Embed Size (px)
Citation preview

[CANCER RESEARCH 44, 734-743, February 1984]
Tumoricidal Response following Perfusion over Immobilized Protein A:
Identification of Immunoglobulin Oligomers in Serum after Perfusionand Their Partial Characterization1
Joseph Balint, Jr.,2 Yusei Ikeda, John J. Langone, William T. Shearer, lerachmiel Daskal, Katheryn Meek,Gary Cook, Joseph Henry, and David S. Terman3
The Cancer Biology Program, Department of Medicine, Baylor College of Medicine, Houston Texas 77030
ABSTRACT
Previously, we showed that perfusion of plasma from hostsbearing breast adenocarcinoma over immobilized staphylococcalprotein A resulted in objective tumor regressions. In the presentstudy, sera perfused in vitro over immobilized staphylococcalprotein A were analyzed by physicochemical and ¡mmunochem-ical methods to characterize newly formed products. Sera fromnormal and breast adenocarcinoma-bearing dogs showed increased levels of Clq-binding IgG after perfusion over a strain
of staphylococcus that is protein A rich (Cowan I), but not proteinA deficient (Woods 46). C1q binding levels were also increasedin normal and tumor-bearing canine or human sera which were
perfused over purified protein A immobilized in collodion charcoal(PACC), and this increase was localized in sucrose densitygradient fractions ranging from 7S to 19S. Polyacrylamide gelelectrophoresis analysis of the high-molecular-weight fraction inpostperfusion canine sera, isolated by G-200 fractionation and
immunoaffinity chromatography, showed predominantly heavyand light immunoglobulin chains of canine IgG. Furthermore,protein A was released from PACC after perfusion with serumor solutions containing IgG or albumin from humans, dogs, andchickens. After serum perfusion over PACC, protein A wasidentified in the effluent by additional studies as follows: (a)polyacrylamide gel electrophoresis analysis showed that eluted125l-protein A comigrated with the protein A marker; (b) postper
fusion C1q-binding complexes, isolated by gel filtration underdissociating conditions and affinity chromatography on IgG-Sepnarose showed a single precipitin band with normal human(protein A reactive) but not chicken (protein A unreactive) serum.Protein A released from PACC which appeared in postperfusionsera was associated with immunoglobulins in macromolecularcomplexes since (a) eluted 125l-protein A was largely (NH4)2S04
and polyethylene glycol precipitable, whereas free protein A wasnot, and it sedimented in sucrose density gradient fractionsdistributed beyond the 7S marker, compared to free protein Awhich localized below 7S; (b) radiolabeled protein A eluting fromPACC after serum perfusion showed 8-fold greater binding toC1q-coated tubes compared to free protein A; and (c) increasedC1q-binding IgG in postperfusion sucrose density gradient frac
tions corresponded to the appearance of protein A in parallel
1Supported by Grant 9R01 CA 28841 from the NIH, from a special fund of The
Methodist Hospital, from the Winograd Memorial Foundation, and from McNeilPharmaceutical, Inc.
2 Present address: IMRE Corporation, 130 Fifth Avenue North, Seattle, WA
98109.3 Recipient of a Research Career Development Award (K04 AI00302) from the
NIH. To whom requests for reprints should be addressed, at Baylor College ofMedicine, Department of Medicine, One Baylor Plaza, Houston, TX 77030.
Received June 22, 1983; accepted November 3, 1983.
gradient fractions. In addition to protein A, analysis of the components of postperfusion complexes in human and canine serabound to solid-phase C1q using radiolabeled monospecificF(ab')2 fragments demonstrated IgG and IgM in canine com
plexes, while human complexes contained IgG, IgA, IgM, andC3. Human and canine serum containing a trace amount of 131I-IgG were passaged over PACC containing 125l-proteinA, and the
effluents were chromatographed on Sepharose 6B. By use ofmolecular weight standards, the molecular weight of complexesranged from 600,000 to 2,000,000. Hence, perfusion of normalor tumor-bearing sera over Staphylococcus aureus Cowan Istrain or PACC results in the formation of C1q binding- andpolyethylene glycol-precipitable oligomers composed predomi
nantly of IgG and protein A. These complexes may activatecellular and humoral inflammatory systems after entering thecirculation and contribute to the tumoricidal effects and toxicitypreviously observed after administration of plasma perfused overimmobilized staphylococcal protein A.
INTRODUCTION
Protein A is a constituent of the cell wall of many strains ofStaphylococcus aureus, which binds selectively to immunoglobulins and immune complexes from many mammalian species atsites located in the IgG-Fc region. These complexes display
diverse biological activity, including complement binding andactivation (6, 9, 11-13, 23). Previously, in studies of dogs with
spontaneous mammary adenocarcinoma, an excellent model ofhuman breast cancer, plasma was circulated over SAC,4 which
were immobilized in a microporous membrane filtration systemand placed on line with plasma emerging from a continuous-flowplasma-cell separator (27). Shortly after perfusion of one plasmavolume over protein A-bearing staphylococci, tumor necrosis
followed by objective tumor regressions were observed. Theseeffects were not seen when plasma was passaged over proteinA-deficient staphylococci (27). Findings of tumor regressions inour experimental canine system were confirmed in an independent study (8). We refined the perfusion system, and similarnecrolytic responses were observed after plasma perfusion overpurified PACC (28). This system was recently used to treat 5consecutive patients with advanced breast adenocarcinoma, andafter repeated plasma perfusion, objective tumor regressionswere noted in 4 patients (29).
In prior studies of patients with breast adenocarcinoma, we
'The abbreviations used are: SAC, Staphylococcus aureus Cowan strain I;
PACC, protein A immobilized in collodion charcoal; SAW, S. aureus Woods strain;PBS, phosphate-buffered saline; AHG, aggregated human IgG; ACG, aggregatedcanine IgG; PEG, polyethylene glycol; SDS, sodium dodecyl sulfate; PAGE, polyacrylamide gel electrophoresis.
734 CANCER RESEARCH VOL. 44
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

Immunoglobulin Oligomers in Serum after Perfusion over Protein A
noted the rapid onset of acute pain, as well as gross andhistopathological changes in chest wall tumors after extracor-
poreal perfusion of small volumes of plasma over PACC (29).These results suggested that these effects might be producedby factors generated in plasma after passage over PACC. Totest this hypothesis, the procedure was technically simplified byeliminating the on-line extracorporeal circulation system and,
instead, small aliquots of autologous or homologous plasma,previously collected by phlebotomy, were perfused over PACCoff-line, followed by direct infusion into patients (29). Patientstreated entirely with the off-line plasma perfusion showed acutetumor-inflammatory responses, and tumor-regressive effectssimilar to those observed with the on-line extracorporeal system
(29). Moreover, the tumoricidal responses in both dogs and manwere accompanied by serological and immunohistochemicalchanges, including increased solid-phase C1q-binding IgG, and
a decline in C3 associated with deposits of IgG and C3 withintumor tissue (27, 29).
From the foregoing observations, it appeared that the interaction of plasma with staphylococcal protein A resulted in theformation of products which were capable of activating hostresponses and producing tumoricidal activity, as well as someof the side effects of the procedure (28-30). Therefore, we
attempted to identify and characterize these products. For thispurpose, we developed an in vitro perfusion system similar tothe off-line system used in our patients, in which sera fromtumor-bearing or normal dogs and patients were passaged over
SAC or PACC. Preperfusion and postperfusion serum sampleswere analyzed by physicochemical and immunochemical methods and compared. The results of the present study show thatperfusion of normal or tumor-bearing serum over SAC or PACCresults in the generation of C1q-binding complexes, composed
predominantly of immunoglobulins and protein A. We postulatethat these complexes may be a major constituent responsiblefor the previously described tumoricidal activity and toxicity ofthis therapeutic system (29, 30).
