Termites (Insecta)Andre Quennedey,* Alexis Peppuy, Annie Courrent, Alain Robert, Claude Everaerts,and Christian Bordereau
UMR CNRS 5548 “Developpement, Communication Chimique,” Universite de Bourgogne, 21000 Dijon, France
ABSTRACT In female alates of Macrotermes annanda-lei, two types of abdominal glands are involved in thesecretion of sex pheromone. Tergal glands are found at theanterior margin of tergites 6–10 and posterior sternalglands (PSGs) are located at the anterior margin of ster-nites 6–7. The cytological features of both types of glandsare quite similar. The fine structural organization of PSGsis studied more precisely and described for the first time.The glandular cuticle is pitted with narrow aperturescorresponding to the openings of numerous subcuticularpouches. Several Class 3 glandular units open in eachpouch. One canal cell and one secretory cell make anindividual glandular unit. The canal cell is enlarged api-cally and is connected with the other canal cells to form acommon pouch. Based on the structural features found inthese glands, we propose a common secretory process forPSGs and tergal glands. During the physiological matu-ration of alates inside the nest, secretory vesicles amass inthe cytoplasm of secretory cells, while large intercellularspaces collapse the cuticular pouches. At the time of dis-persal flight, pouches are filled with the content of secre-tory vesicles while intercellular spaces are sharply re-duced. After calling behavior, no secretion remains in theglands and pouches collapse again, while secretory cellsare drastically reduced in size. The structure and thesecretory processes of PSGs and tergal glands are com-pared to those of abdominal sexual glands known in ter-mites. J. Morphol. 262:683–691, 2004.© 2004 Wiley-Liss, Inc.
KEY WORDS: Isoptera; Termitidae; Macrotermitinae;tergal glands; sex pheromone; secretory process
In termites, the reproduction of colonies is gener-ally mediated through seasonal dispersal flights ofalates. To date, three types of abdominal glandshave been found to be involved in the sexual behav-ior eliciting pairing. The sternal gland secretes a sexattractant pheromone in a few known cases(Pasteels and Bordereau, 1998). Its fine structurehas been described by Quennedey (1977, 1978),Quennedey and Leuthold (1977), and Laduguie(1993). The tergal glands may play different roles.According to Pasteels (1972), tergal secretions allowthe cohesion of the nuptial tandem before mating.They are also responsible for long-range attractionin some species such as Syntermes dirus (Barth,
1955), Trinervitermes bettonianus (Leuthold, 1977),and Cornitermes bequaerti (Bordereau et al., 2002).The fine structure of tergal glands has been de-scribed in some species by Wall (1971), Ampion(1980), and Bordereau et al. (2002). The third type ofabdominal glands, the posterior sternal glands(PSGs), has been reported on sternites 8–9 in malealates of Stolotermes, Porotermes (Termopsidae),and Prorhinotermes (Rhinotermitidae) (Ampion,1980; Ampion and Quennedey, 1981). Recently, Pep-puy (1999) and Peppuy et al. (1999, 2003) haveshown that PSGs located on abdominal sternites6–7 are present in female alates of several species ofAsiatic Macrotermes and are involved in courtship.In this article, we study the ultrastructure and thesecretory process of PSGs in M. annandalei.
MATERIALS AND METHODS
Macrotermes annandalei is a fungus-growing termite fromsoutheastern Asia that builds a massive epigeous nest up to1.50 m in height and 2 m in diameter. The reproductive cyclegenerally starts in late December with the differentiation of firstnymphs. Molting of alate imagoes occurs in early April and dis-persal flights are observed during May and June.
Female alates were collected between 1996 and 1998 in north-ern Vietnam near Tam Dao (Peppuy et al., 1999) at differentphysiological ages: 1) Freshly molted alates and pigmented alatesof several weeks old were taken directly inside the nest. 2) Ma-ture alates of about 2 months old were collected during thedispersal flight. They were fixed immediately or after showingsexual behavior.
For scanning electron microscopy, insects were killed in alco-hol. Abdominal sternites 5–8 and tergites 6–10 were dissected,cleaned in a 1 M aqueous solution of sodium hydroxide, rinsedwith water, dehydrated in absolute ethanol, dried, and coatedwith a mixture of gold-palladium. Specimens were observed witha Philips XL 30 scanning electron microscope at the Centre ofMicroscopy Applied to Biology, University of Burgundy (CMAB).
