Embed Size (px)
Citation preview

Transcriptional Regulation of Sorghum DefenseDeterminants against a Phloem-Feeding Aphid1
Keyan Zhu-Salzman*, Ron A. Salzman, Ji-Eun Ahn, and Hisashi Koiwa
Departments of Entomology (K.Z.-S., J.-E.A.), Biochemistry and Biophysics (R.A.S.), and HorticulturalSciences (H.K.), Texas A&M University, College Station, Texas 77843
When attacked by a phloem-feeding greenbug aphid (Schizaphis graminum), sorghum (Sorghum bicolor) activates jasmonicacid (JA)- and salicylic acid (SA)-regulated genes, as well as genes outside known wounding and SA signaling pathways.A collection of 672 cDNAs was obtained by differential subtraction with cDNAs prepared from sorghum seedlings infestedby greenbug aphids and those from uninfested seedlings. Subsequent expression profiling using DNA microarray andnorthern-blot analyses identified 82 transcript types from this collection responsive to greenbug feeding, methyl jasmonate(MeJA), or SA application. DNA sequencing analyses indicated that these encoded proteins functioning in direct defense,defense signaling, oxidative burst, secondary metabolism, abiotic stress, cell maintenance, and photosynthesis, as well asproteins of unknown function. In response to insect feeding, sorghum increased transcript abundance of numerous defensegenes, with some SA-dependent pathogenesis-related genes responding to greenbug more strongly than to SA. In contrast,only weak induction of MeJA-regulated defense genes was observed after greenbug treatment. However, infestation testsconfirmed that JA-regulated pathways were effective in plant defense against greenbugs. Activation of certain transcriptsexclusively by greenbug infestation was observed, and may represent unique signal transduction events independent of JA-and SA-regulated pathways. Results indicate that plants coordinately regulate defense gene expression when attacked byphloem-feeding aphids, but also suggest that aphids are able to avoid triggering activation of some otherwise potentiallyeffective plant defensive machinery, possibly through their particular mode of feeding.
The greenbug (Schizaphis graminum) has been a se-rious aphid pest on small grains in North Americasince its introduction over a century ago (Porter et al.,1997). Like many other aphids, greenbug inserts itsstylet into phloem sieve elements and consumesphloem sap as its food source (Miles, 1999; Burd,2002). It initially causes red or necrotic spots on cropssuch as sorghum (Sorghum bicolor) and wheat (Triti-cum aestivum), ultimately followed by general necro-sis and plant death (Porter et al., 1997; Miles, 1999).The use of resistant hybrids has been the major inte-grated pest management strategy for managinggreenbug on sorghum, however, very little is knownabout the molecular mechanisms of this resistance.
It has been established that plants respond to insectherbivory by activating an array of defense genes tomount resistance to insects. Much of the informationregarding plant defense gene regulation has beenderived from studies on chewing insects that crushand devour plant cells and tissues, as well as fromthe well-established wound signal transduction path-way (Ryan, 2000; de Bruxelles and Roberts, 2001;Kessler and Baldwin, 2002). Damage to cell mem-branes by mechanical wounding or insect feedingtriggers release of linolenic acid from membrane
phospholipids (Narvaez-Vasquez et al., 1999). Lin-olenic acid is converted, via the octadecanoid path-way, to jasmonic acid (JA), a key intracellular defensesignal. Accumulation of endogenous JA mediates ex-pression of various early defense signals and latefunctioning defense genes (Ryan, 2000). The systemicdefense response is mediated by transport of woundsignals from wounding sites to distant parts of theplant through vascular tissues (de Bruxelles and Rob-erts, 2001; Orozco-Cardenas et al., 2001). The best-characterized long-distance signal systemin, a smallpolypeptide, is transmitted via phloem in tomato(Lycopersicon esculentum; Ryan and Pearce, 1998). Per-ception of systemin by plant cells activates a defensesignal transduction pathway leading to biosynthesisof JA and expression of defense genes in systemictissues.
Despite the commonality between mechanicalwounding and insect feeding damage, differentialplant responses toward different wound stimuli ledto the conclusion that plants can distinguish damagedone mechanically from that done by insects. A DNAmicroarray study with 150 preselected defense-related Arabidopsis genes showed that caterpillar-elicited transcriptional changes differed from thoseof wounding by forceps. In particular, water stress-induced genes were less responsive to caterpillar(Pieris rapae) feeding (Reymond et al., 2000). Insectchewing also accelerated accumulation of wound-induced transcripts in potato (Solanum tuberosum)leaves (Korth and Dixon, 1997). Furthermore, unique
1 This work was partially supported by a grant from USDA-NRI, proposal no. 2000-02914.
* Corresponding author; e-mail [email protected]; fax979 – 862– 4790.
Article, publication date, and citation information can be foundat www.plantphysiol.org/cgi/doi/10.1104/pp.103.028324.
420 Plant Physiology, January 2004, Vol. 134, pp. 420–431, www.plantphysiol.org © 2004 American Society of Plant Biologists www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

to insect feeding, plants are able to initiate indirectdefense by synthesizing specific blends of volatilesthat attract natural enemies of the herbivores (Albornet al., 1997; Pare and Tumlinson, 1997; Schittko et al.,2001), and by forming neoplasmic tissues that im-pede larval entry into the plant tissue (Doss et al.,2000). Discrimination between mechanical woundingand insect herbivory is thought to be due to recog-nition of insect-derived elicitors by plant cells (Al-born et al., 1997; Korth and Dixon, 1997; Halitschke etal., 2001).
Considerably less information is available on mo-lecular mechanisms of plant response to aphids, thelargest group of phloem-feeding insects. A commonfeature among reported studies on plant-aphid inter-actions is that plants activate genes known to mountdefenses against bacterial and fungal pathogens (Fi-dantsef et al., 1999; Moran and Thompson, 2001;Moran et al., 2002). Such a response is generally notstimulated by chewing insects (Reymond et al., 2000).The defense response against pathogens is accompa-nied by elevated cellular salicylic acid (SA) concen-trations and increased expression of pathogenesis-related (PR) proteins that confer disease resistance.Production of reactive oxygen species (ROS), includ-ing H2O2, is one of the earliest plant responses inincompatible interactions between pathogens andplants. ROS induce the accumulation of SA and trig-ger PR protein expression (Wu et al., 1997; Chamnon-gpol et al., 1998). SA is an essential signaling hor-mone for activation of local and systemic defensesagainst pathogens in many plant species (Cao et al.,1998; Dempsey et al., 1999; Zhang et al., 1999). Ara-bidopsis mutants deficient in SA production or signaltransduction exhibit altered disease resistance. NPR1(nonexpresser of PR genes) is a key regulator intransducing the SA signal (Cao et al., 1997). It con-tains an ankyrin-repeat domain involved in protein-protein interactions. Mutations in this domain resultin loss of resistance, whereas overexpression of NPR1gene leads to broad-spectrum disease resistance (Caoet al., 1998). Lipase-like proteins, EDS1 and PAD4,directly interact with each other to control SA accu-mulation, and EDS1 is also engaged early in thehypersensitive response (Feys et al., 2001). Mutantplants eds1-2 and pad4-2 showed reduced SA anddecreased EDS1/PAD4-dependent R gene-mediatedresponse. SA has been shown to suppress woundsignaling and JA function (Doares et al., 1995). How-ever, SA is not the essential mobile signal transmittedthrough plants to initiate systemic defense (Alvarezet al., 1998; Wildermuth et al., 2001). Recent identifi-cation of a lipid transfer protein implies the nature ofthe long distant signaling molecule in Arabidopsismay be lipid derived (Maldonado et al., 2002). Trans-mission of the mobile signal is most likely carried outvia the phloem tissue (de Bruxelles and Roberts,2001).
