Embed Size (px)
Citation preview

www.elsevier.com/locate/jneuroim
Journal of Neuroimmunology 148 (2004) 162–171
Treatment with an acetylcholinesterase inhibitor in Alzheimer patients
modulates the expression and production of the pro-inflammatory
and anti-inflammatory cytokines
Marcella Realea, Carla Iarlorib, Francesco Gambic, Claudio Felicianid, Anatolia Salonec,Lucia Tomab, Giovanna DeLucab, Mirella Salvatoree, Pio Contia, Domenico Gambib,*
a Immunology Unit, Department of Oncology and Neuroscience, University UD’A, Via dei Vestini, 66013 Chieti, ItalybNeurology Unit, Department of Oncology and Neuroscience, University UD’A, Via dei Vestini, 66013 Chieti, ItalycPsychiatry Unit, Department of Oncology and Neuroscience, University UD’A, Via dei Vestini, 66013 Chieti, Italy
dDermatology Unit, Department of Oncology and Neuroscience, University UD’A, Via dei Vestini, 66013 Chieti, ItalyeDepartment MQIE-Faculty of Economics, Italy
Received 20 May 2003; received in revised form 27 October 2003; accepted 3 November 2003
Abstract
Elevated levels of cytokines have been detected in brains of Alzheimer’s disease (AD) patients, and altered peripheral levels of IL-1h,TNFa and IL-6 have been reported in these patients. We studied the ability of PBMC from patients with AD, matched with a control group,
to release pro- and anti-inflammatory cytokines, and the effect of AChEI treatment on cytokine release. Our data indicates that AChEI
treatment down-regulates IL-1, IL-6 and TNF, and up-regulates the expression and production of IL-4 in PBMC in AD patients, and that
AChEI leads to the remodelling of the cytokine network, probably acting on the lymphocytic cholinergic system.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Alzheimer’s disease; IL-1h; IL-6; TNFa; IL-4; PBMC
1. Introduction
Inflammatory mechanisms and immune activation have
been hypothesized to play a role in the pathogenesis of age-
associate disease (O’Mahony et al., 1998; Gerli et al., 2000)
and neurodegenerative processes such as dementia and
atherosclerosis (Benveniste, 1992; De Luca et al., 1998;
Porrini et al., 1998; Bruunsgaard et al., 1999; Iarlori et al.,
2000, 2002; Bruunsgaard and Pedersen, 2003).
Alzheimer’s disease (AD) is a chronic neurodegenerative
disorder causing progressive impairment of memory and
cognitive function. The amyloid cascade hypothesis sug-
gests that unregulated metabolism of the beta-amyloid (Ah)precursor protein (APP) followed by subsequent formation
of non-fibrillar and fibrillar Ah deposits leads to glial
activation and eventually to neurotoxicity, causing cognitive
impairment (Maat-Schieman et al., 1997). Ah up-regulates
0165-5728/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/j.jneuroim.2003.11.003
* Corresponding author. Tel./fax: +39-871-562019.
E-mail address: [email protected] (D. Gambi).
and activates astrocytes, microglia and monocytes to act as
pro-inflammatory cells. These cells release a myriad of pro-
inflammatory cytokines, including TNF, IL-1 and IL-6
(Meda et al., 1999). In AD, immune and inflammatory-
related proteins have been implicated as mediators in
response to brain injury. Elevated levels of cytokines have
been detected in brains of AD patients. Altered peripheral
levels of IL-1h, TNFa and IL-6 have been reported in
patients with Alzheimer’s disease (Licastro et al., 2000).
Since the most recent successful therapeutic approach to AD
involve acetylcholinesterase inhibitors (AChEI) (Giacobini,
2001; Winblad et al., 2001), we investigated the ability of
peripheral blood mononuclear cells from a group of patients
with AD, matched with healthy controls (HC), to release
pro- and anti-inflammatory cytokines, and studied if the
stabilizing effect of AChEI treatment in vivo may relate to
the cytokine release or inhibition.
Biologically, IL-1 and TNF are closely related, because
of their multiple pro-inflammatory properties, these cyto-
kines contribute to many diseases and have strategic impor-
tance to the initiation and progression of inflammation

Table.1
Characteristics of AD patients and healthy controls at blood withdrawal
AD patients Age Gender Medication
1 77 M �2 75 F + (antibiotic)
3 63 F �4 80 F �5 81 M + (anxiolytic)
6 61 F �7 67 M + (analgesic)
8 74 M �9 76 F + (anxiolytic)
10 88 M + (vitamins)
11 77 F �12 70 F + (analgesic)
M. Reale et al. / Journal of Neuroimmunology 148 (2004) 162–171 163
(Dinarello, 1997). IL-1, a key molecule in systemic immune
responses in health and disease, has analogous roles in the
brain where it may contribute to neuronal degeneration
(Sheng et al., 1996) by triggering production of other
cytokines and nitric oxide (Rossi and Bianchini, 1996);
IL-1 promote Ah deposition through increased expression
and processing of amyloid precursor protein (APP) (Roth-
well and Hopkins, 1995) and may facilitate dystrophic
neuritic formation in diffuse non-neuritic plaques (Griffin
et al., 1995). In Alzheimer’s disease, TNF is up-regulated
and increases the production of Ah and inhibits the secre-
tion of neuroprotective, soluble amyloid precursor protein
(sAPPs). The up-regulation of Ah and ApoE by TNF would
then lead to increased neuritic plaque formation.(Perry et al.,
2001). High levels of protease inhibitors induced by IL-1
and IL-6 were responsible for an abnormal processing of
APP leading to high levels of insoluble Ah (Del Bo et al.,
1995). Over-expression of IL-6 in Alzheimer’s microglia are
associated with Ah plaque (Huell et al., 1995), and plasma
IL-6 concentrations in AD subjects was also significantly
higher than that of the control cases (Shibata et al., 2002).
