Embed Size (px)
Citation preview

Journal of Controlled Release 65 (2000) 271–284www.elsevier.com/ locate / jconrel
Tumor vascular permeability and the EPR effect inmacromolecular therapeutics: a review
a , a a b c*H. Maeda , J. Wu , T. Sawa , Y. Matsumura , K. HoriaDepartment of Microbiology, Kumamoto University School of Medicine, Honjo 2-2-1, Kumamoto 860-0811, Japan
bNational Cancer Center Hospital, Tsukiji 5-1-1, Chuo-ku, Tokyo 104, JapancInstitute for Aging and Cancer, Tohoku University, Seiryo-cho, Aoba-ku, Sendai 980-77, Japan
Received 26 July 1999; accepted 20 September 1999
Abstract
Most solid tumors possess unique pathophysiological characteristics that are not observed in normal tissues or organs,such as extensive angiogenesis and hence hypervasculature, defective vascular architecture, impaired lymphatic drainage /recovery system, and greatly increased production of a number of permeability mediators. The phenomenon now known asthe enhanced permeability and retention (EPR) effect for lipid and macromolecular agents has been observed to be universalin solid tumors. Primarily, enhanced vascular permeability will sustain an adequate supply of nutrients and oxygen for rapidtumor growth. The EPR effect also provides a great opportunity for more selective targeting of lipid- or polymer-conjugatedanticancer drugs, such as SMANCS and PK-1, to the tumor. In the present review, the basic characteristics of the EPR effect,particularly the factors involved, are described, as well as its modulation for improving delivery of macromolecular drugs tothe tumor. Tumor-specific vascular physiology is also described. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Tumor vascular permeability; EPR effect; Macromolecular therapeutics; Permeability factors; Tumor targeting
1. Introduction lipids) [1–8]. Furthermore, even targeting of minuteparticles such as liposomes to the tumor appears to
In tumor biology, little is known about tumor- be based on this mechanism.selective or tumor-specific characteristics compared We previously reported that most solid tumorswith those of normal tissues or organs. The concept have elevated levels of vascular permeability factorsof the enhanced permeability and retention (EPR) such as bradykinin, nitric oxide (NO) [9–13], and,
2effect in solid tumors is one of the few tumor- more recently, peroxynitrite (ONOO ) [14].specific characteristics that is becoming a gold Proteinaceous vascular permeability factor (VPF)standard in antitumor drug delivery [1–6]. The EPR [15], which is identical to vascular endothelialeffect is predominantly observed for biocompatible growth factor (VEGF) [16], is also known to bemacromolecules (or macromolecular drugs and produced actively in tumor tissue; its effect is most
likely mediated indirectly by extensive production ofNO [11–13,17]. Enhanced vascular permeability is*Corresponding author. Tel.: 181-96-373-5098; fax: 181-96-also observed in granuloma and inflammatory and362-8362.
E-mail address: [email protected] (H. Maeda) infected tissues [18–21], with resultant extravasation
0168-3659/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0168-3659( 99 )00248-5
272 H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284
Table 1Factors involved in enhanced vascular permeability in solid tumors
Architectural defectivenessAngiogenesis: High vascular densityImpaired lymphatic drainageGeneration of permeability-enhancing factors
3(1) Bradykinin / hydroxyprolyl bradykinin (6) Matrix metalloproteinases?(2) Nitric oxide (NO ) (7) Other proteases (e.g. kallikrein)
2(3) Peroxynitrite (ONOO ) (8) Other cytokines (e.g. tumor necrosis factor)(4) Prostaglandins(5) VPF (VEGF)
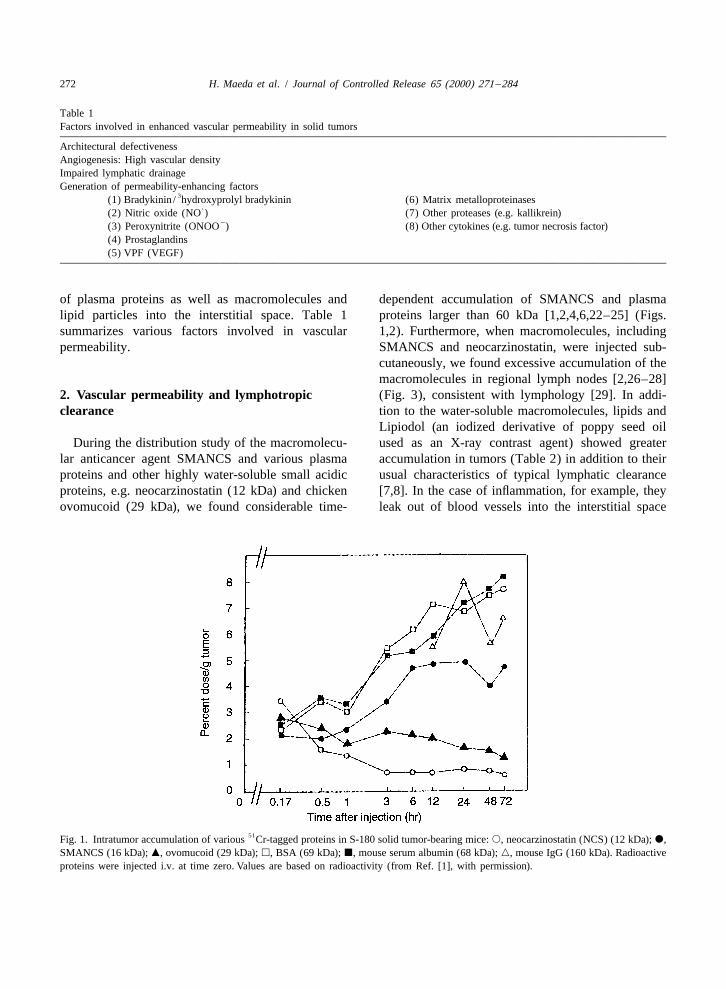
of plasma proteins as well as macromolecules and dependent accumulation of SMANCS and plasmalipid particles into the interstitial space. Table 1 proteins larger than 60 kDa [1,2,4,6,22–25] (Figs.summarizes various factors involved in vascular 1,2). Furthermore, when macromolecules, includingpermeability. SMANCS and neocarzinostatin, were injected sub-
cutaneously, we found excessive accumulation of themacromolecules in regional lymph nodes [2,26–28]
2. Vascular permeability and lymphotropic (Fig. 3), consistent with lymphology [29]. In addi-clearance tion to the water-soluble macromolecules, lipids and
Lipiodol (an iodized derivative of poppy seed oilDuring the distribution study of the macromolecu- used as an X-ray contrast agent) showed greater
lar anticancer agent SMANCS and various plasma accumulation in tumors (Table 2) in addition to theirproteins and other highly water-soluble small acidic usual characteristics of typical lymphatic clearanceproteins, e.g. neocarzinostatin (12 kDa) and chicken [7,8]. In the case of inflammation, for example, theyovomucoid (29 kDa), we found considerable time- leak out of blood vessels into the interstitial space
51Fig. 1. Intratumor accumulation of various Cr-tagged proteins in S-180 solid tumor-bearing mice: s, neocarzinostatin (NCS) (12 kDa); d,SMANCS (16 kDa); m, ovomucoid (29 kDa); h, BSA (69 kDa); j, mouse serum albumin (68 kDa); n, mouse IgG (160 kDa). Radioactiveproteins were injected i.v. at time zero. Values are based on radioactivity (from Ref. [1], with permission).