For transmission electron microscopy, glands were removedand fixed for 16 h at 4°C in a 2% paraformaldehyde / 3% glutar-
*Correspondence to: A. Quennedey, UMR CNRS 5548, Universitede Bourgogne, 6 boulevard Gabriel, F-21000 Dijon, France.E-mail: [email protected]
Published online inWiley InterScience (www.interscience.wiley.com)DOI: 10.1002/jmor.10185
JOURNAL OF MORPHOLOGY 262:683–691 (2004)
© 2004 WILEY-LISS, INC.

aldehyde / 0.1 M sucrose mixture in 0.1 M cacodylate buffer andpostfixed for 1 h in 1% osmium tetroxide in the same buffer. Afterdehydration, specimens were embedded in an epon-araldite mix-ture. Thin sections were stained with 1% Toluidine blue-1% AzurII mixture in 1% sodium borate in distilled water and viewed withlight microscopy. Ultrathin sections were stained with uranylacetate and lead citrate prior to observation with a Hitachi H-600electron microscope (CMAB).
RESULTSSexual Behavior
After the dispersal flight and the landing on avertical surface, female alates of Macrotermes an-nandalei take a calling posture (Fig. 1). While keep-ing still, head down, they spread their wings andextend their last abdominal segments exposing thefive tergal glands of tergites 6–10 and the two PSGsof sternites 6–7. In contrast, the sternal gland lo-cated on sternite 5 is not exposed. Males are rapidlyattracted and soon they touch the abdominal ex-tremity of the female with their palps and antennae.Both sexes shed their wings and undertake a nuptialpromenade in tandem, with the female as the leader.Similar behaviors, except the spreading and shed-ding of the wings, were observed in laboratory withartificially dealated individuals.
Abdominal Glandular Set
Three types of abdominal glands are thus found inMacrotermes annandalei (Fig. 2). The first type cor-responds to the gland located on sternite 5 (200 �m
long and 400 �m wide). That is the unique sternalgland seen in neutral castes of Macrotermes andinvolved in the secretion of a trail pheromone (Pep-puy et al., 2001). It is also present in male imagoesand corresponds to the sternal gland stricto sensuwell known in termites. On the contrary, PSGs andtergal glands are only present in female imagoesand they extend largely on sternites 6–7 and tergi-tes 6–10 (200 �m long and 3,000 �m wide).
Scanning Electron Microcopy
On the anterior part of sternites 6–7 (Fig. 3A,B)and tergites 6–10 (Fig. 3C), the glandular cuticle isornamented with about 1,000 depressions reaching10 �m in diameter. Such depressions are absent onthe anterior part of the other abdominal segments.Generally, in PSGs a single, minute aperture reach-ing about 0.5 �m in diameter is visible on theirbottom (Fig. 3B). The depressions are often morereduced in tergal glands, where up to three aper-tures can be observed (Fig. 3C).
Histological Features of Glands
In swarming female alates numerous cuticularpouches appear beneath the glandular cuticle ofPSGs (Fig. 3D) and tergal glands (Fig. 3E). Suchunusual structures encountered in both types ofglands suggest a common cytological organization.
Transmission Electron Microscopy of PSGsFemale alates collected in the nest. In unpig-
mented freshly molted females, the cuticle of PSGsis mainly made of pre-ecdysial layers and reaches 4�m in thickness. Collapsed and strongly folded cu-ticular pouches extend deeply inside the glandularepithelium (Figs. 4A, 7A). They are made of epicu-ticle lined by very thin cellular septa separated by atiny intercellular space that is almost free of junc-tions (Fig. 4B). Only a few apical junctions are evi-
Fig. 1. Macrotermes annandalei. Female alate in calling pos-ture. After the dispersal flight, the female lands on a verticalsurface, head down, spreading her wings and extending the lastabdominal segments to expose tergal glands and posterior sternalglands (PSGs). �5.