The role of IL-4 in neurodegenerative disorders is still
unclear; however, IL-4, such as other anti-inflammatory
cytokines, regulates microglial responses to Ah in primary
murine microglia and in human monocyte cell line. IL-4 has
been reported to regulate Ah-induced production of the
inflammatory cytokines, IL-1 and IL-6 (Szcepanik et al.,
2001a). IL-4 is able to block or suppress IL-1, TNFa, IL-6
and IL-8 (Conti and Dempsey, 1990; Lee et al., 1995). IL-4
activity may also be associated with the pathophysiology of
AD (Abbas et al., 2002).
Our results demonstrate that, in AD patients the pro- and
anti-inflammatory peripheral cytokine system is affected
when compared with age/sex-matched HC, and that AChEI
treatment modulates cytokine production. Thus, at this stage,
we believe that AChEI treatment, besides restoring the
cholinergic system, reverses the cytokine imbalances ob-
served in AD.
13 68 F + (antibiotics plus vitamins)
14 84 M �15 77 M �16 80 M + (vitamins)
17 75 F �18 80 F + (vitamins plus anxiolytic)
19 67 M �20 72 F �21 73 F �
HC subject
1 77 M �2 69 M �3 81 F + (antibiotic)
4 72 M �5 76 F �6 75 F + (analgesic)
7 73 M �8 82 F �9 77 F �10 74 M + (analgesic)
2. Materials and methods
2.1. Subjects
Twenty-one AD patients, mean age 74.5F 7 (9 men,
42.8%; 12 women, 57.1%) who attended the Neurological
Clinic, Dept. of Oncology and Neuroscience, Chieti Univer-
sity, Chieti, Italy, were matched for age and habits with 10
HC (5 men, 50%; 5 women, 50%; mean age 75.5F 4). All
patients underwent clinical neuro-geriatric assessment and
routine laboratory analysis to exclude any possible influence
of other diseases which could be responsible for immune
activation. All patients were diagnosed with probable AD
according to the National Institute of Neurological and
Communicative Disorders and Stroke-Alzheimer’s Disease
and related Disorders Association (NINCDS-ADRDA) crite-
ria (McKhann et al., 1984). All patients were submitted to
the same research protocol and assessment of mental perfor-
mance and cognitive function was carried out using Fol-
stein’s Mini-Mental State Examination (MMSE) (Folstein et
al., 1975). Both patients and controls were not under med-
ication (especially analgesics and antibiotics) during the 2
weeks prior to testing and were asked not to use alcohol and
nicotine for at least 48 h before testing (Table 1). All patients
underwent early morning blood withdrawal before and after
1 month of AChEI treatment (Donepezil 10 mg/day, Pfeizer)
in the same hospital facility and by the same investigator. All
assays were run in parallel.
2.2. Cell separation and culture conditions
Blood samples for cytokine measurements were col-
lected into 4-ml peripheral blood mononuclear cells
(PBMC) endotoxin-free EDTA tubes (Vacutainer, Becton
Dickinson, NJ, USA). PBMC were isolated by Ficoll-
Hypaque density gradient centrifugation and resuspended
(5� 106/ml) in complete culture media consisting of
RPMI 1640 medium supplemented with 10% fetal calf
serum, 4 mM L-glutamine, 25 mM Hepes buffer, 50 U/ml

Fig. 1. Basal cytokine production (pg/ml/106 cells) in peripheral blood mononuclear cells cultures (incubated at 37 jC overnight) from AD patients (n= 21) and
age-matched HC (10). The cell-free supernatants were stored at � 80 jC until analysis. The ELISA values (in duplicates for each sample) have an error range
within 10%.
Fig. 2. Cytokine production (pg/ml/106 cells) after overnight stimulation with PHA mitogen in peripheral blood mononuclear cells cultures from AD patients
(n= 21) and age-matched healthy individuals (HC= 10). The ELISA values have an error range of within 10%.
M. Reale et al. / Journal of Neuroimmunology 148 (2004) 162–171164

M. Reale et al. / Journal of Neuroim
penicillin and 50 mg/ml streptomycin (all media and
components were purchased from Sigma, Italy). PBMC
were placed in polypropylene culture tubes (Bibby Sterilin
Italia, Italy) in a volume of 1 ml and cultured overnight,
at 37 jC in 95% humidified 5% CO2 cell culture in-
cubator, in complete culture medium, alone or stimulated
with PHA at 20 Ag/ml (Pharmacia, Sweden). Supernatants
were collected at the end of incubation and frozen at � 80
jC until assay. Pellet cells were also kept at � 80 jCuntil analysis. None of the reagents used contained endo-
toxin, as judged by Limulus amebocyte assay (minimum
detection level 0.1 Ag/ml) (E-Toxate).