H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284 273
cancer tissue, and, hence, designing anticancer drugs[4,6,23].
3. Concept of the enhanced permeability andretention (EPR) effect of macromolecules insolid tumors
Our previous data using biocompatible plasmaproteins and synthetic polymers or their variousconjugates showed that these macromolecules areentrapped or accumulate in solid tumors and thatthey are retained there at high concentrations forprolonged periods (more than 100 h) [e.g. 1,2,4,6–Fig. 2. Relationships for molecular weight and tumor uptake and
125clearance of I-Tyr-HPMA-polymer drugs. Mice bearing S-180 8,22–24,28,30]. This phenomenon led to coining of6solid tumor received about 1.8310 cpm per injection i.v.: d, CL, the term enhanced permeability and retention (EPR)
renal clearance rate; m, AUC, area under the concentration curveeffect of macromolecules and lipids in solid tumors.for plasma, both based on 72 h; s, tumor uptake based on 24 h.More recently, the key mechanism for the EPR effectfor macromolecules in solid tumors was found to be
and are cleared via the lymphatic system. Thus, these retention, whereas low-molecular-weight substancesmacromolecular drugs and lipids are highly lymphot- were not retained but were returned to circulatingropic and possess excellent characteristics for blood by diffusion [22], as was shown earlier [1]lymphatic delivery. These characteristics may be [Fig. 4(A), (B), (C)].used to prevent lymphatic metastasis. In addition, We found that macromolecules remain at highone difference between normal inflammatory tissue levels in circulating blood; this phenomenon appliesand tumor tissue is reflected in their clearance to most plasma proteins and biocompatible syntheticvelocities: macromolecules delivered into the inter- polymers or their conjugates [1,2,4,22–24,30–32].stitial space of normal inflammatory tissue will be Here, macromolecules are defined as larger than 40cleared more rapidly than those in tumor tissue, kDa (Fig. 2). Values for the area under the con-although within a few days, and clearance from centration curve (AUC) and tumor uptake increasedtumor tissue is much slower [1,7,8,22,28]. This in parallel, whereas the rate of urinary clearance isphenomenon of vascular permeability has been inversely related to the tumor uptake. These resultsknown and utilized for a long time in clinical are consistent with reports by Seymour et al. [23]radiology as gallium scintigraphy for diagnosis of and Duncan and Sat [24], using doxorubicin conju-various solid tumors and inflammation. gated with HPMA [N-(2-hydroxypropyl)metha-
Maeda et al. realized that the theory behind the crylamide copolymer]. Takakura and Hashida alsouse of the radioactive gallium in scintigraphy for detailed organ biodistribution of macromolecules andtumor detection is based on this EPR mechanism. clearance kinetics with regard to targeting liver,Radioactive gallium binds to transferrin (90 kDa), kidney and tumors [25].and then the gallium–transferrin complex is entrap- The EPR effect for macromolecules has beenped in the tumor site by the mechanism described observed in many experimental and human solidabove, which permits visualization of the tumor tumors, such as S-180 sarcoma, Meth-A, melanomalocation and size using the g-scintillation camera. B16, Ehrlich carcinoma, and colon 38 adenocar-Tumor-selective accumulation of such radioemitting cinoma in mice; Yoshida AH136B, Walker 256macromolecules or macromolecular complexes is carcinoma, and LY tumors in rats; VX-2 carcinomathus utilized in routinely clinical settings. The EPR in rabbits; and many tumors in humans, includingeffect, however, has become a more important hepatoma, renal cancer, lung cancer, and brainprinciple for targeting macromolecular drugs to tumors (see, for example, [33–36]).
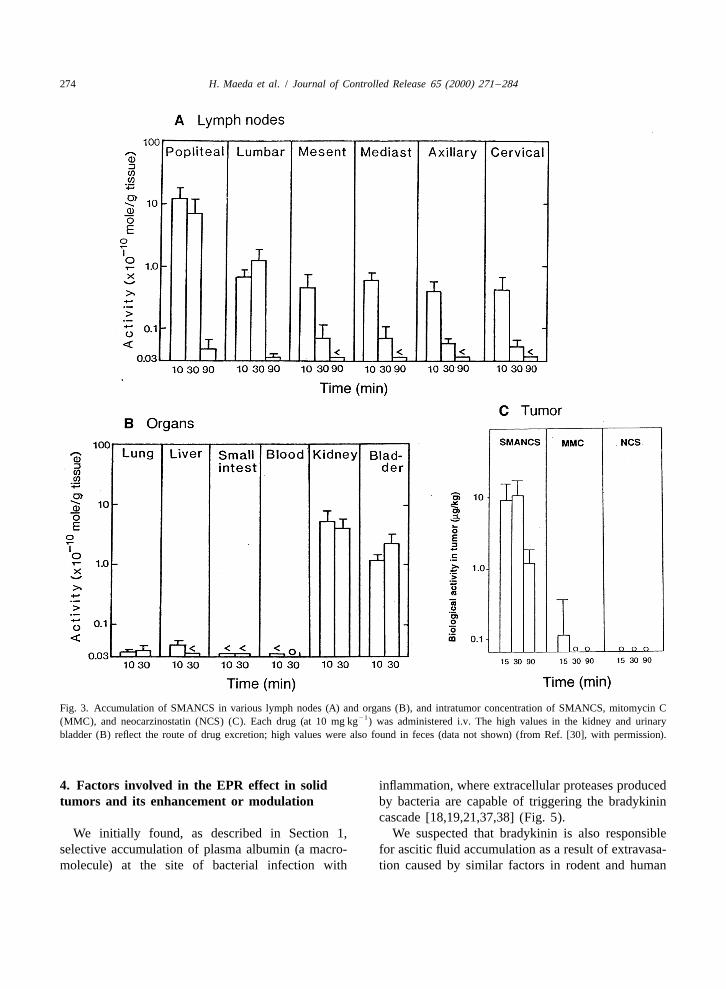
274 H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284
Fig. 3. Accumulation of SMANCS in various lymph nodes (A) and organs (B), and intratumor concentration of SMANCS, mitomycin C21(MMC), and neocarzinostatin (NCS) (C). Each drug (at 10 mg kg ) was administered i.v. The high values in the kidney and urinary
bladder (B) reflect the route of drug excretion; high values were also found in feces (data not shown) (from Ref. [30], with permission).