Similarity in plant responses stimulated byphloem-feeding aphids and by pathogens could bedue to the similar effects insect stylets and fungalhyphae have on their hosts during insect feeding andpathogen infection (Fidantsef et al., 1999). The pathsof aphid stylets penetrating plant epidermal and pa-renchymal cells to reach phloem sieves are mainlyintercellular or intramural (Miles, 1999). Such a feed-ing style results in limited plant damage that is dis-tinct from that of chewing insects. Thus, it is notunexpected that plants activate disparate responseswhen attacked by chewing versus phloem-feedinginsects. However, using the Arabidopsis JA-insensitivemutant coi1-1, Moran and Thompson (2001) demon-strated involvement of the wounding pathway as wellas SA-regulated genes in response to aphids. Further-more, the whitefly (Bemisia argentifolii)-induced SLW3gene from squash did not respond to any knowndefense-signaling molecules, implying that novel de-fense regulation could be elicited by phloem-feedinginsects (van de Ven et al., 2000; Walling, 2000).
In this study, we aimed at understanding the tran-scriptional response of sorghum to infestation by thegreenbug aphid on a broad scale, and at comparingand contrasting this regulation with JA- and SA-regulated gene expression. We first generated acDNA collection enriched in greenbug-responsivesorghum genes by subtractive hybridization. Wethen performed high-throughput DNA microarrayanalyses to identify greenbug-regulated genes, andcompared greenbug-responsive transcript profileswith those after treatments by methyl jasmonate(MeJA) and SA. We also evaluated the effect ofMeJA-regulated defense on infestation by greenbugaphids. To our knowledge, this is the first targetedmicroarray analysis of monocot response againstphloem-feeding insects. Our results suggest thatgreenbug modulates MeJA- and SA-regulated geneexpression, an ability most likely linked to its specialmode of feeding.
RESULTS
Isolation of Defense-Related Genes in Sorghum
To obtain molecular profiles of plant response tophloem-feeding aphids, we used a combination ofsubtractive hybridization and cDNA microarraytechniques to detect greenbug-responsive genes insorghum. After forward and reverse subtractions ofcDNAs from seedlings infested and uninfested bygreenbug, a total of 672 cDNA fragments enriched ingreenbug-induced and -suppressed genes were ob-tained, and their PCR-amplified products were ar-rayed onto glass slides. To determine the roles of SAand JA pathways in regulation of aphid-responsivegenes, the cDNA microarrays were hybridized withprobe pairs prepared from total RNAs of greenbug-infested and -uninfested seedlings, as well as fromseedlings with or without MeJA or SA treatment. A
Sorghum Transcriptional Regulation against Aphids
Plant Physiol. Vol. 134, 2004 421 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

validated expression fold-change ratio cutoff of 1.5was applied to determine the cDNA populations thatwere differentially regulated by the various treat-ments (Fig. 1). When this threshold was used incontrol versus control validation experiments (i.e.Cy3- and Cy5-labeled probes were made from thesame input RNA and were cohybridized to arrays),3.2% of the array spots showed 1.5-fold or greatersignal differences (Fig. 1), i.e. about 20 cDNAs in thetotal collection could be potentially miscategorized.To ensure reproducibility, replicate microarrays foreach treatment were conducted with separate RNApreparations. Furthermore, selected clones showingaltered expression on microarrays were subjected tonorthern-blot analysis (see below). A total of 128cDNA clones exhibiting 1.5-fold or greater inductionor suppression by greenbug aphid, MeJA, or SAtreatment were analyzed by DNA sequencing, andwere found to form 82 contigs (Table I).
Coregulation of Defense-Related Genes
As shown in the Venn diagram, 23, 28, and 27genes were induced, respectively, by greenbug,MeJA, and SA (Fig. 2A; Table I). Among five genesinduced under all three treatments were a number ofwell-known defense-related genes such as variousprotease inhibitor genes. Three genes were inducedonly by aphid and MeJA, seven by aphid and SA, andseven by MeJA and SA, respectively (Fig. 2A). Like-wise, 10, 29, and 13 genes were down-regulated bygreenbug, MeJA, and SA, respectively. Three genes,two photosynthesis-related and one of unknownfunction, were suppressed by all treatments (Fig. 2B).Commonality observed in gene expression profilesamong different treatments suggests that response ofsorghum to aphid infestation is mediated, at leastpartly, by JA and SA signaling processes.
Characterization of Greenbug-ResponsiveGenes in Sorghum
The 82 greenbug-, MeJA-, and SA-responsivecDNA contigs can be categorized into eight groups
encoding proteins functioning in defense, defensesignaling, oxidative burst, secondary metabolism,abiotic stress, cell maintenance, and photosynthesis,as well as proteins of unknown function (Table I;Figs. 3–5).
Defense and Defense-Signaling Genes
The first four groups contain genes known to beinvolved in plant defense responses. Members en-coding defense proteins such as various PRs andprotease inhibitors, and genes in biosynthetic path-ways for phenolics such as flavonone 3�-hydroxylaseand O-methyltransferase (Lo and Nicholson, 1998;Frick and Kutchan, 1999), were up-regulated bygreenbug feeding. Most of them were also positivelyregulated by MeJA or SA (Table I). The cyanogenicglucoside dhurrin is a secondary metabolite whosedegradation products are known to be potent toxinsto herbivore insects (Tattersall et al., 2001). CYP71E1,the P450 gene functioning in the dhurrin biosyntheticpathway, was induced only at earlier time pointsafter greenbug treatment (Fig. 4), and DHUR encod-ing a dhurrin-degrading enzyme, although present inthe subtracted collection, did not respond to green-bug feeding as judged by microarray and northern-blot analyses (Table I; Fig. 4).
Genes encoding antioxidant proteins, such as glu-tathione S-transferase (GST), lactoylglutathionelyase, and catalase (CAT), were up- or down-regulated by greenbug infestation, suggesting thataccumulation and detoxification of ROS simulta-neously occur in greenbug-stressed sorghum seed-lings. Interestingly, CAT was suppressed by green-bug and MeJA, but was induced by SA, indicating adivergence of SA- and aphid-responsive pathways.
Abiotic Stress-Induced Genes
Greenbug infestation induced a drought-, salt-, andlow temperature-responsive gene (DRT) and an alde-hyde oxidase gene involved in abscisic acid (ABA)biosynthesis (Table I; Sekimoto et al., 1997). This
Figure 1. Signal intensity patterns of 672 cDNA fragments after microarray with cDNA probes from mRNA of untreatedversus treated (greenbug, MeJA, or SA) sorghum seedlings. The self versus self (control versus control RNA) array was carriedout by cohybridizing Cy3- and Cy5-labeled probes made from the same RNA (untreated seedlings). Values on x and y axesare signal intensity �10�4. Solid lines correspond to a 1:1 ratio of signal intensity. Upper dotted lines represent a 1.5-foldinduction, and lower dashed lines represent a 1.5-fold suppression of gene expression, respectively. The 1.5-fold ratiothresholds were experimentally established by self versus self experiments and repeatedly captured over 96% of all spots onthe microarray.
Zhu-Salzman et al.
422 Plant Physiol. Vol. 134, 2004 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

a cDNA Sequences of all clones listed in Table I have been submitted to the GenBank dbEST database. Accession nos.are CF675621 to CF675702. b Genes used for northern blot analyses are shown in boldface. c BLASTX searches wereconducted to determine homologous genes and the putative function of the cDNA fragments. The cutoff e-value used was10�3. For those sequences that did not result in any hits using BLASTX, BLASTN searches were employed. d Ratios ofsignal intensity were determined by cDNA microarray hybridization as described in Materials and Methods. Shown are themean � standard deviation of fluorescence ratios of two replicate microarray slides for comparisons of 48 h greenbuginfestation, 24 h MeJA or 24 h SA treatment with untreated control, respectively. Values are highlighted in gray if upregulated�1.5-fold in gene expression by the treatment, or underlined if down-regulated �1.5-fold (expressed as �0.67).
