Embed Size (px)
Citation preview

Transgenerational Epigenetic Inheritance: Prevalence, Mechanisms, and Implications for theStudy of Heredity and EvolutionAuthor(s): Eva Jablonka and Gal RazSource: The Quarterly Review of Biology, Vol. 84, No. 2 (June 2009), pp. 131-176Published by: The University of Chicago PressStable URL: http://www.jstor.org/stable/10.1086/598822 .Accessed: 11/01/2011 00:48
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at .http://www.jstor.org/action/showPublisher?publisherCode=ucpress. .
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access to TheQuarterly Review of Biology.
http://www.jstor.org

TRANSGENERATIONAL EPIGENETIC INHERITANCE:PREVALENCE, MECHANISMS, AND IMPLICATIONS FOR THE
STUDY OF HEREDITY AND EVOLUTION
Eva JablonkaThe Cohn Institute for the History and Philosophy of Science and Ideas, Tel-Aviv University, Tel-Aviv
69978, Israel
e-mail: [email protected]
Gal RazThe Graduate School of Medicine, Tel-Aviv University, Tel-Aviv 69978, Israel
e-mail: [email protected]
keywordscell memory, epigenetics, induced heritable variations, Lamarckism,
microevolution, macroevolution
abstractThis review describes new developments in the study of transgenerational epigenetic inheritance, a
component of epigenetics. We start by examining the basic concepts of the field and the mechanismsthat underlie epigenetic inheritance. We present a comprehensive review of transgenerational cellularepigenetic inheritance among different taxa in the form of a table, and discuss the data containedtherein. The analysis of these data shows that epigenetic inheritance is ubiquitous and suggests linesof research that go beyond present approaches to the subject. We conclude by exploring some of theconsequences of epigenetic inheritance for the study of evolution, while also pointing to the importanceof recognizing and understanding epigenetic inheritance for practical and theoretical issues in biology.
Definitions: Epigenetics andEpigenetic Inheritance
EPIGENETICS has become one of thebuzz words of biology in recent years.
Following the success of genome projectsin defining what genomes are, the empha-sis has shifted to what they do, and there isrenewed interest in understanding the epi-genetic processes of development. Theterm “epigenetics,” however, has under-
gone many transformations since its origi-nal definition by Waddington (see Wad-dington 1968 for a discussion), reflectingthe changing foci of research in develop-mental molecular biology since the secondhalf of the 20th century (Jablonka andLamb 2002, 2007c; Haig 2004; Holliday2006). “Epigenetics” is therefore often em-ployed loosely and inconsistently, and issometimes used as a synonym for “epige-
The Quarterly Review of Biology, June 2009, Vol. 84, No. 2
Copyright © 2009 by The University of Chicago Press. All rights reserved.
0033-5770/2009/8402-0001$15.00
Volume 84, No. 2 June 2009THE QUARTERLY REVIEW OF BIOLOGY
131

netic inheritance.” To avoid misunder-standings, we define both terms as they areused in this review:
Epigenetics is the study of the processesthat underlie developmental plasticity andcanalization and that bring about persistentdevelopmental effects in both prokaryotesand eukaryotes. At the cellular level, theseare the processes involved in cell determina-tion and differentiation. At higher levels ofbiological organization, epigenetic mecha-nisms generate the context-dependent, self-sustaining interactions between groups ofcells that lead to physiological and morpho-logical persistence. The regulatory mecha-nisms that establish and maintain variant cel-lular and organismal states are known asepigenetic control mechanisms, or epigenetic con-trol systems (Nanney 1958).
Epigenetic inheritance is a componentof epigenetics. It occurs when phenotypicvariations that do not stem from variationsin DNA base sequences are transmitted tosubsequent generations of cells or organ-isms. Many of the discoveries about epige-netic inheritance between organisms arederived from studies in developmental bi-ology that look at inheritance in cell lin-eages within an organism. Cell heredity inmitotically dividing cells underlies the per-sistence of determined states in multicellu-lar organisms. That is, an individual’s kid-ney stem cells and skin stem cells generallybreed true, even though their DNA se-quences are identical and the developmen-tal stimuli that led to the different cellphenotypes are long gone. However, thesame cell heredity mechanisms that havebeen found in cell lineages during develop-ment are also observed when epigenetic in-heritance occurs between generations of in-dividuals. In single-celled organisms, such asbacteria and asexually reproducing protists,epigenetic inheritance leads to the clonalpersistence of induced and stochasticallygenerated phenotypic variations. In sexuallyreproducing organisms, heritable epigeneticvariations in germline cells can result in thetransmission of developmentally inducedand stochastically generated phenotypesfrom one generation of individuals to thenext through the gametes. Like “epigenet-
ics,” “epigenetic inheritance” is not alwaysconsistently employed. It is used in both abroad and a narrow sense.
Epigenetic inheritance in the broad sense is theinheritance of developmental variationsthat do not stem from differences in thesequence of DNA or from persistent induc-ing signals in the present environment. Aswell as cell-to-cell transmission of epige-netic variations in unicellular and multicel-lular organisms (as will be explained be-low), the definition covers body-to-body (orsoma-to-soma) information transference thatcan take place through developmental inter-actions between mother and offspring (e.g.,Weaver et al. 2004), through social learning(Avital and Jablonka 2000), and throughsymbolic communication (Richerson andBoyd 2005).
Cellular epigenetic inheritance is a narroweraspect of epigenetic inheritance as dis-cussed in the broad sense. It refers to epi-genetic transmission in sexual or asexualcell lineages, and the unit of this trans-mission is the cell. Following Holliday(1994, 2002, 2006), some biologists restrictthe term epigenetic inheritance solely tothe transmission of chromatin marks andRNAs (e.g., Wu and Morris 2001). How-ever, yeast geneticists use the term epige-netic inheritance to refer to the inheri-tance of protein conformations, such asprions (e.g., Uptain and Lindquist 2001),and the term is also used by biologistsstudying self-sustaining loops and chroma-tin inheritance in bacteria (e.g., Grandjeanet al. 1998; Laurent et al. 2005). We there-fore define cellular epigenetic inheritanceas the transmission from mother cell todaughter cell of variations that are not theresult of differences in DNA base sequenceand/or the present environment. Trans-mission can be through chromatin marks,through RNAs, through self-reconstructingthree-dimensional structures, and throughself-sustaining metabolic loops (Jablonkaet al. 1992; Jablonka and Lamb 1995, 2005,2007a). It occurs following cell division inprokaryotes, mitotic cell division in thesoma of eukaryotes, and sometimes follow-ing the meiotic divisions in the germline.The chromatin and RNA-mediated cellularinheritance systems seem to play an impor-tant role in inheritance through the germ-line in both females and males.
132 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

In Figure 1, we illustrate the differencebetween the broad and narrow sense ofepigenetic inheritance by showing themain routes of between-generation trans-mission in a sexually reproducing multicel-lular organism. As our focus in this reviewis on cellular transgenerational epigenetic inher-itance, we are concentrating on the route ofbetween-generation transmission that in-volves a single-cell “bottleneck,” i.e., trans-mission through a gamete or a spore inmulticellular, sexually reproducing organ-isms, or through a single sexual or asexualcell in unicellular organisms. The environ-ment may induce epigenetic variation bydirectly affecting the germline or by affect-ing germ cells through the mediation ofthe soma, but, in either case, subsequenttransmission is through the germline. Al-though the direct soma-to-soma transmis-sion route of epigenetic variations is ofgreat importance for both developmentand evolution (Jablonka and Lamb 2005,2007a,b), its discussion is beyond the scopeof this review.
Mechanisms of Cellular EpigeneticInheritance (EISs)
Jablonka and Lamb (1989, 1995; Jablonka etal. 1992) suggested that the different mecha-nisms of epigenetic inheritance should beunderstood and studied within a shared evo-lutionary framework that incorporates thedevelopmental construction of heredity andthat acknowledges the Lamarckian aspects ofheredity and evolution. They called the pro-cesses and mechanisms that underlie cellularinheritance “epigenetic inheritance systems”(abbreviated to EISs by Maynard Smith[1990]). Cell heredity may occur when aninduced gene-product is diluted very slowlyby cell divisions, so that its concentrationremains above the threshold required for itsactivity for several cell generations (Zachari-oudakis et al. 2007), but such “memory” isshort-term. For more persistent memory andcell heredity, autocatalysis is necessary, andall the EISs we describe depend on mecha-nisms that enable self-perpetuation. Fourtypes of cellular EISs are recognized today:the EIS based on self-sustaining regulatoryloops, the EIS based on three-dimensional Fi
gu
re1.
Ro
ute
so
fT
ran
smis
sio
no
fE
pig
enet
icV
aria
tio
ns
ina
Mu
ltic
ellu
lar,
Sex
ual
lyR
epro
du
cin
gO
rgan
ism
Rou
tea
show
sth
ege
rmlin
e-to
-ger
mlin
etr
ansm
issi
onof
indu
ced
epig
enet
icva
riat
ion
s(e
.g.,
chro
mat
inm
arks
).A
vari
atio
nca
nbe
indu
ced
inth
ege
rmlin
ean
dca
nth
enbe
tran
smit
ted
from
one
gen
erat
ion
toth
en
ext,
orit
can
firs
tbe
indu
ced
inth
eso
ma,
then
affe
ctth
ege
rmlin
e,an
dth
erea
fter
bein
her
ited
thro
ugh
the
germ
line.
Rou
teb
show
sso
ma-
to-s
oma
tran
smis
sion
(for
exam
ple,
thro
ugh
the
tran
smis
sion
ofsy
mbi
onts
and
para
site
s,or
thro
ugh
the
self
-per
petu
atin
gef
fect
sof
mat
ern
albe
hav
ior,
soci
alle
arn
ing,
and
sym
bolic
com
mun
icat
ion
).A
broa
dvi
ewof
epig
enet
icin
her
itan
ceen
com
pass
esbo
thro
utes
aan
db,
wh
erea
sth
en
arro
wer
,ce
llula
rvi
ewin
clud
eson
lyro
ute
a—tr
ansm
issi
onth
roug
ha
sin
gle-
cell
“bot
tlen
eck,
”in
this
case
,a
gam
ete.
June 2009 133TRANSGENERATIONAL EPIGENETIC INHERITANCE

templating, the chromatin-marking EIS, andthe RNA-mediated EIS. Although these EISsare usually seen as very different types of de-velopmental mechanisms, all can contribute tobetween-generation epigenetic inheritance.Their dual nature as developmental mecha-nisms and as inheritance systems means thatthey can be studied from both perspectives.The developmental perspective is currentlythe dominant one, and the molecular basisof epigenetic control systems is one of themost intensely studied fields in biology today(e.g., see collections of articles in Nature Re-views Genetics 8(4) [Flintoft 2007], Nature447(7143 Insight) [Campbell 2007], and Cell128(4) [Marcus 2007], as well as Allis et al.2007). In this review, we focus on the lessdiscussed aspect of EISs and epigenetic in-heritance—the transmission of cellular epi-genetic states from one generation of organ-isms to the next. We therefore describe thefour EISs from a heredity-focused point ofview. All the examples to which we refer, aswell as many others, are presented here inTable 1 and in our expanded table (availableonline, with accompanying table references,at The Quarterly Review of Biology homepage,www.journals.uchicago.edu/QRB).
inheritance through self-sustainingloops
Self-sustaining feedback loops are metaboliccircuits through which different patternsof activity can be maintained, resulting inalternative heritable cell phenotypes. Thefirst experimental studies of such loopswere those involving the bistability of thelac operon of Escherichia coli (Novick andWeiner 1957), and this system has sincebeen thoroughly analyzed at both the mo-lecular and theoretical levels (Laurentet al. 2005). These studies show that, wheninducer concentrations are low, geneticallyidentical cells can generate two alternative,true-breeding, stable phenotypes.
Many other self-sustaining feedback loopsleading to alternative heritable phenotypeshave been described in bacteria and othertaxa (Dubnau and Losick 2006; Smits et al.2006; Malagnac and Silar 2003, 2006). Onewell-characterized example of positive regu-lation leading to alternative cell phenotypes
is found in the fungus Candida albicans,where an epigenetic switch underlies thetransition between white and opaquecells—two states that are heritable for manygenerations (Zordan et al. 2006). Self-sustaining loops need not be based on tran-scriptional regulation; they can also occur atthe post-translation stage, through proteinself-processing. An example is the enzymevacuolar protease B of Saccharomyces cerevisiae,whose active form is necessary for its ownconversion from an inactive precursor to anactive state. On glycerol media, where theprecursor is synthesized in high amounts,the self-processing of the enzyme is indefi-nitely self-sustaining (Roberts and Wickner2003). As noted by Wickner et al. (2004), itis likely that other protein-modifying pro-teins may also directly or indirectly affecttheir own modification and behave as self-sustaining, cell-transmissible loops.
structural inheritance:spatial templating
Structural inheritance refers to the inher-itance of alternative three-dimensional(3-D) structures through spatial templat-ing: a variant 3-D structure in a mother cellguides the formation of a similar structurefor a daughter cell, leading to the transmis-sion of the architectural variant (Nanney1968). The study of cellular structural in-heritance was initiated by investigations ofthe inheritance of cortical variations inciliates. Beisson and Sonneborn (1965)showed that an experimentally modifiedorganization of the cilia on Paramecium canbe inherited through many asexual andsexual generations. The inheritance of cor-tical variations induced by various physicaland chemical manipulations has also beendemonstrated for other ciliates (Nanney1985; Frankel 1989; Grimes and Aufder-heide 1991).
The propagation of prions is anotherform of structural inheritance. The funda-mental characteristic of a prion—a trans-missible protein—is that it has a conforma-tion that can initiate and sustain the re-production of a similar conformation innewly synthesized proteins. Prusiner (1998)suggested that infectious proteins are the
134 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

causative agents of mammalian neurode-generative diseases such as kuru, scrapie,and mad cow disease and coined the termprion (proteineceous infectious particles)for such agents. He advanced and devel-oped the concept of an infectious proteinwhose propagation is based on the spatial-templating of the variant prion conforma-tion, which, in turn, converts a normally-shaped protein into its own shape. Theconcept was extended when Wickner (1994)characterized the [URE3] variant in yeastas a prion and explained its idiosyncraticbiochemical and hereditary characteristicsin terms of protein-based inheritance.Since then, more prions have been identi-fied in yeast and other fungi (Baxa et al.2006; Tuite and Cox 2006). Although someprions have deleterious effects, othersseem to have important biological func-tions (Wickner et al. 2004; Shorter andLindquist 2005; Benkemoun and Saupe2006).
One of the most interesting findingsabout prions is that a single protein canmisfold into several different conforma-tions that have specific growth dynamics,stabilities, pathologies, and cross-speciesinfectivity (Chien et al. 2004). Differentprions may interact, leading to the forma-tion of many different transmissible (cell-heritable and infectious) phenotypes. There-fore, unicellular organisms, which have thesame genotype and live in the same en-vironment, can exhibit heritably differ-ent morphologies and physiologies thatare the consequence of differently foldedidentical proteins. The differences resultfrom differences in the developmentalhistories of their ancestors.
Cavalier-Smith (2004) studied anotheraspect of spatial templating, an aspectassociated with membrane reproduction.The reproduction of most membranes, in-cluding the plasma membrane, the endo-plasmic reticulum, and the mitochondrialmembrane, requires the presence and tem-plating of pre-existing membrane struc-tures. Cavalier-Smith identified 18 types ofwhat he called “genetic membranes,” thestructures of which are cell-heritablethrough these guided processes, and he
argued that the information embedded inthis “membranome” is as essential for theconstruction of a cell as genomic informa-tion. He suggested that crucial events inthe evolution of cells and major groupswere associated with heritable changes inmembranomes.
the chromatin-marking EISChromatin, the stuff of chromosomes,
includes DNA and all the factors physicallyassociated with it: small chemical groupscovalently attached to DNA (e.g., methylgroups), bound histone and nonhistoneproteins, and associated RNA molecules.The organization of chromatin and chro-mosomes, their localization in the nucleus,and the dynamic interactions among thevarious components of chromatin deter-mine when, where, and to what extentgenes are transcribed, how DNA repair isorchestrated, how different chromosomaldomains are organized, and how chromo-somes, as units, behave during the vari-ous phases of the cell cycle. Patterns ofchromatin are reconstructed followingDNA transcription and replication, and, al-though the processes of reconstruction arenot well understood, there is evidence thatchromatin variations can be transmittedbetween generations of individuals. Thestudy of the chromatin-marking EIS istherefore crucial for the understanding ofboth development and heredity.
DNA methylation, the best-understoodsystem of chromatin inheritance, is an epi-genetic modification found in Eubacteria,Archeabacteria, and Eukaryota. It is in-volved in many important functions (forgeneral reviews see Casadesus and Low2006; Vanyushin 2006a,b), including de-fence against genomic parasites (Kidwelland Lisch 1997), regulation and mainte-nance of gene activity patterns (Barlow andBartolomei 2007; Li and Bird 2007), stabi-lization of chromosomal structure (Karpenand Hawley 2007), and DNA replicationand repair (Mortusewicz et al. 2005; Scher-melleh et al. 2007). In eukaryotes, methyl-ation usually occurs on the cytosines inCpG doublets and also in CpNpG tripletsin plants. Since CpG and CpNpG se-
June 2009 135TRANSGENERATIONAL EPIGENETIC INHERITANCE

quences are palindromic, the two strandsof the duplex are mirror images of eachother. Following replication, two hemi-methylated duplexes are formed, andthese hemimethylated regions are recog-nized by maintenance DNA methylases,which preferentially add methyls to non-methylated C in the new strand (Allshireand Selker 2007; Henderson and Jacobsen2007; Li and Bird 2007). The replication ofmethylation patterns is thus semi-conservative.The fidelity of transmission in cell lineagesranges from about 1% variation per cellgeneration up to the very high fidelity of10-6 variations per cell generation (Gene-reux et al. 2005; Richards 2006).
Variations in a DNA methylation patterncan also be inherited between generations,and examples of this include paramuta-tions in plants (interaction between allelesthat leads to a directed epigenetic herita-ble change in one of the interacting alleles[Brink 1973; Chandler 2007]); silencingof foreign duplicated sequences in fungi(Allshire and Selker 2007); variations intelomeric, centromeric, and rRNA regions(Karpen and Hawley 2007); and variationsin transgenes and endogenous genes. Spe-cific examples of all these types of trans-generational inheritance are presented inTable 1.
How a DNA methylation pattern affectsa cell’s or an organism’s phenotype de-pends on the way it interacts with the pro-tein components of the chromosome,which are also heritable. In eukaryotes, im-portant variations in chromatin are associ-ated with histones, the proteins that makeup the nucleosome core around whichDNA is wrapped. The dynamic nature ofhistones, their variability, and their associ-ation with every conceivable cellular func-tion (see Berger 2007; Groth et al. 2007;Blasco 2007; Morris and Moazed 2007)makes understanding the inheritance oftheir specific structure and organizationboth challenging and urgent. Several mod-els of this process have been constructed,and according to all of them, the nucleo-some variants and the post-transcriptionalmodifications (PTMs) of the parental nu-cleosomes are used as blueprints for the
restoration of the same nucleosome con-figuration. These reconstructions may beassisted by other chromatin componentssuch as DNA methylation patterns (Martinand Zhang 2007), RNAs (Grewal and Jia2007; Ringrose and Paro 2007), and by thelocation of the chromosomal domainswithin the nucleus (Misteli 2007).
In addition to DNA methylation patterns,as well as histone variants and their PTMs,chromatin has various enzymes that associatedynamically with the histones and DNAbound to them and that participate in theirregulatory and structural functions (Bantig-nies and Cavalli 2006). The patterns of asso-ciation of these nonhistone proteins withother chromatin components can also bereconstructed between generations of cellsand organisms (Ringrose and Paro 2004;Bantignies and Cavalli 2006; Schuettengru-ber et al. 2007). Recent data suggest thatRNA produced at regions that bind non-histone chromatin proteins may lead to theinheritance of the bound state throughRNA-DNA pairing (Grewal and Jia 2007;Ringrose and Paro 2007).
the RNA-mediated EISDuring the last decade, it has become
apparent that RNA is central to the regu-lation of cellular dynamics in eukaryotesand is also involved in cell and organismheredity. Gene silencing by small RNAs—RNA interference (RNAi)—has been foundin all eukaryote phyla from yeast to man,although a few species (e.g., buddingyeast) seem to have lost the capacity forthis. In all of the RNAi pathways, double-stranded RNA molecules (dsRNA), whichtrigger the process, are chopped intoshorter dsRNAs (usually between 21-30 nu-cleotides long) by the enzyme Dicer. Af-ter the original dsRNA is chopped, the re-sulting siRNA (small interfering RNA) isloaded onto a complex of proteins, onestrand of the duplex is removed, and theother strand—the guide strand—directs si-lencing. Silencing may occur through anyof the following mechanisms: (i) the siRNAis loaded onto an enzyme complex thatinterferes with the transcription or transla-tion of mRNAs with a homologous se-
136 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

quence (Cullen 2004; Meister and Tuschl2004); (ii) the siRNA is loaded onto anenzyme complex that targets chromatin re-gions with DNA that is homologous to thesiRNA, and alters chromatin into a silentstate (Matzke and Birchler 2005; Ekwall2007; Huettel et al. 2007); or (iii) thesiRNA is loaded onto an enzymatic com-plex that degrades and/or excises the DNAsequences complementary to the siRNAs.The latter processes of RNA-regulatedDNA rearrangement are being intenselystudied in ciliates (Meyer and Chalker2007; Nowacki et al. 2008).
RNA can affect cell and organism hered-ity in at least three different, non-mutuallyexclusive ways. The first is the result of rep-lication of siRNA through RNA-dependentRNA polymerase. This replication is a two-stage process, the details of which differbetween taxa, and it leads to the amplifica-tion of siRNAs that act as repressors ofgene expression (Baulcombe 2007; Pak andFire 2007). These siRNAs are transmittedto daughter cells during cell division andcan migrate to other cells as well (e.g.,Palauqui et al. 1997; Himber et al. 2003).The second way in which RNAs can af-fect cell heredity is by guiding, targeting,and assisting in transmitting variations inchromatin structure that are reconstructedand reproduced in daughter cells throughthe chromatin-marking EIS (Matzke andBirchler 2005; Ekwall 2007; Huettel et al.2007). The third way is by targeting DNAbase sequences and guiding changes inthem that are then replicated by DNA poly-merases (Meyer and Chalker 2007; Nowackiet al. 2008). Heritable variations can be gen-erated and perpetuated through all threeroutes of RNA-mediated heredity, and theformation of a particular dsRNA may be af-fected by local conditions and may be devel-opmentally regulated. Once formed, suchdsRNA may have long-term hereditary ef-fects.
EISs and the germlineIt is obvious from the foregoing descrip-
tions that the different EISs are oftenmechanistically and functionally interre-lated; therefore, the division of EISs into
four categories is somewhat arbitrary andartificial. However, dividing EISs in thismanner highlights the variety and com-plexity of the cellular mechanisms of in-heritance. The details of how epigeneticvariations are transmitted through mitosisin asexual clonal lineages remain a puzzle,but in sexually reproducing organisms,especially multicellular ones, the puzzle iseven greater.
In sexually reproducing organisms, epi-genetic variations have to survive the com-plex process of meiosis in order to be trans-mitted to the next generation, and, inmulticellular organisms, they also have tosurvive gametogenesis and early embryo-genesis—two developmental stages that in-volve significant restructuring of both cellsand chromatin. Although there is as yet noevidence that prions and self-sustainingloops are transmitted between generationsthrough sperm and egg, there is evidencethat chromatin marks and RNAs can betransmitted in this manner, but it is notclear how this occurs. It seems likely thatsome footprints of chromatin marks re-main and lead to the reconstruction ofancestral states, or that some remnants ofancestral states (including some RNAs) areretained. Even in male vertebrates, where acomprehensive replacement of histones byprotamines takes place during gametogen-esis, the erasure of histone marks is notcomplete. In the mouse, for example,about 1% of the DNA remains wrappedaround histones, and two acetylated vari-ants of H4 are maintained through sper-miogenesis (Van der Heijden et al. 2006).Van der Heijden and his colleagues (2006)suggested that these histones, as well asmethylated cytosines in centromeric DNA,are associated with the transgenerationalmaintenance of the structure of centro-meric heterochromatin. Chong et al. (2007)reported that when male mice had muta-tions in genes involved in epigenetic pro-gramming—in a gene that encodes a chro-matin remodeler protein, and in anotherthat encodes DNA methyltransferase—there were phenotypic effects on theiroffspring, even when they did not inheritthe defective gene. Extensive epigenetic
June 2009 137TRANSGENERATIONAL EPIGENETIC INHERITANCE