Fig. 2. Macrotermes annandalei. Diagram of female dealate incalling posture showing the location of abdominal glands. Thesternal gland found on sternite 5 (arrowhead) is not involved insexual behavior and is covered by the posterior margin of sternite4 while the two PSGs found on sternites 6–7 (open arrows) andthe five tergal glands found on tergites 6–10 (arrows) are widelyexhibited.
684 A. QUENNEDEY ET AL.

dent between cells lining a similar flattened pouch(Fig. 4C). Numerous sections of cuticular canals sur-rounded by their canal cell and secretory cell arevisible in the basal part of PSGs (Fig. 4A). Both cellsmake up a glandular unit also classified as Class 3cells (Noirot and Quennedey, 1974, Noirot andQuennedey, 1991). Inside the cytoplasm of secretorycells, the receiving part of the cuticular canal issurrounded by short microvilli. Free ribosomes gath-ered in small aggregates and well-developed Golgibodies secreting numerous small vesicles are abun-dant in these cells (Fig. 4D).
In pigmented females, several modifications areobserved. The intercellular spaces inflate betweenthe collapsed pouches and this movement disruptsthe cell-to-cell organization (Fig. 4E). Narrow inter-cellular spaces remain between the secretory cells,which grow in size, while large secretory vesiclesgather in their cytoplasm (Fig. 4F).
Female alates collected at the dispersalflight. The glandular cuticle of PSGs reaches about6 �m thick. It is lined by flattened epidermal cellsenlarged only in the vicinity of the nucleus. Numer-ous extracellular pouches of about 25 �m in diame-ter are full of loose secretory material; they give avery lacunar appearance to the upper half of theglandular epithelium (Figs. 3D, 5A, 7B). They cor-respond to the collapsed pouches found in alatescaptured in the nest. A short cuticular duct opens ina depression of the glandular cuticle and connectseach pouch with the outside (Fig. 5B). Each extra-cellular pouch is lined with very thin cytoplasmicsepta (less than 0.5 �m thick) covered with epicu-ticle (Fig. 5C).
Sections of cuticular canals, made of cuticulin andinner epicuticle, appear inside the basal part of thesepta (Fig. 5D). No membrane separates these ca-nals from pouches. Therefore, each conducting canaland a part of the pouch are secreted by the same cell,corresponding to the canal cell found in each glan-dular unit. Several conducting canals open into thesame pouch, leading to the presence of several glan-dular units around it. Apical septate junctions arealso observed between adjacent canal cells lining thesame pouch (Fig. 5D).
Fig. 3. Macrotermes annandalei. A–C: Sexual glandular areasobserved on the abdomen (sternites 6–7 and tergites 6–10). SEM.A: Low magnification in the mid anterior part of sternite 6. Ovaldepressions (arrows) are observed among the cuticular pavementof epidermal cells. B: High magnification showing the minuteaperture (arrows) opening in the bottom of cuticular depressions.C: Similar cuticular structures are found on tergite 8. Severalapertures are more often observed in the bottom of depressions(arrows). D,E: Sexual glands of females collected during the dis-persal flight. Semithin sections. D: PSG of sternite 6. E: Tergalgland of tergite 8. In both cases, numerous pouches (arrows)reaching the half of thickness of glands are found close to thecuticle (arrowheads).
685POSTERIOR STERNAL GLANDS OF M. ANNANDALEI

Figure 4
686 A. QUENNEDEY ET AL.

In the secretory cell, the receiving canal comprisesseveral inner epicuticular layers, giving it a felt-likeappearance (Fig. 5D). Dense aggregates of chroma-tin are evident in the secretory cell nucleus whilemitochondria, free ribosomes, Golgi bodies, andabundant large-sized vesicles full of loose materialare found in the cytoplasm.