4. Factors involved in the EPR effect in solid inflammation, where extracellular proteases producedtumors and its enhancement or modulation by bacteria are capable of triggering the bradykinin
cascade [18,19,21,37,38] (Fig. 5).We initially found, as described in Section 1, We suspected that bradykinin is also responsible
selective accumulation of plasma albumin (a macro- for ascitic fluid accumulation as a result of extravasa-molecule) at the site of bacterial infection with tion caused by similar factors in rodent and human

H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284 275
Table 214 14Distribution of C after injection of [ C]Lipiodol (volume, 0.2 ml) via the proper hepatic artery
Sample Radioactivity23(dpm/g310 )
15 min (n53) 3 days (n52) 7 days (n51)
Tumor 444.92 97.18 37.03aLiver 200.67 9.01 4.13bLiver 29.42 3.23 0.75
Small intestine 0.48 2.13 1.60Lung 2.51 0.85 0.41Kidney 0.70 1.03 0.60Stomach 7.78 0.53 0.48
cHeart 1.34 1.72 –Large intestine 0.20 1.06 –Spleen 1.92 3.28 0.55Bladder 0.19 1.31 –Brain 0.10 0.83 –Muscle (hind leg) 0.10 0.46 –Skin (hind leg) 0.10 1.42 –Mesenteric lymph node 0.21 2.21 –Cervical lymph node 0.28 1.61 –Thymus 0.22 0.93 –Plasma 0.37 0.03 –Blood cell fraction 0.64 1.57 –Bone marrow 0.15 2.91 –Urine (fresh) 0.18 1.09 –Urine (stored) – 1.14 –Bile 38.18 2.23 –
a Non-tumorous portion adjacent to the tumor.b Non-tumorous portion distant from tumor.c –, not measured.
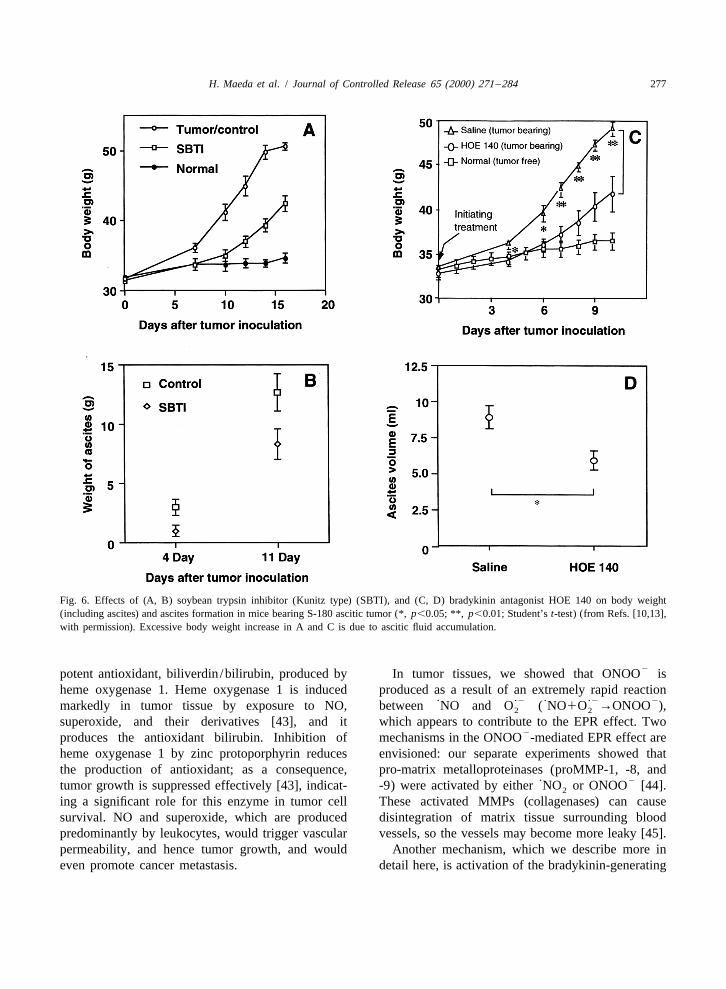
3tumors. Indeed, we identified the presence of brady- [hydroxyprolyl ]bradykinin, is generated more thankinin in ascitic fluid from a patient with gastric bradykinin in ascitic fluid of cancer patients [9,39].cancer [9]. Later, ascitic as well as pleural fluids of Other similarities between inflammation and can-carcinomatosis in cancer patients were found to cer are abundant. At a tissue level, a commoncontain high levels of bradykinin [10,39]. Kinin phenomenon in both cases is increased infiltration atformation was mediated by a serine protease ‘kallik- the disease site of leukocytes, such as neutrophils,rein,’ which specifically cleaves off nonapeptide macrophages and lymphocytes. We later found ex-(bradykinin) from high-molecular-weight kininogen cessive production of superoxide and NO in mice(Fig. 5). Kinin is one of the most potent pain- infected with influenza virus [40,41] and Salmonellainducing agents generated endogenously. A potent typhimurium [42]. This same phenomenon was alsokallikrein inhibitor, soybean trypsin inhibitor (Kunitz found in tumors and appears to contribute greatly totype), suppressed kinin generation and hence ascitic enhanced vascular permeability, because tumorsfluid accumulation [10,13,18] [Fig. 6(A), (B)]. This produce NO and other vascular permeability factors.idea was further investigated more recently by Further detailed study [13] showed involvement ofsuppression of ascitic fluid formation by use of a prostaglandins, perhaps prostaglandin E [Fig. 7(A),2
kinin receptor antagonist (HOE 140), which resulted (B)] (see [13], and unpubl. data). Because the EPRin a similar outcome [Fig. 6(C), (D)] [13]. It was effect involves NO, one can enhance the effect byalso noteworthy that an isoform of bradykinin, i.e. supplying L-arginine, which is the substrate for NO

276 H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284
Fig. 5. Schematic representation of the kinin-generating cascadeand the factors involved. MP, microbial proteases; Bac, bacterialcell wall; LPS, lipopolysaccharide; LTA, lipoteichoic acid; XII,factor XII; HF, Hageman factor; Hfa, active form of Hagemanfactor; XI, factor XI; XIa, active form of XI factor; KK, kallikrein;PK, prekallikrein; UPA, urine-type plasminogen activator; SBTI,soybean trypsin inhibitor (Kunitz type); HKG, high-molecular-weight kininogen; LKG, low-molecular-weight kininogen; KD,kallidin; LK, leukocyte-derived kinin; Gla.KK, glandular kallik-rein; BKR, bradykinin receptor; PMN, polymorphonuclear cells;BK, bradykinin; eNOS, endothelial nitric oxide synthase; iNOS,inducible nitric oxide synthase; NO, nitric oxide; Mf, macro-phages.
production by nitric oxide synthase (NOS), or byusing NO-releasing agents such as NOC-7 or nitro-prusside, which are injected by the arterial route(data not shown). Enhancement can be increased byelevating blood pressure and blood flow (Section 5).