(methylation) variation has been found inhuman germ cells (Flanagan et al. 2006),but whether and to what extent this varia-tion is passed between generations is notknown.
In addition to some chromatin marks,certain RNAs may be transmitted throughthe germline. Cells in the germline containsmall RNAs, known as Piwi-associated inter-fering RNAs (piRNAs), that are importantfor the proper maturation of germ cells(Kim 2006). One function of piRNAs maybe the detection and silencing—or dele-tion—of regions of unpaired DNA duringmeiosis. Unpaired regions are targets ofthe RNAi machinery; RNA transcribedfrom them guides enzymatic complexes tothe unpaired regions, which are then de-leted or heterochromatinized (Shiu et al.2001; Bean et al. 2004; Turner et al. 2005;Costa et al. 2006). Mammalian spermato-cytes are filled with piRNAs, and similarRNAs have been discovered in oocytes too.The abundance of piRNAs suggests that theymay be transmitted to the next generationand lead to transgenerational effects. Thetransmission of an epigenetic modificationin male mice (induced in heterozygotes for avariant Kit gene) is known to be mediatedthrough microRNAs with sequences partiallycomplementary to that of Kit RNA (Rassoul-zadegan et al. 2006; see supplementarymaterial for details, available online at TheQuarterly Review of Biology homepage, www.journals.uchicago.edu/QRB).
Transgenerational EpigeneticInheritance: Prevalence,
Distribution, and InductionThere is no recent review of cellular epi-
genetic inheritance between generationsthat encompasses all four types of EISs andtheir distributions across taxa. The onlycomprehensive survey was made by Jab-lonka and Lamb in 1995. Since then, manymore cases have been described, and ourunderstanding of the molecular mecha-nisms underlying epigenetic inheritancehas deepened and expanded. The table wepresent here (Table 1) brings togetherover one hundred cases of inherited epige-netic variations in bacteria, protists, fungi,
plants, and animals. We have included onlycases where there is convincing evidencefor epigenetic inheritance in the narrowcellular sense, either through a single asex-ual cell (as in bacteria, some protists, somefungi, and some plants) or through a sex-ually generated gamete or spore (most an-imals, plants, and fungi). In most (al-though not all) cases, molecular studieshave revealed the involvement of one ormore of the EISs, but a full molecular char-acterization through all the reproductivestages is not yet available for any organism.More details about the cases are given assupplementary material in an expanded ta-ble form (available online at The QuarterlyReview of Biology homepage, www.journals.uchicago.edu/QRB).
How common is epigenetic inheritance?This question is often raised, and the an-swer, based upon the data we have accu-mulated, is that it may be ubiquitous. Webelieve that epigenetic variants in every lo-cus in the eukaryotic genome can be inher-ited, but in what manner, for how long,and under what conditions has yet to bequalified. In other words, unlike the repli-cation of DNA variations, which is largelycontext insensitive, whether and for howlong a particular mark or cellular elementis transmitted between generations de-pends on genomic, developmental, and ex-ternal conditions. This does not mean thatconditions that allow epigenetic inheri-tance are particularly rare. In fungi, forexample, the widespread occurrence ofepigenetic inheritance is generally ac-knowledged. As long ago as 1949, Linde-gren, reviewing data on inheritance in Neu-rospora, said that two thirds of new variantsdo not show Mendelian segregation andare therefore discarded, and, in their re-cent review of protein inheritance in fungi,Benkemoun and Saupe (2006) comment-ed: “Many of us working with filamentousfungi know how often bizarre looking sectors orsegregates that defy Mendelism appear on ourplates. As a number of pioneering fungal genet-icists have done in the past, maybe we shouldhave a closer look before putting them in theautoclave” (p. 793). They suggested thatmany of these “anomalies” may be caused
138 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

by some form of epigenetic inheritance. Inall organisms, chromatin inheritance cantheoretically occur at every locus in thegenome, and the double-stranded natureof DNA provides a theoretical possibility(when transcription occurs from bothstrands) for every DNA segment to formsmall dsRNA molecules that can lead toheritable silencing. The potential of mostproteins to form � sheets with spatialtemplating properties (Baxa et al. 2006)suggests that a prion-like transfer of con-formation between cells may occur morefrequently than previously thought.
cases included in the tableIn Table 1, we show only a small sample
of representative cases of epigenetic inher-itance in allopolyploid plant hybrids be-cause, in all cases that have been investi-gated, allopolyploidy is accompanied byextensive epigenetic changes, some ofwhich are inherited between generations.In most of the cases presented in the table,the molecular basis of the EIS has beenunravelled, but we included a few cases forwhich the evidence for cellular epigeneticinheritance seemed overwhelming, despitethe fact that molecular studies were lacking(e.g., inheritance of the star phenotype insilver foxes, and some cases of develop-mentally induced variability and inheri-tance in plants and fungi).
As our main goal in this review is to showthat variations in epigenetic marks can beinherited for several generations, we didnot include classical cases of genomic im-printing in the table. With genomic im-printing, the epigenetic marks that are im-posed on parental chromosomes duringoogenesis differ from those imposed dur-ing spermatogenesis; therefore, in the off-spring, a gene’s expression pattern de-pends on whether it was inherited from thefather or from the mother (see Barlow andBartolomei 2007 for genomic imprintingin mammals, Alleman and Doctor 2000 forflowering plants, and for general reviews ofimprinting that include invertebrates, seeJablonka and Lamb 1995; Haig 2002). Bydefinition, the genomic imprints charac-terizing one sex are reversed when chro-
mosomes go through gametogenesis in theopposite sex in the next generation; thus,imprints are inherently reversible. How-ever, molecular studies of imprinting havebeen important in the history of epigeneticinheritance because they have led to a gen-eral recognition of the role of epigeneticcontrol mechanisms, such as changes inDNA methylation in critical regions, andhave opened the way to unraveling casesthat were similar to genomic imprintingbut that were not parent-sex specific (e.g.,the early studies by Hadchouel et al.1987 and Allen et al. 1990, as discussedin Jablonka and Lamb 1995). The onlyimprinting-related case included in the ta-ble is that of an imprint in a human grand-mother that was not erased in her son andthat subsequently led to Prader-Willi syn-drome in her grandchildren (Buiting et al.2003). This is a clear case of an epigeneticimprinting mark being transmitted as a re-sult of a fault in resetting. As yet, we are notaware of any studies showing that new vari-ations in genomic imprints are transmittedover several consecutive generations.
In most of the cases that we included inthe table, the epigenetic variations havebeen transmitted for more than two gen-erations, and we can rule out repeateddirect induction of the variation in eachgeneration. However, when the inducedparent carries an embryo, three genera-tions may be necessary to confirm epige-netic inheritance. For example, when a fe-male mammal is exposed to an inducingagent during pregnancy, direct inductionof the embryo’s germ cells has to be ex-cluded; therefore, three generations oftransmission (from F0 to F3) are requiredto establish that epigenetic inheritance,rather than direct induction of the embryoand its germline, has occurred (Jirtle andSkinner 2007). On the other hand, when amale mammal is exposed to an inducer,two generations of transmission (from F0
to F2) are sufficient to establish that therehas been epigenetic transmission throughthe germline rather than through directinduction. When the epigenetic effects(e.g., patterns of methylation) in the F1
and F2 generations are identical, and when
June 2009 139TRANSGENERATIONAL EPIGENETIC INHERITANCE