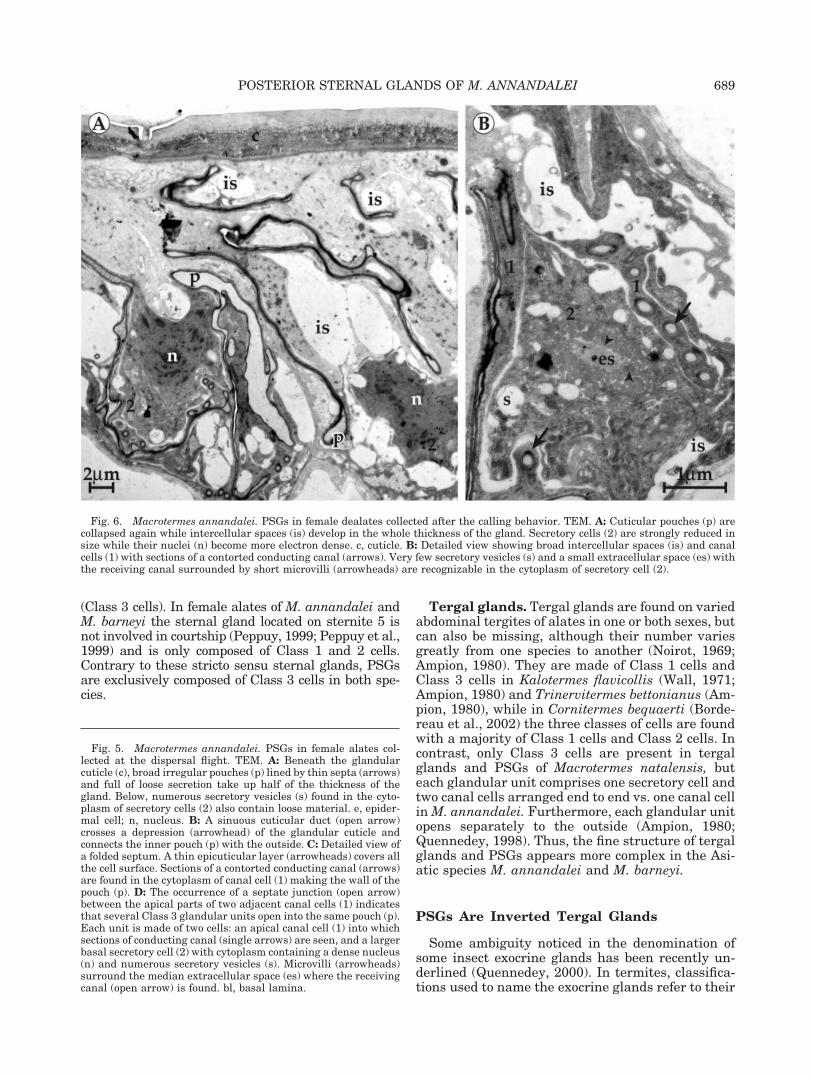
Female dealates collected after the callingbehavior. PSGs are now reduced in thickness: 25�m vs. 50 �m at the time of dispersal flight. Theinflated pouches found in swarming females are col-lapsed again and strongly folded (Figs. 6A, 7C). Withthe drastic reduction in size of secretory cells,pouches extend more deeply into the glandular epi-thelium. A dense nucleus and rare secretory vesiclesare still recognizable into the cytoplasm (Fig. 6B).Broad intercellular spaces are now visible betweencanal cells and between secretory cells.
DISCUSSIONFunctional Peculiarities
Compared to the usual Class 3 glandular unitsalready described, the secretory process found inPSGs and tergal glands of Macrotermes annandaleiappears more complex, as summarized in Figure 7.In freshly molted alates, the small vesicles found inthe cytoplasm of secretory cells are produced bywell-developed Golgi bodies (Fig. 7A). Later, in olderpigmented alates, vesicles are so abundant and sogreatly enlarged that they take up the main part ofsecretory cells. In swarming alates, the loose secre-tion previously contained in the vesicles of secretorycells is now gathered in the swollen cuticularpouches (Fig. 7B). At last, after the calling behavior,no more secretion can be observed, either in thecytoplasm of secretory cells, which are drasticallyreduced in size, or in the cuticular pouches, whichcollapse again (Fig. 7C).
The different steps of the secretory process alsoappear closely associated with the temporary devel-
opment of intercellular spaces full of hemolymph.Initially, narrow spaces exist between canal cells.They gradually inflate while alates remain in thenest but they are finally resorbed just before thedispersal flight. Afterwards, spaces develop again indealates while glandular units regress in size. Thefirst inflation of intercellular spaces maintains thepouches collapsed during the biosynthesis of secre-tory products inside secretory cells (Fig. 7A). Thesubsequent reduction of these spaces makes easieror even brings forth the filling of the pouches (Fig.7B). During calling behavior, a second inflationsqueezes the pouches again, which allows the dis-charge of secretion to the outside (Fig. 7C).