Table I. Genes differentially expressed in response to greenbug, MeJa and SAa
Sorghum Transcriptional Regulation against Aphids
Plant Physiol. Vol. 134, 2004 423 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

Table I. (Continued from previous page.)
Zhu-Salzman et al.
424 Plant Physiol. Vol. 134, 2004 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

implies that ABA and ABA-regulated genes also-played a role in sorghum response to greenbug feed-ing. It has been established that ABA does not di-rectly function in wound signal transduction but mayhelp to maintain healthy physiological conditionsnear wound sites to ensure wound response (Birken-meier and Ryan, 1998). Aphid feeding is known todecrease leaf water potential (Cabrera et al., 1994),
thus dehydration resulting from greenbug feedingmay be the driving force for induction of DRT. Amicroarray study showing that many wounding-inducible genes were also drought-inducible in Ara-bidopsis further supports that the role of wound-induced ABA is to respond to water stress (Reymondet al., 2000). SA increased mRNA levels of aldehydeoxidase (Table I), but it did not activate DRT, thedrought-responsive gene, further evidencing thataphid and SA differentially regulate subsets of stress-related genes.
Cell Maintenance Genes
Among numerous cell maintenance genes, a nitritereductase gene (NiR) functioning in nitrogen-assimilation was significantly induced by greenbuginfestation (Table I). Feeding directly from phloemsap allows aphids to get access to free amino acidsthat do not require protein digestion before assimi-lation. This withdrawal could signal depletion of thenitrogen metabolites Asn and Gln (Sivasankar et al.,1997), leading to the observed induction of NiR.
Genes Involved in Photosynthesis
Photosynthesis-related genes were suppressedstrongly by MeJA, and to a lesser extent by SA andaphids (Table I). Suppression of photosynthesis by JAhas been established (Creelman and Mullet, 1997),and an inverse correlation between photosynthesis-and defense-related gene regulation has also beenobserved in plants subjected to fungal infection, elic-itor treatment, or insect herbivory (Kombrink andHahlbrock, 1990; Hermsmeier et al., 2001). This re-sponse presumably allows energy reallocation to de-fense responses, with suppression of less importantfunctions, upon attack by insects and pathogens.
Figure 2. Venn diagrams showing numbers of overlapping andunique genes induced (A) and suppressed (B) by greenbug (GB),MeJA, or SA at 1.5-fold or higher. Results were based on the meaninductions of two experimental replicates.
Figure 3. Greenbug modulates SA-induced PR genes. Total RNA wasextracted from untreated and mechanical wounding-, MeJA-,greenbug-, or SA-treated sorghum seedlings at various time pointsindicated. cDNA fragments encoding thaumatin-like proteins(THAU1 and THAU2), a chitinase (CHIT1), and PR10 (PR10) werelabeled as probes. C, Untreated control; W, mechanically wounded;J, MeJA treated; GB, greenbug infested; S, SA treated. A representa-tive ethidium bromide-stained total RNA gel is shown to demonstraterelative RNA loading.
Figure 4. Greenbug avoids activation of MeJA-induced defensegenes. Total RNA was extracted from untreated and greenbug-,mechanical wounding-, MeJA-, or SA-treated sorghum seedlings atvarious time points indicated. cDNA fragments encoding a lipooxy-genase (LOX), a cytochrome P450 gene involved in dhurrin biosyn-thesis (CYP71E1), dhurrinase (DHUR), and a Bowman-Birk proteaseinhibitor (BBPI2) were labeled as probes, respectively. C, Untreatedcontrol; W, mechanically wounded; J, MeJA treated; GB, greenbuginfested; S, SA treated.
Sorghum Transcriptional Regulation against Aphids
Plant Physiol. Vol. 134, 2004 425 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

Genes of Unknown Function
Nineteen cDNAs did not match any sequences inGenBank, or matched genes encoding proteins withunknown function. Among the seven greenbug-responsive cDNAs, 2C1 (induced) and 4H3 (sup-pressed) were greenbug specific. Others were co-regulated with MeJA and/or SA.
Greenbug Infestation Activates SA-Induced DefenseGenes Surpassing SA
Strong induction of SA-regulated PR genes, includ-ing chitinase (CHIT), glucanase (BGL), thaumatin-likeproteins (THAU), and PR10 by greenbug infestationsuggested the involvement of an SA signaling path-way. To determine dynamics of PR gene expressionin response to mechanical wounding, greenbug in-festation, and MeJA and SA signals, selected cDNAswere used as probes for northern-blot analyses atmultiple time points. Transcripts of THAUs were in-duced as soon as 6 h after greenbug challenge, com-pared with 72 h after SA treatment. However, a chiti-nase mRNA reached a peak sooner under SAtreatment than under greenbug infestation, and PR10showed a similar expression pattern over time inboth treatments (Fig. 3). The stronger, and in somecases, more rapid response of PR genes to greenbugthan to SA indicated that greenbug was able touniquely modulate some genes in the SA-responsepathway.
Limited Induction of JA-Regulated Defense Genes byGreenbug Aphid Infestation
A time-course northern-blot analysis of gene ex-pression was also conducted for MeJA up-regulatedgenes encoding a LOX, a cytochrome P-450(CYP71E1), DHUR, and a protease inhibitor (BBPI2;Fig. 4). In contrast to the high induction of PR genesby greenbug, the aphid only marginally and tempo-rarily induced JA-regulated defense genes. It is pos-sible that the SA signaling pathway elicited by green-bug suppressed MeJA-responding genes, as in thecase of CYP71E1 and DHUR (Fig. 4). However, itshould be noted that SA also induced the two JA-induced genes, LOX and BBPI2 (Fig. 4), suggesting
that the two pathways are not always exclusive. Analternative explanation for limited expression of JA-induced genes is that tissue damage caused by aphidsmay not generate sufficiently high endogenous JAlevels. Interestingly, however, extensive mechanicalwounding also did not result in significant inductionof JA-responsive genes in most cases (Fig. 4).
Genes Specifically Induced by Greenbug Infestation
cDNA microarray analysis identified eight genessolely responsive to greenbug infestation. However,the single time point selected for the greenbug mi-croarray experiment may not always fall within timeperiod of gene induction by other treatments.THAUs, for instance, responded to greenbug and SA,but SA induction of THAUs (beginning at 72 h, asshown by northern blots) was not reflected by SAmicroarray experiment data at 24 h (Table I; Fig. 3).On the other hand, northern-blot analyses at multipletime points indicated the Leu-rich repeat-containingprotein (SLRR) and a DRP gene were greenbug spe-cific over the entire time course (Fig. 5). SLRR hasbeen postulated to be a signaling molecule involvedin plant-pathogen signal transduction (Hipskind etal., 1996). Accumulation of this transcript could bedetected within several hours after fungal infection(Hipskind et al., 1996) or greenbug infestation (Fig.5), suggesting fungi and phloem-feeding insects mayhave activated at least partly overlapping signalingcascades in sorghum. The second greenbug-specificgene encoded a protein homologous to a maize (Zeamays) DRP, also known to be induced by sugar star-vation (Chevalier et al., 1995). By feeding on phloemsap that contains high concentrations of sugars,aphids could potentially be disturbing sorghum’ssource-sink metabolism. This assumption is sup-ported by the observation of a 4-fold decrease ofsoluble carbohydrates in greenbug-infested barley(Hordeum vulgare) seedlings relative to uninfestedcontrol seedlings (Cabrera et al., 1994). Thus, the DRPgene could be regulated by a sugar-signaling path-way that has been shown to crosstalk with defenseregulatory pathways (Roitsch, 1999).