TA
BL
E1
Tra
nsge
nera
tiona
lep
igen
etic
inhe
rita
nce
inpr
okar
yote
san
deu
kary
otes
Tax
onT
rait
Loc
us/c
ellu
lar
syst
emSt
abili
tyIn
duci
ngco
ndit
ions
EIS
Ref
eren
ce
Bac
teri
aan
dth
eir
viru
ses
�ph
age
ofEs
cher
ichia
coli
Lys
ogen
ic/l
ytic
cycl
eC
lan
dC
roSt
able
Nut
riti
onal
stat
eof
the
hos
t,ph
age
den
sity
Self
-sus
tain
ing
loop
sPt
ash
ne
(200
5)
Bac
illus
subt
ilis
Inac
tiva
tion
ofch
rom
osom
eW
hol
ech
rom
osom
eSt
able
Poly
eth
ylen
glyc
ol(P
EG
)-in
duce
dfu
sion
Ch
rom
atin
-mar
kin
gG
ran
djea
net
al.
(199
8)
Lac
kof
cell
wal
lB
alan
cebe
twee
npe
ptid
ogly
can
syn
thes
isan
dde
stru
ctio
nSt
able
onag
arEx
peri
men
talr
emov
alof
cell
wal
lSt
ruct
ural
inh
erit
ance
Lan
dman
(199
1)
Spor
ulat
ion
Spo0
Aph
osph
orel
aySt
able
Nut
riti
onal
depr
ivat
ion
Self
-sus
tain
ing
loop
sV
een
ing
etal
.(2
005)
Nat
ural
com
pete
nce
(Kst
ate)
Com
Kac
tivi
ty10
%–2
0%in
lab
stra
ins,
1%in
the
wild
Stoc
has
tic,
elev
ated
byst
ress
Self
-sus
tain
ing
loop
sM
aam
aran
dD
ubna
u(2
005)
Esch
eric
hia
coli
Uti
lizat
ion
ofla
ctos
ea
Lac
oper
onac
tivi
tySt
able
unde
rco
ndi
tion
sof
low
indu
cer
con
cen
trat
ion
Stoc
hast
ic,g
row
thin
low
conc
entr
atio
nof
indu
cer
Self
-sus
tain
ing
loop
sC
ohn
and
Hor
ibat
a(1
959a
,b),
Lau
rent
etal
.(2
005)
,Nov
ick
and
Wei
ner
(195
7),O
zbud
aket
al.(
2004
)Fl
uffy
Agn
43Ph
ase
vari
atio
nPr
obab
lyox
idat
ive
stre
ssG
AT
Cm
eth
ylat
ion
Cas
ades
usan
dL
ow(2
006)
Pili
Pap
oper
on10
-4pe
rge
ner
atio
n,
10-3
reve
rsio
nC
han
ged
carb
onso
urce
,te
mpe
ratu
re,
and
spon
tan
eous
GA
TC
met
hyl
atio
nH
ern
day
etal
.(2
002)
Gro
wth
rate
(per
sist
erty
peII
)b
Prob
ably
man
yge
nes
10-6
per
gen
erat
ion
,10
-1pe
rge
ner
atio
nre
vers
ion
Spon
tan
eous
and
anti
biot
ictr
eatm
ent
Self
-sus
tain
ing
loop
sB
alab
anet
al.(
2004
),L
ewis
(200
7),N
.Bal
aban
(per
sona
lco
mm
unic
atio
n)R
esist
ance
toan
tibio
tics
(am
pici
llin,
tetr
acyc
line,
and
nalid
ixic
acid
)
Alt
ered
regu
lati
onof
�
lact
amas
ecr
ypti
cge
ne,
glut
amat
ege
ne,
and
deca
rbox
ylas
ege
ne;
poss
ible
invo
lvem
ent
ofD
NA
met
hyl
ase
gen
es
3%–2
0%su
rviv
al(d
epen
din
gon
con
cen
trat
ion
ofan
tibi
otic
)an
d50
%re
vers
ion
rate
Low
,and
succ
essi
vely
incr
ease
dco
ncen
trat
ions
ofan
tibio
tics
Poss
ibly
self
-sus
tain
ing
loop
san
d/o
rD
NA
met
hyl
atio
n
Ada
met
al.
(200
8)
Pseu
dom
onas
aeru
gino
saT
oxin
inje
ctio
nT
TSS
syst
emSt
able
Cel
lde
nsi
tySe
lf-s
usta
inin
glo
ops
Filo
pon
etal
.(2
006)
140 Volume 84THE QUARTERLY REVIEW OF BIOLOGY
Syne
coco
ccus
elon
gate
s(C
yan
obac
teri
a)
Cir
cadi
anrh
yth
mR
egul
ator
ylo
opin
volv
ing
key
Kai
Cpr
otei
nSe
vera
lda
ysL
igh
tan
dda
rkpu
lses
Self
-sus
tain
ing
loop
sK
ondo
and
Ish
iura
(200
0)
Pro
tist
sO
xytr
icha
trifa
llax
(Cili
ate)
Alt
erat
ion
ofge
ne
orde
r:ab
erra
nt
rear
ran
gem
ents
Gen
esth
atbe
com
eun
scra
mbl
edin
the
som
atic
mac
ronu
cleu
s(i
.e.,
mos
tge
nes)
Stab
leth
roug
has
exua
lre
prod
uctio
n;at
leas
t3ge
nera
tions
follo
win
gse
xual
repr
oduc
tion
Exp
erim
enta
lm
anip
ulat
ion
RN
A-m
edia
ted
rear
ran
gem
ent
Now
acki
etal
.(2
008)
Para
mec
ium
aure
lia(C
iliat
e)
Sero
type
sex
pres
sed
Cili
apr
otei
ns
Stab
leC
han
ges
inpH
,te
mpe
ratu
re,
food
supp
ly,
and
salin
ity
Self
-sus
tain
ing
loop
sL
andm
an(1
991)
Indu
ced
tole
ran
ceto
hea
t,sa
lt,
and
arse
nic
Not
spec
ified
Inh
erit
edfo
rm
any
gen
erat
ion
sbe
fore
fadi
ng
away
grad
ually
Exp
osur
eto
hig
hte
mpe
ratu
re,
hig
hsa
lt,
and
arse
nic
con
cen
trat
ion
s
Not
know
nJo
llos
(192
1);
revi
ewed
inJa
blon
kaet
al.
(199
2)
Para
mec
ium
tetr
aure
lia,
Styl
onch
iale
mna
e,T
etra
hym
ena
ther
mop
hila
(Cili
ates
)c
Var
ious
trai
tsre
late
dto
alte
rnat
ive
gen
etic
orga
niz
atio
npa
tter
ns
inth
em
acro
nuc
leus
Inpr
inci
pal,
any
DN
Ase
quen
cein
the
gen
ome
Stab
leth
roug
hm
acro
nuc
leus
repr
oduc
tion
,bo
thdu
rin
gas
exua
lan
dfo
llow
ing
sexu
alre
prod
ucti
on
Sequ
ence
com
pari
son
ofm
ater
nal
(old
)an
dzy
goti
c(n
ew)
nuc
leus
,ba
sed
onn
on-
expr
esse
dse
quen
ces,
lead
sto
the
inh
erit
ance
ofm
ater
nal
,n
ucle
us-
guid
edge
ne
orga
niz
atio
n
Inhe
rite
dD
NA
rear
rang
emen
t/ed
iting
/pr
ogra
mm
ing
ofth
em
acro
nucl
eus
med
iate
dby
smal
lR
NA
san
dch
rom
atin
mod
ifica
tions
Gar
nie
ret
al.
(200
4),
Jura
nek
etal
.(2
005)
,L
iuet
al.
(200
4),
Mey
eran
dC
hal
ker
(200
7),
Tav
ern
aet
al.
(200
2),
Yao
etal
.(2
003)
Cor
tica
lor
gan
izat
ion
Bas
albo
dyan
dco
rtex
prot
ein
sSt
able
inm
itos
isan
dso
met
imes
inm
eios
is
Exp
erim
enta
lm
anip
ulat
ion
,st
ress
Stru
ctur
alin
her
itan
ce(g
uide
das
sem
bly)
Gri
mes
and
Auf
derh
eide
(199
1)
Plas
mod
ium
falc
ipar
um(M
alar
iapa
rasi
te)
Tel
omer
ein
acti
vati
onT
elom
ere
sequ
ence
sSw
itch
ever
y�
15ge
ner
atio
ns
Spon
tan
eous
Ch
rom
atin
-mar
kin
gR
ober
tset
al.
(199
2) cont
inue
d
June 2009 141TRANSGENERATIONAL EPIGENETIC INHERITANCE
TA
BL
E1
Con
tinue
d
Tax
onT
rait
Loc
us/c
ellu
lar
syst
emSt
abili
tyIn
duci
ngco
ndit
ions
EIS
Ref
eren
ce
Tet
rahy
men
a(C
iliat
e)In
crea
sed
insu
linbi
ndin
gan
dpr
oduc
tion
follo
win
gex
posu
reto
diio
doty
rosin
e(T
2)
Not
spec
ified
Ver
yst
able
—hu
ndre
dsof
gene
ratio
nsIn
duct
ion
byin
sulin
ordi
iodo
tyro
sin
ePr
obab
lym
eth
ylat
ion
;tr
eatm
ent
wit
h5-
azac
ytid
ine
can
abol
ish
resp
onse
Csa
ba(2
008)
,Csa
baet
al.
(199
9),C
saba
and
Kov
acs
(199
0,19
95),
Csa
baet
al.
(198
2a,b
)
Volv
oxca
rter
iSi
lenc
ing
ofin
vitro
met
hyla
ted
tran
sgen
e
C-a
rstr
ansg
ene
Mor
eth
an10
0ge
ner
atio
ns
DN
Atr
ansf
orm
atio
nD
NA
met
hyl
atio
nB
abin
ger
etal
.(2
007)
Fung
iA
scob
olus
imm
erse
sT
ran
sgen
ein
acti
vati
onA
ny
dupl
icat
edtr
ansg
ene
Stab
lePr
e-m
eiot
ical
lyD
NA
met
hyl
atio
nM
artie
nsse
nan
dC
olot
(200
1),R
houn
imet
al.
(199
2)C
andi
daal
bica
nsd
Cel
lmor
phol
ogy,
abili
tyto
form
colo
nies
onva
riou
ssu
bstr
ates
,mat
ing
prop
ertie
s
Mas
ter
regu
lato
rW
OR
1pr
otei
nSw
itch
esev
ery
�
10,0
00ge
ner
atio
ns
Spon
tan
eous
,af
fect
edby
tem
pera
ture
Self
-sus
tain
ing
loop
Hua
ng
etal
.(2
006)
,M
alag
nac
and
Sila
r(2
003)
,Z
orda
net
al.
(200
6)
Cop
rinu
sci
nere
usM
eth
ylat
ion
patt
ern
Cen
trom
ere-
linke
dlo
cus
Stab
lew
hen
hig
hly
met
hyl
ated
Un
know
nC
hro
mat
in-m
arki
ng,
DN
Am
eth
ylat
ion
invo
lved
Zol
anan
dPu
kkila
(198
6)
Podo
spor
aan
seri
na(F
ilam
ento
usfu
ngu
s)
Cri
pple
dG
row
th(C
G)
C—
assu
med
tobe
atr
ansm
issi
ble
self-
sust
aini
ngca
scad
efa
ctor
invo
lvin
ga
Map
kina
sem
odul
e
Stab
lem
itot
icin
her
itan
ceT
rans
form
atio
nof
norm
alce
llsto
CG
cells
indu
ced
bycy
todu
ctio
n,pr
omot
ing
stat
iona
ryst
ate
and
grow
thon
med
ium
supp
lem
ente
dw
ithye
aste
xtra
ct
Self
-sus
tain
ing
loop
Kic
kaet
al(2
006)
,Mal
agna
can
dSi
lar
(200
6),S
ilar
etal
.(19
99)
[Het
-s*]
and
[Het
-s]va
rian
tsaf
fect
ing
vege
tativ
ein
com
patib
ility
het-s
Stab
le;
low
freq
uen
cy[H
et-s
]co
nve
rted
toin
acti
ve[H
et-
s*],
but
the
reve
rse
also
occu
rs,
albe
itat
low
freq
uen
cy
Spon
tan
eous
Stru
ctur
alin
her
itan
ce(p
rion
)M
adde
lein
etal
.(2
002)
142 Volume 84THE QUARTERLY REVIEW OF BIOLOGY
Sacc
haro
myc
esce
revi
siae
Enha
nced
resis
tanc
eto
star
vatio
non
dext
rose
-min
imal
plat
esan
dsu
ppre
ssio
nof
inab
ility
tosp
orul
ate
inm
utan
ts
[�]-
prio
nfo
rmof
the
PrB
vacu
olar
prot
ease
Inde
fin
ite
prop
agat
ion
Em
erge
sin
dele
tion
-mut
ants
;in
duct
ion
isin
crea
sed
asa
resu
ltof
PrB
over
-ex
pres
sion
Prio
n,
thro
ugh
self
-su
stai
nin
glo
opR
ober
tsan
dW
ickn
er(2
003)
Gro
wth
phen
otyp
eFL
Oge
nes
nea
rte
lom
ere
Swit
chev
ery
10to
15ge
ner
atio
ns
Spon
tan
eous
Ch
rom
atin
inh
erit
ance
Hal
me
etal
.(2
004)
Rea
dth
roug
h[P
S]St
able
Var
ious
Stru
ctur
alin
her
itan
ce(p
rion
)T
uite
and
Cox
(200
6)
Nit
roge
nca
tabo
licge
ne
expr
essi
on[U
RE
3]St
able
Spon
tan
eous
and
over
expr
essi
onSt
ruct
ural
inh
erit
ance
(pri
on)
Ben
kem
oun
and
Saup
e(2
006)
Asc
usfo
rmat
ion
[PIN
]�R
nq1
pSt
able
Spon
tan
eous
and
over
expr
essi
onof
Sup3
5St
ruct
ural
inh
erit
ance
(pri
on)
Ben
kem
oun
and
Saup
e(2
006)
Exp
ress
ion
ofex
peri
men
tally
mod
ified
GA
Ln
etw
ork
GA
Ln
etw
ork
Stab
leL
owin
duce
rco
nce
ntr
atio
ns
Self
-sus
tain
ing
loop
Aca
ret
al.
(200
5)
Indu
ctio
nof
GA
L1
and
GA
L7
byga
lact
ose
GA
Lge
nes
Up
to7
gen
erat
ion
sIn
duct
ion
byga
lact
ose
Slow
dilu
tion
ofab
unda
nt
regu
lato
ryG
AL
1pr
otei
n
Zach
ario
udak
iset
al.(
2007
)
An
ti-s
uppr
esso
rIS
P�?
Stab
leSp
onta
neo
usPr
obab
lyba
sed
onst
ruct
ural
inh
erit
ance
Ben
kem
oun
and
Saup
e(2
006)
,Kun
zan
dB
all
(197
7),T
allo
czy
etal
.(2
000)
,Vol
kov
etal
.(2
002)
Glu
cosa
min
ere
sist
ance
GR
?
Con
trol
ofki
ller
viru
sex
pres
sion
KI-
d?(s
uspe
cted
prio
ns)
Slow
grow
than
dad
diti
onal
requ
irem
ent
for
leuc
ine
Stru
ctur
alal
tera
tion
ofth
em
itoc
hon
drio
nM
ore
than
100
gen
erat
ion
sin
the
abse
nce
ofle
ucin
e
Spon
tan
eous
inh
erit
edlo
ssof
mit
och
ondr
ial
DN
A
Stru
ctur
alin
her
itan
ceL
ocks
hon
(200
2)
cont
inue
d
June 2009 143TRANSGENERATIONAL EPIGENETIC INHERITANCE
TA
BL
E1
Con
tinue
d
Tax
onT
rait
Loc
us/c
ellu
lar
syst
emSt
abili
tyIn
duci
ngco
ndit
ions
EIS
Ref
eren
ce
Schi
zosa
ccha
rom
yces
pom
beSu
rviv
alof
mut
ant
stra
inla
ckin
gth
ehc
dre
gion
-en
codi
ng,
hig
hly
con
serv
eddo
mai
nof
the
esse
nti
alch
aper
one
caln
exin
[cif
]C
alne
xin
inde
pend
ence
inhe
rite
dby
88.7
%of
spor
es
Mat
ing
cells
lack
ing
the
gene
codi
ngfo
rca
lnex
inw
ithca
lnex
in-
depe
nden
tce
lls
Stru
ctur
alin
her
itan
ce,
prob
ably
prio
nC
ollin
etal
.(2
004)
Mei
otic
telo
mer
ecl
uste
ring
and
chro
mat
inst
ruct
ure
Inte
ract
ion
ofte
lom
eric
and
subt
elom
eric
regi
ons
with
Taz
1St
able
Nor
mal
;var
iatio
nsh
own
byde
letin
gte
lom
ere
sequ
ence
sin
chro
mos
omes
Ch
rom
atin
-mar
kin
g,st
ruct
ural
inh
erit
ance
Sada
ieet
al.
(200
3)
Rep
orte
rtr
ansg
ene
sile
nci
ng
and
mat
ing
type
swit
chin
g
K�
::ura
4(t
ran
sgen
ein
sert
edat
K-r
egio
n)
At
leas
t30
mit
otic
gen
erat
ion
saf
ter
mei
osis
Tra
nsg
enic
ally
indu
ced
Ch
rom
atin
-org
aniz
ing
fact
ors
prob
ably
invo
lved
Gre
wal
and
Kla
r(1
996)
Pla
nts
Ant
irrhi
num
maj
us(S
napd
rago
n)
Flus
h/g
ran
ulat
edph
enot
ype
(par
amut
atio
n)
nive
alo
cus
Var
iega
ted
inF 1
,m
ore
stab
lein
F 2
Indu
ced
bycr
ossi
ng
Tra
nsp
osit
ion
sugg
este
d;ch
rom
atin
-mar
kin
gpl
ausi
ble
Kre
bber
set
al.
(198
7)
Ara
bido
psis
thal
iana
(Mou
seea
rcr
ess)
Rev
ersi
onof
dwar
fm
orph
olog
yC
pr1–
1ge
ne-
epia
llele
inte
ract
ing
wit
hba
lEf
fect
seen
inF 2
prog
eny
butn
otin
F 1
Indu
ced
bycr
ossi
ng
Un
know
nSt
okes
and
Ric
hard
s(2
002)
Size
,ros
ette
,and
petio
leab
norm
aliti
es;
activ
ityof
GFP
repo
rter
gene
Tra
nsg
enic
loci
E82
and
L91
inte
ract
ing
wit
hC
OP1
endo
gen
e
At
leas
t5
gen
erat
ion
sIn
duce
dby
cros
sin
gIn
volv
emen
tof
RN
A-
med
iate
dD
NA
met
hyl
atio
nsu
gges
ted
Qin
and
von
Arn
im(2
002)
Del
ayed
flow
erin
gfw
a(g
ene-
epia
llele
)V
ery
stab
leE
MS,
fast
neu
tron
trea
tmen
t,or
ddm
1in
duce
d(W
ildty
peD
DM
1re
quir
edto
mai
nta
inD
NA
met
hyl
atio
n)
DN
Am
eth
ylat
ion
,h
isto
ne
H3
met
hyl
atio
n(s
iRN
As
invo
lved
)
Kin
osh
ita
etal
.(2
007)
,L
ippm
anet
al.
(200
4),
Sopp
eet
al.
(200
0)
144 Volume 84THE QUARTERLY REVIEW OF BIOLOGY
Dw
arfis
m,c
onst
itutiv
eac
tivat
ion
ofde
fens
ere
spon
sepa
thw
ay
Bal
locu
sA
tle
ast
5ge
ner
atio
ns
ddm
1ef
fect
even
wh
ense
greg
ated
out
Prob
ably
DN
Am
eth
ylat
ion
Stok
eset
al.
(200
2)
Num
ber
ofre
prod
uctiv
eor
gans
SUP
At
leas
t2
gen
erat
ion
sV
ario
usm
utag
ens
DN
Am
eth
ylat
ion
Jaco
bsen
and
Mey
erow
itz
(199
7)Ex
pres
sion
leve
lsof
the
retr
otra
nspo
son
At2
g104
10
At2
g104
10ep
ialle
leA
tle
ast
8ge
ner
atio
ns
Spon
tan
eous
,fo
und
inn
atur
alpo
pula
tion
sD
NA
met
hyl
atio
nR
angw
ala
etal
.(2
006)
Incr
ease
dle
vels
ofho
mol
ogou
sre
com
bina
tion
inso
ma
Not
spec
ified
Atl
east
4ge
nera
tions
inca
seof
UV
-Cra
diat
ion;
atle
ast2
gene
ratio
nsin
case
ofin
trod
uced
flage
llin
Ult
ravi
olet
radi
atio
nor
intr
oduc
tion
offl
agel
lin
Not
spec
ified
Mol
inie
ret
al.
(200
6),
J.M
olin
ier
(per
son
alco
mm
unic
atio
n)
Los
sof
hyg
rom
ycin
resi
stan
ce(p
aram
utat
ion
)
hpt
tran
sgen
eN
oef
fect
inF 1
;F 2
affe
cted
Tra
nsg
ene-
indu
ced
Invo
lves
DN
Am
eth
ylat
ion
Sch
eid
etal
.(2
003)
Tra
nsc
ript
ion
alac
tivi
tyof
tran
spos
able
elem
ents
Var
ious
tran
spos
able
elem
ents
At
leas
t6
gen
erat
ion
sdd
m1
effe
ct,
even
afte
rse
greg
ated
out
DN
Am
eth
ylat
ion
,ch
rom
atin
mod
ifica
tion
s;R
NA
ipr
obab
lyin
volv
ed
Lipp
man
etal
.(20
03,2
004)
,V
.Col
ot(p
erso
nal
com
mun
icat
ion)
Blu
efl
uore
scen
ce,
tryp
toph
anan
dIA
Ade
fici
ency
,m
orph
olog
ical
abn
orm
alit
ies
PAI
(gen
e-ep
ialle
les)
At
leas
t6
gen
erat
ion
sT
-DN
Am
utag
enes
is,
cros
sin
g;va
riat
ion
foun
din
nat
ural
popu
lati
ons
DN
Am
ethy
latio
n;D
NA
/D
NA
pair
ing
issu
gges
ted
Ben
der
and
Fin
k(1
995)
,L
uff
etal
.(1
999)
Ara
bido
psis
Inte
rspe
cfic
hyb
rids
e
Man
ytr
aits
Man
ylo
ci,
both
codi
ng
and
non
codi
ng
Stab
ility
vari
esac
cord
ing
tolo
cus,
but
man
ylo
cish
owst
able
inh
erit
ance
Indu
ced
byh
ybri
diza
tion
follo
wed
bypo
lypl
oidi
zati
on
Ch
rom
atin
-mar
kin
g,D
NA
met
hyl
atio
n;
RN
Ai
syst
empr
obab
lyal
soin
volv
ed
Com
aiet
al.
(200
0),
Sch
eid
etal
.(2
003)
cont
inue
d
June 2009 145TRANSGENERATIONAL EPIGENETIC INHERITANCE
TA
BL
E1
Con
tinue
d
Tax
onT
rait
Loc
us/c
ellu
lar
syst
emSt
abili
tyIn
duci
ngco
ndit
ions
EIS
Ref
eren
ce
Beta
vulg
aris
(Sug
arbe
et)
Man
ytr
aits
inm
itotic
,ag
amos
perm
ic,a
ndin
bred
lines
incl
udin
g:sin
gle
orm
ultip
leflo
wer
initi
atio
n,se
lf-fe
rtili
ty,
poly
mor
phism
ofm
alic
enzy
me,
and
vari
atio
nin
num
ber
ofch
loro
plas
ts
Mm
,li,
Rf1
,an
dM
e1lo
ciV
aria
ble,
ofte
n2
orm
ore
gen
erat
ion
sM
ode
ofre
prod
ucti
on,
clim
atic
con
diti
ons,
dire
ctio
nof
cros
s
DN
Am
eth
ylat
ion
ispr
obab
lyin
volv
edL
evit
es(2
000)
,L
evit
esan
dM
alet
skii
(199
9),
Mal
etsk
ii(1
999)
Lin
aria
vulg
aris
(Com
mon
toad
flax
)
Flow
ersy
mm
etry
Lcy
c(g
ene-
epia
llele
)A
tle
ast
2ge
ner
atio
ns
Nat
ural
vari
atio
nin
popu
lati
ons
DN
Am
eth
ylat
ion
Cub
aset
al.
(199
9),
J.Pa
rker
(per
son
alco
mm
unic
atio
n)
Lin
um usita
tissi
mum
(Fla
x)
Plan
twei
ght,
heig
ht,
pero
xida
seiso
zym
epa
ttern
,see
dca
psul
ese
pta
hair
num
ber
r-D
NA
gen
esan
dre
peti
tive
sequ
ence
sSt
able
Fert
ilize
ran
dh
eat
regi
mes
Met
hyl
atio
nan
dD
NA
-re
patt
ern
ing
Cul
lis(2
005)
Flow
erin
gag
e,m
ain
stem
heig
htat
mat
urity
,and
num
ber
ofle
aves
Not
spec
ified
,bu
tep
imut
atio
ns
inat
leas
t3
inde
pen
den
t,n
onra
ndo
mlo
ciar
eas
sum
edto
bein
volv
ed
At
leas
t9
gen
erat
ion
s5-
azaC
trea
tmen
tD
NA
met
hyl
atio
n,
poss
ibly
asso
ciat
edch
rom
atin
rem
odel
ing
asw
ell
Fiel
des
and
Am
yot
(199
9),
Fiel
des
etal
.(2
005)
Lyc
oper
sico
nes
cule
ntum
(Tom
ato)
Inhi
bitio
nof
ripe
ning
and
deve
lopm
ento
fa
colo
rles
spe
rica
rp
KeS
PL-C
NR
(gen
eep
ialle
le)
Ver
yst
able
Spon
tan
eous
epim
utat
ion
DN
Am
eth
ylat
ion
Man
nin
get
al.
(200
6)
Col
orva
rieg
atio
n(P
aram
utat
ion)
Sulf
epia
llele
sV
ery
stab
leX
-ray
s;re
gen
erat
ion
Ch
rom
atin
inh
erit
ance
sugg
este
dH
agem
ann
(196
9,19
93),
Hag
eman
nan
dB
erg
(197
7),
Wis
man
etal
.(1
993)
,R
.H
agem
ann
(per
son
alco
mm
unic
atio
n)
146 Volume 84THE QUARTERLY REVIEW OF BIOLOGY
Mel
andr
ium
albu
m(W
hit
eca
mpi
on)
Bis
exua
lity
Dec
reas
ein
CG
met
hyl
atio
nin
man
ylo
ci2
succ
essi
vege
ner
atio
ns,
inh
erit
edth
roug
hth
em
ale
pare
nt
Tre
atm
ent
wit
h5-
azac
ytid
ine
DN
Am
eth
ylat
ion
Jan
ouse
ket
al.
(199
6)
Nico
tiana
taba
cum
(Tob
acco
)
Los
sof
kan
amyc
inre
sist
ance
;pa
ram
utat
ion
-like
effe
cts
Nos
pro
and
nos
At
leas
t2
gen
erat
ion
sT
ran
sgen
icsi
len
cin
gin
doub
lytr
ansf
orm
edpl
ants
DN
Am
eth
ylat
ion
Mat
zke
and
Mat
zke
(199
1),
Mat
zke
etal
.(19
89)
Loss
ofhy
grom
ycin
resis
tanc
eH
pttr
ansg
ene
2ge
ner
atio
ns
Tra
nsg
enic
sile
nci
ng
DN
Am
eth
ylat
ion
Park
etal
.(1
996)
Req
uire
men
tofl
eaf
cell
for
cyto
kini
nN
otsp
ecifi
edA
rise
sat
10-2
per
cell
gen
erat
ion
Subc
ultu
rin
gin
med
iaco
nta
inin
gsu
cces
sive
lylo
wer
con
cen
trat
ion
sof
cyto
kin
in
Un
know
n;
DN
Am
eth
ylat
ion
sugg
este
dM
ein
s(1
986,
1989
a,b)
,M
ein
san
dT
hom
as(2
003)
Ory
zasa
tiva
(Ric
e)C
pGm
ethy
latio
npa
ttern
ssh
owin
heri
ted
culti
var
spec
ifici
ty
Met
hyl
atio
nst
ate
ofcy
tosi
ne
inva
riou
sC
CG
Gsi
tes
acro
ssth
ege
nom
e
At
leas
t6
gen
erat
ion
sC
ross
ing
Nip
pon
bare
and
Kas
alat
hcu
ltiv
ars5
DN
Am
eth
ylat
ion
Ash
ikaw
a(2
001)
Var
ious
trai
tsM
eth
ylat
ion
stat
ein
vari
ous
CC
GG
site
sac
ross
the
gen
ome
At
leas
t2
gen
erat
ion
sIn
trog
ress
ion
and
selfi
ng5
DN
Am
eth
ylat
ion
Don
get
al.
(200
6)
Ker
nel
shap
ean
dti
ller
num
ber
S2,
S3,
and
vari
ous
unsp
ecifi
edm
eth
ylat
ion
site
s3–
6ge
nera
tions
ofse
lfing
Hig
h-p
ress
ure
trea
tmen
tgi
ven
tose
eds
DN
Am
eth
ylat
ion
Shen
etal
.(2
006)
Indu
ced
dwar
fism
;re
sist
ance
topa
thog
en
Gen
eral
chan
gein
DN
Am
ethy
latio
n;X
a21G
prom
oter
At
leas
t9
gen
erat
ion
sIn
duce
dby
5-az
aCD
NA
met
hyl
atio
nin
volv
edA
kim
oto
etal
.(2
007)
,Sa
no
etal
.(1
990)
Petu
nia
hybr
ida
(Pet
unia
)W
hit
e-fl
ower
ing
An3
epia
llele
and
the
dTph
1tr
ansp
oson
At
leas
t3
gen
erat
ion
sof
self
-fe
rtili
zati
on
Indu
ced
byh
eter
ozyg
osit
yU
nkn
own
;ep
igen
etic
inte
ract
ion
betw
een
atle
ast
3dT
ph1
copi
es
Van
Hou
wel
inge
net
al.
(199
9)
Tri
ticum
aest
ivum
(Wh
eat)
Cyt
osin
em
eth
ylat
ion
Glu
ten
inge
ne
At
leas
t2
gen
erat
ion
sIn
bree
din
gC
hro
mat
in-m
arki
ng,
DN
Am
eth
ylat
ion
Flav
ell
and
O’D
ell
(199
0)
Lon
ger
prod
ucti
vesp
ikes
,la
rger
seed
s,an
dot
her
quan
tita
tive
trai
ts
Hl1
and
pclo
ci57
gen
erat
ion
sN
icot
inic
acid
Un
know
nB
ogda
nov
a(2
003)
cont
inue
d
June 2009 147TRANSGENERATIONAL EPIGENETIC INHERITANCE
TA
BL
E1
Con
tinue
d
Tax
onT
rait
Loc
us/c
ellu
lar
syst
emSt
abili
tyIn
duci
ngco
ndit
ions
EIS
Ref
eren
ce
Trit
icum
inte
rspe
cific
synt
hetic
hybr
idse
Man
ytr
aits
�13
%of
the
gen
ome
Stab
ility
depe
nds
onlo
cus;
man
yve
ryst
able
Hyb
ridi
zati
onan
dpo
lypl
oidi
zati
onC
hrom
atin
and
DN
Am
ethy
latio
n;R
NA
isy
stem
prob
ably
invo
lved
Lev
yan
dFe
ldm
an(2
004)
Tri
tical
e(S
tabl
ew
heat
-rye
hybr
id)
Incr
ease
inst
atur
e,n
umbe
rof
tille
rs,
chan
ged
tim
eof
ripe
nin
g
Not
spec
ified
At
leas
t2
gen
erat
ion
s5-
azaC
trea
tmen
tC
hro
mat
in-m
arki
ng;
DN
Am
eth
ylat
ion
invo
lved
Hes
lop-
Har
riso
n(1
990)
Zea
may
s(M
aize
)R
educ
edpi
gmen
tati
on(p
aram
utat
ion
)
B1
(gen
e-ep
ialle
le)
Ver
yst
able
Para
mut
agen
icB�
epia
llele
arise
ssp
onta
neou
slyfr
omB-
Ial
lele
ata
freq
uenc
yof
�1%
to10
%
Chr
omat
inin
heri
tanc
e;siR
NA
-dir
ecte
dch
rom
atin
mod
ifica
tion
isin
volv
ed
Alle
man
etal
.(20
06),
Cha
ndle
r(2
007)
,C
hand
ler
etal
.(20
00),
Coe
(196
6),S
tam
etal
.(2
002)
Red
uced
pigm
enta
tion
(par
amut
atio
n)
r1(c
ompl
exlo
ci)
Var
iabl
eSp
onta
neo
us,
and
expo
sure
tova
ried
ligh
tdu
rati
ons
and
tem
pera
ture
DN
Am
eth
ylat
ion
Ch
andl
eret
al.
(200
0),
Mik
ula
(199
5),W
alke
ran
dPa
nav
as(2
001)
Red
uced
,lig
ht-
depe
nden
tpi
gmen
tatio
n(p
aram
utat
ion)
pl(g
ene-
epia
llele
)V
arie
sfr
omm
etas
tabl
eto
very
stab
lein
her
itan
ce
Spon
tan
eous
Ch
rom
atin
-mar
kin
gsu
gges
ted
Ch
andl
eret
al.
(200
0),
Hol
lick
and
Ch
andl
er(1
998)
,H
ollic
ket
al.
(199
5,20
00)
Red
uced
peri
carp
colo
rbu
tda
rkpi
gmen
tati
onat
the
poin
tof
silk
atta
chm
ent
(par
amut
atio
n)
p1(g
ene-
epia
llele
)A
tle
ast
5ge
ner
atio
ns
Spon
tan
eous
DN
Am
eth
ylat
ion
Coc
ciol
one
etal
.(20
01),
Das
and
Mes
sing
(199
4),
Raj
ande
epet
al.(
2007
),Si
dore
nko
and
Pete
rson
(200
1),S
ekho
net
al.
(200
7)Sp
otti
ng
ofke
rnel
sM
uDR
tran
spos
able
elem
ent
Sile
ncin
gef
fect
ofth
eM
uklo
cus
onM
uDR
was
mai
ntai
ned
over
atle
ast4
gene
ratio
ns,e
ven
whe
nM
ukha
dse
greg
ated
away
Cro
ssin
gle
adin
gto
silen
cing
ofth
ere
gula
tory
tran
spos
onM
uDR
DN
Am
eth
ylat
ion
;si
RN
As
are
invo
lved
inm
ain
tain
ing
her
itab
lem
eth
ylat
ion
stat
esin
Mu1
and
MuD
Rel
emen
ts
Lis
chet
al.
(200
2),
Slot
kin
etal
.(2
003)
,Sl
otki
net
al.
(200
5)
148 Volume 84THE QUARTERLY REVIEW OF BIOLOGY
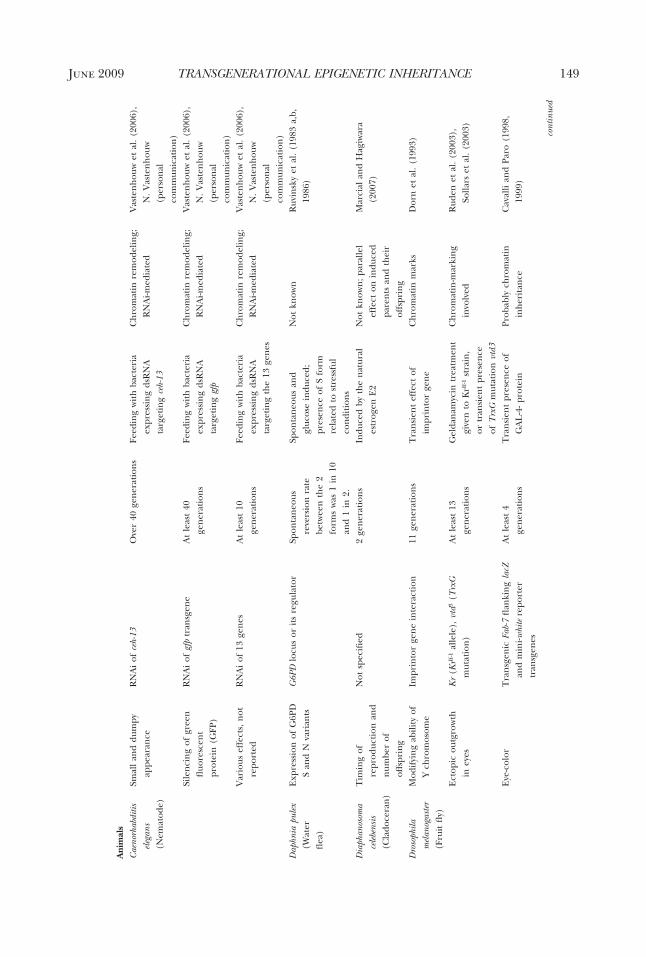
Ani
mal
sC
aeno
rhab
ditis
eleg
ans
(Nem
atod
e)
Smal
lan
ddu
mpy
appe
aran
ceR
NA
iof
ceh-
13O
ver
40ge
ner
atio
ns
Feed
ing
wit
hba
cter
iaex
pres
sin
gds
RN
Ata
rget
ing
ceh-
13
Ch
rom
atin
rem
odel
ing;
RN
Ai-m
edia
ted
Vas
ten
hou
wet
al.
(200
6),
N.
Vas
ten
hou
w(p
erso
nal
com
mun
icat
ion
)Si
len
cin
gof
gree
nfl
uore
scen
tpr
otei
n(G
FP)
RN
Ai
ofgf
ptr
ansg
ene
At
leas
t40
gen
erat
ion
sFe
edin
gw
ith
bact
eria
expr
essi
ng
dsR
NA
targ
etin
ggf
p
Ch
rom
atin
rem
odel
ing;
RN
Ai-m
edia
ted
Vas
ten
hou
wet
al.
(200
6),
N.
Vas
ten
hou
w(p
erso
nal
com
mun
icat
ion
)V
ario
usef
fect
s,n
otre
port
edR
NA
iof
13ge
nes
At
leas
t10
gen
erat
ion
sFe
edin
gw
ith
bact
eria
expr
essi
ng
dsR
NA
targ
etin
gth
e13
gen
es
Ch
rom
atin
rem
odel
ing;
RN
Ai-m
edia
ted
Vas
ten
hou
wet
al.
(200
6),
N.
Vas
ten
hou
w(p
erso
nal
com
mun
icat
ion
)D
aphn
iapu
lex
(Wat
erfl
ea)
Exp
ress
ion
ofG
6PD
San
dN
vari
ants
G6P
Dlo
cus
orit
sre
gula
tor
Spon
tan
eous
reve
rsio
nra
tebe
twee
nth
e2
form
sw
as1
in10
and
1in
2.
Spon
tan
eous
and
gluc
ose
indu
ced;
pres
ence
ofS
form
rela
ted
tost
ress
ful
con
diti
ons
Not
know
nR
uvin
sky
etal
.(1
983
a,b,
1986
)
Dia
phan
osom
ace
lebe
nsis
(Cla
doce
ran
)
Tim
ing
ofre
prod
ucti
onan
dn
umbe
rof
offs
prin
g
Not
spec
ified
2ge
ner
atio
ns
Indu
ced
byth
en
atur
ales
trog
enE
2N
otkn
own
;pa
ralle
lef
fect
onin
duce
dpa
ren
tsan
dth
eir
offs
prin
g
Mar
cial
and
Hag
iwar
a(2
007)
Dro
soph
ilam
elan
ogas
ter
(Fru
itfl
y)
Mod
ifyi
ng
abili
tyof
Ych
rom
osom
eIm
prin
tor
gen
ein
tera
ctio
n11
gen
erat
ion
sT
ran
sien
tef
fect
ofim
prin
tor
gen
eC
hro
mat
inm
arks
Dor
net
al.
(199
3)
Ect
opic
outg
row
thin
eyes
Kr
(KrIf
-1al
lele
),vt
d3(T
rxG
mut
atio
n)
At
leas
t13
gen
erat
ion
sG
elda
nam
ycin
trea
tmen
tgi
ven
toK
rIf-1
stra
in,
ortr
ansi
ent
pres
ence
ofT
rxG
mut
atio
nvt
d3
Ch
rom
atin
-mar
kin
gin
volv
edR
uden
etal
.(2
003)
,So
llars
etal
.(2
003)
Eye
-col
orT
ran
sgen
icFa
b-7
flan
kin
gla
cZan
dm
ini-w
hite
repo
rter
tran
sgen
es
At
leas
t4
gen
erat
ion
sT
ran
sien
tpr
esen
ceof
GA
L-4
-pro
tein
Prob
ably
chro
mat
inin
her
itan
ceC
aval
lian
dPa
ro(1
998,
1999
)
cont
inue
d
June 2009 149TRANSGENERATIONAL EPIGENETIC INHERITANCE
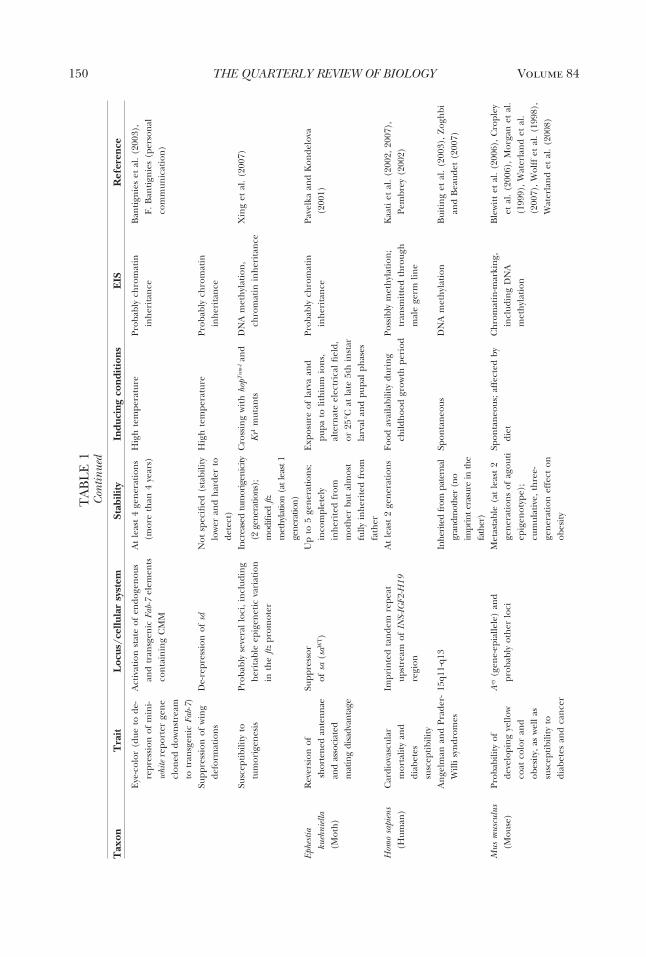
TA
BL
E1
Con
tinue
d
Tax
onT
rait
Loc
us/c
ellu
lar
syst
emSt
abili
tyIn
duci
ngco
ndit
ions
EIS
Ref
eren
ce
Eye
-col
or(d
ueto
de-
repr
essi
onof
min
i-w
hite
repo
rter
gen
ecl
oned
dow
nst
ream
totr
ansg
enic
Fab-
7)
Act
ivat
ion
stat
eof
endo
gen
ous
and
tran
sgen
icFa
b-7
elem
ents
con
tain
ing
CM
M
At
leas
t4
gen
erat
ion
s(m
ore
than
4ye
ars)
Hig
hte
mpe
ratu
rePr
obab
lych
rom
atin
inh
erit
ance
Ban
tign
ies
etal
.(2
003)
,F.
Ban
tign
ies
(per
son
alco
mm
unic
atio
n)
Supp
ress
ion
ofw
ing
defo
rmat
ion
sD
e-re
pres
sion
ofsd
Not
spec
ified
(sta
bilit
ylo
wer
and
har
der
tode
tect
)
Hig
hte
mpe
ratu
rePr
obab
lych
rom
atin
inh
erit
ance
Susc
epti
bilit
yto
tum
orig
enes
isPr
obab
lyse
vera
llo
ci,
incl
udin
gh
erit
able
epig
enet
icva
riat
ion
inth
eftz
prom
oter
Incr
ease
dtu
mor
igen
icity
(2ge
nera
tions
);m
odifi
edftz
met
hyla
tion
(atl
east
1ge
nera
tion)
Cro
ssin
gw
ith
hopT
um-l
and
Kr1
mut
ants
DN
Am
eth
ylat
ion
,ch
rom
atin
inh
erit
ance
Xin
get
al.
(200
7)
Ephe
stia
kueh
niel
la(M
oth
)
Rev
ersi
onof
shor
tene
dan
tenn
aean
das
soci
ated
mat
ing
disa
dvan
tage
Supp
ress
orof
sa(s
aWT)
Up
to5
gen
erat
ion
s;in
com
plet
ely
inh
erit
edfr
omm
oth
erbu
tal
mos
tfu
llyin
her
ited
from
fath
er
Exp
osur
eof
larv
aan
dpu
pato
lith
ium
ion
s,al
tern
ate
elec
tric
alfi
eld,
or25
°Cat
late
5th
inst
arla
rval
and
pupa
lph
ases
Prob
ably
chro
mat
inin
her
itan
cePa
velk
aan
dK
onde
lova
(200
1)
Hom
osa
pien
s(H
uman
)C
ardi
ovas
cula
rm
orta
lity
and
diab
etes
susc
epti
bilit
y
Impr
inte
dta
nde
mre
peat
upst
ream
ofIN
S-IG
F2-H
19re
gion
At
leas
t2
gen
erat
ion
sFo
odav
aila
bilit
ydu
rin
gch
ildh
ood
grow
thpe
riod
Poss
ibly
met
hyl
atio
n;
tran
smit
ted
thro
ugh
mal
ege
rmlin
e
Kaa
tiet
al.
(200
2,20
07),
Pem
brey
(200
2)
An
gelm
anan
dPr
ader
-W
illi
syn
drom
es15
q11-
q13
Inhe
rite
dfr
ompa
tern
algr
andm
othe
r(n
oim
prin
tera
sure
inth
efa
ther
)
Spon
tan
eous
DN
Am
eth
ylat
ion
Bui
tin
get
al.
(200
3),
Zog
hbi
and
Bea
udet
(200
7)
Mus
mus
culu
s(M
ouse
)Pr
obab
ility
ofde
velo
pin
gye
llow
coat
colo
ran
dob
esit
y,as
wel
las
susc
epti
bilit
yto
diab
etes
and
can
cer
Avy
(gen
e-ep
ialle
le)
and
prob
ably
oth
erlo
ciM
etas
tabl
e(a
tle
ast
2ge
ner
atio
ns
ofag
outi
epig
enot
ype)
;cu
mul
ativ
e,th
ree-
gen
erat
ion
effe
cton
obes
ity
Spon
tan
eous
;af
fect
edby
diet
Ch
rom
atin
-mar
kin
g,in
clud
ing
DN
Am
eth
ylat
ion
Ble
wit
tet
al.
(200
6),
Cro
pley
etal
.(2
006)
,M
orga
net
al.
(199
9),
Wat
erla
nd
etal
.(2
007)
,W
olff
etal
.(1
998)
,W
ater
lan
det
al.
(200
8)
150 Volume 84THE QUARTERLY REVIEW OF BIOLOGY
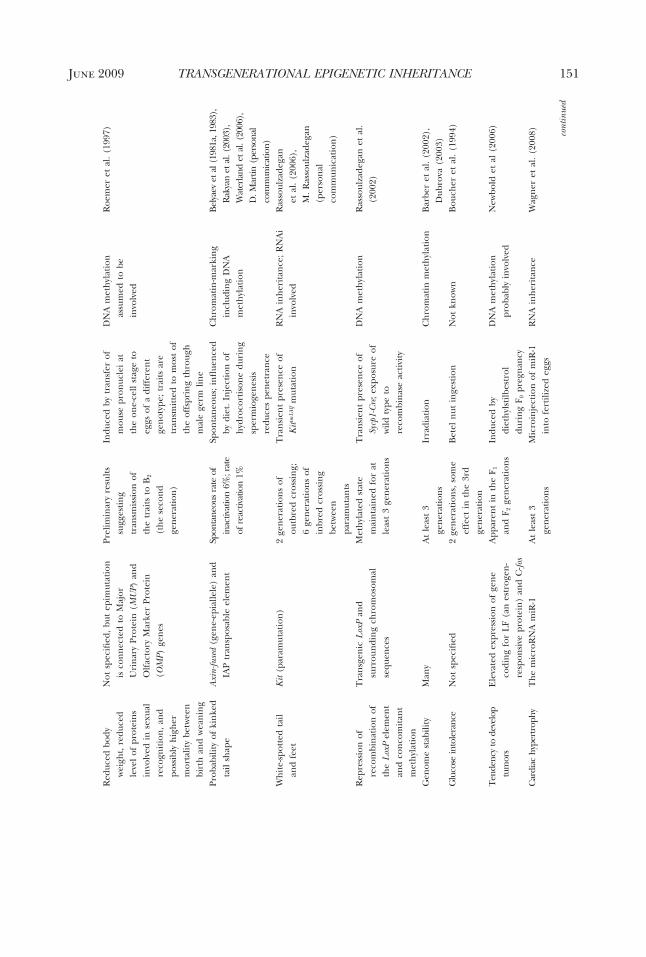
Red
uced
body
wei
ght,
redu
ced
leve
lof
prot
ein
sin
volv
edin
sexu
alre
cogn
itio
n,
and
poss
ibly
hig
her
mor
talit
ybe
twee
nbi
rth
and
wea
nin
g
Not
spec
ified
,bu
tep
imut
atio
nis
con
nec
ted
toM
ajor
Uri
nar
yPr
otei
n(M
UP)
and
Olf
acto
ryM
arke
rPr
otei
n(O
MP)
gen
es
Prel
imin
ary
resu
lts
sugg
esti
ng
tran
smis
sion
ofth
etr
aits
toB
2
(th
ese
con
dge
ner
atio
n)
Indu
ced
bytr
ansf
erof
mou
sepr
onuc
lei
atth
eon
e-ce
llst
age
toeg
gsof
adi
ffer
ent
gen
otyp
e;tr
aits
are
tran
smit
ted
tom
ost
ofth
eof
fspr
ing
thro
ugh
mal
ege
rmlin
e
DN
Am
eth
ylat
ion
assu
med
tobe
invo
lved
Roe
mer
etal
.(1
997)
Prob
abili
tyof
kin
ked
tail
shap
eA
xin-
fuse
d(g
ene-
epia
llele
)an
dIA
Ptr
ansp
osab
leel
emen
tSp
onta
neou
sra
teof
inac
tivat
ion
6%;r
ate
ofre
activ
atio
n1%
Spon
tan
eous
;in
flue
nce
dby
diet
.In
ject
ion
ofh
ydro
cort
ison
edu
rin
gsp
erm
ioge
nes
isre
duce
spe
net
ran
ce
Ch
rom
atin
-mar
kin
gin
clud
ing
DN
Am
eth
ylat
ion
Bel
yaev
etal
(198
1a,1
983)
,R
akya
net
al.(
2003
),W
ater
land
etal
.(20
06),
D.M
artin
(per
sona
lco
mm
unic
atio
n)W
hit
e-sp
otte
dta
ilan
dfe
etK
it(p
aram
utat
ion
)2
gen
erat
ion
sof
outb
red
cros
sin
g;6
gen
erat
ion
sof
inbr
edcr
ossi
ng
betw
een
para
mut
ants
Tra
nsi
ent
pres
ence
ofK
ittm1A
lfm
utat
ion
RN
Ain
her
itan
ce;
RN
Ai
invo
lved
Ras
soul
zade
gan
etal
.(2
006)
,M
.R
asso
ulza
dega
n(p
erso
nal
com
mun
icat
ion
)
Rep
ress
ion
ofre
com
bin
atio
nof
the
Lox
Pel
emen
tan
dco
nco
mit
ant
met
hyl
atio
n
Tra
nsg
enic
Lox
Pan
dsu
rrou
ndi
ng
chro
mos
omal
sequ
ence
s
Met
hyl
ated
stat
em
ain
tain
edfo
rat
leas
t3
gen
erat
ion
s
Tra
nsi
ent
pres
ence
ofSy
cp1-
Cre
;ex
posu
reof
wild
type
tore
com
bin
ase
acti
vity
DN
Am
eth
ylat
ion
Ras
soul
zade
gan
etal
.(2
002)
Gen
ome
stab
ility
Man
yA
tle
ast
3ge
ner
atio
ns
Irra
diat
ion
Ch
rom
atin
met
hyl
atio
nB
arbe
ret
al.
(200
2),
Dub
rova
(200
3)G
luco
sein
tole
ranc
eN
otsp
ecifi
ed2
gen
erat
ion
s,so
me
effe
ctin
the
3rd
gen
erat
ion
Bet
eln
utin
gest
ion
Not
know
nB
ouch
eret
al.
(199
4)
Ten
denc
yto
deve
lop
tum
ors
Ele
vate
dex
pres
sion
ofge
ne
codi
ng
for
LF
(an
estr
ogen
-re
spon
sive
prot
ein
)an
dC
-fos
App
aren
tin
the
F 1an
dF 2
gen
erat
ion
sIn
duce
dby
diet
hyl
stilb
estr
oldu
rin
gF 0
preg
nan
cy
DN
Am
eth
ylat
ion
prob
ably
invo
lved
New
bold
etal
(200
6)
Car
diac
hype
rtro
phy
Th
em
icro
RN
Am
iR-1
At
leas
t3
gen
erat
ion
sM
icro
inje
ctio
nof
miR
-1in
tofe
rtili
zed
eggs
RN
Ain
her
itan
ceW
agn
eret
al.
(200
8) cont
inue
d
June 2009 151TRANSGENERATIONAL EPIGENETIC INHERITANCE

