Embed Size (px)
Citation preview

JOURNAL OF CELLULAR PHYSIOLOGY 153:460-466 (1992)
Tributyltin I s a Potent Inducer of the Heat Shock Response in Human Diploid Fibroblasts
H U A Z H A N G AND ALICE Y.-C. L IU* Depdrtrrieril of Bdogicd/ Science,, Rulgers University, Busch Campus,
Piscataway, New jersey 08855- 1059
Submicromolar concentrations of tributyltin (TBT), a commercially used organotin compound, were found to induce the expression of several stress proteins, most notably HSP89 and HSP70, in IMK-90 human diploid fibroblasts in a time- and dose-dependent manner. This induction can be demowtrated by quantitation of 1) synthesis of the heat 5hock proteins (HSPs), 2) relative abundance of niRNA of hsp70, arid 3 ) Lransient expression of a human hsp70 promoter driven reporter gene. TBT also increased the abundance of mRNA of heme oxygenase, whereas heat shock was without effect. Analysis of protein binding to a consensus heat shock element (HSE) by electrophoretic mobility shift assay suggests that the induction of the heat shock response by TBT was attributable to activation of the heat shock transcription factor (HSTF).
Living cells respond to stress with specific biochemi- cal changes. One of the most profound and best studied biochemical parameters of stress is the rapid induction of a class of proteins known as the heat shock proteins (HSPs; for reviews, see Lindquist, 1986; Morimoto et al., 1990b; Nover, 1984). The heat shock response was first described by Ritossa (1962) in Drosophilu using heat shock and 2,4-dinitrophenol as inducers. Subse- quent studies from a number of laboratories have shown that the heat shock response is ubiquitous, and that the major HSPs are conserved through evolution a t the protein and DNA levels. These findings under- score the biological importance of HSPs (Lindquist, 1986; Morimoto et al., 1990b).
In addition to heat, the heat shock response can be activated by a wide variety of noxious stimuli including heavy metals, inhibitors of electron transport compo- nents, virus infection, and amino acid analogues (Nover, 1984). The mechanisms of action of many of these compoundsiconditions are not entirely clear; in the case of the heavy metal ion, cadmium, and the amino acid analogue, azatidine, the effects are due to activation of the heat shock element (HSEI-binding ac- tivity (Mosser et al., 1988).
Tributyltin (TBT) is one of several organotin com- pounds of industrial, commercial, and environmental importance. The primary utility of organotin com- pounds is 1) as heat stabilizers for polyvinyl chloride polymer, 2) as industrial, aquatic, and agricultural bio- cides, and 3) as industrial catalysts in a variety of chemical reactions (for review, see Snoeji et al., 1987; Zuckerman, 1976). TBT in particular is an effective biocide in antifouling paints. However, the biocidal properties of TBT that provide excellent antifouling action to various target organisms also present risks to nontarget organisms (Boyer, 1989; Gray et al., 1987; Hall, 1988; McMillan and Wenger, 1985).
Given the widespread use of TBT and related com- pounds and their potential ecological and environmen- C 1992 WILEY-LISS, INC
tal impacts, we decided to investigate the effects ofTBT on heat shock gene regulation in the IMR-90 human diploid fibroblasts. We report here on the induction of HSPs by TBT via a transcriptional mechanism.
MATERIALS AND METHODS Materials
Tributyltin chloride (TBT) was obtained from Ald- rich Chemical Co., Inc., Milwaukee, WI; a 1,000 x stock solution of TBT was prepared in dimethylsulfoxide. ["SlTrans-label (a 85:15% mixture of [35Slmethionine:- cysteine), L IX~~PI~CTP and [ySzPIATP were obtained from ICN, Irvine, CA. [14CJChloramphenicol was pro- vided by Amersham Corp., Arlington Heights, IL. Gene Screen Plus membrane was obtained from Dupont-New England Nuclear, North Billerica, MA. Nick transla- tion kit and T4 kinase were obtained from Bethesda Research Laboratories, Gaithersburg, MD. Tissue cul- ture supplies were obtained from GIBCO, Grand Is- land, NY. The plasmids pH2.3, which contains the hu- man hsp70 gene minus 150 base pairs of the 5'- nontranslated and upstream sequences, and pHBCAT, a construct linking the human hsp70 promoter to the coding sequence of chloramphenicol acetyltransferase (CAT), were generous gifts from Dr. Richard I. Mori- mot0 (Wu et al., 1985) of Northwestern University. The plasmid pHHO (2/10), which contains the cDNA of hu- man heme oxygenase, was a generous gift from Dr. Rex M. Tyrrell (Keyse and Tyrrell, 1989) of the Swiss Insti- tute for Experimental Cancer Research, Epalinges, Switzerland.
Cell culture IMR-90 cells were obtained from the Institute of Med-
ical Research, Camden, NJ. Cells were grown as mono-
Received September 23,1991; accepted J u n e 23,1992.