It is intriguing that cancer cells are usuallyequipped with a means to counter the oxidativestress, such as superoxide and NO as well as their125Fig. 4. Distribution of I-labeled HPMA copolymers in micemetabolites, which are generated by infiltratingbearing S-180 solid tumors: (A) plasma; (B) liver; (C) tumor. m,leukocytes as a part of the host defense system700 kDa; d, 500 kDa; ♦, 300 kDa; j, 160 kDa; n, 85 kDa; s,
40 kDa; x, 16.2 kDa; h, 4.5 kDa (from Ref. [22]). against tumor cells. Tumor cells can generate a
H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284 277
Fig. 6. Effects of (A, B) soybean trypsin inhibitor (Kunitz type) (SBTI), and (C, D) bradykinin antagonist HOE 140 on body weight(including ascites) and ascites formation in mice bearing S-180 ascitic tumor (*, p,0.05; **, p,0.01; Student’s t-test) (from Refs. [10,13],with permission). Excessive body weight increase in A and C is due to ascitic fluid accumulation.
2potent antioxidant, biliverdin /bilirubin, produced by In tumor tissues, we showed that ONOO isheme oxygenase 1. Heme oxygenase 1 is induced produced as a result of an extremely rapid reaction
? ?2 ? ?2 2markedly in tumor tissue by exposure to NO, between NO and O ( NO1O →ONOO ),2 2
superoxide, and their derivatives [43], and it which appears to contribute to the EPR effect. Two2produces the antioxidant bilirubin. Inhibition of mechanisms in the ONOO -mediated EPR effect are
heme oxygenase 1 by zinc protoporphyrin reduces envisioned: our separate experiments showed thatthe production of antioxidant; as a consequence, pro-matrix metalloproteinases (proMMP-1, -8, and
? 2tumor growth is suppressed effectively [43], indicat- -9) were activated by either NO or ONOO [44].2
ing a significant role for this enzyme in tumor cell These activated MMPs (collagenases) can causesurvival. NO and superoxide, which are produced disintegration of matrix tissue surrounding bloodpredominantly by leukocytes, would trigger vascular vessels, so the vessels may become more leaky [45].permeability, and hence tumor growth, and would Another mechanism, which we describe more ineven promote cancer metastasis. detail here, is activation of the bradykinin-generating

278 H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284
Fig. 7. Results of the bradykinin antagonist HOE 140, the NO scavenger PTIO, and the cyclooxygenase inhibitor indomethacin, or theircombination, on the EPR effect in mice bearing S-180 solid tumors. PTIO, 2-( p-carboxyphenyl)4,4,5,5-tetramethylimidazoline-1-oxyl3-oxide (*, p,0.05; **, p,0.01; Student’s t-test) (from Ref. [13], with permission).
cascade by MMPs, perhaps involving kallikreinactivation. We previously reported that plasmin can Table 3
2trigger the kallikrein–kinin cascade in tumor tissue, Effect of protease inhibitors in ONOO -induced vascularapermeabilitywhich is induced by urinary-type plasminogen ac-
tivator produced by almost all types of solid tumor Agent Permeabilitycells [39]. Lijnen et al. [46] showed more recently (Evans Blue, mg)
2that MMPs can activate plasminogen to yield minip- Vehicle (with ONOO ) 6.24460.706lasmin. Miniplasmin would probably activate BE 16627B (2.0 mg/head i.p.) 3.71860.184*
SBTI (1.0 mg/head i.v.) 2.25160.128**Hageman factor or prekallikrein. Thus, here againOvomacroglobulin (1 mg/head i.v.) 3.46860.520*the generation of bradykinin may be mediated by
22 Background (without ONOO ) 1.45060.103ONOO via MMP activation. Consistent with this,
a 2ONOO was administered at 100 nmol / injection intradermal-we found that a bradykinin receptor antagonist (HOE2 ly in the normal healthy dorsal skin of mice; BE 16627B is an140) suppressed the ONOO -induced EPR effect in
inhibitor of MMPs; SBTI is a soybean trypsin /kallikrein inhibitor;normal skin tissue (Fig. 7) as well as in tumor tissue ovomacroglobulin is a broad spectrum protease inhibitor from(Table 3). The same effect was also observed in this chicken egg white. *, p,0.05; **, p,0.005 vs. vehicle. See textsetting after injection of kallikrein inhibitor (Kunitz- for details.

H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284 279
type soybean trypsin inhibitor) or of ovomacrog- 5. Unique blood flow in tumor: tumor-specificlobulin (a broad-spectrum protease inhibitor) (J. Wu, vascular pathophysiologyH. Maeda, et al., unpubl.).
In work other than that related to these low- In 1981, Suzuki et al. reported that blood flow inmolecular-weight permeability factors or vascular tumor tissue was quite different from that in all othermediators, Dvorak et al. [47] initially found the VPF normal tissues, in response to hypertension inducedproduced by various cultured tumor cells. This VPF by infusing angiotensin II [50]. Fig. 9 shows thisis more frequently called VEGF [16], an angio- result: tumor blood flow was selectively increased ingenesis factor. As mentioned previously, VPF/VEGF response to angiotensin II-induced hypertensionwas shown to exert its action via generation of NO. when systemic arterial blood pressure was elevatedThus, all these vascular permeability factors facilitate by intravenous administration of this agent. Bloodthe supply of nutrients and oxygen, and thus angio- flow in normal organs such as kidney, bone marrow,genesis and the development of neoplasia. Fig. 8 brain and liver remained constant regardless of theshows an overall view of these events involving elevation of blood pressure, however. Tumor tissuevascular permeability phenomena. seems to lose the capability for autoregulated homeo-
In connection with extravasation and extravascular static control of blood flow. We thus expected that bydissemination, we previously showed, using a model elevating blood pressure and blood flow in tumors byof bacterial infection, that bradykinin enhanced using angiotensin II, one could accomplish moretransvascular dissemination of bacteria (Pseudo- effective delivery of drugs for tumors, particularlymonas sp. and Vibrio sp.) from the local site macromolecular drugs [51] (Fig. 10). Also, tumor(peritoneum) of infection to the general circulation or vessels are passively dilated and the endothelialto the liver from the peritoneal compartment [48,49]. intercellular junctions are opened in the hypertensiveAlthough the role of bradykinin on cancer metastasis state. It is also believed that tumor blood vessels lackstill needs further investigation, it is plausible that both smooth muscle cells surrounding the endothelialthese vascular permeability factors would facilitate cells and angiotensin II receptors.cancer metastasis. Recently, another example of the tumor-selective
2Fig. 8. Schematic representation of the vascular permeability mediators bradykinin, prostaglandins (PGs), NO, ONOO , and MMPs inenhanced vascular permeability in solid tumors. COXs, cyclooxygenases.