TA
BL
E1
Con
tinue
dT
axon
Tra
itL
ocus
/cel
lula
rsy
stem
Stab
ility
Indu
cing
cond
itio
nsE
ISR
efer
ence
Myz
uspe
rsica
e(P
each
pota
toap
hid)
Loss
ofin
sect
icid
ere
sista
nce
Prob
ably
ampl
ified
resi
stan
cege
nes
Stab
lein
her
itan
ceof
lost
resi
stan
cein
clon
esth
ath
ave
ampl
ified
DN
A
Indu
ced
byD
NA
ampl
ifica
tion
DN
Am
eth
ylat
ion
invo
lved
Fiel
det
al.
(198
9)
Rat
tus
norv
egic
us(R
at)
Mod
ified
sero
toni
nco
nten
tin
imm
une
cells
g
Not
spec
ified
2ge
ner
atio
ns
Intr
amus
cula
rad
min
istr
atio
nof
�-e
ndo
rph
indu
rin
g19
thda
yof
preg
nan
cy
Not
spec
ified
Csa
baet
al.
(200
5)
Incr
ease
dex
pres
sion
ofge
nes
codi
ngfo
rm
etab
olic
fact
ors
Prom
oter
sof
PPA
R�
and
GR
inliv
er;
incr
ease
dex
pres
sion
ofot
her
RN
As
At
leas
t2
gen
erat
ion
sPr
otei
n-r
estr
icte
ddi
etdu
rin
gpr
egn
ancy
DN
Am
eth
ylat
ion
Bur
dge
etal
.(2
007)
,G
.B
urdg
e(p
erso
nal
com
mun
icat
ion
)D
ecre
ased
sper
mat
ogen
icca
paci
ty;e
leva
ted
inci
denc
eof
tum
or,
pros
tate
,and
kidn
eydi
seas
es,s
erum
chol
este
roll
evel
s,an
dim
mun
esy
stem
abno
rmal
ities
;pr
emat
ure
agin
gan
dm
ale
mat
ing
disa
dvan
tage
Met
hyl
atio
nst
ate
of15
diff
eren
tD
NA
sequ
ence
s.R
educ
edex
pres
sion
ofan
kyri
n28
,N
cstn
,R
ab12
,L
rrn6
aan
dN
CA
M1
foun
din
vin
cloz
olin
grou
pas
wel
las
incr
ease
dex
pres
sion
ofFa
dd,
Pbm
1b,
snR
P1c
and
Was
pip
Atl
east
3ge
nera
tions
;tr
ansm
issio
nth
roug
hth
em
ale
germ
line
Vin
cloz
olin
orm
eth
oxyc
hlo
rtr
eatm
ent
duri
ng
gest
atio
nD
NA
met
hyl
atio
nA
nway
etal
.(20
05,2
006a ,
2006
b ),C
hang
etal
.(2
006)
,Cre
ws
etal
.(20
07)
Alt
ered
gluc
ose
hom
eost
asis
Not
repo
rted
F 0–F
3ge
ner
atio
ns
Low
-pro
tein
diet
inF 0
,fr
omda
y1
ofpr
egn
ancy
thro
ugh
lact
atio
n
Not
spec
ified
,D
NA
met
hyl
atio
npr
obab
lyin
volv
edin
F 1an
imal
s
Ben
yshe
ket
al.(
2006
),D
.C.B
enys
hek
(per
sona
lco
mm
unic
atio
n)Vu
lpes
vulp
es(F
ox)
Pieb
ald
spot
tin
gA
ctiv
atio
nst
ate
ofSt
arge
ne
Star
(sem
idom
inan
tal
lele
)ac
tivat
edin
�1%
ofdo
mes
ticat
edan
imal
s;in
heri
ted
for
mor
eth
an2
gene
ratio
ns
Spon
tan
eous
inta
med
foxe
sra
ised
infu
rfa
rms;
hor
mon
alst
ress
sugg
este
df
Poss
ibly
her
itab
lech
rom
atin
mod
ifica
tion
Bel
yaev
etal
.(1
981b
),T
rut
etal
.(2
004)
Not
e:T
he
spec
ies
inea
chca
tego
ryar
eor
dere
dal
phab
etic
ally
.R
efer
ence
sto
the
tabl
ear
eav
aila
ble
onlin
eat
The
Qua
rter
lyR
evie
wof
Bio
logy
hom
epag
e,w
ww
.jour
nal
s.uc
hic
ago.
edu/
QR
B.
a Sim
ilar
syst
ems
hav
ebe
ende
scri
bed
for
arab
inos
e-ut
iliza
tion
inE.
coli
(Kh
lebn
ikov
etal
.20
00)
and
the
lact
ose
oper
onin
Salm
onel
laen
teri
caty
phim
uriu
m(T
olke
r-N
iels
enet
al.
1998
).b P
roba
bly
occu
rsin
man
ypa
thog
ens
(Lew
is20
07).
c It
seem
sth
atal
lcili
ates
show
cort
ical
inh
erit
ance
and
guid
edas
sem
bly
ofco
rtic
alst
ruct
ures
(Fra
nke
l198
9).C
iliat
esh
ave
asi
len
tm
icro
nuc
leus
and
anac
tive
mac
ron
ucle
us,f
rom
wh
ich
non
codi
ng
sequ
ence
sar
eex
cise
dan
dco
din
gse
quen
ces
are
ampl
ified
.Fol
low
ing
con
juga
tion
orau
toga
my,
an
ew(z
ygot
ic)
mac
ron
ucle
usis
form
edfr
omth
efu
sed
mei
otic
prod
uct
ofth
em
icro
nuc
leus
,an
dth
eol
d,m
ater
nal
mac
ron
ucle
usde
gen
erat
es.
Th
eco
mpl
expr
oces
ses,
guid
edby
the
mat
ern
aln
ucle
us,
that
lead
toth
ein
her
itan
ceof
patt
ern
sof
chro
mos
omal
rear
ran
gem
ents
inth
ezy
goti
cn
ucle
usse
emto
bech
arac
teri
stic
ofal
lci
liate
spec
ies
(Mey
eran
dC
hal
ker
2007
).d S
imila
rph
enom
ena
are
foun
din
C.
galb
orat
a,C
.tr
opic
alis
,C
.pa
rpsi
losi
s,an
din
the
basi
omyc
etes
fun
gus
Cry
stoc
occu
sne
ofor
man
s.e I
nte
rspe
cifi
cpl
ant
hyb
rids
,an
dso
met
imes
hyb
rids
betw
een
cult
ivar
s,di
spla
yep
igen
etic
vari
atio
ns
that
are
her
itab
leac
ross
gen
erat
ion
s.W
hen
hyb
ridi
zati
onis
follo
wed
bypo
lypl
oidi
za-
tion
,th
isse
ems
tobe
an
orm
alan
din
vari
ant
gen
omic
resp
onse
.In
The
Bio
logi
calJ
ourn
alof
the
Lin
nean
Soci
ety
82(4
)(A
llen
2004
),m
any
exam
ples
ofep
igen
omic
effe
cts
ofh
ybri
diza
tion
inpl
ants
,in
clud
ing
mai
ze,
wh
eat,
rice
,co
tton
,an
dsu
nfl
ower
,ar
ere
view
ed.
f Th
ere
are
sim
ilar,
olde
rst
udie
sof
tran
sgen
erat
ion
alef
fect
sfo
llow
ing
adm
inis
trat
ion
ofh
orm
ones
and
drug
sin
ava
riet
yof
mam
mal
s.T
hes
est
udie
s,w
hic
hdi
dn
otin
clud
em
olec
ular
data
,w
ere
revi
ewed
byC
ampb
ell
and
Perk
ins
(198
8).
152 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

molecular data show that the gametes ofthe F1 generation have acquired alteredepigenetic marks, the transmission of epi-genetic variations rather than the directinduction of such variations is a reasonableassumption. Based on this, we includedsome cases in our table in which only twogenerations of transmission following in-duction were reported. For instance, weincluded a case reported by Burdge et al.(2007) in which protein restriction duringa rat grandmother’s pregnancy led to iden-tical somatic and epigenetic (methylation)variations in her F1 and F2 descendants, aswell as a case reported by Newbold et al.(2006) that showed that F0 female rats ex-posed to diethylstilbestrol had F1 and F2
offspring with an increased susceptibility totumors associated with persistent changesin the DNA methylation and expression ofan oncogene (see Ruden et al. 2005 for anepigenetic model explaining these data).The case of germline inherited epigeneticvariants (and their corresponding pheno-types) that were acquired stochastically inthe Avy locus of the mouse was includedsince transgenerational epigenetic effectson coat color and obesity were reported.However, this is a complex case that seemsto involve epigenetic modifications in theAvy locus as well as in other loci in the ge-nome (Blewitt et al. 2006; Cropley et al.2006; Waterland et al. 2007, 2008). An-other case we included was that reportedby Csaba et al. (2005) in which the modi-fied serotonin content induced in the im-mune cells of mice treated with endomor-phin during pregnancy is subsequentlytransmitted to their grandchildren. We didnot include the many cases of single gen-eration inheritance, even though epige-netic inheritance seems quite plausible insome of them, as, for example, when apredisposition to cancer in humans is re-lated to an epimutation (an epigenetic he-reditary abnormality in gene expression)in a mismatch-repair gene, and this epimu-tation is transmitted from mother to son(Hitchins et al. 2007). We also excluded asimilar case in mice which showed that tu-mor risk was increased following chro-mium III chloride exposure and that this
risk was then transmitted from father toson (Shiao et al. 2005). Other cases ofsingle-generation inheritance, such as pro-tection against type I diabetes in humans(where a paramutation-like process hasbeen reported) (Bennett et al. 1997), hy-drostatic pressure-induced alterations inDNA methylation in Japonica rice (Long etal. 2006), and parallel alterations in geneexpression profiles in White Leghornchickens and their offspring followingstress in the parental generation (Lindqvistet al. 2007), were also excluded, althoughepigenetic inheritance may well have oc-curred and may be revealed when subse-quent generations are studied. However,within the limitations imposed by the re-search designs of the studies we reviewedand the qualifications we have mentioned,we believe that the table provides a fairlyexhaustive overview of the recognized casesof cellular transgenerational epigeneticinheritance that have been described inEnglish-language journals, although, inev-itably, we likely missed some cases.
taxonomic distribution and inducingconditions
The data in the table probably representthe tip of a very large iceberg. What ismissing from the table is important, be-cause the absences point to gaps that needto be filled. For instance, there is no infor-mation about epigenetic inheritance in thekingdom Archea, and most phyla are notrepresented. There are also few data di-rectly addressing epigenetic inheritance inviruses, although it may plausibly be as-sumed that viruses exploit and use the epi-genetic adaptations adopted by their hosts.Data on epigenetic inheritance in chloro-plasts and mitochondria are also veryscant. It is worth noting that the organismsthat show the greatest evidence for epige-netic inheritance are the classical modelorganisms of genetics—E. coli, yeast, Arabi-dopsis, maize, rice, Caenorhabditis, Drosoph-ila, the mouse, and the rat. However, asystematic investigation of epigenetic in-heritance in different conditions is not yetavailable for any of these model organisms.It is also worth noting that all the model
June 2009 153TRANSGENERATIONAL EPIGENETIC INHERITANCE