*To whom reprint requests/correspondcnce should he addressed.
H S P 8 9 -
H S P 7 0 -
H S P 6 4 -
H S P 2 5 -
TBT (nM)
HS I 0 6 6 0 6 0 0 0
ORGANOTIN AND HEAT SHOCK RESPONSE
TBT (0.6 pM)
0 3 6 9 12 24 (htS)
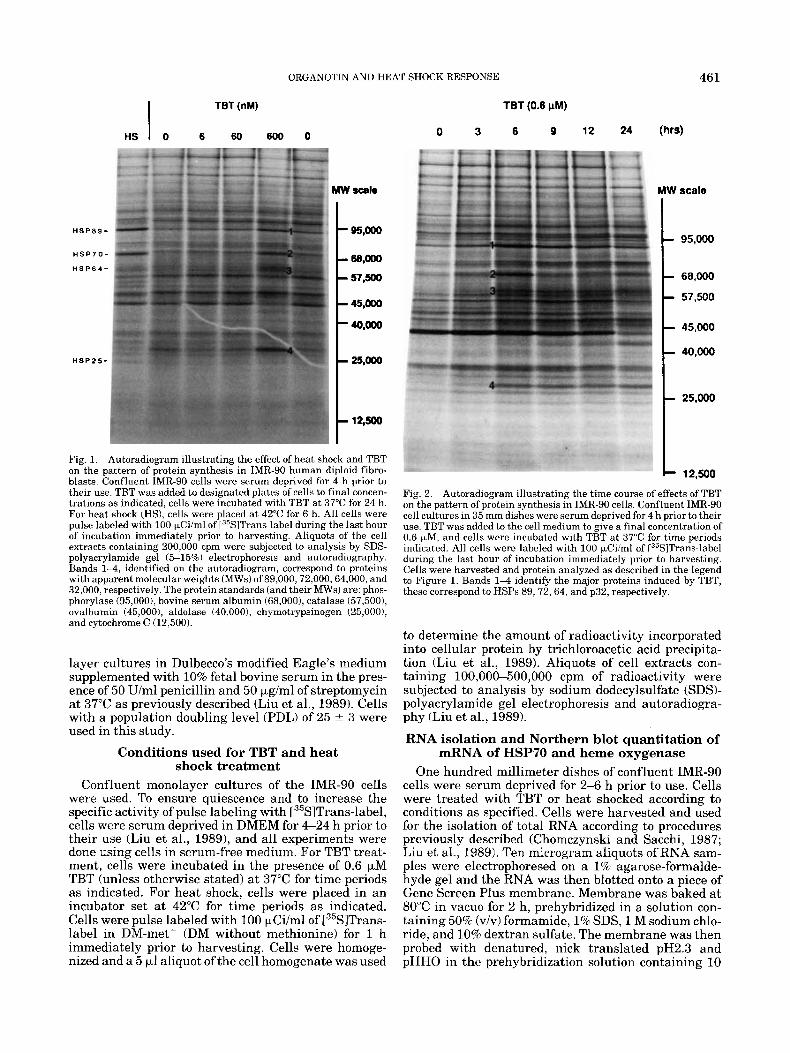
Fig. 1. Autoradiogram illustrating the effect of heat shock and TBT on the pattern of protein synthesis in IMR-90 human diploid fibro- blasts. Confluent IMR-90 cells were serum deprived for 4 h prior to their use. TBT was added to designated plates of cells to final concen- trations as indicated, cells were incubated with TBT at 37°C for 24 h. For heat shock (HS), cells were placed a t 42°C for 6 h. All cells were pulse labeled with 100 kCi/ml of [35S1Trans-label during the last hour of incubation immediately prior to harvesting. Aliquots of the cell extracts containing 200,000 cpm were subjected to analysis by SDS- polyacrylamide gel (5-15%) electrophoresis and autoradiography. Bands 1 4 , identified on the autoradiogram, correspond to proteins with apparent molecular weights (MWs1 of89,000,72,000,64,000, and 32,000, respectively. The protein standards (and their MWs) are: phos- phorylase (95,0001, bovine serum albumin (68,000), catalase (57,5001, ovalbumin (45,0001, aldolase (40,000), chymotrypsinogen (25,0001, and cytochrome C (12,500).
layer cultures in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum in the pres- ence of 50 Uiml penicillin and 50 pgiml of streptomycin at 37°C as previously described (Liu et al., 1989). Cells with a population doubling level (PDL) of 25 2 3 were used in this study.