MATERIALS AND METHODS
Animalsand Sera
Sera obtained from dogs with histologically confirmed mammary ad-
enocarcinoma were used in this study. Normal dog sera were obtainedfrom animals raised in a controlled environment and examined at regularintervals by a veterinarian. Sera from all animals were collected andstored at -70° until use. Normal human sera were obtained from
volunteers with no evidence of active or chronic disease. Tumor-bearing
sera were obtained from patients with breast adenocarcinoma who hada clinically evident tumor burden at the time serum was obtained.
S. aureus, Immobilized Protein A
Cowan I (SAC) (ATCC 2530) and Woods 46 (SAW) (ATCC 1830)strains were cultured, fixed with 0.5% formaldehyde, and stored at 70°
until use, as previously described (27). For in vitro perfusion studies, theheat-killed and formalin-stabilized staphylococci were washed in 0.15 M
NaCI, and 10 g were resuspended to a final concentration of 1% in 0.15M NaCI. The staphylococcal suspension was then pumped at a rate of20 ml/min into a modified 0.2-ntn pleated filter (Gelman Instrument Co.,
Ann Arbor, Ml) until the bacteria were embedded in the membrane andall 0.15 M NaCI had passed through the filter. The filter was then emptiedof residual fluid by air expulsion. In perfusion studies, the filter wasloaded with 100 ml of serum which were then perfused through the
system by forced-air pressure at a rate of 5 ml/min. Effluent fractions (1to 5 ml) were collected and stored at -70° until use. In selected studies,
normal or tumor-bearing sera were simultaneously perfused by dual
channel tubing through filters containing SAC and SAW, which werearranged in parallel. Effluent samples from each system were collectedand stored at -70°. Purified protein A (Pharmacia, Piscataway, NJ; 5
mg), was immobilized in 30 g of collodion charcoal and washed withPBS by a modification of previously described methods (25, 26).
Perfusion Studies
For perfusion studies, 6 g of charcoal containing approximately 1 mgof immobilized protein A were spread in a thin layer between two 20mesh stainless steel screens in a Lexan-stainless steel cartridge. The
PACC was washed with 500 to 1000 ml of 0.01 M PBS, pH 7.5. Afterremoval of residual PBS, 5 to 50 ml of sera were introduced into thecartridge and incubated with PACC for 5 min at 25°. The serum was
evacuated from the chamber by gravity, and 1- to 5-ml effluent fractions
were collected. In selected studies, 1 mg of canine albumin was immobilized in collodion charcoal in a fashion similar to that for protein A andsimilarly incubated with human or canine serum. In other studies, 10 mlof normal human, canine, or chicken serum, solutions of human, canine,or chicken IgG or albumin, were perfused into a 0.15 M NaCI solution-filled Lexan chamber containing PACC with 125l-protein A immobilized
together with unlabeled protein A. Similar experiments were carried outwith collodion charcoal-containing immobilized canine albumin, togetherwith 125l-labeled canine albumin. Effluent fractions (1 ml) were collected
and radioactivity was measured.
Antisera and F(ab')2 Fragments
Purified F(ab')2 fragments from goat IgG specific for human IgG, IgM,
IgA, and C3 (Cappel Laboratories, Cochranville, PA) were used. F(ab')2
fragments of goat or rabbit IgG fractions (Cappel Laboratories) specificfor canine IgG, IgM, IgA, and C3 were prepared as previously described(24). Briefly, IgG fractions of these antisera were digested with pepsin in0.1 M acetate buffer, pH 4.1, using an enzyme/substrate ratio of 1/100(w/w) for 20 hr at 37°.The pH was adjusted to 8.0 by the addition ofsolid Tris salt, and F(ab')2 was separated from the other fragments by
gel filtration on Sephadex G-200 with PBS, pH 7.4, as the eluting buffer.
The preparations were free of intact IgG and Fc fragments, as shown bydouble immunodiffusion.
Radioiodination of Protein
Purified protein A, canine or human IgG, canine albumin, and F(ab')2fragments were labeled with 125Iby the method of McConahey and Dixon
(20). The mean specific activities were as follows: protein A, 0.35 to 0.40MCi/Mg; canine and human IgG, 0.15 to 0.25 MCi/Mg; canine albumin,0.40 MCi/Mg; F(ab'>2 fragments, 0.24 to 061 MCi/Mg. In selected studies,protein A was labeled with 125Iwith a mean specific activity of 0.9 to 1.0
MCi/Mg by the method of Bolton and Hunter (4).
Measurement of C1 q-binding Complexes, Immunoglobulins, and Pro
tein
C1q binding complexes were measured by a standard solid-phaseC1q-binding radioimmunoassay, as previously described (27). In someexperiments, radiolabeled F(ab')2 fragments specific for human or canine
IgG, IgM, IgA, or C3 were used to detect components of C1 q-bindingcomplexes. In preliminary experiments using 125l-C1q, we determined
that approximately 1.2 ¿tgof C1q were bound per tube. In some studies,results were referred to a standard curve using AHG or ACG, and wereexpressed as Mg equivalents AHG or ACG/ml. Mean normal values fornormal canine and human sera were 20.1 ±5.7 (S.D.) and 18.6 ±2.6Mg equivalents ACG or AHG/ml, respectively. In other studies, resultswere given as ng of anti-canine or anti-human IgG, IgM, IgA, or C3
FEBRUARY 1984 735
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

J. Balint, Jr., et al.
bound. For the detection of Clq-binding IgG in sucrose density gradientfractions, or concentrates of postperfusion effluents, 0.2- to 0.5-ml
aliquots were assayed. Preliminary experiments showed that increasingamounts of protein A, tested up to a concentration of 100 M9/ml andadded to normal canine serum, gave a linear increase in the quantity ofcomplexes that bound to C1 q-coated tubes.
Canine or human immunoglobulins were measured in serum by anephelometric assay, as previously described (10). In selected studies,quantitative measurements of canine IgG were performed by the methodof Mancini ef al. (19), using radial immunodiffusion in agarose gels (MilesLaboratories, Elkhart, IN). Protein concentrations of serum were measured by the method of Lowry et al. (18). The protein content ofsolubilized PEG-precipitated protein was determined by the method ofBradford (5) (Bio-Rad Laboratories, Richmond, CA).
Sucrose Density Gradient Fractionation
Serum samples (1.5 ml) obtained before and after perfusion over SACor PACC were layered on 15 ml of 10 to 40% (w/v) sucrose densitygradients in PBS. The samples were centrifugea at 40,000 rpm for 6 hrat 4° with a reorienting TV865B vertical rotor in a Sorvall OTD-65
ultracentrifuge, and 1.2-ml fractions were collected from the bottom of
the gradient. The positions of 7S and 19S were determined with purifiedcanine IgG and IgM (Pel-Freeze Biologicals, Rogers, AK) markers run in
parallel gradients.
Chromatography
Gel filtration Chromatography was carried out on Sepharose 6B orSephadex G-200 (Pharmacia). Buffers used were PBS, and 3-ml fractions
were collected.
SDS-PAGE
The high-molecular-weight IgG isolated by affinity Chromatography
was analyzed by PAGE in 7.5 or 10% gels, using the buffer systems ofLaemmli (14). The samples to be analyzed were mixed in a final solutioncontaining 5 M urea, 2% (w/v) SDS, 5% (v/v) 2-mercaptoethanol, 0.01%bromophenol blue and heated for 1 min at 100° before application to
gels. In other studies, normal canine serum was passaged over PACCwhich contained 125l-protein A (1 to 2 ^iCi), together with unlabeled
protein A (total immobilized protein, 1 mg). The postperfusion serum wasincubated with 5% PEG for 60 min at 25°and then centrifuged at 5000
x g for 15 min. The precipitate was resuspended in PBS and analyzedby SDS disc PAGE. After electrophoresis, the gel was sliced into 2-mm
segments and radioactivity measured.