animals studied belong to taxa in whichthe segregation between germline andsoma occurs early, and, therefore, epige-netic inheritance may be more limitedthan in the non-represented animal taxawhere segregation occurs late if it occurs atall (Buss 1987; Jablonka and Lamb 1995).Although the non-systematic way in whichthe data were collected precludes generalconclusions, it seems as if epigenetic inher-itance in multicellular organisms is mostcommon in plants and fungi. This is prob-ably in part due to the lack of segregationbetween soma and germline in thesegroups that enables developmentally in-duced epigenetic variations occurring insomatic cells to be transferred to the ga-metes when these somatic cells assumegermline functions. However, there aretwo, additional considerations that may berelevant to the difference between animals,on the one hand, and plants and fungi, onthe other. First, the lack of nervous system-directed mobility and activity in plants andfungi means that they cannot adapt tochanging conditions behaviorally; if theconditions experienced by offspring arelikely to be similar to those of their par-ents, then inheriting epigenetic adapta-tions from them is an alternative adaptivestrategy to behavior and is likely to be pos-itively selected in plants and fungi (Jab-lonka et al. 1995; Jablonka and Lamb 1995;Lachmann and Jablonka 1996). Second,mobility and CNS-dependent flexible learn-ing in animals may often limit the predict-ability of the environment in descendentgenerations; therefore, wide-ranging stableepigenetic cellular inheritance throughthe germline may be selected against. Ingeneral, it seems that the difference be-tween the life strategies of plants and ani-mals may account for the observation thatepigenetic inheritance in multicellular or-ganisms is more common in plants andfungi than among animals.
The relative importance—and some-times even the very presence—of particu-lar EISs in different taxa varies. Buddingyeast seems to lack the RNAi system, soepigenetic inheritance based upon it is im-possible, although other types of RNA-
mediated inheritance cannot be ruled out.In some groups of animals, DNA methyl-ation appears to have been lost (Regev etal. 1998) and is not part of chromatinmarking in these organisms. Transgenera-tional structural inheritance and inheri-tance through self-sustaining loops seem tobe more common in unicellular organismsand in fungi where horizontal transfer ofinformation through hyphal interaction iscommon. It may be that the developmentof a germline involves such drastic alter-ations in cellular functions and structuresthat self-sustaining loops and many cellularstructures are destabilized and disrupted.The data presented in Table 1 lend sup-port to this conjecture, although their pau-city precludes decisive conclusions.
To understand why and when cellularepigenetic variants are inherited, we needto know the conditions that promote theirinduction and stability in cell lineages.Their developmental nature requires anapproach that is sensitive to context. How-ever, our knowledge of the chromatin-marking and RNA-mediated systems sug-gests that certain parts of the genome mayexhibit chromatin- and RNA-mediated epi-genetic inheritance more often than oth-ers. Repetitive DNA sequences (especiallyregions that code for RNAs that can formdsRNA or stem-loop structures), DNA re-gions where transcription is likely to startfrom both complementary strands, and re-peated chromosomal segments that pairectopically are all likely to exhibit RNA-mediated epigenetic inheritance under awide range of conditions. These and otherrepeated sequences that cooperatively bindprotein complexes and regions with CG dou-blets are all likely candidates for multigen-erational chromatin- and methylation-basedepigenetic inheritance.
The inducibility and transmissibility of epi-genetic variants depend on developmentalconditions. Conditions of stress seem to beparticularly important as inducers of herita-ble epigenetic variation, and lead to changesin epigenetic and genetic organization thatare targeted to specific genomic sequences.We mentioned earlier that the genomicstresses of allopolyploidization and, to a
154 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

lesser extent, autopolyploidization leadto epigenetic and genetic re-patterning(Grant-Downton and Dickinson 2005,2006; Rapp and Wendel 2005). A well-known epigenetic phenomenon associatedwith hybridization is nucleolar dominance,or the expression in the hybrid of therRNA gene complex (NOR) from only oneparent. The preferential silencing of oneNOR is a large-scale gene-silencing phe-nomenon associated with heritable DNAmethylation and repressive histone modifi-cations (Pikaard 2000, 2003). However, theepigenetic changes occurring in hybridsare not restricted to rRNA genes. An ex-ample we present in the table is that of thegenome-wide changes that occur in syn-thetic wheat hybrids that were formed inorder to simulate the evolution of domes-tic wheat. Levy and Feldman (2004) re-viewed evidence showing that, in these hy-brids, 13% of the genome undergoessignificant methylation changes, whilechanges also occur in genome organiza-tion (e.g., rearrangements and eliminationof some sequences). The methylationchanges affect both low copy numbers andrepetitive DNA sequences, and are associ-ated with heritable transcriptional silenc-ing. In addition, the activation of retroele-ments leads to heritable alterations in geneexpression at other loci, thus resulting inmajor changes in the profile of gene ex-pression. The changes in the epigeneticstate of the genome are region and chro-mosome specific: they are targeted to par-ticular genomic sequences and reoccur,with localized variations, upon repeatedformation of the same type of al-lopolyploid. In species of the cordgrassSpartina, genome-wide epigenetic and ge-netic changes were observed in two re-cently formed, morphologically different,natural hybrids (and an allopolyploid), inwhich 30% of the parental methylation pat-terns were altered, in addition to similarstructural changes in the DNA sequences ofthese two independently formed and genet-ically similar hybrids (Salmon et al. 2005).Many similar effects of allopolyploidizationhave been reviewed in the extensive andgrowing literature on plant polyploidy (e.g.,
see The Biological Journal of the Linnean Society82(4) [Allen 2004]). The overall impressiongained from these studies is that heritableepigenetic changes accompany the firststages of allopolyploidization, and that thetypes of repetitive sequences in the parentalspecies, the amount of divergence betweenthem (especially with regard to elements thatmay be involved in epigenetic control), andthe direction of the cross all play importantroles in specifying the extent and nature ofepigenetic re-patterning.
Another form of genomic stress that maylead to heritable variation is that associatedwith a change in reproductive mode. Forexample, the transition from sexual toa-gametic reproduction in sugar beet leadsto heritable activation of some genes (Le-vites 2000). DNA damage also leads to her-itable epigenetic changes, and researchersare beginning to uncover some of the fac-tors that affect this response. FollowingDNA repair, the epigenetic structure of therepaired region is not fully reconstructedand carries with it a repair-specific chroma-tin signature that can be transmitted tosubsequent generations (Polo et al. 2006).Moreover, the loading of the histone vari-ant �-H2AX (which is associated with re-paired DNA segments) with cohesin leadsto sister-chromatid interactions that maycontribute to the radiation-induced ge-nome instability that arises and is inheritedfor several generations in the progeny ofdamaged cells (Little 2003). This may bethe basis for the heritable genomic insta-bility found in the offspring and grand-offspring of male mice that were exposedto irradiation (Dubrova 2003), as well asfor the increased level of recombinationseen for at least four generations afterUV-C irradiation of Arabidopsis plants (Mo-linier et al. 2006).
Some of the cases included in Table 1show that physiological stresses—for in-stance, nutritional stresses imposed duringsensitive periods in the development offlax (Cullis 2005)—can lead to both ge-netic and epigenetic re-patterning, andboth types of re-patterning seem to be cor-related and share a common mechanisticbasis. It seems likely that other cases in
June 2009 155TRANSGENERATIONAL EPIGENETIC INHERITANCE

which an environmental stressor has tar-geted effects on genome organization,such as the heat-induced changes in rRNA-encoding DNA repeats in Brassica (Watersand Schaal 1996), will also be found tobe associated with heritable epigeneticchanges in the genes or repeated elementsinvolved. In animals, alterations in hor-monal balance, especially those occurringover several generations, may also be fol-lowed by epigenetic changes. This may bethe basis for the pattern of inheritance ofwhite spotting seen in domesticated silverfoxes (as we will discuss later) (Trut et al.2004). However, stressful conditions maynot only affect the chromatin-marking andthe RNA-mediated EISs; if a new prion vari-ant can be generated in stressful condi-tions, it might cross previously existing spe-cies barriers and have novel effects in itsnew “host” species.
epigenetically-based similarities anddifferences between generations:
inducing epigenetic variationsin the germline
Many of the studies of multicellular,sexually reproducing organisms that wepresent in Table 1 show that, as a result ofan inducing stimulus or of changed condi-tions in the F0 (parent) generation, similarchromatin marks and similar phenotypesare reconstructed in subsequent genera-tions. However, the F0 generation itselfmay not show any phenotypic effects;changes in epigenetic marks and associ-ated somatic phenotypes may first appearin the F1 generation and may only then beinherited by subsequent generations. Forexample, in mice, the diethylstilbestrol-induced increase in the probability of de-veloping tumors appeared only in the F1
and F2 generations (Newbold et al. 2006).Understanding the inheritance of in-
duced variation in sexually reproducingmulticellular organisms is an importanttopic of research, not least because, in thepast, much of the debate about the impor-tance of acquired characters in evolutionrevolved around this issue (Delage andGoldsmith 1912). Traditionally, in multi-cellular organisms with a germline, three
types of induced heritable effects havebeen distinguished: direct induction, par-allel induction, and somatic induction.With direct induction, the germline is di-rectly affected without any effect on the F0
parent’s soma, while with parallel induc-tion, similar somatic phenotypic effects areapparent in both the induced ancestor andits descendants, but the induction events inthe somatic and germ lineages are inde-pendent. Finally, with somatic induction, achange is induced in the soma, and thissomatic effect causes a change in the germ-line that reconstructs the somatically-induced parental phenotype in the descen-dants (Fothergill 1952; Jablonka and Lamb1995). There is, however, a fourth possibil-ity: an induced effect on the soma of the F0
generation may cause changes in the germ-line, but the resulting somatic changes indescendants are dissimilar from the effecton the soma in the F0 generation. This is acase of parallel induction with nonparalleleffects. The different types of induced her-itable effects are represented schematicallyin Figure 2.
The literature shows that direct induc-tion, parallel induction, and parallel induc-tion with nonparallel or partially paralleleffects are common. In Table 1, manyof the cases of paramutation and of in-duced, heritable, transposable element ac-tivity can be classified as direct inductionor parallel induction, because the induc-ing conditions directly affect events in thegermline (and sometimes, in parallel, inthe soma as well). Changes in hormonedynamics that specifically target the germ-line will either not affect somatic charac-ters in the induced F0 generation, or willaffect them in a way that is unrelated to theeffect seen in the F1. For example, in onecase, vinclozolin, an androgen suppressor,induced testis disease in at least three gen-erations of males following its administra-tion to a pregnant female ancestor (Anwayet al. 2005, 2006a,b). This is an example ofdirect induction with differing effects inmales and females. Parallel induction isseen in the case in which nutritional andtemperature changes affected morphology
156 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

in the F0 generation of flax, and similarphenotypic effects were found in subse-quent generations (Cullis 2005). Anothercase of parallel induction is that seen in themoth Ephestia kuehniella, where subjectingthe insect to modified temperature condi-tions, lithium ion treatment, or an alterna-tive electric field during the first half of itspupal development resulted in the sup-pression of a mutant short antennae in thetreated generation and five subsequentgenerations (Pavelka and Koudelova 2001).Also, glucose induction of the S form of theG6PD enzyme in Daphnia pulex may be illus-
trative of a further possible case of parallelinduction (Ruvinsky et al. 1983a,b).
The distinction between direct and par-allel induction is not always straightfor-ward, as epigenetic changes may occur inonly some of the somatic cells of the in-duced parent, thus resulting in a varie-gated somatic phenotype. This can be de-scribed as partial parallel induction. It canbe seen in some cases of paramutation inplants where, when the induction rate ishigh, sectors that are the result of paramu-tation in somatic tissues can be seen in theF0 generation (Chandler et al. 2000).
Figure 2. Inducing an Inherited Effect(A) Direct germline induction: An external stimulus induces a germline change from G0 to G1 with no effect on
the parental soma, which remains S0. The G1 state is inherited, and leads to the development of an S1 soma.(B) Parallel induction: An external stimulus induces a change in the parent’s soma from S0 to S1 and in itsgermline from G0 to G1. The G1 state is inherited and causes the development of an S1 soma in descendants.(C) Somatic induction: An external stimulus induces a change in the parent, altering its somatic phenotype fromS0 to S1. The effect is transmitted from the S1 soma to the germline, where G0 is changed into G1; G1 isconsequently inherited and results in the development of an S1 soma. (D) Parallel induction with nonparalleleffects: An external stimulus alters the soma from S0 to S2, and the germline from G0 to G1. The germlinemodification is inherited and leads to the development of S1 soma in subsequent generations. With all fourtypes of induction, S1 could have an effect on G1 in all descendants of the original induced parents (not shown).
June 2009 157TRANSGENERATIONAL EPIGENETIC INHERITANCE

Several cases in the table can be inter-preted as instances of somatic induction inwhich heritable variation is induced in thesoma and is seen as a somatic character,and the resulting effect is then transferredfrom the soma to the germline. The abilityof small RNAs to move from cell to cell mayfacilitate soma to germline informationtransmission and may form the basis ofsuch inheritance. The most impressive ex-ample of somatic induction via small RNAsmay be found in the case discussed by Vas-tenhouw et al. (2006), in which C. eleganswere fed bacteria with DNA sequences cod-ing for dsRNA, and the RNAs migratedfrom the somatic cells of the nematode to itsgerm cells, thus affecting subsequent gener-ations. Steele et al. (1998) suggested anotherroute of transmission that is initiated byRNA: the transfer of RNA transcripts fromthe immune cells of mammals to their germ-line, followed by reversed transcription andincorporation of the reverse transcribedDNA into the germline genome. Zhivotovsky(2002) has modeled the conditions thatcould lead to the evolution of such a system.
Another route of somatic induction is viainducing conditions that affect the secre-tion of hormones, which, in turn, affect thegermline. The effect of a hormone on thesomatic characters of the induced parent(F0) and on its descendants may be similar,but it is unlikely to be identical. This isbecause even if the same genes are affectedin the soma of the adult F0 animals as wellas in the F1 and subsequent generations ofoffspring, it is unlikely that the pattern ofactivity of these genes will be the same. Indescendants, the induced epigenetic changemay be expressed during embryonic as wellas adult stages, whereas, in the F0 animals,the change is induced in adults. Only inrare cases in which the pattern of timingand spatial expression is identical in the F0
and subsequent generations—and is lim-ited to the stage at which it was induced inthe F0—will identical somatic charactersoccur in parent and offspring generations.An additional complication is the sex ofthe F0 animals and their offspring, as sex-limited effects will obviously be transmittedin a sex-specific manner, and epigenetic
variations in an induced F0 parent maytherefore be different for descendants ofthe opposite sex. These considerations arerelevant to any kind of somatic induction,no matter the mechanism behind it, so,although partial similarity is likely, we ex-pect that somatic induction leading toidentical phenotypes in the F0 and subse-quent generations will be rare.
The foregoing discussion suggests thatthe traditional distinctions between direct,parallel, and somatic induction do not sat-isfactorily describe the possible interac-tions between the soma and germline. Thesimilarity between the somatic characteris-tics of an induced ancestor and its descen-dants, which the traditional classificationhighlights, is, of course, of interest, but wethink that the mode and mechanisms ofthe induction of germline variations ratherthan their effect (i.e., the similarity or lackof similarity between the F0 and the F1)should be the focus of study. An importantaspect of the problem is whether externalinducing conditions affect the germline di-rectly or whether their effect is mediatedthrough the soma. External environmentalconditions that are independent of the or-ganism’s activity and its development canhave important heritable epigenetic ef-fects, but when there are somatic mediat-ing signals (e.g., RNAs or hormones) thesesomatic signals may evolve to efficientlycommunicate information to the germ-line. We therefore expect that develop-mentally-mediated somatic effects mayhave adaptive consequences more oftenthan signals that act on the germline di-rectly, and that when there are suchsoma-mediated influences on the germ-line, the effects on the somatic cells ofthe F0 will often differ from those thatwill be seen in the next generations (Fig-ure 2D).
The possibility of a somatic effect on thegermline that is mediated by hormoneswas raised shortly after hormones were dis-covered. The Austrian Lamarckian zoolo-gist Paul Kammerer and the pioneer endo-crinologist Eugen Steinach (1920) foundthat exposing male rats to high tempera-tures led to morphological and physiolog-
158 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

ical changes in their offspring and grand-offspring. They suggested that the presence ofhormone-secreting interstitial cells adjacentto germ cells in the gonads facilitated hor-monal interactions between them, andthey claimed that heat produced a changein hormone production in interstitial cells,thus affecting germline cells and carryinghereditary consequences (see Logan 2007for a discussion of Kammerer and Stein-ach’s work). Although the validity of thisclaim and its possible interpretation interms of epigenetic inheritance is at presentunclear, the possibility that there are hor-monal effects on epigenetic variation is nolonger considered heresy.
The experimental work of Vanyushin etal. (2006a) has shown that methylation pat-terns in the rat genome are controlled byhydrocortisone dynamics, and that phyto-hormones of different classes cause a de-cline in global DNA methylation and therepression of de novo methylation in plants.Moreover, the evidence reviewed by Nazand Sellamuthu (2006) suggests that, de-spite doubts about some of the reportedinformation, there are 8 hormone recep-tors and 16 cytokine/growth receptors inmature ejaculated sperm, thereby allowingfor the possibility that hormones couldexert their effects on male gametes.Hormone and neurotransmitter recep-tors have also been found in oocytes and infemale germline cells. For example, theoestrogen receptor (Wu et al. 1992), sero-tonin receptor 5-HTID (Vesela et al. 2003),Notch1 and Notch2 receptors (Cormier etal. 2004), and the �2-andrenoceptor (Ci-kos et al. 2005) have all been detected inoocytes, and the GH receptor has beendetected in fertilized eggs (Pantaleon et al.1997; Kolle et al. 2001). It is interesting tonote that the changes in expression ofsome of the genes coding for these recep-tors coincide temporally with early waves ofepigenetic re-programming during devel-opment. The presence of hormone recep-tors in gametes, and the modulation ofreceptors’ synthesis during sensitive devel-opmental periods when hormonal changesoccur, suggest that induced variations inhormonal conditions may affect the epige-
netic state of genes within germline cells,and these, in turn, can be transmitted tothe next generation.
The involvement of hormones in the in-duction of heritable epigenetic variationsis no longer a mere speculation: several ofthe mammalian examples presented in Ta-ble 1 suggest that changes in hormonalstimuli induce heritable epigenetic changes.For example, the penetrance of the fusedphenotype is altered in the progeny ofmouse parents treated with hydrocortisone(Belyaev et al. 1983). In silver foxes se-lected for tame behavior, hormonal effectsin the serotonin system that controls ag-gression seem to be involved in the herita-ble activation of the star gene that leads towhite spotting (Belyaev et al. 1981a,b; Trutet al. 2004; Popova 2006). The best inves-tigated case of hormonally-mediated ef-fects on epigenetic marks is that of thetransgenerational effect of the estrogenicandrogen disruptors vinclozolin and me-thoxychlor on testes development in malerats (Anway et al. 2005, 2006a,b; Chang etal. 2006; Crews et al. 2007). With vinclozo-lin, 15 different DNA sequences isolatedfrom sperm were shown to have alteredmethylation patterns, and these patternswere transmitted to the F1-F3 generationsof offspring of treated F0 females. Cru-cially, vinclozolin was effective only whenadministered between 8 and 15 days postcoitum, and had no effect when adminis-tered later, between 15 and 20 days. Thissensitive developmental period coincideswith the epigenetic remethylation phase inthe male (Hajkova et al. 2002), thus sug-gesting that the hormonal effect of andro-gens is developmentally specific (limited tothis period of epigenetic reprogramming)and is not a general toxic effect. If so,modifications in these methylation pat-terns in the soma of F0 vinclozolin-treatedfemales are not expected. In plants, whereno epigenetic reprogramming phase simi-lar to that in mammals is apparent, and thegermline is continuously produced duringdevelopment (Matzke and Scheid 2007),changes in hormonal stimulations duringall phases of somatic development are
June 2009 159TRANSGENERATIONAL EPIGENETIC INHERITANCE

likely to affect epigenetic variations in thegermline.
epigenetic recall and otherdirectional changes in heritable
epigenetic marksWe defined cellular epigenetic inheri-
tance as the transmission from mother cellto daughter cell of variations that are notthe result of differences in DNA sequence,or of persistent inducing signals in thecell’s environment. The examples pre-sented in Table 1 support our assertionthat when epigenetic marks are inherited,the same pattern of marks is more or lessfaithfully reconstructed across generations.If a particular mark—for example, a pat-tern of 5 methylated cytosine sites—is in-duced at a particular locus in the germline,this pattern is then reconstructed (with acertain error rate) in the descendants andhas similar phenotypic effects (Figure 3A).Stable epigenetic inheritance is at oneextreme pole, and total reset to a singledefault state—that of the uninduced par-ents—is at the other pole of developmen-tally influenced inheritance. However, thedevelopmental nature of epigenetic inher-itance and our knowledge of the construc-tion of chromatin marks suggest that weconsider other possibilities; the examplesin the table, therefore, represent only avery small fraction of the types of epige-netic hereditary phenomena that probablyexist.
The first possibility is that of partial re-construction—an intermediate between thetwo extreme poles of complete reset andfaithful reconstruction. For example, ofthe 5 induced methylation sites, only 3 arereconstructed in the offspring, and, in theabsence of the inducing stimulus, if thephenotype of the offspring is the same asthat of an uninduced individual, this wouldnot be seen as a case of epigenetic inheri-tance, even if the threshold for the devel-opmental response was lowered or thespeed of reaction was enhanced in descen-dants (see Figure 3B). The situation is sim-ilar to that found with neural memory,when the original stimulus that leads to theinitial learnt response is required to trigger
the response again, but because memorytraces remain, there is recall—facilitatedreconstruction of the learnt response uponre-induction. We suggest that the inheri-tance of some epigenetic memory tracesmay lead to epigenetic recall—a facilitatedresponse in descendants that requires aninducer. The inherited, partial epigeneticpatterns that facilitate a response arecalled epigenetic engrams. (Engram is a termthat was invented by Richard Semon in1904, to mean, roughly, “memory trace”;see Schacter 2001.) In order to recognizeepigenetic recall, the kinetics of inducedresponses in the F1 and subsequent gener-ations need to be studied. None of theexamples in Table 1 fulfils this require-ment, and, to the best of our knowledge,this kind of investigation—searching forepigenetic engrams and for facilitated butstill inducer-dependent responses—has notbeen part of the research program of epi-genetics. Such responses, however, arelikely to be common, as the mechanismsfor them are all in place, and a systemenabling the reconstruction of epigeneticengrams that allow recall would be selec-tively advantageous in many conditions,just as, in spite of the different timescale,the evolution of neural sensitization hasbeen favored. Agrawal et al. (1999) studiedinduced defenses against predators in wildradish (Raphanus raphanistrum) and the wa-ter flea (Daphnia cucullata), and showedthat induction in the parental generationmade offspring better adapted to preda-tors than the offspring of uninduced par-ents. They suggested that the persistent pa-rental effect in radish plants may be eithera direct maternally-induced effect or theresult of more rapid induction of plantdefenses in the offspring of damagedmothers. If the latter proves correct, it willrepresent a case of epigenetic recall, un-derlain by as yet uncharacterized epige-netic engrams. (For an extended discus-sion of the possibility of learning in cellsand non-neural organisms, see Ginsburgand Jablonka 2008.)
In addition to epigenetic recall based onpartial reconstruction, we must also con-sider the possibility that although induced
160 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