Conditions used for TBT and heat shock treatment
Confluent monolayer cultures of the IMR-90 cells were used. To ensure quiescence and to increase the specific activity of pulse labeling with ["SlTrans-label, cells were serum deprived in DMEM for 4-24 h prior to their use (Liu et al., 1989), and all experiments were done using cells in serum-free medium. For TBT treat- ment, cells were incubated in the presence of 0.6 pM TBT (unless otherwise stated) a t 37°C for time periods as indicated. For heat shock, cells were placed in an incubator set at 42°C for time periods as indicated. Cells were pulse labeled with 100 pCiiml of [35S]Trans- label in DM-met- (DM without methionine) for 1 h immediately prior to harvesting. Cells were homoge- nized and a 5 ~1 aliquot of the cell homogenate was used
461
V scale
- 95,000
- 68,000
- 57,500
- 45,000
- 40,000
- 25,000
- 12,500
Fig. 2. Autoradiogram illustrating the time course of effects of TRT on the pattern of protein synthesis in IMR-90 cells. Confluent IMR-90 cell cultures in 35 mm dishes were serum deprived for 4 h prior to their use. TBT was added to the cell medium to give a final concentration of 0.6 KM, and cells were incubated with TBT a t 37°C for time periods indicated. All cells were labeled with 100 kCi/ml of T35S1Trans-label during the last hour of incubation immediately prior to harvesting. Cells were harvested and protein analyzed as described in the legend to Figure 1. Bands 1 4 identify the major proteins induced by TBT, these correspond to HSPs 89,72,64, and p32, respectively.
to determine the amount of radioactivity incorporated into cellular protein by trichloroacetic acid precipita- tion (Liu et al., 1989). Aliquots of cell extracts con- taining 100,000-500,000 cpm of radioactivity were subjected to analysis by sodium dodecylsulfate (SDSI- polyacrylamide gel electrophoresis and autoradiogra- phy (Liu et al., 1989).
RNA isolation and Northern blot quantitation of mRNA of HSP70 and heme oxygenase
One hundred millimeter dishes of confluent IMR-90 cells were serum deprived for 2-6 h prior to use. Cells were treated with TBT or heat shocked according to conditions as specified. Cells were harvested and used for the isolation of total RNA according to procedures previously described (Chomczynski and Sacchi, 1987; Liu et al., 1989). Ten microgram aliquots of RNA sam- ples were electrophoresed on a 1% agarose-formalde- hyde gel and the RNA was then blotted onto a piece of Gene Screen Plus membrane. Membrane was baked at 80°C in vacuo for 2 h, prehybridized in a solution con- taining 50% (viv) formamide, 1% SDS, 1 M sodium chlo- ride, and 10% dextran sulfate. The membrane was then probed with denatured, nick translated pH2.3 and pHHO in the prehybridization solution containing 10

462 ZHANG AND LIU
TBT (0.6 pM) HS (42 OC)
Tlme (hrs) 0 1 3 6 I 1 3 6
2.6 kb mRNA of hsp 70
1.6 kb mRNA of heme oxygenase
Fig. 3. Northern blot analysis of mRNA of hsp70 and heme oxygen- ase in control, TBT-treated, and heat shocked IMR-90 cells. Confluent cultures of IMR-90 cells were serum deprived for 6 h prior to their use. Cells were incubated with 0.6 p M of TBT for 1,3, and 6 h a t 37°C. Cells heat shocked at. 42°C for 1, 3, and 6 h were used as positive controls. Size fractionated RNA immobilized onto Gene Screen Plus membrane was probed with pH2.3 and pHHO, DNA probes of hsp70 and heme oxygenase, respectively. Quantitation of the 2.6 kb mRNA ofhsp70, by
liquid scintillation counting of excised portions of the membrane, showed that a t the 6-h time point, TBT and heat shock increased the amount of this mRNA by 35- and 49-fold, respectively, over that of the control. Quantitation of the relative abundance of mRNA of heme oxygenase by scanning the autoradiogram with a Joyce Loebl densito- meter showed that there was a 35-fold increase at the 6-h time point of treatment with TBT over that of the control.
mgiml salmon sperm DNA overnight a t 60°C. The membrane was washed and exposed to X-Omat film with intensifying screen at - 80°C for an appropriate time period. Quantitation of mRNA abundance was done either by liquid scintillation counting of excised portions of the membrane or by scanning the autoradio- gram with a densitometer.
scribed (Choi e t al., 1990). The HSE used was a double- stranded 24mer with four GAA blocks in alternat- ing orientations: S'GCCTCGAATGTTCGCGAAGTT- TCG-3' and its complement. To test for specificity of the TBT-induced HSE-binding activity, standard competi- tion assay with a 200 x molar excess of HSE or I-CAP DNA was done (Choi et al., 1990).