280 H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284
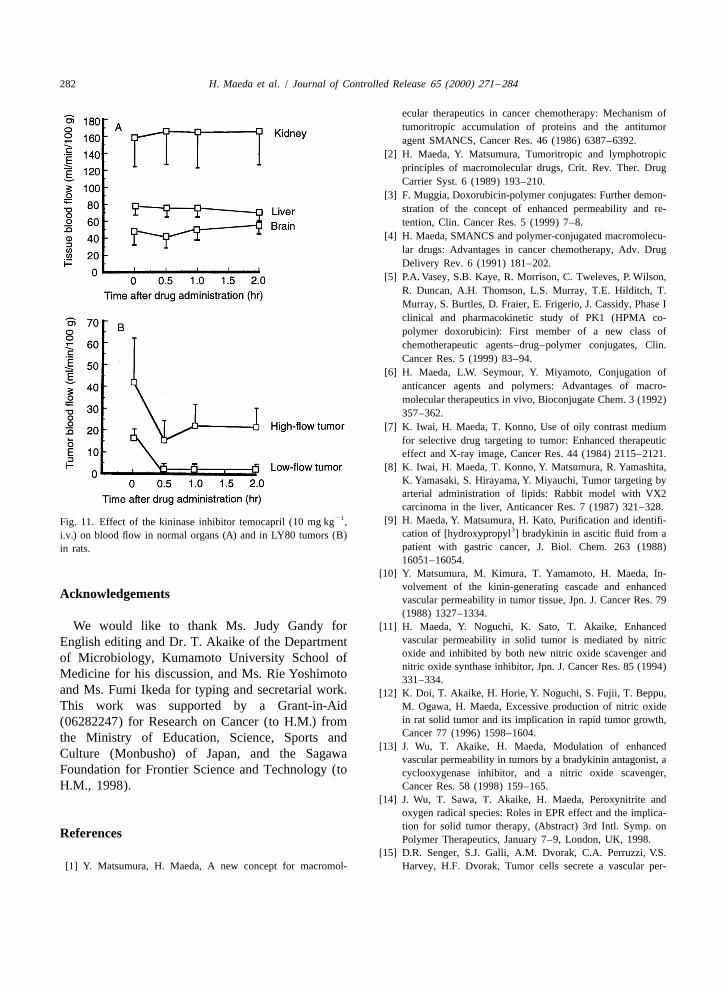
converting enzyme (ACEI) as well as of kininase (abradykinin-degrading enzyme), was injected in-
21travenously at 10 mg kg to rats bearing LY tumor,systemic arterial blood flow and blood flow ofnormal organs were little affected [Fig. 11(A)].However, tumor blood flow, as determined by thehydrogen gas clearance method, was suppressedalmost 80–90% [52] [Fig. 11(B)]. On the contrary,vascular permeability in tumors was found to in-crease significantly. A similar effect was observedafter injection of the prostaglandin I analogue2
beraprost (Dorner , Yamanouchi /Toray, Japan),which exhibits a much longer half-life in vivo thandoes prostaglandin I . Both temocapril and beraprost2
sodium increased vascular permeability to a signifi-cant extent (unpubl. data). Thus, it is speculated thatvery local tumor blood pressure became nil due tovascular dilatation as a result of bradykinin, NO, orprostaglandin I analogues, because such mediators2
and their receptors are more abundant in tumor thanin normal tissue. Blood flow in normal tissuesremained unchanged because such effectors and theirreceptors remain unchanged.
6. Modulation of vascular permeability byinhibitors of vascular mediators or antagonists
By the use of ACEIs such as enalapril or temocap-ril, which also inhibit the degradation of bradykininas described above, one can ultimately activateendothelial NOS because the level of bradykinin isincreased. Thus, the EPR effect becomes moreapparent, and such inhibitors enhance delivery ofmacromolecular drugs or components to the tumor.
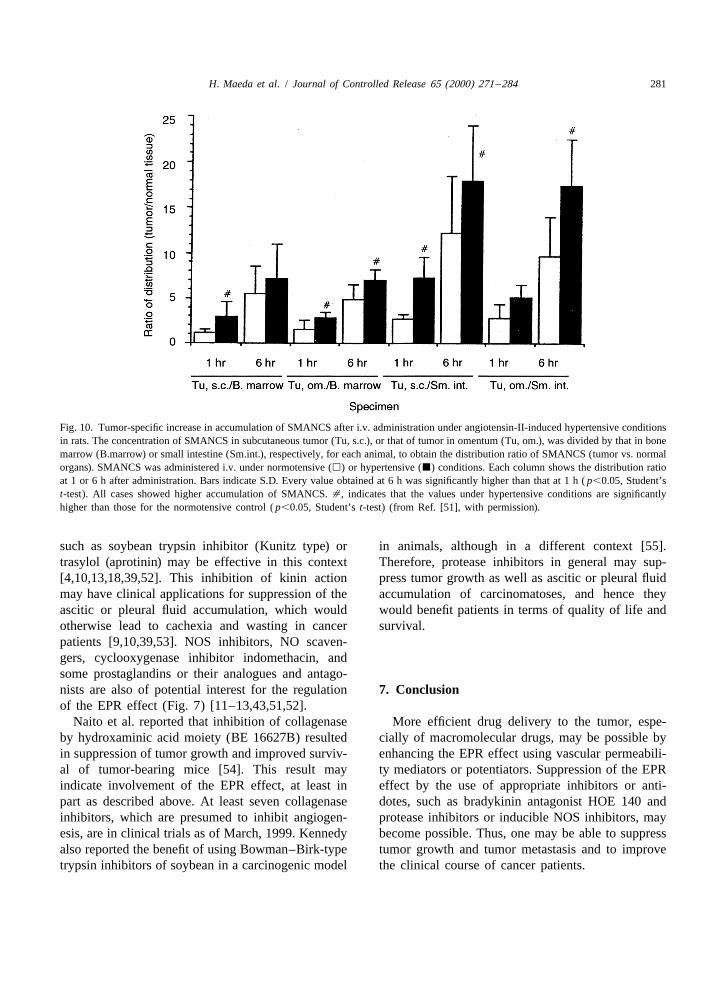
In addition, elevation of blood pressure by infus-ing angiotensin II is another way to enhance drugdelivery to the tumor by two to three times whilereducing delivery to normal organs such as bone
Fig. 9. Relationship between blood pressure and blood flow. marrow to about 60% compared with that in nor-Hypertension was obtained by infusing angiotensin II. (A) Normal
motensive states [51]. Increased differential drugorgans: (1) liver; (2) brain; (3) bone marrow; (4) subcutis. (B)concentration, i.e. in tumor vs. in blood (T/B), ofAH109A tumor implanted in rats. Note the great increase in bloodcourse helps to increase the therapeutic effect andflow-rate in tumor (B), in parallel to blood pressure (from Ref.