Figure 3. Types of Transgenerationally Inherited Epigenetic EffectsAn inactive gene (gray rectangle) and its corresponding phenotype are depicted at the top of the figure. (A)
Epigenetic copying and corresponding phenotypic inheritance: An epigenetic mark consisting of 5 � sites is inducedin the parent and affects the marking of the gene and the phenotype of the induced individual. (The “�” signsindicate an altered methylation or histone modification site, and the inducer is indicated by a curved arrow.)The epigenetic mark is reliably transmitted through the germline, thus leading to a modified heritablemorphology in the uninduced progeny (heritable site-states are indicated by “�” signs within the rectangle, andthe straight black arrows indicate transitions between generations). (B) Epigenetic recall: Partial inheritance ofthe epigenetic pattern (represented by 3 internalized “�” signs) that was established in the induced parentdoes not lead to modified morphology in progeny. However, the amount of inducer needed to re-establish thefull epigenetic pattern (5 � pattern) and the corresponding induced phenotype is much smaller than in theparent (the smaller curved arrow indicates low level stimulation). (C) Reactive but dissimilar effects of inheritedepigenetic patterns: (i) antagonistic - The parental epigenetic mark (5 � signs) is inherited faithfully, but, in amismatched postnatal environment (triangle around progeny), it leads to a different phenotype in theprogeny. (ii) accumulative - Following recurrent induction in each generation, epigenetically modified and“internalized” sites accumulate, and result in correspondingly more extreme phenotypes. When the epigeneticpattern reaches a certain configuration (5 � internalized sites), it is inherited even in the absence of theinducer, and this is a form of epigenetic assimilation. (iii) lingering-fading - Following induction, the mark andits corresponding morphology are established, but fade away gradually in subsequent generations in a nonin-ducing environment.
June 2009 161TRANSGENERATIONAL EPIGENETIC INHERITANCE

marks in one generation may be faithfullyinherited, they might lead to a non-matchingyet predictable phenotype in the subse-quent generation if the environments ofparent and progeny are drastically differ-ent (Figure 3Ci). The offspring’s responsecould be interpreted as a misfired predic-tive response—the consequences of a strat-egy that evolved when the parents’ and theoffspring’s conditions matched. The ef-fects of such mismatches and their medicalsignificance have been discussed by Gluck-man and Hanson (2005; Gluckman et al.2007).
Another possibility worth consideringis that of directional changes in heritableepigenetic marks over the course of gener-ations. For instance, if inducing condi-tions persist for several generations, epige-netic marks may accumulate (Jablonka andLamb 2005). This could lead to a moreextreme phenotype (Figure 3Cii) and, pos-sibly, to a greater fidelity of transmission.Inducing conditions might endure due tothe persistence of external environmen-tal factors (e.g., there is multi-generationalexposure to a chemical), continual trans-mission through one sex (i.e., for severalgenerations a particular epiallele is trans-mitted only through females, or onlythrough males), or continuous transmis-sion through old parents, thus leading tothe Lansing effect (see Lamb 1994). Theopposite (Figure 3Ciii) may also occur;that is, when induction in the parental gen-eration is followed by non-inducing condi-tions in the subsequent offspring genera-tions, the induced epigenetic variationsmay linger and gradually fade away, withsome marks being lost in each generation.This might be the basis of the “lingering”modifications described by Jollos (1921),who found that following exposure to higharsenic or salt concentrations, or to highheat, paramecia showed heritable pheno-types that slowly faded over many genera-tions (Jablonka et al. 1992). We suggestthat the study of epigenetic engrams, andthe study of the kinetics of epigeneticmemory changes in different conditions,will lead to an expansion of the research
agenda of epigenetics (Ginsburg and Jab-lonka 2008).
Implications: Evolutionary,Practical, and Theoretical
Given that epigenetic variations are of-ten less stable than genetic variations, whatevolutionary significance do they hold? Weargue that a view of heredity that incorpo-rates the transmission of epigenetic infor-mation through cellular EISs presents chal-lenges and opportunities to applied andtheoretical research in evolutionary biol-ogy. Since, with few exceptions, the in-corporation of epigenetic inheritance andepigenetic control mechanisms into evolu-tionary models and empirical studies is stillrare, our discussion is, inevitably, some-what speculative.
implications for the study ofevolution
Heritable epigenetic variations and epi-genetic control mechanisms are relevantfor the empirical and theoretical study ofevolution because they affect both the pro-cesses of adaptation and of divergence(Jablonka and Lamb 1995, 2005, 2006a,2007b). Five types of effects are character-ized: (i) evolutionary change occurringthrough selection of epigenetic variants,without involvement of genetic variation;(ii) evolutionary change in which an initialepigenetic modification guides the selec-tion of correlated genetic variations; (iii)evolutionary change stemming from thedirect effects of epigenetic variations andepigenetic control mechanisms on the gen-eration of local and systemic epigenomicvariations; (iv) evolutionary change resultingfrom the constraints and affordances thatepigenetic inheritance imposes on develop-ment; and (v) evolutionary change that leadsto new modes of epigenetic inheritance.
Evolution through Selection ofEpigenetic Variants
Adaptation can occur through the selec-tion of heritable epialleles, without any ge-netic change. This may be of particularimportance when populations are small
162 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

and lack genetic variability (e.g., in situa-tions of intense inbreeding following isola-tion or following changes in reproduc-tive strategies). As the examples listed inTable 1 indicate, epigenetic variants areoften induced when environmental condi-tions change, so several individuals in thepopulation may acquire similar modifica-tions at the same time. This means that ad-aptation through the inheritance of newlyinduced epigenetic variants may be veryrapid (Jablonka and Lamb 1995, 2005; Kus-sell and Leibler 2005; Richards 2006; Boss-dorf et al. 2008), thus leading to the accu-mulation of epigenetic variations. Several ofthe epigenetic variations presented in thetable are beneficial for their carriers, such asincreased epigenetically heritable antibioticresistance in bacteria (Adam et al. 2008) andthe switch between morphotypes in Candidaalbicans (Zordan et al. 2006). Other cases,such as increased mutability as a result ofradiation in mice (Dubrova 2003), increasedrecombination rate in plants (Molinier et al.2006), and alterations in flowering time,color, and flower morphology (see table forseveral specific examples and references) arelikely to be adaptive under some conditions;therefore, positive selection of such variantsis plausible.
In order to assess the role of epigeneticvariation in microevolution, it is importantto evaluate the extent and heritability ofepigenetic variations in natural popula-tions (Yi et al. 2004). In a programmaticpaper that outlines the framework for eco-logical epigenetics, Bossdorf et al. (2008)present some of the fundamental researchquestions that need to be asked about epi-genetic variations in natural populations,regarding the extent and structure of epi-genetic variation, its correlation with phe-notypic variation, its inducibility, and itseffects on fitness.
The mechanisms of epigenetic controlmay play an interesting role in structuringepigenetic variation because they can coor-dinate patterns of gene expression. Zuck-erkandl and Cavalli (2007) believe that re-peated sequences in “junk DNA” might becarriers of epigenetic marks, and thatmarks on these sequences can be commu-
nicated to other regions in the genome.They suggested that an altered mark couldtherefore result in coordinated hereditarychanges in the expression of several differ-ent genes simultaneously, hence accelerat-ing adaptive evolution.
Coordinated hereditary epigenetic changesmay have been involved in the process of do-mestication. For instance, forty-six genera-tions of selection for tameness in silver foxesby Belyaev and his research group in No-vosibirsk resulted in a complex of heritablechanges. The foxes became dog-like in theirbehavior and displayed skeletal, hormonal,and spotting changes, as well as altered tailand ear posture, altered vocalizations, andan increased number of supernumerarychromosomes (Belyaev et al. 1981a,b; Trut etal. 2004). Analysis of the pattern of inheri-tance of white spotting revealed that spottingbehaved like a dominant or semi-dominanttrait, but the rate of appearance and disap-pearance of the character was far too highfor new mutations to be a likely explanation.These reversible changes could not be ex-plained as an effect of inbreeding either,because the coefficient of inbreeding wasonly 0.03 (Trut et al. 2004). A probable ex-planation is that the stress of domesticationand selection for tameness targeted geneswith large effects in the neuro-hormonal sys-tem (Trut et al. 2004; Popova 2006) and mayhave heritably reactivated some of them(Belyaev 1981a,b). This epigenetic interpre-tation, in terms of new epimutations ratherthan new mutations, explains the high rateof appearance and disappearance of somephenotypes, and support for this comesfrom the fact that at least two of the genes(Agouti and C-kit) that seem to be involved inthe changes are known to have heritable epi-genetic variants in mice (Trut et al. 2004).The induction and selection of epigeneticvariations may also have been important inthe domestication of plants: ecological andgenomic stress conditions caused by mov-ing plants to new conditions and crossingdivergent strains induce many epigeneticvariations, and selection of such variationsprobably played a part in domestic plantevolution.
June 2009 163TRANSGENERATIONAL EPIGENETIC INHERITANCE

Epigenetic Change Guiding the Selectionof Genetic Variations
The guiding role of development in evolu-tion has been a subject of discussion eversince the pioneering work of Wadding-ton (1957, 1968, 1975) and Schmalhausen(1949). Their basic idea was that selectioncan lead to a change from a stimulus-dependent to a stimulus-independent (orless dependent) phenotypic response. Theprocess leading to the change from a phe-notype whose expression was dependenton an environmental inducer to a consti-tutive expression was called genetic assim-ilation (Waddington 1957; for recent dis-cussion of the idea and its evaluation, seePigliucci et al. 2006; for suggestions em-phasizing its role in the evolution of behav-ior, see Avital and Jablonka 2000; Gottlieb2002).
Ideas about the significance of develop-mental plasticity have recently been strength-ened and extended to provide a generalframework for evolutionary biology (Pigli-ucci 2001; Schlichting and Pigliucci 1998).West-Eberhard (2003) suggested that environ-mentally-induced changes during develop-ment guide the selection of genetic changesthat simulate, stabilize, and ameliorate any det-rimental effects of induced developmentalchanges. She called this developmental guid-ing process, which includes but is not limitedto genetic assimilation, “genetic accommoda-tion.” Induced epiallelic variations that are epi-genetically inherited may enhance the effec-tiveness of assimilation and accommodationprocesses—something that is likely to be par-ticularly important during conditions of stress(Jablonka et al. 1992; Jablonka and Lamb1995, 2005; Pal 1998; Sangster et al. 2004;Badyaev 2005; Siegal and Bergman 2006). Anexample showing the facilitating evolutionaryeffects of epigenetic inheritance was providedby True and Lindquist’s (2000) study, in whichthey compared pairs of yeast strains differingonly in whether or not they carried [PSI�], theprion form of a protein that is involved inmRNA translation. By growing the pairs ofstrains in a variety of conditions, they uncov-ered strain-specific differences between themin colony morphology and growth characteris-
tics. Since the presence of the [PSI�] prionleads to the suppression of nonsense muta-tions, the production of a variety of new pro-tein products in the [PSI�] containing strains(that arose because translation goes beyondthe normal endpoint of functional genes, orbecause stop-codons in the middle of non-functional genes are ignored) was increasedand was beneficial in some conditions. Theepigenetic, selectable variation that is gener-ated in [PSI�] strains might enable a lineage toadapt and “hold” the adaptation until geneticchanges take over; thus, the heritable epige-netic variations in protein architecture pavethe way for genetic adaptation (True et al.2004; Sangster et al. 2004). As a theoreticalmodel has shown, the adaptive effects of such asystem may lead to its evolution even if theresponse is adaptive only once in a millionyears (Masel and Bergman 2003). Selection ofthe epigenetically-based variation generated bythis type of system would be particularly impor-tant in asexual lineages, where the accumula-tion of mutational changes may be slow.
Epigenetic inheritance-driven accommo-dation has probably been important in chro-mosome evolution as well. It may, forexample, have initiated the evolution of di-morphic X and Y chromosomes. Jablonkaand Lamb (1995) suggested that the initialepigenetic silencing of a sex-determining lo-cus could have produced an epigenetic het-eromorphism between chromosomes, whichled to pairing problems in meiosis and con-sequent heterochromatinization and silenc-ing of the homologous region. This wouldhave reduced recombination frequenciesand driven degeneration of the Y chro-mosome; it could also have led to X-chromosome imprinting and dosage com-pensation in mammals (Jablonka 2004a).
Heritable epigenetic variations may alsoplay an important role in the evolution ofchromosomal structures such as centro-meres. Henikoff et al. (2001) have proposedthat the rapid evolution of centromeric se-quences and some centromere-associatedproteins may be driven by an epigenetically-guided arms race (Talbert et al. 2004; Heni-koff and Smith 2007). They suggested thatcentromeres compete to enter the productof female meiosis that will form a gamete
164 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

(rather than a polar body, which is a deadend). Centromeres with DNA sequences thatresult in more efficient spindle fiber attach-ments out-compete others in the race to theprospective egg, thus leading to centromere-associated meiotic drive. However, meioticdrive often has deleterious effects on the or-ganism’s fitness, because deleterious genesthat are linked to the chromosome with thedriving locus—the “strong” centromere, inthis case—reduce individual fitness. The illeffects of centromere-driven meiotic driveare neutralized by the selection of alleles ofcentromere protein genes with a lower bind-ing affinity for the centromeric sequences;hence, centromere-binding proteins evolverapidly and adaptively to counter the “self-ish” centromere sequences. This evolution isdriven by the centromere DNA sequencesand their attached proteins. It is the func-tional epigenotype that must remain stable,whatever the specific identity of the DNAsequence and the proteins at the centro-meric regions.
An important consequence of the genera-tion and evolution of different epigenotypesacross various populations is that, like ge-netic variations, they may initiate reproduc-tive isolation. Differences in chromatin struc-ture that arise by chance or during localadaptation may result in hybrid offspringthat either fail to develop normally orare sterile because the two sets of parentalchromosomes carry incompatible chromatinmarks (Jablonka and Lamb 1995). For exam-ple, incompatibility between parental marksis thought to be the reason why hybrids be-tween two species in the rodent genus Pero-myscus develop abnormally (Vrana et al.2000), and, in plants, crosses between par-ents of different ploidies fail because of thedysfunction of the hybrid endosperm, a tis-sue exhibiting genomic imprinting (Sokolov2006).
The Effects of Epigenetic Variations andEpigenetic Control Mechanisms on the
Generation of Local and SystemicEpigenomic Variation
Although in practice the biases imposedby epigenetic variations, such as methyl-ation marks, on the generation of local
changes in DNA are intertwined with epi-genetic control mechanisms generatingsystemic genomic changes, we need to dis-cuss them separately, for heuristic reasons.It has been known for some time that therates of mutation, transposition, and re-combination are lower in condensed thanin open chromatin (Belyaev and Borodin1982; Jablonka and Lamb 1995), and thatthe movement of transposable elements,which is recognized as a major cause ofgenomic change (Kidwell and Lisch 1997),is markedly influenced by various types ofinternal (genetic) and external (environ-mental) stresses. It is therefore clear thatepigenetic variations bias genetic changes.However, the effect of epigenetic controlmechanisms can go beyond the more orless localized mutational changes inducedby local chromatin variations. Zufall et al.(2005) have suggested that developmen-tally regulated genome rearrangementsbrought about by epigenetic control mech-anisms are an ancient feature of eu-karyotes. If so, it is possible that, duringperiods of stress, the same epigenetic con-trol mechanisms cause global epigenomicmacro-variations that are inherited be-tween generations and that lead to macro-evolutionary changes (Jablonka and Lamb2008; Lamm and Jablonka 2008). Theseepigenetic control mechanisms may un-derlie the systemic changes in the genomethat Goldschmidt (1940) believed drovemacroevolution. Goldschmidt proposedthat macroevolutionary changes are the re-sult of large changes in the genome thatare based either on macromutations (mu-tations in single genes that have very largephenotypic effects) or on systemic muta-tions (changes in the organization of thegenome, such as chromosomal rearrange-ments). Goldschmidt’s ideas used to bederided, but recent data from many bio-logical fronts are changing this attitude(Shapiro 1999; Bateman and DiMichele2002; Fontdevila 2005). Sequence studieshave shown that during plant and animalphylogeny, many developmental geneshave been duplicated and re-used (Gu etal. 2004), and Rodin et al. (2005) havesuggested how epigenetic silencing may
June 2009 165TRANSGENERATIONAL EPIGENETIC INHERITANCE

play a role in this. Epigenetic controlmechanisms probably have a key role inspeciation through polyploidization andhybridization, which are of central impor-tance in plant evolution (Jorgensen 2004;Rapp and Wendel 2005). As we noted ear-lier, recent studies have shown that inmany naturally occurring and experimen-tally constructed polyploids and hybrids,DNA methylation patterns are dramaticallyaltered, and genes in some of the dupli-cated chromosomes are heritably silenced.Following auto- and allo-polyploidization,there is a burst of selectable variation, withall the opportunities for adaptation thatthis provides. This evidence is very much inline with the suggestions of McClintock(1984), who argued that stress leads to areshaping of the genome.
Although we do not yet know how epi-genetic control systems are involved in thegeneration of such systemic mutations,processes based on pairing, such as themechanisms seen in ciliates (in which pair-ing with scanRNAs determines which se-quences are degraded) and during meioticmis-pairing (in which unpaired regions aredeleted or heterochromatinized) may berecruited under conditions of genomicand ecological stress. Molinier et al. (2006,and personal communication) showed thatexposing Arabidopsis to UV-C radiation inone generation caused a heritable increasein the recombination rate of the wholepopulation of irradiated plants for at leastfour generations. This might be an exam-ple of an induced, pairing-based systemic,epigenetic change. Jablonka and Lamb(1995, 2008) suggested that there may havebeen selection for specific heritable epige-netic responses based on pairing, whichare determined by the type of stress (e.g.,direct radiation-induced damage to DNA,nutritional stress, heat stress), its severity,and its probability of reoccurrence.
Evolutionary Constraints and AffordancesImposed by Epigenetic Inheritance
Cellular EISs were a precondition for theevolution of complex multicellular organ-isms with specialized cell lineages, becausecells in such lineages have to maintain and
transmit their determined states, even whenthe conditions that initiated them are longpast. Since the cells that give rise to the nextgeneration of organisms need to have anuncommitted state, and efficient EISs couldjeopardize this, EISs must have imposed astrong constraint on the evolution of ontog-eny. There are several features of develop-ment that may be outcomes of selection toprevent cells with inappropriate epigeneticlegacies from founding the next generation.For example, the difficulty of reversing someepigenetic states, the early segregation andquiescent state of the germline of many ani-mal groups, and the massive changes inchromatin structure that occur during mei-osis and gamete production, may all be theresult of selection against transmitting theepigenetic “memories” associated with thedevelopmental changes and chance epimu-tations that would prevent a zygote fromstarting its development in a totipotentepigenetic state (Jablonka and Lamb1995, 2005). Recently, Pepper et al. (2007)have suggested that serial differentiation—the sequence of differentiation that startswith self-renewing somatic stem cells andproceeds through several non-self-renewing,transient, amplifying cell stages before end-ing with terminally differentiated cells—isalso a strategy that evolved to avoid thesomatic selection of selfish genetic and epi-genetic variations.
Jablonka and Lamb (2006b) argued thatthe constraints and affordances of epige-netic control systems and epigenetic inher-itance played a crucial role in all eight ofthe major evolutionary transitions identi-fied by Maynard Smith and Szathmary(1995). For example, the transition fromindependent genes to long chromosomeswas probably dependent on epigenetic in-heritance based upon chromatin marking,which maintains patterns of gene activityfollowing DNA replication. Epigenetic con-trol mechanisms may also have been im-portant in the transition from prokaryotesto eukaryotes—a transition that was associ-ated with processes of endosymbiogenesis.It is likely that massive and heritable inac-tivation of large parts of the symbiont-to-begenome, as well as the employment of
166 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

mechanisms of structural inheritance thatenabled the integrity of the basic structureof the symbiont-to-be membranes to per-sist, were involved in this transition.
The Evolution of EISs: Their Origin andSelection
In light of the growing volume of workand the theoretical considerations thatsuggest that nongenetic mechanisms of in-formation transfer play key roles in evolu-tion, the evolutionary origin of nongeneticinheritance systems is of fundamental in-terest. There have been some theoreticaland comparative studies that have ad-dressed the evolution of EISs, but not allaspects have been explored. The evolution-ary origins of DNA methylation have beenconsidered, and several different hypothe-ses for the advantages it conferred havebeen suggested (Bestor 1990; Bird 1995;Regev et al. 1998; Colot and Rossignol1999; Mandrioli 2004). There are also sev-eral comparative studies of histone evolu-tion (Sandman et al. 1998; Felsenfeld andGroudine 2003), and the evolution ofRNAi systems for defence against genomicparasites and as regulators in an ancientRNA world has been suggested (Ceruttiand Casas-Mollano 2006).
Specific developmental processes thatinvolve epigenetic inheritance, such asgenomic imprinting and X-chromosomeinactivation, have also been subjects of evo-lutionary study (e.g., Jablonka and Lamb1995; Lyon 1998; Haig 2002; Jablonka2004a; Wolf and Hager 2006). However,other developmental processes that de-pend on EISs (paramutation and stress-induced epigenomic alterations, for exam-ple) have not yet received much attentionfrom evolutionary biologists, and the adap-tive significance—if any—of epigeneticmechanisms leading to systemic changesduring periods of genomic and ecologicalstresses is at present an open question.
The stability of epigenetic transmissionis likely to be an evolved trait that dependson the relative cost of error and the costof development (Rando and Verstrepen2007). Epigenetic recall may be selectivelysuperior to full epigenetic inheritance in
environments that change every few gener-ations because the cost of response-errorthat occurs when memory is perfect is re-duced, as is the cost of development-from-scratch, which occurs when reset is com-plete and full induction is required. Thetransmission of epigenetic engrams thatlead to an inducer-requiring yet facilitatedresponse may therefore often be an opti-mal compromise between the danger of atyrannically good memory, on the onehand, and the expensive response-delaythat comes with ”forgetting” too thor-oughly, on the other. Direct evidence forepigenetic recall is needed, however, andtheoretical exploration through modelingmight point to biological systems with strat-egies that would qualify them as good tar-gets for empirical research.
Epigenetic inheritance should be fa-vored in fluctuating environmental condi-tions that last for more than one genera-tion (but not for very long) and may beparticularly important in the type of envi-ronments experienced by many microor-ganisms (Lachmann and Jablonka 1996;Balaban et al. 2004; Lewis 2007; Rando andVerstrepen 2007). In such fluctuating en-vironments, efficient epigenetic inheri-tance is likely to evolve (i) if the parentalenvironment carries reliable informationabout the offspring’s environment (Jab-lonka et al. 1995), (ii) when the responseto induction is lengthy and incurs a veryhigh cost (Lachmann and Jablonka 1996),and (iii) when recall is not an option orincurs too high a cost.
theoretical and practicalimplications
Incorporating epigenetic inheritance intoevolutionary theory extends the scope of evo-lutionary thinking and leads to notions ofheredity and evolution that incorporate de-velopment. Dobzhansky’s definition of evo-lution as “a change in the genetic composi-tion of populations” (1937, p.11) appears tobe too narrow because it does not incorpo-rate all sources of heritable variations. Bothevolution and heredity need to be redefined.Jablonka and Lamb (2007a,b,c) suggestedthat evolution should be redefined as the
June 2009 167TRANSGENERATIONAL EPIGENETIC INHERITANCE