DNA transfection and chloramphenicol acetyltransferase (CAT) assay
Cells were transfected with pHBCAT, an hsp70 pro- moter driven reporter gene, essentially according to procedures described (Liu et al., 1989). Thirty-six hours after DNA transfection, cells were rinsed and replen- ished with serum-free medium, TBT was then added, and cells were incubated with TBT at 37°C for a speci- fied time period. In this study, the effects of heat shock on the expression of pHBCAT was used as a positive control. CAT activity was assayed using 50 pg of cell extract protein, 0.1 WCi ~'4C]chloramphenicol, and 0.4 mM acetyl CoA; reaction was run at 37°C for 90 min (Gorman et al., 1982). CAT activity, expressed as per- cent of [14C]chloramphenicol acetylated, was deter- mined by liquid scintillation counting of the appropri- ate spots on the thin layer chromatogram.
Assay of heat shock transcription factor DNA-binding activity by electrophoretic mobility
shift assay Unless otherwise specified, confluent cells in 100 mm
dishes were rinsed and replenished with serum-free medium, TBT was then added, and cells were incubated at 37°C for time periods as specified. Electrophoretic mobility shift assay was performed as previously de-
RESULTS Figure 1 illustrates the effects of increasing concen-
trations of TBT on the pattern of protein synthesis in IMR-90 cells. In this experiment, confluent cells were incubated with 0, 6, 60, and 600 nM of TBT at 37°C for 24 h, and cells were pulse labeled with [35S]Trans-label for 1 h immediately prior to harvesting. Several points can be summarized from the results presented 1) 0.6 p M TBT enhanced the synthesis rate of proteins with apparent molecular weights of 89,72, and 64 KDa. The identity of these proteins with HSPs 89,72, and 64 was demonstrated by the comigration on SDS-polyacryla- mide gel of the TBT-induced proteins and the corre- sponding HSPs induced by heat shock. 2) TBT also in- duced the synthesis of another protein (band 4) with an apparent molecular weight of 32 KDa (p32); p32 was not induced by heat shock. 3) There was a significant decrease in the synthesis rate of the 45 KDa actin in cells treated with 0.6 pM TBT for 24 h; synthesis of other proteins was not significantly affected. It should be noted that while the IMR-90 cells could tolerate sub- micromolar concentrations of TBT for long periods of time, high concentrations of TBT inhibited general pro- tein synthesis and caused cell lysis in a dose- and time- dependent manner. For these reasons, we routinely

ORGANOTIN AND HEAT SHOCK RESPONSE 463
c- TBT (hrs)-
< 3AC
q 1AC
origin
L a n e 1 2 3 4 5 6 7
CAT activity 7.5 19 36 6 0 4 4 4 2 87 (% acetylation)
Fig. 4. Effects of TBT and heat shock on the transient expression of the hsp7O promoter driven reporter gene, pHBCAT. The day before transfection, cells were subcultured and plated at 1:4 dilution in 100 mm dishes. Calcium phosphate-DNA coprecipitate was prepared, and cells were transfected with the precipitate accarding to methods de- scribed previously (Liu et al., 1989). Thirty-six hours after DNA trans- fection, cells were rinsed and replenished with serum-free medium, TBT was added to a final concentration of 0.6 pM, and cells were incubated in the presence of TBT at 37°C for 1 , 2 , 3 , 4 , and 6 h (lanes 2-6). Lane 7 illustrates reporter gene activity of cells heat shocked at 42°C for 3 h. CAT assay was done according to methods described in the text. The percentage of 1'4Clchloramphenicol acetylated, deter- mined by liquid scintillation counting of excised portions of the thin layer chromatogram, is presented at the bottom of the autoradiogram.
used 0.6 pM TBT for the experiments described in this study unless specified otherwise.
Figure 2 illustrates the time course of effect of TBT. The induction of synthesis of HSPs and p32 by TBT appeared rapidly; a maximum induction of the synthe- sis of HSP89,72, and 64 identified as bands 1 through 3, as well as p32 (band 4) occurred a t 6 h. Thereafter, the synthesis of HSP89 and 72 declined somewhat, whereas the induced synthesis of HSP64 and p32 was sustained for at least 24 h, the longest time period stud- ied.
We were interested in the identity of the TBT-in- duced 32 kDa protein as well as the mechanism by which TBT induced the synthesis of HSPs. We note that a 32 dKa protein has been reported by many investiga- tors to be induced in human cell lines by thiol-reactive agents andlor oxidant stress; this protein has since been identified as heme oxygenase (Caltabiano et al., 1986; Keyse and Tyrrell, 1987, 1989; Keyse et al., 1990; Ta- ketani et al., 1989). Given the SH-reactivity of TBT (Gould, 1978; Gray et al., 1987) and its ability to uncou- ple oxidative phosphorylation (Aldridge, 1976), we evaluated the possibility that TBT may also induce the expression of heme oxygenase. In the experiment illus-
trated in Figure 3, total RNA isolated from control, TBT-treated, and heat shocked cells was hybridized with DNA probes of the human hsp70 and heme oxyge- nase genes. We showed that both TBT (0.6 pM) and heat shock (42°C) increased the abundance of the 2.6 kb mRNA hsp70. On the other hand, the 1.6 kb mRNA of heme oxygenase was induced by TBT but not by heat shock. The induction of the mRNA of hsp70 and heme oxygenase by TBT was time dependent; at 6 h of incu- bation with TBT, we observed a 35-fold increase in the mRNA of both hsp70 and HO over that of the control.