[50], with permission). reduce the toxicity (Fig. 10).Inhibition or suppression of the EPR effect by
suppressive effect of blood flow was demonstrated inhibitors of the kinin cascade or by a bradykininwith the use of temocapril by Hori et al. When receptor antagonist such as HOE 140 (Hoechst AG)temocapril, which is an inhibitor of angiotensin- is also possible [13,18]. Some protease inhibitors
H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284 281
Fig. 10. Tumor-specific increase in accumulation of SMANCS after i.v. administration under angiotensin-II-induced hypertensive conditionsin rats. The concentration of SMANCS in subcutaneous tumor (Tu, s.c.), or that of tumor in omentum (Tu, om.), was divided by that in bonemarrow (B.marrow) or small intestine (Sm.int.), respectively, for each animal, to obtain the distribution ratio of SMANCS (tumor vs. normalorgans). SMANCS was administered i.v. under normotensive (h) or hypertensive (j) conditions. Each column shows the distribution ratioat 1 or 6 h after administration. Bars indicate S.D. Every value obtained at 6 h was significantly higher than that at 1 h ( p,0.05, Student’st-test). All cases showed higher accumulation of SMANCS. [, indicates that the values under hypertensive conditions are significantlyhigher than those for the normotensive control ( p,0.05, Student’s t-test) (from Ref. [51], with permission).
such as soybean trypsin inhibitor (Kunitz type) or in animals, although in a different context [55].trasylol (aprotinin) may be effective in this context Therefore, protease inhibitors in general may sup-[4,10,13,18,39,52]. This inhibition of kinin action press tumor growth as well as ascitic or pleural fluidmay have clinical applications for suppression of the accumulation of carcinomatoses, and hence theyascitic or pleural fluid accumulation, which would would benefit patients in terms of quality of life andotherwise lead to cachexia and wasting in cancer survival.patients [9,10,39,53]. NOS inhibitors, NO scaven-gers, cyclooxygenase inhibitor indomethacin, andsome prostaglandins or their analogues and antago-nists are also of potential interest for the regulation 7. Conclusionof the EPR effect (Fig. 7) [11–13,43,51,52].
Naito et al. reported that inhibition of collagenase More efficient drug delivery to the tumor, espe-by hydroxaminic acid moiety (BE 16627B) resulted cially of macromolecular drugs, may be possible byin suppression of tumor growth and improved surviv- enhancing the EPR effect using vascular permeabili-al of tumor-bearing mice [54]. This result may ty mediators or potentiators. Suppression of the EPRindicate involvement of the EPR effect, at least in effect by the use of appropriate inhibitors or anti-part as described above. At least seven collagenase dotes, such as bradykinin antagonist HOE 140 andinhibitors, which are presumed to inhibit angiogen- protease inhibitors or inducible NOS inhibitors, mayesis, are in clinical trials as of March, 1999. Kennedy become possible. Thus, one may be able to suppressalso reported the benefit of using Bowman–Birk-type tumor growth and tumor metastasis and to improvetrypsin inhibitors of soybean in a carcinogenic model the clinical course of cancer patients.
282 H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284
ecular therapeutics in cancer chemotherapy: Mechanism oftumoritropic accumulation of proteins and the antitumoragent SMANCS, Cancer Res. 46 (1986) 6387–6392.
[2] H. Maeda, Y. Matsumura, Tumoritropic and lymphotropicprinciples of macromolecular drugs, Crit. Rev. Ther. DrugCarrier Syst. 6 (1989) 193–210.
[3] F. Muggia, Doxorubicin-polymer conjugates: Further demon-stration of the concept of enhanced permeability and re-tention, Clin. Cancer Res. 5 (1999) 7–8.
[4] H. Maeda, SMANCS and polymer-conjugated macromolecu-lar drugs: Advantages in cancer chemotherapy, Adv. DrugDelivery Rev. 6 (1991) 181–202.
[5] P.A. Vasey, S.B. Kaye, R. Morrison, C. Tweleves, P. Wilson,R. Duncan, A.H. Thomson, L.S. Murray, T.E. Hilditch, T.Murray, S. Burtles, D. Fraier, E. Frigerio, J. Cassidy, Phase Iclinical and pharmacokinetic study of PK1 (HPMA co-polymer doxorubicin): First member of a new class ofchemotherapeutic agents–drug–polymer conjugates, Clin.Cancer Res. 5 (1999) 83–94.
[6] H. Maeda, L.W. Seymour, Y. Miyamoto, Conjugation ofanticancer agents and polymers: Advantages of macro-molecular therapeutics in vivo, Bioconjugate Chem. 3 (1992)357–362.
[7] K. Iwai, H. Maeda, T. Konno, Use of oily contrast mediumfor selective drug targeting to tumor: Enhanced therapeuticeffect and X-ray image, Cancer Res. 44 (1984) 2115–2121.
[8] K. Iwai, H. Maeda, T. Konno, Y. Matsumura, R. Yamashita,K. Yamasaki, S. Hirayama, Y. Miyauchi, Tumor targeting byarterial administration of lipids: Rabbit model with VX2carcinoma in the liver, Anticancer Res. 7 (1987) 321–328.
21 [9] H. Maeda, Y. Matsumura, H. Kato, Purification and identifi-Fig. 11. Effect of the kininase inhibitor temocapril (10 mg kg ,3cation of [hydroxypropyl ] bradykinin in ascitic fluid from ai.v.) on blood flow in normal organs (A) and in LY80 tumors (B)
patient with gastric cancer, J. Biol. Chem. 263 (1988)in rats.16051–16054.
[10] Y. Matsumura, M. Kimura, T. Yamamoto, H. Maeda, In-volvement of the kinin-generating cascade and enhanced
Acknowledgements vascular permeability in tumor tissue, Jpn. J. Cancer Res. 79(1988) 1327–1334.