set of processes that lead to changes in the natureand frequency of heritable types in a population,and heredity as the developmental reconstruc-tion processes that link ancestors and descen-dants and lead to similarity between them.These deliberately broad redefinitions al-low evolutionary possibilities denied by the“Modern Synthesis” version of evolutionarytheory, which states that variations areblind, are genetic (nucleic acid-based), andthat saltational events do not significantly con-tribute to evolutionary change (Mayr 1982).The epigenetic perspective challenges all theseassumptions, and it seems that a new extendedtheory, informed by developmental studiesand epigenetic inheritance, and incorporatingDarwinian, Lamarckian, and saltational frame-works, is going to replace the Modern Synthe-sis version of evolution (Jablonka and Lamb2005, 2007c). We believe, therefore, that theimpact of epigenetics and epigenetic inheri-tance on evolutionary theory and the philoso-phy of biology will be profound.
As we noted earlier, it is now recognizedthat epigenetic inheritance is relevant forecology, and new methods and approachesto the research questions to which it pointsshould be developed (Bossdorf et al.2008). The relevance of epigenetic varia-tions to biodiversity in our rapidly chang-ing world is also of obvious interest andclearly has to be explored.
A discussion of the implications of epige-netic studies for medicine is beyond thescope of this review, but since epigenetic de-fects can be transmitted between genera-tions of cells and individuals, we direct thereader’s attention to some recent reviews.Baylin and Jones (2007) review the epigenet-ics of cancer, and Zoghbi and Beaudet(2007) review diseases caused by defects inchromatin marking and imprinting. The epi-genetic aspects of metabolic diseases andtheir transgenerational effects are also beingintensely studied (see Bateson et al. 2004;Gluckman and Hanson 2005; Gluckman et
al. 2007; Petronis 2004, 2006). The epidemi-ological aspects of epigenetic inheritancewere reviewed by Jablonka (2004b), and theimportance of epigenetics for aging researchhas been discussed by Vanyushin (1973),Holliday (1984), Lamb (1994), and Issa(2000). The recently reported ability ofpathogenic microorganisms to evolve herita-ble epigenetic resistance to medication (e.g.,antibiotics) may be of major medical impor-tance (Adam et al 2008), and the relevanceof epigenetic inheritance for therapeuticcloning and nuclear transplantation in ani-mals, including humans, is self-evident (seeJaenisch and Gurdon 2007).
Heredity is a fundamental property of liv-ing organisms. It is therefore not surprisingthat, in the beginning of the last century, therediscovery of Mendel’s laws and the chro-mosomal mechanisms underlying them ledto profound changes in all branches of biol-ogy. Today, at the dawn of the 21st century,another aspect of heredity—epigenetic in-heritance and the epigenetic control mech-anisms underlying it—is being unravelled.Like the early 20th-century discoveries, it,too, is driving a great expansion and trans-formation in our understanding of biology.
acknowledgments
We would like to thank the many colleagues whogenerously provided us with greatly needed informa-tion and feedback: Nathalie Q. Balaban, Daniel Beny-shek, Renee Borges, Graham C. Burdge, GiacomoCavalli, Vincent Colot, Gyorgy Csaba, Scott Gilbert,Janine Guespin-Michel, Rudolf Hagemann, LuisaHirschbein, David Martin, Marjorie Matzke, FredMeins, Jean Molinier, John Parker, Minoo Rassoulza-degan, Wolf Reik, Yih-Horng Shiao, Michael Skinner,Nadine Vastenhouw, Robert A. Waterland, andGuangtian Zou. We also thank Maya Raz for her con-tribution to the graphics. We are especially grateful toMarion Lamb for reading (and re-reading) the wholemanuscript, and for her invaluable critical commentsand constructive suggestions. Finally, we would like tothank Massimo Pigliucci and two anonymous review-ers for their useful and critical comments.
REFERENCES
Adam M., Murali B., Glenn N. O., Potter S. S. 2008.Epigenetic inheritance based evolution of antibi-
otic resistance in bacteria. BMC Evolutionary Biol-ogy 8:52.
168 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

Agrawal A. A., Laforsch C., Tollrian R. 1999. Trans-generational induction of defences in animals andplants. Nature 401(6748):60–63.
Alleman M., Doctor J. 2000. Genomic imprinting inplants: observations and evolutionary implica-tions. Plant Molecular Biology 43(2–3):147–161.
Allen J. A., editor. 2004. Issue on plant polyploidy.Biological Journal of the Linnean Society 82(4):409–700.
Allen N. D., Norris M. L., Surani M. A. 1990. Epige-netic control of transgene expression and imprint-ing by genotype-specific modifiers. Cell 61(5):853–861.
Allis C. D., Jenuwein T., Reinberg D., Caparros M.-L.,editors. 2007. Epigenetics. Cold Spring Harbor(NY): Cold Spring Harbor Laboratory Press.
Allshire R. C., Selker E. U. 2007. Fungal models forepigenetic research: Schizosaccharomyces pombe andNeurospora crassa. Pages 101–125 in Epigenetics, ed-ited by C. D. Allis et al. Cold Spring Harbor (NY):Cold Spring Harbor Laboratory Press.
Anway M. D., Cupp A. S., Uzumcu M., Skinner M. K.2005. Epigenetic transgenerational actions of en-docrine disruptors and male fertility. Science308(5727):1466–1469.
Anway M. D., Leathers C., Skinner M. K. 2006a. En-docrine disruptor vinclozolin induced epigenetictransgenerational adult-onset disease. Endocrinol-ogy 147(12):5515–5523.
Anway M. D., Memon M. A., Uzumcu M., SkinnerM. K. 2006b. Transgenerational effect of the en-docrine disruptor vinclozolin on male spermato-genesis. Journal of Andrology 27(6):868–879.
Avital E., Jablonka E. 2000. Animal Traditions: Behav-ioural Inheritance in Evolution. Cambridge (UK):Cambridge University Press.
Badyaev A. V. 2005. Stress-induced variation in evolu-tion: from behavioural plasticity to genetic assim-ilation. Proceedings of the Royal Society of London,Series B: Biological Sciences 272(1566):877–886.
Balaban N. Q., Merrin Q. J., Chait R., Kowalik L.,Leibler S. 2004. Bacterial persistence as a pheno-typic switch. Science 305(5690):1622–1625.
Bantignies F., Cavalli G. 2006. Cellular memory anddynamic regulation of polycomb group proteins.Current Opinion in Cell Biology 18(3):275–283.
Barlow D. P., Bartolomei M. S. 2007. Genomic im-printing in mammals. Pages 357–373 in Epigenetics,edited by C. D. Allis et al. Cold Spring Harbor(NY): Cold Spring Harbor Laboratory Press.
Bateman R. M., DiMichele W. A. 2002. Generatingand filtering major phenotypic novelties: neo-Goldschmidtian saltation revisited. Pages 109–159in Developmental Genetics and Plant Evolution, editedby Q. C. B. Cronk et al. London (UK): Taylor &Francis.
Bateson P., Barker D., Clutton-Brock T., Deb D.,
D’Udine B., Foley R. A., Gluckman P., et al. 2004.Developmental plasticity and human health. Na-ture 430(6998):419–421.
Baulcombe D. C. 2007. Amplified silencing. Science315(5809):199–200.
Baxa U., Cassese T., Kajava A. V., Steven A. C. 2006.Structure, function, and amyloididogenesis of fun-gal prions: filament polymorphisms and prionsvariants. Advances in Protein Chemistry 73:125–180
Baylin S. B., Jones P. A. 2007. Epigenetic determi-nants of cancer. Pages 457–476 in Epigenetics, ed-ited by C. D. Allis et al. Cold Spring Harbor (NY):Cold Spring Harbor Laboratory Press.
Bean C. J., Schaner C. E., Kelly W. G. 2004. Meioticpairing and imprinted X chromatin assembly inCaenorhabditis elegans. Nature Genetics 36(1):100–105.
Beisson J., Sonneborn T. M. 1965. Cytoplasmic inher-itance of the organization of the cell cortex inParamecium aurelia. Proceedings of the National Acad-emy of Sciences USA 53(2):275–282.
Belyaev D. K., Borodin P. M. 1982. The influence ofstress on variation and its role in evolution. Biolo-gisches Zentralblatt 100:705–714.
Belyaev D. K., Ruvinsky A. O., Borodin P. M. 1981a.Inheritance of alternative states of the fused genein mice. Journal of Heredity 72:107–112.
Belyaev D. K., Ruvinsky A. O., Trut L. N. 1981b. In-herited activation-inactivation of the star gene infoxes: its bearing on the problem of domestica-tion. Journal of Heredity 72(4):267–274.
Belyaev D. K., Ruvinsky A. O., Agulnik A. I., AgulnikS. I. 1983. Effect of hydrocortisone on the fre-quency and inheritance of the fused gene in mice.Genetika 64(4):275–281.
Benkemoun L., Saupe S. J. 2006. Prion proteins asgenetic material in fungi. Fungal Genetics and Biol-ogy 43(12):789–803.
Bennett S. T., Wilson A. J., Esposito L., Bouzekri N.,Undlien D. E., Cucca F., Nistico L., et al. 1997.Insulin VNTR allele-specific effect in type 1 diabe-tes depends on identity of untransmitted paternalallele. Nature Genetics 17(3):350–352.
Berger S. L. 2007. The complex language of chromatinregulation during transcription. Nature 447(7143):407–412.
Bestor T. H. 1990. DNA methylation: evolution ofa bacterial immune function into a regulatorof gene expression and genome structure inhigher eukaryotes. Philosophical Transactions of theRoyal Society of London, Series B: Biological Sciences326(1235):179–187.
Bird A. P. 1995. Gene number, noise reduction andbiological complexity. Trends in Genetics 11(3):94–100.
Blasco M. A. 2007. The epigenetic regulation of mam-
June 2009 169TRANSGENERATIONAL EPIGENETIC INHERITANCE

malian telomeres. Nature Reviews Genetics 8(4):299–309.
Blewitt M. E., Vickaryous N. K., Paldi A., Koseki H.,Whitelaw E. 2006. Dynamic reprogramming ofDNA methylation at an epigenetically sensitive al-lele in mice. PLoS Genetics 2(4):399–405.
Bossdorf O., Richards C. L., Pigliucci M. 2008. Epige-netics for ecologists. Ecology Letters 11(2):106–115.
Brink R. A. 1973. Paramutation. Annual Review of Ge-netics 7:129–152.
Buiting K., Gro� S., Lich C., Gillessen-Kaesbach G.,El-Maarri O., Horsthemke B. 2003. Epimutationsin Prader-Willi and Angelman syndromes: a mo-lecular study of 136 patients with an imprintingdefect. American Journal of Human Genetics 72(3):571–577.
Burdge G. C., Slater-Jefferies J., Torrens C., PhillipsE. S., Hanson M. A., Lillycrop K. A. 2007. Dietaryprotein restriction of pregnant rats in the F0 gen-eration induces altered methylation of hepaticgene promoters in the adult male offspring in theF1 and F2 generations. British Journal of Nutrition97(3):435–439.
Buss L. W. 1987. The Evolution of Individuality. Prince-ton (NJ): Princeton University Press.
Campbell P., editor. 2007. Follow the curve: the forcesthat shape cell membranes. Nature 447(7143 In-sight):353–500.
Casadesus J., Low D. 2006. Epigenetic gene regulationin the bacterial world. Microbiology and MolecularBiology Reviews 70(3):830–856.
Cavalier-Smith T. 2004. The membranome and mem-brane heredity in development and evolution.Pages 335–351 in Organelles, Genomes and EukaryotePhylogeny, edited by R. P. Hirt and D. S. Horner.Boca Raton (FL): CRC Press.
Cerutti H., Casas-Mollano J. A. 2006. On the originand functions of RNA-mediated silencing: fromprotists to man. Current Genetics 50(2):81–99.
Chandler V. L. 2007. Paramutation: from maize tomice. Cell 128(4):641–645.
Chandler V. L., Eggleston W. B., Dorweiler J. E. 2000.Paramutation in maize. Plant Molecular Biology43(2–3):121–145.
Chang H.-S., Anway M. D., Rekow S. S., Skinner M. K.2006. Transgenerational epigenetic imprinting ofthe male germline by endocrine disruptor expo-sure during gonadal sex determination. Endocri-nology 147(12):5524–5541.
Chien P., Weissman J. S., DePace A. H. 2004. Emerg-ing principles of conformation-based prion inher-itance. Annual Review of Biochemistry 73:617–656.
Chong S., Vickaryous N., Ashe A., Zamudio N.,Youngson N., Hemley S., Stopka T., et al. 2007.Modifiers of epigenetic reprogramming show pa-ternal effects in the mouse. Nature Genetics 39(5):614–622.
Cikos S., Vesela J., Il’kova G., Rehak P., Czikkova S.,Koppel J. 2005. Expression of beta adrenergic re-ceptors in mouse oocytes and preimplantationembryos. Molecular Reproduction and Development71(2):145–153.
Colot V., Rossignol J. L. 1999. Eukaryotic DNA meth-ylation as an evolutionary device. BioEssays 21(5):402–411.
Cormier S., Vandormael-Pournin S., Babinet C.,Cohen-Tannoudji M. 2004. Developmental ex-pression of the Notch signaling pathway genesduring mouse preimplantation development. GeneExpression Patterns 4(6):713–717.
Costa Y., Speed R. M., Gautier P., Semple C. A., Ma-ratou K., Turner J. M. A., Cooke H. J. 2006. MouseMAELSTROM: the link between meiotic silencingof unsynapsed chromatin and microRNA path-way? Human Molecular Genetics 15(15):2324–2334.
Crews D., Gore A. C., Hsu T. S., Dangleben N. L.,Spinetta M., Schallert T., Anway M. D., SkinnerM. K. 2007. Transgenerational epigenetic imprintson mate preference. Proceedings of the NationalAcademy of Sciences USA 104(14):5942–5946.
Cropley J. E., Suter C. M., Beckman K. B., MartinD. I. K. 2006. Germ-line epigenetic modificationof the murine Avy allele by nutritional supplemen-tation. Proceedings of the National Academy of SciencesUSA 103(46):17308–17312.
Csaba G., Karabelyos C., Inczefi-Gonda A., PallingerE. 2005. Three-generation investigation on seroto-nin content in rat immune cells long after�-endorphin exposure in late pregnancy. Hormoneand Metabolic Research 37:172–177.
Cullen B. R. 2004. Transcription and processingof human microRNA precursors. Molecular Cell16(6):861–865.
Cullis C. A. 2005. Mechanisms and control of rapidgenomic changes in flax. Annals of Botany 95(1):201–206.
Delage Y., Goldsmith M. 1912. The Theories of Evolution.Trans. Andre Tridon. New York: B. W. Huebsch.
Dobzhansky T. 1937. Genetics and the Origin of Species.New York: Columbia University Press.
Dubnau D., Losick R. 2006. Bistability in bacteria.Molecular Microbiology 61(3):564–572.
Dubrova Y. E. 2003. Radiation-induced transgenera-tional instability. Oncogene 22(45):7087–7093.
Ekwall K. 2007. ‘Arc’ escorts siRNAs in heterochro-matin assembly. Nature Structural and Molecular Bi-ology 14(3):178–179.
Felsenfeld G., Groudine M. 2003. Controlling thedouble helix. Nature 421(6921):448–453.
Flanagan J. M., Popendikyte V., Pozdniakovaite N.,Sobolev M., Assadzadeh A., Schumacher A., Zan-geneh M., et al. 2006. Intra- and interindividualepigenetic variation in human germ cells. Ameri-can Journal of Human Genetics 79(1):67–84.
170 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

Flintoft L., editor. 2007. Focus on: epigenetics. NatureReviews Genetics 8(4):245–314.
Fontdevila A. 2005. Hybrid genome evolution bytransposition. Cytogenetic and Genome Research110(1):49–55.
Fothergill P. G. 1952. Historical Aspects of Organic Evo-lution. London (UK): Hollis and Carter.
Frankel J. 1989. Pattern Formation Ciliate Studies andModels. New York: Oxford University Press.
Genereux D. P., Miner B. E., Bergstrom C. T., LairdC. D. 2005. A population-epigenetic model to in-fer site-specific methylation rates from double-stranded DNA methylation patterns. Proceedings ofthe National Academy of Sciences USA 102(16):5802–5807.
Ginsburg S., Jablonka E. 2008. Epigenetic learning innon-neural organisms. Journal of Biosciences 33(4).
Gluckman P., Hanson M. 2005. The Fetal Matrix: Evo-lution, Development and Disease. Cambridge (UK):Cambridge University Press.
Gluckman P. D., Hanson M. A., Beedle A. S. 2007.Non-genomic transgenerational inheritance ofdisease risk. BioEssays 29(2):145–154.
Goldschmidt R. B. 1940. The Material Basis of Evolution.New Haven (CT): Yale University Press.
Gottlieb G. 2002. Developmental-behavioral initiationof evolutionary change. Psychological Review 109(2):211–218.
Grandjean V., Hauck Y., Beloin C., Le Hegarat F.,Hirschbein L. 1998. Chromosomal inactivation ofBacillus subtilis exfusants: a prokaryotic model ofepigenetic regulation. Biological Chemistry 379(4–5):553–557.
Grant-Downton R. T., Dickinson H. G. 2005. Epige-netics and its implications for plant biology. 1.The epigenetic network in plants. Annals of Botany96(7):1143–1164.
Grant-Downton R. T., Dickinson H. G. 2006. Epige-netics and its implications for plant biology. 2.The ‘epigenetic epiphany’: epigenetics, evolutionand beyond. Annals of Botany 97(1):11–27.
Grewal S. I. S., Jia S. T. 2007. Heterochromatin revis-ited. Nature Reviews Genetics 8(1):35–46.
Grimes G., Aufderheide K. 1991. Cellular aspects ofpattern formation: the problem of assembly.Monographs in Developmental Biology 22:1–94.
Groth A., Rocha W., Verreault A., Almouzni G. 2007.Chromatin challenges during DNA replicationand repair. Cell 128(4):721–733.
Gu Z., Rifkin S. A., White K. P., and Li W.-H. 2004.Duplicate genes increase gene expression diversitywithin and between species. Nature Genetics 36(6):577–579.
Hadchouel M., Farza H., Simon D., Tiollais P., Pour-cel C. 1987. Maternal inhibition of hepatitis Bsurface antigen gene expression in transgenic
mice correlates with de novo methylation. Nature329(6138):454–456.
Haig D. 2002. Genomic Imprinting and Kinship. NewBrunswick (NJ): Rutgers University Press.
Haig D. 2004. The (dual) origin of epigenetics. ColdSpring Harbor Symposia on Quantitative Biology 69:67–70.
Hajkova P., Erhardt S., Lane N., Haaf T., El-Maarri O.,Reik W., Walter J., Surani M. A. 2002. Epigeneticreprogramming in mouse primordial germ cells.Mechanisms of Development 117(1–2):15–23.
Henderson I. R., Jacobsen S. E. 2007. Epigenetic in-heritance in plants. Nature 447(7143):418–424.
Henikoff S., Ahmad K., Malik H. S. 2001. The centro-mere paradox: stable inheritance with rapidlyevolving DNA. Science 293(5532):1098–1102.
Henikoff S., Smith M. M. 2007. Histone variants andepigenetics. Pages 249–264 in Epigenetics, editedby C. D. Allis et al. Cold Spring Harbor (NY): ColdSpring Harbor Laboratory Press.
Himber C., Dunoyer P., Moissiard G., RitzenthalerC., Voinnet O. 2003. Transitivity-dependent and-independent cell-to-cell movement of RNA si-lencing. EMBO Journal 22(17):4523– 4533.
Hitchins M. P., Wong J. J. L., Suthers G., Suter C. M.,Martin D. I. K., Hawkins N. J., Ward R. L. 2007.Brief report: inheritance of a cancer-associatedMLH1 germ-line epimutation. New England Journalof Medicine 356(7):697–705.
Holliday R. 1984. The significance of DNA methyl-ation in cellular aging. Pages 269–283 in MolecularBiology of Aging, edited by A. D. Woodhead et al.New York: Plenum Press.
Holliday R. 1994. Epigenetics—an overview. Develop-mental Genetics 15:453–457.
Holliday R. 2002. Epigenetics come of age in thetwenty-first century. Journal of Genetics 81(1):1–4.
Holliday R. 2006. Epigenetics: a historical overview.Epigenetics 1(2):76–80.
Huettel B., Kanno T., Daxinger L., Bucher E., van derWinden J., Matzke A. J., Matzke M. 2007. RNA-directed DNA methylation mediated by DRD1 andPol IVb: a versatile pathway for transcriptional genesilencing in plants. Biochimica et Biophysica Acta-GeneStructure and Expression 1769(5–6):358–374.
Issa J. P. 2000. CpG-island methylation in aging andcancer. Current Topics in Microbiology and Immunol-ogy 249:101–118.
Jablonka E. 2004a. The evolution of the peculiaritiesof mammalian sex chromosomes: an epigeneticview. BioEssays 26(12):1327–1332.
Jablonka E. 2004b. Epigenetic epidemiology. Interna-tional Journal of Epidemiology 33(5):929–935.
Jablonka E., Lachmann M., Lamb M. J. 1992. Evi-dence, mechanisms and models for the inheri-tance of acquired characters. Journal of TheoreticalBiology 158(2):245–268.
June 2009 171TRANSGENERATIONAL EPIGENETIC INHERITANCE