To further ascertain the mechanism of action of TBT on the induction of HSPs, we analyzed the effects of TBT on the transient expression of a human hsp7O pro- moter driven reporter gene, pHBCAT. In the experi- ment illustrated in Figure 4, IMR-90 cells were trans- fected with pHBCAT, and CAT activity present in extracts of control, TBT-treated, and heat shocked cells was determined. We showed that TBT increased re- porter gene expression in a time-dependent manner; a sustained maximal increase, which measured approxi- mately six-fold over that of the control, was observed at 3-6 h of treatment with TBT. In the same experiment, we showed that heat shock (42"C, 3 h) gave an 11.6-fold increase in reporter gene activity.
The promoter of the human hsp7O gene is equipped with multiple basal and inducible control elements (Morimoto et al., 1990a). In particular, we note that the effect of heat shock is mediated through a consensus DNA sequence known as the HSE consisting of alter- nating GAA or TTC blocks arranged at 2-nucleotide intervals (Amin et al., 1988; Xiao and Lis, 1988). Heat shock promotes the activation of a preexisting HSTF (Zimarino and Wu, 19871, the activated HSTF then binds to the HSE, and facilitates the transcription of hsp genes (Morimoto et al., 1990a,b; Mosser et al., 1988). Significantly, the effects of some of the other elicitors of heat shock response, including heavy metal ions and amino acid analogues, also appear to be attrib- utable to activation of the HSE-binding activity (Mosser et al., 1988). This being given, we asked if TBT would activate the DNA-binding activity of HSTF, and if this activation could subserve the effects of TBT on heat shock gene expression.
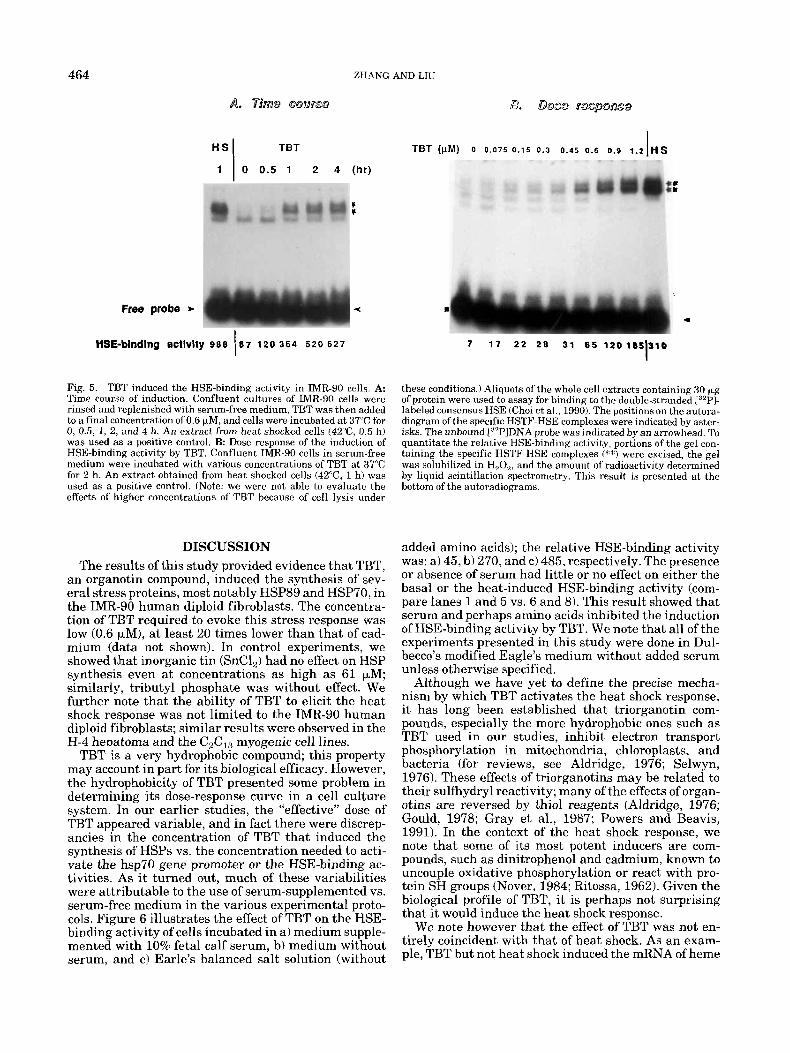
In Figure 5, we showed that TBT activated the HSE- binding activity in a time- and dose-dependent manner. The induction of HSE-binding activity by TBT was in- dependent of new protein synthesis in that it was ob- served both in the absence and presence of cyclohexi- mide, suggesting a direct effect of TBT (data not shown). We note that the mobility of the TBT-induced HSE-protein complex was identical to that induced by heat shock. Further, the TBT-induced HSE-binding ac- tivity (a doublet indicated by asterisks) was displaced specifically by HSE and not by unrelated DNA se- quence (data not shown). Analysis of the time course and dose dependence of the induction of HSE-binding activity by TBT suggest that this increase can account for some of the down stream events, including promoter activation, increased abundance of mRNA, and synthe- sis of the HSPs that we have observed and demon- strated in Figures 1 through 4 of this study. These results suggest that the effects of TBT on HSP induc- tion is transcriptional.