Fig. 3. (A) RT-PCR shows IL-1h mRNA expression in PBMC isolated from AD pa
(T0) and from HC. Cells were stimulated with PHA (20 Ag/ml) overnight. The fi
expressed IL-1h mRNA levels relative to G3PDH mRNA. Date are meanF S.E.M
plots, where we have the data of IL-1h production (ELISA method) (basal and PH
distributed under the diagonal, that means after the treatment we have a reductio
2.3. Quantitation of chemokine and cytokine levels
Cytokine concentration in culture supernatants was
determined by a solid phase sandwich ELISA kit
(Endogen, Woburn, MA, USA) with monoclonal anti-
human IL-1h, IL-6, IL-4 and TNFa, according to the
manufacturers’ instructions. Samples from culture super-
natants were diluted in complete medium 1:10 for IL-1
h and IL-6, 1:2 for TNFa and undiluted for IL-4. All
steps were performed at room temperature. The color
reaction was stopped at 30 min and the OD was read at
450 nm within 30 min. Cytokine levels were then
munology 148 (2004) 162–171 165
tients after 1-month treatment with AChEI (T1), from untreated AD patients
gure shows one RT-PCR representative experiment. Densitometric analysis
. values for 21 (T0 and T1) to 10 (HC) individual samples. (B) In the scatter
A-stimulated) for untreated AD patients vs. treated AD patients, the data are
n in the quantities of these cytokines for all 21 AD patients.
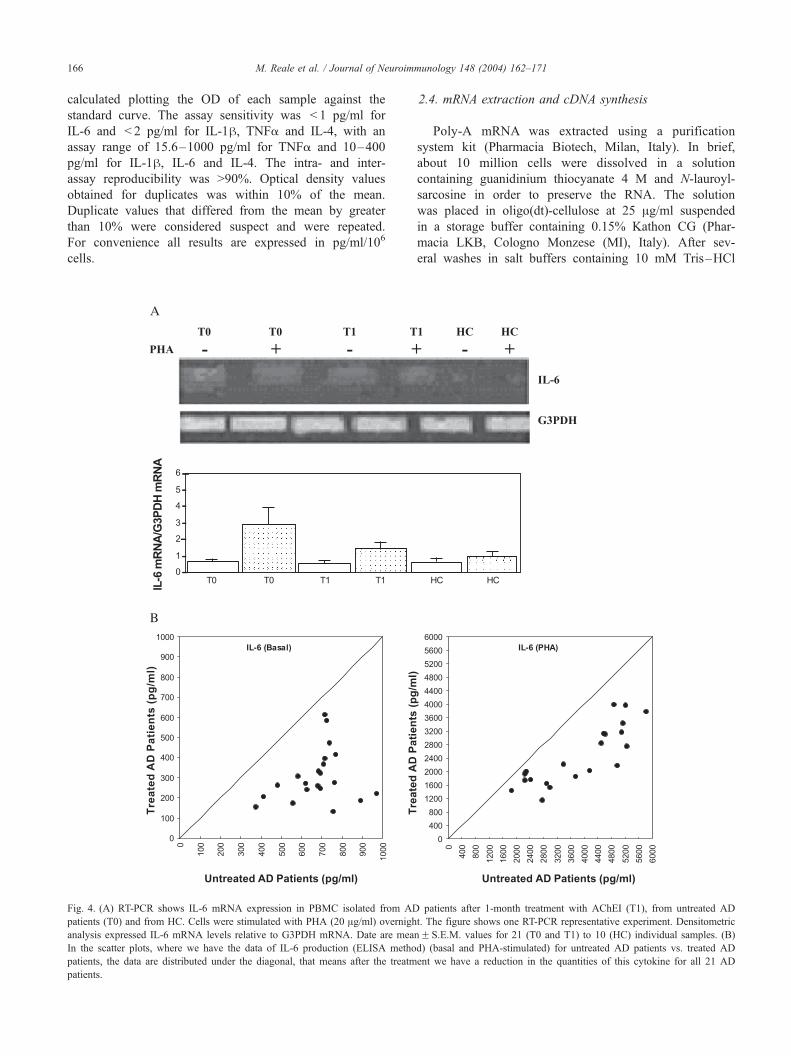
M. Reale et al. / Journal of Neuroim166
calculated plotting the OD of each sample against the
standard curve. The assay sensitivity was < 1 pg/ml for
IL-6 and < 2 pg/ml for IL-1h, TNFa and IL-4, with an
assay range of 15.6–1000 pg/ml for TNFa and 10–400
pg/ml for IL-1h, IL-6 and IL-4. The intra- and inter-
assay reproducibility was >90%. Optical density values
obtained for duplicates was within 10% of the mean.
Duplicate values that differed from the mean by greater
than 10% were considered suspect and were repeated.
For convenience all results are expressed in pg/ml/106
cells.
Fig. 4. (A) RT-PCR shows IL-6 mRNA expression in PBMC isolated from AD
patients (T0) and from HC. Cells were stimulated with PHA (20 Ag/ml) overnigh
analysis expressed IL-6 mRNA levels relative to G3PDH mRNA. Date are mea
In the scatter plots, where we have the data of IL-6 production (ELISA metho
patients, the data are distributed under the diagonal, that means after the treatm
patients.
2.4. mRNA extraction and cDNA synthesis
Poly-A mRNA was extracted using a purification
system kit (Pharmacia Biotech, Milan, Italy). In brief,
about 10 million cells were dissolved in a solution
containing guanidinium thiocyanate 4 M and N-lauroyl-
sarcosine in order to preserve the RNA. The solution
was placed in oligo(dt)-cellulose at 25 Ag/ml suspended
in a storage buffer containing 0.15% Kathon CG (Phar-
macia LKB, Cologno Monzese (MI), Italy). After sev-
eral washes in salt buffers containing 10 mM Tris–HCl
munology 148 (2004) 162–171
patients after 1-month treatment with AChEI (T1), from untreated AD
t. The figure shows one RT-PCR representative experiment. Densitometric
nF S.E.M. values for 21 (T0 and T1) to 10 (HC) individual samples. (B)
d) (basal and PHA-stimulated) for untreated AD patients vs. treated AD
ent we have a reduction in the quantities of this cytokine for all 21 AD

M. Reale et al. / Journal of Neuroim
(pH 7.4), 1 mM EDTA, 0.5 M NaCl or 0.1 M NaCl in
the last two washes, the oligo(dt)-cellulose containing
the mRNA was placed in filter columns and the mRNA
was eluted in warm Tris–HCl 10 mM and precipitated
in chilled 95% ethanol overnight. After centrifugation,
the pellet was dissolved in 14 Al of DEPC-treated
sterile water and quantitated by spectrophotometric anal-
ysis. A total of 0.5 Ag of mRNA was transcribed into
cDNA using 200 U of superscript reverse transcriptase
(GIBCO BRL, Milan, Italy) and 50 ng of Random
Examers.