We would like to thank Ms. Judy Gandy for [11] H. Maeda, Y. Noguchi, K. Sato, T. Akaike, Enhancedvascular permeability in solid tumor is mediated by nitricEnglish editing and Dr. T. Akaike of the Departmentoxide and inhibited by both new nitric oxide scavenger andof Microbiology, Kumamoto University School ofnitric oxide synthase inhibitor, Jpn. J. Cancer Res. 85 (1994)
Medicine for his discussion, and Ms. Rie Yoshimoto 331–334.and Ms. Fumi Ikeda for typing and secretarial work. [12] K. Doi, T. Akaike, H. Horie, Y. Noguchi, S. Fujii, T. Beppu,This work was supported by a Grant-in-Aid M. Ogawa, H. Maeda, Excessive production of nitric oxide
in rat solid tumor and its implication in rapid tumor growth,(06282247) for Research on Cancer (to H.M.) fromCancer 77 (1996) 1598–1604.the Ministry of Education, Science, Sports and
[13] J. Wu, T. Akaike, H. Maeda, Modulation of enhancedCulture (Monbusho) of Japan, and the Sagawa vascular permeability in tumors by a bradykinin antagonist, aFoundation for Frontier Science and Technology (to cyclooxygenase inhibitor, and a nitric oxide scavenger,H.M., 1998). Cancer Res. 58 (1998) 159–165.
[14] J. Wu, T. Sawa, T. Akaike, H. Maeda, Peroxynitrite andoxygen radical species: Roles in EPR effect and the implica-tion for solid tumor therapy, (Abstract) 3rd Intl. Symp. on
References Polymer Therapeutics, January 7–9, London, UK, 1998.[15] D.R. Senger, S.J. Galli, A.M. Dvorak, C.A. Perruzzi, V.S.
[1] Y. Matsumura, H. Maeda, A new concept for macromol- Harvey, H.F. Dvorak, Tumor cells secrete a vascular per-

H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284 283
meability factor that promotes accumulation of ascites fluid, Conjugation of Cu,Zn-superoxide dismutase with suc-Science 219 (1983) 983–985. cinylated gelatin: Pharmacological activity and cell-lubricat-
[16] D.W. Leung, G. Cachianes, W.J. Kuang, D.V. Goeddel, N. ing function, Bioconjugate Chem. 4 (1993) 490–498.Ferrara, Vascular endothelial growth factor is a secreted [32] Y. Kojima, T. Akaike, K. Sato, H. Maeda, T. Hirano,angiogenic mitogen, Science 246 (1989) 1306–1309. Polymer conjugation to Cu,Zn-SOD and suppression of
[17] A. Papapetropoulos, G. Garcia-Cardena, J.A. Madri, W.C. hydroxyl radical generation on exposure to H O : Improved2 2Sessa, Nitric oxide production contributes to the angiogenic stability of SOD in vitro and in vivo, J. Bioactive Compat.properties of vascular endothelial growth factor in human Polymers 11 (1996) 169–190.endothelial cells, J. Clin. Invest. 100 (1997) 3131–3139. [33] T. Konno, H. Maeda, K. Iwai, S. Tashiro, S. Maki, T.
[18] H. Maeda, J. Wu, T. Okamoto, K. Maruo, T. Akaike,Morinaga, M. Mochinaga, T. Hiraoka, I. Yokoyama, Effect
Kallikrein–kinin in infection and cancer, Immunopharmacol-of arterial administration of high-molecular-weight anti-
ogy 12 (1999).cancer agent SMANCS with lipid lymphographic agent on[19] H. Maeda, Role of microbial proteases in pathogenesishepatoma: A preliminary report, Eur. J. Cancer Clin. Oncol.(Review), Microbiol. Immunol. 40 (1996) 685–699.19 (1983) 1053–1065.[20] H.I. Peterson, K.L. Apergren, Experimental studies on the
[34] T. Konno, H. Maeda, K. Iwai, S. Maki, S. Tashiro, M.uptake and retention of labelled proteins in a rat tumor, Eur.Uchida, Y. Miyauchi, Selective targeting of anti-cancer drugJ. Cancer 9 (1973) 543–547.and simultaneous image enhancement in solid tumors by[21] A. Molla, T. Yamamoto, T. Akaike, S. Miyoshi, H. Maeda,arterially administered lipid contrast medium, Cancer 54Activation of Hageman factor and prekallikrein and genera-(1984) 2367–2374.tion of kinin by various microbial proteinases, J. Biol. Chem.
[35] S. Maki, T. Konno, H. Maeda, Image enhancement in264 (1989) 10589–10594.computerized tomography for sensitive diagnosis of liver[22] Y. Noguchi, J. Wu, R. Duncan, J. Strohalm, K. Ulbrich, T.cancer and semiquantitation of tumor selective drug targetingAkaike, H. Maeda, Early phase tumor accumulation ofwith oily contrast medium, Cancer 56 (1985) 751–757.macromolecules: A great difference between the tumor vs.
[36] L.W. Seymour, S.P. Olliff, C.J. Poole, P.G. De Takats, R.normal tissue in their clearance rate, Jpn. J. Cancer Res. 89(1998) 307–314. Orme, D.R. Ferry, H. Maeda, D.J. Kerr, A novel dosage
[23] L.W. Seymour, K. Ulbrich, P.S. Steyger, M. Brereton, V. approach for evaluation of SMANCS in the treatment ofSubr, J. Strohalm, R. Duncan, Tumour tropism and anti- primary hepatocellular carcinoma, Int. J. Oncol. 12 (1998)cancer efficacy of polymer-based doxorubicin prodrugs in 1217–1223.subcutaneous murine B16F10 melanoma, Br. J. Cancer 70 [37] K. Matsumoto, T. Yamamoto, R. Kamata, H. Maeda, Patho-(1997) 636–641. genesis of serratial infection: Activation of the Hageman
[24] R. Duncan, Y.-N. Sat, Tumor targeting by enhanced per- factor-prekallikrein cascade by serratial protease, J. Bio-meability and retention (EPR) effect, Ann. Oncol. 9 (2) chem. 96 (1984) 739–749.(1998) 39. [38] A. Molla, Y. Matsumura, T. Yamamtoto, R. Okamura, H.
[25] Y. Takakura, M. Hashida, Macromolecular carrier systems Maeda, Pathogenetic capacity of proteases from Serratiafor targeting drug delivery: Pharmacokinetic considerations marcescens and Pseudomonas aeruginosa and their suppres-on biodistribution, Pharm. Res. 13 (1996) 820–831. sion by chicken egg white ovomacroglobulin, Infect. Immun.