Treatment of Sera with PEG and Ammonium Sulfate
Normal or tumor-bearing sera (50 ml) were passaged into a 0.15 MNaCI-filled Lexan chamber containing PACC with 125l-protein A immobi
lized together with unlabeled protein A (total immobilized protein A,approximately 1 mg). An additional 200 ml of 0.15 M NaCI was perfusedthrough the chamber to ensure removal of serum. Effluent fractionscontaining the peak 125I(50 ml) were pooled and treated as follows: (a)
to 15 ml of effluent an equal volume of saturated (NHi^SCv was added,incubated for 30 min at 4°,and centrifuged at 1500 x g for 30 min at4°;and (b) to 15 ml of effluent, 5 ml of 20% PEG were added, incubatedfor 60 min at 25°, and centrifuged at 5000 x g for 15 min at 4°.In all
samples, radioactivity in the precipitates was measured. In all instances,free 125Iprotein A used in the immobilization process was similarly
treated.
Evaluation of Stability of Immobilized Protein A
To evaluate the possible release of protein A from PACC duringperfusion of serum over it, the following studies were performed. Radio-
labeled protein A (1 to 2 /¿Ci)was mixed with 1 mg of unlabeled protein
A and immobilized in collodion charcoal, as described above. Greaterthan 95% of the radiolabeled protein A was incorporated into the collodion charcoal. The PACC was placed in a Lexan chamber and washedwith 3000 ml of 0.15 M NaCI. Normal human or chicken sera (10 ml)were then perfused through the chamber at a flow rate of 2 ml/min.Radioactivity which eluted from the PACC after perfusion with serum orsolutions containing IgG or albumin was measured in the effluent samples. The quantity of protein A released was calculated as a percentageof the total radioactivity added to the PACC after correction for theamount lost in the immobilization and washing procedures.
Isolation of Complexes and Identification of Components
Method 1. High-molecular-weight canine IgG in postperfusion sera
was isolated by a modification of a previously described procedure (1).Postperfusion sera (3 ml) were passaged through Sephadex G-200, and
fractions in the void volume peak were collected. The void volume peakwas passaged through Sepharose 4B to which goat IgG specific for 7chain of canine IgG was bound. The complexed protein was eluted with0.1 M glycine-HCI buffer, pH 2.5. The eluate was adjusted to pH 7.5 with5 N NaOH and concentrated to a final volume of 0.2 to 0.3 ml by negative-pressure dialysis. This sample was then examined by SDS-PAGE and
immunodiffusion.Method 2. Postperfusion sera were incubated with 5% PEG and
centrifuged, and the precipitate was resuspended and passaged overSephadex G-100 at pH 3.0, as in Method 1, above. The effluent wasneutralized and concentrated as above. To isolate protein A, the effluentwas treated further as follows: to remove small quantities of contaminating IgG, the included fractions were incubated with an equal volumeof saturated (NH4)2SO4 at pH 3.0 for 30 min at 4°,then centrifuged at5000 x g for 15 min at 4°C.The supernatant was dialyzed extensivelyagainst H2O at 4°and lyophilized. The protein was resuspended in PBS,
passaged over IgG-Sepharose 4B, and bound protein was eluted with0.1 M glycine-HCI buffer, pH 2.5. The pH was then adjusted to 7.5 with5 N NaOH, concentrated to a volume of 0.1 ml by negative-pressure
dialysis, and analyzed for the presence of protein A by ¡mmunodiffusionin 1% agarose, using 20 /¿Iof normal human or chicken sera per well,with continuous addition of 20-j/l aliquots of the test sample containing
putative protein A.Method 3. Pre- and postperfusion sera were subjected to sucrose
density gradient ultracentrifugation, as described above. Each fractionwas incubated with C1 q-coated tubes for 18 hr at 4°.The tubes were
washed twice with PBS containing 1% bovine serum albumin. To determine the presence of canine IgG, IgM, IgA, or C3,125l-labeled goat F(ab')2
fragments specific for canine IgG, IgM, IgA, or C3 were added, the tubeswere washed, and radioactivity was measured, as described above. Inother studies, 10 ml of effluent serum from PACC or canine albumincollodion charcoal columns were collected. The eluted 125l-protein A or
125l-labeled canine albumin was concentrated by ultrafiltration, using an
Amicon XM10 filter (Amicon, Lexington, MA), to a volume of 1 ml, and0.5-ml samples were incubated with C1q-coated tubes. The tubes werewashed and radioactivity was measured. As a control, free 125l-protein A
or 125l-labeled canine albumin solution was similarly incubated with C1q-
coated tubes and radioactivity was measured.
RESULTS
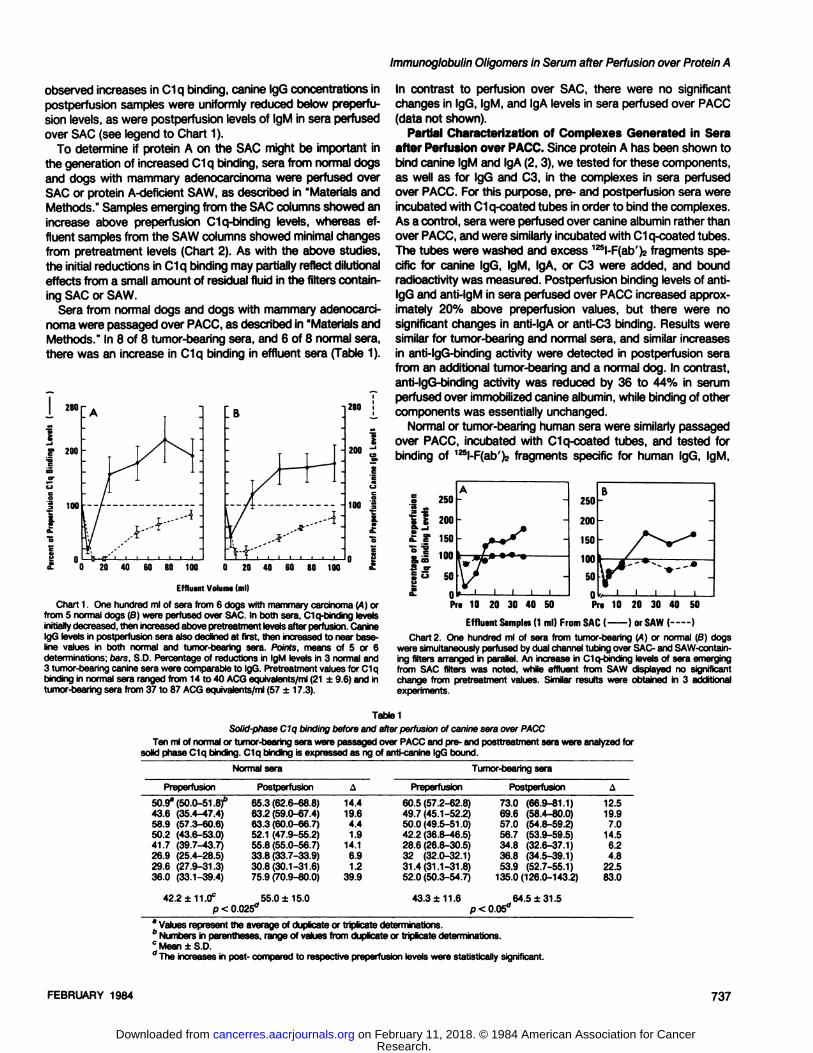
Changes in C1q Binding in Normal and Tumor-bearing Sera
after Perfusion over SAC, SAW, or PACC. Sera from 5 normaldogs or 6 dogs with mammary adenocarcinoma were perfusedover SAC, and pre- and postperfusion samples were assayedfor C1q-binding activity by radioimmunoassay. As shown in Chart1, C1q-binding levels in postperfusion samples declined at first,then increased above pretreatment values. The initial decreasein C1q-binding levels may represent a dilutional effect from smallamounts of residual 0.15 M NaCI in the filter. In contrast to the
736 CANCER RESEARCH VOL. 44
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
observed increases in C1q binding, canine IgG concentrations inpostperfusion samples were uniformly reduced below preperfu-
sion levels, as were postperfusion levels of IgM in sera perfusedover SAC (see legend to Chart 1).