Structural Comparison With Other SexualGlands
Posterior sternal glands. Compared to PSGs ofMacrotermes annandalei, we must point out threestriking differences between the three species, Poro-termes, Stolotermes, and Prorhinotermes, wherePSGs have been previously recorded (Ampion,1980). First, they are only found in male imagoes vs.female imagoes in M. annandalei. Second, they arelocated on sternites 8–9 vs. sternites 6–7. Finally,they are made of Class 1 cells and Class 3 cells vs.only Class 3 cells. However, more accurate behav-ioral and structural studies are needed to establishpotential similarities between all PSGs of termites.
It must be noted that PSGs are also present inAsiatic Macrotermes barneyi, M. latignathus, M.malaccensis, and African M. natalensis species (un-publ. data). In African species, ultrastructural datashow that the cell organization is very similar inPSGs and tergal glands but differs from that ofPSGs and tergal glands found in M. annandalei andM. barneyi (unpubl. data).
Sternal gland stricto sensu. Except in the an-cestral termite Mastotermes darwiniensis, where threesternal glands are found on sternites 3–5, a singlesternal gland stricto sensu located on sternite 4 or 5 isalways present in all castes of other termites (Noirot,1969, 1995). This gland is generally involved in thesecretion of a trail pheromone in workers and soldiersbut can also be involved in sexual behavior in alates.When the sternal gland is the only source of sexpheromone, it often becomes highly enlarged, as inmales of Hodotermes mossambicus (Leuthold, 1977;Quennedey, 1978), Anacanthotermes ochraceus (Noi-rot, 1969; Quennedey, 1978), and in females of Reticu-litermes santonensis (Noirot, 1969; Quennedey, 1977,1978), Trinervitermes bettonianus (Leuthold and Lus-cher, 1974; Quennedey and Leuthold, 1977) andPseudacanthotermes spiniger (Laduguie, 1993). Ac-cording to the cytological classification of gland cellsproposed by Noirot and Quennedey (1974, 1991), thehypertrophied sternal gland found on sternite 4 or 5 ismade of modified epidermal cells (Class 1 cells),oenocyte-like cells (Class 2 cells), and more or lessabundant glandular units with a cuticular canal
Fig. 4. Macrotermes annandalei. PSGs in female alates col-lected inside the nest. TEM. The glandular cuticle is alwaysarranged at the top. A–D: Few-day-old females. A: Low magnifi-cation of the glandular epithelium. Cuticular pouches (p) arestrongly collapsed. Numerous sections of conducting canals (ar-rows) are seen, while secretory cells (2) have a dense cytoplasm.Note also the depression of the cuticle (open arrow). B: Narrowintercellular spaces (is) are seen beneath canal cells (1) lining acollapsed pouch (p). C: A septate junction (arrow) links two ad-jacent canal cells (1). The irregular appearance of the cuticlelining the pouch (arrowheads) cannot be confused with the regu-lar section of a conducting canal. D: Detailed view of secretory cellcytoplasm showing a Golgi body (open arrow), small secretoryvesicles (s), and a receiving canal (arrow) located within a reducedextracellular space limited by short microvilli (arrowheads).E,F: Pigmented females. E: Intercellular spaces (is) found be-tween canal cells (1) become inflated while pouches (p) are col-lapsed. F: The intercellular space (open arrow) found betweensecretory cells (2) is reduced. In the cell cytoplasm, numeroussecretory vesicles (s) full of loose material are now bigger.
687POSTERIOR STERNAL GLANDS OF M. ANNANDALEI

Figure 5
688 A. QUENNEDEY ET AL.
(Class 3 cells). In female alates of M. annandalei andM. barneyi the sternal gland located on sternite 5 isnot involved in courtship (Peppuy, 1999; Peppuy et al.,1999) and is only composed of Class 1 and 2 cells.Contrary to these stricto sensu sternal glands, PSGsare exclusively composed of Class 3 cells in both spe-cies.