464
TBT (pM) o 0.075 0 .15 0.3 0.45 0.6 0.9 1.2 HS I TBT
1 0 0.5 1 2 4 (hr) c *
V r
H S I
Free probe b <
7 1 7 2 2 2 8 31 6 5 120185310 I HSE-binding activity 988 67 120 364 520 627 I Fig. 5. TBT induced the HSE-binding activity in IMR-90 cells. A: Time course of induction. Confluent cultures of IMR-90 cells were rinsed and replenished with serum-free medium, TBT was then added to a final concentration of 0.6 kM, and cells were incubated at 37°C for 0, 0.5, 1, 2, and 4 h. An extract from heat shocked cells (42"C, 0.5 h) was used as a positive control. €3: Dose response of the induction of HSE-binding activity by TBT. Confluent TMR-90 cells in serum-free medium were incubated with various concentrations of TBT at 37°C for 2 h. An extract obtained from heat shocked cells (42"C, 1 h) was used as a positive control. (Note: we were not able to evaluate the effects of higher concentrations of TBT because of cell lysis under
DISCUSSION The results of this study provided evidence that TBT,
an organotin compound, induced the synthesis of sev- eral stress proteins, most notably HSP89 and HSP70, in the IMR-90 human diploid fibroblasts. The concentra- tion of TBT required to evoke this stress response was low (0.6 KM), a t least 20 times lower than that of cad- mium (data not shown). In control experiments, we showed that inorganic tin (SnC1,) had no effect on HSP synthesis even at concentrations as high as 61 kM; similarly, tributyl phosphate was without effect. We further note that the ability of TBT to elicit the heat shock response was not limited to the IMR-90 human diploid fibroblasts; similar results were observed in the H-4 hepatoma and the C2CI3 myogenic cell lines.
TBT is a very hydrophobic compound; this property may account in part for its biological efficacy. However, the hydrophobicity of TBT presented some problem in determining its dose-response curve in a cell culture system. In our earlier studies, the "effective" dose of TBT appeared variable, and in fact there were discrep- ancies in the concentration of TBT that induced the synthesis of HSPs vs. the concentration needed to acti- vate the hsp70 gene promoter or the HSE-binding ac- tivities. As it turned out, much of these variabilities were attributable to the use of serum-supplemented vs. serum-free medium in the various experimental proto- cols. Figure 6 illustrates the effect of TBT on the HSE- binding activity of cells incubated in a) medium supple- mented with 10% fetal calf serum, b) medium without serum, and c) Earle's balanced salt solution (without
these conditions.) Aliquots of the whole cell extracts containing 30 pg of protein were used to assay for binding to the double-stranded [32P]- labeled consensus HSE (Choi et al., 1990). The positions on the autora- diogram of the specific HSTF-HSE complexes were indicated by aster- isks. The unbound L"P1DNA probe was indicated by an arrowhead. To quantitate the relative HSE-binding activity, portions of the gel con- taining the specific HSTF-HSE complexes (**) were excised, the gel was solubilized in H,O,, and the amount of radioactivity determined by liquid scintillation spectromet.ry. This result is presented at the bottom of the autoradiograms.
added amino acids); t.he relative HSE-binding activity was: a) 45, bl270, and c) 485, respectively. The presence or absence of serum had little or no effect on either the basal or the heat-induced HSE-binding activity (com- pare lanes 1 and 5 vs. 6 and 8). This result showed that serum and perhaps amino acids inhibited the induction of HSE-binding activity by TBT. We note that all of the experiments presented in this study were done in Dul- becco's modified Eagle's medium without added serum unless otherwise specified.
Although we have yet to define the precise mecha- nism by which TBT activates the heat shock response, it has long been established that triorganotin com- pounds, especially the more hydrophobic ones such as TBT used in our studies, inhibit electron transport phosphorylation in mitochondria, chloroplasts, and bacteria (for reviews, see Aldridge, 1976; Selwyn, 1976). These effects of triorganotins may be related to their sulfhydryl reactivity; many of the effects of organ- otins are reversed by thiol reagents (Aldridge, 1976; Gould, 1978; Gray et al., 1987; Powers and Beavis, 1991). In the context of the heat shock response, we note that some of its most potent inducers are com- pounds, such as dinitrophenol and cadmium, known to uncouple oxidative phosphorylation or react with pro- tein SH groups (Nover, 1984; Ritossa, 1962). Given the biological profile of TBT, it is perhaps not surprising that it would induce the heat shock response.