MeJA-Induced Plant Defense Response DetersGreenbug Infestation
To directly test whether the MeJA-induced defenseresponse has any impact on greenbug infestation, weconducted a choice test where greenbugs wereequally exposed to MeJA-treated sorghum seedlingsand untreated control seedlings and were allowed tofreely choose which plants to infest. The frequency ofgreenbug infestation on control plants was signifi-cantly higher than that on the MeJA-treated plants(chi square � 343.6, df � 1, P � 0.0001). The meanproportion of aphids observed on control plants was0.647 � 0.007 (sd) compared with on MeJA plants,
Figure 5. Transcripts of a defense-related protein (DRP) and anLRR-containing protein (SLRR) respond specifically to greenbug in-festation. Total RNA was extracted from untreated and greenbug-,mechanical wounding-, MeJA-, or SA-treated sorghum seedlings atvarious time points indicated. cDNA fragments DRP and SLRR werelabeled as probes. C, untreated control; W, mechanically wounded;J, MeJA treated; GB, greenbug infested; S, SA treated.
Zhu-Salzman et al.
426 Plant Physiol. Vol. 134, 2004 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.
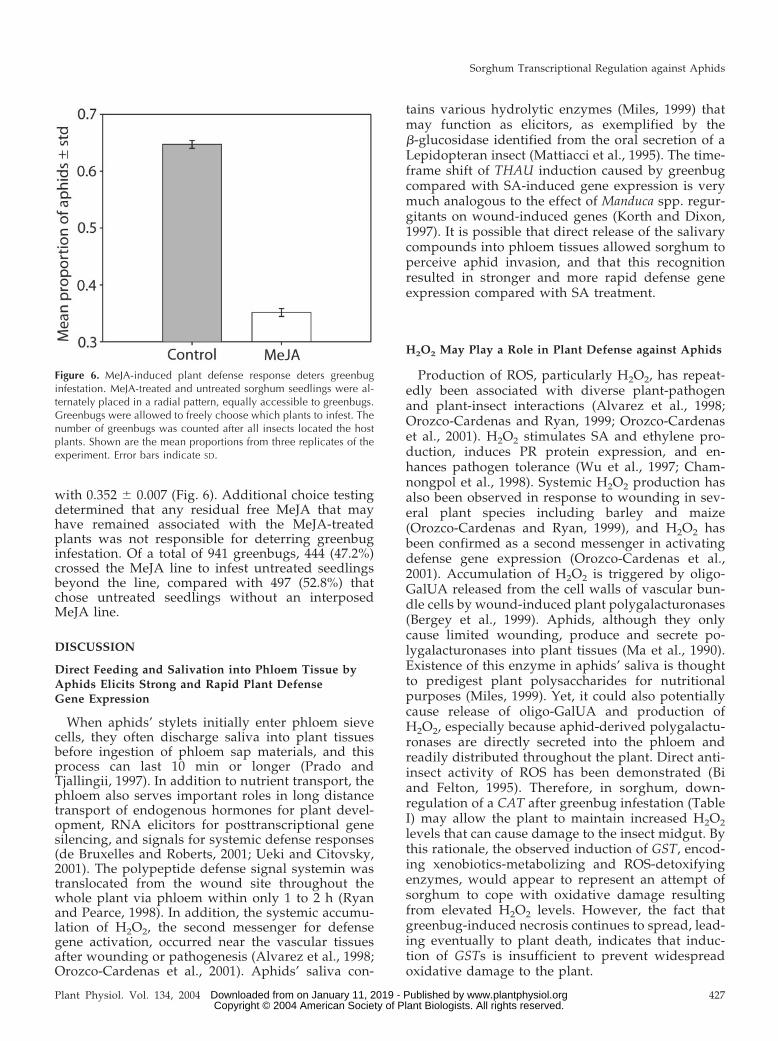
with 0.352 � 0.007 (Fig. 6). Additional choice testingdetermined that any residual free MeJA that mayhave remained associated with the MeJA-treatedplants was not responsible for deterring greenbuginfestation. Of a total of 941 greenbugs, 444 (47.2%)crossed the MeJA line to infest untreated seedlingsbeyond the line, compared with 497 (52.8%) thatchose untreated seedlings without an interposedMeJA line.
DISCUSSION
Direct Feeding and Salivation into Phloem Tissue byAphids Elicits Strong and Rapid Plant DefenseGene Expression
When aphids’ stylets initially enter phloem sievecells, they often discharge saliva into plant tissuesbefore ingestion of phloem sap materials, and thisprocess can last 10 min or longer (Prado andTjallingii, 1997). In addition to nutrient transport, thephloem also serves important roles in long distancetransport of endogenous hormones for plant devel-opment, RNA elicitors for posttranscriptional genesilencing, and signals for systemic defense responses(de Bruxelles and Roberts, 2001; Ueki and Citovsky,2001). The polypeptide defense signal systemin wastranslocated from the wound site throughout thewhole plant via phloem within only 1 to 2 h (Ryanand Pearce, 1998). In addition, the systemic accumu-lation of H2O2, the second messenger for defensegene activation, occurred near the vascular tissuesafter wounding or pathogenesis (Alvarez et al., 1998;Orozco-Cardenas et al., 2001). Aphids’ saliva con-
tains various hydrolytic enzymes (Miles, 1999) thatmay function as elicitors, as exemplified by the�-glucosidase identified from the oral secretion of aLepidopteran insect (Mattiacci et al., 1995). The time-frame shift of THAU induction caused by greenbugcompared with SA-induced gene expression is verymuch analogous to the effect of Manduca spp. regur-gitants on wound-induced genes (Korth and Dixon,1997). It is possible that direct release of the salivarycompounds into phloem tissues allowed sorghum toperceive aphid invasion, and that this recognitionresulted in stronger and more rapid defense geneexpression compared with SA treatment.
H2O2 May Play a Role in Plant Defense against Aphids
Production of ROS, particularly H2O2, has repeat-edly been associated with diverse plant-pathogenand plant-insect interactions (Alvarez et al., 1998;Orozco-Cardenas and Ryan, 1999; Orozco-Cardenaset al., 2001). H2O2 stimulates SA and ethylene pro-duction, induces PR protein expression, and en-hances pathogen tolerance (Wu et al., 1997; Cham-nongpol et al., 1998). Systemic H2O2 production hasalso been observed in response to wounding in sev-eral plant species including barley and maize(Orozco-Cardenas and Ryan, 1999), and H2O2 hasbeen confirmed as a second messenger in activatingdefense gene expression (Orozco-Cardenas et al.,2001). Accumulation of H2O2 is triggered by oligo-GalUA released from the cell walls of vascular bun-dle cells by wound-induced plant polygalacturonases(Bergey et al., 1999). Aphids, although they onlycause limited wounding, produce and secrete po-lygalacturonases into plant tissues (Ma et al., 1990).Existence of this enzyme in aphids’ saliva is thoughtto predigest plant polysaccharides for nutritionalpurposes (Miles, 1999). Yet, it could also potentiallycause release of oligo-GalUA and production ofH2O2, especially because aphid-derived polygalactu-ronases are directly secreted into the phloem andreadily distributed throughout the plant. Direct anti-insect activity of ROS has been demonstrated (Biand Felton, 1995). Therefore, in sorghum, down-regulation of a CAT after greenbug infestation (TableI) may allow the plant to maintain increased H2O2levels that can cause damage to the insect midgut. Bythis rationale, the observed induction of GST, encod-ing xenobiotics-metabolizing and ROS-detoxifyingenzymes, would appear to represent an attempt ofsorghum to cope with oxidative damage resultingfrom elevated H2O2 levels. However, the fact thatgreenbug-induced necrosis continues to spread, lead-ing eventually to plant death, indicates that induc-tion of GSTs is insufficient to prevent widespreadoxidative damage to the plant.