Tergal glands. Tergal glands are found on variedabdominal tergites of alates in one or both sexes, butcan also be missing, although their number variesgreatly from one species to another (Noirot, 1969;Ampion, 1980). They are made of Class 1 cells andClass 3 cells in Kalotermes flavicollis (Wall, 1971;Ampion, 1980) and Trinervitermes bettonianus (Am-pion, 1980), while in Cornitermes bequaerti (Borde-reau et al., 2002) the three classes of cells are foundwith a majority of Class 1 cells and Class 2 cells. Incontrast, only Class 3 cells are present in tergalglands and PSGs of Macrotermes natalensis, buteach glandular unit comprises one secretory cell andtwo canal cells arranged end to end vs. one canal cellin M. annandalei. Furthermore, each glandular unitopens separately to the outside (Ampion, 1980;Quennedey, 1998). Thus, the fine structure of tergalglands and PSGs appears more complex in the Asi-atic species M. annandalei and M. barneyi.
PSGs Are Inverted Tergal Glands
Some ambiguity noticed in the denomination ofsome insect exocrine glands has been recently un-derlined (Quennedey, 2000). In termites, classifica-tions used to name the exocrine glands refer to their
Fig. 6. Macrotermes annandalei. PSGs in female dealates collected after the calling behavior. TEM. A: Cuticular pouches (p) arecollapsed again while intercellular spaces (is) develop in the whole thickness of the gland. Secretory cells (2) are strongly reduced insize while their nuclei (n) become more electron dense. c, cuticle. B: Detailed view showing broad intercellular spaces (is) and canalcells (1) with sections of a contorted conducting canal (arrows). Very few secretory vesicles (s) and a small extracellular space (es) withthe receiving canal surrounded by short microvilli (arrowheads) are recognizable in the cytoplasm of secretory cell (2).
Fig. 5. Macrotermes annandalei. PSGs in female alates col-lected at the dispersal flight. TEM. A: Beneath the glandularcuticle (c), broad irregular pouches (p) lined by thin septa (arrows)and full of loose secretion take up half of the thickness of thegland. Below, numerous secretory vesicles (s) found in the cyto-plasm of secretory cells (2) also contain loose material. e, epider-mal cell; n, nucleus. B: A sinuous cuticular duct (open arrow)crosses a depression (arrowhead) of the glandular cuticle andconnects the inner pouch (p) with the outside. C: Detailed view ofa folded septum. A thin epicuticular layer (arrowheads) covers allthe cell surface. Sections of a contorted conducting canal (arrows)are found in the cytoplasm of canal cell (1) making the wall of thepouch (p). D: The occurrence of a septate junction (open arrow)between the apical parts of two adjacent canal cells (1) indicatesthat several Class 3 glandular units open into the same pouch (p).Each unit is made of two cells: an apical canal cell (1) into whichsections of conducting canal (single arrows) are seen, and a largerbasal secretory cell (2) with cytoplasm containing a dense nucleus(n) and numerous secretory vesicles (s). Microvilli (arrowheads)surround the median extracellular space (es) where the receivingcanal (open arrow) is found. bl, basal lamina.
689POSTERIOR STERNAL GLANDS OF M. ANNANDALEI

location on the body wall and their role in the soci-ety, both criteria being generally linked. However,in a few cases the same sternal gland produces thetrail pheromone in neutral castes and the sex pher-omone in imagoes.
In the present study, we made the distinctionbetween the sternal gland stricto sensu found onsternite 5 and PSGs found on sternites 6–7 becausetheir structure and function are very different. Inaddition, because of their ultrastructural similari-ties, PSGs can be considered as the equivalent oftergal glands located on the opposite side of theabdomen. Yet, in this first study we kept the denom-ination previously given for these glands by Ampion(1980) before studying PSGs and tergal glands inancestral species (Mastotermes, Porotermes, Stolot-ermes, and Prorhinotermes) more accurately and re-considering their denomination.