Fig. 5. (A) RT-PCR shows TNFa mRNA expression in PBMC isolated from A
patients (T0) and from HC. Cells were stimulated with PHA (20 Ag/ml)
Densitometric analysis expressed TNFa mRNA levels relative to G3PDH mR
individual samples. (B) In the scatter plots, where we have the data of TNFa p
patients vs. treated AD patients, the data are distributed under the diagonal, tha
cytokine for all 21 AD patients.
2.5. Reverse transcriptase PCR amplification (RT-PCR)
cDNAwas amplified with 2.5 U Taq polymerase (Perkin
Elmer Cetus, Milan, Italy) using 1.5 pM of the primers
specific for IL-1h, IL-6, IL-4 and TNFa and G3PDH. IL-1,
IL-6, IL-4, TNFa and G3PDH primers were purchased from
Clonetech Laboratory (Palo Alto, CA, USA).
Each sample was divided in half, one part was used for
the cytokine under investigation and the other half for the
G3PDH for semi-quantitative analysis. RT-PCR was con-
ducted with the following protocol: (1) predenaturation at
munology 148 (2004) 162–171 167
D patients after 1-month treatment with AChEI (T1), from untreated AD
overnight. The figure shows one RT-PCR representative experiment.
NA. Date are meanF S.E.M. values for 21 (T0 and T1) to 10 (HC)
roduction (ELISA method) (basal and PHA-stimulated) for untreated AD
t means after the treatment we have a reduction in the quantities of this

M. Reale et al. / Journal of Neuroimmunology 148 (2004) 162–171168
94 jC for 5 min, (2) denaturation at 94 jC for 2 min, (3)
annealing at 55 jC for 2 min, (4) extension at 72 jC for 2
min and (5) denaturation at 94 jC for 2 min.
The linear range of signal strength for each cytokine was
determined by performing titration for cDNA and cycle
numbers to obtain non saturated PCR reactions.
Five microliters of amplified products were electropho-
retically separated in a 2% agarose gel containing ethidium
bromide and finally analyzed for molecular size.
The following controls were used: cDNA without pri-
mers and cDNA from peripheral blood mononuclear cells
from HC.
Signals were analyzed using the Bio-profile software
(Vilber Lourmat, Nice, France) and semi-quantitative anal-
ysis was performed comparing the amplified product signals
with the G3PDH signal.
2.6. Statistical analysis
Treated and untreated AD patients were analyzed by two-
sample paired Student’s t-test to determine whether sample’s
means are distinct. We also compared the mean of the
Fig. 6. (A) RT-PCR shows IL-4 mRNA expression in PBMC isolated from ACh
patients (T0 = no treatment) and from age-matched HC. Cells were incubated with
experiment. Densitometric analysis expressed IL-4 mRNA levels relative to G3PD
individual samples. (B) Illustrates the production of interleukin-4 (ELISA method)
of IL-4. The data are all distributed above the diagonal, meaning that after the trea
patients.
untreated AD patients and the HC, and the AD patients
treated and the HC.
The t-test is significant, either comparing the untreated
AD patients and the HC in basal and the AD patients treated
or the HC in PHA-stimulated PBMC cultures.
3. Results
3.1. Cytokine production in unstimulated PBMC
Cytokine production was examined directly in periph-
eral blood mononuclear cells cultures of HC subjects and
AD patients. Immunoreactive IL-1h, IL-6, TNFa and IL-
4 were measured in an attempt to determine more closely
any possible correlation between the in vitro productions
of cytokines in the HC vs. the AD group of subjects. In
the unstimulated PBMC of the AD group, there was a
significant increase in IL-1h, IL-6 and TNFa production
when compared with the HC group. On the contrary,
levels of IL-4 released by unstimulated PBMC did not
differ between AD patients and HC ( p = 0.427) (Fig. 1).
EI-treated AD patients after 1 month (T1 = 1 month), from untreated AD
or without PHA (20 Ag/ml) overnight. The figure shows one representative
H mRNA. Date are meanF S.E.M. values for 21 (T0 and T1) to 10 (HC)
in basal and PHA stimulation from PBMC. The scatterplot shows the results
tment we have an increase in the quantities of this cytokines for all 21 AD

uroimmunology 148 (2004) 162–171 169
3.2. Cytokine production by PBMC after PHA stimulation
The ability of PBMC to produce IL-1h, IL-6, TNFa and
IL-4 when exposed to PHA was evaluated. The PHA
mitogen was chosen for its ability to aspecifically stimulate
the release of these cytokines. The results show that cyto-
kine production in AD patients after stimulation with PHA
was higher than in their matched HC. The difference
between the levels was significant ( p < 0.001) for each
cytokine studied (Fig. 2).