Figure 6. MeJA-induced plant defense response deters greenbuginfestation. MeJA-treated and untreated sorghum seedlings were al-ternately placed in a radial pattern, equally accessible to greenbugs.Greenbugs were allowed to freely choose which plants to infest. Thenumber of greenbugs was counted after all insects located the hostplants. Shown are the mean proportions from three replicates of theexperiment. Error bars indicate SD.
Sorghum Transcriptional Regulation against Aphids
Plant Physiol. Vol. 134, 2004 427 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

Are Plants Misled by Aphids?
It has been proposed that phloem-feeding insectsare perceived as pathogens due to similarities in themanner of penetration of plant tissues by fungalhyphae and aphid stylets, and to some extent, by thesimilar hydrolytic enzymes released during fungalgrowth and insect feeding (Fidantsef et al., 1999;Walling, 2000). In accordance with this view, weobserved a strong induction of SA-regulated PRgenes by greenbug feeding, but a weak induction ofwounding and JA-regulated genes. Although up-regulation of PR genes by several aphid species hasbeen reported, no direct role has been established forPR proteins in defense against phloem-feeding in-sects. Higher chitinase and glucanase activities didnot occur in greenbug-resistant sorghum (Krishnav-eni et al., 1999). Likewise, induction of SA-dependentresponses did not confer resistance to green peachaphid in Arabidopsis (Moran and Thompson, 2001).In contrast, control of insect pests including phloem-feeders by JA-regulated genes such as protease inhib-itors has been established (Rahbe and Febvay, 1993;Tran et al., 1997). In our study, significantly lessgreenbug infestation was observed on MeJA-pretreated seedlings, suggesting the effectiveness ofplant defense elicited by MeJA against aphid inva-sion. We also found CYP71E1, one of the sorghumcytochrome P450s involved in dhurrin biosynthesis(a cyanogenic glucoside derived from Tyr), to behighly induced by MeJA (Fig. 4). Upon damage ofplant tissue by insect herbivory, dhurrin is hydro-lyzed by DHUR to produce toxic hydrogen cyanide.Integration of dhurrin biosynthesis into Arabidopsisconferred resistance to flea beetle (Phyllotreta nemo-rum; Tattersall et al., 2001). Thus, activation of geneexpression involved in dhurrin biosynthesis and deg-radation could negatively impact aphids. Anothereffective anti-insect gene, LOX, regulates plant de-fense by its role in the octadecanoid pathway for JAbiosynthesis (Royo et al., 1999). Yet, greenbug onlyweakly induced the accumulation of these mRNAscompared with the levels of induction by MeJA.Thus, it appears that sorghum tended to activateineffective, but suppress effective, defense gene setswhen attacked by aphids. Supporting this conclu-sion, some aphids such as Aphis fabae accepted pre-infested wheat as host plants more readily than un-infested wheat (Prado and Tjallingii, 1997). It seemsaphid feeding not only avoided provoking plant de-fense, but also stimulated changes in phloem compo-nents that were nutritionally advantageous to aphidsthemselves.
Several scenarios for inadequate activation of de-fense gene expression are possible. First, minimaltissue damage by aphid infestation could produceinsufficient JA levels for gene induction. Second,strong induction of SA-dependent responses mayhave suppressed the JA-signaling pathway due topathway crosstalk (Doares et al., 1995). However, we
noticed that mechanically wounded sorghum seed-lings in our experiments, which exhibited significanttissue damage, also did not induce JA-responsivegenes. Thus, there is a third possibility that wound-induced endogenous ethylene may have suppressedJA-induced genes. This suppression has been ob-served in numerous plant species (Zhu-Salzman etal., 1998; Rojo et al., 1999; Stotz et al., 2000; Winz andBaldwin, 2001). Even though greenbug infestationentailed minimal wounding, infestation was found tocause ethylene production in barley (Argandona etal., 2001), and this ethylene evolution, elicited bytissue damage or by aphid salivary compounds,could potentially block JA-induced gene expression.
Plant response to herbivore and pathogen attacksoften involves the interaction of multiple signal path-ways that are controlled by a small number of globalsignals (Reymond and Farmer, 1998). However, it isquestionable whether the outcome of the crosstalkalways leads to the most effective plant defense forthe specific challenge. Unnecessary SA productionand SA-mediated defense responses were also elic-ited by a nonhost pathogen in Arabidopsis (van Weesand Glazebrook, 2003). In our study, the observedgene regulation profile in response to greenbug in-festation builds toward a case for subversion of po-tentially effective plant defense measures to a mis-guided anti-microbial defense pathway in sorghum.Aphid-vectored viruses and endosymbionts could beresponsible for this elicitation of pathogen-responsive gene expression. Infestation by whitefliesvectoring tomato mottle virus induced much stron-ger PR protein expression than the virus-free white-flies (McKenzie et al., 2002). Accordingly, higher ac-cumulation of PR proteins did not negatively impactthe whitefly reproduction. On the contrary, moreeggs and nymphs were associated with high PR-expressing plants (McKenzie et al., 2002). More stud-ies are needed to fully unravel phloem feedinginsect-host plant interactions; however, evidencefrom our study indicates the phloem-feeding insectsmay currently hold an important advantage.
MATERIALS AND METHODS
Growing Sorghum (Sorghum bicolor) Seedlings andRearing Greenbugs (Schizaphis graminum)
Sorghum seeds (ATx399 � RTx430) were planted in potting soil in plasticpots (4.5-inch diameter and 3.5-inch depth), and seedlings were grown in aninsect-free growth chamber (30°C, 60% relative humidity, and a 13-h light/11-h dark photoperiod). Two-week-old sorghum seedlings were used forgreenbug culturing, and infested plants were maintained in a separatechamber under the same growth conditions.
Plant Treatments with Mechanical Wounding, MeJA,SA, and Greenbug Infestation
One-week-old seedlings were used for all treatments. Wounding wasdone by pressing a rounded file against seedling leaves and stems until thewhole area above the soil of each seedling was covered by wounding spots.MeJA solution (200 �m) was sprayed onto seedling leaves to run-off with a
Zhu-Salzman et al.
428 Plant Physiol. Vol. 134, 2004 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

spray bottle. The plants were then grown under constant light in a sealedcontainer. Greenbugs were brushed onto seedlings at a density of approx-imately 20 per seedling. Before harvest, greenbugs were removed fromgreenbug-infesting seedlings by flushing the plants with deionized water.Seedlings were gently dried using paper towels before freezing in liquidnitrogen. Plant materials above the soil from wounding, MeJA, and green-bug treatments were harvested after 0-, 6-, 12-, 24-, and 48-h time points,wrapped in aluminum foil, frozen in liquid nitrogen, and stored at �80°Cuntil use. For the SA treatment, sorghum seedlings were grown hydropon-ically. Briefly, sorghum seeds were germinated for 3 d as described byFinlayson et al. (1998). The seedlings were then transferred into truncatedpipette tips placed in plastic pegboard sheets that were suspended overplastic buckets filled with 0.5� Hoagland solution. An airstone was used toaerate the growth medium. After an additional 5-d growth period with onemedium exchange, SA was added to a final concentration of 2 mm. Tissuesabove the roots were harvested at 0, 6, 24, 48, and 72 h, as described above.Approximately 20 seedlings were collected at each time point for eachtreatment. Control plants not subjected to treatments were also harvested ateach time point for each treatment.