To determine if protein A on the SAC might be important inthe generation of increased C1q binding, sera from normal dogsand dogs with mammary adenocarcinoma were perfused overSAC or protein A-deficient SAW, as described in "Materials andMethods." Samples emerging from the SAC columns showed an
increase above preperfusion C1q-binding levels, whereas ef
fluent samples from the SAW columns showed minimal changesfrom pretreatment levels (Chart 2). As with the above studies,the initial reductions in C1q binding may partially reflect dilutionaleffects from a small amount of residual fluid in the filters containing SAC or SAW.
Sera from normal dogs and dogs with mammary adenocarcinoma were passaged over PACC, as described in "Materials andMethods." In 8 of 8 tumor-bearing sera, and 6 of 8 normal sera,
there was an increase in C1q binding in effluent sera (Table 1).
Immunoglobulin Oligomers in Serum after Perfusion over Protein A
In contrast to perfusion over SAC, there were no significantchanges in IgG, IgM, and IgA levels in sera perfused over PACC(data not shown).
Partial Characterization of Complexes Generated in Seraafter Perfusion over PACC. Since protein A has been shown tobind canine IgM and IgA (2, 3), we tested for these components,as well as for IgG and C3, in the complexes in sera perfusedover PACC. For this purpose, pre- and postperfusion sera wereincubated with C1q-coated tubes in order to bind the complexes.
As a control, sera were perfused over canine albumin rather thanover PACC, and were similarly incubated with C1q-coated tubes.The tubes were washed and excess 125l-F(ab')2fragments spe
cific for canine IgG, IgM, IgA, or C3 were added, and boundradioactivity was measured. Postperfusion binding levels of anti-IgG and anti-IgM in sera perfused over PACC increased approx
imately 20% above preperfusion values, but there were nosignificant changes in anti-lgA or anti-C3 binding. Results weresimilar for tumor-bearing and normal sera, and similar increasesin anti-lgG-binding activity were detected in postperfusion serafrom an additional tumor-bearing and a normal dog. In contrast,anti-lgG-binding activity was reduced by 36 to 44% in serum
perfused over immobilized canine albumin, while binding of othercomponents was essentially unchanged.
Normal or tumor-bearing human sera were similarly passagedover PACC, incubated with C1q-coated tubes, and tested forbinding of 125l-F(ab')2 fragments specific for human IgG, IgM,
20 40 60 80 100
Effluent Volume (ml)
Chart 1. One hundred ml of sera from 6 dogs with mammary carcinoma ¡A)orfrom 5 normal dogs (B) were perfused over SAC. In both sera, Clq-binding levelsinitiallydecreased,then increasedabove pretreatment levelsafter perfusion.CanineIgG levels in postperfusion sera also declined at first, then increasedto near baseline values in both normal and tumor-bearing sera. Points, means of 5 or 6determinations; oars, S.D. Percentageof reductions in IgM levels in 3 normal and3 tumor-bearingcanine sera were comparableto IgG. Pretreatmentvalues for C1qbinding in normal sera ranged from 14 to 40 ACG equivalents/ml(21 ±9.6) and intumor-bearing sera from 37 to 87 ACG equivalents/ml (57 ±17.3).
12505'S^
ä200s--1£
e150E
i'«s=•
Susoe°-
0A-.**~*'*^,
/ ••-»•^7"i
i i i i75020015010050nB—f~~^v^*
~/V"^"'
-!11¡Pre
10 20 30 40 50 Pre 10 20 30 40 50
Effluent Samples(I ml) From SAC ( 1or SAW ( )Chart 2. One hundred ml of sera from tumor-bearing (A) or normal (B) dogs
were simultaneouslyperfused by dual channel tubing over SAC- and SAW-contain-ing filters arranged in parallel. An increase in C1q-binding levels of sera emergingfrom SAC filters was noted, while effluent from SAW displayed no significantchange from pretreatment values. Similar results were obtained in 3 additionalexperiments.
Table1Solid-phaseC1q binding before and after perfusion of canine sera over PACC
Ten ml of normalor tumor-bearingsera were passagedover PACC and pre- and posttreatment sera were analyzedforsolid phase Clq binding. C1q binding is expressed as ng of anti-canine IgG bound.
NormalseraPreperfusion50.9"
(50.0-51. 8)"
43.6 (35.4-47.4)58.9 (57.3-60.6)50.2 (43.6-53.0)41.7 (39.7-43.7)26.9 (25.4-28.5)29.6 (27.9-31.3)36.0 (33.1-39.4)Postperfusion65.3
(62.6-68.8)63.2 (59.0-67.4)63.3 (60.0-66.7)52.1 (47.9-55.2)55.8 (55.0-56.7)33.8 (33.7-33.9)30.8(30.1-31.6)75.9 (70.9-80.0)A14.4
19.64.41.9
14.16.91.2
39.9Tumor-bearing
seraPreperfusion60.5
(57.2-62.8)49.7(45.1-52.2)50.0(49.5-51.0)42.2 (36.8-46.5)28.6 (26.8-30.5)32 (32.0-32.1)31.4(31.1-31.8)52.0 (50.3-54.7)Postperfusion73.0
(66.9-81.1)69.6 (58.4-80.0)57.0 (54.8-59.2)56.7 (53.9-59.5)34.8 (32.6-37.1)36.8 (34.5-39.1)53.9 (52.7-55.1)
135.0(126.0-143.2)A12.5
19.97.0
14.56.24.8
22.583.0
42.2 ±11.0° 55.0 ±15.0 43.3 ±11.6p < 0.025° p < 0.05°_64.5 ±31.5
* Values represent the average of duplicate or triplicate determinations.6 Numbers in parentheses, range of values from duplicate or triplicate determinations.c Mean ±S.D.'' The increasesin post- compared to respective preperfusion levels were statistically significant.
FEBRUARY 1984 737
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

J.Balint, Jr., étal.
Table 2Components ol C1q-binding complexes in human serum after passage over PACC or human albumin
Ten ml of normal or tumor-bearing serum were passaged over PACC or human albumin immobilized in collodion charcoal. Pre- and posttreatment samples weretested for C1q binding using F(ab').. fragments specific for human immunoglobulins.