We note however that the effect of TBT was not en- tirely coincident with that of heat shock. As an exam- ple, TBT but not heat shock induced the mRNA of heme

ORGANOTIN AND HEAT SHOCK RESPONSE
t TBT - C I S b c l H S
465
C TBT H S
Free probe- - Lane 1 2 3 4 5 6 7 8
HSE-bindfng 8CtiVity 4 0 4 5 2 7 0 4 8 5 624 4 2 295 6 5 0
Fig. 6. Effects of medium composition on the induction of HSE-bind- ing activity by TBT. Lanes 1-5: Confluent monolayer cultures of IMR-90 cells in a 10% fetal bovine serum-supplemented medium were used. The effects of 0.6 pM TBT (1 h, 37°C) on the activation of HSE- binding activity was evaluated using a) cells maintained in the 10% serum-supplemented medium, b) cells rinsed and replenished with serum-free medium, and c) cells rinsed and replenished with Earle’s balanced salt solution, as represented by lanes 2, 3, and 4, respec- tively. Lanes 1 and 5 represent, respectively, samples from control
oxygenase in IMR-90 cells, It is possible that binding of HSTF to the HSE is, by itself, not sufficient to confer transcriptional activation to the heme oxygenase gene; this activation may require the corporation of HSTF that binds to HSE and another transacting factor, such as the HO transcription factor (HOTF) proposed by Sat0 et al. (1989).
TBT has long been known to be extremely toxic to a wide variety of terrestrial and aquatic organisms (Boyer, 1989; Gray et al., 1987; Hall, 1988; McMillan and Wenger, 1985; Snoeji et al., 1987; Zuckerman, 1976). Given the many important and diverse functions of HSPs (Morimoto, 1990b), the induction of HSPs by TBT in human diploid cells may provide a basis to bet- ter understand the effects of TBT at a molecular and cellular level.
ACKNOWLEDGMENTS We thank Drs. Morimoto and Tyrrell for providing us
with the DNA probes. We thank Dr. K. Y. Chen of the Chemistry Department at Rutgers University for help- ful discussions. This work was supported in part by grants from the American Cancer Society (CD 406 and CB 28B) and National Science Foundation (DCB 90- 19808). Support from the Busch Memorial Fund is ac- knowledged.
LITERATURE CITED Aldridge, W.N. (1976) The influence of organotin compounds on mito-
chondrial functions. In: Organotin Compounds: New Chemistry and
(37°C) and heat shocked (42”C, 1 h) cells incubated in medium supple- mented with 10% fetal bovine serum. Lanes 6 4 HSE-binding activ- ity of control, TBT-treated (0.6 pM, 1 h), and heat shocked (42OC, 1 h) cells maintained in serum-free medium. Cells were thoroughly rinsed and incubated in serum-free medium for 2 h prior to subjecting them to treatment with TBT or heat shock. The amount of 1”’PIHSE specifi- cally bound (indicated by **) was determined by liquid scintillation counting of excised portions of the gel, and the result is illustrated at the bottom of Ihe autoradiogram.
Applications. J. J. Zuckerman, ed., Advances in Chemistry Series 157, American Chemical Society, Washington, D.C., pp. 186196.
Amin, J., Ananthan, J., and Voellmy, R. (1988) Key features of heat shock regulatory elements. Mol. Cell. Biol., 8,37613769,
Boyer, I.J. (1989) Toxicity of dibutyltin, tributyltin, and other organ- otin comoounds to humans and to exuerimental animals. Toxieol- ogy, 55.2’53-298.
Caltabiano, M.M., Koestler, T.P., Poste, G., and Greia, R. G. (1986) Induction of 32- and 34-KDa stress proteins by sodium arsenite, heavy metals, and thiol-reactive agents. J. Biol. Chem., 261r13381- 13386.
Choi, H-S., Lin, Z., Li, B., and Liu, A.Y.-C. (1990) Age-dependent decrease in the heat inducible DNA sequence-specific binding activ- ity in human diploid fibroblasts. J. Biol. Chem., 265:18005-18011.
Chomczynski, P., and Sacchi, N. (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform ex- traction. Anal. Biochem., 162:15&159.
Gorman, C.M., Moffat, L.F.M., and Howard, B.H. (1982) Recombinant genomes which express chloramphenicol acetyltransferase in mam- malian cells. Mol. Cell. B i d , 2:1044-1051.