Construction of the Subtractive cDNA Library
Sorghum seedlings infested with greenbugs for 48 h and control seed-lings (respectively) were ground into fine powder in liquid nitrogen. mR-NAs from each sample were isolated using a QuickPrep micro-mRNApurification kit (Amersham Pharmacia Biotech, Piscataway, NJ). The PCR-select cDNA subtraction kit (Clontech, Palo Alto, CA) was used to obtaincDNAs corresponding to genes differentially regulated by greenbug infes-tation. Forward and reverse subtractive hybridizations were performed. Theforward subtraction used tester cDNA obtained from mRNA of greenbug-infested seedlings and driver cDNA from control seedlings, and vice versain the reverse subtraction. Subtracted cDNAs were subjected to two roundsof PCR amplification to normalize cDNA populations. The PCR productswere ligated into pT-Adv vector (Clontech) and transformed into Escherichiacoli cells. Bacterial clones harboring cDNA inserts were inoculated, ampli-fied, and maintained in freezer medium [50 mm potassium phosphate, 2 mmsodium citrate, 0.8 mm MgSO4, and 7 mm (NH4)2SO4, pH 7.5] with carben-icillin (50 �g mL�1) in 96-well microplates.
DNA Microarrays and Probe Preparation
cDNA inserts of the subtracted collections were PCR amplified in 150-�Lreactions in microplate format, using M13 forward and M13 reverse uni-versal primers. PCR products confirmed by gel electrophoresis were pre-cipitated by addition of 15 �L of 3 m NaOAc, pH 5.2, and 150 �L ofisopropanol reaction�1 at �20°C overnight. Microplates were centrifuged at3,200 rpm for 2 h at 4°C. Pellets were washed with 70% (w/v) ethanol,vacuum dried, and resuspended in 35 �L of spotting solution (2� SSC and0.1% [w/v] Sarkosyl) each. DNA clones were printed onto poly-L Lys-coated glass slides (CEL Associates, Houston) using an Affymetrix 417arrayer. After printing, DNA was UV crosslinked at 550 mJ, and slides werefurther processed by blocking in 0.2% (w/v) SDS for 10 min at 25°C,followed by DNA denaturation in boiling water for 2 min, and treatmentwith �20°C ethanol (95%, w/v) for 2 min.
Microarray probes were synthesized from equal amounts of total RNAextracted from sorghum seedlings subjected to MeJA (24 h), SA (24 h), orgreenbug infestation (48 h), as well as from their respective controls, usingthe 3DNA expression array system (Genisphere, Hatfield, PA) according tomanufacturer’s instructions. Briefly, primers comprising oligo dT and aproprietary “capture sequence” for Cy3 or Cy5, respectively, were usedwith reverse transcriptase to synthesize cDNA probes. These probes (de-rived from 5 �g of input RNA channel�1 for each microarray) were thenmixed with an equal volume of hybridization buffer (Express-hyb; Clon-tech), added to the microarray and covered with a coverslip, and the slideswere sealed in aluminum hybridization chambers (Monterrey Industries,Monterrey, CA) for hybridization at 65°C overnight. After washing (2�SSC/0.1% [w/v] SDS for 10 min at 65°C, followed by 10 min in 2� SSC andthen 10 min in 0.2� SSC, both at 25°C with shaking), a second hybridizationwas conducted to incorporate Cy3 or Cy5 fluor, respectively, which wascoupled to an oligonucleotide complementary to the “capture sequence.”
Thus, cDNA probes bound to the microarray were fluor labeled byhybridization.
Data Acquisition and Analysis
Slides were scanned with a four-laser confocal scanner (Packard Scanar-ray 5000; Packard BioChip Technologies, Billerica, MA) using the Scanarrayprogram. Scanning parameters were adjusted to obtain balanced signals onthe two channels using the line-scan function. Image analysis was done withthe Quantarray Program (Packard BioScience, Downers Grove, IL). Datafrom both channels were background subtracted and normalized. Normal-ization factors were generated using spots composing the middle 75% ofsignal intensity values. Means and standard deviations of normalized andbackground-subtracted fold-change values from replicate experiments werederived in Excel (Microsoft, Redmond, WA), and are presented in Table I.For each given cDNA spot, gene expression was considered to be changedby treatments if the spot had an average fold change ratio of �1.50 or �0.67over the two treatments, and gave a signal intensity of �3,000 in one or bothof the two channels (control or treatment) in at least one replicate of at leastone treatment. Fold-cutoffs for valid induction and suppression ratios wereestablished by a series of self versus self microarray experiments in whichCy3- and Cy5-labeled probes made from the same RNA were cohybridizedto the DNA arrays. The labeling system used repeatedly gave a false-positive rate of 3% to 4% when using 1.5-fold induction and suppressionratio cutoffs and a signal intensity cutoff of 3,000. This characteristic behav-ior was also observed in self versus self tests using RNAs from diversetreatments (data not shown). A series of preliminary dye-swap experimentsdemonstrated that the labeling system used resulted in negligible dye bias,as has been reported elsewhere (Yu et al., 2002). Therefore, we adopted theconvention of using Cy5 fluor to label the control probe in all experiments.
DNA Sequencing and Data Analyses
Differentially expressed cDNAs were subjected to dideoxy terminatorcycle sequencing using the ABI BigDye sequencing kit (PE Biosystems,Foster City, CA), and were analyzed on an ABI Prism 3100 DNA sequencer(PE Biosystems). Sequencher software (Gene Codes, Ann Arbor, MI) wasused to trim the vector sequence from raw sequence data, and to assemblecontigs. cDNA identities were determined by sequence comparison with theGenBank database using BLASTX.
Northern-Blot Analyses and Signal Quantitation
Twenty micrograms of total RNA from each selected sample was sepa-rated on 1.2% (w/v) agarose formaldehyde gels, transferred to Hybond-Nnylon membrane, and hybridized with 32P-labeled cDNA probes. Blots werewashed (2� SSC/0.1% [w/v] SDS, 1� SSC/0.1% [w/v] SDS, and 0.1%SSC/0.1% SDS [w/v] at 65°C) and exposed to x-ray film. Ethidium bromide-stained total RNA gels were used to visually ensure even sample loading.
Infestation Choice Test
One-week-old sorghum seedlings were subjected to MeJA vapor treat-ment in a 10-L air-tight container for 15 h. Twenty microliters of 10� dilutedMeJA solution was applied to a 1- � 1-cm filter paper placed in the sealedcontainer. The final concentration of MeJA was 900 nmol L�1 air. After 15 h,the shoots from MeJA-treated and untreated plants were excised and eachwas placed in an Eppendorf tube filled with distilled water at the cut end.The tube openings were sealed around the stems with parafilm. This systemprotected the seedling from dehydration for several days. Several hundredgreenbug aphids were brushed onto the center of a large piece of filterpaper, surrounded in a radial pattern by 10 pairs of excised sorghumseedlings (MeJA-treated and untreated control) several centimeters away.Alternating MeJA-treated and untreated plants were equally accessible tothe aphids. After 5 to 8 h, all aphids were able to locate host plants,MeJA-treated, or untreated control. The number of aphids on each seedlingwas counted at the 8-h time point. The experiment was performed threetimes. Chi-square tests were used to determine statistical significance ofdifferences between treatments (Sokal and Rohlf, 1995). Count data weretransformed to proportions for visual display.
Sorghum Transcriptional Regulation against Aphids
Plant Physiol. Vol. 134, 2004 429 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

To determine whether any residual MeJA vapor potentially remainingassociated with the treated plants could deter greenbug infestation, un-treated sorghum shoots were arranged in two straight rows on a piece offilter paper, one row on either side of, and equidistant from, the greenbugintroduction zone. A thin line of MeJA (400 �m) was painted on the filterpaper in front of one of the two rows of seedlings, so that the insects wouldneed to cross the line to get to that row of plants. The greenbugs were thenimmediately introduced. Eight hours later, the number of aphids on eachseedling was counted, and total greenbugs on each of the two groups werecompared.