3.3. IL-1� , IL-6 and TNF� expression and production by
PBMC in AChEI-treated, untreated AD patients and HC
To determine the effect of the in vivo AChEI-treat-
ment, cells from treated and untreated patients were
cultured overnight with or without 20 Ag/ml PHA and
than extracted for mRNA. As shown in the representative
experiments (Figs. 3A, 4A and 5A) RT-PCR amplification
demonstrates that the three cytokines are detectable in
small quantities in HC subjects, but are expressed in
greater quantities in AD untreated patients. Treatment
with AChEI reduces the expression of pro-inflammatory
cytokines in AD patients. The densitometric analysis
confirmed the decreases in TNFa, IL-1h and IL-6 mRNA
expression relative to G3PDH mRNA expression in
AChEI-treated AD patients.
The treatment with AChEI reduces the production of
pro-inflammatory cytokines in AD patients: levels of IL-
1h, IL-6 and TNFa were reduced in AD patients after 30
days of therapy with AChEI (Figs. 3B, 4B and 5B). In all
21 patients, the following percent decreases of the cyto-
kines were seen: IL-1h in basal conditions decreased
57%, in PHA-treated cells 54%, IL-6 decreased 62% in
basal conditions and 34% in PHA-treated cells, and TNFa
decreased 32% in basal conditions, 54% in PHA-treated
cells.
3.4. IL-4 expression and production by PBMC in Alzheim-
er’s patients receiving AChEI or not
The expression of IL-4 in AD patients, HC and AChEI-
treated AD patients is shown in Fig. 6. RT-PCR shows that
IL-4 mRNA expression is increased in AChEI-treated AD
patients respect to untreated AD patients. Determination of
IL-4 in cell-free supernatants harvested from cultures of
PBMC from AD patients treated with AChEI revealed
increased levels of biologically active IL-4, compared with
cultures of PBMC from untreated AD patients. In basal
conditions, IL-4 increase 197% after treatment; after PHA
stimulation, IL-4 increased 200%; and in both cases, the
values were higher than the HC. When we compared HC
subject with AChEI-treated AD patients, the spontaneous
release of IL-4 was higher in AChEI-treated group ( p <
0.001), while PHA-induced IL-4 production was not signif-
icantly different ( p= 0.601) (data not shown).
M. Reale et al. / Journal of Ne
4. Discussion
There is an emerging concept that the net biological
response of pro and anti-inflammatory cytokines affects
the outcome of certain diseases such as neurodegenerative
disorders (O’Shea and Lipsky, 2002; Sredni-Kenigsbuch,
2002). Modified production of cytokines, with conflicting
data on circulating serum levels of IL-1h, TNFa and IL-
6 in AD patients, have been reported (Fillit et al., 1991;
Kalman et al., 1997; Lanzrein et al., 1998; Engelborghs
et al., 1999; Licastro et al., 2000). Several studies have
addressed the issue of peripheral inflammatory molecules
as possible markers for AD and other dementia condi-
tions (Paganelli et al., 2002), but it is still an open issue.
Cytokines found in plasma and/or serum may be pro-
duced by blood cells, endothelium or may originate from
the brain. We hypothesize that the inflammatory condition
in AD patients, such as Ah accumulation, may influence
the PBMC and drive these cells to an ‘‘active state’’,
producing high levels of cytokines, these activated T cells
can cross the blood brain barrier (BBB) and contribute to
neurodegeneration (Yates et al., 2000; Szczepanik et al.,
2001b).
Evaluation of the ability of PBMC of AD patients to
produce cytokines when exposed to mitogenic stimuli
may clarify the role of these cells in active immune
response occurring in AD and if changes in the respon-
siveness of these cells are related to changes in circulating
cytokines.
Experiments designed to determine the constitutive and
PHA-inducible production, of IL-1h, IL-6, TNFa and IL-4
by PBMC from AD patients were performed both at the
mRNA and protein levels and provide evidence that PBMC
from AD release more IL-1h, IL-6, TNFa and less IL-4 than
PBMC from HC, and that such production is regulated at the
mRNA level. Our study supports the hypothesis of a
systemic production of cytokines, changes in circulating
cytokines may contribute to the pathophysiology of AD and
pro-inflammatory cytokines may affect neuronal function.
Cholinesterase inhibitors (ChEI) are used in AD patients
to maintain cognitive function at a constant level. Recent
studies have postulated the effect of ChEI on Ah metabo-
lism to explain the stabilising effect of the drugs. Cells
exposed to pro-inflammatory cytokines have elevated levels
of APP, which may result in increased production of Ah,further exacerbating plaque deposition, which is a crucial
event in pathophysiology of AD (Goldgaber et al., 1989;
Chao et al., 1995).
In order to gain insight in the potential benefits of oral
administration of AChEI in AD patients, we have examined
its role in immunological responses. The interesting results
that emerge from this study is that transcription and pro-
duction of pro-inflammatory cytokines in PBMC of AD
patients are markedly enhanced, with respect to HC sub-
jects, and IL-1h, IL-6 and TNFa over-expression and
production, are in accordance with other studies (i.e. Lom-

M. Reale et al. / Journal of Neuroimmunology 148 (2004) 162–171170
bardi et al., 1999) that reported an increase of different pro-
inflammatory cytokines, including TNFa, IL-6 and IL-1h,in AD patients. We hypothesize that increased production of
multiple cytokines from AD patients could produce a pre-
activation state in circulating PBMC.
Interestingly, we found that oral AChEI-treatment in AD
patients reduces expression and secretion of the pro-inflam-
matory cytokines IL-1h, IL-6 and TNFa and up-regulates
the expression and production of IL-4 under basal and
stimulated conditions in PBMC.