Anti-F(ab'), bound (ng)
Anti-IgGImmobilized
proteinNormal
serumProtein AProtein AProteinAHuman
albuminTumorserum
Protein AProtein AProtein AProteinAHuman
albuminSampleno.1
23412
345Preperfusion93.7
±1.2a-"96.4 ±1.6"
111.6±9.513.9±1.541
.8 ±2.4"78.2 ±3.1°59.9 ±2.3"19.4±0.7"44.8
±0.7Postperfu-ston100.4
±3.0118.5 ±2.3114.7±7.210.8±1.199.2
±4.195.6 ±7.478.1 ±2.328.1 ±0.645.1
±0.7AntMgMPreperfusion144.4
+ 10.0122.5± 5.0153.2±5.9144.8
±5.391
.9 ± 4.8"
145.7± 5.2110.0± 3.283.6 ±2.8"136.5±
2.5Postperfu-sion146.8
±2.6121.0 ±7.8142.8 ±0.8142.5±2.7147.4
±5.0137.4 ±2.8104.7 ±3.0108.0±2.9139.4
±2.5AntMgAPreperfusion90.0
± 3.1"112.3± 3.6"
81.5 ±6.482.3±5.0169.3
±24.0122.1 ± 1.7"
109.4± 2.1241.5 ±10.4278.0
± 3.4Postperfusion114.4+
2.8230.5 ±15.279.0 ±5.382.6
±4.9169.1
± 5.2185.9± 8.194.5 ± 2.3
268.6 ±4.4274.5± 9.7Anti-C3Preperfu
sion18.3
±2.0"13.9 ±3.5"13.6±0.7*34.3
±1.316.9
±2.9"
23.0 ±2.424.4 ±3.6"
49.5 ±2.416.3±3.3Postperfuston28.6
± 1.2253.4+15.2
17.3+1.336.9±2.157.5+
2.129.5 ± 2.829.3 ± 3.155.8 ±4.418.0
± 1.7
o
MEo.
i•a
" Mean ±S.D. of triplicate determinations." Postperfusion values were significantly increased above pretreatment levels (p = <0.05).
IgA, and C3. Results shown in Table 2 demonstrate (a) increasedanti-IgG binding in effluents of 2 of 3 normal sera, and of 4 of 4 'tumor-bearing sera; (b) increased anti-lgA binding in effluentsfrom 2 of 3 normal sera, and from 1 of 4 tumor-bearing sera; (c)increased anti-C3 binding in all normal sera, and in 2 of 4 tumor-bearing sera tested; and (d) increased anti-IgM binding in 2 of 4tumor-bearing sera. In contrast, there were no significant increases in anti-IgG, anti-lgA, anti-IgM, or anti-C3 binding innormal or in tumor-bearing sera perfused over immobilized hu
man albumin (Table 2).To determine the size characteristics of the C1q-binding IgG
generated in sera after perfusion over PACC, pre- and postperfusion effluent samples from normal or tumor-bearing sera were
analyzed by sucrose density gradient centrifugaron, and fractions were assayed for C1q-binding IgG. Both sera showedincreased levels of Clq-binding IgG in fractions greater than 7S,compared to corresponding fractions of preperfusion sera (Chart3). Similar increases in C1q binding in these gradient fractionswere noted when F(ab')2 fragments specific for y chains of
canine IgG were used to detect IgG bound to C1q.The high-molecular-weight IgG complexes in postperfusion
canine sera were then isolated by fractionation on G-200 columnchromatography. The void volume peak fractions were rechro-matographed over an immunoadsorbent containing y chain-specific rabbit anti-canine IgG, as described in "Materials andMethods." The retained protein was eluted, neutralized, concen
trated, and reduced before analysis by SDS-PAGE. PAGE pro
files stained with Coomassie blue revealed distinct bands withmolecular weights of 50,000 and 25,000 comigrating with y andlight chains of canine IgG (Fig 1). A lightly stained M, 75,000band comigrating with the u.immunoglobulin chain of canine IgMwas also observed, and probably represents canine IgM presentin the complex isolate. Ten ^9 of the isolated high-molecular-
weight IgG complexes examined in double diffusion studiesshowed a single precipitin band with goat anti-whole canineserum which formed a line of identity with y chain-specific rabbitanti-canine IgG (data not shown). These studies indicated thathigh-molecular-weight complexes containing canine IgG and IgMwere formed during perfusion of sera over PACC.
Release of Protein A form PACC by Serum and SerumComponents. Although high-molecular-weight C1q-binding IgGwas formed during perfusion of serum over PACC, PAGE anal-
's B
T
itst
7S
123456789 10 123456789 10
Bottom Top Bottom Top
Gradient Fraction Numbet
Chart 3. Serum samples from 2 normal W and 5 tumor-bearing (B) dogs beforeand after perfusion over PACC were layered on 10 to 40% sucrose densitygradients and ultracentrifuged; each gradient fraction was assayed for Clq-bindingIgG. The results are expressed as the mean change in Clq-binding IgG in postperfusion gradient fractions compared to corresponding gradient fractions of preperfusion serum; bars. S.D. Comparable increases in C1q-binding IgG were noted inFractions 1 through 5 (>19S) for normal and tumor-bearing sera. However, unlikenormal sera, tumor-bearing sera showed less C1q-binding activity in Fractions 6through 10, compared to respective preperfusion levels. Pretreatment values forradioactivity bound to C1q were as follows: normal sera ranged from 8,317 to22,943 cpm in Fractions 1 to 3, and from 25,517 to 40,223 cpm in Fractions 4 to10; tumor-bearing sera ranged from 13,707 to 22,943 cpm in Fractions 1 to 3, andfrom 23,441 to 40,079 cpm in Fractions 4 to 10.
ysis of eluates failed to reveal a band that could be unequivocallyidentified as protein A, since authentic protein A migrates to aposition nearly coincident with canine y chains. Therefore, release of protein A into serum during perfusion was studied, usingtrace-labeled 125l-protein A in the PACC as a marker. The resultsin Chart 4 show that 125l-protein A was released from PACC by
perfusion of sera as well as from solutions of albumin or IgGfrom 3 different species. In addition, the elution was not specificfor protein A, since the same solutions also released 125l-labeled
canine albumin that had been immobilized in collodion charcoalin a fashion similar to that for protein A. In this model system,the amount of protein A released from 1 mg of immobilizedprotein A after perfusion of 10 ml of serum ranged from 2 to 16
MQ.In further studies, canine serum was perfused over PACC
containing 125l-labeled protein A as a marker. The postperfusionserum was treated with 5% PEG to precipitate any 125l-protein
A-IgG complexes and then analyzed by PAGE. A radioactive
738 CANCER RESEARCH VOL. 44
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

Immunoglobulin Oligomers in Serum after Perfusion over Protein A
68K
50K
25K
1Fig. 1. Coomassie blue-stained PAGE profile of isolated high-molecular-weight
postperfusion canine IgG is shown. The high-molecular-weight IgG was isolated bySephadex G-200 and affinity chromatography, using 7 chain-specific anti-canineIgG. Note the distinct y and light immunoglobulin polypeptide chains in the isolate(Lane 3). A lightly stained polypeptide comigrating with the \i immunoglobulin chainis also stained. Marker proteins are: BSA (Lane 1), canine IgG (Lane 4), and canineIgM (Lane 2).
peak corresponding to the protein A marker was observed. Somelower-molecular-weight radioactive protein A species were alsonoted which might represent 125l-protein A fragments (Chart 5).
In additional studies, normal canine sera perfused over PACCwere treated with 5% PEG to precipitate protein A-IgG com
plexes. The solubilized precipitate was fractionated on SephadexG-100 under dissociating conditions, and the included fraction
was precipitated with (NH„)2SO4to remove IgG, as described in"Materials and Methods." The protein A-containing supernatant
was passaged over cyanogen bromide-activated Sepharose 4B
to which human IgG was coupled. The bound material waseluted and concentrated as described in "Materials and Methods." The presence of protein A in the eluate was suggested by
double-diffusion studies, which showed a precipitin band with
normal human serum but not with normal chicken serum, whichdoes not bind to protein A (data not shown) (15).
Identification of Protein A in High-Molecular-Weight Complexes. Postperfusion normal canine sera containing eluted 125I-
protein A were fractionated on sucrose density gradients andthe profiles were compared with that of purified free protein A.