[26] H. Maeda, J. Takeshita, R. Kanamaru, H. Sato, J. Katoh, 55 (1987) 2509–2517.Antimetastatic and antitumor activity of a derivative of [39] Y. Matsumura, K. Maruo, M. Kimura, T. Yamamoto, T.neocarzinostatin: An organic solvent- and water-soluble Konno, H. Maeda, Kinin-generating cascade in advancedpolymer-conjugated protein, Gann 70 (1979) 601–606. cancer patients and in vitro study, Jpn. J. Cancer Res. 82
[27] J. Takeshita, H. Maeda, R. Kanamaru, In vitro mode of (1991) 732–741.action, pharmacokinetics, and organ specificity of poly(mal- [40] T. Akaike, Y. Noguchi, S. Ijiri, K. Setoguchi, M. Suga, Y.M.eic acid–styrene)-conjugated neocarzinostatin, SMANCS, Zheng, B. Dietzschold, H. Maeda, Pathogenesis of influenzaGann 73 (1982) 278–284. virus-induced pneumonia: Involvement of both nitric oxide
[28] H. Maeda, T. Matsumoto, T. Konno, K. Iwai, M. Ueda, and oxygen radicals, Proc. Natl. Acad. Sci. USA 93 (1996)Tailor-making of protein drugs by polymer conjugation for 2448–2453.tumor targeting: A brief review on SMANCS, J. Protein [41] T. Akaike, M. Suga, H. Maeda, Free radicals in viralChem. 3 (1984) 181–193. pathogenesis: Molecular mechanisms involving superoxide
[29] F.C. Curtice, The origin of lipoprotein in lymph, in: H.S. and NO, Proc. Soc. Exp. Biol. Med. 217 (1998) 64–73.Meyersen (Chairman), Lymph and the Lymphatic System, [42] K. Umezawa, T. Akaike, S. Fujii, M. Suga, K. Setoguchi, A.Charles C. Thomas, Springfield, IL, 1963, pp. 89–126. Ozawa, H. Maeda, Induction of nitric oxide synthesis and
[30] H. Maeda, Y. Matsumura, Cancer selective macromolecular xanthine oxidase and their role in the antimicrobial mecha-therapeusis: Tailoring of an antitumor protein drug, in: R.E. nism against Salmonella typhimurium in mice, Infect.Feeney, J.R. Whitaker (Eds.), Protein Tailoring for Food and Immun. 65 (1997) 2932–2940.Medical Uses, Marcel Dekker, New York, 1985, pp. 353– [43] K. Doi, T. Akaike, S. Fujii, S. Tanaka, N. Ikebe, T. Beppu,382. K. Setoguchi, M. Takeya, S. Shibahara, M. Ogawa, H.
[31] Y. Kojima, A. Haruta, T. Imai, M. Otagiri, H. Maeda, Maeda, Induction of haeme oxygenase-1 by nitric oxide and

284 H. Maeda et al. / Journal of Controlled Release 65 (2000) 271 –284
ischemia in experimental solid tumours and implications for bradykinin in intravascular dissemination of Vibrio vulnificustumour growth, Br. J. Cancer 80 (1999) 1945–1954. and prevention of invasion by a bradykinin antagonist,
[44] T. Okamoto, T. Akaike, T. Nagano, S. Miyajima, M. Suga, Infect. Immun. 66 (1998) 866–869.M. Ando, K. Ichimori, H. Maeda, Activation of human [50] M. Suzuki, K. Hori, I. Abe, H. Saito, H. Sato, A newneutrophil procollagenase by nitrogen dioxide and perox- approach to cancer chemotherapy: Selective enhancement ofynitrite: A novel mechanism for procollagenase activation tumor blood flow with angiotensin II, J. Natl. Cancer Inst. 67involving nitric oxide, Arch. Biochem. Biophys. 342 (1997) (1981) 663–669.261–274. [51] C.J. Li, Y. Miyamoto, Y. Kojima, H. Maeda, Augmentation
[45] N. Utoguchi, H. Mizuguchi, A. Dantakean, H. Makimoto, Y. of tumor delivery of macromolecular drugs with reducedWakai, Y. Tsutsumi, S. Nakagawa, T. Mayumi, Effect of bone marrow delivery by elevating blood pressure, Br. J.tumor cell-conditioned medium on endothelial macromolecu- Cancer 67 (1993) 975–980.lar permeability and its correlation with collagen, Br. J. [52] K. Hori, S. Saito, H. Takahashi, H. Sato, H. Maeda, Y. Sato,Cancer 73 (1996) 24–28. Tumor selective blood flow decrease induced by an angioten-
[46] H.R. Lijnen, F. Ugwu, A. Bini, D. Collen, Generation of an sin converting enzyme inhibitor, temocapril hydrochloride,angiostatin-like fragment from plasminogen by stromelysin-1 Jpn. J. Cancer Res. (2000), in press.(MMP-3), Biochemistry 14 (1998) 4699–4702. [53] D.S. Zimmon, M. Oratz, R. Kessler, S.S. Schreiber, M.A.
[47] H.F. Dvorak, L.F. Brown, M. Detmar, A.M. Dvorak, Vascu- Rothschild, Albumin to ascites: Demonstration of a directlar permeability factor /vascular endothelial growth factor, pathway bypassing the systemic circulation, J. Clin. Invest.microvascular hyperpermeability, and angiogenesis, J. Natl. 48 (1969) 2074–2078.Cancer Inst. 146 (1996) 1029–1039. [54] K. Naito, N. Kanbayashi, S. Nakajima, T. Murai, K.
[48] Y. Sakata, T. Akaike, M. Suga, S. Ijiri, M. Ando, H. Maeda, Arakawa, S. Nishimura, A. Okuyama, Inhibition of growthBradykinin generation triggered by Pseudomonas proteases of human tumor cells in nude mice by a metalloproteasefacilitates invasion of the systemic circulation by Pseudo- inhibitor, Int. J. Cancer 58 (1994) 730–735.monas aeruginosa, Microbiol. Immunol. 40 (1996) 415–423. [55] A.R. Kennedy, Chemopreventive agents: Protease inhibitors,
[49] K. Maruo, T. Akaike, T. Ono, H. Maeda, Involvement of Pharmacol. Ther. 78 (1998) 167–209.



![Complement-Mediated Neutralization of Dengue Virus ... · rhage and vascular permeability syndrome (dengue hemorrhagic fever/dengue shock syndrome [DHF/DSS]) (2). Although the](https://img.dokumen.tips/doc/110x75/5cae414a88c9938f4d8c97e1/complement-mediated-neutralization-of-dengue-virus-rhage-and-vascular-permeability.jpg)
![Research Paper Zwitterionic-to-cationic charge conversion ...achieving improved drug delivery through the enhanced permeability and retention (EPR) effect [5-13]. Polyprodrug strategy,](https://img.dokumen.tips/doc/110x75/5ed95f94f59b0f56f45f5f00/research-paper-zwitterionic-to-cationic-charge-conversion-achieving-improved.jpg)
