These data indicates that AChEI plays a role in the
regulation of immune response by reducing the production
of functionally important inflammatory cytokines that may
contribute to the immunopathology associated with AD, and
promoting the Th2 response, leading to the remodelling of
the cytokine network, probably acting on the lymphocytic
cholinergic system. Recently, it has been demonstrated that
in vitro IL-4 inhibit almost 100% the secretion of IL-1h and
TNF, while the secretion of IL-6 was found to be reduced
from 70% to 85%. Pre-treatment of co-culture cells with IL-
4, an immunosuppressive cytokine, prevented neuronal cell
injury induced by activated microglia, in a dose-dependent
manner (Ledeboer et al., 2000). Moreover, co-treatment of
stimulated cells with IL-4 blocked the increase levels of
TNFa and IL-1h, the inhibition was manifest also at the
mRNA level and hypothesized that IL-4 could be involved
and/or responsible for an aberrant regulation of cytokines
which are crucial in regulating brain and endocrine function
under normal physiological conditions (Hart et al., 1989; te
Velde et al., 1990). Indeed, recent investigations have
provided evidence that PBMC possess most of the essential
components needed to constitute (Chao et al., 1993) a
cholinergic system and that ACh synthesized and released
from PBMC acts as an immunomodulator (Kawashima and
Fujii, 2003, Tayebati et al., 2001).
In the light of these data, we suggest that the mecha-
nism(s) whereby AChEI reduces the level of inflammatory
cytokines probably is represented by the increased levels of
IL-4. The mechanism by which IL-4 exerts its neuropro-
tective effect was found to involve the inhibition of IFNa
priming of microglia with a subsequent decrease in the
production of TNFa and nitric oxide (Chao et al., 1995).
These findings may have implications for the control of the
pathogenic mechanisms in neurodegeneration of Alzheim-
er’s disease and provide a basis for future studies related to
systemic cytokines and AD.
Acknowledgements
Thanks are due to Renato Barbacane for his technical
skills. This work was supported by MURST grants, prot.
CE00538159 ‘‘Biological and immunological markers in
neurodegenerative disorders’’ from the Ministry of Univer-
sity, Scientific and Technological Research, Italy for the
year 2000.
References
Abbas, N., Bednar, I., Mix, E., Marie, S., Paterson, D., Ljungberg, A.,
Morris, C., 2002. Up-regulation of the inflammatory cytokines IFN-g
and IL-12 and down-regulation of IL-4 in cerebral cortex regions of
APPSWE transgenic mice. J. Neuroimmunol. 126, 50–57.
Benveniste, E.N., 1992. Inflammatory cytokines within the central nervous
system: source, function, and mechanism of action. Am. J. Physiol. 363,
C1–C16.
Bruunsgaard, H., Pedersen, B.K., 2003. Age-related inflammatory cyto-
kines and disease. Immunol. Allergy Clin. North Am. 23, 15–39.
Bruunsgaard, H., Andersen-Ranberg, K., Jeune, B., Pedersen, A.N., Skin-
hoj, P., Pedersen, B.K., 1999. A high plasma concentration of TNF-
alpha is associated with dementia in centenarians. J. Gerontol., Ser. A,
Biol. Sci. Med. Sci. 54, M357–M364.
Chao, C.C., Molitor, T.W., Hu, S., 1993. Neuroprotective role of IL-4
against activated microglia. J. Immunol. 151, 1473–1481.
Chao, C.C., Hu, S., Ehrlich, L., Peterson, P.K., 1995. Interleukin-1 and
tumour necrosis factor-alpha synergistically mediate neurotoxicity: in-
volvement of nitric oxide and of N-methyl-D-aspartate receptors. Brain
Behav. Immun. 9, 355–365.
Conti, P., Dempsey, R.A., 1990. Interleukin-4 and interleukin-1 receptor
antagonist down-regulate interleukin-1 and tumour necrosis factor
(TNF) generation in macrophages. Int. J. Immunopathol. Pharmacol.
2, I – II.
Del Bo, R., Angeretti, N., Lucca, E., De Simoni, M.G., Forloni, G., 1995.
Reciprocal control of inflammatory cytokines, IL-1 and IL-6, and h-amyloid production in cultures. Neurosci. Lett. 188, 70–74.
De Luca, G., Lugaresi, A., Iarlori, C., Marzoli, F., Uncini, A., Gambi, D.,
1998. Interferon beta normalizes suppressor cell function in dysimmune
neuropathies. J. Neuroimmunol. 82, 1–4.
Dinarello, C.A., 1997. Role of pro- and anti-inflammatory cytokines during
inflammation: experimental and clinical findings. J. Biol. Regul. Ho-
meost. Agents 11, 91–103.
Engelborghs, S., De Brabander, M., De Cree, J., D’Hooge, R., Geerts, H.,
Verhaegen, H., De Deyn, P.P., 1999. Unchanged levels of interleukins,
neopterin, interferon-gamma and tumour necrosis factor-alpha in cere-
brospinal fluid of patients with dementia of the Alzheimer type. Neuro-
chem. Int. 34, 523–530.
Fillit, H., Ding, W., Buee, L., Kalman, J., Altstiel, L., Lawlor, B., Wolf-
Klein, G., 1991. Elevated circulating tumour necrosis factor levels in
Alzheimer’s disease. Neurosci. Lett. 129, 318–320.
Folstein, M.F., Folstein, S.E., McHug, P.R., 1975. Mini-mental state: a
practical methods for grading the cognitive state of patients for the
clinician. J. Psychiatr. Res. 12, 189–198.