Phylogenetic Significance of PSGs
Until now PSGs have been found in several speciesof Asiatic Macrotermes, i.e., M. barneyi, M. latig-nathus, M. malacensis and in the African species M.natalensis (Peppuy, 1999). The Macrotermes genus be-longs to the Macrotermitinae subfamily and includesall fungus-growing termites which are phylogeneti-cally considered at a basal position in the Termitidae,the most derivative and most diversified family (Kam-bhampati and Eggleton, 2000; Noirot, 2001). In theancestral genera Porotermes and Stolotermes belong-ing to Termopsidae a single sternal gland stricto sensuis found on sternite 4 and PSGs are present in malesand are located on sternites 8–9 (Ampion, 1980; Fig.8). Their role remains unknown. PSGs also exist inmale alates, on sternites 8–9 in Prorhinotermes, occu-pying a basal position in Rhinotermitidae, which is akey taxon in the evolution of termites (Parmentier andRoisin, 2003). PSGs could be considered as relics of anancestral situation, since in the most basal termite,Mastotermes darwiniensis, seven sternal glands (in abroad sense) are found in males on sternites 3–9 andfive sternal glands are found in females on sternites
Fig. 7. Macrotermes annandalei. Diagrams summarizing themain structural features involved in the secretory process ofPSGs. A: Freshly molted female alate found in the nest: the apicalparts of several canal cells (1) form a common infolded pouch (p)opening to the outside by a short cuticular duct (open arrow).Intercellular spaces (is) are well developed between canal cellsbut are tiny between secretory cells (2) into which Golgi bodies(arrows) secrete small vesicles. B: Female alate collected at thedispersal flight: secretory cells (2) contain large secretory vesicles(s) whereas Golgi bodies (arrows) regress in size. The pouch (p) isfull of secretion (not drawn), strongly inflated and next to theneighboring pouches. Intercellular spaces are very reduced.C: Female dealate collected after the calling behavior: the secre-tion previously contained in secretory cells (2) and pouches (p)has been released to the outside. The cell cytoplasm drasticallyregresses in size, pouches collapse again while intercellularspaces (is) develop everywhere. c, cuticle; e, epidermal cell; nucleiin black.
690 A. QUENNEDEY ET AL.

3–7. According to Ampion (1980), the structure ofPSGs found on sternites 6–9 in males and on sternites6–7 in females of Mastotermes would be similar to thatof PSGs in Porotermes, Stolotermes, and Prorhinot-ermes. However, a more detailed study would be nec-essary to know whether all these PSGs are homolo-gous.
ACKNOWLEDGMENTS
The authors thank Josette Relot (Centre of Mi-croscopy Applied to Biology) for technical assistance,Profs. Charles Noirot and Jean Deligne for usefulcomments on the manuscript and Jessica Meredithfor kindly revising the English manuscript.
LITERATURE CITED
Ampion M. 1980. Les glandes tergales des imagos de Termites:etude comparative et signification evolutive. Third Cycle thesis,University of Dijon.
Ampion M, Quennedey A. 1981. The abdominal epidermal glandsof termites and their phylogenetic significance. In: Howse PE,Clement JL, editors. Biosystematics of social insects. London:Academic Press. p 249–261.
Barth R. 1955. Uber die tergitalen Druesenfelder von Syntermesdirus (Isoptera). Rev Brasil Biol 15:257–263.
Bordereau C, Cancello EM, Semon E, Courrent A, Quennedey B.2002. Sex pheromone identified after solid phase microextrac-tion from tergal glands of female alates in Cornitermes be-quaerti (Isoptera, Nasutitermitinae). Insectes Soc 43:209–215.
Kambhampati S, Eggleton P. 2000. Taxonomy and phylogeny oftermites. In: Abe T, Bignell DE, Higashi M, editors. Termites:evolution, sociality, symbioses, ecology. Dordrecht: Kluwer Ac-ademic. p 1–23.
Ladugie N. 1993. Pheromones de piste et pheromones sexuelleschez les termites. Etude chez Reticulitermes santonensisFeytaud, Pseudacanthotermes spiniger (Sjostedt) et Nasuti-termes lujae (Wasmann). Ph.D. thesis, University of Burgundia.
Leuthold R. 1977. Postflight communication in two termite spe-cies, Trinervitermes bettonianus and Hodotermes mossambicus.Proc VIIIth Int Congr IUSSI, Wageningen. p 62–64.
Leuthold R, Luscher M. 1974. An unusual caste polymorphism ofthe sternal gland and its trail pheromone production in thetermite Trinervitermes bettonianus. Insectes Soc 21:335–341.