Distribution of Materials
The expressed sequence tag clones generated in this project will be madeavailable upon request for noncommercial research purposes.
Received June 10, 2003; returned for revision July 24, 2003; accepted October9, 2003.
LITERATURE CITED
Alborn T, Turlings TCJ, Jones TH, Stenhagen G, Loughrin JH, TumlinsonJH (1997) An elicitor of plant volatiles from beet armyworm oral secre-tion. Science 276: 945–949
Alvarez ME, Pennell RI, Meijer PJ, Ishikawa A, Dixon RA, Lamb C (1998)Reactive oxygen intermediates mediate a systemic signal network in theestablishment of plant immunity. Cell 92: 773–784
Argandona VH, Chaman M, Cardemil L, Munoz O, Zuniga GE, CorcueraLJ (2001) Ethylene production and peroxidase activity in aphid-infestedbarley. J Chem Ecol 27: 53–68
Bergey DR, Orozco-Cardenas M, de Moura DS, Ryan CA (1999) A wound-and systemin-inducible polygalacturonase in tomato leaves. Proc NatlAcad Sci USA 96: 1756–1760
Bi JL, Felton GW (1995) Foliar oxidative stress and insect herbivory: pri-mary compounds, secondary metabolites, and reactive oxygen species ascomponents of induced resistance. J Chem Ecol 21: 1511–1530
Birkenmeier GF, Ryan CA (1998) Wound signaling in tomato plants: evi-dence that ABA is not a primary signal for defense gene activation. PlantPhysiol 117: 687–693
Burd JD (2002) Physiological modification of the host feeding site by cerealaphids (Homoptera: Aphididae). J Econ Entomol 95: 463–468
Cabrera HM, Argandona VH, Corcuera LJ (1994) Metabolic changes inbarley seedlings at different aphid infestation levels. Phytochemistry 35:317–319
Cao H, Glazebrook J, Clarke JD, Volko S, Dong XN (1997) The ArabidopsisNPR1 gene that controls systemic acquired resistance encodes a novelprotein containing ankyrin repeats. Cell 88: 57–63
Cao H, Li X, Dong XN (1998) Generation of broad-spectrum disease resis-tance by overexpression of an essential regulatory gene in systemicacquired resistance. Proc Natl Acad Sci USA 95: 6531–6536
Chamnongpol S, Willekens H, Moeder W, Langebartels C, SandermannH, Van Montagu A, Inze D, Van Camp W (1998) Defense activation andenhanced pathogen tolerance induced by H2O2 in transgenic tobacco.Proc Natl Acad Sci USA 95: 5818–5823
Chevalier C, Bourgeois E, Pradet A, Raymond P (1995) Molecular cloningand characterization of 6 cDNAs expressed during glucose starvation inexcised maize (Zea mays L.) root tips. Plant Mol Biol 28: 473–485
Creelman RA, Mullet JE (1997) Biosynthesis and action of jasmonates inplants. Annu Rev Plant Physiol Plant Mol Biol 48: 355–381
de Bruxelles GL, Roberts MR (2001) Signals regulating multiple responsesto wounding and herbivores. Crit Rev Plant Sci 20: 487–521
Dempsey DA, Shah J, Klessig DF (1999) Salicylic acid and disease resis-tance in plants. Crit Rev Plant Sci 18: 547–575
Doares SH, Narvaez-Vasquez J, Conconi A, Ryan CA (1995) Salicylic acidinhibits synthesis of proteinase-inhibitors in tomato leaves induced bysystemin and jasmonic acid. Plant Physiol 108: 1741–1746
Doss RP, Oliver JE, Proebsting WM, Potter SW, Kuy SR, Clement SL,Williamson RT, Carney JR, DeVilbiss ED (2000) Bruchins: insect-derived plant regulators that stimulate neoplasm formation. Proc NatlAcad Sci USA 97: 6218–6223
Feys BJ, Moisan LJ, Newman MA, Parker JE (2001) Direct interactionbetween the Arabidopsis disease resistance signaling proteins, EDS1 andPAD4. EMBO J 20: 5400–5411
Fidantsef AL, Stout MJ, Thaler JS, Duffey SS, Bostock RM (1999) Signalinteractions in pathogen and insect attack: expression of lipoxygenase,proteinase inhibitor II, and pathogenesis-related protein P4 in the tomato,Lycopersicon esculentum. Physiol Mol Plant Pathol 54: 97–114
Finlayson SA, Lee IJ, Morgan PW (1998) Phytochrome B and the regulationof circadian ethylene production in sorghum. Plant Physiol 116: 17–25
Frick S, Kutchan TM (1999) Molecular cloning and functional expression ofO-methyltransferases common to isoquinoline alkaloid and phenylpro-panoid biosynthesis. Plant J 17: 329–339
Halitschke R, Schittko U, Pohnert G, Boland W, Baldwin IT (2001) Mo-lecular interactions between the specialist herbivore Manduca sexta (Lep-idoptera, Sphingidae) and its natural host Nicotiana attenuata: Fatty acid-amino acid conjugates in herbivore oral secretions are necessary andsufficient for herbivore-specific plant responses. Plant Physiol 125:711–717
Hermsmeier D, Schittko U, Baldwin IT (2001) Molecular interactions be-tween the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae)and its natural host Nicotiana attenuata: large-scale changes in the accu-mulation of growth- and defense-related plant mRNAs. Plant Physiol125: 683–700
Hipskind JD, Nicholson RL, Goldsbrough PB (1996) Isolation of a cDNAencoding a novel leucine-rich repeat motif from Sorghum bicolor inocu-lated with fungi. Mol Plant-Microbe Interact 9: 819–825
Kessler A, Baldwin IT (2002) Plant responses to insect herbivory: theemerging molecular analysis. Annu Rev Plant Biol 53: 299–328
Kombrink E, Hahlbrock K (1990) Rapid, systemic repression of the synthe-sis of ribulose 1,5-bisphosphate carboxylase small-subunit messenger-RNA in fungus-infected or elicitor-treated potato leaves. Planta 181:216–219
Korth KL, Dixon RA (1997) Evidence for chewing insect-specific molecularevents distinct from a general wound response in leaves. Plant Physiol115: 1299–1305
Krishnaveni S, Muthukrishnan S, Liang GH, Wilde G, Manickam A (1999)Induction of chitinases and �-1,3-glucanases in resistant and susceptiblecultivars of sorghum in response to insect attack, fungal infection andwounding. Plant Sci 144: 9–16
Lo SCC, Nicholson RL (1998) Reduction of light-induced anthocyaninaccumulation in inoculated sorghum mesocotyls: implications for a com-pensatory role in the defense response. Plant Physiol 116: 979–989
Ma R, Reese JC, Black WC, Bramel-Cox P (1990) Detection of pectinesteraseand polygalacturonase from salivary secretions of living greenbugs,Schizaphis graminum (Homoptera, Aphididae). J Insect Physiol 36:507–512
Maldonado AM, Doerner P, Dixon RA, Lamb CJ, Cameron RK (2002) Aputative lipid transfer protein involved in systemic resistance signallingin Arabidopsis. Nature 419: 399–403
Mattiacci L, Dicke M, Posthumus MA (1995) �-Glucosidase: an elicitor ofherbivore-induced plant odor that attracts host-searching parasiticwasps. Proc Natl Acad Sci USA 92: 2036–2040
McKenzie CL, Shatters RG, Doostdar H, Lee SD, Inbar M, Mayer RT(2002) Effect of geminivirus infection and Bemisia infestation on accu-mulation of pathogenesis-related proteins in tomato. Arch Insect Bio-chem Physiol 49: 203–214
Miles PW (1999) Aphid saliva. Biol Rev 74: 41–85Moran PJ, Cheng YF, Cassell JL, Thompson GA (2002) Gene expression
profiling of Arabidopsis thaliana in compatible plant-aphid interactions.Arch Insect Biochem Physiol 51: 182–203
Moran PL, Thompson GA (2001) Molecular responses to aphid feeding inArabidopsis in relation to plant defense pathways. Plant Physiol 125:1074–1085
Narvaez-Vasquez J, Florin-Christensen J, Ryan CA (1999) Positional spec-ificity of a phospholipase A activity induced by wounding, systemin, andoligosaccharide elicitors in tomato leaves. Plant Cell 11: 2249–2260
Orozco-Cardenas M, Ryan CA (1999) Hydrogen peroxide is generatedsystemically in plant leaves by wounding and systemin via the octade-canoid pathway. Proc Natl Acad Sci USA 96: 6553–6557
Orozco-Cardenas ML, Narvaez-Vasquez J, Ryan CA (2001) Hydrogen per-oxide acts as a second messenger for the induction of defense genes intomato plants in response to wounding, systemin, and methyl jasmonate.Plant Cell 13: 179–191
Zhu-Salzman et al.