Gerli, R., Monti, D., Bistoni, O., Mazzone, A.M., Peri, G., Cossarizza, A.,
Di Gioacchino, M., Cesarotti, M.E., Doni, A., Mantovani, A., France-
schi, C., Paganelli, R., 2000. Chemokines, sTNF-Rs and sCD30 serum
levels in healthy aged people and centenarians. Mech. Ageing Dev. 121,
37–46.
Giacobini, E., 2001. Is anti-cholinesterase therapy of Alzheimer’s disease
delaying progression? Aging 13, 247–254.
Goldgaber, D., Harris, H.W., Hla, T., Maciag, T., Donnelly, R.J., Jacobsen,
J.S., Vitek, M.P., Gajdusek, D.C., 1989. Interleukin 1 regulates syn-
thesis of amyloid beta-protein precursor mRNA in human endothelial
cells. Proc. Natl. Acad. Sci. U. S. A. 86, 7606–7610.
Griffin, W.S.T., Sheng, J.G., Roberts, G.W., Mrak, R.E., 1995. Interleukin-
1 expression in different plaque types in AD: significance in plaque
evolution. J. Neuropathol. Exp. Neurol. 54, 276–281.
Hart, P.H., Vitti, G.F., Buegess, D.R., Whitty, G.A., Piccoli, D.S., Hamilton,
J.A., 1989. Potential anti-inflammatory effects of interleukin 4: suppres-
sion of human monocyte tumour necrosis factor alpha, interleukin 1 and
prostaglandin E2. Proc. Natl. Acad. Sci. U. S. A. 86, 3803–3807.
Huell, M., Strauss, S., Volk, B., Berger, M., Bauer, J., 1995. Interleukin-6 is
present in early stage of plaque formation and is restricted to the brains of
Alzheimer’s disease patients. Acta Neuropathol. (Berl.) 89, 544–551.

M. Reale et al. / Journal of Neuroimmunology 148 (2004) 162–171 171
Iarlori, C., Reale, M., Lugaresi, A., De Luca, G., Bonanni, L., Di Iorio,
C., Feliciani, C., Conti, P., Gambi, D., 2000. RANTES production
and expression is reduced in relapsing-remitting multiple sclerosis
patients treated with interferon-beta-1b. J. Neuroimmunol. 107,
100–107.
Iarlori, C., Reale, M., De Luca, G., Di Iorio, A., Feliciani, C., Tulli, A.,
Conti, P., Gambi, D., Lugaresi, A., 2002. Interferon beta-1h, modulates
MCP-1 expression and production in relapsing-remitting multiple scle-
rosis. J. Neuroimmunol. 123, 170–179.
Kalman, J., Juhasz, A., Laird, G., Dickens, P., Jardanhazy, T., Rimanoczy
Boncz, I., Parry-Jones, W.L., Janka, Z., 1997. Serum interleukin-6 lev-
els correlate with the severity of dementia in Down syndrome and
Alzheimer’s disease. Acta Neurol. Scand. 96, 236–240.
Kawashima, K., Fujii, T., 2003. The lymphocytic cholinergic system and its
biological function. Life Sci. 72, 2101–2109.
Lanzrein, A.S., Johnston, C.M., Perry, V.H., Jobst, K.A., King, E.M.,
Smith, A.D., 1998. Longitudinal study of inflammatory factors in se-
rum, cerebrospinal fluid, and brain tissue in Alzheimer disease: inter-
leukin-1beta, interleukin-6, interleukin-1 receptor antagonist, tumour
necrosis factor-alpha, the soluble tumour necrosis factor receptors I
and II, and alpha1-antichymotrypsin. Alzheimer Dis. Assoc. Disord.
12, 215–227.
Ledeboer, A., Breve, J.J., Poole, S., Tilders, F.J., Van Dam, A.M., 2000.
Interleukin-10, interleukin-4, and transforming growth factor-beta dif-
ferentially regulate lipopolysaccharide-induced production of pro-in-
flammatory cytokines and nitric oxide in co-cultures of rat astroglial
and microglial cells. Glia 30, 134–142.
Lee, J.D., Rhoades, K., Economou, J.S., 1995. Interleukin-4 inhibits
the expression of tumour necrosis factors alpha and beta, interleu-
kins-1 beta and -6 and interferon-gamma. Immunol. Cell Biol. 73,
57–61.
Licastro, F., Pedrini, S., Caputo, L., Annoni, G., Davis, L.J., Ferri, C.,
Casadei, V., Grimaldi, L.M., 2000. Increased plasma levels of interleu-
kin-1, interleukin-6 and alpha-1-antichymotrypsin in patients with Alz-
heimer’s disease: peripheral inflammation or signals from the brain?
J. Neuroimmunol. 103, 97–102.
Lombardi, V.R.M., Garcia, M., Rey, L., Cacabelos, R., 1999. Character-
ization of cytokine production, screening of lymphocyte subset patterns
and in vitro apoptosis in healthy an Alzheimer disease (AD) individuals.
J. Neuroimmunol. 97, 163–171.
Maat-Schieman, M.L., van Duinen, S.G., Rozemuller, A.J., Haan, J., Ross,
R.A., 1997. Association of vascular amyloid beta and cells of the
mononuclear phagocyte system in hereditary cerebral hemorrhage with
amyloidosis (Dutch) and Alzheimer disease. J. Neuropathol. Exp. Neu-
rol. 56, 273–284.
McKhann, G., Drachman, D., Folstein, M., Katzman, R., Price, D., Stadlan,
E.M., 1984. Clinical diagnosis of Alzheimer’s disease: report of
NINCDS-ADRDA work group. Neurology 34, 939–944.