Jablonka E., Lamb M. J. 1989. The inheritance ofacquired epigenetic variations. Journal of Theoreti-cal Biology 139(1):69–83.
Jablonka E., Lamb M. J. 1995. Epigenetic Inheritance andEvolution: The Lamarckian Dimension. Oxford (UK):Oxford University Press.
Jablonka E., Lamb M. J. 2002. The changing conceptof epigenetics. Annals of the New York Academy ofSciences 981:82–96.
Jablonka E., Lamb M. J. 2005. Evolution in Four Dimen-sions: Genetic, Epigenetic, Behavioral, and SymbolicVariation in the History of Life. Cambridge (MA):MIT Press.
Jablonka E., Lamb M. J. 2006a. Evolutionary epigenet-ics. Pages 252–264 in Evolutionary Genetics: Conceptsand Case Studies, edited by C. W. Fox and J. B. Wolf.New York: Oxford University Press.
Jablonka E., Lamb M. J. 2006b. The evolution ofinformation in the major transitions. Journal ofTheoretical Biology 239(2):236–246.
Jablonka E., Lamb M. J. 2007a. Precis of Evolution inFour Dimensions. Behavioral and Brain Sciences 30:353–365.
Jablonka E., Lamb M. J. 2007b. Bridging the gap: thedevelopmental aspects of evolution. Behavioral andBrain Sciences 30:378–392.
Jablonka E., Lamb M. J. 2007c. The expanded evolu-tionary synthesis—a response to Godfrey-Smith,Haig, and West-Eberhard. Biology and Philosophy22(3):453–472.
Jablonka E., Lamb M. J. 2008. The epigenome inevolution: beyond the modern synthesis. VOGisHerald 12:242–254.
Jablonka E., Oborny B., Molnar I., Kisdi E., HofbauerJ., Czaran T. 1995. The adaptive advantage of phe-notypic memory in changing environments. Philo-sophical Transactions of the Royal Society of London,Series B: Biological Sciences 350(1332):133–141.
Jaenisch R., Gurdon J. 2007. Nuclear transplantationand the reprogramming of the genome. Pages415–434 in Epigenetics, edited by C. D. Allis et al.Cold Spring Harbor (NY): Cold Spring HarborLaboratory Press.
Jirtle R. L., Skinner M. K. 2007. Environmental epi-genomics and disease susceptibility. Nature ReviewsGenetics 8(4):253–262.
Jollos V. Experimentelle Protistenstudien 1. 1921. Un-tersuchungen uber Variabilitat und Vererbungbei Infusorien. Archiv fur Protistenkunde 43:1–222.
Jorgensen R. A. 2004. Restructuring the genome inresponse to adaptive challenge: McClintock’s boldconjecture revisited. Cold Spring Harbor Symposia onQuantitative Biology 69:349–354.
Karpen G. H., Hawley R. S. 2007. Epigenetic regula-tion of chromosome inheritance. Pages 265–289in Epigenetics, edited by C. D. Allis et al. Cold
Spring Harbor (NY): Cold Spring Harbor Labora-tory Press.
Kidwell M. G., Lisch D. 1997. Transposable elementsas sources of variation in animals and plants. Pro-ceedings of the National Academy of Sciences USA94(15):7704–7711.
Kim V. N. 2006. Small RNAs just got bigger: Piwi-interacting RNAs (piRNAs) in mammalian testes.Genes and Development 20(15):1993–1997.
Kolle S., Stojkovic M., Prelle K., Waters M., Wolf E.,Sinowatz F. 2001. Growth hormone (GH)/GH re-ceptor expression and GH-mediated effects dur-ing early bovine embryogenesis. Biology of Repro-duction 64:1826–1834.
Kussell E., Leibler S. 2005. Phenotypic diversity, pop-ulation growth, and information in fluctuating en-vironments. Science 309(5743):2075–2078.
Lachmann M., Jablonka E. 1996. The inheritance ofphenotypes: an adaptation to fluctuating environ-ments. Journal of Theoretical Biology 181:1–9.
Lamb M. J. 1994. Epigenetic inheritance and aging.Reviews in Clinical Gerontology 4(2):97–105.
Lamm E., Jablonka E. 2008. The nurture of nature:hereditary plasticity in evolution. Philosophical Psy-chology 21(3):305–319.
Laurent M., Charvin G., Guespin-Michel J. 2005. Bi-stability and hysteresis in epigenetic regulation ofthe lactose operon. Since Delbruck, a long seriesof ignored models. Cellular and Molecular Biology51(7):583–594.
Levites E. V. 2000. Epigenetic variability as a source ofbiodiversity and a factor of evolution. Pages 73–75in Biodiversity and Dynamics of Ecosystems in NorthEurasia, Volume 1, Part 3. Novosibirsk (Russia):Institute of Cytology and Genetics SB RAS.
Levy A. A., Feldman M. 2004. Genetic and epigeneticreprogramming of the wheat genome upon al-lopolyploidization. Biological Journal of the LinneanSociety 82(4):607–613.
Li E., Bird A. 2007. DNA methylation in mammals.Pages 341–356 in Epigenetics, edited by C. D. Alliset al. Cold Spring Harbor (NY): Cold Spring Har-bor Laboratory Press.
Lindegren C. C. 1949. The Yeast Cell, its Genetics andCytology. Saint Louis (MO): Educational Publishers.
Lindqvist C., Janczak A. M., Natt D., Baranowska I.,Lindqvist N., Wichman A., Lundeberg J., Lind-berg J., Torjesen P. A., Jensen P. 2007. Transmis-sion of stress-induced learning impairment andassociated brain gene expression from parents tooffspring in chickens. PLoS ONE 2(4):e364.
Little J. B. 2003. Genomic instability and bystandereffects: a historical perspective. Oncogene 22(45):6978–6987.
Logan C. A. 2007. Overheated rats, race, and thedouble gland: Paul Kammerer, endocrinology and
172 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

the problem of somatic induction. Journal of theHistory of Biology 40(4):683–725.
Long L., Lin X., Zhai J., Kou H., Yang W., Liu B. 2006.Heritable alteration in DNA methylation patternoccurred specifically at mobile elements in riceplants following hydrostatic pressurization. Bio-chemical and Biophysical Research Communications340(2):369–376.
Lyon M. F. 1998. X-chromosome inactivation: a re-peat hypothesis. Cytogenetics and Cell Genetics 80:133–137.
Malagnac F., Silar P. 2003. Non-Mendelian determi-nants of morphology in fungi. Current Opinion inMicrobiology 6(6):641–645.
Malagnac F., Silar P. 2006. Regulation, cell differen-tiation and protein-based inheritance. Cell Cycle5(22):2584–2587.
Mandrioli M. 2004. Epigenetic tinkering and evolu-tion: is there any continuity in the role of cytosinemethylation from invertebrates to vertebrates? Cel-lular and Molecular Life Sciences 61(19–20):2425–2427.
Marcus E., editor. 2007. Issue on epigenetics. Cell128(4):627–802.
Martin C., Zhang Y. 2007. Mechanisms of epigeneticinheritance. Current Opinion in Cell Biology 19(3):266–272.
Masel J., Bergman A. 2003. The evolution of the evolv-ability properties of the yeast prion [PSI�]. Evolu-tion 57(7):1498–1512.
Matzke M. A., Birchler J. A. 2005. RNAi-mediatedpathways in the nucleus. Nature Reviews Genetics6(1):24–35.
Matzke M. A., Scheid O. M. 2007. Epigenetic regula-tion in plants. Pages 167–189 in Epigenetics, editedby C. D. Allis et al. Cold Spring Harbor (NY): ColdSpring Harbor Laboratory Press.
Maynard Smith J. 1990. Models of a dual inheritancesystem. Journal of Theoretical Biology 143(1):41–53.
Maynard Smith J., Szathmary E. 1995. The Major Tran-sitions in Evolution. Oxford (UK) and New York:W. H. Freeman Spektrum.
Mayr E. 1982. The Growth of Biological Thought: Diver-sity, Evolution, and Inheritance. Cambridge (MA):Belknap Press.
McClintock B. 1984. The significance of responses ofthe genome to challenge. Science 226(4676):792–801.
Meister G., Tuschl T. 2004. Mechanisms of gene silenc-ing by double-stranded RNA. Nature 431(7006):343–349.
Meyer E., Chalker D. L. 2007. Epigenetics of ciliates.Pages 127–150 in Epigenetics, edited by C. D. Alliset al. Cold Spring Harbor (NY): Cold Spring Har-bor Laboratory Press.
Misteli T. 2007. Beyond the sequence: cellular orga-nization of genome function. Cell 128:787–800.
Molinier J., Ries G., Zipfel C., Hohn B. 2006. Trans-generation memory of stress in plants. Nature 442:1046–1049.
Morris C. A., Moazed D. 2007. Centromere assemblyand propagation. Cell 128(4):647–650.
Mortusewicz O., Schermelleh L., Walter J., CardosoM. C., Leonhardt H. 2005. Recruitment of DNAmethyltransferase I to DNA repair sites. Proceedingsof the National Academy of Sciences USA 102(25):8905–8909.
Nanney D. L. 1958. Epigenetic control systems. Pro-ceedings of the National Academy of Sciences USA44(7):712–717.
Nanney D. L. 1968. Cortical patterns in cellular mor-phogenesis: differences in cortical patterns in cil-iates may be hereditary, but independent of genicdifferences. Science 160(3827):496–502.
Nanney D. L. 1985. Heredity without genes: ciliateexplorations of clonal heredity. Trends in Genetics1:295–298.
Naz R. K., Sellamuthu R. 2006. Receptors in sperma-tozoa: are they real? Journal of Andrology 27(5):627–636.
Newbold R. R., Padilla-Banks E., Jefferson W. N. 2006.Adverse effects of the model environmental estro-gen diethylstilbestrol are transmitted to subse-quent generations. Endocrinology 147:S11–S17.
Novick A., Weiner M. 1957. Enzyme induction as anall-or-none phenomenon. Proceedings of the Na-tional Academy of Sciences USA 43(7):553–566.
Nowacki M., Vijayan V., Zhou Y., Schotanus K., DoakT. G., Landweber L. F. 2008. RNA-mediated epige-netic programming of a genome-rearrangementpathway. Nature 451(7175):153–159.
Pak J., Fire A. 2007. Distinct populations of primaryand secondary effectors during RNAi in C. elegans.Science 315(5809):241–244.
Pal C. 1998. Plasticity, memory and the adaptive land-scape of the genotype. Proceedings of the Royal Societyof London, Series B: Biological Sciences 265(1403):1319–1323.
Palauqui J. C., Elmayan T., Pollien J. M., Vaucheret H.1997. Systemic acquired silencing: transgene-specific post-transcriptional silencing is transmit-ted by grafting from silenced stocks to non-silenced scions. EMBO Journal 16(15):4738–4745.
Pantaleon M., Whiteside E. J., Harvey M. B., BarnardR. T., Waters M. J., Kaye P. L. 1997. Functionalgrowth hormone (GH) receptors and GH are ex-pressed by preimplantation mouse embryos: a rolefor GN in early embryogenesis? Proceedings of theNational Academy of Sciences USA 94(10):5125–5130.
Pavelka J., Koudelova J. 2001. Inheritance of atemperature-modified phenotype of the short an-tennae (sa) mutation in a moth, Ephestia kuehniella(Lepidoptera: Pyralidae). Journal of Heredity 92(3):234–242.
June 2009 173TRANSGENERATIONAL EPIGENETIC INHERITANCE

Pepper J. W., Sprouffske K., Maley C. C. 2007. Animalcell differentiation patterns suppress somatic evo-lution. PloS Computational Biology 3(12):e250.
Petronis A. 2004. The origin of schizophrenia: geneticthesis, epigenetic antithesis, and resolving synthe-sis. Biological Psychiatry 55(10):965–970.
Petronis A. 2006 Epigenetics and twins: three variationson the theme. Trends in Genetics 22(7):347–350.
Pigliucci M. 2001. Phenotypic Plasticity: Beyond Natureand Nurture. Baltimore (MD): John Hopkins Uni-versity Press.
Pigliucci M., Murren C. J., Schlichting C. D. 2006.Phenotypic plasticity and evolution by genetic as-similation. Journal of Experimental Biology 209(12):2362–2367.
Pikaard C. S. 2000. The epigenetics of nucleolar dom-inance. Trends in Genetics 16(11):495–500.
Pikaard C. S. 2003. Nucleolar dominance. Pages 399–402 in Nature Encyclopedia of the Human Genome,Volume 4, edited by D. N. Cooper. London (UK):Nature Publishing Group.
Polo S. E., Roche D., Almouzni G. 2006. New histoneincorporation marks sites of UV repair in humancells. Cell 127(3):481–493.
Popova N. K. 2006. From genes to aggressive behav-ior: the role of serotonergic system. BioEssays28(5):495–503.
Prusiner S. B. 1998. Prions. Proceedings of the NationalAcademy of Sciences USA 95(23):13363–13383.
Rando O. J., Verstrepen K. J. 2007. Timescales ofgenetic and epigenetic inheritance. Cell 128(4):655–668.
Rapp R. A., Wendel J. F. 2005. Epigenetics and plantevolution. New Phytologist 168(1):81–91.
Rassoulzadegan M., Grandjean V., Gounon P., Vin-cent S., Gillot I., Cuzin F. 2006. RNA-mediatednon-mendelian inheritance of an epigeneticchange in the mouse. Nature 441(7092):469–474.
Regev A., Lamb M. J., Jablonka E. 1998. The role ofDNA methylation in invertebrates: developmentalregulation or genome defense? Molecular Biologyand Evolution 15(7):880–891.
Richards E. J. 2006. Inherited epigenetic variation—revisiting soft inheritance. Nature Reviews Genetics7(5):395–401.
Richerson P. J., Boyd R. 2005. Not by Genes Alone: HowCulture Transformed Human Evolution. Chicago(IL): University of Chicago Press.
Ringrose L., Paro R. 2004. Epigenetic regulation ofcellular memory by the polycomb and trithoraxgroup proteins. Annual Review of Genetics 38:413–443.
Ringrose L., Paro R. 2007. Polycomb/trithorax re-sponse elements and epigenetic memory of cellidentity. Development 134(2):223–232.
Roberts B. T., Wickner R. B. 2003. Heritable activity:
a prion that propagates by covalent autoactivation.Genes and Development 17(17):2083–2087.
Rodin S. N., Parkhomchuk D. V., Riggs A. D. 2005.Epigenetic changes and repositioning determinethe evolutionary fate of duplicated genes. Biochem-istry (Moscow) 70(5):559–567.
Ruden D. M., Xiao L., Garfinkel M. D., Lu X. 2005.Hsp90 and environmental impacts on epigeneticstates: a model for the trans-generational effects ofdiethylstibesterol on uterine development andcancer. Human Molecular Genetics 14(Review Issue1):R149–R155.
Ruvinsky A. O., Lobkov Y. I., Belyaev D. K. 1983a.Spontaneous and induced activation of genes af-fecting the phenotypic expression of glucose6-phosphate dehydrogenase in Daphnia pulex. 1.Intraclonal variations in the electrophoretic mo-bility of G6PD. Molecular & General Genetics 189(3):485–489.
Ruvinsky A. O., Lobkov Y. I., Belyaev D. K. 1983b.Spontaneous and induced activation of genes af-fecting the phenotypic expression of glucose6-phosphate dehydrogenase in Daphnia pulex 2.Glucose-induced changes in the electrophoreticmobility of G6PD. Molecular & General Genetics 189:490–494.
Salmon A., Ainouche M. L., Wendel J. F. 2005. Ge-netic and epigenetic consequences of recent hy-bridization and polyploidy in Spartina (Poaceae).Molecular Ecology 14(4):1163–1175.
Sandman K., Pereira S. L., Reeve J. N. 1998. Diversityof prokaryotic chromosomal proteins and the or-igin of the nucleosome. Cellular and Molecular LifeSciences 54(12):1350–1364.
Sangster T. A., Lindquist S., Queitsch C. 2004. Undercover: causes, effects and implications of Hsp90-mediated genetic capacitance. BioEssays 26(4):348–362.
Sano H., Kamada I., Youssefian S., Katsumi M.,Wabiko H. 1990. A single treatment of rice seed-lings with 5-azacytidine induces heritable dwarf-ism and undermethylation of genomic DNA. Mo-lecular and General Genetics 220(3):441–447.
Schacter D. L. 2001. Forgotten Ideas, Neglected Pioneers:Richard Semon and the Story of Memory. Philadelphia(PA): Psychology Press.
Schermelleh L., Haemmer A., Spada F., Rosing N.,Meilinger D., Rothbauer U., Cardoso M., Leon-hardt H. 2007. Dynamics of Dnmt1 interactionwith the replication machinery and its role inpostreplicative maintenance of DNA methylation.Nucleic Acids Research 35(13):4301–4312.
Schlichting C. D., Pigliucci M. 1998 Phenotypic Evolu-tion: A Reaction Norm Perspective. Sunderland (MA):Sinauer.
Schmalhausen I. I. 1949. Factors of Evolution: The The-
174 Volume 84THE QUARTERLY REVIEW OF BIOLOGY

ory of Stabilizing Selection. Philadelphia (PA): Black-iston.
Schuettengruber B., Chourrout D., Vervoort M., Le-blanc B., Cavalli G. 2007. Genome regulation bypolycomb and trithorax proteins. Cell 128(4):735–745.
Shapiro J. A. 1999. Genome system architecture andnatural genetic engineering in evolution. Annals ofthe New York Academy of Sciences 870:23–35.
Shiao Y. H., Crawford E. B., Anderson L. M., Patel P.,Ko K. 2005. Allele-specific germ cell epimutationin the spacer promoter of the 45S ribosomal RNAgene after Cr(III) exposure. Toxicology and AppliedPharmacology 205(3):290–296.
Shiu P. K. T., Raju N. B., Zickler D., Metzenberg R. L.2001. Meiotic silencing by unpaired DNA. Cell107(7):905–916.
Shorter J., Lindquist S. 2005. Prions as adaptive con-duits of memory and inheritance. Nature ReviewsGenetics 6(6):435–450.
Siegal M. L., Bergman A. 2006. Canalization. Pages235–251 in Evolutionary Genetics, edited by C. W.Fox and J. B. Wolf. New York: Oxford UniversityPress.
Smits W. K., Kuipers O. P., Veening J. W. 2006. Phe-notypic variation in bacteria: the role of feedbackregulation. Nature Reviews Microbiology 4(4):259–271.
Sokolov V. A. 2006. Imprinting in plants. Russian Jour-nal of Genetics 42(9):1043–1052.
Steele E. J., Lindley R. A., Blanden R. V. 1998.Lamarck’s Signature: How Retrogenes are ChangingDarwin’s Natural Selection Paradigm. Reading (MA):Perseus Books.
Talbert P. B., Bryson T., Henikoff S. 2004. Adaptiveevolution of centromere proteins in plants andanimals. Journal of Biology 3:18.
True H. L., Lindquist S. L. 2000. A yeast prion pro-vides a mechanism for genetic variation and phe-notypic diversity. Nature 407(6803):477–483.
True H. L., Berlin I., Lindquist S. L. 2004. Epigeneticregulation of translation reveals hidden geneticvariation to produce complex traits. Nature431(7005):184–187.
Trut L. N., Plyusnina I. Z., Oskina I. N. 2004. Anexperiment on fox domestication and debatableissues of evolution of the dog. Russian Journal ofGenetics 40(6):644–655.
Tuite M. F., Cox B. S. 2006. The [PSI�] prion of yeast:a problem of inheritance. Methods 39(1):9–22.
Turner J. M. A., Mahadevaiah S. K., Fernandez-Capetillo O., Nussenzweig A., Xu X. L., DengC.-X., Burgoyne P. S. 2005. Silencing of unsyn-apsed meiotic chromosomes in the mouse. NatureGenetics 37(1):41–47.
Uptain S. M., Lindquist S. 2002. Prions as protein-
based genetic elements. Annual Review of Microbi-ology 56:703–741.
Van der Heijden G. W., Derijck A., Ramos L., GieleM., Van der Vlag J., de Boer P. 2006. Transmissionof modified nucleosomes from the mouse malegermline to the zygote and subsequent remodel-ing of paternal chromatin. Developmental Biology298(2):458–469.
Vanyushin B. F. 2006a. DNA methylation and epige-netics. Russian Journal of Genetics 42(9):985–997.
Vanyushin B. F. 2006b. DNA methylation in plants.Current Topics in Microbiology and Immunology 301:67–122.
Vanyushin B. F., Nemirovsky L. E., Klimenko V. V.,Vasiliev V. K., Belozersky A. N. 1973. The5-methylcytosine in DNA of rats. Tissue and agespecificity and the changes induced by hydrocor-tisone and other agents. Gerontologia 19(3):138–152.
Vastenhouw N. L., Brunschwig K., Okihara K. L., Mul-ler F., Tijsterman M., Plasterk R. H. A. 2006. Long-term gene silencing by RNAi. Nature 442(7105):882.
Vesela J., Rehak P., Mihalik J., Czikkova S., Pokorny J.,Koppel J. 2003. Expression of serotonin receptorsin mouse oocytes and preimplantation embryos.Physiological Research 52(2):223–228.
Vrana P. B., Fossella J. A., Matteson P., del Rio T.,O’Neill M. J., Tilghman S. M. 2000. Genetic andepigenetic incompatibilities underlie hybrid dys-genesis in Peromyscus. Nature Genetics 25(1):120–124.
Waddington C. H. 1957. The Strategy of the Genes: ADiscussion of Some Aspects of Theoretical Biology. Lon-don (UK): Allen & Unwin.
Waddington C. H. 1968. The basic ideas of biology.Pages 1–32 in Towards a Theoretical Biology: Prolego-mena, edited by C. H. Waddington. Edinburgh(UK): Edinburgh University Press.
Waddington C. H. 1975. The Evolution of an Evolution-ist. Edinburgh (UK): Edinburgh University Press.
Waterland R. A., Travisano M., Tahiliani K. G. 2007.Diet-induced hypermethylation at agouti viable yel-low is not inherited transgenerationally throughthe female. FASEB Journal 21(12):3380–3385.
Waterland R. A., Travisano M., Tahiliani K. G.,Rached M. T., Mirza S. 2008. Methyl donor sup-plementation prevents transgenerational amplifi-cation of obesity. International Journal of Obesity 32:1373–1379.
Waters E. R., Schaal B. A. 1996. Heat shock induces aloss of rRNA-encoding DNA repeats in Brassicanigra. Proceedings of the National Academy of SciencesUSA 93(4):1449–1452.
Weaver I. C. G., Cervoni N., Champagne F. A.,D’Alessio A. C., Sharma S., Seckl J. R., Dymov S.,Szyf M., Meaney M. J. 2004. Epigenetic program-
June 2009 175TRANSGENERATIONAL EPIGENETIC INHERITANCE

ming by maternal behavior. Nature Neuroscience7(8):847–854.
West-Eberhard M. J. 2003. Developmental Plasticity andEvolution. New York: Oxford University Press.
Wickner R. B. 1994. [URE3] as an altered URE2protein: evidence for a prion analog in Saccharo-myces cerevisiae. Science 264(5158):566–569.
Wickner R. B., Edskes H. K., Ross E. D., Pierce M. M.,Baxa U., Brachmann A., Shewmaker F. 2004.Prion genetics: new rules for a new kind of gene.Annual Review of Genetics 38:681–707.
Wolf J. B., Hager R. 2006. A maternal–offspring co-adaptation theory for the evolution of genomicimprinting. PLoS Biology 4(12):2238–2243.
Wu C.-T., Morris J. R. 2001. Genes, genetics, andepigenetics: a correspondence. Science 293(5532):1103–1105.
Wu T. C. J., Wang L., Wan Y.-J. Y. 1992. Expression ofestrogen receptor gene in mouse oocyte and dur-ing embryogenesis. Molecular Reproduction and De-velopment 33(4):407–412.
Yi H., Riddle N. C., Stokes T. L., Woo H.-R., RichardsE. J. 2004. Induced and natural epigenetic varia-tion. Cold Spring Harbor Symposia on QuantitativeBiology 69:155–160.
Zacharioudakis I., Gligoris T., Tzamarias D. 2007. Ayeast catabolic enzyme controls transcriptionalmemory. Current Biology 17(23):2041–2046.
Zaratiegui M., Irvine D. V., Martienssen R. A. 2007. Noncoding RNAs and gene silencing. Cell 128(4):763–776.
Zhivotovsky L. A. 2002. A model of the early evolutionof soma-to-germline feedback. Journal of TheoreticalBiology 216(1):51–57.
Zoghbi H. Y., Beaudet A. L. 2007. Epigenetics andhuman disease. Pages 435–456 in Epigenetics, ed-ited by C. D. Allis et al. Cold Spring Harbor (NY):Cold Spring Harbor Laboratory Press.
Zordan R. E., Galgoczy D. J., Johnson A. D. 2006. Epi-genetic properties of white-opaque switching in Can-dida albicans are based on a self-sustaining transcrip-tional feedback loop. Proceedings of the NationalAcademy of Sciences USA 103(34):12807–12812.
Zuckerkandl E., Cavalli G. 2007. Combinatorial epi-genetics, “junk DNA,” and the evolution of com-plex organisms. Gene 390(1–2):232–242.
Zufall R. A., Robinson T., Katz L. A. 2005. Evolutionof developmentally regulated genome rearrange-ments in eukaryotes. Journal of Experimental Zoology,Part B: Molecular and Developmental Evolution304B(5):448–455.
176 Volume 84THE QUARTERLY REVIEW OF BIOLOGY



![JUDr. Branislav Jablonka, PhD. - uniba.sk · 2020. 6. 22. · ADF05 Jablonka, Branislav [UKOPAHP] (100%) : Podpis starostu na zmenke z pohľadu súkromného a ústavného práva v](https://img.dokumen.tips/doc/110x75/60c51f9f19cf8307a4538382/judr-branislav-jablonka-phd-unibask-2020-6-22-adf05-jablonka-branislav.jpg)

