Gould, J.M. (1978) Dithiol-specific reversal of triphenyltin inhibition of CP,-catalyzed transmembrane proton transfer in chloroplasts. FEBS Lett., 94r90-94.
Gray, B.H., Porvaznik, M., Flemming, C., and Lee, L.H. (1987) Tri-n- butyltin: A membrane toxicant. Toxicology, 47:35-54.
Hall, L.W. (1988) Tributyltin environmental studies in Chesapeake Bay. Marine Pollution Bull., I9:431-438.
Keyse, S.M., and Tyrrell, R.M. (1987) Both near UV radiation and the oxidizing agent hydrogen peroxide induce a 32-KDa stress protein in normal human skin fibroblasts. J. Biol. Chem., 262r14821- 14825.
Keyse, S.M., and Tyrrell, R.M. (1989) Heme oxygenase is the major 32-KDa stress protein induced in human skin fibroblasts by UV- radiation, hydrogen peroxide and sodium arsenite. Proc. Natl. Acad. Sci., U.S.A., 86:99-103.
Keyse, S.M., Applegate, L.A., Tromvoukis, Y ., and Tyrrell, R.M. (1990) Oxidant stress leads to transcriptional activation of the hu- man heme oxygenase gene in cultured skin fibroblasts. Mol. Cell. Biol., 10:4967-4969.

466 ZHANG AND LIU
Lindquist, S. (1986) The hcat shock response. Ann. Rev. Biochem., 55,1151-1191.
Liu, A.Y.-C., Lin, Z., Choi, H.-S., Sorhage, F., and Li, €3. (1989) Atten- uated induction of heat shock gene expression in aging diploid fibro- blasts. J. Biol. Chem., 264:12037-12045.
McMillan, D.E., and Wenger, G.R. (1985) Neurobehavioral toxicology of trialkyltins. Pharmacol. Rev: 37t365-379.
Morimoto, R. I., Abravaya, K., Mosser, D., and Williams, G.T. (1990a) Transcription of the human hsp7O gene: Cis-acting elements and trans-acting factors involved in basal, adenovirus Ela . and stress- induced expression. In: Stress Proteins, Induction and Function. M.J. Schlesinger, M.G. Santoro, and E. Garaci, eds. Springer-Ver- lag, Berlin/HeidelbergNew York, pp. 1-18.
Morimoto, R.I., Tissieres, A., and Georgopoulos, C. (1990b) Stress Pro- tein in Biology and Medicine. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, p. 449.
Mosser, D.D., Theodorakis, N.G., and Morimoto, R.I. (1988) Coordi- nate changes in heat shock element-binding activity and HSP'IO gene transcription rates in human cells. Mol. Cell. Biol., 8,4736- 4744.
Nover, L. (1984) Heat Shock Response of Eukaryotic Cells. Springer- Verlag, BerliniHeidelberglNew York, p. 82.
Powers, M.F., and Beavis, A.D. (1991) Triorganotins inhibit the mito- chondrial inner membrane anion channel. J. Biol. Chem., 266:17250-17256.
Ritossa, F. (1962) A new puffing pattern induced by temperature shock and DNP in Drosophilu. Experientia, 18:571-573
Sato, M., Fukushi, Y.. Ishizama, S., Okinaga, S., Muller, R.M., and Shibahara, S. (1989) Transcriptional control of the rat heme oxyge- nase gene by a nuclear protein that interacts with adenovirus 2 major late promoter. J. Biol. Chem., 264,10251-10260.
Selwyn, M.J. (1976) Triorganotin compounds as ionophores and inhib- itors of ion translocating ATPase. In: Organotin Compounds: New Chemistry and Applications. J.J. Zuckerman, ed. Advances in Chemistry Series 157, American Chemical Society, Washington, D.C., pp. 204-226.
Snoeji, N.J., Penninks, A.H., and Seinen, W. (1987) Biological activity of organotin compounds-An overview. Environ. Res., 44t335-353.
Taketani, S., Kohno, H., Yoshinaga, T., and Tokunaga, R. (1989) The human 32-KDa stress protein induced by exposure to arsenite and cadmium ions is heme oxygenase. FEES Lett., 245:173-176.
Wu, B., Hunt, C., and Morimoto, R.1. (1985) Structure and expression of the human gene encoding major heat shock protein hsp70. Mol. Cell. Biol., 5:330-342.
Xiao, H., and Lis, J.T. (1988) Germline transformation used to define key features of heat-shock response elements. Science, 239:1139- 1142.
Zimarino, V., and Wu, C. 11987) Induction of sequence-specific binding of' Drosophilu heat shock activator protein without prior protein synthesis. Nature, 327t727-730.
Zuckerman, J.J . (1976) Organotin Compounds: New Chemistry and Applications. Advances in Chemistry Series 157, American Chemi- cal Society, Washington, D.C., pp, 1-299.





