430 Plant Physiol. Vol. 134, 2004 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.

Pare PW, Tumlinson JH (1997) De novo biosynthesis of volatiles induced byinsect herbivory in cotton plants. Plant Physiol 114: 1161–1167
Porter DR, Burd JD, Shufran KA, Webster JA, Teetes GL (1997) Greenbug(Homoptera: Aphididae) biotypes: selected by resistant cultivars or pre-adapted opportunists? J Econ Entomol 90: 1055–1065
Prado E, Tjallingii WF (1997) Effects of previous plant infestation on sieveelement acceptance by two aphids. Entomol Exp Appl 82: 189–200
Rahbe Y, Febvay G (1993) Protein toxicity to aphids: an in vitro test onAcyrthosiphon pisum. Entomol Exp Appl 67: 149–160
Reymond P, Farmer EE (1998) Jasmonate and salicylate as global signals fordefense gene expression. Curr Opin Plant Biol 1: 404–411
Reymond P, Weber H, Damond M, Farmer EE (2000) Differential geneexpression in response to mechanical wounding and insect feeding inArabidopsis. Plant Cell 12: 707–720
Roitsch T (1999) Source-sink regulation by sugar and stress. Curr OpinPlant Biol 2: 198–206
Rojo E, Leon J, Sanchez-Serrano JJ (1999) Cross-talk between wound sig-nalling pathways determines local versus systemic gene expression inArabidopsis thaliana. Plant J 20: 135–142
Royo J, Leon J, Vancanneyt G, Albar JP, Rosahl S, Ortego F, Castanera P,Sanchez-Serrano JJ (1999) Antisense-mediated depletion of a potatolipoxygenase reduces wound induction of proteinase inhibitors and in-creases weight gain of insect pests. Proc Natl Acad Sci USA 96: 1146–1151
Ryan CA (2000) The systemin signaling pathway: differential activation ofplant defensive genes. Biochim Biophys Acta 1477: 112–121
Ryan CA, Pearce G (1998) Systemin: a polypeptide signal for plant defen-sive genes. Annu Rev Cell Dev Biol 14: 1–17
Schittko U, Hermsmeier D, Baldwin IT (2001) Molecular interactions be-tween the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae)and its natural host Nicotiana attenuata: accumulation of plant mRNAs inresponse to insect-derived cues. Plant Physiol 125: 701–710
Sekimoto H, Seo M, Dohmae N, Takio K, Kamiya Y, Koshiba T (1997)Cloning and molecular characterization of plant aldehyde oxidase. J BiolChem 272: 15280–15285
Sivasankar S, Rothstein S, Oaks A (1997) Regulation of the accumulationand reduction of nitrate by nitrogen and carbon metabolites in maizeseedlings. Plant Physiol 114: 583–589
Sokal RR, Rohlf FJ (1995) Biometry. The Principles and Practice of Statisticsin Biological Research, Ed 3. W. H. Freeman and Company, New York
Stotz HU, Pittendrigh BR, Kroymann J, Weniger K, Fritsche J, Bauke A,Mitchell-Olds T (2000) Induced plant defense responses against chewinginsects: Ethylene signaling reduces resistance of Arabidopsis against Egyp-
tian cotton worm but not diamondback moth. Plant Physiol 124:1007–1017
Tattersall DB, Bak S, Jones PR, Olsen CE, Nielsen JK, Hansen ML, Hoj PB,Moller BL (2001) Resistance to an herbivore through engineered cyano-genic glucoside synthesis. Science 293: 1826–1828
Tran P, Cheesbrough TM, Keickhefer RW (1997) Plant proteinase inhibi-tors are potential anticereal aphid compounds. J Econ Entomol 90:1672–1677
Ueki S, Citovsky V (2001) RNA commutes to work: regulation of plant geneexpression by systemically transported RNA molecules. Bioessays 23:1087–1090
van de Ven WTG, LeVesque CS, Perring TM, Walling LL (2000) Local andsystemic changes in squash gene expression in response to silverleafwhitefly feeding. Plant Cell 12: 1409–1423
van Wees SCM, Glazebrook J (2003) Loss of non-host resistance of Arabi-dopsis NahG to Pseudomonas syringae pv. phaseolicola is due to degrada-tion products of salicylic acid. Plant J 33: 733–742
Walling LL (2000) The myriad plant responses to herbivores. J Plant GrowthRegul 19: 195–216
Wildermuth MC, Dewdney J, Wu G, Ausubel FM (2001) Isochorismatesynthase is required to synthesize salicylic acid for plant defence. Nature414: 562–565
Winz RA, Baldwin IT (2001) Molecular interactions between the specialistherbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural hostNicotiana attenuata: Insect-induced ethylene reduces jasmonate-inducednicotine accumulation by regulating putrescine N-methyltransferasetranscripts. Plant Physiol 125: 2189–2202
Wu GS, Shortt BJ, Lawrence EB, Leon J, Fitzsimmons KC, Levine EB,Raskin I, Shah DM (1997) Activation of host defense mechanisms byelevated production of H2O2 in transgenic plants. Plant Physiol 115:427–435
Yu J, Othman M, Farjo R, Zareparsi S, MacNee S, Yoshida S, Swaroop A(2002) Evaluation and optimization of procedures for target labeling andhybridization of cDNA microarrays. Mol Vision 8: 130–137
Zhang YL, Fan WH, Kinkema M, Li X, Dong XN (1999) Interaction of NPR1with basic leucine zipper protein transcription factors that bind se-quences required for salicylic acid induction of the PR-1 gene. Proc NatlAcad Sci USA 96: 6523–6528
Zhu-Salzman K, Salzman RA, Koiwa H, Murdock LL, Bressan RA, Hase-gawa PM (1998) Ethylene negatively regulates local expression of plantdefense lectin genes. Physiol Plant 104: 365–372
Sorghum Transcriptional Regulation against Aphids
Plant Physiol. Vol. 134, 2004 431 www.plantphysiol.orgon January 11, 2019 - Published by Downloaded from Copyright © 2004 American Society of Plant Biologists. All rights reserved.
![RESEARCH Open Access Genotypic variation in sorghum [Sorghum … · 2018-06-03 · RESEARCH Open Access Genotypic variation in sorghum [Sorghum bicolor (L.) Moench] exotic germplasm](https://img.dokumen.tips/doc/110x75/5f03a3af7e708231d40a0b15/research-open-access-genotypic-variation-in-sorghum-sorghum-2018-06-03-research.jpg)