As shown in Chart 6, radiolabeled protein A in the postperfusionsamples were distributed in high-molecular-weight fractions of
the gradient compared to free protein A. Parallel sucrose densitygradient fractions of postperfusion sera showed increased C1q-
binding IgG compared to pretreatment sera. This suggested thatprotein A which eluted from PACC into postperfusion sera wasbound in complexes with immunoglobulins, and may have contributed to the increased C1q binding observed in postperfusionsucrose density gradient fractions.
To determine if 131l-protein A eluted from PACC after serum
perfusion could bind to C1q-coated tubes, the following experiment was carried out. Ten ml of normal or tumor-bearing canine
serum were perfused over PACC containing 1 mg of unlabeledprotein A, together with 131l-protein A, followed by 200 ml ofPBS. The effluents (200 ml) containing eluted 131l-protein A were
concentrated to a volume of 10 ml, and 0.5-ml aliquots wereincubated with C1q-coated tubes. Free 131l-protein A was usedas a control. Effluent serum samples containing 131l-protein A
demonstrated approximately 8-fold greater binding to C1q-coated tubes compared to free 131l-protein A. Binding of eluted125l-protein A to C1q-coated tubes represented 34 and 37%
(mean of triplicate samples) of total added radioactivity fromnormal and tumor-bearing effluents, respectively, compared to4.3% for free protein A. Hence, some of the protein A elutedfrom PACC after serum perfusion appeared to acquire C1q-
binding properties and was in a form probably complexed toimmunoglobulins.
In further studies, 50 ml of normal or tumor-bearing caninesera were perfused through columns containing 125l-protein A or125l-labeledcanine albumin, together with unlabeled protein A or
canine albumin, respectively, immobilized in collodion charcoal.The effluent fractions containing the peaks of the radioactivityreleased from the charcoal were pooled. To 10-ml aliquots of
these peaks, an equal volume of saturated (NH4)2SO4 or 5%PEG was added. Results demonstrate that 70 to 80% of thereleased 125l-protein A was precipitatale with (NH4)2SO4and PEG,
compared to only 5 to 10% of free protein A. In contrast, lessthan 10% of released albumin was (NhU^SCu or PEG precipitatale. These findings suggest that protein A but not albuminemerges in postperfusion effluent complexed with immunoglobulins.
Molecular Weight of Protein A-containing Complexes. Todetermine if IgG from canine serum and staphylococcal proteinA were bound together in C1q-binding complexes present in
postperfusion serum, the following experiment was carried out:10 ml of normal canine serum containing 131l-labeledcanine IgGwere perfused over PACC containing 125l-protein A. The effluent
was concentrated and then incubated with C1q-coated tubes.
Based on the specific activities of the labeled protein A (PA) andIgG, the empirical formula of C1q-binding complexes was cal
culated to be close to lgG2PA (Table 3). The approximate molecular weight of the protein A-containing complexes ranged from
600,000 to 2,000,000 by chromatography over Sepharose 6B(Chart 7). Hence, it appears that in addition to IgG and proteinA, other molecules such as IgA and C3 may be present in thecomplexes to give a molecular weight greater than 700,000. Inthe case of human serum, in addition to the high-molecular-
weight protein A peak corresponding to the complexes found incanine serum, a second protein A peak appeared close to themolecular weight of IgG. When this smaller peak was rechro-
FEBRUARY 1984 739
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

J.Balint, Jr., étal.
350
- 300
U
I 250
£ 200e
l ISOÌ
20 40 60 80 100 120140160180200
(ml)
600
500
400
300
200
S 10°
0
lE
l
Humin IgG
Chicken IgG
CanineIgG
f—Wash*] Flux
o | 20 40 60 80 100120140 160 180200IM n
(ml)-Flux-
100
i 6003;S 500
£ 400
300
200lS 100
HumanalbuminChickenalbuminCaninealbumin
20 40 60 80 100 120 140160180200
(ml)S S
20 40 60 80 100120140160180200
Flux-(ml)
-Flux-
Chart 4. PACC was loaded into a Lexan chamber and washed with 3000 ml of 0.15 u NaCI solution. Serum or solutions of purified albumin or IgG from variousspecies were then perfused through the columns at 2 ml/min, and radioactivity in the effluent was measured. Perfusion with (A) normal serum, (8) immunoglobulinsolutions, or (C) albumin solutions obtained from humans, dogs, or chickens resulted in release of radioactivity from PACC. The release effect by normal sera was notselective for immobilized protein A, since similar leaching of immobilized canine albumin was noted after perfusion with sera from the same 3 species (D). WHS, normalhuman serum; NCS, normal chicken serum; NOS, normal dog serum.
matographed on Sephadex G-200, it still corresponded to a
molecular weight of 160,000.
DISCUSSION
In previous studies, we showed that extracorporeal perfusionof plasma from hosts bearing breast adenocarcinoma over SACor PACC, resulted in acute tumoricidal responses and objectivetumor regressions in both dogs and humans (27-29). In our
initial studies in humans, we noted that these tumoricidal effectsand tumor regressions were also induced by plasma which waspassaged over PACC off-line and administered i.v. Hence, wedeveloped an in vitro system, similar to the off-line perfusionsystem used in our patients, in which sera from normal or tumor-
bearing hosts were passaged over SAC or PACC. The effluentsera from these systems were collected and analyzed by physi-
cochemical and immunochemical methods. The original perfusionsystem which we used in human studies (29, 30) contained alarge internal fluid volume and additional attachments, making itcumbersome for study of products emerging in sera passagedover PACC. In order to conveniently test undiluted samples in
GammiChains
lSfA
LightChains
DyeFront
1 1
10
'
JT 2
20 40 60 80mm ofGel
Chart 5. PAGE profile of effluent of normal canine sera perfused over PACCcontaining 125l-protein A. Effluent was incubated with 5% PEG, and resulting
precipitate was dissolved in PBS before analysis. A distinct peak corresponding toprotein A as well as other unidentified components, perhaps corresponding tosmaller fragments of protein A, are noted. SpA, staphylococcal protein A.
large numbers and evaluate compositional changes in the effluent, we designed a model perfusion system using quantitiesof perfusate and immobilized protein A similar to those utilizedin clinical studies (29, 30).
740 CANCER RESEARCH VOL. 44
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

Immunoglobulin Oligomers in Serum after Perfusion over Protein A
1000r -i2500
- 2000
I;îl
- 1500
- 1000 1!
Bottom
7 9
Fraction Number
15
TopChart 6. Postperfusion normal canine sera containing eluted 12Sl-proteinA was
fractionated on 10 to 40% sucrose density gradients. Radiolabeled protein A wasdistributed in high-molecular-weight fractions. Increased Clq-binding IgG in post-perfusion was also noted in parallel gradient fractions, suggesting that protein Awhich eluted from PACC in postperfusion sera may have contributed to theincreased C1q binding.
TabledRatio of IgG to protein A molecules in postperfusion Clq-binding complexes
Refer to text for experimental details.