Noirot Ch. 1969. Glands and secretions. In: Krishna K, WeesnerFM, editors. Biology of termites. New York: Academic Press.p 89–123.
Noirot Ch. 1995. The sternal glands of termites: segmental pat-tern, phylogenetic implications. Insectes Soc 42:321–323.
Noirot Ch. 2001. The gut of termites (Isoptera). Comparativeanatomy, systematics, phylogeny. II. Higher termites (Termiti-dae). Ann Soc Ent Fr 37:431–471.
Noirot Ch, Quennedey A. 1974. Fine structure of insect epidermalglands. Annu Rev Entomol 19:61–80.
Noirot Ch, Quennedey A. 1991. Glands, gland cells, glandularunits: some comments on terminology and classification. AnnSoc Ent Fr 27:123–128.
Parmentier D, Roisin Y. 2003. Caste morphology and develop-ment in Termitogeton nr. planus (Insecta, Isoptera, Rhinoter-mitidae). J Morphol 255:69–79.
Pasteels JM. 1972. Sex-specific pheromones in a termite. Experi-entia 28:105–106.
Pasteels JM, Bordereau C. 1998. Releaser pheromones in ter-mites. In: Van der Meer RK, Breed MD, Winston ML, EspelieKE, editors. Pheromone communication in social insects. Boul-der, CO: Westview Press. p 193–215.
Peppuy A. 1999. Termites du nord-Vietnam, communicationchimique et isolement specifique. Ph.D. thesis, University ofParis VI.
Peppuy A, Robert A, Bordereau C. 1999. Comportement sexuel etpheromones chez deux especes sympatriques de Macrotermesasiatiques, M. annandalei et M. barneyi (Isoptera, Macroter-mitinae). Mise en evidence de nouvelles glandes sexuelles.Actes Colloq Insectes Soc 12:143–150.
Peppuy A, Robert A, Semon E, Ginies C, Lettere M, Bonnard O,Bordereau C. 2001. (Z)-Dodec-3-en-1-ol, a novel termite trailpheromone identified after solid phase microextraction fromMacrotermes annandalei. J Insect Physiol 47:445–453.
Quennedey A. 1977. An ultrastructural study of the polymorphicsternal gland in Reticulitermes santonensis (Isoptera, Rhinot-ermitidae); another way of looking at the true termite trailpheromone. Proc VIII Congr IUSSI, Wageningen. p 48–49.
Quennedey A. 1978. Les glandes exocrines des termites. Ultra-structure comparee des glandes sternales et frontales. Doctoralthesis, University of Dijon.
Quennedey A. 1998. Insect epidermal gland cells: ultrastructureand morphogenesis. In: Harrison FW, Locke M. editors. Micro-scopic anatomy of invertebrates, vol. 11A. Insecta. New York:Wiley-Liss. p 177–207.
Quennedey A. 2000. Perspectives on four decades oftransmission-electron microscopy on insect exocrine glands.Atti Accad Naz Ital Ent Rend 48:85–116.
Quennedey A, Leuthold R. 1977. Fine structure and pheromonalproperties of the polymorphic sternal gland in Trinervitermesbettonianus (Isoptera, Termitidae). Insectes Soc 25:153–162.
Wall M. 1971. Zur Geschletsbiologie des Termite Kalotermes fla-vicollis (Fabr.) (Isoptera). Acta Tropica 28:17–60.
Fig. 8. Phylogenetic relationships of termites (from Kamb-hampati and Eggleton, 2000). Only subfamilies with generashowing PSGs are considered. Their occurrence in one or bothsexes and their location on the abdomen are given. Continuouslines: families; 1, Mastotermitidae; 2, Hodotermitidae; 3, Termop-sidae; 4, Kalotermitidae; 5, Serritermitidae; 6, Rhinotermitidae;7, Termitidae; dotted lines: subfamilies; 3b, Porotermitinae; 3c,Stolotermitinae; 6a, Prorhinotermitinae; 7a, Macrotermitinae.
691POSTERIOR STERNAL GLANDS OF M. ANNANDALEI





