Meda, L., Baron, P., Prat, E., Scarpini, E., Scarlato, G., Cassatela, M.A.,
Rossi, F., 1999. Proinflammatory profile of cytokine production by
human monocytes and murine microglia stimulated with h-amyloid
[25–35]. J. Neuroimmunol. 93, 45–52.
O’Mahony, L., Holland, J., Jackson, J., Feighery, C., Hennessy, T.P.,
Mealy, K., 1998. Quantitative intracellular cytokine measurement:
age-related changes in proinflammatory cytokine production. Clin.
Exp. Immunol. 113, 213–219.
O’Shea, J.J., Ma, A., Lipsky, P., 2002. Cytokines and autoimmunity. Nat.
Rev., Immunol. 2, 37–45.
Paganelli, R., Di Iorio, A., Patricelli, L., Ripani, F., Sparvieri, E., Faricelli,
R., Iarlori, C., Porreca, E., Di Gioacchino, M., Abate, G., 2002. Proin-
flammatory cytokines in sera of elderly patients with dementia: levels in
vascular injury are higher than those of mild-moderate Alzheimer’s
disease patients. Exp. Gerontol. 37, 257–263.
Perry, R.T., Collins, J.S., Wiener, H., Acton, R., Go, R.C.P., 2001. The role
of TNF and its receptors in Alzheimer’s disease. Neurobiol. Aging 22,
873–883.
Porrini, A.M., De Luca, G., Gambi, D., Reder, A.T., 1998. Effects of an
anti-IL-10 monoclonal antibody on rIFNbeta-1b-mediated immune
modulation. Relevance to multiple sclerosis. J. Neuroimmunol. 81,
109–115.
Rossi, F., Bianchini, E., 1996. Synergistic induction of nitric oxide by h-amyloid and cytokines in astrocytes. Biochem. Biophys. Res. Commun.
225, 474–478.
Rothwell, N.J., Hopkins, S.J., 1995. Cytokines and the nervous system: II.
Actions and mechanisms of action. Trends Neurosci. 18, 130–136.
Sheng, J.G., Ito, K., Skinner, R.D., Mrak, R.E., Rovnaghi, C.R., van Eldik,
L.J., Griffin, W.S.T., 1996. In vivo, and in vitro evidence supporting a
role for the inflammatory cytokine interleukin-1 as a driving force in
Alzheimer pathogenesis. Neurobiol. Aging 17, 761–766.
Shibata, N., Ohnuma, T., Takahashi, T., Baba, H., Ishizuka, T., Ohtsuka,
M., Ueki, A., Nagao, M., Arai, H., 2002. Effect of IL-6 polymorphism
on risk of Alzheimer disease: genotype-phenotype association study in
Japanese cases. Am. J. Med. Genet. 114, 436–439.
Sredni-Kenigsbuch, D., 2002. TH1/TH2 cytokines in the central nervous
system. Int. J. Neurosci. 112, 665–703.
Szcepanik, A.M., Funes, S., Petko, W., Ringheim, G.E., 2001a. IL-4, IL-10
and IL-13 modulate A-beta(1–42)-induced cytokine and chemokines
production in primary murine microglia and a human cell line. J. Neuro-
immunol. 113, 49–62.
Szczepanik, A.M., Rampe, D., Ringheim, G.E., 2001b. Amyloid-beta pep-
tide fragment p3 and p4 induce pro-inflammatory cytokine and chemo-
kine production in vitro and in vivo. J. Neurochem. 77, 304–317.
Tayebati, S.K., Amenta, F., Amici, S., El-Assouad, D., Gallai, V., Ricci, A.,
Parnetti, L., 2001. Peripheral blood lymphocytes muscarinic cholinergic
receptor subtypes in Alzheimer’s disease: a marker of cholinergic dys-
function? J. Neuroimmunol. 121, 126–131.
te Velde, A.A., Huijbens, R.J., Heije, K., de Vries, J.E., Figdor, C.G.,
1990. Interleukin-4 (IL-4) inhibits secretion of IL-1beta, tumour
necrosis factor alpha, and IL-6 by human monocytes. Blood 76,
1392–1397.
Winblad, B., Engedal, K., Soininen, H., 2001. A 1-year, randomized, pla-
cebo-controlled study of Donepezil in patients with mild to moderate
AD. Neurology 57, 489–495.
Yates, S.L., Burgess, L.H., Kocsis-Angle, J., Antal, J.M., Dority, M.D.,
Embury, P.B., Piotrkowski, A.M., Brunden, K.R., 2000. Amyloid beta
and amylin fibrils induce increases in proinflammatory cytokine and
chemokine production by THP-1 cells and murine microglia. J. Neuro-
chem. 74, 1017–1025.
![Inflammatory Cytokines Induce DNA damage and …...[CANCER RESEARCH 60, 184–190, January 1, 2000] Inflammatory Cytokines Induce DNA damage and Inhibit DNA repair in Cholangiocarcinoma](https://img.dokumen.tips/doc/110x75/5e9271986a3d48445f29f8e8/inflammatory-cytokines-induce-dna-damage-and-cancer-research-60-184a190.jpg)













![RadiationInhibitsInterleukin-12Production viaInhibitionofC ...ir.ymlib.yonsei.ac.kr/bitstream/22282913/146674/1/T201601065.pdf · inflammatory cytokines and chemokines [1–3].In](https://img.dokumen.tips/doc/110x75/5ec96bd5adb7620946361fb8/radiationinhibitsinterleukin-12production-viainhibitionofc-irymlib-inflammatory.jpg)