Sample1
2Protein
A bound(ng)5.25(12.38)"
7.41 (17.64)8.62(20.52)3.4
(8.1)3.6 (8.5)IgG
bound(ng)26.83
(16.77)52.24 (32.65)
101.70(63.56)32.38
(20.24)41.43(25.59)Molar
ratio IgG/pro-teinA2.1
±0.73"
2.74 ±0.25C
8 Numbers in parentheses, nmol x 10"" Mean ±S.D. of 3 determinations.c Average of 2 determinations.
The present investigation focuses on the identification andanalysis of products which are formed in sera of normal andtumor-bearing dogs after perfusion over SAC or PACC. Perfusionof canine sera over SAC resulted in an increase of Clq-bindingIgG in effluent samples, whereas perfusion of canine sera overprotein A-deficient SAW did not. Hence, the presence of proteinA on heat-killed and formalin-stabilized SAC appeared to berequired to produce the increased C1q-binding activity. This wassupported by the observation that similar increases in Clq-binding IgG were noted in effluent canine serum after perfusionover purified protein A. The increased C1q-binding IgG emergingfrom SAC columns could not be ascribed to nonspecific increases in IgG in effluent samples, since the concentration ofthis major immunoglobulin class was decreased in the samepostperfusion samples which showed increased levels of C1q-
M M tOO IZO 140
ElutionVolume(ml)
Chart 7. Sepharose CL-6B chromatography of human (A) or canine (B) serumcontaining 131l-labeledhuman or canine IgG after passage over PACC containing 1mg of unlabeled protein A, together with 125l-protein A. Human serum was similarlypassaged over unlabeled human albumin with 125l-labeled human serum albumin
immobilized in collodion charcoal (C). In human and canine serum passaged overPACC, eluted protein A appears in a high-molecular-weight peak, suggesting thatit is present in a form complexed to other macromolecules (A and B). In contrast,the human serum effluent from the albumin collodion charcoal did not demonstrateeluted albumin in a high-molecular-weight peak (C). Free staphylococcal protein A
(SpA) emerged at an elution volume corresponding to a molecular weight of-150,000 which is greater than its true value of 42,000, and is consistent with itsbehavior as a rod-like structure, rather than as a typical globular protein (15).
binding IgG. However, a decrease in IgG levels did not appearnecessary for the formation of C1q-binding IgG, since effluent
sera from PACC column showed no significant reductions incanine immunoglobulins.
Pre- and postperfusion sera from normal or tumor-bearingdogs were further analyzed by sucrose density gradient ultra-centifugation. Increased levels of C1q-binding IgG sedimenting
in fractions greater than 7S were noted in postperfusion serafrom both groups. This increase was not due to Fc binding ofthe anti-canine IgG antibody indicator to any protein A in thecomplexes, since F(ab')2 fragments of anti-canine IgG also de
tected elevated C1q binding in similar postperfusion gradient
FEBRUARY 1984 741
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

J. Balint, Jr., et al.
fractions. Moreover, PAGE analysis of the high-molecular IgGisolated by G-200 fractionation and affinity chromatography re
vealed predominantly 7 and light immunoglobulin chains of canine IgG. Immunodiffusion studies confirmed the presence ofcanine IgG in the isolate, since a precipitin line formed betweenthe isolate and anti-canine IgG y chain antisera. Further analysisof the Clq-binding complexes using heterologous and F(ab')2
fragments to human or canine IgG, IgM, IgA, or C3 showedincreased binding of anti-IgG and anti-IgM in canine complexesand anti-IgG, IgA, IgM, and C3 in human complexes. Hence, the
complexes formed in serum after perfusion over PACC appearedto be of high molecular weight and consisted predominantly ofIgG, together with other serum components.
Since we detected an increase in C1q-binding activity in sera
after perfusion, we considered the possibility that protein Areleased from PACC had combined with canine immunoglobulinsin sera to produce high-molecular-weight Clq-binding com
plexes. Using radiolabeled protein A immobilized on PACC withunlabeled protein A, we found that 2 to 16 ng of protein A weredesorbed from PACC during serum perfusion. Moreover, proteinA was identified in serum effluent emerging from the PACC bythe following additional studies: (a) PAGE analysis of postperfu-sion effluent after Sepharose G-100 separation showed a radio
active peak migrating at the protein A marker; and (b) postperfusion complexes isolated by gel filtration and affinity chromatography and then dissociated under acid conditions showed aprecipitin line with normal human but not with chicken serum.
The latter experiments showed that protein A was releasedfrom PACC and present in serum perfused over PACC. Furtherstudies demonstrated that the released protein A was present inthe high-molecular-weight complexes since, (a) sucrose densitygradient analysis of postperfusion sera showed that 125l-protein
A released from PACC was distributed in high-molecular-weight
fractions beyond the 7S marker and was distinct from free proteinA; (b) radiolabeled protein A emerging in effluent samples boundto C1q-coated tubes to a greater extent than did free protein A;(c) the increase of released 125l-protein A from PACÃ’ in high-
molecular-weight sucrose density gradient fractions corresponded to augmentation in C1q-binding IgG in parallel gradientfractions; and (d) a larger percentage of 125l-protein A eluted
from PACC and emerging in postperfusion effluent was precipi-table with (NH4)2SO4and PEG, compared to free 125l-protein A.
In prior studies, using gross techniques such as pyrogen andLimulus testing of effluent samples emerging from SAC or PACC,leaching of protein A was not detected (27-29). Moreover, we
did not detect release of immobilized protein A, probably due tothe low mean specific activity of labeled protein A on the collodioncharcoal or SAC. The present model system, using a high meanspecific activity of radiolabeled protein A immobilized in PACCenabled us to detect leaching. Indeed, release of protein A fromheat-killed and formalin-fixed SAC and of proteins immobilized
on collodion membranes has been previously observed by others(7, 21, 22). The present studies show that release of protein Afrom PACC occurred not only with serum, but also with solutionsof purified serum proteins (Chart 5). Furthermore, this phenomenon was not specific for protein A, since 125l-labeled canine
albumin immobilized in collodion charcoal was also eluted bysera from 3 species. These results suggest that solubilization ofimmobilized proteins by serum or its components may play animportant role in the release of proteins from collodion charcoal.
Additional studies are under way to further dissect the mechanism of this effect.
Using 125Iprotein A in the PACC and 131l-lgGadded to serum
perfused over it, we showed that the empirical formula of theC1q-binding complexes corresponded to [lgG2PA]. With the useof molecular weight standards, the protein A-containing com
plexes appeared between M, 680,000 and 2,000,000. Previousstudies have shown that rabbit IgG and protein A form complexeswith same empirical formula, but with a molecular formula corresponding to the dimeric structure [(lgG)2PA]2 with a molecularweight of approximately 700,000. These complexes behavedlike IgM in being relatively efficient in binding and activating C1(16, 17). The complexes we detected may, in fact, be predominantly dimeric, and therefore correspond to the previously described rabbit IgM-like complexes. The higher molecular weight
of the former may be due to the presence of components otherthan IgG and protein A in these complexes, such as IgG, IgM,and C3, as we demonstrated with the use of monospecific F(ab')2
fragments (Table 2). We are currently studying this possibility.Having isolated and partially characterized complexes that
contain protein A, IgG, and other serum components in effluentfrom PACC, we are currently testing the activity of these complexes in vitro and in vivo, prepared by adding protein A to serumor to monomeric IgG. We have shown that these model complexes may activate Fc-bearing leukocytes and complement
system to generate oxidant and anaphylatoxin activity, respectively, and may have tumoricidal activity in experimental animals.We are currently attempting to prepare these complexes on apreparative scale for testing in vivo. The basis for the selectivityof the tumoricidal effects after infusion may involve their deposition in tumor microcirculation, recruitment of inflammatory cells,and local activation of mediators that may be directly or indirectlytumoricidal.
These studies show that perfusion of normal or tumor-bearing
sera over SAC or PACC results in the appearance of increasedC1q-binding activity. The high-molecular-weight material con
sists predominantly of IgG and protein A. After serum or serumcomponent perfusion, protein A desorbs from PACC, appears inthe postperfusion (NH^SCv and PEG-precipitable fractions,binds to solid-phase C1q, and coincides with increased Clq-binding IgG in heavy-sedimenting sucrose density gradient frac
tions. Finally, results to be reported elsewhere suggest thatthese protein A-IgG complexes play a significant role in the
genesis of the antitumor effects and toxicity previously observedin dogs and humans after receiving plasma perfused over PACCor SAC (27-30).
ACKNOWLEDGMENTS
The authors wish to thank Dr. Antonio Gotto for his support, patience, andmany valuable comments. We are also indebted to Yolanda Olgine and SharonEllsworth for their secretarial assistance.
REFERENCES
1. Balint, J. P., Jr. Immune complexes with antiglobulin activity in sera of Moloneysarcoma bearing rats. Clin. Exp. Immunol., 48: 70-78,1982.
2. Balint, J. P., Jr., Ikeda, Y., Nagai, T., and Terman, D. S. Isolation of humanand canine IgM utlizing protein A affinity chromatography. Immunol. Commun.,Õ0:553-540,1981.
3. Balint, J. P., Jr., Ikeda, Y., Nagai, T., and TemÃan, D. S. Isolation of humanIgA utilizing protein A affinity chromatography. Immunol. Commun., 11: 283-
291,1982.
742 CANCER RESEARCH VOL. 44
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

¡mmunoglobulinOligomers in Serum after Perfusion over Protein A
4. Bolton, A. E., and Hunter, W. M. The labelling of proteins to high specificradioactivities by conjugation to a 1î5l-containingacrylating agent. Biochem.
J., Õ33:529-539, 1973.5. Bradford, M. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal.Biochem., 72: 248-254,1976.
6. Forsgren, A., and Sjoquist, J. "Protein A" from Staphylococcus aureus I.
Pseudo-immune reaction with human gamma globulin. J. Immunol., 97: 822-
827,1966.7. Goldman, R., and Lenhoff, H. M. Glucose-6-phosphate dehydrogenase ad
sorbed on collodion membranes. Biochem. Biophys. Acta, 242: 514-518,
1971.8. Holohan, T. V., Phillips, T. M., Bowles, C., and Deisseroth, A. Regression of
canine mammary carcinoma after ¡mmunoadsorption therapy. Cancer Res.,42: 3663-3668,1982.
9. Kessler, S. W. Rapid isolation of antigens from cells with a staphylococcalprotein A-antibody adsorbent: parameters of the interaction of antibody-antigencomplexes with protein A. J. Immunol., 115: 1617-1624, 1975.
10. Killingsworth, L M., and Savoy, J. Manual nephelometric methods for immu-nochemical determination of immunoglobulins IgG. IgA and IgM in humanserum. Clin. Chem., 18: 335-339, 1972.
11. Kronvall, G., and Frommal, D. Definition of staphylococcal protein A reactivityfor human immunoglobulin G fragments. Immunochemistry, 7:124-127,1970.
12. Kronvall, G., and Gewürz,H. Activation and inhibition of IgG mediated complement fixation by staphylococcal protein A. Clin. Exp. Immunol., 7: 211-219,
1970.13. Kronvall, G., Seal, U. S., Finstad, J., and Williams, R. C., Jr. Phylogenetic
insight into evolution of mammalian Fc fragment of gamma globulin usingstaphylococcal protein A. J. Immunol., 104:140-147,1970.
14. Laemmli, U. K. Cleavage of structural proteins during the assembly of the headof bacteriophage T4. Nature (Lond.), 227: 680-685,1970.
15. Langone, J. J. Protein A of Staphylococcus aureus and related immunoglobulinreceptors produced by streptococci and pneumonococci. Adv. Immunol., 32:157-252,1982.
16. Langone, J. J., Boyle, M. D. P., and Borsos, T. Studies on the interactionbetween protein A and immunoglobulin G.I. Effect of protein A on the functionalactivity of IgG. J. Immunol., 727: 327-332,1978.
17. Langone, J. J., Boyle, M. D. P., and Borsos. T. Studies on the interactionbetween protein A and immunoglobulin G. II. Composition and activity ofcomplexes formed between protein A and IgG. J. Immunol., 727: 333-338,1978.
18. Lowry, O. H., Rosenbrough, N. R., Fair, A. L., and Randall, R. J. Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem., 793: 265-275,
1951.19. Mancini, G., Carbonara, A. 0., and Heremans, J. F. Immunochemical quanti
tation of antigens by single radial immunodiffusion. Immunochemistry, 2: 235-254,1965.
20. McConahey, P. J., and Dixon, F. J. A method of trace iodination of proteinsfor immunologie studies. Int. Arch. Allergy Appi. Immunol., 29:185-189,1966.
21. O'Keefe, E., and Bennett, V. Use of immunoglobulin-loaded protein A-bearing
staphylococci as a primary solid phase immunoadsorbent in radioimmunoas-say. J. Biol. Chem., 255: 561-568, 1980.
22. Schuurman, R. K. B., Gellfand, E. W., and Dosen, H. M. Polyclonal activationof human lymphocytes in vitro. I. Characterization of the lymphocyte responseto a T-cell independent B Cell mitogen. J. Immunol., 725: 820-826,1980.
23. Statenheim, G., Götze,0., Cooper, N. R. Sjoquist, J., and Müller-Eberhard,H.J. Consumption of human comlement components by complexes of IgG withprotein A of Staphylococcus aureus. Immunochemistry, 70: 501-507, 1973.
24. Stanworth, D. R., and Turner, M. W. Immunochemical analysis of immunoglobulins and their sub-units. In: D. M. Weir (ed.), Immunochemistry, pp. 6.1-6.102.Oxford: Alden Press, 1978.
25. Terman, D. S. Extracorporeal immunoadsorbents for extraction of circulatingimmune reactants. In: C. Giordano (ed.), Sorbents in Medicine, pp. 469-491.New York: Academic Press, Inc., 1980.
26. Terman, D. S., Durante, D., Buffaloe, G., and Mclntosh, R. Attenuation ofcanine nephrotoxic glomerulonephritis with an extracorporeal immunoadsorbent. Scand. J. Immunol., 6:195-202,1977.
27. Terman, D. S., Yamamoto, T., Mattioli, M., Cook, G., Tillquist, R., Henry, J.,Poser, R., and Daskal, Y. Extensive necrosis of spontaneous canine mammaryadenocarcinoma after extracorporeal perfusion over Staphy/ococcus aureusCowans 1.1.Description of acute tumoricidal response: Morphologic, histologie,immunohistochemical, immunologie and serologie findings. J. Immunol., 724:795-805,1980.
28. Terman, D. S., Yamamoto, T., Tillquist, R. L., Henry, J. F., Cook, G. L., Silvers,A., and Shearer, W. T. Tumoricidal response induced by cytosine arabinoskteafter plasma perfusion over protein A. Science (Wash. D. C.), 209: 1257-1259, 1980.
29. Terman, D. S., Young, J. B., Shearer, W. T., Ayus, C. A., Lehane, D., Mattioli,C., Espada, R., Howell, J. F., Yamamoto, T., Zaleski, H. I., Miller, L., Frommer,P., Feldman, L., Henry, J. F., Tillquist, R., Cook, G., and Daskal, Y. Preliminaryobservations of the effects on breast adenocarcinoma of plasma perfused overimmobilized protein A. N. Engl. J. Med., 305: 1195-1200, 1981.
30. Young, J. B., Ayus, J. C., Miller, L K., Divine, G. W., Frommer, J. P., Miller, R.R., and Terman, D. S. Cardiopulmonary toxicity in breast carcinoma patientsduring plasma perfusion over immobilized protein A: pathophysiology of reaction and attenuating methods. Am. J. Med., 75: 278-288,1983.
FEBRUARY 1984 743
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

1984;44:734-743. Cancer Res Joseph Balint, Jr., Yusei Ikeda, John J. Langone, et al. after Perfusion and Their Partial CharacterizationProtein A: Identification of Immunoglobulin Oligomers in Serum Tumoricidal Response Following Perfusion over Immobilized
Updated version
http://cancerres.aacrjournals.org/content/44/2/734
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/44/2/734To request permission to re-use all or part of this article, use this link
Research. on February 11, 2018. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from





























